Submitted:
11 December 2024
Posted:
11 December 2024
You are already at the latest version
Abstract
The analyzed data set on the problem of geophagy-lithophagy shows that the instinctive desire, preserved in many groups of animals and humans, to eat mineral-crystalline substances - products of hypergenic transformation of various rocks in the most general form, is a manifestation of evolutionarily conditioned universal way of regulating the material composition of the internal environment, as well as many biological and physiological processes in the body. The effect of minerals on the organism can be divided into their main and secondary functions. The main cause of geophagy-lithophagy in animals and humans all over the world is the regulation of concentration and ratio of rare earth elements in the neuroimmunoendocrine system, which controls metabolic processes and provides immune defense of the body. More than ten secondary causes of consumption of rare earth elements have been identified.
Keywords:
geophagy
; lithophagy
; rare earth elements
; animals
; humans
In this review, we have attempted to summarize the data that I and a team of my associates have collected on the causes of the earth substances consumption phenomenon that we have identified in humans and various animal groups.
About the Terminology
Before dealing with the terminology, we should emphasize that there are two forms of consumption of earthy substances: accidental and instinctive, both occurring in humans and animals. In this article, only the instinctive forms will be discussed.
There are several terms used in the scientific literature to describe the phenomenon of human and animal consumption of earthy substances. Two of the most commonly used terms are geophagy (from Greek, literally "earth eating") and lithophagy (from Greek, "stone eating"). In the English-speaking world, the terms "soil consumption" and "pica" (from Latin "pica" – a bird that picks everything) are also sometimes used to refer to distorted food cravings, including mineral consumption. Such cravings usually occur either in pregnant women, which can be considered an instinctive form of geophagy, or in both sexes in the context of psychotic disorders or mental retardation. In relation to domestic and farm animals, another term close in meaning to "pica" is used – "allotrophagia" (literally meaning "food perversions" in Greek).
In this article, we will use two of the most popular terms in science: "geophagy" and "lithophagy," with the meanings accepted in English-language scientific texts. We will use the term geophagy when we talk about the consumption by humans and mammals of loosened mineral, organomineral, or predominantly organic substances of soil and rock weathering crust derivatives. When we speak of lithophagy, we mean only cases of ingestion of stone material ("pebbles") and only by animals that have a muscular stomach. In this case, the ingestion of "pebbles" of approximately 1 to 10 mm in size is characteristic mainly of birds and small reptiles, and stones of more significant size are characteristic of large reptiles, including extinct species. Unlike mammals, which ingest earthy substances only in certain areas and at certain times of the annual life cycle, in birds and their closest relatives reptiles "pebbles" are almost always present in the digestive tract, especially their many accumulate in the muscular stomach, from which they received the appropriate name – "gastroliths" (literally from Greek "stomach stones").
Areas where hoofed animals and other large mammals regularly consume earthy substances usually form landscape complexes easily recognizable by their specific appearance, which in Russian popular and sometimes scientific literature are called "animal solonetz", and in English literature "salt lick" or "mineral lick", which are close in meaning. After we were repeatedly convinced that these objects are far from being directly related to soluble salts, as well as to licks, and besides, the term "solonetz" with a completely different meaning has existed in soil science in Russia for a long time, we proposed and have long been using the term "kudur" (from the Turkic "kudyur" – a place where animals eat soil), borrowing it from the lexicon of Turkic shepherds (Panichev et al., 2013). We use the derived term "kudurit" to refer to all types of earthy substances consumed by animals at kudurs.
When we talk about human geophagy, we only mean cases with a long, often millennia-long history of mass consumption of earthy substances by humans in certain areas of the world. As a rule, mass geophagy among animals at kudurs is also common in the same areas. Geophagy among humans can also occur among natives of the areas where geophagy was originally widespread who have moved to new locations. In contrast to kudurits, lithogenic substances eaten by humans will be called "edible earths". We would like to emphasize that when we talk about geophagy in relation to people, we are not talking about all cases of internal use of mineral substances by people. Some minerals can be widely used in traditional healing practices, and their list, unlike "edible earths," is quite long.
Geophagy
Humans
The study of geophagy in humans began almost 200 years ago, not by natural scientists, but by physicians who encountered the phenomenon against the background of an unusual form of dystrophy in black Africans on the American continent, called "Cachexia Africana" (Mason, 1833). Later, F.W. Cragin (1836) concluded that the tendency to eat earth was a consequence rather than a cause of the disease. A similar symptomatic disease called Loeffler endocarditis and endomyocardial fibrosis (EMF) was described in Africa only in 1946 (Bukhman, 2008). Its localization is most characteristic of the tropical forest zone with predominantly rural populations. Importantly, in Uganda, the disease is more prevalent among immigrants from neighboring Rwanda and Burundi, who settle only in certain areas. Other regions of the world with high prevalence include some South American countries (Brazil and Colombia), with many cases reported in southern China (Guangxi Province) (Bukhman, 2008). All of these areas of EMF prevalence, as will be shown later, are areas where geophagy is also prevalent, both in humans and animals.
A really systematic collection of data on geophagy among people began after ethnographic studies were carried out all over the world at the turn of the XIX-XX centuries (Ross, 1895; Hooper, Mann, 1906; Laufer, 1930; Sternberg, 1933). The most thorough summaries of the collected data on the geophagic traditions of people in different regions of the world were the books by Laufer (1930) and Аnell and Lagercrantz (1958). As a result of archaeologists’ excavations of ancient human burials in Africa and South America, it was revealed that the historical roots of geophagy traditions are many thousands of years old (Vermeer, 1984; Browman, Gundersen, 1993; Rowland, 2002).
The general current state of the study of geophagy in humans is relatively well reflected in a review article by English researchers from the University of Wales, written on the basis of works dealing with various aspects of the phenomenon in question (Abrahams, Parsons, 1996). This review shows that until the mid-1990s, science had made little progress in studying this issue. According to the analysis of the available data, the main purpose of the consumption of earthy substances was and still is related either to the needs of pregnant women and children for some mineral substances, or to the needs of people of different sexes and ages for the treatment of specific diseases characteristic of certain areas where people live. There are also described cases when the consumption of earth was related to hunger, as well as during magical rituals. It is important to note that the traditions of earth eating in Africa continue to this day. Modern clay eaters obtain the necessary product either by buying it in markets or by collecting it themselves from a variety of sources, which in some places are quite numerous.
As mineralogical studies of the "edible earth" used by humans on all continents have shown (Gebel, 1862; Vermeer et al., 1985; Ferrell et al., 1985; Browman, Gundersen, 1993; Young et al., 2010; Mahaney et al., 1990, 1997, 1999, 2000; Wilson, 2993; Ngole, Ekosse, 2012; Duplex et al., 2018) their composition is dominated by clay minerals from the kaolinite and smectite groups, less frequently from the hydrous mica group (illite and vermiculite). Chlorites, sulfates, iron oxides (hematite and goethite), and aluminum hydroxides (gibbsite) occur as trace elements. A significant amount of carbonates is observed in some "edible earths". Quartz and feldspars are always present, sometimes dominant in the composition of the primary phases (magmatic crystalline) of "edible earths". Any geologist will immediately notice that this set of minerals includes the most common species typical of the weathering crusts of various rocks.
In the works on the medical aspects of human geophagy, in order to better explain the causes of the phenomenon under study, attempts have been made to link the iron and vitamin deficiencies that develop in the organism to the consumption of clay (Kurtlis et al., 1963; Mengel et al., 1964), but without success. Researchers who have studied geophagy in humans have shown that the consumption of clays removed iron from the body, which could even lead to iron deficiency anemia (Minnich et al., 1968; Nchito et al., 2004; Young et al., 2011; Ngole, Ekosse, 2012). Similar results have been obtained in rat experiments (Edwards et al., 1983; Dreyer et al., 2004; Hooda et al., 2004; Seim et al., 2013). Attempts to attribute the causes of iron deficiency to deficiencies of elements such as zinc, copper, and calcium in addition to iron have also been unconvincing (Cheek et al., 1981; Feildman, 1986; Abrahams, 1997). A case study of a woman has also been described showing that excessive use of clays can lead to potassium deficiency (Severans et al., 1988). Other adverse effects associated with geophagy have been identified, including the association of worm infestations in people who have eaten contaminated soil (Robinson et al., 1990; Geissler et al., 1998; Hinz, 1999; Rowland, 2002). There have also been articles linking geophagy to the consumption of poisonous plants. In this case, "edible earths" were considered as probable detoxicants of the organism (Johns, 1986; Johns, Duquette, 1991; Reid, 1992).
A very informative example of a comprehensive study of "edible earths" used by people in Africa is provided by Young et al. (2010). The researchers examined the types of earths consumed by people on the islands of Pemba and Zanzibar in southeastern Africa. Laboratory studies showed that the pH of the water extract in samples of "edible earths" could vary widely, from 5 to 10.5. The results of particle size distribution, chemical and mineralogical analyses showed that "edible earths" can be both predominantly siliceous mineral complexes with a low (sometimes not more than 3%) content of clay minerals with mostly sandy-silty dimension of quartz and feldspar grains, and in the form of almost purely clayey-silt deposits with predominance of kaolinite and halloysite, sometimes enriched with iron oxides. The origin of all these sediments is connected with the processes of deep weathering of various rocks with their subsequent transfer by water in the form of muddy suspensions with deposition in the mouth part of a relatively large valley river. The content of trace elements in the "edible earths" fluctuates at the level of the minimum values for the soils of the investigated area. In general, such contents are common for highly weathered residual weathering crusts, which are widespread in Africa.
Similar results in terms of physicochemical properties, mineralogy, and geochemistry have also been demonstrated in studies of "edible earths" in mainland South Africa, particularly in the Eastern Cape Province (Ngole, Ekosse, 2012).
S.L. Young et al. (2010) hypothesize that the reason for the consumption of such earthy substances is the therapeutic effect of kaolinite and halloysite, which is supposed to be consistent with recent data on the use of these minerals as pharmaceuticals to remove toxic substances from the body and to protect the mucous membranes of the gastrointestinal tract. However, the consumption of earthy substances containing less than 3% of clay and the rest of quartz-feldspar sands clearly refutes this assumption. Rather, the revealed facts mean that the reasons why people eat these mineral complexes are simply unknown. However, despite the differences in mineral and chemical composition, it is obvious that all of them have the same property sought by their consumers.
Unfortunately, most researchers who have studied "edible earths" have not paid attention to rare earth elements (REE). Why this is important will become clear in the text below. For now, let us review some of the published data on the geochemistry of "edible earths" that includes the results of REE analysis.
The first paper (Mahany et al., 2000) covers the study of 5 soil samples, including 3 eaten by humans and 2 not eaten for comparison, collected on Java Island (Indonesia). Their mineral and chemical compositions were examined. All samples were found to be highly altered volcanics represented by hydrated halloysite and kaolinite with admixture of smectites. In terms of trace element content, only slightly higher concentrations of Co and Cr were found in the edible varieties. The authors concluded that people ate these minerals because of the high content of clay minerals, which help to sorb toxic plant substances in the digestive tract. Meanwhile, according to the data on the composition of REE in the studied soils, some of them show unusually high levels of several REEs (La up to 172 g/t, Ce up to 336 g/t, and Nd up to 128 g/t).
The second paper (Ekosse et al., 2011) examines samples of soils consumed by humans in an area of the Congo in central Africa. Whitish, light gray, and sometimes brown "edible earths" belonged to clay, clay-silt, and loamy types. X-ray structure of the samples revealed kaolinite and smectite as trace elements, along with quartz, microcline, muscovite, talc, goethite, hematite, siderite, gibbsite, anatase. In the rock-forming oxides, the contents of Na and Ca were very low, Al was abundant, and sometimes Fe was abundant. Among trace elements, many samples also had elevated REE concentrations (La up to 100 g/t, Ce up to 162 g/t, and Nd up to 125 g/t).
Having drawn the reader’s attention to the unusually high levels of REE in some clays consumed by humans, let us continue the consideration of geophagy in animals and simultaneously in humans.
Humans and Animals
One of the first researchers to suggest that the main cause of geophagy in animals and humans might be the same was the Russian geologist P.L. Dravert (1922). P.L. Dravert’s reflections on geophagy are interesting because they were written before the first scientific publications on geophagy in animals, which began to appear only in the 1930s (Mure, 1936; Nasimovich, 1938), and the territorial connection of consumers of earthy substances among humans and at the same time among animals, according to the publications, became clearly visible only in the second half of the XX century, and only in the countries of the equatorial zone.
The analysis of the mineral composition of kudurits, which is presented in detail on the basis of our own research and review of data published in books (Panichev, 1990; 2011), shows that it is similar to the composition of the earth eaten by humans. It is usually the same clay minerals from the kaolinite, smectite, and hydrous mica groups, in some cases with a significant amount of zeolites and carbonates. The primary crystals are the same quartz and feldspars. Among the trace elements, iron and manganese oxides and hydroxides are common, with occasional significant content (more than 1%) of chloride salts or sodium hydrogen carbonates.
Since the first studies of the phenomenon of geophagy in animals, many experts from different disciplines have tried to determine the cause that makes animals regularly ingest earthy substances. However, only a series of hypotheses have been proposed so far, both for animal and for human geophagy. The best known among them are the "sodium" hypotheses: the need for sodium during the periods of lowest sodium intake (Mure, 1936; Stockstad et al. 1953; Staaland, 1980), as well as the need for sodium and mineral sorbents necessary to normalize the electrolyte balance in the digestive tract during periods of seasonal changes in the diet (Kreulen, 1985; Panichev, 1987; Panichev, 1990; Klaus, Schmid, 1998). The second group of hypotheses is about the replenishment of deficiencies of a number of chemical elements in the body (Nasimovich, 1938; Klaus et al.1998; Jones et al. 2000; Mincher et al. 2008; José et al. 2017). The third group is about the use of bacteriostatic properties of clay minerals in the control of pathogenic microflora against diarrhea and intestinal parasites, replenishment of symbiont microorganisms and regulation of pH in the digestive tract (Robertson, 1996; Knezevich, 1998; Abrahams, 1999; Ketch et al. 2001; Wilson, 2003; Banenzoue et al. 2014; Lebedeva et al. 2020). The fourth group is based on the elimination of toxic chemical elements and compounds, including organic ones such as phenols, terpenes, tannins, alkaloids and cyanogenic glycosides from the body with the help of mineral sorbents (Johns, Duquette, 1991; Gilardi et al. 1999; Houston et al. 2001; Dominy et al. 2004; Ekosse et al. 2020). Based on experimental studies, an explanation for geophagy has been proposed as the desire of animals to use clay minerals to influence nutrient assimilation and to protect the intestinal mucosa (Reichardt, 2008). It has even been suggested that geophagy is not related to a physiological need, but to a sense of gustatory pleasure (Krishnamani, Mahaney, 2000; Hladik, Gueguen, 1974).
One of the relatively recent attempts to unify hypotheses on the causes of geophagy in humans and animals is the work of Young et al. (2011), based on the analysis of 482 publications on human geophagy and 330 on animal geophagy. As a main result, it was proposed to explain human geophagy in terms of protection from harmful and toxic substances, parasites and pathogens, while animal geophagy, in addition to such protection, may also be aimed at the absorption of a number of chemical elements, primarily sodium.
In 2015, after analyzing the published data on the geochemistry of earthy substances eaten by animals in different regions of the world (our own and other researchers’ data), as well as taking into account the emerging data on the effects of REE in the body on the nervous, immune, and hormonal spheres, most fully shown in the review by K. Redling (2006), we had the idea that one of the main reasons for the desire for geophagy could be due to the disturbance of REE metabolism in the body (Panichev, 2015; 2016).
In the period from 2020 to 2023, based on the results of special landscape geochemical studies conducted with the financial support of the Russian Science Foundation, we have convincingly demonstrated the relationship between geophagy and REE on the material of studies in a number of regions of southern Siberia and the Far East of Russia (Panichev et al. 2021; 2023a; 2023b).
A reliable connection between mass cases of geophagy and anomalous REE contents in landscape components was first revealed in the territory of the Southern Sikhote-Alin, in the area of the Vanchin graben (the area of the upper Milogradovka River). In this area of about 50 km2 we found 14 places where wild animals (mainly red deer) regularly eat kaolinite and smectite clays in large quantities (kilograms per visit), sometimes with admixture of fine zeolites. As a result of detailed landscape-geochemical studies conducted in the area, we were able to detect anomalously high REE contents, often with a predominance of heavy REE subgroup elements (HREE), in various types of Early Cenozoic volcanogenic rocks, soils, surface waters, vegetation, and even in brain tissues of local reindeer (Panichev et al. 2021). Comparison of the composition of ingested clays and animal feces containing clays showed that clays in the digestive tract actively sorb REE, especially HREE, and remove them from the body. Based on the data obtained, it was concluded that the desire of reindeer to consume clays may be caused by disturbances of REE metabolism in the body due to their high content in feed and drinking water.
We were able to substantiate this conclusion after analyzing the published data on the biological properties of REE. The accumulated experience in the study of the biological properties of REE presented in the literature (Powis at al., 1994; Palasz, 2000; He, Xia, 2001; Jiesheng at al., 2002; Feng at al., 2006; Redling, 2006; Zhao at al., 2012; Panichev, 2015; 2016; Brouziotis et al., 2022), as well as the experience of using this group of elements in animal feeding (Wang et al., 2003; Rambeck et al., 2004; Xun et al., 2014; Tariq et al., 2020), indicate that among these elements, the biologically significant ones are predominantly representatives of the light subgroup (LREE), including: Sc, La, Ce, Pm, and Nd. It seems that among the representatives of HREE only yttrium (Y) performs biological functions in the body. However, all REE without exception can be integrated into biological structures and replace each other, but at the same time almost all HREE except Y are unable to perform the functions required by the body, unlike LREE. Judging by the distribution of REE in the tissues of mammals, critical disorders in the animal bodies may occur in the nervous, immune, and endocrine systems. Thus, we have come to the conclusion that excessive intake of REE into the internal environment of the body may disturb the neuroimmunoendocrine system, which controls all metabolic processes in the body, stimulating animals to consume mineral sorbents in order to regulate the composition and ratio of REE in it. From the point of view of such ideas, the regular consumption of clays is an inevitable necessity for the survival of animals in landscapes with an excess of REE, especially with an excess of HREE. In other words, in this case we are dealing with hypermicroelementosis according to the understanding of A.P. Avtsyn et al. (1991), in which 17 trace elements of the REE group act as the main agent.
Studies in other areas of the Sikhote-Alin (Panichev, 1990; Panichev et al. 2021), as well as in the Altai Mountains (Panichev et al. 2023a), have shown that a wide variety of sorbents are suitable for survival in such landscapes. These may include not only clays of various mineral compositions, but also zeolitic and opal "earths," peat soils, and other varieties of mineral and organomineral sorbents based on diatomites, silica clays, or rotten-stone. According to published data, both animals (Struhsaker et al. 1997) and humans (Chung et al. 2019) can also use purely "carbon-based" sorbents in the form of charcoal. We have recently found that animals can even consume bituminous coal, selecting only REE-enriched varieties (Panichev et al. 2024). Why they do this will become clear in the text below.
In 2022, we conducted detailed landscape geochemical studies on Olkhon Island in Lake Baikal (Panichev et al. 2023b), where the geological and geochemical situation is quite different from that in the Sikhote-Alin and Altai Mountains. Kaolinite clays eaten by wild reindeer and livestock on Olkhon turned out to be significantly enriched in LREE against the background of anomalously low concentrations of this group of elements in all components of the island landscapes. Comparison of the composition of clayey feces of reindeer and the clays eaten by them showed that a part of LREE consumed with clays remains in the body. As a result, it was concluded that the consumption of clays by animals on Olkhon is also caused by disturbances in the composition and concentration of REE in the organism, but against the background of abnormally low content of these elements in feed and drinking water. Thus, in this case we encountered manifestations of typical REE hypomicroelementosis in reindeer. Such endemic metabolic disorders in the bodies of wild and domestic animals should be most common when they live in areas where sedimentary chemogenic and organogenic carbonate rocks predominate, which are known to be characterized by very low REE concentrations (Dubinin, 2006). Meanwhile, REE hypermicroelementoses will be common in the areas of development of magmatic carbonate rocks (carbonatites) characterized by unusually high REE concentrations. These situations are observed, for example, in the territory of the same East African rift system, where REE-enriched rocks such as carbonatites and alkaline basites are known to be widespread. Areas with mass manifestations of geophagy in both animals (Mahaney and Hancock, 1990; Mahaney et al. 1990, 1997) and humans (Annel and Lagerkranz, 1958; Abrahams and Parsons, 1996; Knudsen, 2002) are found there. However, our more detailed study of the landscape-geochemical conditions in this region based on the published data (Panichev et al. 2023b) indicates the possibility of a high contrast in REE distribution in landscapes, caused not only by the contrast in chemical composition of the rocks, but also by the contrast in terrain and climatic conditions.
It should be noted that in the equatorial part of Africa there is almost everywhere a high contrast of REE concentrations in rocks and soils. According to Temga et al. (2021), the total REE content in soils of Cameroon (Central Africa) varies from 0.482 to 5926 mg/kg (5 orders of magnitude difference) with the average value of 297 mg/kg, which is currently the highest among soils in the world.
After the work on Olkhon Island it became clear to us that all areas where geophagy is widespread in mass form are to some extent REE-anomalous, with increased or decreased content of mobile REE forms in landscape components compared to local background values. Some animals living in such conditions, mainly herbivores, to a lesser extent omnivores and carnivores, may suffer from disturbances in the metabolism of this group of elements in the neuroimmunoendocrine system of the body. The hormonal stress that develops in this case most likely triggers the instinctive mechanism of searching for sorbents that, depending on the geochemical conditions of the habitat, are preferred by the animals either in the REE-enriched or in the REE-depleted form. The universal character of the revealed regularities is indicated by our analysis of geological and geochemical situations in a number of regions of the equatorial zone of the Earth, which shows that the connection of mass cases of geophagy with REE-anomalous landscapes in this part of the world is more pronounced than in the middle latitudes, and with a vivid manifestation of REE-endemic diseases in humans.
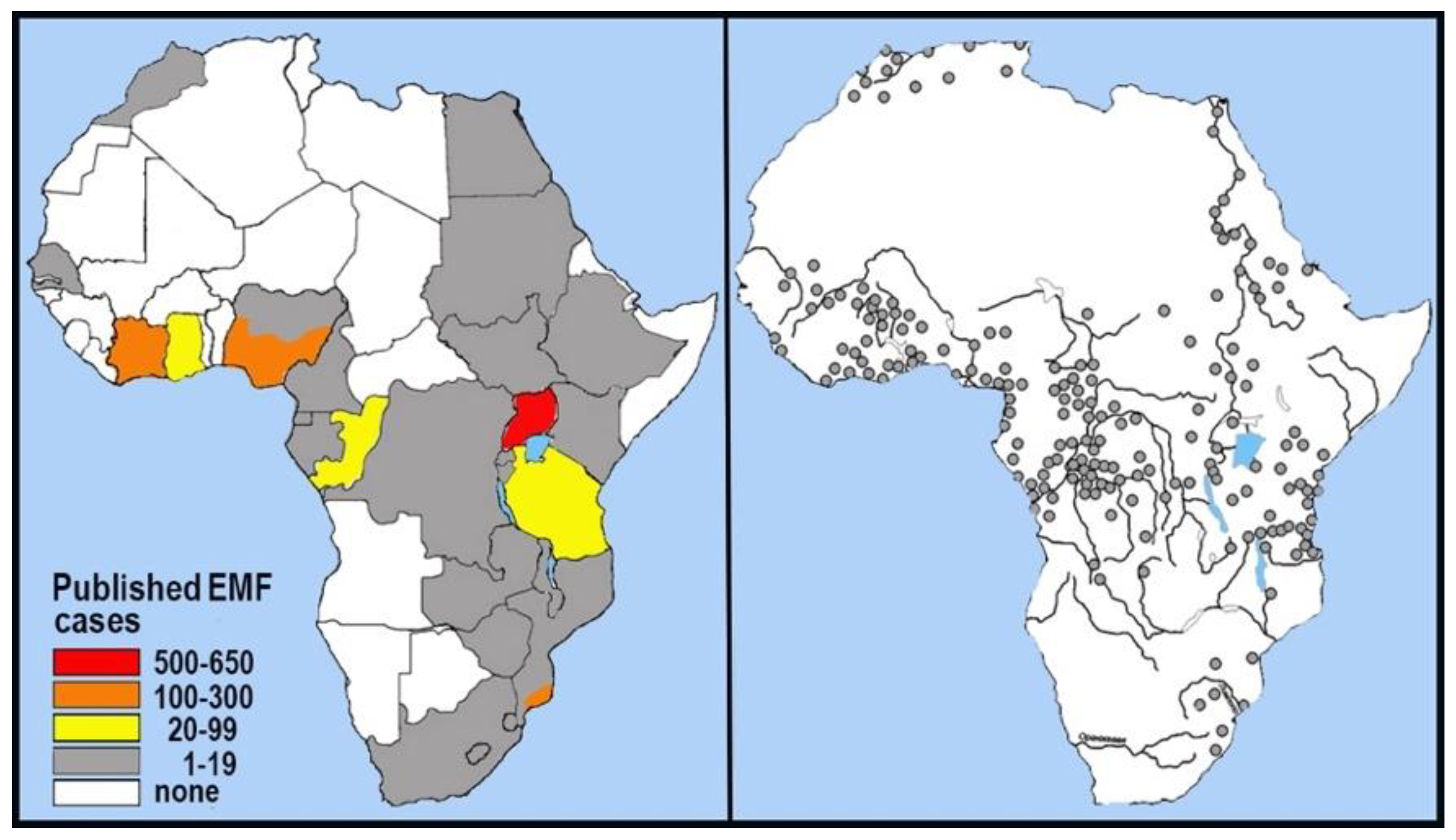
One such disease is the aforementioned Loeffler endomyocardial fibrosis, which has been shown to be directly associated with excess cerium (Ce) in the heart muscle tissue of diseased individuals (Valiathanet al., 1986), as well as with excess Ce in the plant diet of affected individuals (Eapen, 1998) and in monazite-enriched soils in EMF-endemic areas of India (Kutty et al. 1996). A similar pattern of disease association with REE-enriched soils has been found in Africa (Smith et al., 1998). Our comparison of the areas where EMF diseases are prevalent in Africa according to Bukhman et al. (2008) with the areas where geophagy is historically prevalent among humans according to (Annel, Lagerkranz, 1958) showed good convergence (Figure 1).
In the southern EMF-affected states of India, as well as in Africa, there is an association with areas of animal geophagy. The presence of typical kudurs in Kerala, frequently visited by local ungulates, has been described in particular in the Chinnar Nature Park (Ramachandran et al. 1995), as well as in the Marakkanam Reserved Forest in the neighboring state of Tamil Nadu (Voros et al., 2001). Chinnar Park is located on a mountain plateau with elevations up to 2500 m, composed of metamorphic rocks of Precambrian age, predominantly crystalline schists and gneisses, including charnockite series, enriched in REE, mostly concentrated in monazite (Anitha et al., 2020). From this plateau, rivers flow along the banks and at the mouths of which human biogeochemical endemics have been identified, associated with excess REE in monazite-bearing sands (Kutty et al. 1996). Similar rocks are common not only in the south, but also in the eastern parts of India, i.e. exactly where, according to B. Laufer (1930), geophagy in humans was already widespread at the beginning of the 20th century.
Figure 1.
Left – EMF incidence in African countries (Bukhman et al. 2008); right – human geophagy prevalence in Africa (Annel, Lagerkranz, 1958).
Figure 1.
Left – EMF incidence in African countries (Bukhman et al. 2008); right – human geophagy prevalence in Africa (Annel, Lagerkranz, 1958).
Previously, referring to the publication of Bukhman et al. (2008), we noted numerous cases of EMF illness in southern China (especially in Guangxi Province). According to Aufreiter et al. (1997), geophagy was common in humans in the neighboring province of Hunan not so long ago. According to Laufer (1930), geophagy was much more widespread in China in the past. In southeastern China, geophagy is also found in animals (Li et al., 2018). At the same time, REE-enriched carbonatite volcanic complexes are common in central and southeastern China, and the world’s largest ion-adsorption-type REE deposit in clayey weathering crusts is located there. That the REE content in soil and products grown on it in China affects human health and longevity is indicated by the REE content in the hair of long-lived Chinese people (Zhang et al., 2020), which is on average 100 times lower than in the hair of people living in areas with high background REE content (Meryem et al., 2016).
It should be noted that the association of endemic diseases with landscape REE anomalies is not limited to EMF. According to Zhu et al. (2005), people living in Jangxi Province in southern China in areas with high background REE levels suffer from gastric disorders and anorexia, with markedly reduced levels of total protein, globulins, albumin, and serum glutamic pyruvic transaminase with higher levels of total immunoglobulin. A paper by Chinese researchers (Liu et al., 2019) also shows that exposure of pregnant women to Ce and Yb leads to lower levels of thyrotropic hormone in infants (this hormone is secreted in the thyroid and epiphysis and largely regulates the body’s growth and development). Henríquez-Hernández et al. (2017) demonstrated that in the inhabitants of sub-Saharan Africa, elevated levels of REE in the blood are somehow related to the development of anemia, since the occurrence of this pathology in them often does not depend on the level of iron in blood. An article by Collignon (1992), citing some researchers, discusses the possibility of a link between geophagy and a blood disorder known as "sickle cell anemia". According to Gómez-Aracena et al. (2006), Ce levels in toenails are associated with an increased risk of acute myocardial infarction. It has also been shown (Zhu et al., 1997) that REE can accumulate in the cerebral cortex with prolonged intake of small amounts from the environment and cause subclinical damage up to oncology. Brain tumor tissue (astrocytoma) has been found to contain significantly higher levels of REE, especially La, Ce, Gd, and Lu, compared to similar normal tissue (Zhuang et al., 1996). Using electron microscopy, we have detected Ce-containing mineral phases in brain tissue from wild deer harvested in an area with high background REE levels in the Sikhote-Alin (Baranovskaya et al., 2024).
In recent years, a number of U.S. researchers have studied unusual pathologies in wild and domestic animals, such as "chronic wasting disease" (CWD), which is associated with atrophy of bone marrow fat and hematopoietic cells (Gilbertson et al., 2022), and "transmissible spongiform encephalopathies", which are associated with damage to the brain and throughout the nervous system (Lavelle et al., 2014; Plummer et al., 2018). This family of diseases, common in both animals and humans, is considered to be caused by misfolded proteins - prions. The term "prion", from fragments of the English words protein and infection, was coined by Stanley Prusiner, who discovered such proteins in 1987. The causes of these diseases remain largely unknown. In wildlife, CWD most commonly affects members of the deer family. In the natural environment, according to the most widespread hypothesis (Plummer et al., 2018), such diseases are transmitted from diseased animals to healthy ones, including at kudurs, when animals eat prion-infected earthy substances. However, it seems to us that the cause of all diseases with prion formation, as well as blood anomalies, which are clearly triggered by the malfunction of the medullary component of the hematopoietic system, is directly related to REE microelementoses. Similarly, this is the main cause of such a neurodegenerative human disease as Alzheimer’s disease.
What is our basis for associating the named diseases with REE elementoses?
As mentioned above, interest in the use of LRE in the fattening of various animals has been actively growing during the last decades. Several researchers (Eisele et al., 1980; He, Rambeck, 2000; Fiddler et al., 2003; Rambeck et al., 2004; Hutchison, Albaaj, 2005) found that only small amounts of mineral forms of these elements are absorbed in the gastrointestinal tract. Nevertheless, due to the apparent ergotropic effects of LRRE supplements, some researchers (He, Rambeck, 2000; He, Xia, 2001) believe that even low doses of LRRE in the blood can cause significant biological effects. The most complete list of biological effects of LREE use that have been identified to date (illustrative in terms of dose dependence) is reported by Chinese researchers (Wang et al., 2003). This list includes stimulation or inhibition of body growth, cell proliferation, apoptosis; antioxidant or pro-oxidant activity; stabilization or destabilization of cytoskeleton; increase or inhibition of cell permeability; positive or negative regulation of cell signaling system; increase or decrease of hemoglobin affinity to oxygen; and enhancement of mineralization or demineralization.
From the brief review of biological effects associated with LREE food supplements, it is clear that the problem of assimilation of such elements in the body is positively solved. However, the mechanism of such assimilation remains unclear.
The fact is that the absorption of metal ions, according to Sullivan et al. (1984) and Kostial et al. (1989), occurs in the small intestine, but not for REE ions. They can only exist in a highly acidified aqueous solution. Once in the intestine, where the pH of the medium rises sharply to 8.0 and above, REE ions are inevitably converted to deionized mineral forms that cannot enter the body. Some researchers (Szasz et al., 1978; Cheng et al., 1999) have suggested that REEs are not absorbed in the intestine in ionic form, but as part of macromolecules capable of penetrating the cell membrane. In support of this idea in general, we propose to develop this hypothesis.
The mechanism of REE entry into the internal environment of the body most likely involves some microorganisms that are able to actively assimilate REE into their protein molecules. Such microorganisms may live in the gastric microbiome, where acidic digestion of food takes place, or in the border area of the stomach and small intestine (in mammals, probably, their maximum number lives in the duodenum). It is possible that assimilation of REE by bacteria occurs not only at the level of amino acids in proteins, but also at the level of DNA. If such microorganisms and products of their metabolism periodically enter the intestine, they decompose there, and REEs in proteins or their components are assimilated by the body. It should be noted that, according to the thesis of A.I. Belyanovskaya (2019), the maximum content of REEs in the tissues of the digestive tract of pigs is found in the large intestine (colon), i.e. exactly there, where the substances synthesized by bacteria are absorbed, including such a fat-soluble vitamin as phylloquinone (Metelsky, 2009), which is necessary for protein synthesis and bone mineralization.
If such a mechanism of REE assimilation exists, it should be universal for all multicellular animals. The specific REE-assimilating microorganisms should include the most ancient representatives, because normal functioning of the earliest multicellular animals would be impossible without them. The most likely candidates for "REE-bacteria" in the microbiome of the digestive tract of animals could be from the group of methanotrophs. Several types of methanol dehydrogenase (MeDH) enzymes have been found in the representatives of this group, in particular in Methylosinus trichosporium OB3b, and the expression of each of them is regulated by different REEs of the light subgroup (Ce, La, Pr, Nd) and sometimes Sm (Gu et al., 2016). Regarding the fact that no significant effect of REEs on the microbial composition in the gastrointestinal tract of pigs, poultry and bovine rumen has been demonstrated during animal fattening, as described in the theses of S. Schuller (2001) and S. Knebel (2004), it can only indicate that the proportion of "REE-bacteria" in the microbiome of the digestive tract of any animal is insignificant both quantitatively and species-wise, and that their localization there is spatially limited.
Since microorganisms participate in REE assimilation in the digestive tract of animals, and specific protein molecules are produced whose expression is regulated by different rare earth elements, it is quite possible that under the combination of some factors, protein molecules with incorrect chirality, i.e. prions, may be produced in the digestive tract.
In evaluating the published information we have collected on human geophagy, it is important to note also that both currently and historically, people practicing geophagy in Europe and the United States are predominantly either African Americans (more often women and children) from the United States itself and other countries in the Americas, or recent immigrants from Africa, as well as from India and Pakistan (Henry, Cring, 2014). All of these individuals come from areas of the world where geophagy was historically prevalent. This fact suggests that the genetic factor (the fixation in the genome of one or another norm of the level of REE content in the body) plays an important role in the impact of landscape REE anomalies on humans and animals. Only this can explain the persistent tendency to geophagy in natives of regions of the world with high levels of REE in landscapes when they move to new places of residence where REE levels differ greatly from their ancestral places. Significant differences in REE levels between human locations are well illustrated by data from Brown et al. (2004). The researchers compared La and Ce concentrations in baby teeth from Uganda and the United Kingdom. Teeth from Uganda had 4 times higher La concentrations than teeth from children in the UK, and Ce concentrations were almost 10 times higher.
Summarizing the review of studies on the relationship between mass geophagy and landscape REE anomalies, we can state with a high degree of certainty that the main cause of geophagy in mammals, including humans, worldwide is due to REE elementosis, which can be caused by both excess and deficiency of REE in feed and drinking water, reaching upper or lower thresholds of their concentrations beyond which the body cannot maintain metabolic processes in normal mode. By positioning the "rare earth" cause of geophagy as the main one, we imply that it is more widespread than the "sodium" cause, which is still the most popular in the scientific community in relation to mammals. As for the other ideas expressed by various authors about the causes of geophagy, we consider them, as well as the "sodium" cause, to be quite reasonable, but of secondary, auxiliary importance. Nevertheless, some seemingly typical kudurs may be visited by animals for the sole purpose of obtaining sodium. A particular craving for sodium is known to be most characteristic of ruminants, in which a large volume of this element is involved in the digestive cycle.
For almost 50 years we have visited and studied dozens of areas with mass manifestations of animal geophagy in the vast expanses of Russia, from the Caucasus Mountains in the west to the Sikhote-Alin Mountains in the east, and from the Verkhoyansk Mountains in the north to the Altai-Sayan Mountains in the south. More than a hundred kudurs were studied and tested in detail. There were only a few cases when animals visited kudurs only for sodium.
Lithophagy-Geophagy
Birds and Mammals
For what reason reptiles, as well as some aquatic mammals, e.g., sea lions, swallow relatively large pebbles from 5 cm in diameter and more (Fleming, 1951), we will not judge here because of the insufficiency of collected facts on these groups of animals. In this part of the paper we will deal mainly with birds, implying at the same time that in reptiles, which like birds have a muscular stomach, the functions of the swallowed stone material are most likely close to those of the gastroliths in birds.
The modern scientific community is still dominated by the belief that the main function of pebbles ingested by birds is "grinding", i.e. pebbles are supposed to help animals that have no chewing apparatus to grind their food in their muscular stomachs. Not only does such a hypothesis seem wrong to us, it is a classic example of a persistent scientific misconception that has lasted for decades.
We will begin the discussion of this topic with the reliable fact that in the composition of gastroliths of all bird species on all continents, 80 to 90% of the mineral grains are represented either by pure silica (quartz, chalcedony, opal) or by highly siliceous minerals, among which feldspars predominate. Very rarely, carbonate minerals (calcite or aragonite) may predominate in the composition of gastrolites. All authors of articles devoted to the study of mineral composition of gastroliths in birds point to such mineral composition (May, Braun, 1973; Lopez-Calleja et al., 2000; Sanders, Jarvis, 2000; Lee et al., 2004; Wings, 2007; Spragens et al., 2013; Sanders, Braun, 2021; Panichev, Seryodkin, 2022). Here we should note another important fact that has been reliably established: the size of most gastroliths in bird stomachs is sometimes less than 1 mm; in such cases, clay minerals may appear and even predominate in their composition. This unusual variety of minerals consumed by birds is found, for example, in southeastern Africa (Downs, 2006), in Papua New Guinea (Symes et al. 2006), and in northeastern South America, especially in the territories of Peru, Bolivia, and Ecuador (Brightsmith et al., 2008). In fact, instead of typical gastroliths in these areas, herbivorous forest birds actively ingest mineral associations of clay-silt fractions in mass form, the composition of which is often dominated by clay minerals, the same smectite, kaolinite, and illite. Such mineral complexes are in fact full analogs of kudurits.
One of the first to doubt the "grinding" purpose of gastrolites was V.I. Bgatov, again a geologist and again a Russian. He not only doubted, but also performed a highly competent experiment on chickens (Bgatov et al., 1987), the results of which were very important for understanding the role of silica minerals in the physiology of birds. This experiment was described in detail earlier (Panichev, Seryodkin, 2022). Here we will note the main point.
Applied by V.I. Bgatov three types of mineral additives in the menu of chickens in the form of gravel from quartz, rock crystal and zeolite tuff calibrated to 4 mm, after staying in the muscular stomach of chickens for one day were destroyed (rubbed against each other) by about 50% for quartz and quartz glass and 100% for much less strong zeolite tuff into crumbs of size from 3 to 0. 01 mm and smaller, which, as it turned out, are constantly excreted in the intestine, cloaca and from there outside as part of the droppings. In terms of the gross chemical composition, the silica crumbs at the "input" and "output" remained practically unchanged, unlike the zeolite crumbs, at the same time about twice as many water-soluble salts were found in bird droppings in experiments with quartz and glass than in experiments with zeolites (7.24 and 3.42%, respectively). However, taking into account the chemical elements involved in ion-exchange interactions on zeolites in the digestive tract, the total amount of elements excreted from the body by both zeolites and quartz was comparable. The set of elements excreted was approximately the same, including, in descending order, P, K, Na, Ca, Mg, S, Si, Mn, Al, and Fe. The only difference was that in the experiments with quartz and glass slightly more phosphorus and manganese were excreted from the birds’ bodies, and with zeolites it was potassium in the absorbed form.
The results on feed digestibility were also interesting in the experiment with chickens. It turned out that the use of mineral additives significantly increased the digestibility of proteins (by 2-3%), fats (by 10-20%), and fiber (by 10-50%). At the same time, the results on digestibility of proteins and fats were slightly better in the experiment with quartz and glass, and on digestibility of fiber – with zeolites.
V.I. Bgatov, with reference to R. Iler (1982), discussing the mechanism of removal of elements from the body of birds using silica gastroliths, explained it by the formation of negatively charged silica gel in the intestine in the conditions of neutral and slightly alkaline reaction of the environment. Such a gel in the biological electrolyte binds various cations and excretes them as part of the droppings. Based on the results obtained, he concluded that gastric stones from silicon oxides are actively involved in the regulation of mineral metabolism in the body of birds.
The ability of silica gastrolites to influence feed digestibility has been explained by the participation of "silica gel" sorbents in enzyme hydrolysis reactions. The increased activity of a number of digestive enzymes in the presence of clay sorbents was noted by scientists as early as the 1950s (Triven, 1983). In the late 1980s and 1990s, the effects of increasing feed digestibility by activating and prolonging the action of digestive enzymes in the presence of siliceous rocks (Makaridze, 1986) and zeolites (Kalyuzhnov et al., 1988) were also demonstrated in the fattening of birds and cattle.
As for the "grinding" function of gastroliths, it was not confirmed experimentally by V.I. Bgatov or his followers in industrial experiments of feeding birds with silica gastroliths (Lantseva and Motovilov, 2003; Lantseva, 2009). This hypothesis was finally rejected after it was shown that different mineral types of gastroliths, including those of the same strength, had significantly different effects on the digestibility of different dietary components, while the growth performance of chickens with and without gastroliths was often identical.
Thus, the available indisputable facts indicate that fragments of quartz and other highly siliceous minerals and rocks in the composition of gastroliths are subjected to mechanical crushing against each other under the action of compressive forces in the muscular stomach. When crushed products of gastroliths get into the intestine, there is an active production of silica gel from them, which by its negatively charged colloidal particles captures a significant part of various cations and removes them from the body as part of the droppings, thus providing regulation of mineral balance in the bird’s body. Silica mineral crumbs and gel particles as sorbents also participate in reactions of enzymatic hydrolysis of food. Speaking of which, in recent years such silica gel sorbents have been widely used in medicine (drugs on the Russian market: "Asorb", "Alesorb", "Alfasorb", "Novosorb", "Polysorb", etc.).
It should be noted that the most active grinding of mineral grains in the muscular stomach of birds seems to occur during certain periods of their diurnal cycle when there is almost no food in the stomach. This is due to the unusually high rate of food processing and removal from the digestive tract that is characteristic of birds. Almost complete absence of food in gastrolith-filled muscular stomachs of wild ducks and geese caught by hunters in the early morning has been repeatedly recorded in the Primorsky Krai (Panichev, Seryodkin, 2022).
Concluding the analysis of the published information on the functions of gastroliths in the body of birds, we should mention another of their secondary functions - "cleansing", which is evident only in domesticated ostriches. According to Wings (2007), these birds die of constipation without access to stones. If we take into account that in the stomach of these birds there are constantly more than 1 kg of "stones", and the same amount of them in the abnormally long intestine, we can assume that in the absence of such a weighty stone supplement, the system of evacuation of the food lump is forced to work in significantly different conditions. A severe physiological imbalance in the food evacuation system may well lead to constipation and death of birds.
Analyzing the published data on lithophagy in birds from the perspective of the "rare-earth" hypothesis, we had the idea to conduct an experiment to determine the ability of silica gels to bind REE ions in a solution with physicochemical parameters close to those of the animal intestine (including birds). A solution of REE salts in ammonium acetate buffer with pH 8.6 was prepared as a model of intestinal electrolyte. The experiment methodology is described in (Panichev et al., 2023a). Then 5.0 g of minerals (quartz, albite and calcite) ground to 1-10 μm were poured into flasks, then 50 ml of the prepared buffer solution with REE salts were added to each flask. Then the samples were shaken for 12 h. Separation of liquid from minerals was performed by centrifuge. The analysis was carried out by ICP-MS method.
The results showed that all three varieties of ground minerals added to an alkaline buffer with REE salts bind up to 98% of REE ions provided the concentration of elements is in the range from 5 to 15 ppb. This means that silica gels produced from gastroliths in the muscular stomachs of birds permanently bind and remove REE ions from the biological electrolyte in the digestive tract if their concentration does not exceed certain limits. Based on this fact, it can be concluded that one of the functions of gastroliths is to regulate the concentration and ratio of REE entering the body as part of the feed and drinking water. At normal, slightly elevated REE concentrations in feed and water, silica gel will continuously filter REE entering the digestive tract. In the case of inadequate intake of the light subgroup of REE, birds must seek out and periodically consume mineral substances with elevated REE concentrations in order to maintain their neuroimmunoendocrine systems in a state of stable equilibrium.
If we develop the "rare earth" hypothesis in relation to birds, taking into account the fact that in some habitats they ingest not typical gastroliths, but finely dispersed substances with a significant content of clays, then logic suggests that these are the cases in which REE should not be excreted from the body, which is what ordinary silica gastroliths do well. On the contrary, the necessary REE should be introduced into the body through the consumption of mineral sorbents saturated with them, and the elements in the composition of such sorbents should, as it is obvious, be in bioavailable form.
This most likely explains the mass aggregations of parrots and other large forest birds that consume clay sediments along the banks of the Tambopata, Madre de Dios, and other rivers in the Amazon basin. These Tertiary and Quaternary clay sediments are most likely enriched with bioavailable forms of REE as products of the destruction of Proterozoic and Early Paleozoic crystalline schists and granitoids, which, according to the geological map of the area, are distributed in the headwaters of the listed rivers. The revealed high content of sodium salts in clays eaten by birds (Powell, Brightsmith, 2009) can hardly be evaluated from the point of view of the reason of their eating. Rather, this fact explains the formation and preservation of smectite clays in such an unsuitable modern local climate. Unfortunately, we have not yet found clear analytical data on REE in bird teeth in this part of the world. Nevertheless, the presence of typical kudurits in the same areas frequented by numerous ungulates and other animals (Tobler, 2009; Griffiths et al., 2022; Ghanem, 2012), as well as evidence of geophagy in domestic animals (José et al., 2017), may suggest a "rare earth" source of their consumption. It is worth mentioning here that the study of geophagy in bats in Ecuador (Ghanem, 2012) revealed significant contents of Al, Ti, Y, Ce, La, and Nd in the feces of herbivores that visited kudurs, in contrast to the feces of insectivorous species that did not visit kudurs. The same areas are characterized by human geophagy, both currently (Johns, 1986; Browman, Gundersen, 1993) and historically (Laufer, 1930).
To consume clays, large forest birds come from the surrounding and remote floodplains of the Amazon, where REE deficiency in their diet is caused by the peculiarities of the geochemistry of forest landscapes in humid tropical climates. The hypothesis that clays sorb toxic substances of organic origin, which have been identified in some plant foods of forest birds in the tropics (Gilardi et al., 1999), is also not unfounded in our opinion, but we believe that such a function of clays consumed by birds should be considered only as one of the auxiliary ones.
Long-distance migrations of large forest birds to places where suitable gastroliths can be found are characteristic not only of the tropical zone, but also of the temperate zone. Periodic long-distance migrations are observed, for example, in grouse in some areas of Western Siberia. The same migrations of upland birds are also known for Eastern Siberia (Nazarov et al., 1975). According to the publication of A.M. Khokhlov (1969), similar phenomena are characteristic for a number of localities in the Russian Plain.
It should be noted that when we talk about the consumption of clay sorbents by birds, it is not a coincidence that we are talking about large birds (larger than a pigeon). Smaller birds have not been observed to consume clay materials. It seems that for small birds the problem of REE deficiency simply does not exist. Most likely, they require too small amounts of REE to meet their needs. Based on this logic, the most likely consumers of clays in REE-deficient landscapes among birds should be ostriches, cassowaries, bustards, and cranes. The main need of small birds in terms of REE is to constantly excrete their excess from the organism. The same seems to be true for small animals, including insects, such as termites, which constantly pass mineral soils through their digestive tract and at the same time remove REE from the organism, concentrating them in the products of their life activity and, first of all, in the composition of specific microorganisms that are active assimilators of REE.
Termites are mentioned because in many parts of Africa various animals tend to occasionally eat the mineral matter of termite structures. Soil samples from termite nests used by monkeys (chimpanzees) have been studied in Mahale Mountain National Park in Tanzania (Mahaney et al., 1999). An increased content of clay minerals of the kaolinite group was found in the eaten soils compared to the non-eaten ones, with an almost complete coincidence of macro- and micro-elements in their composition (except REE). This led the authors to suggest that the reason for this type of geophagy in monkeys is the desire to absorb toxic substances of plant origin with the help of clay minerals from termite constructions. Meanwhile, a simple comparison of the REE data given in the article indicates that the ingested soils are enriched in almost all trace elements of this group. Let us give some REE values for eaten and not eaten soils (averaged for 10 samples from termite nests and 9 from not eaten soils near termite nests): Ce content in the first case – 115 g/t, in the second – 74; La – 67 and 39; Nd – 34 and 18; Eu – 2.2 and 1.3; Dy – 5.0 and 3.0. As it is obvious, the interest of animals in soils of termite constructions could be explained from the point of view of the "rare-earth" hypothesis.
Another example regarding termite nests are the data from a research in Namibia (Sako, Mills, 2009). In this case, it was found that the contents of B, Fe, Mn, Ni, Cu, Zn, Se, Mo, and Cd, as well as REE, with a clear enrichment of elements of the light subgroup, were significantly higher in termite nest soils compared to the upper layer of surrounding soils. At the same time, as in other similar cases, the authors did not consider REE as the reason for the consumption of soils by animals.
Furthermore, we will consider the question of the occurrence of gastroliths in birds according to the seasons of the year. The analysis of literature data on the annual dynamics of gastrolith abundance in the stomachs of different bird species shows that annual maximums and minimums occur during the main seasonal changes in the diet of certain species, as well as during the breeding periods. At the same time, the minimum of gastroliths in the territory of the middle latitudes in upland birds is usually observed in spring, and the maximum in autumn (Panichev, Seryodkin, 2022). In the tropical forests of the Amazon basin, the maximum consumption of mineral substances varies for different species of birds, coinciding with their nesting period (Brightsmith et al., 2017).
In the most general form, these observations can be explained by the fact that in spring in the middle latitudes, on the threshold of rapid renewal and growth of all living things, all mineral substances are always in deficit, and birds are faced with the task of supplying the body with chemical elements to the greatest extent, so that gastrolites are not particularly necessary. In autumn, on the contrary, it is necessary to get rid of accumulated excesses of a number of chemical elements, which is possible only with plenty of silicon gastrolites. In tropical forests with an active leaching regime of soils and the upper part of the weathering crust of the underlying rocks, the situation is different. In the period before hatching, large birds need to restore the normal balance of REE and other elements in their bodies. Clay sediments enriched with various sorbed cations, including REE, are quite suitable for this purpose.
Thus, the main reason for the ingestion of mineral grains by birds is an instinctive way of adjusting the chemical composition of the digestive electrolyte in order to free it from excessive and toxic elements. It is not excluded that such regulation of electrolyte composition is also connected with maintenance of pH level in the digestive tract, thus providing optimal conditions for biochemical and microbiological processes. Among the main elements whose concentration is regulated by birds in the electrolytes of the digestive tract and, consequently, in the whole body, are undoubtedly the rare earth elements.
Gastrolith functions of lesser importance probably include: detoxification, activation and prolongation of digestive enzymes, cleansing function, perhaps to some extent the "grinding" function is also manifested in some bird species. All of these functions may be important for some bird species, but not for others. Thus, the secondary importance of the "enzyme" function is indicated by the fact of significant difference in the degree of digestibility of feed components in different species of birds, up to almost complete absence of effect. Such facts are shown on the basis of the analysis of literature on commercial fattening of birds in the article of Panichev and Seryodkin (2022).
Evaluating all the information we have collected on geophagy-lithophagy, we should note the following important idea: it seems very likely that birds and mammals (and perhaps all animals) are not only able to sense hormonal stress caused by REE imbalance in their neuroimmunoendocrine system. All of them should also be endowed with the ability to "sense" the REE they need in natural substances (including minerals) and to compensate for their deficiency in the body using them. In case of accumulation of excessive amounts of REE in the body, animals are able to instinctively get rid of them by consuming various types of natural, mainly mineral sorbents. In case of insufficient intake of necessary REE into the body, animals have to search for and consume mineral substances with high REE content, which may include not only clay-based sorbents, but also loams and even sands with REE minerals. The need to restore the normal functioning of the main control and protective system in the body should be most pronounced in young animals during the periods of active growth of the body, in females during the periods of carrying the fetus, in males after the rut, when a significant part of REE is lost with sperm. Finally, the above applies to all animals in the state of some diseases, including those due to disorders in the composition of the digestive microflora. As it is obvious, this list covers almost all known cases of geophagy, both in humans and animals, which is reflected in the extensive scientific literature on this problem. We have summarized our generalized version of the main and secondary causes of geophagy-lithophagy in Table 1.
Conclusions
The extensive complex of geological, mineralogical, landscape-geochemical and ecological studies in areas with manifestations of active geophagy in animals on the territory of Russia, as well as the analysis of published data on the consumption of earthy substances by humans and animals in different regions of the world, allow us to draw some generalizing conclusions about the problem of geophagy-lithophagy.
- The instinctive desire to eat earthy substances, products of hypergenic transformation of various rocks, preserved in many groups of animals and partly in humans, is, in its most general form, a manifestation of the evolutionarily conditioned way of regulating the material composition of the internal environment, as well as some biological and physiological processes in the body.
- There are two varieties of instinctive geophagy-lithophagy, which define two types of self-regulation of organisms with the help of natural minerals and earthy substances of complex composition.
All animals with muscular stomachs, especially herbivorous birds, are characterized by a more or less constant intake of silicon-rich, sand-sized mineral aggregates. In the muscular stomach, under the influence of compressive forces, the gastroliths are crushed by rubbing against each other with the formation of micro-sized crumbs which, upon entering the weakly alkaline environment of the intestine, form a silica gel with high sorption properties for cations and some molecules. The cations captured by the gel are excreted as part of the droppings, regulating the mineral balance in the bird’s body. Due to such mechanism of action in the body of this group of animals, silica minerals are permanently included in their homeostasis system.
For terrestrial placental mammals and humans living in some regions of the world, there is a characteristic periodic consumption of earthy substances, in the composition of which finely dispersed siliceous-clay mineral complexes are most often predominant. Such mineral substances, similar to silica gels, have the property of binding various cations and, in addition to their own wide spectrum of bioavailable chemical elements, mainly in the form of absorbed cations, are used only when necessary, mainly to restore the mineral balance in the body, which has been lost under the influence of external geochemically determined factors.
- 3.
- The main cause of geophagy-lithophagy in humans and animals is to maintain the required concentration and ratio of rare earth elements in the neuroimmunoendocrine system of the body. This control center of metabolic and immune defense processes in any organism is largely based on the unique physicochemical properties of f-electron atoms of REE, not all seventeen, but at best a few, perhaps six of them. The role of such atoms in the organism is only beginning to be studied. According to the available data, it is determined by the effects on ionotropic receptors. The tendency to regulate REE in the body, in fact, to geophagy, is controlled by the development of specific hormonal stress.
- 4.
- The complexity of maintaining the necessary ratio and concentration of REE in the body consists not only in the fact that all these elements without exception can be integrated into biological structures, but not all of them are capable of performing the functions required by the body. The complexity also lies in the fact that all REE necessary for the body, as well as all other vital elements, follow the law of "threshold" concentrations according to V.V. Kovalsky (1974), i.e. such concentrations above or below which metabolic regulation is disturbed, which is accompanied by dysfunction of life-support systems and, as a result, persistent metabolic disorders with the transition of the body to the state of endemic geochemically determined disease. The nature of REE endemias seems to be determined on the one hand by the ratios and concentrations of REE entering the internal environment of the body, and on the other hand by the genetic characteristics of organisms that have recently settled or have lived for a long time in landscapes that are REE-anomalous for a given organism.
- 5.
- The effect of minerals on any living body is multifactorial and not always positive. The complete list of mineral effects on living systems is presented in a separate table (see Table 1).
- 6.
- Sometimes the need for geophagy in animals (possibly also in humans) may not be related to the regulation of rare earth elements in the body, but is determined by the need to regulate their internal environment based on other functions that the mineral-crystalline substances have.
- 7.
- Instinctive geophagy in animals (especially in mammals) usually acquires traditional forms with long visits to the same places over centuries and millennia, resulting in the formation of specific landscape complexes, which we call by the term "kudurs". Kudurs, as places of wildlife concentration, have always been used by humans for hunting animals.
Acknowledgments
The author would like to thank N.V. Baranovskaya, I.Yu. Chekryzhov, I.V. Seryodkin, N.Y. Popov, A.V. Ruslan, E.A. Vakh, A.V. Vetoshkina, A.S. Kholodov, who participated in the collection and processing of factual information during the landscape geochemical studies in the areas of active geophagy in Siberia and the Russian Far East in the period from 2020 to 2024.
References
- Abrahams P.W. (1997) Geophagy (soil consumption) and iron supplementation in Uganda // Trop. Med. Int. Health. Vol. 2. No 7. Р. 23–617.Abrahams P.W. The Chemistry and Mineralogy of Three Savanna Lick Soils // Journal of Chemical Ecology,1999, V. 25, Is. 10, pp. 2215-2228.
- Abrahams P.W., Parsons J.A. (1996) Geophagy in the Tropics: a literature eview // Geogr. J. Vol. 162. P. 63–72.
- Аiler R.K. (1982) The Chemistry of Silica: Solubility, Polymerization, Colloid and Surface Properties and Biochemistry of Silica. Part I. Moscow: Mir. 416 p. (Russian Edition translated from Iler R.K., 1979).
- Аnell B., Lagercrantz S. (1958) Gefagical customs // Stud. ethnogr. upsal. Vol. 17. 98 p.
- Anitha J.K., Joseph Sabu, Rejith R.G., Sundararajan M. (2020) Monazite chemistry and its distribution along the coast of Neendakara-Kayamkulam belt, Kerala, India // SN Applied Sciences 2:812. [CrossRef]
- Aufreiter S., Hancock R.G.V., Mahanev W.C., Stambolic-Robb A., Sanmugadas K. (1997) Geochemistry and mineralogy of soils eaten by humans // J. Food Sciences and Nutrition V.48. P.293-305.
- Avtsyn A.P., Zhavoronkov A.A., Rish M.A., Strochkova L.S. (1991) Human microelementoses. Moscow: Medicine. 496 p. (In Russian).
- Banenzoue C., Signing P., Mbey Jean-Aimé, Njopwouo D. (2014) Antacid power and their enhancements in some edible clays consumed by geophagia in Cameroon // Journal of Chemical and Pharmaceutical Research, 6 (10): 668-676. https://www.researchgate.net/publication/282154824.
- Baranovskaya N.V., Mazukhina S.I., Panichev A.M. et al. (2024) Features of chemical elements migration in natural waters and their deposition in the form of neocrystallisations in living organisms (physico-chemical modeling with animal testing) // Bulletin of the Tomsk Polytechnic University. Geo Аssets Engineering. V. 335. No. 2. P. 187-201. [CrossRef]
- Belyanovskaya A.I. (2019) Elemental composition of the organism of mammals of natural and man-made territories and their ranking using the USETOX model // Dissertation for the degree of candidate of geological and mineralogical sciences. Tomsk-Bordeaux, 22 c. (In Russian).
- Bgatov V.I., Motovilov K.Ya., Speshilova M.A. (1987) Functions of natural minerals in metabolic processes of agricultural poultry. Agricultural Biology, No 7, pp. 98-102. (In Russian).
- Brightsmith D.J., Taylor J., Phillips T.D. (2008) The Roles of Soil Characteristics and Toxin Adsorption in Avian Geophagy // Biotropica, V. 40(6). P.766-774. [CrossRef]
- Brightsmith D.J., Hobson E.A., Martinez G. (2017) Food availability and breeding season as predictors of geophagy in Amazonian parrots // Ibis 18 p. [CrossRef]
- Brouziotis A.A., Giarra A., Libralato G., Pagano G., Guida M., Trifuoggi M. (2022) Toxicity of rare earth elements: An overview on human health impact // Frontiers in Environmental Science. [CrossRef]
- Browman D.L., Gundersen J.N. (1993) Altiplano Comestible Earths: Prehistoric and Historie Geophagy of Highland Peru and Bolivia // Geoarchaeology: An International Journal, Vol. 8, No. 5, 413-425.
- Brown C.J., Chenery S.R., Smith B., Mason C., Tomkins A., Roberts G.J. et al. (2004) Environmental influences on the trace element content of teeth – Implications for disease and nutritional status // Arch. Oral Biol. Vol. 49. Is. 9. P. 705-717.
- Bukhman G., Ziegler J., Parry E. (2008) Endomyocardial Fibrosis: Still a Mystery after 60 Years //www.plosntds.org 2008 Volume 2 Issue 2 e97.
- Cheek D.B., Smith R.M., Spargo R.M., Francis N. (1981) Zinc, copper and environmental factors in the Aboriginal peoples of the north-west // Aust. N. J. Med. Vol. 2. P. 508–512.
- Cheng Y., You L., Rongchang L., Wang K., Yao H. (1999) The uptake of cerium by erythrocytes and the changes of membrane permeability in CeCl3 feeding rats // Progress in Natural Science, 9:610-616.
- Chung E.O., Mattah B., Hickey M.D., Salmen C.R., Milner E.M., Bukusi E.A., Brashares J.S., Young S.L., Fernald L.C.H., Fiorella K.J. (2019) Characteristics of Pica Behavior among Mothers around Lake Victoria, Kenya: A Cross-Sectional Study// Int. J. Environ. Res. Public Health, 16, 2510. [CrossRef]
- Collignon R. (1992) А propos des troubles des conduites alimentaires du pica des médecins à la géophagie des géographes, des voyageurs et des ethnologues // Psychopathologie africaine, Vol. XXIV. Is. 3. P. 385-396.
- Cragin F.W. (1836) Observations on Cachexia Africana or dirt-eating //Am. J. Med. Sci. Vol. 17. P. 356–364.
- Curtlius M.F., Millican F.K., Layman E.M. et. al. (1963) Treatment of Rica with a vitamin and mineral supplement // Amer. J. Clin. Nutr. Vol. 12. P. 388–393.
- Dominy N.J., Davoust E., Minekus M. (2004) Adaptive function of soil consumption: An in vitro study modelling the human stomach and small intestine. Journal of Experimental Biology, 207: 319-324.
- Downs C.T (2006) Geophagy in the African Olive Pigeon (Columba arquatrix) // Ostrich, V. 77(1-2) P.40-44. [CrossRef]
- Dravert P.L. (1922) On lithophagy // Siberian Nature. No 1. P. 3-6. (In Russian).
- Dreyer M.J., Chaushev P.G., Gledhill R.F. (2004) Biochemical investigations in geophagia // J. Royal Soc. Med. Vol. 97 P. 48.
- Dubinin A.V. (2006) Geochemistry of rare earth elements in the ocean. Moscow: Nauka. 360 p. (In Russian).
- Duplex K. K. E., Wouatong Armand Sylvain Ludovic, Njopwouo Daniel, Ekosse Georges Ivo. (2018) Physico-chemical Characterization of Clayey Materials Consumed by Geophagism in Locality of Sabga (North-western Cameroon): Health Implications // International Journal of Applied Science and Technology, V. 8, No.3. P.57-68. [CrossRef]
- Eapen J.T. (1998) Elevated Levels of Cerium in Tubers from Regions Endemic for Endomyocardial Fibrosis (EMF) // Bull. Environ. Contam. Toxicol. 60:168-170.
- Edwards A.A., Mathura C.B., Edwards C.H. (1983) Effects of maternal geophagia on infant and juvenile rats // J. Nat. Medical Assoc., Vol. 75, №. 9. P. 895-902.
- Eisele G.R., Mraz F.R., Woody M.C. (1980) Gastrointestinal uptake and 144Ce in the neonatal mouse, rat and pig. Health Physics, 39:185-192.
- Ekosse G.E., Ngole V.M., Longo-Mbenza B. (2011) Mineralogical and geochemical aspects of geophagic clayey soils from the Democratic Republic of Congo // International Journal of the Physical Sciences, Vol. 6. Is. 31. P. 7302-7313.
- Ekosse G.I., Chistyakov K.V., Rozanov A.B. et al., (2020) Landscape Settings and Mineralogy of Some Geophagic Clay Occurrences in South Africa /O.V. Frank-Kamenetskaya et al. (eds.), Processes and Phenomena on the Boundary Between Biogenic and Abiogenic Nature, Lecture Notes in Earth System Sciences, pp 785-801. [CrossRef]
- Feidman M.D. (1986) Pica: current perspectives // Psychosomat. No 27. P. 519–523.
- Feng L., Xiao H., He X., et al. (2006) Neurotoxicological consequence of long-term exposure to lanthanum // Toxicology Letters. Vol. 165. P. 112-120.
- Fiddler G., Tanaka T., Webster I. Low systemic adsorption and excellent tolerability during administration of Lanthanum carbonate (FosrenolTM) for 5 days. In 9th Asian Pacific Congress of Nephrology, 19- 20. Februar 2003, Pattaya, Thailand, 2003.
- Fleming C.A. (1951) Sea Lions as Geological Agents // Journal of Sedimentary Petrology, V.21, No. 1. P. 22-25.
- Ferrell R.E., Vermeer D.E., LeBlanc W.S. (1985) Chemical and mineralogical composition of geophagical materials // Trace substances in environ. health XIX. Univ. Missouri, Р. 47–55.
- Gebel A.D. (1862) About earthy substances used for food in Persia // Notes of the Imperial Academy of Sciences. St. Petersburg. Vol. 2. P. 126-135.
- Geissler P.W., Shulman C.E., Prince R.J., Mutemi W., Mnazi C., Friis H., Lowe B. (1998): Geophagy, iron status and anaemia among pregnant woman on the coast of Kenya // Trans. R. Soc. Trop. Med. Hyg., 92. P.549-553.
- Ghanem S.J. Geophagy of tropical fruit-eating bats – mineral licks as a link between ecology and conservation // Dissertation zur Erlangung des akademischen Grades des Doktors der Naturwissenschaften (Dr. rer. nat.). 2012. 127 p.
- Gilardi J.D., Duffey S.S., Munn C.A., Tell L. (1999). Biochemical functions of geophagy in parrots: detoxification of dietary toxins and cytoprotective effects // J. Chem Ecol., 25. Р.897-922.
- Gilbertson M.L., Brandell E.E., et al. (2022) Сause of death, pathology, and chronic wasting disease status of white-tailed deer (Оdocoileus virginianus) mortalities in Wisconsin, USA // Journal of Wildlife Diseases, 58(4), pp. 803–815. [CrossRef]
- Gómez-Aracena, J., Riemersma, R. A., Gutiérrez-Bedmar, M., Bode, P., Kark, J. D., Garcia-Rodríguez, A., et al. (2006). Toenail cerium levels and risk of a first acute myocardial infarction: The EURAMIC and heavy metals study. Chemosphere 64, 112–120. [CrossRef]
- Griffiths B.M., Jin Yan, Griffiths L.G., Gilmore M.P. (2022) Physical, landscape, and chemical properties of Amazonian interior forest mineral licks // Environ Geochem Health. [CrossRef]
- Gu W., Muhammad Farhan Ul Haque, DiSpirito A.A., Semrau J.D. (2016) Uptake and effect of rare earth elements on gene expression in Methylosinus trichosporium OB3b // FEMS Microbiol Lett. 363(13): fnw129. Epub 2016 May 12. [CrossRef]
- Henríquez-Hernández, L. A., Boada, L. D., Carranza, C., Pérez-Arellano, J. L., González-Antuña, A., Camacho, M., et al. (2017). Blood levels of toxic metals and rare Earth elements commonly found in e-waste may exert subtle effects on hemoglobin concentration in sub-Saharan immigrants. Environ. Int. 109, 20–28. [CrossRef]
- Henry J.M., Cring F.D. (2014) Geophagy аn Anthropological Perspective. Р. 179-198. https://www.researchgate.net/publication/261362058.
- He M.L., Rambeck W.A. (2000) Rare earth elements – a new generation of growth promoters for pigs? Archives of Animal Nutrition, 53(4):323–334.
- He R., Xia Z. (2001) Effect of rare earth compounds added to diet on performance of growing- finishing pigs. World Wide Web, http://www.rare-earth-agri.com.cn/eng5.htm, 2001.accessed 15. July 2001.
- Hinz E. (1999) Formen der Geophagie und ihre Bedeutung für die Parasitologie // Tropenmed. Parasitol. Vol. 21. P. 1-14.
- Hladik, C. M., Gueguen, L. (1974) Géophagie et nutrition minérale chez les primates sauvages. Comptes Rendus de L’académie des Sciences Serie D, 279, 1393-1396.
- Hooda P.S., Henry C.J.K., Seyoum T.A., Armstrong L.D.M., Fowler M.B. (2004) The potential impact of soil ingestion on human mineral nutrition // Science of the Total Environment 333 75–87. [CrossRef]
- Hooper D., Mann H.H. (1906) Earth eating and the earth eating habit in India // Mem. Asiat. Soc. Bengal.Calcutta. Vol. 1. P. 249–270.
- Houston D.C., Gilardi J.D., Hall A.J. (2001). Soil consumption by elephants might help to minimize the toxic effects of plant secondary compounds in forest browse. Mammal. Review. Vol. 31. Is. 3-4. P. 249-254.
- Hutchison A.J., Albaaj F. (2005) Lanthanum carbonate for the treatment of hyperphosphataemia in renal failure and dialysis patients. Expert Opinion on Pharmacotherapy, 6(2): 319–328.
- Khokhlov A.M. (1969) Arrangement of artificial pebble beds for upland game. In: Proceedings of the Zavidovsky reserve hunting farm. Vol. 1. Moscow: Voenizdat, pp. 291-297. (In Russian).
- Jiesheng L., Shen Zhiguo, Yang Weidong et al. (2002) Effact of Long-term Intake of Rare Earht in Drinking Water on Trace Elements in Brains of Mice // J. Rare Earths. Vol. 20. № 5.
- Johns T. (1986) Detoxification function of geophagy and domestication of the potato // J. Chem. Ecol. 12. P. 635–646.
- Johns T., Duquette M. (1991) Detoxification and mineral supplementation as functions of geophagy // American Journal of Clinical Nutrition, 53: 448-456.
- Jones, Krishnamani, R., Mahaney W.C. (2000) Geophagy among primates: Adaptive significance and ecological consequences. Animal Behaviour, 59 (5), 899-915.
- José C.A., Donicer M.V., Mastoby M.M. Clinical diagnosis of allotrophagia in cattle of the department of Sucre Colombia // Colombiana Cienc Anim 2017; 9(2):141-146. [CrossRef]
- Kalyuzhnov V.T., Zlobina I.E., Nikulina P.G. (1988) Physiological justification of inclusion of zeolites in birds’ food. In: Use of zeolites from Siberia and the Far East in agriculture. Novosibirsk: Siberian Branch of the All-Union Academy of Agricultural Sciences, pp. 15-20. (In Russian).
- Ketch L.A., Malloch D., Mahaney W.C., Huffman M.A. (2001) Comparative microbial analysis and clay mineralogy of soil eaten by chimpanzees (Pan troglodytes Schweinfurhii) in Tanzania. Soil Biology and Biochemistry. Vol. 33. № 2. P. 199-203.
- Klaus G., Klaus-Hügi C., Schmid B. (1998) Geophagy by large mammals at natural licks in the rain forest of the Dzanga National Park, Central African Republic. Journal of Tropical Ecology, 14: 829-839.
- Klaus, G., Schmid B. (1998) Geophagy at natural licks and mammal ecology: a review. Mammalia, 62(4): 482-498.
- Knebel C. Untersuchungen zum Einfluss Seltener Erd-Citrate auf Leistungsparameter beim Schwein und die ruminale Fermentation im künstlichen Pansen (RUSITEC). Dissertation, Ludwig-Maximilians-Universität, München, 2004.
- Knezevich, M. (1998) Geophagy as a therapeutic mediator of endoparasitism in a free-ranging group of rhesus macaques (Macaca mulatta) // American Journal of Primatology, 44(1),71-82.
- Knudsen, J.W. (2002) Kula Udongo (earth eating habits): a social and cultural practice among Chagga Women on the slopes of Mount Kilimanjaro // Indilinga African Journ. Of Indigenous Knowledge Systems. V. 1. p.19-25. [CrossRef]
- Kostial K., Kargacin B., Landeka M. (1989) Gut retention of metals in rats. Biological Trace Element Research, 21:213-218.
- Kovalsky V.V. (1974) Geochemical ecology. Essays. Moscow: Nauka, 229 p. (In Russian).
- Kreulen D.A. (1985). Lick use by large herbivores: a review of benefits and banes of soil consumption. Mammal Rev. 15: 107-123.
- Krishnamani, R., Mahaney W.C. (2000) Geophagy among primates: Adaptive significance and ecological consequences. Animal Behaviour, 59 (5), 899–915. [CrossRef]
- Kutty V.R., Abraham S., Kartha C.C. (1996) Geographical distribution of endomyocardial fibrosis in South Kerala // Int. J. Epi. Vol. 25. No 6. P. 1202–1207.26.
- Lantseva N.N., Motovilov K.Ya. (2003) Influence of different high silica additives on the quality of poultry products. Advances in current natural sciences, No. 8, pp. 21-24. (In Russian).
- Lantseva, N.N. (2009) Experimental substantiation of the system of use of natural minerals-kudurits in feeding of agricultural poultry. Abstract of dissertation for the degree of Doctor of Agricultural Sciences. Novosibirsk, Altai State Agrarian University, 41 p. (In Russian).
- Laufer B. (1930) Geophagy Publications of the Field Museum of Natural History. Anthropological Series, Vol. 18, No. 2, pp. 99, 101-198.
- Lavelle M.J., Phillips G.E., Fischer J.W., Burke P.W., Seward N.W., Stahl R.S., Nichols T.A., Wunder B.A., VerCauteren K.C. (2014) Mineral licks: motivational factors for visitation and accompanying disease risk at communal use sites of elk and deer // Environ Geochem Health, V. 36(6). P. 1049-1061. [CrossRef]
- Lebedeva E., Panichev A., Kharitonova N., Kholodov A. Golokhvast К. (2020) Diversity, Abundance, and Some Characteristics of Bacteria Isolated from Earth Material Consumed by Wild Animals at Kudurs in the Sikhote-Alin Mountains, Russia // International Journal of Microbiology, Article ID 8811047, 9 pages. [CrossRef]
- Lee, D.E., Hamman, M.G., Black, J.M. (2004) Grit-site selection of black brant: Particle size or calcium content? Wilson Bulletin, V.116, no. 4, pp. 304–313. [CrossRef]
- Li W., Li C., Jiang Z., Guo R. & Ping X. (2018) Daily rhythm and seasonal pattern of lick use in sika deer (Cervus nippon) in China// Biological Rhythm ReseaRch, 2018. [CrossRef]
- Liu, Y., Wu, M., Zhang, L., Bi, J., Song, L., Wang, L., et al. (2019). Prenatal exposure of rare Earth elements cerium and ytterbium and neonatal thyroid stimulating hormone levels: Findings from a birth cohort study. Environ. Int. 133, 105222. [CrossRef]
- Lopez-Calleja, M.V., Soto-Gamboa, A., Rezende, E.L. (2000) The role of gastrolites on feeding behavior and digestive efficiency in the rufous-collared sparrow. The Condor, vol. 102, no. 2, pp. 465-469. [CrossRef]
- Mahaney W.C. Hancock R.G.V. (1990) Geochemical nalysis of African buffalo geophagic sites and dung on Mount Kenya, East Africa // Mammalia, Vol. 54. Is.1. P. 25-32.
- Mahaney W.С., Milner M.W., Sunmugadas K. et al. (1997) Analysis of geophagy soils in Kibale Forest, Uganda // Primates, Vol. 38. Is. 2. P. 159-176.
- Mahaney W.C., Watts D., Hancock R.G.V. (1990) Geophagia by Mountain Gorillas (Gorilla gorilla beringei) in the Virunga Mountains, Rwanda // Primates, Vol. 31. Is.1. P. 113-120.
- Mahaney W.C., Zippin, J., Hancock R.G.V., Aufreiter S., Campbell S., Malloch D., Wink M., Huffman M.A. (1999) Chemistry, mineralogy and microbiology of termite mound soils eaten by the chimpanzees of the Mahale mountains, Western Tanzania // J. Tropical Ecology, Vol. 15. P. 565-588.
- Mahaney W.C., Milner M.W., Muliono H., Hancock R.G.V., Aufreiter S. (2000). Mineral and chemical analyses of soil eaten by humans in Indonesia // Int. J. Environm. Healht Research, Vol. 10. P. 93-109. [CrossRef]
- Makaridze N.G. (1986) Adsorption ability of zeolites to some proteolytic enzymes. Communications of the Academy of Sciences of the Georgian Soviet Socialist Republic, Vol. 122, No 3, pp. 621-623. (In Russian).
- Mason D. (1833) On atrophia a ventriculo (mal d’Estomac) or dirt-eating // Rdlnb. Med. S. J. Vol. 39. P. 95–289.
- May, T.A., Braun C.E. (1973) Gizzard stones from adult white-tailed ptarmigan (Lagopus leucurus) in Colorado. Arctic and Alpine Research 5:49-57.
- Mengel G.E., Garter W.A., Horton E.S. (1964) Geophagia with iron deficiency and hypokalemia // Arch.lntern. Med. Vol. 114. P. 470–474.
- Meryem B., Hongbing J.I., Gao Yang, Ding Huajian, Li Cai (2016) Distribution of rare earth elements in agricultural soil and human body (scalp hair and urine) near smelting and mining areas of Hezhang, China //J. of Rare Earths, Vol. 34, No. 11. P.1156-1167. [CrossRef]
- Metelsky S.T. (2009) Physiological mechanisms of absorption in the intestine // National School of Gastroenterologists, Hepatologists, 3. P. 51-56. (In Russian).
- Mincher B.J., Ball R.D., Houghton T.P. et al. (2008) Some aspects of geophagia in Wyoming bighorn sheep (Ovis canadensis). European Journal of Wildlife Research, 54: 193-198.
- Minnich V., Okcuoglu A., Tarcon Y., Arcasov A., Cin S., Yorukoglu O., Renda, F., Demirag B. (1968) Pica in Turkey II. Effect of clay upon iron absorption. Am. J. Clin. Nutr. 21, 78–86.
- Mure A.I. (1934) The moose of isle Royale // Misc. Publ. Mus. Zool. Univ. Mich. No 25. P. 1–44.
- Nasimovich А.А. (1938) In regard to the knowledge of the mineral nutrition of wild animals of the Caucasian Reserve // Proceedings of the Caucasus Nature Reserve. Iss. 1. Moscow. 1938. P. 103-150. (In Russian).
- Nazarov A.A., Shubnikova O.N., Kirikov, S.V. (1975) Northern taiga. In: Grouse Birds. Moscow: Nauka, pp. 31-40.
- Nchito M., Geissler P. W., Mubila L., Friis H., Olsen A. (2004) Effects of iron and multimicronutrient supplementation on geophagy: a two-by-two factorial study among Zambian schoolchildren in Lusaka // Transactions of the Royal Society of Tropical Medicine and Hygiene, Vol. 98. P. 218-227.
- Ngole V.M., Ekosse G.E. (2012) Physico-chemistry, mineralogy, geochemistry and nutrient bioaccessibility of geophagic soils from Eastern Cape, South Africa // Scientific Research and Essays Vol. 7(12), pp. 1319-1331. [CrossRef]
- Palasz P.C. (2000) Toxicological and cytophysiological aspects of lanthanides action // Acta Biochimica Polonica, Vol. 47 No. 4. P. 1107-1114.
- Panichev A.M. (1987) Natural mineral ion exchangers - regulators of ionic equilibrium in the body of lithophagous animals // Dokl. of the USSR Academy of Sciences. Vol. 292. No 4. P. 1016-1019. (In Russian).
- Panichev A.M. (1990). Geophagia in the worlds of animals and humans. 223 p. Moscow: Nauka (In Russian).
- Panichev A. M. (2011). Lithophagy (geological, ecological and biomedical aspects). Moscow: Nauka. 149 p. (in Russian).
- Panichev A.M., Golokhvast K.S., Gulkov A.N., Сhekryzhov I.Yu. (2013) Geophagy and geology of mineral licks (kudurs): a review of Russian publications // Environmental Geochemistry and Health. № 1. Р.133-152.
- Panichev A.M. (2015) Rare Earth Elements: Review of Medical and Biological Properties and Their Abundance in the Rock Materials and Mineralized Spring Waters in the Context of Animal and Human Geophagia Reasons Evaluation // Achievements in the Life Sciences 9. P. 95-103. [CrossRef]
- Panichev A.M. (2016) Lithophagy: causes of the phenomenon // Priroda. No. 4. P. 25-35. (In Russian).
- Panichev A.M., Popov V.K., Chekryzhov I.Yu et al. (2016) Rare earth elements upon assessment of reasons of the geophagy in Sikhote-Alin region (Russian Federation), Africa and other world regions // Environ Geochem Health. 38. P.1255–1270. [CrossRef]
- Panichev A.M., Baranovskaya N.V., Seryodkin I.V., et al. (2021) Landscape REE anomalies and the cause of geophagy in wild animals at kudurs (mineral salt licks) in the Sikhote-Alin // Environmental Geochemistry and Health. [CrossRef]
- Panichev, A.M., Seryodkin, I.V. (2022) The mineral composition of gastroliths in the stomachs of Anatidae in Primorsky Region and the importance of silicon minerals in the physiology of birds. Amurian Zoological Journal, vol. XIV, no. 3, pp. 469–491. [CrossRef]
- Panichev A.M., Baranovskaya N.V., Seryodkin I. et al. (2023a) Excess of REE in plant foods as a cause of geophagy in animals in the Teletskoye Lake basin, Altai Republic, Russia // World Academy of Sciences Journal, № 5 V. 6. Р.1-22.
- Panichev A.M., Baranovskaya N.V., Seryodkin I.V et al. (2023b) The Main Cause of Geophagy According to Extensive Studies on Olkhon Island, Lake Baikal // Geosciences, 13, 211. [CrossRef]
- Panichev A.M., Baranovskaya N.V., Chekrizhov I.Yu., Ivanov V.V., Tsatska A.N. (2024) An Unusual Variety of Geophagy: Coal Consumption by Snow Sheep in the Transbaikalia Mountains // Doklady Earth Sciences,. [CrossRef]
- Plummer I.H., Johnson C.J., Chesney A.R., et al. (2018) Mineral licks as environmental reservoirs of chronic wasting disease prions PLoS One. May 2;13(5):e0196745. eCollection 2018. [CrossRef]
- Powell T.U., Brightsmith D. (2009) Parrots Take it with a Grain of Salt: Available Sodium Content May Drive Collpa (Clay Lick) Selection in Southeastern Peru // Biotropica V.41(3):279-282. [CrossRef]
- Powis D.A., Clark C.L., O’Brien K.J. (1994) Lanthanum can be transported by the sodium-calcium exchange pathway and directly triggers catecholamine release from bovine chromaffin cells. Cell Calcium, 16(5):377-390.
- Ramachandran K.K., Balagopalan M., Vijayakumaran Nayr P. (1995). Use pattern and chemical characterization of the natural salt licks in Chinnar wildlife sanctuary (Research report 94). Kerala Forest Research Institute Peechi, Thrissur, 18 p.
- Rambeck W.A., He M.L., Wehr U. (2004) Influence of the alternative growth promoter "Rare Earth Elements" on meat quality in pigs. In Proceedings of the British Society of Animal Science pig and poultry meat quality - genetic and non-genetic factors, 14-15. October 2004, Krakow, Poland 2004.
- Redling K. (2006). Rare earth elements in agriculture with emphasis on animal husbandry. Dissertation. Munich: University of Munich. https://edoc.ub.uni-muenchen.de/5936/1/Redling_Kerstin.pdf.
- Reichardt F. (2008) Ingestion spontanée d’argiles chez le rat: Rôle dans la physiologie intestinale/ Thèse de doctorat. Université Louis Pasteur – Strasbourg. 222 p. https://theses.fr/2008STR13206.
- Reid R.M. (1992) Cultural and medical perspectives on geophagia // Med. Anthropol. Vol. 13. No 4. P. 337–351.
- Robertson R.H.S. 1996.Sadavers, cholera and clays//Mineralogical Society Bulletin,No.113.Р. 3-7.
- Robinson B.A., Tolan P.W. (1990) Golding-Beecher О. Childhood pica: some aspects of the clinical profile in Manchester, Jamaica // W. lnd. Med. J. Vol. 39. P. 20-26.
- Ross T. (1895) Personal narrative of travels to the equinoctial regiom of America during the years 1799–1804 by Alexander von Humholdt and Aime Bonpland. London: Routledge, Vol. 2.
- Rowland, M.J. (2002) Geophagy: an assessment of implications for the development of Australian Indigenous plant processing technologies // Australian Aboriginal Studies, https://www.thefreelibrary.com/Geophagy%3A+an+assessment+of+implications+for+the+development+of...-a0127621929.
- Sako A., Mills A.J., Roychoudhury A.N. Rare earth and trace element geochemistry of termite mounds in central and northeastern Namibia: Mechanisms for micro-nutrient accumulation // Geoderma, 2009. Vol. 153. Is. 1-2. P. 217-230.
- Sanders T.A., Braun C.E. Minerals in gastroliths and foods consumed by band-tailed pigeons // J Wildl Manag. 2021;1–24. [CrossRef]
- Sanders, T. A., Jarvis R.L. 2000. Do band-tailed pigeons seek a calcium supplement at mineral sites? Condor 10: 855-863.
- Schuller S. Seltene Erden als Leistungsförderer beim Geflügel. Untersuchungen an Broilern und Japanischen Wachteln. Dissertation, Ludwig-Maximilians-Universität, München, 2001.
- Seim G.L., Ahn C.I., Bodis M.S., Luwedde F., Miller D.D., Hillier S.E, Tako E.C., Glahn R.P., Young S.L. (2013) Bioavailability of iron in geophagic earths and clay minerals, and their effect on dietary iron absorption using an in vitro digestion // Food and Function, Vol. 4. Is. 8. P.1263-1270.
- Severance H.W., Holt T., Patrone N.A., Chapman L. (1988) Profound muscle weakness and hypokalemia due to clay ingestion. South // Med. J. Vol. 81. P. 272-284.
- Smith B., Chenery S.R.N., Cook J.M. et al. (1998) Geochemical and environmental factors controlling exposure to cerium and magnesium in Uganda // J. Geochem. Explor. Vol. 65. Is.1. P. 1-15.
- Spragens, K. A., Bjerre, E. R., Black, J. M. (2013). Black brant Branta bernicla nigricans grit acquisition at Humboldt Bay, California, USA. Wildfowl, no. 3, pp. 104-115.
- Staaland H., White R. G., Luick J. R., Holleman D.F. (1980) Dietary influences on sodium and potassium metabolism of reindeer. Canadian Journal of Zoology 58:1728-1734.
- Stockstad D.S., Morris M.S., Lory E.C. (1953) Chemical characteristics of natural licks used by big game animals in western Montana // Trans.N. Amer. Wildlife Conf. Vol. 18. P. 247-257.
- Struhsaker T.T., Cooney D.O., Siex K.S. (1997) Charcoal Consumption by Zanzibar Red Colobus Monkeys: Its Function and Its Ecological and Demographic Consequences //International Journal of Primatology, V.18, Р. 61–72.
- Sternberg L.Y. (1933) Gilyaks, Orochs, Golds, Negidals, Ainu. Habarovsck: Dalgiz. 740 p.
- Sullivan M.E., Miller B.M., Goebel J.C. (1984) Gastrointestinal absorption of metals (51Cr. 65 Zn, 95mTc, 109Cd, 113Sn, 147Pm and 238Pu) by rats and swine. Environmental Research, 35 (2):439- 453.
- Symes C.T., Hughes J C., Mack A.L., Marsden S.J (2006) Geophagy in birds of Crater Mountain Wildlife Management Area, Papua New Guinea// Journal of Zoology. V. 268. P. 87-96. [CrossRef]
- Szasz B., Sarkadi, Schubert A., Gardos G. (1978) Effects of lanthanum on calcium dependent phenomena in human red cells // Biochimica et Biophysica acta, 512:331-340.
- Tariq H., Sharma A., Sarkar S., Ojha L., Pal R.P., Mani V. (2020) Perspectives of rare earth elements as feed additive in livestock: A review //Asian-Australas J Anim Sci. Mar.33(3):373-381. [CrossRef]
- Temga J.P., Sababa E., Mamdem L.E., Ngo Bijeck M.L., Azinwi P.T. et al. (2021) Rare earth elements in tropical soils, Cameroon soils (Central Africa) // Geoderma Regional, V. 25 (2021) e00369. [CrossRef]
- Tobler M.W., Carrillo-Percastegui S.E., Powell G. (2009) Habitat use, activity patterns and use of mineral licks by five species of ungulate in south-eastern Peru // Journal of Tropical Ecology, V.25. P.261-270. [CrossRef]
- Triven M., (1983) Immobilized Enzymes. Moscow: Mir. 211 p. (Russian Edition translated from Triven M., 1980).
- Valiathan M.S., Kartha C.C., Eapen J.T. et al. A geochemical basis for endomyocardial fibrosis // Cardiovasc Res. 1989. Vol. 23. No 7. Р. 647–648.
- Vermeer D.E. (1984) Geophagical clays from the Benin area of Nigeria: a major source for the markets of West Africa // Nat. Geogr. Soc. Res. Rep. No 16. P. 705-711.
- .
- Vermeer D.E., Ray E., Ferrell J. (1985) Nigerian Geophagical Clay: A Traditional Antidiarrheal Pharmaceutical // Science. Vol. 227. P. 634-636.
- Voros J., Mahaney W.C., Milner M.W. (2001) Geophagy by the Bonnet Macaques (Macaca radiata) of Southern India: A Preliminary Analysis // Primates, V.42. Is.4. P. 327-344.
- Wang M.Q., Xu Z.R. (2003) Effect of Supplemental Lanthanum on the Growth Performance of Pigs // Asian-Aust. J. Anim. Sci. Vol. 16. No. 9. Р. 1360-1363.
- Wilson M.J. (2003) Сlay mineralogical and related characteristics of geophagic materials // Journal of Chemical Ecology, Vol. 29, No. 7, Р. 1525-1547.
- Wings, O. (2007) A review of gastrolith function with implications for fossil vertebrates and a revised classification. Acta Palaeontologica Polonica, vol. 52, no. 1, pp. 1-16.
- Xun W., Shi L., Hou G., et al. (2014) Effect of rare earth elements on feed digestibility, rumen fermentation, and purine derivatives in sheep //Italian Journal of Animal Science 2014; V.13. Р. 357-362.
- Young S.L., Wilson J., Hillier S. et al., (2010) Differences and Commonalities in Physical, Chemical and Mineralogical Properties of Zanzibari Geophagic Soils // Journal of Chemical Ecology 36(1):129-40. [CrossRef]
- Young S.L., Sherman P.W., Lucks J.B., Pelto G.H. (2011) Why on earth?: evaluating hypotheses about the physiological functions of human geophagy // The quarterly review of biology,.V.86, No. 2. Р.97-120. http://www.jstor.org/stable/10.1086/659884.
- Zhao H., Cheng Z., Cheng J., et al. (2012) Erratum to: The Toxicological Effects in Brain of Mice Following Exposure to Cerium Chloride // Biol Trace Elem Res, 148:134. [CrossRef]
- Zhuang, G., Zhou, Y., Lu, H., Lu, W., Zhou, M., Wang, Y., et al. (1996) Concentration of rare Earth elements, As, and Th in human brain and brain tumors, determined by neutron activation analysis. Biol. Trace Elem. Res. 53, 45–49. [CrossRef]
- Zhang Ru, Wang Li, Yonghua Li et al. (2020) Distribution Characteristics of Rare Earth Elements and Selenium in Hair of Centenarians Living in China Longevity Region // Biological Trace Element Research, V.197(3). P.15-24. [CrossRef]
- Zhu, W., Xu, S., Shao, P., Zhang, H., Wu, D., Yang, W., et al. (2005) Investigation on liver function among population in high background of rare Earth area in South China. Biol. Trace Elem. Res. 104, 001–008. [CrossRef]
- Zhu, W., Xu, S., Shao, P., Zhang, H., Wu, D., Yang, W., et al. (1997) Bioelectrical activity of the central nervous system among populations in a rare Earth element area. Biol. Trace Elem. Res. 57, 71–77. [CrossRef]

Table 1.
Constituents and mechanisms of action of mineral-crystalline substances of "edible earths", kudurits, and gastroliths in the body of humans and animals.
Table 1.
Constituents and mechanisms of action of mineral-crystalline substances of "edible earths", kudurits, and gastroliths in the body of humans and animals.
| No. | Biological effects | Implementation mechanisms |
| Main effect | ||
| 1 |
Regulation of the composition and concentration of rare earth elements (REE) in the body, primarily in the neuroimmunoendocrine system |
In REE-rich landscapes by sorption in the digestive tract of excess REE on various types of mineral, organomineral and organic sorbents (including silica gels produced in the muscular stomachs of birds from siliceous minerals) and their removal from the body. |
|
In REE-poor landscapes by delivering the necessary elements to the body as part of REE-enriched minerals, clays, and other types of sorbents. | ||
| Minor effects | ||
| 2 | Supply of Na and other chemical elements to the body | Through salts and in sorbed form in various types of natural sorbents. In a number of cases in relation to mammals, especially from the ruminant group, Na can be positioned as the main element sought in kudurits. |
| 3 | Detoxification effects | By sorption and inactivation of toxic elements and compounds. |
| 4 | Participation in enzymatic reactions, especially hydrolytic reactions | By prolonging the action of enzymes when sorbed by silica gel or other natural sorbents. |
| 5 | Mechanical effects in the digestive tract | Increased peristalsis, cleansing of the digestive tract; in muscular stomachs, some mechanical processing of food is possible |
| 6 | Antacid effect | Regulation of pH in different parts of the digestive tract through the consumption of minerals with amphoteric, acidic or alkaline properties. |
| 7 | Binds excess water in the bowel | Due to hydration of ions, molecules and absorption of water by various types of sorbents. |
| 8 | Strengthening of antioxidant protection of body cells | By enhancing the antioxidant properties of a number of enzymes with some chemical elements. |
| 9 | Immune system training | By constant influx of antigenic material into the body as part of mineral adjuvants. |
| 10 | Effect on digestive microflora | Introduction of beneficial associations of microorganisms into the digestive tract; activation of fermentative forms; binding and removal of atypical microorganisms, including fungi and actinomycetes, from the digestive tract. |
| 11 | Energy-information effects | By the appearance of stable holograms of crystalline substances in the body, as well as by other, not yet studied energy-information interactions. |
| 12 | Negative biological effects are determined by the presence of toxic mineral and organic impurities in the ingested earthy substances, their dosage and the degree of their dispersion, as well as by the presence of harmful microorganisms, protozoa, helminth eggs and various toxins. | |
Note. Biological effects and their implementation mechanisms numbered 6-12 are discussed in the book (Panichev, 2011).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



