
Submitted:
10 December 2024
Posted:
11 December 2024
You are already at the latest version
Abstract
For photosynthesis, phototrophic organisms necessarily contain not only chlorophylls but also carotenoids. Various carotenoids have been identified in algae and taxonomic studies of algae have been conducted. In this review, the relationship between the distribution of chlorophylls and carotenoids and the phylogeny of sea and fresh water oxygenic phototrophs, including cyanobacteria, red algae, brown algae, and green algae, is summarized. These phototrophs contain division- or class-specific carotenoids, such as fucoxanthin, peridinin, diadinoxanthin, and siphonaxanthin. The distribution of -carotene and its derivatives, including -carotene, zeaxanthin, violaxanthin, neoxanthin, diadinoxanthin, fucoxanthin, and peridinin (-branch carotenoids), are limited to divisions of a part of Rhodophyta, Cryptophyta, Heterokontophyta, Haptophyta, and Dinophyta. Whereas, the distribution of α-carotene and its derivatives, such as lutein, loroxanthin, and siphonaxanthin (α-branch carotenoids), are limited to divisions of a part of Rhodophyta (macrophytic type), Cryptophyta, Euglenophyta, Chlorarachniophyta, and Chlorophyta. In addition, carotenogenesis pathways are also discussed based on the chemical structures of carotenoids and the known characteristics of carotenogenesis enzymes in other organisms. The specific genes and enzymes for carotenogenesis in algae are not yet known. Most carotenoids bind to membrane-bound pigment-protein complexes, such as reaction centers and light-harvesting complexes. Some carotenoids function in photosynthesis and are briefly summarized. Water-soluble peridinin-chlorophyll a-protein (PCP) and orange carotenoid protein (OCP) have also been characterized. This review is a summary and update from the previous review (Takaichi, S. Carotenoids in Algae. Mar. Drugs 2011, 9, 1101-1118) on the distribution of major carotenoids, primary carotenogenesis pathways, and the characteristics of carotenogenesis enzymes and genes.
Keywords:
algal phylogeny
; biosynthesis of carotenoids
; distribution of carotenoids
; distribution of chlorophylls
; function of carotenoids
; pigment-protein complexes
1. Introduction
Oxygenic phototrophs are classified throughout many divisions of the plant kingdom and bacteria, including cyanobacteria, red algae, brown algae, green algae, and land plants (Table 1). The sizes of algae range from single cells of picophytoplankton—the smallest of which is less than 1 µm—to macrophytic seaweeds—the largest of which is greater than 50 m. Attempts have been made to cultivate single-cell algae and cyanobacteria for a long time, but the numbers of species that can be cultured was initially limited. With the recent development of culture techniques, some single cell may be cultured, and enabling the study of their characteristics, including pigments and phylogeny. The development of taxonomic technologies, such as DNA base sequences of 16S or 18S rRNA, some genes, and both genomic and chloroplast DNA, algae phylogenetics, and also precise morphological characteristics, has advanced algal phylogenetics [1,2].
More than 850 structurally defined natural carotenoids have been identified in bacteria including cyanobacteria and anoxygenic phototrophic bacteria, archaea, fungi, algae, land plants, and animals [2,3]. Except for animals, these organisms can synthesize a variety of carotenoids from commonly isopentenyl pyrophosphate (IPP) and carotenoids are synthesized through diverse carotenogenesis pathways. Their distribution is division- and/or class-specific (Table 1) [1,2,4,5,6,7]. In addition, the characteristics of carotenogenesis genes and enzymes have been examined. Some carotenogenesis genes exhibit high 4amino acid sequence similarity from bacteria to land plants, whereas some have low similarity. Some homologous genes have been proposed, but some carotenogenesis genes and enzymes, particularly in algae, have not been found.
In contrast, the number of chlorophyll (Chl) molecules is limited. Their distribution is division- and/or class-specific, and their biosynthesis has been established (Table 1). The biosynthetic pathways of chlorophylls are common, and each chlorophyll and bacteriochlorophyll is branched from the main pathway [8,9].
Takaichi [10] provided a summary of “Carotenoids in Algae: Distribution, Biosyntheses and Functions” more than one decade ago. In this updated review, oxygenic phototrophs, including algae and cyanobacteria found in both seawater and fresh water, as well as land plants are discussed. This review summarizes the distribution of carotenoids and chlorophylls, carotenogenesis enzymes and pathways, and the function of carotenoids in photosynthesis in oxygenic phototrophs (Table 1).

Table 1.
Distribution of carotenoids and chlorophylls in oxygenic phototrophs. This table is an updated version presented in the previous review [10].
Table 1.
Distribution of carotenoids and chlorophylls in oxygenic phototrophs. This table is an updated version presented in the previous review [10].
| Division | Carotene | Xanthophyll | Chlorophyll | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | β | α | Ze | Vi | Ne | Da | Dd | Fx | Va | Lu | Lo | Sx | Others | a | b | c | ||
| Cyanophyta | H | L | H | No, L; Ec, H; My, H; Sy, L | H | L | ||||||||||||
| Glaucophyta | H | H | H | |||||||||||||||
| Rhodophyta (red algae) | ||||||||||||||||||
| ZEA-type | H | H | H | |||||||||||||||
| ANT-type | H | H | An, H | H | ||||||||||||||
| LUT-type | H | L | L | H | H | |||||||||||||
| Cryptophyta | L | H | L | Al, H; Cr, L; Mo, L | H | c2 | ||||||||||||
| Heterokontophyta (brown algae) | ||||||||||||||||||
| Chrysophyceae | H | L | L | L | L | H | H | c1, c2 | ||||||||||
| Raphidophyceae | H | H | L | L | L | H | H | c1, c2 | ||||||||||
| Bacillariophyceae | H | L | L | L | H | H | c1, c2, c3 | |||||||||||
| Phaeophyceae | H | H | H | L | L | H | H | c1, c2, c3 | ||||||||||
| Xanthophyceae | H | L | L | H | H | L | He, L; Va-FA, L | H | c1, c2 | |||||||||
| Eustigmatophyceae | H | L | H | L | Va-FA, L | H | ||||||||||||
| Haptophyta | H | L | L | H | H | L | Fx-FA, L | H | c1, c2 | |||||||||
| Dinophyta | L | L | L | L | H | L | Pe, H; P457, L | H | c2 | |||||||||
| Euglenophyta | H | L | L | L | H | H | H | |||||||||||
| Chlorarachniophyta | H | L | L | L | L | L | L | Lo-FA, L | H | H | ||||||||
| Chlorophyta (green algae) | ||||||||||||||||||
| Prasinophyceae | H | L | L | H | H | L | L | H | Pr, L; Lo-FA, L; Sx-FA, H | H | H | |||||||
| Chlorophyceae | H | H | L | H | H | H | L | L | Sx-FA, L | H | H | |||||||
| Ulvophyceae | H | L | L | H | H | L | L | L | Sx-FA, H | H | H | |||||||
| Trebouxiophyceae | H | L | L | H | H | H | H | H | ||||||||||
| Charophyceae | H | L | L | H | H | H | H | H | ||||||||||
| Land Plants | H | L | L | H | H | H | An,L | H | H | |||||||||
Red, a-Branch carotenoids; H, Major catenoid in most species of the class; L, Low content in most species or major carotenoid in limited species. α,α-carotene; β,β-carotene; Al, alloxanthin; An, antheraxanthin; Cr, crocoxanthin; Da, diatoxanthin; Dd, diadinoxanthin; Ec, echinenone; -FA, fatty acid ester; Fx, fucoxanthin; He, heteroxanthin; Lo, loroxanthin; Lu, lutein; Mo, monadoxanthin; My, myxol 2’-glycosides and 2,2’-oscillol diglycosides; Ne, neoxanthin; No, nostoxanthin; Pe, peridinin; Pr, prasinoxanthin; Sx, siphonaxanthin; Sy, synechoxanthin; Va, vaucheriaxanthin; Vi, violaxanthin; Ze, zeaxanthin. Some cyanobacteria contain chlorophyll d, chlorophyll f, divinyl-chlorophyll a, or divinyl-chlorophyll b.
2. Distribution of Carotenoids and Chlorophylls
Many different carotenoids are found in oxygenic phototrophs, including cyanobacteria, red algae, brown algae, green algae, and land plants. The chemical structures of several important carotenoids in the oxygenic phototrophs are illustrated in the figures of this review. Of these, approximately 30 to 50 types are believed to have direct roles in photosynthesis, whereas others may be intermediates in carotenogenesis or as accumulated carotenoids. Some carotenoids are unique to specific algal divisions and/or classes, making them valuable chemotaxonomic markers (Table 1) [1,2,4,5,6,7].
Carotenoids contain several unique functional groups. The allene group (C=C=C) is a unique structure in natural products and is primarily found in carotenoids [11], such as fucoxanthin in brown algae and diatoms, peridinin only in Dinophyta, and 9′-cis neoxanthin in green algae, Chlorarachniophyta, and land plants. The acetylene group (C≡C) is a unique structure, and acetylenic carotenoids occur only in algae, such as alloxanthin, crocoxanthin, and monadoxanthin in Cryptophyta as well as diadinoxanthin and diatoxanthin in Heterokontophyta, Haptophyta, Dinophyta, and Euglenophyta. Acetylated carotenoids (-O-CO-CH3), such as fucoxanthin, peridinin, and dinoxanthin, are also primally found in algae, such as Heterokontophyta, Haptophyta, and Dinophyta. These carotenoids are specific to certain algal divisions and classes (Table 1) based on our findings [2,10,12,13,14,15,16] and others [1,3,4,5,6,17,18,19]. Their structures are illustrated in the figures of this review.
In contract, the number of chlorophylls is limited. Their structures are relatively similar, and their distribution is also division- and/or class-specific. Consequently, the distribution of these carotenoids, carotenogenesis pathways, and chlorophylls may be used as valuable chemotaxonomic markers. Their distribution in the oxygenic phototrophs is summarized in Table 1. The structures of chlorophylls are illustrated in Figure 1 and their biosynthetic pathways are summarized in [8,9].
It is believed that oxygenic cyanobacteria evolved from two types of anoxygenic phototrophic bacteria through evolution and endosymbiosis. Whole genome analysis of organisms has provided new insight into the evolution between cyanobacteria and anoxygenic phototrophic bacteria [20,21]. Furthermore, all known plastids, including those in algal and land plant chloroplasts, originated from a single primary endosymbiotic event involving a cyanobacterium. This led to eukaryotic cells and the formation of three divisions, including Glaucophyta, Rhodophyta, and Chlorophyta. Other algae, such as Cryptophyta, Heterokontophyta, Haptophyta, Euglenophyta, and Chlorarachniophyta evolved through secondary endosymbiosis of these algae, whereas Dinophyta represents a tertiary endosymbiosis involving some algae [22,23].
Although chlorophylls and their synthetic pathways [8,9] and the peptide components of the photosynthetic pigment–protein complexes in oxygenic phototrophs from cyanobacteria, algae, and land plants, as well as anoxygenic phototrophic bacteria, have remained relatively consistent throughout evolution, the carotenoid compositions have critically changed at each step of endosymbiosis. This suggests that certain pathways and genes for carotenoid synthesis are lost or acquired discontinuously at each evolutionary stage (Table 1).
In this review, cyanobacteria include both seawater and freshwater environments and both contain similar pigments. Many cyanobacteria contain β-carotene, zeaxanthin, echinenone, and myxol methylpentosides (myxoxanthophyll), whereas some species lack one or more of these carotenoids. Some contain additional carotenoids, such as nostoxanthin, canthaxanthin, synechoxanthin, and oscillol dimethylpentoside (Table 1) [14]. The carotenoid compositions in cyanobacteria differs from that in chloroplasts of algae. Consequently, during the symbiosis of a cyanobacterium to a eukaryotic cell, carotenoids may undergo considerable restructuring. Notably, since the name “myxoxanthophyll” does not specify the glycoside moieties, we have proposed the name of “myxol glycoside” to specify the glycosides, such as myxol 2′-α-l-fucoside, 4-ketomyxol 2′-rhamnoside, and oscillol dichinovoside [14,24]. The glycoside attached to myxol and oscillol is usually one of three methylpentoses: fucose, rhamnose, or chinovose. The carotenogenic pathways are discussed in Section 3.3.1. Most cyanobacteria contain only Chl a, whereas some species also contain additional Chl b, Chl d, or Chl f (Table 1).
This review describes both microalgae and macroalgae in seawater and freshwater environments without distinction since they share similar pigment compositions. Algae are commonly categorized into red algae, brown algae, and green algae, as described below. The carotenoids in algae are highly diverse and their compositions vary according to divisions and/or classes (Table 1).
In red algae (Rhodophyta), carotenoid composition particularly varies. These algae contain only Chl a as chlorophyll and can be classified into three types based on carotenoid composition and phylogenetics: an unicellular group, which contains only β-carotene and zeaxanthin (ZEA-type), a macrophytic group, which also includes additional antheraxanthin (ANT-type), and Bangiophyceae as well as others, which contain α-carotene and lutein in addition to ZEA-type carotenoids (LUT-type) (Table 1) [16,25]. The carotenogenic pathways are discussed in Section 3.3.2.
Cryptophyta contains Chl a, Chl c2, α-carotene, zeaxanthin, and alloxanthin (Table 1). The major carotenoid is acetylenic (C≡C) alloxanthin. In addition, Cryptophyta contains α-carotene and its unique acetylenic derivatives, crocoxanthin and monadoxanthin, which are found only in this division [16]. The carotenogenic pathways are discussed in Section 3.3.2.
Brown algae, including Heterokontophyta, Haptophyta, and Dinophyta, contain β-carotene and its derivatives (b-branch carotenoids) as well as Chl a and Chl c (Table 1, Figure 2). As Chl c pigments, Chl c1, Chl c2, and Chl c3 are known (Figure 1) and their distribution is specific to divisions and classes (Table 1). Brown algae contain unique acetylenic carotenoids, such as diadinoxanthin and diatoxanthin. Fucoxanthin and its derivatives are found only in four classes of Heterokontophyta (Chrysophyceae, Raphidophyceae, Bacillariophyceae, and Phaeophyceae), Haptophyta, and Dinophyta. Peridinin and its derivatives, as well as P457 [26], are unique to Dinophyta. Both fucoxanthin and peridinin are characterized by their unique structures (Figure 2) and class-specific distributions (Table 1) [2]. The carotenogenic pathways are discussed in Section 3.3.3. Notably, brown algae lack lutein and its derivatives (α-branch carotenoids), whereas Eustigmatophyceae lacks Chl c, fucoxanthin, diadinoxanthin, and diatoxanthin, but contains β-carotene, violaxanthin, and vaucheriaxanthin.
Green algae, including Euglenophyta, Chlorarachniophyta, and Chlorophyta, contain the same carotenoids, including β-carotene, violaxanthin, 9′-cis neoxanthin [12], and lutein, as well as Chl a and Chl b, all of which are present in land plants (Table 1, Figure 2). Some classes contain additional carotenoids, such as loroxanthin, siphonaxanthin, and prasinoxanthin, which are derivatives of lutein (α-branch carotenoids) and specific to particular classes. The carotenogenic pathways are discussed in Section 3.3.4. Notably, Euglenophyta lacks lutein and its derivatives (α-branch carotenoids), but contains diadinoxanthin and diatoxanthin.
Land plants, such as moss, fern, gymnosperm, and angiosperm, primarily contain only four types of carotenoids: β-carotene, violaxanthin, 9’-cis neoxanthin [12], and lutein. These carotenoids are located in the main synthetic pathways and are used for photosynthesis in the pigment-protein complexes (Table 1, Figure 2). The carotenogenic pathways are discussed in Section 3.3.5. Only Chl a and Chl b are present.
It is important to note that the identifications of some carotenoids may be incomplete because of insufficient analysis. Moreover, some algae names have been updated in response to developments in taxonomic technology and phylogenetic classification [27].
3. Carotenogenesis Pathways, Enzymes, and Genes
Carotenogenesis pathways and their enzymes have been primarily examined in cyanobacteria [14] and land plants among the oxygenic phototrophs [2,18,19]. The carotenogenesis pathways and enzyme characteristics for land plants are presented in Figure 2. Algae share common pathways with land plants but also contain additional algae-specific pathways, which have been proposed based on the chemical structures of carotenoids (Figure 2). Some common carotenogenesis genes in algae have been identified through gene homology, whereas many genes and enzymes specific to algal pathways remain unknown (Figure 2). In cyanobacteria, carotenoid compositions are different from those in algae and land plants. Therefore, their carotenogenesis pathways and enzymes are also different from those presented in Figure 2.
3.1. Lycopene Synthesis
3.1.1. Isopentenyl Pyrophosphate to Phytoene Synthesis
Isopentenyl pyrophosphate (IPP), a C5-compound, is the precursor for isoprenoids, terpenes, quinones, sterols, phytol of chlorophylls and bacteriochlorophylls, and carotenoids. IPP is synthesized via two distinct pathways: the classical mevalonate (MVA) pathway and the alternative, non-mevalonate, 1-deoxy-d-xylulose-5-phosphate (DOXP) pathway [28,29]. In the MVA pathway, acetyl-coenzyme A is converted to IPP through mevalonate, and the genes and enzymes involved have been characterized. This pathway occurs in the cytoplasm of algae and land plants, yeast, animals, and some bacteria, including green filamentous bacteria [28,30]. In contrast, the DOXP pathway was discovered in the 1990s, which involves the conversion of pyruvate and glyceraldehyde into IPP. The DOXP pathway is found in cyanobacteria, the plastids of algae and land plants, and certain bacteria, including purple bacteria, green sulfur bacteria, and E. coli. Although carotenoids are synthesized in plastids, the genes responsible for their synthesis are encoded in nuclear DNA. Unique to oxygenic phototrophs, Euglenophyta only contains the MVA pathway, whereas Chlorophyceae only has the DOXP pathway [28,29].
All carotenoids in oxygenic phototrophs contain eight IPP units. Farnesyl pyrophosphate (C15) is synthesized from three IPP units. Subsequently, one IPP unit is added to farnesyl pyrophosphate by geranylgeranyl pyrophosphate synthase (CrtE, GGPS) to yield geranylgeranyl pyrophosphate (C20). With a head-to-head condensation of the two C20-compounds, the first carotene, phytoene (C40), is synthesized by phytoene synthase (CrtB, Pys, Psy) using ATP [31,32]. This pathway was confirmed by cloning the genes from two species of Rhodobacter capsulatus and Cereibacter sphaeroides (previously Rhodobacter, purple bacteria), and two species of Pantoea [31,32,33]. Among the oxygenic phototrophs, the functions of CrtE and CrtB have also been confirmed. CrtE occurs in Thermosynechococcus elongates [34], Porphyra umbilicalis [35], Euglena gracilis [36], and Arabidopsis thaliana [37], and CrtB is present in Gloeobacter violaceus [38], Phaeodactylum tricomutum [39], Chlamydomonas reinhardtii [40], Haematococcus pluvialis [41], and Euglena gracilis [36]. The crtE and crtB genes exhibit high sequence homology from bacteria to land plants, respectively [2].
3.1.2. Phytoene to Lycopene Synthesis
Four desaturation steps are needed for the conversion of phytoene to lycopene. Oxygenic phototrophs, including cyanobacteria, algae, and land plants, require four enzymes for this process: phytoene desaturase (plant type) (CrtP, Pds), ζ-carotene desaturase (CrtQ, Zds), ζ-carotene isomerase (Z-ISO), and cis-carotene isomerase (CrtH, CrtISO). CrtP catalyzes the first two desaturation steps from phytoene to ζ-carotene through phytofluene. CrtQ then catalyzes two additional desaturation steps from ζ-carotene to lycopene through neurosporene. During desaturation by CrtP, 9,1100’-tri-cis ζ-carotene is produced and subsequently converted to 9,9’-di-cis ζ-carotene by Z-ISO. This reaction is also catalyzed with light, non-enzymatically. In addition, during the desaturation by CrtQ, neurosporene and lycopene are isomerized to poly-cis forms, followed by isomerization to all-trans forms by CrtH. Light is also effective for their photoisomerization to all-trans forms, non-enzymatically. The functions of these enzymes have been mainly established in cyanobacteria, green algae, and land plants: CrtP from Synechocystis sp. PCC 6803 [42], Synechococcus elongatus PCC 7942 [43], Phaeodactylum tricornutum [39], Euglena gracilis [44], Chlamydomonas reinhardtii [45], and Chlorella zofingiensis [46], CrtQ from Anabaena sp. PCC 7120 (CrtQa, crtI-like sequence) [47], Synechocystis sp. PCC 6803 (CrtQb, plant crtQ-like) [48], Phaeodactylum tricornutum [39] and Euglena gracilis [44], Z-ISO from Arthrospira and Euglena [49] and Arabidopsis and Zea [50], and CrtH from Synechocystis sp. PCC 6803 [51,52], Arabidopsis [53], and Lycopersicon [54]. CrtP of S. elongatus PCC 7942 is stimulated by NAD(P) and oxygen as a potential final electron acceptor [55]. CrtQa shows sequence homology with bacterial phytoene desaturase (CrtI) and CrtH, whereas CrtQb shows sequence homology with CrtP. In addition, the genes homologous to crtQa are not found in cyanobacteria. Therefore, among the oxygenic phototrophs, Anabaena sp. PCC 7120 is the only species with functional CrtQa.
In contrast, the bacterial type requires only one enzyme, phytoene desaturase (bacterial type) (CrtI), to convert from phytoene to lycopene. Notably, this type of CrtI is only found in the primitive cyanobacterium Gloeobacter violaceus, whereas the homologous genes of crtP, crtQ and crtH are not present in the genome [38,56]. Among anoxygenic phototrophs, purple bacteria, green filamentous bacteria, and heliobacteria use CrtI, whereas green sulfur bacteria use CrtP, CrtQ, and CrtH, but do not use Z-ISO [7,49,57]. CrtI shares amino acid sequence homology with CrtP, CrtQ, CrtH, CrtO, and CrtD.
3.2. β-Carotene and α-Carotene Synthesis by Lycopene Cyclases
All carotenoids in oxygenic phototrophs are derived from lycopene; dicyclic carotenoids, β-carotene and α-carotene (β- and α-branch carotenoids, respectively), and their derivatives (Figure 2). In cyanobacteria, exceptionally, myxol methylpentosides and oscillol di-methylpentosides are present as monocyclic and acyclic carotenoids, respectively. C30- and C50-carotenoids and derivatives of neurosporene are not found in oxygenic phototrophs.
Lycopene is cyclized into either β-carotene through γ-carotene, or α-carotene through δ-carotene. Three distinct families of lycopene cyclases have been identified in carotenogenic organisms [14,58,59,60]. One large family includes CrtY, which is present in some bacteria except cyanobacteria and green sulfur bacteria, and CrtL (CrtL-b, Lcy-b), which is found in certain cyanobacteria, algae, and land plants. Lycopene ε-cyclases (CrtL-e, Lcy-e) from green algae and land plants and lycopene β-monocyclases (CrtYm, CrtLm) from bacteria are also included. The amino acid sequences for these lycopene cyclases contain five significantly conserved regions [58,61,62] and have an NAD(P)/FAD-binding motif [63]. Note that Sandmann [58,61] and Takaichi [60] consider these enzymes belong to the same family, whereas Bryant [59] classes them into two separate CrtY and CrtL families.
Two cyanobacteria also express CrtL-type enzymes. Synechococcus elongatus PCC 7942 contains a functional CrtL [64]. Prochlorococcus marinus MED4 contains two lycopene cyclases, both of which have sequence homology to CrtL. CrtL-b only exhibits lycopene β-cyclase activity, whereas CrtL-e is a bifunctional enzyme with both lycopene ε-cyclase and lycopene β-cyclase activities. The combination of these two cyclases enables the production of β-carotene, α-carotene and ε-carotene. Both enzymes may have originated from the duplication of a single gene [65]. The characteristics of this CrtL-e differ somewhat from those in land plants [66]. P. marinus produces the usual (6R)-α-carotene, whereas Acaryochloris marina MBIC 11017 produces (6S)-α-carotene, which displays the opposite chirality to the usual (6R)-α-carotene and contains only one crtL-like gene based on its genome sequence [15]. Among algae, certain CrtL enzymes have been functionally confirmed including Cyanidioschyzon merolae [67], Phaeodactylum tricomutum [39], Dunaliella salina [62], and Haematococcus pluvialis [68]. CrtL-b and CrtL-e from Porphyra umbilicalis [69] and Chloromochloris zofingliensis [70] also have these functions.
The second family of lycopene cyclases includes a heterodimer consisting of CrtYc and CrtYd found in bacteria, a fused monomer (CrtYc-Yd) present in both bacteria and archaea, and a fused 39bifunctional CrtYB found in fungi [71,72,73], but not in phototrophs.
A new family of functional lycopene cyclases, CruA, has been found in Chlorobaculum tepidum (green sulfur bacterium). Homologous genes, cruA and cruP, have also been in the genome of Synechococcus sp. PCC 7002, with their primary product being γ-carotene when expressed in lycopene-producing E. coli [59]. However, Bradbury et al. [74] was not able to detect cruP activity in Synechococcus sp. PCC 7002 in lycopene-producing E. coli. More recently, Sugiyama et al. [75] demonstrated that CruA from Arthrospira platensis exhibits lycopene cyclase activity in lycopene-producing E. coli. In addition, Xiong et al. [76] demonstrated that cruA from Synechocystis sp. PCC 6803 exhibits lycopene cyclase activity in the cruA-less mutant of Synechococcus sp. PCC 7002, which requires bound Chl a; however, the functions of bound Chl a are unclear.
The homologous genes cruA and cruP are widely distributed across the genomes of certain cyanobacteria; however, confirmation of the activities of these cruA- and cruP-like genes remains limited, as noted above. Phylogenetic analysis of functional CruA, CruP, and CrtL-type lycopene cyclases, along with their homologs in cyanobacteria, revealed two distinct clusters [49], however, further studies are needed to determine the distribution and functionality of lycopene cyclases, such as CrtL-, CruA-, and CruP-like types, in cyanobacteria.
The distribution of α-carotene and its derivatives (α-branch carotenoids), such as CrtL-e, is restricted to certain red algae, green algae, and land plants (Table 1). The characteristics of the genes and enzymes of CrtL-e have been investigated and they show sequence homology with crtL-b. Lycopene is initially converted to δ-carotene by CrtL-e, and then to α-carotene by CrtL-b. γ-Carotene produced by CrtL-b is not a suitable substrate for CrtL-e [60,66]. Their functions have been confirmed in red alga Porphyra umbilicalis [69], green alga Chloromochloris zofingliensis [70], and some species of land plants [66],
3.3. Xanthophyll Synthesis
3.3.1. Cyanobacteria
Most cyanobacteria produce zeaxanthin, whereas some produce both zeaxanthin and nostoxanthin (Figure 3, hydroxy pathway). The hydroxy group is first introduced into the C-3 and C-3′ positions of β-carotene by β-carotene hydroxylase (CrtR) to produce zeaxanthin via β-cryptoxanthin. Subsequently, at the C-2 and C-2′ positions of zeaxanthin, the hydroxy group is introduced by 2,2′-β-hydroxylase (CrtG) to produce nostoxanthin via caloxanthin [17,77,78,79,80,81]. The same enzymes, CrtR and CrtG, can also introduce hydroxy groups to deoxymyxol and myxol to produce myxol and 2-hydroxymyxol, respectively (myxol pathway) [78,81]. Thus, CrtR and CrtG are involved in two different pathways. CrtR is a non-heme di-iron enzyme, whereas CrtG is a cytochrome P450-type enzyme, but do not share sequence homology [14].
Cyanobacteria contain some ketocarotenoids, including echinenone, canthaxanthin, 3’-hydroxyechinenone, and 4-ketomyxol. Two distinct β-carotene ketolases, CrtO and CrtW, have been identified. Although both enzymes catalyze ketolation of the β-end group, they have different characteristics (Figure 3, keto pathway) [14]. CrtO is almost twice the size of CrtW and shows little amino acid sequence homology with CrtW. CrtO usually converts β-carotene to echinenone; however, the final product of canthaxanthin is low or absent in Synechocystis sp. PCC 6803, Anabaena sp. PCC 7120, and Gloeobacter violaceus [38,80,82,83]. In contrast, CrtW introduces one or two keto groups into β-carotene, zeaxanthin, and myxol to produce canthaxanthin, astaxanthin, and 4-ketomyxol, respectively, in Anabaena sp. PCC 7120, Nostoc punctiforme, and Gloeobacter violaceus [38,56,80,83,84]. Taken together, these ketolases function in different pathways, β-carotene and myxol, depending on the species [14,80].
The pathways and the enzymes involved in the production of the right half of myxol 2′-methylpentoside remain unclear (Figure 3, myxol pathway) [2,14]; however, two enzymes from Synechococcus sp. PCC 7002 have been functionally confirmed: carotene 1’,2’-hydratase (CruF) and glycosyltransferase (CruG) [85].
Synechoxanthin contains an aryl end group with a carboxy group, and both structures are rare in carotenoids (Figure 3, synechoxanthin pathway). The chemical structure has been identified in 2008 [86]. Some cyanobacteria, such as Synechococcus sp. PCC 7002, Synechocystis sp. PCC 6803, and Anabaena sp. PCC 7120, produce this carotenoid; however, its distribution in cyanobacteria has not been fully examined. It is produced from β-carotene, and two biosynthetic enzymes (CruE and CruH) have been functionally identified, although one is unknown [87].
3.3.2. Red Algae
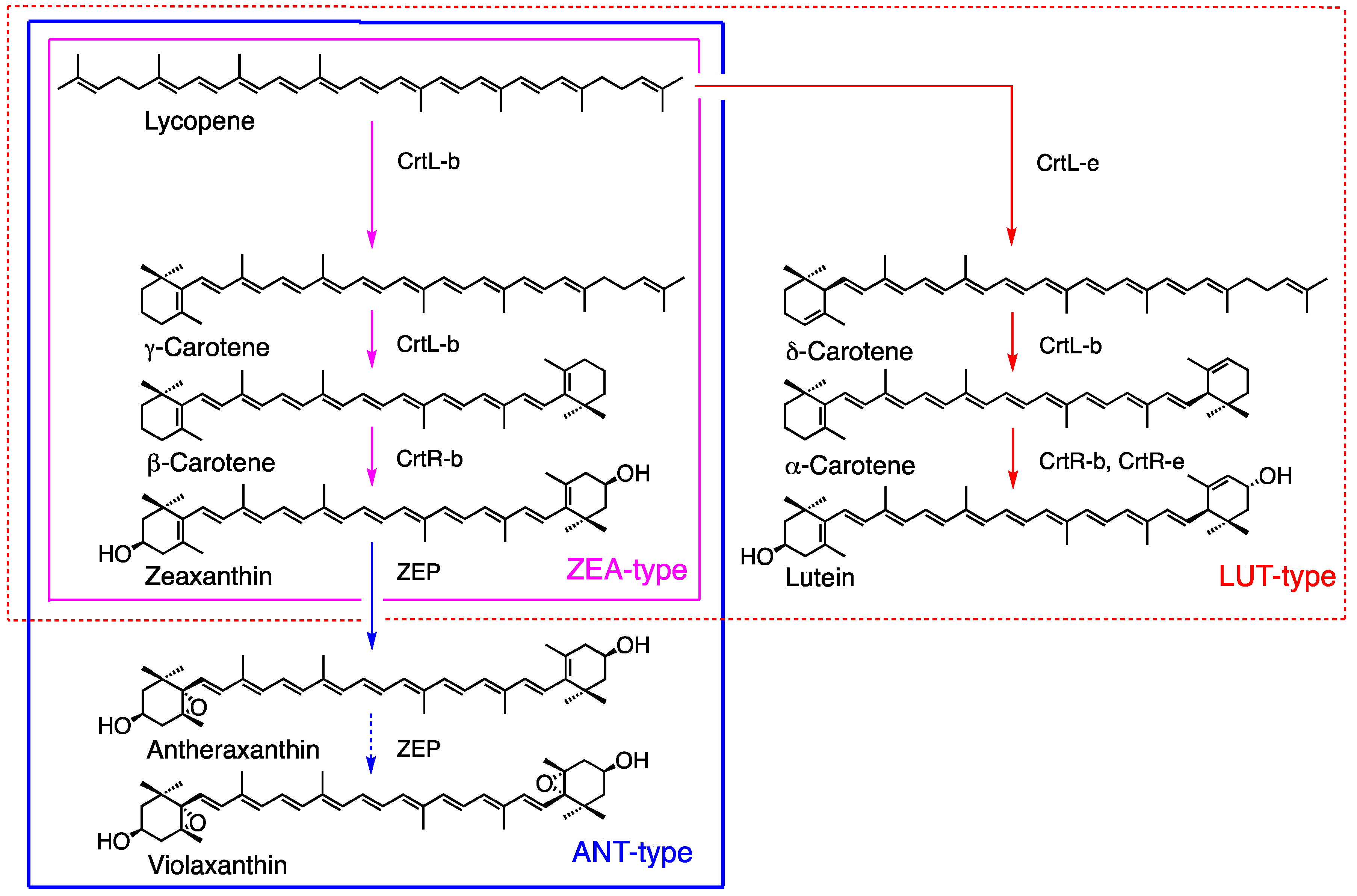
Red algae (Rhodophyta) produce a wide variety of carotenoids and may be categorized into three types based on carotenoid composition and phylogenetics, including the ZEA-type (Figure 4, magenta line), ANT-type (blue line), and LUT-type (red line) as described in Section 2 (Table 1) [16,25]. The major carotenoids of the ZEA-type are β-carotene and zeaxanthin, and this type contains homologous genes, such as crtL-b and crtR-b, which are involved in zeaxanthin synthesis. The ANT-type includes antheraxanthin in addition to the ZEA-type and features the enzyme of zeaxanthin epoxidase ZEP for antheraxanthin synthesis, whose function is confirmed in Madagascaria erythrocladioides [88]. Antheraxanthin is the major carotenoid, whereas violaxanthin is either absent or present in minor amounts, and it may have an important role in photosynthesis. The LUT-type is characterized by the presence of α-carotene and lutein and includes crtL-e and crtR-e for lutein biosynthesis (Figure 4). The function of the P450-type β-carotene hydroxylase (CYP97) has been confirmed in Porphyra umbilicalis [89].
Figure 4.
Carotenogenesis pathways in red algae categorized into three types based on carotenoids and phylogeny [16].
Figure 4.
Carotenogenesis pathways in red algae categorized into three types based on carotenoids and phylogeny [16].
Cryptophyta contains α-carotene, zeaxanthin, and alloxanthin (Table 1). In organic chemistry, the C-7,8 and C-7’,8’ double bonds of zeaxanthin can be oxidized to form the triple bond (acetylenic group, C≡C) present in alloxanthin [18]. Therefore, in Cryptophyta, alloxanthin may be enzymatically synthesized from zeaxanthin since there are no epoxy and allenic carotenoids as diadinoxanthin (Figure 2).
3.3.3. Brown Algae
Brown algae, including Heterokontophyta, Haptophyta, and Dinophyta, primarily contain carotenoids, such as β-carotene, zeaxanthin, diadinoxanthin, fucoxanthin, and peridinin, depending on the divisions and classes. Notably, the α-branch carotenoids are absent. Little is known regarding the carotenogenesis pathways of brown algae; however, some have been proposed based on the chemical structures of the carotenoids present, and the functions of some enzymes have been confirmed as below (Table 1, Figure 2).
In the cell-free preparations of Amphidinium carterae (Dinophyta), 14C-labeled zeaxanthin was converted into allenic neoxanthin, which was then converted into acetylenic (C≡C) diadinoxanthin and C37 peridinin (Figure 2). Moreover, the three carbon atoms at C-13′,14′,20′ of peridinin are eliminated from neoxanthin (C-13,14,20) [90,91]. Allenic carotenoids (C=C=C) are rare in algae. Based on their chemical structures, all-trans neoxanthin may be converted into fucoxanthin, diadinoxanthin, dinoxanthin, peridinin, and vaucheriaxanthin; however, the specific pathways and enzymes involved are still unclear (Figure 2). Diadinoxanthin and diatoxanthin have also contain a triple bond (acetylene group) and may be synthesized from the epoxy and allene groups of neoxanthin since they are asymmetric structures. Consequently, the formation of the acetylene group in diadinoxanthin in brown algae and alloxanthin in Cryptophyta occur differently (Figure 2).
For fucoxanthin synthesis, a simplified biosynthetic pathway involving ketolation and acetylation of trans neoxanthin has been proposed based on chemical structures (Figure 5, blue line); however, the specific genes and enzymes have not yet been identified. Recently, an alternative pathway has been discovered in Phaeodactylum tricomutum (Figure 5). In this pathway, β-carotene is hydroxylated to zeaxanthin by the P450-type enzyme PtCYP97B2 [92]. Zeaxanthin and diatoxanthin are epoxidated to violaxanthin and diadinoxanthin by the epoxidases PtZEP2 and PtZEP3, respectively [93,94]. Subsequently, both violaxanthin and diadinoxanthin are converted back to zeaxanthin and diatoxanthin, respectively, by the same violaxanthin de-epoxidase (PtVDE), which is activated under high-light conditions [95]. These processes are known as the violaxanthin cycle and diadinoxanthin cycle, respectively [96]. The violaxanthin de-epoxidase-like1 (PtVDL1) enzyme from P. tricumutum and some related species exhibits neoxanthin synthase activity in violaxanthin-producing E. coli; however, its homologous gene is absent in land plants [95,97]. Neoxanthin is then dehydrated to diadinoxanthin by an unknown dehydratase. Diadinoxanthin is converted to allenoxanthin by violaxanthin de-epoxydase-like2 (PtVDL2), whose function is confirmed using a green mutant of P. tricumutum [98]. Allenoxanthin is subsequently acetylated to form haptoxanthin. The final hydration or ketolation step is catalyzed by PtCrtISO5 [99]. The specific dehydrase and acetylase involved have not yet been unidentified. In addition, homologous genes to VDL1 and VDL2 are found only in a limited number of brown algal species. Therefore, the universality of this pathway for fucoxanthin production in brown algae has not yet been confirmed.
3.3.4. Green Algae
Green algae encompass several divisions, such as Euglenophyta, Chlorarachniophyta, and Chlorophyta (Table 1). Their carotenoids primarily include β-carotene, zeaxanthin, violaxanthin, 9’-cis neoxanthin, lutein, and lutein derivatives. Functionally confirmed enzymes have mainly been identified in Chlorophyceae including Chlorella, Chlamydomonas, Dunaliella, and Haematococcus for CrtB, CrtP, CrtL-b, CrtR-b, ZEP, VED, and CrtW for the β-branch carotenoids. The α-branch carotenoids, such as siphonaxanthin [13], loroxanthin, prasinoxanthin, and monadoxanthin, may be derived from lutein based on their chemical structures, although the specific pathways and enzymes remain unknown (Figure 2, Table 1). Recently, an intermediate of siphonaxanthin has been identified, and siphonaxanthin may be synthesized from lutein via loroxanthin and/or deoxysiphonaxanthin [100]. Functionally confirmed enzymes include CrtR-b from Haematococcus pluvialis [101], ZEP from Chlamydomonas reinhardtii [102], and VDE from Mantonilla squamata [103].
Under stressful conditions, including high-light irradiation, UV irradiation, and nutrient stress, some Chlorophyceae, such as Haematococcus, Chlorella, and Scenedesmus, accumulate ketocarotenoids, such as canthaxanthin and astaxanthin. These carotenoids are synthesized by a combination of CrtR-b and β-carotene ketolase (CrtW, BKT) [104,105,106,107,108]. Notably, while the β-carotene ketolase in Haematococcus and Chlorella was initially named CrtO [104,106,107], it is actually CrtW-type, not CrtO-type, based on amino acid sequences.
3.3.5. Land Plants
Land plants include moss, fern, gymnosperm, and angiosperm, and primarily contain only four types of carotenoids: β-carotene, violaxanthin, 9’-cis neoxanthin [12], and lutein. These carotenoids are located in the main synthetic pathways (Figure 2).
In land plants, most of the carotenogenesis pathways and the functionally confirmed enzymes are known (Figure 2). Hydroxy groups are introduced into β-carotene to produce zeaxanthin by β-carotene hydroxylase (CrtR, CrtR-b, BCH). Arabidopsis thaliana contains four hydroxylases. β-Carotene is hydroxylated mainly by the non-heme di-iron enzymes, BCH1 and BCH2 (CrtR-b), to produce zeaxanthin, whereas α-carotene is primarily hydroxylated by the cytochrome P450-type enzymes, CYP97A3 for the β-end group and CYP97C1 for the β- and ε-end groups, which results in the production of lutein [109]. Epoxy groups are introduced into zeaxanthin by zeaxanthin epoxidase (ZEP, NPQ) to produce violaxanthin through antheraxanthin. Under high-light conditions, violaxanthin is converted back to zeaxanthin by violaxanthin de-epoxidase (VDE) to disperse the excess energy from the excited chlorophylls. This process is known as the violaxanthin cycle as described above. One end group of violaxanthin is converted to an allene group in neoxanthin by neoxanthin synthase (NSY). Three proteins from land plants and one from P. tricumutum [95,97] have been reported to have NSY activity; however, their homologous genes are not distributed in all land plants and brown algae. Since neoxanthin in chloroplasts exists exclusively in the 9′-cis form, unknown 9′-isomerase for all trans neoxanthin to 9′-cis neoxanthin should be present [12].
The structures of the carotenoids are also shown in Figure 6.
4. Function of Carotenoids
For photosynthesis, most carotenoids and chlorophylls are bound to peptides to form the photosynthetic pigment-protein complexes and super complexes located in the thylakoid membrane of phototrophs. Certain complexes have been isolated from cyanobacteria, some algae, and land plants such as photosystem I (PSI), light-harvesting complex I (LHCI), PSI-LHCI super complex, photosystem II (PSII), light-harvesting complex II (LHCII), PSII-LHCII super complex, and the cytochrome b6f complex between PSII and PSI. Their 3D structures, characteristics, functions including excitation-energy transfer, and the pigment compositions have been examined. The primary functions of carotenoids are light-harvesting and then transferring the energy to Chls, protection of the Chls and peptides from high-light radiation, and the assembly and stability of the pigment-protein complexes. Fucoxanthin-chlorophyll a/c-proteins (FCP) from brown algae [110], violaxanthin-chlorophyll-a-proteins (VCP) from Nannochloropsis (Eustigmatophyceae) [111], PSI-LHCI supercomplex from Chlorella sorokinian [112], and PSII-LHCII supercomplex from Chaetoeros gracilis [113] have been isolated and characterized.
β-Carotene is presented in the most divisions of the PSI and LHCI as well as the PSII and core LHCII. Zeaxanthin is exceptionally presented in some red algae of the LHCI. Whereas, in the peripheral LHCII, the bound carotenoids are heterogeneous depending on the classes. The major carotenoids are alloxanthin (Cryptophyta), fucoxanthin (Chrysophyceae, Raphidophyceae, Bacillariophyceae, Phaeophyceae, and Haptophyta), diadinoxanthin and vaucheriaxanthin (Xanthophyceae), violaxanthin and vaucheriaxanthin (Eustigmatophyceae), peridinin (Dinophyta), diadinoxanthin (Euglenophyta), siphonaxanthin (Chlorophyceae and Ulvophyceae), and lutein, violaxanthin, and 9′-cis neoxanthin (land plants) (Figure 2, Table 1). β-Carotene in both PSI and PSII may exert protective functions, and carotenoids in the peripheral LHCII may have light-harvesting functions. Myxol glycosides and some carotenoids are located in the cytoplasmic membrane, stroma, or lumen to protect from the high-light in phototrophs, whereas astaxanthin is accumulated in the cytoplasm of certain green algae under high-light conditions.
The keto groups at C-8 of fucoxanthin [114], siphonaxanthin [115,116], and prasinoxanthin [117], which are found only in algae, represent the single-bond trans-conformation for the conjugated double bond (Figure 2). From the femtosecond time-resolved fluorescence spectroscopy of the purified carotenoids in organic solvents and the LHC in solution, these ketocarotenoids along with peridinin transfer the harvested energy highly efficiently from the S1 state, but not the S2 state, of carotenoids to Chls. A comparison of the structures of other carotenoids revealed that these keto groups are essential for high transfer efficiency [118,119]. These ketocarotenoids may primarily have light-harvesting functions.
The water-soluble peripheral LHC of peridinin-chlorophyll-protein (PCP) isolated from Amphidinium carterae and Symbiodinium sp. CS-156 (Dinophyta) exhibit a trimeric structure, and the monomer contains eight peridinin and two Chl a molecules [120,121]. The water-soluble orange carotenoid protein (OCP) isolated from the cyanobacterium Arthrospira maxima forms a homodimer with two 3′-hydroxyechinenone molecules [122]. OCP is also produced in some cyanobacteria and its function might regulate the energy dissipation from phycobilisomes to PSII [123,124].
The xanthophyll cycle has a role in protecting from the potentially harmful effects of excess light by enhancing the dissipation of excess energy as heat. It may be divided into the violaxanthin and diadinoxanthin cycles. The violaxanthin cycle is the cyclical interconversion of violaxanthin, antheraxanthin, and zeaxanthin in green algae and land plants, whereas the diadinoxanthin cycle is that of diadinoxanthin and diatoxanthin in brown algae (Figure 2) [96,125]. These have been primarily investigated in land plants. Zeaxanthin epoxidase (ZEP) converts zeaxanthin to violaxanthin through antheraxanthin during biosynthesis. Violaxanthin is found in the peripheral LHC of PSII. Under high-light conditions, the lumen inside of the thylakoid, is acidified by proton movement to the lumen, and then violaxanthin de-epoxidase (VDE) in the lumen is activated by acidic pH and ascorbate. VDE catalyzes the de-epoxidation of violaxanthin to zeaxanthin through antheraxanthin. Zeaxanthin dissipates excess energy from excited Chls. ZEP from Chlorophyceae Chlamydomonas reinhardtii [102,126] and Chlorella zofingiensis [127], and VDE from Pracinophyceae Mantonilla squamata [103] and C. reinhardtii [126] have been functionally confirmed. Similarly, the diadinoxanthin cycle occurs in Heterokontophyta, Haptophyta, and Dinophyta, which contain diadinoxanthin and diatoxanthin. The diadinoxanthin and diatoxanthin contents are not normally low and a portion may be used for the diadinoxanthin cycle. For the brown alga of Phaeodactylum tricomutum, functional zeaxanthin and diatoxanthin cycles are discussed above (Section 3.3.3).
Certain carotenoids are essential for the assembly and structural stability of pigment-protein complexes, such as Synechocystis sp. PCC 6803 and Thermosynechococcus elongatus (cyanobacteria) [128,129], whereas lutein is essential for LHCII of land plants.
Water-soluble astaxanthin-binding glycoprotein (AstaP) and its homologs are widely distributed in Scenedesmaceae. They are accumulated under stress conditions, such as high-salt concentration and high-light irradiation. They may have scavenging activity for reactive oxygen species, particularly 1O2, and may function as a sunscreen [130,131].
References
- Jeffrey, S.W.; Wright, S.W.; Zapata, M. Chlorophylls and carotenoids. In Phytoplankton Pigments; Roy, S., Llewellyn, C.A., Egeland, E.S., Johnsen, G., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 3–77. [Google Scholar]
- Takaichi, S. Carotenoids in carotenogenic organisms: distribution, biosynthesis, and functions. Ramawat, K.G.; Mérillon, J.M. (Eds.) Natural Products, Springer, Berlin, Germany. 2024; In press.
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Handbook; Birkhäuser: Basel, Switzerland, 2004. [Google Scholar]
- Liaaen-Jensen, S. Marine carotenoids. New J. Chem. 1990, 14, 747–759. [Google Scholar]
- Liaaen-Jensen, S. Carotenoids in chemosystematics. In Carotenoids: Biosynthesis and Metabolism; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 1998; Volume 3, pp. 217–247. [Google Scholar]
- Mackey, M.D.; Mackey, D.J.; Higgins, H.W.; Wright, S.W. CHEMTAX-a program for estimating class abundances from chemical markers: Application to HPLC measurements of phytoplankton. Mar. Ecol. Prog. Ser. 1996, 144, 265–283. [Google Scholar] [CrossRef]
- Shimada, K.; Takaichi, S. Anoxygenic Phototrophic Bacteria; Academic Press, London, UK, 2024.
- Bryant, D.A.; Hunter, C.N.; Warren, M.J. Biosynthesis of the modified tetrapyrroles—the pigments of life. J. Biol. Chem. 2020, 295, 6888–6925. [Google Scholar] [CrossRef]
- Jiang, Y.; Cao, T.; Yang, Y.; Zhang, H.; Zhang, J.; Li, X. A chlorophyll c synthase widely co-opted by phytoplankton. Science 2023, 382, 92–98. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in algae: Distribution, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- 11Dembitsky, V.M.; Maoka, T. Allenic and cumulenic lipids. Prog. Lipid Res. 2007, 46, 328–375. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Mimuro, M. Distribution and geometric isomerism of neoxanthin in oxygenic phototrophs: 9′-cis, a sole molecular form. Plant Cell Physiol. 1998, 39, 968–977. [Google Scholar] [CrossRef]
- Yoshii, Y.; Takaichi, S.; Maoka, T.; Suda, S.; Sekiguchi, H.; Nakayama, T.; Inouye, I. Variation of siphonaxanthin series among the genus Nephroselmis (Prasinophyceae, Chlorophyta), including a novel primary methoxy carotenoid. J. Phycol. 2005, 41, 827–834. [Google Scholar] [CrossRef]
- Takaichi, S.; Mochimaru, M. Carotenoids and carotenogenesis in cyanobacteria: Unique ketocarotenoids and carotenoid glycosides. Cell. Mol. Life Sci. 2007, 64, 2607–2619. [Google Scholar] [CrossRef]
- Takaichi, S.; Mochimaru, M.; Uchida, H.; Murakami, A.; Hirose, E.; Maoka, T.; Tsuchiya, T.; Mimuro, M. Opposite chilarity of α-carotene in unusual cyanobacteria with unique chlorophylls, Acaryochloris and Prochlorococcus. Plant Cell Physiol. 2012, 53, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Yokoyama, A.; Mochimaru, M.; Uchida, H.; Murakami, A. Carotenogenesis diversification in phylogenetic lineages of Rhodophyta. J. Phycol. 2016, 52, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, J.; Chamovitz, D. Carotenoids in cyanobacteria. In The Molecular Biology of Cyanobacteria; Bryant, D.A. Ed.: Kluwer, Dordrecht, The Netherland, 1994; pp. 559–579. [Google Scholar]
- Britton, G. Overview of carotenoid biosynthesis. In Carotenoids: Biosynthesis and Metabolism; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser: Basel, Switzerland, 1998; Volume 3, pp. 13–147. [Google Scholar]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of coevolution towards plant photosynthesis. New Phytologist 2021, 232, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F.; Bryant, D.A.; Beatty, J.T. A physiological perspective on the origin and evolution of photosynthesis. FEMS Microbiol. Rev. 2018, 42, 205–231. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, A.; Tsukatani, Y.; Azai, C.; Nobu, M. Illuminating the coevolution of photosynthesis and bacteria. Proc. Natl. Acad. Sci. USA 2024, 121, e2322120121. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Membrane heredity and early chloroplast evolution. Trends Plant Sci. 2000, 5, 174–182. [Google Scholar] [CrossRef] [PubMed]
- McFadden, G.I. Primary and secondary endosymbiosis and the origin of plastids. J. Phycol. 2001, 37, 951–959. [Google Scholar] [CrossRef]
- Takaichi, S.; Maoka, T.; Masamoto, K. Myxoxanthophyll in Synechocystis sp. PCC 6803 is myxol 2′-dimethyl-fucoside, (3R,2′S)-myxol 2′-(2,4-di-O-methyl-α-l-fucoside), not rhamnoside. Plant Cell Physiol. 2001, 42, 756–762. [Google Scholar] [CrossRef]
- Schubert, N.; García-Mendoza, E. Photoinhibition in red algal species with different carotenoid profiles. J. Phycol. 2008, 44, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Wakahama, T.; Laza-Martinez, A.; Taha, A. I,B.H.M.; Okuyama, H.; Yoshida, K.; Kogame, K.; Awai, K.; Kawachi, M.; Maoka, T.; Takaichi, S. Structural confirmation of a unique carotenoid lactoside, P457, in Symbiodinium sp. strain NBRC 104787 isolated from a sea anemone and its distribution in dinoflagellates and various marine organisms. J. Phycol. 2012, 48, 1392–1402. [Google Scholar]
- Takaichi, S. General methods for identification of carotenoids. Biotechnol. Lett. 2014, 36, 1127–1128. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. The 1-deoxy-d-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Eisenreich, W.; Bacher, A.; Arigoni, D.; Rohdich, F. Biosynthesis of isoprenoids via the non-mevalonate pathway. Cell. Mol. Life Sci. 2004, 61, 1401–1426. [Google Scholar] [CrossRef] [PubMed]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoid biosynthesis in microorganisms and plants. Eur. J. Biochem. 1994, 223, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.A. Genetics of eubacterial carotenoid biosynthesis: A colorful tale. Annu. Rev. Microbiol. 1997, 51, 629–659. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Nakagawa, M.; Kobayashi, K.; Yamano, S.; Izawa, Y.; Nakamura, K.; Harashima, K. Elucidation of the Erwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J. Bacteriol. 1990, 172, 6704–6712. [Google Scholar] [CrossRef]
- Ohto, C.; Ishida, C.; Nakane, H.; Muramatsu, M.; Nishino, T.; Obata, S. A thermophilic cyanobacterium Synechococcus elongatus has three different Class I prenyltransferase genes. Plant Mol. Biol. 1999, 40, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Huang, X. : Lu, Q.; Zhu, J.; Lu, S. Cloning and characterization of the geranylgeranyl diphosphate synthase (GGPS) responsible for carotenoid biosynthesis in Pyropia umbilicalis. J. Appl. Phycol. 2016, 28, 671–678. [Google Scholar] [CrossRef]
- Kato, S.; Takaichi, S.; Ishikawa, T.; Asahina, M.; Takahashi, S.; Shinomura, T. Identification and functional analysis of the geranylgeranyl pyrophosphate synthase gene (crtE) and phytoene synthase gene (crtB) for carotenoid biosynthesis in Euglena gracilis. BMC Plant Biol. 2016, 67, e4. [Google Scholar]
- Beck, G.; Coman, D.; Herren, E.; Ruiz-Sola, M.Á.; Rodríguez-Concepción, M.; Gruissem, W.; Vranová, E. Characterization of the GGPP synthase gene family in Arabidopsis thaliana. Plant Mol. Biol. 2013, 82, 393–416. [Google Scholar] [CrossRef]
- Steiger, S.; Jackisch, Y.; Sandmann, G. Carotenoid biosynthesis in Gloeobacter violaceus PCC4721 involves a single crtI-type phytoene desaturase instead of typical cyanobacterial enzymes. Arch. Microbiol. 2005, 184, 207–214. [Google Scholar] [CrossRef]
- Dambek, M.; Eilers, U.; Breitenbach, J.; Steiger, S.; Buchel, C.; Sandmann, G. Biosynthesis of fucoxanthin and diadinoxanthin and function of initial pathway genes in Phaeodactylum tricornutum. J. Experi. Botany 2012, 63, 5607–5612. [Google Scholar] [CrossRef]
- McCarthy, S.S.; Kobayashi, M.C.; Niyogi, K.K. White mutants of Chlamydomonas reinhardtii are defective in phytoene synthase. Genetics 2004, 168, 1249–1257. [Google Scholar]
- Steinbrenner, J.; Linden, H. Regulation of two carotenoid biosynthesis genes coding for phytoene synthase and carotenoid hydroxylase during stress-induced astaxanthin formation in the green alga Haematococcus pluvialis. Plant Physiol. 2001, 125, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Férez, I.M.; Vioque, A. Nucleotide sequence of the phytoene desaturase gene from Synechocystis sp. PCC 6803 and characterization of a new mutation which confers resistance to the herbicide norflurazon. Plant Mol. Biol. 1992, 18, 981–983. [Google Scholar] [CrossRef]
- Chamovitz, D.; Misawa, N.; Sandmann, G.; Hirschberg, J. Molecular cloning and expression in Escherichia coli of a cyanobacterial gene coding for phytoene synthase, a carotenoid biosynthesis enzyme. FEBS Lett. 1992, 296, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Tanno, Y.; Takaichi, S.; Shinomura, T. Low temperature stress alters the expression of phytoene desaturase genes (crtP1 and crtP2) and the -carotene desaturase gene (crtQ) together with the cellular carotenoid content of Euglena gracilis. Plant Cell Physiol. 2019, 60, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Vila, M.; Couso, I.; León, R. Carotenoid content in mutants of the chlorophyte Chlamydomonas reinhardtii with low expression levels of phytoene desaturase. Process Biochem. 2008, 43, 1147–1152. [Google Scholar] [CrossRef]
- Huang, J.; Liu, J.; Li, Y.; Chen, F. Isolation and characterization of the phytoene desaturase gene as a potential selective marker for genetic engineering of the astaxanthin-producing green alga Chlorella zofingiensis (Chlorophyta). J. Phycol. 2008, 44, 684–690. [Google Scholar] [CrossRef]
- Linden, H.; Vioque, A.; Sandmann, G. Isolation of a carotenoid biosynthesis gene coding for ζ-carotene desaturase from Anabaena PCC 7120 by heterologous complementation. FEMS Microbiol. Lett. 1993, 106, 99–104. [Google Scholar]
- Breitenbach, J.; Fernández-González, B.; Vioque, A.; Sandmann, G. A higher-plant type ζ-carotene desaturase in the cyanobacterium Synechocystis PCC6803. Plant Mol. Biol. 1998, 36, 725–732. [Google Scholar] [CrossRef]
- Sugiyama, K.; Takahashi, K.; Nakazwa, K.; Yamada, M.; Kato, S.; Shinomura, T.; Nagashima, Y.; Suzuki, H.; Ara, T.; Harada, J.; Takaichi, S. Oxygenic phototrophs need -carotene isomerase (Z-ISO) for carotene synthesis: functional analysis in Arthrospira and Euglena. Plant Cell Physiol. 61, 276–282. [CrossRef]
- Chen, Y.; Li, F.; Wurtzel, E.T. Isolation and characterization of the Z-ISO gene encoding a missing component of carotenoid biosynthesis in plants. Plant Physiol. 2010, 153, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Masamoto, K.; Wada, H.; Kaneko, T.; Takaichi, S. Identification of a gene required for cis-to-trans carotene isomerization in carotenogenesis of the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2001, 42, 1398–1402. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, J.; Vioque, A.; Sandmann, G. Gene sll0033 from Synechocystis 6803 encodes a carotene isomerase involved in the biosynthesis of all-E lycopene. Z. Naturforsch. 2001, 56c, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kreunen, S.S.; Cuttriss, A.J.; DellaPenna, D.; Pogson, B.J. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis. Plant Cell 2002, 14, 321–332. [Google Scholar] [CrossRef]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Böger, P.; Sandmann, G. Phytoene desaturase: Heterologous expression in an active state, purification, and biochemical properties. Protein Expr. Purif. 1997, 10, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Takaichi, S.; Misawa, N.; Maoka, T.; Miyashita, H.; Mimuro, M. The cyanobacterium Gloeobacter violaceus PCC 7421 uses bacterial-type phytoene desaturase in carotenoid biosynthesis. FEBS Lett. 2005, 579, 2125–2129. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Distribution and biosynthesis of carotenoids. In The Purple Phototrophic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 97–117. [Google Scholar]
- Krubasik, P.; Sandmann, G. Molecular evolution of lycopene cyclases involved in the formation of carotenoids with ionone end groups. Biochem. Soc. Trans. 2000, 28, 806–810. [Google Scholar] [CrossRef]
- Maresca, J.A.; Graham, J.E.; Wu, M.; Eisen, J.A.; Bryant, D.A. Identification of a fourth family of lycopene cyclases in photosynthetic bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 11784–11789. [Google Scholar] [CrossRef]
- Sugiyama, K.; Takaichi, S. Carotenogenesis in cyanobacteria: lycopene cyclases of CruA/CruP-type and CrtL-type. J. Gen. Appl. Microbiol. 2020, 66, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Molecular evolution of carotenoid biosynthesis from bacteria to plants. Physiol. Plant. 2002, 116, 431–440. [Google Scholar] [CrossRef]
- Ramos, A.; Coesel, S.; Marques, A.; Rodrigues, M.; Baumgartner, A.; Noronha, J.; Rauter, A.; Brenig, B.; Varela, J. Isolation and characterization of a stress-inducible Dunaliella salina Lyc-β gene encoding a functional lycopene β-cyclase. Appl. Microbiol. Biotechnol. 2008, 79, 819–828. [Google Scholar] [CrossRef]
- Harker, M.; Hirschberg, J. Molecular biology of carotenoid biosynthesis in photosynthetic organisms. Methods Enzymol. 1998, 297, 244–263. [Google Scholar]
- Cunningham, F.X., Jr.; Sun, Z.; Chamovitz, D.; Hirschberg, J.; Gantt, E. Molecular structure and enzymatic function of lycopene cyclase from the cyanobacterium Synechococcus sp. strain PCC7942. Plant Cell 1994, 6, 1107–1121. [Google Scholar]
- Stickforth, P.; Steiger, S.; Hess, W.R.; Sandmann, G. A novel type of lycopene ε-cyclase in the marine cyanobacterium Prochlorococcus marinus MED4. Arch. Microbiol. 2003, 179, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X., Jr.; Gantt, E. One ring or two? Determination of ring number in carotenoids by lycopene ε-cyclases. Proc. Natl. Acad. Sci. USA 2001, 98, 2905–2910. [Google Scholar] [CrossRef]
- Cunningham, F.X., Jr.; Lee, H.; Gantt, E. Carotenoid biosynthesis in the primitive red alga Cyanidioschyzon merolae. Eukaryot. Cell 2007, 6, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, J.; Linden, H. Light induction of carotenoid biosynthesis genes in the green alga Haematococcus pluvialis: Regulation by photosynthetic redox control. Plant Mol. Biol. 2003, 52, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.-Y.; Cheng, L.; Wang, Q.; Ge, Z.-H.; Zheng, H.; Cao, T.-J.; Lu, Q.-Q.; Yang, L.-E.; Lu, S. Functional characterization of lycopene cyclases illustrates the metabolic pathway toward lutein in red algal seaweeds. J. Agric. Food Chem. 2020, 68, 1354–1363. [Google Scholar] [CrossRef]
- Cordero, B.F.; Couso, I.; Leon, R.; Rodrigguez, H.; Vargas, M.A. Isolation and characterization of a lycopene e-cyclase gene of Chlorella (Chromochloris) zofingiensis. Regulation of the carotenogenic pathway by nitrogen and light. Mar. Drugs 2012, 10, 2069–2088. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Ikejiri, S.; Nakayama, T.; Nishino, T. Fusion-type lycopene β-cyclase from a thermoacidophilic archaeon Sulfolobus solfataricus. Biochem. Biophys. Res. Commun. 2003, 305, 586–591. [Google Scholar] [CrossRef]
- Iniesta, A.A.; Cervantes, M.; Murillo, F.J. Conversion of the lycopene monocyclase of Myxococcus xanthus into a bicyclase. Appl. Microbiol. Biotechnol. 2008, 79, 793–802. [Google Scholar] [CrossRef]
- Göttl, V.L.; Pucker, B.; Wendisch, V.F.; Henke, N.A. Screening of structurally distinct lycopene b-cyclases for production of the cyclic C40 carotenoids b-carotene and astaxanthin by Corynebacterium glutamicum. J. Agric. Food Chem. 2023, 71, 7765–7776. [Google Scholar] [CrossRef]
- Bradbury, L.M.T.; Shumskaya, M.; Tzfadia, O.; Wu, S.B.; Kennkelly, E.J.; Wurtzel, E.T. 2012. Lycopene cyclase paralog CruP protects against reactive oxygen species in oxygenic photosynthetic organisms. Proc. Natl. Acad. Sci. USA 2012, 109, E1888–E1897. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, K.; Ebisawa, M.; Yamada, M.; Nagashima, Y.; Suzuki, H.; Maoka, T.; Takaichi, S. Functional lycopene cyclase (CruA) in cyanobacterium, Arthrospira platensis NIES-39, and its role in carotenoid synthesis. Plant Cell Physiol. 2017, 58, 831–838. [Google Scholar] [CrossRef]
- Xiong, W.; Shen, G.; Bryant, D.A. Synechocystis sp. PCC 6803 CruA (sll0147) encodes lycopene cyclase and requires bound chlorophyll a for activity. Photosynth. Res. 2017, 131, 276–280. [Google Scholar] [CrossRef]
- Masamoto, K.; Misawa, N.; Kaneko, T.; Kikuno, R.; Toh, H. β-Carotene hydroxylase gene from the cyanobacterium Synechocystis sp. PCC6803. Plant Cell Physiol. 1998, 39, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Vermaas, W. The zeaxanthin biosynthesis enzyme β-carotene hydroxylase is involved in myxoxanthophyll synthesis in Synechocystis sp. PCC 6803. FEBS Lett. 1999, 454, 247–251. [Google Scholar]
- Mochimaru, M.; Msukawa, H.; Maoka, T.; Mohamed, H.E.; Vermaas, W.F.J.; Takaichi, S. Substrate specificities and availability of fucosyltransferase and β-carotene hydroxylase for myxol 2′-fucoside synthesis in Anabaena sp. strain PCC 7120 compared with Synechocystis sp. strain PCC 6803. J. Bacteriol. 2008, 190, 6726–6733. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Harada, H.; Ikenaga, H.; Matsuda, S.; Takaichi, S.; Shindo, K.; Sandmann, G.; Ogata, T.; Misawa, N. Characterization of cyanobacterial carotenoid ketolase CrtW and hydroxylase CrtR by complementation analysis in Escherichia coli. Plant Cell Physiol. 2008, 49, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Iwai, M.; Maoka, T.; Ikeuchi, M.; Takaichi, S. 2,2′-β-Hydroxylase (CrtG) is involved in carotenogenesis of both nostoxanthin and 2-hydroxymyxol 2′-fucoside in Thermosynechococcus elongatus strain BP-1. Plant Cell Physiol. 2008, 49, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Fernández-González, B.; Sandmann, G.; Vioque, A. A new type of asymmetrically acting β-carotene ketolase is required for the synthesis of echinenone in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 1997, 272, 9728–9733. [Google Scholar] [CrossRef]
- Mochimaru, M.; Msukawa, H.; Takaichi, S. The cyanobacterium Anabaena sp. PCC 7120 has two distinct β-carotene ketolase: CrtO for echinenone and CrtW for ketomyxol synthesis. FEBS Lett. 2005, 579, 6111–6114. [Google Scholar] [CrossRef]
- Steiger, S.; Sandmann, G. Cloning of two carotenoid ketolase genes from Nostoc punctiforme for the heterologous production of canthaxanthin and astaxanthin. Biotechnol. Lett. 2004, 26, 813–817. [Google Scholar] [CrossRef]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for myxol-2’ fucoside (myxoxanthophyll) in the cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 2009, 191, 3292–3300. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Lecomte, J.T.J.; Bryant, D.A. Synechoxanthin, an aromatic C40 xanthophyll that is a major carotenoid in the cyanobacterium Synechococcus sp. PCC 7002. J. Nat. Prod. 2008, 71, 1647–1650. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for synechoxanthin, an aromatic carotenoid synthesized by the euryhaline, unicellular cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 2008, 190, 7966–7974. [Google Scholar] [CrossRef]
- Dautermann, O.; Lohr, M. A functional zeaxanthin epoxidase from red algae shedding light on the evolution of light-harvesting carotenoids and the xanthophyll cycle in photosynthetic eukaryotes. Plant J. 2017, 92, 897–891. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Huang, X.; Hang, Y.; Deng, Y.; Lu, Q.; Lu, S. The P450-type carotene hydroxylase PuCHY1 from Porphyra suggests the evolution of carotenoid metabolism in red algae. J. Integr. Plant Biol. 2014, 56, 902–915. [Google Scholar] [CrossRef]
- Swift, I.E.; Milborrow, B.V. Stereochemistry of allene biosynthesis and the formation of the acetylenic carotenoid diadinoxanthin and peridinin (C37) from neoxanthin. Biochem. J. 1981, 199, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Swift, I.E.; Milborrow, B.V.; Jeffrey, S.W. Formation of neoxanthin, diadinoxanthin and peridinin from [14C]zeaxanthin by a cell-free system from Amphidinium carterae. Phytochemistry 1982, 21, 2859–2864. [Google Scholar] [CrossRef]
- Cui, H.; Ma, H.; Cui, Y.; Zhu, X.; Qin, S.; Li, R. Cloning, identification and functional characterization of two cytochrome P450 carotenoids hydroxylases from the diatom Phaeodactylum tricornutum. J. Biosci. Bioeng. 2019, 128, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Eilers, U.; Dietzel, L.; Breitenbach, J.; Buchel, C.; Sandmann, G. Identification of genes coding for functional zeaxanthin epoxidases in the diatom Phaeodactylum tricornutum. J. Plant Physiol. 2016, 192, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Græsholt, C.; Brembu, T.; Volpe, C.; Bartosova, Z.; Serif, M.; Winge, P.; Nymark, M. Zeaxanthin epoxidase 3 knockout mutants of the model diatom Phaeodactylum tricornutum enable commercial production of the bioactive carotenoid diatoxanthin. Mar Drugs 2024, 22, 185. [Google Scholar] [CrossRef]
- Dautermann, O.; Lysjka, D.; Andersen-Ranberg, J.; Becker, M.; Fröhlich-Nowoisky, J.; Gartmann, H.; Kärmer, L.C.; Mayr, K.; Pirper, D.; Rij, L.M.; Wiph, M.-L.; Niyogi, K.K.; Lohr, M. An algal enzyme required for biosynthesis of the most abundant marine carotenoids. Sci. Adv. 2020, 6, eaaw9183. [Google Scholar] [CrossRef] [PubMed]
- Goss, R.; Jakob, T. Regulation and function of xanthophyll cycle-dependent photoprotection in algae. Photosynth. Res. 2010, 106, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Iha, M.; Maoka, T.; Misawa, N.; Takemura, M. Synthetic-biological approach for production of neoxanthin in Escherichia coli. Plant Biotechnol. 2023, 40, 15–20. [Google Scholar] [CrossRef]
- Bai, Y.; Cao, T.; Dautermann, O.; Buschbeck, P.; Cantrell, M.B.; Chen, Y.; Lein, C.D.; Shi, X.; Ware, M.A.; Yng, F.; Zhang, H.G.; Zhang, L.; Peers, G.; Li, X.; Lohr, M. Green diatom mutants reveal ana intricate biosynthetic pathway of fucoxanthin. Proc. Natl. Acad. Sci. USA 2022, 119, e2203708119. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Bai, Y.; Buschbeck, P.; Tan, Q.; Cantrell, M.B.; Chen, Y.; Jiang, Y.; Liu, R.-Z.; Ries, N.K.; Shi, X.; Sun, Y.; Ware, M.A.; Yang, F.; Zhang, H.; Han, J.; Zhang, L.; Huang, J.; Lohr, M.; Peers, G.; Li, X. An unexpected hydratase synthesizes the green light-absorbing pigment fucoxanthin. Plant Cell 2023, 35, 3035–3072. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Yamano, Y.; Oka, N.; Kamei, Y.; Fujii, R. Discovery of a novel siphonaxanthin biosynthetic precursor in Codium fragile that accumulates only by exposure to blue-green light. FEBS Lett. 2022, 596, 1544–1555. [Google Scholar] [CrossRef]
- Linden, H. Carotenoid hydroxylase from Haematococcus pluvialis: cDNA sequence, regulation and functional complementation. Biochim. Biophys. Acta 1999, 1446, 203–212. [Google Scholar] [CrossRef]
- Baroli, I.; Do, A.D.; Yamane, T.; Niyogi, K.K. Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress. Plant Cell 2003, 15, 992–1008. [Google Scholar] [CrossRef] [PubMed]
- Goss, R. Substrate specificity of the violaxanthin de-epoxidase of the primitive green alga Mantoniella squamata (Prasinophyceae). Planta 2003, 217, 801–812. [Google Scholar] [CrossRef]
- Lotan, T.; Hirschberg, J. Cloning and expression in Escherichia coli of the gene encoding β-C-4-oxygenase, that converts β-carotene to the ketocarotenoid canthaxanthin in Haematococcus pluvialis. FEBS Lett. 1995, 364, 125–128. [Google Scholar]
- Kajiwara, S.; Kakizono, T.; Saito, T.; Kondo, K.; Ohtani, T.; Nishio, N.; Nagai, S.; Misawa, N. Isolation and functional identification of a novel cDNA from astaxanthin biosynthesis from Haematococcus pluvialis, and astaxanthin synthesis in Escherichia coli. Plant Mol. Biol. 1995, 29, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-C.; Wang, Y.; Sandmann, G.; Chen, F. Isolation and characterization of a carotenoid oxygenase gene from Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2006, 71, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-C.; Chen, F.; Sandmann, G. Stress-related differential expression of multiple β-carotene ketolase genes in the unicellular green alga Haematococcus pluvialis. J. Biotechnol. 2006, 122, 176–185. [Google Scholar] [CrossRef]
- Lemoine, Y.; Schoefs, B. Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Smith, J.J.; Tian, L.; DellaPenna, D. The evolution and function of carotenoid hydroxylases in Arabidopsis. Plant Cell Physiol. 2009, 50, 463–479. [Google Scholar] [CrossRef] [PubMed]
- Büchel, C. Light harvesting complexes in chlorophyll c-containing algae. BBA-Bioenerg, 2020; 1861, 148027. [Google Scholar]
- Llansola-Portoles, M.J.; Litvin, R.; Ilioaia, C.; Pascal, A.A.; Bina, D.; Robert, B. Pigment structure in the violaxanthin-chlorophyll-a-binding protein VCA. Photosynth. Res. 2017, 134, 51–58. [Google Scholar] [CrossRef]
- Watanabe, A.; Minagawa, J. Structural characterization of the photosystems in the green alga Chlorella sorokiniana. Planta. 2020, 252, 79. [Google Scholar] [CrossRef]
- Nagao, R.; Kato, K.; Kumazawa, M.; Ifuka, K.; Yokono, M.; Suzuki, T.; Dohmae, N.; Akita, F.; Akimoto, S.; Miyazaki, N.; Shen, J.-R. Structural basis for different types of hetero-tetrameric light-harvesting complexes in a diatom PSII-FCPII supercomplex. Nat. Comun. 2022, 3, 1764. [Google Scholar] [CrossRef]
- Englert, G.; Bjørnland, T.; Liaaen-Jensen, S. 1D and 2D NMR study of some allenic carotenoids of the fucoxanthin series. Magn. Reson. Chem. 1990, 28, 519–528. [Google Scholar] [CrossRef]
- Egeland, E.S.; Liaaen-Jensen, S. Ten minor carotenoids from Prasinophyceae (Chlorophyta). Phytochemistry 1995, 40, 515–520. [Google Scholar] [CrossRef]
- Yoshii, Y.; Takaichi, S.; Maoka, T.; Hanada, S.; Inouye, I. Characterization of two unique carotenoid fatty acid esters from Pterosperma cristatum (Prasinophyceae, Chlorophyta). J. Phycol. 2002, 38, 297–303. [Google Scholar] [CrossRef]
- Egeland, E.S.; Guillard, R.R.L.; Liaaen-Jensen, S. Additional carotenoid prototype representatives and a general chemosystematic evaluation of carotenoids in Prasinophyceae (Chlorophyta). Phytochemistry 1997, 44, 1087–1097. [Google Scholar] [CrossRef]
- Mimuro, M.; Nagashima, U.; Takaichi, S.; Nishimura, Y.; Yamazaki, I.; Katoh, T. Molecular structure and optical properties of carotenoids for the in vivo energy transfer function in the algal photosynthetic pigment system. Biochim. Biphys. Acta 1992, 1098, 271–274. [Google Scholar] [CrossRef]
- Akimoto, S.; Yokono, M.; Higuchi, M.; Tomo, T.; Takaichi, S.; Murakami, A.; Mimuro, M. Solvent effects on excitation relaxation dynamics of a keto-carotenoid, siphonaxanthin. Photochem. Photobiol. Sci. 2008, 7, 1206–1209. [Google Scholar] [CrossRef]
- Hofmann, E.; Wrench, P.M.; Sharples, F.P.; Hiller, R.G.; Welte, W.; Diederichs, K. Structural basis of light harvesting by carotenoids: Peridinin-chlorophyll-protein from Amphidinium carterae. Science 1996, 272, 1788–1791. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, H.; Lu, X.; Lu, Y.; Cuneo, M.J.; O’Neill, M.J.; Urban, V.; Lo, C.S.; Blankenship, R.E. Oligomerization state and pigment binding strength of the peridinin-Chl a-protein. FEBS Lett. 2015, 589, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Kerfeld, C.A.; Sawaya, M.R.; Brahmandam, V.; Cascio, D.; Ho, K.K.; Trevithick-Sutton, C.C.; Krogmann, D.W.; Yeates, T.O. The crystal structure of a cyanobacterial water-soluble carotenoid binding protein. Structure 2003, 11, 55–65. [Google Scholar] [CrossRef]
- Kirilovsky, D. Modulating energy arriving at photochemical reaction centers: orange carotenoid protein-related photoprotection and state transitions. Photosynth. Res. 2015, 126, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Muzzopappa, F.; Kirilovsky, D. Changing color for photoprotection: the orange carotenoid protein. Trends Plant Sci. 2020, 25, 92–104. [Google Scholar] [CrossRef]
- Yamamoto, H.Y.; Bugos, R.C.; Hieber, A.D. Biochemistry and molecular biology of the xanthophyll cycle. In The Phytochemistry of Carotenoids; Frank, H.A., Young, A.J., Britton, G., Cogdell, R.J., Eds.; Kluwer: Dordrecht, The Netherlands, 1999; pp. 293–303. [Google Scholar]
- Niyogi, K.K.; Björkman, O.; Grossman, A.R. Chlamydomonas xanthophyll cycle mutants identified by video imaging of chlorophyll fluorescence quenching. Plant Cell 1997, 9, 1369–1380. [Google Scholar] [CrossRef]
- Couso, I.; Cordero, B.F.; Vargas, M.Á.; Rodríguez, H. Efficient heterologous transformation of Chlamydomonas reinhardtii npq2 mutant with the zeaxanthin epoxidase gene isolated and characterized from Chlorella zofingiensis. Mar. Drugs 2012, 10, 1955–1976. [Google Scholar] [CrossRef] [PubMed]
- Tóth, T.N.; Chukhutsina, V.; Domonkos, I.; Knoppová, J.; Komenda, J.; Kis, M.; Lénárt, Z.; Garab, G.; Kovács, L.; Gombos, Z.; van Amerongen, H. Carotenoids are essential for the assembly of cyanobacterial photosynthetic complexes. Biochim Biophys Acta 2015, 1847, 1153–1165. [Google Scholar] [CrossRef]
- Zakar, T.; Laczko-Dobos, H.; Toth, T.N.; Gombos, Z. Carotenoids assist in cyanobacterial photosystem II assembly and function. Front Plant Sci 2016, 7, 295. [Google Scholar]
- Toyoshima, H.; Miyata, A.; Yoshida, R.; Ishige, T.; Takaichi, S.; Kawasaki, S. Distribution of the water-soluble astaxanthin binding carotenoprotein (AstaP) in Scenedismaceae. Mar Drugs 2021, 19, 349. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, S.; Mitsui, T.; Omori, K.; Tsuboya, T.; Bader, A.; Toyoshima, H.; Takaichi, S. Microalgal AstaP is a natural solubilizer of astaxanthin with broad binding ability for carotenoids and related compounds. Algal Res. 2023, 70, 102982. [Google Scholar] [CrossRef]
Figure 1.
Structures of the chlorophylls.
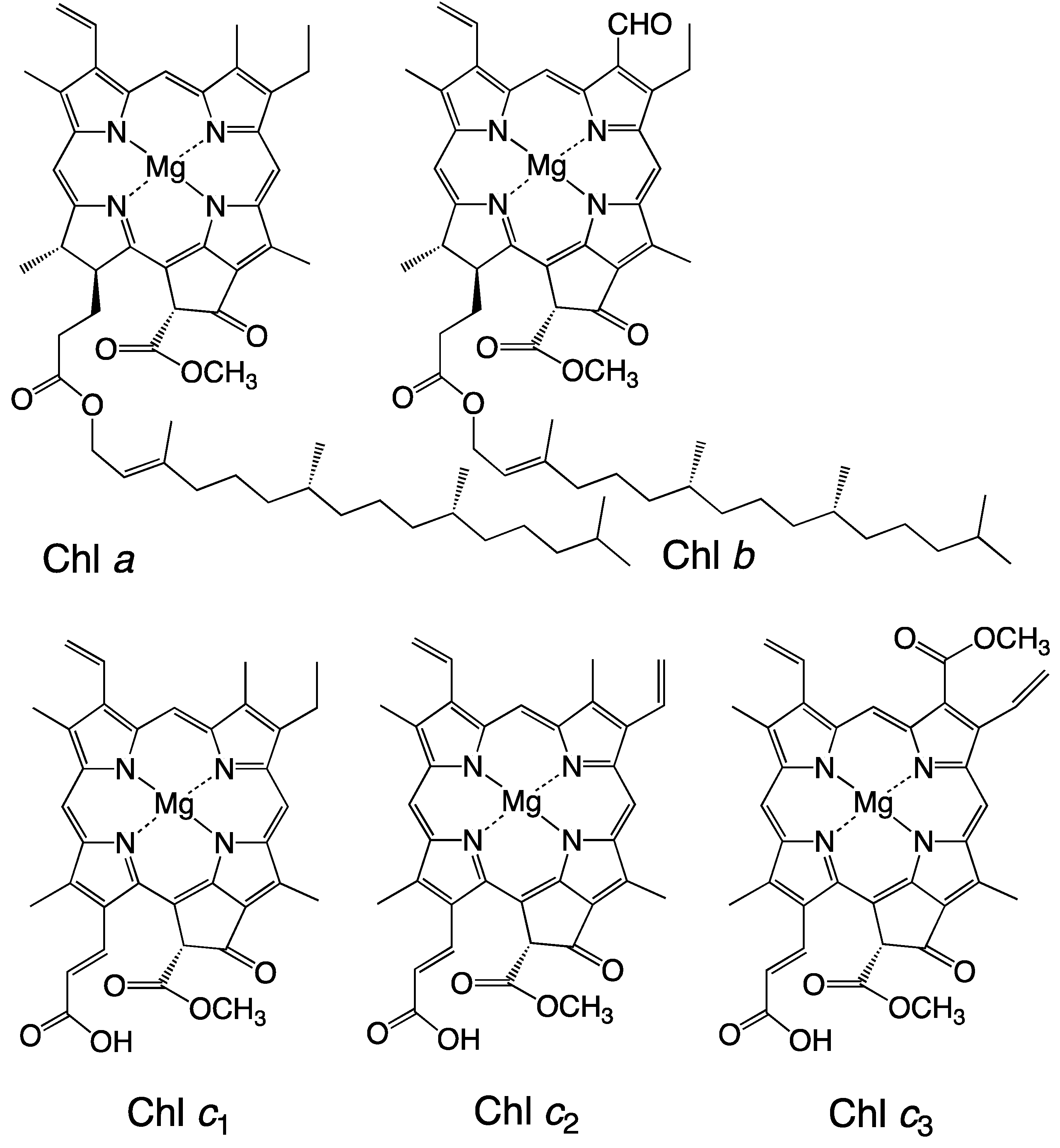
Figure 2.
Carotenogenesis pathways and chlorophylls in the chloroplasts of algae and land plants, highlighting key differences and functionally confirmed enzymes involved. The pathways, enzymes, and chlorophyll content vary across algal divisions and classes. This figure is an updated version presented in the previous review [10]. For the recently proposed fucoxanthin pathway, refer to Figure 4.
Figure 2.
Carotenogenesis pathways and chlorophylls in the chloroplasts of algae and land plants, highlighting key differences and functionally confirmed enzymes involved. The pathways, enzymes, and chlorophyll content vary across algal divisions and classes. This figure is an updated version presented in the previous review [10]. For the recently proposed fucoxanthin pathway, refer to Figure 4.
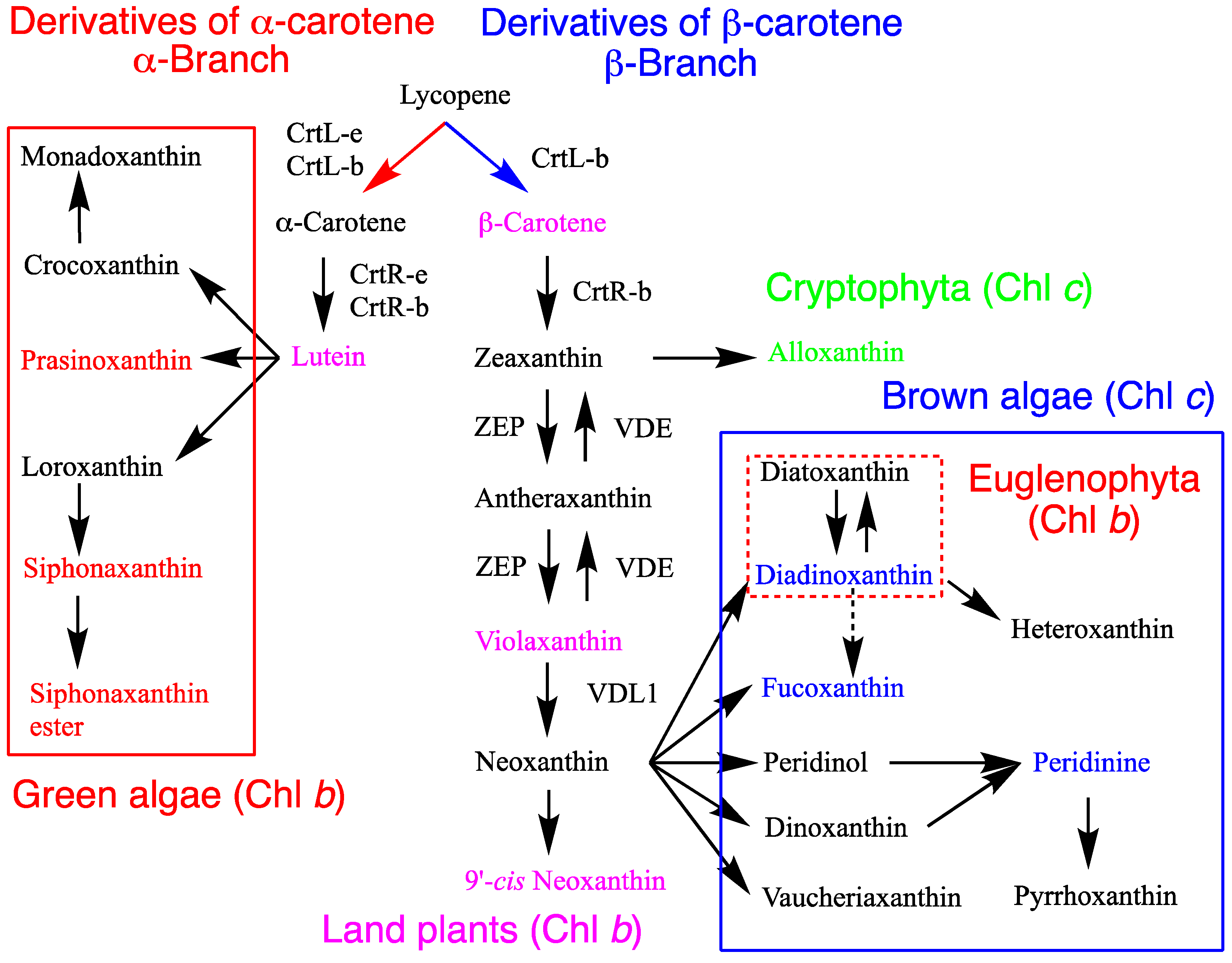
Figure 3.
Major carotenogenesis pathways in cyanobacteria along with the functionally confirmed enzymes involved. These pathways and enzymes vary across cyanobacterial species. Two lycopene cyclases, CruA and CrtL, have been identified. The glycoside attached to myxol and oscillol may be one of fucose, rhamnose, or chinovose. This figure is an updated version presented in the previous review [10].
Figure 3.
Major carotenogenesis pathways in cyanobacteria along with the functionally confirmed enzymes involved. These pathways and enzymes vary across cyanobacterial species. Two lycopene cyclases, CruA and CrtL, have been identified. The glycoside attached to myxol and oscillol may be one of fucose, rhamnose, or chinovose. This figure is an updated version presented in the previous review [10].
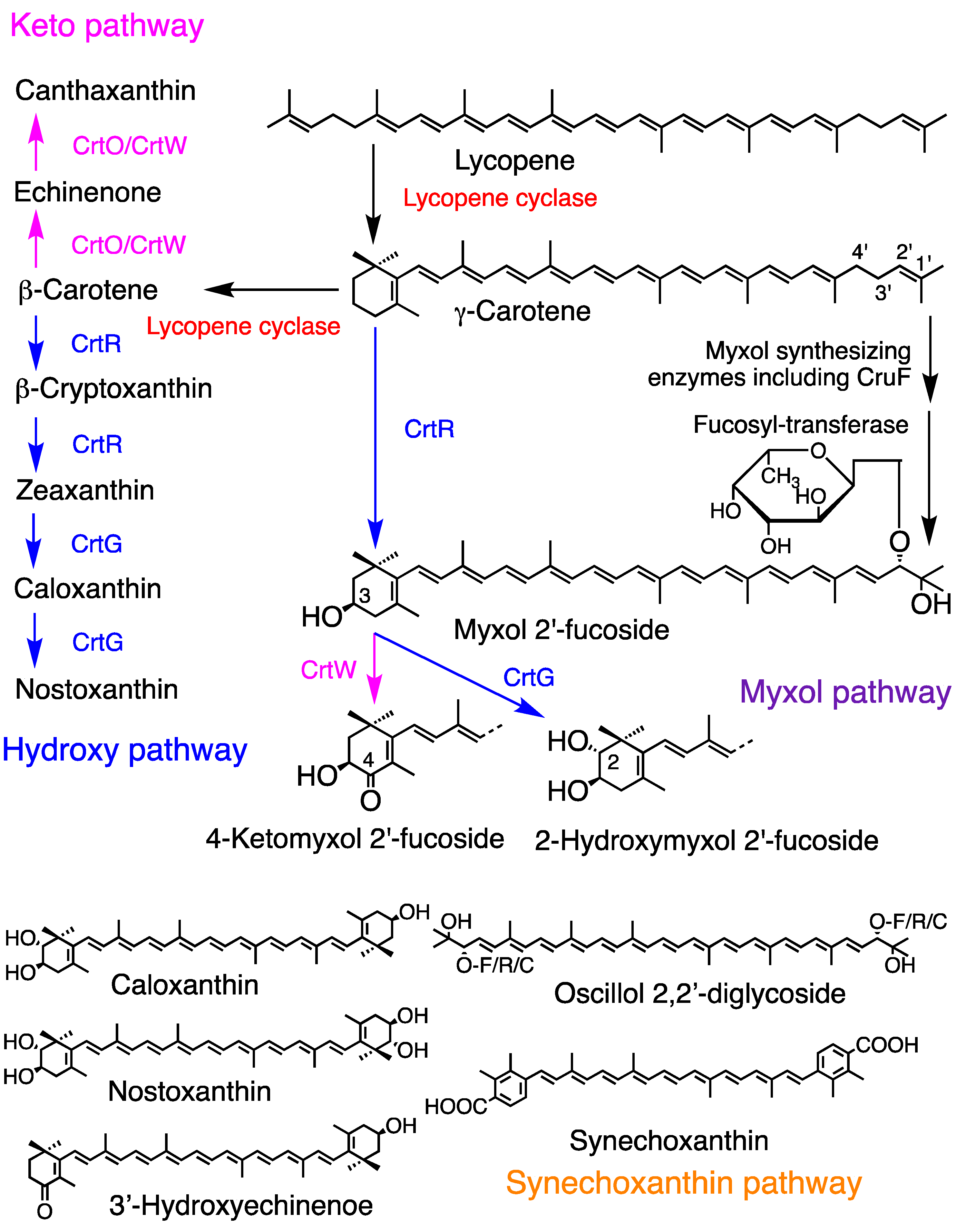
Figure 5.
Carotenogenesis pathway from b-carotene to fucoxanthin, as well as violaxanthin and diadinoxanthin cycles in the brown alga Phaeodactylum tricornutum. VDE is activated by high-light. The dotted blue line highlights a simple pathway (Fig. 1). The structures of fucoxanthin and fucoxanthinol are reversed for the right and left sides according to the IUPAC-IUB semi-systematic rule.
Figure 5.
Carotenogenesis pathway from b-carotene to fucoxanthin, as well as violaxanthin and diadinoxanthin cycles in the brown alga Phaeodactylum tricornutum. VDE is activated by high-light. The dotted blue line highlights a simple pathway (Fig. 1). The structures of fucoxanthin and fucoxanthinol are reversed for the right and left sides according to the IUPAC-IUB semi-systematic rule.
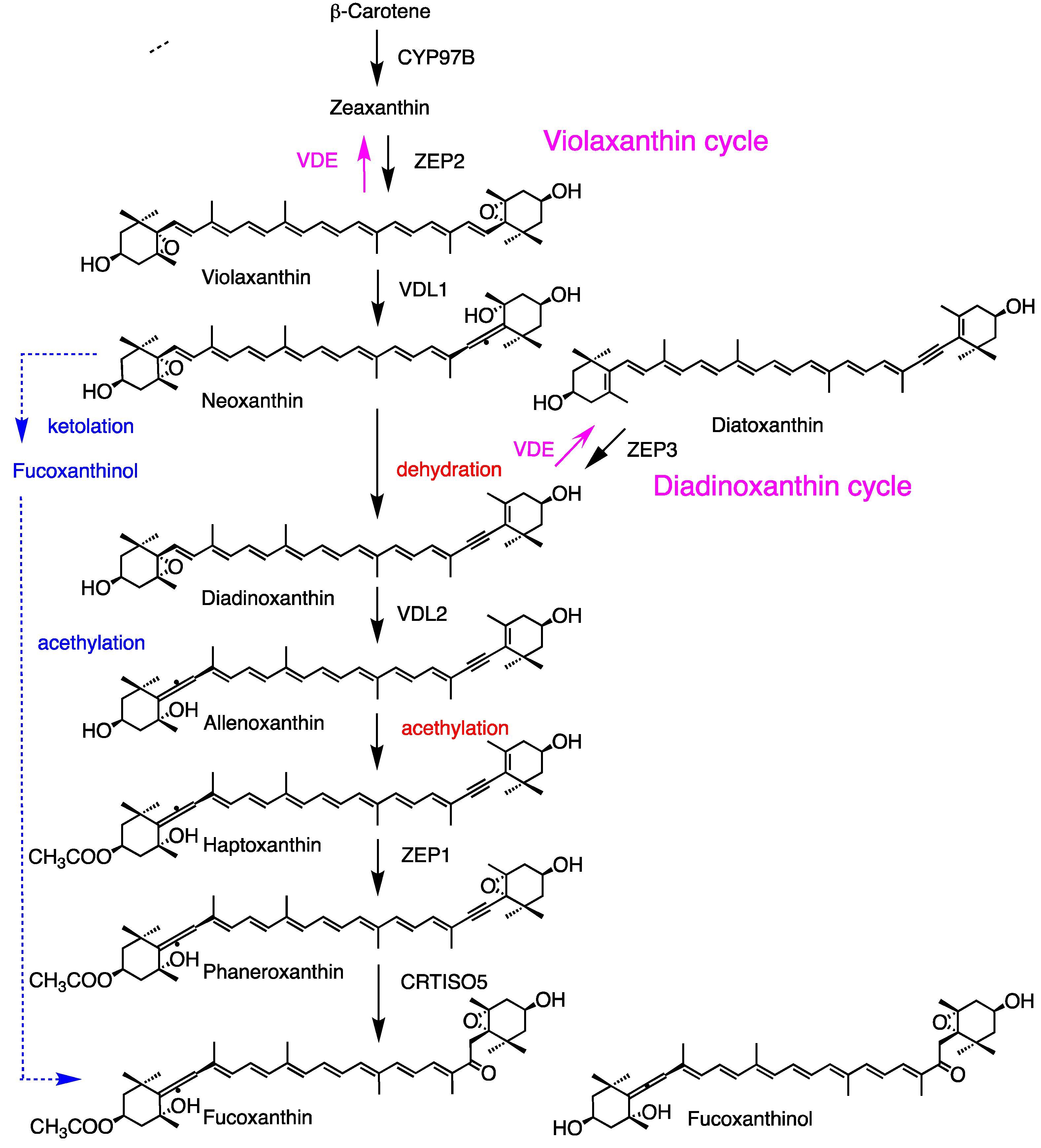
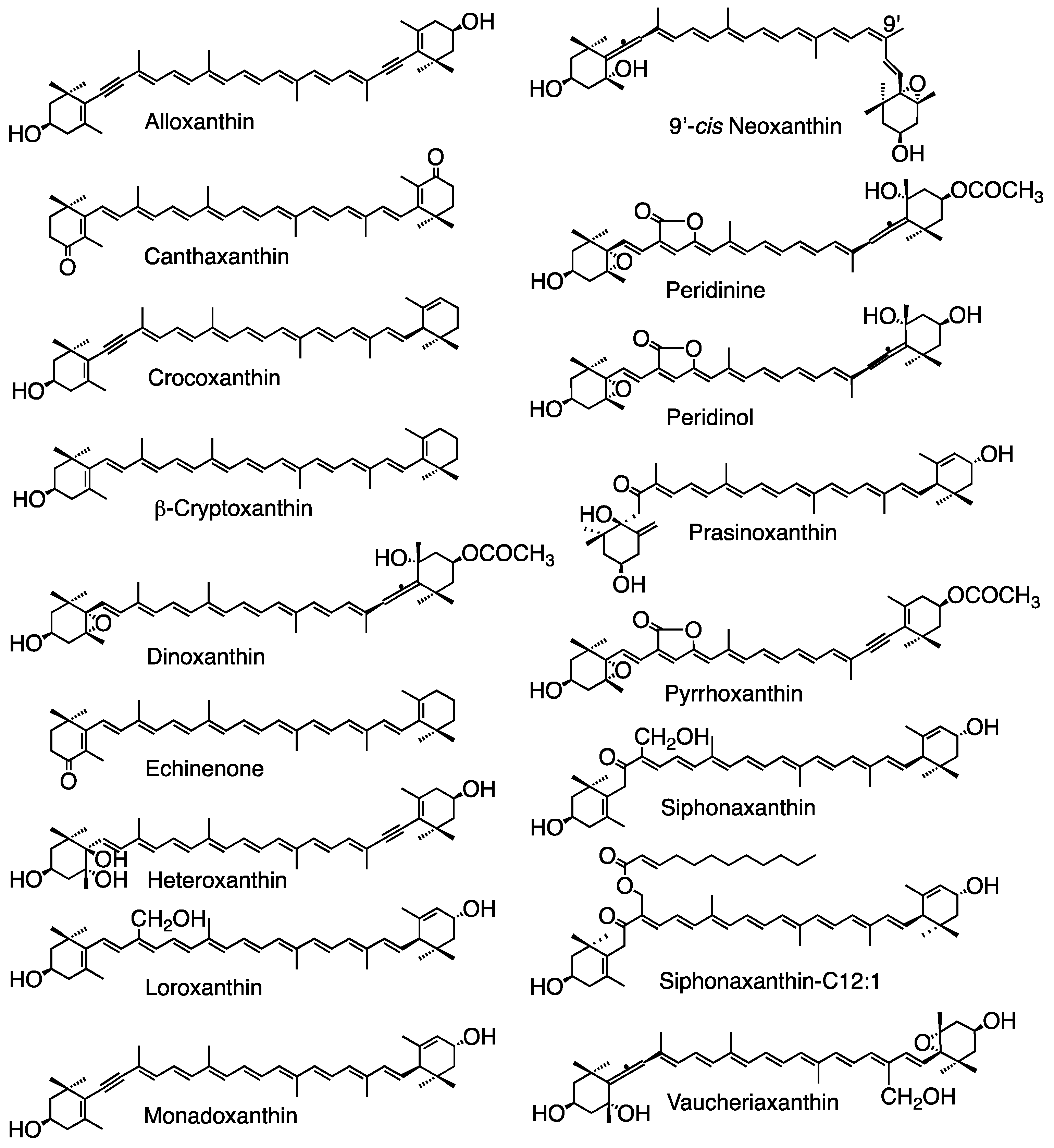
Figure 6.
Structures of some carotenoids mentioned in this review.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



