
Submitted:
11 December 2024
Posted:
12 December 2024
You are already at the latest version
Abstract
Background: Streptococcus pneumoniae serotype 1 is one of the most prevalent serotypes commonly associated with invasive pneumococcal disease cases and outbreaks worldwide. Several sequence types of this serotype have been identified globally, including those exhibiting both virulence potential and antimicrobial resistance profiles. This systematic review presents the global distribution of clones of pneumococcal serotype 1, describing their circulating patterns in various regions in the world. Methods: A database search was conducted in Google Scholar, PubMed, Scopus, ScienceDirect, and Web of Science using keywords related to Streptococcus pneumoniae serotype 1. The inclusion criteria entailed peer-reviewed studies published in English describing the utilization of at least one molecular genotyping tool to identify S. pneumoniae serotype 1 clones based on their sequence types. Data extracted were managed and analyzed using Microsoft Excel 365. Results: Forty-three studies were finally included in the systematic review. A total of 103 MLST serotype 1 sequence types were identified in 48 countries. These clones were widely reported to be associated with invasive pneumococcal diseases. Globally, ST217 and ST306 clonal complexes (CC217 and CC306) were the predominant lineages of serotype 1 sequence types, exhibiting distinct continental distribution patterns. CC217 characterized by ST217, ST303, ST612, ST618, and ST3081 was predominant in Africa and Asia. ST306 clonal complex group into ST306, ST304, and ST227 were mostly found in Europe, Oceania, North America, and some countries in South America. ST615 was predominant in Chile, Peru, and Argentina. Conclusion: The hypervirulence nature of serotype 1 coupled with its complex genetic diversity, poses a significant public health threat. Consequently, our findings emphasize the need for enhanced surveillance and targeted interventions to mitigate the spread of these hypervirulent clones, ultimately informing evidence-based strategies for disease prevention and control.
Keywords:
Streptococcus pneumoniae
; serotype 1
; sequence type
; clonal complex
; invasive pneumococcal disease
1. Introduction
Streptococcus pneumoniae is a Gram-positive, opportunistic commensal bacterium that colonizes the nasopharynx of the upper respiratory tract of humans [1]. Its ability to spread to other parts of the body and evade host immune response has marked S. pneumoniae as the leading cause of invasive pneumococcal diseases (IPD) in the world, resulting in significant morbidity and mortality, especially among vulnerable populations such as children, older adults, immunocompromised patients [2,3,4,5]. The commonly reported invasive pneumococcal diseases associated with this bacterium are pneumonia, bacteremia, and meningitis. According to the World Health Organization (WHO), S. pneumoniae is responsible for about 300,000 deaths in younger children annually, with the majority of these casualties occurring in developing countries [6]. The epidemiology of S. pneumoniae varies geographically and is influenced by several factors, including carriage patterns, vaccination strategies, antibiotic use, and the emergence of multidrug-resistant strains [7].
To date, one hundred pneumococcal serotypes have been discovered throughout the world [8], with a small number of these serotypes responsible for most invasive diseases. Serotype 1 is among the most prevalent and invasive pneumococcal serotypes globally [9], accounting for a substantial proportion of IPD cases and outbreaks in many regions, including sub-Saharan Africa, Asia, South and North America, and Europe [10]. Although serotype 1 is associated with high invasive potential, it is rarely found in the nasopharyngeal carriage of healthy individuals due to limited carriage duration, capsular characteristics, and genetic factors [9,10]. The global distribution of serotype 1 S. pneumoniae is complex, with different clones circulating in various geographic regions. Understanding the clonal composition of serotype 1 S. pneumoniae is crucial to inform vaccine development, antibiotic treatment, and disease surveillance. Molecular genotyping methods including Multilocus sequence typing (MLST), pulsed-field gel electrophoresis (PFGE), whole genome sequencing (WGS), and a multilocus variable number of tandem repeat analysis (MLVA) have provided a vivid understanding of the genetic structure of S. pneumoniae populations [11]. Studies utilizing these methods have identified several sequence types (STs) associated with serotype 1, highlighting the existence of clonal complexes that demonstrate both virulence potential and antimicrobial resistance profiles.
The introduction of pneumococcal conjugate vaccines (PCVs) has been a significant public health achievement, leading to a substantial reduction in the incidence of invasive pneumococcal disease, especially those caused by vaccine-type serotypes [12,13]. However, the widespread use of these vaccines has been associated with the phenomenon of serotype replacement, where non-vaccine serotypes become more predominant [8]. This has been observed in many countries following the implementation of PCVs, with non-vaccine types such as serotype 1 becoming more prevalent [2]. The emergence of serotype 1 as a major cause of IPD in the post-PCV era, highlights the need for continuous surveillance and monitoring of pneumococcal serotypes, particularly in low-and middle-income countries, where the burden of pneumococcal disease is highest and access to diagnostic tools and vaccines may be limited. Despite the growing body of research on S. pneumoniae serotype 1, a comprehensive synthesis to assess the distribution of its clones is lacking. By consolidating current available data on the continent, this systematic review aims to describe the circulation of serotype 1 clones and their clonal complexes.
2. Results
2.1. Search Results and Selection Process
A total of 1,234 records were identified from all five electronic databases. Two hundred and nine duplicate records were removed, leaving 1,025 unique records to be screened. Of the 1,025 unique records, 813 were excluded based on their title, abstract, and language, and the remaining 212 records were sought for full-text retrieval. After full-text retrieval, 157 articles were excluded following the full-text eligibility assessment. Finally, 43 studies were included in the systematic review (Figure 1).
2.2. Description of Included Studies
Most of the data analyzed in this study were obtained from single-center studies conducted mainly in 19 countries (n = 38 studies). The majority of the studies were found in the United Kingdom (n = 5), Spain (n = 5), the United States (n = 5), and Gambia (n = 5). Five articles were multicenter studies including two intra-continental surveys that grouped data from countries in sub-Saharan Africa and West Africa respectively, one intra-continental survey that described data from the individual countries, and two inter-continental multicenter studies that also reported data from the individual countries. Of the 43 studies, 19 studies (44.2%) focused on participants of all ages (children to adults), 16 studies (37.2) on children only, and 2 studies (4.7%) on adults only. Six studies did not specify the age group of the included participants. Most studies (n = 29, %) described invasive pneumococcal diseases (IPD) among participants while four described pneumococcal carriage. Ten studies reported on two or more isolate types (IPD and NIPD, n = 6; IPD and carriage, n = 3; all three types, n = 1). The PCVs evaluated includes PCV7, PCV9, PCV10, PCV13, PPV23, and PPV24. Studies employed polymerase chain reaction (PCR) or whole genome sequencing (WGS) or a combination thereof, alongside multilocus sequence typing (MLST) analysis for S. pneumoniae to determine their sequence types.
2.3. Pneumococcal Serotype 1 Clones Reported Worldwide
Table S1 describes the molecular characteristics of 103 pneumococcal serotype 1 clones identified globally. Based on the BURST analysis, three main clonal complexes (CCs) were identified: CC217, CC306, and CC615.
2.3.1. Africa
Our search identified seventeen (17) studies describing pneumococcal serotype 1 STs from African countries (Table 3). BURST analysis revealed that clonal complex 217 (CC217) is the largest group, comprising five main clusters CC217-ST217, ST303, ST612, ST618, and ST3081. This group was predominantly reported in Africa. The ancestral clone, ST217, was predominantly associated with pneumococcal meningitis in Niger [52], Ghana [32,33], Burkina Faso [49], and Egypt [61]. Additionally, ST217 was widely responsible for several IPD cases described in South Africa [27,39], Kenya [27,40], Ethiopia [59], Mozambique [61], Malawi [56], and Gambia [25,48,61]. Carriage of serotype 1 ST217 was reported in Gambia [25], Kenya [40], Ethiopia [61], and Nigeria [61]. Single locus variants of ST217 such as ST613 and ST614 were identified in Kenya as associated with IPD cases and, to a lesser extent, carriage [27,40]. Other SLVs including ST1316, ST1325, ST2034, ST2206, ST2839, ST3575, ST5012, ST5632, ST8158, ST8314, ST9067, and ST9529, as well as DLVs like ST12150 and ST12739 were identified in countries in sub-Saharan Africa [20,25,33,39,61].
ST303, a triple locus variant of 217 was recovered from IPD cases in Gambia [35,57,61], Ghana [32,33,61], Malawi [61], Niger [61], and Togo [58]. Its SLVs, ST1322, ST1323, and ST4755 were reported in Ghana [33,61] while others including ST11104, ST11736, ST12305, and ST12803 were identified in sub-Saharan African countries [20]. ST3081 (TLV of 217) and its SLVs, ST11779, and ST12197 were identified in Gambia [25,35,61], Senegal [61], and other countries in sub-Saharan Africa [20]. ST612 (TLV of 217) was described in studies from Ghana [33], Gambia [25], and South Africa [27,39,61] while its SLVs including ST1327, ST1328 and ST3570, were found in Ghana [33] and Gambia [25]. Another TLV of 217, ST618 was reported in Burkina Faso [49], Egypt [61], Gambia [25,35,48,57,61], South Africa [39], and Togo [58]. SLV of ST618 such as ST1331 was only reported in Ghana [33], ST2084 in Ghana and Gambia [25,61], and ST3336, ST3573, ST3574, ST3577, and ST3581 in Gambia [25]. Some studies from South Africa reported ST304. ST306, and ST611 [27,39].
2.3.2. Europe
A total of 16 studies from the European region were identified (Table 3). Clonal complex 306 was the second largest group based on the BURST analysis. It was divided into three main clusters: CC306-ST306, ST304, and ST227. This group was widespread and diverse across Europe, with a significant presence in cases of IPD reported in the 16 studies. The ancestral clone, ST306 was the predominant clone in the region frequently associated with IPD cases in the United Kingdom (UK) [36,44,45,55], Spain [23,24,43,46,51], Sweden [61], Portugal [29,60], Poland [27], Norway [27], Netherland [27], Germany [27], France [27], Czech Republic [27,42], and Croatia [61]. Alongside ST306, its double locus variant, ST304 was notably reported in France [27], Netherlands [27], Norway [27], Portugal [60], Slovenia [61], Spain [23,24,43,46,51,61], and the UK [36,44,45,55]. ST227 was reported in Denmark [27] and the UK [27,36,45,53,55] while ST228 was described in Portugal [60], France [27], and Croatia [61]. Both ST227 and ST228 were identified in Spain [23,24,43,46,61] and the Netherlands [27]. Other STs with CC306, including ST1247 (DLV of 306), ST1310 (SLV of 306), and ST2376 (SLV of 306) were reported in the UK [36] in Spain [23,46]. Additionally, ST617 (SLV of 306), ST1239 (SLV of 227), and ST3861 (SLV of 304) were reported in Norway [27], UK [36] and Spain [24] respectively. Clonal complex 217 (CC217) was also identified in European countries, with ST217 widely documented in studies reporting IPD cases in the UK [36], Denmark [27], France [27], Germany [27], and Spain [23,24]. ST618 was only reported in the Netherlands [27] but identified along with ST614 (SLV of 217) in Spain [43]. To a lesser extent ST300 (DLV), ST616 (SLV), and ST3860 (DLV) of ST615 were documented in reports from the UK [27,55], Poland [27], and Spain [24] respectively. Regarding serotype 1 carriage, ST306, ST304, ST350, ST228, and ST217 were identified in nasopharyngeal samples from healthy children attending nursery school in Belgium [28] and Portugal [22].
2.3.3. Asia
Pneumococcal serotype 1 clones reported in five studies conducted in Asia (Table 3) revealed a predominant clustering of member clones of CC217. This group encompasses subgroups such as CC303, CC618, and CC3081. In India, clones belonging to this group including ST217 and its variants ST2839, and ST5002, were identified along with ST5044 related to ST618 and ST5672 (SLV of 303) [61]. Following the introduction of PCV7 in Singapore, serotype 1 strains causing meningitis and sepsis were primarily ST217, ST303, ST5044, and ST5254 [31]. In other Asian countries, such as the Philippines, Bangladesh, Nepal, and Thailand, STs belonging to CC217 including ST217, ST303, and ST3081 were detected [61]. Notably, ST615, part of the CC615 group, was also identified in clinical isolates in Thailand [61]. In Qatar, clinical isolates from blood specimens were represented by ST217, ST303, and ST3081, as well as ST306 (CC306) [61] (Figure 4). Porat et al. found that serotype 1 ST217 was the predominant clone in the southern region of Israel, accounting for cases of acute otitis media (AOM) and invasive pneumococcal disease (IPD) in children under 5 years old [54]. Notably, ST304 was also identified in children with IPD in this study. In China, ST2296 was predominant along with singletons such as ST81, ST242, ST342, ST1263, ST3397, and ST13362 identified in isolates from invasive and non-invasive pneumococcal disease [38].
2.3.4. Americas
Six studies reported pneumococcal serotype 1 clones circulating in North American countries (Table 3). Related clones of clonal complex 306 were widely reported in this region. Studies conducted in the United States from 1990 to 2002 consistently reported ST227 (DLV of 306) as the dominant ST (Figure 5). For instance, a study assessing the clonal compositions of invasive pneumococcal serotypes in the United States in 1999, 2001, and 2002 found that ST227 was the prevalent serotype 1 isolates [47]. This was consistent with the findings of Brueggemann and Spratt [27] and Gonzalez et al. [26] confirming ST227 as the most prevalent serotype 1 clone associated with cases of IPD. However, from 2003 onwards, multiple STs closely related to ST227, including ST304, 306, 2126, 4290 (SLV 306), 4288, and 4289, were reported in cases of Pediatric Pneumococcal Empyema (PPE) in Utah [19] (Figure 5). Like in the United States, ST227 was the predominant ST in Canada [27]. In contrast, in Mexico, cases of serotype 1 IPD were predominantly associated with ST304 [42].
Three studies reported pneumococcal serotype 1 clones in six countries from South America (Table). These studies documented the widespread of member clones of CC306 and CC615. In Argentina and Uruguay [42,61], the dominant clones identified in isolates of IPD cases were ST615 and ST304. ST615 clone was reported in Chile [27] and Peru [61] in isolates from blood and pleural fluid. According to Zemlicková et al. [42], serotype 1 IPD in Brazil and Colombia were caused by ST304 and ST306 respectively.
2.3.5. Oceania
Our search identified six studies from Oceania that reported clones of pneumococcal serotype 1 (Table 3). In this region, most reported STs belonged to clonal complex 306. In Australia, ST304 and ST227 were widely reported in serotype 1 IPD cases and carriage in the north according to Smith-Vaughan et al. [30]. An outbreak of serotype 1 IPD in Australia between 2008 and 2012 was predominantly caused by ST306 clones [34,37]. The proportion of ST304 isolates reported in Australia significantly decreased during this period, attributed to the clonal expansion of CC306, characterized by an increase in ST306 isolates (Figure 6). In New Caledonia (NC), ST306 emerged as the most predominant serotype 1 clone responsible for outbreaks reported [21]. Genomic characterization of serotype 1 isolates from outbreaks in NC discovered ST306 strains (62/67, 92.53%) and their single locus variants, ST3717 (5/67, 7.46%) [21]. Additionally, ST306 serotype 1 isolates were reported in cases of IPD 61 in New Zealand. ,
3. Discussion
Streptococcus pneumonaie serotype 1 has been a leading cause of severe pneumococcal disease worldwide, affecting both children and adults. Notably, this serotype is seldom found in healthy individuals, suggesting that while it may not be easily transmitted, it has a high tendency to progress to invasive disease once carried. This implies that serotype 1 is remarkably hypervirulent, corroborating its frequent association with outbreaks of severe pneumococcal infection. Furthermore, several clones, including ST217 and ST306, have been linked to outbreaks of IPD globally. In light of these findings, we conducted a systematic review to elucidate the molecular epidemiology of S. pneumoniae serotype 1 clones circulating worldwide. This comprehensive analysis seeks to identify high-risk clones with the potential to trigger outbreaks in various countries or continents, ultimately informing targeted evidence-based strategies for disease prevention and control.
The multilocus sequence typing (MLST) technique is a widely accepted method for bacterial genotyping, leveraging seven housekeeping genes (aroE, gdh, gki, recP, spi, xpt, and ddl) to characterize S. pneumoniae isolates. As the golden standard for S. pneumoniae genotyping, MLST was mainly used in most studies to elucidate the genetic diversity of this pathogen. We identified one hundred and three (103) different MLST sequence types associated with serotype 1. Most of these STs shared genetic relationships with ST217 followed by ST306.
The S. pneumoniae ST217 clonal complex is a hypervirulent lineage of significant public health concern. In our study, this lineage was mostly reported in studies from Africa and Asia. The ancestral clone, ST217 was predominantly associated with pneumococcal meningitis across several countries, including, Nigeria, Niger, Israel, India, Thailand, Singapore, and the Philippines. ST217 was also linked to numerous IPD cases in Kenya, South Africa, Ethiopia, Mozambique, and Malawi. Although not the most common clone, ST217 was associated with a serotype 1 pneumococcal meningitis outbreak in northern Ghana [33]. This finding is not surprising, since the predominance of ST217 in sub-Saharan African countries has been previously reported in several studies [62]. This widespread presence of ST217 may be attributed to intra and intercontinental movements of people between countries, leading to the transfer of clones. Other clones within CC217, such as ST618, were prevalent among serotype 1 isolates from both invasive and carriage isolates in Gambia. Also, ST618 was responsible for pneumococcal meningitis in patients from Burkina Faso [49] and Egypt [61]. Additionally, studies from neighboring countries like Ghana and Togo, as well as Nepal and Bangladesh, mostly described ST303 as associated with bacterial meningitis. Several other STs within CC217 were also identified in Africa and Asia (Table 3) indicating an extensive clonal expansion and diversity of the ST217 clonal complex, particularly in Africa. The propensity of ST217 and its clonal complex 217 to cause pneumococcal meningitis poses a significant threat to these regions, emphasizing the importance of implementing mitigation strategies to curb their spread. Fortunately, many isolates of CC217 are sensitive to antibiotics, making their treatment easy and effective [33]. While the ST217 clonal complex is predominantly reported in Africa and Asia, its presence has also been noted in other parts of the world. For example, studies have reported STs from the clonal complex in regions like Australia, the US, and specific countries in Europe.
ST306 clonal complex is another lineage of serotype 1 of public health importance. This group emerged as the prominent group in Europe, Oceania, North America, and some countries in South America characterized by its division into three main subgroups: ST306, ST304, and ST227. ST306 (ancestral clone), was the most predominant ST among this group widely dispersed in most European countries. ST304 was common in Mexico, Brazil, Uruguay, and Slovenia. STs of CC306 were more associated with bacteremia than other pneumococcal diseases. Our analysis revealed that ST227 was the predominant clone in Canada and the US, supporting the findings of Brueggemann & Spratt [27]. However, our results diverged from theirs regarding the UK, where they reported ST227 as the dominant clone. This discrepancy suggests that a clonal shift may have occurred in the UK over time, potentially replacing ST227 with ST306 as discovered in our study. Like CC217, the genetic variation of ST306 clonal complex is also illustrated by the identification of several other related STs.
One key finding of our study is the geographic structure among major serotype 1 lineages, revealing distinctive regional differences in circulating clones. This observation may be attributed to the rarity of serotype 1 carriage. Unlike other serotypes that can easily be carried and transferred between populations via human movement, the rarity of serotype 1 carriage restricts its geographic spread and may contribute to unique clonal distributions. Also, this unique clonal distribution may be attributed to geographic weather conditions in the various regions. For instance, it might be that the ST217 clonal complex thrives in humid and tropical areas which may explain why they are widely found in Africa and Asia. This possibly applies to ST306 and ST615 clonal complexes that are common in temperate climate areas.
While our study provides valuable insight into the global circulation of serotype 1 S. pneumoniae, it is not without limitations. One notable constraint is the geographical representation of the included studies, predominantly clustered in forty-eight (48) countries. The uneven distribution leaves a significant knowledge gap in many countries, highlighting the need for further research to understand better the epidemiological landscape of pneumococcal serotype 1 clones in these underrepresented regions.
4. Materials and Methods
This systematic review adhered to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines 14.
4.1. Literature Search Strategy and Selection Process
An extensive literature search was conducted in Google Scholar, PubMed, Scopus, ScienceDirect, and Web of Science using keywords related to Streptococcus pneumoniae serotype 1. The search was conducted until October 2024 and restricted to free full-text or open-access research articles published in English (Table 1). Retrieved records were exported in RIS and CSV formats.
Citations of the retrieved records were imported to the Rayyan application [15] (a web-based tool for systematic reviews) for screening. Duplicate records were first detected and removed, followed by title and abstract screening of the remaining unique records, where all unrelated records were excluded. After title and abstract screening, the remaining articles were assessed for full-text eligibility based on the abovementioned inclusion criteria. Included studies were imported to Zotero 7.0.8 for data extraction. O.K.N. and E.S.D. assessed the eligibility of the retrieved records.
4.2. Eligibility Criteria
This study included English-published peer-reviewed reports that utilized at least one molecular genotyping tool to describe S. pneumoniae serotype 1 clones based on their sequence types (STs). Multicenter studies describing serotype 1 clones in specific geographic regions (e.g., West Africa, sub-Saharan Africa) or those reporting clone data for individual countries were eligible for inclusion. Exclusions included non-peer-reviewed sources (Theses, dissertations, editorials, comments, preprints, etc.), studies not reporting serotype 1 clones, studies that selected specific serotype 1 clones for genetic analysis, and studies that focused or reported on other serotypes or their clones.
4.3. Data Extraction and Analysis
The relevant data were extracted from each included study as described in Table 2 by O.K.N. and E.S.D. The sequence types (STs) of S. pneumoniae serotype 1 clones were extracted and stratified by country/region. Microsoft Excel 365 was used to extract and manage the extracted data.
4.4. BURST Cluster Analysis
The allelic profiles of identified STs were downloaded from the Streptococcus pneumoniae database on the MLST website (https://pubmlst.org/organisms/streptococcus-pneumoniae). STs were grouped into clonal complexes according to their allelic profiles using the BURST algorithm [16]. Furthermore, the geoBURST algorithm in the Phyloviz online application (https://online.phyloviz.net/index) was used to construct the minimum spanning tree, providing a visual representation of the genetic relationship among the STs.
4.5. Risk of Bias Assessment
Two investigators (O.K.N. and E.S.D) assessed the risk of bias of the included studies using the STROBE checklist for reporting observational studies [17], as adapted according to Sanderson et al. [18] The assessment tool comprised of five key domains, method of selecting study participants, method of measuring exposure and outcome variables, method of control confounding, design-specific source of bias, and statistical methods (Supplementary table S2). Studies were graded into three categories: High risk of bias (≥1 of any criteria classified as H, or ≥2 major criteria as M or U); Moderate risk of bias (≥2 of any criteria classified as M or U (<2 major criteria)); Low risk of bias (all major criteria classified as L and <2 minor criteria as M or U.
5. Conclusion
This systematic review highlights the complex genetic diversity of S. pneumoniae serotype 1, characterized by distinct geographic structures among major lineages. A striking continental distribution emerged, with the ST217 clonal complex predominantly clustered in Africa and Asia, whereas the ST306 clonal complex was widespread across Europe, the Americas, and Oceania. Notably, invasive pneumococcal disease (IPD) was primarily linked to clones from these clonal complexes, with few instances of carriage. The findings of this study emphasize the need for enhanced surveillance and targeted interventions to mitigate the spread of hypervirulent clones, ultimately informing evidence-based strategies for disease prevention and control. To address existing knowledge gaps, further research is warranted, particularly in countries with limited or no data on serotype 1 S. pneumoniae clones.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: E.S.D. and O. K. N.; Data curation: E.S.D. and O. K. N.; Formal analysis: E.S.D. and O. K. N.; Investigation: E.S.D. and O. K. N.; Methodology: E.S.D. and O. K. N.; Software: E.S.D. and O. K. N.; Resource: E.S.D. and O. K. N.; Project administration: O.K.N. and E.S.D; Supervision: E.S.D. Validation: E.S.D. and O. K. N.; Funding acquisition: E.S.D.; Writing – original draft: O.K.N.; Writing – review & editing: O.K.N., and E.S.D.
Funding
This review paper was supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health through the PneumoSIC Study (Invasive Pneumococcal Disease and Carriage among Children with Sickle Cell Disease in Ghana: A Post-Vaccination Study) hosted at the Department of Medical Microbiology, University of Ghana Medical School (Award Number: R01AI169674). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Acknowledgments
Not applicable.
Conflicts of interest
Authors have no competing interest to declare.
Ethics approval and consent to participate
Not applicable.
Availability of data and materials
Data and materials used for the analysis are available upon request. Table S1 is available as a supplementary material.
Consent for publication
Not applicable.
References
- Bogaert, D.; De Groot, R.; Hermans, P.W.M. Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect Dis. 2004 Mar;4(3):144–54.
- Løchen, A.; Croucher, N.J.; Anderson, R.M. Divergent serotype replacement trends and increasing diversity in pneumococcal disease in high income settings reduce the benefit of expanding vaccine valency. Sci Rep. 2020 Nov 4;10(1):18977.
- Lynch, J.P.; Zhanel, G.G. Streptococcus pneumoniae: epidemiology, risk factors, and strategies for prevention. Semin Respir Crit Care Med. 2009 Apr;30(2):189–209.
- O’Brien, K.L.; Wolfson, L.J.; Watt, J.P.; Henkle, E.; Deloria-Knoll, M.; McCall, N. , et al. Burden of disease caused by Streptococcus pneumoniae in children younger than 5 years: global estimates. Lancet Lond Engl. 2009 Sep 12;374(9693):893–902.
- von Mollendorf, C.; von Gottberg, A.; Tempia, S.; Meiring, S.; de Gouveia, L.; Quan, V. , et al. Increased risk for and mortality from invasive pneumococcal disease in HIV-exposed but uninfected infants aged .
- CDC Pneumococcal Disease Surveillance and Trends [Internet]. Pneumococcal Disease. 2024 [cited 2024 Dec 3]. Available from: https://www.cdc.gov/pneumococcal/php/surveillance/index.html.
- Lynch, J.P.; Zhanel, G.G. Streptococcus pneumoniae: epidemiology and risk factors, evolution of antimicrobial resistance, and impact of vaccines. Curr Opin Pulm Med. 2010 May;16(3):217–25.
- Ganaie, F.; Saad, J.S.; McGee, L.; van Tonder, A.J.; Bentley, S.D.; Lo, S.W. , et al. A New Pneumococcal Capsule Type, 10D, is the 100th Serotype and Has a Large cps Fragment from an Oral Streptococcus. mBio. 2020 May 19;11(3):e00937-20.
- Brueggemann, A.B.; Griffiths, D.T.; Meats, E.; Peto, T.; Crook, D.W.; Spratt, B.G. Clonal relationships between invasive and carriage Streptococcus pneumoniae and serotype- and clone-specific differences in invasive disease potential. J Infect Dis. 2003 May 1;187(9):1424–32.
- Chaguza, C.; Yang, M.; Jacques, L.C.; Bentley, S.D.; Kadioglu, A. Serotype 1 pneumococcus: epidemiology, genomics, and disease mechanisms. Trends Microbiol. 2022 Jun 1;30(6):581–92.
- Rayner, R.E.; Savill, J.; Hafner, L.M.; Huygens, F. Genotyping Streptococcus pneumoniae. Future Microbiol. 2015;10(4):653–64.
- Horn, E.K.; Wasserman, M.D.; Hall-Murray, C.; Sings, H.L.; Chapman, R.; Farkouh, R.A. Public health impact of pneumococcal conjugate vaccination: a review of measurement challenges. Expert Rev Vaccines. 2021 Oct 3;20(10):1291–309.
- Mackenzie, G.A.; Hill, P.C.; Jeffries, D.J.; Ndiaye, M.; Sahito, S.M.; Hossain, I. , et al. Impact of the introduction of pneumococcal conjugate vaccination on invasive pneumococcal disease and pneumonia in The Gambia: 10 years of population-based surveillance. Lancet Infect Dis. 2021 Sep 1;21(9):1293–302.
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D. , et al. PRISMA 2020 explanation and elaboration: updated guidance and exemplars for reporting systematic reviews. The BMJ. 2021 Mar 29;372:n160.
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan-a web and mobile app for systematic reviews. Syst Rev. 2016 Dec 5;5(1):210.
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018;3:124.
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P. , et al. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: guidelines for reporting observational studies. J Clin Epidemiol. 2008 Apr;61(4):344–9.
- Sanderson, S.; Tatt, I.D.; Higgins, J.P.T. Tools for assessing quality and susceptibility to bias in observational studies in epidemiology: a systematic review and annotated bibliography. Int J Epidemiol. 2007 Jun;36(3):666–76.
- Byington, C.; Hulten, K.G.; Ampofo, K.; Shen, X.; At, P.; Aj, B. , et al. Molecular epidemiology of pediatric pneumococcal empyema from 2001 to 2007 in Utah. J Clin Microbiol [Internet]. 2010 Feb [cited 2024 Nov 4];48(2). Available from: https://pubmed.ncbi.nlm.nih.gov/20018815/.
- Chaguza, C.; Yang, M.; Cornick, J.E.; du Plessis, M.; Gladstone, R.A.; Ba, K.A. , et al. Bacterial genome-wide association study of hyper-virulent pneumococcal serotype 1 identifies genetic variation associated with neurotropism. Commun Biol [Internet]. 2020 Oct 8 [cited 2024 Nov 4];3(1). Available from: https://pubmed.ncbi.nlm.nih.gov/33033372/.
- Hanachi, M.; Kiran, A.M.; Cornick, J.E.; Harigua-Souiai, E.; Everett, D.; Benkahla, A. , et al. Genomic Characteristics of Invasive Streptococcus pneumoniae Serotype 1 in New Caledonia Prior to the Introduction of PCV13. Bioinforma Biol Insights [Internet]. 2020 Sep 29 [cited 2024 Nov 4];14. Available from: https://pubmed.ncbi.nlm.nih.gov/33088176/.
- Almeida, S.; de Lencastre, H.; Sá-Leão, R. Epidemiology and population structure of serotypes 1, 5 and 7f carried by children in Portugal from 1996-2010 before introduction of the 10-valent and 13-valent pneumococcal conjugate vaccines. PloS One [Internet]. 2013 Sep 18 [cited 2024 Nov 4];8(9). Available from: https://pubmed.ncbi.nlm.nih.gov/24058686/.
- Esteva, C.; Selva, L.; de Sevilla, M.F.; Garcia-Garcia, J.J.; Pallares, R.; Muñoz-Almagro, C. Streptococcus pneumoniae serotype 1 causing invasive disease among children in Barcelona over a 20-year period (1989-2008). Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis [Internet]. 2011 Sep [cited 2024 Nov 4];17(9). Available from: https://pubmed.ncbi.nlm.nih.gov/21729192/.
- Marimon, J.M.; Ercibengoa, M.; Alonso, M.; Zubizarreta, M.; Pérez-Trallero, E. Clonal structure and 21-year evolution of Streptococcus pneumoniae serotype 1 isolates in northern Spain. Clin Microbiol Infect Off Publ Eur Soc Clin Microbiol Infect Dis [Internet]. 2009 Sep [cited 2024 Nov 4];15(9). Available from: https://pubmed.ncbi.nlm.nih.gov/19702591/.
- Antonio, M.; Hakeem, I.; Awine, T.; Secka, O.; Sankareh, K.; Nsekpong, D. , et al. Seasonality and outbreak of a predominant Streptococcus pneumoniae serotype 1 clone from The Gambia: expansion of ST217 hypervirulent clonal complex in West Africa. BMC Microbiol. 2008 Nov 17;8:198.
- Gonzalez, B.E.; Hulten, K.G.; Kaplan, S.L.; Mason, E.O. US Pediatric Multicenter Pneumococcal Surveillance Study Group. Clonality of Streptococcus pneumoniae serotype 1 isolates from pediatric patients in the United States. J Clin Microbiol. 2004 Jun;42(6):2810–2.
- Brueggemann, A.B.; Spratt, B.G. Geographic distribution and clonal diversity of Streptococcus pneumoniae serotype 1 isolates. J Clin Microbiol. 2003 Nov;41(11):4966–70.
- Jourdain, S.; Drèze, P.A.; Verhaegen, J.; Van Melderen, L.; Smeesters, P.R. Carriage-associated Streptocccus pneumoniae serotype 1 in Brussels, Belgium. Pediatr Infect Dis J. 2013 Jan;32(1):86–7.
- Horácio, A.N.; Silva-Costa, C.; Diamantino-Miranda, J.; Lopes, J.P.; Ramirez, M.; Melo-Cristino, J. , et al. Population Structure of Streptococcus pneumoniae Causing Invasive Disease in Adults in Portugal before PCV13 Availability for Adults: 2008-2011. PloS One. 2016;11(5):e0153602.
- Smith-Vaughan, H.; Marsh, R.; Mackenzie, G.; Fisher, J.; Morris, P.S.; K, H.; et al. Age-specific cluster of cases of serotype 1 Streptococcus pneumoniae carriage in remote indigenous communities in Australia. Clin Vaccine Immunol CVI [Internet]. 2009 Feb [cited 2024 Nov 4];16(2). Available from: https://pubmed.ncbi.nlm.nih.gov/19091995/.
- Jauneikaite, E.; Jefferies, J.M.; Churton, W.V.; Lin, R.P.T.; Hibberd, M.L.; Clarke, C.S. Genetic diversity of Streptococcus pneumoniae causing meningitis and sepsis in Singapore during the first year of PCV7 implementation. Emerg Microbes Infect [Internet]. 2014 Jun [cited 2024 Nov 4];3(6). Available from: https://pubmed.ncbi.nlm.nih.gov/26038742/.
- Kwambana-Adams, B.A.; Asiedu-Bekoe, F.; Sarkodie, B.; Afreh, O.K.; Kuma, G.K.; Owusu-Okyere, G. , et al. An outbreak of pneumococcal meningitis among older children (≥5 years) and adults after the implementation of an infant vaccination programme with the 13-valent pneumococcal conjugate vaccine in Ghana. BMC Infect Dis. 2016 Oct 18;16(1):575.
- Leimkugel, J.; Adams Forgor, A.; Gagneux, S.; Pflüger, V.; Flierl, C.; Awine, E. , et al. An outbreak of serotype 1 Streptococcus pneumoniae meningitis in northern Ghana with features that are characteristic of Neisseria meningitidis meningitis epidemics. J Infect Dis. 2005 Jul 15;192(2):192–9.
- Lai, J.Y.; Cook, H.; Yip, T.W.; Berthelsen, J.; Gourley, S.; Krause, V. , et al. Surveillance of pneumococcal serotype 1 carriage during an outbreak of serotype 1 invasive pneumococcal disease in central Australia 2010-2012. BMC Infect Dis. 2013 Sep 3;13:409.
- Ebruke, C.; Roca, A.; Egere, U.; Darboe, O.; Hill, P.C.; Greenwood, B. , et al. Temporal changes in nasopharyngeal carriage of Streptococcus pneumoniae serotype 1 genotypes in healthy Gambians before and after the 7-valent pneumococcal conjugate vaccine. PeerJ. 2015;3:e903.
- Kirkham, L.A.S.; Jefferies, J.M.C.; Kerr, A.R.; Jing, Y.; Clarke, S.C.; Smith, A. , et al. Identification of Invasive Serotype 1 Pneumococcal Isolates That Express Nonhemolytic Pneumolysin. J Clin Microbiol. 2006 Jan;44(1):151.
- Staples, M.; Graham, R.M.A.; Jennison, A.V.; Ariotti, L.; Hicks, V.; Cook, H. , et al. Molecular characterization of an Australian serotype 1 Streptococcus pneumoniae outbreak. Epidemiol Infect. 2015 Jan;143(2):325–33.
- Zhou, H.; Guo, J.; Qin, T.; Ren, H.; Xu, Y.; Wang, C. , et al. Serotype and MLST-based inference of population structure of clinical Streptococcus pneumoniae from invasive and noninvasive pneumococcal disease. Infect Genet Evol J Mol Epidemiol Evol Genet Infect Dis. 2017 Nov;55:104–11.
- du Plessis, M.; Allam, M.; Tempia, S.; Wolter, N.; de Gouveia, L.; von Mollendorf, C. , et al. Phylogenetic Analysis of Invasive Serotype 1 Pneumococcus in South Africa, 1989 to 2013. J Clin Microbiol. 2016 May;54(5):1326–34.
- Brueggemann, A.B.; Muroki, B.M.; Kulohoma, B.W.; Karani, A.; Wanjiru, E.; Morpeth, S. , et al. Population genetic structure of Streptococcus pneumoniae in Kilifi, Kenya, prior to the introduction of pneumococcal conjugate vaccine. PloS One. 2013;8(11):e81539.
- Donkor, E.S.; Adegbola, R.A.; Wren, B.W.; Antonio, M. Population biology of Streptococcus pneumoniae in West Africa: multilocus sequence typing of serotypes that exhibit different predisposition to invasive disease and carriage. PloS One. 2013;8(1):e53925.
- Zemlicková, H.; Crisóstomo, M.I.; Brandileone, M.C.; Camou, T.; Castañeda, E.; Corso, A. , et al. Serotypes and clonal types of penicillin-susceptible streptococcus pneumoniae causing invasive disease in children in five Latin American countries. Microb Drug Resist Larchmt N. 2005;11(3):195–204.
- Muñoz-Almagro, C.; Ciruela, P.; Esteva, C.; Marco, F.; Navarro, M.; Bartolome, R. , et al. Serotypes and clones causing invasive pneumococcal disease before the use of new conjugate vaccines in Catalonia, Spain. J Infect. 2011 Aug;63(2):151–62.
- Cooke, B.; Smith, A.; Diggle, M.; Lamb, K.; Robertson, C.; Inverarity, D. , et al. Antibiotic resistance in invasive Streptococcus pneumoniae isolates identified in Scotland between 1999 and 2007. J Med Microbiol. 2010 Oct;59(Pt 10):1212–8.
- Clarke, S.C.; Jefferies, J.M.C.; Smith, A.J.; McMenamin, J.; Mitchell, T.J.; Edwards, G.F.S. Pneumococci causing invasive disease in children prior to the introduction of pneumococcal conjugate vaccine in Scotland. J Med Microbiol. 2006 Aug;55(Pt 8):1079–84.
- Muñoz-Almagro, C.; Jordan, I.; Gene, A.; Latorre, C.; Garcia-Garcia, J.J.; Pallares, R. Emergence of invasive pneumococcal disease caused by nonvaccine serotypes in the era of 7-valent conjugate vaccine. Clin Infect Dis Off Publ Infect Dis Soc Am. 2008 Jan 15;46(2):174–82.
- Beall, B.; McEllistrem, M.C.; Gertz, R.E.; Wedel, S.; Boxrud, D.J.; Gonzalez, A.L. , et al. Pre- and postvaccination clonal compositions of invasive pneumococcal serotypes for isolates collected in the United States in 1999, 2001, and 2002. J Clin Microbiol. 2006 Mar;44(3):999–1017.
- Antonio, M.; Dada-Adegbola, H.; Biney, E.; Awine, T.; O’Callaghan, J.; Pfluger, V. , et al. Molecular epidemiology of pneumococci obtained from Gambian children aged 2-29 months with invasive pneumococcal disease during a trial of a 9-valent pneumococcal conjugate vaccine. BMC Infect Dis. 2008 Jun 11;8:81.
- Yaro, S.; Lourd, M.; Traoré, Y.; Njanpop-Lafourcade, B.M.; Sawadogo, A.; Sangare, L. , et al. Epidemiological and molecular characteristics of a highly lethal pneumococcal meningitis epidemic in Burkina Faso. Clin Infect Dis Off Publ Infect Dis Soc Am. 2006 Sep 15;43(6):693–700.
- Zähner, D.; Gudlavalleti, A.; Stephens, D.S. Increase in Pilus Islet 2–encoded Pili among Streptococcus pneumoniae Isolates, Atlanta, Georgia, USA. Emerg Infect Dis. 2010 Jun;16(6):955.
- Grau, I.; Ardanuy, C.; Calatayud, L.; Rolo, D.; Domenech, A.; Liñares, J. , et al. Invasive pneumococcal disease in healthy adults: increase of empyema associated with the clonal-type Sweden(1)-ST306. PloS One. 2012;7(8):e42595.
- Kourna Hama, M.; Khan, D.; Laouali, B.; Okoi, C.; Yam, A.; Haladou, M. , et al. Pediatric Bacterial Meningitis Surveillance in Niger: Increased Importance of Neisseria meningitidis Serogroup C, and a Decrease in Streptococcus pneumoniae Following 13-Valent Pneumococcal Conjugate Vaccine Introduction. Clin Infect Dis Off Publ Infect Dis Soc Am. 2019 Sep 5;69(Suppl 2):S133–9.
- Jefferies, J.M.; Smith, A.J.; Edwards, G.F.S.; McMenamin, J.; Mitchell, T.J.; Clarke, S.C. Temporal analysis of invasive pneumococcal clones from Scotland illustrates fluctuations in diversity of serotype and genotype in the absence of pneumococcal conjugate vaccine. J Clin Microbiol. 2010 Jan;48(1):87–96.
- Porat, N.; Benisty, R.; Trefler, R.; Givon-Lavi, N.; Dagan, R. Clonal distribution of common pneumococcal serotypes not included in the 7-valent conjugate vaccine (PCV7): marked differences between two ethnic populations in southern Israel. J Clin Microbiol. 2012 Nov;50(11):3472–7.
- Foster, D.; Knox, K.; Walker, A.S.; Griffiths, D.T.; Moore, H.; Haworth, E. , et al. Invasive pneumococcal disease: epidemiology in children and adults prior to implementation of the conjugate vaccine in the Oxfordshire region, England. J Med Microbiol. 2008 Apr;57(Pt 4):480–7.
- Chaguza, C.; Cornick, J.E.; Andam, C.P.; Gladstone, R.A.; Alaerts, M.; Musicha, P. , et al. Population genetic structure, antibiotic resistance, capsule switching and evolution of invasive pneumococci before conjugate vaccination in Malawi. Vaccine. 2017 Aug 16;35(35 Pt B):4594–602.
- Sanneh, B.; Okoi, C.; Grey-Johnson, M.; Bah-Camara, H.; Kunta Fofana, B.; Baldeh, I. , et al. Declining Trends of Pneumococcal Meningitis in Gambian Children After the Introduction of Pneumococcal Conjugate Vaccines. Clin Infect Dis Off Publ Infect Dis Soc Am. 2019 Sep 5;69(Suppl 2):S126–32.
- Tsolenyanu, E.; Bancroft, R.E.; Sesay, A.K.; Senghore, M.; Fiawoo, M.; Akolly, D. , et al. Etiology of Pediatric Bacterial Meningitis Pre- and Post-PCV13 Introduction Among Children Under 5 Years Old in Lomé, Togo. Clin Infect Dis Off Publ Infect Dis Soc Am. 2019 Sep 5;69(Suppl 2):S97–104.
- Sharew, B.; Moges, F.; Yismaw, G.; Mihret, A.; Lobie, T.A.; Abebe, W. , et al. Molecular epidemiology of Streptococcus pneumoniae isolates causing invasive and noninvasive infection in Ethiopia. Sci Rep. 2024 Sep 13;14(1):21409.
- Serrano, I.; Melo-Cristino, J.; Carriço, J.A.; Ramirez, M. Characterization of the genetic lineages responsible for pneumococcal invasive disease in Portugal. J Clin Microbiol. 2005 Apr;43(4):1706–15.
- Cornick, J.E.; Chaguza, C.; Harris, S.R.; Yalcin, F.; Senghore, M.; Kiran, A.M. , et al. Region-specific diversification of the highly virulent serotype 1 Streptococcus pneumoniae. Microb Genomics. 2015 Aug;1(2):e000027.
- Chaguza, C.; Cornick, J.E.; Harris, S.R.; Andam, C.P.; Bricio-Moreno, L.; Yang, M. , et al. Understanding pneumococcal serotype 1 biology through population genomic analysis. BMC Infect Dis. 2016 Nov 8;16(1):649.
Figure 1.
PRISMA flow diagram of the study selection process.
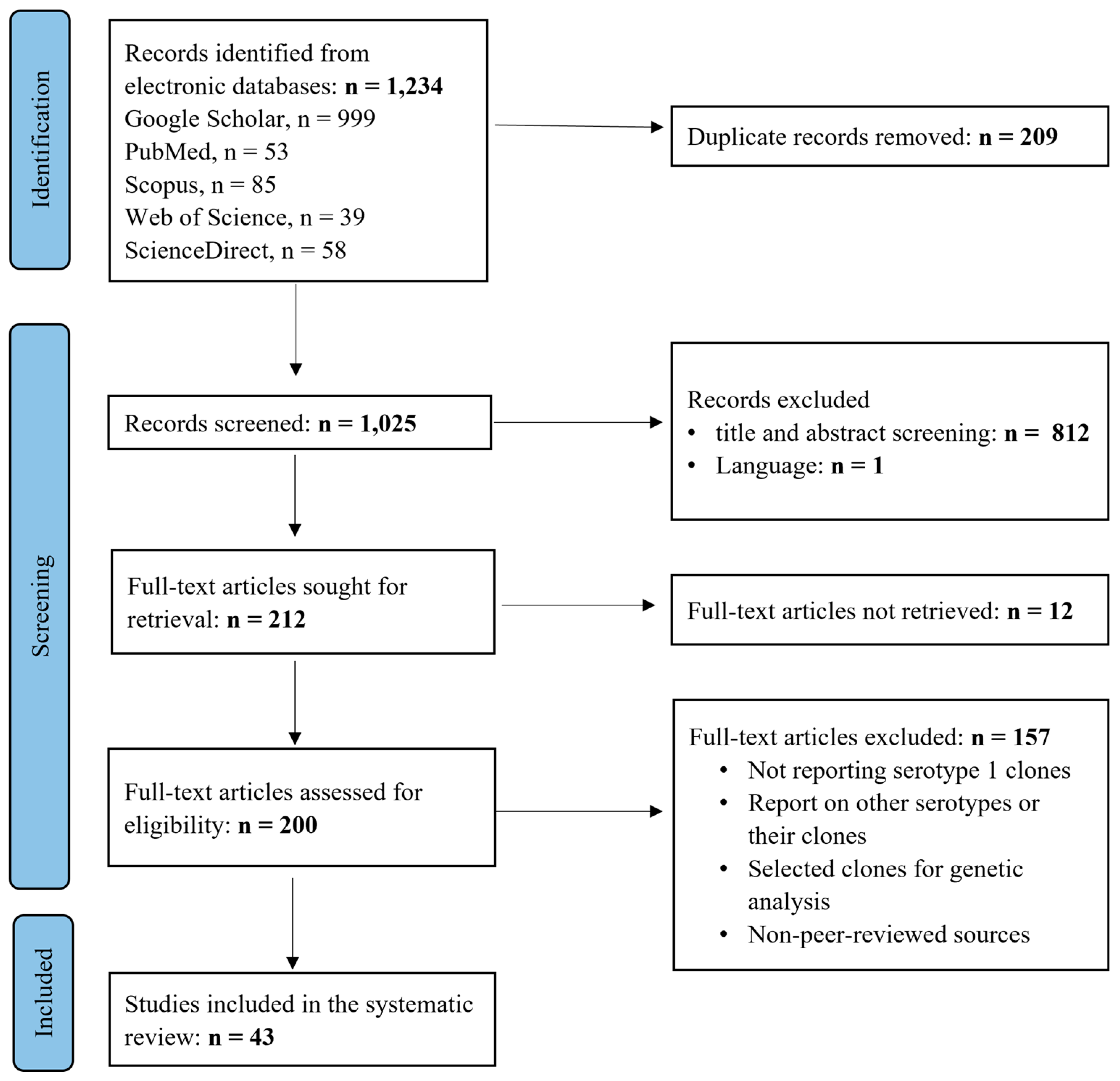
Figure 2.
Distribution of serotype 1 isolates identified in Africa.
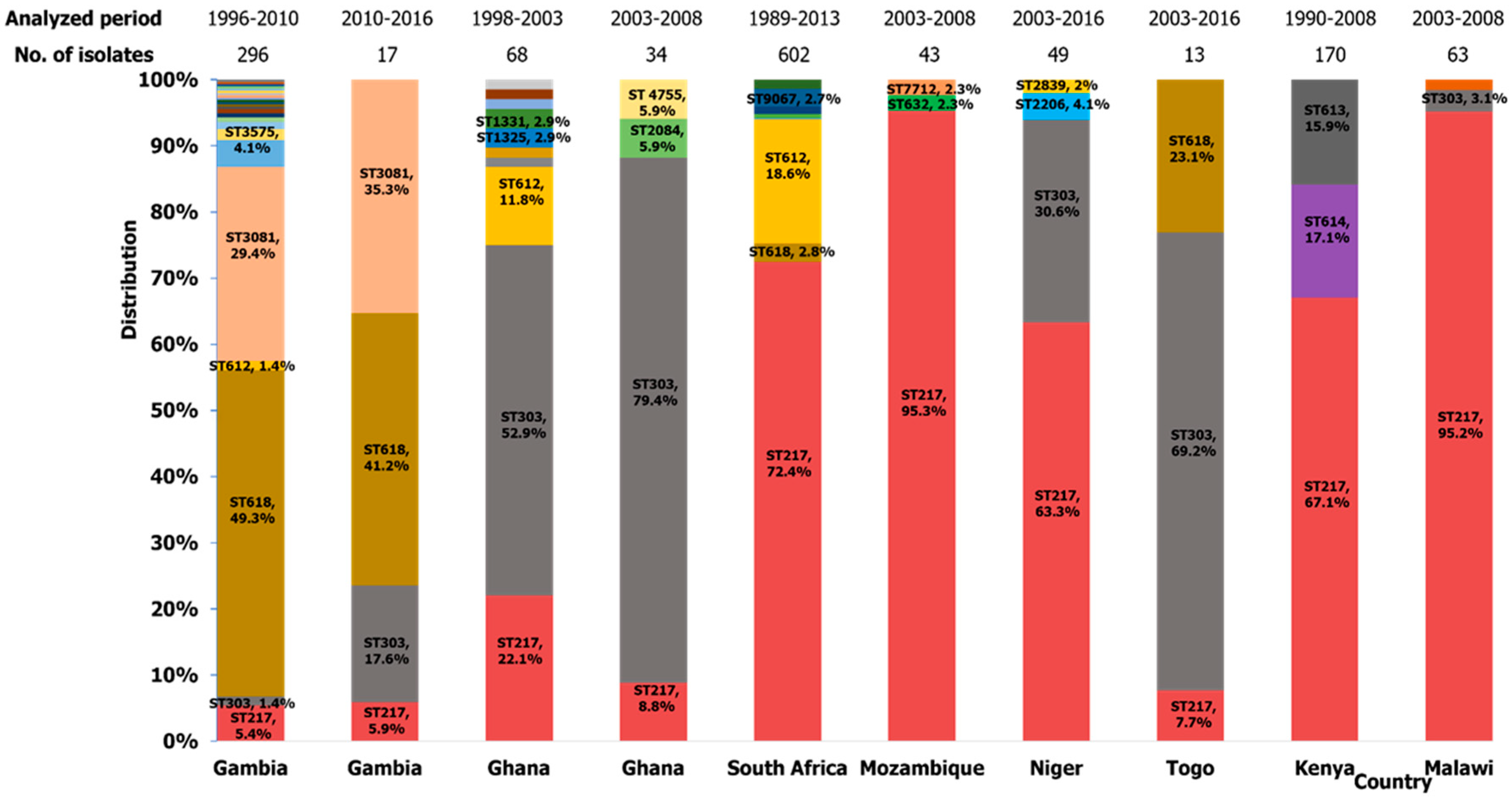
Figure 3.
Distribution of serotype 1 isolates identified in Europe.
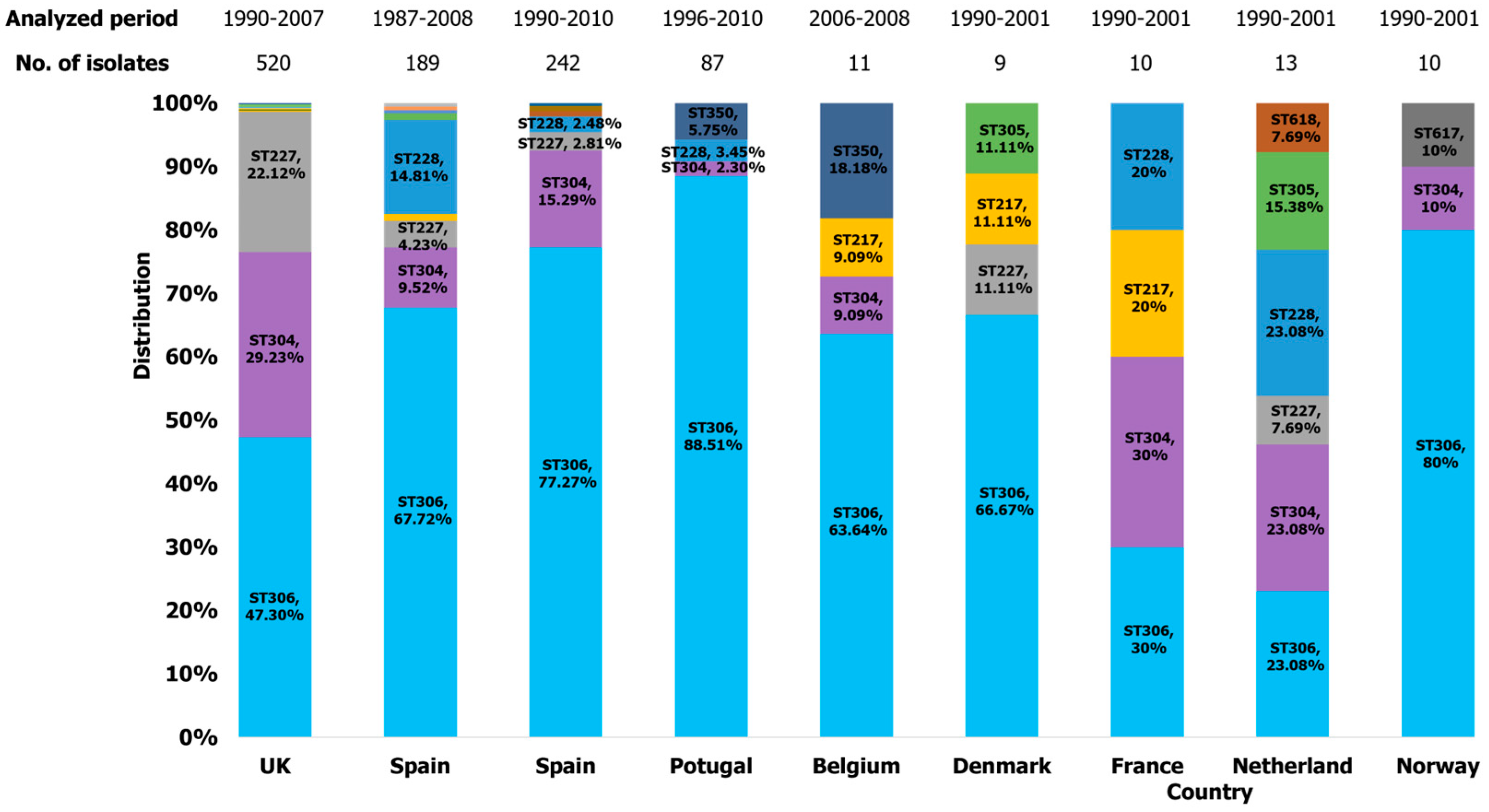
Figure 4.
Distribution of serotype 1 isolates identified in Asia.
Figure 5.
Distribution of serotype 1 isolates identified in the Americas.
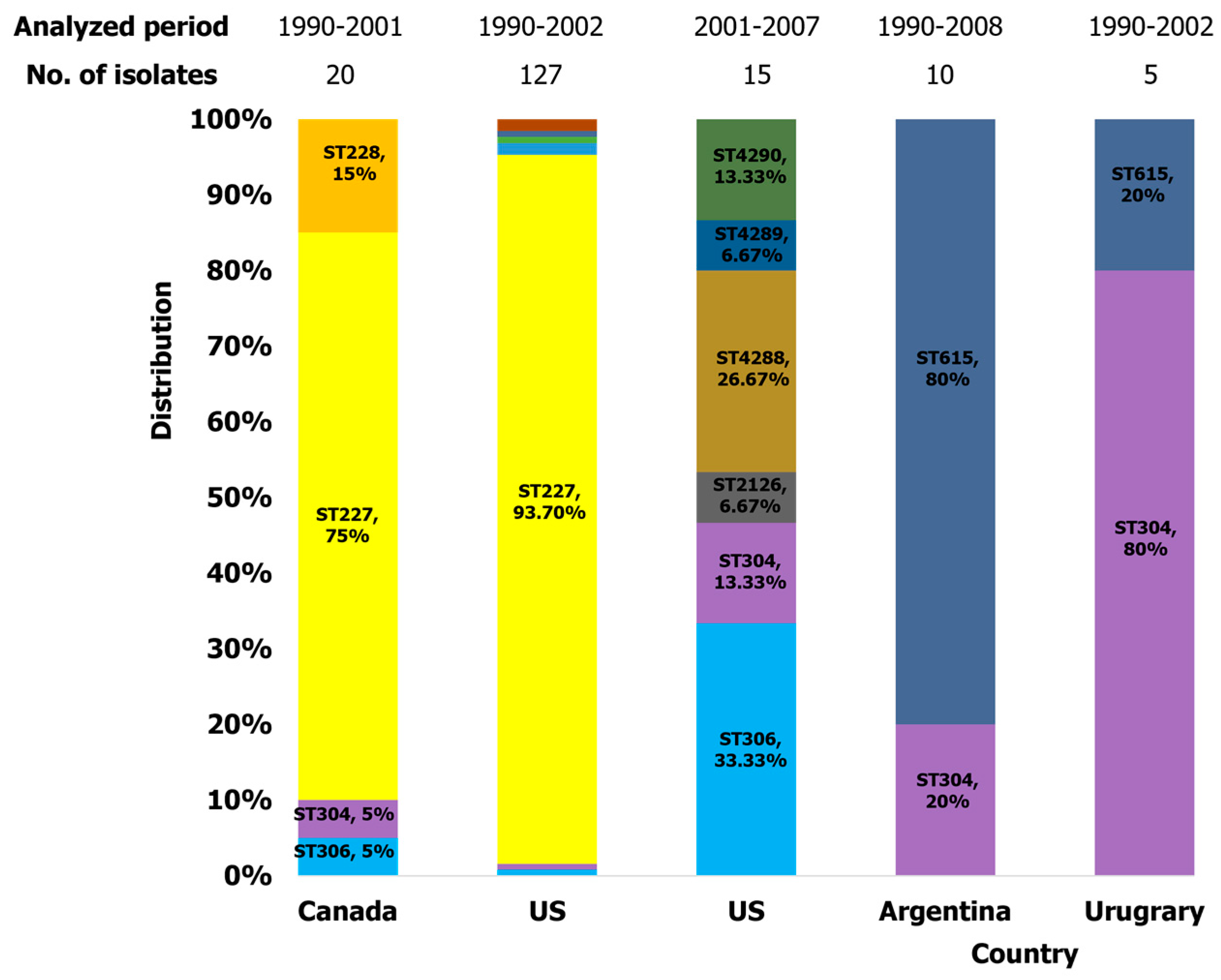
Figure 6.
Distribution of serotype 1 isolates identified in Oceania.
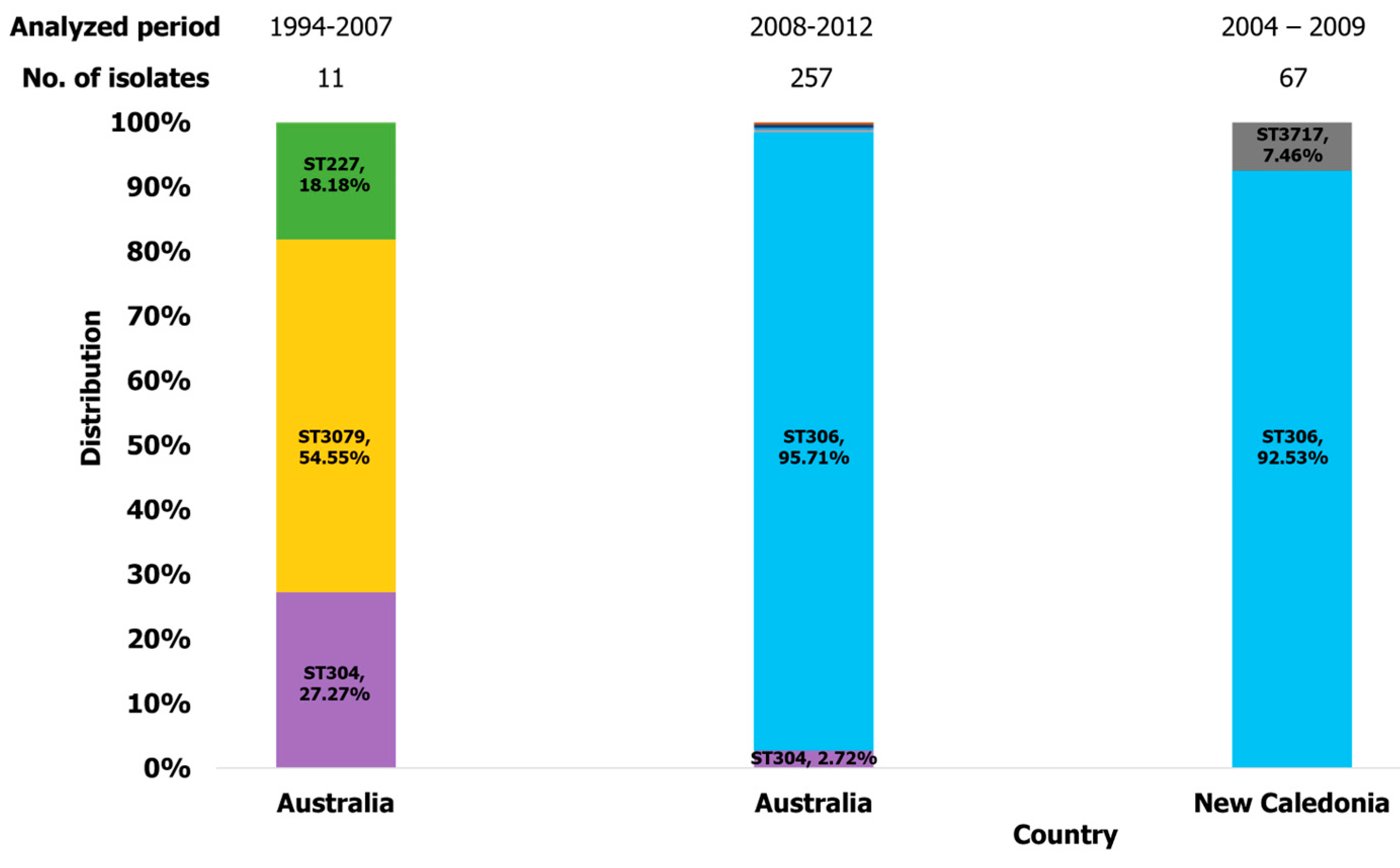
Figure 7.
The global circulation of dominant clones of S. pneumoniae serotype 1.
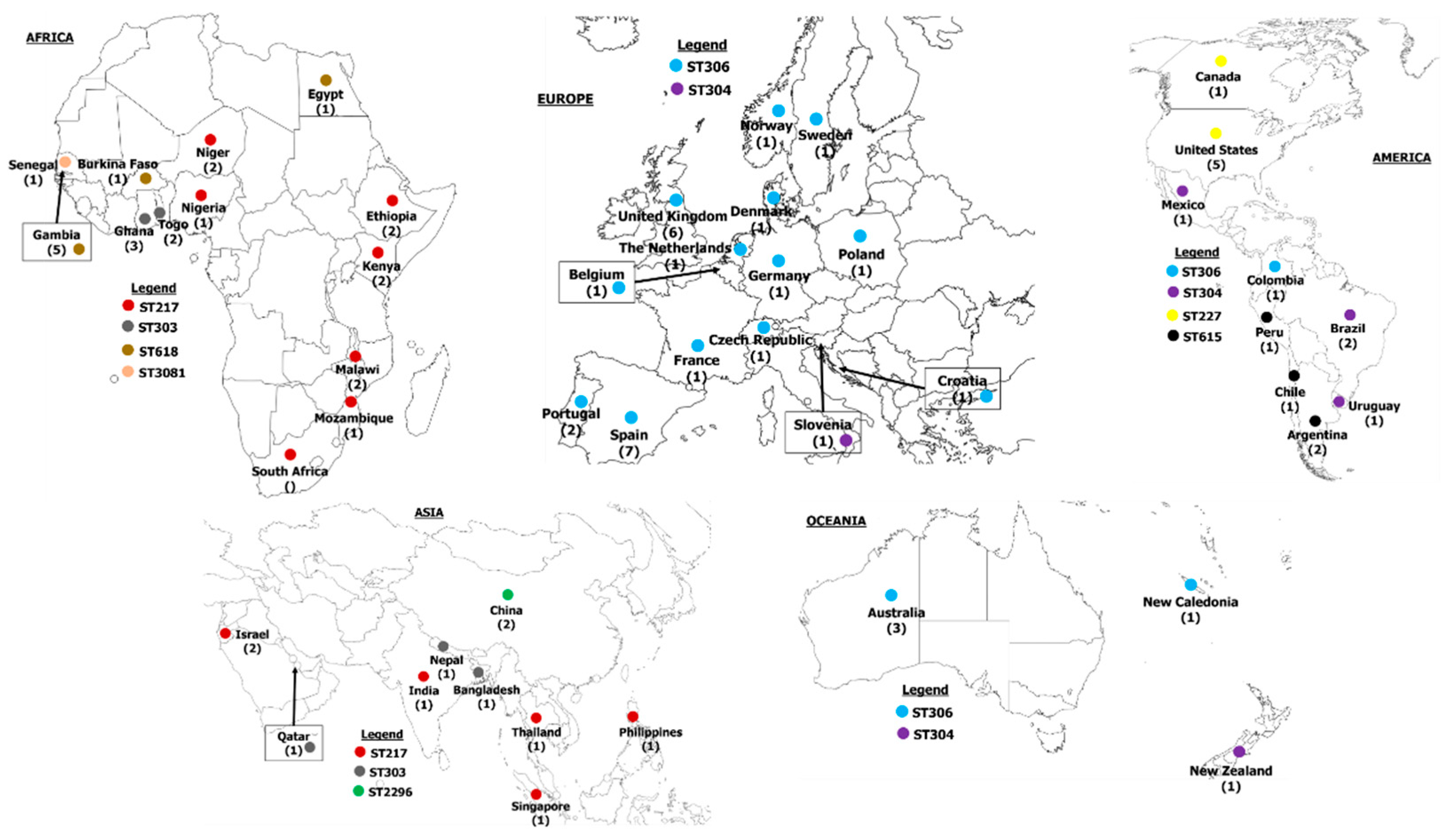
Figure 8.
Minimum spanning tree describing the relationships between the various clones identified in this review.
Figure 8.
Minimum spanning tree describing the relationships between the various clones identified in this review.
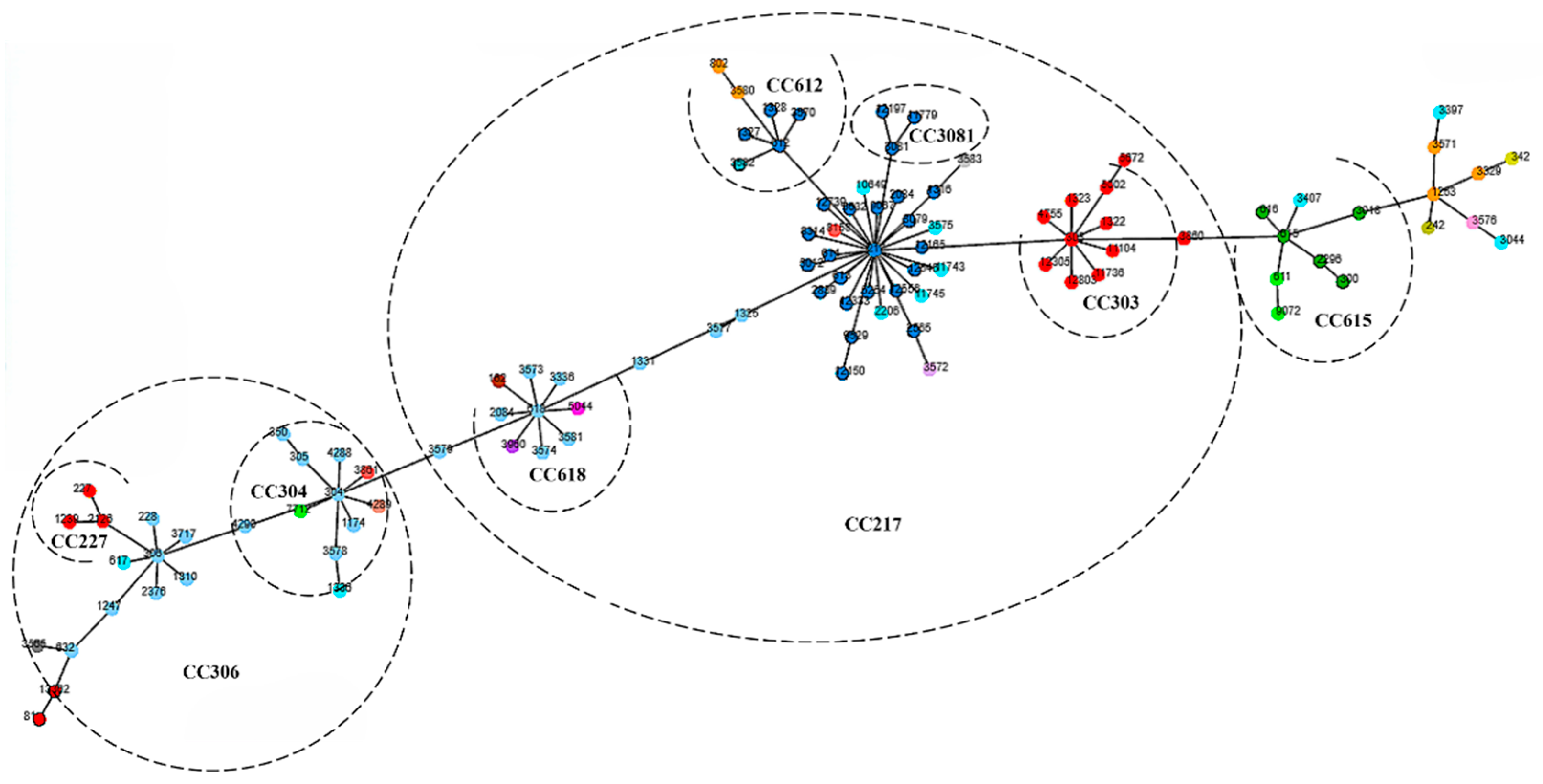

Table 2.
Characteristics of forty-three included studies.
| Author, year (ref) | Country | Year of study | Study population | Study group | Isolate type | No. of S. Pneumoniae serotype 1 | Molecular identification method | Genotyping tools | |
|---|---|---|---|---|---|---|---|---|---|
| MLST | WGS | ||||||||
| Byington et al., 2010 [19] | United States | 2001 - 2007 | Children with Pediatric pneumococcal empyema | Children | IPD | 17 | Unspecified | √ | |
| Chaguza et al., 2020 [20] | sub-Saharan Africa | 1996 - 2016 | Invasive pneumococcal serotype 1 isolates from sub-Saharan Africa | All ages | IPD | 909 | Whole genome sequencing | √ | √ |
| Hanachi et al., 2020 [21] | New Caledonia | 2004 - 2009 | Pneumococcal serotype 1 samples | Unspecified | IPD & NIPD | 67 | Whole genome sequencing | √ | √ |
| Almeida et al., 2013 [22] | Portugal | 1996 - 2010 | Healthy children attending day care | Children | Carriage | 21 | PCR | √ | |
| Esteva et al., 2011 [23] | Spain | 1989 - 2008 | Children with IPD | Children | IPD | 56 | Unspecified | √ | |
| Marimon et al., 2009 [24] | Spain | 1987 - 2007 | Pneumococcal serotype 1 samples | All ages | IPD & NIPD | 135 | Unspecified | √ | |
| Antonio et al., 2008 [25] | Gambia | 1996 - 2006 | Healthy Gambians and IPD cases | All ages | IPD & Carriage | 163 | PCR | √ | |
| Gonzalez et al., 2004 [26] | United States | 1993 - 2002 | Pediatric patients | Children | IPD & NIPD | 55 | Rep-PCR | √ | |
| Brueggemann & Spratt, 2003 [27] | Multicenter | 1990 - 2001 | Serotype 1 IPD isolates | All ages | IPD | 166 | PCR | √ | |
| Jourdian et al., 2013 [28] | Belgium | 2006 - 2008 | Nursery school children | Children, 3 - 6 years | Carriage | 11 | PCR | √ | |
| Horacia et al., 2016 [29] | Portugal | 2008 - 2011 | Adult patients with IPD | Adults, 18 years and above | IPD | 66 | Unspecified | √ | |
| Smith-Vaughan et al., 2009 [30] | Australia | 1992 - 2007 | Individuals in remote Indigenous communities in Australia | All ages | IPD & Carriage | 26 | PCR | √ | |
| Jauneikaite et al., 2014 [31] | Singapore | 2009 - 2010 | IPD cases | Unspecified | IPD | 4 | PCR | √ | |
| Kwambana-Adams et al., 2016 [32] | Ghana | Cases of suspected meningitis | All ages | IPD | 38 | Triplex qPCR | √ | ||
| Leimkugel et al., 2005 [33] | Ghana | 1998 - 2003 | Patients with suspected meningitis | All ages | IPD | 58 | PCR | √ | |
| Lai et al., 2013 [34] | Australia | 2010 - 2012 | participants presenting to or visiting the Alice Springs Hospital Emergency Department | All ages, 1 - 77 years | Carriage | 4 | multiplex PCR | √ | |
| Ebruke et al., 2015 [35] | Gambia | 2003 - 2004 | Healthy Gambians | All ages | Carriage | 81 | multiplex PCR | √ | |
| Kirkham et al., 2006 [36] | United Kingdom | 2000 - 2004 | Serotype 1 IPD isolates | All ages | IPD | 34 | PCR | √ | |
| Staples et al., 2014 [37] | Australia | 2008 - 2012 | Serotype 1 IPD isolates | Unspecified | IPD | 253 | PCR | √ | |
| Zhou et al., 2017 [38] | China | 2006 - 2012 | Patients in a hospital in Shanghai, China | Children, < 10 years | IPD & NIPD | 15 | PCR | √ | |
| du Plessis et al., 2016 [39] | South Africa | 1989 - 2013 | IPD cases | All ages | IPD | 534 | Whole genome sequencing | √ | √ |
| Brueggemann et al., 2013 [40] | Kenya | 1994 - 2008 | Healthy persons and ill children | Children, > 5 | IPD & Carriage | 161 | PCR | √ | |
| Donkor et al., 2013 [41] | West Africa | 1996 - 2007 | Pneumococcal carriage and IPD isolates | Children, < 15 years | IPD & Carriage | 7 | PCR | √ | |
| Zemlicková et al, 2005 [42] | Multicenter | 1990 - 2002 | Children with IPD | Children, < 5 years | IPD | 26 | PCR | √ | |
| Muñoz-Almagro et al., 2011 [43] | Spain | 2009 | Patients with IPD | All ages | IPD | 137 | multiplex PCR | √ | |
| Cooke et al., 2010 [44] | United Kingdom | 1999 - 2007 | IPD cases | Unspecified | IPD | 225 | Unspecified | √ | |
| Clarke et al., 2006 [45] | United Kingdom | 2000 - 2004 | Children with IPD | Children, < 5 years | IPD | 7 | Unspecified | √ | |
| Muñoz-Almagro et al., 2008 [46] | Spain | 1997 - 2006 | Patients with IPD | All ages | IPD | 34 | Unspecified | √ | |
| Beall et al., 2006 [47] | United States | 1999 - 2002 | IPD cases | Unspecified | IPD | 63 | Unspecified | √ | |
| Antonio, Dada-Adegbola, et al., 2008 [48] | Gambia | 2000 - 2004 | Children investigated for possible IPD | Children, 2 - 29 months | IPD | 8 | Box PCR | √ | |
| Yaro et al., 2006 [49] | Burkina Faso | 2002 - 2005 | Persons with suspected acute bacterial meningitis | All ages, 2 - 29 years | IPD | 21 | PCR | √ | |
| Zahner et al., 2010 [50] | United States | 1994 - 2006 | IPD cases | Unspecified | IPD | 5 | Unspecified | √ | |
| Grau et al., 2012 [51] | Spain | 1996 - 2010 | Patients with IPD | Adults, 18 - 64 years | IPD | 76 | PCR | √ | |
| Kourna Hama et al., 2019 [52] | Niger | 2010 - 2016 | Children with suspected meningitis | Children, < 5 years | IPD | 10 | qPCR, Whole genome sequencing | √ | |
| Jefferies et al., 2010 [53] | United Kingdom | 2001 - 2006 | IPD cases | All ages | IPD | 261 | PCR | √ | |
| Porat et al., 2012 [54] | Israel | 1999 - 2008 | Cases of AOM & IPD | Children | IPD & NIPD | 92 | Unspecified | √ | |
| Foster et al., 2008 [55] | United Kingdom | 1996 - 2005 | IPD cases | All ages | IPD | 203 | Unspecified | √ | |
| Chaguza et al., 2017 [56] | Malawi | 2004 - 2010 | IPD cases | All ages | IPD | 113 | PCR, whole genome sequencing | √ | √ |
| Sanneh et al., 2019 [57] | Gambia | 2010 - 2016 | Suspected cases of meningitis | Children, < 5 years | IPD | 17 | qPCR, multiplex PCR, Whole genome sequencing | √ | |
| Tsolenyanu et al., 2019 [58] | Togo | 2010 - 2016 | Suspected cases of meningitis | Children, < 5 years | IPD | 7 | RT-PCR, whole genome sequencing | √ | |
| Sharew et al., 2024 [59] | Ethiopia | 2018 - 2019 | Patients with IPD and NIPD | All ages | IPD & NIPD | 1 | Whole genome sequencing | √ | √ |
| Serrano et al., 2005 [60] | Portugal | 1999 - 2002 | IPD cases | Children | IPD | 470 | Unspecified | √ | |
| Cornick et al., 2015 [61] | Multicenter | 2003 - 2008 | serotype 1 pneumococci recovered from hospital, surveillance and carriage studies | All ages | IPD, NIPD & Carriage | 448 | Whole genome sequencing | √ | √ |
Table 1.
Detailed search keywords and filters used in each database.
| Database | Keywords | Filters | ||
|---|---|---|---|---|
| Language | Document Type | Open access | ||
| PubMed | “Streptococcus pneumoniae serotype 1” OR “serotype 1 streptococcus pneumoniae” OR “pneumococcal serotype 1” | English | - | Free full-text |
| Scopus | English | Article | All Open Access | |
| Web of Science | English | Article | Open Access | |
| ScienceDirect | English | Research article | Open Access & Open archives | |
| Google Scholar | English | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



