
Submitted:
18 December 2024
Posted:
19 December 2024
You are already at the latest version
Abstract
(1) Background: in this study, bee pollen, fresh and aged beebread, collected in the central Italy during the spring/summer 2024, were analysed as reservoir of potential new probiotic yeast strains. (2) Methods: culture dependent methods and molecular analyses were used to quantify and identify bacteria, molds and yeasts populations (3) Results: microbiological analyses of pollen showed a clear dominance of molds and bacteria over yeasts in all samples. In mature beebread the presence of lactic acid or other bacterial metabolites preserved the development of molds that were almost absent. As a general abundance, yeasts were about ten times less than bacteria, in particular the osmophilic yeasts were more abundant in pollen. Specifically, four yeast genera were identified in bee pollen, Cryptococcus, Starmerella, Bullera, Microstroma and five in the beebread, Starmerella, Zygosaccharomyces, Metschnikowia, Aureobasidium, Kodamaea and Moniliella. (4) Conclusions: out of 58 assayed yeasts, 9 strains exhibited the ability to resist to gastrointestinal physicochemical condition and 4 possessed all probiotic traits tested, demonstrating the effectiveness of pollen and beebread as natural source for new bioactive and functional yeasts.
Keywords:
pollen
; probiotic yeasts
; fresh beebread
; aged beebread functional food
1. Introduction
Bee pollen is a ball or pellet of field-gathered flower pollen packed by worker honeybees and used as the primary food source for the hive [1]. It consists of simple sugars, protein, minerals and vitamins, fatty acids, and a small percentage of other components (average content 54.22% carbohydrates, 21.30% proteins, 5.31% and 2.91% lipids). Bee pollen is stored in brood cells, mixed with saliva, and sealed with a drop of honey. In food industry, bee pollen is manually harvested from hives with the use of device called pollen traps [2] and commercialized as natural healthy product for humans as having various, but not completely demonstrated and controversial health benefits [3]. Several studies agree in demonstrating wide range of bioactivities of the bee pollen, such as antimicrobial, antioxidant, antiradical, anticancer, anti-inflammatory, hepatoprotective, anti-atherosclerotic, and immunomodulatory activities [3]. Moreover, bee pollen improves blood supply to the nerve tissue, thereby increasing mental performance and eliminating the state of fatigue. Research works have also shown a positive effect on some diseases of the liver, heart and prostate [3,4,5]. However, plant allergy problems related to pollen consumption together with the lack of legislation guaranteeing the microbiological safety of this food, make it susceptible to controversy [6]. In beekeeping, after pollen collection by forager bees the next step is the storage into honeycomb cells [7], and further enriched with honey, as well as digestive enzymes and organic acids that are contained in the secretions of the salivary glands of bees[8]. At this time, a spontaneous fermentation conducted by Lactic Acid bacteria, that spontaneously present in the honeycombs occurred. Although there are few studies to support it, the transformation into beebread is also mediated by yeasts that intervene halfway through the process, when the pH drops.
The microbial occurrence in bee-collected pollen has been evidenced of about 105 UFC/g of bacteria which is lower of about 3/4 Log orders if compared to other fermented food matrices, due to the low water activity [9]. However, a microbial bacteria-yeasts succession occurs and starting from the second day the concentration of yeasts becomes comparable with that of bacteria; in aged beebread yeasts concentration decreases [10].
It is assumed that fermentation of the beebread mixture and the pre-digestion of pollen grains by added bee enzymes as well as the beebread microbiota preserve beebread and promote its nutritional value [11]. Indeed, pollen and beebread also contain a wide variety of other health promoting compounds present in functional foods, such as prebiotics, probiotics, fiber, lignans, triterpenes, carotenoids, bioactive peptides and organic acids. Moreover, the microbial community of beebread seems to produce their own antimicrobial compounds thus further contributing to its bioactivity. Some authors determined the occurrence and highlighted the involvement of lactic acid bacteria suggesting their functional role during beebread fermentation producing vitamins and other metabolites [10,11,12]
Starting from the assumption that pollen fermentation has a functional key role and considering the lack of studies regarding yeasts occurrence and involvement in pollen and beebread production, the aim of this work was to monitor the mycobiota and search for possible new strains with probiotic traits.
2. Materials and Methods
2.1. Bee pollen collected and beebread sampling
A total of 24 samples of ivy bee pollen (monofloral Hedera helix L.), fresh and aged beebread (1-4 and 25-30 days hive stored, respectively) were sterilely collected from Val di Castro and Argignano locations (43°21’53.6”N, 13°02’56.5”W, 930 m a.s.l; 43°32’47.45”N and 12°95’07.91’’W, respectively) in Marche region, Italy, during May-September 2024. Samples were immediately transported to the laboratory and processed.
2.2. Processing of samples
The harvested samples were subjected to microbial analyses through viable cell counts: the samples were weighed (30 g) and serial dilution in 0.9% sterile peptone water was carried out, homogenized for 1 min and spread on WL nutrient agar plates (Wallerstein Laboratories, Oxoid, Hampshire, UK) supplemented with 0.005% chloramphenicol (Sigma‒Aldrich, Saint Louis, MO, USA) to count and isolate yeasts and moulds. The plates were incubated at 25 °C for 3-4 days and the yeasts population was expressed as colony-forming unit (CFU) per grams of each bee product. Malt yeast agar (Oxoid, Oxford, MYA) added with 50% of D-glucose was used as selective medium for osmophilic yeasts, PCA (Liofhilchem, Roseto degli Abruzzi, Teramo) was used for mesophilic bacterial growth.
2.3. Isolation and identification of yeasts
The yeast isolation was carried out based on macro- and micromorphological characteristics, in proportion to the frequency of each yeast morphotype by plates containing between 30 and 300 colonies. Approximately 10 colonies per plate were purified on YPD agar plates (1% yeast extract, 2% glucose, 2% peptone, 2% agar) and stored for long term period in YPD broth added with 40% glycerol at -80 °C.
Purified yeast strains were then identified through ITS1- 5.8S rRNA-ITS2 region analyses. Briefly, the DNA of each isolate was extracted following the method described by Stringini et al. (2008) [12] and amplified by PCR using the primer pairs ITS1 (5′-TCCGTAGGT GAACCTCGCG-3′)-ITS4 (5′-TCCTCCGCTTTATTG ATATGC-3′) [13]. PCR products were separated by horizontal electrophoresis (Bio-Rad, Hercules, CA, USA) in a 1.5% (w/v) agarose gel using 0.5×TBE buffer and used for identification by sequencing. The genomic sequences obtained were compared with those already present in the data library using the BLAST program [14] and the GenBank database. A total of 17 sequences were included in the NCBI GenBank data library under accession numbers PQ571343 to PQ571359. To exclude the possibility of clones inside the same yeast species, the ISSR-PCR protocol was applied following the procedure reported by [15].
2.4. Potential probiotic features of isolated yeasts
The yeasts were screened for potential probiotic behaviour via in vitro assays, and a first selection was made excluding those not able to growth at 37 °C and in presence of low pH and pepsin (like stomach conditions), and then pH 7.0 and bile salts (like intestinal conditions) following the procedure reported by Agarbati et al., 2024 [15]. Only surviving yeast strains were subsequently tested for hydrophobicity, auto-aggregation, adhesion to Caco-2 cells and biofilm formation to analyse properties involved in intestinal mucosae interactions. Briefly, hydrophobicity property was analysed spectrophotometrically as the ability of hexadecane to catch cells when added in a cell suspension. Water was added to the cell suspension instead hexadecane as control [16]. Auto-aggregation was analysed spectrophotometrically as the ratio between the turbidity of a cell suspension under vigorous agitation and immediately after its vortex [15]. The adhesion of selected yeasts to Caco-2 cells (derived from human colon adenocarcinoma) was carried out seeding 4.5 × 105 cell/mL Caco-2 cells in 24-well plates and then adding 1 mL of yeast suspension having a concentration of 4.5 × 106 cell/mL. After 1 h of incubation time, non-adherent yeast cells were removed and the adherent yeast cells were collected and enumerated to know the adhesion ability of yeasts tested to Caco-2 cells [15]. The yeast ability to produce biofilm was investigated applying the protocols proposed by Speranza et al. [17] inoculation Log 5 cells and shortening the incubation time, reading the results after 3 days.
The % of biofilm production was calculated as the ratio between sessile and total cells (sessile cells and free cells).Phytase activity was evaluated following the protocol proposed by Ogunremi et al. (2020)[18] with some modifications: overnight pre-culture of yeasts was inoculate in 250 ml flask containing 150 ml of minimal salt medium and Na-phytate, then incubated at 30°C for 24 h in a shaking system (120 rpm). Culture supernatant with extracellular phytase was obtained after centrifugation and used for phytase assay as suggested by Kim and Lei (2005) [19].
The selected yeasts were analysed also for the ability to inhibit the growth of five human pathogenic bacteria such as Escherichia coli, Listeria monocytogenes, Salmonella enterica, Staphylococcus aureus and Candida albicans following the double layer method in the plate [20]. A first layer of media containing yeasts was covered with a second layer of media containing pathogens and the antimicrobial activity of yeasts was represented by a clear zone of pathogens growth inhibition.
Probiotic microorganisms must be GRAS for humans, for this reason the potential probiotic yeasts were subjected to safety tests, including haemolytic, gelatinase and DNase activities [21] following FAO/WHO guidelines.
All the results were compared with those of the commercial probiotic S. cerevisiae var. boulardii (CODEX, Zambon Italia S.r.l., Bresso, Italy) used as control. The trials were conducted in triplicate.
3. Results
3.1. Yeast, bacterial and mold occurrence
A culture-dependent approach using selective or differential media was applied to detect and quantify the microbial community associated with pollen and fresh or aged beebread samples (Table S1, Supplementary materials). Within each sample the variability of bacterial, yeast and mold populations isolated were compared between them. Indeed, total bacterial population varied from LOG 4.7 in beebread and LOG 6.1 in pollen samples; molds concentrations decreased from LOG 5.6, LOG 3.8 and LOG 1.0 in pollen, fresh and aged beebread respectively.
The results of the evaluation of cultivable non-osmophilic yeasts revealed very comparable trends in all samples but osmophilic yeasts, as expected, were absent in bee pollen (Figure 1).
3.2. Yeast isolates identification
All 58 pure cultures obtained from 24 samples (see Table 1 Suppl mat) were clustered matching micro-macro morphologies characteristics and 26S-rDNA ITS profiles, obtaining 17 groups. A representative yeast from each group was undergone to sequencing as shown in Table 1.
Four groups were excluded since they belonged to Basidiomycetes (Bullera alba, Cryptococcus aureus, Microstroma album, Moniliella spp.), instead, Ascomycetes showed a total of 6 strains belonging to Starmerella spp. recovered both in pollen and beebread, 2 cultures of Zygosaccharomyces spp. were isolated only in fresh beebread, 2 cultures to Metschnikowia spp., and 1 strain to Kodameae ohmeri in beebread were identified. The yeast-like A. pullulans were isolated only in agedbee bread. Overall results clearly showed a lower yeast variability in pollen if compared with the matrix beebread.
3.2. Probiotic features, antimicrobial activity and safety tests
All 14 identified yeasts belonging to Ascomycetes phyla were analysed for the main probiotic characteristics. Firstly, they were evaluated for their ability to growth at 37 °C pH 2.0, then in presence of pepsin, finally at pH 7.0 and bile salts. Results reported in Table 2 showed that 9 of the yeasts analysed had been able to survive in these conditions.
The yeasts specie 54BB, 62BB and 65BB dead in acidic pH and in presence of pepsin. The yeast 65BB dead also in presence of bile salts. Subsequently, only the yeasts that passed the first tests were analysed for their properties linked to intestinal mucosa interactions. Out of the 9 yeasts that passed the first screening, four of them 15BP (Starmerella spp.), 18BB (Starmerella spp.), 67BB (Starmerella magnoliae), 88BB (Metschnikowia viticola) showed percentages of hydrophobicity, auto-aggregation and Caco-2 adhesion greater or equal to 50% and for this reason they were selected for biofilm formation and phytase analyses. In particular, the yeast 18BB showed the highest percentage of biofilm formation, much higher than all other yeasts tested and the control strain, instead no yeasts showed phytase activity.
An ISSR-PCR RAPD analysis excluded the possibility that the three Starmerella strains are clones (Figure 1s supplemental material).
The best four yeasts selected were evaluated also for antimicrobial activity against five common human pathogens. All of them showed antimicrobial action on E. coli, L. monocytogenes and C. albicans. Moreover, 18BB, 67BB and 88BB showed antagonistic activity also against S. enterica. A general poor activity was observed versus S. aureus.
All the new potential probiotic yeasts did not show DNase, proteolytic and haemolytic activities, thus resulting to be safe.
4. Discussion
Yeasts that are present in gastrointestinal traits have evolved to survive hostile environments that are characterized by very low pH, high salt content, high temperature and high concentrations of inhibitory compounds. Therefore, these isolated yeasts could hide interesting characteristics for biotechnological applications. Most well-characterized probiotic microbes are bacteria, such as Bifidobacteria and Lactobacillus [22], certain yeasts have been shown to have health benefits [23] although lower studied. Indeed, the knowledge of probiotic yeasts in research on pharmaceutical fields is currently very limited. On the other side, the growing demand for functionalised or probiotic-rich food forces humans to search alternatives to probiotic bacteria where the technology related to the formulation often shows limitations due to high costs or low viability of the strain [24]. Probiotic preparations come in various forms: capsules, suspensions, powders, and combined into probiotic food. All these forms could be applied for yeasts to reduce the issue of a loss of viability during both processing and storage [25]. Moreover, yeasts are among the preferred candidates due to their easy management. In this regard, yeasts show exploitable technological characteristics that include the ability to easily produce large quantities of biomass, resistance to conservation procedures such as freeze-drying with high shelf life in the finished product, genetic stability and no deterioration of the organoleptic characteristics of the final products. The present study aimed to identify indigenous yeast strains that can be used as new probiotic candidates. Bee products such as pollen and beebread, owing to their nutritional and medicinal properties, are considered as an important food supplement for humans, rich in macro-, micro-, and phytonutrients. Recently, due to the high content of phytonutrients such as phenolic compounds, anthocyanins, volatiles, and carotenoids and unsaturated fatty acids (USFAs) of improved lipid profile such as linoleic, linolenic, and oleic pollen and beebread are classified as super-foods [26]. For these reasons it appears to be of fundamental importance to expand knowledge regarding microorganisms colonizing beebread and those involved in beebread production, with particular attention on yeasts population, helping to fill the lack of information on this regard [27]. At the same time, beebread, as natural fermented product, could exploit a source of isolation of yeasts with peculiar biotechnological characteristics, such as probiotic features, representing a starting point of this work. The first works on the isolation of probiotic yeasts appeared in literature at the end of 1990 [28,29] and the matrices firstly used for isolation were principally medical samples such as oropahringeo biofilms, mammalian intestine or infant feces. Later, some food related products such as dairy products, worldwide naturally fermented foods, fermented table olives or other beverages [30,31]. However, attention was mainly focused on lactic acid bacteria, which were more beneficial. The difficulty in maintaining the stability of probiotic LAB, from production to consumption, has been the key to the food industry’s research and development of new probiotic yeast strains.
There is a lack of recent studies that address yeasts in honeybee-collected pollen and beebread (Agarbati et al., 2023). However, a yeast-species-dependent affinity on bee agro-ecosystem was recognised by the addition of different yeast species to artificial nectar fed to bumble bees [32]. Physiologically and ecologically different groups of yeasts may also be involved in pollen resources of honeybees with potentially different effects on bee fitness, and it seems appropriate to address the yeast diversity during the storage and maturation of beebread by culture-based techniques.
Our findings reveal the presence of Basidiomycetes and Ascomycetes in the samples analysed, paying more attention to Ascomycetes where Metschnikowia, Starmerella, and Zygosaccharomyces genera were the most abundant yeasts detected, in accordance with Detry et al. (2020) [10] whose described the same genera as the most abundant in pollen, fresh and aged beebread during a study focused on yeasts population occurrence in the different stages of beebread maturation. Yeasts belonging to Starmerella genus are well known for their association with honeybees and their relative products such as pollen and beebread [10,27,33].
It was demonstrated that some strains of Metschnikowia viticola isolated from aquaculture environment, exhibited certain probiotic properties in fishes. Specifically, [34] tested the probiotic properties of yeasts in aquaculture founding a protective effect of a wild strain of M. viticola, previously isolated from the gut of local fish [35], in the Vibrium anguillarum infection model. The positive involvement of Metschnikowia as probiotic has been studied to inhibit various infectious, inflammatory diseases or cancer inactivation. Indeed, possible cellular and molecular mechanisms of this probiotic yeast, such as influencing pathogenic bacteria, inactivation of carcinogenic compounds, especially those derived from food, improvement of intestinal barrier function, modulation of immune responses, antitoxic function, apoptosis, and anti-proliferative effects are on the base of human healthy [36].
Regarding the yeast strains belonging to the Starmerella genus here results as potential probiotics, any studies currently highlighted the effectiveness of this genus as healthy microorganism [37].
5. Conclusions
This work highlighted the effectiveness of pollen and beebread matrices as reservoir for new possible functional yeasts with also probiotic traits, confirming the relevance of naturally fermented substrates as source of them. In conclusion, the results of this study show that new yeasts isolate belonging to Starmerella spp. (15BP, 18BB), S. magnoliae (67BB) and M. viticola (88BB) were found to possess the desirable in vitro probiotic properties which may serve to plan further in vivo studies. Moreover, demonstrated promising probiotic attributes suggests the potential application of yeasts as for the functional food industry.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, F.C. and L.C,; methodology, S.G. and A.A.; software, A.A.; validation, M.C., F.C. and L.C.; formal analysis, A.A..; investigation, F.C., L.C., M.C., S.G. and A.A..; data curation, A.A..; writing—original draft preparation, M.C., L.C., F.C..; writing—review and editing, M.C.; visualization, A.A.; supervision, M.C., L.C., F.C.; project administration, M.C.; All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
References
- Gilliam, M. MICROBIOLOGY OF POLLEN AND BEE BREAD : THE GENUS BACILLUS. Apidologie 1979, 10, 269–274. [Google Scholar] [CrossRef]
- Campos, M.G.R.; Bogdanov, S.; De Almeida-Muradian, L.B.; Szczesna, T.; Mancebo, Y.; Frigerio, C.; Ferreira, F. Pollen Composition and Standardisation of Analytical Methods. J. of Api. Res 2008, 47, 154–161. [Google Scholar] [CrossRef]
- Denisow, B.; Denisow-Pietrzyk, M. Biological and Therapeutic Properties of Bee Pollen: A Review. J Sci Food Agric 2016, 96, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Mărgăoan, R.; Stranț, M.; Varadi, A.; Topal, E.; Yücel, B.; Cornea-Cipcigan, M.; Campos, M.G.; Vodnar, D.C. Bee Collected Pollen and Bee Bread: Bioactive Constituents and Health Benefits. Antioxidants 2019, 8, 568. [Google Scholar] [CrossRef]
- Zakaria, Z.; Othman, Z.A.; Suleiman, J.; Nna, V.; Mohamed, M. Pollen and Bee Bread and Liver Health. In; 2022; pp. 283–314 ISBN 978-0-323-85400-9.
- Végh, R.; Csóka, M.; Sörös, C.; Sipos, L. Food Safety Hazards of Bee Pollen – A Review. Trends in Food Science & Technology 2021, 114, 490–509. [Google Scholar] [CrossRef]
- Fuenmayor B, C.; Zuluaga D, C.; Díaz M, C.; Quicazán De C, M.; Cosio, M.; Mannino, S. Evaluation of the Physicochemical and Functional Properties of Colombian Bee Pollen. Rev MVZ Córdoba 2014, 19, 4003–4014. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and Bee Bread as New Health-Oriented Products: A Review. Trends in Food Science & Technology 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-stored Pollen of Honey Bees: Many Lines of Evidence Are Consistent with Pollen Preservation, Not Nutrient Conversion. Molecular Ecology 2014, 23, 5904–5917. [Google Scholar] [CrossRef] [PubMed]
- Detry, R.; Simon-Delso, N.; Bruneau, E.; Daniel, H.-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms 2020, 8, 1789. [Google Scholar] [CrossRef]
- Gilliam, M. Identification and Roles of Non-Pathogenic Microflora Associated with Honey Bees. FEMS Microbiology Letters 1997, 155, 1–10. [Google Scholar] [CrossRef]
- Stringini, M.; Comitini, F.; Taccari, M.; Ciani, M. Yeast Diversity in Crop-Growing Environments in Cameroon. International Journal of Food Microbiology 2008, 127, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Zarzoso, B. Identification of Yeasts by RFLP Analysis of the 5.85 rRNA Gene and the Two Ribosomal Internal Transcribed Spacers. International Journal of Systematic Bacteriology.
- Altschul, S. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Research 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Agarbati, A.; Moretti, L.; Canonico, L.; Ciani, M.; Comitini, F. Agro-Ecosystem of Honeybees as Source for Native Probiotic Yeasts. World J Microbiol Biotechnol 2024, 40, 147. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Altieri, C.; Sinigaglia, M.; Corbo, M. Challenges for the Production of Probiotic Fruit Juices. Beverages 2015, 1, 95–103. [Google Scholar] [CrossRef]
- Speranza, B. Biofilm Formation by Potentially Probiotic Saccharomyces Cerevisiae Strains. Food Microbiology 2020. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.R.; Agrawal, R.; Sanni, A. Production and Characterization of Volatile Compounds and Phytase from Potentially Probiotic Yeasts Isolated from Traditional Fermented Cereal Foods in Nigeria. Journal of Genetic Engineering and Biotechnology 2020, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Lei, X.G. An Improved Method for a Rapid Determination of Phytase Activity in Animal Feed1. Journal of Animal Science 2005, 83, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Marini, E.; Zannini, E.; Ciani, M.; Comitini, F. Potential Probiotic Yeasts Sourced from Natural Environmental and Spontaneous Processed Foods. Foods 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Pereira, W.A.; Mendonça, C.M.N.; Urquiza, A.V.; Marteinsson, V.Þ.; LeBlanc, J.G.; Cotter, P.D.; Villalobos, E.F.; Romero, J.; Oliveira, R.P.S. Use of Probiotic Bacteria and Bacteriocins as an Alternative to Antibiotics in Aquaculture. Microorganisms 2022, 10, 1705. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann Nutr Metab 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Czerucka, D.; Piche, T.; Rampal, P. Review Article: Yeast as Probiotics – Saccharomyces Boulardii. Aliment Pharmacol Ther 2007, 26, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Kiepś, J.; Dembczyński, R. Current Trends in the Production of Probiotic Formulations. Foods 2022, 11, 2330. [Google Scholar] [CrossRef]
- Staniszewski, A.; Kordowska-Wiater, M. Probiotic and Potentially Probiotic Yeasts—Characteristics and Food Application. 2021.
- Baky, M.H.; Abouelela, M.B.; Wang, K.; Farag, M.A. Bee Pollen and Bread as a Super-Food: A Comparative Review of Their Metabolome Composition and Quality Assessment in the Context of Best Recovery Conditions. Molecules 2023, 28, 715. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Gattucci, S.; Canonico, L.; Ciani, M.; Comitini, F. Yeast Communities Related to Honeybees: Occurrence and Distribution in Flowers, Gut Mycobiota, and Bee Products. Appl Microbiol Biotechnol 2024, 108, 175. [Google Scholar] [CrossRef]
- Giger-Reverdin, S.; Bezault, N.; Sauvant, D.; Bertin, G. Effects of a Probiotic Yeast in Lactating Ruminants: Interaction with Dietary Nitrogen Level. Animal Feed Science and Technology 1996, 63, 149–162. [Google Scholar] [CrossRef]
- Busscher, H.J. Effect of Probiotic Bacteria on Prevalence of Yeasts in Oropharyngeal Bio®lms on Silicone Rubber Voice Prostheses in Vitro.
- Gusils, C.; Chaia, A.P.; González, S.; Oliver, G. Lactobacilli Isolated from Chicken Intestines: Potential Use as Probiotics. Journal of Food Protection 1999, 62, 252–256. [Google Scholar] [CrossRef]
- Jakobsen, M.; Narvhus, J. Yeasts and Their Possible Beneficial and Negative Effects on the Quality of Dairy Products. International Dairy Journal 1996, 6, 755–768. [Google Scholar] [CrossRef]
- Pozo, M.I.; Van Kemenade, G.; Van Oystaeyen, A.; Aledón-Catalá, T.; Benavente, A.; Van Den Ende, W.; Wäckers, F.; Jacquemyn, H. The Impact of Yeast Presence in Nectar on Bumble Bee Behavior and Fitness. Ecological Monographs 2020, 90, e01393. [Google Scholar] [CrossRef]
- Rosa, C.; Lachance, M.; Silva, J.; Teixeira, A.; Marini, M.; Antonini, Y.; Martins, R. Yeast Communities Associated with Stingless Bees. FEMS Yeast Research 2003, 4, 271–275. [Google Scholar] [CrossRef]
- Caruffo, M.; Navarrete, N.; Salgado, O.; Díaz, A.; López, P.; García, K.; Feijóo, C.G.; Navarrete, P. Potential Probiotic Yeasts Isolated from the Fish Gut Protect Zebrafish (Danio Rerio) from a Vibrio Anguillarum Challenge. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Raggi, P.; Lopez, P.; Diaz, A.; Carrasco, D.; Silva, A.; Velez, A.; Opazo, R.; Magne, F.; Navarrete, P.A. D Ebaryomyces Hansenii and R Hodotorula Mucilaginosa Comprised the Yeast Core Gut Microbiota of Wild and Reared Carnivorous Salmonids, Croaker and Yellowtail. Environmental Microbiology 2014, 16, 2791–2803. [Google Scholar] [CrossRef]
- Saber, A.; Alipour, B.; Faghfoori, Z.; Yari Khosroushahi, A. Cellular and Molecular Effects of Yeast Probiotics on Cancer. Critical Reviews in Microbiology 2017, 43, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Bai, X.; Zhang, Y.; Gao, Q.; Bu, X.; Xu, Y.; Guo, N. Evaluation of the Potential Probiotic Yeast Characteristics with Anti-MRSA Abilities. Probiotics & Antimicro. Prot. 2022, 14, 727–740. [Google Scholar] [CrossRef]
Figure 1.
Microbial population occurrence in pollen, fresh beebread and aged beebread. Results report quantitatively bacteria ( ), molds (
), molds ( ) and yeasts as osmophilic (
) and yeasts as osmophilic ( ) or non-osmophilic (
) or non-osmophilic ( ). Bubble marked with * indicate the group to which the probiotic yeasts belong.
). Bubble marked with * indicate the group to which the probiotic yeasts belong.
), molds () and yeasts as osmophilic () or non-osmophilic (). Bubble marked with * indicate the group to which the probiotic yeasts belong.
Figure 1.
Microbial population occurrence in pollen, fresh beebread and aged beebread. Results report quantitatively bacteria (), molds () and yeasts as osmophilic () or non-osmophilic (). Bubble marked with * indicate the group to which the probiotic yeasts belong.
), molds () and yeasts as osmophilic () or non-osmophilic (). Bubble marked with * indicate the group to which the probiotic yeasts belong.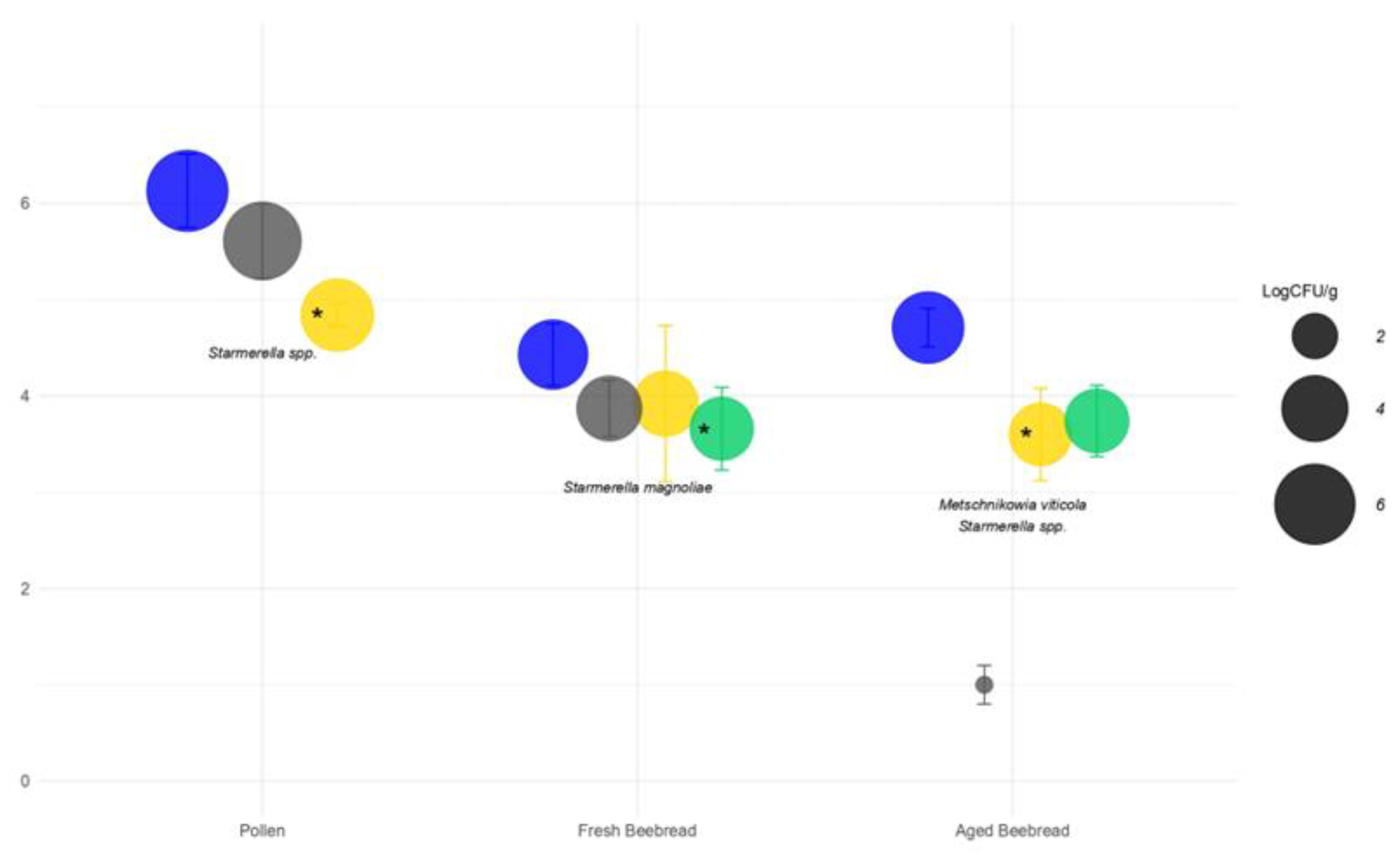

Table 1.
Identification, source and GeneBank accession number of yeast isolates.
| Source | Isolate’s code | Yeast species identification | GeneBank accession number | Grouping | |
|---|---|---|---|---|---|
| Bee pollen | 1BP | Cryptococcus aureus | PQ571343 | Group 1 | |
| Bee pollen | 3BP | Starmerella spp. | PQ571344 | Group 2 | |
| Bee pollen | 9BP | Bullera alba | PQ571345 | Group 3 | |
| Bee pollen | 15BP | Starmerella spp. | PQ571346 | Group 4 | |
| Bee pollen | 17BP | Microstroma album | PQ571347 | Group 5 | |
| Fresh beebread | 52BB | Starmerella magnoliae | PQ571351 | Group 6 | |
| Fresh beebread | 54BB | Zygosaccharomyces pseudorouxii | PQ571352 | Group 7 | |
| Fresh beebread | 55BB | Metschnikowia rancensis | PQ571353 | Group 8 | |
| Fresh beebread | 65BB | Zygosaccharomyces siamensis | PQ571354 | Group 9 | |
| Fresh beebread | 67BB | Starmerella magnoliae | PQ571355 | Group 10 | |
| Aged beebread | 18BB | Starmerella spp. | PQ571348 | Group 11 | |
| Aged beebread | 20BB | Aureobasidium pullulans | PQ571349 | Group 12 | |
| Aged beebread | 88BB | Metschnikowia viticola | PQ571356 | Group 13 | |
| Aged beebread | 91BB | Kodamaea ohmeri | PQ571357 | Group 14 | |
| Aged beebread | 93BB | Starmerella spp. | PQ571358 | Group 15 | |
| Aged beebread | 94BB | Moniliella spp. | PQ571359 | Group 16 | |
| Aged beebread | 21H | Aureobasidium pullulans | PQ571350 | Group 17 | |
Table 2.
Probiotic features, antimicrobial activity and safety of yeasts. The results of the ability of yeasts to growth/survive at 37 °C, pH 2.0 and in presence of pepsin or bile salts were reported as Log CFU/ml. Results of hydrophobicity, auto-aggregation and Caco-2 cells adhesion were reported as percentage value. Quantitative data are reported as mean values ± SD. Phytase activity, antimicrobial results and safety tests were reported using a semiquantitative method: “+” indicates the presence of activity, “++” indicates a strong presence of activity, “+/-” indicates a low activity and “-” indicates absence of activity.
Table 2.
Probiotic features, antimicrobial activity and safety of yeasts. The results of the ability of yeasts to growth/survive at 37 °C, pH 2.0 and in presence of pepsin or bile salts were reported as Log CFU/ml. Results of hydrophobicity, auto-aggregation and Caco-2 cells adhesion were reported as percentage value. Quantitative data are reported as mean values ± SD. Phytase activity, antimicrobial results and safety tests were reported using a semiquantitative method: “+” indicates the presence of activity, “++” indicates a strong presence of activity, “+/-” indicates a low activity and “-” indicates absence of activity.
| Probiotic features | 3BP | 15BP | 18BB | 20BB | 21H | 52BB | 54BB | 55BB | 65BB | 67BB | 88BB | 91BB | 93BB | Codex |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pollen | Pollen | Aged beebread | Aged beebread | Aged beebread | Fresh beebread | Fresh beebread | Fresh beebread | Fresh beebread | Fresh beebread | Aged bebread | Fresh beebread | Fresh beebread | Commercial strain | |
| 37 °C pH 2.0 (Log CFU/ml) |
5.74 ± 0.04 | 5.18 ± 0.15 | 5.73 ± 0.10 | 0 | 0 | 5.88 ± 0.03 | 0 | 5.24 ± 0.00 | 0 | 5.28 ± 0.03 | 4.68 ± 0.38 | 5.37 ± 0.17 | 6.44 ± 0.09 | 5.24 ± 0.26 |
| Pepsin (Log CFU/ml) |
6.10 ± 0.07 | 5.48 ± 0.15 | 5.67 ± 0.03 | 0 | 0 | 5.89 ± 0.03 | 0 | 5.47 ± 0.01 | 0 | 5.49 ± 0.10 | 4.53 ± 0.26 | 5.32 ± 0.31 | 5.93 ± 0.19 | 5.47 ± 0.25 |
| Bile salts (Log CFU/ml) |
5.74 ± 0.04 | 5.82 ± 0.08 | 6.00± 0.14 | 0 | 0 | 5.67 ± 0.04 | 0 | 5.56 ± 0.23 | 0 | 5.36 ± 0.33 | 4.28 ± 0.03 | 5.52 ± 0.13 | 4.20 ± 0.15 | 4.20 ± 0.15 |
| Hydrophobicity (%) | 59.86 ± 4.03 | 50.00 ± 4.17 | 6.21 ± 1.12 | 12.40±0.80 | 50.71 ± 1.94 | 61.34 ± 4.89 | 5.09±1.33 | 11.71±2.48 | 62.30 ± 0.18 | |||||
| Auto-aggregation (%) | 20.70 ± 0.03 | 74.56 ± 0.01 | 54.43 ± 0.02 | 35.54 ± 0.05 | 72.98 ± 0.02 | 37.51 ± 0.04 | 88.18 ± 0.04 | 81 ± 0.01 | 55.37 ± 0.03 | 91.99 ± 0.02 | ||||
| Caco-2 adhesion (%) | 36.43 ± 0.17 | 61.46 ± 0.02 | 89.92± 0.58 | 47.42 ± 0.00 | 77.33 ± 0.06 | 63.76 ± 0.15 | 84.99 ± 0.06 | 26.50 ± 0.01 | 31.70 ± 0.03 | 90.28 ± 0.01 | ||||
| Biofilm formation (%) | 0.62 ± 0.06 | 12.69 ± 0.01 | 0.26 ± 0.02 | 0.02 ± 0.03 | 0.03 ± 0.03 | |||||||||
| Phytase activity | - | - | - | - | + | |||||||||
| Antimicrobial activity | ||||||||||||||
| E. coli | ++ | + | + | - | + | ++ | + | |||||||
| L. monocytogenes | + | + | - | - | + | + | + | |||||||
| S. enterica | - | + | + | - | + | + | + | |||||||
| S. aureus | - | - | +/- | - | +/- | - | - | |||||||
| C. albicans | + | + | - | - | + | + | + | |||||||
| Safety tests | ||||||||||||||
| Haemolytic activity | - | - | - | - | - | - | - | - | - | - | - | - | ||
| Gelatinase activity | - | - | - | - | - | - | - | - | - | - | - | - | ||
| DNase activity | - | - | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



