Submitted:
23 December 2024
Posted:
24 December 2024
You are already at the latest version
Abstract
Ischemia-reperfusion (IR) injury is a major cause of multiple organ failure. The purpose of this study was to look into the role of Tephrosia purpurea (TEP) and its active constituent pseudosemiglabrin (PS) in allevi-ating severe acute pancreatitis and its associated acute lung injury. We established a rat pancreatic IR model. Rats were treated with TEP (200 mg/kg and 400 mg/kg), and PS (20 and 40 mg/kg). In addition to IR-control and sham groups. The results showed that the respiratory parameters including inspiratory time (Ti), expiratory time (Te), duration (Dr), and respiratory rate (RR) were comparable among all groups while peak inspiratory flow (PIF), forced vital capacity (FVC), and forced expiratory volume at 0.1 sec-onds (FEV0.1) were significantly impaired. Notably, PS at 40 mg/kg showed normal PIF, FVC and FEV0.1/FVC compared to IR group indicating an improved lung function. Additionally, TEP and PS showed protective effects on pancreatic and lung tissues by alleviating the pathological damage, reducing serum levels of trypsinogen activation peptide (TAP), lipase, and amylase, decreasing oxidative stress markers MDA, GPx, and MPO, suppressing inflammatory markers TNF-α, IL-6, and NF-κB, downregulating HMGB1 gene in pancreatic tissue, and upregulating IL-22 and Th22 genes in lung tissues compared with the IR control group. In conclusions, the obtained findings demonstrate that oral supplementation of TEP and PS to rats with pancreatic IR alleviates pancreatic and lung injuries by reducing oxidative stress and modulation of inflammatory processes, which offers an attractive therapeutic option for severe acute pancreatitis and its associated acute lung injury.
Keywords:
Pulmonary function
; Tephrosia purpurea
; Pseudosemiglabrin
; iNOS
; Pancreas
; Ischemia
; Reperfusion
1. Introduction
Ischemia is the lack of blood flow to a tissue as a result of constricted or blocked arteries, which leaves the tissue devoid of nutrients and oxygen needed for cellular metabolism. It is commonly recognized that ischemia, or reduced blood flow, can cause tissue damage and malfunction of the organs. The length and intensity of the ischemia insult dictate the tissue's ability to recover from its harm and survive in the end [1]. The vulnerability of various organs to ischemia harm varies. Pancreatic ischemia/reperfusion (IR) injury is a major clinical concern, especially in cases involving pancreatic surgery and organ transplantation [2]. Systemic inflammatory responses resulting from pancreatic ischemia/reperfusion contribute to increased white blood cell counts, production of oxygen free radicals, and release of cytokines [3]. These inflammatory responses can lead to conditions such as acute pancreatitis (AP) where there is an increase in leukocyte activity and the release of pro-inflammatory cytokines, leading to systemic inflammatory response syndrome (SIRS) [4]. Consequently, ischemia/reperfusion can affect other organs in addition to the ischemic organ itself, potentially resulting in multi-system organ failure [5].
The main cause of pancreatic injury is oxidative stress, which can happen in a number of clinical situations, such as heart surgery, hemorrhagic shock, hypothermia, and pancreas transplantation [6]. According to Hoffmann et al. [7], and Rolim et al. [8], the generation of reactive oxygen species (ROS) is able to induce AP that overwhelm the body's antioxidant defenses by inducing inflammatory cell infiltration, interstitial edema, enzymatic elevation, and pro-inflammatory cytokine activation. Additionally, oxidant chain reaction raises the permeability of the pancreatic microcirculation, which results in microvascular leakage as well as a lack of perfusion because of the intravascular no-reflow phenomenon brought on by the swelling of endothelial cells, neutrophil adhesion, thromboxane A2, and other vasoactive mediators [9]. This impairment of the microcirculation of the pancreas in the early stage of acute pancreatitis plays an important role in the pathogenesis of this disease [10].
The lungs are generally the most commonly targeted organ in AP, and pulmonary injury resulting from pancreatitis is the leading cause of early death in patients with AP [11]. One of the main causes of AP-induced lung injury is the production of inflammatory mediators, as well as ROS. These mediators aid in the build-up of neutrophils and macrophages, which in turn sets off a series of pathological alterations in the lungs' pulmonary microcirculation that eventually result in the development and exacerbation of lung injury caused by AP [12].
Tephrosia purpurea (TEP) (Leguminosae) has traditionally been used to treat diabetes mellitus and is thought to be helpful for conditions affecting the kidneys, liver, and spleen [13]. Numerous pharmacological effects, including antioxidant, anti-inflammatory [14], hepato-protective [15], immunomodulatory [16], and anti-helicobacter pylori [17], are exhibited by the plant extracts. According to Gokhale and Saraf [18], phytochemical screening of TEP showed the presence of polyphenols, sterols, retinoids like rotenone, flavonoids like purpurin, and flavonoid glycosides like rutin and osyritin. Pseudosemiglabrin (PS), a flavone derived from TEP [19], has demonstrated anti-inflammatory properties, as evidenced by its impact on interleukin-1 (IL-1), tumor necrosis factor-α (TNF-α), and nitric oxide (NO) levels in vitro. In vivo studies have further supported its anti-inflammatory potential by inhibiting granuloma tissue formation and reducing cytokine production [20,21].
TEP extract appears to be useful in encouraging pancreatic regeneration due to its cytoprotective, anti-inflammatory, and antioxidant properties [22]. Nevertheless, there is a dearth of experimental evidence in this area, so the current study was planned to investigate the possible protective effects of TEP and its active constituent, PS, against AP induced by IR in a rat model.
2. Results
2.1. Respirometer
The inspiratory time (Ti), expiratory time (Te), duration (D), and respiratory rate (RR) of the IR control, TEP-200, TEP-400, PS-20, and PS-40 groups were comparable to the sham group (Figure 1).
On the contrary, the peak inspiratory flow (PIF), Forced vital capacity (FVC), and FEV0.1 were hampered in IR control compared to the sham group. Similarly, TEP-200, TEP-400, and PS-20 groups were comparable to the IR control. At the same time, the PS-40 group showed a substantially higher FEV0.1 and PIF value compared to the IR control group, with a comparable value to the sham group (Figure 2).
The IR PEF, FVC, FEV0.1 and FEV0.1/FVC were reduced compared to the sham group. Meanwhile, only rats administered the TEP-400 and PS-40 significantly increased the PEF Flow and FEV0.1 /FVC compared to the IR control group. Similarly, only PS-40 groups showed an appreciable increment in the FVC ratio compared to the IR control group, as well as comparable values of FEV0.1 to both the IR and Sham groups.
2.2. Serum Parameters
As shown in Table 1, serum amylase and lipase levels in the IR control group increased more than 4-fold and the TAP level increased more than 7-fold compared to the sham rats. The TEP-200 or PS-20 groups showed a decrease in serum amylase levels by about 36%, while the TEP-400 or PS-40 groups showed a decrease of about 65% compared to the IR control group. Similarly, serum lipase levels decreased by 70% in the TEP-400 or PS-40 groups and by about 40% in the TEP-200 or PS-20 groups compared to the IR control group. Further, treatment with TEP-200, TEP-400, PS-20 or PS-40 reduced TAP in the serum of the rats by 35, 68, 29 or 55% compared to the IR control group, respectively.
2.3. Assessment of Inflammatory Markers in Pancreatic and Lung Homogenates
The results showed that the pancreatic and lung TNF-α, IL-1β and NF-κB contents were extremely higher in the IR control group than in the sham group (Table 2 and Table 3). The TEP-200 or TEP-400 and PS-20 or PS-40 groups showed a 40, 77, 23, or 69% reduction in pancreatic TNF-α levels compared to the IR control group, respectively. Similarly, administration of TEP-200 or TEP-400 and PS-20 or PS-40 reduced lung TNF-α levels by 59, 82, 46, or 60% compared to the IR control group, respectively. In addition, the pancreatic and lung contents of IL-1β in rats exposed to TEP-200 or TEP-400 and PS-20 or PS-40 were significantly reduced as compared to the IR control group. Moreover, the pancreatic NF-κB in the TEP-200 or TEP-400 and PS-20 or PS-40 groups were reduced by 46, 75, 46 or 75% compared to the IR control group, respectively. At the same time, the TEP-200 or TEP-400 and PS-20 or PS-40 lowered the lung NF-κB by 44, 63, 32 or 62% versus the IR control group, respectively.
2.4. Assessment of Pancreatic Oxidative Stress Markers
Pancreatic MDA was elevated 6-fold in the IR control group compared to sham rats. Pancreatic MDA was reduced by 41, 63, 26 or 55% in the TEP-200 or TEP-400 and PS-20 or PS-40 groups versus IR control group, respectively (Table 4). At the same time, activities of GPx and MPO in the pancreatic tissue were reduced by 75 and 86% in the IR control group compared to sham rats. TEP-200 or TEP-400 and PS-20 or PS-40 increased GPx by 2.9, 4.5, 2.8 or 3.9-fold compared to IR control rats, respectively. Similarly, pancreatic MPO was replenished by 3.1, 5.3, 3.2, or 5.6% compared to the IR control group, respectively.
2.5. Gene Expression Analyses
This study further explored some of the mechanisms by which TEP and PS could regulate the inflammatory response. As detected by RT-qPCR, a marked overexpression in the pancreatic HMGB1 (4.99-fold) was evident in IR group compared to sham (Figure 3). Interestingly, treatment with TEP and PS significantly decreased HMGB1 expression to about 2.2, 1.5, 2.8, 1.9-fold compared to sham in TEP-200, TEP-400, PS-20 and PS-40 groups, respectively. On the other hand, the obtained results demonstrated that, following pancreatic IR induction, the expression levels of IL-22 and Th22 in the lung tissues were markedly downregulated to about 0.3-fold compared to sham group. Meanwhile, administration of TEP-200, TEP-400, PS-20 and PS-40 were efficient to restore the mRNA expression of IL-22 (0.8, 1.2, 0.9 or 1.1-fold, respectively) and Th22 (0.8, 1.1, .9 and 1.2-fold, respectively) in the lung compared to sham group.
2.6. Histopathological Examination of Pancreatic Tissues
As represented in Figure 4, the pancreas tissue of the sham group showed a normal histological structure, while pancreatic sections of IR controls showed hemorrhage and edema between pancreatic acini. TEP and PS-treated groups showed congestion and slight edema between pancreatic acini which was dose dependent.
2.7. Histopathological Examination of Lung Tissues
Figure 5 depicts the histopathologic investigation of the lung tissue. The IR control group showed congestion of perialveolar blood vessels with infiltration of interstitial tissue by mononuclear inflammatory cells. TEP-treated group showed emphysematous alveoli with infiltration of interstitial tissue by mononuclear inflammatory cells, while PS-treated groups revealed infiltration of interstitial tissue by low number of mononuclear inflammatory cells.
2.8. Immunohistochemical Examination of iNOS in Pancreatic Tissue
The immunohistochemical study of the pancreas in the sham group showed negative reaction for nitric oxide synthase (iNOS) in pancreatic acini (Figure 6). However, strong positive reaction for iNOS in nuclei of pancreatic acini was detected in the IR control. All treated groups showed strong reaction for iNOS, however it was mild reaction in cytoplasm of some pancreatic acini of PS-40 treated group.
2.9. Immunohistochemical Examination of iNOS in Lung Tissue
The immunohistochemical investigation of the lung tissue of sham group showed negative reaction for iNOS in pneumocytes (Figure 7). However, strong positive reaction for iNOS in cytoplasm of pneumocytes was spotted in the IR control. On the other hand, low dose of TEP and PS-treated groups showed strong reaction for iNOS, and very mild reaction in groups treated with high dose TEP and PS in few numbers of pneumocytes.
3. Discussion
Pancreatic IR injury is a serious clinical concern. The length and intensity of the ischemia insult dictate the tissue's ability to recover from its harm and survive in the end [23,24]. The general explanation for ischemic tissue damage is a prolonged, severe tissue hypoxia followed by cellular ATP depletion. The present study investigated the role of TEP and its active ingredient, PS in protecting the pancreas and lung from IR injury in rats.
In this study, we found that IR following splenic artery clamping resulted in severe histological damage in pancreatic tissue in rats, which is associated with systemic oxidative stress, pro-inflammatory cytokine release, and loss of the cellular integrity in the lung tissue. Exposure of an organ to IR leads to inflammatory activation in other non-ischemic organs, leading to failure of these organs due to elevated levels of oxidative stress and inflammatory reaction in the re-perfused tissues [25,26]. In the process of severe AP, the lung is the most easily affected organ aside the pancreas [27]. The generation of inflammatory mediators, including cytokines and chemokines is a fundamental cause of AP-induced lung injury. These mediators contribute to the accumulation of macrophages and neutrophils and, later, trigger a cascade of pathological changes in microcirculation of the lungs, leading to the occurrence of AP-induced lung injury [12]. Oxidative stress plays a crucial role in the development of lung injury in acute pancreatitis. An excessive amount of ROS can be generated by activating neutrophils, leading to severe oxidative stress damage [28]. Oxidative stress damage not only plays a role in the local damage of the pancreas but also plays a dominant role in damage to other organs [29].
Evaluating respiratory parameters provides critical insights into the impact of pancreatic IR injury on lung function; it also aids in assessing the treatment efficacy of TEP or PS. In the current work, the respiratory parameters that were selected for analysis are inspiration time (Ti), and expiration time (Te) which are the time elapsed during each breath cycle from inhale to exhale/ or from exhale to inhale, respectively. Interestingly, during our study Te, Ti D (duration of respiratory cycle) and RR (i.e., the number of breaths per minute) showed no significant deviations across all groups from their normal values, while other critical measurements changed in an appreciable manner after an IR insult. The fall in peak inspiratory flow (PIF) and peak expiratory flow (PEF) denoting the maximum amount of gas inhaled/ or per unit time, respectively were observed in IR control group implies poor airway function which hints at a possible presence bronchial obstruction accompanied by decreased strength in respiratory muscles like diaphragm and intercostals among others along reference line [30]. This alteration has great importance because it reflects a general impact of pancreatic IR on respiratory mechanics (increased alveolar permeability pulmonary edema and decreased blood oxygen saturation) and agrees with those seen during acute lung injury in severe pancreatitis [27].
Similarly, decrease in forced vital capacity (FVC) as well as forced expiratory volume at 0.1 seconds (FEV0.1) in the IR control group typically indicates development of lung disease secondary to inflammatory mediated changes of lung compliance and elastic recoil [27,31]. Hence, those levels are prime in appreciating integration of pulmonary organs as well as indicating anomalies prevalent in the lung. Treatment with TEP-400 and PS-40 exhibited potential cure hence PS-40 displayed significant improvements on PIF, FVC, and FEV0.1/FVC ratios comparable to sham group. Restoration of respiratory function was therefore done by PS-40 which helps reduce lung injuries due to pancreatic I/R. Moreover, dose-dependent manner observed on both TEP and PS implies that treatment dose has direct correlation with therapeutic effectiveness. Furthermore, TEP-400 treatment significantly improved the FEV0.1/FVC ratio implying that the airways are more patent than before and the lung is more compliant. This indicates that these drugs could potentially prevent or reverse both constrictive and obstructive diseases involving lung parenchyma with I/R injury caused by pancreatic IR.
Amylase and lipase enzymes are synthesized in the pancreatic acinar cells. They are vital enzymes produced by the pancreas and are the most commonly used biochemical marker in the diagnosis of acute pancreatitis [32]. In the present study, serum amylase and lipase levels in the IR control group increased more than 4-fold compared to the sham rats. In this respect, a raised level of serum amylase activity, at least three times the upper limit of normal, supports the diagnosis of acute pancreatitis [33]. Treatment with TEP-400 and PS-40 accelerated normalization of serum lipase activity. Among the evidence that may support the therapeutic properties of the plant in the course of IR-induced AP is its ability to attenuate serum lipase activity. This enzyme is released by pancreatic acinar cells into the interstitial tissue during AP, and its concentration in serum serves as an indicator of AP severity with high sensitivity and specificity [34].
Furthermore, the reduction in inflammatory markers such as TNF-α, IL-6, and Nuclear factor kappa B (NF-κB) in both the pancreas and lungs of treated groups demonstrates the anti-inflammatory effect of TEP and PS. These findings are consistent with a previous in vitro study that reported the role of PS in reducing the inflammatory markers TNF-α, IL-1, and NO in rat granuloma as well as human macrophages [21].
Small, nonstructural proteins called cytokines are mainly involved in the way the body reacts to injury, illness, and ischemia [35]. A pleiotropic cytokine, TNF-α can result in tissue damage following tissue ischemia and leukocyte infiltration. TNF-α damages ischemic tissue by altering the permeability of vascular endothelial cells, causing vascular dysfunction, and by interacting with endothelial cells to trigger coagulation events [36].
TNF-α inhibition has been shown to have the potential to be a treatment method to lessen the severity of IR injury. In a rat model of IR-induced lung injury, anti-TNF-α antibody prevented microvascular damage [5].
IL-1β is a well-known mediator of acute inflammation which plays a crucial role in the release of other members of pro-inflammatory cytokine cascade including tumor necrosis factor alpha, platelet-activating factor, prostaglandins and pro-inflammatory interleukins, and consequently stimulates the development of AP [37,38]. Moreover, it has been demonstrated that early and sustained activation of inflammatory cells with successive release of IL-1β and other cytokines is responsible for the intense local and systemic inflammatory response in AP [34]. Contrarily, inhibition of the cytokine cascade at the level of the IL-1 receptor, before or soon after induction of pancreatitis, markedly attenuates the rise in these cytokines and is associated with decreased severity of pancreatitis and reduced pancreatic damage [37,39].
TEP and PS used in our study have been shown to modulate the inflammatory response by reducing the release of pro-inflammatory cytokines in both pancreatic and lung tissues compared to the IR control group. By activating the cytokine cascade and iNOS, IR has been shown to cause severe acute necrotizing pancreatitis with a high mortality rate and to cause a systemic inflammatory response. This implies a connection between the apoptotic process in the lung and pancreas and the excess NO produced by iNOS [40]. Those effects were alleviated on treatment with TEP and PS that was evident by results of the immunohistochemical examination of both pancreatic and lung tissues, in a dose dependent manner.
In the current study, pancreatic malondialdehyde (MDA) was elevated 6-fold in the IR control group compared to sham rats. TEP-200 or TEP-400 and PS-20 or PS-40 groups revealed significant reduction in pancreatic MDA versus IR control group. Additionally, activities of glutathione peroxidase (GPx) and myeloperoxidase (MPO) in the pancreatic tissue were reduced in the IR control group compared to sham rats, and were significantly elevated in TEP-200 or TEP-400 and PS-20 or PS-40 compared to IR control rats, respectively. The restoration of pancreatic oxidative stress indicators as MDA, GPx, and MPO in the therapy groups supports antioxidant capabilities. This is critical in the context of IR injury, because oxidative stress is a major contributor to tissue damage.
ROS and microvascular blood flow failure appear to be important pathological mechanisms of acute pancreatitis caused by IR [41]. Several studies have shown that oxidative stress is the primary mechanism of IR injury in the pancreas [42]. Another main effect of ROS is lipid peroxidation [43]. Lipid peroxidation is brought on by tissue ischemia and reperfusion [44]. It is a series of events that result in the oxidation of polyunsaturated fatty acids. This process damages cellular membranes and generates hazardous byproducts including MDA [45]. MDA is a useful measure of the degree of lipid peroxidation, which is a free radical generating mechanism that may be strongly linked to IR-induced tissue damage with an increase at reperfusion [46]. Further, the body has an inbuilt anti-oxidation system that keeps the production and removal of free radicals in a state of dynamic equilibrium, safeguarding the integrity and functionality of cells. GPx, MPO, and other enzyme systems are among them [47]. MPO is considered to be a marker of local neutrophil activity causing tissue damage in various inflammatory diseases including AP [48]. In addition, in patients with AP, it has been shown that MPO contributes to the production of reactive oxygen metabolites and its level depends on the severity of AP, as well as on cytokine blood level [49].
In this study, pancreatic IR resulted in severe oxidative stress characterized by increased free radical products MDA and MPO activity as well as decreased GPx activity in pancreatic tissue of IR control rats. These parameters correlate well with disease severity [50,51]. Administration of TEP-200, TEP-400, PS-20 and PS-40 showed significant protection against the oxidative status of pancreatic tissue of rats, via reducing free radical-derived products (MDA), and increasing the activities of GPx and MPO in comparison to IR control group. TEP-400 and PS-40 restored GPx activities in the pancreas to levels similar to those in the sham group. These findings are corroborated by the fact that TEP's antioxidant potential is primarily due to its high polyphenol and flavonoid contents. This is consistent with previous report that found a substantial link between plant antioxidant potential and its polyphenols content [52].
Accumulating evidence highlighted the linking interaction between HMGB1 and various inflammatory signaling pathways [53]. HMGB1 is a nuclear protein belongs to the damage-associated molecular pattern family (DAMP) and acts as a late proinflammatory mediator in tissue injury [54]. Once released, HMGB1 activates toll-like receptors (TLRs) which subsequently stimulates NF-κB signaling [55]. Consistent with previous studies, the present study demonstrated a marked increased pancreatic HMGB1 and expression in the IR group compared with sham group [56]. In our study, TEP and PS downregulated HMGB1 gene expressions with concomitant decrease in the pancreatic and pulmonary of the inflammatory mediators on the pancreatic inflammatory NF-κB, TNF-α, and IL-1β. This multipronged mechanism verifies the anti-inflammatory potential of TEP and its PS derivative.
Evidence has also highlighted the pro-inflammatory and anti-inflammatory roles of Th22 cells in the pathogenesis of inflammatory and autoimmune diseases. Th22 cells are identified as CD4+ cells where IL-22 primarily mediates their function [57]. IL-22 is one of the IL-10 family members acts through IL-22 receptor 1 to coordinate immune cells, regulate inflammation and tissue repair [58]. Previous studies reported that IR induced activation of L-22/IL-22R1/STAT3 signaling which subsequently regulates mitochondrial apoptosis [59]. In addition, IL-22 has been reported to induce the Bcl-2 which binds to Beclin-1 and suppress autophagy [60]. Several animal model studies have reported a significant reduction of IL-22 expression and Th22 cells in the lung tissues following acute pancreatitis [61,62]. Based on previous studies, we suggested that upregulation of lung IL-22 expression in TEP and PS treated groups could mitigates acute lung injury [58].
The regulatory effects of TEP and PS on the pancreatic HMGB1 and pulmonary IL-22 and Th22 gene expression profiles suggest a plausible explanation for the observed protective benefits. Modification of these genes may contribute to TEP and PS's anti-inflammatory and tissue-protective properties.
The histopathological examination further supported the protective effects of TEP and PS treatments on both pancreatic and lung tissues. These observations align with the functional improvements in respiratory parameters and the modulation of inflammatory and oxidative stress markers.
4. Materials and Methods
4.1. Plant Material
The plants of Tephrosia purpurea L. (Pers.) were collected from Gazan, Southern part Saudi Arabia in November 2016. Details of plant identification, method of extraction, isolation and identification of (-)-pseudosemiglabrin were recently described [63].
4.2. Animals
After the approval of the animal experimental protocol by the Ethical Committee for Medical Research, National Research Centre, Egypt, about thirty-six adult Male Wistar rats weighing 180-200 g were purchased from the Animal Facility of the National Research Centre, Egypt. Animals were kept in standard cages, under pathogen-free conditions, and maintained under controlled room temperature and under normal dark –light cycles. Animals were provided with standard food and water ad libitum. Rats were allowed to adapt to these conditions for 1 weeks before beginning the experimental protocol. Experiments were performed according to the National Regulations of Animal Welfare and the Institutional Animal Ethical Committee (IAEC), Approval #2416072022.
4.3. Experimental Design
Thirty-six adult male Sprague Dawley rats were randomly divided into 6 equal groups.
- Sham group: Rats received 1 mL of the vehicle (2% Tween 80 in sterile saline).
- IR control group: Rats received orally 1 mL of the vehicle kg.
- TEP-200 group: Rats treated orally with T. purpurea at 200 mg/kg.
- TEP-400 group: Rats treated orally with T. purpurea at 400 mg/kg.
- PS-20 group: Rats treated orally with pseudosemiglabrin at 20 mg/kg.
- PS-40 group: Rats treated orally with pseudosemiglabrin at 40 mg/kg.
The vehicle, TP and PS were given to rats daily for 5 days before the induction of pancreatic ischemia, and once after 24 hours of reperfusion.
4.4. Induction of Pancreatic Ischemia
One hour after the fifth day dose of the vehicle, TEP extract or PS, rats were anesthetized with ketamine (50 mg/kg i.p). A midline abdominal incision was done to expose the splenic artery. The splenic artery was occluded with a microvascular clamp for 60 minutes. Ischemia was confirmed by the blanching of the pancreas. After 60 minutes, the clamp was removed and pancreatic reperfusion was observed visually. The skin incision was sutured with silk suture and the rats were allowed to recover [55]. The sham-operated group were subjected to the same conditions but without splenic artery occlusion. Another oral dose of the vehicle, TEP extract or PS was given to rats, 24 h after the beginning of pancreatic reperfusion.
One hour later, assessment of respiratory functions of rats using Respirometer was done. Later, blood samples were collected from the retro-orbital venous plexus of rats for estimating serum biochemical parameters. The serum was separated by centrifugation for 1,538 g for 10 minutes.
Rats were euthanized by decapitation; the pancreas and lung were separated for histopathological immunohistochemical, molecular, and biochemical investigations. Tissues of three animals in each group were fixed in 10% formalin buffer for 24 h for histological examination. While the other tissue samples of the rest of animals in each group were dissected and divided into two portions. The first portion was immediately snap-frozen in liquid nitrogen and stored at −80 °C for gene expression analysis. While, the second portion was immediately homogenized in ice-cold 10% (w/v) phosphate buffer. The homogenate was centrifuged at 1800 xg for 10 min at 4˚C. The supernatant was used for different biochemical analyses.
4.5. Assessment of Respiratory Functions Using Respirometer
Inspiratory time (Ti), expiratory time (Te), duration (D), peak inspiratory flow (PIF), peak expiratory flow (PEF), Forced vital capacity (FVC), forced expiratory volume 1 (FEV1), the ratio of FEV1/FVC and respiratory rate (RR) were estimated by putting the rats in a 1.5-liter chamber connected to spirometer with an MTL1 flow meter and AID instruments® Power Lab gear. After calibrating and acclimatizing for 5 min, labChart 8 was used to analyze the respiratory wave and to detect forced expiration for 15 min. The respiratory parameters were estimated by putting the rats in a 1.5-liter chamber connected to spirometer with an MTL1 flow meter and AID instruments® Power Lab gear. After calibrating and acclimatizing for 5 min, labChart 8 was used to analyze the respiratory wave and to detect forced expiration for 15 min.
4.6. Serum Analysis
Serum amylase, lipase, trypsinogen activated peptide (TAP), were analyzed according to the manufacturer’s protocol of the used kit.
4.7. Assessment of Inflammatory Markers in Pancreatic and Lung Homogenate
TNF-α, IL-1B, NF-κB were assessed in lung homogenates using ELISA kits procured from BioVision Inc. (Milpitas, CA, USA).
4.8. Assessment of Pancreatic Oxidative Stress Markers
Different oxidative stress markers such as MDA, GPx, and MPO were assessed in pancreatic homogenates using ELISA technique according to the manufacturer’s protocol (BioVision Inc., Milpitas, CA, USA).
4.9. Real-Time Polymerase Chain Reaction (RT PCR)
Total RNA from the pancreas and lungs samples was purified with the use of TRIzol reagent (Thermo Fisher Scientific, USA) in accordance with the manufacturer’s protocol. RNA quantity, quality, and integrity were measured with the spectrophotometric analysis of A260/A280 ratio, A260/A230 ratio, and 1% agarose gel electrophoresis. RNA was used as a template in the reverse transcription reaction to form cDNA with the use of RevertAid First Strand cDNA Synthesis Kit (Fermentas, Thermofsher Scientifc, USA). Then, PCR quantitative analyses of target genes expression were performed adopting a QuantiTect SYBR Green Kit (Qiagen, Germany) on a Real-Time PCR Detection System (Bio-Rad Laboratories, USA). The cycling program was as follow: Pre-denaturation (94 °C, 5 min); 40 cycles of denaturation (94 °C, 45 s), primer annealing (58 °C, 45 s), primer extension (72 °C, 55 s); and a final extension (72 °C, 7 min). Specific primers for detection of HMGB1, IL-22, Th22 were used: HMGB1, forward 5′- GAGTACCGCCCAAAAATCAA -3′, reverse 5′- TTCATCCTCCTCGTCGTCTT -3’ (XM_039100280.1); IL-22, forward 5′- AGTCACCTCAGCCCCTGTCAC -3′, reverse 5′- CCCGATCGCTTTAATCTCTCC -3’(NM_001191988.1); Th22, forward 5’-GGGCTTCCAGGGTGCTTCGC-3′, reverse 5’-CCTCAGTTCACCGAGAACCCCA-3’ (NM_201270.1); and GAPDH, forward 5'- CCTCGTCTCATAGACAAGATGGT -3', and reverse 5'- GGGTAGAGTCATACTGGAACATG -3' (XM_039082880.1). The relative expression of target genes was normalized to GAPDH as an endogenous control gene, calculated using the 2−ΔΔCt formula, and expressed as a fold change of the normal control group [64].
4.10. Histopathological Examination of Pancreatic and Lung Tissues
Pancreatic and lung tissues were fixed in 10% neutral-buffered formalin for 24 h, washed with tap water, prepared and stained for light microscopy. For dehydration, serial dilutions of alcohol were applied and thereafter the specimens were cleared in xylene and embedded in paraffin wax in hot air oven at 56 ºC for 6 h. Paraffin wax tissue blocks were sectioned by using microtome at 5-6 microns thickness. Then, sections were collected on glass slides and deparaffinized. They were stained for routine histological examination using Hematoxylin and Eosin stain (H&E), according to Bancroft and Gamble [65].
4.11. Immunohistochemical Examination of iNOS in Pancreatic and Lung Tissues
Paraffin sections were mounted on positively charged slides by using avidin biotin- peroxidase complex (ABC) method. Rabbit iNOS polyclonal Antibody (Gene tex, Cat.No. GTX130246, Dil.: 1:100). Sections from each group were incubated with these antibodies, then the reagents required for ABC method were added (Vectastain ABC-HRP kit, Vector laboratories). Marker expression was labeled with peroxidase and colored with diaminobenzidine (DAB, produced by Sigma) to detect antigen-antibody complex. Negative controls were included using non-immune serum in place of the primary or secondary antibodies. IHC stained sections were examined via using Olympus microscope (BX-53). Scoring was performed by determination of reaction area percent using ImageJ 1.53K, national institute of health, USA.
4.12. Statistical Analysis
Data were presented as mean ± S.E, and analyzed by one-way ANOVA followed by the Tukey–Kramer Post hoc test in case of normality, and were analyzed using the Kruskal-Wallis test followed by Dunn's multiple comparisons test in case of non-normality (GraphPad Software, version 9, USA). The significance limit was set to p≤0.05.
5. Conclusions
In conclusion, the results of this study suggest that TEP and PS have promising protective benefits in reducing IR-induced damage in both the respiratory system and the pancreas. The proposed mechanism of protection involves reducing cytokine response and suppressing oxidative stress. Additionally, the mechanism may involve modulating the HMGB1 and IL-22 pathway. These findings may help provide a potential preventive and therapeutic alternative for pancreatic I/R injury and associated lung complications. However, further studies are required to explore other possible mechanisms of TEP and PS.
Author Contributions
Conceptualization, G.A.S., R.F.A.-R., M.A.E. and M.S.A.-K.; methodology, M.A.A., R.F.A.-R., H.A.O., M.A.E. and R.M.A.-E.; software, M.A.A. and R.M.A.-E.; validation, M.A.E., H.A.O. and M.A.A.; formal analysis, M.A.A., R.F.A.-R. and R.M.A.-E.; investigation, G.A.S., R.F.A.-R., M.A.A., H.A.O. and M.A.E.; resources, M.S.A.-K.; data curation, G.A.S., M.A.A., R.F.A.-R. and M.S.A.-K.; writing—original draft preparation, M.A.A., M.A.E., R.F.A.-R., H.A.O. and R.M.A.-E.; writing—review and editing, G.A.S. and R.F.A.-R.; visualization, A.S.S., R.F.A.-R., H.A.O., R.M.A.-E. and M.A.E.; supervision, G.A.S. and M.S.A.-K.; project administration, G.A.S. and M.S.A.-K.; funding acquisition, M.S.A.-K. All authors have read and agreed to the published version of the manuscript
Funding
The project was funded by Prince Sattam bin Abdulaziz University through the project number (PSAU/2024/03/31988).
Institutional Review Board Statement
Experiments were performed according to the National Regulations of Animal Welfare and the Institutional Animal Ethical Committee (IAEC), Approval #2416072022.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
The authors extend their appreciation to Prince Sattam bin Abdulaziz University for funding this research work through the project number (PSAU/2024/03/31988).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IR | Ischemia-reperfusion |
| AP | Acute pancreatitis |
| SIRS | Systemic inflammatory response syndrome |
| ROS | Reactive oxygen species |
| TEP | Tephrosia purpurea |
| PS | Pseudosemiglabrin |
| IL-1 | Interleukin 1 |
| TNF-α | Tumor necrosis factor-α |
| NO | Nitric oxide |
| Ti | Inspiratory time |
| Te | Expiratory time |
| D | Duration |
| RR | Respiratory rate |
| PIF | Peak inspiratory flow |
| FVC | Forced vital capacity |
| FEV0.1 | Forced expiratory volume at 0.1 seconds |
| HMGB1 | High mobility group box 1 protein |
| IL-22 | Interleukin 22 |
| Th22 | T helper cell 22 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| iNOS | Nitric oxide synthase |
| NF-κB | Nuclear factor kappa B |
| MDA | Malondialdehyde |
| GPx | Glutathione peroxidase |
| MPO | Myeloperoxidase |
| DAMP | Damage- associated molecular pattern family |
| TLRs | Toll-like receptors |
| CD4+ | Cluster of differentiation 4-positive |
| STAT3 | Signal transducer and activator of transcription 3 |
| Bcl-2 | B-cell leukemia/lymphoma 2 protein |
| i.p. | Intraperitoneal |
| TAP | Trypsinogen activation peptide |
| ABC | Avidin biotin peroxidase complex |
| DAB | Diaminobenzidine |
References
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox Biol 2015, 6, 524–551. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.O.S.; Losada, D.M.; Jordani, M.C.; Évora, P.; Castro-E-Silva, O. Ischemia/reperfusion injury revisited: An overview of the latest pharmacological strategies. Int J Mol Sci 2019, 20, 5034. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Hoffmann, T.F.; Leiderer, R.; Haeris, A.C.; Messmer, K. Ischemia and reperfusion in pancreas. Microsc Res Tech 1997, 37, 557–571. [Google Scholar] [CrossRef]
- Robert, F.; Matthew, T. Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists; University of Adelaide Press: South Australia, 2011. [Google Scholar]
- Halliwell, B.; Gutteridge, J. Free radicals in biology and medicine; Oxford University Press: New York, 1999. [Google Scholar]
- Hoffmann, T.F.; Leiderer, R.; Waldner, H.; Arbogast, S.; Messmer, K. Ischemia reperfusion of the pancreas: a new in vivo model for acute pancreatitis in rats. Res Exp Med 1995, 195, 125–144. [Google Scholar] [CrossRef]
- Rolim, M.F.; Riger, C.J.; Eleutherio, E.C.A.; Cola˜o, C.F.; Pereira, G.C.; Schanaider, A. Colonic healing after portal ischemia and reperfusion: an experimental study with oxidative stress biomarkers. Redox Rep 2007, 12, 267–274. [Google Scholar] [CrossRef]
- Schanaider, A.; de Carvalho, T.P.; de Oliveira Coelho, S.; Renteria, J.M.; Eleuthério, E.C.A.; Castelo-Branco, M.T.L.; Madi, K.; Baetas-da-Cruz, W.; de Souza, H.S.P. Ischemia–reperfusion rat model of acute pancreatitis: protein carbonyl as a putative early biomarker of pancreatic injury. Clin Exp Med 2015, 15, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Li, Z.J.; Zhang, J. Inflammatory mediators and microcirculatory disturbance in acute pancreatitis. Hepatobiliary Pancreat Dis Int 2009, 4, 351–357. [Google Scholar]
- Samanta, J.; Singh, S.; Arora, S.; Muktesh, G.; Aggarwal, A.; Dhaka, N.; Sinha, S.K.; Gupta, V.; Sharma, V.; Kochhar, R. Cytokine profile in prediction of acute lung injury in patients with acute pancreatitis. Pancreatology 2018, 18, 878–84. [Google Scholar] [CrossRef]
- Ge, P.; Luo, Y.; Okoye, C.S.; Chen, H.; Liu, J.; Zhang, G.; Xu, C.; Chen, H. Intestinal barrier damage, systemic inflammatory response syndrome, and acute lung injury: a troublesome trio for acute pancreatitis. Biomed Pharmacother 2020, 132, 110770. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.K.; Verma, P.R.; Itankar, P.R.; Prasad, S.K.; Nakhate, K.T. Evaluation of pancreatic regeneration activity of Tephrosia purpurea leaves in rats with streptozotocin-induced diabetes. J Tradit Complement Med 2021, 11, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.; Shwetha, K.; Prabhu, K.; Maradi, R.; Bairy, K.L.; Shanbhag, T. Evaluation of antiinflammatory activity of Tephrosia purpurea in rats. Asian Pac J Trop Med 2010, 3, 193–195. [Google Scholar] [CrossRef]
- Khatri, A.; Garg, A.; Agrawal, S.S. Evaluation of hepatoprotective activity of aerial parts of Tephrosia purpurea L. and stem bark of Tecomella undulata. J Ethnopharmacol 2009, 122, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Damre, A.S.; Gokhale, A.B.; Phadke, A.S.; Kulkarni, K.R.; Saraf, M.N. Studies on the immunomodulatory activity of flavonoidal fraction of Tephrosia purpurea. Fitoterapia 2003, 74, 257–261. [Google Scholar] [CrossRef]
- Chinniah, A.; Mohapatra, S.; Goswami, S. On the potential of Tephrosia purpurea as anti-Helicobacter pylori agent. J Ethnopharmacol 2009, 124, 642–645. [Google Scholar] [CrossRef]
- Gokhale, A.B.; Saraf, M.N. Tephrosia Purpurea: a review of contemporary literature and medicinal properties. Indian Drugs 2000, 37, 553–560. [Google Scholar]
- Hassan, L.E.A.; Ahamed, M.B.K.; Majid, A.S.A.; Iqbal, M.A.; Suede, F.S.R.A.; Haque, R.A.; Ismail, Z.; Ein, O.C.; Majid, A.M.S.A. Crystal structure elucidation and anticancer studies of (-)-pseudosemiglabrin: A flavanone isolated from the aerial parts of tephrosia apollinea. PLoS One 2014, 9, e90806. [Google Scholar] [CrossRef]
- Balaha, M.F.; Alamer, A.A.; Abdel-Kader, M.S.; Alharthy, K.M. Ameliorative Potential of (-) Pseudosemiglabrin in Mice with Pilocarpine-Induced Epilepsy: Antioxidant, Anti-Inflammatory, Anti-Apoptotic, and Neurotransmission Modulation. Int J Mol Sci 2023, 24, 10773. [Google Scholar] [CrossRef]
- Hassan, L.E.A.; Dahham, S.S.; Fadul, S.M.; Umar, M.I.; Majid, A.S.A.; Khaw, K.Y.; Majid, A.M.S.A. Evaluation of in vitro and in vivo anti-inflammatory effects of (−)-pseudosemiglabrin, a major phytoconstituent isolated from Tephrosia apollinea (Delile) DC. J Ethnopharmacol 2016, 193, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, S.; Pawar, R.S.; Jain, A.P.; Singhai, A.K. Wound healing potential of Tephrosia purpurea (Linn.) Pers. in rats. J Ethnopharmacol 2006, 108, 204–210. [Google Scholar] [CrossRef]
- Carden, D.; Granger, D. Pathophysiology of ischaemia-reperfusion injury. J Pathol 2000, 190, 255–266. [Google Scholar] [CrossRef]
- Raedschelders, K.; Ansley, D.; Chen, D. The cellular and molecular origin of reactive oxygen species generation during myocardial ischemia and reperfusion. Pharmacol Ther 2012, 133, 230–255. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Kim, M.; Brown, K.M.; D’Agati, V.D.; Lee, H.T. Paneth cell-derived interleukin-17A causes multiorgan dysfunction after hepatic ischemia and reperfusion injury. Hepatology 2011, 53, 1662–75. [Google Scholar] [CrossRef] [PubMed]
- Nishikata, R.; Kato, N.; Hiraiwa, K. Oxidative stress may be involved in distant organ failure in tourniquet shock model mice. Leg Med 2014, 16, 70–5. [Google Scholar] [CrossRef]
- Liu, D.; Wen, L.; Wang, Z.; Hai, Y.; Yang, D.; Zhang, Y.; Bai, M.; Song, B.; Wang, Y. The Mechanism of Lung and Intestinal Injury in Acute Pancreatitis: A Review. Front Med (Lausanne) 2022, 7, 904078. [Google Scholar] [CrossRef]
- Levy, B.D.; Hickey, L.; Morris, A.J.; Larvie, M.; Keledjian, R.; Petasis, N.A.; Bannenberg, G.; Serhan, C.N. Novel polyisoprenyl phosphates block phospholipase D and human neutrophil activation in vitro and murine peritoneal inflammation in vivo. Br J Pharmacol 2005, 146, 344–51. [Google Scholar] [CrossRef] [PubMed]
- Que, R.S.; Cao, L.P.; Ding, G.P.; Hu, J.A.; Mao, K.J.; Wang, G.F. Correlation of nitric oxide and other free radicals with the severity of acute pancreatitis and complicated systemic inflammatory response syndrome. Pancreas 2010, 39, 536–40. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, N.; Oto, J.; Ueno, Y.; Nakataki, E.; Itagaki, T.; Nishimura, M. Change in diaphragm and intercostal muscle thickness in mechanically ventilated patients: a prospective observational ultrasonography study. J Intensive Care 2019, 7, 56. [Google Scholar] [CrossRef]
- Zhou, M.T.; Chen, C.S.; Chen, B.C.; Zhang, Q.Y.; Andersson, R. Acute lung injury and ARDS in acute pancreatitis: mechanisms and potential intervention. World J Gastroenterol 2010, 16, 2094–2099. [Google Scholar] [CrossRef]
- Banks, P.A.; Bollen, T.L.; Dervenis, C.; Gooszen, H.G.; Johnson, C.D.; Sarr, M.G.; Tsiotos, G.G.; Vege, S.S. Classification of acute pancreatitis—2012: revision of the Atlanta classification and definitions by international consensus. Gut 2013, 62, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Matull, W.R.; Pereira, S.P.; O'Donohue, J.W. Biochemical markers of acute pancreatitis. J Clin Pathol 2006, 59, 340–4. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.P.; Dawra, R.K.; Saluja, A.K. New insights into the pathogenesis of pancreatitis. Curr Opin Gastroenterol 2013, 29, 523–30. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. The Role of the Interleukin-1–Receptor Antagonist in Blocking Inflammation Mediated by Interleukin-1. N Engl J Med 2000, 343, 732–734. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. TNF-alpha as a therapeutic target in inflammatory diseases, ischemia-reperfusion injury and trauma. Curr Med Chem 2009, 16, 3152–67. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin-1 and interleukin-1 antagonism. Blood 1991, 77, 1627–1652. [Google Scholar] [CrossRef]
- Kingsnorth, A. Role of cytokines and their inhibitors in acute pancreatitis. Gut 1997, 40, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.; Franz, M.; Messina, J.; Riker, A.; Fabri, P.J.; Rosemurgy, A.S.; Gower Jr., W. R. Interleukin-1 receptor antagonist decreases severity of experimental acute pancreatitis. Surgery 1995, 117, 648–55. [Google Scholar] [CrossRef]
- Li, B.F.; Liu, Y.F.; Cheng, Y.; Zhang, K.Z.; Li, T.M.; Zhao, N. Protective effect of inducible nitric oxide synthase inhibitor on pancreas transplantation in rats. World J Gastroenterol 2007, 13, 6066–71. [Google Scholar] [CrossRef] [PubMed]
- Sakorafas, G.; Tsiotos, G.; Sarr, M. Ischemia/Reperfusion-Induced Pancreatitis. Dig Surg 2000, 17, 3–14. [Google Scholar] [CrossRef]
- Muñoz-Casares, F.; Padillo, F.; Briceño, J.; Collado, J.; Muñoz-Castañeda, J.; Ortega, R.; Cruz, A.; Túnez, I.; Montilla, P.; Pera, C.; Muntané, J. Melatonin Reduces Apoptosis and Necrosis Induced by Ischemia/Reperfusion Injury of the Pancreas. J Pineal Res 2006, 40, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int J Mol Sci 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Halladin, N.L.; Ekeløf, S.; Jensen, S.E.; Aarøe, J.; Kjærgaard, B.; Heegaard, P.M.H.; Lykkesfeldt, J.; Rosenberg, J.; Gögenur, I. Melatonin does not affect oxidative and inflammatory biomarkers in a closed-chest porcine model of acute myocardial infarction. In Vivo 2014, 28, 483–88. [Google Scholar] [PubMed]
- Valenzuela, A. The biological significance of malondialdehyde determination in the assessment of tissue oxidative stress. Life Sci 1991, 48, 301–9. [Google Scholar] [CrossRef]
- Lips, J.; de Haan, P.; Bodewits, P.; Vanicky, I.; Dzoljic, M.; Jacobs, M.J.; Kalkman, C.J. Neuroprotective effects of riluzole and ketamine during transient spinal cord ischemia in the rabbit. Anesthesiology 2000, 93, 1303–11. [Google Scholar] [CrossRef] [PubMed]
- Kryl’skii, E. D.; Popova, T. N.; Safonova, O. A.; Stolyarova, A. O.; Razuvaev, G. A.; de Carvalho, M. A. P. Transcriptional regulation of antioxidant enzymes activity and modulation of oxidative stress by melatonin in rats under cerebral ischemia / reperfusion conditions. Neuroscience 2019, 406, 653–666. [Google Scholar] [CrossRef]
- Chooklin, S.; Pereyaslov, A.; Bihalskyy, I. Pathogenic role of myeloperoxidase in acute pancreatitis. Hepatobiliary Pancreat Dis Int 2009, 8, 627–31. [Google Scholar]
- Bukowczan, J.; Warzecha, Z.; Ceranowicz, P.; Kuśnierz-Cabala, B.; Tomaszewska, R. Obestatin Accelerates the Recovery in the Course of Ischemia/Reperfusion-Induced Acute Pancreatitis in Rats. PLoS ONE 2015, 10, e0134380. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Bonham, M.; Buss, H.; Abu-Zidan, F.; Windsor, J.A. Elevated Protein Carbonyls as Plasma Markers of Oxidative Stress in Acute Pancreatitis. Pancreatology 2003, 3, 375–382. [Google Scholar] [CrossRef]
- Hernández, V.; Miranda, M.; Pascual, I.; Sanchiz, V.; Almela, P.; Añón, R.; Cuadrado, E.; Sanz, M.I.; Mínguez, M.; Mora, F.; Romero, F.J.; Benages, A. Malondialdehyde in Early Phase of Acute Pancreatitis. Rev Esp Enferm Dig 2011, 103, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid Med Cell Longev 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Febriza, A.; Idrus, H. H. Linking Interaction between Antimicrobial Peptide and High Motility Group Box-1 (HMBG-1) in Bacterial Infection. Open Biochem J 2024, 18, e1874091X277312. [Google Scholar] [CrossRef]
- Wulandari, S.; Nuryastuti, T.; Oktoviani, F.N.; Daniwijaya, M.E.W.; Supriyati, E.; Arguni, E.; Wibawa, T. The association between high mobility group box 1 (HMGB1) and Interleukin-18 (IL-18) serum concentrations in COVID-19 inpatients. Heliyon 2024, 10, e26619. [Google Scholar] [CrossRef]
- Abdel-Kader, M.S.; Abdel-Rahman, R.F.; Soliman, G.A.; Ogaly, H.A.; Alamri, M.A.; Alharbi, A.G. Oleuropein Relieves Pancreatic Ischemia Reperfusion Injury in Rats by Suppressing Inflammation and Oxidative Stress through HMGB1/NF-κB Pathway. Int J Mol Sci 2024, 25, 10171. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Q.; Hao, W.A.; Liu, Y.L.; Yang, D.; Wang, H.L.; Zhao, L.; Chen, H.; Li, L.; Jiang, C.L.; Zhou, X.; Fu, J.; Li, Z. The efficacy and active compounds of Chaihuang Qingyi Huoxue granule to Ameliorate intestinal mucosal barrier injury in rats with severe acute pancreatitis by suppressing the HMGB1/TLR4/NF-κB signaling pathway. Int Immunopharmacol 2025, 144, 113632. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. T helper cell differentiation, heterogeneity, and plasticity. Cold Spring Harb perspect biol 2018, 10, a030338. [Google Scholar] [CrossRef]
- Liu, Q.; Zhu, X.; Guo, S. From pancreas to lungs: The role of immune cells in severe acute pancreatitis and acute lung injury. Immun Inflamm Dis 2024, 12, e1351. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Jiang, H.; Bai, C.; Yu, S.; Pan, Y.; Wang, C.; Li, H.; Li, M.; Sheng, Y.; Chu, F.; Wang, J.; Chen, Y.; Li, J.; Jiang, J. Ac2-26 attenuates hepatic ischemia-reperfusion injury in mice via regulating IL-22/IL-22R1/STAT3 signaling. PeerJ 2022, 10, e14086. [Google Scholar] [CrossRef]
- Feng, D.; Park, O.; Radaeva, S.; Wang, H.; Yin, S.; Kong, X.; Zheng, M.; Zakhari, S.; Kolls, J.K.; Gao, B. Interleukin-22 ameliorates cerulein-induced pancreatitis in mice by inhibiting the autophagic pathway. Int J Biol Sci 2012, 8, 249–257. [Google Scholar] [CrossRef]
- Stojanovic, B.; Jovanovic, I.P.; Stojanovic, M.D.; Jovanovic, M.; Vekic, B.; Milosevic, B.; Cvetkovic, A.; Spasic, M.; Stojanovic, B.S. The emerging roles of the adaptive immune response in acute pancreatitis. Cells 2023, 12, 1495. [Google Scholar] [CrossRef] [PubMed]
- Huai, J. P.; Sun, X.C.; Chen, M.J.; Jin, Y.; Ye, X.; Wu, J.S.; Huang, Z.M. Melatonin attenuates acute pancreatitis-associated lung injury in rats by modulating interleukin 22. World J Gastroenterol 2012, 18, 5122–8. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Kader, M.S.; Saeedan, A.S.; Rehman, N.U.; Faqihi, H.M.; Soliman, G.A. New flavonoids with multiple bronchodilator activity pathways from Tephrosia purpurea L. (Pers.) growing in Saudi Arabia. Saudi Pharm J 2024, 32, 101992. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill Livingstone: China, 2008. [Google Scholar]
Figure 1.
Effect of TEP and PS on the lung function (A) Ti, (B) Te, (C) Duration, (D) RR of pancreatic IR rats. No significant difference.
Figure 1.
Effect of TEP and PS on the lung function (A) Ti, (B) Te, (C) Duration, (D) RR of pancreatic IR rats. No significant difference.
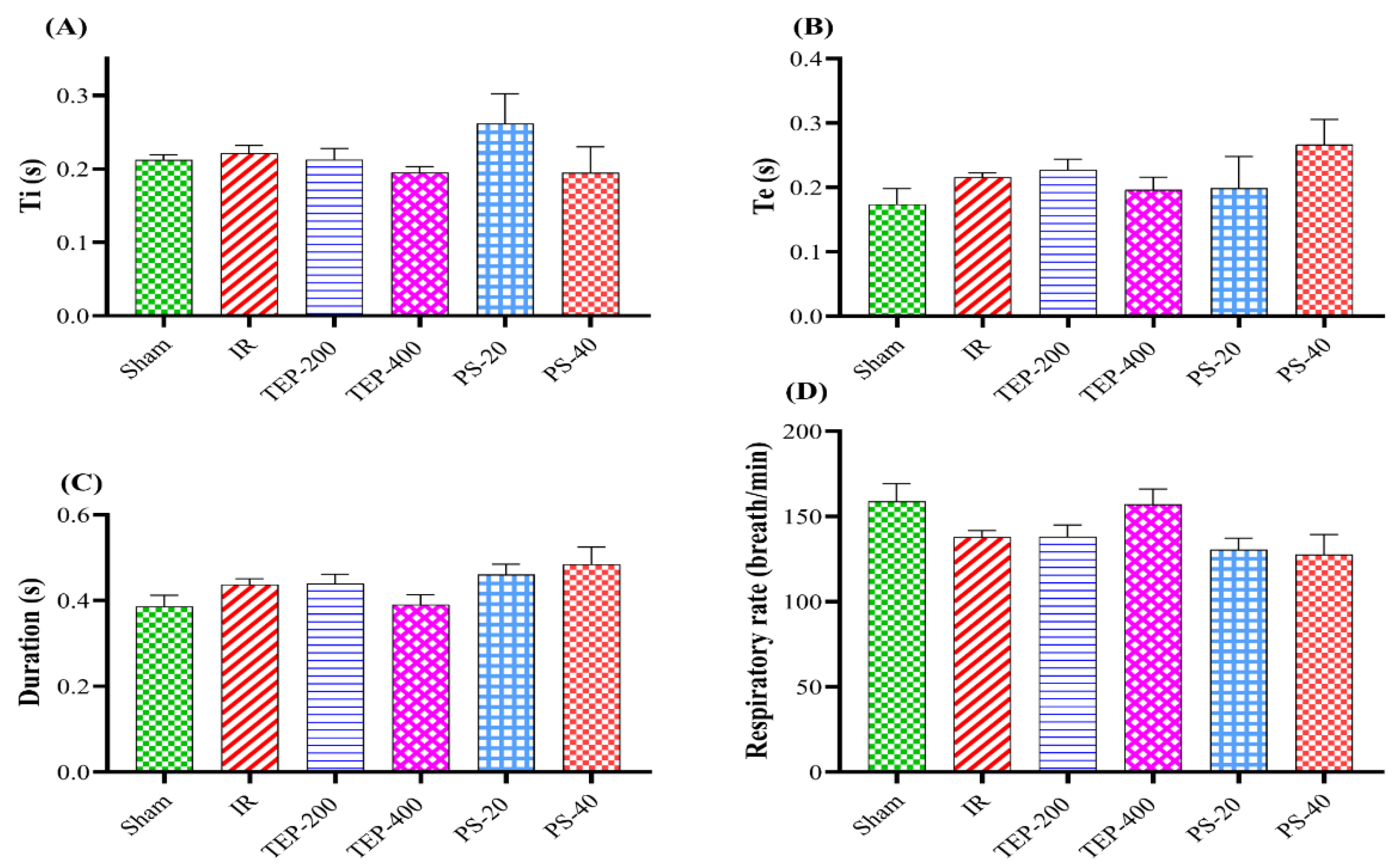
Figure 2.
Effect of TEP and PS on the lung functions: (A) PIF, (B) PEF, (C) FVC, (D) FEV0.1, and (E) FEV0.1 /FVC ratio of pancreatic IR rats. a p ≤0.05 versus sham group, b p≤0.05 versus IR control group.
Figure 2.
Effect of TEP and PS on the lung functions: (A) PIF, (B) PEF, (C) FVC, (D) FEV0.1, and (E) FEV0.1 /FVC ratio of pancreatic IR rats. a p ≤0.05 versus sham group, b p≤0.05 versus IR control group.
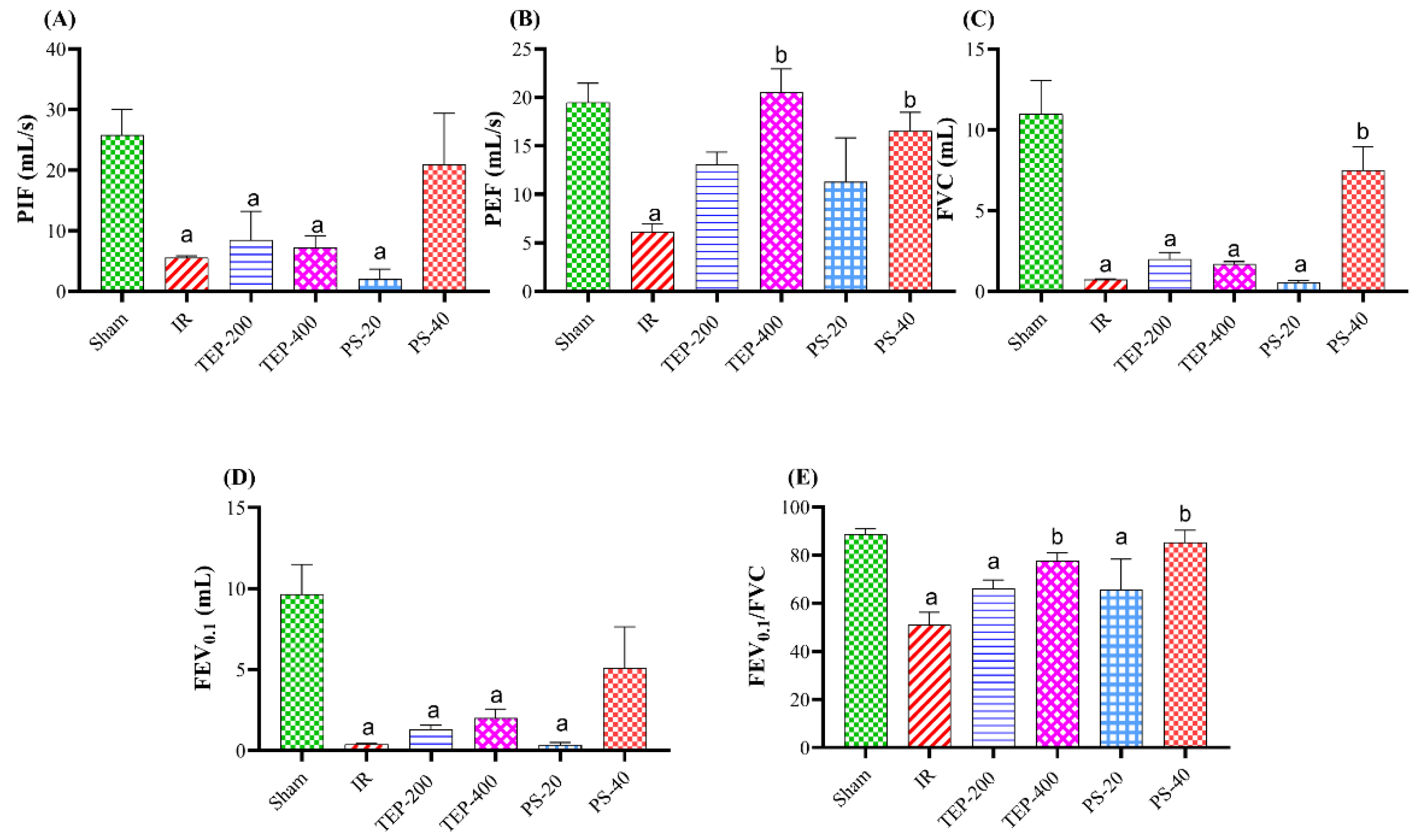
Figure 3.
Effects of TEP and PS administration on the expression of High mobility group box 1 protein (HMGB1), interleukin 22 (IL-22), and T helper cell 22 (Th22) genes in pancreatic IR-induced injury and associated lung injury by real-time qRT-PCR. (A) Pancreatic mRNA expression of HMGB1. (B) Lung mRNA expression of IL-22. (C) Lung mRNA expression of Th22. For all genes, mRNA levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA level. Bars represent mean values of each group ± SEM. a p≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
Figure 3.
Effects of TEP and PS administration on the expression of High mobility group box 1 protein (HMGB1), interleukin 22 (IL-22), and T helper cell 22 (Th22) genes in pancreatic IR-induced injury and associated lung injury by real-time qRT-PCR. (A) Pancreatic mRNA expression of HMGB1. (B) Lung mRNA expression of IL-22. (C) Lung mRNA expression of Th22. For all genes, mRNA levels were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA level. Bars represent mean values of each group ± SEM. a p≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
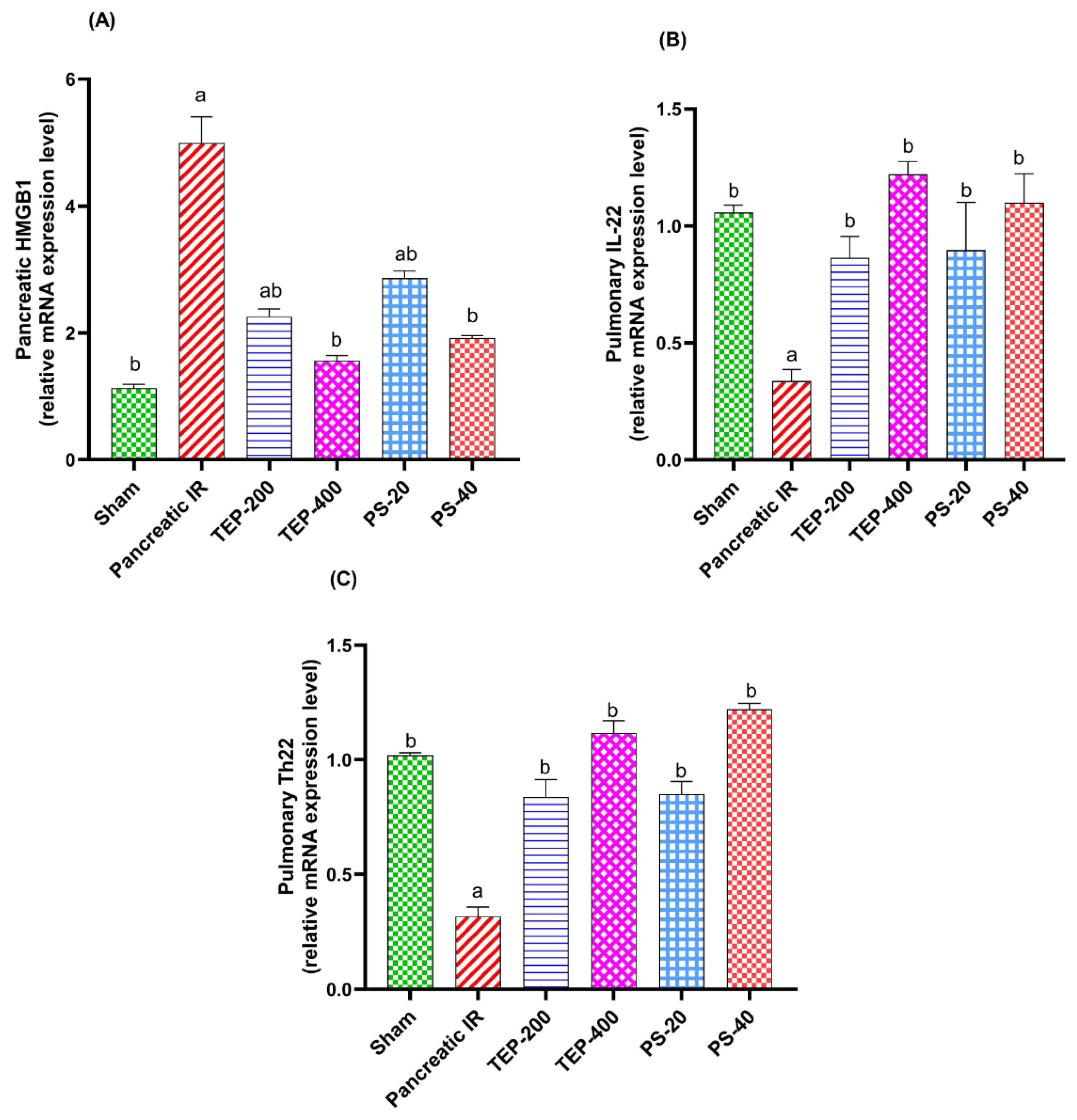
Figure 4.
(i) photomicrograph of sham rat showing normal histological structure of pancreas; (ii) photomicrograph of IR control showing hemorrhage (H) and edema (E) between pancreatic acini; (iii) photomicrograph of TEP-200 treated rat showing congestion of pancreatic blood vessels (C) with edema (E) between pancreatic acini; (iv) photomicrograph of TEP-400 treated rat showing edema between pancreatic acini; (v) photomicrograph of PS-20 treated rat showing hemorrhage (H) and edema (E) between pancreatic acini; (vi) photomicrograph of PS-40 treated rat showing edema (E) between pancreatic acini (H&E).
Figure 4.
(i) photomicrograph of sham rat showing normal histological structure of pancreas; (ii) photomicrograph of IR control showing hemorrhage (H) and edema (E) between pancreatic acini; (iii) photomicrograph of TEP-200 treated rat showing congestion of pancreatic blood vessels (C) with edema (E) between pancreatic acini; (iv) photomicrograph of TEP-400 treated rat showing edema between pancreatic acini; (v) photomicrograph of PS-20 treated rat showing hemorrhage (H) and edema (E) between pancreatic acini; (vi) photomicrograph of PS-40 treated rat showing edema (E) between pancreatic acini (H&E).
Figure 5.
(i) photomicrograph of sham rat showing normal histological structure of alveoli; (ii) photomicrograph of IR control rat showing congestion of perialveolar blood vessels (C) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (iii) photomicrograph of TEP-200 treated rat showing congestion of perialveolar blood vessels (C) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (iv) photomicrograph of TEP-400 treated showing emphysematous alveoli (Em) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (v) photomicrograph of PS-20 treated rat showing congestion of perialveolar blood vessels (C) and infiltration of interstitial tissue by mononuclear inflammatory cells (I) with emphysematous alveoli (Em); (vi) photomicrograph of PS-40 treated rat showing infiltration of interstitial tissue by low number of mononuclear inflammatory cells (I) (H&E).
Figure 5.
(i) photomicrograph of sham rat showing normal histological structure of alveoli; (ii) photomicrograph of IR control rat showing congestion of perialveolar blood vessels (C) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (iii) photomicrograph of TEP-200 treated rat showing congestion of perialveolar blood vessels (C) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (iv) photomicrograph of TEP-400 treated showing emphysematous alveoli (Em) with infiltration of interstitial tissue by mononuclear inflammatory cells (I); (v) photomicrograph of PS-20 treated rat showing congestion of perialveolar blood vessels (C) and infiltration of interstitial tissue by mononuclear inflammatory cells (I) with emphysematous alveoli (Em); (vi) photomicrograph of PS-40 treated rat showing infiltration of interstitial tissue by low number of mononuclear inflammatory cells (I) (H&E).
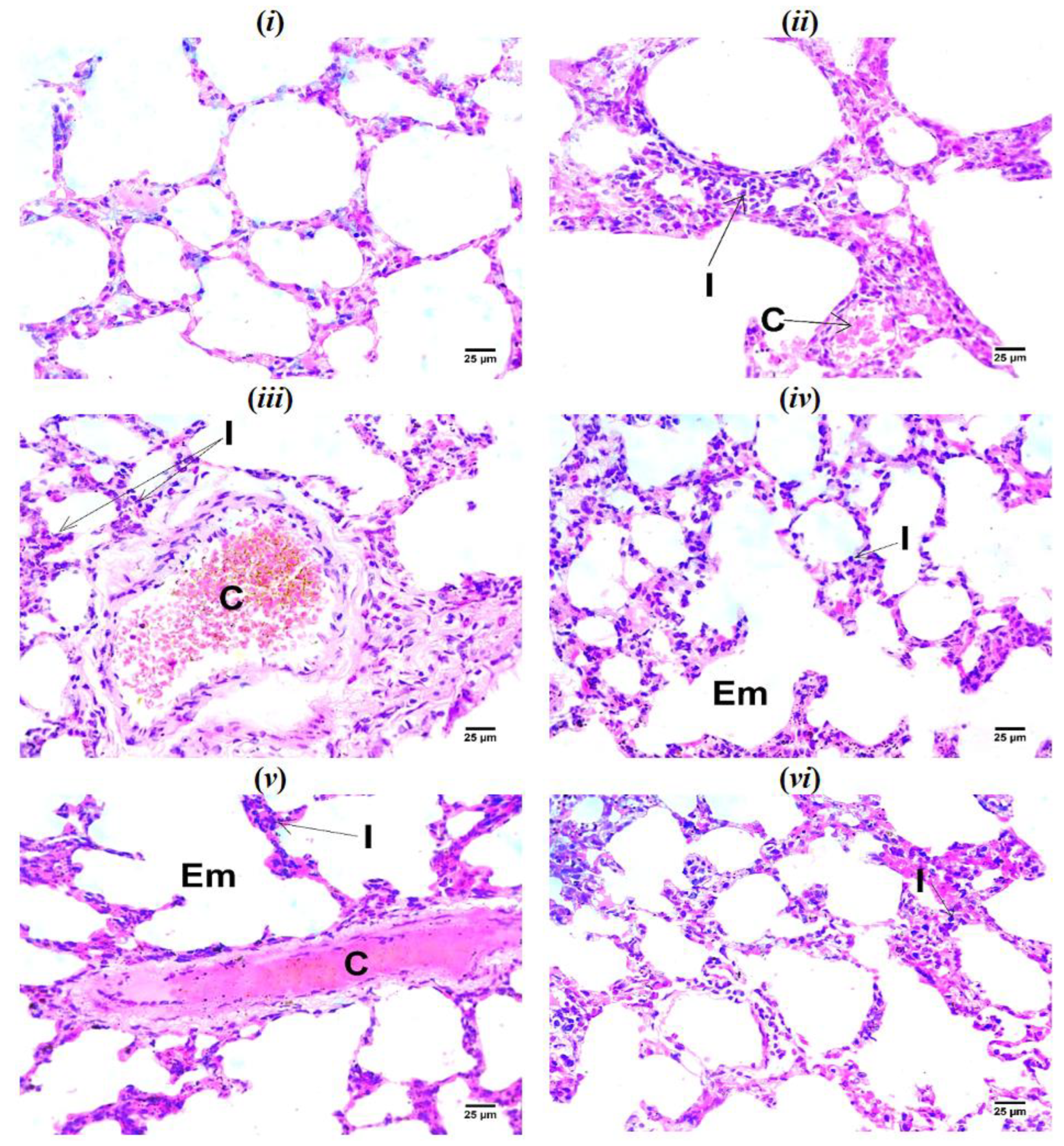
Figure 6.
(i) photomicrograph of sham rat showing negative reaction for iNOS in pancreatic acini; (ii) photomicrograph of IR control showing strong positive reaction for iNOS in nuclei of pancreatic acini (arrows); (iii) photomicrograph of TEP-200 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pancreatic acini (arrows); (iv) photomicrograph of TEP-400 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pancreatic acini (arrows); (v) photomicrograph of PS-20 treated rat showing strong positive reaction for iNOS in nuclei and cytoplasm of pancreatic acini (arrows); (vi) photomicrograph of PS-40 treated rat showing mild positive reaction for iNOS in cytoplasm of some pancreatic acini (arrows) (IHC-peroxidase-DAB). (vii) Bars represent mean values of each group ± SEM. a p≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
Figure 6.
(i) photomicrograph of sham rat showing negative reaction for iNOS in pancreatic acini; (ii) photomicrograph of IR control showing strong positive reaction for iNOS in nuclei of pancreatic acini (arrows); (iii) photomicrograph of TEP-200 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pancreatic acini (arrows); (iv) photomicrograph of TEP-400 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pancreatic acini (arrows); (v) photomicrograph of PS-20 treated rat showing strong positive reaction for iNOS in nuclei and cytoplasm of pancreatic acini (arrows); (vi) photomicrograph of PS-40 treated rat showing mild positive reaction for iNOS in cytoplasm of some pancreatic acini (arrows) (IHC-peroxidase-DAB). (vii) Bars represent mean values of each group ± SEM. a p≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
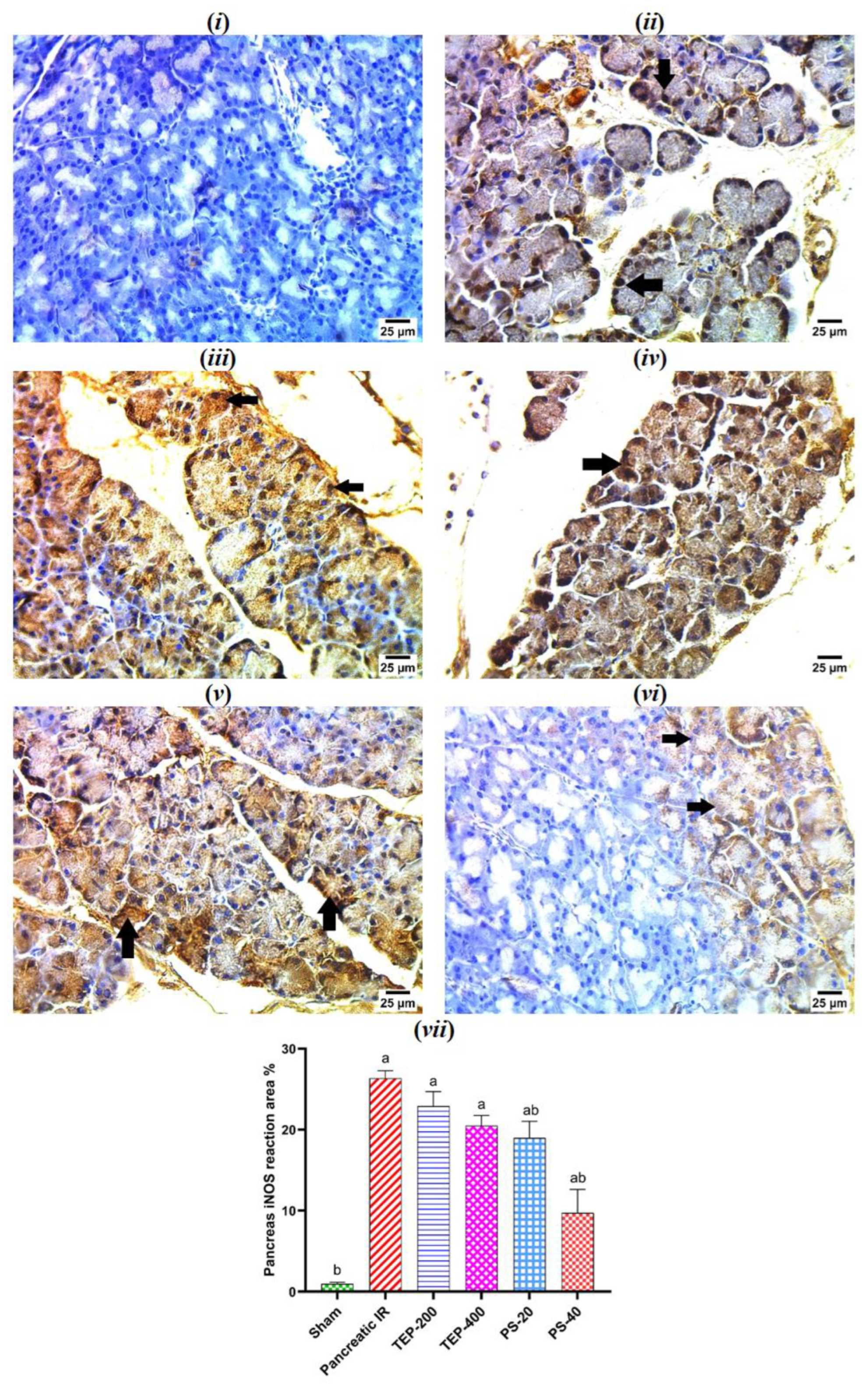
Figure 7.
(i) photomicrograph of sham rat showing negative reaction for iNOS in pneumocytes; (ii) photomicrograph of IR control rat showing strong positive reaction for iNOS in cytoplasm of pneumocytes; (iii) photomicrograph of TEP-200 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pneumocytes (arrows); (iv) photomicrograph of TEP-400 treated rat showing very mild positive reaction for iNOS in cytoplasm of few numbers of pneumocytes (arrows); (v) photomicrograph of PS-20 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pneumocytes (arrows); (vi) photomicrograph of PS-40 treated rat showing very mild positive reaction for iNOS in cytoplasm of pneumocytes (arrows) (IHC-peroxidase-DAB). (vii) Bars represent mean values of each group ± SEM. a p ≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
Figure 7.
(i) photomicrograph of sham rat showing negative reaction for iNOS in pneumocytes; (ii) photomicrograph of IR control rat showing strong positive reaction for iNOS in cytoplasm of pneumocytes; (iii) photomicrograph of TEP-200 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pneumocytes (arrows); (iv) photomicrograph of TEP-400 treated rat showing very mild positive reaction for iNOS in cytoplasm of few numbers of pneumocytes (arrows); (v) photomicrograph of PS-20 treated rat showing strong positive reaction for iNOS in cytoplasm and nuclei of pneumocytes (arrows); (vi) photomicrograph of PS-40 treated rat showing very mild positive reaction for iNOS in cytoplasm of pneumocytes (arrows) (IHC-peroxidase-DAB). (vii) Bars represent mean values of each group ± SEM. a p ≤0.05 versus sham, b p≤0.05 versus pancreatic IR.
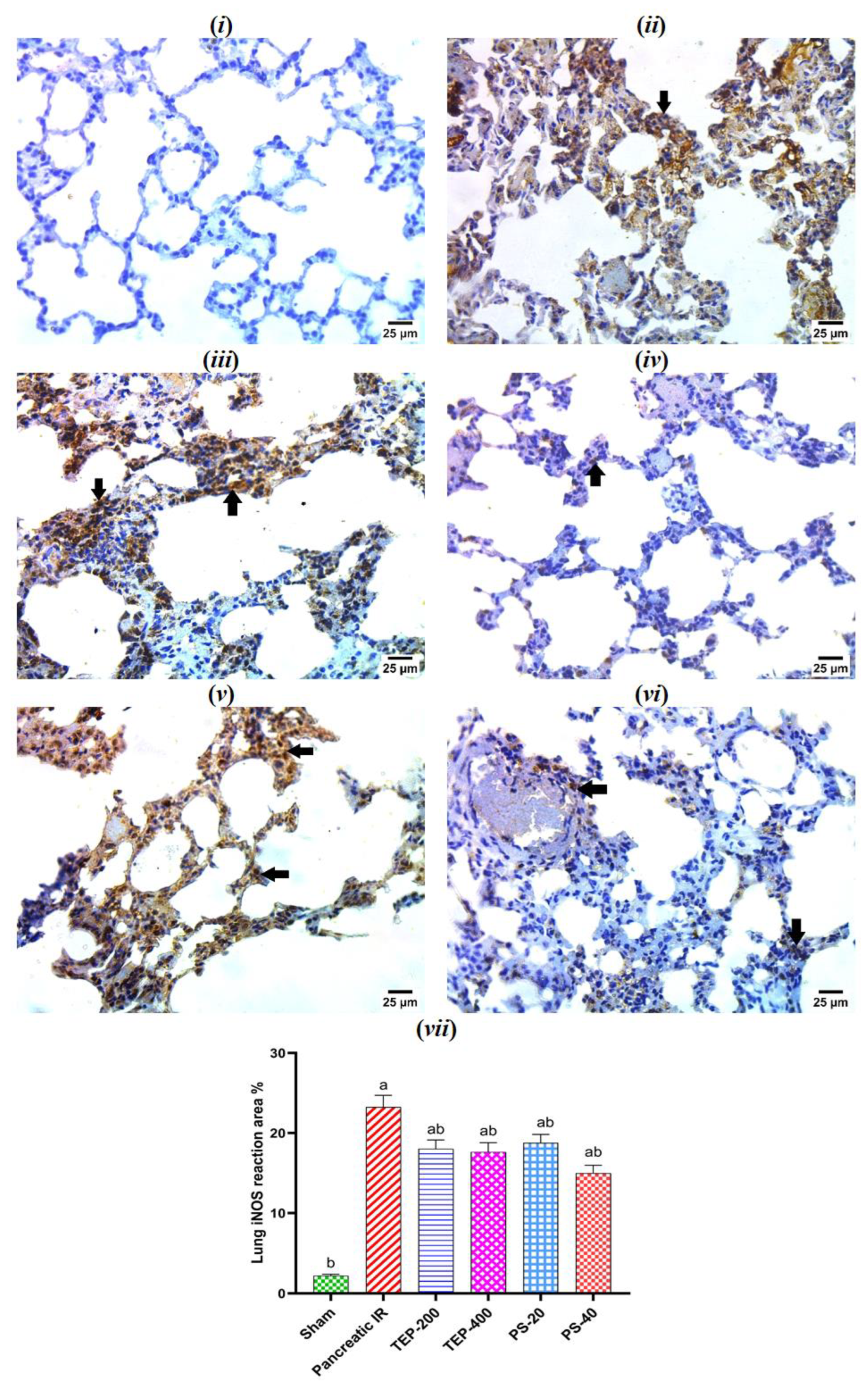

Table 1.
Effect of TEP and PS on the serum pancreatic enzymes of pancreatic IR rats.
| Group | Amylase (ng/ml) | Lipase (pg/ml) | TAP (ng/ml) |
|---|---|---|---|
| Sham | 5.0 b ±0.46 | 32.4 b ±1.57 | 0.4 b ±0.04 |
| IR control | 21.9 a ±1.11 | 144.4 a ±6.23 | 3.1 a ±0.09 |
| TEP-200 | 14.5 ab ±0.66 | 91.4 ab ±4.64 | 2.0 ab ±0.05 |
| TEP-400 | 7.6 b ±0.44 | 45.5 b ±1.85 | 1.0 ab ±0.07 |
| PS-20 | 14.1 ab ±0.55 | 90.4 ab ±4.15 | 2.2 ab ±0.10 |
| PS-40 | 7.3 b ±0.40 | 48.1 b ±1.87 | 1.4 ab ±0.06 |
Results are presented as Mean±SEM (n=6). a Significantly different in comparison with sham group (p≤0.05). b Significantly different in comparison with IR control group (p≤0.05).
Table 2.
Effect of TEP and PS on the pancreatic inflammatory markers of pancreatic IR rats.
| Group | Pancreatic TNF-α (pg/mg protein) | Pancreatic IL-1β (pg/mg protein) | Pancreatic NF-κB (ng/mg protein) |
|---|---|---|---|
| Sham | 30.3 b ±1.00 | 40.5 b ±1.56 | 29.6 b ±1.18 |
| IR control | 324.5 a ±5.37 | 212.9 a ±4.71 | 287.7 a ±15.34 |
| TEP-200 | 194.6 ab ±2.47 | 163.3 ab ±3.98 | 156.1 ab ±9.85 |
| TEP-400 | 72.2 ab ±6.50 | 56.5 b ±4.83 | 72.9 ab ±2.45 |
| PS-20 | 248.6 ab ±11.03 | 159.5 ab ±8.22 | 155.6 ab ±3.44 |
| PS-40 | 100.5 ab ±7.22 | 59.2 b ±3.60 | 73.6 ab ±3.84 |
Results are presented as Mean±SEM (n=6). a Significantly different in comparison with sham group (p≤0.05). b Significantly different in comparison with IR control group (p≤0.05).
Table 3.
Effect of TEP and PS on the pulmonary inflammatory markers of pancreatic IR rats.
| Group | Lung TNF-α (pg/mg protein) | Lung IL-1β (pg/mg protein) | Lung NF-κB (ng/mg protein) |
|---|---|---|---|
| Sham | 33.5 b ±2.77 | 31.5 b ±1.03 | 18.4 b ±0.95 |
| IR control | 177.3 a ±5.48 | 205.6 a ±3.88 | 90.6 a ±3.47 |
| TEP-200 | 72.1 ab ±4.39 | 136.6 ab ±3.43 | 50.9 ab ±2.18 |
| TEP-400 | 31.2 b ±1.28 | 51.7 ab ±4.11 | 33.1 ab ±2.77 |
| PS-20 | 96.0 ab ±4.98 | 148.6 ab ±3.73 | 62.0 ab ±2.12 |
| PS-40 | 70.3 ab ±2.67 | 53.8 ab ±3.69 | 34.3 ab ±1.71 |
Results are presented as Mean±SEM (n=6). a Significantly different in comparison with sham group (p≤0.05). b Significantly different in comparison with IR control group (p≤0.05).
Table 4.
Effect of TEP and PS on the pancreatic oxidation markers of pancreatic IR rats.
| Group | Pancreatic MDA (nmol/mg protein) | Pancreatic GPx (nmol/mg protein) | Pancreatic MPO (ng/mg protein) |
|---|---|---|---|
| Sham | 0.3 b ±0.01 | 3.0 b ±0.23 | 6.4 b ±0.31 |
| IR control | 1.8 a ±0.03 | 0.7 a ±0.03 | 0.9 a ±0.09 |
| TEP-200 | 1.1 ab ±0.05 | 1.9 ab ±0.12 | 2.9 ab ±0.08 |
| TEP-400 | 0.7 ab ±0.03 | 3.0 b ±0.09 | 4.9 ab ±0.18 |
| PS-20 | 1.3 ab ±0.05 | 1.9 ab ±0.09 | 3.0 ab ±0.23 |
| PS-40 | 0.8 ab ±0.03 | 2.6 b ±0.11 | 5.2 ab ±0.27 |
Results are presented as Mean±SEM (n=6). a Significantly different in comparison with sham group (p≤0.05). b Significantly different in comparison with IR control group (p≤0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



