
Submitted:
16 December 2024
Posted:
17 December 2024
You are already at the latest version
Abstract
The first immune response controls many bacterial and viral inflammatory diseases. Oral immunization with cholera toxin (CT) elicits antibodies and can prevent cholerae in endemic environments. While the IgG immune response to the toxin is well-documented, the IgA and IgM epitopes responsible for the initial immune reaction to the toxin remained uncharted. In this study, our objective was to identify and characterize immunologically and structurally these IgA and IgM epitopes. Materials and Methods: We conducted SPOT synthesis to create two libraries, each containing one hundred twenty-two 15-mer peptides, encompassing the entire sequence of the three chains of the CT protein. We could map continuous IgA and IgM epitopes by testing these membrane-bound peptides with sera from mice immunized with an oral vaccine (Schankol™). Our approach involved topological studies, peptide synthesis, and the development of an ELISA. Results: We successfully identified seven IgA epitopes, two in CTA, two in CTB, and three in protein P. Additionally, we discovered eleven IgM epitopes, all situated within CTA. Three IgA-specific and three IgM-specific epitopes were synthesized as MAP4 and validated using ELISA. We then used two chimeric 45-mer peptides, which included these six epitopes, to coat ELISA plates and screened them with sera from immunized mice. This yielded sensitivities and specificities of 100%. Conclusion: Our findings have unveiled a significant collection of IgA and IgM-specific peptide epitopes from cholera toxins A, B, and P. These epitopes, along with those IgG previously identified by our group, reflect the immunoreactivity associated with the dynamic of the immunoglobulins switching associated with the cholera toxin vaccination.
Keywords:
Cholera toxin
; IgA epitope
; IgM epitopes
; Peptide microarray
; Synthetic peptide
; MAPs
; ELISA-peptide
1. Introduction
The Gram-negative bacterium Vibrio cholerae is the causative agent of cholera, a life-threatening diarrheal disease primarily transmitted through contaminated food or water. This pathogen continues to pose a global threat, predominantly affecting developing nations with inadequate sewage treatment and clean water infrastructure, notably in Southeast Asia, Latin America, and parts of Africa. Annually, cholera leads to 1.3–4 million cases and 20,000–140,000 cholera-related fatalities worldwide, with nearly half of the victims being children under the age of five [1,2]. Furthermore, the expanding geographical reach of V. cholerae due to climate change and rising temperatures underscores the pressing need for further research to enhance our understanding of this pathogen [3,4].
V. cholerae strains responsible for disease outbreaks and pandemics fall under the O1 or O139 serogroups, while most V. cholerae is considered "non-O1/O139" environmental strains, which may or may not cause gastroenteritis [5,6]. Cholerae is highly contagious and is characterized by severe watery diarrhea, often leading to extreme dehydration, shock, and even death [6]. According to World Health Organization (WHO) reports, more than 80% of cholerae patients can be effectively treated with oral rehydration therapy [7].
The cholerae toxin (CT) is an 85 kDa protein in the AB class of bacterial toxins. It comprises the A subunit (CTA), a 28 kDa heterodimer, and the B subunit (CTB5), with a molecular weight of 55 kDa. CT is transported via cell receptors and converts ATP into cyclic AMP while transferring ADP-ribose from NAD (Nicotinamide Adenine Dinucleotide) to a specific arginine residue in the target protein (Gsa) [8]. This sequential process initiates adenyl cyclase activation, ultimately leading to increased intracellular cAMP levels, which, in turn, triggers the activation of cAMP-dependent protein kinase. This cascade results in protein phosphorylation, altered ion transport, and diarrhea with significant fluid loss [9]. Consequently, public health interventions are of utmost importance to limit the spread of cholera, particularly in regions lacking proper sanitation and clean water infrastructure, where outbreaks can easily occur.
Immunization through cholera vaccines is pivotal in controlling cholerae epidemics and pandemics. Key virulence factors of V. cholerae, including CT [10,11], toxin-coregulated pili [12], lipopolysaccharide (LPS) [13], and outer membrane proteins (Omps) [14,15,16], are considered ideal candidates for the development of cholera vaccines. Currently, two types of oral vaccines are in use: attenuated oral cholera vaccines (aOCVs) and inactivated oral cholera vaccines (kOCVs). aOCVs are composed of attenuated whole V. cholerae cells and have demonstrated direct effectiveness in protecting populations in endemic regions [17,18]. They have also been deployed in 'reactive' vaccination campaigns during outbreaks in non-endemic areas to contain the spread of cholerae [19,20]. In contrast, kOCVs provide herd protection for at least three years and offer short-term immunity with a single dose, which is valuable for outbreak management. However, their optimal efficacy necessitates two doses administered 14 days apart, and these vaccines require refrigeration [21]. These characteristics may limit the ability of kOCVs to swiftly control ongoing outbreaks in unstable or resource-limited settings. Nevertheless, single-dose live aOCVs have demonstrated satisfactory efficacy in challenge studies [22] and early-phase clinical trials in endemic regions [23]. Utilizing live aOCVs in reactive vaccination initiatives may be the most effective approach for reducing cholerae incidence during outbreaks [24,25].
While available oral-killed cholera vaccines are a valuable addition to control efforts, they may offer limited long-term protection, particularly in individuals with no prior exposure to the pathogen, such as children under five years of age, who bear a significant proportion of the global cholerae burden [26]. In contrast, cholerae survivors, including young children, develop high-level protective immunity that lasts three to four years [7,27].
Oral cholera vaccination effectively prevents outbreaks in high-risk areas and manages cholerae in endemic regions [28,29]. However, these vaccines may provide only limited and temporary protection, especially in individuals lacking prior exposure to the pathogen, such as children under five, who represent a significant portion of the global cholerae-affected population [30]. In contrast, cholerae survivors, including young children, develop high-level protective immunity that persists for years [31].
As the supply and utilization of aOCVs expand, crucial questions must be addressed regarding coverage and the vaccination schedule required to establish herd immunity [32]. Additionally, mucosal vaccines, such as nasal vaccines, can activate immune cells located in the mucosal tissues of the upper and lower respiratory tract. This dual immune system stimulation, combined with needle-free administration, supports the development of novel nasal vaccines designed to provide long-lasting immunity [33].
Previously, we characterized the secondary immune response following oral cholera vaccination, identifying eighteen IgG B linear epitopes, with six cross-reacting with several bacteria [34]. This study focuses on the primary IgA and IgM immune response epitopes induced by aOCVs with CT. Serological diagnostic assays can be used in large population studies [35] to determine the duration of the immune response, which can potentially be enhanced by identifying CT epitopes recognized by vaccine-induced antibodies. This advancement could improve our understanding of the duration of the immune response and herd protection.
Efficient high-content screening for epitope mapping involves screening peptide libraries representing target coding sequences. By synthesizing peptides directly onto cellulose membranes, it is possible to create refined maps of the entire epitome and numerous epitopes within a large protein [36]. SPOT synthesis analysis [37] was employed in this study to identify the IgA and IgM epitopes in three toxins from V. cholerae. Eighteen epitopes (7 IgA and 11 IgM) distinct from the previously identified IgG [34] were identified. ELISA selected and evaluated three of these as multiepitope peptides using sera from mice orally vaccinated.
2. Results
2.1. Identification of the Immunodominant IgA and IgM Epitopes in Cholera Toxin Subunits
In this section, we aimed to identify the immunodominant epitopes within the three components of the CT, namely CTA (258 aa), CTB (124 aa), and CTP (221 aa). To achieve this, we synthesized two libraries of peptides and assessed their recognition by antibodies from mice immunized with an oral V. choleraee vaccine (15 days post-vaccination). Figures 1A and 1C present the positions of these peptides and their respective measured intensities, as detected through chemiluminescent methods for mouse IgA and IgM antibodies present in the sera of mice that received the aOCV vaccination. Additionally, Figure 1B shows each peptide-epitope's hierarchical position in terms of its reactivity, arranged from the top left to the bottom right position. Epitopes were defined based on the overlap of spots with signal strengths exceeding 30%, establishing the minimum sequence for these epitopes. The comprehensive list of the synthetic peptides and their positions on the membrane is in Figure S1. This analysis identified 18 IgG epitopes, with sizes ranging from 4 to 14 amino acids, which were generated because of vaccination with aOCV (Table 1).
2.2. Spatial Localization of the Major IgA and IgM Epitopes Within the Three Chain of the Toxin
The seven IgA and eleven IgM linear epitopes identified by Spot-synthesis analysis were distributed throughout TxA, TxB, and TxP (Table 1). The enterotoxin chain A contains 2 IgA epitopes (TxA-1A and TxA-2A) and all 11 IgM epitopes (TxA-1M-TxA-11. Enterotoxin B houses 2 IgA epitopes (TxA-3A and TxA-4A) and no IgM epitopes. Enterotoxin P has three IgA epitopes (TxB-5A to TxB-7A) and no IgM epitopes (Table 1).
2.3. Spatial Distribution of the Reactive Epitopes of Enterotoxin A, B, and P
The analysis of the seven IgA and eleven IgM epitopes, as determined by the SPOT synthesis approach, revealed their distribution throughout the V. cholera toxin proteins (Figure 2). The V. cholera protein gene can be divided into three distinct segments: a short signal peptide in the N-terminal extension (aa 1–18), the A1 subunit chain segment (aa 19–212), the A2 subunit chain segment (aa 213–258), the B subunit (aa 1–124), and the P protein (aa 1–221).
To pinpoint the location of these epitopes, we accessed the enterotoxin A, B, and P structures from the Protein Data Bank (PDB) or utilized AlphaFold DB predictions. Alternatively, we deduced the tertiary structure using the I-TASSER server (http://zhanggroup.org/I-TASSER/, accessed on 11 May 2023). Figures 2A–C showcase the resulting predicted structural models for the enterotoxin A, B, and P proteins. These models offer insights into the spatial localization of the eight reactive epitopes in CTA, three epitopes in CTB, and seven epitopes in CTP. Notably, most of the identified epitopes were situated within loop/coil structures, making them accessible to the solvent and located on the protein's surface (Table 1 and Figure 2).
2.4. Specific and Cross-Immune IgA and IgM Epitopes
In exploring potential cross-immunity conferred by CT proteins, we employed CTA, CTB, and CTP sequences as templates in a multi-peptide matching search within the UniProtKB database. The criteria for this search involved the identification of peptides with four or more identical amino acids. Our analysis revealed that ten epitopes from V. cholera were specific (Table 1), while eight epitopes (TxA-1A, TxB-4A, TxP-5A, TxP-7A, TxA-1M, TxA-4M, TxA-5M, TxA-7M) exhibited cross-reactivity with E. coli or various bacteria. Specifically, the V. cholera-specific epitopes in CTA included TxA-2A, TxB-6A, TxA-2M, TxA-3M, TxA-6M, and TxA-8M to TxA-11M. Within CTP, only the epitope TxP-6A was identified as specific; in CTB, the specific epitope was TxB-3A. The epitopes TxP-5A and TxP-6A were determined to be exact within CTP (Table 1).
2.5. Reactivity of MAP4 and Chimeric Peptides via ELISA
It is well-known that serological assays based on bacterial proteins may exhibit cross-reactivity, underscoring the need to identify individual linear epitopes in the CT involved in the vaccination process. By restricting the antigens employed in an assay to capture reactive antibodies definitively specific to V. cholera through sequence analysis, developing a diagnostic ELISA that eliminates the potential for false positive results becomes feasible. To achieve this, three IgA and three IgM epitopes were carefully selected, synthesized, and subjected to ELISA analysis, both as individual peptides (Figure S3) and within multi-antigen peptides (MAP4) (Figure 3). A single IgA 45-mer and an IgM 45-mer chimeric peptide (Figure 3) encompassing the three chosen peptides were also synthesized. The sequences of the synthetic peptides are provided in Figure S2.
Upon examination, the sera from each immunized mouse showed reactivity to either the three synthetic peptides derived from IgA epitopes or the IgM epitopes, with no response to the negative control peptide QEVRKYFCV (Vaccinia virus; Figure 1A, G9 and G17). This reactivity was detected 15 days after vaccination (Figure 3A) and 30 days later (Figure 3B), with no significant difference in antibody levels observed when using single peptides versus MAPs. These results, as shown in Figure 3 and Figure S3, were found to be statistically significant (p < 0.001).
Based on ROC curve analysis (Figure S4), the Area under the curve (AUC) for the epitope Tx45-IgA and Tx45-IgM was 0.9987 and 0.9896 (p < 0.0001), as detected by ELISA with a confidence interval of 95%, demonstrating high diagnostic accuracy for both the peptides. The Tx45-IgM peptide presented a lower reactivity than the Tx45-IgA, but both demonstrated 100% sensitivity and specificity.
3. Discussion
In this study, we identified linear B epitopes in CT recognized by IgA and IgM antibodies from mice vaccinated with an inactivated vaccine using peptide microarray analysis. CT serves as the primary virulence factor of V. cholerae, capable of inducing cholera symptoms with as little as 5 µg ingestion [38]. The serum reactivity against specific regions in CT demonstrated the vaccine's high antigenic potential with a single dose. We identified two IgA and eleven IgM linear epitopes in CTA, two in CTB, and three in protein CTP, all accessible to the immune system due to their exposure on the molecular surface.
CTA is initially synthesized as a single polypeptide with a signal peptide (aa1-18). The enzymatic activity begins after proteolytic cleavage, "nicking" at residue Arg-192, resulting in two functional domains: A1 (~21.8 kDa, aa 19–204) and A2 (~5.4 kDa, aa 206–258), covalently linked by a disulfide bond. The B subunit comprises five chains forming a pentameric ring around a central pore structure. CTA1 is responsible for ADP-ribosyltransferase activity, while the α-helical CTA2 anchors CTA1 and CTB5 together [39,40,41]. Despite the extensive potential for robust IgG response in CTB, our study did not confirm this, as only two IgA epitopes were identified among the seven detected.
Recent studies have shown that IgG anti-CTA antibodies can neutralize the toxin at very low concentrations [9], attributed to the cross-reactivity of CTA epitopes [42,43]. However, none of these IgG sequences [34] are shared with the IgA or IgM epitopes identified in this study.
During the immune response, isotype change is a protective property that has the function of the organism producing antibodies with greater affinity to neutralize the antigen. After antigen recognition, recombination occurs between DNA segments of the genes that encode the constant regions of the heavy chains, determining the isotype [44]. The production time of these isotypes is variable and appears to depend on the antigen. Consequently, some types of antibodies have a short half-life while others take a long time to disappear from circulation. Therefore, it is important to know which epitopes induce antibodies of longer duration and neutralizing capacity [44].
In mouse models, it has been demonstrated that the primary immune response to cholerae is dominated by IgG against CTB, which is protective [45,46]. However, our study indicates that CTB also elicits IgA and IgM responses, but these responses appear weak compared to the number of identified IgG epitopes [34], therefore these responses may be of very short duration. On the other hand, no detailed studies on the immunogenic importance of CTP immunoglobulin isotypes were found.
Each isotype is designed to tackle specific challenges, especially involving affinity. Normally, the IgM is the first antibody to emerge during embryonic development, and the humoral immune response is followed by IgA and IgG [47,48]. However, some IgM is also produced in secondary and subsequent responses, and after somatic hypermutation, although other isotypes dominate the later phases of the antibody response. The IgM can exist under distinct forms, including monomeric, membrane-bound IgM within the B cell receptor (BCR) complex, pentameric and hexameric IgM in serum can provide effective anti-microbial immunity [49,50], and secretory IgM on the mucosal surface. However, the regulation of humoral Immune responses and B cell tolerance by the IgM Fc Receptor (FcμR) is poorly understood [41,51].
Each isotype might prefer certain epitopes or residues of amino acids, which may be better suited to deal with specific types of invaders [52]. However, these epitopes possess different residues (right or left) but shared residues indicating distinct response preferences and evolution of the immune response, providing immunoglobulins with a higher binding or neutralization ability. Therefore, changes in the affinity epitopes will allow the immune system to be highly adaptable and effective in fine tuning, ready to handle diverse threats or situations [53].
Several IgG rapid antigen detection-based tests are available for cholerae diagnosis, such as immunochromatographic lateral flow devices like Crystal VC and Cholkit, which detect the presence of the O1 or O139 antigen in watery diarrheal stools and others [54,55,56]. However, limitations exist, and these tests may only sometimes offer high specificity [46,57]. Molecular testing using polymerase chain reaction (PCR) has also been used for diagnosis but primarily for research and surveillance [58,59]. Darkfield microscopy is another diagnostic approach for detecting V. cholerae, but it needs the required sensitivity for reliable diagnosis.
Thus, more precise, rapid diagnostic assays are needed. The IgA and IgM epitopes identified in this study can be applied in an ELISA assay to identify the primary immune response and provide insights into serological immunity. These unique IgA and IgM epitopes hold the potential for developing chimeric polyproteins for diagnostic purposes.
Vaccine production is typically a labor-intensive and costly experimental process. Nonetheless, these immunobiological molecules/agents are the most potent defenses against infectious diseases.
Presently, there are three pre-qualified oral cholera vaccines (OCV): Dukoral® Crucell, Netherlands), Shanchol™ (Shantha, Biotechnics-Sanofi Pasteur, India), and Euvichol-Plus® (EuBiologics Co., Seoul, South bbbgb, Korea). While both categories have demonstrated protective effects against cholerae [4,60,61,62], the immunological mechanisms involving B-cell and/or T-cell immunity remain unclear [63,64]. It is well established that the key stimulator of innate immunity and subsequent adaptive immune responses is the LPS O antigen [6,10], which promotes long-term mucosal protection [8,13,65,66,67]. Hence, the pressing question is how to create more effective cholera vaccines, as current vaccine efficacies hover around 60% [27,68]. Therefore, identifying immunodominant epitopes that interact with antibodies generated in response to vaccination can contribute to selecting better antigens and improve our understanding.
4. Conclusions
A resolutive assessment of the immune response was conducted using a peptide array that was directly generated on a cellulose membrane. This approach facilitated the identification of the major antigenic determinants in enterotoxins A, B, and P, which were recognized by antibodies from mice that had been orally vaccinated with a single dose of the aOCV. In total, we identified seven IgA and eleven IgM epitopes that were distributed throughout the bacterial CT protein. Specifically, six epitopes were in the A1 chain of the enterotoxin, two in the A2 chain, three in enterotoxin B, and seven in enterotoxin P. Notably, six of these epitopes exhibited a degree of similarity to proteins in other pathogens, indicating a high potential for cross-reactivity. Furthermore, we defined ten V. cholera-specific epitopes (3 IgA and 7 IgM) and elucidated their spatial locations within protein structural models. The efficacy of three of these IgA epitopes (TxA-2A, TxB-3A, and TxP-6A) and three IgM epitopes (TxA-2M, TxA-3M, and TxA-6M) in detecting antibodies produced in response to vaccination in mice was confirmed using an indirect ELISA. This comprehensive molecular characterization of linear IgA and IgM epitopes in CT holds significant promise for developing a poly epitope chimeric protein, which could be employed in next-generation rapid diagnostic tests. Additionally, our findings underscore the value of epitope mapping in enhancing our understanding of the immune response to current and future vaccines, ultimately leading to improved production of specific neutralizing antibodies.
5. Materials and Methods
5.1. Immunization of Mice
We orally immunized thirty Balb Cmice, each weighing between 15 and 21 g, with 20 µL of the SchancolTM cholerae vaccine (lot SCN021A15; Shantha Biotechnics Ltd. in Muppiriddipalli, Telangana, India). Fifteen days following vaccination, we collected blood samples and performed a complete bleed at the 30-day mark [34]. In parallel, blood samples were collected from 30 healthy, unvaccinated mice to serve as controls. The serum from each group was separately collected, divided into 0.5 mL aliquots in Eppendorf tubes, and stored at -20 °C.
5.2. Synthesis of the Cellulose Membrane-Bound Peptide Array
Two libraries encompassing 122 peptides, each spanning the entire coding sequences of cholera enterotoxin A (P01555, 258 amino acids), enterotoxin B (P01556, 124 amino acids), and Toxin P (P29485, 221 amino acids) of serotype O1 was synthesized on amino-PEG500-UC540 cellulose membranes (Intavis Bioanalytical Instruments, Köln, Germany) following standard SPOT synthesis protocols [70].
These peptides, each comprising 15 amino acids in length with a ten-amino-acid overlap, were prepared using an Auto-Spot Robot ASP-222 (Intavis Bioanalytical Instruments AG, Köln, Germany). To enhance the libraries, we introduced a GSGSG spacer sequence at each protein's amino and carboxy termini (designated as points A1, D5, D6, C4, C5, and F2). The library construction and program execution were carried out using the MultiPep program (Intavis). In addition to the libraries, a negative control peptide QEVRKYFCV (from Vaccinia virus, located at spots G9 and G17) was included, as well as positive control peptides, such as KEVPALTAVETGATN (from Poliovirus, at spots G3 and G11), GYPKDGNAFNNLDRI (from Clostridium tetani, at spots G5 and G13), and YDYDVPDYAGYPYDV (from the hemagglutinin of Influenza virus, located at spots G7, G15, and G24).
Coupling reactions were followed by a blocking step involving acetylation with acetic anhydride (4%, v/v) in N, N-dimethylformamide. The F-moc protecting group was removed from the N-terminus of each peptide by introducing a 20% piperidine solution in DMF. This cycle of coupling, blocking, and deprotection was repeated for each successive amino acid until the final peptide was produced. After the addition of the last amino acid, the side chains of the amino acids were deprotected using a dichloromethane-trifluoroacetic acid-triisopropylsilane solution (1:1:0.05, v/v/v), followed by ethanol washing. The membranes containing the synthetic peptides were then immediately subjected to probing.
5.3. Screening of SPOT Membranes
The SPOT membranes underwent a 10-min wash with TBS-T (50 mM Tris, 136 mM NaCl, 2 mM KCl, and 0.05% Tween® 20, pH 7.4), followed by blocking in TBS-T containing 1.5% BSA for 90 min at 8°C with agitation. After thorough TBS-T washing, the membranes were incubated for 12 h with a pool of sera from ten vaccinated mice, diluted (1:150 for IgA or IgM detection) in TBS-T with 0.75% BSA. Subsequent washes with TBS-T were carried out. The membranes were then incubated with goat anti-mouse IgM and goat anti-mouse IgA alkaline phosphatase labeled (Sigma Chemical Co, Saint Louis, MO, U.S.A.; diluted 1:5000) for 1 h, prepared in TBS-T with 0.75% BSA, and washed with TBS-T and CBS (50 mM citrate-buffered saline). Finally, the chemiluminescent substrate Nitro-Block II Enhance (Applied Biosystems, Waltham, MA, USA) was added to complete the reaction. The integrity of the library synthesis was confirmed through the reactivity of the control peptides to human sera.
5.4. Scanning and Measurement of Spot Signal Intensities
Chemiluminescent signals were detected using an Odyssey FC (LI-COR Bioscience, Lincoln, NE, USA) with minor modifications to previously described conditions [71]. A digital image file was generated at a 5 MP resolution, and signal intensities were quantified using TotalLab TL100 (v 2009, Nonlinear Dynamics, Newcastle-Upon-Tyne, UK) software. The signal intensities (SI) were exported to Excel (Microsoft Corp., Redmond, WA, USA), where background signals (signal at the negative control) were subtracted. SI values were then normalized as a percentage of the highest measured signal. An epitope was defined in the sequences of two or three contiguous spots presenting a normalized SI of 30% or greater. For epitopes involving three or more contiguous spots with SI values of 30% or higher, identification was based on visual assessment.
According to the approach described previously, a comparative analysis of the reactivity index of the Spots normalized on a dimensional hierarchical level was realized [71].
5.5. Preparation of Single and Multi-Antigen Peptides (MAPs)
The single and multi-antigen peptides were synthesized following the standard solid-phase synthesis protocol [73]. The Wang and tetrameric Fmoc2-Lys-B-Ala Wang resins were used for the synthesis (Figure S3). These constructs were produced in an automated peptide synthesizer (MultiPep-1, CEM Corp, Charlotte, NC, USA). The side chains of tetrafunctional F-moc amino acids were protected with TFA-labile protecting groups as needed. After the assembly of the peptide sequences, the F-moc groups were removed, and the peptide resin was cleaved and fully deprotected using TFA/H2O/EDT/ TIS (94/2.5/2.5/1.0, v/v) for 90 min. The peptides were then precipitated by adding chilled diethyl ether and centrifugation (30,000× g, 10 min at 4 °C). The resulting pellet was dissolved in aqueous AcOH (10% v/v), dried, and stored as a lyophilized powder. When required, the MAPs were dissolved in water and subjected to centrifugation (10,000× g, 60 minutes at 15 °C), and the supernatant was filtered through a Centricon™ (Merck Millipore, Burlington, MA, USA) ten filter. The MAPs were purified using an XBridge BEH C18 (2.7 μ, 5 cm × 4.6 mm) column coupled to a water auto-purify HPLC system (Water Corporation, Newcastle, NS, Australia). This purification was performed at a flow rate of 1.2 mL/min, using mobile phases A [0.05% formic acid in water (18 MΩ × cm)] and B [0.05% formic acid in ACN (acetonitrile acid)] (v/v) in water with a gradient of 0-97 B over 40 min. During the purification, the detection was carried out at 200–300 nm using a diode array.
For the ESI-TOF analysis, the peptides were dissolved in deionized water until a final concentration of 10 µg/mL was reached. Subsequently, formic acid was added to achieve a final concentration of 0.1%. The mass spectrometer used for this analysis was the Water UPLC model Acquity-I Class (Water Corp., Newcastle, Australia), with sample injections performed at 1 µL/min. The ion detection range covered values between 1,000 and 11,500 m/z.
5.6. Preparation of 45-mer Chimeric Peptides
Two chimeric multiepitope peptides (Tx45-IgA and Tx45-IgM) were synthesized sequentially and contained two to six glycine spacers (Figure S3). The F-moc protocol and TentaGel-S-NH2 resin [73] were used for the synthesis. An automated peptide synthesizer (MultiPep-1, CEM Corp, Charlotte, NC, USA) was employed, along with F-moc amino acids and necessary protecting groups. After synthesis, the peptides were cleaved, deprotected, and precipitated. Their identities were confirmed through mass spectrometry (MALDI-TOF or electrospray).
5.7. In House ELISA
Peptide-based ELISA (pELISA) assays were conducted as previously described [73]. Briefly, 4HB NUNC plates for ELISAs were coated with 80 ng of each peptide in 100 µL of coating buffer (Na2CO3–NaHCO3, pH 9.6) overnight at 4 °C. The plates underwent a series of steps, including three washes with PBS-T washing buffer (PBS with 0.1% Tween® 20, pH 7.2). They were then blocked in PBS-T containing 2.5% BSA for 2 h at 37 °C. Subsequently, 100 µL of diluted mouse sera in coating buffer was applied and incubated for an additional 2 h at 37 °C. After thorough washing with PBS-T, the plates were subjected to a 2-h incubation at 37 °C with 100 µL of goat anti-mouse IgA-HRP (#62-672) or goat anti-mouse IgM-HPP (#PAL 84383; Life Technology, Carlsbad, California, U.S.A.), and diluted in coating buffer. Another round of washing was performed, followed by exposure to the p-nitro phenyl phosphate (pNPP) substrate (Sigma, St. Louis, MO, USA). Absorbance was measured at 405 nm on a Hidex Sense Microplate Reader, Turku, Finland). The optimal serum dilution value of 1:100 was initially determined by a titration series of vaccinated mouse sera and the construction of the receiver operating characteristic (ROC) curve.
5.8. Structural Localization of IgG Epitopes and Bioinformatics Tools
To determine the location of the epitopes within the 3D molecular structure of enterotoxin A, B, and P proteins from V. cholerae, in silico protein models were constructed using the I-TASSER server (http://zhanglab.ccmb, accessed on 10 April 2023). Models were selected based on the best C-score and TM-score (topological evaluation value) [74]. The resulting 3D structural models were also analyzed in the AlphaFold database [75]. Searches for V. cholerae sequence homologies were performed using previously identified sequences and annotated proteins from other organisms in the UniProt database (http://www.uniprot.org/, accessed on 12 April 2023) and multiple peptide match (https://research.bioinformatics.udel.edu/peptide match/index.jsp, accessed on 10 May 2023).
5.9. Statistical Analysis
The data were analyzed using the R program (version 3.6.0) and R Studio. To assess the statistical significance between the two samples, a paired t-test was employed, with significance considered at p ≤ 0.05. GraphPad Prism version 5.0 was used to analyze the receiver operating characteristic curve.
Supplementary Materials
The following supporting information can be downloaded at website of this paper post on Preprints.org, Figure S1: List of synthesized peptides covering the entire sequence of Vibrio cholera toxin A (P01555), toxin B (P01556); Figure S2: List of single (TxA2-A, TxB3-A, Tx-P-6A), multi-antigen peptides (MAP4) (TxA-2M, TxA-3M, TxA-6M) and tripeptides synthetic peptides (Tx45-IgA, Tx45-IgM) and physicochemical properties.; Figure S3: ELISA using as antigen the Tx45-IgA-peptide (A, B) and Tx45-IgM-peptide (C, D) derived cholerae proteins and sera of mice immunized with an oral vaccine (15 days after vaccination (A, C) and 30 days after vaccination (B, D).
Author Contributions
Conceptualization, S.G.D.-S. and F.R.d.S.; methodology, P.N.-P., F.R.d.S., P.S.G., SVC, A.O.S. and GCL; validation, S.G.D.-S.; writing-original draft, S.G.D.-S.; writing-review and editing, S.G.D.-S.; funding acquisition, S.G.D.-S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brazilian Council for Scientific Research (CNPq #30515- 2020-5) and the Carlos Chagas Filho Foundation of Research Support of the State of Rio de Janeiro (FAPERJ #200.960-2022).
Institutional Review Board Statement
The study was approved by the UNIGRANRIO (CEUA 052/2021) study center ethics committee and conducted under good clinical practice and all applicable regulatory requirements, including the Declaration of Helsinki.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the National Institute of Quality Control (INCQs) from FIOCRUZ for their support with the vaccine and with animal immunization, and Dr. Ulisses C Araujo from the Multiuser Research Laboratory in Mass Spectrometry applied to Biomolecules (MultiMasses/ IBRAG/UERJ). P.N.-P. and G.C.L. are postdoc fellows from the CAPES-FIOCRUZ program. P.S.G. is a CAPES fellow from the Post-Graduation Program on Science and Biotechnology at Federal Fluminense University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ali, M.; Nelson, A.R.; Lopez, A.L. ; Sack, DA Updated global burden of cholera in endemic countries. PLoS Negl Trop Dis 2015, 9, e0003832. [Google Scholar] [CrossRef]
- Yuki, Y.; Nojima., M.; Hosono, O.; Tanaka, H.; Kimura, Y.; Satoh, T.; Imoto, S.; Uematsu, S.; Kurokawa, S.; Kashima, K.; et al. Oral MucoRice-CTB vaccine for safety and microbiota-dependent immunogenicity in humans: A phase 1 randomized trial. Lancet Microbe 2021, 2, e429–e440. [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc Natl Acad Sci USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Deeb, R.; Tufford, D.; Scott, G.I.; Moore, J.G.; Dow, K. Impact of climate change on Vibrio vulnificus abundance and exposure risk. Estuaries Coasts 2018, 41, 2289–2303. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. Infections. Nat Rev Dis Prim 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Legros, D. Legros, D.; Partners of the Global Task Force on Cholerae Control. Global cholerae epidemiology: Opportunities to reduce the burden of cholera by 2030. J Infect Dis 2018, 218, S137–S140; Erratum in J Infect Dis 2019, 219, 509. [CrossRef]
- WHO. Cholerae; World Health Organization: Geneva, Switzerland, 2022. https://www.who.int/news-room/fact-sheets/detail/cholera? gadsource =1&gclid=EAIaIQobChMIqYXdh PW1hgMVgUVIAB0qbwipEAAYASAAEgJ 1CfD_BwE (Accessed on 21 March 2023).
- Banerjee, T.; Grabon, A.; Taylor, M.; Teter, K. cAMP-Independent activation of the unfolded protein response by cholera toxin. Infect Immun 2021, 89, e00447–e20. [Google Scholar] [CrossRef] [PubMed]
- Clemens, J.D.; Nair, G.B.; Ahmed, T.; Qadri, F.; Holmgren, J. Cholerae. Lancet 2017, 390, 1539–1549. [Google Scholar] [CrossRef]
- Bourque, D.L.; Bhuiyan, T.R.; Genereux, D.P.; Rashu, R.; Ellis, C.N.; Chowdhury, F.; Khan, A.I.; Alam, N.H.; Paul, A.; Hossain, L.; et al. Analysis of the human mucosal response to cholerae reveals sustained activation of innate immune signaling pathways. Infect Immun 2018, 86, e00594–17. [Google Scholar] [CrossRef]
- Sánchez, J.; Holmgren, J. Cholera toxin-a foe & a friend. Indian J. Med. Res. 2011, 133, 153. [Google Scholar]
- Taylor, R.K.; Miller, V.L.; Furlong, D.B.; Mekalanos, J.J. Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc Natl Acad Sci. USA 1987, 84, 2833–2837. [Google Scholar] [CrossRef]
- Waldor, M.K.; Colwell, R.; Mekalanos, J.J. The Vibrio cholerae O139 serogroup antigen includes an O-antigen capsule and lipopolysaccharide virulence determinants. Proc Natl Acad Sci USA 1994, 91, 11388–11392. [Google Scholar] [CrossRef]
- Sperandio, V.; Giron, J.A.; Silveira, W.D.; Kaper, J.B. The OmpU outer membrane protein, a potential adherence factor of Vibrio cholerae. Infect Immun 1995, 63, 4433–4438. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.K.; Sengupta, T.K.; Ghose, A.C. Major outer membrane proteins of Vibrio cholerae and their role in induction of protective immunity through inhibition of intestinal colonization. Infect Immun 1992, 60, 4848–4855. [Google Scholar] [CrossRef]
- Nandi, B.; Nandy, R.K.; Sarkar, A.; Ghose, A.C. Structural features, properties, and regulation of the outer-membrane protein W (OmpW) of Vibrio cholerae. Microbiology 2005, 151, 2975–2986. [Google Scholar] [CrossRef]
- Qadri, F.; Ali, M.; Chowdhury, F.; Khan, A.I.; Saha, A.; Khan, I.A.; Begum, Y.A.; Bhuiyan, T.R.; Chowdhury, M.I.; Uddin, M.J.; et al. Feasibility and effectiveness of oral cholerae vaccine in an urban endemic setting in Bangladesh: A cluster randomized open-label trial. Lancet 2015, 386, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Bi, Q.; Ferreras, E.; Pezzoli, L.; Legros, D.; Ivers, L.C.; Date, K.; Qadri, F.; Digilio, L.; Sack, D.A.; Ali, M.; et al. Protection against cholera from killed whole-cell oral cholerae vaccines: A systematic review and meta-analysis. Oral cholerae vaccine working group of the global task force on cholerae control. Lancet Infect Dis 2017, 17, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Peak, CM; Reilly, A.L.; Azman, AS; Buckee, C.O. Prolonging herd immunity to cholerae via vaccination: Accounting for human mobility and waning vaccine effects. PLoS Negl Trop Dis 2018, 12, e0006257. [CrossRef]
- Royal, J.M.; Reeves, M.A.; Matoba, N. Repeated oral administration of a KDEL-tagged recombinant cholerae toxin B subunit effectively mitigates DSS colitis despite a robust immunogenic response. Toxins 2019, 11, 678. [Google Scholar] [CrossRef]
- Kabir, S. Critical analysis of compositions and protective efficacies of oral killed cholera vaccines. Clin Vaccine Immunol 2014, 21, 1195–1205. [Google Scholar] [CrossRef]
- Chen, W.H.; Cohen, M.B.; Kirkpatrick, B.D.; Brady, R.C.; Galloway, D.; Gurwith, M.; Hall, R.H.; Kessler, R.A.; Lock, M.; Haney, D.; et al. Single-dose live oral cholera vaccine CVD 103-HgR protects against human experimental infection with Vibrio cholerae El Tor. Clin. Infect. Dis. 2016, 62, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.J. Modern history of cholera vaccines and the pivotal role of ICDDR. Infect Dis 2021, 224, S742–S748. [CrossRef]
- Wierzba, T.F. Oral cholera vaccines and their impact on the global burden of disease. Hum Vaccines Immunother 2019, 15, 1294–1301. 2019; 15, 1294–1301. [CrossRef] [PubMed]
- Qadri, F.; Chowdhury, M.I.; Faruque, S.M.; Salam, M.A.; Ahmed, T.; Begum, Y.A.; Saha, A.; Al Tarique, A.; Seidlein, L.V.; Park, E.; et al. PXV Study Group, Peru-15, a live attenuated oral cholera vaccine, is safe and immunogenic in Bangladeshi toddlers and infants. Vaccine 2007, 25, 231–238. [Google Scholar] [CrossRef]
- Khan, A.I.; Islam, M.T.; Khan, Z.H.; Tanvir, N.A.; Amin, M.A.; Khan, I.I.; Bhuiyan, A.T.M.R.H.; Hasan, A.S.M.M.; Islam, M.S.; 0 Bari, T.I.A.; et al. Implementation and delivery of oral cholera vaccination campaigns in humanitarian crisis settings among Rohingya Myanmar nationals in Cox's Bazar, Bangladesh. Vaccines (Basel) 2023, 11, 843. [Google Scholar] [CrossRef] [PubMed]
- Song, KR; Lim, J.K.; Park, S.E.; Saluja, T.; Cho, S.I.; Wartel, T.A.; Lynch, J. Oral cholera vaccine efficacy and effectiveness. Vaccines 2021, 9, 1482. [CrossRef]
- Bi, Q.; Ferreras, E.; Pezzoli, L.; Legros, D.; Ivers, L.C.; Date, K.; Qadri, F.; Digilio, L.; Sack, DA; Ali, M.; et al. Oral cholerae vaccine working group of the global task force on cholerae control. Protection against cholerae from killed whole-cell oral cholerae vaccines: a systematic review and meta-analysis. Lancet Infect Dis 2017, 17, 1080–1088. [CrossRef]
- Koelle, K.; Rodo, X.; Pascual, M.; Yunus, M.; Mostafa, G. Refractory periods and climate forcing in cholerae dynamics. Nature 2005, 436, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Khatib, A.M.; Ali, M.; von Seidlein, L.; Kim, D.R.; Hashim, R.; Reyburn, R.; Ley, B.; Thriemer, K.; Enwere, G.; Hutubessy, R.; et al. Effectiveness of an oral cholerae vaccine in Zanzibar: Findings from a mass vaccination campaign and observational cohort study. Lancet Infect Dis 2012, 1, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J. An update on cholerae immunity and current and future cholerae vaccines. Trop. Med. Infect. Dis. 2021, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Sur, D.; You, Y.A.; Kanungo, S.; Sah, B.; Manna, B.; Puri, M.; Wierzba, T.F.; Donner, A.; Nair, G.B.; et al. Herd protection by a bivalent killed whole-cell oral cholerae vaccine in the slums of Kolkata, India. Clin Infect Dis 2013, 56, 1123–1131. [CrossRef]
- Naidu, A.; Lulu, S.S. Mucosal and systemic immune responses to Vibrio cholerae infection and oral cholerae vaccines (OCVs) in humans: a systematic review. Expert Rev Clin Immunol 2022, 18, 1307–1318. [Google Scholar] [CrossRef]
- Kanungo, S.; Azman, AS; Ramamurthy, T.; Deen, J.; Dutta, S. Cholerae. Lancet 2022, 399, 1429–1440. [CrossRef]
- De-Simone, S.G.; Napoleão-Pêgo, P.; Gonçalves, P.S.; Provance-Jr, D.W.; Morel, C.M.; Silva, F.R. B Cell epitope mapping of the Vibrio cholerae toxins A, B and P. Int. J. Mol. Sci. 2023, 24, 531. [Google Scholar] [CrossRef]
- Winkler, D.F.H. Automated solid-phase peptide synthesis. Methods Mol Biol 2020, 2103, 59–94. [Google Scholar] [CrossRef] [PubMed]
- Prim, D.; Rebeaud, F.; Cosandey, V.; Marti, R.; Passeraub, P.; Pfeifer, M.E. ADIBO-based "click" chemistry for diagnostic peptide micro-array fabrication: physicochemical and assay characteristics. Molecules 2013, 18, 9833–49. [Google Scholar] [CrossRef] [PubMed]
- Pinkhasov, J.; Alvarez, M.L.; Pathangey, L.B.; Tinder, TL; Mason, H.S.; Walmsley, A.M.; Gendler, S.J.; Mukherjee, P. Analysis of a cholerae toxin B subunit (CTB) and human mucin 1 (MUC1) conjugate protein in a MUC1-tolerant mouse model. Cancer Immunol Immunother 2010, 59, 1801–1811. [CrossRef]
- Sánchez, J.; Holmgren, J. Cholerae toxin structure, gene regulation, and pathophysiological and immunological aspects. Cell. Mol. Life Sci. 2008, 65, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Bharati, K.; Ganguly, N.K. Cholerae toxin: A paradigm of a multifunctional protein. Indian J. Med. Res. 2011, 133, 179. [Google Scholar]
- Sikora, A.E. Proteins secreted via the type II secretion system: Smart strategies of Vibrio cholerae to maintain fitness in different ecological niches. PLoS Pathog 2013, 9, e1003126. [Google Scholar] [CrossRef] [PubMed]
- Mekalanos, J.; Collier, R.; Romig, W. Enzymic activity of cholerae toxin. II. Relationships to proteolytic processing, disulfide bond reduction, and subunit composition. J. Biol. Chem. 1979, 254, 5855–5861. [CrossRef]
- Mayo, S.; Royo, F.; Hau, J. Correlation between adjuvanticity and immunogenicity of cholerae toxin B subunit in orally immunized young chickens. APMIS 2005, 113, 284–287. [Google Scholar] [CrossRef]
- Price, GA; Holmes, R.K. Evaluation of TcpF-A2-CTB chimera and evidence of additive protective efficacy of immunizing with TcpF and CTB in the suckling mouse model of cholerae. PLoS ONE 2012, 7, e42434. [CrossRef]
- Adachi, M.; Kurihara, Y.; Nojima, H.; Takeda-Shitaka, M.; Kamiya, K.; Umeyama, H. Interaction between the antigen and antibody is controlled by the constant domains: normal mode dynamics of the HEL-HyHEL-10 complex. Protein Sci. 2003, 12, 2125–31. [Google Scholar] [CrossRef]
- Hou, J.; Liu, Y.; Tao, R.; Hsi, J.; Shao, Y.; Wang, H. Cholerae Toxin B subunit acts as a potent systemic adjuvant for HIV-1 DNA vaccination intramuscularly in mice. Hum. Vaccines Immunother 2014, 10, 1274–1283. [Google Scholar] [CrossRef] [PubMed]
- Mayo, S.; Royo, F.; Hau, J. Correlation between adjuvanticity and,immunogenicity of cholera toxin B subunit in orally immunized young chickens. APMIS 2005, 113, 284–287. [Google Scholar] [CrossRef]
- Keyt, B.A.; Baliga, R.; Sinclair, A.M.; Carroll, S.F.; Peterson, M.S. Structure, function, and therapeutic use of IgM antibodies. Antibodies (Basel). 2020, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, H.; Zhang, R.; Ji, C.; Wang, Y.; Su, C.; Xiao, J. Immunoglobulin M perception by FcμR. Nature. 2023, 615, 907–912. [Google Scholar] [CrossRef]
- Hiramoto, E.; Tsutsumi, A.; Suzuki, R.; Matsuoka, S.; Arai, S.; Kikkawa, M.; Miyazaki, T. The IgM pentamer is an asymmetric pentagon with an open groove that binds the AIM protein. Sci Adv. 2018, 4, eaau1199. [Google Scholar] [CrossRef]
- Wang Y, Su C, Ji C, Xiao J. CD5L associates with IgM via the J chain. Nat Commun. 2024, 15, 8397. [CrossRef]
- Liu, J.; Wang, Y.; Min, Q.; Xiong, E.; Heyman, B.; Wang, J.Y. Regulation of humoral immune responses and B cell tolerance by the IgM Fc R,eceptor (FcμR). Adv Exp Med Biol. 2020, 1254, 75–86. [Google Scholar] [CrossRef] [PubMed]
- De-Simone, S.G.; Napoleão-Pêgo, P.; Gonçalves, P.S.; Lechuga, G.C.; Cardoso, S.V.; Provance, D.W. Jr.; Morel, C.M.; da Silva, F.R. B-Cell epitope mapping of the Vibrio cholera Toxins A, B, and P and an ELISA assay. Int J Mol Sci. 2022, 24, 531. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Janda, A. Immunoglobulin isotype influences affinity and specificity. Proc Natl Acad Sci USA 2012, 109, 12272–12273. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, M.A.; Islam, K.; Hossain, M.; Akter, N.J.; Alam, M.N.; Sultana, N.; Khanam, F.; Kelly, M.; Charles, R.C.; Kováč, P.; et al. Development of a new dipstick (Cholkit) for rapid detection of Vibrio cholerae O1 in acute watery diarrheal stools. PLoS Negl Trop Dis 2018, 12, e0006286. [Google Scholar] [CrossRef] [PubMed]
- Heyman, B.; Shulman, M.J. in Encyclopedia of Immunobiology (ed. DeFranco A.L and Ratcliffe, M.J.H.) 2016, p1–14. Academic Press, ISBN 012374282X, 9780123742827.
- Li, Y.; Shen, H.; Zhang, R.; Ji, C.; Wang, Y.; Su, C.; Xiao, J. Immunoglobulin M perception by FcμR. Nature 2023, 615, 907–912. [Google Scholar] [CrossRef]
- Wang, Y.; Min, Q.; Xiong, E.; Heyman, B.; Wang, J.Y.; Liu, J. Regulation of Humoral Immune Responses and B Cell Tolerance by the IgM Fc Receptor (FcμR). Adv Exp Med Biol 2020, 1254, 75–86. [Google Scholar] [CrossRef]
- De-Simone, S.G.; Gomes, L.R.; Napoleão-Pêgo, P.; Lechuga, G.C.; Pina, J.C.; Silva, F.R. Identification of linear B epitopes liable for the protective immunity of diphtheria toxin. Vaccines 2021, 9, 313. [Google Scholar] [CrossRef]
- Muzembo, B.; Kitahara, K.; Ohno, A.; Debnath, A.; Okamoto, K.; Miyoshi, S.I. Cholerae rapid diagnostic tests for the detection of Vibrio cholerae O1: An updated meta-analysis. Diagnostics (Basel) 2021, 11, 2095. [Google Scholar] [CrossRef]
- Muzembo, B.A.; Kitahara, K.; Debnath, A.; Okamoto, K.; Miyoshi, S.I. Accuracy of cholera rapid diagnostic tests: a systematic review and meta-analysis. Clin Microbiol Infect. 2022, 28, 155–162. [Google Scholar] [CrossRef]
- Nelson, E.J.; Grembi, J.A.; Chao, D.L.; Andrews, J.R.; Alexandrova, L.; Rodriguez, P.H.; Ramachandran, V.V.; Sayeed, MA; Wamala, J.F.; Debes, A.K.; et al. Gold standard cholera diagnostics are tarnished by lytic bacteriophage and antibiotics. J Clin Microbiol 2020, 58. e00412-20. [CrossRef]
- Ramamurthy, T.; Das, B.; Chakraborty, S.; Mukhopadhyay, A.K.; Sack, DA Diagnostic techniques for rapidly detecting Vibrio cholerae O1/O139. Vaccine. 2020, 38, A73-A82. [CrossRef]
- Chakraborty, S.; Velagic, M.; Connor, S. Development of a simple, rapid, and sensitive molecular diagnostic assay for cholera. PLoS Negl Trop Dis. 2023, 17, e0011113. [Google Scholar] [CrossRef]
- Zareitaher, T.; Sadat, T.; Seyed, AS; Gargari, L.M. Immunogenic efficacy of DNA and protein-based vaccine from a chimeric gene consisting of OmpW, TcpA, and CtxB, of Vibrio cholerae. Immunobiology 2022, 227, 152190. [CrossRef]
- Zereen, F.; Akter, S.; Sobur, M.A.; Hossain, M.T.; Rahman, M.T. Molecular detection of Vibrio cholerae from human stool collected from SK Hospital, Mymensingh, and their antibiogram. J Adv Vet Anim Res. 2019, 6, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Velagic, M.; Connor, S. Development of a simple, rapid, and sensitive molecular diagnostic assay for cholera. PLoS Negl Trop Dis. 2023, 17, e0011113. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Nelson, A.R.; Lopez, A.L.; Sack, D.A. Updated global burden of cholera in endemic countries. PLoS Negl Trop Dis. 2015, 9, e0003832. [Google Scholar] [CrossRef] [PubMed]
- Pezzoli, L. Oral cholera vaccine working group of the global task force on cholera control. Global oral cholera vaccine use, 2013-2018. Vaccine 2020, 38 Suppl 1, A132-A140. [CrossRef]
- Silva, F.R.; Napoleão-Pêgo, P.; De-Simone, S.G. Identification of linear B epitopes of pertactin of Bordetella pertussis induced by immunization with whole and acellular vaccine. Vaccine 2014, 32, 6251–6258. [Google Scholar] [CrossRef] [PubMed]
- De-Simone, S.G.; Napoleão-Pêgo, P.; Lechuga, G.C.; Carvalho, J.P.R.S.; Gomes, L.R.; Cardoso, S.V.; Morel, C.M.; Provance-Jr, D.W.; Silva, F.R.S. High-throughput IgG epitope mapping of tetanus neurotoxin: implications for immunotherapy and vaccine design. Toxins 2023, 15, 239. [Google Scholar] [CrossRef]
- Moutsinas, G.; Shuaib, C.; Guo, W.; Jarvis, S. Graph hierarchy: a novel framework to analyze hierarchical structures in complex networks. Sci Rep 2021, 11, 13943. [Google Scholar] [CrossRef] [PubMed]
- De-Simone, S.; Souza, A.L.A.; Melgarejo, A.R.; Aguiar, A.S.; Provance, D.W., Jr. Development of elisa assay to detect specific human IgE anti-therapeutic horse sera. Toxicon 2017, 138, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Jumper, J.; Evansm, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
IgA (I) and IgM (II) epitope mapping of V. cholerae toxin A (P01555), B (P01556), and P (P29485). A membrane-bound peptide library representing the three chains of the toxin was probed with a pool of vaccinated mouse sera (n=15), and reactivity was detected using goat anti-mouse IgA (I) and IgM (II) alkaline phosphatase labeled secondary antibody and chemiluminescence substrate. Panels (A) present an image of the peptide array showing reactivity as dark circles. The panels in (B) show the hierarchy position of each positive peptide and, in (C), the percent signal after normalizing the signals to the positive and negative controls. The sequences of peptides in each position are listed in Figure S1.
Figure 1.
IgA (I) and IgM (II) epitope mapping of V. cholerae toxin A (P01555), B (P01556), and P (P29485). A membrane-bound peptide library representing the three chains of the toxin was probed with a pool of vaccinated mouse sera (n=15), and reactivity was detected using goat anti-mouse IgA (I) and IgM (II) alkaline phosphatase labeled secondary antibody and chemiluminescence substrate. Panels (A) present an image of the peptide array showing reactivity as dark circles. The panels in (B) show the hierarchy position of each positive peptide and, in (C), the percent signal after normalizing the signals to the positive and negative controls. The sequences of peptides in each position are listed in Figure S1.
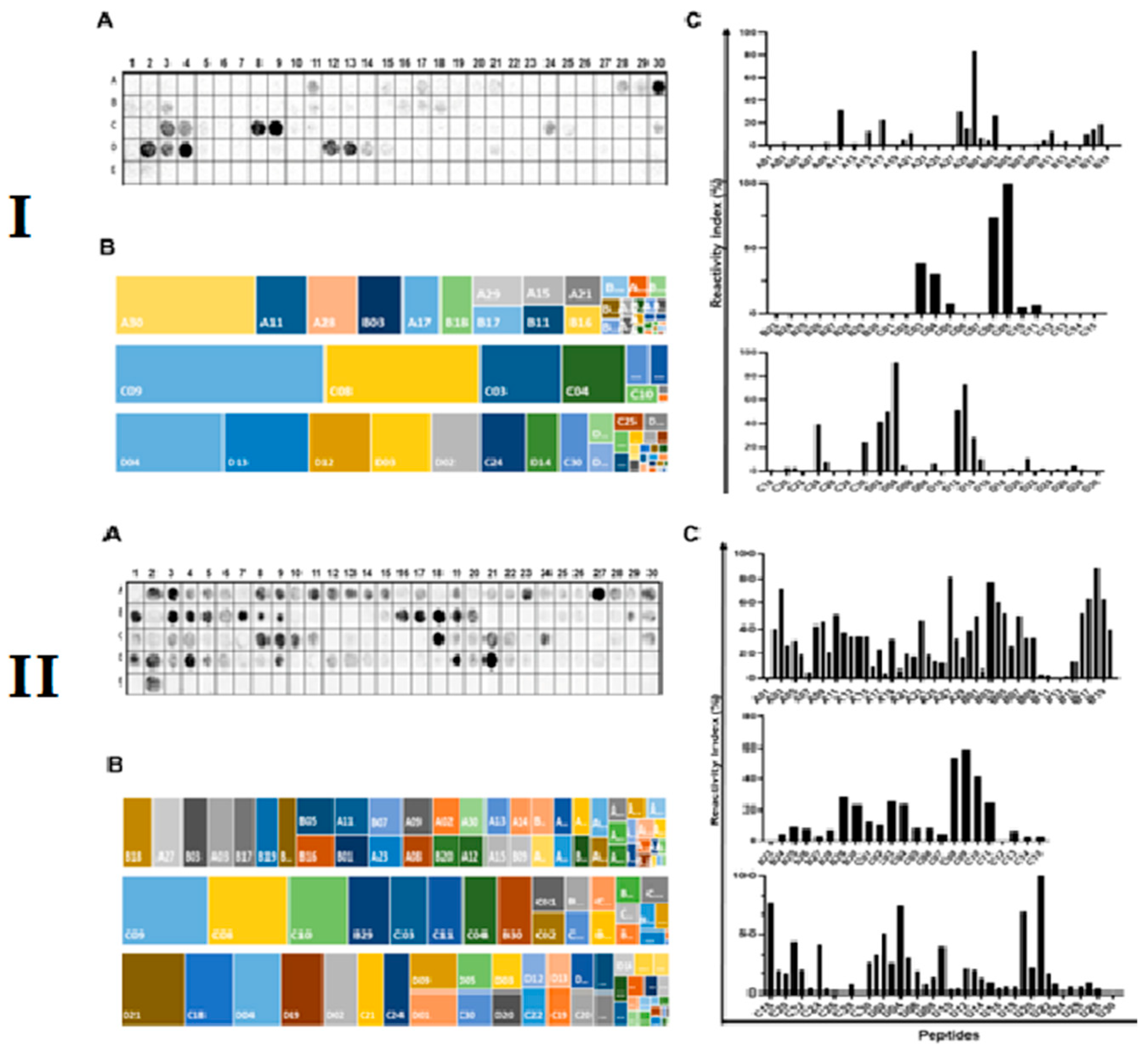
Figure 2.
Epitope localization in three-dimensional structures of V. choleraee toxins. (A) IgA epitopes in V. choleraee toxins A (TxA), B (TxB), and P (TxP) epitopes are presented and colored within models constructed using the structure of toxin A, B, and P predicted by AlphaFold. (B) IgM epitope mapping within V. choleraee toxin A. Images were created using ChimeraX.
Figure 2.
Epitope localization in three-dimensional structures of V. choleraee toxins. (A) IgA epitopes in V. choleraee toxins A (TxA), B (TxB), and P (TxP) epitopes are presented and colored within models constructed using the structure of toxin A, B, and P predicted by AlphaFold. (B) IgM epitope mapping within V. choleraee toxin A. Images were created using ChimeraX.
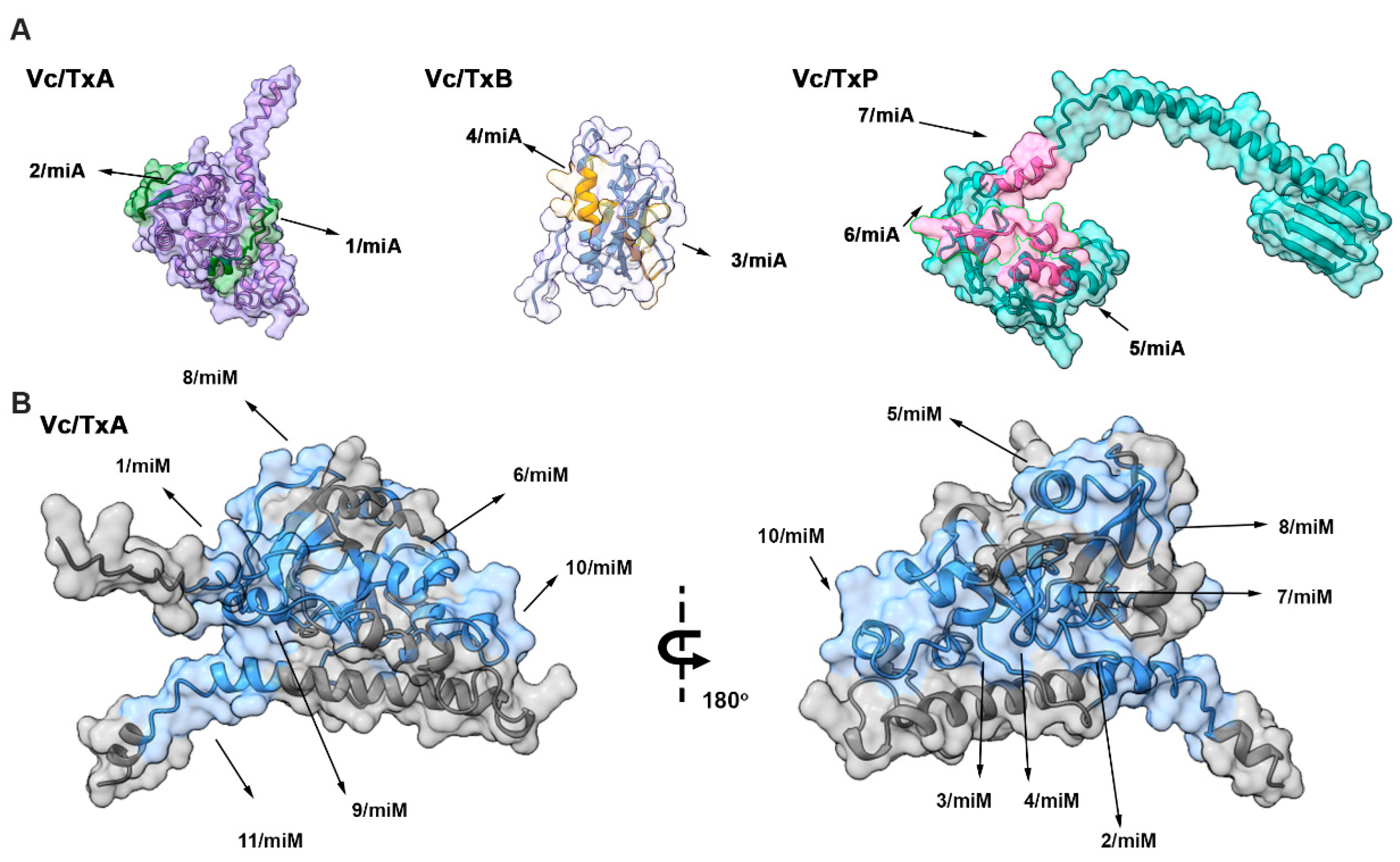
Figure 3.
Reactivity of vaccinated mice (n = 12) sera 15-(A) and 30 (B) days post vaccination against the MAP4 peptides (TxA-2A, TxB-3A, TxP-6A, TxA-2M, TxA-3M, and TxA-6M) and chimeric tripeptides (Tx45-IgA and Tx45-IgM) by ELISA. As controls were used, sera of unvaccinated mice (n = 30). The relative index was calculated using the cut-off of each MAP to normalize the measured value from the sera of 12 vaccinated mice. The ROC curve was employed to establish the peptide cut-off values (Tx45-IgA and Tx45-IgM; 0.053=0.056), evaluate reactivity (99-100%), assess specificity (100%), and calculate the Area Under the Curve (AUC) values (ranging from 0.9987 to 0.9896. (****) Statistical significance at p < 0.0001.
Figure 3.
Reactivity of vaccinated mice (n = 12) sera 15-(A) and 30 (B) days post vaccination against the MAP4 peptides (TxA-2A, TxB-3A, TxP-6A, TxA-2M, TxA-3M, and TxA-6M) and chimeric tripeptides (Tx45-IgA and Tx45-IgM) by ELISA. As controls were used, sera of unvaccinated mice (n = 30). The relative index was calculated using the cut-off of each MAP to normalize the measured value from the sera of 12 vaccinated mice. The ROC curve was employed to establish the peptide cut-off values (Tx45-IgA and Tx45-IgM; 0.053=0.056), evaluate reactivity (99-100%), assess specificity (100%), and calculate the Area Under the Curve (AUC) values (ranging from 0.9987 to 0.9896. (****) Statistical significance at p < 0.0001.
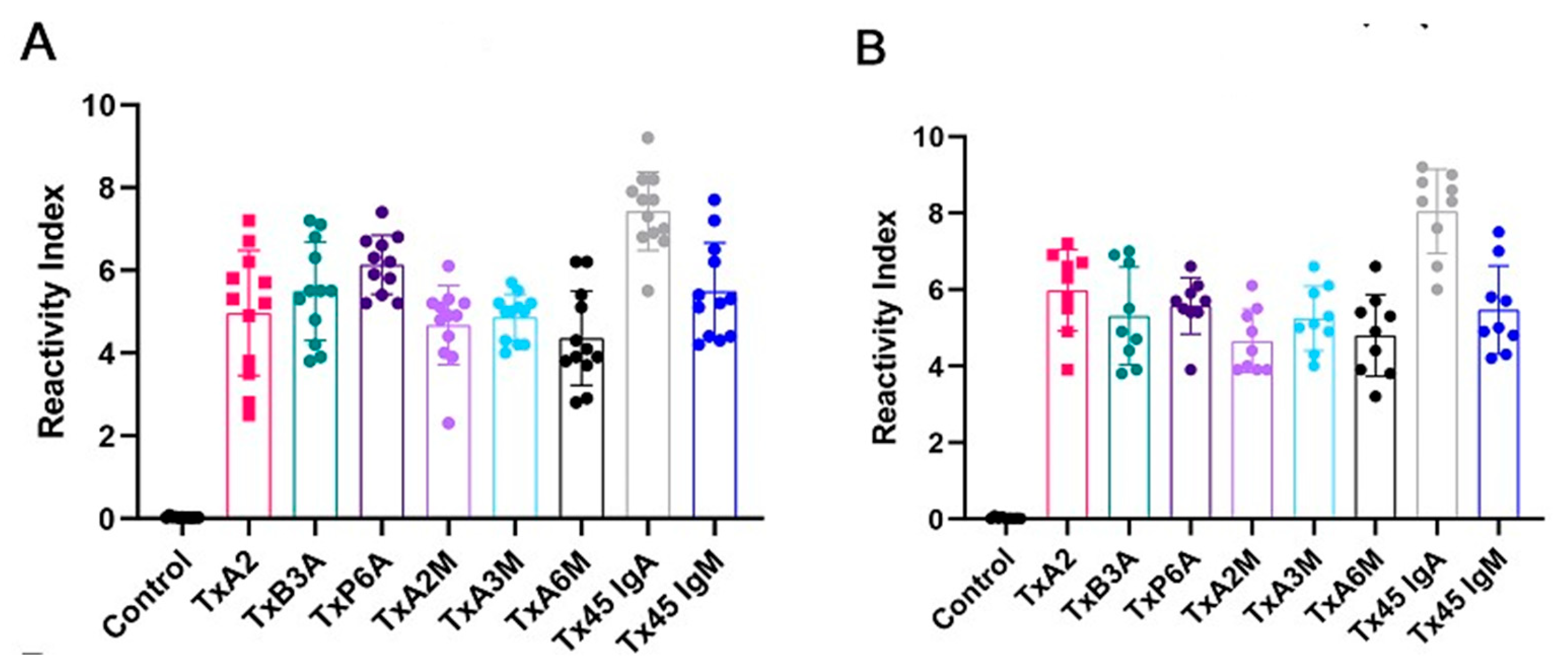

Table 1.
A list of B-cell linear IgA and IgM epitopes identified in TxA, TxB, and P was deduced from the overlap of consecutive positive peptides with signal intensities greater than 30%.
Table 1.
A list of B-cell linear IgA and IgM epitopes identified in TxA, TxB, and P was deduced from the overlap of consecutive positive peptides with signal intensities greater than 30%.
| Protein Code | Code | aa | Sequence | 2nd Structure * | Peptide Search ** |
| P01555 | TxA-1A | 51 | RGTQMNINLYDHARG | C | E. coli |
| TxA-2A | 146-160 | YRVHFGVLDEQLHRN | C | Sp | |
| P01556 | TxB-3A | 66-75 | REMAIITFKN | C+H | Sp |
| TxB-4A | 81-90 | SQKKAIERMK | H | E. coli | |
| P29485 | TxP-5A | 31-45 | KPERLIGTPSIIQT | C+H | Sp |
| TxP-6A | 81-90 | AIKRTRDFLN | C+H | Sp | |
| TxP-7A | 126-135 | QKKSVKERIK | C+H | Various bacteria | |
| P01555 | TxA-1M | 11-25 | FLSSFSYANDDKLYR | C | Various bacteria |
| TxA-2M | 41-50 | MPRGQSEYFD | C | Sp | |
| TxA-3M | 56-64 | NINLYDHAR | C+H | Sp | |
| TxA-4M | 71-80 | VRHDDGYVST | C | E. coli | |
| TxA-5M | 91-105 | GQTILSGHSTYYIYV | C+H | Various bacteria | |
| TxA-6M | 111-125 | NMFNVNDVLGAYSPH | C | Sp | |
| TxA-7M | 131-145 | VSALGGIPYSQIYGW | C | Various bacteria | |
| TxA-8M | 151-160 | GVLDEQLHRN | C | Sp | |
| TxA-9M | 171-185 | RGYRDRYYSNLDIAP | C | Sp | |
| TxA-10M | 191-205 | GLAGFPPEHRAWREE | C | Sp | |
| TxA-11M | 236-250 | VKRQIFSGYQSDIDT | C+H | Sp |
Sp, specific epitopes; C, coil; H, helix; S, strand; * based on an I-TASSER analysis; ** UNIPROT.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



