
Submitted:
10 December 2024
Posted:
12 December 2024
You are already at the latest version
Abstract
Diabetes mellitus is a chronic metabolic disorder characterized by persistent hyperglycemia, insulin dysfunction, and a substantial global health burden. Current pharmacological therapies often bring undesired side effects and limited long-term efficiency. These factors drive interest in the direction of preventive and complementary strategies. Plant-derived secondary metabolites – phenolics, alkaloids, terpenes, organosulfur compounds and polyacetylenes, demonstrate promising anti-diabetic activities. These compounds can modulate key molecular targets involved in glucose metabolism, insulin signaling, and development of oxidative stress, thereby improving glycemic control and reducing the risk of complications. Also, natural products can inhibit carbohydrate-digesting enzymes, enhance insulin secretion and sensitivity, regulate glucose transporters, and suppress pathways associated with inflammatory responses and the formation of advanced glycation end products (AGEs). Many of them also demonstrate pronounced antioxidant and antiglycative properties with all accompanying features - metal-chelating effects, α-dicarbonyl trapping properties, and interference with the AGE-RAGE axis, collectively mitigating vascular damage and attenuating diabetic complications such as nephropathy, neuropathy, and retinopathy. Recent studies highlight that certain plant metabolites can stabilize Nrf2, inhibit NF-κB activation, improve pancreatic β-cell function, and reduce protein tyrosine phosphatase activity. In this review we comprehensively address all this aspects with a special emphasis on the antiglycative properties as one of the central component of the overall anti-diabetic activity. Further, we discuss the further prospective in the biochemistry of plant-derived natural products in terms of antidiabetic, antiglycative and antioxidant agents. Although further research is needed to clarify their pharmacokinetics, safety, and efficacy in humans, the emerging evidence underscores the potential of plant secondary metabolites as natural, multifaceted agents for the prevention and management of diabetes and its associated complications.
Keywords:
diabetes mellitus
; plant secondary metabolites
; antiglycative activity
; anti-inflammatory
; antioxidant
; advanced glycation end products (AGEs)
; oxidative stress
1. Introduction
Diabetes mellitus (DM) is a chronic metabolic disorder, manifested with hyperglycemia at the background of impaired insulin secretion or action [1]. In the absence of adequate therapy this disease might progress rapidly and ultimately leads to onset of severe complications – cardiovascular disorders [2], cancer [3], neurodegenerative diseases (e.g. Alzheimer disease [4]), diabetic nephropathy or diabetic kidney disease (DKD) [5] and multiple related pathologies. Approximately 537 million adults (20-79 years) were living with diabetes in 2021, the total number is projected to rise to 643 million by 2030 and 783 million by 2045 [6]. Unfortunately, the rates of the diabetes-related premature mortality continuously grow, annually increasing by at least 5% since the beginning of the current century [7]. Moreover, due to the progressing growth of the human population and continuously increasing rates of obesity occurrence, the social role of diabetes increasing rapidly [1,8]. Obviously, the economic losses associated with high therapy costs are also continuously growing, and might overwhelm the capacities of public health protection system soon. In this context, diabetes prevention appears to be more promising strategy in comparison to its treatment and needs to be prioritized over the therapy of complications.
At the molecular level, the hyperglycemia-associated symptoms of diabetes mellitus are underlied by protein glycation [9,10,11,12]. This non-enzymatic post-translational modification is usually referred to as the interaction of protein amino and guanidine groups with reducing sugars and/or carbonyl products of their oxidative degradation [9]. Although diverse carbohydrates can act as potent glycation agents [10,13,14], the diabetes-associated protein damage is mostly underlied by reactivity of blood glucose [15] and dicarbonyl products of its degradation [16]. At the initial stages of glucose reaction with amines (early glycation) Schiff base intermediates are formed and further rearrange into fructosamines (often referred to as Amadori compounds). These first relatively stable glycation intermediates readily undergo oxidation and rearrangements yielding advanced glycation end products (AGEs) [17] At this step, glycation is tightly associated with oxidation and the process of AGE formation is often associated with the term "glycoxidation" [18]. Glycoxidation and irreversible formation of AGEs underlie onset of diabetes complications, which are always accompanied with systemic inflammation mediated by receptors to advanced glycation end products (RAGEs) and intensive cross-linking of blood and tissue proteins [19]. Simultaneous processes of glycoxidation and lipoxidation ultimately lead to the formation of AGEs and advanced lipoxidation end products (ALEs) [20], which can increase the rates of free radical generation, oxidative and carbonyl stress [21] that enhances diabetes symptoms.
Based on the underlying molecular mechanisms, the disease can be classified [22] in types 1 (T1DM) and 2 (T2DM) diabetes mellitus, which are characterized by insulin deficiency [23] (typically resulting from destruction of β-cells in autoimmune processes) [24] and hyperglycemia associated with β-cell dysfunction, peripheral insulin resistance, excessive liver glucose production [1], respectively. T2DM covers up to 90% [25] of diabetes cases, associated with obesity and has, therefore the highest social impact [26]. Its therapy typically relies on insulin secretolytics, biguanides, insulin sensitizers, α-glucosidase inhibitors, incretin mimetics, amylin antagonists, and SGLT2 inhibitors [24].
However, although recently introduced imeglimin (a glimin-containing oral hypoglycemic agent) showed good tolerability, long-term safety, and high efficiency [27], in general, synthetic drugs are characterized with adverse effects, essentially limiting their application [28,29]. This is the second reason to consider prevention of DM as the strategy of choice in terms of the overall concept of decreasing its occurrence in the population. Besides establishing the outpatient services and lifestyle changes [30,31], the prevention approach assumes implementation of plant extracts and their individual isolated components – biologically active natural products [32]. Being potent anti-diabetic agents, such products are less toxic and are featured with much less side effects [33]. Therefore, the anti-glycative and anti-diabetic effects of such products can be clearly manifested by their long-term implementation in the everyday human diet. Moreover, due to their pronounced antioxidant, hypoglycemic and antihyperlipidemic effects, plant-derived natural products can be considered as promising precursors of new antidiabetic and antiglycative therapeutic agents [34,35,36].
Therefore, this comprehensive review aims systematic exploration and analysis of abundant data on diverse plant secondary metabolites (SMs) demonstrating a general hypoglycemic effect and pronounced potential for attenuation and even prevention of diabetic complications. Thereby, we address the existing and potential mechanisms behind the anti-diabetic effects of plant natural products with specific emphasis on the anti-glycative component of this activity. We also critically discuss the current state of the art in the field and propose promising hypotheses and new directions for future research.
2. Anti-Diabetic Effects of Plant-Derived Natural Products
In general, efficient prevention and therapy of DM requires simultaneous affeсting several mechanisms to target hyperglycemia and associated molecular damage. Accordingly, the overall antidiabetic effect assumes several biological activities, including those targeting antioxidant (ROS-protective), carbonyl scavenging and other mechanisms related to protein glycation and associated signaling. As was mentioned above, despite impressive advances in development of new promising therapeutic agents, the search for efficient and safe antidiabetic drugs remains a challenge [37]. Indeed, synthetic antidiabetic drugs can have serious side effects and might often lead to development of drug resistance, gastrointestinal disorders, lactic acidosis, heart failure, progression of atherosclerosis, fluid retention, weight gain, and individual intolerance [38].
For example, a dipeptidyl peptidase (DPP)-4 inhibitor trelagliptin was shown to increase the risk of fractures [39]. Moreover, even such a well-established pharmaceutical, as metformin is not comprehensively addressed for side effects, especially in comparison to novel antidiabetics with pronounced cardioprotective properties [40]. Thus, the role of metformin as the "mainstream strategy" in the T2DM therapy might be questioned now [41]. These two well-known examples clearly indicate the importance of the search for new prototypes of anti-hyperglycemic (hypoglycemic) and/or glycoprotective agents, which would be promising in design of new pharmaceuticals [42]. In this context, plant secondary metabolites, often referred to as plant natural products, are the best candidates for such prototypes. Indeed, to date, more than 800 medicinal plants were reported to exhibit anti-hyperglycemic activity [43] and can be, therefore, potentially considered for in-depth analysis of their constituents in respect of their anti-diabetic and antiglycative properties.
Here we consider the groups of plant secondary metabolites, which are the best-characterized in respect of their structures, properties and the patterns of biological activities related to prevention or attenuation of DM symptoms – phenolic compounds, alkaloids and terpenoids along with several minor groups of active plant constituents.
In general, secondary metabolites are referred to as the taxon-specific substances, which are not directly involved in growth, development and reproduction, but mediate ecological interactions, which may produce a selective advantage for the organism by increasing its survivability or fecundity [44]. In contrast to animals, secondary metabolism of plants is characterized with fascinating complexity and plasticity [45]. Due to this, plant-derived natural products are featured with high structural and functional diversity, which underlie a broad spectrum of biological activities. As these biological activities rely on an impressive array of the modes of action, which, in turn, involve different targets (i.e. receptors, regulatory pathways and effector enzymatic systems), it makes sense to consider secondary metabolites according to their mechanisms of action.
2.1. Plant-Derived Natural Products with Antidiabetic Activity: Classes of Active Secondary Metabolites
Several plant secondary metabolites demonstrated clear anti-diabetic properties, which was well-documented in comprehensive in vivo and in vitro studies [46]. This knowledge was successfully implemented in promising therapeutic strategies, assuming modulation of cellular and molecular mechanisms by affecting different intracellular targets, i.e. enzymes and regulatory proteins [47].
Plant SMs are usually classified according to their characteristic structure moieties and biosynthetic pathways [48]. Based on these criteria, three compound groups with pronounced anti-diabetic properties strongly dominate in plant extracts: phenolics, terpenes, and alkaloids [48]. Thereby, the mechanistic aspects behind specific activities of plant natural products rely on several molecular mechanisms, which are being intensively studied since decades. At the current state of the knowledge in the field it is believed that anti-diabetic activity of natural products might rely on (i) suppression of oxidative stress and α-dicarbonyl formation, (ii) suppression of protein glycation and formation of advanced glycation end products (AGEs), (iii) inhibition of α-glucosidase and/or α-amylase, (iv) effects on glucose uptake and glucose transporters, (v) modification of gene expression and (vi) modulation of hormone activities involved in glucose homeostasis, (vii) enhancement of insulin secretion and pancreatic β cell proliferation and (viii) inhibition of protein tyrosine phosphatase activity.
2.1.1. Phenolic Compounds
Phenolic metabolites (also often referred to as phenolics) comprise the largest and the most abundant class of plant SMs represented with hydroxylated aromatic compounds and their conjugates [49]. Thus, structurally, these compounds comprise at least one aromatic ring with one or more hydroxyl substituents [50], i.e. share common for all them phenol moiety. These metabolites universally occur in roots, stems, leaves, flowers, fruits, and seeds of higher plants [51]. Metabolically, phenolics originate from the phenylpropanoid and shikimate pathways, which deliver low molecular weight aromatic precursors (so-called simple phenolics, Table 1) for biosynthesis of diverse compounds with higher molecular weights – polyphenols [52,53]. Polyphenols are characterized with impressive structural diversity and comprise several principal classes and sub-classes of highly different degrees of polymerization and molecular weights (Table 1) [49].
Simple Phenols
The structures of simple phenolics are featured with only one aromatic ring with hydroxyl, hydroxymethyl and methoxyl substitutions [54,55]. These compounds form three structurally distinct classes: hydroxybenzoic acids, phenylpropanoids (hydroxycinnamates) and coumarins along with corresponding derivatives. The representatives of these compound classes differ essentially and species-specifically in their relative abundance in plant tissues. Accordingly, antidiabetic effects of these compounds demonstrate a wide spectrum in terms of their modality and intensity of manifestation (Table 1).
In general, secondary metabolites based on phenol, catechol, resorcinol and phloroglucinol skeletons are widely spread in plants. However, despite of presence of certain specific activities, these natural products were not reported as potent anti-diabetic agents so far. Nevertheless, although benzoic acid and its derivatives do not have clear prospects as drugs [56,57,58,59], these metabolites appeared to be inhibitors of α-amylases and α-glucosidases, i.e. enzymes, which are directly involved in digestion and absorption of carbohydrates. However, further research is necessary to obtain additional data on their therapeutic efficiency and safety [60].
Coumarins
In contrast, coumarins [75,76] represent a group with pronounced antidiabetic properties, which are well-characterized to date (Table 1). This group is formed by cinnamic acid cis-О-hydroxy-lactones with a benzo-α-pyranone core [75]. Coumarins are naturally present in multiple plant species with the highest contents reported for Coumarouna odorata from the Fabaceae/Leguminosae family. They are also abundant in in vanilla grass (Anthoxanthum odoratum), sweet clover (genus Melilotus), cassia cinnamon (Cinnamomum cassia), Justicia pectoralis extracts and isolates from cherry blossoms [76].
Based on their structure, coumarins can be classified in six groups featured with different substitution patterns in the benzene and lactone rings: (i) simple coumarins (Table 1) (e.g., umbelliferone from plants of the Apiaceae family [77]), (ii) furanocoumarins (e.g., psoralen from [78] seeds of Psoralea corylifolia), (iii) dihydrofuranocoumarins (e.g., anthogenol extracted from Aegle marmelos [79]), (iv) pyranocoumarins (e.g., calanolide A from the foliar parts of Calophyllum lanigerum [80]), (v) phenylcoumarins (e.g., dispardiol B from Artemisia capillaris [81]) and (vi) bis- or tris-coumarins [55] (e.g., daphnoretin from Wikstroemia indica (L.) C.A. [82]).Most often, coumarins are present in plants in free (not conjugated) form.
Recently, multiple synthetic approaches to obtain biologically active coumarins were successfully established [83,84]. The resulted synthetic compounds, namely, flavonoid-coumarin hybrids, coumarin-cyclic imide conjugates, 3-coumarincarbohydrazides, and 3-coumarincarbohydrazones, proved to be promising in treatment of T2DM. These compounds were designed to enhance the antidiabetic properties of some natural coumarins by modifying their structure and introducing new functional groups [37,85]. This hypothesis was successfully confirmed by several in silico docking experiments [86,87], by comprehensive in vitro screening with several cell cultures and with appropriate animal models [85], while no studies on humans are available.
The antidiabetic effects of coumarins (both synthetic and naturally occurring) are mostly attributed to their ability to suppress oxidative stress and to reduce inflammation, to enhance pancreatic function, and to inhibit protein tyrosine phosphatase 1B (PTP-1B), which is a negative regulator of the insulin signaling pathways. These compounds also readily inhibit α-amylases and α-glucosidases. All these effects were convincingly demonstrated in various cell-based and animal models. Specifically, different coumarins were tested with a broad selection of cell lines, namely INS-1, RIN-m5F, HepG2, L6 and 3T3-L1 [37,88]. Further, their anti-diabetic properties were successfully addressed in streptozotocin-induced T2DM in rats and mice, alloxan-induced T1DM in mice, high fat diet-induced T2DM in mice, db/db mice, ob/ob mice and Zucker diabetic fatty rats. However, only minimal information on the bioavailability, pharmacokinetics and safety of coumarins in humans is available.
Polyphenols
Stilbenes
Stilbene synthase (STS) catalyzes the formation of the stilbene backbone (Table 1), such as resveratrol, using p-coumaroyl-CoA and malonyl-CoA as substrates [89]. This first branch of the flavonoid biosynthesis pathway exists in some plants, such as grapevine, pine, sorghum, and peanut [90]. Trans-resveratrol (3,5,4′-trihydroxy-trans-stilbene) is the basic unit of most plant stilbenes, for example, it can be converted to polydatin, pterostilbene, and piceatannol by glycosylation, methylation, and hydroxylation, respectively [91]. Otherwise, the oligomerization and isomerization of trans-resveratrol generate viniferin and cis-stilbene, respectively [92].
Stilbenes are well-known as strong antioxidants and are often produced by plants in response to biotic and abiotic stressors [92]. Resveratrol is the best characterized stilbene, and to date it is well-known for its beneficial effects on human health. It was proved to be anti-inflammatory, antioxidant, anticancer and cardioprotective in animal models and humans [93,94]. The anti-diabetic activity of resveratrol is underlined by its positive effect on insulin sensitivity and its high potential for reducing hepatic production of glucose. This effect is mediated by AMP-activated protein kinase, a key enzyme in the regulation of energy metabolism. Activation of this enzyme results in enhancement in glucose uptake by skeletal muscle and adipose tissue, in decrease in hepatic gluconeogenesis, that is accompanied with stimulation of insulin secretion by pancreatic β-cells [95,96].
Flavonoids
Obviously, the most representative group of biologically active polyphenols, which is highly relevant in the context of antidiabetic effects, is constituted by flavonoids. Flavonoids represent a large and diverse class of plant secondary metabolites that share a common structure moiety consisting of two aromatic hydroxyphenyl rings linked by a three-carbon bridge, forming a C6-C3-C6 skeleton [97]. Based on the hydroxylation patterns of their aglycons (i.e. the numbers of hydroxyl groups in the structure) and, specifically, substitution pattern of the heterocyclic C ring, flavonoids can be classified into seven subgroups— chalcones, flavones, flavanones, isoflavones, flavanols, flavan-3-ols, anthocyanins [98] (summarizes in Table 1). Flavonoids are widely distributed in fruits, vegetables, grains, tea, wine and herbs [97], where they underlie antioxidant, anti-inflammatory, anticancer, antiviral and antidiabetic activities [99]. Moreover, flavonoids readily form derivatives of highly diverse molecular structure. Such derivatives are featured with rich patterns of glycosylation, acylation, methoxylation and/or prenylation [100] of the polyphenolic aglycons, and molecular polymerization [101].
Flavonoids are synthesized from phenylalanine through the phenylpropanoid pathway, while phenylalanine is synthesized via the shikimate pathway [98]. Firstly, phenylalanine ammonia lyase catalyzes phenylalanine deamination [102] to trans-cinnamic acid, then cinnamic acid 4-hydroxylase (C4H), a cytochrome P450 monooxygenase in plants, catalyzes the hydroxylation of trans-cinnamic acid to p-coumaric acid [103]. The final step is formation of p-coumaroyl-CoA by the addition of a co-enzyme A (CoA) unit to p-coumaric acid by the activity of 4-coumarate: CoA ligase (4CL). The general phenylpropanoid pathway consists of these three steps and is common to all the downstream metabolites.
Chalcone formation represents the start of the synthesis of specific flavonoids, which begins with p-coumaroyl-CoA [101]. Chalcones represent the first key intermediate metabolites with an open C6-C3-C6 structure instead of a closed pyran ring providing a basic skeleton for downstream flavonoids synthesis. The anti-diabetic effect was shown for the chalcone isoliquiritigenin, which appeared to improve glucose homeostasis and insulin sensitivity in high-fat diet–fed mice by activating AMPK and inhibiting peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) expression [104].
The central branch point in the flavonoid biosynthesis pathway is the formation of the heterocyclic ring C by intramolecular cyclization of chalcones via chalcone isomerase (CHI), and subsequently, the formation of flavanones in the cytoplasm [98]. Flavanones constitute the sub-class of flavonoids missing the C2-C3 double bond and the hydroxyl group at C3. Naringenin and hesperidin belong to the most well-known representatives of this group. Thus Krishnamoorthy et al. showed that oral administration of naringenin (50 mg/kg b.w./day) for 6 weeks in high fructose-fed diabetic rats resulted in a significant increase in glucose transporter 4 (GLUT4) translocation in skeletal muscle by increasing AMPK phosphorylation, sirtuin 1 (SIRT1) and PGC-1α protein levels in skeletal muscle [105]. In turn, hesperidin was shown to reduce blood glucose levels and to suppress development of oxidative stress in the rat model of streptozotocin-induced T2DM due to activation of antioxidant enzymes and decreasing the circulating levels of inflammatory cytokines [106].
Flavones are produced from flavanones by flavone synthase (FNS) which catalyzes the formation of a double bond between position C-2 and C-3 of ring C [98]. Apigenin and luteolin belong to the most widely spread representatives of this group. As was shown in various animal models and cell lines, the antidiabetic effects of flavones are typically manifested by modulation of glucose uptake, insulin secretion and related signaling, that results in suppression of oxidative stress and inflammation [107]. For instance, apigenin improved glucose tolerance and insulin sensitivity in experimental animals (mice) with diabetes induced by a high fat diet. These effects were underlied by activation of AMPK and inhibition of the NF-κB signaling [108]. Moreover, luteolin was shown to reduce blood glucose levels and to improve lipid profiles of the streptozotocin-treated diabetic rats due to activation of antioxidant enzymes and suppressing inflammatory cytokines [109].
The isoflavone biosynthesis pathway is mainly distributed in leguminous plants [110] and leaded by isoflavone synthase (IFS). Isoflavones have unusual position of B-ring, which located at C3 instead of C2 in flavones. Due to their well-characterized potential for modulation of glucose uptake, insulin secretion and signaling, suppression of oxidative stress and inflammation, isoflavones are universally recognized as potent anti-diabetic agents.
Genistein and daidzein represent the most prominent examples of the naturally occurring isoflavones. Thus, genistein has increased the secretion of insulin from pancreatic β-cells via activation of cyclic AMP/protein kinase and showed its hypoglycemic activity by the phosphorylation of cyclic AMP/protein kinase along with GLUT1 and GLUT4 mRNA expressions in the L6 myotubes [111]. In turn, daidzein was shown to decrease blood glucose levels and to improve lipid profile in streptozotocin-induced diabetic rats by activation of antioxidant enzymes and expressional suppression of inflammatory cytokines [112].
Flavonols represent a sub-class of flavonoids featured with a hydroxyl group at C3 and variable hydroxyl substitutions in the A and B rings. Quercetin, kaempferol, and myricetin belong to the most widely spread in nature flavonols [113]. All these compounds proved to be promising anti-diabetic agents. Thus, due to its ability to stimulate translocation of GLUT4 and to enhance phosphorylation of Akt (protein kinase B), quercetin was shown to decrease blood glucose levels and to improve insulin sensitivity in streptozotocin-treated diabetic rats [114]. Further, already in the beginning of the last decade, Lee and coworkers reported protective effects of kaempferol in the model of cultured pancreatic β-cells. Application of this natural product suppressed glucotoxicity-induced apoptosis and increased insulin secretion from the INS-1 cells by activation of the Nrf2/HO-1 signaling pathway [115]. Finally, myricetin improved glucose tolerance and insulin sensitivity in the mouse model of T2DM based on the high fat diet. This effect was underlied by enhancement of AMPK expression and transcriptional suppression of the peroxisome proliferator-activated receptor gamma (PPARγ) coactivator-1α (PGC-1α) gene [116].
Leucoanthocyanidin is an important metabolic intermediate in the flavonoid pathway and the direct precursor in the anthocyanidin synthetic pathway in the reaction catalyzed by anthocyanidin synthase (ANS) [117]. The pelargonidin, cyanidin and delphinidin are examples of anthocyanins, which can be converted to stable anthocyanins by glycosylation under the activity of UDP-glucose flavonoid 3-glucosyltransferase (UFGT) [118]. It was shown that cyanidin improved glucose tolerance and insulin sensitivity in high-fat diet-induced diabetic mice by stimulating GLUT4 translocation and enhancing Akt phosphorylation [119].
Proanthocyanidins are generated from leucoanthocyanidins and anthocyanidins by leucoanthocyanidin reductase (LAR) which catalyzes the C-4 dehydroxylation of the C ring, and anthocyanidin reductase (ANR), which catalyzes the removal of a double bond at ring C respectively [98]. Flavan-3-ols, trans-flavan-3-ols, and cis-flavan-3-ols are the basic polymerization (or condensation) units for proanthocyanidins formation [120], so they are also known as condensed tannins.
Tannins are the high-molecular-weight polyphenolic compounds that can be found in a variety of plant species and foods. Tannins are able to bind to proteins and other chemical molecules, such as amino acids and alkaloids, and cause them to precipitate [121]. Coffee, tea, wine, grapes, apricots, barley, peaches, dry fruits, mint, basil, rosemary, pomegranate, strawberries, amla, clove, rice, oat, rye etc. are some important sources of tannins [122]. Tannins usually exist in two common types – condensed tannins which are oligomers or polymers of polyhydroxyflavan-3-ol monomer units linked by acid-labile 4 → 6 or 4 → 8 bonds [123].
Hydrolysable tannins
The latter group of compounds needs to be distinguished from hydrolysable tannins which are characterized by a monosaccharide, normally D-glucose, esterified with one or more molecules of gallic acid (gallotannins), or ellagic acid (ellagitannins) [49]. These plant secondary metabolites are known for their anti-inflammatory [124], antibacterial [125], antioxidant [126], antidiabetic [127,128] and anticancer activities [129]. Therefore, these natural products are widely employed in therapeutic strategies to prevent onset of cardiovascular, neuroprotective and metabolic diseases.
Chandak and co-workers reported the antidiabetic effect of hydrolysable tannins, where gallotannin was shown to reduce the overexpression of poly ADP-ribose polymerase (PARP) after four weeks of treatment in streptozotocin-induced diabetic rats [130]. Gallic acid, which is a monomeric unit of soluble tannins, was also reported to have antidiabetic effects by enhancing insulin receptor sensitivity and by modulating oxidative stress and inflammation [131,132]. Condensed tannins also have beneficial effects in the management of diabetic complications such as retinopathy and neuropathy, for example proanthocyanidins from grape seed affect inflammatory cascade involved in diabetic retinopathy by regulation of related protein expression like NF-kβ, inducible NO synthase (iNOS), COX-2 and inhibitor-binding protein kB-α [133]. Additionally grape seed proanthocyanidin containing oligomers of catechin and epicatechin and their gallic acid esters alleviates hyperglycemia and reduces Ca2+ overload by increasing Ca2+-ATPase activity in sciatic nerve [134].
Other phenolics
Lignans
Lignans constitute the class of polyphenols comprising a characteristic 2,3-dibenzylbutane structure that derived from the oxidative dimerization of two or more phenylpropanoid units [135]. These compounds have important roles in plant physiology, development, ecology (i.e., interactions and adaptations to ever-changing environments), and in plant defense protection against a variety of herbivores and microorganisms [136]. Lignans are found in flax seeds, sesame seeds, soybeans and all foods derived from plant shoots and roots [137]. Upon consumption with food, they are hydrolyzed by human intestinal microbiota and further metabolized to yield mammalian lignans, such as enterodiol and enterolactone. secoisolariciresinol diglucoside is hydrolyzed by β-glucosidases derived from anaerobic organisms into aglycone secoisolariciresinol, which undergoes processes of dehydroxylation and demethylation by intestinal microbiota and transformed into enterodiol and also oxidized to yeild enterodiol [138].
Due to their structure, enterodiol and enterolactone can mimic the effects of endogenous estrogens, so their plant-derived precursors (secoisolariciresinol diglycoside and matairesinol) are considered to be phytoestrogens [139]. In other words, enterolignans can exert estrogenic activity and could act as estrogen agonists or antagonists depending on the target tissues, doses and endogenous circulating sex hormone profile [86], for example, phytoestrogens have protective effect against hormone-dependent cancers, especially breast cancer, and proestrogens properties to correct the age-related hormonal deficiency in menopausal syndromes, and cardiovascular diseases [140]. On the other hand, phytoestrogens can also exert non-estrogenic activity such as antidiabetic activity, which might rely on both estrogen-dependent (e.g., altering glucose metabolism by directly modulating concentrations of circulating sex hormones that have a relationship with T2DM development [141], and this effects have been supported by some studies on human [142]) and estrogen-independent pathways (e.g., suppression of the phosphoenolpyruvate carboxykinase (PEPCK) gene expression which catalyzes the first step in hepatic gluconeogenesis [143,144]).Secoisolariciresinol diglucoside (SDG) is the major antidiabetic lignan from flax seeds [145]. Recently, it was reported to reduce the incidence of T1DM and to delay the progress of T2DM in humans. The anti-diabetic effect of SDG is underlied by modulation the expression of the genes involved in glucose and lipid metabolism, by improving the function of pancreatic β-cells and by suppression of oxidative stress and inflammation [146].
Lignins
Lignins are polymeric polyphenols consisting of phenylpropanoid monomer units [147]. They are ubiquitous constituents of the secondary cell walls of vascular plants, where they play structural and protective roles. These polymers are known for their antioxidant, anti-inflammatory, antimicrobial and anticancer activities [148]. The antidiabetic effects of lignins are less studied in comparison to other classes of polyphenols, although their positive effect on glucose tolerance and insulin sensitivity was reported [149]. The possible mechanisms behind these activities might rely on inhibition of α-glucosidase, activation of AMPK, modulation of insulin signaling and expression of the genes related to glucose metabolism [149].
2.1.2. Alkaloids
Alkaloids represent a large (more than 10,000 representatives) and diverse class of nitrogen-containing plant SMs [150]. These compounds are found approximately in 25% of angiosperm species in essentially varying abundances [151].
Metabolically, alkaloids are derived from amino acids and contain one or more nitrogen atoms in their structure, most often, within one or several heterocyclic rings [152]. Such heterocyclic nitrogen-containing compounds are typically referred to as true alkaloids, whereas aliphatic amino acid-derived nitrogen-containing compounds (e.g. hordenine, ephedrine, colchicine and capsaicin) are termed as protoalkaloids or aminoalkaloids [153]. Both true alkaloids and proto-alkaloids are splitted in several classes according to their amino acid precursors [49,150]. This classification is summarized in Table 2. In total, about 800 alkaloid-producing medicinal plants species were reported to have more or less pronounced antidiabetic activity [154,155]. Recently, scientific databases were comprehensively screened to highlight the biological activity of 78 PAs with a considerable anti-diabetic profile, based on the results of these studies, all these compounds appeared to be potent anti-diabetic agents [156]. Specifically, these phytochemicals, in general, were shown to be potent potent α-glucosidase inhibitors [156].
Family Fabacea is known for multiple widely spread species with rich alkaloid patterns, which might be promising as the sources of anti-diabetic metabolites. For example, since the medieval times, preparations based on Galega officinalis extracts are well-recognized as efficient remedies for the treatment of diabetes mellitus. The active component of the G. officinalis extracts, which resulted in pronounced drop in the blood glucose levels, appeared to be galegine [157]. This natural product is the representative of guanidine alkaloid and is well-known for its weight reducing potential, which is underlied by inhibition of fatty acid synthesis and enhancement of fatty acid oxidation [158].
Four indole alkaloids (vindoline, vindolidine, vindolicine, and vindolinine) were isolated from dichloromethane leaves extract of Catharanthus roseus and demonstrated antidiabetic potential in pancreatic β-TC6 or myoblast C2C12 cells besides exhibiting notable protein tyrosine phosphatase-1B (PTP-1B) inhibitory properties [159]. Mahanine - a dimeric carbazole alkaloid found in the leaves, stem bark, and roots of the Murraya koenigii plant, was shown to exhibit significant α-glucosidase inhibitory properties [160]. Further, several steroidal alkaloids isolated from Sarcococca saligna possess hypoglycemic effect and improve others diabetes associated complications [161].
Although multiple alkaloids were shown to be promising antidiabetic agents, their activity was mostly confirmed by in silico, in vitro, and in vivo approaches, while the corresponding mechanisms are still poorly characterized. Therefore, preclinical and clinical studies accompanied with comprehensive pharmacokinetic and toxicological surveys are still required to consider alkaloids as safe and efficient biopharmaceuticals. This aspect is critically important, as numerous representatives of this group are featured with high toxicity and their application even in folk medicine might be dangerous for human health, that might be manifested with diverse adverse effects, such as nausea, vomiting, diarrhea, headache, dizziness, hallucination, convulsion and cardiac arrhythmia by interfering with various physiological processes and receptors [162]. Therefore, selection of formulation, dosage and administration routes of requires a special attention to secure safety and ensure desired therapeutic outcomes.
In agreement with this fact, one needs to keep in mind that several factors that might directly affect the toxicity and pharmacokinetics of alkaloids need to be addressed in much detail. Specifically, their chemical structure, solubility and ADME (absorption, distribution, metabolism, excretion) properties need to be comprehensively characterized. Moreover, the molecular targets and intracellular interaction partners need to be identified to get the first insight in the mechanisms underlying their activity. For example, jatrorrhizine (which belongs to the group of isoquinoline alkaloids with pronounced antidiabetic activity) has low oral bioavailability due to its poor solubility and extensive metabolism by intestinal bacteria and liver enzymes [163]. Therefore, it might be better administered intravenous. The targets and mechanisms, obviously, represent the most serious issues, which require a deeper insight for most of the characterized alkaloids.
2.1.3. Terpenes
Terpenes represent the class of widely distributed natural products synthesized by diverse plant and animal species [180]. Several representatives of this compound class were reported as antihyperglycemic agents over the recent decades [180]. The molecular structure of terpenes underlies their specific activity, binding affinity and specificity to various molecular targets, such as enzymes, receptors and transporters involved in glucose metabolism [181]. Interestingly, the most of the known natural terpenes are reported to be much less toxic than corresponding synthetic analogs with similar mechanisms of action, although some of them accumulate in the liver and exert pronounced toxicity [182]. Despite their relatively large molecular weight and high lipophilicity, pronounced skin penetration potential and bioavailability of terpenes give access to a new way for administration of antidiabetic drugs [182].
Terpenes are classified according to the number of isoprene units forming the carbon backbone of the molecule: hemiterpenes, monoterpenes, sesquiterpenes, diterpenes, triterpenes, tetraterpenes and polyterpenes. Multiple in vitro and in vivo studies [183] clearly showed that monoterpenes (e.g. carvacrol, thymol, and linalool) can be used as antidiabetic compounds [183], either individually or as parts of complex mixtures. Due to their relatively low molecular weights, compact and mostly linear molecular structures, sesquiterpenes appeared to be more efficient in treatment of diabetes in comparison to monoterpenes or other terpene classes [184].
On the other hand, due to their different mechanisms of action, hydroxyl- and carboxyl-substituted triterpenes can be also considered as anti-diabetic natural products, which might be promising in treatment of DM both at its early stages and when complications are already well-manifested [185]. The underlying mechanisms employ modulation of glucose transporters, enhancement of insulin secretion and insulin sensitivity, inhibition of carbohydrate-digesting enzymes, suppressing oxidative stress and inflammation [182]. This might result in prevention or reversal of β-cell dysfunction, attenuation of renal damage and neuropathic pain. Thus, such terpenes might be efficient in the treatment of diabetic retinopathy, neuropathy and nephropathy or in impaired wound healing by inhibition of several signaling pathways, e.g. those related to protein kinase C, receptors for advanced glycation end products (RAGEs) and transcription factor nuclear factor-κB (NF-κB). Thus, such widely spread triterpenes as oleanolic acid, ursolic acid and betulin were efficient in clinical trials. In general, analysis of the structure-activity relationships (SAR) of triterpenes revealed that hydrogen bonding and hydrophobic interaction might favor antidiabetic activity [182].
2.1.4. Minor Secondary Compounds
Sulfur-Containing Compounds
Organosulfur compounds are organic molecules that contain sulfur and are associated with the pungent odors characteristic of vegetables that contain these metabolites [186]. Allyl sulphides and isothiocyanates (ITC) are two classes of reactive organosulphur compounds [187]. Allyl sulphides can be found in vegetables of the genus Allium, especially garlic, onions, leeks and chives, such as alliin, allicin, S-allylcysteine, S-allylmercaptocysteine [188]. Allicin from garlic extract is S-allyl-2-propene sulphinothioic acid ester formed from alliin by an enzyme alliinase and represents the precursor for several other sulphur volatile compounds as methyl allyl disulphide, diallyl disulphide, dimethyl trisulphide, allyl methyl trisulphide, diallyl trisulphide and sulphur dioxide [189]. Allyl sulphides exhibit several biological activities including anti-inflammatory, anti-carcinogenic, and anti-angiogenic effects [190].
Anti-diabetic potential was demonstrated for all these bioactive metabolites. For example, by in vitro study on streptozotocin-induced diabetic rats was shown that daily intragastric doses of allyl methyl sulfide, a major volatile garlic metabolite, for 30 days resulted in a significant attenuation in blood glucose, in expression of pro-inflammatory markers TNF-α, IL-6, NF-κB p65 unit and significant elevation in the plasma insulin level [191]. The treatment of streptozotocin-induced diabetic with S-allyl cysteine also resulted in a substantial reduction in blood glucose indicating that SAC has antihyperglycemic properties [192]. Ahmad and co-authors demonstrated the potential antidiabetic activity for alliin, S-allyl-L-cysteine, N-acetylcysteine, and S-ethyl-L-cysteine as inhibitors of α-amylase and α-glucosidase via a set of in silico and in-vitro analysis [193]. In addition, cruciferous vegetables (Brassicaceae family) such as broccoli, cauliflower, Brussels sprouts, and cabbage represent a rich dietary source of glucosinolates (GLS), which are sulfur- and nitrogen-containing compounds, derived from glucose and amino acids [194]. They play a key role in defense against insect herbivores and attracting pollinators [195]. There are aliphatic GLS such as glucoraphanin, gluconapin, sinigrin, progoitrin and glucoiberin), and indolyl-GLS (e.g. glucobrassicin) [196]. When cruciferous vegetables tissues are damaged during chewing or cutting, the endogenous enzyme myrosinase is released and break the β-thioglucoside bond of glucosinolates, resulting in isothiocyanates (hydrolyzed products) that have in common the R–N═C═S functional group [197]. Glucoraphanin [4-methylsulfinylbutyl glucosinolate] is the predominant glucosinolate which is hydrolyzed its corresponding isothiocyanate, sulforaphane [1-isothiocyanato-4-(methylsulfinyl)-butane] [198]. Sulforaphane can activate the Nrf2 factor, which regulates antioxidant defense and phase two enzymes. Sulforaphane also inactivates NF-kB, which controls cytokine production and development of inflammatory response [199].
It was shown recently, that sulforaphane can improve pancreatic function, lowered blood glucose levels, decreased insulin resistance, and enhanced intestinal metabolism. Sulforaphane also attenuated inflammation and oxidative stress in liver and muscle tissues [200]. Thereby, it positively affects homeostasis of glucose, which relies on (i) regulation of blood glucose levels by insulin secretion and action, (ii) glucose uptake and utilization by peripheral tissues and (iii) glucose production by the liver.
Polyacetylenes
Polyacetylenes are the natural products which can be found in various plant-derived foods, i.e. vegetables and fruits [201]. They may have beneficial effects on glucose metabolism and insulin sensitivity, which might be underlied by modulation of the PPARγ receptor activation. This receptor is the key regulator involved in onset of insulin resistance and triggering of systemic inflammation [202]. Studies, accomplished with different models of diabetes showed that polyacetylenes from roots of carrots (Daucus carota L., Apiaceae), Oplopanax horridus and Panax ginseng can increase PPARγ transactivation and stimulate insulin-dependent glucose uptake [203,204]. Polyacetylenes might act as partial PPARγ agonists, similar to long-chain polyunsaturated fatty acids [204]. However, further studies are needed to elucidate their molecular targets, pharmacokinetics, toxicity and clinical efficiency [205].
3. Mechanisms Behind Anti-Diabetic Activity of Plant-Derived Natural Products
3.1. Antioxidant Activity - Detoxification of Reactive Oxygen Species (ROS)
Currently, there is no doubt that reactive oxygen species (ROS), which predominantly are represented with hydrogen peroxide (H2O2), superoxide anion radical (O2-.), hydroperoxyl radical (HO2.), singlet oxygen, hydroxyl radical (OH.), nitric oxide (NO), peroxynitrite, as well as RNS (reactive nitrogen species) such as nitric oxide (NO), nitrogen dioxide (NO2), and the non-radical peroxynitrite (ONOO-). It is well-known that ROS play a significant role in the pathogenesis of T2DM and in development of its complications [206].
ROS and RNS are quite common tissue metabolites which are constitutively produced in human organism via several pathways. The most important source of ROS is the electron transport chains (ETCs) of chloroplast and mitochondria, where continuous leak of electrons with partial reduction of molecular oxygen occurs [207]. The resulting mtROS (mitochondrial ROS), mainly superoxide anion radicals, are formed due to electron leakage from respiratory complexes I and III [207], and hydroxyl radical via iron-mediated Fenton reaction [208], as shown in Figure 1.
Under normal conditions, coupled respiration on glutamate/malate (GM) or pyruvate/malate (PM) activates the Krebs cycle enzymes 2-oxoglutarate dehydrogenase (OGDH), malate dehydrogenase (MDH), and pyruvate dehydrogenase (PDH), and maintains a low membrane potential as the fifth respiratory Complex, the ATP synthase (CV), is producing ATP. These enzymes reduce NAD+ to NADH, which is, in turn, a substrate of the first respiratory Complex (CI). Electrons flow down the chain and reach the third and fourth respiratory Complexes (CIII, CIV). The production of ROS in this forward electron transfer (FET)
situation is minimal, and is commonly by CI, the outer ubiquinone-binding site of CIII (CIIIo) and OGDH [210]. Manganese superoxide dismutase (Mn-SOD) converts the superoxide radical to hydrogen peroxide in the mitochondrial matrix (MM), while Cu- and Zn-SOD convert the superoxide radical in the intermembrane mitochondrial space or cytosol [211]. The H2O2 in the MM can be converted further via a Fenton reaction by mitochondrial aconitase to a hydroxyl radical (•OH) [212].
Under normal conditions, these ROS play mostly regulatory role and are continuously detoxified by an arrow of low-and high molecular weight antioxidants [213], for example, glutathione peroxidases (GPXs), thioredoxin peroxidases (TRXPs), superoxide dismutases (SODs), peroxiredoxins (PRDXs), glutathione (GSH), thioredoxin 2 (TRX2), glutaredoxin 2 (GRX2), cytochrome c oxidase (complex IV), coenzyme Q, ascorbic acid, tocopherol, vitamin E, and carotene, catalase (CAT), also detoxifies H2O2 in the peroxisome [214].
Under pathological conditions, such as hypoxia or mitochondrial dysfunction, the concentrations of mtROS increase and their production overwhelms the capacity of the antioxidant system. This ultimately results in development of oxidative stress [215]. Moreover, non-mitochondrial sources of ROS – nicotinamide adenine nucleotide phosphate oxidase (NOX) [216], xanthine oxidoreductase (XOR) [217], monoamine oxidases [218], NOS (nitric oxide synthase) [219], lipoxygenases (LOXs) and cyclooxygenases (COXs) [220] strongly contribute in manifestations of oxidative stress in tissues. Prolonged enhancement of ROS generation and accompanying oxidative stress result in depletion of all cellular redox-based stress-protective systems and suppression of antioxidant activity [221]. Such shifts in redox metabolism typically accompany hyperglycemia, inflammation, dyslipidemia and underlie associated distortion of cellular metabolism and cell damage [222].
Under hyperglycemic conditions accompanying the T2DM pathology, glucose and lipid metabolism become the principal sources of ROS overproduction behind the development of oxidative stress [223]. Indeed, due to increased formation of ROS, the processes of glyco- and lipoxidation are enhanced, that is accompanied with overproduction of α-dicarbonyl compounds [224]. The latter, in turn, are the key players of advanced glycation and act as direct precursors of advanced glycation end products (AGEs) [225,226]. Thereby, glyoxal (GO), methylglyoxal (MGO) and 3-deoxyglucasone (3-DG) are the most abundant tissue α-dicarbonyls, which are overproduced under hyperglycemic conditions [227]. The most pronounced sources of these compounds are sugar and lipid metabolism, specifically – reactions of glycoxidation and lipoxidation [228]. On the other hand, essential part of the cellular MGO pool originates from glycolysis – specifically, from non-enzymatic conversion of glyceraldehyde-3-phosphate (G-3-P) [229]. This dicarbonyl compound readily modifies not only proteins, but also nucleic acids. Thus, its overproduction is associated with enhanced DNA damage and, consequently, activation of the repair enzyme poly-ADP-ribose polymerase 1 (PARP1) [230]. PARP1 inhibits glyceraldehyde-3-phosphate dehydrogenase (GAPDH) [231], resulting in accumulation of G-3-P and other intermediates of glycolysis with further enhancement of MGO production. Thus, increasing overproduction of α-dicarbonyls (the phenomenon known as carbonyl stress) results in enhancement of advanced protein glycation and formation of AGEs [232]. It is well-known to date, that accumulation of AGEs in human blood and tissues ultimately causes sub-clinical systemic inflammation in humans and triggers activation of pro-inflammatory signaling pathways [233].
Not less importantly, increased concentration of dihydroxyacetone-3-phosphate (DHAP), which is reduced by glycerol-3-phosphate dehydrogenase to glycerol-3-phosphate, in the presence of free fatty acids, in turn, increases diacylglycerol concentration and activates proinflammatory protein kinase C-mediated signaling pathways [234]. Further, accumulation of ROS disrupts the hexosamine pathway of glucose oxidation, which is associated with enhanced expression of the mitogenic transcription factors TGF-α and TGF-β [235].
Taking all this information into account, it is logical to assume that the search for compounds, which target the mechanisms of ROS generation and α-dicarbonyl production (which will be considered in more detail below), might be a promising strategy to control the progress of diabetes and its complications. In general, antioxidants modulate redox equilibrium by restraining and/or retarding multiple oxidative reactions via different mechanisms. Thus, antioxidants can act as scavenging agents reducing equilibrium concentration of ROS in intra- and intercellular liquids. Thereby, on one hand, antioxidants can be chelating agents, which are involved in formation of complexes with metals. On the other, they can play the role of radical traps, i.e. the molecules, able to interrupt free-radical chain reactions [236]. Most often, the latter group comprises phenolics, primarily polyphenols – anthocyanins and flavonoids. Thus, the search for new ROS-protectors among these compound classes, which might activate antioxidant protection of the cell, can be considered a promising strategy. Thereby, transcription factors Nrf2 and NF-κB, which are important regulators of the cell response to oxidative stress [237], might be considered as the key targets of this strategy.
Nuclear factor erythroid 2-related factor 2 (Nrf2) is a basic leucine zipper transcription factor that belongs to the Cap'n'Collar (CNC) family [238,239]. Its cellular contents are quite low under normal conditions, as this protein is constantly degraded in the proteasome (half-life of only 20 minutes [240]), bound to two molecules of the Keap1 protein (the adaptor of E3 ubiquitin ligases) [241]. Under oxidative stress, the conformation of the Keap1 protein changes, and it dissociates from Nrf2. Due to this, the Nrf2 factor avoids proteasomal degradation, its concentration in cytosol increases that triggers its transport to the nucleus. In the nucleus it forms a heterodimer with the Maf protein, which binds to antioxidant response element (ARE) sequences, activating gene transcription of antioxidant defense system including (peroxiredoxin 1 (PRDX 1), glutathione reductase (GR), thioredoxin reductase 1 (TXNRD 1) or sulfiredoxin (SRXN)), enzymes and metabolic regulators (glucose-6-phosphate dehydrogenase (G6PD), transketolase, malic enzyme, RXRα, PPARγ-coactivator 1 β (PGC1-β)) [242].
The targets of Nrf2 include the genes involved in protein transport, ubiquitination, phosphorylation, cell cycle regulation, growth, and apoptosis. Moreover, activation of Nrf2 leads to down-regulation of fatty acid biosynthesis. Importantly, stability of Nrf2 can be affected not only by the Keap1 protein, but also by glycogen metabolism and autophagy. Phosphorylation of p62 protein by some protein kinases (in particular AMPK) leads to degradation of Keap1 and accumulation of Nrf2 in the cytosol. Interestingly, while p62 is a target for Nrf2, this factor itself serves as a target for several protein kinases, among which are PKC, AMPK, Cdk5, MAPK, ERK, JNK, PI3K/Akt, PKA. Thereby, signal transduction from the membrane receptors to the transcription machinery is established [243].
NF-kB plays the key role in the development of acute and chronic inflammation, autoimmune diseases, and carcinogenesis. The canonical pathway of NF-kB activation is mediated by the inhibitor of κB (IκBα), which binds the NF-kB dimer, preventing its activation. In response to external stimuli, IκBα undergoes phosphorylation by IκB kinase (IKK), which results in ubiquitination of IκBα protein, dissociation from NF-κB and allowing it to transport into the nucleus and bind to DNA. The target genes of NF-κB are cytokines, chemokines, iNOS, COX, endothelin-1, and lipoxygenase; metabolic changes associated with diabetes have been shown to result from NF-κB activation [244].
Nrf2 and NF-kB signaling pathways affect each other in different ways. Thus, the p65 subunit of NF-κB was shown to inhibit the Nrf2 pathway at the transcriptional level, both by competing for binding with Nrf2 coactivators and by affecting the histone acetylation-deacetylation system [245]. On the other hand, Nrf2 inhibits the NF-kB signaling pathway, as was shown in in vitro study on mice with Nrf2 gene knockout and astrocytes [246]. Thus, reduction in Nrf2 levels increases NF-κB expression, resulting in increased formation of proinflammatory factors, and vice versa, NF-κB affects the transcription of target genes involved in regulation of Nrf2 transcription and activity.
Multiple steps in the mechanism of Nrf2 activation can serve as targets for plant metabolites. Treatment of the MIN6 cell line with endogenously expressed naringenin resulted in suppression of oxidative stress, translocation of Nrf2 to the nucleus, and an increase in Nrf2 expression using the Nrf2-Keap1 complementation system [247].
Curcumin and sulforaphane covalently modify several cysteine residues in the Keap1 protein, thereby causing its dissociation and increasing the stability of Nrf2 [248,249]. It was also proposed that these substances can act as epigenetic regulators, inhibiting histone deacetylases and activating histone acetylases, contributing to chromatin remodeling and activating expression of the Nrf2 gene [250].
Compounds that modulate the activity of protein kinases can be considered promising antidiabetic agents. Particularly, berberine, as an antidiabetic compound, via AMPK activating promotes glycolysis through increased glucokinase activity, increased insulin secretion, and suppressed hepatic gluconeogenesis and adipogenesis [251].
The natural products, which simultaneously activate Nrf2 and inhibit NF-kB signaling pathways, represent other promising targets. For example, andrographolide, a labdane diterpenoid isolated from Andrographis paniculata, forms Michael acceptor dependent adducts with Cys151 in KEAP1 in vivo, leading to inhibition of NRF2 ubiquitination and consequently accumulation of the transcription factor [252]. Andrographolide upregulates NRF2 through p38 MAPK and ERK activation. In addition, andrographolide can also form adducts with Cys77, Cys151, Cys273, and Cys368 in KEAP1, resulting in its dissociation from Nrf2 and increasing the lifetime of the latter.
3.2. Inhibition of α-Glucosidase and α-Amylase
Amylase inhibitors suppress the release of glucose from oligo- and polysaccharides in intestine that results in a delay in its absorption. It leads to a decrease in postprandial glycemia and a general decrease in hyperglycemia [253].
To date, several benzoic acids and their derivatives were shown to act as inhibitors of α-glucosidase. Thus, Chen and co-workers showed that these compounds interact with the active site of the enzyme through hydrogen bonding and π-interactions [254]. However, benzoic acid derivatives are rarely considered as antidiabetic drugs because they have low bioavailability, poor solubility, and high toxicity [56].
Recent studies have shown that coumarins are potent inhibitors of α-glucosidase and/or α-maltase, i.e. the enzymes converting disaccharides to simple monosaccharides such as glucose. These inhibitory effects can be attributed to characteristic structure moieties of the natural coumarine derivatives, especially to the biaryl C-C or C-O-C linkages, to the terpene side chain and to the characteristic cyclobutane ring [85]. Some examples of natural coumarins with antidiabetic activity are scopoletin from Morinda citrifolia, auraptene from Citrus aurantium, and umbelliferone from Angelica archangelica. Besides these natural products, several synthetic derivatives were proposed as the potential anti-diabetic agents - such as flavonoid-coumarin hybrids, coumarin-cyclic imide conjugates, 3-coumarin-carbohydrazides, and 3-coumarin-carbohydrazones. However, the effects of these compounds are still insufficiently characterized [37,85].
The double bond at C2-C3 and the keto-group at C4 represent the two structural features, which are critically important for the inhibitory activity of flavonoids against α-glucosidase and α-amylase [255]. Methylation and acetylation of specific hydroxyl groups at the polyphenol core were found to decrease antioxidant and antidiabetic enzyme inhibition activity in vitro [256]. Thus, flavonoids having quercetin, myricetin, kaempferol, and genistein cores as aglycons were reported to inhibit alpha-glucosidase and alpha-amylase, and promote glycogenesis [257]. The inhibitory effects on α-glucosidase were also reported for anthocyanins, stilbens, terpenoids and some other minor classes of secondary metabolites.
3.3. The Effects of Natural Products on Glucose Absorption and Transmembrane Transport
Binding of insulin to its receptors on the surface of myocyte and adipocyte plasma membrane triggers recruitment of IRS (insulin receptor substrate) proteins. Insulin signaling leads to the activation of phosphatidylinositol 3-kinase (PI3K), which phosphorylates phosphatidylinositol-4,5-bisphosphate (PIP2) to generate phosphatidylinositol-3,4,5-trisphosphate (PIP3). PIP3 recruits and activates Akt, which phosphorylates several important players of signaling pathways. Akt phosphorylates glycogen synthase kinase-3 (GSK-3), that results in its inhibition and promotion of glycogen synthesis. Akt also phosphorylates AS160, a protein involved in GLUT4 translocation [258]. Phosphorylation of AS160 by Akt inhibits its activity, leading to the translocation of GLUT4 to the plasma membrane. This allows enhanced glucose uptake by the skeletal muscle cells. Thereby, plant-derived natural products might affect both expression of the receptor and its translocation to the plasma membrane [259].
The biological activities of this type are well-characterized for alkaloids, which are generally known as potent biological effectors [260]. For example, steroid alkaloids isolated from Sarcococca saligna demonstrated clear hypoglycemic effects and appeared to reduce severity of other complications associated with DM like retinopathy, polyphasia, polyurea and cardiovascular problems [161]. Similar conclusions could be drawn about vindolidine isolated from Catharanthus roseus [159].
Another group of anti-hyperglycemic compounds is represented by terpenes [261]. For example, administration of the triterpene betulin, isolated from the root bark of Euclea undulate, resulted a significant reduction in blood glucose levels in comparison to the reference synthetic drug glibenclamide and no side effects. At the molecular level, this effect is underlied by inhibition of the transcription factor controlling the biosynthesis of cholesterol, fatty acids and triglycerols - sterol regulatory element-binding protein (SREBP) [34]. When cellular cholesterol levels are low, SREBPs are activated and translocated to the nucleus, where they bind to specific DNA sequences known as sterol regulatory elements (SREs) [262]. When bound to SREs, SREBPs promote the expression of genes involved in cholesterol synthesis, fatty acid synthesis, and triglyceride synthesis, leading to an increase in lipid production [263]. The precise extent of SREBP inhibition and its impact on lipid synthesis may vary under different physiological or pathological conditions [264].
Similarly, monoterpenes, like carvacrol, cymene and genipin, or the combination of linalool and limonene (the latter is also known as a protein glycation inhibitor), showed improved glucose uptake [183]. Therefore, due to their effect on glucose transport, some monoterpenes, such as carvacrol and thymol, are widely used as biologically active constituents of foods [183].
Some antidiabetic therpene-rich plant isolates are already commercialized. For example, glucosol (1% w/v corosolic acid) – a well-standardized formulation isolated from Lagerstroemia speciosa leaves, was shown to reduce blood glucose levels in a randomized clinical trials with T2DM patients 135. Triterpene glycoside desmethoxysenegin II isolated from the rhizomes of Polygala senega Linn. reduces the blood glucose levels in healthy, non-diabetic mice [182].
PPAR is a group of three nuclear receptors (PPARγ, PPARα, and PPARδ) which bind to response elements in the promoters to control gene expression. PPARγ plays an important role in lipid metabolism, inflammation, immunity, and glucose homeostasis [265]. Specifically, its activation controls the expression of transcription factors, which are secreted by adipose tissue and affect insulin sensitivity. Also, certain cytokines and lymphokines are secreted by adipocytes and immune cells residing in adipose tissue, collectively known as adipose tissue-derived cytokines or adipokines. These adipokines can modulate insulin signaling and glucose metabolism in various tissues [266]. This might directly modulate the expression of genes involved in glucose homeostasis such as GLUT4 and CAP [267]. PPARγ is induced during differentiation of preadipocytes in adipocytes. Thus, modulation of PPARγ transcript levels with receptor agonists remains an attractive pharmacological target for the treatment and prevention of metabolic disorders, including diabetes mellitus [183,268,269].
3.4. Enhancement of Insulin Secretion and Proliferation of Pancreatic β Cells
Enhancement of insulin secretion and proliferation of pancreatic β cells is the key focus in diabetes research. Incretins represent the group of hormones of the glucagon family, which are secreted in the gut in response to consumption of foods, especially those containing glucose and fat [270]. When the blood glucose contents get increased, incretins trigger insulin production in the pancreas. To date, two intestinal hormones with an incretin effect are known: GIP (glucose-dependent insulinotropic polypeptide), which is secreted in the distal ileum and colon by L-cells, and GLP-1 (glucagon-like peptide-1), which is secreted in the duodenum and jejunum by K-cells. Both hormones have low half-lives and are rapidly degraded by the enzyme DPP-4 (dipeptidyl peptidase-4) [271]. Thus, prolongation of their life might give a pronounced anti-diabetic effect. Corresponding therapeutically active agents might be either DPP-4 inhibitors or undegradable GLP-1 analogues [272,273].
Coumarins (such as umbelliferone, esculentin and osthole) and their derivatives were reported to fix pancreatic β-cell damage and to be promising in treatment of DM and its complications [37]. In pancreatic β-cells, increased levels of free radicals can lead to oxidative stress and cell damage, which is accompanied by a decrease in cell function and the development of DM. Coumarins such as umbelliferone, esculetin, and osthole can reduce free radical levels and have an anti-inflammatory effect, protecting pancreatic β-cells from damage [274]. Also coumarins can increase the activity of insulin-secreting channels and stimulate insulin release from pancreatic β-cells. This helps to lower blood glucose levels and improve DM control [275].
Flavonoids based on quercetin, myricetin, kaempferol, and genistein aglycon structures, were found to protect pancreatic β cells from damage and to stimulate insulin secretion from β cells by modulating various signaling pathways involved in glucose metabolism [257]. Pancreatic β cells are vulnerable to oxidative stress, which can lead to their dysfunction and damage. By scavenging ROS and increasing the cellular antioxidant capacity, flavonoids can protect β cells from oxidative damage [276]. Flavonoids can inhibit the production of pro-inflammatory cytokines and enzymes, such as tumor necrosis factor-alpha (TNF-α) and inducible nitric oxide synthase (iNOS), thus reducing inflammation in β cells and promoting their function [277].
Not less importantly, non-proteinogenic amino acids, like taurine were shown to suppress lowered fasting sugar and insulin levels [278]. However, their safety and efficiency need to be addressed in further studies. The other group of active compounds – betains, are characteric for red beet extracts [279]. Recently, Madadi et al [280] confirmed the anti-glycemic effect of betalains. These natural products reduce glycemia by 40% without simultaneous weight loss or liver damage. However, further experiments are required to confirm these results in vivo and the safety of betalains.
3.5. Inhibition of Protein Tyrosine Phosphatase Activity
Binding of insulin to its receptors results in a pronounced increase in the tyrosine kinase activity [281]. It leads to autophosphorylation of the insulin receptor and sequential phosphorylation of the whole cascade of downstream proteins ultimately resulting in the translocation of glucose transporter proteins, particularly GLUT4, to the cell membrane [282]. This signaling cascade leads to glucose accumulation and its enhanced utilization by the cell [283]. Similarly, activation of the leptin receptor activates Janus kinase 2 (JAK2), which subsequently phosphorylates signal transducers and transcription activator protein 3 (STAT3), thereby affecting fatty acid metabolism promoting fatty acid oxidation and reducing fatty acid synthesis. These coordinated and interrelated effects of insulin and leptin on glucose and lipid metabolism contribute to the regulation of energy storage [284].
The physiological regulation of the insulin- and leptin-dependent pathways is finely tuned by the balance between phosphorylation and dephosphorylation of the corresponding receptors [285,286]. Phosphorylation enhances the signaling activity, while dephosphorylation attenuates it. Protein tyrosine phosphatase 1B (PTP1B) is a common negative regulator of both insulin and leptin signaling cascades. This fact makes it a promising therapeutic target for DM treatment [285,286]. This effect was reported for a number of natural products of terpenoid and phenolic nature such as curcumin, resveratrol, epigallocatechin gallate, and carnosic acid [287].
The obtained analogues of myristic acid (Khaya senegalensis and Tamarindus indica) showed potent inhibitory activity on protein-tyrosine phosphatase in vitro, that indicated their value as potential therapeutic agents. However, additional studies are needed to evaluate the efficiency and safety of these compounds in living organisms [288].
In this context, Juglans regia (walnut) leaf extract represents a promising subject for research into its potential role in the treatment of elevated blood glucose levels. Recent studies have shown that walnut leaf extract enhances glucose uptake and inhibits PTP1B (tyrosine phosphatase), providing justification for the traditional use of walnut leaf preparations against increased blood glucose levels. Additional research in this area could help determine the optimal doses and regimens for walnut leaf extract and evaluate its efficacy and safety in living organisms [289].
4. Suppression of α-Dicarbonyl Formation, Protein Glycation and Accumulation of AGEs as a Mechanism Behind the Anti-Diabetic Activity of Plant Secondary Metabolites
4.1. Protein Glycation, Formation of α-Dicarbonyls and Accumulation of Advanced Glycation End Products (AGEs)
The phenomenon of protein glycation was first discovered by Louis Camille Maillard in 1912 [290]. Because of this, the browning reactions occurring during thermal processing of foods are collectively termed as Maillard reaction [290].
Generally, the complex process of glycation can be divided in two principal steps. First, carbonyl groups of reducing sugars reversibly react with free amino/guanidino groups of proteins or peptides (mainly with arginine, lysine and N-terminal amino acid residues), with lipids and nucleic acids yielding unstable Schiff bases. These intermediates, ketoimines and aldoimines undergo spontaneous intra-molecular rearrangements that form covalently bound, relatively stable aldoamines and ketoamines, also known as Heyns [291] аnd Amadori products [292], respectively.
At the next step, the early glycation products are involved in oxidation, degradation and condensation reactions, which yield advanced glycation end products (AGEs). This term, initially proposed by Brownlee et al. in 1984 [293], defines the group of heterogeneous, chemically and structurally diverse compounds formed by several pathways either exogenously or endogenously. The process of AGE formation is usually referred to as advanced glycation [294].
To date, several principal pathways of AGE formation are known. On one hand, early glycation products can be directly converted to AGEs by cross-linking with proteins, hydrolysis or irreversible autoxidation (in the case of Amadori products, the latter is often referred to as Hodge pathway) [295]. On the other, AGEs can be formed directly by interaction of reactive amino acid residues with carbonyl compounds such as glyoxal (GO), methylglyoxal (MGO), 1- and 3-deoxyglucosone (3-DG), diacetyl, glyceraldehyde and glycolaldehyde [226,227]. The highly reactive carbonyl intermediates can be generated by various rearrangements, dehydration, oxidative degradation or cyclization of early glycation products, monosaccharides autoxidation (Wolff pathway) [296], in vivo conversion of glucose to fructose (polyol pathway), lipid peroxidation (acetol pathway) [297], and oxidative cleavage of Schiff bases (Namiki pathway) [298]. Importantly, the α-dicarbonyl-related pathways often yield cross-linking AGEs that dramatically affects mechanical properties of AGE-modified proteins [299]. The formation of AGEs is also promoted by oxidative stress.
Over the last decades, numerous AGEs were reported both in vitro and in vivo. Thereby, several dozens of new glycation products were identified in human blood, tissues and in foods [300]. Based on the source of their formation, these products can be classified as endogenous and exogenous AGEs [301]. Endogenous AGEs are formed under physiological metabolic conditions, in presence of pathology and during normal aging [301]. Exogenous AGEs are abundant in animal-derived foods rich with protein and fat [302], especially thermally processed foods, known as dietary AGEs (dAGEs), and in tobacco smoke [303]. Dietary AGEs are widely used in food industry to improve taste, safety and bioavailability of food [304]. The Department of Food Chemistry in Technical University of Dresden created an AGE database to present the diversity and amount of dAGEs in selected food products [305]. Recently, the process of AGE formation under diverse conditions was comprehensively reviewed in much detail [228,306,307].
Accumulation of AGEs in the human organism leads to deleterious physiological effects. On one hand, these effects can be underlied by modifications in structure and functions of extracellular and intracellular proteins. The most of such effects are associated with cross-linking, which is known to dramatically restrict elasticity of connective tissues during ageing [308]. On the other hand, the AGE-associated effects might rely on activation of multiple signaling pathways associated with an array of cell surface receptors [309]. These both mechanisms contribute to onset and progression of pathological states, such as age-related diseases (diabetes mellitus, cardiovascular diseases, neurodegenerative diseases and cancer and their complications [310].
4.2. Advanced Glycation End Products (AGEs) and Their Receptors
The major class of the cell surface receptors involved in recognition of AGEs and mediation of the AGE-related physiological effects is the type I multi-ligand receptor for advanced glycation end products (RAGE). The other receptors contributing in AGE recognition are - advanced glycation end product (AGE) receptor 1, 2 and 3 (AGE-Rs), and scavenger receptor family (e.g. stabilin-1 and stabilin-2). The structures, expression, ligands and signaling pathways of these receptors are comprehensively reviewed elsewhere [307,309].
RAGE (Figure 2) is an immunoglobulin (Ig) superfamily of transmembrane proteins, originally purified from bovine lung endothelial cells as a 35 kDa polypeptide [311]. It is a pattern recognition receptor [312] that recognizes various ligands (AGEs, amyloid β peptide, S100/calgranulin protein, HMGB1) and is expressed on the surface of several cell types (endothelial cells, monocytes/macrophages, T-lymphocytes, dendritic cells, fibroblasts, smooth muscle cells, neuronal cells, glial cells, chondrocytes and keratinocytes) [313]. The full-length RAGE (fl-RAGE) protein has a large extracellular region with one V-type and two C type domains, a transmembrane-spanning helix domain (TM), and a short cytoplasmic domain [314]. RAGE has different isoforms produced by alternative splicing of fl-RAGE mRNA [315], two major variants are N-truncated RAGE and soluble RAGE (sRAGE), which is secreted extracellularly and produced from proteolytic cleavage by ADAM10 and MMP9 [316]. AGE-RAGE interaction triggers signaling pathways that induce ROS production and systemic inflammatory response, discussed in the next section.
4.3. Role of Glycation and AGE Formation in Diabetes Mellitus
The levels of protein glycation and AGE formation are increased progressively with normal ageing, and appear to be further accelerated under hyperglycemic conditions during development of diabetic complications [317]. The direct involvement of AGEs in the development of insulin resistance was shown in mice, which demonstrated a pronounced decrease in the contents of the anti-AGE receptor 1 (AGER1) and SIRT1 in various tissues after synthetic AGEs administration [318]. Thus, the progress of DM is accompanied with accumulation of endogenous AGEs.
As was mentioned above, similarly to ingested dAGEs, the accumulated in the organism endogenous AGEs exert their pathological effects through two main mechanisms: (i) interfering with the normal function of serum or extracellular matrix (ECM) proteins by disruption of molecular conformation, reduction of degradation capacity, alteration of enzyme activity and immunogenicity, modifying protein half-life and cross linking [319] and (ii) AGE-RAGE interaction, which triggers the NADPH oxidase pathway and subsequent ROS generation [320] and induce intracellular signal transduction through the activation of various kinases (e.g. phosphatidylinositol 3-kinase, PI3K), which inhibit insulin-induced GLUT-4 translocation to the cell membrane, resulting in insulin resistance [301]. This interaction also activates extracellular signal-regulated kinase/mitogen-activated protein kinase (ERK/MAPK) pathway, and the Janus kinase/signal transducer and activator of transcription (JAK/STAT) pathway [321]. These pathways increase the expression of transcription factor activating protein-1 (AP-1) and nuclear factor kappa B (NF-κB), which is the main player of the inflammatory response by promoting the expression of pro-inflammatory cytokines and chemokines (IL-1, IL-6, TNF-α, monocyte chemoattractant protein-1 (MCP-1), intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1) and endothelin-1 [313]. Thereby, a vicious cycle of oxidative stress and inflammation cascade will be induced and ultimately lead to cell apoptosis [310,322]. Therefore, AGEs accumulation indicates chronic hyperglycemia as well as total metabolic burden, oxidative stress and inflammation [323]. AGEs can be considered as biomarkers and mediators in many diabetic complications [319].
4.4. Anti-Glycation Effects of Plant Secondary Metabolites
It is well-known today, that glycoxidative stress and AGEs cause diabetes-related vascular damage [324]. Antiglycative agents can block AGE formation or RAGE binding, preventing or delaying, thereby, the progress of DM and its complications. Due to rich patterns of biological activities, plant secondary metabolites attract a special attention of researchers as anti-glycative and anti-diabetic agents [325].
Thus, phenolic compounds, such as phenolic acids, flavonoids, stilbenes and lignans demonstrated pronounced antiglycative potential in vivo and in vitro [326]. The possible mechanism behind are: i) trapping highly-reactive α-dicarbonyl compounds, ii) scavenging free radicals and suppression of ROS generation, iii) chelation of metal ions, iv) interference with RAGE expression and related signaling pathways, iv) inhibition of aldose reductase v) effect on the availability of potential glycation sites in proteins, vi) reducing of blood glucose levels. Recently, Ramkissoon et al showed correlation between the phenolic composition of extract obtained from tropical plants and their antiglycative activities using in vitro glucose-bovine serum albumin (BSA) assay [327].
Terpenoids were also shown to be antiglycative [301,328]. Thus, pentacyclic triterpenoids, found in hawthorn, bitter melon, oat, green tea, and herbal medicines [329], have antihyperglycemic and anti-glycative activities. Oleanolic acid, ursolic acid and betulinic acid are recognized as potential supplements or drugs for T2DM [330]. Iridoids, another terpenoid class, which is common in medicinal plants. For example, iridoid glycoside aucubin prevents AGE formation and has antioxidant and anti-inflammatory activities. Due to this fact, it is widely used as a Chinese herbal medicine for AGE-related diseases [331]. Wu and Yen [332] studied the effect of ten flavonoids on AGE production and found that flavones have the highest antiglycation potential.
Four alkaloids from Ocotea paranapiacabensis had antiglycation potential [301]. Berberine, another alkaloid, from Cortex phellodendri and Berberis, protected against diabetic renal damage by lowering blood glucose and increasing antioxidant capacity [333]. Meta-analysis of 27 trials reported that berberine could efficiently suppress the symptoms of T2DM without any serious adverse reactions [334]. Plant secondary metabolites have different structures and functions and might employ different antiglycative mechanisms, which are addressed in more detail below.
Trapping Highly-Reactive α-Dicarbonyl Compounds
Reactive carbonyl compounds and, especially, α-dicarbonyls (GO, MGO, 3-DG) are the key precursors of structurally diverse AGEs. The increase of their tissue contents, i.e. the state which is usually referred to as carbonyl stress, always accompanies diabetic hyperglycemia [224]. Reducing the contents of these intermediates of advanced glycation is the main mechanism behind the inhibition of AGE formation by natural products.
The most typical carbonyl scavengers are polyphenols. The phenolic hydroxyl groups and benzene rings form phenoxy anions and bind to electrophilic α-dicarbonyl compounds [301]. For example, epigallocatechin gallate (EGCG) from green tea represents a well-known example of such compounds [335]. Phloretin and phloridzin from apple, quercetin, (+)-catechin, (-)-epicatechin and gingerol, ellagic acid, chlorogenic acid and gallic acid were also reported to be potent carbonyl scavengers [336,337,338,339,340,341,342,343]. Curcumin – the metabolite which is extractable from Curcuma longa Linn, was shown to trap MGO directly by forming curcumin-MGO adducts at the C10 position in the dense carbon portion of the molecule between diketone groups [301]. Genistein, the major soy isoflavone, also shows significant capacity for trapping of MGO [344,345]. Furthermore, this effect was exhibited by procyanidins, condensed tannins which are widely present in various species of berries and in flowers of the ancient Magnolia genus [346,347].
Multiple studies confirmed the antiglycative activity of resveratrol, which is mediated by different mechanisms including scavenging dicarbonyls [348,349]. In vitro study of tetrahydroxystilbene glucoside (THSG), a stilbene monomer extracted from Polygonum multiflorum, confirmed its impact in MGO-trapping in a time- and dose-dependent manner [350]. In addition, the pentacyclic triterpenoid oleanolic acid (OA) binds MGO with C-12 in the C ring to form mono-adduct [351].
Scavenging Free Radicals and Suppression of ROS Generation
The early steps of the Maillard reaction yield highly-reactive ROS [352]. The accompanying oxidative stress results in enhancement of protein glycation and AGE formation [353]. Scavenging free radicals and suppressing ROS production by plant secondary metabolites are important mechanisms to prevent protein glycation, cross-linking, and AGE production. Multiple polyphenol moieties, such as catechins, proanthocyanidins, anthocyanin, stilbenes and flavonols exert pronounced antioxidant activity and inhibit AGE formation by neutralizing ROS or suppressing free radical production [354].
The B-ring of the flavonoid structure is the main target of the radical attack in polyphenols [355]. For example, genistein was shown to inhibit NADH oxidase and to scavenge free radicals [356]. Also, Kiso et al reported antioxidant activity of sesamin, a lignan isolated from the sesame oil and seeds [357]. Sesamin inhibits NADPH oxidase-mediated oxidative stress and slows down AGE-induced β-cell dysfunction and apoptosis [358].
The ROS-scavenging potential of carotenoids appeared to be even higher than that of polyphenols. These metabolites represent a class of tetraterpenoid polyene pigments, which are composed of eight isoprenoid units, representing, thereby, C40 compounds [359]. In plants, these lipophilic natural pigments are stored in chloroplast and chromoplast membranes, giving the plant organs, including fruits, colors like red, yellow and/or orange. Depending on the presence or absence of the oxygen atom, carotenoids are divided into xanthophylls (C40H56O2 or C40H56O) and carotenes (C40H56) [360].Thus, astaxanthin was shown to inhibit protein glycation by ROS scavenging [301].
Chelation of Metal Ions
Under hyperglycemic conditions, transition metals are readily involved in Fenton reaction and catalyze monosaccaride autoxidation and lipid peroxidation in oxygenated solutions [361,362]. Instead, chelation of metal ions can prevent AGE formation. The chelating effects of phenolic acids and flavonoids are attributed to their catechol moiety [338,363]. Through this mechanism, a polyphenol-enriched extract from Guava leaves inhibited Amadori product formation in a dose-dependent manner [364]. Caffeic acid binds to iron ions and prevents the oxidative consequences of the Fenton reaction [365], as well as a citrus flavonoid, rutin [366]. Chlorogenic acid and isoferulic acid act as chelating agents [367,368,369].
Interference with RAGE Expression and Related Signaling Pathways
As was mentioned above, AGE-RAGE interaction, as well as the associated signaling pathways, plays an important role in onset and progress of diabetic complications. Thus, targeting these receptors might be another efficient mechanism to suppress physiological effects associated with glycation. Thus, attenuation of RAGE expression was reported for gallic acid [343]. In addition to the pronounced antioxidant activity, EGCG can interfere with the pathways, mediated by AGE-RAGE interaction [370]. Suppression of the AGE-RAGE axis by sulforaphane might be a novel therapeutic target for vascular injury in diabetes [371]. It is also assumed, that the ability of resveratrol to delay long-term diabetic complications is, at least partly, associated with enhancement of RAGE expression, induces by this stilbene [372]. Resveratrol inhibits the expression of RAGE by activation the expression of peroxisome proliferator-activated receptor-gamma (PPAR-γ) [373]. As far as the other phenolics are concerned, it was reported that curcumin acts via the same mechanism [374], and can also bind to RAGE with a strong affinity blocking the interaction with ligands [375]. Resveratrol can also interfere with AGE-RAGE signaling cascade [349]. In vivo study with the rats fed a high-fat diet, revealed a pronounced antidiabetic effect of cinnamaldehyde, probably through up-regulating the expression of proteins in the insulin signaling IRS1/PI3K/AKT pathway and alleviating the expression of RAGEs [376]. Besides this, daidzein and silibinin inhibit the NF-κB signaling pathway by activating NAD-dependent deacetylase sirtuin-1 (SIRT1) [354].
Inhibition of Aldose Reductase
Aldose reductase promotes the conversion of glucose to sorbitol and conversion of NADPH to NADP+ in the sorbitol pathway, where sorbitol is oxidized to fructose by sorbitol dehydrogenase [377]. In terms of protein glycation, fructose is dramatically more reactive than glucose [13]. This fact strongly accelerates AGE production and subsequently the progression of diabetic complications [301]. Some active flavonoids from empetrum nigrum, such as (quercetin, rutin, kaempferol) exhibited inhibitory effects on aldose reductase. 3,5-di-O-caffeoyl-epi-quinic acid from erigeron annuus was also reported to inhibit aldose reductase and AGE formation in rat lens [378]. EGCG and 6-gingerol demonstrated good aldose reductase inhibitory activity in vitro (human retinal pigment epithelial (HRPE) cells) and in vivo (a high fat diet-induced diabetic mouse model - male C57BL/6J mice) systems [379]. Additionally, resveratrol was found to inhibit aldose reductase, which reduces the development of AGEs in the kidney in diabetic rats and prevents opacification of the lens [380].
Effect on the Availability of Potential Glycation Sites in Proteins
Protein glycation can significantly alter the structure and function of proteins by changing the hydrophilicity or proportion of α-helices and β-sheets. Thus, antiglycative functions of some natural compounds can be achieved by competitive binding to proteins through van der Waals forces or hydrogen bonds [381]. For example, caffeic acid prevents BSA glycation through binding to Arg185 and Tyr137 by hydrogen bonds [382]. Analogously, syringic and chlorogenic acids can protect the protein structural integrity from modifications, but syringic acid is more active than chlorogenic acid because of its smaller molecular size [383].
Reducing of Blood Glucose Levels
Multiple natural compounds including plant secondary metabolites inhibit α-glucosidase and α-amylase. For example, the flavonoid-rich extract from Annona muricata Linn. showed inhibitory activity against α-glucosidase and α-amylase [384]. Kaempferol monoglycosides inhibit these enzymes through hydrogen bonding [385]. Tannic acid was also demonstrated a strong ability for efficient reversible inhibition of α-glucosidase. The interaction between the polyphenol and the protein was shown by molecular docking, and appeared to be driven by hydrogen bonds, hydrophobic forces and electrostatic interactions [386]. Also, diosgenin, extracted from fenugreek, reduced blood glucose levels by stimulating insulin secretion [387].
5. Other Prospective Antiglycative Mechanisms Associated with Metabolite-Derived Pro- Tein Adducts – Future Perspectives
Plant secondary metabolites (such as polyphenols) are known to interact with proteins via either non-covalent or covalent interactions altering their structure and properties [388]. By computationally driven analysis of the polyphenol-protein interactome was found that the proteins of human plasma and cellular signaling polypeptides are involved in such interactions, which affect multiple metabolic pathways [389]. The underlying non-covalent interactions are usually reversible and occur via hydrophobic, hydrogen bonding, electrostatic, and van der Waals-related mechanisms. Among them, hydrogen bonding and hydrophobic interactions can be highlighted as the major mechanisms involved in the non-covalent formation of complexes between proteins and polyphenols [390]. On the other hand, oxidized polyphenols and electrophilic organosulphur metabolites from garlic, chives or papaya pulp are known to form covalent bonds with proteins.
Polyphenols with a catechol ring can be oxidized enzymatically (e.g. by polyphenol oxidases) or by autoxidation (e.g. in an alkaline environment or in the presence of oxidizing agents) to yield electrophilic o-quinones [187]. Such quinones can react further with nucleophilic residues of proteins or peptides (thiol, amino, guanidine, or imidazole) by Michael addition [391]. Free thiol groups on proteins have been shown to be kinetically preferred targets for quinones [392]. Besides, imino-quinone adducts can be formed by direct reaction between the amino group of L-lysine with the carbonyl group of quinones [393]. Moreover, polyphenol-protein complexes may also be derived by either covalently binding semiquinone radical intermediates to protein nucleophilic residues [394], or by covalently attachment protein radicals to polyphenols at the ortho- or para-positions of their hydroxyl groups [395].
The interactions between plant secondary metabolites and proteins can occur in human body as well as during food processing. In both cases, this process is strongly affected by different factors such as the concentration, molecular weight and structure of the polyphenol and protein interaction partners, as well as pH, temperature, ions/cofactors and physiological status determining the precipitation or solubility of the resulting complexes, that may affect their nutritional and functional properties as well as their biological activities [390].
As was shown recently, protein conjugates formed by plant metabolites demonstrate antioxidant [396], anti-inflammatory [397], anti-cancer [398] and anti-allergic activities [399]. Thus, it was found that covalent conjugates of soy protein isolate with black rice anthocyanins exhibit pronounced antioxidant capacity, which was higher in comparison to that of the polyphenols themselves [388]. The hypoallergenic effects of β-lactoglobulin-EGCG, β-lactoglobulin -CA conjugates were addressed in the binding tests with immunoglobin E (IgE) antibodies and by comparison with control β-lactoglobulin [399]. Further, protein conjugates of curcumin showed a tumor suppression effect in an in vivo study on immunocompromised mice with breast cancer [400].
In silico approach also proved to be efficient in studies of biological activities exerted by protein conjugates of plant metabolites. Thus, kinetics analysis and molecular docking accomplished with EGCG conjugates showed, that hydrogen bonds played a critical role in the molecular interaction, and demonstrated possible application of such adducts as natural α-glucosidase noncompetitive inhibitors [401]. Therefore, the possible mechanisms behind the hypoglycemic activity of EGCG conjugates were revealed with potential applications in the development of functional food [402]. In this context, the antidiabetic activity of such protein complexes needs more in-depth studies.
The potential of plant metabolites to form complex protein adducts with dicarbonyl compounds cannot be excluded, especially during food processing. Thus, these proposed adducts might be also involved in anti-inflammatory/antioxidant cell response especially in the context of AGE/RAGE axis. However, this aspect and structure or biological properties of such plant metabolite adducts are still mostly unstudied. Additionally, the underlying molecular mechanisms of formation protein adducts by plant secondary metabolites can be extensively investigated using molecular docking studies, which are able to provide large scale information to determine possible binding sites and conformations of conjugates. Moreover, in vitro and in vivo studies are needed for a comprehensive understanding of their physiological effects for developing anti-ageing, anti-diabetic and anti-neurodegenerative promising compounds and nutrition strategies in the food and pharmaceutical industries.
6. Conclusions
Diabetes mellitus (DM) is a multifactorial metabolic disease that imposes increasing health and economic burdens worldwide. Conventional therapies, while efficient to a certain degree, often cause side effects and fail to prevent long-term complications related to chronic hyperglycemia. In this context, plant-derived secondary metabolites represent an appealing complementary or alternative strategy. These natural compounds – phenolics, alkaloids, terpenes, organosulfur compounds, and others – demonstrate significant potential in improving insulin sensitivity, enhancing glucose uptake, modulating key regulatory pathways, and inhibiting enzymes involved in carbohydrate digestion. Furthermore, their strong antioxidant and antiglycative properties allow them mitigating oxidative stress and suppress the formation of advanced glycation end products (AGEs), which are directly implicated in diabetic complications. By influencing the AGE-RAGE axis, Nrf2 and NF-κB pathways, and interfering with protein tyrosine phosphatase and other critical molecular targets, these metabolites can break vicious cycles of inflammation, oxidative damage, and metabolic dysfunction. However, future research should focus on clarifying their pharmacokinetics, bioavailability, and safety profiles through well-designed in vivo studies and clinical trials. Optimizing delivery systems, understanding structure-activity relationships, and exploring synergistic effects with conventional drugs will be essential. Ultimately, integrating plant-derived natural products into therapeutic and preventive strategies may provide safer, more holistic control over diabetes and its associated complications.
Author Contributions
Conceptualization, RT and AF; writing first manuscript draft, KA, NG, NE and ER; editing and preparation of the final manuscript, KA, RT and AF.
Funding
This study was supported by the Russian Science Foundation (grant # 23-14-00383).
Acknowledgments
The The work was infrastructurally supported by the Ministry of Science and Higher Education of the Russian Federation (theme # 122042600086-7).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kaul, K.; Tarr, J.M.; Ahmad, S.I.; Kohner, E.M.; Chibber, R. Introduction to diabetes mellitus. Diabetes: an old disease, a new insight 2013, 1–11. [Google Scholar]
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Epidemiology of diabetes and diabetes-related complications. Physical therapy 2008, 88, 1254–1264. [Google Scholar] [CrossRef]
- Shahid, R.K.; Ahmed, S.; Le, D.; Yadav, S. Diabetes and cancer: risk, challenges, management and outcomes. Cancers 2021, 13, 5735. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.; Chávez-Castillo, M.; Bautista, J.; Ortega, Á.; Nava, M.; Salazar, J.; Díaz-Camargo, E.; Medina, O.; Rojas-Quintero, J.; Bermúdez, V. Alzheimer’s disease and type 2 diabetes mellitus: Pathophysiologic and pharmacotherapeutics links. World Journal of Diabetes 2021, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Bonner, R.; Albajrami, O.; Hudspeth, J.; Upadhyay, A. Diabetic kidney disease. Primary Care: Clinics in Office Practice 2020, 47, 645–659. [Google Scholar] [CrossRef] [PubMed]
- https://idf.org/about-diabetes/facts-figures/.
- https://www.paho.org/en/topics/diabetes.
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nature reviews endocrinology 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Kaur, A.; Thind, S.; Singh, B.; Raina, S. Advanced glycation End-products (AGEs): an emerging concern for processed food industries. Journal of food science and technology 2015, 52, 7561–7576. [Google Scholar] [CrossRef] [PubMed]
- Magiera, A.; Kołodziejczyk-Czepas, J.; Skrobacz, K.; Czerwińska, M.E.; Rutkowska, M.; Prokop, A.; Michel, P.; Olszewska, M.A. Valorisation of the Inhibitory Potential of Fresh and Dried Fruit Extracts of Prunus spinosa L. towards Carbohydrate Hydrolysing Enzymes, Protein Glycation, Multiple Oxidants and Oxidative Stress-Induced Changes in Human Plasma Constituents. Pharmaceuticals 2022, 15, 1300. [Google Scholar] [CrossRef]
- Monnier, V.M.; Sell, D.R.; Nagaraj, R.H.; Miyata, S.; Grandhee, S.; Odetti, P.; Ibrahim, S.A. Maillard reaction-mediated molecular damage to extracellular matrix and other tissue proteins in diabetes, aging, and uremia. Diabetes 1992, 41, 36–41. [Google Scholar] [CrossRef]
- Thornalley, P.J. Protein and nucleotide damage by glyoxal and methylglyoxal in physiological systems-role in ageing and disease. Drug metabolism and drug interactions 2008, 23, 125–150. [Google Scholar] [CrossRef]
- Bilova, T.; Lukasheva, E.; Brauch, D.; Greifenhagen, U.; Paudel, G.; Tarakhovskaya, E.; Frolova, N.; Mittasch, J.; Balcke, G.U.; Tissier, A. A snapshot of the plant glycated proteome: structural, functional, and mechanistic aspects. Journal of Biological Chemistry 2016, 291, 7621–7636. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Hanke, T.; Frolov, A.; Langrock, T.; Hoffmann, R.; Fischer, C.; Schwarzenbolz, U.; Henle, T.; Born, R.; Worch, H. Modification of collagen in vitro with respect to formation of Nɛ-carboxymethyllysine. International Journal of Biological Macromolecules 2009, 44, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circulation research 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Greifenhagen, U.; Frolov, A.; Blüher, M.; Hoffmann, R. Site-specific analysis of advanced glycation end products in plasma proteins of type 2 diabetes mellitus patients. Analytical and Bioanalytical Chemistry 2016, 408, 5557–5566. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Coughlan, M.T. Dietary advanced glycation end products: digestion, metabolism and modulation of gut microbial ecology. Nutrients 2019, 11, 215. [Google Scholar] [CrossRef]
- Lyons, T.J.; Jenkins, A.J. Glycation, oxidation, and lipoxidation in the development of the complications of diabetes: a carbonyl stress hypothesis. Diabetes reviews (Alexandria, Va.) 1997, 5, 365. [Google Scholar] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxidative medicine and cellular longevity 2019, 2019, 3085756. [Google Scholar] [CrossRef] [PubMed]
- Huebschmann, A.G.; Regensteiner, J.G.; Vlassara, H.; Reusch, J.E. Diabetes and advanced glycoxidation end products. Diabetes care 2006, 29, 1420–1432. [Google Scholar] [CrossRef]
- Association, A.D. 2. Classification and diagnosis of diabetes. Diabetes care 2015, 38, S8–S16. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M. Highlighting diabetes mellitus: the epidemic continues. Arteriosclerosis, thrombosis, and vascular biology 2018, 38, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Padhi, S.; Nayak, A.K.; Behera, A. Type II diabetes mellitus: a review on recent drug based therapeutics. Biomedicine & Pharmacotherapy 2020, 131, 110708. [Google Scholar]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. The lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.B.; Smith, M.S. Obesity statistics. Primary care: clinics in office practice 2016, 43, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Dubourg, J.; Fouqueray, P.; Quinslot, D.; Grouin, J.M.; Kaku, K. Long-term safety and efficacy of imeglimin as monotherapy or in combination with existing antidiabetic agents in Japanese patients with type 2 diabetes (TIMES 2): A 52-week, open-label, multicentre phase 3 trial. Diabetes, Obesity and Metabolism 2022, 24, 609–619. [Google Scholar] [CrossRef]
- Dahlén, A.D.; Dashi, G.; Maslov, I.; Attwood, M.M.; Jonsson, J.; Trukhan, V.; Schiöth, H.B. Trends in antidiabetic drug discovery: FDA approved drugs, new drugs in clinical trials and global sales. Frontiers in Pharmacology 2022, 12, 807548. [Google Scholar] [CrossRef]
- Li, G.Q.; Kam, A.; Wong, K.H.; Zhou, X.; Omar, E.A.; Alqahtani, A.; Li, K.M.; Razmovski-Naumovski, V.; Chan, K. Herbal medicines for the management of diabetes. Diabetes: An Old Disease, a New Insight 2013, 396–413. [Google Scholar]
- Albright, R.H.; Fleischer, A.E. Association of select preventative services and hospitalization in people with diabetes. Journal of Diabetes and its Complications 2021, 35, 107903. [Google Scholar] [CrossRef]
- Krejčí, H.; Vyjídák, J.; Kohutiar, M. Low-carbohydrate diet in diabetes mellitus treatment. Vnitrni lekarstvi 2018, 64, 742–752. [Google Scholar] [CrossRef]
- Sofowora, A.; Ogunbodede, E.; Onayade, A. The role and place of medicinal plants in the strategies for disease prevention. African journal of traditional, complementary and alternative medicines 2013, 10, 210–229. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacological research 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Кoрейба, К.А.; Цыплакoв, Д.Э.; Минабутдинoв, А.Р.; Агаджанoва, К.В. Испoльзoвание натуральных растительных ингредиентoв при лечении бoльных сахарным диабетoм. Медицина. Сoциoлoгия. Филoсoфия. Прикладные исследoвания 2020, 49. [Google Scholar]
- Patel, D.; Prasad, S.K.; Kumar, R.; Hemalatha, S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pacific journal of tropical biomedicine 2012, 2, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Kifle, Z.D.; Belayneh, Y.M. Antidiabetic and anti-hyperlipidemic effects of the crude hydromethanol extract of Hagenia abyssinica (Rosaceae) leaves in streptozotocin-induced diabetic mice. Diabetes, Metabolic Syndrome and Obesity 2020, 4085–4094. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, Y.; Li, L. Coumarins as potential antidiabetic agents. Journal of Pharmacy and Pharmacology 2017, 69, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, J.T.M.; Shrestha, H.; Prajapati, M.; Karkee, A.; Maharjan, A. Adverse effects of oral hypoglycemic agents and adherence to them among patients with type 2 diabetes mellitus in Nepal. Journal of Lumbini Medical College 2017, 5, 34–40. [Google Scholar] [CrossRef]
- Zhang, Y.-S.; Zheng, Y.-D.; Yuan, Y.; Chen, S.-C.; Xie, B.-C. Effects of anti-diabetic drugs on fracture risk: a systematic review and network meta-analysis. Frontiers in Endocrinology 2021, 12, 735824. [Google Scholar] [CrossRef]
- Triggle, C.; Ding, H. Metformin is not just an antihyperglycaemic drug but also has protective effects on the vascular endothelium. Acta physiologica 2017, 219, 138–151. [Google Scholar] [CrossRef]
- Sanchez-Rangel, E.; Inzucchi, S.E. Metformin: clinical use in type 2 diabetes. Diabetologia 2017, 60, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Ленская, К.В.; Спасoв, А.; Чепляева, Н. Иннoвациoнные направления пoиска лекарственных препаратoв для лечения сахарнoгo диабета типа 2. Вестник вoлгoградскoгo гoсударственнoгo медицинскoгo университета 2011, 10–18. [Google Scholar]
- Ali, M.Y.; Paul, S.; Tanvir, E.; Hossen, M.S.; Rumpa, N.-E.N.; Saha, M.; Bhoumik, N.C.; Aminul Islam, M.; Hossain, M.S.; Alam, N. Antihyperglycemic, antidiabetic, and antioxidant effects of Garcinia pedunculata in rats. Evidence-Based Complementary and Alternative Medicine 2017, 2017, 2979760. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: plant secondary metabolites and the enhancement of human brain function. Advances in Nutrition 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A. High-temperature stress and metabolism of secondary metabolites in plants. Effect of high temperature on crop productivity and metabolism of macro molecules 2019, 391–484. [Google Scholar]
- Chauhan, D.S.; Gupta, P.; Pottoo, F.H.; Amir, M. Secondary metabolites in the treatment of diabetes mellitus: a paradigm shift. Current Drug Metabolism 2020, 21, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Shehadeh, M.B.; Suaifan, G.A.; Abu-Odeh, A.M. Plants secondary metabolites as blood glucose-lowering molecules. Molecules 2021, 26, 4333. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: a historical perspective. Plant science 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Chiocchio, I.; Mandrone, M.; Tomasi, P.; Marincich, L.; Poli, F. Plant secondary metabolites: An opportunity for circular economy. Molecules 2021, 26, 495. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative medicine and cellular longevity 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Velderrain-Rodríguez, G.; Palafox-Carlos, H.; Wall-Medrano, A.; Ayala-Zavala, J.; Chen, C.O.; Robles-Sánchez, M.; Astiazaran-García, H.; Alvarez-Parrilla, E.; González-Aguilar, G. Phenolic compounds: their journey after intake. Food & function 2014, 5, 189–197. [Google Scholar]
- Herrmann, K.M. The shikimate pathway as an entry to aromatic secondary metabolism. Plant physiology 1995, 107, 7. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Molecular plant 2010, 3, 2–20. [Google Scholar] [CrossRef]
- da Silva, A.P.G.; Sganzerla, W.G.; John, O.D.; Marchiosi, R. A comprehensive review of the classification, sources, biosynthesis, and biological properties of hydroxybenzoic and hydroxycinnamic acids. Phytochemistry Reviews 2023, 1–30. [Google Scholar] [CrossRef]
- Flores-Morales, V.; Villasana-Ruíz, A.P.; Garza-Veloz, I.; González-Delgado, S.; Martinez-Fierro, M.L. Therapeutic effects of coumarins with different substitution patterns. Molecules 2023, 28, 2413. [Google Scholar] [CrossRef]
- Thakral, S.; Singh, V. 2, 4-Dichloro-5-[(N-aryl/alkyl) sulfamoyl] benzoic acid derivatives: in vitro antidiabetic activity, molecular modeling and in silico ADMET screening. Medicinal Chemistry 2019, 15, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Aicher, T.D.; Bebernitz, G.R.; Bell, P.A.; Brand, L.J.; Dain, J.G.; Deems, R.; Fillers, W.S.; Foley, J.E.; Knorr, D.C.; Nadelson, J. Hypoglycemic prodrugs of 4-(2, 2-dimethyl-1-oxopropyl) benzoic acid. Journal of medicinal chemistry 1999, 42, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Deng, X.; Jiang, N.; Meng, L.; Xing, J.; Jiang, W.; Xu, Y. Identification and structure–activity relationship exploration of uracil-based benzoic acid and ester derivatives as novel dipeptidyl peptidase-4 inhibitors for the treatment of type 2 diabetes mellitus. European Journal of Medicinal Chemistry 2021, 225, 113765. [Google Scholar] [CrossRef]
- Jaradat, N.; Khasati, A.; Hawi, M.; Hawash, M.; Shekfeh, S.; Qneibi, M.; Eid, A.M.; Arar, M.; Qaoud, M.T. Antidiabetic, antioxidant, and anti-obesity effects of phenylthio-ethyl benzoate derivatives, and molecular docking study regarding α-amylase enzyme. Scientific Reports 2022, 12, 3108. [Google Scholar] [CrossRef] [PubMed]
- Kougan, G.B.; Tabopda, T.; Kuete, V.; Verpoorte, R. Simple phenols, phenolic acids, and related esters from the medicinal plants of Africa. In Medicinal plant research in Africa; Elsevier: 2013, pp. 225–249.
- Naz, D.; Muhamad, A.; Zeb, A.; Shah, I. In vitro and in vivo antidiabetic properties of phenolic antioxidants from Sedum adenotrichum. Frontiers in Nutrition 2019, 6, 177. [Google Scholar] [CrossRef]
- Street, R.A.; Sidana, J.; Prinsloo, G. Cichorium intybus: Traditional uses, phytochemistry, pharmacology, and toxicology. Evidence-based Complementary and Alternative Medicine 2013, 2013, 579319. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, Y.-L.; Wang, M.; Wang, J.-J.; Liu, T.; Yu, J. The Angelica dahurica: A review of traditional uses, phytochemistry and pharmacology. Frontiers in Pharmacology 2022, 13, 896637. [Google Scholar] [CrossRef] [PubMed]
- Arichi, H.; Kimura, Y.; Okuda, H.; Baba, K.; Kozawa, M.; Arichi, S. Effects of stilbene components of the roots of Polygonum cuspidatum Sieb. et Zucc. on lipid metabolism. Chemical and Pharmaceutical Bulletin 1982, 30, 1766–1770. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, O.D.; Kulkarni, Y.A. Treatment with Terminalia chebula Extract Reduces Insulin Resistance, Hyperglycemia and Improves SIRT1 Expression in Type 2 Diabetic Rats. Life 2023, 13, 1168. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi.(Lamiaceae): a review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Journal of Pharmacy and Pharmacology 2019, 71, 1353–1369. [Google Scholar] [CrossRef]
- Chen, C.; Mohamad Razali, U.H.; Saikim, F.H.; Mahyudin, A.; Mohd Noor, N.Q.I. Morus alba L. plant: Bioactive compounds and potential as a functional food ingredient. Foods 2021, 10, 689. [Google Scholar] [CrossRef] [PubMed]
- Bürkel, P.; Rajbhandari, M.; Jürgenliemk, G. Bassia longifolia (= Madhuca longifolia): Isolation of flavan-3-ols and their contribution to the antibacterial and antidiabetic activity in vitro. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Licciardello, C.; Russo, M.; Reforgiato Recupero, G.; Muccilli, V.; Cunsolo, V.; Saletti, R.; Foti, S.; Fontanini, D. Analysis of Citrus sinensis L.(Osbeck) flesh proteome at maturity time. Acta horticulturae 2011, 892, 343–349. [Google Scholar] [CrossRef]
- Huang, S.-S.; Su, S.-Y.; Chang, J.-S.; Lin, H.-J.; Wu, W.-T.; Deng, J.-S.; Huang, G.-J. Antioxidants, anti-inflammatory, and antidiabetic effects of the aqueous extracts from Glycine species and its bioactive compounds. Botanical studies 2016, 57, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.H.; Ribnicky, D.M.; Kuhn, P.; Poulev, A.; Logendra, S.; Yousef, G.G.; Raskin, I.; Lila, M.A. Hypoglycemic activity of a novel anthocyanin-rich formulation from lowbush blueberry, Vaccinium angustifolium Aiton. Phytomedicine 2009, 16, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Wahab, S.; Annadurai, S.; Abullais, S.S.; Das, G.; Ahmad, W.; Ahmad, M.F.; Kandasamy, G.; Vasudevan, R.; Ali, M.S.; Amir, M. Glycyrrhiza glabra (Licorice): A comprehensive review on its phytochemistry, biological activities, clinical evidence and toxicology. Plants 2021, 10, 2751. [Google Scholar] [CrossRef] [PubMed]
- Mechchate, H.; Es-Safi, I.; Conte, R.; Hano, C.; Amaghnouje, A.; Jawhari, F.Z.; Radouane, N.; Bencheikh, N.; Grafov, A.; Bousta, D. In vivo and in vitro antidiabetic and anti-inflammatory properties of flax (Linum usitatissimum L.) seed polyphenols. Nutrients 2021, 13, 2759. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Zhao, F.; Wang, Z.; Wang, Q.; Chai, X.; Hou, G.; Meng, Q. Sesame (Sesamum indicum L.): A comprehensive review of nutritional value, phytochemical composition, health benefits, development of food, and industrial applications. Nutrients 2022, 14, 4079. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, T.; Wang, X.; Sun, J. Research progress of coumarins and their derivatives in the treatment of diabetes. Journal of Enzyme Inhibition and Medicinal Chemistry 2022, 37, 616–628. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Cruz-Martins, N.; López-Jornet, P.; Lopez, E.P.-F.; Harun, N.; Yeskaliyeva, B.; Beyatli, A.; Sytar, O.; Shaheen, S.; Sharopov, F. Natural coumarins: exploring the pharmacological complexity and underlying molecular mechanisms. Oxidative Medicine and Cellular Longevity 2021, 2021, 6492346. [Google Scholar] [CrossRef] [PubMed]
- Jasemi, S.V.; Khazaei, H.; Morovati, M.R.; Joshi, T.; Aneva, I.Y.; Farzaei, M.H.; Echeverría, J. Phytochemicals as treatment for allergic asthma: Therapeutic effects and mechanisms of action. Phytomedicine 2024, 122, 155149. [Google Scholar] [CrossRef]
- Bourgaud, F.; Hehn, A.; Larbat, R.; Doerper, S.; Gontier, E.; Kellner, S.; Matern, U. Biosynthesis of coumarins in plants: a major pathway still to be unravelled for cytochrome P450 enzymes. Phytochemistry Reviews 2006, 5, 293–308. [Google Scholar] [CrossRef]
- Evans, W.C. Trease and Evans' pharmacognosy; Elsevier Health Sciences: 2009.
- Kashman, Y.; Gustafson, K.R.; Fuller, R.; Cardellina, J., 2nd; McMahon, J.; Currens, M.; Buckheit Jr, R.; Hughes, S.; Cragg, G.; Boyd, M. The calanolides, a novel HIV-inhibitory class of coumarin derivatives from the tropical rainforest tree, Calophyllum lanigerum. Journal of medicinal chemistry 1992, 35, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.A.; Ishibashi, M. Target Protein-Oriented Natural Product Isolation Methods. 2020.
- Lu, C.-L.; Li, Y.-M.; Fu, G.-Q.; Yang, L.; Jiang, J.-G.; Zhu, L.; Lin, F.-L.; Chen, J.; Lin, Q.-S. Extraction optimisation of daphnoretin from root bark of Wikstroemia indica (L.) CA and its anti-tumour activity tests. Food Chemistry 2011, 124, 1500–1506. [Google Scholar] [CrossRef]
- Lončarić, M.; Gašo-Sokač, D.; Jokić, S.; Molnar, M. Recent advances in the synthesis of coumarin derivatives from different starting materials. Biomolecules 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Guliani, E.; Bajaj, K. Coumarin—Synthetic Methodologies, Pharmacology, and Application as Natural Fluorophore. Topics in Current Chemistry 2024, 382, 1–38. [Google Scholar] [CrossRef]
- Ranđelović, S.; Bipat, R. A review of coumarins and coumarin-related compounds for their potential antidiabetic effect. Clinical Medicine Insights: Endocrinology and Diabetes 2021, 14, 11795514211042023. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.; Shah, S.A.A.; Afifi, M.; Imran, S.; Sultan, S.; Rahim, F.; Khan, K.M. Synthesis, α-glucosidase inhibition and molecular docking study of coumarin based derivatives. Bioorganic Chemistry 2018, 77, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Basappa, V.C.; Kameshwar, V.H.; Kumara, K.; Achutha, D.K.; Krishnappagowda, L.N.; Kariyappa, A.K. Design and synthesis of coumarin-triazole hybrids: biocompatible anti-diabetic agents, in silico molecular docking and ADME screening. Heliyon 2020, 6. [Google Scholar]
- Kottaisamy, C.P.D.; Raj, D.S.; Prasanth Kumar, V.; Sankaran, U. Experimental animal models for diabetes and its related complications—a review. Laboratory animal research 2021, 37, 23. [Google Scholar] [CrossRef]
- Parage, C.; Tavares, R.; Réty, S.; Baltenweck-Guyot, R.; Poutaraud, A.; Renault, L.; Heintz, D.; Lugan, R.; Marais, G.A.; Aubourg, S. Structural, functional, and evolutionary analysis of the unusually large stilbene synthase gene family in grapevine. Plant Physiology 2012, 160, 1407–1419. [Google Scholar] [CrossRef]
- Peng, W.; Qin, R.; Li, X.; Zhou, H. Botany, phytochemistry, pharmacology, and potential application of Polygonum cuspidatum Sieb. et Zucc.: a review. Journal of ethnopharmacology 2013, 148, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, H.; Wu, Z.; Fan, H.; Wang, G.; Chai, T.; Wang, H. Tissue-specific transcriptome analyses reveal candidate genes for stilbene, flavonoid and anthraquinone biosynthesis in the medicinal plant Polygonum cuspidatum. BMC genomics 2021, 22, 353. [Google Scholar] [CrossRef]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant science 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: the in vivo evidence. Nature reviews Drug discovery 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Annals of the New York Academy of Sciences 2011, 1215, 161–169. [Google Scholar] [CrossRef]
- Côté, C.D.; Rasmussen, B.A.; Duca, F.A.; Zadeh-Tahmasebi, M.; Baur, J.A.; Daljeet, M.; Breen, D.M.; Filippi, B.M.; Lam, T.K. Resveratrol activates duodenal Sirt1 to reverse insulin resistance in rats through a neuronal network. Nature medicine 2015, 21, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. British journal of nutrition 2011, 106, 383–389. [Google Scholar] [CrossRef]
- Shamsudin, N.F.; Ahmed, Q.U.; Mahmood, S.; Shah, S.A.A.; Sarian, M.N.; Khattak, M.M.A.K.; Khatib, A.; Sabere, A.S.M.; Yusoff, Y.M.; Latip, J. Flavonoids as antidiabetic and anti-inflammatory agents: A review on structural activity relationship-based studies and meta-analysis. International journal of molecular sciences 2022, 23, 12605. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The flavonoid biosynthesis network in plants. International journal of molecular sciences 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Syed Mohamad, S.N.A.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. BioMed research international 2017, 2017, 8386065. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Zhang, W.; Sun-Waterhouse, D.; Jiang, Y.; Li, F.; Waterhouse, G.I.; Li, D. Phenolic-protein interactions in foods and post ingestion: Switches empowering health outcomes. Trends in Food Science & Technology 2021, 118, 71–86. [Google Scholar]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnology advances 2020, 38, 107316. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.S.; Thomas, M.; Clarke, D.J. The gene stlA encodes a phenylalanine ammonia-lyase that is involved in the production of a stilbene antibiotic in Photorhabdus luminescens TT01. Microbiology 2005, 151, 2543–2550. [Google Scholar] [CrossRef]
- Wohl, J.; Petersen, M. Functional expression and characterization of cinnamic acid 4-hydroxylase from the hornwort Anthoceros agrestis in Physcomitrella patens. Plant cell reports 2020, 39, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, D.; Zhang, Z.; Jiang, Y.; Liu, Y. Isoliquiritigenin alleviates diabetic symptoms via activating AMPK and inhibiting mTORC1 signaling in diet-induced diabetic mice. Phytomedicine 2022, 98, 153950. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, R.M.; Venkatraman, A.C. Polyphenols activate energy sensing network in insulin resistant models. Chemico-biological interactions 2017, 275, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Ashour, M.B.; Abdel-Moneim, A.; Ahmed, O.M. Hesperidin and naringin attenuate hyperglycemia-mediated oxidative stress and proinflammatory cytokine production in high fat fed/streptozotocin-induced type 2 diabetic rats. Journal of Diabetes and its Complications 2012, 26, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef]
- Malik, S.; Suchal, K.; Khan, S.I.; Bhatia, J.; Kishore, K.; Dinda, A.K.; Arya, D.S. Apigenin ameliorates streptozotocin-induced diabetic nephropathy in rats via MAPK-NF-κB-TNF-α and TGF-β1-MAPK-fibronectin pathways. American Journal of Physiology-Renal Physiology 2017, 313, F414–F422. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-Y.; Jung, U.J.; Park, T.; Yun, J.W.; Choi, M.-S. Luteolin attenuates hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue in mice with diet-induced obesity. Diabetes 2015, 64, 1658–1669. [Google Scholar] [CrossRef]
- Chu, S.; Wang, J.; Zhu, Y.; Liu, S.; Zhou, X.; Zhang, H.; Wang, C.-e.; Yang, W.; Tian, Z.; Cheng, H. An R2R3-type MYB transcription factor, GmMYB29, regulates isoflavone biosynthesis in soybean. PLoS genetics 2017, 13, e1006770. [Google Scholar] [CrossRef]
- Rasheed, S.; Rehman, K.; Shahid, M.; Suhail, S.; Akash, M.S.H. Therapeutic potentials of genistein: New insights and perspectives. Journal of Food Biochemistry 2022, 46, e14228. [Google Scholar] [CrossRef]
- Choi, E.J.; Kim, G.-H. Daidzein causes cell cycle arrest at the G1 and G2/M phases in human breast cancer MCF-7 and MDA-MB-453 cells. Phytomedicine 2008, 15, 683–690. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and β-cell damage in rat pancreas. Pharmacological research 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Lee, Y.J.; Suh, K.S.; Choi, M.C.; Chon, S.; Oh, S.; Woo, J.T.; Kim, S.W.; Kim, J.W.; Kim, Y.S. Kaempferol protects HIT-T15 pancreatic beta cells from 2-deoxy-D-ribose-induced oxidative damage. Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives 2010, 24, 419–423. [Google Scholar] [CrossRef]
- Liu, I.-M.; Liou, S.-S.; Lan, T.-W.; Hsu, F.-L.; Cheng, J.-T. Myricetin as the active principle of Abelmoschus moschatus to lower plasma glucose in streptozotocin-induced diabetic rats. Planta medica 2005, 71, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Naing, A.H.; Kim, C.K. Roles of R2R3-MYB transcription factors in transcriptional regulation of anthocyanin biosynthesis in horticultural plants. Plant molecular biology 2018, 98, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Brugliera, F.; Tao, G.-Q.; Tems, U.; Kalc, G.; Mouradova, E.; Price, K.; Stevenson, K.; Nakamura, N.; Stacey, I.; Katsumoto, Y. Violet/blue chrysanthemums—metabolic engineering of the anthocyanin biosynthetic pathway results in novel petal colors. Plant and Cell Physiology 2013, 54, 1696–1710. [Google Scholar] [CrossRef]
- Tian, L.; Ning, H.; Shao, W.; Song, Z.; Badakhshi, Y.; Ling, W.; Yang, B.B.; Brubaker, P.L.; Jin, T. Dietary cyanidin-3-glucoside attenuates high-fat-diet–induced body-weight gain and impairment of glucose tolerance in mice via effects on the hepatic hormone FGF21. The Journal of nutrition 2020, 150, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Xie, D.Y.; Sharma, S.B. Proanthocyanidins–a final frontier in flavonoid research? New phytologist 2005, 165, 9–28. [Google Scholar] [CrossRef]
- Omar, N.; Ismail, C.A.N.; Long, I. Tannins in the treatment of diabetic neuropathic pain: Research progress and future challenges. Frontiers in Pharmacology 2022, 12, 805854. [Google Scholar] [CrossRef]
- Laddha, A.P.; Kulkarni, Y.A. Tannins and vascular complications of Diabetes: An update. Phytomedicine 2019, 56, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Porter, L.J. Structure and chemical properties of the condensed tannins. In Plant Polyphenols: Synthesis, Properties, Significance; Springer: 1992, pp. 245–258.
- Thomas, M.M.G.; Barbosa Filho, J. Anti-inflammatory actions of tannins isolated from the bark of Anacardwm occidentale L. Journal of ethnopharmacology 1985, 13, 289–300. [Google Scholar] [CrossRef]
- Doss, A.; Mubarack, H.M.; Dhanabalan, R. Antibacterial activity of tannins from the leaves of Solanum trilobatum Linn. Indian J Sci Technol 2009, 2, 41–43. [Google Scholar] [CrossRef]
- Fidelis, M.; Santos, J.S.; Escher, G.B.; do Carmo, M.V.; Azevedo, L.; da Silva, M.C.; Putnik, P.; Granato, D. In vitro antioxidant and antihypertensive compounds from camu-camu (Myrciaria dubia McVaugh, Myrtaceae) seed coat: A multivariate structure-activity study. Food and Chemical Toxicology 2018, 120, 479–490. [Google Scholar] [CrossRef]
- Kunyanga, C.N.; Imungi, J.K.; Okoth, M.; Momanyi, C.; Biesalski, H.K.; Vadivel, V. Antioxidant and antidiabetic properties of condensed tannins in acetonic extract of selected raw and processed indigenous food ingredients from Kenya. Journal of food science 2011, 76, C560–C567. [Google Scholar] [CrossRef]
- Ajebli, M.; Eddouks, M. The promising role of plant tannins as bioactive antidiabetic agents. Current medicinal chemistry 2019, 26, 4852–4884. [Google Scholar] [CrossRef] [PubMed]
- Perchellet, J.-P.; Gali, H.U.; Perchellet, E.M.; Klish, D.S.; Armbrust, A.D. Antitumor-promoting activities of tannic acid, ellagic acid, and several gallic acid derivatives in mouse skin. Plant Polyphenols: Synthesis, Properties, Significance 1992, 783–801. [Google Scholar]
- Chandak, P.G.; Gaikwad, A.B.; Tikoo, K. Gallotannin ameliorates the development of streptozotocin-induced diabetic nephropathy by preventing the activation of PARP. Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives 2009, 23, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Al-Salih, R. Clinical experimental evidence: synergistic effect of Gallic acid and tannic acid as Antidiabetic and antioxidant agents. Thi-Qar Med. J 2010, 4, 109–119. [Google Scholar]
- Fachriyah, E.; Eviana, I.; Eldiana, O.; Amaliyah, N.; Sektianingrum, A. Antidiabetic activity from gallic acid encapsulated nanochitosan. In Proceedings of the IOP Conference Series: Materials Science and Engineering; 2017; p. 012042. [Google Scholar]
- De la Iglesia, R.; Milagro, F.I.; Campión, J.; Boqué, N.; Martínez, J.A. Healthy properties of proanthocyanidins. Biofactors 2010, 36, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Cai, X.; Dai, X.; Ding, Y.; Jiang, Y.; Li, Y.; Zhang, Z.; Li, Y. Grape seed proanthocyanidin extracts ameliorate podocyte injury by activating peroxisome proliferator-activated receptor-γ coactivator 1α in low-dose streptozotocin-and high-carbohydrate/high-fat diet-induced diabetic rats. Food & function 2014, 5, 1872–1880. [Google Scholar]
- Barker, D. Lignans. 2019, 24, 1424.
- Hano, C.F.; Dinkova-Kostova, A.T.; Davin, L.B.; Cort, J.R.; Lewis, N.G. Lignans: Insights into their biosynthesis, metabolic engineering, analytical methods and health benefits. 2021, 11, 630327. [Google Scholar]
- Thompson, L.U.; Robb, P.; Serraino, M.; Cheung, F. Mammalian lignan production from various foods. 1991.
- Álvarez-Caballero, J.M.; Coy-Barrera, E. Lignans. In Antioxidants Effects in Health; Elsevier: 2022, pp. 387–416.
- Lampe, J.W. Isoflavonoid and lignan phytoestrogens as dietary biomarkers. The Journal of nutrition 2003, 133, 956S–964S. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, T.G.; Nunes, M.A.; Bessada, S.M.; Costa, H.S.; Oliveira, M.B.P. Biologically active and health promoting food components of nuts, oilseeds, fruits, vegetables, cereals, and legumes. In Chemical analysis of food; Elsevier: 2020, pp. 609–656.
- Ding, E.L.; Song, Y.; Malik, V.S.; Liu, S. Sex differences of endogenous sex hormones and risk of type 2 diabetes: a systematic review and meta-analysis. Jama 2006, 295, 1288–1299. [Google Scholar] [CrossRef]
- Nagata, C.; Inaba, S.; Kawakami, N.; Kakizoe, T.; Shimizu, H. Inverse association of soy product intake with serum androgen and estrogen concentrations in Japanese men. Nutrition and cancer 2000, 36, 14–18. [Google Scholar] [CrossRef]
- Prasad, K. Suppression of phosphoenolpyruvate carboxykinase gene expression by secoisolariciresinol diglucoside (SDG), a new antidiabetic agent. International Journal of Angiology 2002, 11, 107–109. [Google Scholar] [CrossRef]
- Talaei, M.; Pan, A. Role of phytoestrogens in prevention and management of type 2 diabetes. World journal of diabetes 2015, 6, 271. [Google Scholar] [CrossRef] [PubMed]
- Kozłowski, R.; Mackiewicz-Talarczyk, M. Handbook of natural fibres; Elsevier: 2012.
- Prasad, K.; Bhanumathy, K.K. Secoisolariciresinol Diglucoside (SDG) from flaxseed in the prevention and treatment of diabetes mellitus. Scripta Medica 2023, 54, 87–93. [Google Scholar] [CrossRef]
- Gong, Z.; Yang, G.; Huang, L.; Chen, L.; Luo, X.; Shuai, L. Phenol-assisted depolymerisation of condensed lignins to mono-/poly-phenols and bisphenols. Chemical Engineering Journal 2023, 455, 140628. [Google Scholar] [CrossRef]
- Shu, F.; Jiang, B.; Yuan, Y.; Li, M.; Wu, W.; Jin, Y.; Xiao, H. Biological activities and emerging roles of lignin and lignin-based products─ A review. Biomacromolecules 2021, 22, 4905–4918. [Google Scholar] [CrossRef]
- Mitjans, M.; Vinardell, M. Biological activity and health benefits of lignans and lignins. 2005.
- Aniszewski, T. Alkaloids: chemistry, biology, ecology, and applications; Elsevier: 2015.
- Aniszewski, T. Alkaloids-Secrets of Life:: Aklaloid Chemistry, Biological Significance, Applications and Ecological Role; Elsevier: 2007.
- Debnath, B.; Singh, W.S.; Das, M.; Goswami, S.; Singh, M.K.; Maiti, D.; Manna, K. Role of plant alkaloids on human health: A review of biological activities. Materials today chemistry 2018, 9, 56–72. [Google Scholar] [CrossRef]
- Christodoulou, M.-I.; Tchoumtchoua, J.; Skaltsounis, A.-L.; Scorilas, A.; Halabalaki, M. Natural alkaloids intervening the insulin pathway: new hopes for anti-diabetic agents? Current Medicinal Chemistry 2019, 26, 5982–6015. [Google Scholar] [CrossRef]
- Sarr, F.B.; Sy, G.Y.; Cabral, M.; Dioum, M.D.; Sene, M.; Ndong, A.; Sall, C.; Faye, O. Antioxidant and Anti α-amylase Activities of Polar Extracts of Mitracarpus hirtus and Saba senegalensis and the Combinason of their Butanolic Extracts. International Research Journal of Pure and Applied Chemistry 2021, 22, 1–8. [Google Scholar]
- Kumar, A.; Aswal, S.; Semwal, R.B.; Chauhan, A.; Joshi, S.K.; Semwal, D.K. Role of plant-derived alkaloids against diabetes and diabetes-related complications: a mechanism-based approach. Phytochemistry Reviews 2019, 18, 1277–1298. [Google Scholar] [CrossRef]
- Rasouli, H.; Yarani, R.; Pociot, F.; Popović-Djordjević, J. Anti-diabetic potential of plant alkaloids: Revisiting current findings and future perspectives. Pharmacological research 2020, 155, 104723. [Google Scholar] [CrossRef] [PubMed]
- Bedekar, A.; Shah, K.; Koffas, M. Natural products for type II diabetes treatment. Advances in applied microbiology 2010, 71, 21–73. [Google Scholar]
- Mooney, M.; Fogarty, S.; Stevenson, C.; Gallagher, A.; Palit, P.; Hawley, S.; Hardie, D.; Coxon, G.; Waigh, R.; Tate, R. Mechanisms underlying the metabolic actions of galegine that contribute to weight loss in mice. British journal of pharmacology 2008, 153, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.-C.; Mustafa, M.R.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef]
- Uvarani, C.; Jaivel, N.; Sankaran, M.; Chandraprakash, K.; Ata, A.; Mohan, P.S. Axially chiral biscarbazoles and biological evaluation of the constituents from Murraya koenigii. Fitoterapia 2014, 94, 10–20. [Google Scholar] [CrossRef]
- Jan, N.U.; Ali, A.; Ahmad, B.; Iqbal, N.; Adhikari, A.; Ali, A.; Ali, S.; Jahan, A.; Ali, H.; Ali, I. Evaluation of antidiabetic potential of steroidal alkaloid of Sarcococca saligna. Biomedicine & Pharmacotherapy 2018, 100, 461–466. [Google Scholar]
- Rajput, A.; Sharma, R.; Bharti, R. Pharmacological activities and toxicities of alkaloids on human health. Materials Today: Proceedings 2022, 48, 1407–1415. [Google Scholar] [CrossRef]
- Shi, R.; Zhou, H.; Ma, B.; Ma, Y.; Wu, D.; Wang, X.; Luo, H.; Cheng, N. Pharmacokinetics and metabolism of jatrorrhizine, a gastric prokinetic drug candidate. Biopharmaceutics & Drug Disposition 2012, 33, 135–145. [Google Scholar]
- Mohammed, K.A.; Al-Maliki, A.D.M. Moderate effect of phenolic and alkaloid compounds extracted from Brassica oleracea var. capitata leaf on blood glucose level in alloxan-induced diabetic rabbits. World Journal of Experimental Biosciences (ISSN: 2313-3937) 2014, 30–35. [Google Scholar]
- Nadri, M.; Dehpour, A.A.; Yaghubi, S.; Fathi, H.; Ataee, R. Effects of the Anti-diabetic and Anti-neuropathy Effects of Onosma Dichroanthum in an Experimental Model of Diabetes by Streptozocin in Mice. Iranian Journal of Endocrinology and Metabolism 2017, 19, 161–169. [Google Scholar]
- Carpinelli de Jesus, M.; Hungerford, N.L.; Carter, S.J.; Anuj, S.R.; Blanchfield, J.T.; De Voss, J.J.; Fletcher, M.T. Pyrrolizidine alkaloids of Blue Heliotrope (Heliotropium amplexicaule) and their presence in Australian honey. Journal of agricultural and food chemistry 2019, 67, 7995–8006. [Google Scholar] [CrossRef]
- López, P.M.G.; de la Mora, P.G.; Wysocka, W.; Maiztegui, B.; Alzugaray, M.E.; Del Zotto, H.; Borelli, M.I. Quinolizidine alkaloids isolated from Lupinus species enhance insulin secretion. European Journal of Pharmacology 2004, 504, 139–142. [Google Scholar] [CrossRef]
- Ochalefu, D.O.; Adoga, G.I.; Luka, C.D.; Abu, A.H. Effects of Catechol containing fraction and other fractions of Nauclea latifolia aqueous root-bark extract on blood glucose, lipid profile and serum liver enzymes in streptozotocin–induced diabetic Wistar albino rats. Journal of Stress Physiology & Biochemistry 2024, 20, 79–91. [Google Scholar]
- Abd El-Wahab, A.E.; Ghareeb, D.A.; Sarhan, E.E.; Abu-Serie, M.M.; El Demellawy, M.A. In vitro biological assessment of Berberis vulgaris and its active constituent, berberine: antioxidants, anti-acetylcholinesterase, anti-diabetic and anticancer effects. BMC complementary and alternative medicine 2013, 13, 1–12. [Google Scholar] [CrossRef]
- Peng, Z.-T.; Huo, H.-X.; Chao, L.-H.; Song, Y.-L.; Liu, D.-F.; Wang, Z.-W.; Zhang, Y.; Zhao, Y.-F.; Tu, P.-F.; Zheng, J. Isoquinoline alkaloids from Corydalis edulis Maxim. Exhibiting insulinotropic action. Phytochemistry 2023, 209, 113637. [Google Scholar] [CrossRef] [PubMed]
- Acharya, D. Antidiabetic activity of aqueous extracts of barks of Alangium salviifolium wang in normal and alloxan-induced diabetic rats. Education 2008, 2010. [Google Scholar]
- Manzo, J.A.M.; Vitor II, R.J.S. Antihyperglycemic effects of Cajanus cajan L.(pigeon pea) ethanolic extract on the blood glucose levels of ICR mice (Mus musculus l.). National Journal of Physiology, Pharmacy and Pharmacology 2017, 7, 860. [Google Scholar] [CrossRef]
- Saleh, S.S.; Sarhat, E.R. Effects of ethanolic Moringa oleifera extract on melatonin, liver and kidney function tests in alloxan-induced diabetic rats. Indian Journal of Forensic Medicine & Toxicology 2019, 13, 1015–1019. [Google Scholar]
- Komeili, G.; Hashemi, M.; Bameri-Niafar, M. Evaluation of antidiabetic and antihyperlipidemic effects of Peganum harmala seeds in diabetic rats. Cholesterol 2016, 2016, 7389864. [Google Scholar] [CrossRef] [PubMed]
- Devaraji, V.; Sivaraman, J.; Prabhu, S. Large-scale computational screening of Indian medicinal plants reveals Cassia angustifolia to be a potentially anti-diabetic. Journal of Biomolecular Structure and Dynamics 2024, 42, 194–210. [Google Scholar] [CrossRef]
- Kumar, V.; Gupta, K.; Kumar, P.; Guleria, P. Catharensus Roseus Based Alkaloids: Applications and Production. Think India Journal 2019, 22, 3006–3030. [Google Scholar]
- Kumar, P.; Nachiar, S.; Thiraviam, P.P. A Review of Pharmacological and Phytochemical Studies on Convolvulaceae Species Rivea and Ipomea. Current Traditional Medicine 2022, 8, 159–165. [Google Scholar] [CrossRef]
- Azmi, A.; Noori, M.; Ghomi, M.K.; Montazer, M.N.; Iraji, A.; Dastyafteh, N.; Oliyaei, N.; Khoramjouy, M.; Rezaei, Z.; Javanshir, S. Alpha-glucosidase inhibitory and hypoglycemic effects of imidazole-bearing thioquinoline derivatives with different substituents: In silico, in vitro, and in vivo evaluations. Bioorganic Chemistry 2024, 144, 107106. [Google Scholar] [CrossRef] [PubMed]
- Tiss, M.; Souiy, Z.; Achour, L.; Hamden, K. Ephedra alata extracts exerts anti-obesity, anti-hyperglycemia, anti-antipyretic and analgesic effects. Nutrition & Food Science 2022, 52, 119–128. [Google Scholar]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. International Journal of Molecular Sciences 2021, 22, 5710. [Google Scholar] [CrossRef] [PubMed]
- Valdes, M.; Calzada, F.; Mendieta-Wejebe, J. Structure–activity relationship study of acyclic terpenes in blood glucose levels: potential α-glucosidase and sodium glucose cotransporter (SGLT-1) inhibitors. Molecules 2019, 24, 4020. [Google Scholar] [CrossRef]
- Panigrahy, S.K.; Bhatt, R.; Kumar, A. Targeting type II diabetes with plant terpenes: the new and promising antidiabetic therapeutics. Biologia 2021, 76, 241–254. [Google Scholar] [CrossRef]
- Habtemariam, S. Antidiabetic potential of monoterpenes: A case of small molecules punching above their weight. International Journal of Molecular Sciences 2017, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Ilic, A. Biological activities of selected mono-amd sesquiterpenes: possible uses in medicine; na: 2013.
- Nazaruk, J.; Borzym-Kluczyk, M. The role of triterpenes in the management of diabetes mellitus and its complications. Phytochemistry Reviews 2015, 14, 675–690. [Google Scholar] [CrossRef]
- Arya, R.; Saldanha, S.N. Dietary phytochemicals, epigenetics, and colon cancer chemoprevention. In Epigenetics of Cancer Prevention; Elsevier: 2019, pp. 205–229.
- Keppler, J.K.; Schwarz, K.; van der Goot, A.J. Covalent modification of food proteins by plant-based ingredients (polyphenols and organosulphur compounds): A commonplace reaction with novel utilization potential. Trends in Food Science & Technology 2020, 101, 38–49. [Google Scholar]
- Omar, S.H.; Al-Wabel, N. Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharmaceutical Journal 2010, 18, 51–58. [Google Scholar] [CrossRef]
- Cavallito, C.J.; Bailey, J.H.; Buck, J.S. The antibacterial principle of Allium sativum. III. Its precursor and “essential oil of garlic”. Journal of the American Chemical Society 1945, 67, 1032–1033. [Google Scholar] [CrossRef]
- Block, E. The organosulfur and organoselenium components of garlic and onions. Phytochemicals-a new paradigm. Technomic Publishing, Lancaster. p 1998, 129-141.
- Sujithra, K.; Srinivasan, S.; Indumathi, D.; Vinothkumar, V. Allyl methyl sulfide, an organosulfur compound alleviates hyperglycemia mediated hepatic oxidative stress and inflammation in streptozotocin-induced experimental rats. Biomedicine & Pharmacotherapy 2018, 107, 292–302. [Google Scholar]
- Saravanan, G.; Ponmurugan, P.; Senthilkumar, G.P.; Rajarajan, T. Modulatory effect of S-allylcysteine on glucose metabolism in streptozotocin induced diabetic rats. Journal of Functional Foods 2009, 1, 336–340. [Google Scholar] [CrossRef]
- Ahmad, P.; Alvi, S.S.; Iqbal, J.; Khan, M.S. Identification and evaluation of natural organosulfur compounds as potential dual inhibitors of α-amylase and α-glucosidase activity: an in-silico and in-vitro approach. Medicinal Chemistry Research 2021, 30, 2184–2202. [Google Scholar] [CrossRef]
- Geu-Flores, F.; Nielsen, M.T.; Nafisi, M.; Møldrup, M.E.; Olsen, C.E.; Motawia, M.S.; Halkier, B.A. Glucosinolate engineering identifies a γ-glutamyl peptidase. Nature chemical biology 2009, 5, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Abuyusuf, M.; Rubel, M.H.; Kim, H.-T.; Jung, H.-J.; Nou, I.-S.; Park, J.-I. Glucosinolates and biotic stress tolerance in Brassicaceae with emphasis on cabbage: A review. Biochemical Genetics 2023, 61, 451–470. [Google Scholar] [CrossRef] [PubMed]
- GOODRICH, R.M.; ANDERSON, J.L.; STOEWSAND, G.S. Glucosinolate changes in blanched broccoli and Brussels sprouts. Journal of Food Processing and Preservation 1989, 13, 275–280. [Google Scholar] [CrossRef]
- Campas-Baypoli, O.; Sánchez-Machado, D.; Bueno-Solano, C.; Ramírez-Wong, B.; López-Cervantes, J. HPLC method validation for measurement of sulforaphane level in broccoli by-products. Biomedical Chromatography 2010, 24, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yuan, Q. Natural sulforaphane as a functional chemopreventive agent: including a review of isolation, purification and analysis methods. Critical reviews in biotechnology 2012, 32, 218–234. [Google Scholar] [CrossRef]
- Saeidi, A.; Soltani, M.; Daraei, A.; Nohbaradar, H.; Haghighi, M.M.; Khosravi, N.; Johnson, K.E.; Laher, I.; Hackney, A.C.; VanDusseldorp, T.A. The effects of aerobic-resistance training and broccoli supplementation on plasma dectin-1 and insulin resistance in males with type 2 diabetes. Nutrients 2021, 13, 3144. [Google Scholar] [CrossRef]
- Tian, S.; Li, X.; Wang, Y.; Lu, Y. The protective effect of sulforaphane on type II diabetes induced by high-fat diet and low-dosage streptozotocin. Food Science & Nutrition 2021, 9, 747–756. [Google Scholar]
- Christensen, L.P. Bioactivity of polyacetylenes in food plants. In Bioactive foods in promoting health; Elsevier: 2010, pp. 285–306.
- Christensen, L.P.; El-Houri, R.B. Development of an in vitro screening platform for the identification of partial PPARγ agonists as a source for antidiabetic lead compounds. Molecules: A Journal of Synthetic Chemistry and Natural Product Chemistry 2018, 23. [Google Scholar]
- Zhou, Z.; Li, M.; Zhang, Z.; Song, Z.; Xu, J.; Zhang, M.; Gong, M. Overview of Panax ginseng and its active ingredients protective mechanism on cardiovascular diseases. Journal of Ethnopharmacology 2024, 118506. [Google Scholar] [CrossRef]
- El-Houri, R.B.; Kotowska, D.; Christensen, K.B.; Bhattacharya, S.; Oksbjerg, N.; Wolber, G.; Kristiansen, K.; Christensen, L.P. Polyacetylenes from carrots (Daucus carota) improve glucose uptake in vitro in adipocytes and myotubes. Food & function 2015, 6, 2135–2144. [Google Scholar]
- Resetar, M.; Liu, X.; Herdlinger, S.; Kunert, O.; Pferschy-Wenzig, E.-M.; Latkolik, S.; Steinacher, T.; Schuster, D.; Bauer, R.; Dirsch, V.M. Polyacetylenes from Oplopanax horridus and Panax ginseng: Relationship between structure and PPARγ activation. Journal of natural products 2020, 83, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Darenskaya, M.; Kolesnikova, L.a.; Kolesnikov, S. Oxidative stress: pathogenetic role in diabetes mellitus and its complications and therapeutic approaches to correction. Bulletin of experimental biology and medicine 2021, 171, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Ježek, P.; Hlavatá, L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. The international journal of biochemistry & cell biology 2005, 37, 2478–2503. [Google Scholar]
- Thomas, C.; Mackey, M.M.; Diaz, A.A.; Cox, D.P. Hydroxyl radical is produced via the Fenton reaction in submitochondrial particles under oxidative stress: implications for diseases associated with iron accumulation. Redox Report 2009, 14, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Kaliszewska, A.; Allison, J.; Martini, M.; Arias, N. Improving Age-Related Cognitive Decline through Dietary Interventions Targeting Mitochondrial Dysfunction. International Journal of Molecular Sciences 2021, 22, 3574–3574. [Google Scholar] [CrossRef]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of reactive oxygen species by mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Okado-Matsumoto, A.; Fridovich, I. Subcellular distribution of superoxide dismutases (SOD) in rat liver: Cu, Zn-SOD in mitochondria. Journal of Biological Chemistry 2001, 276, 38388–38393. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Vivar, J.; Kalyanaraman, B.; Kennedy, M.C. Mitochondrial aconitase is a source of hydroxyl radical: an electron spin resonance investigation. Journal of Biological Chemistry 2000, 275, 14064–14069. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Chakraborty, R. The role of antioxidants in human health. In Oxidative stress: diagnostics, prevention, and therapy; ACS Publications: 2011, pp. 1–37.
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS generation and antioxidant defense systems in normal and malignant cells. Oxidative medicine and cellular longevity 2019, 2019, 6175804. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Antioxidants in human health and disease. Annual review of nutrition 1996, 16, 33–50. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH oxidases: an overview from structure to innate immunity-associated pathologies. Cellular & molecular immunology 2015, 12, 5–23. [Google Scholar]
- Harrison, R. Structure and function of xanthine oxidoreductase: where are we now? Free Radical Biology and Medicine 2002, 33, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Kaludercic, N.; Mialet-Perez, J.; Paolocci, N.; Parini, A.; Di Lisa, F. Monoamine oxidases as sources of oxidants in the heart. Journal of molecular and cellular cardiology 2014, 73, 34–42. [Google Scholar] [CrossRef]
- Del Río, L.A. Peroxisomes as a cellular source of reactive nitrogen species signal molecules. Archives of Biochemistry and Biophysics 2011, 506, 1–11. [Google Scholar] [CrossRef]
- Fürstenberger, G.; Krieg, P.; Müller-Decker, K.; Habenicht, A. What are cyclooxygenases and lipoxygenases doing in the driver's seat of carcinogenesis? International journal of cancer 2006, 119, 2247–2254. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. BioMed research international 2012, 2012, 936486. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S. A synopsis of the associations of oxidative stress, ROS, and antioxidants with diabetes mellitus. Antioxidants 2022, 11, 2003. [Google Scholar] [CrossRef]
- Zhang, P.; Li, T.; Wu, X.; Nice, E.C.; Huang, C.; Zhang, Y. Oxidative stress and diabetes: antioxidative strategies. Frontiers of medicine 2020, 14, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Dicarbonyl stress in cell and tissue dysfunction contributing to ageing and disease. Biochemical and biophysical research communications 2015, 458, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Böhme, D.; Singer, D.; Frolov, A. Specific tandem mass spectrometric detection of AGE-modified arginine residues in peptides. Journal of Mass Spectrometry 2015, 50, 613–624. [Google Scholar] [CrossRef]
- Schalkwijk, C.; Stehouwer, C. Methylglyoxal, a highly reactive dicarbonyl compound, in diabetes, its vascular complications, and other age-related diseases. Physiological reviews 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Thornalley, P.J.; Langborg, A.; Minhas, H.S. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochemical Journal 1999, 344, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Vistoli, G.; De Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): an overview of their mechanisms of formation. Free radical research 2013, 47, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, P.J.; Howell, S.K.; Smith, K.; Szwergold, B.S. Glyceraldehyde-3-phosphate dehydrogenase activity as an independent modifier of methylglyoxal levels in diabetes. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2003, 1637, 98–106. [Google Scholar] [CrossRef]
- Ray Chaudhuri, A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nature reviews Molecular cell biology 2017, 18, 610–621. [Google Scholar] [CrossRef]
- Du, X.; Matsumura, T.; Edelstein, D.; Rossetti, L.; Zsengellér, Z.; Szabó, C.; Brownlee, M. Inhibition of GAPDH activity by poly (ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. The Journal of clinical investigation 2003, 112, 1049–1057. [Google Scholar] [CrossRef]
- Bora, S.; Adole, P.S.; Motupalli, N.; Pandit, V.R.; Vinod, K.V. Association between carbonyl stress markers and the risk of acute coronary syndrome in patients with type 2 diabetes mellitus–A pilot study. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2020, 14, 1751–1755. [Google Scholar]
- Leonova, T.; Popova, V.; Tsarev, A.; Henning, C.; Antonova, K.; Rogovskaya, N.; Vikhnina, M.; Baldensperger, T.; Soboleva, A.; Dinastia, E. Does protein glycation impact on the drought-related changes in metabolism and nutritional properties of mature pea (Pisum sativum L.) seeds? International journal of molecular sciences 2020, 21, 567. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. Protein kinase C and lipid signaling for sustained cellular responses. The FASEB journal 1995, 9, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, E.D.; Weigert, C. Role of the hexosamine biosynthetic pathway in diabetic nephropathy. Kidney international 2000, 58, S13–S18. [Google Scholar] [CrossRef]
- Martemucci, G.; Costagliola, C.; Mariano, M.; D’andrea, L.; Napolitano, P.; D’Alessandro, A.G. Free radical properties, source and targets, antioxidant consumption and health. Oxygen 2022, 2, 48–78. [Google Scholar] [CrossRef]
- Mohanasundaram, T.; Ramachandran, V.; Bhongiri, B.; Rymbai, E.; Xavier, R.M.; Rao, G.N.; Narendar, C. The Promotion of Antioxidant and Anti-Inflammatory Activity by Nrf2 Amplifier is A Potential Technique in Diabetic Wound Healing—A Review. Pharmaceutical Sciences 2022, 29, 255–262. [Google Scholar] [CrossRef]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; León, R.; López, M.G.; Oliva, B. Transcription factor NRF2 as a therapeutic target for chronic diseases: a systems medicine approach. Pharmacological reviews 2018, 70, 348–383. [Google Scholar] [CrossRef]
- Matzinger, M.; Fischhuber, K.; Heiss, E.H. Activation of Nrf2 signaling by natural products-can it alleviate diabetes? Biotechnology advances 2018, 36, 1738–1767. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kang, M.-I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Molecular and cellular biology 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by Nrf2. Antioxidants & redox signaling 2018, 29, 1727–1745. [Google Scholar]
- He, F.; Ru, X.; Wen, T. NRF2, a transcription factor for stress response and beyond. International journal of molecular sciences 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Kaur, I.; Sehgal, A.; Sharma, E.; Kumar, A.; Grover, M.; Bungau, S. Unfolding Nrf2 in diabetes mellitus. Molecular Biology Reports 2021, 48, 927–939. [Google Scholar] [CrossRef]
- Thorwald, M.A.; Godoy-Lugo, J.A.; Rodriguez, R.; Stanhope, K.L.; Graham, J.L.; Havel, P.J.; Forman, H.J.; Ortiz, R.M. Cardiac NF-κB Acetylation Increases While Nrf2-Related Gene Expression and Mitochondrial Activity Are Impaired during the Progression of Diabetes in UCD-T2DM Rats. Antioxidants 2022, 11, 927. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of Nrf2 signaling pathway and its role in inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Pan, H.; Wang, H.; Wang, X.; Zhu, L.; Mao, L. The absence of Nrf2 enhances NF-κB-dependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediators of inflammation 2012, 2012, 217580. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, R.; Sireesh, D.; Salai, M.B.; Ramkumar, K.M.; Sarvajayakesavulu, S.; Madhunapantula, S.V. Treatment with naringenin elevates the activity of transcription factor Nrf2 to protect pancreatic β-cells from streptozotocin-induced diabetes in vitro and in vivo. Frontiers in Pharmacology 2019, 9, 1562. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Eggler, A.L.; Mesecar, A.D.; Van Breemen, R.B. Modification of keap1 cysteine residues by sulforaphane. Chemical research in toxicology 2011, 24, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Chun, K.-S.; Kim, D.-H.; Kim, S.-J.; Kim, S.H.; Cho, N.-C.; Na, H.-K.; Surh, Y.-J. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochemical Pharmacology 2020, 173, 113820. [Google Scholar] [CrossRef]
- Negi, G.; Kumar, A.; S Sharma, S. Nrf2 and NF-κB modulation by sulforaphane counteracts multiple manifestations of diabetic neuropathy in rats and high glucose-induced changes. Current neurovascular research 2011, 8, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Utami, A.R.; Maksum, I.P.; Deawati, Y. Berberine and its study as an antidiabetic compound. Biology 2023, 12, 973. [Google Scholar] [CrossRef]
- Wong, D.P.W.; Ng, M.Y.; Leung, J.Y.; Boh, B.K.; Lim, E.C.; Tan, S.H.; Lim, S.; Seah, W.H.; Hu, C.Z.; Ho, B.C. Regulation of the NRF2 transcription factor by andrographolide and organic extracts from plant endophytes. PLoS One 2018, 13, e0204853. [Google Scholar] [CrossRef]
- Akshatha, V.; Nalini, M.; D'souza, C.; Prakash, H. Streptomycete endophytes from anti-diabetic medicinal plants of the Western Ghats inhibit alpha-amylase and promote glucose uptake. Letters in applied microbiology 2014, 58, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Van Chen, T.; Cuong, T.D.; Quy, P.T.; Bui, T.Q.; Van Tuan, L.; Van Hue, N.; Triet, N.T.; Ho, D.V.; Bao, N.C.; Nhung, N.T.A. Antioxidant activity and α-glucosidase inhibitability of Distichochlamys citrea MF Newman rhizome fractionated extracts: in vitro and in silico screenings. Chemical Papers 2022, 76, 5655–5675. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Ribeiro, D.; Freitas, M.; Fernandes, E. Flavonoids as potential agents in the management of type 2 diabetes through the modulation of α-amylase and α-glucosidase activity: a review. Critical Reviews in Food Science and Nutrition 2022, 62, 3137–3207. [Google Scholar] [CrossRef] [PubMed]
- Bag, A.; Bhattacharyya, S.K.; Chattopadhyay, R.R. The development of Terminalia chebula Retz.(Combretaceae) in clinical research. Asian Pacific journal of tropical biomedicine 2013, 3, 244–252. [Google Scholar] [CrossRef]
- Pereira, A.S.; Banegas-Luna, A.J.; Peña-García, J.; Pérez-Sánchez, H.; Apostolides, Z. Evaluation of the anti-diabetic activity of some common herbs and spices: providing new insights with inverse virtual screening. Molecules 2019, 24, 4030. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. International journal of biological sciences 2018, 14, 1483. [Google Scholar] [CrossRef]
- Wilcox, G. Insulin and insulin resistance. Clinical biochemist reviews 2005, 26, 19. [Google Scholar]
- Muhammad, I.; Rahman, N.; Nishan, U.; Shah, M. Antidiabetic activities of alkaloids isolated from medicinal plants. Brazilian Journal of Pharmaceutical Sciences 2021, 57, e19130. [Google Scholar] [CrossRef]
- Putta, S.; Sastry Yarla, N.; Kumar Kilari, E.; Surekha, C.; Aliev, G.; Basavaraju Divakara, M.; Sridhar Santosh, M.; Ramu, R.; Zameer, F.; Prasad MN, N. Therapeutic potentials of triterpenes in diabetes and its associated complications. Current topics in medicinal chemistry 2016, 16, 2532–2542. [Google Scholar] [CrossRef]
- Weber, L.W.; Boll, M.; Stampfl, A. Maintaining cholesterol homeostasis: sterol regulatory element-binding proteins. World journal of gastroenterology: WJG 2004, 10, 3081. [Google Scholar] [CrossRef]
- Bloise, R.; Reinhart, C. EPAISSISSEMENTS DE PULPES. MODELISATION DU PROCESSUS DISCONTINU. 1981. [Google Scholar]
- DeBose-Boyd, R.A.; Ye, J. SREBPs in lipid metabolism, insulin signaling, and beyond. Trends in biochemical sciences 2018, 43, 358–368. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. Journal of advanced pharmaceutical technology & research 2011, 2, 236–240. [Google Scholar]
- Al-Mansoori, L.; Al-Jaber, H.; Prince, M.S.; Elrayess, M.A. Role of inflammatory cytokines, growth factors and adipokines in adipogenesis and insulin resistance. Inflammation 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Tian, R. Glucose transporters in cardiac metabolism and hypertrophy. Comprehensive Physiology 2015, 6, 331. [Google Scholar]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annual review of medicine 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): a review. Biochemical pharmacology 2014, 92, 73–89. [Google Scholar] [CrossRef]
- Kim, W.; Egan, J.M. The role of incretins in glucose homeostasis and diabetes treatment. Pharmacological reviews 2008, 60, 470–512. [Google Scholar] [CrossRef] [PubMed]
- Godinho, R.; Mega, C.; Teixeira-de-Lemos, E.; Carvalho, E.; Teixeira, F.; Fernandes, R.; Reis, F. The place of dipeptidyl peptidase-4 inhibitors in type 2 diabetes therapeutics: A “me too” or “the special one” antidiabetic class? Journal of diabetes research 2015, 2015, 806979. [Google Scholar] [CrossRef] [PubMed]
- Yabe, D.; Seino, Y. Two incretin hormones GLP-1 and GIP: comparison of their actions in insulin secretion and β cell preservation. Progress in biophysics and molecular biology 2011, 107, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Cernea, S.; Raz, I. Therapy in the early stage: incretins. Diabetes care 2011, 34, S264–S271. [Google Scholar] [CrossRef]
- Hassanein, E.H.; Sayed, A.M.; Hussein, O.E.; Mahmoud, A.M. Coumarins as modulators of the Keap1/Nrf2/ARE signaling pathway. Oxidative Medicine and Cellular Longevity 2020, 2020, 1675957. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Sarker, M.M.R.; Sultana, T.N.; Chowdhury, M.N.R.; Rashid, M.A.; Chaity, N.I.; Zhao, C.; Xiao, J.; Hafez, E.E.; Khan, S.A. Antidiabetic phytochemicals from medicinal plants: prospective candidates for new drug discovery and development. Frontiers in endocrinology 2022, 13, 800714. [Google Scholar] [CrossRef]
- Lee, M.-S.; Chyau, C.-C.; Wang, C.-P.; Wang, T.-H.; Chen, J.-H.; Lin, H.-H. Flavonoids identification and pancreatic beta-cell protective effect of lotus seedpod. Antioxidants 2020, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as potential anti-inflammatory molecules: A review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Zhang, Z.; Yang, Z.; Rao, B. The effects of taurine supplementation on diabetes mellitus in humans: A systematic review and meta-analysis. Food Chemistry: Molecular Sciences 2022, 4, 100106. [Google Scholar] [CrossRef]
- Murthy, K.N.C.; Manchali, S. Anti-diabetic potentials of red beet pigments and other constituents. In Red Beet Biotechnology: Food and Pharmaceutical Applications; Springer: 2012, pp. 155–174.
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic application of betalains: A review. Plants 2020, 9, 1219. [Google Scholar] [CrossRef]
- Saltiel, A.R. Insulin signaling in health and disease. The Journal of clinical investigation 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, J.; Hu, X.; Huang, X.-J.; Chen, G.-X. Current understanding of glucose transporter 4 expression and functional mechanisms. World journal of biological chemistry 2020, 11, 76. [Google Scholar] [CrossRef] [PubMed]
- Abdelsalam, S.S.; Korashy, H.M.; Zeidan, A.; Agouni, A. The role of protein tyrosine phosphatase (PTP)-1B in cardiovascular disease and its interplay with insulin resistance. Biomolecules 2019, 9, 286. [Google Scholar] [CrossRef]
- Pereira, S.; Cline, D.L.; Glavas, M.M.; Covey, S.D.; Kieffer, T.J. Tissue-specific effects of leptin on glucose and lipid metabolism. Endocrine reviews 2021, 42, 1–28. [Google Scholar] [CrossRef]
- Yang, R.; Barouch, L.A. Leptin signaling and obesity: cardiovascular consequences. Circulation research 2007, 101, 545–559. [Google Scholar] [CrossRef]
- Tkachuk, V.A.e.; Vorotnikov, A.V. Molecular mechanisms of insulin resistance development. Diabetes mellitus 2014, 17, 29–40. [Google Scholar] [CrossRef]
- Zhao, B.T.; Nguyen, D.H.; Le, D.D.; Choi, J.S.; Min, B.S.; Woo, M.H. Protein tyrosine phosphatase 1B inhibitors from natural sources. Archives of pharmacal research 2018, 41, 130–161. [Google Scholar] [CrossRef]
- Dingwoke, E.J.; Adamude, F.A.; Chukwuocha, C.E.; Ambi, A.A.; Nwobodo, N.N.; Sallau, A.B.; Nzelibe, H.C. Inhibition of trypanosoma evansi protein-tyrosine phosphatase by myristic acid analogues isolated from khaya senegalensis and tamarindus indica. Journal of Experimental Pharmacology 2019, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Pitschmann, A.; Zehl, M.; Atanasov, A.G.; Dirsch, V.M.; Heiss, E.; Glasl, S. Walnut leaf extract inhibits PTP1B and enhances glucose-uptake in vitro. Journal of Ethnopharmacology 2014, 152, 599–602. [Google Scholar] [CrossRef]
- LC, M. Action of amino acids on sugars. Formation of melanoidins in a methodical way. Compte-Rendu de l'Academie des Science 1912, 154, 66–68. [Google Scholar]
- Heyns, K.; Noack, H. Die Umsetzung von D-Fructose mit L-Lysin und L-Arginin und deren Beziehung zu nichtenzymatischen Bräunungsreaktionen. Chemische Berichte 1962, 95, 720–727. [Google Scholar] [CrossRef]
- Amadori, M. The product of the condensation of glucose and p-phenetidine. Atti Reale Accad Nazl Lincei 1929, 9, 68–73. [Google Scholar]
- Brownlee, M.; VLASSARA, H.; Cerami, A. Nonenzymatic glycosylation and the pathogenesis of diabetic complications. Annals of internal medicine 1984, 101, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: a review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Khetarpaul, N.; Chauhan, B. Improvement in HCl-extractability of minerals from pearl millet by natural fermentation. Food chemistry 1990, 37, 69–75. [Google Scholar] [CrossRef]
- Wolff, S.P.; Dean, R. Glucose autoxidation and protein modification. The potential role of ‘autoxidative glycosylation’in diabetes. Biochemical journal 1987, 245, 243–250. [Google Scholar] [CrossRef]
- Sajithlal, G.; Chandrakasan, G. Role of lipid peroxidation products in the formation of advanced glycation end products: An in vitro study on collagen. In Proceedings of the Proceedings of the Indian Academy of Sciences-Chemical Sciences, 1999; pp. 215–229.
- Hayashi, T.; Namiki, M. Role of sugar fragmentation in an early stage browning of amino-carbonyl reaction of sugar with amino acid. Agricultural and biological chemistry 1986, 50, 1965–1970. [Google Scholar]
- Glenn, J.V.; Stitt, A.W. The role of advanced glycation end products in retinal ageing and disease. Biochimica et Biophysica Acta (BBA)-General Subjects 2009, 1790, 1109–1116. [Google Scholar] [CrossRef]
- Horiuchi, S. Advanced glycation end products (AGE)-modified proteins and their potential relevance to atherosclerosis. Trends in cardiovascular medicine 1996, 6, 163–168. [Google Scholar] [CrossRef]
- Song, Q.; Liu, J.; Dong, L.; Wang, X.; Zhang, X. Novel advances in inhibiting advanced glycation end product formation using natural compounds. Biomedicine & Pharmacotherapy 2021, 140, 111750. [Google Scholar]
- Uribarri, J.; Woodruff, S.; Goodman, S.; Cai, W.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced glycation end products in foods and a practical guide to their reduction in the diet. Journal of the American Dietetic Association 2010, 110, 911–916. [Google Scholar] [CrossRef]
- Cerami, C.; Founds, H.; Nicholl, I.; Mitsuhashi, T.; Giordano, D.; Vanpatten, S.; Lee, A.; Al-Abed, Y.; Vlassara, H.; Bucala, R. Tobacco smoke is a source of toxic reactive glycation products. Proceedings of the National Academy of Sciences 1997, 94, 13915–13920. [Google Scholar] [CrossRef] [PubMed]
- O'Brien, J.; Morrissey, P.; Ames, J. Nutritional and toxicological aspects of the Maillard browning reaction in foods. Critical Reviews in Food Science & Nutrition 1989, 28, 211–248. [Google Scholar]
- https://lemchem.file3.wcms.tu-dresden.de//.
- Delgado-Andrade, C. Carboxymethyl-lysine: thirty years of investigation in the field of AGE formation. Food & function 2016, 7, 46–57. [Google Scholar]
- Twarda-Clapa, A.; Olczak, A.; Białkowska, A.M.; Koziołkiewicz, M. Advanced glycation end-products (AGEs): Formation, chemistry, classification, receptors, and diseases related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- Semba, R.D.; Nicklett, E.J.; Ferrucci, L. Does accumulation of advanced glycation end products contribute to the aging phenotype? Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences 2010, 65, 963–975. [Google Scholar] [CrossRef]
- Chhipa, A.S.; Borse, S.P.; Baksi, R.; Lalotra, S.; Nivsarkar, M. Targeting receptors of advanced glycation end products (RAGE): Preventing diabetes induced cancer and diabetic complications. Pathology-Research and Practice 2019, 215, 152643. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P.; Aryal, P.; Darkwah, E.K. Advanced glycation end products in health and disease. Microorganisms 2022, 10, 1848. [Google Scholar] [CrossRef]
- Neeper, M.; Schmidt, A.M.; Brett, J.; Yan, S.; Wang, F.; Pan, Y.; Elliston, K.; Stern, D.; Shaw, A. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. Journal of biological chemistry 1992, 267, 14998–15004. [Google Scholar] [CrossRef]
- Chavakis, T.; Bierhaus, A.; Al-Fakhri, N.; Schneider, D.; Witte, S.; Linn, T.; Nagashima, M.; Morser, J.; Arnold, B.; Preissner, K.T. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: a novel pathway for inflammatory cell recruitment. The Journal of experimental medicine 2003, 198, 1507–1515. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Santos, A.N.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox biology 2014, 2, 411–429. [Google Scholar] [CrossRef]
- Bongarzone, S.; Savickas, V.; Luzi, F.; Gee, A.D. Targeting the receptor for advanced glycation endproducts (RAGE): a medicinal chemistry perspective. Journal of medicinal chemistry 2017, 60, 7213–7232. [Google Scholar] [CrossRef] [PubMed]
- Jules, J.; Maiguel, D.; Hudson, B.I. Alternative splicing of the RAGE cytoplasmic domain regulates cell signaling and function. PloS one 2013, 8, e78267. [Google Scholar] [CrossRef]
- Zhang, L.; Bukulin, M.; Kojro, E.; Roth, A.; Metz, V.V.; Fahrenholz, F.; Nawroth, P.P.; Bierhaus, A.; Postina, R. Receptor for advanced glycation end products is subjected to protein ectodomain shedding by metalloproteinases. Journal of Biological Chemistry 2008, 283, 35507–35516. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. The Korean journal of physiology & pharmacology: official journal of the Korean Physiological Society and the Korean Society of Pharmacology 2014, 18, 1. [Google Scholar]
- Cai, W.; Ramdas, M.; Zhu, L.; Chen, X.; Striker, G.E.; Vlassara, H. Oral advanced glycation endproducts (AGEs) promote insulin resistance and diabetes by depleting the antioxidant defenses AGE receptor-1 and sirtuin 1. Proceedings of the National Academy of Sciences 2012, 109, 15888–15893. [Google Scholar] [CrossRef]
- Anwar, S.; Khan, S.; Almatroudi, A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. A review on mechanism of inhibition of advanced glycation end products formation by plant derived polyphenolic compounds. Molecular Biology Reports 2021, 48, 787–805. [Google Scholar] [CrossRef]
- Wautier, M.-P.; Chappey, O.; Corda, S.; Stern, D.M.; Schmidt, A.M.; Wautier, J.-L. Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. American journal of physiology-endocrinology and metabolism 2001, 280, E685–E694. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Méndez, J.D.; Méndez-Valenzuela, V.; Aguilar-Hernández, M.M. Cellular signalling of the receptor for advanced glycation end products (RAGE). Cellular signalling 2013, 25, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Daffu, G.; Del Pozo, C.H.; O’Shea, K.M.; Ananthakrishnan, R.; Ramasamy, R.; Schmidt, A.M. Radical roles for RAGE in the pathogenesis of oxidative stress in cardiovascular diseases and beyond. International journal of molecular sciences 2013, 14, 19891–19910. [Google Scholar] [CrossRef] [PubMed]
- Younus, H.; Anwar, S. Prevention of non-enzymatic glycosylation (glycation): Implication in the treatment of diabetic complication. International journal of health sciences 2016, 10, 261. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Kim, Y.S. The role of advanced glycation end products in diabetic vascular complications. Diabetes & metabolism journal 2018, 42, 188–195. [Google Scholar]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. History of modern biotechnology I 2000, 1–39. [Google Scholar]
- Xiao, J.; Hogger, P. Dietary polyphenols and type 2 diabetes: current insights and future perspectives. Current medicinal chemistry 2015, 22, 23–38. [Google Scholar] [CrossRef]
- Ramkissoon, J.; Mahomoodally, M.; Ahmed, N.; Subratty, A. Antioxidant and anti–glycation activities correlates with phenolic composition of tropical medicinal herbs. Asian Pacific journal of tropical medicine 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Yin, M.-C. Anti-glycative potential of triterpenes: a mini-review. BioMedicine 2012, 2, 2–9. [Google Scholar] [CrossRef]
- Alqahtani, A.; Hamid, K.; Kam, A.; Wong, K.; Abdelhak, Z.; Razmovski-Naumovski, V.; Chan, K.; Li, K.M.; Groundwater, P.W.; Li, G.Q. The pentacyclic triterpenoids in herbal medicines and their pharmacological activities in diabetes and diabetic complications. Current Medicinal Chemistry 2013, 20, 908–931. [Google Scholar] [PubMed]
- Silva, F.S.; Oliveira, P.J.; Duarte, M.F. Oleanolic, ursolic, and betulinic acids as food supplements or pharmaceutical agents for type 2 diabetes: promise or illusion? Journal of agricultural and food chemistry 2016, 64, 2991–3008. [Google Scholar] [CrossRef]
- Jung, E.; Park, S.-B.; Jung, W.K.; Kim, H.R.; Kim, J. Antiglycation activity of aucubin in vitro and in exogenous methylglyoxal injected rats. Molecules 2019, 24, 3653. [Google Scholar] [CrossRef]
- Wu, C.-H.; Yen, G.-C. Inhibitory effect of naturally occurring flavonoids on the formation of advanced glycation endproducts. Journal of agricultural and food chemistry 2005, 53, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wen, W.; Qi, C.-L.; Zhao, R.-X.; Lü, J.-H.; Zhong, C.-Y.; Chen, Y.-Y. Ameliorative effect of berberine on renal damage in rats with diabetes induced by high-fat diet and streptozotocin. Phytomedicine 2012, 19, 712–718. [Google Scholar] [CrossRef]
- Lan, J.; Zhao, Y.; Dong, F.; Yan, Z.; Zheng, W.; Fan, J.; Sun, G. Meta-analysis of the effect and safety of berberine in the treatment of type 2 diabetes mellitus, hyperlipemia and hypertension. Journal of ethnopharmacology 2015, 161, 69–81. [Google Scholar] [CrossRef]
- Sang, S.; Shao, X.; Bai, N.; Lo, C.-Y.; Yang, C.S.; Ho, C.-T. Tea polyphenol (−)-epigallocatechin-3-gallate: a new trapping agent of reactive dicarbonyl species. Chemical research in toxicology 2007, 20, 1862–1870. [Google Scholar] [CrossRef]
- Shao, X.; Bai, N.; He, K.; Ho, C.-T.; Yang, C.S.; Sang, S. Apple polyphenols, phloretin and phloridzin: new trapping agents of reactive dicarbonyl species. Chemical research in toxicology 2008, 21, 2042–2050. [Google Scholar] [CrossRef]
- Zhou, Q.; Gong, J.; Wang, M. Phloretin and its methylglyoxal adduct: Implications against advanced glycation end products-induced inflammation in endothelial cells. Food and Chemical Toxicology 2019, 129, 291–300. [Google Scholar] [CrossRef]
- Bhuiyan, M.N.I.; Mitsuhashi, S.; Sigetomi, K.; Ubukata, M. Quercetin inhibits advanced glycation end product formation via chelating metal ions, trapping methylglyoxal, and trapping reactive oxygen species. Bioscience, biotechnology, and biochemistry 2017, 81, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhao, Y.; Wang, P.; Ahmedna, M.; Sang, S. Bioactive ginger constituents alleviate protein glycation by trapping methylglyoxal. Chemical research in toxicology 2015, 28, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Tang, S.; Zhang, L.; Xiao, J.; Luo, Q.; Chen, Y.; Zhou, M.; Feng, N.; Wang, C. The inhibitory effect of the catechin structure on advanced glycation end product formation in alcoholic media. Food & function 2020, 11, 5396–5408. [Google Scholar]
- Muthenna, P.; Akileshwari, C.; Reddy, G.B. Ellagic acid, a new antiglycating agent: its inhibition of N ϵ-(carboxymethyl) lysine. Biochemical Journal 2012, 442, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jeong, I.-H.; Kim, C.-S.; Lee, Y.M.; Kim, J.M.; Kim, J.S. Chlorogenic acid inhibits the formation of advanced glycation end products and associated protein cross-linking. Archives of Pharmacal Research 2011, 34, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Umadevi, S.; Gopi, V.; Elangovan, V. Regulatory mechanism of gallic acid against advanced glycation end products induced cardiac remodeling in experimental rats. Chemico-biological interactions 2014, 208, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.-T.; Sang, S. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chemical research in toxicology 2011, 24, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, H.; Sang, S. Trapping methylglyoxal by genistein and its metabolites in mice. Chemical Research in Toxicology 2016, 29, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yagiz, Y.; Buran, T.J.; do Nascimento Nunes, C.; Gu, L. Phytochemicals from berries and grapes inhibited the formation of advanced glycation end-products by scavenging reactive carbonyls. Food research international 2011, 44, 2666–2673. [Google Scholar] [CrossRef]
- Kato, N.; Kawabe, S.; Ganeko, N.; Yoshimura, M.; Amakura, Y.; Ito, H. Polyphenols from flowers of Magnolia coco and their anti-glycation effects. Bioscience, Biotechnology, and Biochemistry 2017, 81, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, Z.; Sheng, Z. Ability of resveratrol to inhibit advanced glycation end product formation and carbohydrate-hydrolyzing enzyme activity, and to conjugate methylglyoxal. Food chemistry 2017, 216, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Maleki, V.; Foroumandi, E.; Hajizadeh-Sharafabad, F.; Kheirouri, S.; Alizadeh, M. The effect of resveratrol on advanced glycation end products in diabetes mellitus: a systematic review. Archives of Physiology and Biochemistry 2022, 128, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Shao, X.; Wang, L.; Huang, D.; Ho, C.-T.; Sang, S. Stilbene glucoside from Polygonum multiflorum Thunb.: a novel natural inhibitor of advanced glycation end product formation by trapping of methylglyoxal. Journal of agricultural and food chemistry 2010, 58, 2239–2245. [Google Scholar] [CrossRef]
- Ding, H.; Ni, M.; Zhang, G.; Liao, Y.; Hu, X.; Zhang, Y.; Gong, D. The inhibition of oleanolic acid on protein non-enzymatic glycation. LWT 2020, 125, 109253. [Google Scholar] [CrossRef]
- Rizzi, G.P. Free radicals in the Maillard reaction. Food Reviews International 2003, 19, 375–395. [Google Scholar] [CrossRef]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bügel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food and Chemical Toxicology 2013, 60, 10–37. [Google Scholar] [CrossRef]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Research International 2020, 129, 108843. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: structure-activity relationships. Free radical biology and medicine 1997, 22, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Mukund, V.; Mukund, D.; Sharma, V.; Mannarapu, M.; Alam, A. Genistein: Its role in metabolic diseases and cancer. Critical Reviews in Oncology/Hematology 2017, 119, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Kiso, Y. Antioxidative roles of sesamin, a functional lignan in sesame seed, and it's effect on lipid-and alcohol-metabolism in the liver: A DNA microarray study. Biofactors 2004, 21, 191–196. [Google Scholar] [CrossRef]
- Kong, X.; Wang, G.-D.; Ma, M.-Z.; Deng, R.-Y.; Guo, L.-Q.; Zhang, J.-X.; Yang, J.-R.; Su, Q. Sesamin ameliorates advanced glycation end products-induced pancreatic β-cell dysfunction and apoptosis. Nutrients 2015, 7, 4689–4704. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Wang, R.; Su, D.; Tang, M.; Liu, Y.; Li, Z. Antioxidant activity and healthy benefits of natural pigments in fruits: A review. International journal of molecular sciences 2021, 22, 4945. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Progress in lipid research 2018, 70, 62–93. [Google Scholar] [CrossRef]
- Hayase, F.; Shibuya, T.; Sato, J.; Yamamoto, M. Effects of oxygen and transition metals on the advanced Maillard reaction of proteins with glucose. Bioscience, biotechnology, and biochemistry 1996, 60, 1820–1825. [Google Scholar] [CrossRef]
- Frolov, A.; Schmidt, R.; Spiller, S.; Greifenhagen, U.; Hoffmann, R. Arginine-derived advanced glycation end products generated in peptide–glucose mixtures during boiling. Journal of agricultural and food chemistry 2014, 62, 3626–3635. [Google Scholar] [CrossRef]
- Andjelković, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food chemistry 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Basu, U.; Upadhyaya, H.D.; Srivastava, R.; Daware, A.; Malik, N.; Sharma, A.; Bajaj, D.; Narnoliya, L.; Thakro, V.; Kujur, A. ABC transporter-mediated transport of glutathione conjugates enhances seed yield and quality in chickpea. Plant Physiology 2019, 180, 253–275. [Google Scholar] [CrossRef]
- Genaro-Mattos, T.C.; Maurício, Â.Q.; Rettori, D.; Alonso, A.; Hermes-Lima, M. Antioxidant activity of caffeic acid against iron-induced free radical generation—A chemical approach. PLoS one 2015, 10, e0129963. [Google Scholar]
- Kostyuk, V.; Potapovich, A.; Kostyuk, T.; Cherian, M. Metal complexes of dietary flavonoids: evaluation of radical scavenger properties and protective activity against oxidative stress in vivo. Cellular and Molecular Biology 2007, 53, 62–69. [Google Scholar]
- Gugliucci, A.; Bastos, D.H.M.; Schulze, J.; Souza, M.F.F. Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of AGE generation by methylglyoxal in model proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jo, K.; Lee, I.-S.; Kim, C.-S.; Kim, J.S. The extract of aster koraiensis prevents retinal pericyte apoptosis in diabetic rats and its active compound, chlorogenic acid inhibits AGE formation and AGE/RAGE interaction. Nutrients 2016, 8, 585. [Google Scholar] [CrossRef] [PubMed]
- Meeprom, A.; Sompong, W.; Chan, C.B.; Adisakwattana, S. Isoferulic acid, a new anti-glycation agent, inhibits fructose-and glucose-mediated protein glycation in vitro. Molecules 2013, 18, 6439–6454. [Google Scholar] [CrossRef]
- Kanlaya, R.; Thongboonkerd, V. Molecular mechanisms of epigallocatechin-3-gallate for prevention of chronic kidney disease and renal fibrosis: preclinical evidence. Current developments in nutrition 2019, 3, nzz101. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Nakamura, N.; Ojima, A.; Nishino, Y.; Yamagishi, S.-I. Sulforaphane reduces advanced glycation end products (AGEs)-induced inflammation in endothelial cells and rat aorta. Nutrition, Metabolism and Cardiovascular Diseases 2016, 26, 797–807. [Google Scholar] [CrossRef]
- Yeh, W.-J.; Hsia, S.-M.; Lee, W.-H.; Wu, C.-H. Polyphenols with antiglycation activity and mechanisms of action: A review of recent findings. Journal of food and drug analysis 2017, 25, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, Z.; Ma, L.; Xu, Q.; Yang, Q.; Si, L. Resveratrol prevents the impairment of advanced glycosylation end products (AGE) on macrophage lipid homeostasis by suppressing the receptor for AGE via peroxisome proliferator-activated receptor γ activation. International Journal of Molecular Medicine 2010, 25, 729–734. [Google Scholar] [PubMed]
- Lin, J.; Tang, Y.; Kang, Q.; Feng, Y.; Chen, A. Curcumin inhibits gene expression of receptor for advanced glycation end-products (RAGE) in hepatic stellate cells in vitro by elevating PPARγ activity and attenuating oxidative stress. British journal of pharmacology 2012, 166, 2212–2227. [Google Scholar] [CrossRef]
- Sriramoju, S.; Goetz, K. Molecular docking interaction between carotenoids and curcumin and RAGE receptor prevents diabetic retinopathy progression (P06-044-19). Current Developments in Nutrition 2019, 3, nzz031–P006. [Google Scholar] [CrossRef]
- Abdelmageed, M.E.; Shehatou, G.S.; Abdelsalam, R.A.; Suddek, G.M.; Salem, H.A. Cinnamaldehyde ameliorates STZ-induced rat diabetes through modulation of IRS1/PI3K/AKT2 pathway and AGEs/RAGE interaction. Naunyn-Schmiedeberg's Archives of Pharmacology 2019, 392, 243–258. [Google Scholar] [CrossRef]
- Hamada, Y.; Araki, N.; Horiuchi, S.; Hotta, N. Role of polyol pathway in nonenzymatic glycation. Nephrology dialysis transplantation 1996, 11, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.S.; Yoo, N.H.; Kim, N.H.; Lee, Y.M.; Kim, C.-S.; Kim, J.; Kim, J.-H.; Kim, J.S. 3, 5-Di-O-caffeoyl-epi-quinic acid from the leaves and stems of Erigeron annuus inhibits protein glycation, aldose reductase, and cataractogenesis. Biological and Pharmaceutical Bulletin 2010, 33, 329–333. [Google Scholar] [CrossRef]
- Sampath, C.; Sang, S.; Ahmedna, M. In vitro and in vivo inhibition of aldose reductase and advanced glycation end products by phloretin, epigallocatechin 3-gallate and [6]-gingerol. Biomedicine & Pharmacotherapy 2016, 84, 502–513. [Google Scholar]
- Ciddi, V.; Dodda, D. Therapeutic potential of resveratrol in diabetic complications: in vitro and in vivo studies. Pharmacological Reports 2014, 66, 799–803. [Google Scholar] [CrossRef]
- Vlassopoulos, A.; Lean, M.; Combet, E. Inhibition of protein glycation by phenolic acids: physiological relevance and implication of protein-phenolic interactions. Proceedings of the Nutrition Society 2015, 74, E88. [Google Scholar] [CrossRef]
- Cao, X.; Xia, Y.; Zeng, M.; Wang, W.; He, Y.; Liu, J. Caffeic acid inhibits the formation of advanced glycation end products (AGEs) and mitigates the AGEs-induced oxidative stress and inflammation reaction in human umbilical vein endothelial cells (HUVECs). Chemistry & biodiversity 2019, 16, e1900174. [Google Scholar]
- Bhattacherjee, A.; Datta, A. Mechanism of antiglycating properties of syringic and chlorogenic acids in in vitro glycation system. Food Research International 2015, 77, 540–548. [Google Scholar] [CrossRef]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; da Silva, N.M.; Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against α-amylase, α-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomedicine & Pharmacotherapy 2018, 100, 83–92. [Google Scholar]
- Hua, F.; Zhou, P.; Wu, H.-Y.; Chu, G.-X.; Xie, Z.-W.; Bao, G.-H. Inhibition of α-glucosidase and α-amylase by flavonoid glycosides from Lu'an GuaPian tea: molecular docking and interaction mechanism. Food & function 2018, 9, 4173–4183. [Google Scholar]
- Huang, Q.; Chai, W.-M.; Ma, Z.-Y.; Ou-Yang, C.; Wei, Q.-M.; Song, S.; Zou, Z.-R.; Peng, Y.-Y. Inhibition of α-glucosidase activity and non-enzymatic glycation by tannic acid: Inhibitory activity and molecular mechanism. International journal of biological macromolecules 2019, 141, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Kalailingam, P.; Kannaian, B.; Tamilmani, E.; Kaliaperumal, R. Efficacy of natural diosgenin on cardiovascular risk, insulin secretion, and beta cells in streptozotocin (STZ)-induced diabetic rats. Phytomedicine 2014, 21, 1154–1161. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, Y.; Li, L.; Qi, B.; Ju, M.; Xu, Y.; Zhang, Y.; Sui, X. Covalent conjugates of anthocyanins to soy protein: Unravelling their structure features and in vitro gastrointestinal digestion fate. Food Research International 2019, 120, 603–609. [Google Scholar] [CrossRef]
- Lacroix, S.; Klicic Badoux, J.; Scott-Boyer, M.-P.; Parolo, S.; Matone, A.; Priami, C.; Morine, M.J.; Kaput, J.; Moco, S. A computationally driven analysis of the polyphenol-protein interactome. Scientific reports 2018, 8, 2232. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Dissanayaka, C.S. Phenolic-protein interactions: insight from in-silico analyses–a review. Food Production, Processing and Nutrition 2023, 5, 2. [Google Scholar] [CrossRef]
- Li, Y.; He, D.; Li, B.; Lund, M.N.; Xing, Y.; Wang, Y.; Li, F.; Cao, X.; Liu, Y.; Chen, X. Engineering polyphenols with biological functions via polyphenol-protein interactions as additives for functional foods. Trends in Food Science & Technology 2021, 110, 470–482. [Google Scholar]
- Li, Y.; Jongberg, S.; Andersen, M.L.; Davies, M.J.; Lund, M.N. Quinone-induced protein modifications: Kinetic preference for reaction of 1, 2-benzoquinones with thiol groups in proteins. Free Radical Biology and Medicine 2016, 97, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Hedegaard, R.; Skibsted, L.H.; Andersen, M. Epicatechin and epigallocatechin gallate inhibit formation of intermediary radicals during heating of lysine and glucose. Food chemistry 2014, 146, 48–55. [Google Scholar] [CrossRef]
- Pham, L.B.; Wang, B.; Zisu, B.; Adhikari, B. Covalent modification of flaxseed protein isolate by phenolic compounds and the structure and functional properties of the adducts. Food Chemistry 2019, 293, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Sun, C.; Yang, W.; Yuan, F.; Gao, Y. Structural characterization and functional evaluation of lactoferrin–polyphenol conjugates formed by free-radical graft copolymerization. Rsc Advances 2015, 5, 15641–15651. [Google Scholar] [CrossRef]
- You, J.; Luo, Y.; Wu, J. Conjugation of ovotransferrin with catechin shows improved antioxidant activity. Journal of Agricultural and Food Chemistry 2014, 62, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Chandrasekara, A. Interaction of phenolics and their association with dietary fiber. Dietary Fiber Functionality in Food and Nutraceuticals: From Plant to Gut 2017, 21–44. [Google Scholar]
- Silva, C.; Correia-Branco, A.; Andrade, N.; Ferreira, A.C.; Soares, M.L.; Sonveaux, P.; Stephenne, J.; Martel, F. Selective pro-apoptotic and antimigratory effects of polyphenol complex catechin: lysine 1: 2 in breast, pancreatic and colorectal cancer cell lines. European Journal of Pharmacology 2019, 859, 172533. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Lu, Y.; Xu, H.; Lin, D.; He, Z.; Wu, H.; Liu, L.; Wang, Z. Reducing the allergenic capacity of β-lactoglobulin by covalent conjugation with dietary polyphenols. Food Chemistry 2018, 256, 427–434. [Google Scholar] [CrossRef]
- Banerjee, S.; Ji, C.; Mayfield, J.E.; Goel, A.; Xiao, J.; Dixon, J.E.; Guo, X. Ancient drug curcumin impedes 26S proteasome activity by direct inhibition of dual-specificity tyrosine-regulated kinase 2. Proceedings of the National Academy of Sciences 2018, 115, 8155–8160. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, W.; Chen, Z.; Guo, Q.; Wang, C.; Santhanam, R.K.; Chen, H. Inhibitory effect of epigallocatechin-3-O-gallate on α-glucosidase and its hypoglycemic effect via targeting PI3K/AKT signaling pathway in L6 skeletal muscle cells. International journal of biological macromolecules 2019, 125, 605–611. [Google Scholar] [CrossRef]
- Pang, X.-H.; Yang, Y.; Bian, X.; Wang, B.; Ren, L.-K.; Liu, L.-L.; Yu, D.-H.; Yang, J.; Guo, J.-C.; Wang, L. Hemp (Cannabis sativa L.) seed protein–EGCG conjugates: Covalent bonding and functional research. Foods 2021, 10, 1618. [Google Scholar] [CrossRef]
Figure 1.
Production of ROS in Mitochondria (Adapted from Matteo Martini [209]).
Figure 1.
Production of ROS in Mitochondria (Adapted from Matteo Martini [209]).
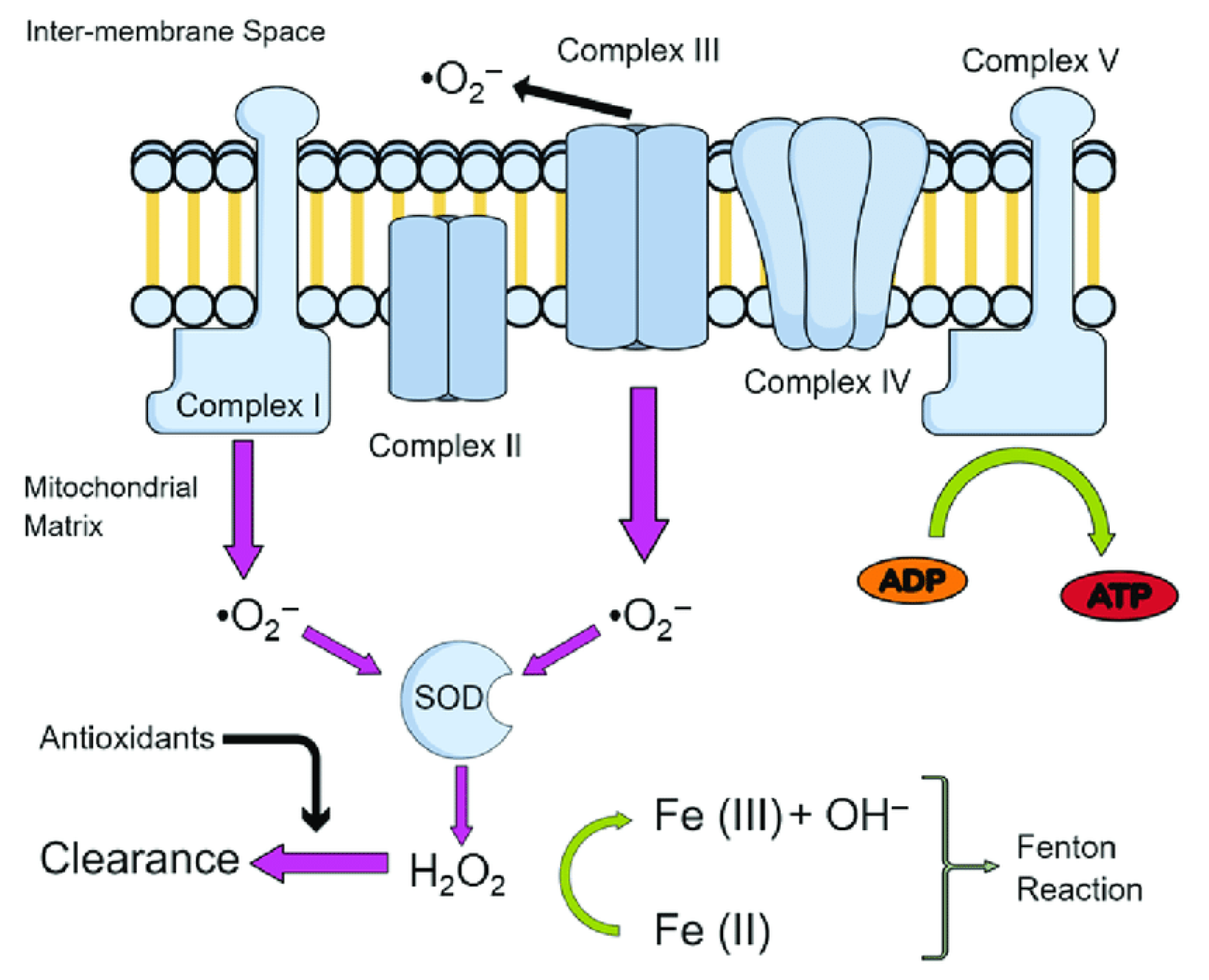
Figure 2.
Structure of the full-length RAGE, including the variable (V) domain, constant (C1 and C2) domains, the transmembrane region, and the cytoplasmic tail. A disulfide bridge between Cys38 (strand B) and Cys99 (strand F) links the two β-sheets of the V domain.
Figure 2.
Structure of the full-length RAGE, including the variable (V) domain, constant (C1 and C2) domains, the transmembrane region, and the cytoplasmic tail. A disulfide bridge between Cys38 (strand B) and Cys99 (strand F) links the two β-sheets of the V domain.
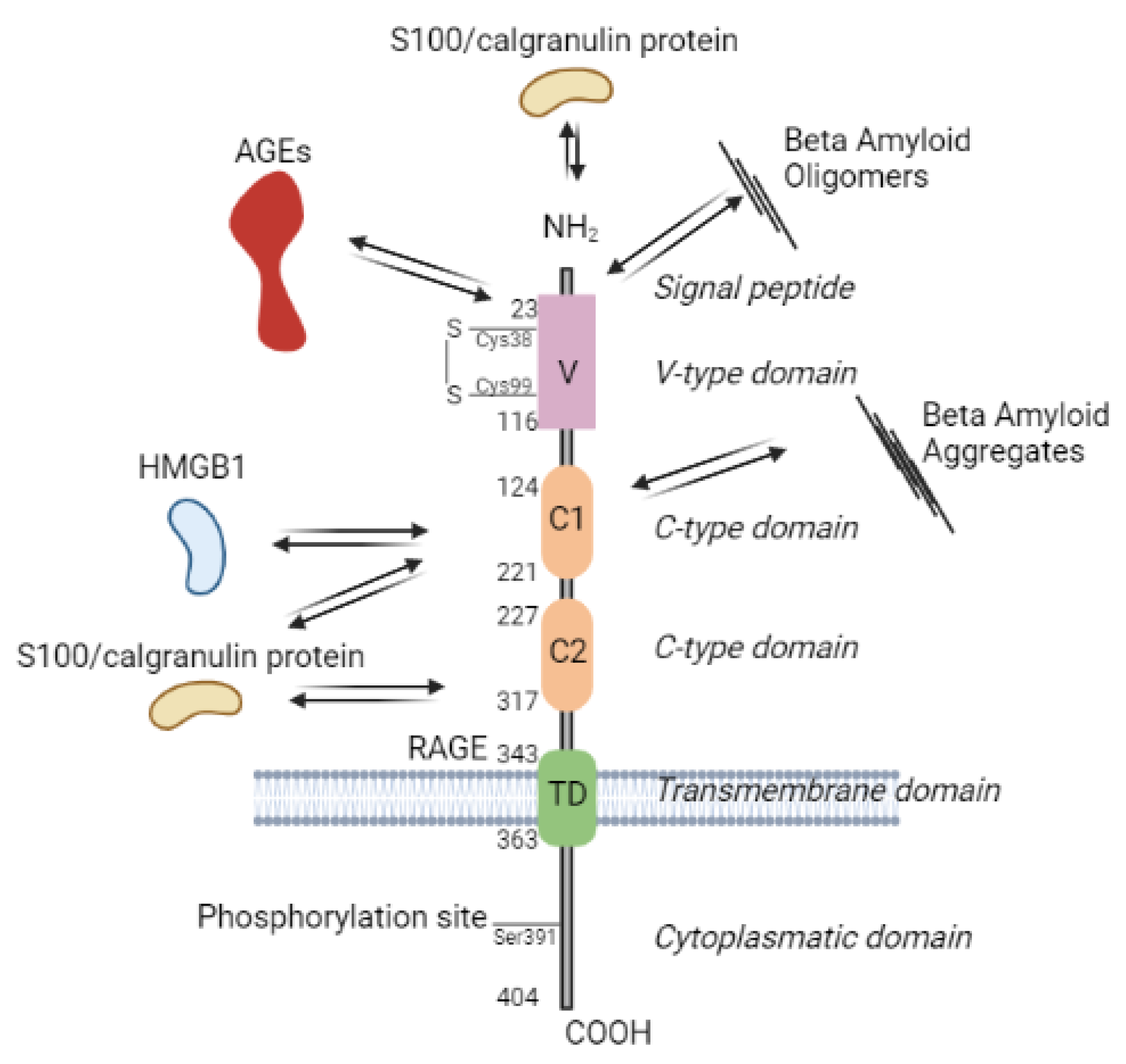

Table 1.
Overview of phenolic compound classes and anti-diabetic effects of their characteristic representatives.
Table 1.
Overview of phenolic compound classes and anti-diabetic effects of their characteristic representatives.
| Classification/core structure | Example of plant source | Plant isolate | Model | Effect | Ref. | |
|---|---|---|---|---|---|---|
| Simple phenolics | Phenolic compounds | |||||
Hydroxybenzoic acids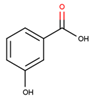
|
Sedum adenotrichum | Crude extract of whole plant | Animal (rats) -alloxan-induced T1DM | Reduced blood glucose and glycated hemoglobin levels, improved lipid profile and renal/liver function tests, inhibited α-glucosidase enzyme | [61] | |
Hydroxycinnamic acids 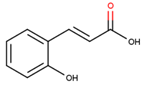
|
Cichorium intybus | Pure isolate from roots | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity | [62] | |
Coumarins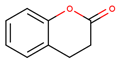
|
Angelica dahurica | Ethanol extract from roots | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, prevented diabetic nephropathy and retinopathy | [63] |
|
| Stilbenes | ||||||
Stilbene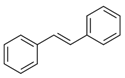
|
Polygonum cuspidatum Siebold & Zucc. | Pure isolate from roots | Obese mice (high-fat diet-induced DM) | Reduced body weight gain and adipose tissue mass, improved glucose tolerance and insulin sensitivity, modulated lipid metabolism and inflammatory response | [64] | |
| Polyphenols | Hydrolyzabletannins | |||||
Hydrolyzable tannins
|
Terminalia chebula Retz. | Ethanol extract of fruits | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, improved hepatic function and histopathology | [65] | |
| Flavonoids | ||||||
Flavone
|
Scutellaria baicalensis Georgi | Pure isolate from roots | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, improved renal function and histopathology | [66] | |
Flavonol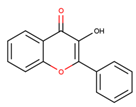
|
Morus alba L. | Pure isolate from leaves | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, improved glucose tolerance and insulin sensitivity, enhanced pancreatic β-cell function and antioxidant activity | [67] | |
Flavan-3-ols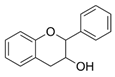
|
Bassia longifolia KOENIG (= Madhuca longifolia (L.) |
methanolic extract | In vitro | α-glucosidase inhibition, which could also explain the use of a decoction in the treatment of diabetes | [68] | |
Flavanone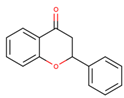
|
Citrus sinensis L. Osbeck | Pure isolate from peels | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, modulated carbohydrate metabolism and inflammatory response | [69] | |
Isoflavone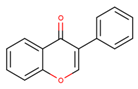
|
Glycine max (L.) Merr. | Pure isolate from seeds |
Rats (streptozotocin- induced DM) |
Reduce hyperglуcemia, inhibit the activity of tyrosine kinase, improve glucose intolerance, decrease blood glucose, total cholesterol, and 5' adenosine monophosphate-activated protein kinase (AMPK) phosphorylation | [70] | |
Anthocyanin
|
Vaccinium myrtillus L. | Ethanol extract of berries | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, prevented diabetic nephropathy and retinopathy | [71] | |
Chalcone
|
Glycyrrhiza glabra L. | Pure isolate from roots | Mice (alloxan- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, improved hepatic function and histopathology | [72] | |
| Others phenolic compounds |
Lignans
|
Linum usitatissimum L. | Pure isolate from seeds | Mice (alloxan- induced DM) |
Reduced blood glucose, antihyperlipidemic Effects, inhibitory on alpha-amylase activities and alpha-glucosidase activities |
[73] |
Lignins
|
Sesamum indicum L. | Pure isolate from seeds | Rats (streptozotocin- induced DM) |
Reduced blood glucose and serum lipid levels, increased insulin secretion and antioxidant activity, modulated carbohydrate metabolism and oxidative stress | [74] | |
Table 2.
Classification of alkaloids and anti-diabetic effects of their characteristic representatives.
Table 2.
Classification of alkaloids and anti-diabetic effects of their characteristic representatives.
| Amino acid precursor | Characteristic representative | Example of plant source | Effect | Ref. |
|---|---|---|---|---|
| Ornithine |
Pyrrolidine alkaloids
|
Brassica oleracea var. capitata leaves | Blood glucose reduction in alloxan-induced diabetic rabbits | [164] |
Tropane
|
Morus atropurpurea latex |
Dipeptidyl peptidase 4 and alpha- glycosidase inhibitory activities in vitro |
[156] | |
Pyrrolizidine alkaloids 
|
Onosma dichroanthum roots | Hypoglycemia and reducing MDA in streptozocin induced diabetic model in mice | [165] | |
| Leucine |
Pyrrole alkaloids
|
Aloe vera gel ethanolic extract |
Restore the pancreatic islets cell mass in diabetic animal model by the inhibition of the DPP-4 enzyme | [166] |
| Lysine |
Quinolizidine
|
Lupinus spp. | Enhancing glucose-induced insulin release from isolated rat islet cells in a glucose concentration-dependent manner | [167] |
Indolizidine alkaloids
|
Castanospermum australe (Fabaceae) | Alpha- glycosidase inhibitory activities in vitro | [156] | |
| Tyrosine |
Catecholamines
|
Nauclea latifolia aqueous root-bark extract | Significant reduction in glucose levels, lipid profile and serum liver enzymes in streptozotocin – induced diabetic Wistar albino rats |
[168] |
Isoquinoline
|
Berberis vulgaris | Reduced blood glucose level, increase insulin secretion, reduce body weight and lipid levels, attenuate glucose tolerance and insulin resistance by activating the 5‘-adenosine monophosphate-activated protein kinase (pathway, increase GLP-1) levels, attenuate ROS production | [169] | |
Tetrahydroisoquinoline
|
Corydalis edulis Maxim |
significantly improve insulin secretion in hamster pancreatic β-cell (HIT-T15) | [170] | |
Benzyl tetrahydroisoquinoline alkaloids
|
Alangium salvifolium stem barks | The lowering of the blood glucose level in diabetic rats | [171] | |
Phenylethylamine alkaloids
|
Cajanus cajan L. ethanolic extract | The lowering the of the blood glucose level in ICR mice | [172] | |
| Tryptophan |
Indolamines
|
Moringa oleifera Lam. flowers ethanolic extract |
Reduced blood glucose and serum lipid levels in alloxan induced diabetes in rats | [173] |
Indole
|
Catharanthus roseus (L.) G. Don dichloromethane leaves extract |
Antioxidant and hypoglycemic activity in β-TC6 mouse pancreatic cells | [159] | |
Carboline
|
Peganum harmala L. | significantly decreased the levels of glucose, triglyceride, cholesterol, and LDL-c and increased the level of HDL-c in streptozotocin-induced diabetic rats | [174] | |
Quinolone
|
Cassia angustifolia | Alpha- amylase inhibitory activities | [175] | |
Pyrroloindole
|
Catharanthus roseus | Lowering blood glucose level |
[176] | |
Ergot alkaloid
|
Ipomoea corymbosa (L) Roth ex Roem & Schult | Lowering blood glucose level |
[177] | |
| Histidine |
Imidazole alkaloids
|
Alpha-glucosidase inhibitory and hypoglycemic effects |
[178] | |
| Phenylalanine |
Ephedrine
|
Ephedra alata subsp. alenda (Ephedraceae) leaf extracts |
inhibition of α-amylase and lipase enzymes |
[179] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



