
Submitted:
17 December 2024
Posted:
19 December 2024
You are already at the latest version
Abstract
Advances in sequencing techniques have revealed extreme chromosomal diversity andunderscored the importance of chromoanagenesis in macroevolution, speciation, and the contextof cancer and tumor progression. Evolution is essential for adapting to the environment, preparingfor future pressures, ensuring survival, and generating diversity. Here we discuss some events likechromothripsis and other chromosomal rearrangements, emphasizing their roles as driving forcesin genomic evolution from cancer to speciation.
Keywords:
chromoanagenesis
; chromothripsis
; chromosomal instability
; micronucleus
; genome evolution
; macroevolution
; speciation
; gametogenesis
; embryogenesis
; cancer
1. Introduction on Evolution and Chromoagenesis
Evolution is intricately linked to the capacity to change and adapt. It manifests along a spectrum, with some species evolving rapidly while others exhibit notably slower rates of change. A few animal species are known for their highly efficient DNA repair mechanisms and are referred to as 'living fossils' due to their incredibly low species diversity and minimal differences from their fossil relatives [1,2]. In the other side, some species exhibit extreme cases of genomic evolution, with chromosomal evolution being one of the most visible and impactful changes. One of the mysteries of structural genome evolution is its extreme temporal irregularity. For many organisms, it is known that periods of very slow genome changes alternate with explosions of chromosomal rearrangements, resulting in rapid and complete reshuffling of the linkage groups and gene orders [3]. These types of evolution are known as rapid chromosomal change [4,5], extensive chromosome evolution [6], fast karyotype evolution [7], karyotypic mega-evolution [8], or runaway chromosomal evolution [9]. Events like whole genome doubling favor rapid and drastic changes in karyotype due to their genomic instability [10,11]. Furthermore, such explosive evolution is now known to be associated with chromoanagenesis, a process of chromosome rebirth from chaos, resulting from a catastrophic event. Chromoanagenesis is an umbrella term that to describe the full spectrum of large-scale rearrangement [12]. The most frequent and well-studied example is chromothripsis, where hundreds of chromosome fragments are reorganized, duplicated or removed. In addition, other complex rearrangements include chromoplexy, chromoanasynthesis. These drastic chromosomal evolutions have recently been well described in disease such as cancer [12,13,14] or in the case of rare diseases [15,16] but is also known in evolution [17] from numerous groups of organisms like mammals [5], plankton [18], fungi [19] or plants [20,21,22].
2. Chromoagenesis, a Chromosome Rebirth
Chromoanagenesis is a term describing the complex chromosomal rearrangements chromothripsis, chromoplexy and chromoanagenesis (Figure 1).
Chromothripsis is a chromosomal instability phenomenon, a genomic catastrophe, where hundreds of chromosomal rearrangements occur during one single event in a localized region of one or two chromosomes. It was first described by Stephens et al [23] in a patient with chronic lymphocytic leukemia. It is now found in 40-80% of all tumors [24]. It was also rapidly identified in patients with congenital malformations, developmental disorders or carrying apparently balanced rearrangements [15,16,17]. Chromothripsis involve a pulverization of the chromosome in several fragments that can reorganized in a different order during the next mitosis [25,26]. Chromothripsis is known to drive tumorigenesis by disruption or loss of tumor suppressor, generation of oncogenic fusions and amplification of oncogene through extrachromosomal DNA (ecDNA) [12,27].
Chromoplexy, in another hand, tends to involve less pieces of chromosome but implicates more chromosomes, frequently 5 or even more [28]. It leads to multiple interchromosomal and intrachromosomal translocations and deletions.
Chromoanasynthesis is replication-based process with local rearrangement and altered gene copy number due to serial fork stalling or template switching, or microhomology-mediated break-induced replication [29]. It involves duplications and deletions and it is associated with replication stress.
3. Molecular Mechanisms to Generate Genome Rearrangements, Source for Evolution
The origin of chromoanagenesis is not fully understood yet. However, concerning chromothripsis, it seems clear that nuclear envelope rupture (NER) from a micronucleus is responsible for the chromosome pulverization. During mitosis, a lagging chromosome or a chromosome fragment resulting from an unrepaired dsDNA break (DSBs), can produce the formation of micronuclei (Figure 2). When a full or piece of chromosome is late during the nuclear envelope (NE) formation, it is not clustered with the other chromosomes. Then a proper NE forms around this one, forming a separate small nucleus close to the main nucleus, known as micronuclei (MN) [33].
Micronuclei formation is now recognized as a major event that triggers chromosome shattering, followed by the aberrant reassembly of the fragments [25,26,34]. NER happens on micronuclei, where the envelope is fragile and tends to disrupt without possibility of proper repair [35,36]. The nuclear envelope of micronuclei often lacks sufficient nuclear pores and normally is associated with is delayed DNA replication and repair [25,37,38]. This envelope can rupture exposing the chromatin to cytosolic factors [36,37]. Factors contributing to this rupture include the loss or mislocalization of the nuclear lamina component lamin B1 [36,39], depletion of nuclear envelope proteins (Figure 3) [37] or activation of ATR pathway to clear damaged DNA [40]. Experimental evidence suggests that chromosomes from ruptured micronuclei undergo chromothripsis, a process of chromosome shattering and rearrangement [25,26].
In another case, NER happens in cells with a chromatin bridge, where telomere fusion connects two daughter cells [41]. This implies the generation of additional tension forces affecting the NE during movement, leading to NER that can last up to two minutes [42]. Cells can also experience transient NER of their nuclei during migration [43,44,45,46]. Interestingly, immune attack can fail and contribute to mutagenesis. Cytotoxic T lymphocytes often fail to kill target cells during one-on-one interaction, leading to transient NER and subsequent DNA damage [47]. Such ‘failed’ attacks then contribute to the generation of genomic diversity and progression toward resistance. Additionally, certain viruses, such as human papillomavirus (HPV) and Epstein–Barr virus (EBV), have been implicated in the formation of micronuclei, leading to chromothripsis in infected cells. These viral interactions might have played a role in chromosomal rearrangements, evolution, and speciation [48,49].
Remarkably, a study using CRISPR-Cas9 gene editing has shown that a single DSB can trigger a cascade of events resulting in the formation of micronuclei and chromosome bridges [50]. Such simple DSBs can be amplified into far more extensive genomic alterations during subsequent mitosis, leading to a myriad of genomic diversity.
During NER, DSBs can occur due to the entry of cytoplasmic nucleases such as TREX1 [41,51], errors in DNA repair or replication [12,52]. Such breaks can lead to chromosome pulverization and fragmentation. Several mechanisms have been proposed to explain the complexity of new genomic rearrangements. One of such mechanisms is non-homologous end joining (NHEJ), which can lead to reciprocal translocations following DSBs [53,54]. Studies indicate that translocations are more frequent in mammalian cells lacking NHEJ components like Ku70 and ligase Xrcc4, suggesting that alternative end-joining (alt-EJ) mechanisms might facilitate these translocations [55]. These processes often involve modifications at break sites, typically resulting in deletions. Other common features of DNA junctions formed by end-joining could include microhomology [55]. A novel model, fork stalling and template switching (FoSTeS), has been introduced to explain the complexity and microhomologies at breakpoints of non-recurrent duplications and deletions associated with genomic disorders [56]. This model is related to another called microhomology-mediated break-induced replication (MMBIR), which is based on break-induced replication and accounts for various complex rearrangements, mainly involving duplications and deletions [56,57,58].
Interestingly, samples with events of whole genome doubling (WGD) are associated with the presence of chromothripsis [12]. WGD has been shown to induce replication stress as well as DSBs [59]. Furthermore, during proliferation, WGD+ cells, which have extra centrosomes, exhibit chromosomal instability during proliferation, leading to an elevated rate of chromosome mis-segregation and the rapid accumulation of both numerical and structural abnormalities [60].
4. Chromosomal Instability and Macroevolution
In the wild, organisms can show an impressive change in number of chromosomes between closely related species. This enormous karyotypic diversification indicates that chromosomal evolution is an active and recurrent process. Such changes are also associated with chromoanagenesis-like rearrangements. Recent advances in sequencing technology and bioinformatics software, along with their reduced costs, have enabled the detailed characterization of highly complex chromosomal rearrangements mostly in human tumor samples. While the human genome is well-mapped, reference templates for other species are necessary to fully understand genomic complexity. There is no doubt that in the near future, chromoanagenesis will be described in the genomic evolution of several species, especially those showing huge karyotype diversification. In fact, a recent study has shown the importance of chromoanagenesis in adaptation as observed between marine annelids and the terrestrial clitellates. This study provides evidence of punctuated genomic change via chromoanagenesis leading not only to the origin of a new animal lineage but also to adaptive genomic changes facilitating the colonization of new habitats [61].
The case of tuco-tuco, a small neotropical rodent in the family Ctenomyidae found in South America, shows an extraordinary chromosomal variability. With 70 species with a diploid number varying from 10 to 70 chromosomes, it is one of the most speciose and chromosomally variable rodent genera of the world [62]. Within the genus, several types of chromosomal rearrangements were described such as Robertsonian translocations, inversions, heterochromatin additions and deletions, as well as complex rearrangements [62,63,64]. In Corydoras fish genus (Siluriformes: Callichthyidae) diploid numbers range from n = 44 to n = 102 and have extensive chromosomal rearrangements [65]. Similar diversity is found in Muntjac deer (Artiodactyla: Cervidae) with karyotypes ranging from n = 3 to n = 23 [66] and drastic chromosomal rearrangements [5]. In plants, the greatest range of within-genus karyotype variation is found in Carex, where haploid chromosome numbers range from n = 6 to n = 66 [67,68]. Lepidoptera is an order of winged insects that includes butterflies and moths. Despite this general stability, some genera of butterflies and moths exhibit remarkable variation in chromosome numbers. For instance, in the clade comprising the genera ((Polyommatus + Neolysandra) + Lysandra) (Lepidoptera, Lycaenidae), an array of chromosomally diverse species emerged within less than 5 million years of evolution. In these species, the haploid chromosome numbers range from n = 10 to n = 226 [3]. Lepidoptera evolution involves fission and fusion of chromosomes, but also some species present much more complex rearrangements of their chromosomes [3,69].
The selective breeding of new species with enhanced production and improved flavor is foundational to modern agriculture and livestock farming and such new species are often associated with chromosomal aberrations. In plants as coffee [70] or grape [71], in cattle [72] or in yeast [73], selective pressure applied during selection is associated with massive changes in their chromosome organization. However, such selection is often associated with a decrease in resilience and a loss of advantages in the wild. For example, cattle breeding is associated with dysfunction in fertility, notably due to Robertsonian translocation, a common type of translocation [72]. In the case of black and white tempranillo, different grape varieties, a genomic study has shown events of complex chromosomal exchanges between three chromosomes [71], that are likely a chromoplexy pattern. However, even though these new species were selected for their gustative values, in term of stiffness, such chromosomal changes are associated with issues in viability of gametes [74]. In yeast, the 'Guinness strains' exhibit polyploidy and chromosomal aberrations. Presumably due to the effects of aneuploidy, all 16 known Guinness strains show poor sporulation [73].
Extreme conditions found in the laboratory, such as the use of haploid induction in plants, biolistic transformation, mutagenesis, or irradiation, can result in both chromothripsis and chromoplexy. In plant, such events were observed in Arabidopsis thaliana [75,76], poplar trees [21,77], maize [78], rice [78] or potatoes [79]. In C. elegans, chromoanasynthesis was observed after a mutagenesis screen [80,81]. Interestingly, ecDNA known to be associated with chromothripsis can serve as a mechanism of resistance for organisms such as yeast for copper resistance or plants for glyphosate resistance [82].
5. Chromosomal Instability in Cancer
An intriguing aspect lies in the similarity and parallelism between the history of genomic complexity in tumors and genome evolutionary changes. Both are shaped by drastic events like chromosome rearrangements and the gain or loss of chromosomes, which drive their evolution and adaptation to new environments and pressures. WGD is an early event in tumorigenesis [83,84,85,86,87], promoting genome diversity and chromosomal instability [88], similar to what has been observed before the diversification of vertebrates [89,90] and in plants [91]. Interestingly, genome duplication can occur multiple times during tumor development, resulting in highly complex cancer genomes [84,92]. Furthermore, chromothripsis increased with whole-genome duplications in most cancer types [93].
Cancer genomes are complex, heterogeneous, and highly rearranged. Between 80-97% of tumors exhibit detectable CIN and complex chromosomal rearrangements [92,94]. Notably, chromothripsis is observed in 40-80% of tumors, chromoplexy in 10%, and chromoanasynthesis in 5% [23,24,28,30,95,96]. Thus, chromoanagenesis plays a crucial role in driving tumor evolution. Furthermore, tumor progression, in another word, evolution, is associated with increased complexity and genomic large-scale reorganization within the metastatic lesions [83,84,85,86,87,97], accelerating the odds of developing resistance to anti-cancer therapies [98]. Importantly, chromothripsis is associated with the formation of circular extrachromosomal DNA (ecDNA) as well as with segmental deletion, supporting the amplification of oncogenes as well as the disruption or loss of tumor suppressor, highlighting their pivotal role in tumor evolution [12,27,34,99].
6. Mitotic and Meiotic Programs as Drivers for Genome Evolution
The genome must balance stability and variability to preserve essential information while allowing adaptations to environmental changes. Accurate DNA repair and replication, during mitosis and meiosis, are crucial for maintaining and transmitting DNA through chromosomes during development and from parents to offspring. Despite programmed DSBs and meiotic recombination mixing genome information, these processes are not entirely error-free [13]. It could lead in de novo genomic rearrangements, such as inversions, duplications, insertions/deletions (indels), and large-scale structural changes like Robertsonian translocations and Whole Arms Translocations (WARTs), or chromoanagenesis processes, and aneuploidies. These structural changes significantly impact chromosomal and nuclear architecture [102,103]. These chromosomal rearrangements can occur during gamete formation, zygote formation, and embryo development, affecting the individual's genome, and may vary with each stage of mitosis and meiosis [104,105]. The evolutionary impact of de novo mutations depends on their characteristics and the context of their emergence, as tissues or time of development appearance.
Chromatin domains and compartment patterns differ among cell types, with topologically associating domains (TADs) undergoing reorganization throughout the cell cycle [106,107]. Chromothripsis reshapes gene regulation, leading to coordinated changes in the epigenetic landscape, transcription, and TADs domains. These reorganizations are crucial for genome functionality and inheritance. In cancer, TADs are commonly disturbed by chromothripsis, leading to changes in gene expression profiles [108]. Chromosomal alterations disrupting TAD architecture can lead from meiotic recombination defects to developmental deficiencies due to improper enhancer-promoter interactions [109]. Additionally, research has examined conserved chromosome territory arrangements and species-specific changes contributing to reproductive isolation and speciation [110,111,112].
7. Rearrangements of Chromosomes During the Initial Stages of Development
Although chromosomal rearrangements are known to occur during meiosis, the majority of documented chromoanagenesis events in animals [26,34,57,113], and all reported events in plants [75,78,114], are associated with mitotic processes, particularly during the early stages of embryonic development [75,115]. Chromosomal instability may occur during one of the initial mitotic divisions of early embryonic development, and such errors can either lead to the embryo's elimination or result in mosaicism. Mosaicism occurs when an individual has two or more genetically distinct sets of cells. Typically, it originates from a single cell that becomes the progenitor of a clone, which can sometimes cause genetic diseases, including cancer, more commonly in children than in adults [116,117]. Mosaicism, which is common in mammals including humans, can arise from unequal chromosomal division of blastomeres or the presence of different cell clones [118]. Approximately 50–70% of human cleavage embryos are aneuploid, highlighting the high levels of chromosomal instability during the generation of offspring [119,120].
Another chromosomal anomaly, the formation of a dicentric Y chromosome, leads to sterility in humans and is believed to occur early in development due to recombination between sister chromatids or delayed joining after a break or deletion. This can cause mosaic disorders, such as sterile males with karyotypes lacking a Y chromosome [121,122]. Dicentric chromosomes are also observed in livestock such as buffaloes, sheep, and goats [123]. Despite their formation, these chromosomes are often eliminated within a few generations due to centromeric drive, meaning they usually do not have an evolutionary impact since they are not passed on to gametes.
Most aneuploid embryos fail to develop to term, making aneuploidy in embryos a leading cause of miscarriages and infertility. Nonetheless, if chromosomal alterations occur during the formation of primordial germ cells and do not severely impair gametogenesis and fertility, these changes could be inherited and may have evolutionary significance [124]. Another key aspect to consider is the reorganization of the zygotic genome for subsequent activation from the previous maternal control. Chromosomal rearrangements impact both the physical arrangement of chromatin and the regulation of gene activity [125,126,127]. Research in mice has demonstrated that chromatin architecture is reprogrammed during germ cell development [128]. If the chromatin architecture is not correctly established at the onset of embryonic development, the genetic program cannot proceed, leading to the termination of the embryo's development [129,130]. Chromosomal rearrangements can also impact the genetic program beyond the directly affected sequence. However, these rearrangements can introduce new combinations of genetic patterns that contribute to genome evolution and speciation. Studies have shown that functional plasticity under a higher-order genomic organization is inherited by offspring, leading to the formation of new allelic variants on which natural selection and evolution can act [127].
8. Chromosomal Instability Processes in the Germ Cells
Genome instability can have significant evolutionary consequences, leading either to extinction or rapid speciation. This process is closely tied to the emergence and establishment of chromosome rearrangements within populations. The spread and fixation of these rearrangements can be driven by their impact on gametogenesis, spermatogenesis in males or oogenesis in females. For instance, rearrangements that result in altered gametes during spermatogenesis or oogenesis, as the ones taking place in the germline, can facilitate the rapid establishment of these changes within a few generations [13]. Models of population genetics suggest that even slight reductions in viability or fertility due to these rearrangements can promote the development of divergent genetic lines, even in populations that coexist geographically. This can lead to reproductive isolation and eventually speciation.
Germline rates of de novo meiotic deletions and duplications are a significant source of genetic diversity, contributing to several genomic disorders. Chromosomal evolution, as deep chromosomal rearrangements or reciprocal translocations in germ lines are particularly important, as they have been observed across multiple phyla, including humans. A study in a trio family has identify de novo balanced rearrangements in a set of three chromosomes in a patient, mediated by simultaneous breakpoints accumulated in specific chromosomal regions and joined, resulting in a chromothripsis pattern, presumably in the germline. These translocations play a crucial role in generating genetic diversity, which is essential for the adaptive potential of populations [131,132,133,134].
Mosaicism in germ cells can have profound effects on genetic diversity and health. When chromothripsis occurs in germ cells, the resultant rearrangements can lead to significant genetic variations in gametes, potentially causing severe health problems [135]. Interestingly, many of these rearrangements, although stably transmitted in an unbalanced form from a healthy mother to her child with congenital abnormalities, are likely due to de novo copy-number changes of dosage-sensitive genes.
A major challenge in chromosome evolution is that new large-scale chromosome mutations must successfully go through meiosis. Even balanced chromosomal changes can harm the fertility of heterozygotes because they can mess up meiotic pairing and produce unbalanced gametes with incorrect gene dosage. However, a study has documented a case of balanced germline chromothripsis that was transmitted through three generations involving 11 healthy carriers. This suggests that chromothripsis does not always result in an abnormal phenotype. Specifically, the study hypothesized that truncation of the gene encoding the ataxia–telangiectasia and Rad3-related protein kinase (ATR), a vital component of the DNA damage response, might be linked to the chromothripsis event. This finding implies that germline chromothripsis may be more prevalent than currently acknowledged, underscoring its potential role in evolution and species divergence [136].
9. Gametogenesis as a Source of Chromoanagenesis
The relationship between fertility and chromosomal instability is complex and multifaceted. Abnormal chromosomes can disrupt meiosis, resulting in the production of aneuploid sperm or eggs. When these aneuploid gametes are involved in fertilization, the resulting embryos often have chromosomal abnormalities, which can lead to miscarriages or developmental issues. Additionally, chromosomal instability can impair fertility by affecting the mitotic self-renewing cells in the germ line, which are essential for the formation of gametes.
In multicellular diploid organisms like mammals, the creation of new life begins with the fusion of two haploid gametes, restoring the diploid state in the zygote. The differences between male and female meiosis and their respective products influence meiotic drive, a mechanism of natural selection [13]. Each step of gametogenesis must be meticulously regulated to ensure proper development. Male and female meiosis differ significantly in both timing and the resulting products. In males, meiosis produces four haploid spermatids, which differentiate into four spermatozoa. In females, meiosis involves asymmetric cytokinesis, resulting in one functional oocyte and two discarded polar bodies. This asymmetry during the first and second meiotic divisions affects selection processes in forming the final haploid gamete, the egg. The first division produces an oocyte and a first polar body, both with a haploid set of chromosomes but doubled DNA content. This occurs after synapsis and chromosome recombination, leading to distinct genomes for the oocyte and the polar body. The second division results in another polar body and the mature egg, each with a haploid content. In mammals, the appearance of polar bodies occurs after fertilization, as female gametes are arrested in Prophase I until triggered by fertilization. Properly clustering and separating polar bodies from parental genomes is crucial, as errors can lead to aneuploidy and the formation of micronuclei, which are common issues. Micronuclei can ultimately result in aneuploidy or chromoanagenesis events (Figure 4). Thus, polar bodies may enable rapid changes in the chromosome set, making meiotic drive a significant evolutionary force [137].
Chromothripsis has been observed during spermatogenesis and in the initial cleavages of preimplantation embryos, but not often during oogenesis [57,138]. Notably, the majority of identified chromoanagenesis events are linked with male gametogenesis [77]. Specifically, chromoanagenesis has been demonstrated to occur during meiotic divisions in spermiogenesis in humans [57,139]. While chromosomal rearrangements are also likely to arise due to errors in female meiosis, there are relatively few studies on this topic [138,140]. It has been reported that the DNA repair capacity in the male germ line is relatively low compared to somatic cells, and the mutation rate is higher during spermatogenesis than during oogenesis. The extent of these rearrangements can be substantial, potentially leading to significant evolutionary consequences [141,142]. The extensive genome reshuffling observed in rodents may be explained by chromoanagenesis events occurring in their germ lines [143,144]. The result of different meiotic segregation patterns can also be disrupted by chromosomal reorganizations involving different translocations, creating complex structures between homologous chromosomes that are difficult to resolve. This increases genome instability during meiosis. Studies in patients have described improper disjunction of chromosomes [131,145,146], which is a source of chromosomal variation and speciation.
Chromoanagenesis have also been observed during haploid induction in Arabidopsis thaliana [75]. Due to haploidy, chromosome mis-segregation can occur during gametogenesis, leading to the formation of a micronucleus containing a lagging chromosome. This chromosome, trapped within the micronucleus, undergoes fragmentation and reorganization, ultimately resulting in a shattered chromosome during meiosis (Figure 4). The viability of the meiotic process and survival of the next generation of plants can be attributed to the presence of three copies in a diploid background, where the shattered chromosome is buffered by the intact copies. Similar chromoanagenesis-like rearrangements have been identified as potentially influencing the genome structure of Camelina sativa and the genus Cucumis [22,147].
Although few cases of chromoanagenesis related to defects during gametogenesis have been described, especially in the wild [76], chromoanagenesis may be more common than currently recognized, similar to other rearrangements like Robertsonian translocations. These translocations involve the entire arms of chromosomes and occur in the acrocentric chromosomes 13, 14, 15, 21, and 22 in humans, making them the most frequent type of chromosome rearrangement in the human population and the most prevalent form of chromosomal changes observed in mammals [148]. In situations involving partial or chained-monobrachial homology, or when multiple Robertsonian translocations (Rbs) are present, synapsis can be extended but leading to meiotic failure in some cells and reduced fertility [149,150]. This phenomenon has been observed in species such as bats, shrews, mole voles, mice, and rock-wallabies, where monobrachial homology appears to restrict gene flow and possibly drive speciation [144,151,152,153]. In mice, various forms likely originated from whole-arm reciprocal translocations (WARTs), creating complex hybridization patterns through parental lines [154]. WARTs have been noted in the same groups as Rbs, suggesting a shared origin mechanism for these rearrangements. Occasionally, extensive rearrangements, including tandem fusions, can cause a surge in intraspecific diversity, as seen in the Evoron vole [134].
10. Meiotic Specific Processes as Drivers for Chromosomal Rearrangements
Recombination is a key aspect of meiosis that increases genetic diversity during inheritance. This process is carefully programmed and controlled, particularly through synapsis, where homologous chromosomes pair up via the synaptonemal complex. Any defects in these processes can lead to chromosomal structure changes. Heterologous synapsis, for example, can reduce recombination, cause sterility, or result in improper recombination [155,156]. These alterations can lead to new chromosomal rearrangements and change the organization of topologically associating domains (TADs), both crucial for chromosome evolution [157].
In Arabidopsis thaliana, a case of chromoanagenesis was reported in an asy1 mutant background, where defective meiosis led to significant chromosomal changes [76]. The ASY1 gene is a component of the synaptonemal complex that helps regulate crossover assurance and interference during meiosis. In this mutant, a dense breakpoint region on chromosome 1, associated with open euchromatin, was observed. Various types of rearrangements were identified, including those with microhomology, minor base pair additions or deletions, and perfect junctions without modifications. These rearrangements led to the loss of function in several genes and the emergence of novel genes, highlighting the evolutionary impact of such changes. The ASY1 mutation likely altered recombination patterns and caused unbalanced chromosome segregation during meiosis, especially in meiosis I. Additionally, micronuclei were observed during male sporogenesis in asy1 mutants, serving as a source of chromoanagenesis [158].
DNA recombination is a crucial process during meiosis, leading to physical attachment between homologous chromosomes in a step called crossing-over (CO). This universal feature is essential for accurate chromosome segregation during meiosis and sexual reproduction, thereby maintaining the genome integrity of species. Moreover, meiotic recombination shuffles alleles along homologous chromosomes, generating genetic diversity in sexual gametes and offspring [159]. Understanding the mechanisms and factors involved in meiotic CO formation and resolution is important for insights into biodiversity. However, if not properly regulated, this process can lead to repair errors, resulting in sterility or miscarriages. Conversely, proper regulation of new meiotic CO formation and resolution can be adaptive, supporting species evolution and the emergence of new species. Disruptions in chromosome architecture, such as inversions, fusions, or translocations, are often linked to genetic instability and cancer due to oncogene activation and novel gene functions. Similarly, intra- or interchromosomal alterations in the germ line or during meiosis can alter normal segregation patterns, contributing to organismal evolution [160,161]. Another important aspect is that meiotic recombination between highly similar duplicated sequences can cause various genome disorders, including deletions, duplications, inversions, and translocations [133]. Although there have been no documented cases of chromoanagenesis directly linked to defects in meiotic recombination so far, it is plausible that such defects could play a role in this process, and connected with evolution.
11. Concluding Remarks
The use of new technologies such as long read DNA sequencing has allowed the discovery of another level of complexity in the genome organization.
It is undoubted that several species have not been annotated for chromosomal rearrangement events because the complexity of the chromosomal rearrangements they have experienced is beyond our current ability to resolve [69]. Furthermore, the lack of proper species mapping limits the discovery of chromoanagenesis in the evolution genome.
Evolutionary biologists have found that polyploid organisms often face a competitive disadvantage compared to their diploid counterparts, which raises questions about the persistence of this trait. One answer could be that polyploidy may help species weather catastrophic environmental changes. Increased genetic flexibility provided by multiple genomes lets polyploids make quick adjustments to new stresses and ride out catastrophic events that wipe out most normal plants and animals [162]. As for WGD, aneuploidy and CIN also are disadvantageous at first for a cell, but also offer the opportunity to adapt to new environment. CIN can offer results in the evolution of large-scale structures such as chromoanagenesis, and can dramatically change the structure and functions of proteins, leading to the evolution of entirely new structures, adaptations, or increased fitness. As indicated by the persistence of germline chromothripsis through three generations in 11 healthy individuals, chromothripsis does not invariably lead to abnormal phenotypes [136]. This insight suggests that germline chromothripsis might be more widespread than we currently assumed, emphasizing its potential impact on evolution and species diversity. To conclude, the origin, mechanism, and potential effects of chromoanagenesis are just starting to be deciphered, marking an exciting frontier in our understanding of genetic evolution and its broader implications.
References
- Brownstein CD, MacGuigan DJ, Kim D, Orr O, Yang L, David SR, Kreiser B, Near TJ (2024) The genomic signatures of evolutionary stasis. Evolution. [CrossRef]
- Gazo I, Franek R, Sindelka R, Lebeda I, Shivaramu S, Psenicka M, Steinbach C (2020) Ancient Sturgeons Possess Effective DNA Repair Mechanisms: Influence of Model Genotoxicants on Embryo Development of Sterlet, Acipenser ruthenus. Int J Mol Sci 22 (1). [CrossRef]
- Pazhenkova EA, Lukhtanov VA (2023) Chromosomal conservatism vs chromosomal megaevolution: enigma of karyotypic evolution in Lepidoptera. Chromosome Res 31 (2):16. [CrossRef]
- Mudd AB, Bredeson JV, Baum R, Hockemeyer D, Rokhsar DS (2020) Analysis of muntjac deer genome and chromatin architecture reveals rapid karyotype evolution. Commun Biol 3 (1):480. [CrossRef]
- Yin Y, Fan H, Zhou B, Hu Y, Fan G, Wang J, Zhou F, Nie W, Zhang C, Liu L, Zhong Z, Zhu W, Liu G, Lin Z, Liu C, Zhou J, Huang G, Li Z, Yu J, Zhang Y, Yang Y, Zhuo B, Zhang B, Chang J, Qian H, Peng Y, Chen X, Chen L, Li Z, Zhou Q, Wang W, Wei F (2021) Molecular mechanisms and topological consequences of drastic chromosomal rearrangements of muntjac deer. Nature Communications 12 (1):6858. [CrossRef]
- Baker RJ, Bickham JW (1980) Karyotypic Evolution in Bats: Evidence of Extensive and Conservative Chromosomal Evolution in Closely Related Taxa. Systematic Biology 29 (3):239-253. [CrossRef]
- Carbone L, Harris RA, Gnerre S, Veeramah KR, Lorente-Galdos B, Huddleston J, Meyer TJ, Herrero J, Roos C, Aken B, Anaclerio F, Archidiacono N, Baker C, Barrell D, Batzer MA, Beal K, Blancher A, Bohrson CL, Brameier M, Campbell MS, Capozzi O, Casola C, Chiatante G, Cree A, Damert A, de Jong PJ, Dumas L, Fernandez-Callejo M, Flicek P, Fuchs NV, Gut I, Gut M, Hahn MW, Hernandez-Rodriguez J, Hillier LW, Hubley R, Ianc B, Izsvak Z, Jablonski NG, Johnstone LM, Karimpour-Fard A, Konkel MK, Kostka D, Lazar NH, Lee SL, Lewis LR, Liu Y, Locke DP, Mallick S, Mendez FL, Muffato M, Nazareth LV, Nevonen KA, O'Bleness M, Ochis C, Odom DT, Pollard KS, Quilez J, Reich D, Rocchi M, Schumann GG, Searle S, Sikela JM, Skollar G, Smit A, Sonmez K, ten Hallers B, Terhune E, Thomas GW, Ullmer B, Ventura M, Walker JA, Wall JD, Walter L, Ward MC, Wheelan SJ, Whelan CW, White S, Wilhelm LJ, Woerner AE, Yandell M, Zhu B, Hammer MF, Marques-Bonet T, Eichler EE, Fulton L, Fronick C, Muzny DM, Warren WC, Worley KC, Rogers J, Wilson RK, Gibbs RA (2014) Gibbon genome and the fast karyotype evolution of small apes. Nature 513 (7517):195-201. [CrossRef]
- Bell DM, Hamilton MJ, Edwards CW, Wiggins LE, MartÍnez RM, Strauss RE, Bradley RD, Baker RJ (2001) Patterns of Karyotypic Megaevolution in Reithrodontomys: Evidence From a Cytochrome-b Phylogenetic Hypothesis. Journal of Mammalogy 82 (1):81-91. [CrossRef]
- Vershinina AO, Lukhtanov VA (2017) Evolutionary mechanisms of runaway chromosome number change in Agrodiaetus butterflies. Sci Rep 7 (1):8199. [CrossRef]
- Van de Peer Y, Mizrachi E, Marchal K (2017) The evolutionary significance of polyploidy. Nat Rev Genet 18 (7):411-424. [CrossRef]
- Comai L (2005) The advantages and disadvantages of being polyploid. Nat Rev Genet 6 (11):836-846. [CrossRef]
- Krupina K, Goginashvili A, Cleveland DW (2024) Scrambling the genome in cancer: causes and consequences of complex chromosome rearrangements. Nat Rev Genet 25 (3):196-210. [CrossRef]
- Comaills V, Castellano-Pozo M (2023) Chromosomal Instability in Genome Evolution: From Cancer to Macroevolution. Biology (Basel) 12 (5). [CrossRef]
- Holland AJ, Cleveland DW (2012) Chromoanagenesis and cancer: mechanisms and consequences of localized, complex chromosomal rearrangements. Nat Med 18 (11):1630-1638. [CrossRef]
- Schuy J, Grochowski CM, Carvalho CMB, Lindstrand A (2022) Complex genomic rearrangements: an underestimated cause of rare diseases. Trends Genet 38 (11):1134-1146. [CrossRef]
- Zepeda-Mendoza CJ, Morton CC (2019) The Iceberg under Water: Unexplored Complexity of Chromoanagenesis in Congenital Disorders. Am J Hum Genet 104 (4):565-577. [CrossRef]
- Pellestor F, Gatinois V (2020) Chromoanagenesis: a piece of the macroevolution scenario. Mol Cytogenet 13:3. [CrossRef]
- Blanc-Mathieu R, Krasovec M, Hebrard M, Yau S, Desgranges E, Martin J, Schackwitz W, Kuo A, Salin G, Donnadieu C, Desdevises Y, Sanchez-Ferandin S, Moreau H, Rivals E, Grigoriev IV, Grimsley N, Eyre-Walker A, Piganeau G (2017) Population genomics of picophytoplankton unveils novel chromosome hypervariability. Sci Adv 3 (7):e1700239. [CrossRef]
- Croll D, Zala M, McDonald BA (2013) Breakage-fusion-bridge cycles and large insertions contribute to the rapid evolution of accessory chromosomes in a fungal pathogen. PLoS Genet 9 (6):e1003567. [CrossRef]
- Burchardt P, Buddenhagen CE, Gaeta ML, Souza MD, Marques A, Vanzela ALL (2020) Holocentric Karyotype Evolution in Rhynchospora Is Marked by Intense Numerical, Structural, and Genome Size Changes. Front Plant Sci 11:536507. [CrossRef]
- Guo W, Comai L, Henry IM (2023) Chromoanagenesis in plants: triggers, mechanisms, and potential impact. Trends Genet 39 (1):34-45. [CrossRef]
- Zhao Q, Meng Y, Wang P, Qin X, Cheng C, Zhou J, Yu X, Li J, Lou Q, Jahn M, Chen J (2021) Reconstruction of ancestral karyotype illuminates chromosome evolution in the genus Cucumis. Plant J 107 (4):1243-1259. [CrossRef]
- Stephens PJ, Greenman CD, Fu B, Yang F, Bignell GR, Mudie LJ, Pleasance ED, Lau KW, Beare D, Stebbings LA, McLaren S, Lin ML, McBride DJ, Varela I, Nik-Zainal S, Leroy C, Jia M, Menzies A, Butler AP, Teague JW, Quail MA, Burton J, Swerdlow H, Carter NP, Morsberger LA, Iacobuzio-Donahue C, Follows GA, Green AR, Flanagan AM, Stratton MR, Futreal PA, Campbell PJ (2011) Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell 144 (1):27-40. [CrossRef]
- Cortes-Ciriano I, Lee JJ, Xi R, Jain D, Jung YL, Yang L, Gordenin D, Klimczak LJ, Zhang CZ, Pellman DS, Group PSVW, Park PJ, Consortium P (2020) Comprehensive analysis of chromothripsis in 2,658 human cancers using whole-genome sequencing. Nat Genet 52 (3):331-341. [CrossRef]
- Crasta K, Ganem NJ, Dagher R, Lantermann AB, Ivanova EV, Pan Y, Nezi L, Protopopov A, Chowdhury D, Pellman D (2012) DNA breaks and chromosome pulverization from errors in mitosis. Nature 482 (7383):53-58.
- Zhang C-Z, Spektor A, Cornils H, Francis JM, Jackson EK, Liu S, Meyerson M, Pellman D (2015) Chromothripsis from DNA damage in micronuclei. Nature 522 (7555):179-184.
- Shoshani O, Brunner SF, Yaeger R, Ly P, Nechemia-Arbely Y, Kim DH, Fang R, Castillon GA, Yu M, Li JSZ, Sun Y, Ellisman MH, Ren B, Campbell PJ, Cleveland DW (2021) Chromothripsis drives the evolution of gene amplification in cancer. Nature 591 (7848):137-141. [CrossRef]
- Baca SC, Prandi D, Lawrence MS, Mosquera JM, Romanel A, Drier Y, Park K, Kitabayashi N, MacDonald TY, Ghandi M, Van Allen E, Kryukov GV, Sboner A, Theurillat JP, Soong TD, Nickerson E, Auclair D, Tewari A, Beltran H, Onofrio RC, Boysen G, Guiducci C, Barbieri CE, Cibulskis K, Sivachenko A, Carter SL, Saksena G, Voet D, Ramos AH, Winckler W, Cipicchio M, Ardlie K, Kantoff PW, Berger MF, Gabriel SB, Golub TR, Meyerson M, Lander ES, Elemento O, Getz G, Demichelis F, Rubin MA, Garraway LA (2013) Punctuated evolution of prostate cancer genomes. Cell 153 (3):666-677. [CrossRef]
- Liu P, Erez A, Nagamani SC, Dhar SU, Kolodziejska KE, Dharmadhikari AV, Cooper ML, Wiszniewska J, Zhang F, Withers MA, Bacino CA, Campos-Acevedo LD, Delgado MR, Freedenberg D, Garnica A, Grebe TA, Hernandez-Almaguer D, Immken L, Lalani SR, McLean SD, Northrup H, Scaglia F, Strathearn L, Trapane P, Kang SH, Patel A, Cheung SW, Hastings PJ, Stankiewicz P, Lupski JR, Bi W (2011) Chromosome catastrophes involve replication mechanisms generating complex genomic rearrangements. Cell 146 (6):889-903. [CrossRef]
- Baker TM, Waise S, Tarabichi M, Van Loo P (2024) Aneuploidy and complex genomic rearrangements in cancer evolution. Nat Cancer 5 (2):228-239. [CrossRef]
- Rosswog C, Bartenhagen C, Welte A, Kahlert Y, Hemstedt N, Lorenz W, Cartolano M, Ackermann S, Perner S, Vogel W, Altmuller J, Nurnberg P, Hertwig F, Gohring G, Lilienweiss E, Stutz AM, Korbel JO, Thomas RK, Peifer M, Fischer M (2021) Chromothripsis followed by circular recombination drives oncogene amplification in human cancer. Nat Genet 53 (12):1673-1685. [CrossRef]
- Umbreit NT, Zhang CZ, Lynch LD, Blaine LJ, Cheng AM, Tourdot R, Sun L, Almubarak HF, Judge K, Mitchell TJ, Spektor A, Pellman D (2020) Mechanisms generating cancer genome complexity from a single cell division error. Science 368 (6488). [CrossRef]
- Gauthier BR, Comaills V (2021) Nuclear Envelope Integrity in Health and Disease: Consequences on Genome Instability and Inflammation. Int J Mol Sci 22 (14). [CrossRef]
- Ly P, Brunner SF, Shoshani O, Kim DH, Lan W, Pyntikova T, Flanagan AM, Behjati S, Page DC, Campbell PJ (2019) Chromosome segregation errors generate a diverse spectrum of simple and complex genomic rearrangements. Nature genetics 51 (4):705-715.
- Vietri M, Schultz SW, Bellanger A, Jones CM, Petersen LI, Raiborg C, Skarpen E, Pedurupillay CRJ, Kjos I, Kip E, Timmer R, Jain A, Collas P, Knorr RL, Grellscheid SN, Kusumaatmaja H, Brech A, Micci F, Stenmark H, Campsteijn C (2020) Unrestrained ESCRT-III drives micronuclear catastrophe and chromosome fragmentation. Nat Cell Biol 22 (7):856-867. [CrossRef]
- Hatch EM, Fischer AH, Deerinck TJ, Hetzer MW (2013) Catastrophic nuclear envelope collapse in cancer cell micronuclei. Cell 154 (1):47-60.
- Liu S, Kwon M, Mannino M, Yang N, Renda F, Khodjakov A, Pellman D (2018) Nuclear envelope assembly defects link mitotic errors to chromothripsis. Nature 561 (7724):551-555.
- Krupina K, Goginashvili A, Cleveland DW (2021) Causes and consequences of micronuclei. Current opinion in cell biology 70:91-99.
- Vargas JD, Hatch EM, Anderson DJ, Hetzer MW (2012) Transient nuclear envelope rupturing during interphase in human cancer cells. Nucleus 3 (1):88-100.
- Joo YK, Black EM, Trier I, Haakma W, Zou L, Kabeche L (2023) ATR promotes clearance of damaged DNA and damaged cells by rupturing micronuclei. Mol Cell 83 (20):3642-3658 e3644. [CrossRef]
- Maciejowski J, Chatzipli A, Dananberg A, Chu K, Toufektchan E, Klimczak LJ, Gordenin DA, Campbell PJ, de Lange T (2020) APOBEC3-dependent kataegis and TREX1-driven chromothripsis during telomere crisis. Nature genetics 52 (9):884-890.
- Maciejowski J, Li Y, Bosco N, Campbell PJ, de Lange T (2015) Chromothripsis and Kataegis Induced by Telomere Crisis. Cell 163 (7):1641-1654. [CrossRef]
- Denais CM, Gilbert RM, Isermann P, McGregor AL, te Lindert M, Weigelin B, Davidson PM, Friedl P, Wolf K, Lammerding J (2016) Nuclear envelope rupture and repair during cancer cell migration. Science 352 (6283):353-358. [CrossRef]
- Comaills V, Kabeche L, Morris R, Buisson R, Yu M, Madden MW, LiCausi JA, Boukhali M, Tajima K, Pan S, Aceto N, Sil S, Zheng Y, Sundaresan T, Yae T, Jordan NV, Miyamoto DT, Ting DT, Ramaswamy S, Haas W, Zou L, Haber DA, Maheswaran S (2016) Genomic Instability Is Induced by Persistent Proliferation of Cells Undergoing Epithelial-to-Mesenchymal Transition. Cell Rep 17 (10):2632-2647. [CrossRef]
- Raab M, Gentili M, de Belly H, Thiam HR, Vargas P, Jimenez AJ, Lautenschlaeger F, Voituriez R, Lennon-Dumenil AM, Manel N, Piel M (2016) ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death. Science 352 (6283):359-362. [CrossRef]
- Gauthier BR, Lorenzo PI, Comaills V (2021) Physical Forces and Transient Nuclear Envelope Rupture during Metastasis: The Key for Success? Cancers (Basel) 14 (1). [CrossRef]
- Weigelin B, den Boer AT, Wagena E, Broen K, Dolstra H, de Boer RJ, Figdor CG, Textor J, Friedl P (2021) Cytotoxic T cells are able to efficiently eliminate cancer cells by additive cytotoxicity. Nat Commun 12 (1):5217. [CrossRef]
- Dacus D, Stancic S, Pollina SR, Rifrogiate E, Palinski R, Wallace NA (2022) Beta human papillomavirus 8 E6 induces micronucleus formation and promotes chromothripsis. Journal of Virology 96 (19):e01015-01022.
- Li JSZ, Abbasi A, Kim DH, Lippman SM, Alexandrov LB, Cleveland DW (2023) Chromosomal fragile site breakage by EBV-encoded EBNA1 at clustered repeats. Nature 616 (7957):504-509.
- Leibowitz ML, Papathanasiou S, Doerfler PA, Blaine LJ, Sun L, Yao Y, Zhang CZ, Weiss MJ, Pellman D (2021) Chromothripsis as an on-target consequence of CRISPR-Cas9 genome editing. Nature genetics. [CrossRef]
- Nader GPF, Aguera-Gonzalez S, Routet F, Gratia M, Maurin M, Cancila V, Cadart C, Palamidessi A, Ramos RN, San Roman M, Gentili M, Yamada A, Williart A, Lodillinsky C, Lagoutte E, Villard C, Viovy JL, Tripodo C, Galon J, Scita G, Manel N, Chavrier P, Piel M (2021) Compromised nuclear envelope integrity drives TREX1-dependent DNA damage and tumor cell invasion. Cell 184 (20):5230-5246 e5222. [CrossRef]
- Tang S, Stokasimov E, Cui Y, Pellman D (2022) Breakage of cytoplasmic chromosomes by pathological DNA base excision repair. Nature 606 (7916):930-936. [CrossRef]
- Lieber MR (2010) The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annual review of biochemistry 79:181-211.
- McVey M, Lee SE (2008) MMEJ repair of double-strand breaks (director’s cut): deleted sequences and alternative endings. Trends in Genetics 24 (11):529-538.
- Simsek D, Jasin M (2010) Alternative end-joining is suppressed by the canonical NHEJ component Xrcc4–ligase IV during chromosomal translocation formation. Nature structural & molecular biology 17 (4):410-416.
- Lee JA, Carvalho CM, Lupski JR (2007) A DNA replication mechanism for generating nonrecurrent rearrangements associated with genomic disorders. cell 131 (7):1235-1247.
- Kloosterman WP, Guryev V, van Roosmalen M, Duran KJ, de Bruijn E, Bakker SCM, Letteboer T, van Nesselrooij B, Hochstenbach R, Poot M, Cuppen E (2011) Chromothripsis as a mechanism driving complex de novo structural rearrangements in the germline†. Human Molecular Genetics 20 (10):1916-1924. [CrossRef]
- Zhang F, Khajavi M, Connolly AM, Towne CF, Batish SD, Lupski JR (2009) The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nature genetics 41 (7):849-853.
- Gemble S, Wardenaar R, Keuper K, Srivastava N, Nano M, Mace AS, Tijhuis AE, Bernhard SV, Spierings DCJ, Simon A, Goundiam O, Hochegger H, Piel M, Foijer F, Storchova Z, Basto R (2022) Genetic instability from a single S phase after whole-genome duplication. Nature 604 (7904):146-151. [CrossRef]
- Vittoria MA, Quinton RJ, Ganem NJ (2023) Whole-genome doubling in tissues and tumors. Trends Genet 39 (12):954-967. [CrossRef]
- Vargas-Chávez C, Benítez-Álvarez L, Martínez-Redondo GI, Álvarez-González L, Salces-Ortiz J, Eleftheriadi K, Escudero N, Guiglielmoni N, Flot J-F, Novo M, Ruiz-Herrera A, McLysaght A, Fernández R (2024) A punctuated burst of massive genomic rearrangements by chromosome shattering and the origin of non-marine annelids. bioRxiv:2024.2005.2016.594344. [CrossRef]
- Buschiazzo LM, Caraballo DA, Labaroni CA, Teta P, Rossi MS, Bidau CJ, Lanzone C (2022) Comprehensive cytogenetic analysis of the most chromosomally variable mammalian genus from South America: (Rodentia: Caviomorpha: Ctenomyidae). Mamm Biol 102 (5-6):1963-1979. [CrossRef]
- Buschiazzo LM, Caraballo DA, Calcena E, Longarzo ML, Labaroni CA, Ferro JM, Rossi MS, Bolzan AD, Lanzone C (2018) Integrative analysis of chromosome banding, telomere localization and molecular genetics in the highly variable Ctenomys of the Corrientes group (Rodentia; Ctenomyidae). Genetica 146 (4-5):403-414. [CrossRef]
- Caraballo DA, Abruzzese GA, Rossi MS (2012) Diversity of tuco-tucos (Ctenomys, Rodentia) in the Northeastern wetlands from Argentina: mitochondrial phylogeny and chromosomal evolution. Genetica 140 (4-6):125-136. [CrossRef]
- C. Oliveira LFA-T, L. Mori, S. A. Toledo-Filho (1992) Extensive chromosomal rearrangements and nuclear DNA content changes in the evolution of the armoured catfishes genus Corydoras (Pisces, Siluriformes, Callichthyidae). [CrossRef]
- Wang W, Lan H (2000) Rapid and parallel chromosomal number reductions in muntjac deer inferred from mitochondrial DNA phylogeny. Mol Biol Evol 17 (9):1326-1333. [CrossRef]
- Hipp AL (2007) Nonuniform processes of chromosome evolution in sedges (Carex: Cyperaceae). Evolution 61 (9):2175-2194. [CrossRef]
- Chung KS, Hipp AL, Roalson EH (2012) Chromosome number evolves independently of genome size in a clade with nonlocalized centromeres (Carex: Cyperaceae). Evolution 66 (9):2708-2722. [CrossRef]
- Chen X, Wang Z, Zhang C, Hu J, Lu Y, Zhou H, Mei Y, Cong Y, Guo F, Wang Y, He K, Liu Y, Li F (2023) Unraveling the complex evolutionary history of lepidopteran chromosomes through ancestral chromosome reconstruction and novel chromosome nomenclature. BMC Biol 21 (1):265. [CrossRef]
- Scalabrin S, Magris G, Liva M, Vitulo N, Vidotto M, Scaglione D, Del Terra L, Ruosi MR, Navarini L, Pellegrino G, Berny Mier YTJC, Toniutti L, Suggi Liverani F, Cerutti M, Di Gaspero G, Morgante M (2024) A chromosome-scale assembly reveals chromosomal aberrations and exchanges generating genetic diversity in Coffea arabica germplasm. Nat Commun 15 (1):463. [CrossRef]
- Carbonell-Bejerano P, Royo C, Torres-Perez R, Grimplet J, Fernandez L, Franco-Zorrilla JM, Lijavetzky D, Baroja E, Martinez J, Garcia-Escudero E, Ibanez J, Martinez-Zapater JM (2017) Catastrophic Unbalanced Genome Rearrangements Cause Somatic Loss of Berry Color in Grapevine. Plant Physiol 175 (2):786-801. [CrossRef]
- Jennings RL, Griffin DK, O'Connor RE (2020) A new Approach for Accurate Detection of Chromosome Rearrangements That Affect Fertility in Cattle. Animals (Basel) 10 (1). [CrossRef]
- Kerruish DWM, Cormican P, Kenny EM, Kearns J, Colgan E, Boulton CA, Stelma SNE (2024) The origins of the Guinness stout yeast. Commun Biol 7 (1):68. [CrossRef]
- Tello J, Royo C, Baroja E, García-Escudero E, Martínez-Zapater JM, Carbonell-Bejerano P (2021) Reduced gamete viability associated to somatic genome rearrangements increases fruit set sensitivity to the environment in Tempranillo Blanco grapevine cultivar. Sci Hortic-Amsterdam 290. [CrossRef]
- Tan EH, Henry IM, Ravi M, Bradnam KR, Mandakova T, Marimuthu MP, Korf I, Lysak MA, Comai L, Chan SW (2015) Catastrophic chromosomal restructuring during genome elimination in plants. Elife 4. [CrossRef]
- Guo WE, Comai L, Henry IM (2023) Chromoanagenesis in the asy1 meiotic mutant of Arabidopsis. G3-Genes Genom Genet 13 (2). [CrossRef]
- Guo W, Comai L, Henry IM (2021) Chromoanagenesis from radiation-induced genome damage in Populus. PLoS genetics 17 (8):e1009735.
- Liu J, Nannas NJ, Fu FF, Shi J, Aspinwall B, Parrott WA, Dawe RK (2019) Genome-Scale Sequence Disruption Following Biolistic Transformation in Rice and Maize. Plant Cell 31 (2):368-383. [CrossRef]
- Fossi M, Amundson K, Kuppu S, Britt A, Comai L (2019) Regeneration of Solanum tuberosum Plants from Protoplasts Induces Widespread Genome Instability. Plant Physiol 180 (1):78-86. [CrossRef]
- Itani OA, Flibotte S, Dumas KJ, Moerman DG, Hu PJ (2015) Chromoanasynthetic Genomic Rearrangement Identified in a N-Ethyl-N-Nitrosourea (ENU) Mutagenesis Screen in Caenorhabditis elegans. G3 (Bethesda) 6 (2):351-356. [CrossRef]
- Meier B, Cooke SL, Weiss J, Bailly AP, Alexandrov LB, Marshall J, Raine K, Maddison M, Anderson E, Stratton MR, Gartner A, Campbell PJ (2014) C. elegans whole-genome sequencing reveals mutational signatures related to carcinogens and DNA repair deficiency. Genome Res 24 (10):1624-1636. [CrossRef]
- Zhao Y, Yu L, Zhang S, Su X, Zhou X (2022) Extrachromosomal circular DNA: Current status and future prospects. Elife 11. [CrossRef]
- Notta F, Chan-Seng-Yue M, Lemire M, Li Y, Wilson GW, Connor AA, Denroche RE, Liang SB, Brown AM, Kim JC, Wang T, Simpson JT, Beck T, Borgida A, Buchner N, Chadwick D, Hafezi-Bakhtiari S, Dick JE, Heisler L, Hollingsworth MA, Ibrahimov E, Jang GH, Johns J, Jorgensen LG, Law C, Ludkovski O, Lungu I, Ng K, Pasternack D, Petersen GM, Shlush LI, Timms L, Tsao MS, Wilson JM, Yung CK, Zogopoulos G, Bartlett JM, Alexandrov LB, Real FX, Cleary SP, Roehrl MH, McPherson JD, Stein LD, Hudson TJ, Campbell PJ, Gallinger S (2016) A renewed model of pancreatic cancer evolution based on genomic rearrangement patterns. Nature 538 (7625):378-382. [CrossRef]
- Zack TI, Schumacher SE, Carter SL, Cherniack AD, Saksena G, Tabak B, Lawrence MS, Zhang CZ, Wala J, Mermel CH, Sougnez C, Gabriel SB, Hernandez B, Shen H, Laird PW, Getz G, Meyerson M, Beroukhim R (2013) Pan-cancer patterns of somatic copy number alteration. Nat Genet 45 (10):1134-1140. [CrossRef]
- Yates LR, Knappskog S, Wedge D, Farmery JHR, Gonzalez S, Martincorena I, Alexandrov LB, Van Loo P, Haugland HK, Lilleng PK, Gundem G, Gerstung M, Pappaemmanuil E, Gazinska P, Bhosle SG, Jones D, Raine K, Mudie L, Latimer C, Sawyer E, Desmedt C, Sotiriou C, Stratton MR, Sieuwerts AM, Lynch AG, Martens JW, Richardson AL, Tutt A, Lonning PE, Campbell PJ (2017) Genomic Evolution of Breast Cancer Metastasis and Relapse. Cancer Cell 32 (2):169-184 e167. [CrossRef]
- Stachler MD, Taylor-Weiner A, Peng S, McKenna A, Agoston AT, Odze RD, Davison JM, Nason KS, Loda M, Leshchiner I, Stewart C, Stojanov P, Seepo S, Lawrence MS, Ferrer-Torres D, Lin J, Chang AC, Gabriel SB, Lander ES, Beer DG, Getz G, Carter SL, Bass AJ (2015) Paired exome analysis of Barrett's esophagus and adenocarcinoma. Nat Genet 47 (9):1047-1055. [CrossRef]
- Shen H, Shih J, Hollern DP, Wang L, Bowlby R, Tickoo SK, Thorsson V, Mungall AJ, Newton Y, Hegde AM, Armenia J, Sanchez-Vega F, Pluta J, Pyle LC, Mehra R, Reuter VE, Godoy G, Jones J, Shelley CS, Feldman DR, Vidal DO, Lessel D, Kulis T, Carcano FM, Leraas KM, Lichtenberg TM, Brooks D, Cherniack AD, Cho J, Heiman DI, Kasaian K, Liu M, Noble MS, Xi L, Zhang H, Zhou W, ZenKlusen JC, Hutter CM, Felau I, Zhang J, Schultz N, Getz G, Meyerson M, Stuart JM, Cancer Genome Atlas Research N, Akbani R, Wheeler DA, Laird PW, Nathanson KL, Cortessis VK, Hoadley KA (2018) Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep 23 (11):3392-3406. [CrossRef]
- Dewhurst SM, McGranahan N, Burrell RA, Rowan AJ, Gronroos E, Endesfelder D, Joshi T, Mouradov D, Gibbs P, Ward RL, Hawkins NJ, Szallasi Z, Sieber OM, Swanton C (2014) Tolerance of whole-genome doubling propagates chromosomal instability and accelerates cancer genome evolution. Cancer Discov 4 (2):175-185. [CrossRef]
- Holland LZ, Ocampo Daza D (2018) A new look at an old question: when did the second whole genome duplication occur in vertebrate evolution? Genome Biol 19 (1):209. [CrossRef]
- Sacerdot C, Louis A, Bon C, Berthelot C, Roest Crollius H (2018) Chromosome evolution at the origin of the ancestral vertebrate genome. Genome Biol 19 (1):166. [CrossRef]
- Clark JW, Donoghue PCJ (2018) Whole-Genome Duplication and Plant Macroevolution. Trends Plant Sci 23 (10):933-945. [CrossRef]
- Steele CD, Abbasi A, Islam SMA, Bowes AL, Khandekar A, Haase K, Hames-Fathi S, Ajayi D, Verfaillie A, Dhami P, McLatchie A, Lechner M, Light N, Shlien A, Malkin D, Feber A, Proszek P, Lesluyes T, Mertens F, Flanagan AM, Tarabichi M, Van Loo P, Alexandrov LB, Pillay N (2022) Signatures of copy number alterations in human cancer. Nature 606 (7916):984-991. [CrossRef]
- Consortium ITP-CAoWG (2020) Pan-cancer analysis of whole genomes. Nature 578 (7793):82-93. [CrossRef]
- Drews RM, Hernando B, Tarabichi M, Haase K, Lesluyes T, Smith PS, Morrill Gavarro L, Couturier DL, Liu L, Schneider M, Brenton JD, Van Loo P, Macintyre G, Markowetz F (2022) A pan-cancer compendium of chromosomal instability. Nature 606 (7916):976-983. [CrossRef]
- Shen MM (2013) Chromoplexy: a new category of complex rearrangements in the cancer genome. Cancer Cell 23 (5):567-569. [CrossRef]
- Voronina N, Wong JKL, Hubschmann D, Hlevnjak M, Uhrig S, Heilig CE, Horak P, Kreutzfeldt S, Mock A, Stenzinger A, Hutter B, Frohlich M, Brors B, Jahn A, Klink B, Gieldon L, Sieverling L, Feuerbach L, Chudasama P, Beck K, Kroiss M, Heining C, Mohrmann L, Fischer A, Schrock E, Glimm H, Zapatka M, Lichter P, Frohling S, Ernst A (2020) The landscape of chromothripsis across adult cancer types. Nat Commun 11 (1):2320. [CrossRef]
- Durante MA, Rodriguez DA, Kurtenbach S, Kuznetsov JN, Sanchez MI, Decatur CL, Snyder H, Feun LG, Livingstone AS, Harbour JW (2020) Single-cell analysis reveals new evolutionary complexity in uveal melanoma. Nat Commun 11 (1):496. [CrossRef]
- Lukow DA, Sausville EL, Suri P, Chunduri NK, Wieland A, Leu J, Smith JC, Girish V, Kumar AA, Kendall J, Wang Z, Storchova Z, Sheltzer JM (2021) Chromosomal instability accelerates the evolution of resistance to anti-cancer therapies. Dev Cell 56 (17):2427-2439 e2424. [CrossRef]
- Yan X, Mischel P, Chang H (2024) Extrachromosomal DNA in cancer. Nat Rev Cancer 24 (4):261-273. [CrossRef]
- Davoli T, Uno H, Wooten EC, Elledge SJ (2017) Tumor aneuploidy correlates with markers of immune evasion and with reduced response to immunotherapy. Science 355 (6322). [CrossRef]
- Bakhoum SF, Ngo B, Laughney AM, Cavallo JA, Murphy CJ, Ly P, Shah P, Sriram RK, Watkins TBK, Taunk NK, Duran M, Pauli C, Shaw C, Chadalavada K, Rajasekhar VK, Genovese G, Venkatesan S, Birkbak NJ, McGranahan N, Lundquist M, LaPlant Q, Healey JH, Elemento O, Chung CH, Lee NY, Imielenski M, Nanjangud G, Pe'er D, Cleveland DW, Powell SN, Lammerding J, Swanton C, Cantley LC (2018) Chromosomal instability drives metastasis through a cytosolic DNA response. Nature 553 (7689):467-472. [CrossRef]
- Winick-Ng W, Kukalev A, Harabula I, Zea-Redondo L, Szabó D, Meijer M, Serebreni L, Zhang Y, Bianco S, Chiariello AM (2021) Cell-type specialization is encoded by specific chromatin topologies. Nature 599 (7886):684-691.
- Salari H, Di Stefano M, Jost D (2022) Spatial organization of chromosomes leads to heterogeneous chromatin motion and drives the liquid-or gel-like dynamical behavior of chromatin. Genome research 32 (1):28-43.
- Kim S, Peterson SE, Jasin M, Keeney S Mechanisms of germ line genome instability. In: Seminars in Cell & Developmental Biology, 2016. Elsevier, pp 177-187.
- Hattori A, Fukami M (2020) Established and novel mechanisms leading to de novo genomic rearrangements in the human germline. Cytogenetic and Genome Research 160 (4):167-176.
- Bhat P, Honson D, Guttman M (2021) Nuclear compartmentalization as a mechanism of quantitative control of gene expression. Nature Reviews Molecular Cell Biology 22 (10):653-670.
- Glaser J, Mundlos S (2022) 3D or not 3D: shaping the genome during development. Cold Spring Harbor Perspectives in Biology 14 (5):a040188.
- Akdemir KC, Le VT, Chandran S, Li Y, Verhaak RG, Beroukhim R, Campbell PJ, Chin L, Dixon JR, Futreal PA, Akdemir KC, Alvarez EG, Baez-Ortega A, Boutros PC, Bowtell DDL, Brors B, Burns KH, Campbell PJ, Chan K, Chen K, Cortés-Ciriano I, Dueso-Barroso A, Dunford AJ, Edwards PA, Estivill X, Etemadmoghadam D, Feuerbach L, Fink JL, Frenkel-Morgenstern M, Garsed DW, Gerstein M, Gordenin DA, Haan D, Haber JE, Hess JM, Hutter B, Imielinski M, Jones DTW, Ju YS, Kazanov MD, Klimczak LJ, Koh Y, Korbel JO, Kumar K, Lee EA, Lee JJ-K, Li Y, Lynch AG, Macintyre G, Markowetz F, Martincorena I, Martinez-Fundichely A, Meyerson M, Miyano S, Nakagawa H, Navarro FCP, Ossowski S, Park PJ, Pearson JV, Puiggròs M, Rippe K, Roberts ND, Roberts SA, Rodriguez-Martin B, Schumacher SE, Scully R, Shackleton M, Sidiropoulos N, Sieverling L, Stewart C, Torrents D, Tubio JMC, Villasante I, Waddell N, Wala JA, Weischenfeldt J, Yang L, Yao X, Yoon S-S, Zamora J, Zhang C-Z, Aaltonen LA, Abascal F, Abeshouse A, Aburatani H, Adams DJ, Agrawal N, Ahn KS, Ahn S-M, Aikata H, Akbani R, Akdemir KC, Al-Ahmadie H, Al-Sedairy ST, Al-Shahrour F, Alawi M, Albert M, Aldape K, Alexandrov LB, Ally A, Alsop K, Alvarez EG, Amary F, Amin SB, Aminou B, Ammerpohl O, Anderson MJ, Ang Y, Antonello D, Anur P, Aparicio S, Appelbaum EL, Arai Y, Aretz A, Arihiro K, Ariizumi S-i, Armenia J, Arnould L, Asa S, Assenov Y, Atwal G, Aukema S, Auman JT, Aure MRR, Awadalla P, Aymerich M, Bader GD, Baez-Ortega A, Bailey MH, Bailey PJ, Balasundaram M, Balu S, Bandopadhayay P, Banks RE, Barbi S, Barbour AP, Barenboim J, Barnholtz-Sloan J, Barr H, Barrera E, Bartlett J, Bartolome J, Bassi C, Bathe OF, Baumhoer D, Bavi P, Baylin SB, Bazant W, Beardsmore D, Beck TA, Behjati S, Behren A, Niu B, Bell C, Beltran S, Benz C, Berchuck A, Bergmann AK, Bergstrom EN, Berman BP, Berney DM, Bernhart SH, Beroukhim R, Berrios M, Bersani S, Bertl J, Betancourt M, Bhandari V, Bhosle SG, Biankin AV, Bieg M, Bigner D, Binder H, Birney E, Birrer M, Biswas NK, Bjerkehagen B, Bodenheimer T, Boice L, Bonizzato G, De Bono JS, Boot A, Bootwalla MS, Borg A, Borkhardt A, Boroevich KA, Borozan I, Borst C, Bosenberg M, Bosio M, Boultwood J, Bourque G, Boutros PC, Bova GS, Bowen DT, Bowlby R, Bowtell DDL, Boyault S, Boyce R, Boyd J, Brazma A, Brennan P, Brewer DS, Brinkman AB, Bristow RG, Broaddus RR, Brock JE, Brock M, Broeks A, Brooks AN, Brooks D, Brors B, Brunak S, Bruxner TJC, Bruzos AL, Buchanan A, Buchhalter I, Buchholz C, Bullman S, Burke H, Burkhardt B, Burns KH, Busanovich J, Bustamante CD, Butler AP, Butte AJ, Byrne NJ, Børresen-Dale A-L, Caesar-Johnson SJ, Cafferkey A, Cahill D, Calabrese C, Caldas C, Calvo F, Camacho N, Campbell PJ, Campo E, Cantù C, Cao S, Carey TE, Carlevaro-Fita J, Carlsen R, Cataldo I, Cazzola M, Cebon J, Cerfolio R, Chadwick DE, Chakravarty D, Chalmers D, Chan CWY, Chan K, Chan-Seng-Yue M, Chandan VS, Chang DK, Chanock SJ, Chantrill LA, Chateigner A, Chatterjee N, Chayama K, Chen H-W, Chen J, Chen K, Chen Y, Chen Z, Cherniack AD, Chien J, Chiew Y-E, Chin S-F, Cho J, Cho S, Choi JK, Choi W, Chomienne C, Chong Z, Choo SP, Chou A, Christ AN, Christie EL, Chuah E, Cibulskis C, Cibulskis K, Cingarlini S, Clapham P, Claviez A, Cleary S, Cloonan N, Cmero M, Collins CC, Connor AA, Cooke SL, Cooper CS, Cope L, Corbo V, Cordes MG, Cordner SM, Cortés-Ciriano I, Covington K, Cowin PA, Group PSVW, Consortium P (2020) Disruption of chromatin folding domains by somatic genomic rearrangements in human cancer. Nature Genetics 52 (3):294-305. [CrossRef]
- Lupiáñez DG, Kraft K, Heinrich V, Krawitz P, Brancati F, Klopocki E, Horn D, Kayserili H, Opitz JM, Laxova R (2015) Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell 161 (5):1012-1025.
- Tanabe H, Müller S, Neusser M, von Hase J, Calcagno E, Cremer M, Solovei I, Cremer C, Cremer T (2002) Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proceedings of the National Academy of Sciences 99 (7):4424-4429.
- Guerrero RF, Kirkpatrick M (2014) Local adaptation and the evolution of chromosome fusions. Evolution 68 (10):2747-2756.
- Matveevsky S, Tretiakov A, Kashintsova A, Bakloushinskaya I, Kolomiets O (2020) Meiotic nuclear architecture in distinct mole vole hybrids with Robertsonian translocations: Chromosome chains, stretched centromeres, and distorted recombination. International Journal of Molecular Sciences 21 (20):7630.
- Kneissig M, Keuper K, de Pagter MS, van Roosmalen MJ, Martin J, Otto H, Passerini V, Campos Sparr A, Renkens I, Kropveld F, Vasudevan A, Sheltzer JM, Kloosterman WP, Storchova Z (2019) Micronuclei-based model system reveals functional consequences of chromothripsis in human cells. Elife 8. [CrossRef]
- Guo W, Comai L, Henry IM (2021) Chromoanagenesis from radiation-induced genome damage in Populus. PLoS Genet 17 (8):e1009735. [CrossRef]
- Papathanasiou S, Markoulaki S, Blaine LJ, Leibowitz ML, Zhang CZ, Jaenisch R, Pellman D (2021) Whole chromosome loss and genomic instability in mouse embryos after CRISPR-Cas9 genome editing. Nat Commun 12 (1):5855. [CrossRef]
- Roosen M, Odé Z, Bunt J, Kool M (2022) The oncogenic fusion landscape in pediatric CNS neoplasms. Acta Neuropathologica 143 (4):427-451.
- van Belzen IA, Cai C, van Tuil M, Badloe S, Strengman E, Janse A, Verwiel ET, van der Leest DF, Kester L, Molenaar JJ (2023) Systematic discovery of gene fusions in pediatric cancer by integrating RNA-seq and WGS. BMC cancer 23 (1):618.
- Dai X, Guo X (2021) Decoding and rejuvenating human ageing genomes: Lessons from mosaic chromosomal alterations. Ageing Res Rev 68:101342. [CrossRef]
- Vera-Rodriguez M, Chavez SL, Rubio C, Pera RAR, Simon C (2015) Prediction model for aneuploidy in early human embryo development revealed by single-cell analysis. Nature communications 6 (1):7601.
- Vanneste E, Voet T, Le Caignec C, Ampe M, Konings P, Melotte C, Debrock S, Amyere M, Vikkula M, Schuit F (2009) Chromosome instability is common in human cleavage-stage embryos. Nature medicine 15 (5):577.
- Tuck-Muller CM, Chen H, Martínez JE, Shen CC, Li S, Kusyk C, Batista DA, Bhatnagar YM, Dowling E, Wertelecki W (1995) Isodicentric Y chromosome: cytogenetic, molecular and clinical studies and review of the literature. Hum Genet 96 (1):119-129. [CrossRef]
- Chandley AC, Ambros P, McBeath S, Hargreave TB, Kilanowski F, Spowart G (1986) Short arm dicentric Y chromosome with associated statural defects in a sterile man. Hum Genet 73 (4):350-353. [CrossRef]
- Iannuzzi A, Parma P, Iannuzzi L (2021) Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals (Basel) 11 (3). [CrossRef]
- Yang X, Breuss MW, Xu X, Antaki D, James KN, Stanley V, Ball LL, George RD, Wirth SA, Cao B, Nguyen A, McEvoy-Venneri J, Chai G, Nahas S, Van Der Kraan L, Ding Y, Sebat J, Gleeson JG (2021) Developmental and temporal characteristics of clonal sperm mosaicism. Cell 184 (18):4772-4783.e4715. [CrossRef]
- Lieberman-Aiden E, Van Berkum NL, Williams L, Imakaev M, Ragoczy T, Telling A, Amit I, Lajoie BR, Sabo PJ, Dorschner MO (2009) Comprehensive mapping of long-range interactions reveals folding principles of the human genome. science 326 (5950):289-293.
- Rao SS, Huntley MH, Durand NC, Stamenova EK, Bochkov ID, Robinson JT, Sanborn AL, Machol I, Omer AD, Lander ES (2014) A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 159 (7):1665-1680.
- Vara C, Paytuví-Gallart A, Cuartero Y, Le Dily F, Garcia F, Salvà-Castro J, Gómez HL, Julià E, Moutinho C, Aiese Cigliano R, Sanseverino W, Fornas O, Pendás AM, Heyn H, Waters PD, Marti-Renom MA, Ruiz-Herrera A (2019) Three-Dimensional Genomic Structure and Cohesin Occupancy Correlate with Transcriptional Activity during Spermatogenesis. Cell Rep 28 (2):352-367.e359. [CrossRef]
- Alavattam KG, Maezawa S, Sakashita A, Khoury H, Barski A, Kaplan N, Namekawa SH (2019) Attenuated chromatin compartmentalization in meiosis and its maturation in sperm development. Nat Struct Mol Biol 26 (3):175-184. [CrossRef]
- Ing-Simmons E, Rigau M, Vaquerizas JM (2022) Emerging mechanisms and dynamics of three-dimensional genome organisation at zygotic genome activation. Curr Opin Cell Biol 74:37-46. [CrossRef]
- Ghosh RP, Meyer BJ (2021) Spatial Organization of Chromatin: Emergence of Chromatin Structure During Development. Annu Rev Cell Dev Biol 37:199-232. [CrossRef]
- Oliver-Bonet M, Benet J, Sun F, Navarro J, Abad C, Liehr T, Starke H, Greene C, Ko E, Martin RH (2005) Meiotic studies in two human reciprocal translocations and their association with spermatogenic failure. Hum Reprod 20 (3):683-688. [CrossRef]
- Liu P, Carvalho CM, Hastings PJ, Lupski JR (2012) Mechanisms for recurrent and complex human genomic rearrangements. Curr Opin Genet Dev 22 (3):211-220. [CrossRef]
- Turner DJ, Miretti M, Rajan D, Fiegler H, Carter NP, Blayney ML, Beck S, Hurles ME (2008) Germline rates of de novo meiotic deletions and duplications causing several genomic disorders. Nat Genet 40 (1):90-95. [CrossRef]
- Bazykin AD (1969) HYPOTHETICAL MECHANISM OF SPECIATION. Evolution 23 (4):685-687. [CrossRef]
- Biesecker LG, Spinner NB (2013) A genomic view of mosaicism and human disease. Nat Rev Genet 14 (5):307-320. [CrossRef]
- Bertelsen B, Nazaryan-Petersen L, Sun W, Mehrjouy MM, Xie G, Chen W, Hjermind LE, Taschner PE, Tümer Z (2016) A germline chromothripsis event stably segregating in 11 individuals through three generations. Genet Med 18 (5):494-500. [CrossRef]
- Lindholm AK, Dyer KA, Firman RC, Fishman L, Forstmeier W, Holman L, Johannesson H, Knief U, Kokko H, Larracuente AM, Manser A, Montchamp-Moreau C, Petrosyan VG, Pomiankowski A, Presgraves DC, Safronova LD, Sutter A, Unckless RL, Verspoor RL, Wedell N, Wilkinson GS, Price TAR (2016) The Ecology and Evolutionary Dynamics of Meiotic Drive. Trends Ecol Evol 31 (4):315-326. [CrossRef]
- Pellestor F, Gatinois V, Puechberty J, Geneviève D, Lefort G (2014) Chromothripsis: potential origin in gametogenesis and preimplantation cell divisions. A review. Fertility and sterility 102 (6):1785-1796.
- Chiang C, Jacobsen JC, Ernst C, Hanscom C, Heilbut A, Blumenthal I, Mills RE, Kirby A, Lindgren AM, Rudiger SR, McLaughlan CJ, Bawden CS, Reid SJ, Faull RL, Snell RG, Hall IM, Shen Y, Ohsumi TK, Borowsky ML, Daly MJ, Lee C, Morton CC, MacDonald ME, Gusella JF, Talkowski ME (2012) Complex reorganization and predominant non-homologous repair following chromosomal breakage in karyotypically balanced germline rearrangements and transgenic integration. Nat Genet 44 (4):390-397, s391. [CrossRef]
- Pardo-Manuel de Villena F, Sapienza C (2001) Nonrandom segregation during meiosis: the unfairness of females. Mammalian Genome 12 (5):331-339. [CrossRef]
- Eisfeldt J, Pettersson M, Petri A, Nilsson D, Feuk L, Lindstrand A (2021) Hybrid sequencing resolves two germline ultra-complex chromosomal rearrangements consisting of 137 breakpoint junctions in a single carrier. Hum Genet 140 (5):775-790. [CrossRef]
- Hurst LD, Ellegren H (1998) Sex biases in the mutation rate. Trends Genet 14 (11):446-452. [CrossRef]
- Capilla L, Garcia Caldés M, Ruiz-Herrera A (2016) Mammalian Meiotic Recombination: A Toolbox for Genome Evolution. Cytogenet Genome Res 150 (1):1-16. [CrossRef]
- Romanenko SA, Perelman PL, Trifonov VA, Graphodatsky AS (2012) Chromosomal evolution in Rodentia. Heredity (Edinb) 108 (1):4-16. [CrossRef]
- Scriven PN, Handyside AH, Ogilvie CM (1998) Chromosome translocations: segregation modes and strategies for preimplantation genetic diagnosis. Prenat Diagn 18 (13):1437-1449.
- Zhang S, Lei C, Wu J, Sun H, Zhou J, Zhu S, Wu J, Fu J, Sun Y, Lu D, Sun X, Zhang Y (2018) Analysis of segregation patterns of quadrivalent structures and the effect on genome stability during meiosis in reciprocal translocation carriers. Hum Reprod 33 (4):757-767. [CrossRef]
- Mandáková T, Pouch M, Brock JR, Al-Shehbaz IA, Lysak MA (2019) Origin and Evolution of Diploid and Allopolyploid Camelina Genomes Were Accompanied by Chromosome Shattering. Plant Cell 31 (11):2596-2612. [CrossRef]
- Hochstenbach R, van Binsbergen E, Engelen J, Nieuwint A, Polstra A, Poddighe P, Ruivenkamp C, Sikkema-Raddatz B, Smeets D, Poot M (2009) Array analysis and karyotyping: workflow consequences based on a retrospective study of 36,325 patients with idiopathic developmental delay in the Netherlands. Eur J Med Genet 52 (4):161-169. [CrossRef]
- Bogdanov Yu F, Kolomiets OL, Lyapunova EA, Yanina I, Mazurova TF (1986) Synaptonemal complexes and chromosome chains in the rodent Ellobius talpinus heterozygous for ten Robertsonian translocations. Chromosoma 94 (2):94-102. [CrossRef]
- Volleth M, Heller KG, Yong HS, Müller S (2014) Karyotype evolution in the horseshoe bat Rhinolophus sedulus by whole-arm reciprocal translocation (WART). Cytogenet Genome Res 143 (4):241-250. [CrossRef]
- Nunes AC, Catalan J, Lopez J, Ramalhinho Mda G, Mathias Mda L, Britton-Davidian J (2011) Fertility assessment in hybrids between monobrachially homologous Rb races of the house mouse from the island of Madeira: implications for modes of chromosomal evolution. Heredity (Edinb) 106 (2):348-356. [CrossRef]
- Tapisso JT, Gabriel SI, Cerveira AM, Britton-Davidian J, Ganem G, Searle JB, Ramalhinho MDG, Mathias MDL (2020) Spatial and Temporal Dynamics of Contact Zones Between Chromosomal Races of House Mice, Mus musculus domesticus, on Madeira Island. Genes (Basel) 11 (7). [CrossRef]
- Potter S, Bragg JG, Blom MP, Deakin JE, Kirkpatrick M, Eldridge MD, Moritz C (2017) Chromosomal Speciation in the Genomics Era: Disentangling Phylogenetic Evolution of Rock-wallabies. Front Genet 8:10. [CrossRef]
- Giménez MD, Förster DW, Jones EP, Jóhannesdóttir F, Gabriel SI, Panithanarak T, Scascitelli M, Merico V, Garagna S, Searle JB, Hauffe HC (2017) A Half-Century of Studies on a Chromosomal Hybrid Zone of the House Mouse. J Hered 108 (1):25-35. [CrossRef]
- Chandley AC, McBeath S, Speed RM, Yorston L, Hargreave TB (1987) Pericentric inversion in human chromosome 1 and the risk for male sterility. J Med Genet 24 (6):325-334. [CrossRef]
- León-Ortiz AM, Panier S, Sarek G, Vannier JB, Patel H, Campbell PJ, Boulton SJ (2018) A Distinct Class of Genome Rearrangements Driven by Heterologous Recombination. Mol Cell 69 (2):292-305.e296. [CrossRef]
- Vara C, Paytuví-Gallart A, Cuartero Y, Álvarez-González L, Marín-Gual L, Garcia F, Florit-Sabater B, Capilla L, Sanchéz-Guillén RA, Sarrate Z (2021) The impact of chromosomal fusions on 3D genome folding and recombination in the germ line. Nature communications 12 (1):1-17.
- Pochon G, Henry IM, Yang C, Lory N, Fernández-Jiménez N, Böwer F, Hu B, Carstens L, Tsai HT, Pradillo M, Comai L, Schnittger A (2023) The Arabidopsis Hop1 homolog ASY1 mediates cross-over assurance and interference. PNAS Nexus 2 (3):pgac302. [CrossRef]
- Veller C, Kleckner N, Nowak MA (2019) A rigorous measure of genome-wide genetic shuffling that takes into account crossover positions and Mendel's second law. Proc Natl Acad Sci U S A 116 (5):1659-1668. [CrossRef]
- Lam I, Keeney S (2014) Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb Perspect Biol 7 (1):a016634. [CrossRef]
- Mattioli K, Oliveros W, Gerhardinger C, Andergassen D, Maass PG, Rinn JL, Melé M (2020) Cis and trans effects differentially contribute to the evolution of promoters and enhancers. Genome biology 21 (1):1-22.
- Pennisi E (2023) Stress responders. Science 381 (6660):825-829. [CrossRef]
Figure 1.
Types of chromoanagenesis.
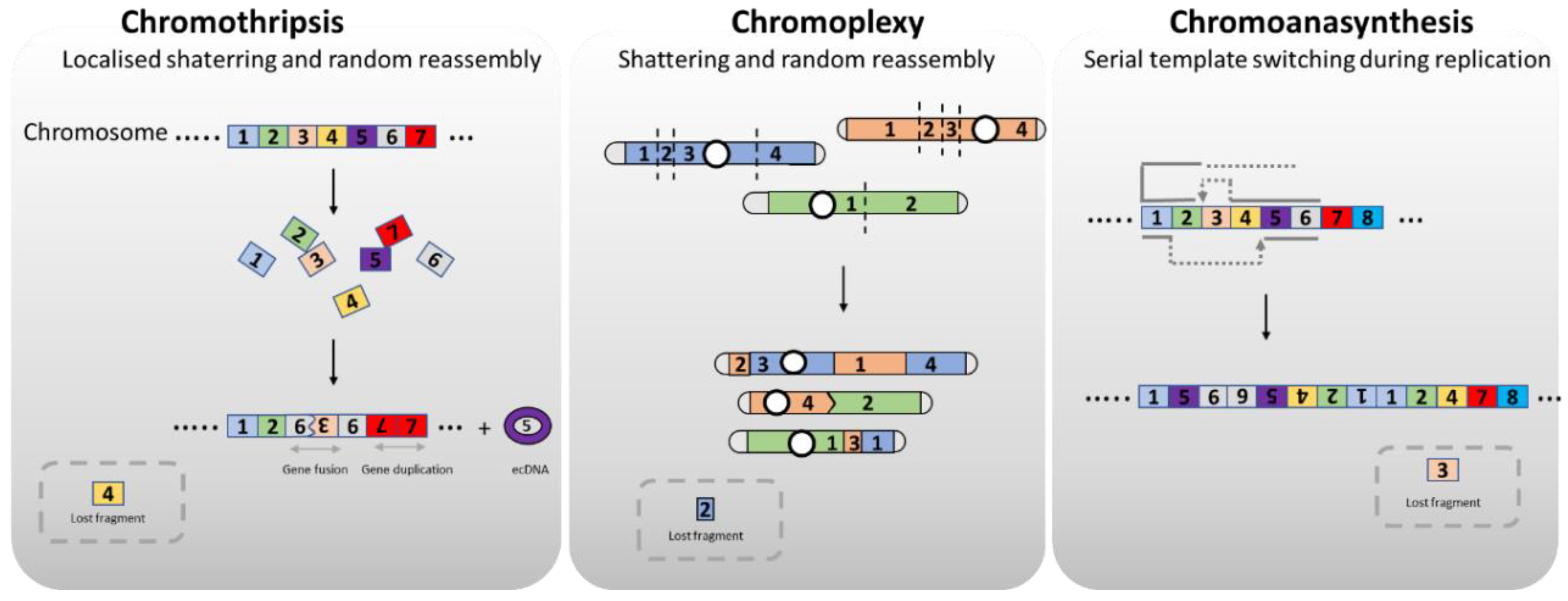
Figure 2.
Origin of chromoanagenesis. Error in cell division, which involve physical separation of chromosomes, can lead to the formation of micronuclei, a separate nucleus containing the lagging chromosome. A fragment of chromosome resulting from double-strand DNA damage can also lead to micronuclei formation. Micronuclei present a defective nuclear envelope that tend to disrupt. NE can also disrupt due to chromatin bridges. During nuclear envelope rupture, DNA is exposed to cytoplasmic DNAse or to the endonuclease TREX1 associate with the reticulum endoplasmic, resulting in chromosome fragmentation.
Figure 2.
Origin of chromoanagenesis. Error in cell division, which involve physical separation of chromosomes, can lead to the formation of micronuclei, a separate nucleus containing the lagging chromosome. A fragment of chromosome resulting from double-strand DNA damage can also lead to micronuclei formation. Micronuclei present a defective nuclear envelope that tend to disrupt. NE can also disrupt due to chromatin bridges. During nuclear envelope rupture, DNA is exposed to cytoplasmic DNAse or to the endonuclease TREX1 associate with the reticulum endoplasmic, resulting in chromosome fragmentation.
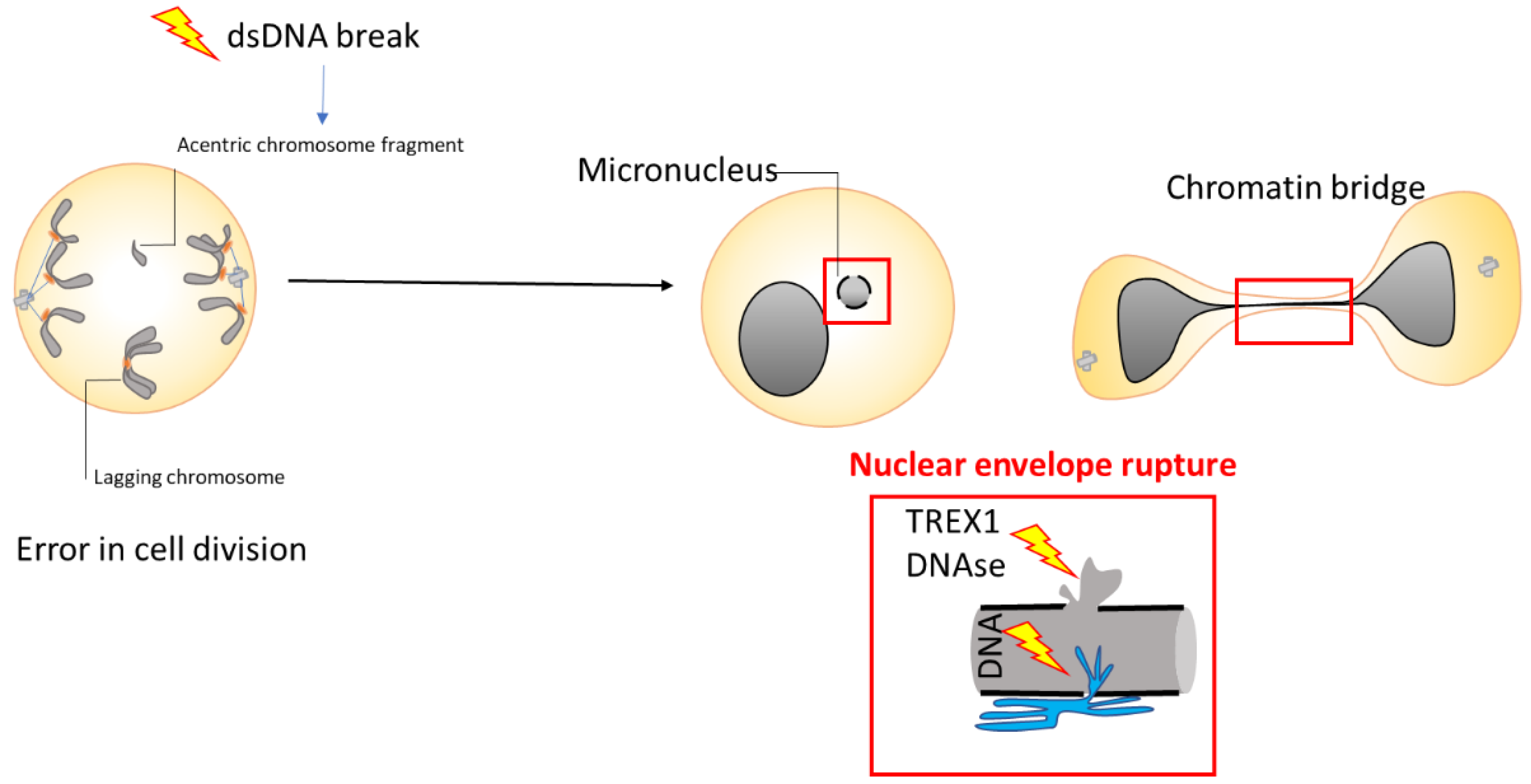
Figure 3.
DNA metabolism in the micronuclei. Micronuclei exhibit a nuclear envelope (NE) that lacks sufficient nuclear pore complexes and shows abnormal lamina formation. These abnormalities cause issues with nuclear protein import and frequent NE ruptures. When the NE ruptures, cytoplasmic components, as nuclease TREX1, invade the micronucleus, while soluble nuclear components and DNA can leak out.
Figure 3.
DNA metabolism in the micronuclei. Micronuclei exhibit a nuclear envelope (NE) that lacks sufficient nuclear pore complexes and shows abnormal lamina formation. These abnormalities cause issues with nuclear protein import and frequent NE ruptures. When the NE ruptures, cytoplasmic components, as nuclease TREX1, invade the micronucleus, while soluble nuclear components and DNA can leak out.
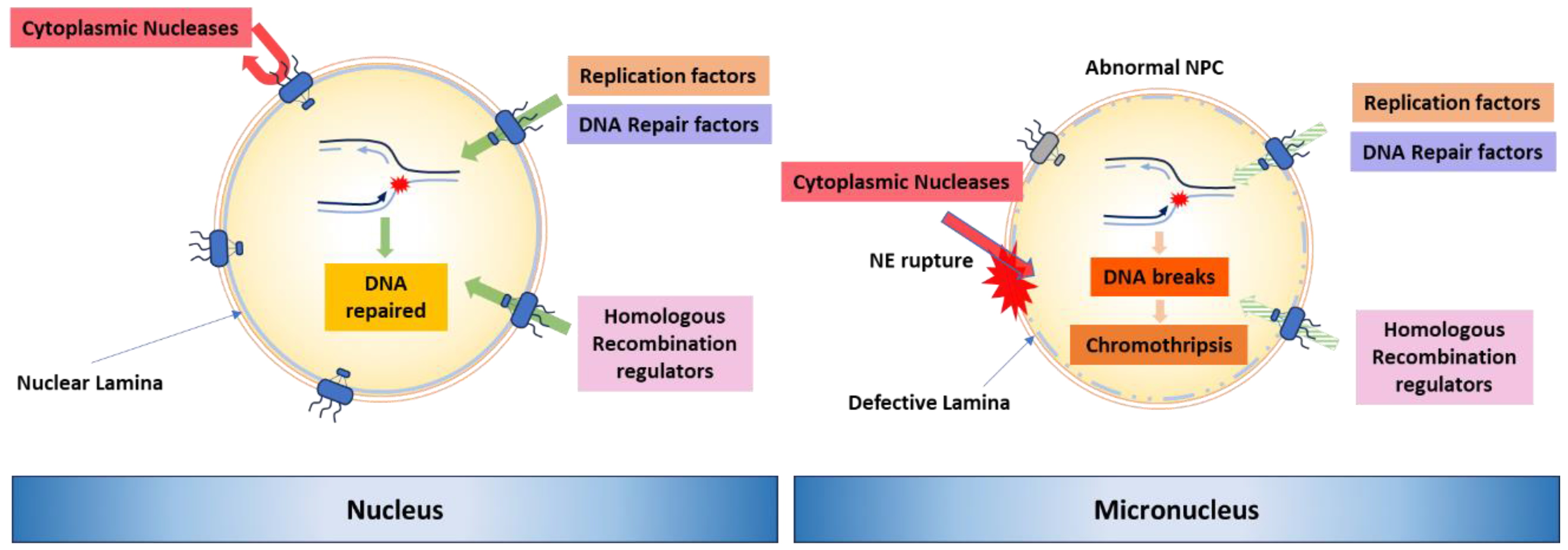
Figure 4.
Proposed mechanism for chromoanagenesis during meiosis. In some cases, chromosome fragmentation, synapsis defects, spindle failures, or chromosome missegregation leads to lagging chromosomes or acentric chromosome fragments during meiosis. During meiotic division II, these missegregated chromosomes or fragments become incorporated into a compartmentalized micronucleus. DNA damage mediation, non-homologous end joining (NHEJ) repair, and restitution of the micronucleus to the euploid pole nucleus can result in aneuploidy in the zygote or during embryogenesis. Alternatively, shattered chromosomes could result from chromothripsis (represented here) and chromoanasynthesis, due to the micronucleus failing to synchronize with the mitotic divisions of the main nucleus. Chromothripsis involves DNA fragmentation and random reassembly, while chromoanasynthesis involves replication fork collapses and microhomology-mediated strand switching. Subsequently, the pulverized and reassembled chromosome forms a single unit and can be meiotically inherited by the next generation, carrying forward the clustered structural variations.
Figure 4.
Proposed mechanism for chromoanagenesis during meiosis. In some cases, chromosome fragmentation, synapsis defects, spindle failures, or chromosome missegregation leads to lagging chromosomes or acentric chromosome fragments during meiosis. During meiotic division II, these missegregated chromosomes or fragments become incorporated into a compartmentalized micronucleus. DNA damage mediation, non-homologous end joining (NHEJ) repair, and restitution of the micronucleus to the euploid pole nucleus can result in aneuploidy in the zygote or during embryogenesis. Alternatively, shattered chromosomes could result from chromothripsis (represented here) and chromoanasynthesis, due to the micronucleus failing to synchronize with the mitotic divisions of the main nucleus. Chromothripsis involves DNA fragmentation and random reassembly, while chromoanasynthesis involves replication fork collapses and microhomology-mediated strand switching. Subsequently, the pulverized and reassembled chromosome forms a single unit and can be meiotically inherited by the next generation, carrying forward the clustered structural variations.
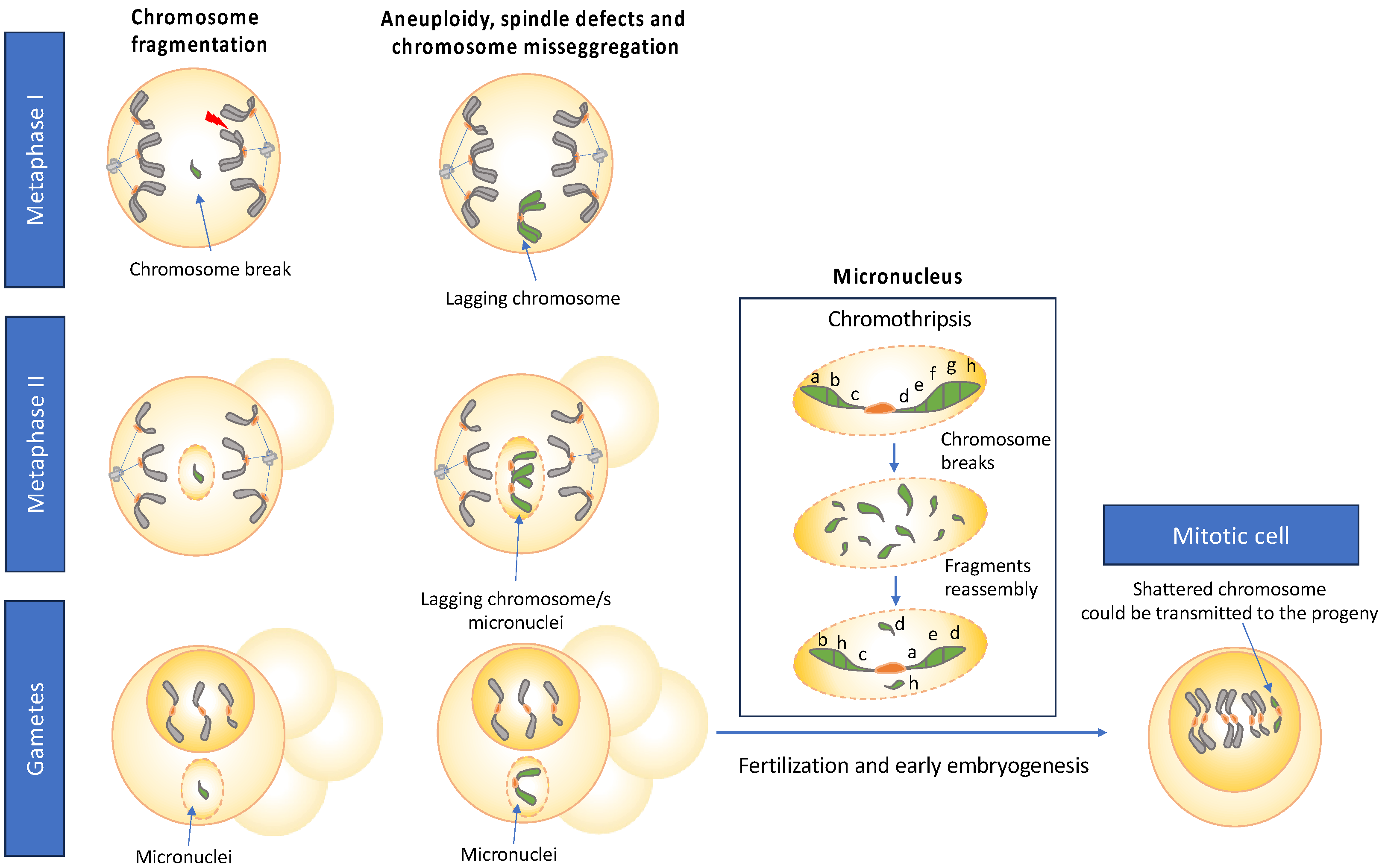
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



