
Submitted:
21 December 2024
Posted:
23 December 2024
You are already at the latest version
Abstract
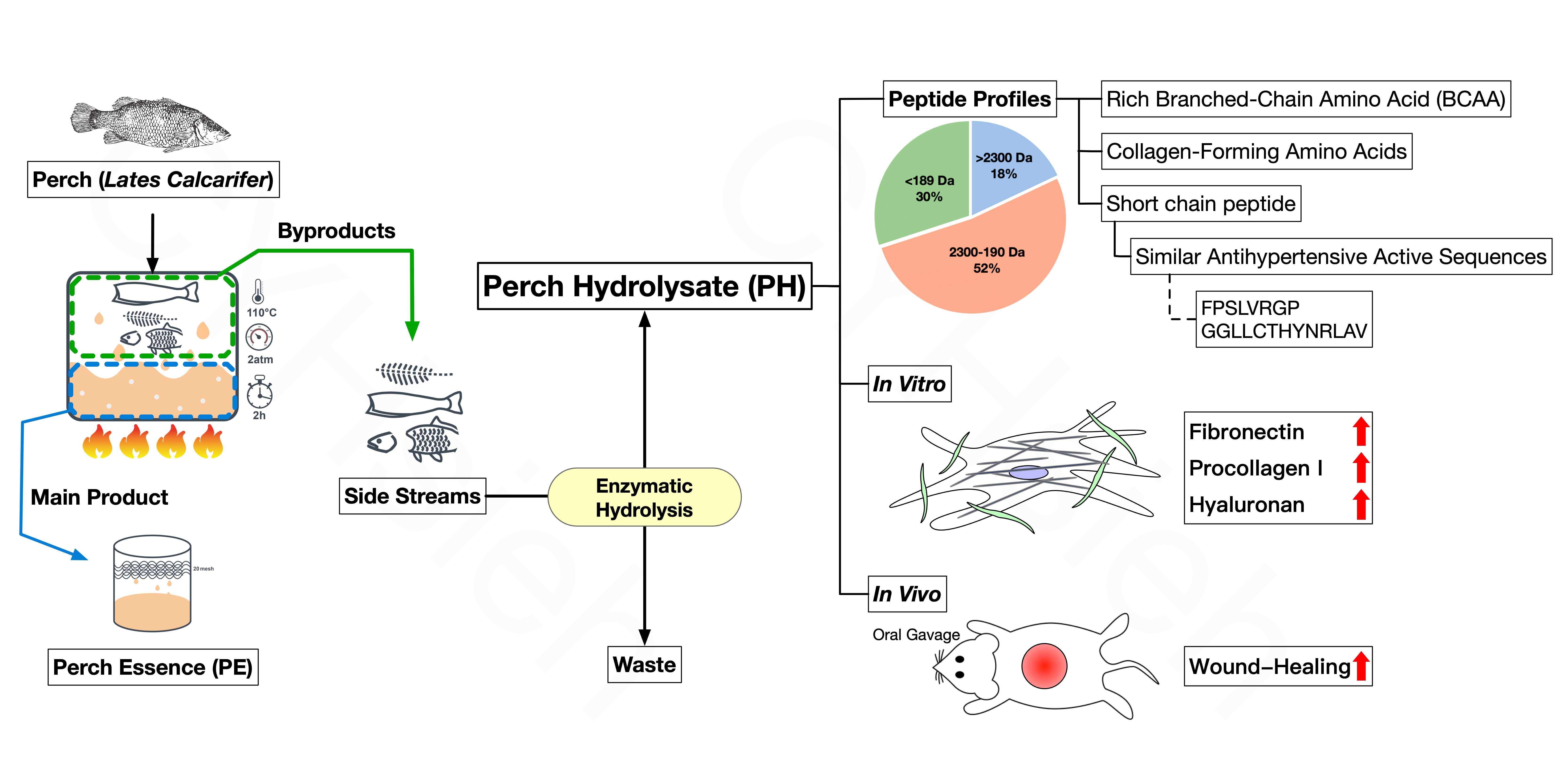
Wound healing incurs various challenges, making it a paramount topic in medicine. Short chain peptides from fish protein hydrolysates pose wound healing properties that may bring a solution. In this study, perch hydrolysates were produced from perch side steams using a designed commercial complex enzyme via a proprietary pressure extraction technique. The average molecular weight of the perch peptides was 1,289 kDa, and 62.60% of the peptides had a low molecular weight (≤ 1 kDa). The beneficial amino acid sequence FPSLVRGP with antihypertensive effects accounted for 6.21% abundance. The concentrations of collagen composition and branched chain amino acids was 1183 and 1122 mg/100g, respectively. In fibroblast model, active perch peptides accelerated wound healing mainly by increasing the secretion of procollagen I, fibronectin, and hyaluronan. In SD rat model to mimic human wound, orally perch hydrolysates with a molecular weight below 2.3 kDa accelerating wound healing mainly resulted from collagen-forming amino acids, branched-chain amino acids, and matrikine-like peptides. Collectively, the residue of the perch extract can be upcycled via hydrolysis technique to produce not only bioactive sequences but also short chain peptides. Considering the therapeutic potential to promote wound healing, such by-products are of great value that may develop as dietary nutraceuticals.
Keywords:
collagen
; hydrolysate
; perch
; short chain peptides
; upcycle
; wound healing
1. Introduction
Fish soup is a conventional oriental tonic, and fish is an important source of protein for humans. Furthermore, fish soup is ideal for individuals unable to consume fish easily, such as the sick or post-surgery patients. The decomposition and spoilage of fish protein after capture, leading to quick decay, can be attributed to endogenous enzymes and external microbial activity. Extraction technology is extensively used in Taiwan to produce fish essence by extracting the protein from the fish body and sterilizing it. However, after the extraction, a significant amount of side-stream and waste is generated, and the fish body cannot be entirely recycled [1]. The aquatic industry considers it essential to improve product value from aquatic side-streams and minimize waste [2,3]. Enzymatic hydrolysis is a highly efficient technology to retrieve 80% protein from the heads of cod (Gadus morhua). The extracted protein can be used as an ingredient in dietary supplements intended for human consumption [4].
Peptides typically remain inactive within the parent protein sequence. However, bioactive peptides can be generated through proteolysis using methods like enzymatic hydrolysis or fermentation [5,6]. Numerous food-derived peptides have demonstrated diverse beneficial impacts on human health based on their structure, encompassing antioxidant, anticancer, antihypertensive, antimicrobial, and immunomodulatory effects [7,8,9,10,11]. Studies illustrate that peptides obtained from different types of fish proteins exhibit a range of biological actions [12]. Two marine hydrolysates, extracted respectively from blue whiting (Micromesistius poutassou) and brown shrimp (Penaeus aztecus), elicited a high level of cholecystokinin secretion in endocrine cells [13]. Furthermore, the peptide originating from the fish protein hydrolysate of croaker (Otolithes ruber) revealed antioxidative properties, increased endurance capacity, and facilitated the recovery from oxidative stress in Wistar rats [14]. Consuming fish hydrolysates, consisting primarily of dipeptides and tripeptides, has demonstrated multiple advantages in maintaining skeletal muscle health and metabolism amidst the aging process [15,16]. Enzymatically hydrolyzed peptides extracted from the Pacific oyster (Crassostrea gigas), with molecular weights ranging from 550 Da to 2300 Da and peptide chain lengths between 11 and 16 amino acids, have been observed to exhibit anti-inflammatory properties whilst promoting fibroblast proliferation upon topical application [17].
Reducing waste is a worldwide issue for promoting the reduction of carbon emissions within ESG. Collagen peptide and flesh protein peptide extracted from Alaska Pollock showed a significant increase in the wound healing rate in a rat wound model [18]. The process of extracting perch essence, using high temperature and pressure, results in about 66% residue consisting of flesh, bones, head, scales, skin, and fins. Despite numerous investigations into wound healing activity associated with fish byproducts and their hydrolysates [18,19], few studies have analyzed the protein profile of perch hydrolysates as a byproduct of whole perch extraction and their wound healing activity as a dietary supplement for waste reduction. Thus, the purpose of this study is to evaluate the composition characteristics of perch hydrolysate and the effectiveness of its impact on wound healing.
2. Materials and Methods
2.1. Perch Hydrolysate Preparation
The perch (Lates calcarifer) used in this study was supplied by Yilan Anyong Lohas Co., Ltd. (Yilan, Taiwan). The fish were subjected to thorough cleaning procedures that involved removal of the viscera, followed by extracting of the flesh, bone, and scales with RO water (w/v=1:1) under high pressure. The resulting extract was filtered to obtain the commercial perch essence. Perch byproduct was mixed with deionized water in a ratio of 1:3 (w/w). Commercial enzymes, including Protamex, Neutrase, Pepsin, Papain, Trysin, Flavourzyme, and Protana Prime (Novozymes A/S Enzyme Inc., Bagsvaerd, Denmark), were added at a concentration of 0.6% (the enzyme to perch byproduct weight ratio was 0.6:100, w/w). This was succeeded by hydrolysis under an optimal temperature of enzymes (40~60°C) (YC-EC-600, Yenchen Machinery Co., Ltd., Taoyuan, Taiwan). The enzymatic reaction was halted at 100°C for ten minutes. Following this, the resulting hydrolysate was subjected to centrifugation and filtering to obtain perch hydrolysate (PH). The PH underwent filtration using a tangential flow filtration system (Advanced Biotechnology Laboratories Co., Ltd., New Taipei City, Taiwan) with a 30 kDa ultrafiltration membrane (Microdyn-Nadir Ltd., Wiesbaden, Germany) to yield PH30 with MW< 30 kDa. The resulting filtrate was then freeze-dried to produce a powder for experimental analysis.
2.2. Determination of Free and Hydrolyzed Amino Acid Compositions
Free and hydrolyzed amino acid compositions were analyzed according to ISO 13903:2005 using an amino acid analyzer (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The perch hydrolysate is extracted with sulphosalicylic acid to precipitate any protein. The resultant sample was brought to volume with a loading buffer and then transferred into a vial. The free amino acid concentration in mg/100g is calculated by separating amino acids in an amino acid analyzer, followed by detection through post column derivatization with ninhydrin reagent utilizing 440 and 570 nm. Finally, compare the absorbance intensity of the sample and standard to calculate the free amino acid concentration, and the unit is mg/100g.
The composition of hydrolyzed amino acids is analyzed in a similar way to that of free amino acids, except that the process of hydrolysis is involved. Drying perch hydrolysate are oxidized with hydrogen peroxide and formic acid at cold temperature, followed by acid hydrolysis using aqueous hydrochloric acid. The amino acids are separated by ion exchange chromatography and determined by reaction with ninhydrin, using photometric detection at 570 nm (440 nm for proline) with amino acid analyzer (Biochrom Ltd., Cambridge, UK).
2.3. Analysis of Soluble Protein Content
Soluble protein content was determined using a modified Folin-Lowry method [20]. First, 0.5 mL of diluted sample solution was combined with 0.5 mL reagent A and 4 mL reagent B (Bio-Rad Dc Protein Assay Kit, Bio-Rad Laboratories, Inc., Berkeley, CA, USA). The solution was then shaken and allowed to stand at room temperature for 15 minutes. Finally, the absorbance was assessed at 540 nm using the SP-UV1100 spectrophotometer from DLAB Scientific Inc. in Beijing, China. Bovine serum albumin was utilized as an analytical standard, and the result was transformed into soluble protein content, measured in mg/mL.
2.4. Determination of Peptide Content by α-Amino Group Content
Peptide levels were assessed with minor adjustments to the approach employed by [21]. The reaction comprised adding 2 mL of o-phthaldialdehyde (OPA) reagent to 50 µL of the sample solution, which had been diluted. After mixing, the sample solution was allowed to settle for 2 minutes at room temperature, followed by measuring the absorbance at 340 nm using a spectrophotometer (SP-UV1100, DLAB Scientific Inc., Beijing, China). The α-amino group levels were evaluated using Leu-Gly as a standard (unit: mg/mL).
2.5. Determination of Degree of Hydrolysis
The degree of hydrolysis of PH was determined by the OPA method [22]. To fully hydrolyze the perch hydrolysate, 6 N HCl was used in a dry bath at 110°C for 24 hours. The total content of α-amino groups in the perch hydrolysate, fully hydrolyzed perch by-product and raw material background (the perch by-product was added to deionized water, homogenized for 5 minutes, then heated in boiling water for 10 minutes) was determined via the OPA method. A standard calibration curve was prepared with L-leucine standard substance and the α-amino group content was converted to mg/mL. Furthermore, utilize the subsequent formula to calculate the degree of hydrolysis.
Formula:DH (%) = (α-ni) / (nT-ni)
α: α-amino groups content in the perch hydrolysate.
nT: α-amino groups content in the fully hydrolyzed perch byproduct.
ni: α-amino groups content in the perch byproduct.
2.6. Analysis of Protein Profile
The sample solution underwent filtration through a 0.22-µm filter membrane, and 100 µL of the filtrate served as the injection volume. Subsequently, gel filtration chromatography (TSKgel GMPWxL Column, Tosoh Bioscience Inc., South San Francisco, CA, USA) was performed for purification and separation, equilibrated with 0.05 M NaNO3. Data analysis was conducted with the Viscotek GPC System (1122 pump, 717 Auto Injector, 270 Dual Detector/Differential Viscometer & Laser Light Scattering Detector, 3580 RI detector, and OmniSEC 4.6 Station, Malvern Panalytical Ltd., Malvern, UK).
2.7. Identification of Peptide Sequence by LC-MS/MS Analysis
The peptides were diluted in HPLC buffer A (0.1% formic acid) and loaded onto a reverse-phase column (Zorbax 300SB-C18, 0.3 × 5 mm; Agilent Technologies). The desalted peptides were then separated on a homemade column (Waters BEH 1.7 μm, 100 μm I.D. × 10 cm with a 15 μm tip) using a multi-step gradient of HPLC buffer B (99.9% acetonitrile/0.1% formic acid) for 70 minutes with a flow rate of 0.3 μ l/min. The LC apparatus was coupled with a 2D linear ion trap mass spectrometer (Orbitrap Elite ETD; Thermo Fisher) operated using Xcalibur 2.2 software (Thermo Fisher). The full-scan MS was performed in the Orbitrap over a range of 400 to 2,000 Da and a resolution of 120,000 at m/z 400. Internal calibration was performed using the ion signal protonated dodecamethylcyclohexasiloxane ion at m/z 536.165365 as lock mass. The 12 data-dependent MS/MS scan events were followed by one MS scan for the 12 most abundant precursor ions in the preview MS scan. The m/z values selected for MS/MS were dynamically excluded for 60 seconds with a relative mass window of 15 ppm. The electrospray voltage was set to 2.0 kV, and the temperature of the capillary was set to 200°C. MS and MS/MS automatic gain control was set to 1,000 ms (full scan) and 300 ms (MS/MS), or 3 × 106 ions (full scan) and 30,000 ions (MS/MS) for maximum accumulated time or ions, respectively. De Novo Peptide sequencing was carried out using PEAKS studio (version 10.5, Bioinformatics Solutions Inc.). The MS/MS spectra were analyzed using de novo function. For peptide identification, 10 ppm mass tolerance was permitted for intact peptide masses and 0.1 Da for HCD fragment ions; oxidized methionine, acetyl (protein N-terminal), and deamination (asparagine) as variable modifications. Peptide-spectrum match (PSM) was then filtered based on 50% of average local confidence (de novo score).
2.8. Wound Healing Assay in Animal Model
Six-week-old male Sprague-Dawley (SD) rats (264.6 ± 7.0 g) were acquired from Bio LASCO Taiwan Co., Ltd. (Taipei, Taiwan) to assess the wound-healing potential of perch hydrolysate. These rats were kept in standard cages with relative humidity maintained at 30-70%, temperature controlled to 22 ± 4 °C, and subjected to a 12-hour light-dark cycle. They had free access to a commercial pellet diet and water ad libitum. The protocol was approved by the Institutional Animal Care and Use Committee of the Agricultural Technology Research Institute under codes 109093 and 110005. Preliminary groups of rats were studied to confirm the oral dosage of the treatment groups using varying doses of PH. A group of five SD rats were orally administered PH30 at a dose of 1972 mg/kg body weight for 21 days, with a daily single dose by gavage for 7 days. The control group was administered RO water.
The rats were given an intraperitoneal injection of 50 mg/kg bw Zoletil 50 for anesthesia. The fur surrounding each wound was shaved. A circular mark with a diameter of 2 cm was drawn on the back of each rat and a full-thickness wound was made with a sterilized instrument to the depth of the dermis. The wounds were then covered with 3M Tegaderm dressing. The dressings were taken off on days 1, 2, 3, 5, 7, and 21 post-surgery, and the wounds were observed and photographed using Image J (National Institutes of Health, Maryland, U.S.A) to measure the wound size. The percentage of wound contraction at varying days after the wound rate (%) was computed by applying the following formula:
Wound Healing Rate (%)
x is the corresponding time point.
2.9. Analytical Assays for Fibronectin, Procollagen I and Hyaluronan in Human Fibroblasts
Human dermal fibroblast cells (890510-01F ATIT) were seeded at 5 × 105 cells/well in a 6-well microplate. Once the cells had adhered, the medium was replaced with DMEM medium devoid of fetal bovine serum. Simultaneously, the sample was added, and the cells were incubated for 2 days at 37 °C in a humidified atmosphere consisting of 95% air and 5% CO2. The supernatant was subsequently collected and used to detect fibronectin, procollagen I and hyaluronan levels quantitatively by enzyme-linked immunosorbent assay (R&D systems, U.S). Measure the absorbance value at a wavelength of 450 nm with a microplate reader, plot it against the standard curve of known concentrations and their respective absorbance, and calculate the concentration of the sample. The negative control group (NC) was cultured in DMEM medium, the positive control group (PC) was treated with 5% fetal bovine serum, and all samples were analyzed in triplicate.
2.10. Statistical Analysis of Data
The statistical analysis in this study was performed using SAS software (Statistical Analysis System, SAS; Version 9.4 TSIM5, SAS Institute Inc., Cary, NC, USA). The mean ± standard error (mean ± SEM) was used to present the values of experimental results. To identify differences between groups, the experimental data was analyzed using one-way analysis of variance (one-way ANOVA) and compared using Duncan’s multiple range test.
3. Results
3.1. Upcycled Perch Hydrolysates from Perch Side Steams
After high-pressure extraction of gutted perch, the resulting side streams of perch essence possess a sludge-like consistency and can undergo hydrolysis through the addition of enzymes without the need for any further grinding or homogenization. Table 1 presents the α-amino group amount and its degree at different times using the designed complex enzyme. The side streams of perch essence enhance the amino-group amount with time of hydrolysis, thereby increasing the degree of hydrolysis. Table 1 demonstrates a higher rate of hydrolysis for up to five hours, subsequently slowing down. For subsequent analysis, the hydrolysate gained after five hours of partial hydrolysis was selected due to its operating efficiency and anticipated bioactive peptide availability. The PH of perch side streams can be achieved with a hydrolysis time of five hours, resulting in a high degree hydrolysis of 43.1%.
3.2. Protein Profiles and Amino Acid Composition in PH
Table 2 shows the molecular weight distribution of perch essence and PH using gel filtration chromatography. While perch essence was predominantly protein, with approximately 62.9% of the proteins having molecular weights greater than 2.3 kDa, the PH was predominantly peptides and amino acids, accounting for 81.6% of the total proteins. Peptides with molecular weights (MW) ranging from 190 to 2300 Dalton account for 52% of the PH, while peptides or amino acids with molecular weights below 190 Da account for 29.6%.
The composition of free amino acids in PH is shown in Table 3. In addition to the highly abundant amount of leucine and valine in PH, lysine and arginine are the primary amino acids that are cleaved by the enzymes, as they are positively charged amino acid residues and typically found on the surface of proteins [23].
The hydrolyzed amino acid composition of protein in PH consists primarily of glutamic acid, aspartic acid, glycine, lysine, and alanine (Table 4). Fish proteins generally have elevated levels of essential amino acids such as lysine and leucine, as well as non-essential amino acids including aspartic acid, glutamic acid, and alanine [24], which is consistent with the findings of this study (Table 4). The hydrolyzed amino acids found in the perch essence have a composition nearly identical to the abundance order of the perch hydrolysate content in this study. This suggests that enzyme hydrolysis could be a valuable method to recover essential amino acids that would otherwise be discarded as waste.
3.3. Effect of PH on Wound Healing Rate in the Animal Model
Many fish peptides have been shown to promote wound healing [25,26]. However, most studies have been conducted in vitro or on the effects of topical application. To determine whether orally the recovered peptide had any effect on wound repair, three different doses of PH were administered to SD rats. Although the oral administration of PH seemed to have a wound-healing activity after three days, there was only a statistically significant difference at 7 days (data was not shown). Small 7-29 amino-acid peptides obtained from the skin of silver carp contain significant amounts of peptides that may provide beneficial effects in patients requiring reconstructive treatment [27]. Therefore, there is an expectation that smaller proteins or peptides may have a more pronounced effect [28]. A further statistically significant increase in the rate of wound healing was observed on the second day of feeding with PH30 obtained by tangential flow filtration (Figure 1).
3.4. Effects of PH on Fibroblasts Through Secreting Fibronectin, Procollagen I and Hyaluronan
Spent yeast waste streams containing bioactive peptides from fermentation processes have been shown to enhance the production of metabolites, such as fibronectin, procollagen I and hyaluronic acid [29]. Although there is no statistical difference, it appears that PH enhances fibroblast viability as shown in Figure 2a. In addition, it can be concluded that PH is not cytotoxic to fibroblasts. As expected, the PH effect on fibroblasts with an incubation time for 48 hours significantly increased the secretion of fibronectin (Figure 2b), procollagen I (Figure 2c) and hyaluronan (Figure 2d), which are major components of the skin's extracellular matrix (ECM).
3.5. Relative Abundance of Peptide Sequence in Perch Hydrolysates
Table 5 shows the amino acid sequence of the more abundant peptides in PH, almost all of which have MW below 1200 kDa. Among them, there are relatively high abundances of FPSLVRGP and GGLLCTHYNRLAV. These two more abundant peptides in PH are characterized by more residues of electron-rich groups (e.g. aromatic groups, imidazole and pyrrole rings), more hydrophobic groups, hydrophobic amino acid residues at the N end, and groups which are easy to form hydrogen bond at the C end.
4. Discussion
Wound healing incurs various challenges, making it a paramount topic in medicine. Short chain peptides from fish protein hydrolysates pose wound healing properties that may bring a solution. Previous research has shown that perch essence has measurable amounts of soluble protein and α-amino groups at 66.80±0.56 mg/ml and 10.63±0.09 mg/ml, respectively [30]. The protein analysis results indicate that residual side streams of perch essence are hydrolyzed to PH, which still contain many proteins, peptides, and amino acids. The potential benefits of the fish by-products are derived from the transformation of the biomass by means of exogenous enzyme hydrolysis, which avoids the potential risks of biological and chemical contamination [31]. From the perspective of upcycling, the resulting PH recovered about 9.22% of solids from the side streams of perch essence following enzymatic hydrolysis. Of the solids recovered, 715.7 ± 19.4 mg/g were soluble proteins. Subsequently, the PH was concentrated by ultrafiltration through a 30 kDa membrane, yielding PH30 with 887.2 ± 6.3 mg/g of soluble protein.
A wide range of bioactive properties such as antioxidant, antihypertensive, anticoagulant and immunomodulatory properties have been demonstrated in peptides obtained by enzymatic hydrolysis [32,33,34,35]. Numerous studies indicate that bioactive fish peptides tend to have low molecular weights and are composed of oligo-peptides containing between 2 to 10 amino acid residues with a molecular weight of less than 1.2 kDa [15,36,37]. With an average molecular weight of 110 Da per amino acid, the PH comprises 36.8% oligopeptides that contain less than 10 amino acid residues whose molecular weights range from 190 Da to 1.2 kDa, as presented in Table 2.
In a systematic review and meta-analysis, supplementation with either arginine or glutamine was shown to benefit wound healing [38]. High levels of glutamic acid and arginine in the PH (Table 3, 4) can contribute to the same effect. Furthermore, collagen peptides from chum salmon skin could accelerate the process of wound-healing in rats [39]. Low molecular weight (<3KDa) collagen peptides with in vitro wound-healing activities were obtained from fish skin of unicorn leatherjacket (Aluterus monoceros) [40]. The majority of peptides and amino acids in PH was shown in Table 2, the high wound repair ability of oral administration might be related to the presence of these peptides with a molecular weight of less than 2.3 kDa except amino acids.
Short sequences with potential biological activities were identified in 19 peptide sequences in prior research, including the antihypertensive sequence of the FP [41]. As shown in Table 5, our peptide sequences beginning with FP suggest that PH may also exert similar effects. Rémond et al. found that the FPSIVGRP fragment sequence had antihypertensive effects [42]. This sequence is very similar to the FPSLVRGP found in our study. These two peptides are almost identical except for the difference between leucine and isoleucine and the difference in order between glycine and arginine.
In Figure 2, the higher concentration of PH added induced more secretion in a dose-dependent manner. Signal peptides from many sources have been shown to trigger signaling pathways that stimulate fibroblasts to produce collagen, fibronectin and laminin to improve wound healing [43]. Our signal peptides from the side stream of the perch essence are also effective in promoting the secretion of these substrates that are associated with wound repair or skin conditions.
Inflammation, cell proliferation, and remodeling are recognized as the crucial stages of wound healing. The intricate process of tissue repair is driven by the proliferation of parenchymal cells induced by specific growth factors, involving mediators and hemocytes, as well as the production of ECM [44]. The ECM performs a crucial role in regulating cellular activities, including cell growth, proliferation, and migration. It can trigger apoptosis through signalling molecules generated by the proteolysis of the ECM to form soluble peptides, which are known as matrikines [45,46]. Matrikines regulate inflammatory, fibrogenic or reparative actions by modifying the phenotype and function of fibroblasts during wound healing [47]. The primary element of the skin's ECM is hyaluronan, an unsulphated glycosaminoglycan that plays a role in inflammatory responses and tissue regeneration processes and is frequently applied as a wound dressing [48]. The use of topical or oral matrikine-like substances may improve the ability of wounds to heal. In our study, the PH containing substances are similar to matrikines. These branched-chain amino acids (leucine, isoleucine and valine) play a pivotal role in promoting protein synthesis. Also, glycine, proline and hydroxyproline in PH contribute to collagen formation. Most importantly, matrikine-like peptides in PH induce the production of ECM substances such as hyaluronan.
Peptides applied orally should avoid gastrointestinal digestion to preserve their bioactivities and structures for wound repair. Peptides with C-terminal proline or hydroxyproline residues can be absorbed through the intestine intact by paracellular and transcellular routes [49]. It has been reported that peptides containing proline are typically impervious to degradation by digestive enzymes [41,50,51]. The bioactive peptide FPSIVGRP can be transported into the bloodstream by releasing mucosal peptidases, and withstand plasma peptidases, resulting in an antihypertensive impact [42]. Therefore, the C-terminal proline-containing FPSLVRGP and other peptides containing proline in our PH may have the ability to pass through the intestines and display wound healing properties. PH could be applied for not only topical purposes but also dietary supplements to speed up the wound healing process by providing amino acids and non-digestible matrikine-like peptides.
5. Conclusions
Through a designed commercial complex enzyme via a proprietary pressure extraction technique, perch hydrolysates upcycled from perch side steams contain various beneficial peptides and amino acids. Short chain peptides are necessary for wound repair and matrikine-like peptides prompt fibroblasts to generate fibronectin, procollagen I and hyaluronan. Our upcycling technique is effective on recovering the proteins left over after perch extraction in the production of perch essence. In light of this, our perch hydrolysates could develop as dietary nutraceuticals or functional foods to promote wound healing, particularly in undernourished subjects, post-surgical patients or post-partum women.
Author Contributions
Conceptualization, C.-C. Lin and J.-F. Chang; methodology, J.-F. Chang; software, C.-Y. Hsieh; validation, M.-H. Lee, and Y.-H. Ting; formal analysis, C.-Y. Yang; investigation, L.-N. Chen and C.-Y. Yang; resources, L.-N. Chen; data curation, M.-H. Lee, and Y.-H. Ting; writing—original draft preparation, C.-Y. Hsieh and C.-C. Lin; writing—review and editing, C.-C. Lin; supervision, C.-C. Lin. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Anyong Biotechnology Inc. (Taipei, Taiwan) for supplying perch sample for the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ohshima, T. By-products and seafood production in japan. Journal of Aquatic Food Product Technology 1996, 5, 27–42. [Google Scholar] [CrossRef]
- Karayannakidis, P.D.; Zotos, A. Fish processing by-products as a potential source of gelatin: a review. Journal of Aquatic Food Product Technology 2016, 25, 65–92. [Google Scholar] [CrossRef]
- González-González, D.C.; Rodríguez-Félix, D.E.; García-Sifuentes, C.O.; Castillo-Ortega, M.M.; Encinas-Encinas, J.C.; Ortega, H.D.C.S.; Romero-García, J. Collagen scaffold derived from tilapia (Oreochromis niloticus) skin: obtention, structural and physico-chemical properties. Journal of Aquatic Food Product Technology 2022, 31, 374–387. [Google Scholar] [CrossRef]
- Remme, J.; Tveit, G.M.; Toldnesb, B.; Slizyte, R.; Carvajal, A.K. Production of protein hydrolysates from cod (Gadus morhua) heads: lab and pilot scale studies. Journal of Aquatic Food Product Technology 2022, 31, 114–127. [Google Scholar] [CrossRef]
- Kehinde, B.A.; Sharma, P. Recently isolated antidiabetic hydrolysates and peptides from multiple food sources: a review. Critical Reviews in Food Science and Nutrition 2020, 60, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. International Journal of Molecular Sciences 2022, 23, 23–1445. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.-P. S.; Hancock, R.E.W. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: from research to food applications. Current Opinion in Biotechnology 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: a review on correlation between structure of bioactive peptides and their functional properties. Journal of Food Science and Technology 2019, 56, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Sosalagere, C.; Kehinde, B.A.; Sharma, P. Isolation and functionalities of bioactive peptides from fruits and vegetables: A reviews. Food Chemistry, 2022; 366, 130494. [Google Scholar]
- Guo, Q.; Chen, P.; Chen, X. Bioactive peptides derived from fermented foods: Preparation and biological activities. Journal of Functional Foods 2023, 101, 105404. [Google Scholar] [CrossRef]
- Senevirathne, M.; Kim, S.-K. Development of bioactive peptides from fish proteins and their health promoting ability. Advances in Food and Nutrition Research 2012, 65, 235–248. [Google Scholar]
- Cudennec, B.; Ravallec-Plé, R.; Courois, E.; Fouchereau-Peron, M. Peptides from fish and crustacean by-products hydrolysates stimulate cholecystokinin release in STC-1 cells. Food Chemistry 2008, 111, 970–975. [Google Scholar] [CrossRef]
- Nazeer, R.A.; Sampath Kumar, N.S.; Jai Ganesh, R. In vitro and in vivo studies on the antioxidant activity of fish peptide isolated from the croaker (Otolithes ruber) muscle protein hydrolysate. Peptides 2012, 35, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Dale, H.F.; Madsen, L.; Lied, G.A. Fish–derived proteins and their potential to improve human health. Nutrition Reviews 2019, 77, 572–583. [Google Scholar] [CrossRef]
- Lees, M.J.; Carson, B.P. The potential role of fish-derived protein hydrolysates on metabolic health, skeletal muscle mass and function in ageing. Nutrients, 2020; 12, 2434. [Google Scholar]
- Yu, D.; Cui, S.; Chen, L.; Zheng, S.; Zhao, D.; Yin, X.; Yang, F.; Chen, J. Marine-Derived bioactive peptides self-assembled multifunctional materials: antioxidant and wound healing. Antioxidants 2023, 12, 12–1190. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, K.; Li, B.; Hou, H. Effects of oral administration of peptides with low molecular weight from Alaska Pollock (Theragra chalcogramma) on cutaneous wound healing. Journal of Functional Foods 2018, 48, 682–691. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Li, C.X.; Shapawi, R.; Mokhtar, R.A.M.; Noordin, W.N.M.; Huda, N. Extraction and characterization of bioactive fish by-product collagen as promising for potential wound healing agent in pharmaceutical applications: Current Trend and Future Perspective. International Journal of, 2022; 9437878. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. The Journal of Biological and Chemistry 1951, 193, 265–75. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins, Journal of Dairy Science 1983, 66, 1219-1227.
- Linarès, E.; Larré, C.; Popineau, Y. Freeze- or spray-dried gluten hydrolysates. 1. Biochemical and emulsifying properties as a function of drying process, Journal of Food Engineering 2001, 48, 127–135. [Google Scholar] [CrossRef]
- Shah, D.; Shaikh, A.R. Interaction of arginine, lysine, and guanidine with surface residues of lysozyme: implication to protein stability. Journal of Biomolecular Structure and Dynamics 2016, 34, 104–114. [Google Scholar] [CrossRef]
- Ryu, B.; Shin, K.H.; Kim, S.K. Muscle Protein Hydrolysates and Amino Acid Composition in Fish. Marine Drugs 2021, 19, 377. [Google Scholar] [CrossRef] [PubMed]
- Felician, F.F.; Yu, R.H.; Li, M.Z.; Li, C.J.; Chen, H.Q.; Jiang, Y.; Tang, T.; Qi, W.-Y.; Xu, H.M. The wound healing potential of collagen peptides derived from the jellyfish Rhopilema esculentum. Chinese Journal of Traumatology 2019, 22, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Verma, A.K.; Patel, R. Collagen extraction and recent biological activities of collagen peptides derived from sea-food waste: a review. Sustainable Chemistry and Pharmacy 2020, 18, 100315. [Google Scholar] [CrossRef]
- Wojtkowiak, D.; Frydrychowski, A.F.; Hadzik, J.; Dominiak, M. Identification of small peptides of acidic collagen extracts from silver carp skin and their therapeutic relevance. Advances in Clinical and Experimental Medicine 2016, 25, 227–35. [Google Scholar] [CrossRef]
- Hu, Z.; Yang, P.; Zhou, C.; Li, S.; Hong, P. Marine collagen peptides from the skin of Nile Tilapia (Oreochromis niloticus): Characterization and wound healing evaluation. Marine drugs 2017, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.M.; Oliveira, A.S.; Silva, S.; Ribeiro, A.B.; Pereira, C.F.; Ferreira, C.; Casanova, F.; Pereira, J.O.; Freixo, R.; Pintado, M.E.; Carvalho, A.P.; Ramos, O.L. Spent yeast waste streams as a sustainable source of bioactive peptides for skin applications. International Journal of Molecular Sciences 2023, 24(3), 24–3. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Chang, J.-H.; Hung, Y.-T.; Lin, C.-C. The protein profile of perch essence and its improvement of metabolic syndrome in vitro. MOJ Food Processing & Technology 2022, 10, 10–12. [Google Scholar]
- Siddiqui, S.A.; Schulte, H.; Pleissner, D.; Schönfelder, S.; Kvangarsnes, K.; Dauksas, E.; Rustad, T.; Cropotova, J.; Heinz, V.; Smetana, S. Transformation of seafood side-streams and residuals into valuable products. Foods 2023, 12, 12–422. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Lee, W.W.; Lee, J.H.; Sanjeewa, K.K.A.; Fernando, I.P.S.; Ko, S.-C.; Lee, S.-H.; Kim, Y.-T.; Jeon, Y.-J. Purification and identification of an antioxidative peptide from digestive enzyme hydrolysis of cutlassfish muscle. Journal of Aquatic Food Product Technology 2018, 27, 934–944. [Google Scholar] [CrossRef]
- Phadke, G.G.; Rathod, N.B.; Ozogul, F.; Elavarasan, K.; Karthikeyan, M.; Shin, K.-H.; Kim, S.-K. Exploiting of secondary raw materials from fish processing industry as a source of bioactive peptide-rich protein hydrolysates. Marine Drugs 2021, 19, 480. [Google Scholar] [CrossRef] [PubMed]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and bioactive properties of peptides derived from marine side streams. Marine Drugs 2021, 19, 19–71. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Wu, Y.; Wang, Z.-G.; Wang, K.; Zhang, Z.- K.; Wang, B.; Luo, H.- Y. Evaluation of the anti-inflammatory activity of enzymatic hydrolysis peptide sep-3 from Skipjack (Katsuwonus pelamis) based on NF-κB and MAPK pathways. Journal of Aquatic Food Product Technology 2022, 31, 1128–1141. [Google Scholar] [CrossRef]
- Peng, X.; Xu, J.; Tian, Y.; Liu, W.; Peng, B. Marine fish peptides (collagen peptides) compound intake promotes wound healing in rats after cesarean section. Food & Nutrition Research 2020, 64, 4247. [Google Scholar]
- Abachi, S.; Pilon, G.; Marette, A.; Bazinet, L.; Beaulieu, L. Beneficial effects of fish and fish peptides on main metabolic syndrome associated risk factors: Diabetes, obesity and lipemia. Critical Reviews in Food Science and Nutrition 2022, 63, 7896–7944. [Google Scholar] [CrossRef]
- Arribas-López, E.; Zand, N.; Ojo, O.; Snowden, M.J.; Kochhar, T. The effect of amino acids on wound healing: a systematic review and meta-analysis on arginine and glutamine. Nutrients 2021, 13, 13–2498. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, M.; Liang, R.; Zhao, M.; Zhang, Z.; Li, Y. Oral administration of marine collagen peptides prepared from chum salmon (Oncorhynchus keta) improves wound healing following cesarean section in rats. Food & Nutrition Research 2015, 59, 26411. [Google Scholar]
- Kumar, L.V.; Shakila, R.J.; Jeyasekaran, G. In vitro anti-cancer, anti-diabetic, anti-inflammation and wound healing properties of collagen peptides derived from unicorn leatherjacket (Aluterus Monoceros) at different hydrolysis. Turkish Journal of Fisheries and Aquatic Sciences 2019, 19, 551–560. [Google Scholar]
- Bauchart, C.; Morzel, M.; Chambon, C.; Mirand, P.; Reynès, C.; Buffière, C.; Rémond, D. Peptides reproducibly released by in vivo digestion of beef meat and trout flesh in pigs. British Journal of Nutrition 2007, 98, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Rémond, D.; Savary-Auzeloux, I.; Gatellier, P.; Santé-Lhoutellier, V. Nutritional properties of meat peptides and proteins: impact of processing. SCIENCES DES ALIMENTS 2008, 28, 389–401. [Google Scholar] [CrossRef]
- Skibska, A.; Renata, P. Signal peptides - Promising ingredients in cosmetics. Current Protein & Peptide Science 2021, 22, 22–716. [Google Scholar]
- Gonzalez, A.C.; Costa, T.G.; Andrade, Z.A.; Medrado, A.R.A.P. Wound healing –A literature review. Anais Brasileiros de Dermatologia 2016, 91, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Shanthi, C. Cryptic peptides from collagen: a critical review. Protein & Peptide Letters, 2016; 23, 664–672. [Google Scholar]
- Sivaraman, K.; Shanthi, C. Matrikines for therapeutic and biomedical applications. Life Science 2018, 214, 22–33. [Google Scholar] [CrossRef] [PubMed]
- de Castro Brás, L.E.; Frangogiannis, N.G. Extracellular matrix-derived peptides in tissue remodeling and fibrosis. Matrix Biology, 2020; 91–92, 176–187. [Google Scholar]
- Graça, M.F.P.; Miguel, S.P.; Cabral, C.S.D.; Correia, I.J. Hyaluronic acid-Based wound dressings: A review. Carbohydrate Polymers 2020, 1, 116364. [Google Scholar] [CrossRef] [PubMed]
- Vermeirssen, V.; Camp, J.; Verstraete, W. Bioavailability of angiotensin I converting enzyme inhibitory peptides. British Journal of Nutrition 2004, 92, 357–66. [Google Scholar] [CrossRef]
- Sosulski, F.W.; Imafidon, G.I. Amino acid composition and nitrogen-to-protein conversion factors for animal and plant foods. Journal of Agricultural and Food Chemistry 1990, 38, 38–1351. [Google Scholar] [CrossRef]
- Hausch, F.; Shan, L.; Santiago, N.A.; Gray, G.M.; Khosla, C. Intestinal digestive resistance of immunodominant gliadin peptides. American Journal Physiology-Gastrointestinal and Liver Physiology 2002, 283, G996–G1003. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Changes in wound healing rate after oral feeding of perch hydrolysate with molecular weight less than 30 kDa in SD rats. The experiment was carried out by feeding SD rats with PH30. Data were presented as mean±S.D., n=5. *p <0.05 (one-way ANOVA test).
Figure 1.
Changes in wound healing rate after oral feeding of perch hydrolysate with molecular weight less than 30 kDa in SD rats. The experiment was carried out by feeding SD rats with PH30. Data were presented as mean±S.D., n=5. *p <0.05 (one-way ANOVA test).
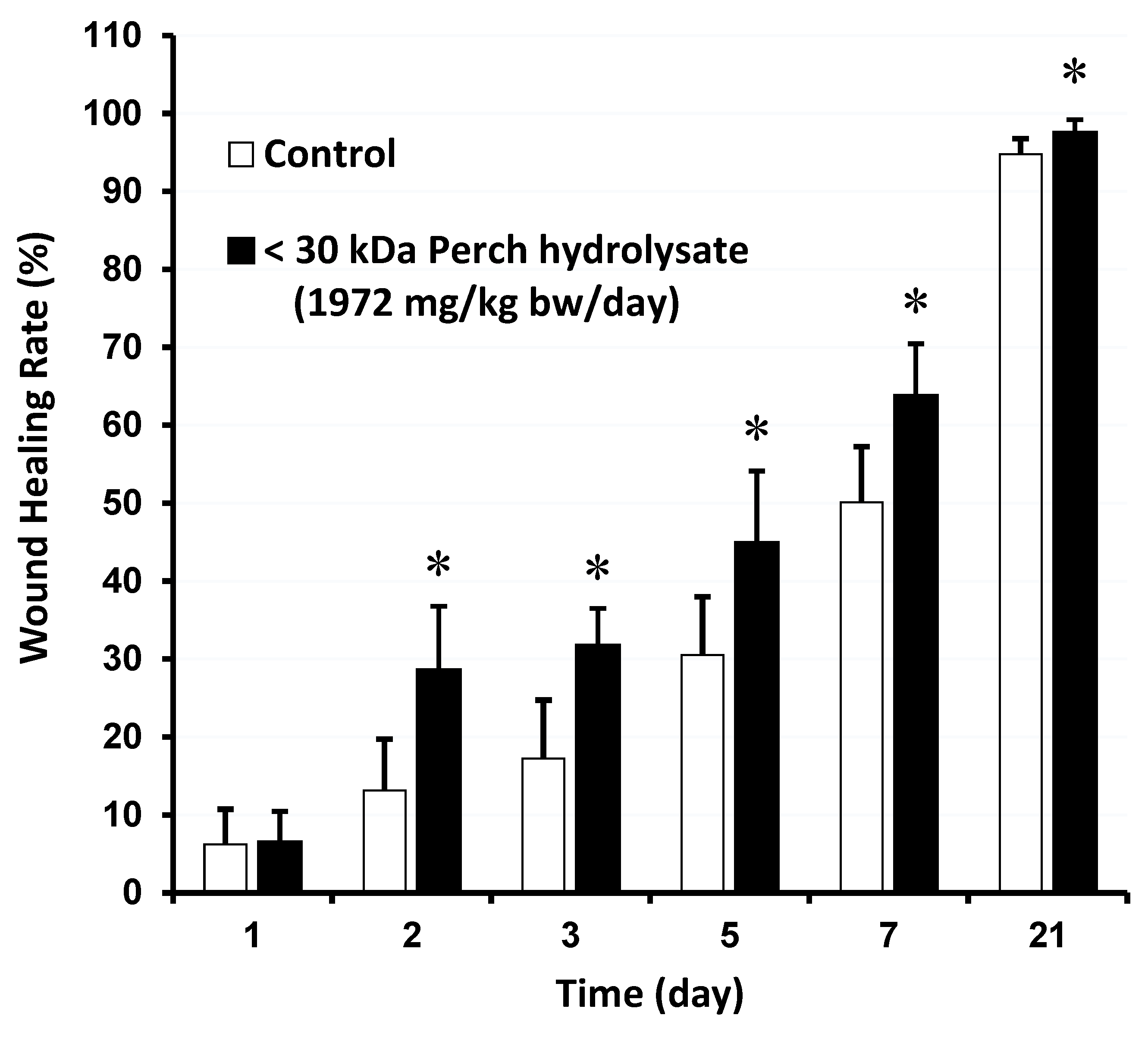
Figure 2.
Effect of perch hydrolysates after addition to fibroblasts for 48 hours. (a) fibroblast viability; (b) Fibronectin secretion; (c) Procollagen I secretion; (d)Hyaluronan secretion. NC means negative control, which is not added. Data were presented as mean±S.D., n=6. *P <0.05, ** P <0.01, *** P <0.001 and **** P <0.0001 (one way ANOVA test).
Figure 2.
Effect of perch hydrolysates after addition to fibroblasts for 48 hours. (a) fibroblast viability; (b) Fibronectin secretion; (c) Procollagen I secretion; (d)Hyaluronan secretion. NC means negative control, which is not added. Data were presented as mean±S.D., n=6. *P <0.05, ** P <0.01, *** P <0.001 and **** P <0.0001 (one way ANOVA test).
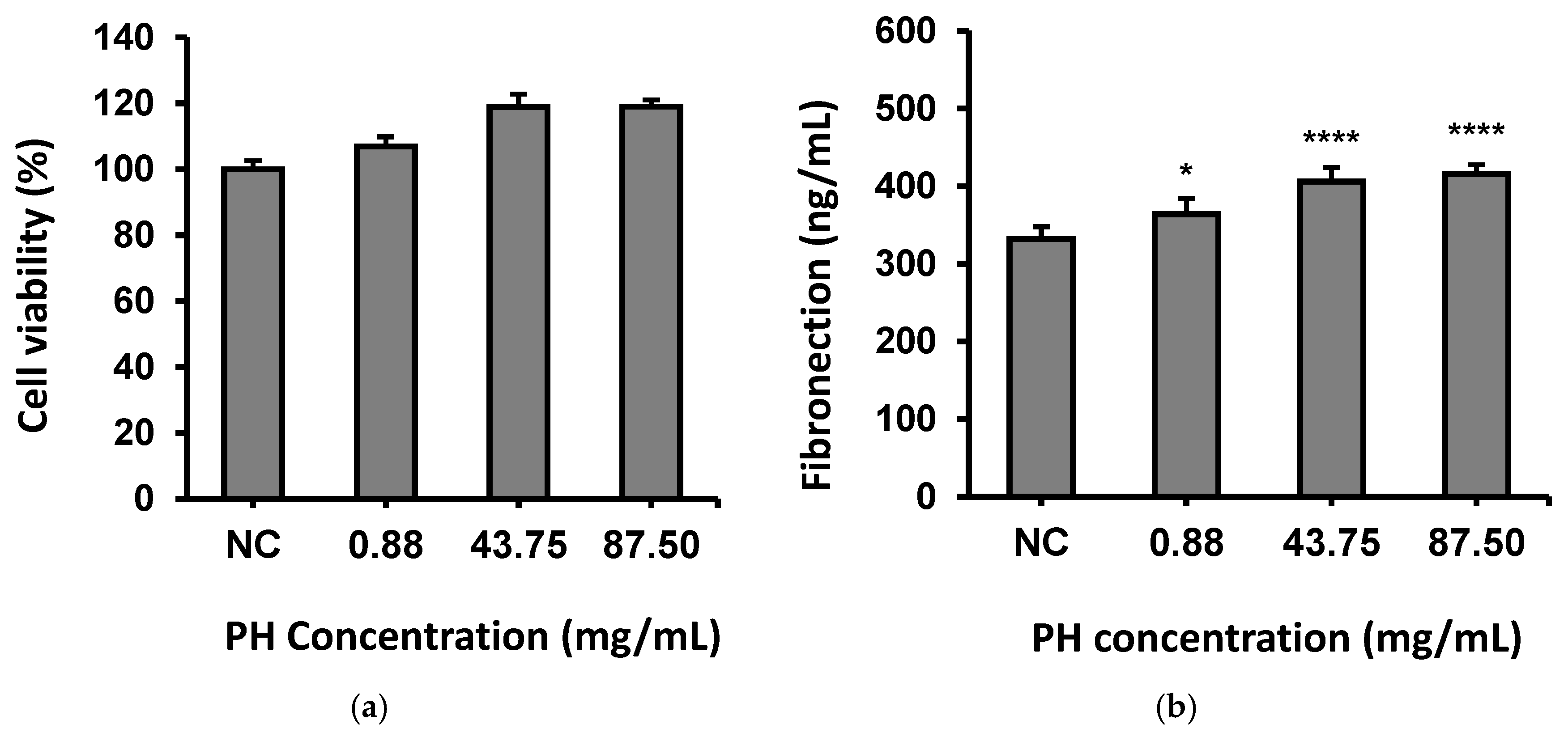
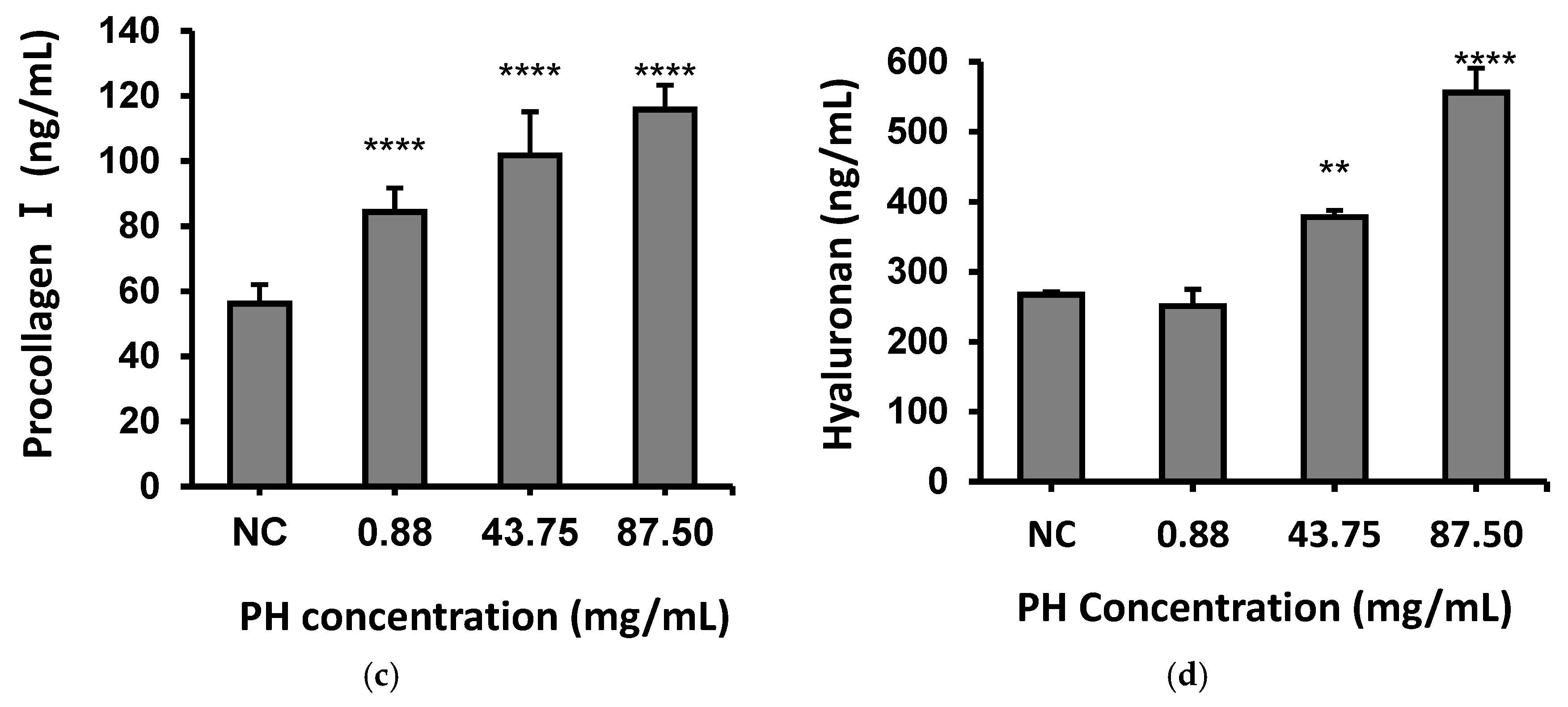

Table 1.
α-Amino group concentration and the hydrolysis degree during enzymatic hydrolysis at different times.
Table 1.
α-Amino group concentration and the hydrolysis degree during enzymatic hydrolysis at different times.
| Hydrolysis time (hr) |
α-Amino group concentration (mg/mL) |
Hydrolysis degree (%) |
|---|---|---|
| 0 | 4.62 ± 0.34 | 10.68 |
| 2 | 9.63 ± 0.28 | 22.28 |
| 4 | 12.43 ± 0.17 | 28.75 |
| 5 | 18.66 ± 0.32 | 43.14 |
| 6 | 18.88 ± 0.64 | 43.67 |
| 10 | 21.49 ± 0.09 | 49.69 |
| 24 | 24.84 ± 0.23 | 57.45 |
Note: Table shows α-amino group concentration and hydrolysis degree in Perch hydrolysates during enzymatic hydrolysis. Data are mean ± SD from four experiments (n=4), showing increasing trends indicative of protein hydrolysis by gel filtration chromatography, highlighting protein breakdown during hydrolysis.
Table 2.
Analysis of molecular weight distribution in perch essence and perch hydrolysates using gel filtration chromatography.
Table 2.
Analysis of molecular weight distribution in perch essence and perch hydrolysates using gel filtration chromatography.
| Molecular weight (Da) | Percentage (%) | |
|---|---|---|
| Perch essence | Perch hydrolysate (PH) | |
| >2300 | 62.9 | 18.4 |
| 2300-1200 | 14.5 | 15.2 |
| 1199-580 | 10.7 | 15.8 |
| 579-240 | 7.9 | 17.0 |
| 239-189 | 1.3 | 4.0 |
| <189 | 2.7 | 29.6 |
| Average | 5843 | 1289 |
Note: Table displays molecular weight distribution in perch essence and hydrolysates (PH) analyzed.
Table 3.
Concentrations of free amino acids in perch hydrolysates.
| Amino Acid | Concentration (mg/100g) |
|---|---|
| Leucine | 212.84 |
| Lysine | 113.29 |
| Valine | 96.22 |
| Arginine | 94.60 |
| Phenylalanine | 84.75 |
| Isoleucine | 87.41 |
| Methionine | 62.24 |
| Tyrosine | 50.58 |
| Proline | 20.02 |
| Carnosine | 8.80 |
| Tryptophan | 4.80 |
| β-Aminoisobutyric Acid | 4.75 |
| DL-plus allo-δ-Hydroxylysine | 4.52 |
| Cystathionine | 4.47 |
| Cysteine | 3.44 |
| Ornithine | 2.31 |
| γ-Aminobutyric Acid | 2.32 |
| Hydroxyproline | N.D. |
| Total | 890.58 |
Note: Free amino acid concentrations in Perch hydrolysates demonstrates Leucine is the most abundant. 'N.D.' stands for 'Not Detected.'.
Table 4.
Concentration of hydrolyzed amino acids in perch hydrolysate.
| Hydrolyzed Amino Acid Profiles | Concentration (mg/100g) |
|---|---|
| Glutamic acid | 957 |
| Aspartic acid | 660 |
| Glycine | 631 |
| Lysine | 574 |
| Alanine | 500 |
| Leucine | 499 |
| Arginine | 443 |
| Proline | 383 |
| Threonine | 332 |
| Valine | 332 |
| Isoleucine | 290 |
| Serine | 283 |
| Phenylalanine | 256 |
| Methionine | 202 |
| Hydroxyproline | 169 |
| Histidine | 166 |
| Total | 6677 |
Note: Table shows concentrations of hydrolyzed amino acids in perch hydrolysates, highlighting high levels of glutamic acid, aspartic acid, and glycine.
Table 5.
Relative abundance of peptide sequence in perch hydrolysates.
| Sequence | Relative abundance (%) |
|---|---|
| FPSLVRGP | 6.21% |
| GGLLCTHYNRLAV | 5.16% |
| GALGMLGDYSLV | 2.89% |
| LNLAMNALDLYL | 2.87% |
| MGLLCKGSPATP | 2.79% |
| TSEGTRVAPW | 2.66% |
| QLGMLMYGPGLTGQ | 1.99% |
| PEDVLLDAFKVLDPKYHRT | 1.90% |
| MGLGTPWLNQF | 1.72% |
| GMTGLWPW | 1.49% |
Note: The table shows FPSLVRGP being the most prevalent in the relative abundance of peptide sequences in perch hydrolysates.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



