
Submitted:
23 December 2024
Posted:
24 December 2024
You are already at the latest version
Abstract
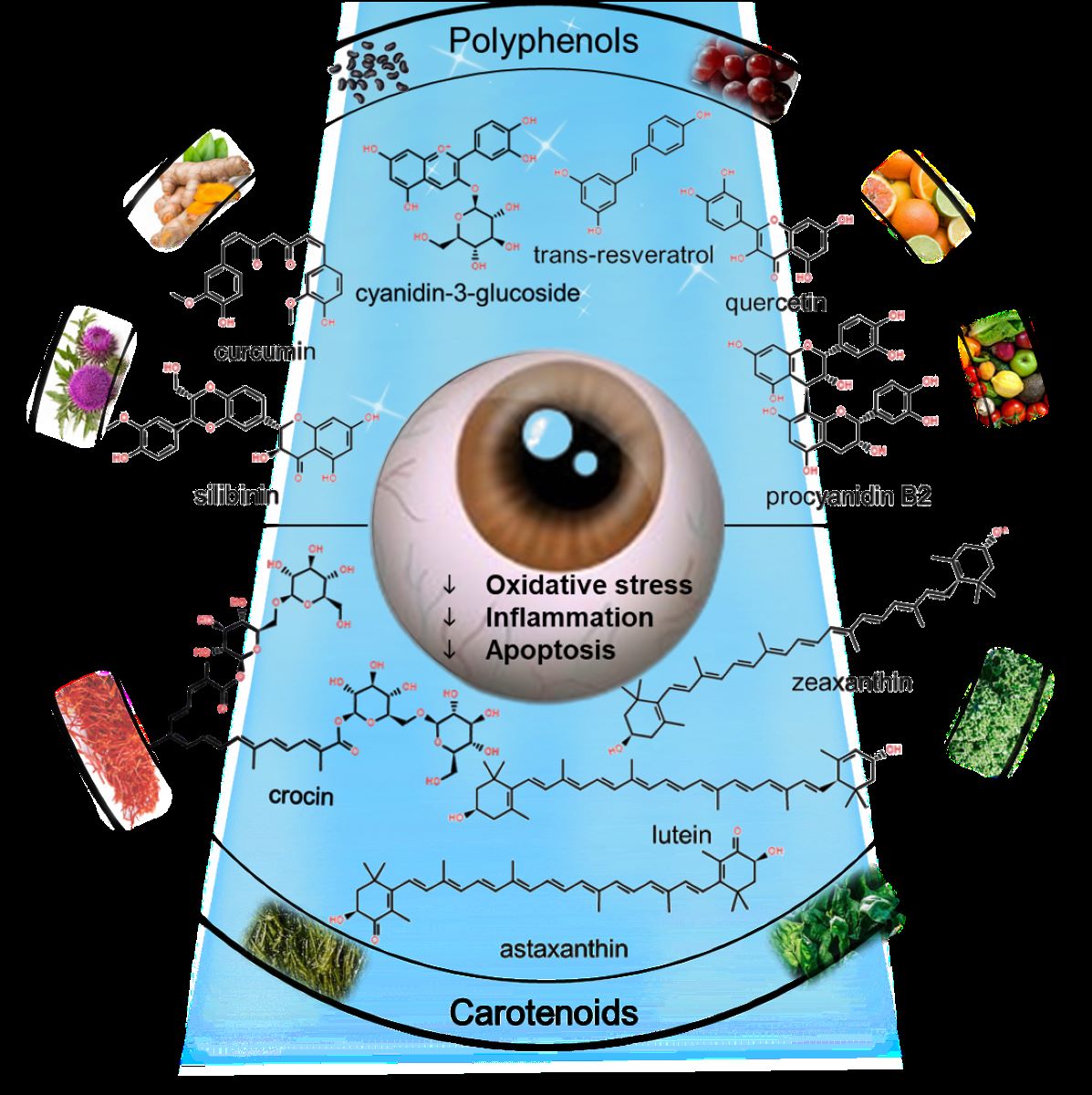
The increasing prevalence of blue light (BL) exposure from modern light-emitting diodes (LEDs) has raised significant concerns regarding its potential adverse effects on ocular health. Prolonged exposure to BL has been implicated in the pathogenesis of various retinal disorders, including age-related macular degeneration (AMD), primarily through mechanisms involving oxidative stress and inflammation mediated by the overproduction of reactive oxygen species (ROS). This review synthesizes current evidence on the photoprotective properties of plant extracts and di-etary bioactive compounds, with a focus on their potential to mitigate BL-induced retinal damage. Accumulating research suggests that dietary antioxidants, particularly carotenoids and poly-phenols, may offer photoprotective benefits. These phytochemicals act by neutralizing ROS and enhancing the retina's endogenous antioxidant capacity. Based on these findings, this review advocates for a food-first approach in future investigations, emphasizing the development of evidence-based dietary recommendations to bolster retinal health and mitigate the risk of BL-related ocular diseases.
Keywords:
blue light
; light-emitting diodes
; phototoxicity
; phytochemical
; polyphenol
; retina
1. Introduction
Blue light (BL), defined as high-energy visible light with wavelengths ranging from 400 to 500 nm, has become increasingly prevalent in contemporary society due to the widespread use of electronic devices and energy-efficient lighting solutions such as light-emitting diodes (LEDs) [1]. Unlike traditional incandescent bulbs, which emit a broader spectrum of light, LEDs are characterized by their high intensity and significant emission of BL [2]. This shift in lighting technology has raised critical concerns about the potential health implications of prolonged BL exposure, particularly in relation to visual function and retinal integrity [3].
Research has demonstrated that excessive exposure to BL can pass through the retina and lead to photochemical damage, contributing to the development of various ocular conditions, including age-related macular degeneration (AMD) and other forms of retinopathy [4]. The underlying mechanisms of BL-induced retinal damage are primarily associated with the generation of reactive oxygen species (ROS), which result from the phototoxic effects of blue light on retinal tissues [5]. These ROS can induce oxidative stress, leading to cellular apoptosis and inflammation, ultimately compromising retinal function [6].
The retinal pigment epithelium (RPE) plays a crucial role in maintaining retinal health by providing essential nutrients, assisting in phagocytosis of oxidized photoreceptor outer segments and regeneration of visual pigments to support visual function [7]. However, under conditions of elevated oxidative stress, the RPE may become dysfunctional, exacerbating retinal damage [8]. In this context, dietary bioactive compounds, particularly those with antioxidant properties, have garnered increasing attention for their potential protective effects against BL-induced toxicity. Polyphenols found in various fruits and vegetables have demonstrated significant antioxidant capacities that may effectively counteract the harmful effects of BL [9,10].
This review aims to comprehensively evaluate the current understanding of the protective roles of dietary bioactive compounds against BL toxicity. Certain antioxidants have been shown to effectively combat photodamage, suggesting that naturally occurring phytochemicals may also possess anti-BL toxicity properties [11,12]. Consumption of polyphenols may help prevent AMD [13]. The age-related eye disease study (AREDS) [14] and AREDS2 [15] trials have both revealed that taking an antioxidant supplement could reduce the risk of developing advanced AMD. Thus, phytogenic products and phytochemicals with a high potential for reducing BL toxicity need further investigation.
In this review, we summarize and discuss the photoprotective properties of dietary bioactive compounds against BL damage in vitro and in vivo, along with the underlying mechanisms.
2. Naturally Occurring Phytochemicals and Plant Extracts with Protective Potential Against BL-Induced Retinal Damage
Given the importance of photooxidative stress in retinal pathology, supplementing antioxidants in response to light challenges could be a possible strategy to avoid BL risks [16]. This review highlights current information on using naturally occurring phytochemicals (Table 1) and plant extracts (Table 3) to protect against BL photodamage and explores the possible mechanisms that underlie them in the following section.
2.1. Polyphenols
2.1.1. Resveratrol and Resveratrol Analogs
Resveratrol is a polyphenol compound abundant in grape skin. It is known for its antioxidant properties, beneficial biological functions, and low toxicity. In humans, resveratrol can be rapidly absorbed and metabolized into glucuronide and sulfated derivatives, which are eliminated via the urine [17]. Although peak plasma resveratrol levels may reach 25 ng/mL (0.11 μM), this is still below the quantities effective in vitro (> 1,000 ng/mL or 5–50 μM) [17,18]. The oral bioavailability of resveratrol is < 1% [19].
The effects of resveratrol on BL phototoxicity in humans and animals have yet to be elucidated. Resveratrol protects human ARPE-19 [20] and keratinocyte HaCaT cells [21] against oxidative damage caused by ultraviolet (UV) A. Kang et al. [22] demonstrated that resveratrol and its analog piceatannol protected A2E-laden ARPE-19 cells that were exposed to BL at a concentration range of 7.5–60 μM. Treatment with resveratrol and piceatannol reduces oxidized A2E product formation and intracellular A2E accumulation. However, the precise exact mechanism of action remains unclear. Among the compounds tested, resveratrol-3, 4′-O-β-d-diglucoside had the strongest photoprotective properties in ARPE-19 cells [22]. A case report of an 82-year-old patient with dry AMD revealed that a combination of nonblue LED light exposure at 590, 660, and 850 nm and resveratrol supplementation for 6 months improved retinal morphology and visual acuity [23]. Combining 100 mg microencapsulated trans-resveratrol with quercetin, ferulic acid, vitamin D3, and inositol hexaphosphate (a copper/iron/calcium chelator) improved choroidal blood flow and RPE function in patients with treatment-resistant macular degeneration [24]. Additionally, intravitreal injection or topical instillation of resveratrol at a dose of 5–50 mg/kg/d suppressed the development of retinal angiogenesis in mice [25].
Resveratrol is a phytoalexin produced by plants in response to environmental injuries including solar radiation or pathogen infection [26]. Resveratrol can be photoactivated using blue LED light and may be used in photodynamic therapy as an antimicrobial photosensitizer [27]. This may result in singlet oxygen photogeneration, raising health and safety concerns for persons who are frequently exposed to high-BL environments while taking high-dose resveratrol supplements. As a result, resveratrol may play a dual role as a photoprotective or phototoxic agent, depending on the action spectrum caused by light exposure.
2.1.2. Curcumin
Curcumin is a natural yellow pigment extracted from the rhizomes of Curcuma longa L. In a rodent model of acute photodamage, Wistar rats were exposed to 1,000 lux of cool white light for 3 h. Photoreceptor function and retinal morphology were assessed using ERG and quantitative histology. Mandal et al. [28] showed that dietary supplementation with 0.2% curcumin for 2 weeks prevented the loss of photoreceptor function and the thinning of the ONL from light-induced retinal degeneration. Curcumin also inhibited H2O2-induced cell death in 661 W photoreceptor and ARPE-19 cell lines by upregulating the expression of HO-1 and thioredoxin. Furthermore, 70% ethanol extracts of Curcuma longa L. and its curcuminoid compounds (curcumin, demethoxycurcumin, and bisdemethoxycurcumin) have conferred photoprotection by suppressing proapoptotic c-abl and p53 genes in A2E-laden ARPE-19 cells exposed to BL (420–470 nm; 9.4 mW/cm2). However, at concentrations over 20 μM, curcumin is cytotoxic to ARPE-19 cells [29].
Curcumin, which is phytochemically similar to resveratrol, is emerging as a photosensitizer capable of producing ROS when exposed to blue LED light and has been applied in antimicrobial photodynamic therapy [30,31] and food preservation [32]. The working concentrations of curcumin, ranging from 0.5 µM to 6.1 mM, have been effectively used in antimicrobial photodynamic therapy studies [33]. However, curcumin is phototoxic to mammalian cells [34,35]. Curcumin absorption spectra in aqueous buffers show a distinctive broad peak at 430 nm and a small shoulder at 355 nm. When curcumin is converted to its ketone form, the maximum absorption in the UV region occurs at approximately 389 nm [36]. Curcumin has drawbacks due to photodegradability, poor water solubility, and instability at physiological pH, resulting in rapid degradation and low bioavailability [37]. Curcumin conjugates and analogs have been developed to improve water dispersibility, chemical stability, absorption, and overall bioavailability [38]. However, when compared to other natural photosensitizers such as riboflavin, anthraquinones, and porphyrins, this fact may raise ocular safety concerns [33]. Several studies have addressed these questions, but the exact photobiological responses in the retina have yet to be determined.
2.1.3. Anthocyanins
Anthocyanins are water-soluble pigments abundant in red, blue, and purple fruits and vegetables such as berries, plums, and eggplants [39]. According to data from the Phenol-Explorer database between 2013 and 2016, the average dietary flavonoid intake among US adults is approximately 379.1 mg per 1,000 kcal/d [40]. Moreover, daily anthocyanin intake is estimated to range from 3–43 mg/d among nations [41]. The dietary intakes of the six major anthocyanins, including cyanidin, delphinidin, malvidin, pelargonidin, peonidin, and petunidin, among Australian adults, are estimated to be 9.33, 5.46, 14.01, 0.22, 1.97, and 1.96 mg/d, respectively [41].
The therapeutic potential of anthocyanins in ophthalmology has been demonstrated in in vivo and in vitro studies, including rhodopsin regeneration, smooth muscle relaxation, and blood circulation improvement studies [42]. In pigmented rabbits, oral administration of cyanidin-3-glucoside (50 mg/kg/d) and its metabolite ferulic acid (21.61 mg/kg/d) for 3 weeks ameliorated photoreceptor structural and functional abnormalities during short-term high-intensity light exposure. The activation of the Nrf2/HO-1 pathway and suppression of inflammatory responses are postulated to be protective mechanisms [43]. More than 250 known natural anthocyanins are found in glycoside form because free anthocyanins are unstable. A human 13C-tracer study conducted by Czank et al. [44] showed that the bioavailability of isotopically labeled cyanidin-3-glucoside was 12.38%, and the peaked serum concentration of ferulic acid was low (0.94 μM), 48 h after an intake of 500 mg cyanidin-3-glucoside. In a cellular model [45], cyanidin-3-glucoside at a concentration range of 10–50 μM was found to protect cells against BL damage from scavenging ROS, suppress A2E photooxidation, and prevent GSH reaction with photooxidized-A2E, leading to cell survival.
Nine monoglycosylated anthocyanins were identified in bilberry extracts, including cyanidin, delphinidin, malvidin, and petunidin. Malvidin-3-glucoside (oenin) provides the most potent photoprotection, and all other identified anthocyanins can also increase resistance to A2E-mediated membrane disruption [46]. Yacout and Gaillard [47] reported that, compared with pelargonidin 3-glucoside (callistephin), malvidin-3-glucoside more effectively protected ARPE-19 cells from BL-induced mitochondrial damage and inhibited ROS production. Additionally, treatment with anthocyanin-rich blueberry extracts delays cellular senescence in white LED light-exposed ARPE-19 cells [48] and protects 661 W cells from BL-induced cell death [49].
Grape skin contains a variety of polyphenols, including resveratrol, catechin, epicatechin, anthocyanin, quercetin, and phenolic acid; consequently, it possesses potent antioxidant properties [50]. According to Zhao et al. [51], grape skin extracts prevent BL-induced apoptosis in A2E-laden ARPE-19 cells. As grape skin extracts have no therapeutic action in GRP78 knockdown cell lines, the protective mechanisms are likely associated with the inhibition of the ER stress-mediated intrinsic apoptotic pathway. Yu et al. [52] observed that administering grape or lutein/zeaxanthin-rich marigold extracts prevented lipofuscin accumulation, hydroxynonenal-adduct formation, actin destabilization, and age-related photoreceptor dysfunction in β5−/− mice. These data suggest that a diet rich in polyphenol antioxidants benefits retinal health.
Previous studies have examined the anthocyanin levels in eyes. The total anthocyanin concentration in the ocular tissue of pigs fed 1–4% blueberries for 4 weeks was calculated to be 709 pg/g tissue (1.58 pmol/g tissue). Furthermore, of the 11 anthocyanins identified in the eye, malvidin glycosides were the most abundant [53]. In an ocular distribution study with Wistar rats given 100 mg/kg body weight (b.w.) of blackcurrant anthocyanins orally, the maximum concentration in the entire eye reached 115 ng/g tissue. The anthocyanin level in the retina following i.p. injection (500 mg/kg b.w.) was as high as 6.89 μg/g tissue, which was similar to that in plasma [54]. These results demonstrate that natural anthocyanins can cross the BRB, accumulate in the retina, and may act as neuroprotective agents against light damage.
2.1.4. Quercetin, quercetin-3-O-α-L-arabinopyranoside, and myricetin
A decade of research has focused on the use of quercetin in ophthalmic diseases, including cataracts, glaucoma, autoimmune Graves’ orbitopathy, keratoconus, retinoblastoma, and various other ocular surface diseases [55]. Systemic administration of quercetin (50 mg/kg b.w.) to rats for 6 d prior to a light insult protected the retinal tissue from photooxidative damage caused by high-intensity white fluorescent light (3,000 lux). The levels of 8-OHdG in the RPE and ONL, as well as the number of TUNEL-positive cells in the photoreceptors, were decreased in quercetin-treated rats. Moreover, quercetin restored ERG-based scotopic a- and b-wave losses [56]. At a dose of 50 μM quercetin protected A2E-laden ARPE-19 cells from BL cytotoxicity. Protective mechanisms are postulated to involve ROS scavenging and diminishing A2E-epoxide formation [45]. Given that reactive dicarbonyl species, including glyoxal and methylglyoxal, can be released during bis-retinoid photodegradation [57], Wang’s study found that quercetin reduced methylglyoxal-derived hydroimidazolone (MG-H1) protein adduct levels and RAGE gene expression in A2E-laden cells exposed to 430 nm BL irradiation [45]. Quercetin also protects ARPE-19 [58] and RGC-5 ganglion cells [59] from H2O2 oxidative damage and apoptotic cell death. However, in Ccl2/Cx3cr1−/− mice, which spontaneously develop AMD-like retinal lesions, the systemic administration of quercetin (25 mg/kg/d) did not alter the course of retinal lesions [58].
Quercetin-3-O-α-L-arabinopyranoside is a plant flavonoid found in Vaccinium uliginosum L. extracts and is abundant in berries such as guava. Kim et al. [60] used this flavone in a series of investigations. In vivo experiments revealed that orally administering quercetin-3-O-α-L-arabinopyranoside (25–100 mg/kg b.w.) reduced the thinning of the INL, OPL, and ONL in the retina, protecting against short-term high-intensity blue–green light (480 nm)-induced retinal damage in BALB/c mice. In the cellular model, quercetin-3-O-α-L-arabinopyranoside inhibited A2E internalization, protecting A2E-laden cells against BL-induced apoptosis.
Myricetin was first isolated from the bark of Myrica nagi Thunb. (Myricaceae); it is structurally related to quercetin, kaempferol, morin, and fisetin. Vegetables, berries, apples, oranges, nuts, and tea are the most common dietary sources of myricetin. The estimated daily consumption of myricetin in European and Korean adults is between 0.8 and 4 mg/d [61]. A recent in vitro study by Ortega et al. [62] suggested that myricetin and quercetin upregulate rhodopsin gene expression and maintain the proper folding and stability of rod opsins. HPLC revealed that myricetin and quercetin bind to unliganded opsins, facilitating retinal binding to such opsin proteins. The study demonstrated that giving myricetin and quercetin (0.02–2 mg/kg b.w.) before BL exposure protected the retina from morphologic abnormalities and preserved its function in Abca4-Rdh8−/− mice exposed to short-term high-intensity BL (10,000 lux for 45 min). Myricetin was found at a concentration of approximately 30 pmol in the ocular tissue 30 minutes after intraperitoneal injection (using HPLC-MS), showing that this substance can cross the BRB However, quercetin is not easily detected, possibly owing to its detection limit. The protective effects of myricetin and quercetin are probably linked to the upregulation of photoreceptor-specific proteins (rhodopsin and cone opsins) and downregulation of proinflammatory chemokines and cytokines, leading to BL-mediated apoptosis suppression [63]. A daily oral dose of myricetin (25–100 mg/kg b.w./d) for 6 weeks prevented intraocular pressure elevation and reduced oxidative stress in rats with hyaluronic acid-induced glaucoma, according to a different study on myricetin eye protection [64]. In rats, a single intraperitoneal injection of high-dose myricetin (20 mg/kg b.w.) altered circadian rhythms by reducing nocturnal serum melatonin levels and locomotor activity by decreasing acetyl coenzyme A: arylalkylamine N-acetyltransferase activity [65].
Laabich et al. [66] found that when primary bovine retinal cells were exposed to BL (420 nm; 500 lux, 17.4 W/m2) for 20 h, 75% of the photoreceptors perished. This phototoxicity is remarkedly attenuated in cells pretreated with myricetin (5‒40 μM), whereas quercetin and kaempferol have only mild beneficial effects at the same concentrations. The number of hydroxyl groups in their structures may be associated with the antioxidant activity of flavonoids (that is, their ability to scavenge free radicals) [66]. Myricetin has three hydroxyl groups in its B ring and is hypothesized to be more effective than quercetin and kaempferol in response to BL challenge. Furthermore, myricetin and quercetin have greater free radical-scavenging properties than those of kaempferol [67]. Additionally, A2E causes dose-dependent primary bovine photoreceptor and bipolar cell apoptosis. In the absence of BL, treatment with myricetin, quercetin, and kaempferol protects cells from A2E toxicity.
2.1.5. Cynaroside
Cynaroside, also known as luteolin 7-glucoside or luteoloside, is a glycosyloxyflavone of luteolin found in numerous fruits and cruciferous vegetables, such as berries, lemons, oranges, grapes, red cabbage, kale, spinach, lettuce, and carrots. According to the scientific literature, cynaroside exhibits antioxidant and antibacterial properties [68]. In the A2E-laden ARPE-19 cell model, pretreatment with 10 and 20 μM of cynaroside inhibits NF-κB signaling and NLRP3 inflammasome activation, normalizes oxidative stress-related markers, and reduces apoptotic cell death caused by BL (430 nm, 2500 lux for 2 h) [69]. Cynaroside also prevents H2O2-induced cytotoxicity in ARPE-19 cells [70]. Similar antiapoptotic and antioxidative effects were observed in rats administered 5 μg cynaroside intravitreally while exposed to high-intensity BL irradiation (10,000 lux, 2 h/d) for 7 d.
2.1.6. Procyanidin B2
One of the principal proanthocyanidins found in grape seeds is procyanidin B2, a B-type proanthocyanidin consisting of two (−)-epicatechin molecules [71]. Presently, there are no in vivo studies on procyanidin B2 focusing on photodamage, and data on the distribution of procyanidins in the retina are unavailable. However, Yamakoshi et al. [72] reported that procyanidin-rich grape seed extracts exhibited anticataract activity in hereditary cataractous rats. The antiglycation properties of procyanidin B2 prevented the onset of cataracts and retinopathy in rats with streptozotocin-induced diabetes. Procyanidin-B2 reduced the expression of GFAP and vascular endothelial growth factor (VEGF) in the diabetic retina [73]. These findings suggest that procyanidins or their active metabolites can cross the BRB and enter ocular tissues.
In an A2E-laden ARPE-19 cell model, treatment with procyanidin B2 [74] or grape skin extracts containing resveratrol, catechin, epicatechin, anthocyanins, quercetin, oligomeric procyanidins, and phenolic acids [51] prevented BL-induced ROS generation, ER stress, and mitochondria-dependent apoptosis. Mice fed a grape-enriched diet showed improved photoreceptor survival and ERG-based functions in a model of retinal degeneration induced by subretinal paraquat injection [75]. Among the 19 natural compounds isolated from ginkgo biloba extracts, procyanidin B2 is particularly effective in preventing tert-butyl hydroperoxide-induced oxidative stress and ARPE-19 cell death at a concentration of 10 μM [76]. Sea buckthorn (Hippophae rhamnoides L.) extracts, which contain 39% proanthocyanidins, protected the retina of pigmented rabbits from extremely high-intensity visible light (18,000 lux)-induced photoreceptor lesions and suppressed lipid peroxidation and proinflammatory cytokine levels [77]. Without being conjugated or methylated, procyanidin B2 is only marginally absorbed [78]. As a result, the health effects of procyanidin B2 may be attributed, at least in part, to its metabolites rather than to procyanidin B2 itself. Spencer et al. [79], determined that (−)-epicatechin monomer is the predominant metabolite of procyanidin B2 in an isolated procyanidin B2-perfused small intestine. Furthermore, 53 metabolites exhibiting methylation, sulfation, hydration, hydroxylation, hydrogenation, and glucuronidation were detected using UPLC-DAD-ESI-IT-TOF in mice administered pure procyanidin B2 [80].
2.1.7. Phloroglucinol
Phloroglucinol is abundant in the edible alga Ecklonia cava. Piao et al. [81] demonstrated that phloroglucinol protects human HaCaT keratinocytes from UVB radiation by modulating MAPK-AP-1-MMP-1 signaling. Furthermore, Cia et al. [82] reported that phloroglucinol protects primary cultures of RPE and photoreceptor cells against all-trans-retinal-induced ROS formation and cytotoxicity. In cotreatment culture media, ~60% of all-trans-retinal can be trapped by phloroglucinol to produce a stable chromene adduct in an equimolar ratio, as demonstrated by 1H, 13C NMR spectra, and mass analysis. Additionally, phloroglucinol inhibits A2E synthesis in vitro by competing with ethanolamine and all-trans-retinal for binding to the retinal carbonyl group. Because phloroglucinol has low bioavailability and poor lipid solubility, it remains to be determined whether the concentration used in these tests (50 μg/mL) can be detected in a physiologically relevant context. Taveau et al. [83] synthesized a phloroglucinol-based lipophenol derivative, isopropyl-phloroglucinol-DHA, to investigate its protective efficacy against acute photodamage in albino Abca4−/− mice. They found that intravenous injection of isopropyl-phloroglucinol-DHA (5–30 mg/kg b.w.) inhibited photoreceptor degradation and ERG-based reduction of a- and b-wave amplitudes caused by 470 nm cool white fluorescent lamps. These photoprotective effects persisted for 3 months following a light insult and are comparable to the beneficial effects of emixustat, a therapeutic drug used for treating Stargardt disease [84].
2.1.8. Silibinin
Silibinin, also known as silybin, is the primary bioactive flavonoid compound found in 60–70% of Silybum marianum L. (milk thistle) varieties. Silibinin is composed of two diastereomers, silybin A and silybin B, in an approximately equimolar ratio and has been reported to exhibit multiple health benefits such as antioxidant, anti-inflammatory, hepatoprotective, neuroprotective, and antiviral activities [85]. Under hypoxic conditions, treating immortalized RPE cells with silibinin inhibited VEGF secretion by modulating the PI3K-Akt-mTOR-p70S6K signaling pathway [86]. Silibinin protects retinal ganglion cells against BL-induced inflammatory responses and apoptosis via the MEK-ERK-CREB pathway [87]. According to Tvrdy et al. [88], the oral bioavailability of silymarin flavonolignans in humans and rats ranges from 0.45% to 5%, likely owing to their poor aqueous solubility. Nevertheless, in vitro biological effects observed at silibinin concentrations greater than 20–30 μM have little therapeutic relevance because this flavonoid’s maximal achievable plasma concentration is around 10 μM [85].

Table 1.
Protective effects of polyphenolic compounds against blue and white LED light-induced retinal photodamage in vitro and in vivo.
Table 1.
Protective effects of polyphenolic compounds against blue and white LED light-induced retinal photodamage in vitro and in vivo.
| Compounds | Phototoxic lighting | Experimental models | Effects | References |
|---|---|---|---|---|
| Resveratrol | A2E (10 or 20 µM) + BL (430 nm, 4.02 J/cm2, 7 min for 3 d) | ARPE-19 cells | ↓ Oxidized A2E formation and intracellular A2E accumulation | [22] |
| Curcumin | Cool white LED (1,000 lux, 3 h, 09:00–12:00) | Wistar rats | Inhibits NF-κB activation and downregulates the inflammatory gene expression | [28] |
| A2E (20 µM) + BL (450 nm, 9.4 mW/cm2) | ARPE-19 cells | Inhibits c-Abl and p53 mRNA expression to prevent apoptosis | [29] | |
| Cyanidin-3-glucoside | A2E (3 μM) + BL (430 nm, 1.5 mW/cm2, 30 min) | ARPE-19 cells | Scavenges ROS and inhibits A2E photooxidation | [45] |
| Malvidin-3-glucoside | BL for 30 min | ARPE-19 cells | Inhibits mitochondrial damage and ROS production | [47] |
| Quercetin | White fluorescent light (3,000 lux) | Sprague–Dawley rats | ↓ 8-OHdG level & TUNEL-positive cells | [56] |
| A2E (3 μM) + BL (430 nm, 1.5 mW/cm2, 30 min) | ARPE-19 cells | Inhibits proinflammatory cytokines release and cell apoptosis Inhibits MG-H1 levels & RAGE mRNA expression |
[45] | |
| Quercetin-3-O-α-L-arabinopyranoside | Blue-green light (480 nm, 4,000 lux, 10 min) | ARPE-19 cells Balb-c mice |
Inhibits NF-κB, AP-1, and C3 expression Inhibits the pyrolysis of poly polymerases Inhibits the thinning of the INL, OPL, and ONL in the mouse retina |
[60] |
| Myricetin and quercetin | Bright light (10,000 lux, 45 min) | Abca4−/−Rdh8−/− mice | Inhibits inflammatory responses and photoreceptor apoptosis | [63] |
| Myricetin | BL (420 nm, 500 lux, 17.4 W/m2, 20 h); A2E (30 μM) | Primary bovine retinal cells | Prevents BL-induced and A2E-induced death of primary photoreceptor cells in the retina | [66] |
| Cynaroside | A2E (25 μM) + BL (430 nm, 2500 lux, 2 h) | ARPE-19 cells | Impedes NF-κB signaling and NLRP3 inflammasome activation Normalizes oxidative stress-related markers Inhibits apoptotic cell death |
[69] |
| Procyanidin B2 | A2E (25 μM) + BL (2000 lux, 30 min) | ARPE-19 cells | Prevents ROS generation, ER stress, and mitochondria-dependent apoptosis | [74] |
| Silibinin | BL (530 nm with a peak of 470 nm, 12.08 W/m2, 24 h) | Retinal ganglion cells | Inhibits apoptosis | [87] |
A2E, N-retinylidene-N-retinylethanolamine; BL, blue light; ARPE, adult retinal pigment epithelial; LED, light-emitting diodes; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; ROS, reactive oxygen species; 8-OHdG, 8-hydroxy-2-deoxyguanosine; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; MG-H1, methylglyoxal-derived hydroimidazolones; RAGE, receptor for advanced glycation end products; AP-1, activator protein 1; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer.
2.2. Lipoic Acid
Lipoic acid is an amphiphilic molecule and metabolic enzyme cofactor with potent antioxidant, anti-inflammatory, and metal-chelating properties. According to clinical investigations, a single oral dose of 2,400 mg or daily doses of 1,800 mg of lipoic acid for 6 months had no adverse effects on individuals [89]. Because lipoic acid can cross the blood–brain barrier, it may exhibit neuroprotective effects in the central nervous system and eyes [90]. Supplementation with lipoic acid suppressed malondialdehyde (MDA) production, enhanced antioxidant enzyme activities, and lowered corneal and scleral damage induced by UVA [91] and UVB [92] in rabbits and mice. In streptozocin-induced diabetic rats, lipoic acid treatment normalized intraretinal manganese ion uptake [93] and oxygenation response [94] without losing ONL thickness or BRB integrity. By reducing apoptosis, inflammation, oxidative stress, and iron-related gene expression, intraperitoneal injection of lipoic acid at a dose of 100 mg/kg b.w. preserved ERG functions and photoreceptor survival in BALB/c mice with acute light damage (10,000 lux cool white LED light for 4 h) [95]. These findings further support the hypothesis that lipoic acid-rich extracts, such as those from Ginkgo biloba, red berries, and white willow bark, may have photoprotective potential that requires further investigation.
2.3. Prunella vulgaris L. Extracts
Prunella vulgaris L. is a widely grown perennial plant that is commonly used as an ingredient in foods, drinks, and traditional medicines. P. vulgaris extracts enhance wound healing, alleviate sore throat, fever, and skin allergies, and exert antioxidant, anti-inflammatory, and antiallergic effects; thus, they possess substantial therapeutic potential [96]. According to Kim et al. [97], water extracts of P. vulgaris prevent BL-induced apoptosis in A2E-laden ARPE-19 cells and reduce the internalization of A2E. In addition, P. vulgaris extracts (50–200 mg/kg bw) activate Nrf2/HO-1 signaling and inhibit NF-κB nuclear translocation in mice exposed to short-term, high-intensity BL. The thickness of ONL and the levels of oxidative markers (ROS, MDA, and GSH), inflammatory cytokines (IL-1, IL-6, and MCP-1), and angiogenesis factors (HIF-1α and VEGF) are improved in P. vulgaris extracts-treated groups compared to those in non-treated groups.
Table 3.
Protective effects of plant extracts against blue LED light-induced retinal photodamage in vitro and in vivo.
Table 3.
Protective effects of plant extracts against blue LED light-induced retinal photodamage in vitro and in vivo.
| Plant extracts | Phototoxic lighting | Experimental models | Effects | References |
|---|---|---|---|---|
| Blueberry anthocyanin-rich extracts | White LED (420–800 nm, 2,500 lux for 12 h) | ARPE-19 cells | Inhibits cellular senescence | [48] |
| Vaccinium myrtillus L. (bilberry) and Vaccinium vitis-idaea (lingonberry) | BL (460–470 nm, 2500 lux, 6 h) | Photoreceptor 661W cells | Inhibits the generation of ROS and regulates the activation of NF-κB, p38, MAPK, and caspase-3/7 to protect retinal photoreceptor cells | [49] |
| Grape skin | BL (2000 lux, 30 min) | ARPE-19 cells | Inhibits the ER-stress-mediated intrinsic apoptotic pathway | [51] |
| Prunella vulgaris var. L | A2E (20 μM) + BL (430 nm, 4000 lux, 10 min) | ARPE-19 cells | Activates Nrf2/HO-1 signaling, inhibits ROS and MDA production ↓ inflammation |
[97] |
| BL (430 nm, 10000 lux, 1 h/d for 14 d) | BALB/c mice |
BL, blue light; ROS, reactive oxygen species; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; MAPK, mitogen-activated protein kinase; ARPE, adult retinal pigment epithelial; Bax, Bcl-2 associated X; Bcal-2, B-cell lymphoma 2; FasL, Fas ligand; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; ONL, outer nuclear layer; ER, endoplasmic reticulum; A2E, N-retinylidene-N-retinylethanolamine; Nrf2, NFE2-related factor 2; HO-1, heme oxygenase-1; MDA, malondialdehyde.
5. Conclusions
Prolonged exposure to the screen can lead to visual impairments, as the chronic phototoxic effects of repeated BL exposure contribute to the deterioration of retinal functions and morphological integrity. Given the pivotal role of photooxidative stress in the pathogenesis of retinal disorders, the consumption of antioxidant-rich foods or adherence to phytochemical-rich diet may offer a strategic approach to mitigating the detrimental effects of blue light exposure. This perspective emphasizes identifying naturally occurring phytochemicals with potential photoprotective properties as a promising avenue for reducing light-induced retinal damage.
Author Contributions
Wan-Ju Yeh: Writing–original draft, Investigation, Conceptualization. Cin Yan: Investigation. Chi-Hao Wu: Writing–review & editing, Writing–original draft, Validation, Supervision, Investigation, Conceptualization.
Acknowledgments
Wan-Ju Yeh: Writing–original draft, Investigation, Conceptualization. Cin Yan: Investigation. Chi-Hao Wu: Writing–review & editing, Writing–original draft, Validation, Supervision, Investigation, Conceptualization. This research work was supported by the grants NSTC 112-2320-B-003-001 and NSTC 113-2320-B-003-002-MY3 from the National Science and Technology Council, Taiwan.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cougnard-Gregoire, A.; Merle, B.M.J.; Aslam, T.; Seddon, J.M.; Aknin, I.; Klaver, C.C.W.; Garhöfer, G.; Layana, A.G.; Minnella, A.M.; Silva, R.; Delcourt, C. Blue light exposure: ocular hazards and prevention-A narrative review. Ophthalmol. Ther. 2023, 12, 755–788. [Google Scholar] [CrossRef]
- Behar-Cohen, F.; Martinsons, C.; Viénot, F.; Zissis, G.; Barlier-Salsi, A.; Cesarini, J.P.; Enouf, O.; Garcia, M.; Picaud, S.; Attia, D. Light-emitting diodes (LED) for domestic lighting: any risks for the eye? Prog. Retin. Eye Res. 2011, 30, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.A.; Bahmani, H. A review of the current state of research on artificial blue light safety as it applies to digital devices. Heliyon 2022, 8, e10282. [Google Scholar] [CrossRef] [PubMed]
- Yeh, W.J.; Chien, P.T.; Wen, Y.T.; Wu, C.H. A comprehensive review of experimental models for investigating blue light-induced ocular damage: Insights into parameters, limitations, and new opportunities. Exp. Eye Res. 2024, 249, 110142. [Google Scholar] [CrossRef]
- Ratnayake, K.; Payton, J.L.; Meger, M.E.; Godage, N.H.; Gionfriddo, E.; Karunarathne, A. Blue light-triggered photochemistry and cytotoxicity of retinal. Cell. Signal. 2020, 69, 109547. [Google Scholar] [CrossRef] [PubMed]
- Marie, M.; Bigot, K.; Angebault, C.; Barrau, C.; Gondouin, P.; Pagan, D.; Fouquet, S.; Villette, T.; Sahel, J.A.; Lenaers, G.; Picaud, S. Light action spectrum on oxidative stress and mitochondrial damage in A2E-loaded retinal pigment epithelium cells. Cell. Death. Dis. 2018, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Bao, X.L.; Cong, Y.Y.; Fan, B.; Li, G.Y. Autophagy in age-related macular degeneration: A regulatory mechanism of oxidative stress. Oxid. Med. Cell. Longev. 2020, 2020, 2896036. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, D.; Zhang, Y.; Zhang, L.; Liao, Z.; Aihemaitijiang, S.; Hou, Y.; Zhan, Z.; Xie, K.; Zhang, Z. Lutein protected the retina from light induced retinal damage by inhibiting increasing oxidative stress and inflammation. J. Funct. Foods 2020, 73, 104107. [Google Scholar] [CrossRef]
- Ouyang, X.; Yang, J.; Hong, Z.; Wu, Y.; Xie, Y.; Wang, G. Mechanisms of blue light-induced eye hazard and protective measures: a review. Biomed. Pharmacother. 2020, 130, 110577. [Google Scholar] [CrossRef]
- Cabrera, M.P.; Chihuailaf, R.H. Antioxidants and the integrity of ocular tissues. Vet. Med. Int. 2011, 2011, 905153. [Google Scholar] [CrossRef]
- Bungau, S.; Abdel-Daim, M.M.; Tit, D.M.; Ghanem, E.; Sato, S.; Maruyama-Inoue, M.; Yamane, S.; Kadonosono, K. Health benefits of polyphenols and carotenoids in age-related eye diseases. Oxid. Med. Cell Longev. 2019, 2019, 9783429. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study Research, G. The Age-Related Eye Disease Study (AREDS): design implications. AREDS report no. 1. Control Clin. Trials 1999, 20, 573–600. [Google Scholar]
- Group, A.R.; Chew, E.Y.; Clemons, T.; SanGiovanni, J.P.; Danis, R.; Domalpally, A.; McBee, W.; Sperduto, R.; Ferris, F.L. The Age-Related Eye Disease Study 2 (AREDS2): study design and baseline characteristics (AREDS2 report number 1). Ophthalmology 2012, 119, 2282–2289. [Google Scholar]
- Choo, P.P.; Woi, P.J.; Bastion, M.C.; Omar, R.; Mustapha, M.; Md Din, N. Review of evidence for the usage of antioxidants for eye aging. Biomed. Res. Int. 2022, 2022, 5810373. [Google Scholar] [CrossRef]
- Pintea, A.; Rugină, D.: Resveratrol and the human retina. In Handbook of Nutrition, Diet, and the Eye. Elsevier; 2019: 127-145.
- Alarcon De La Lastra, C.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Chan, C.M.; Huang, C.H.; Li, H.J.; Hsiao, C.Y.; Su, C.C.; Lee, P.L.; Hung, C.F. Protective effects of resveratrol against UVA-induced damage in ARPE19 cells. Int. J. Mol. Sci. 2015, 16, 5789–5802. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chan, F.; Sun, H.; Yan, J.; Fan, D.; Zhao, D.; An, J.; Zhou, D. Resveratrol protects human keratinocytes HaCaT cells from UVA-induced oxidative stress damage by downregulating Keap1 expression. Eur. J. Pharmacol. 2011, 650, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Choung, S.Y. Protective effects of resveratrol and its analogs on age-related macular degeneration in vitro. Arch. Pharm. Res. 2016, 39, 1703–1715. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, R.; Bertelli, M.; Scaffidi, E.; Bumah, V.V.; Biagioni, F.; Busceti, C.L.; Puglisi-Allegra, S.; Fornai, F. The neurobiology of nutraceuticals combined with light exposure, a case report in the course of retinal degeneration. Arch. Ital. Biol. 2021, 159, 134–150. [Google Scholar] [CrossRef]
- Richer, S.; Stiles, W.; Ulanski, L.; Carroll, D.; Podella, C. Observation of human retinal remodeling in octogenarians with a resveratrol based nutritional supplement. Nutrients 2013, 5, 1989–2005. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Zhang, X.Y.; Leung, K.W.; Duan, R.; Dong, T.T.; Qin, Q.W.; Tsim, K.W. Resveratrol, an inhibitor binding to VEGF, restores the pathology of abnormal angiogenesis in retinopathy of prematurity (ROP) in mice: Application by intravitreal and topical instillation. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Shakibaei, M.; Harikumar, K.B.; Aggarwal, B.B. Resveratrol addiction: to die or not to die. Mol. Nutr. Food Res. 2009, 53, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, D.P.; Soares Lopes, D.P.; de Moraes, R.C.J.; Vieira Goncalves, C.; Pereira Rosa, L.; da Silva Rosa, F.C.; da Silva, R.A.A. Photoactivated resveratrol against Staphylococcus aureus infection in mice. Photodiagnosis Photodyn. Ther. 2019, 25, 227–236. [Google Scholar] [CrossRef]
- Mandal, M.N.; Patlolla, J.M.; Zheng, L.; Agbaga, M.P.; Tran, J.T.; Wicker, L.; Kasus-Jacobi, A.; Elliott, M.H.; Rao, C.V.; Anderson, R.E. Curcumin protects retinal cells from light-and oxidant stress-induced cell death. Free Radic. Biol. Med. 2009, 46, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Lee, E.H.; Kim, S.R.; Jang, Y.P. Anti-apoptotic effects of Curcuma longa L. extract and its curcuminoids against blue light-induced cytotoxicity in A2E-laden human retinal pigment epithelial cells. J. Pharm. Pharmacol. 2017, 69, 334–340. [Google Scholar] [PubMed]
- Souza, E.Q.M.; da Rocha, T.E.; Toro, L.F.; Guiati, I.Z.; Freire, J.d.O.A.; Ervolino, E.; Brandini, D.A.; Garcia, V.G.; Theodoro, L.H. Adjuvant effects of curcumin as a photoantimicrobial or irrigant in the non-surgical treatment of periodontitis: Systematic review and meta-analysis. Photodiagnosis Photodyn. Ther. 2021, 34, 102265. [Google Scholar] [CrossRef] [PubMed]
- Muniz, I.P.R.; Galantini, M.P.L.; Ribeiro, I.S.; Goncalves, C.V.; Dos Santos, D.P.; Moura, T.C.; Silva, E.S.; Silva, N.R.; Cipriano, B.P.; Correia, T.M.L.; et al. Antimicrobial photodynamic therapy (aPDT) with curcumin controls intradermal infection by Staphylococcus aureus in mice with type 1 diabetes mellitus: a pilot study. J. Photochem. Photobiol. B. 2021, 224, 112325. [Google Scholar] [CrossRef] [PubMed]
- Cossu, M.; Ledda, L.; Cossu, A. Emerging trends in the photodynamic inactivation (PDI) applied to the food decontamination. Food. Res. Int. 2021, 144, 110358. [Google Scholar] [CrossRef] [PubMed]
- Polat, E.; Kang, K. Natural photosensitizers in antimicrobial photodynamic therapy. Biomedicines 2021, 9. [Google Scholar] [CrossRef]
- Dahl, T.A.; Bilski, P.; Reszka, K.J.; Chignell, C.F. Photocytotoxicity of curcumin. Photochem. Photobiol. 1994, 59, 290–294. [Google Scholar] [CrossRef]
- Banerjee, S.; Prasad, P.; Hussain, A.; Khan, I.; Kondaiah, P.; Chakravarty, A.R. Remarkable photocytotoxicity of curcumin in HeLa cells in visible light and arresting its degradation on oxovanadium(IV) complex formation. Chem. Commun. (Camb) 2012, 48, 7702–7704. [Google Scholar] [CrossRef]
- Seidi Damyeh, M.; Mereddy, R.; Netzel, M.E.; Sultanbawa, Y. An insight into curcumin-based photosensitization as a promising and green food preservation technology. Compr. Rev. Food Sci. 2020, 19, 1727–1759. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Noureddin, S.A.; El Shishtawy, R.M.; Al Footy, K.O. Curcumin analogues and their hybrid molecules as multifunctional drugs. Eur. J. Med. Chem. 2019, 182, 111631. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Heinonen, M. Berry phenolics and their antioxidant activity. J. Agric. Food Chem. 2001, 49, 4076–4082. [Google Scholar] [CrossRef]
- Huang, Q.; Braffett, B.H.; Simmens, S.J.; Young, H.A.; Ogden, C.L. Dietary polyphenol intake in US adults and 10-year trends: 2007-2016. J. Acad. Nutr. Diet 2020, 120, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- Igwe, E.O.; Charlton, K.E.; Probst, Y.C. Usual dietary anthocyanin intake, sources and their association with blood pressure in a representative sample of Australian adults. J. Hum. Nutr. Diet 2019, 32, 578–590. [Google Scholar] [CrossRef]
- Nomi, Y.; Iwasaki-Kurashige, K.; Matsumoto, H. Therapeutic effects of anthocyanins for vision and eye health. Molecules 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huo, Y.; Zhao, L.; Lu, F.; Wang, O.; Yang, X.; Ji, B.; Zhou, F. Cyanidin-3-glucoside and its phenolic acid metabolites attenuate visible light-induced retinal degeneration in vivo via activation of Nrf2/HO-1 pathway and NF-kappaB suppression. Mol. Nutr. Food Res. 2016, 60, 1564–1577. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: a (13)C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kim, H.J.; Sparrow, J.R. Quercetin and cyanidin-3-glucoside protect against photooxidation and photodegradation of A2E in retinal pigment epithelial cells. Exp. Eye Res. 2017, 160, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.P.; Zhou, J.; Nakanishi, K.; Sparrow, J.R. Anthocyanins protect against A2E photooxidation and membrane permeabilization in retinal pigment epithelial cells. Photochem. Photobiol. 2005, 81, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Yacout, S.M.; Gaillard, E.R. The Anthocyanins, oenin and callistephin, protect RPE cells against oxidative stress. Photochem. Photobiol. 2017, 93, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, X.; Zhang, D.; Zhou, F.; Wang, D.; Wei, Y.; Gao, F.; Xie, L.; Jia, G.; Wu, W.; Ji, B. Blueberry anthocyanins: protection against ageing and light-induced damage in retinal pigment epithelial cells. Br. J. Nutr. 2012, 108, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Kuse, Y.; Tsuruma, K.; Kobayashi, S.; Shimazawa, M.; Hara, H. Protective effects of bilberry and lingonberry extracts against blue light-emitting diode light-induced retinal photoreceptor cell damage in vitro. BMC Complement. Altern. Med. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Katalinić, V.; Možina, S.S.; Skroza, D.; Generalić, I.; Abramovič, H.; Miloš, M.; Ljubenkov, I.; Piskernik, S.; Pezo, I.; Terpinc, P.; Boban, M. Polyphenolic profile, antioxidant properties and antimicrobial activity of grape skin extracts of 14 Vitis vinifera varieties grown in Dalmatia (Croatia). Food Chem. 2010, 119, 715–723. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, T.; Jiang, Y.; Wu, L.; Cai, X.; Sun, X.; Sun, X. Photooxidative damage in retinal pigment epithelial cells via GRP78 and the protective role of grape skin polyphenols. Food Chem. Toxicol. 2014, 74, 216–224. [Google Scholar] [CrossRef]
- Yu, C.C.; Nandrot, E.F.; Dun, Y.; Finnemann, S.C. Dietary antioxidants prevent age-related retinal pigment epithelium actin damage and blindness in mice lacking alphavbeta5 integrin. Free Radic. Biol. Med. 2012, 52, 660–670. [Google Scholar] [CrossRef]
- Kalt, W.; Blumberg, J.B.; McDonald, J.E.; Vinqvist-Tymchuk, M.R.; Fillmore, S.A.; Graf, B.A.; O'Leary, J.M.; Milbury, P.E. Identification of anthocyanins in the liver, eye, and brain of blueberry-fed pigs. J. Agric. Food Chem. 2008, 56, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Nakamura, Y.; Iida, H.; Ito, K.; Ohguro, H. Comparative assessment of distribution of blackcurrant anthocyanins in rabbit and rat ocular tissues. Exp. Eye Res. 2006, 83, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, H.; Du, X. The therapeutic use of quercetin in ophthalmology: recent applications. Biomed. Pharmacother. 2021, 137, 111371. [Google Scholar] [CrossRef]
- Koyama, Y.; Kaidzu, S.; Kim, Y.C.; Matsuoka, Y.; Ishihara, T.; Ohira, A.; Tanito, M. Suppression of light-induced retinal degeneration by quercetin via the AP-1 pathway in rats. Antioxidants (Basel) 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ueda, K.; Zhao, J.; Sparrow, J.R. Correlations between photodegradation of bisretinoid constituents of retina and dicarbonyl adduct deposition. J. Biol. Chem. 2015, 290, 27215–27227. [Google Scholar] [CrossRef]
- Cao, X.; Liu, M.; Tuo, J.; Shen, D.; Chan, C.C. The effects of quercetin in cultured human RPE cells under oxidative stress and in Ccl2/Cx3cr1 double deficient mice. Exp. Eye Res. 2010, 91, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Maher, P.; Hanneken, A. Flavonoids protect retinal ganglion cells from oxidative stress–induced death. Invest. Ophthalmol. Vis. Sci. 2005, 46, 4796–4803. [Google Scholar] [CrossRef]
- Kim, J.; Jin, H.L.; Jang, D.S.; Jeong, K.W.; Choung, S.Y. Quercetin-3-O-α-l-arabinopyranoside protects against retinal cell death via blue light-induced damage in human RPE cells and Balb-c mice. Food Funct. 2018, 9, 2171–2183. [Google Scholar] [CrossRef] [PubMed]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin bioactive effects: moving from preclinical evidence to potential clinical applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar]
- Ortega, J.T.; Parmar, T.; Jastrzebska, B. Flavonoids enhance rod opsin stability, folding, and self-association by directly binding to ligand-free opsin and modulating its conformation. J. Biol. Chem. 2019, 294, 8101–8122. [Google Scholar] [CrossRef]
- Ortega, J.T.; Parmar, T.; Golczak, M.; Jastrzebska, B. Protective Effects of flavonoids in acute models of light-induced retinal degeneration. Mol. Pharmacol. 2021, 99, 60–77. [Google Scholar] [CrossRef]
- Yang, Q.; Li, Y.; Luo, L. Effect of myricetin on primary open-angle glaucoma. Transl. Neurosci. 2018, 9, 132–141. [Google Scholar] [CrossRef]
- Shin, J.C.; Jung, H.Y.; Harikishore, A.; Kwon, O.D.; Yoon, H.S.; Kim, K.T.; Choi, B.H. The flavonoid myricetin reduces nocturnal melatonin levels in the blood through the inhibition of serotonin N-acetyltransferase. Biochem. Biophys. Res. Commun. 2013, 440, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Laabich, A.; Manmoto, C.C.; Kuksa, V.; Leung, D.W.; Vissvesvaran, G.P.; Karliga, I.; Kamat, M.; Scott, I.L.; Fawzi, A.; Kubota, R. Protective effects of myricetin and related flavonols against A2E and light mediated-cell death in bovine retinal primary cell culture. Exp. Eye Res. 2007, 85, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: structure-activity relationships. Free Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Caporali, S.; De Stefano, A.; Calabrese, C.; Giovannelli, A.; Pieri, M.; Savini, I.; Tesauro, M.; Bernardini, S.; Minieri, M.; Terrinoni, A. Anti-inflammatory and active biological properties of the plant-derived bioactive compounds luteolin and luteolin 7-glucoside. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.H.; Dong, X.W.; Yu, H.L.; Shen, W.; Lv, X.Y.; Wang, R.; Cheng, X.X.; Xiong, F.; Hu, X.L.; Wang, H. Cynaroside protects the blue light-induced retinal degeneration through alleviating apoptosis and inducing autophagy in vitro and in vivo. Phytomedicine 2021, 88, 153604. [Google Scholar] [CrossRef]
- Yu, H.; Li, J.; Hu, X.; Feng, J.; Wang, H.; Xiong, F. Protective effects of cynaroside on oxidative stress in retinal pigment epithelial cells. J. Biochem. Mol. Toxicol. 2019, 33, e22352. [Google Scholar] [CrossRef]
- Rodriguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martin, M.A. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur J Nutr 2012, 51, 881–892. [Google Scholar] [CrossRef]
- Yamakoshi, J.; Saito, M.; Kataoka, S.; Tokutake, S. Procyanidin-rich extract from grape seeds prevents cataract formation in hereditary cataractous (ICR/f) rats. J. Agric. Food Chem. 2002, 50, 4983–4988. [Google Scholar] [CrossRef]
- Reddy, G.B.; Muthenna, P.; Akileshwari, C.; Raghu, G.; Suryanarayana, P. Antiglycating potential of procyanidin-B2 isolated from cinnamon bark: Prevention or treatment of diabetic ocular complications (cataract & retinopathy). Invest. Ophthalmol. Vis. Sci. 2013, 54, 1945–1945. [Google Scholar]
- Li, W.; Jiang, Y.; Sun, T.; Yao, X.; Sun, X. Supplementation of procyanidins B2 attenuates photooxidation-induced apoptosis in ARPE-19 cells. Int. J. Food Sci. Nutr. 2016, 67, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.K.; Davis, A.; Rodriguez, M.E.; Agron, S.; Hackam, A.S. Protective effects of a grape-supplemented diet in a mouse model of retinal degeneration. Nutrition 2016, 32, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, Z.; Wang, K.; Zhu, X.; Ali, Y.; Shu, W.; Bao, X.; Zhu, L.; Fan, X.; Murray, M.; Zhou, F. Procyanidin B2 and rutin in Ginkgo biloba extracts protect human retinal pigment epithelial (RPE) cells from oxidative stress by modulating Nrf2 and Erk1/2 signalling. Exp. Eye Res. 2021, 207, 108586. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, L.; Huo, Y.; Zhou, F.; Wu, W.; Lu, F.; Yang, X.; Guo, X.; Chen, P.; Deng, Q. Protective effect of proanthocyanidins from sea buckthorn (Hippophae rhamnoides L.) seed against visible light-induced retinal degeneration in vivo. Nutrients 2016, 8, 245. [Google Scholar]
- Stoupi, S.; Williamson, G.; Viton, F.; Barron, D.; King, L.J.; Brown, J.E.; Clifford, M.N. In vivo bioavailability, absorption, excretion, and pharmacokinetics of [14C] procyanidin B2 in male rats. Drug Metab. Dispos. 2010, 38, 287–291. [Google Scholar] [CrossRef]
- Spencer, J.P.; Schroeter, H.; Shenoy, B.; Srai, S.K.; Debnam, E.S.; Rice-Evans, C. Epicatechin is the primary bioavailable form of the procyanidin dimers B2 and B5 after transfer across the small intestine. Biochem. Biophys. Res. Commun. 2001, 285, 588–593. [Google Scholar] [CrossRef]
- Xiao, Y.; Hu, Z.; Yin, Z.; Zhou, Y.; Liu, T.; Zhou, X.; Chang, D. Profiling and distribution of metabolites of procyanidin B2 in mice by UPLC-DAD-ESI-IT-TOF-MSn technique. Front. Pharmacol. 2017, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Zhang, R.; Lee, N.H.; Hyun, J.W. Phloroglucinol attenuates ultraviolet B radiation-induced matrix metalloproteinase-1 production in human keratinocytes via inhibitory actions against mitogen-activated protein kinases and activator protein-1. Photochem. Photobiol. 2012, 88, 381–388. [Google Scholar] [CrossRef]
- Cia, D.; Cubizolle, A.; Crauste, C.; Jacquemot, N.; Guillou, L.; Vigor, C.; Angebault, C.; Hamel, C.P.; Vercauteren, J.; Brabet, P. Phloroglucinol protects retinal pigment epithelium and photoreceptor against all-trans-retinal-induced toxicity and inhibits A2E formation. J. Cell Mol. Med. 2016, 20, 1651–1663. [Google Scholar] [CrossRef] [PubMed]
- Taveau, N.; Cubizolle, A.; Guillou, L.; Pinquier, N.; Moine, E.; Cia, D.; Kalatzis, V.; Vercauteren, J.; Durand, T.; Crauste, C.; Brabet, P. Preclinical pharmacology of a lipophenol in a mouse model of light-induced retinopathy. Exp. Mol. Med. 2020, 52, 1090–1101. [Google Scholar] [CrossRef]
- Zhang, J.; Kiser, P.D.; Badiee, M.; Palczewska, G.; Dong, Z.; Golczak, M.; Tochtrop, G.P.; Palczewski, K. Molecular pharmacodynamics of emixustat in protection against retinal degeneration. J. Clin. Invest. 2015, 125, 2781–2794. [Google Scholar] [CrossRef] [PubMed]
- Kren, V.; Valentova, K. Silybin and its congeners: from traditional medicine to molecular effects. Nat. Prod. Rep. 2022, 39, 1264–1281. [Google Scholar] [CrossRef]
- Lin, C.H.; Li, C.H.; Liao, P.L.; Tse, L.S.; Huang, W.K.; Cheng, H.W.; Cheng, Y.W. Silibinin inhibits VEGF secretion and age-related macular degeneration in a hypoxia-dependent manner through the PI-3 kinase/Akt/mTOR pathway. Br. J. Pharmacol. 2013, 168, 920–931. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, H.; Wang, Z.; Guan, W.; Kang, X.; Tai, X.; Sun, Y. Silibinin declines blue light-induced apoptosis and inflammation through MEK/ERK/CREB of retinal ganglion cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4059–4065. [Google Scholar] [CrossRef] [PubMed]
- Tvrdy, V.; Pourova, J.; Jirkovsky, E.; Kren, V.; Valentova, K.; Mladenka, P. Systematic review of pharmacokinetics and potential pharmacokinetic interactions of flavonolignans from silymarin. Med. Res. Rev. 2021, 41, 2195–2246. [Google Scholar] [CrossRef] [PubMed]
- Theodosis-Nobelos, P.; Papagiouvannis, G.; Tziona, P.; Rekka, E.A. Lipoic acid. Kinetics and pluripotent biological properties and derivatives. Mol. Biol. Rep. 2021, 48, 6539–6550. [Google Scholar] [PubMed]
- Gilgun-Sherki, Y.; Melamed, E.; Offen, D. Oxidative stress induced-neurodegenerative diseases: the need for antioxidants that penetrate the blood brain barrier. Neuropharmacology 2001, 40, 959–975. [Google Scholar] [CrossRef] [PubMed]
- Demir, U.; Demir, T.; Ilhan, N. The protective effect of alpha-lipoic acid against oxidative damage in rabbit conjunctiva and cornea exposed to ultraviolet radiation. Ophthalmologica 2005, 219, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.Y.; Lin, D.P.; Chang, L.S.; Huang, T.P.; Liu, H.J.; Luk, C.P.; Lo, Y.L.; Chang, H.H. Dietary alpha-lipoic acid prevents UVB-induced corneal and conjunctival degeneration through multiple effects. Invest. Ophthalmol. Vis. Sci. 2013, 54, 6757–6766. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, B.A.; Roberts, R.; Stemmler, A.; Luan, H.; Gradianu, M. Impaired apparent ion demand in experimental diabetic retinopathy: correction by lipoic acid. Invest. Ophthalmol. Vis. Sci. 2007, 48, 4753–4758. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Luan, H.; Berkowitz, B.A. alpha-lipoic acid corrects late-phase supernormal retinal oxygenation response in experimental diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2006, 47, 4077–4082. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, C.; Song, D.; Li, Y.; Song, Y.; Su, G.; Dunaief, J.L. Systemic administration of the antioxidant/iron chelator alpha-lipoic acid protects against light-induced photoreceptor degeneration in the mouse retina. Invest. Ophthalmol. Vis. Sci. 2014, 55, 5979–5988. [Google Scholar] [CrossRef]
- Psotová, J.; Kolár, M.; Sousek, J.; Svagera, Z.; Vicar, J.; Ulrichová, J. Biological activities of Prunella vulgaris extract. Phytother. Res. 2003, 17, 1082–1087. [Google Scholar] [CrossRef]
- Kim, J.; Cho, K.; Choung, S.Y. Protective effect of Prunella vulgaris var. L extract against blue light induced damages in ARPE-19 cells and mouse retina. Free. Radic. Biol. Med. 2020, 152, 622–631. [Google Scholar] [PubMed]
- Barker, F.M., 2nd; Snodderly, D.M.; Johnson, E.J.; Schalch, W.; Koepcke, W.; Gerss, J.; Neuringer, M. Nutritional manipulation of primate retinas, V: effects of lutein, zeaxanthin, and n-3 fatty acids on retinal sensitivity to blue-light-induced damage. Invest. Ophthalmol. Vis. Sci. 2011, 52, 3934–3942. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Gao, S.; Zhou, J.; Qin, J.; Taylor, A.; Johnson, E.J.; Tang, G.; Sparrow, J.R.; Gierhart, D.; Shang, F. Lutein and zeaxanthin supplementation reduces photooxidative damage and modulates the expression of inflammation-related genes in retinal pigment epithelial cells. Free. Radic. Biol. Med. 2012, 53, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Yang, C.M.; Yang, C.H. Protective Effect of astaxanthin on blue light light-emitting diode-induced retinal cell damage via free radical scavenging and activation of PI3K/Akt/Nrf2 pathway in 661W cell model. Mar. Drugs 2020, 18. [Google Scholar] [CrossRef] [PubMed]
- Laabich, A.; Vissvesvaran, G.P.; Lieu, K.L.; Murata, K.; McGinn, T.E.; Manmoto, C.C.; Sinclair, J.R.; Karliga, I.; Leung, D.W.; Fawzi, A.; Kubota, R. Protective effect of crocin against blue light- and white light-mediated photoreceptor cell death in bovine and primate retinal primary cell culture. Invest. Ophthalmol. Vis. Sci. 2006, 47, 3156–3163. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, M.; Zhang, X.; Chen, Q.; Chen, H.; Sun, L.; Liu, G. Protective Effect of fucoxanthin isolated from Laminaria japonica against visible light-induced retinal damage both in vitro and in vivo. J. Agric. Food Chem. 2016, 64, 416–424. [Google Scholar] [CrossRef]
- Fontaine, V.; Monteiro, E.; Brazhnikova, E.; Lesage, L.; Balducci, C.; Guibout, L.; Feraille, L.; Elena, P.P.; Sahel, J.A.; Veillet, S.; Lafont, R. Norbixin protects retinal pigmented epithelium cells and photoreceptors against A2E-mediated phototoxicity in vitro and in vivo. PLoS One 2016, 11, e0167793. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



