
Submitted:
18 December 2024
Posted:
19 December 2024
You are already at the latest version
Abstract
Vaccines represent an essential tool in the prevention of infectious diseases. Upon administration, a complex interaction occurs between the vaccine formulation and the recipient's immune system, ultimately resulting in protection against disease. Significant variability exists in individual and population responses to vaccination, and these differences remain the focus of ongoing research. Notably, well-documented factors such as age, gender, and genetic predisposition influence immune responses. In contrast, the effects of overweight and obesity have not been as thoroughly investigated. Evidence indicates that high body mass index (BMI) constitutes a significant risk factor for infections in general, with adipose tissue playing a crucial role in modulating the immune response. Furthermore, suboptimal levels of vaccine seroconversion have been observed among individuals with obesity. This review provides a plausible examination of the immunity and protection conferred by various vaccines in individuals with overweight status, offering a comprehensive analysis of the mechanisms to enhance vaccination efficiency.
Keywords:
gender
; obesity
; immune response
; adipokines
; vaccine response
; inactivated vaccine
; recombinant vaccines
; mRNA vaccines
1. Introduction
The response to vaccination constitutes a multifaceted phenomenon that necessitates the proper activation of the immune response, facilitating an adequate defense against infection or disease [1]. In recent years, particularly during the vaccination campaign against the SARS-CoV-2 virus, the issues of overweight and obesity have gathered significant attention due to observations indicating that individuals with obesity have an increased risk of severe disease [2,3] and may exhibit a diminished response to vaccines [4,5,6]. Nonetheless, the literature presents conflicting reports on obesity and vaccine response, highlighting the necessity for a comprehensive understanding of the underlying mechanisms [4,5,6,7]. This article seeks to review the relationship between adipose tissue, obesity, and the immune response, examine existing vaccine literature on the topic, and explore the potential mechanisms implicated in this phenomenon.
2. Overview of Adipose Tissue Physiology and Physiopathology
Adipose tissue is a connective tissue characterized by an absence of fibroblasts and a minimal presence of fibrous structures [8]. It falls under the category of loose connective tissue instead of dense connective tissue [8,9]. Its primary cellular component is adipocytes, which store triglycerides crucial for energy metabolism. Adipocytes are the primary cell type found in adipose tissue [8,9,10]. When energy is abundant, they store it as lipid droplets and release it when the body requires it [8,9,10]. Figure 1 describes the different types of adipose cells and their location. Most scholars categorize body fat based on the predominant effects on overall health, offering a coherent framework for understanding their implications.
Statistically, women tend to possess a higher percentage of adipose tissue than men, with a tendency to accumulate fat in the subcutaneous layer [11]. In contrast, men often exhibit an accumulation of adipose tissue in the abdominal region, particularly within the visceral compartment [11]. Visceral fat is correlated with an increased risk of developing metabolic diseases.
Beyond their role in energy storage, adipocytes also have endocrine functions. They secrete a variety of adipokines, which are crucial in the homeostasis of adipose tissue and the link between adipose cells and the immune cells within the tissue. Table 1 summarizes the most relevant adipokines [12,13,14,15,16,17,18,19,20,21,22,23,24,25,26].
Other intermediates involved in adipose tissue physiological and physio-pathological responses have been identified as playing a significant role in the physiological responses of adipose tissue (Table 2). Some of these intermediates have been hypothesized to function as adipokines; however, discrepancies remain in this area of research.
The role of adipokines and related factors is critical in adipocyte and adipose tissue responses. However, these factors are not independent of other processes (endocrine, immune cell migration, and others) in the complex process of adipose tissue homeostasis.
2.1. Adipose Tissue, Gender, and Immune Response
There is an increased prevalence of overweight and obese women [44]. Gender differences are observed in the prevalence and phenotype of obesity, body fat distribution, drug efficacy, representation in clinical trials, and the varied secondary effects associated with bariatric surgery. Hence, gender emerges as a critical variable in the analysis of obesity [45].
Sex hormones significantly influence gender differences in body composition. Research indicates that women typically exhibit better insulin sensitivity despite having higher adiposity levels than men [46]. This phenomenon may be related to decreased insulin sensitivity observed after menopause, with estrogen therapy shown to enhance insulin sensitivity [45,46]. Furthermore, androgens have distinct effects on adipose tissue and insulin resistance that vary between the sexes [45,46]. Elevated androgen levels in women correlate with increased insulin resistance, while lower testosterone levels in men are linked to insulin resistance; this condition tends to improve with testosterone replacement therapy [46]. The levels of adipokines also differ between genders [47,48,49], and these variations are associated with the risk of developing type 2 diabetes [50]. It is important to note that type 2 diabetes is a chronic metabolic disease, and its effects on adipose tissue and immune response are complex.
Obesity has been associated with the aging process due to an increased subclinical inflammatory response; however, variations in hormonal conditions may lead to differing experiences of aging between genders [51]. Estrogen is particularly linked to a preferential increase in subcutaneous adipose tissue, as opposed to visceral adipose tissue, primarily induced by testosterone. This distinction also correlates with the risk of various associated diseases, notably cardiovascular conditions and cancer [51]. Despite this indirect evidence, it is imperative to recognize the significance of the hypothalamus-pituitary-adrenal axis [48]. This axis is critical not only for the regulation of sex hormones but also for the management of adipokines that play a vital role in maintaining adipose tissue homeostasis and growth.
Luo and coworkers [52] analyzed a population of children and adolescents, finding that the prevalence of inflammatory markers was higher in overweight and obese individuals. They also observed gender differences. However, the difference was less prevalent in the adolescent population. Silva and Iwasaki [53] summarized published data that post-acute infection syndromes are prevalent in females.
A recent bioinformatic analysis [54] revealed that chromosome interactions in the Eurasian admixed population revealed that the X chromosome acted on autosomal immunity-associated genes. Consequently, the immune response of admixed populations should not differ between ethnic groups. However, Persons and coworkers [55] showed differences in obesity among different races in the USA.
Figure 2 delineates the distinctions between lean and obese adipose tissue, focusing on the role of immune cells, cytokines, and sex hormones associated with visceral and subcutaneous adipose tissue. It is essential to highlight that, notwithstanding the variations presented in the figure, stable adipose tissue may exhibit immunological tolerance regardless of its size. However, under conditions of stress induced by peripheral or localized factors, the cells and cytokines can provoke an inflammatory response within the tissue. This response may result in the deregulation of metabolic processes and peripheral function, ultimately contributing to the development of insulin resistance and an exacerbated inflammatory response.
Numerous authors have documented variations in the immune response based on sex [56,57]. However, the mechanisms underlying certain responses remain undefined, and some researchers inappropriately extrapolate findings from rodent studies to humans. Popotas and colleagues [58] examined Toll-like receptors (TLRs) as potential mechanisms for elucidating responses to pathogens and other stimuli. Their rationale is grounded in the fact that the X chromosome influences TLR regulation [58]. Notably, there are disparities in TLR receptor expression and functionality when comparing humans and mice. Specifically, TLR7 and TLR8 are expressed at higher levels in female immune cells than in male cells, while TLR4 exhibits the opposite pattern [58]. Furthermore, TLR9 signaling is significantly elevated in females, correlating with increased production of interferon types I and II [58]. It can be proposed that the immune response may depend on the initial signaling that induces cellular activation. Further research is warranted to explore this matter comprehensively.
Recently, Wang P et al. showed in the murine model that Tlr9 deficiency leads to obesity [59]. This receptor has also been associated with autoimmune diseases [60]. How obesity, gender, and autoimmune diseases are connected is still under investigation.
Layug and coworkers [61] also reviewed the difference in CD8+ lymphocyte response, showing that females have more CD8+ effector and memory lymphocytes to respond to pathogens. In addition, the CD8+ lymphocyte response in males with autoimmunity and cancer is characterized by cell exhaustion compared to the females, in which the effector cells are responsible for the process. Forsyth and coworkers partially share the proposal since they envision the responses to the genetic impact of the X chromosome [62]. Several points on the X chromosome have been involved in innate and adaptative immune responses, and, according to the authors [62], these events are responsible for the increased susceptibility of males to infections. There are still many enigmas to solve in the puzzle. Still, it can be concluded that gender, sex hormones, and adipose tissue endocrine functions influence the efficiency of the immune response.
2.2. Adipocytes as Antigen-Presenting Cells
Recent research has highlighted the involvement of adipocytes in immune responses, as they can recruit and activate immune cells [63,64]. They are antigen-presenting cells (APCs) expressing CD1d and MHC class I and II molecules [65,66]. Adipocytes can directly activate CD4+ T lymphocytes through the antigen: MHCII complex in a contact-dependent manner [65,66]. A recent study has shown that adipocytes also express MHC class II molecules, along with co-stimulatory molecules CD80 and CD86, and their expression is significantly heightened in response to high-fat diets [67]. While adipocytes display MHC class I molecules like other nucleated cells, there is still inconclusive evidence regarding direct interactions with CD8+ T lymphocytes via the antigen: MHCI complex [68]. Conversely, studies have demonstrated that CD1d expressed in adipocytes can present lipid antigens to invariant natural killer T (iNKT) cells, effectively stimulating their activation [69,70].
In obesity, both local and systemic immune dysfunction arise from metabolic stress [71]. In adipose tissue, the immune cells that are normally anti-inflammatory and immune-regulatory—such as M2-type macrophages, regulatory T cells (Treg), Th2, and type 2 innate lymphoid cells (ILC2)—are replaced by a higher number of pro-inflammatory immune cells. These include M1 macrophages, Th1, Th17, Th22 and CD8+ T lymphocytes, which secrete pro-inflammatory cytokines like IL-1β, IL-6, IL-17, and IFN-γ [72,73]. This pro-inflammatory response may be exacerbated by intestinal inflammation associated with obesity [74]. In addition to the local immune changes within adipose tissue, systemic immune adaptations are also evident in obesity, characterized by increased circulating numbers of monocytes, neutrophils, and lymphocytes (Th1, Th17, and Th22), along with a decrease in circulating Treg lymphocytes and elevated levels of pro-inflammatory cytokines [72,73,74]. Collectively, these alterations create a pro-inflammatory state of the immune system in obese individuals, marked by heightened cytokine levels both locally in adipose tissue and systemically [72]. This chronically elevated inflammatory condition is believed to stimulate regulatory pathways that ultimately restrict the immune response to acute infections. A notable example is the compromised type I interferon antiviral response observed in individuals with obesity [31].
Table 3 illustrates the different cells directly and indirectly involved in adipose tissue physiology and physiopathology. The difference with Figure 2 derives from a comprehensive analysis of all the possible cells described in the white adipose tissue increase, and stress response is present [75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102]. It is important to note that the role of mesenchymal stem cells in tissue repair and remodeling is recent and is still under investigation, as well as the possible role of follicular B and T cells in the link between lymphoid organs, leukocyte migration, immune response, inflammation, and autoimmunity [103].
The mechanisms by which leptin exerts its effects on immune cells are complex, partially due to the presence of multiple isoforms of the leptin receptor generated through alternative splicing, each with distinct signaling capabilities [104]. For example, T lymphocytes predominantly express the long form of the leptin receptor, particularly following activation, whereas neutrophils primarily express the short form. On the other hand, NK cells express short and long form receptors. Individuals with genetic mutations that impair the synthesis of leptin are often morbidly obese and exhibit compromised immune defenses [104]. Obesity leads to hyperleptinemia, which can adversely affect the immune response [104,105]. Moreover, obesity has been associated with increased thymic senescence and a reduction in the diversity of the T cell repertoire, potentially impacting immune surveillance [104,105]. Numerous studies have highlighted that obesity constitutes a significant risk factor for postoperative and surgical nosocomial infections, reviewed by Muscogiuri and coworkers [45].
Deng and colleagues [106] showed that low serum leptin levels in young and elderly healthy subjects are associated with lower antibody responses to influenza and hepatitis B (HBV) vaccines. Leptin stimulates the differentiation and function of human and mouse TFH cells in culture and is also required to maintain TFH function and sustained effective humoral immunity [106]. TFH is necessary to support and maintain effective humoral immunity to infection and immunization in mice. The mechanism of action of leptin is regulated in part by activation of the Stat3 and mTOR (mechanistic target of rapamycin) pathways [106]. Their results suggest that leptin is a physiological regulator of TFH function and that leptin deficiency may serve as a biomarker to identify the risk of low vaccine efficacy. Moreover, serum leptin levels did not always correlate positively with absolute antibody titers after vaccination or changes in antibody titers in adults vaccinated against influenza or HBV. Overall, their data [106] support the notion that leptin is a natural regulator of TFH cells in the general population. This should not be interpreted to mean that higher levels of leptin are associated with higher vaccine responses; on the contrary, in their view, leptin constitutes a metabolically mediated threshold factor that is needed to mount normal vaccine responses.
Investigating the inflammatory response within adipose tissue presents a complex scenario [107,108,109]. The infiltration of various cell types into this tissue, driven by metabolic demands or stressors, increases chemokine production [107,108,109]. This increase in chemokines subsequently facilitates the migration of cells, thereby fostering an inflammatory environment [108]. The activation of macrophages by external stimuli results in a proinflammatory profile characterized predominantly by M1 macrophages, in contrast to the M2 macrophages found in stable adipose tissue. A parallel is also observed between CD8 and CD4 T lymphocyte infiltration (Th1, Th17, Th22), in which T regulatory lymphocytes are displaced. Consequently, there is a lack of tolerogenic tissue response and a high local inflammatory response. Moreover, neutrophil migration appears to be influenced by IL-17 production [108,109]. B cells' role seems to depend upon the infiltration of cells and the presence of T lymphocytes. The polyclonal stimulation of B cells may generate the formation of autoantibodies in the tissues [86,87,88]. The inhibition or resolution of the inflammatory response passes by the inhibition of IL-1β signaling by the production of IL-1RA and the secretion of TGFβ [107,108,109]. The process can be facilitated by the production of IL-10 by the local immune cells. It is also possible that the secretion of lipids from activated adipocytes modulates the inflammatory response. The saturated lipids may enhance the production of lipid intermediates, facilitating the inflammatory process [110]. On the contrary, the presence of ω3 fatty acids and resolving production [111,112].
It is important to note that the inflammatory response in adipose tissue is not uniform across different types [10]. Specifically, the increase in visceral adipose tissue correlates more strongly with insulin resistance than the increase in subcutaneous adipose tissue [45]. This suggests that subcutaneous fat may be less stable and more inflammation-resistant than visceral fat. Additionally, alterations in energy demands or surgical interventions may affect the dynamics of visceral adipose tissue, potentially enhancing the local immune cell response by stabilizing the inflammatory environment [45,46].
Cellular senescence is characterized by an irreversible arrest of the cell cycle, typically initiated by various forms of cellular stress [113,114,115]. Cells that undergo senescence exhibit a senescence-associated secretory phenotype (SASP), which includes the secretion of proinflammatory cytokines, chemokines, growth factors, and proteases [115,116,117,118]. Immunosenescence represents a complex process associated with aging, involving significant changes in the architecture and functionality of immune organs, ultimately leading to compromised innate and adaptive immune responses [115,116,117,118]. Although the precise molecular and cellular mechanisms are not fully elucidated, several prominent features of immunosenescence have been identified [115,116,117,118,119,120]. These include thymic involution, dysfunction of hematopoietic stem cells, disruption in T and B lymphocyte homeostasis, chronic low-grade inflammation (often referred to as inflammaging), accumulation of senescent cells, impaired antigen response, mitochondrial dysfunction, genomic instability, and enhanced stress responses [115,116,117,118,119,120].
Obesity contributes to the accelerated aging of adipose tissue, promoting the premature senescence of adipocytes [115,116,117,118,119,120]. Senescent adipocytes release increased amounts of free fatty acids (FFAs) and adipokines, including leptin, TNF-α, and IL-6 [115,116,117,118,119,120]. The SASP phenomenon can potentially induce senescence in adjacent tissues, particularly within the immune system [115,116,117,118,119,120]. Furthermore, adipose tissue in individuals with obesity is markedly infiltrated by B cells [118]. These adipose tissue-resident B cells are either recruited or activated by the byproducts of altered lipolysis, and the adipokines are secreted by expanding adipose tissue as they express the corresponding receptors [118]. Interestingly, Valentino [121] and coworkers analyzed the role of autoantibody formation, cell senescence, and aging, providing a fascinating insight into the process and suggesting possible therapeutic targets. The role of B1 and B2 in adipose tissue, normal immune response, and autoimmunity is still under research.
2.3. Adipocyte-Derived Extracellular Vesicles
Circulating extracellular vesicles (EVs) are recognized as significant mediators of cell-to-cell communication and the exchange of biological messages [122,123]. These lipid bilayer nanoparticles range in size from 50 to 1,000 nanometres and can be released by nearly all cell types [122]. They are present in various body fluids, including blood, saliva, urine, breast milk, and amniotic fluid. Notably, adipose tissue serves as a crucial source of circulating EVs. Research indicates that individuals with obesity generally exhibit elevated levels of EVs in their serum compared to those without [123,124]. The underlying cause of this increase remains unclear; however, it has been suggested that fatty tissue in the context of obesity may produce EVs at a higher rate or exhibit a reduced capacity for EV elimination by the liver. Importantly, interventions such as bariatric surgery or caloric restriction have been shown to decrease the number of circulating EVs, implying that a reduction in adipose tissue mass correlates with diminished EV secretion [124]. Recently, EVs have been acknowledged as effective messengers for intercellular communication. Emerging evidence highlights that adipose-derived EVs play a vital role in the interactions among macrophages, adipocytes, and adipose tissue-derived stem cells, significantly influencing immunometabolism in healthy and obese states [121,122,123,124].
The significance of microRNAs (miRNAs) in the context of adipose tissue and inflammation is noteworthy. EV derived from adipocytes and immune cells play an instrumental role in differentiating various cell types within the tissue. Rakib and colleagues [125] have reviewed a potential mechanism involving miRNA-34a, which is secreted by activated adipose tissue and functions to inhibit the transcription factor KLF4, thereby obstructing the transformation of M2 macrophages [125]. Conversely, miRNA-326 secreted by M1 macrophages enhances the expression of RORC2, resulting in the upregulation of Th17 cells, which facilitates the inflammatory response [125].
Additionally, the miRNA-34 family is implicated in cellular senescence. In conjunction with miRNA-155, miRNA-34 contributes to telomere shortening [125]. Furthermore, miRNA-146 and miRNA-181 promote cellular senescence, while miRNA-335 is involved in "inflammaging" [125]. These molecular mechanisms lead to an exacerbated local inflammatory response due to increased cell senescence and mortality, which can, in turn, promote peripheral inflammation. In conclusion, EVs derived from adipose tissue may be pivotal in mediating tissue and multi-organ senescence and contributing to peripheral inflammatory responses.
Current investigations involving seemingly healthy, obese individuals indicate notable variations in the size, quantity, and composition of EVs. Compared to non-obese individuals, these variations appear to correlate with specific metabolic parameters, such as glucose levels, insulin sensitivity, and serum lipid profiles. Considering these observations, it is hypothesized that EVs play a significant role in progressing metabolic and cardiovascular complications associated with obesity. However, the precise mechanisms underlying this process remain to be elucidated [126].
3. Obesity and Infectious Diseases
Research regarding the interaction between obesity and various infectious agents remains contentious and presents a highly intricate scenario [127,128]. The increased susceptibility to numerous types of infections among individuals with obesity is not yet fully understood. Obese individuals can have micronutrient deficiency [129], which may affect their response to viral infections, as observed in SARS-CoV-2 [130,131]. For example, the Edmonton obesity staging system reported that the impairment of vitamin D nutritional status and metabolic profile was associated with worsened obesity [132,133]. Thus, vitamin D deficiency may be linked to the impaired immune response observed in SARS-CoV-2 infection in obese individuals, and EV may play an essential role in the severity of the disease.
Additional cofactors frequently linked to obesity may indirectly contribute to the development or exacerbation of infectious diseases, even in the absence of a clear causal relationship [127,128]. These cofactors encompass modifications in respiratory physiology, skin, and soft tissue integrity changes, co-morbidities such as type 2 diabetes mellitus and cardiovascular disease, pharmacological interventions, and inadequate antimicrobial treatment [127,128].
The outcomes of infections in obese individuals and animal models appear to vary depending on the extent of the infection, likely due to differential impacts on the metabolic pathways of immune cells [10]. Obesity is a significant disruptor of bodily homeostasis, leading to alterations in immune metabolic pathways, often resulting in a diminished protective immune response to infections. The specific modifications in immune response due to obesity are still being fully elucidated. As documented in Table 2, the decreased production of IFN type I and the high secretion of IL-1RA may be related to a reduced effective response in obese individuals [31,36]. Pugliese et al. [132] analyzed the most relevant infection sites for obese patients. Upper respiratory tract infections are most commonly associated with pharyngitis, sinusitis, laryngotracheitis, lower respiratory infections, bronchitis, bronchiolitis, and pneumonia. Since sleep apnea in obese individuals increases, the risk of respiratory infections increases [134]. Hypoxia may jeopardize the response to treatment. Then, urinary tract infections (cystitis, urethritis, pyelonephritis), skin infections (high incidence of cellulitis, candida, erysipelas, onychomycosis), and surgical site infections.
A growing body of research demonstrates that women living with HIV experience a significantly elevated risk of developing metabolic disorders in comparison to their male counterparts [135]. These metabolic disorders encompass weight gain and obesity, type 2 diabetes mellitus, dyslipidemia, bone loss, and sarcopenia. Conversely, men diagnosed with HIV exhibit a greater susceptibility to conditions such as hepatic steatosis and fibrosis [135].
Table 4 summarizes the relationship between viral infections, adipose tissue, obesity, and interferon response in different reports [136,137,138,139,140,141,142,143,144,145,146,147,148,149,150]. The Table aims to provide the reader with insight concerning the pathogens, the possible role of adipose tissue to be altered upon the inflammatory response generated by the infection, and the effect of the virus on the IFN response. The decreased antiviral response and chronicity can be potentiated by the intermediates generated by adipose tissue as part of the response to the infection; however, the role of adipose tissue as a reservoir of virus or the role of adipose tissue on viral escape cannot be overlooked. More research is required in this area.
Hornung et al. [151] revised different in vivo and in vitro models to study the role of adipose tissue in bacterial and viral infections; however, most of the effort involved murine models, which are informative but do not necessarily follow the same response as humans. Hales and coworkers [152] explored the role of leptin in Streptococcus pneumonia infections and the difference between the results in humans and mice, showing the importance of the hormone in immune cell activation and response.
However, critical elements have received insufficient attention in the existing literature. The first pertains to the increase in cell death among underweight individuals with sepsis in the intensive care unit and the differences in gender responses observed in overweight and obese populations [153]. These observations prompt new inquiries regarding the incidence of infections and the immune response in obese individuals and probably better therapeutic strategies to protect individuals from severe infections.
4. Impact of Obesity on Vaccination
While substantial progress has been made through vaccination in protecting against infectious diseases, certain subpopulations exhibit suboptimal responses to these interventions. This inadequacy increases the vulnerability of these groups to vaccine-preventable illnesses. Recent evidence suggests that obesity may significantly influence vaccine immunogenicity and efficacy, potentially exacerbating the likelihood of an inadequate immune response among individuals with obesity. The negative impact of obesity on immune system functionality raises essential concerns regarding the effectiveness of vaccine-induced immunity within this demographic. The initial studies indicating a potential correlation between obesity and compromised immune response to vaccinations were published in 1985, focusing on a cohort of obese hospital employees who demonstrated a poor response to the hepatitis B vaccine [154]. Therefore, it is imperative to explore strategies to enhance the protection of this at-risk population.
4.1. Inactivated or Subunit Vaccines
Obesity may impede an individual's capacity to generate an effective immune response to vaccination or infection, a phenomenon attributable to increased body fat and elevated leptin levels. Callahan et al. [155] undertook a comprehensive analysis of pooled data from three independently conducted, NIH-supported Phase 2 clinical trials assessing monovalent, unadjuvanted, split-virus pandemic H1N1 vaccines administered at eight Vaccine and Treatment Evaluation Units (VTEUs) between August 2009 and March 2010. One trial was conducted with children and adolescents (6 months to 17 years), utilizing the Sanofi Pasteur vaccine. The other two trials, designed with identical methodologies, recruited non-pregnant adults (ages ≥18 years) and used vaccines manufactured by Sanofi Pasteur or CSL Biotherapies. Participants were randomly assigned to receive two intramuscular injections, which contained either 15 or 30 International Units (IU) of hemagglutinin (HA), measured by high-performance liquid chromatography, administered 21 days apart. The final potency evaluation of the Sanofi Pasteur vaccine, conducted using the single radial immunodiffusion (SRID) assay, indicated an HA content of 22-25 IU for the 15 IU dose (analyzed across two different batches) and 47 IU for the 30 IU dose. Among adult subjects, nearly 30% were classified as obese or morbidly obese, 37% as overweight, and only 1% as underweight. The findings concluded that a single dose of the vaccine prompted higher hemagglutination inhibition (HAI) geometric mean titers (GMT) on day 21 in obese adults compared to individuals in other BMI categories [154].
Furthermore, Clarke et al. [156] examined the effects of obesity on responses to the quadrivalent influenza vaccine in children. This study enrolled children classified as having obesity (BMI) ≥ 95th percentile for age and gender) and those without obesity (BMI < 95th percentile). Blood samples were collected before vaccination and at one- and six months post-vaccination to evaluate antibody responses utilizing the hemagglutination inhibition (HI) assay. The immunogenicity of the vaccine was compared across the two groups of children. Both groups, those with and without obesity, demonstrated robust and sustained antibody responses to the tetravalent influenza vaccine six months post-vaccination. Sheridan et al. [157] reported that a higher BMI initially correlated with an enhanced antibody response following vaccination with the inactivated trivalent influenza vaccine. Nonetheless, twelve months after vaccination, a higher BMI was associated with a more significant decline in antibody levels and a decreased presence of specific CD8+ T lymphocytes and IFNγ production in obese individuals [157].
Huang et al. [158] conducted a study involving children aged 8 to 18 who had completed their routine childhood immunizations. Serum samples were analyzed using ELISA to assess antibody levels against diphtheria, tetanus, Haemophilus influenzae type B, and Streptococcus pneumoniae, in addition to measuring serum HbA1c levels. BMI percentiles and HbA1c levels were utilized as continuous variables about antibody titer levels. The study revealed that 43% of the children had a BMI at or above the 95th percentile (n=69). A notable negative correlation was observed between BMI and the antibody titers for pneumococcal, diphtheria, and tetanus vaccines, with a significant correlation identified explicitly for the S. pneumoniae serotype P3 titer (p=0.037). The findings indicate that increased BMI and HbA1c levels are associated with lower overall vaccine titers. Additionally, the study highlighted that obese child (BMI ≥ 95%) exhibited a higher likelihood of having impaired pneumococcal titers compared to their non-obese counterparts (BMI 25-94%) in a prospective, population-based cohort study [158].
To address the significant challenges posed to global public health by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), innovative technology platforms have successfully completed their research and development efforts, including mRNA and adenoviral vector vaccines. The primary objective of these advancements was the large-scale production of vaccines targeting coronavirus disease 2019 (COVID-19). These novel methodologies facilitated a rapid and adaptable vaccine design process, ensuring the timely availability of effective vaccines to combat the pandemic. However, clinical trials examining BMI and central obesity have indicated that individuals with obesity exhibited lower antibody titers in response to vaccination compared to those of healthy weight [4,5,6,160]. This observation highlights a potential early decline in vaccine-induced antibody levels correlated with higher obesity rates. Consequently, the anticipated protective effects of SARS-CoV-2 vaccination may be diminished in individuals with obesity relative to their healthy-weight counterparts [4]. The results were challenged by other researchers [7].
Ou et al. [161] performed a meta-analysis of the literature, examining antibody responses to COVID-19 vaccinations among individuals with and without obesity. This meta-analysis incorporated findings from eleven studies, five of which provided absolute values of antibody titers for both the obese and non-obese groups. The results indicated that the obese population exhibited a statistically significant association with lower antibody titers following COVID-19 vaccination [161]. Similarly, Faizo et al. [162] reported comparable findings. Their study analyzed sera from vaccinated obese individuals (n = 73) alongside controls with a normal BMI (n = 46). The samples were analyzed for total anti-S protein and neutralizing antibodies. Additionally, a nucleocapsid ELISA was employed to differentiate between immunity obtained solely through vaccination and acquired through a combination of vaccination and recovery from infection. This study also revealed a decrease in vaccine-induced neutralizing humoral immunity among obese participants, a phenomenon observed regardless of sex, previous infection recovery, and the time elapsed since the last vaccination [162].
Well-controlled assays are essential for analyzing the effects of obesity, gender, and malnutrition—such as vitamin D deficiency—and the humoral, innate, and adaptive responses elicited by the vaccine.
4.2. Live-attenuated vaccines
Dumrisilp and colleagues [163] conducted a prospective study between November 2019 and February 2020. The participants, aged between seven and twenty-five years, were from Bangkok and the obesity outpatient center at King Chulalongkorn Memorial Hospital. A total of 212 subjects were included in the immunogenicity analysis. The enrolled individuals were vaccinated with MEVAC™-A (Hepatitis A live attenuated virus). Blood samples were collected to assess the levels of anti-HAV antibodies one day before vaccination and 8–9 weeks post-vaccination. According to prior studies, an anti-HAV IgG titer of 20 mIU/mL is deemed seroprotective. Statistical analysis revealed that a single administration of the live attenuated hepatitis A vaccine is both safe and highly immunogenic in subjects classified as either underweight/normal weight or overweight/obese during the brief follow-up period. Truncal obesity and female gender were identified as factors associated with an enhanced immune response; however, no significant differences in anti-HAV titers were observed between the non-obese and obese groups, nor between the child and young adult cohorts. It is important to note that the duration of follow-up for evaluating safety and immunogenicity was relatively brief, limited to only nine weeks [162]. Thereafter, Soponkanabhorn et al. ([164] carried out a retrospective study utilizing blood samples from Dumrisilp et al. [163]. The results of this study suggest that obesity does not influence the short-term cellular immune response to HAV live attenuated virus vaccination. However, this clinical trial had several limitations, one of which was the absence of data regarding cardiometabolic risk factors, specifically in obese participants, and the other limitation of the clinical trial is the relatively long interval between vaccination and subsequent immunogenicity testing. The long period between vaccination and evaluation may explain the absence of substantial improvement in vaccine-induced cell-mediated immunity in most participants [164].
Fonzo et al. [165] conducted a clinical trial involving 2,185 students at the School of Medicine, University of Padua (815 males and 1,370 females). The objective of this study was to examine the relationship between BMI and current antibody levels following vaccinations for measles, mumps, and rubella (MMR) and a recombinant hepatitis B virus (HBV), which were administered during childhood. The BMI was classified based on the World Health Organization (WHO) criteria. There is no significant association between BMI and the persistence of immune response after HBV and MMR vaccinations. Furthermore, no noteworthy sex-related differences were observed in the results [165].
Using SARS-CoV-2 inactivated vaccines, a decreased production of antibody titers was observed in individuals with severe obesity and BMI ≥ 40 [166], suggesting that the responses are similar to the other vaccines. A study using recombinant SARS-CoV-2 vaccine and inactivated influenza virus generated identical results; a low response was observed in obese individuals [167]. In a small observational study by Frasca and coworkers [168], the authors showed differences in B defects in obesity and an improvement when individuals successfully lost weight. These results suggest that weight reduction may decrease B lymphocyte impairment; however, well-defined critical trials should assess the improvement of patient response with strict weight loss conditions.
5. Future Perspectives
Chronic unresolved systemic and adipose tissue inflammation significantly contributes to the onset of obesity-related cardiometabolic diseases. While pharmaceuticals targeting pro-inflammatory cytokines or inflammasome activation have received clinical approval, their widespread application is often limited by severe adverse effects, including weight gain and increased susceptibility to infections. These factors hinder their broader clinical implementation. There remains a pronounced gap in the availability of biomarkers that can effectively differentiate between acute and chronic inflammation and assess the functionality of distinct leukocyte populations. Developing such biomarkers would enhance personalized treatment approaches and facilitate the monitoring of therapeutic interventions. The resolution phase of inflammation is an active and regulated process governed by specialized pro-resolving mediators, which have demonstrated efficacy in alleviating obesity-related inflammation and systemic disease in experimental models. This area represents a significant opportunity for therapeutic advancement.
Recent technological advancements are facilitating the development of more effective and innovative vaccines and adjuvants. The primary objective of these efforts is to restore inflammatory and immune homeostasis while maintaining other essential physiological processes. This aim may be accomplished by enhancing leptin sensitivity through leptin-based therapeutic strategies, encompassing synthetic or modified leptin and pharmaceuticals that selectively target leptin-induced pathways [169,170].
The use of GLP-1 therapies for obesity has generated a new area of research. The GLP-1 receptor is a negative costimulatory receptor, leading to possible use in cancer and other diseases. However, to our knowledge, the role of these treatments on vaccine effectiveness has not been published. One may assume that it will impact future therapeutics [171].
It is also critical to analyze the possibility of autoimmune disease in the process of adipose tissue dysregulation that could lead to a less effective response [121]. Prologued SARS-CoV-2 infection and other chronic viral diseases [86,167,172,173] have been related to an increased incidence of autoimmunity, but the mechanism by which obesity is linked to the generation of autoimmune disease and circulating autoantibodies is still a matter of research.
6. Conclusions
In the current review, summarized in Figure 3, we have outlined the most significant aspects of adipose tissue physiology, the influence of sex hormones, adipokines, associated cytokines, and the presence of local immune cells under normal conditions and during inflammatory responses. Additionally, we have examined the role of sex hormones in these processes. Despite our thorough investigation, we have only partially addressed the critical components necessary for understanding the reasons behind the diminished vaccine responses observed in specific obese individuals. Further research is essential in this domain, particularly considering the growing prevalence of overweight and obesity globally.
From our perspective, resting adipose tissue is characterized by a tolerogenic environment, which does not influence insulin resistance or lipid metabolism and, consequently, does not impact the immune response to vaccinations. In contrast, subclinical inflamed adipose tissue, marked by immune cell migration and the production of local and peripheral pro-inflammatory cytokines and adipokines, does influence responses to infections and vaccines. To better delineate this difference, it is essential to identify specific biomarkers that account for variables such as gender, sex hormones, the hypothalamic-pituitary-adrenal axis, nutritional factors, and both genetic and epigenetic modifications.
The vaccines developed to combat the SARS-CoV-2 virus have introduced a novel framework for analyzing the vaccination process. Nevertheless, further research is essential to understand the limitations of these vaccines, particularly regarding their safety, efficacy, and overall effectiveness within the population.
Author Contributions
Conceptualization, A.H.G., and J.B.D.S..; investigation, A.H.G., G.B.N., and J.B.D.S.; resources, A.H.G. and J.B.D.S..; writing—original draft preparation, A.H.G.; writing—review and editing, , A.H.G., G.B.N., and J.B.D.S.; supervision, A.H.G., and J.B.D.S; project administration, A.H.G., and J.B.D.S; funding acquisition, A.H.G., and J.B.D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed by the National Fund for Science, Technology, and Innovation (FONACIT), an entity attached to the Ministry of Popular Power for Science and Technology of the Bolivarian Republic of Venezuela (MINCYT). JBDS is partially financed by the National Institute of Virology and Bacteriology [Program EXCELES, ID Project No. LX22NPO5103]—Funded by the European Union—Next Generation EU from the Ministry of Education, Youth, and Sports of the Czech Republic (MEYS)]. Also partially supported by a grant from the Ministry of Education, Youth, and Sport, Czech Republic: Molecular and Cellular Clinical Approach to Healthy Ageing, ENOCH (European Regional Development Fund Project No. CZ.02.1.01/0.0/0.0/16_019/0000868, IMTM #869/V19).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- https://www.cdc.gov/vaccines/basics/explaining-how-vaccines-work.html. Assessed December 8, 2024.
- Petrakis, D.; Margină, D.; Tsarouhas, K.; Tekos, F.; Stan, M.; Nikitovic, D.; et al. Obesity - a risk factor for increased COVID-19 prevalence, severity, and lethality. Mol Med Rep. 2020 Jul;22(1):9-19. [CrossRef]
- Russo, A.; Pisaturo, M.; Zollo, V.; Martini, S.; Maggi, P.; Numis, F.G., et al. Obesity as a Risk Factor of Severe Outcome of COVID-19: A Pair-Matched 1:2 Case-Control Study. J Clin Med. 2023 Jun 15;12(12):4055. [CrossRef]
- Nasr, M.C.; Geerling, E.; Pinto, A.K. Impact of Obesity on Vaccination to SARS-CoV-2. Front Endocrinol (Lausanne). 2022 Jun 20; 13:898810. [CrossRef]
- Chauvin, C.; Retnakumar, S.V.; Bayry, J. Obesity negatively impacts maintenance of antibody response to COVID-19 vaccines. Cell Rep Med. 2023 Jul 18;4(7):101117. [CrossRef]
- van der Klaauw, A. A.; Horner, E. C.; Pereyra-Gerber, P.; Agrawal, U.; Foster, W. S.; et al. Accelerated waning of the humoral response to COVID-19 vaccines in obesity. Nature Med, 2023; 29(5), 1146–1154. [CrossRef]
- D'Souza, M.; Keeshan, A.; Gravel, C.A.; Langlois, M.A, Cooper CL. Obesity does not influence SARS-CoV-2 humoral vaccine immunogenicity. NPJ Vaccines. 2024 Nov 18;9(1):226. [CrossRef]
- Zwick, R.K.; Guerrero-Juarez, C.F,; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018 Jan 9;27(1):68-83. [CrossRef]
- Hagberg, C.E.; Spalding, K.L. White adipocyte dysfunction and obesity-associated pathologies in humans. Nat Rev Mol Cell Biol 2024; 25, 270–289. [CrossRef]
- Richard, A.J.; White, U.; Elks, C.M, et al. Adipose Tissue: Physiology to Metabolic Dysfunction. [Updated 2020 Apr 4]. In: Feingold KR, Anawalt B, Blackman MR, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK555602/.
- Gavin, K.M.; Bessesen D.H. Sex Differences in Adipose Tissue Function. Endocrinol Metab Clin North Am. 2020 Jun;49(2):215-228. [CrossRef]
- Luo, L.; Liu, M. Adiponectin: friend or foe in obesity and inflammation. Medical Review 2022, 2(4), 349–362. [CrossRef]
- Baldelli, S.; Aiello, G.; Mansilla Di Martino, E.; Campaci, D.; Muthanna, F.M.S.; Lombardo, M. The Role of Adipose Tissue and Nutrition in the Regulation of Adiponectin. Nutrients. 2024 Jul 26;16(15):2436. [CrossRef]
- Dare, A.; Chen, S. Y. Adipsin in the pathogenesis of cardiovascular diseases. Vascular pharmacology, 2024; 154, 107270. [CrossRef]
- Han, R.; Huang, H.; Zhu, J.; Jin, X.; Wang, Y.; Xu, Y.; Xia, Z. Adipokines and their potential impacts on susceptibility to myocardial ischemia/reperfusion injury in diabetes. Lipids health dis, 2024; 23(1), 372. [CrossRef]
- Boucher, J.; Masri, B.; Daviaud, D.; Gesta, S.; Guigné, C.; Mazzucotelli, A.; et al. Apelin, a newly identified adipokine up-regulated by insulin and obesity. Endocrinology, 2005; 146(4), 1764–1771. [CrossRef]
- Tan, L; Lu, X.; Danser, A.H.J.; Verdonk. K. The Role of Chemerin in Metabolic and Cardiovascular Disease: A Literature Review of Its Physiology and Pathology from a Nutritional Perspective. Nutrients. 2023 Jun 25;15(13):2878. [CrossRef]
- Münzberg, H.; Heymsfield, S. B.; Berthoud, H. R.; Morrison, C. D. History and future of leptin: Discovery, regulation and signaling. Metabolism: clinical and experimental, 2024; 161, 156026. [CrossRef]
- Perakakis, N.; Mantzoros, C. S. Evidence from clinical studies of leptin: current and future clinical applications in humans. Metabolism: clinical and experimental, 2024; 161, 156053. [CrossRef]
- Li, Z.; Gao, Z.; Sun, T.; Zhang, S.; Yang, S.; Zheng, M.; Shen, H. Meteorin-like/Metrnl, a novel secreted protein implicated in inflammation, immunology, and metabolism: A comprehensive review of preclinical and clinical studies. Front Immunol, 2023; 14, 1098570. [CrossRef]
- Shi, R.; He, M.; Peng, Y.; Xia, X. Homotherapy for heteropathy: Interleukin-41 and its biological functions. Immunology 2024, 173(1), 1–13. [CrossRef]
- Sena, C.M. Omentin: A Key Player in Glucose Homeostasis, Atheroprotection, and Anti-Inflammatory Potential for Cardiovascular Health in Obesity and Diabetes. Biomedicines. 2024 Jan 26;12(2):284. [CrossRef]
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N. Z. Resistin in metabolism, inflammation, and disease. The FEBS journal, 2020; 287(15), 3141–3149. [CrossRef]
- Radzik-Zając, J.; Wytrychowski, K.; Wiśniewski, A.; Barg, W. The role of the novel adipokines vaspin and omentin in chronic inflammatory diseases. Pediatr Endocrinol Diabetes Metab. 2023;29(1):48-52. [CrossRef]
- Dimova, R.; Tankova, T. The role of vaspin in the development of metabolic and glucose tolerance disorders and atherosclerosis. BioMed research international, 2015, 823481. [CrossRef]
- Adeghate, E. Visfatin: structure, function and relation to diabetes mellitus and other dysfunctions. Current Med Chem, 2008; 15(18), 1851–1862. [CrossRef]
- Wu, Y.; Ma, Y. CCL2-CCR2 signaling axis in obesity and metabolic diseases. J Cell Physiol. 2024 Apr;239(4):e31192. [CrossRef]
- Chan, P. C.; Lu, C. H.; Chien, H. C.; Tian, Y. F.; Hsieh, P. S. Adipose Tissue-Derived CCL5 Enhances Local Pro-Inflammatory Monocytic MDSCs Accumulation and Inflammation via CCR5 Receptor in High-Fat Diet-Fed Mice. Inter J Mol Sci, 2022; 23(22), 14226. [CrossRef]
- Yuan, Y.; Hu, R.; Park, J.; Xiong, S.; Wang, Z.; Qian, Y.; et al. Macrophage-derived chemokine CCL22 establishes local LN-mediated adaptive thermogenesis and energy expenditure. Science Advances, 2024; 10(26), eadn5229. [CrossRef]
- Wueest, S.; Konrad, D. The role of adipocyte-specific IL-6-type cytokine signaling in FFA and leptin release. Adipocyte. 2018;7(3):226-228. [CrossRef]
- Huang, L.Y.; Chiu, C.J.; Hsing, C.H.; Hsu Y.H. Interferon Family Cytokines in Obesity and Insulin Sensitivity. Cells. 2022 Dec 14;11(24):4041. [CrossRef]
- Sewter, C. P.; Digby, J. E.; Blows, F.; Prins, J.; O'Rahilly, S. Regulation of tumour necrosis factor-alpha release from human adipose tissue in vitro. J Endocrinol, 1999; 163(1), 33–38. [CrossRef]
- Engin, A. Reappraisal of Adipose Tissue Inflammation in Obesity. Adv Exper Med Biol, 2024; 1460, 297–327. [CrossRef]
- Ghanbari, M.; Momen Maragheh, S.; Aghazadeh, A.; Mehrjuyan, S. R.; Hussen, B. M.; Abdoli Shadbad, M.; Dastmalchi, N.; Safaralizadeh, R. Interleukin-1 in obesity-related low-grade inflammation: From molecular mechanisms to therapeutic strategies. International Immunopharmacol, 2021; 96, 107765. [CrossRef]
- Hofwimmer, K.; de Paula Souza, J.; Subramanian, N. et al. IL-1β promotes adipogenesis by directly targeting adipocyte precursors. Nat Commun 2024; 15, 7957. [CrossRef]
- Juge-Aubry, C. E.; Somm, E.; Giusti, V.; Pernin, A.; Chicheportiche, R.; Verdumo, C.; et al. Adipose tissue is a major source of interleukin-1 receptor antagonist: upregulation in obesity and inflammation. Diabetes, 2003; 52(5), 1104–1110. [CrossRef]
- Frühbeck, G; Catalán, V.; Ramírez, B.; Valentí, V.; Becerril, S.; et al. Serum Levels of IL-1 RA Increase with Obesity and Type 2 Diabetes in Relation to Adipose Tissue Dysfunction and are Reduced After Bariatric Surgery in Parallel to Adiposity. J Inflamm Res. 2022 Feb 24;15:1331-1345. [CrossRef]
- Barchetta, I.; Cimini, F.A.; Dule, S.; Cavallo, M.G. Dipeptidyl Peptidase 4 (DPP4) as A Novel Adipokine: Role in Metabolism and Fat Homeostasis. Biomedicines. 2022 Sep 16;10(9):2306. [CrossRef]
- Cuevas-Ramos, D.; Mehta, R.; Aguilar-Salinas, C. A. Fibroblast Growth Factor 21 and Browning of White Adipose Tissue. Frontiers in physiology, 2019; 10, 37. [CrossRef]
- Flores-Cortez, Y. A.; Barragán-Bonilla, M. I.; Mendoza-Bello, J. M.; González-Calixto, C.; Flores-Alfaro, E.; Espinoza-Rojo, M. Interplay of retinol binding protein 4 with obesity and associated chronic alterations. Molecular Medic rep, 2022; 26(1), 244. [CrossRef]
- Zhang, J.; Wu, Y.; Zhang, Y.; Leroith, D.; Bernlohr, D.A.; Chen, X. The Role of Lipocalin 2 in the Regulation of Inflammation in Adipocytes and Macrophages, Molecular Endocrinology, 2008; 22, (6), 1416–1426. [CrossRef]
- Moschen, A. R.; Adolph, T. E.; Gerner, R. R.; Wieser, V.; Tilg, H. Lipocalin-2: a master mediator of intestinal and metabolic inflammation. Trends Endocrinol. Metab. 2017; 28, 388–397.
- Lee, M. J. Transforming growth factor beta superfamily regulation of adipose tissue biology in obesity. Biochimica et biophysica acta. Molecular basis of disease, 2018; 1864(4 Pt A), 1160–1171. [CrossRef]
- Flegal, K.M.; Kruszon-Moran, D.; Carroll, M.D.; Fryar, C.D.; Ogden CL. Trends in obesity among adults in the United States, 2005 to 2014. JAMA. 2016;315(21):2284–2291. [CrossRef]
- Muscogiuri, G.; Verde, L.; Vetrani, C.; Barrea, L.; Savastano, S.; Colao, A. Obesity: a gender-view. J Endocrinol Invest. 2024 Feb;47(2):299-306. [CrossRef]
- Tramunt, B.; Smati, S.; Grandgeorge, N.; Lenfant, F.; Arnal, J.F.; Montagner, A, et al. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia. 2020;63(3):453–461. [CrossRef]
- Guerra, B.; Fuentes, T.; Delgado-Guerra, S.; Guadalupe-Grau, A.; Olmedillas, H.; Santana, A.; Ponce-Gonzalez, J. G.; Dorado, C.; Calbet, J. A. Gender dimorphism in skeletal muscle leptin receptors, serum leptin and insulin sensitivity. PloS one, 2008; 3(10), e3466. [CrossRef]
- Rak, A.; Mellouk, N.; Froment, P.; Dupont, J. Adiponectin and resistin: potential metabolic signals affecting hypothalamo-pituitary gonadal axis in females and males of different species. Reproduction (Cambridge, England), 2017; 153(6), R215–R226. [CrossRef]
- Sanchez-Rebordelo, E.; Cunarro, J.; Perez-Sieira, S.; Seoane, L.M.; Diéguez, C.; Nogueiras, R.; Tovar, S. Regulation of Chemerin and CMKLR1 Expression by Nutritional Status, Postnatal Development, and Gender. Int J Mol Sci. 2018 Sep 25;19(10):2905. [CrossRef]
- Kautzky-Willer, A.; Leutner, M.; Harreiter, J. Sex differences in type 2 diabetes. Diabetologia, 2023; 66(6), 986–1002. [CrossRef]
- Koceva, A.; Herman, R.; Janez, A.; Rakusa, M.; Jensterle, M. Sex- and Gender-Related Differences in Obesity: From Pathophysiological Mechanisms to Clinical Implications. Int. J. Mol. Sci. 2024, 25, 7342. [CrossRef]
- Luo, L.; Chen, L.; Song, J.; Ma, X.; Wang, X. Association between systemic immune-inflammatory index and systemic inflammatory response index with body mass index in children and adolescents: a population-based study based on the National Health and Nutrition Examination Survey 2017-2020. Front Endocrinol 2024, 15, 1426404. [CrossRef]
- Silva, J.; Iwasaki, A. Sex differences in postacute infection syndromes. Science Translational Medicine, 2024; 16(773), eado2102. [CrossRef]
- Cheng, S.; Ning, Z.; Huang, K.; Yuan, Y.; Tan, X.; Pan, Y.; et al. Analysis of sex-biased gene expression in a Eurasian admixed population. Briefings in bioinformatics, 2024; 25(5), bbae451. [CrossRef]
- Persons, P. A.; Williams, L.; Fields, H.; Mishra, S.; Mehta, R. Weight gain during midlife: Does race/ethnicity influence risk? Maturitas, 2024; 185, 108013. [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat Rev Immunol. 2016; 16:626–38. 6: 16. [CrossRef]
- Wilkinson, N.M.; Chen, H.C.; Lechner, M.G.; Su M.A. Sex differences in immunity. Annu Rev Immunol. 2022 40: 75-94. [CrossRef]
- Popotas, A.; Casimir, G. J.; Corazza, F.; Lefèvre, N. Sex-related immunity: could Toll-like receptors be the answer in acute inflammatory response? Frontiers in immunology, 2024; 15, 1379754. [CrossRef]
- Wang, P.; Yang, X.; Zhang, L.; Sha, S.; Huang, J. Peng, J.; et al. (). Tlr9 deficiency in B cells leads to obesity by promoting inflammation and gut dysbiosis. Nature comm, 2024; 15(1), 4232. [CrossRef]
- Hamerman, J. A.; Barton, G. M. The path ahead for understanding Toll-like receptor-driven systemic autoimmunity. Current opinion in immunology, 2024; 91, 102482. [CrossRef]
- Layug, P. J.; Vats, H.; Kannan, K.; Arsenio, J. Sex differences in CD8+ T cell responses during adaptive immunity. WIREs mechanisms of disease, 2024; 16(5), e1645. [CrossRef]
- Forsyth, K.S.; Jiwrajka, N.; Lovell, C.D.; Toothacre, N.E.; Anguera, M.C. The conneXion between sex and immune responses. Nat Rev Immunol. 2024 Jul;24(7):487-502. [CrossRef]
- Kirichenko, T.V.; Markina, Y.V.; Bogatyreva, A.I.; Tolstik, T.V.; Varaeva, Y.R.; Starodubova, A.V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int J Mol Sci. 2022 Nov 29;23(23):14982. [CrossRef]
- Trim, W.V.; Lynch, L. Immune and non-immune functions of adipose tissue leukocytes. Nat Rev Immunol 22, 371–386 (2022). [CrossRef]
- Xiao, L.; Yang, X.; Lin, Y.; Li, S.; Jiang, J.; Qian, S.; Tang, Q.; He, R.; Li, X. Large adipocytes function as antigen-presenting cells to activate CD4(+) T cells via upregulating MHCII in obesity. International journal of obesity 2016; (2005), 40(1), 112–120. [CrossRef]
- Chan, C.C.; Damen, M.S.M.A.; Alarcon, P.C.; Sanchez-Gurmaches, J.; Divanovic, S. Inflammation and Immunity: From an Adipocyte's Perspective. J Interferon Cytokine Res. 2019 Aug;39(8):459-471. [CrossRef]
- Castoldi, A.; Sanin, D. E.; van Teijlingen Bakker, N.; Aguiar, C. F.; de Brito Monteiro, L.; et al. Metabolic and functional remodeling of colonic macrophages in response to high-fat diet-induced obesity. iScience, 2023; 26(10), 107719. [CrossRef]
- Chen, X.; Wang, S.; Huang, Y.; Zhao, X.; Jia, X.; Meng, G.; et al. Obesity Reshapes Visceral Fat-Derived MHC I Associated-Immunopeptidomes and Generates Antigenic Peptides to Drive CD8+ T Cell Responses. iScience, 2020; 23(4), 100977. [CrossRef]
- Satoh, M.; Iizuka, M.; Majima, M.; Ohwa, C.; Hattori, A.; Van Kaer, L.; Iwabuchi, K. Adipose invariant NKT cells interact with CD1d-expressing macrophages to regulate obesity-related inflammation. Immunology, 2022; 165(4), 414–427. [CrossRef]
- Satoh, M.; Iwabuchi, K. Contribution of NKT cells and CD1d-expressing cells in obesity-associated adipose tissue inflammation. Frontiers in immunology, 2024; 15, 1365843. [CrossRef]
- Andersen, C.J.; Murphy, K.E.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv Nutr. 2016 Jan 15;7(1):66-75. [CrossRef]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021 Mar 1;320(3):C375-C391. [CrossRef]
- Valentine, Y.; Nikolajczyk, B. S. T cells in obesity-associated inflammation: The devil is in the details. Immunological reviews, 2024; 324(1), 25–41. [CrossRef]
- Ding, S.; Lund, PK. Role of intestinal inflammation as an early event in obesity and insulin resistance. Curr Opin Clin Nutr Metab Care. 2011 Jul;14(4):328-33. [CrossRef]
- Brotfain, E.; Hadad, N.; Shapira, Y.; Avinoah, E.; Zlotnik, A.; Raichel, L.; Levy, R. Neutrophil functions in morbidly obese subjects. Clin Exp Immunol. 2015 Jul;181(1):156-63. [CrossRef]
- Gomez-Casado, G.; Jimenez-Gonzalez, A.; Rodriguez-Muñoz, A.; Tinahones, F.J.; González-Mesa, E.; Murri, M.; Ortega-Gomez, A. Neutrophils as indicators of obesity-associated inflammation: A systematic review and meta-analysis. Obes Rev. 2024 Nov 29:e13868. [CrossRef]
- Shantaram, D.; Hoyd, R.; Blaszczak, A. M.; Antwi, L.; Jalilvand, A.; Wright, V. P.; et al. Obesity-associated microbiomes instigate visceral adipose tissue inflammation by recruitment of distinct neutrophils. Nature Comm 2024, 15(1), 5434. [CrossRef]
- Hu, Y.; Chakarov, S. Eosinophils in obesity and obesity-associated disorders. Discov Immunol. 2023 Nov 14;2(1):kyad022. [CrossRef]
- Divoux, A.; Moutel, S.; Poitou, C.; Lacasa, D.; Veyrie. N.; Aissat, A.; Arock, M.; Guerre-Millo, M.; Clément, K. Mast cells in human adipose tissue: link with morbid obesity, inflammatory status, and diabetes. J Clin Endocrinol Metab. 2012 Sep;97(9): E1677-85. [CrossRef]
- Mukherjee, S.; Skrede, S.; Haugstøyl, M.; López, M.; Fernø, J. Peripheral and central macrophages in obesity. Frontiers in endocrinology, 2023; 14, 1232171. [CrossRef]
- Wilkin, C.; Piette, J.; Legrand-Poels, S. Unravelling metabolic factors impacting iNKT cell biology in obesity. Biochem Pharmacol. 2024 Oct; 228:116436. [CrossRef]
- Cui, G.; Abe, S.; Kato, R.; Ikuta, K. Insights into the heterogeneity of iNKT cells: tissue-resident and circulating subsets shaped by local microenvironmental cues. Front Immunol. 2024 Feb 19;15:1349184. [CrossRef]
- Canter, R.J.; Judge, S.J.; Collins, C.P.; Yoon, D.J.; Murphy, W.J. Suppressive effects of obesity on NK cells: is it time to incorporate obesity as a clinical variable for NK cell-based cancer immunotherapy regimens? J Immunother Cancer. 2024 Mar 13;12(3):e008443. [CrossRef]
- De Barra, C.; O'Shea, D.; Hogan, A. E. NK cells vs. obesity: A tale of dysfunction & redemption. Clinical immunology 2023, 255, 109744. [CrossRef]
- Goldberg, E.L.; Shchukina, I.; Asher, J.L.; Sidorov, S.; Artyomov, M.N.; Dixit, V.D. Ketogenesis activates metabolically protective γδ T cells in visceral adipose tissue. Nat Metab. 2020 Jan;2(1):50-61. [CrossRef]
- Frasca, D.; Romero, M.; Blomberg, B.B. Similarities in B Cell Defects between Aging and Obesity. J Immunol. 2024 Nov 15;213(10):1407-1413. [CrossRef] [PubMed]
- Gao, F.; Litchfield, B.; Wu, H. Adipose tissue lymphocytes and obesity. J Cardiovasc Aging. 2024 Jan;4(1):5. [CrossRef]
- Meher, A.K.; McNamara, C.A. B-1 lymphocytes in adipose tissue as innate modulators of inflammation linked to cardiometabolic disease. Immunol Rev. 2024 Jul;324(1):95-103. [CrossRef]
- Liu, R.; Nikolajczyk, B. S. Tissue Immune Cells Fuel Obesity-Associated Inflammation in Adipose Tissue and Beyond. Frontiers in immunology, 2019; 10, 1587. [CrossRef]
- McLaughlin, T.; Liu, L. F.; Lamendola, C.; Shen, L.; Morton, J.; Rivas, H.; et al. T-cell profile in adipose tissue is associated with insulin resistance and systemic inflammation in humans. Arteriosclerosis, thrombosis, and vascular biology, 2014; 34(12), 2637–2643. [CrossRef]
- Zi, C.; Wang, D.; Gao, Y.; He, L. The role of Th17 cells in endocrine organs: Involvement of the gut, adipose tissue, liver and bone. Front in Immunol , 2023; 13, 1104943. [CrossRef]
- Kochumon, S.; Hasan, A.; Al-Rashed, F.; Sindhu, S.; Thomas, R.; Jacob, T.; et al. Increased Adipose Tissue Expression of IL-23 Associates with Inflammatory Markers in People with High LDL Cholesterol. Cells, 2022; 11(19), 3072. [CrossRef]
- Fabbrini, E.; Cella, M.; McCartney, S.A.; Fuchs, A.; Abumrad, N.A.; et al. Association between specific adipose tissue CD4+ T-cell populations and insulin resistance in obese individuals. Gastroenterology. 2013 Aug;145(2):366-74.e1-3. [CrossRef]
- Wang, Q.; Wang, Y.; Xu, D. The roles of T cells in obese adipose tissue inflammation. Adipocyte, 2021; 10(1), 435–445. [CrossRef]
- Delacher, M.; Schmidleithner, L.; Simon, M.; Stüve, P.; Sanderink, L.; Hotz-Wagenblatt, A.; et al. The effector program of human CD8 T cells supports tissue remodeling. J Exp Med, 2024; 221(2), e20230488. [CrossRef]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; et al. Mucosal-associated invariant T cell alterations in obese and type 2 diabetic patients. J Clin Invest, 2015; 125(4), 1752–1762. [CrossRef]
- Kedia-Mehta, N.; Hogan, A. E. MAITabolism2 - the emerging understanding of MAIT cell metabolism and their role in metabolic disease. Front Immunol 2022; 13, 1108071. [CrossRef]
- Sage, P. T.; Sharpe, A. H. T follicular regulatory cells in the regulation of B cell responses. Trends in immunology, 2015; 36(7), 410–418. [CrossRef]
- Hildreth, A. D.; Ma, F.; Wong, Y. Y.; Sun, R.; Pellegrini, M.; O'Sullivan, T. E. Single-cell sequencing of human white adipose tissue identifies new cell states in health and obesity. Nature immunology, 2021; 22(5), 639–653. [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Vazquez, T.; Blomberg, B. B. Obesity induces pro-inflammatory B cells and impairs B cell function in old mice. Mechanisms of ageing and development, 2017; 162, 91–99. [CrossRef]
- Park, M. J.; Kwok, S. K.; Lee, S. H.; Kim, E. K.; Park, S. H.; Cho, M. L. Adipose tissue-derived mesenchymal stem cells induce expansion of interleukin-10-producing regulatory B cells and ameliorate autoimmunity in a murine model of systemic lupus erythematosus. Cell transplantation, 2015; 24(11), 2367–2377. [CrossRef]
- Hong, C.; Li, X.; Zhang, K.; Huang, Q.; Li, B.; Xin, H.; et al. Novel perspectives on autophagy-oxidative stress-inflammation axis in the orchestration of adipogenesis. Front Endocrinol, 2024, 15, 1404697. [CrossRef]
- Yang, Q.; Zhang, F.; Chen, H.; Hu, Y.; Yang, N.; Yang, W.; Wang, J.; Yang, Y.; Xu, R.; Xu, C. The differentiation courses of the Tfh cells: a new perspective on autoimmune disease pathogenesis and treatment. Bioscience reports, 2024; 44(1), BSR20231723. [CrossRef]
- Zhang, Y.; Chua, S. Jr. Leptin Function and Regulation. Compr Physiol. 2017 Dec 12;8(1):351-369. 3: 12;8(1). [CrossRef]
- Kiernan, K.; MacIver, N. J. The Role of the Adipokine Leptin in Immune Cell Function in Health and Disease. Front Immunol 2021, 11, 622468. [CrossRef]
- Deng, J.; Chen, Q.; Chen, Z.; Liang, K.; Gao, X.; Wang, X.; et al. The metabolic hormone leptin promotes the function of TFH cells and supports vaccine responses. Nat Commun. 2021 May 24;12(1):3073. [CrossRef]
- Park, J.; Sohn, J.H.; Han, S.M.; Park, Y.J.; Huh, J.Y.; Choe, S.S.; Kim, J.B. Adipocytes Are the Control Tower That Manages Adipose Tissue Immunity by Regulating Lipid Metabolism. Front Immunol. 2021 Jan 28; 11:598566. [CrossRef]
- Shaikh, S.R.; Beck, M.A.; Alwarawrah, Y.; MacIver NJ. Emerging mechanisms of obesity-associated immune dysfunction. Nat Rev Endocrinol 2024; 20, 136–148. [CrossRef]
- Soták, M.; Clark, M.; Suur, B.E.; Börgeson, E. Inflammation and resolution in obesity. Nat Rev Endocrinol 2025; 21, 45–61. [CrossRef]
- Lee, M.J.; Kim, J. The pathophysiology of visceral adipose tissues in cardiometabolic diseases. Biochem Pharmacol. 2024 Apr;222:116116. [CrossRef]
- McTavish, P.V.; Mutch, D.M.. Omega-3 fatty acid regulation of lipoprotein lipase and FAT/CD36 and its impact on white adipose tissue lipid uptake. Lipids Health Dis. 2024 Nov 20;23(1):386. [CrossRef]
- Lima, G.B.; Figueiredo, N.; Kattah, F.M.; Oliveira, E.S.; Horst, M.A.; Dâmaso, A.R.; et al. Serum Fatty Acids and Inflammatory Patterns in Severe Obesity: A Preliminary Investigation in Women. Biomedicines. 2024 Oct 3;12(10):2248. [CrossRef]
- Childs, B.G.; Gluscevic, M.; Baker, D.J.; Laberge, R.M.; Marquess, D.; Dananberg, J.; van Deursen, J.M. Senescent cells: an emerging target for diseases of ageing. Nat Rev Drug Discov. 2017 Oct;16(10):718-735. [CrossRef]
- Liu, Z.; Liang, Q.; Ren, Y.; Guo, C.; Ge, X.; Wang, L, et al. Immunosenescence: molecular mechanisms and diseases. Signal Transduct Target. 2023;8(1):200. [CrossRef]
- Shirakawa, K.; Sano, M. T Cell Immunosenescence in Aging, Obesity, and Cardiovascular Disease. Cells. 2021 Sep 15;10(9):2435. [CrossRef]
- Wang, Y.; Dong, C.; Han, Y.; Gu, Z.; Sun, C. Immunosenescence, aging and successful aging. Front Immunol. 2022 Aug 2;13:942796. [CrossRef]
- Shimi, G.; Sohouli, M.H.; Ghorbani, A.; Shakery, A.; Zand, H. The interplay between obesity, immunosenescence, and insulin resistance. Immun Ageing. 2024 Feb 5;21(1):13. [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Garcia, D.; Blomberg, B.B. B Cell Immunosenescence. Annu Rev Cell Dev Biol. 2020 Oct 6;36:551-574. [CrossRef]
- Garmendia, J.V.; Moreno, D.; Garcia, A.H.; De Sanctis, J.B. Metabolic syndrome and asthma. Recent Pat Endocr Metab Immune Drug Discov. 2014 Jan;8(1):60-6. [CrossRef]
- Kudlova, N.; De Sanctis, J.B.; Hajduch, M. Cellular Senescence: Molecular Targets, Biomarkers, and Senolytic Drugs. Int J Mol Sci. 2022 Apr 10;23(8):4168. [CrossRef]
- Valentino, T.R.; Chen, N.; Makhijani, P.; Khan, S.; Winer, S.; Revelo, X.S.; Winer, DA. The role of autoantibodies in bridging obesity, aging, and immunosenescence. Immun Ageing. 2024 Nov 30;21(1):85. [CrossRef]
- Zhou, Z.; Tao, Y.; Zhao, H.; Wang, Q. Adipose Extracellular Vesicles: Messengers from and to Macrophages in Regulating Immunometabolic Homeostasis or Disorders. Front Immunol. 2021 May 24; 12:666344. [CrossRef]
- Kwan, H.Y.; Chen, M.; Xu, K.; Chen, B. The impact of obesity on adipocyte-derived extracellular vesicles. Cell Mol Life Sci. 2021 Dec;78(23):7275-7288. [CrossRef]
- Matilainen, J.; Berg, V.; Vaittinen, M.; Impola, U.; Mustonen, A. M.; Männistö, V.; et al. Increased secretion of adipocyte-derived extracellular vesicles is associated with adipose tissue inflammation and the mobilization of excess lipid in human obesity. Journal of Translational Medicine 2024, 22(1), 623. [CrossRef]
- Rakib, A.; Kiran, S.; Mandal, M.; Singh, U. P. (). MicroRNAs: a crossroad that connects obesity to immunity and aging. Immunity & ageing 2022; 19(1), 64. [CrossRef]
- Mendivil-Alvarado, H.; Sosa-León, L.A.; Carvajal-Millan, E.; Astiazaran-Garcia, H. Malnutrition and Biomarkers: A Journey through Extracellular Vesicles. Nutrients. 2022 Feb 27;14(5):1002. [CrossRef]
- Leocádio, P.C.L.; Oriá, R.B.; Crespo-Lopez, M.E.; Alvarez-Leite, J.I. Obesity: More Than an Inflammatory, an Infectious Disease? Front Immunol. 2020 Jan 14; 10:3092. [CrossRef]
- Pugliese, G.; Liccardi, A.; Graziadio, C.; Barrea, L.; Muscogiuri, G.; Colao, A Obesity and infectious diseases: pathophysiology and epidemiology of a double pandemic condition. Int J Obes 2022; 46, 449–465. [CrossRef]
- Cristancho, C.; Mogensen, K.M.; Robinson, M.K. Malnutrition in patients with obesity: An overview perspective. Nutr Clin Pract. 2024 Dec;39(6):1300-1316. [CrossRef]
- Crespo, F.I.; Mayora, S.J.; De Sanctis, J.B.; Martínez, W.Y.; Zabaleta-Lanz, M.E.; Toro, F.I.; Deibis, L.H.; García A.H. SARS-CoV-2 Infection in Venezuelan Pediatric Patients-A Single Center Prospective Observational Study. Biomedicines. 2023 May 9;11(5):1409. [CrossRef]
- García, A.H.; Crespo, F.I.; Mayora, S.J.; Martinez, W.Y.; Belisario, I.; Medina, C.; De Sanctis, J.B. Role of Micronutrients in the Response to SARS-CoV-2 Infection in Pediatric Patients. Immuno 2024, 4, 211-225. [CrossRef]
- Cordeiro, A.; Luna, M.; Pereira, S. E.; Saboya, C. J.; Ramalho, A. Impairment of Vitamin D Nutritional Status and Metabolic Profile Are Associated with Worsening of Obesity According to the Edmonton Obesity Staging System. Intl J Mol Sci, 2022; 23(23), 14705. [CrossRef]
- Bennour, I.; Haroun, N.; Sicard, F.; Mounien, L.; Landrier, J.-F. Vitamin D and Obesity/Adiposity—A Brief Overview of Recent Studies. Nutrients 2022, 14, 2049. [CrossRef]
- Keto, J.; Feuth, T.; Linna, M.; Saaresranta, T. Lower respiratory tract infections among newly diagnosed sleep apnea patients. BMC Pulm Med. 2023 Sep 8;23(1):332. [CrossRef]
- Alvarez, J.A.; Yang, C.A.; Ojuri, V.; Buckley, K.; Bedi, B.; Musonge-Effoe, J.; Soibi-Harry, A.; La-hiri, C.D. Sex Differences in Metabolic Disorders of Aging and Obesity in People with HIV. Curr HIV/AIDS Rep. 2024 Nov 21;22(1):3. [CrossRef]
- Cancelier, A.C.L.; Schuelter-Trevisol, F.; Trevisol, D.J.; Atkinson, R.L. Adenovirus 36 infection and obesity risk: current understanding and future therapeutic strategies. Expert Rev Endocrinol Metab. 2022 Mar;17(2):143-152. [CrossRef]
- Hameed, M.; Geerling, E.; Pinto, A.K.; Miraj, I.; Weger-Lucarelli, J. Immune response to arbovirus infection in obesity. Front Immunol. 2022 Nov 18; 13:968582. [CrossRef]
- Tian, Y.; Jennings, J.; Gong, Y.; Sang, Y. Viral Infections and Interferons in the Development of Obesity. Biomolecules. 2019 Nov 12;9(11):726. [CrossRef]
- Gallagher, P.; Chan, K.R.; Rivino, L.; Yacoub, S. The association of obesity and severe dengue: possible pathophysiological mechanisms. J Infect. 2020 Jul;81(1):10-16. [CrossRef]
- Chen, C.Y.; Chiu, Y.Y.; Chen, Y.C.; Huang, C.H.; Wang, W.H.; Chen, Y.H.; Lin C.Y. Obesity as a clinical predictor for severe manifestation of dengue: a systematic review and meta-analysis. BMC Infect Dis. 2023 Jul 31;23(1):502. [CrossRef]
- Molokwu, J. C.; Penaranda, E.; Lopez, D. S.; Dwivedi, A.; Dodoo, C.; & Shokar, N. Association of Metabolic Syndrome and Human Papillomavirus Infection in Men and Women Residing in the United States. Cancer epidemiology, biomarkers & prevention, 2017; 26(8), 1321–1327. [CrossRef]
- Huang, X.; Zhao, Q.; Yang, P.; Li, Y.; Yuan, H.; Wu, L.; Chen, Z. Metabolic Syndrome and Risk of Cervical Human Papillomavirus Incident and Persistent Infection. Medicine, 2016; 95(9), e2905. [CrossRef]
- Shin, H.S.; Jun, B.G.; Yi, S.W. Impact of diabetes, obesity, and dyslipidemia on the risk of hepatocellular carcinoma in patients with chronic liver diseases. Clin Mol Hepatol. 2022 Oct;28(4):773-789. [CrossRef]
- Markakis, K.; Tsachouridou, O.; Georgianou, E.; Pilalas, D.; Nanoudis, S.; Metallidis, S. Weight Gain in HIV Adults Receiving Antiretroviral Treatment: Current Knowledge and Future Perspectives. Life (Basel, Switzerland), 2024; 14(11), 1367. [CrossRef]
- Cáceres, C.; Castillo, M.; Carrillo, K.; Tapia, C.V.; Valderrama, G.; Maquilón, C.; Toro-Ascuy, D.; Zorondo-Rodríguez, F.; Fuenzalida, L.F. Overnutrition as a risk factor for more serious respiratory viral infections in children: A retrospective study in hospitalized patients. Endocrinol Diabetes Nutr (Engl Ed). 2023 Aug-Sep;70(7):476-483. [CrossRef]
- Ramaswamy, M.; Shi, L.; Monick, M. M.; Hunninghake, G. W.; Look, D. C. Specific inhibition of type I interferon signal transduction by respiratory syncytial virus. American Journal of Respiratory Cell And Molecular Biology, 2004; 30(6), 893–900. [CrossRef]
- Mîndru, D. E.; Țarcă, E.; Adumitrăchioaiei, H.; Anton-Păduraru, D. T.; Ștreangă, V.; Frăsinariu, O. E.; Sidoreac, A.; Stoica, C.; Bernic, V.; Luca, A. C. Obesity as a Risk Factor for the Severity of COVID-19 in Pediatric Patients: Possible Mechanisms-A Narrative Review. Children (Basel, Switzerland), 2024; 11(10), 1203. [CrossRef]
- Jang, S.; Hong, W.; Moon, Y. Obesity-compromised immunity in post-COVID-19 condition: a critical control point of chronicity. Front Immunol, 2024; 15, 1433531. [CrossRef]
- Miron, V. D.; Drăgănescu, A. C.; Pițigoi, D.; Aramă, V.; Streinu-Cercel, A.; Săndulescu, O. The Impact of Obesity on the Host-Pathogen Interaction with Influenza Viruses - Novel Insights: Narrative Review. Diabetes, metabolic syndrome and obesity: targets and therapy, 2024; 17, 769–777. [CrossRef]
- Chiang, C.H.; Huang, KC. Association between metabolic factors and chronic hepatitis B virus infection. World J Gastroenterol. 2014 Jun 21;20(23):7213-6. [CrossRef]
- Hornung, F.; Rogal, J.; Loskill, P.; Löffler, B.; Deinhardt-Emmer, S. The Inflammatory Profile of Obesity and the Role on Pulmonary Bacterial and Viral Infections. Int J Mol Sci. 2021 Mar 26;22(7):3456. [CrossRef]
- Hales, C.; Burnet, L.; Coombs, M.; Collins, A. M.; Ferreira, D. M. Obesity, leptin and host defence of Streptococcus pneumoniae: the case for more human research. European Respiratory Review, 2022; 231(165), 220055. [CrossRef]
- Li, C.; Huang, H.; Xia, Q.; Zhang, L. Correlation between body mass index and gender-specific 28-day mortality in patients with sepsis: a retrospective cohort study. Front Med (Lausanne). 2024 Oct 8;11:1462637. [CrossRef]
- Weber, D.J.; Rutala, W.A.; Samsa, G.P.; Santimaw, J.E.; Lemon, S.M. Obesity as a predictor of poor antibody response to hepatitis B plasma vaccine. JAMA. 1985 Dec 13;254(22):3187-9.
- Callahan, S.T.; Wolff, M.; Hill, H.R.; Edwards, K.M.; NIAID Vaccine and Treatment Evaluation Unit (VTEU) Pandemic H1N1 Vaccine Study Group. Impact of body mass index on immunogenicity of pandemic H1N1 vaccine in children and adults. J Infect Dis. 2014 Oct 15;210(8):1270-4. [CrossRef]
- Clarke, M.; Mathew, S.M.; Giles, L.C.; Pena, A.S.; Barr, I.G.; Richmond, P.C.; Marshall, H.S. A Prospective Study Investigating the Impact of Obesity on the Immune Response to the Quadrivalent Influenza Vaccine in Children and Adolescents. Vaccines (Basel). 2022 Apr 29;10(5):699. [CrossRef]
- Sheridan, P.A.; Paich, H.A.; Handy, J.; Karlsson, E.A.; Hudgens, M.G.; Sammon, A.B.; Holland, L.A.; Weir, S; Noah, T.L.; Beck, M.A. Obesity is associated with impaired immune response to influenza vaccination in humans. Int J Obes 2012. 36:1072–1077. [CrossRef]
- Huang, J.Y.; Kaur, B.P.; Seth, D.; Pansare, M.V.; Kamat, D.; McGrath, E, et al Can Obesity Alter the Immune Response to Childhood Vaccinations? Journal of Allergy and Clinical Immunology, 2019, Volume 143, Issue 2, AB299.
- Huang, J.; Kaur, B.; Farooqi, A.; Miah, T.; McGrath, E.; et al. Elevated Glycated Hemoglobin Is Associated with Reduced Antibody Responses to Vaccinations in Children. Pediatr Allergy Immunol Pulmonol. 2020 Dec;33(4):193-198. [CrossRef]
- Xue, P.; Merikanto, I.; Delale, E. A.; Bjelajac, A.; Yordanova, J.; Chan, R. N. Y.; et al. Associations between obesity, a composite risk score for probable long COVID, and sleep problems in SARS-CoV-2 vaccinated individuals. International journal of obesity 2024; (2005), 48(9), 1300–1306. [CrossRef]
- Ou, X.; Jiang, J.; Lin, B.; Liu, Q.; Lin, W.; Chen, G.; Wen, J. Antibody responses to COVID-19 vaccination in people with obesity: A systematic review and meta-analysis. Influenza Other Respir Viruses. 2023 Jan;17(1): e13078. [CrossRef]
- Faizo, A.A.; Qashqari, F.S.; El-Kafrawy, S.A.; Barasheed, O.; Almashjary, M.N.; Alfelali, M.; Bawazir, A.A.; et al. A potential association between obesity and reduced effectiveness of COVID-19 vaccine-induced neutralizing humoral immunity. J Med Virol. 2023 Jan;95(1): e28130. [CrossRef]
- Dumrisilp, T.; Wongpiyabovorn, J.; Buranapraditkun, S.; Tubjaroen, C.; Chaijitraruch, N.; Prachuapthunyachart, S.; Sintusek, P.; Chongsrisawat, V. Impact of Obesity and Being Overweight on the Immunogenicity to Live Attenuated Hepatitis A Vaccine in Children and Young Adults. Vaccines (Basel). 2021 Feb 6;9(2):130. [CrossRef]
- Soponkanabhorn, T.; Suratannon, N.; Buranapraditkun, S.; Tubjareon, C.; Prachuapthunyachart, S.; Eiamkulbutr, S.; Chongsrisawat, V. Cellular immune response to a single dose of live attenuated hepatitis a virus vaccine in obese children and adolescents. Heliyon. 2024 Aug 20;10(16):e36610. [CrossRef]
- Fonzo, M.; Nicolli, A.; Maso, S.; Carrer, L.; Trevisan, A.; Bertoncello, C. Body Mass Index and Antibody Persistence after Measles, Mumps, Rubella and Hepatitis B Vaccinations. Vaccines (Basel). 2022 Jul 20;10(7):1152. [CrossRef]
- Kara, Z.; Akçin, R.; Demir, A. N.; Dinç, H. Ö.; Taşkın, H. E.; Kocazeybek, B.; Yumuk, V. D. Antibody Response to SARS-CoV-2 Vaccines in People with Severe Obesity. Obesity surgery, 2022, 32(9), 2987–2993. [CrossRef]
- Drożdżyńska, J.; Jakubowska, W.; Kemuś, M.; Krokowska, M.; Karpezo, K.; Wiśniewska, M.; Bogdański, P.; Skrypnik, D. SARS-CoV-2 and Influenza Vaccines in People with Excessive Body Mass-A Narrative Review. Life (Basel, Switzerland), 2022; 12(10), 1617. [CrossRef]
- Frasca, D.; Romero, M.; Diaz, A.; Blomberg, B. B. Obesity accelerates age defects in B cells, and weight loss improves B cell function. Immunity & ageing: I & A, 2023; 20(1), 35. [CrossRef]
- García, A.; De Sanctis, J.B. An overview of adjuvant formulations and delivery systems. APMIS. 2014 Apr;122(4):257-67. [CrossRef]
- White, S.J.; Taylor, M.J.; Hurt, R.T.; Jensen, M.D.; Poland, G.A. Leptin-based adjuvants: an innovative approach to improve vaccine response. Vaccine. 2013 Mar 25;31(13):1666-72. [CrossRef]
- Ben Nasr, M.; Usuelli, V.; Dellepiane, S.; Seelam, A.J.; Fiorentino, T.V.; D'Addio, F.; et al. Glucagon-like peptide 1 receptor is a T cell-negative costimulatory molecule. Cell Metab. 2024 Jun 4;36(6):1302-1319.e12. [CrossRef]
- Garmendia, J. V.; García, A. H.; De Sanctis, C. V.; Hajdúch, M.; De Sanctis, J. B. Autoimmunity and Immunodeficiency in Severe SARS-CoV-2 Infection and Prolonged COVID-19. Current Issues In Molecular Biology, 2022; 45(1), 33–50. [CrossRef]
- García, A. H.; De Sanctis, J. B. Exploring the Contrasts and Similarities of Dengue and SARS-CoV-2 Infections During the COVID-19 Era. Inter J Mol Sci, 2024; 25(21), 11624. [CrossRef]
Figure 1.
The figure represents the characteristics of the different adipose cells. The white adipose cells contain the highest amount of stored lipids and are low responders to stimuli; the brown adipose cells are involved in thermogenesis to control body temperature, and the beige adipose cells also serve as lipid deposits but control temperature upon stimulation. Beige adipose cells are found along white adipose cells in the adipose tissue. Cell comp refers to organelle cell composition. The white circles refer to lipid deposits, the blue circles refer to mitochondria, and the black circles refer to the nucleus.
Figure 1.
The figure represents the characteristics of the different adipose cells. The white adipose cells contain the highest amount of stored lipids and are low responders to stimuli; the brown adipose cells are involved in thermogenesis to control body temperature, and the beige adipose cells also serve as lipid deposits but control temperature upon stimulation. Beige adipose cells are found along white adipose cells in the adipose tissue. Cell comp refers to organelle cell composition. The white circles refer to lipid deposits, the blue circles refer to mitochondria, and the black circles refer to the nucleus.
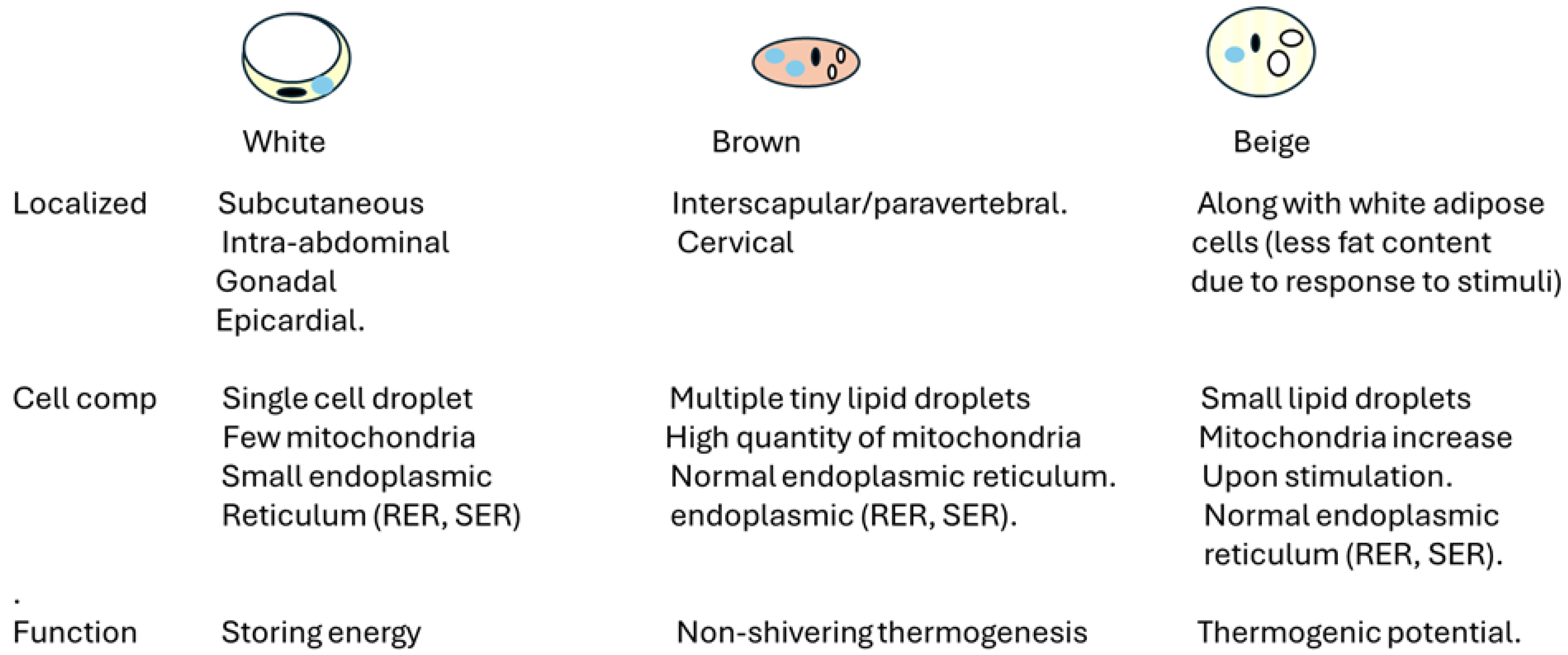
Figure 2.
illustrates the distinct differences between lean and obese adipose tissue. The obese tissue is characterized by the loss of a tolerogenic environment and the infiltration of immune cells. Additionally, it highlights the impact of sex hormones on the localization, metabolic processes, and inflammatory responses associated with these tissues.
Figure 2.
illustrates the distinct differences between lean and obese adipose tissue. The obese tissue is characterized by the loss of a tolerogenic environment and the infiltration of immune cells. Additionally, it highlights the impact of sex hormones on the localization, metabolic processes, and inflammatory responses associated with these tissues.
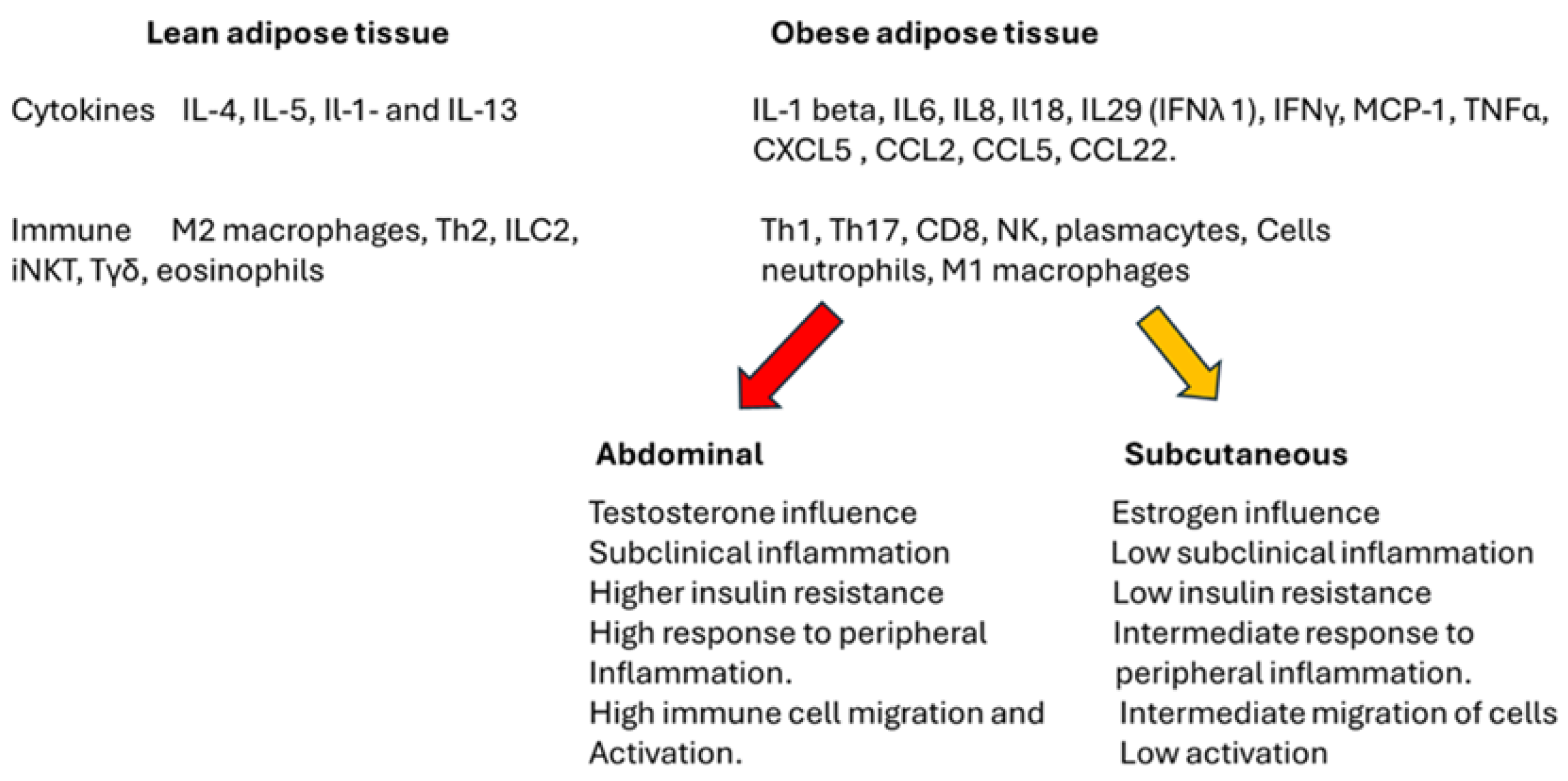
Figure 3.
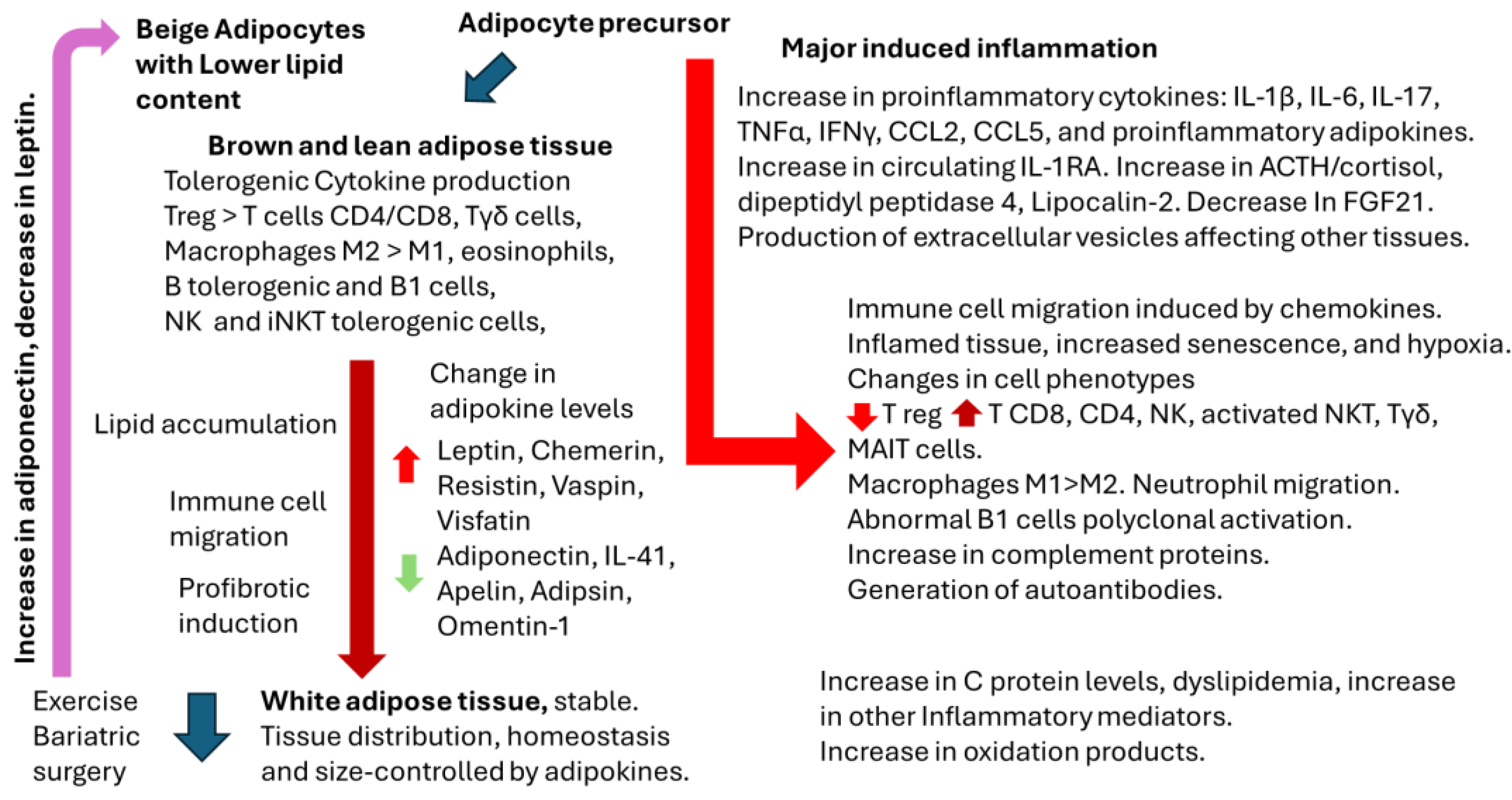

Table 1.
Adipokines and their role in inflammatory response.
| Adipokine | Pro-inflammatory | Anti-inflammatory | Reference |
| Adiponectin | No | Yes | [12,13] |
| Adipsin (complement factor-D) | No | Yes | [14,15] |
| Apelin | No | Yes | [16] |
| Chemerin | Yes | No | [17] |
| Leptin | Yes | Yes | [18,19] |
| Meteorin like (IL41) | No | Yes | [20,21] |
| Omentin-1 | No | Yes | [22] |
| Resistin | Yes | No | [23] |
| Vaspin | Yes | Yes | [24,25] |
| Visfatin | Yes | No | [26] |
Table 2.
Other cytokines and factors involved in adipose tissue responses.
| Effect | References | |
| CCL2 (MCP-1) | Monocyte migration to adipose tissue | [27] |
| CCL5 | Monocyte migration to adipose tissue | [28] |
| CCL22 | Thermogenesis induction. | [29] |
| IL-6 | Local activation of immune cells. Metabolic dysregulation. | [30] |
| IFN | IFNα induces apoptosis in adipocytes. IFNβ regulates metabolism. IFNγ proinflammatory response, reduction of adipose tissue. IFNλ1 enhances inflammatory response. IFNτ reduces inflammatory response. |
[31] |
| TNFα | Activation of tissue immune cells. Metabolic dysregulation. | [32,33] |
| IL-1 and IL-RA | IL-1 α hypertrophy of white adipose tissue IL-1 β promotes adipogenesis in murine and human adipose-derived stem cells. IL-RA is upregulated in white adipose tissue, and high circulating levels in obesity |
[34,35,36,37] |
| Dipeptidyl peptidase 4 | It plays a role in metabolic homeostasis and inflammatory response. Inhibition of the enzyme plus metformin induces a significant decrease in visceral adipose tissue. | [38] |
| Fibroblast growth factor 21 | Anti-inflammatory. | [39] |
| Retinol binding protein 4 | Induction of inflammatory response. Inhibition of insulin signaling. | [40] |
| Lipocalin-2 | Produced by white adipocytes. Increase adipose tissue. Involved in neutrophil chemoattraction | [41,42] |
| TGFβ | Involved in tissue fibrosis and insulin resistance. | [43] |
Table 3.
Summary of the effects of immune cells and mesenchymal stem cells on adipose tissue.
| Cell type | Effect | Reference |
| Neutrophils | Retain phagocytic activity, increase basal superoxide, and chemotaxis. Absolute neutrophil counts and neutrophil to lymphocyte ratio may indicate adipose tissue inflammation. Relationship of microbiota with neutrophil infiltration in adipose tissue. |
[75,76,77] |
| Eosinophils | Eosinophils protect adipose tissue inflammation. | [78] |
| Mast cells | Mast cells are activated in human adipose tissue and localized preferentially in fibrosis depots. | [79] |
| Macrophages | In lean tissue M2 macrophages and M1 in inflammatory tissues. | [80] |
| iNKT cells | Present in lean adipose tissue can be activated by CD1 and can incorporate lipids, generating a local inflammatory response | [69,70,82] |
| NK | Present in adipose tissue. Tolerogenic response in adipose tissue? Different responses in gender. | [83,84] |
| Tγδ | Inhibit inflammatory response | [85] |
| B cells | Dysfunctional B cells in obese individuals. The lean adipose tissue contains B regulatory and B1 cells. The B1 produces IgM antibodies for primary innate immunity. B2 cells usually generate protective antibodies in lymphoid organs. However, they participate in local inflammation and promote insulin resistance after migrating to white adipose tissue. |
[86,87,88] |
| Th1 cells | Promote obesity-associated inflammation. | [87,89] |
| Th2 | Stabilize adipose tissue and induce M2 polarization. The decrease in Th2 cells in the tissue is due to increased local IFNγ and inflammation. | [87,90] |
| Th17 | Proinflammatory role. Related to IL-23 secretion in adipose tissue |
[91,92] |
| Th22 | IL-22 is produced by innate lymphocyte cells upon tissue inflammation. It is related to insulin resistance. | [93] |
| CD8 cells | Cytotoxic response. Adipose tissue inflammation. Tissue remodeling. | [94,95] |
| Mucosal-associated invariant T (MAIT) cells | Secretes IL-17, inducing local tissue inflammation. | [96,97] |
| T follicular cells (TF). TFh Helper and TFreg Regulatory. | Modulate the response of B cells in adipose tissue. Impairment of TFregulatory cells is related to autoimmunity. | [98,99] |
| Follicular B cells | In adipose tissue, it induces inflammation depending on the cytokine milieu. Mesenchymal adipose stem cells induce expansion of IL-10-producing B cells—possible role in autoimmunity. |
[100] |
| Mesenchymal stem cells | Anti-inflammatory in the presence of Treg and Th2 milieu. Pro-inflammatory in the presence of inflammatory cytokines. | [101,102] |
Table 4.
Viral infection, adipose tissue involvement, and IFN responses.
| Virus | Adipose tissue involvement | IFN responses | Reference |
| Adenoviruses |
Yes |
Suppression. Chronic infection. Obesity-induced viral infection? | [135] |
| Arboviruses | Yes | Suppression. Chronic infection. | [136] |
| Herpesviridae | Yes | HSV-1 suppression through miRNA CMV-multiple antagonistic mechanisms |
[137] |
| Slow Virus (Prion) | Yes |
Inhibition of IFN signaling | [137] |
| Dengue | Yes | Inhibition of INF signaling | [138,139] |
| Papillomavirus | Yes | IFN signaling decreased | [140,141] |
| HCV | Yes | Antagonism of IFN signaling. Chronicity | [142] |
| HIV | Yes | Antagonism of IFN signaling. Chronicity | [143] |
| RSV | Yes | Virus inhibits IFN signaling | [144,145] |
| Coronavirus | Yes | IFN signaling is inhibited | [146,147] |
| Influenza | Yes | IFN signaling is inhibited | [148] |
| Hepatitis B virus | Yes | IFN response impaired | [149] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



