
Submitted:
06 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
Lyme-borreliosis (LB) is a tick-borne disease prevalent in many parts of the world, yet it remains largely unknown. This lack of awareness poses significant challenges for surveillance and diagnosis, requiring integrating epidemiological and clinical information. Additionally, the limited specificity and low titers of serological tests further complicate the diagnostic process. To address these issues, we investigated the molecular fine specificity of IgM antibody responses to five surface proteins of B. burgdorferi. Materials and Methods: We employed SPOT synthesis to screen for IgM epitopes and assess cross-reactivity through databank searches and ELISA. Validation was performed using ELISA and ROC curve analysis. Results: We identified ten IgM epitopes, of which four were classified as specific. The ELISA-peptide assay demonstrated a sensitivity of ≥ 87.3%, specificity of ≥ 56.2%, and accuracy of ≥ 66.6%. Two B. burgdorferi-specific epitopes, Bburg/02/huG, and Bburg/06/huG, were synthesized as polypeptides and reevaluated using ELISA with a panel of patient sera representing different pathologies. The results showed a sensitivity of 85.0% and a specificity of 100.0%, with significant differences in cross-reactivity between the leptospirosis and syphilis groups. Conclusions: These findings indicate that the identified peptide combinations could facilitate the development of new, highly specific serodiagnostic assays, thereby enhancing public health initiatives and epidemiological studies.
Keywords:
Lyme Disease
1. Introduction
The introduction should briefly place the study in a broad context and highlight its importance. Lyme-Borreliosis (LB) disease (LBD) is a bacterial zoonosis caused by spirochetes of the Borrelia burgdorferi sensu lato complex. Transmitted by the bite of ticks of the genus Ixodes, this condition is one of the most common vector-borne diseases in temperate regions of the Northern Hemisphere [1,2,3], with increasing recognition in other parts of the world including Brazil, which received the name of Baggio–Yoshinari Syndrome [4,5,6].
Recently, the recognition of the existence of various species of different parties do Mundo with different genomes (genospecies) has been conducted by researchers at the Borreliae Lyme Group (BLG) for better diagnosis of LB [7,8,9,10,11]. The genospecies reported inside the BLG differ regarding clinical manifestation and dissemination inside the human body and genome/antigens expression [12]. Consequently, these facts pose enormous difficulty in diagnosis and the complexity of clinical management, especially in advanced stages.
This multi-systemic disease is preventable and treatable when identified early. Awareness, early diagnosis, and appropriate treatment are crucial to reducing disease burden and preventing long-term complications. However, the diagnosis of this condition can be challenging due to the wide range of symptoms and its multiphasic nature, which includes localized, disseminated, and chronic initial stages and the antigenic and genetic heterogeneity of B. burgdorferi populations.
The clinical diagnosis of the LB is essential, especially in the early stages of the disease, which can be confused with other infectious or autoimmune conditions. In addition, inappropriate diagnosis leads to therapeutic wandering, inducing potential chronic infection with a strong antibody response that fails to clear the infection.
Detection of antibodies against B. burgdorferi is the most common laboratory method; however, the standard approach involves a two-tier test: ELISA (Enzyme-Linked Immunosorbent Assay) and Western Blot (confirmatory to identify specific antibodies). Although widely used, this method is fastidious and has limitations, including false-negative results in the early stages (before antibody production) and false-positive results due to cross-reactions with other infections [13]. PCR is an alternative, and although highly specific, its sensitivity varies depending on the sample and the stage of the disease [14,15,16].
Therefore, the development and implementation of new diagnostic platforms, based on single or multiple subunit antigens of unique Borrelia epitopes, may increase specificity and sensitivity, thus reducing the false positive rate in human borreliosis [17,18].
Compared to crude parasite antigens, the peptide subunit antigens offer greater consistency, purity, and standardization, making them a preferred approach in research and clinical settings. They also reduce the likelihood of cross-reactivity with antibodies raised against other agents [19]. Among these, the outer surface proteins (Osp) [20,21,22,23] and flagellar hooks (flag) [24,25,26] are conserved across Borrelia isolates and function as a universal antigen, playing a pivotal role in host immunomodulation and cell colonization, inducing a strong antibody response. These proteins play a critical role in the transmission and infection of LB disease spirochetes, as they help bacteria adapt to different hosts and evade the host immune system [27,28,29].
The microorganisms have the potential to propagate throughout the body, thereby stimulating an immune response that produces immunoglobulins G and M [30]. In contrast to IgG, IgM normally is the first antibody to activate the classic immune system pathway and is commonly found in the early stages of infection [31]. After activation of a mature B cell via its membrane-bound antibody molecule by antigen, these B cells proliferate. Through specific signaling via CD40 and cytokine receptors (both modulated by T helper cells), they undergo antibody class switching to produce IgG or IgA antibodies. During class switching, the constant region of the immunoglobulin heavy chain changes, but the variable areas do not, and therefore, antigenic specificity remains the same, but the immunoglobulins present higher affinities [32].
Our group recently identified the epitopes of these five outer membrane proteins and found some specific epitopes for some IgG genera [33]. Identifying IgM epitopes is important to understanding the changes in the humoral immune system and thus identifying the best single epitopes present from the acute to the chronic phase to constitute a diagnostic test.
In some cases, patients who consistently test positive for anti-Borrelia IgM may develop persistent or recurrent symptoms of Lyme disease, which can be classified as either chronic LB disease or post-treatment Lyme disease syndrome (PTLDS) [34,35].
As a result, antibodies generated by infection with B. burgdorferi have been of great interest in confirming infection. Serological diagnosis of LB disease based on CDC criteria has been used [36]; however, in Brazil, it can be problematic and lead to false-negative diagnoses [37]. In addition to detecting antibodies against acute and chronic infections, serological tests can be used for interventions with antimicrobials and to understand the epidemiology and surveillance of LB-like syndrome, which can guide public health or health, given that it is a zoonotic disease [38]. Other rapid serological tests have been developed, and epitope-specific antibody serological assays have improved specificity [39,40,41].
Conversely, high-throughput biomarker analysis methods have been developed to identify disease risk and diagnostic, prognostic, and therapeutic targets in human clinical samples [42]. Protein microarrays are perfectly suited to fill the gap between biomarker discovery and diagnostics on the same platforms. Disease signatures can be identified, validated, and used for routine diagnostics.
Thus, this study aims to elucidate the IgM epitopes of five flagellar and outer surface proteins of B. burgdorferi, offering essential insights for developing new diagnostic tests aimed at early detection and current challenges imposed by Borreliosis.
2. Materials and Methods
2.1. Human Serum Samples
Serum samples were collected from a panel of 21 seropositive or suspected cases, with samples taken either more than 3 months prior (n=11) or less than 3 months prior (n=13). Patients clinically diagnosed with Lyme disease-like syndrome were confirmed by Western blot and ELISA tests and obtained from the Rheumatology Division at the University of São Paulo, School of Medicine (USP), and the Laboratory of Biodiversity in Entomology at Instituto Oswaldo Cruz (IOC/FIOCRUZ). Additionally, the study included 39 seronegative samples from healthy blood donors provided by HEMORIO.
2.2. Preparation of the Cellulose Membrane-Bound Peptide Array
The complete sequence of B. burgdorferi (Q44767, P11089, O51173, Q44849, and P0CL66) was covered by synthesizing 15-residue peptides with a 10-residue overlap, automatically prepared on cellulose membranes (Amino-PEG 500-UC540) following the standard SPOT synthesis protocol using an Auto-Spot Robot ASP-222 (Intavis Bioanalytical Instruments AG, Köln, Germany) [43]. Positive control peptides, including [IHLVNNESSEVIVHK and GYPKDGNAFNNLDRI] (Clostridium tetani, spots P20 and P21), KEVPALTAVETGATN (Polio virus, spot P22), and YPYDVPDYAGYPYDV (H. influenza virus hemagglutinin, spot P23), were included, and programming was conducted using Multipep software (Intavis Bioanalytical Instruments AG, Köln, Germany). The library contained 371 peptides plus 4 positive control peptides (Table S1). Coupling reactions were followed by acetylation with 4% acetic anhydride in N, N-dimethylformamide to make the peptides N-reactive for subsequent steps. After acetylation, F-moc protecting groups were removed with piperidine to activate the peptides. Additional amino acids were added through coupling, blocking, and deprotection until the desired peptide was obtained. Once the final amino acid was added, sidechain deprotection was performed using a solution of dichloromethane-trifluoracetic acid-triisopropylsilane (1:1:0.05, v/v/v), followed by ethanol washing as described previously [44]. The membranes containing the synthetic peptides were immediately probed.
2.3. Evaluation of SPOT Membranes
SPOT membranes were washed for 10 min with TBS-T buffer (50 mM Tris, 136 mM NaCl, 2 mM KCl, and 0.05% Tween, pH 7.4) and then blocked with TBS-T containing 1.5% BSA for 90 min at 8°C with agitation. After extensive washing with TBS-T, the membranes were incubated for 12 h with a pool (n = 7) of Lyme disease patient sera (1:150) in TBS-T + 0.75% BSA, followed by another wash with TBS-T. The membranes were then incubated with goat anti-human IgM (µ-chain specific) conjugated with alkaline phosphatase (anti-human IgM, 1:5000; Sigma-Merck, St Louis, Mo, USA) for 1 h and washed with TBS-T and CBS (50 mM citrate-buffered saline). Finally, chemiluminescent CDP-Star® substrate (Cytiva, Marlborough, MA, USA) (0.25 mM) with Nitro-Bloc-II™ Enhancer (Cat # T2218, Applied Biosystems, USA) was added to complete the reaction for 5 min.
2.4. Scanning and Quantification of Spot Signal Intensities
Chemiluminescent signals were detected using an Odyssey FC (LI-COR Bioscience, Lincoln, NE, USA) under previously established conditions [45], with minor modifications. In brief, a digital image file with a 5 MP resolution was generated, and signal intensities were quantified using TotalLab TL100 software (v 2009, Nonlinear Dynamics, Newcastle-Upon-Tyne, UK). This software includes an automatic grid search for 384 spots but does not identify potential epitope sequences automatically. Therefore, the data were further analyzed using Microsoft Excel. For a sequence to be considered an epitope, two or more positive contiguous spots must have a signal intensity (SI) equal to or greater than 30% of the highest value from the set of spots on the corresponding membrane. Signal intensity (SI) for the background was defined by a set of negative controls on each membrane.
2.5. Peptide preparation
The selected individual peptides were synthesized via the solid-phase chemical method using the 9-fluorenyl methoxycarbonyl (F-moc) strategy on an automated peptide synthesizer (MultiPep-1, CEM Corp, Charlotte, NC, USA), as previously described [43]. Peptide concentrations were determined by measuring optical density and applying the molar extinction coefficient calculated by the PROTPARAM software package [http://www.expasy.ch]. The peptide sequences were confirmed using mass spectrometry (MALDI-TOF MS) (Matrix-Assisted Laser Desorption Ionization Time-of-Flight).
2.6. Synthesis of Bi-Specific Antigen Peptides
The individual epitope peptides Bburg/02/huG and Bburg/06/huG were synthesized in tandem, incorporating a GSGG interpeptide spacer. For the preparation of the multi-antigen peptide (MAPs4), a polyethylene glycol grafted TentaGel®M NH2 resin (RAPP Polymer, Tübingen, Land Baden-Württemberg, Germany) and the standard solid-phase synthesis protocol was used. The constructs were synthesized on an automated peptide synthesizer (MultiPep1, CEM Corp, Charlotte, NC, USA), following the established method [45]. HPLC-purified Peptides and their identities were confirmed by MS (MALDI-TOF or electrospray).
2.7. Immunoenzymatic Assay (ELISA)
The ELISA was conducted as previously outlined, with slight adjustments [46]. ELISA plates (Immulon 4HB flat-bottom 96-well microtiter, Corning ) were coated with 100 µL (0.5 µg/well) of a synthetic single peptide or polypeptide dissolved in coating buffer (0.1 M sodium carbonate-bicarbonate, pH 9.6) and incubated overnight at 4°C. Following each incubation step, the plates (Immulon 4HBx) were washed three times with PBS-T (phosphate-buffered saline with 0.1% Tween 20, pH 7.2) and blocked with 200 µL of PBS-T containing 2.5% BSA for 2 h at 37°C.
Subsequently, the plates were incubated for 1 h at 37°C with 50 µL of serum from patients with Lyme disease-like syndrome, diluted in the blocking buffer (1:100 for single peptides and 1:50 for polypeptides). After additional washes, the plates were treated with 100 µL of goat anti-human IgM (μ-chain specific) −HRP (1:20,000, Sigma‒Aldrich, St Louis, Mo, USA) for 1 h at 37°C. The reaction was developed using Chemiluminescent CDP-Star® Substrate with Nitro-Block-II™ Enhancer (Applied Biosystems, Waltham, MA, U.S.A.) substrate for 15 min, and absorbance at 405 nm was recorded with a Hidex Sense Microplate Reader (Turku, Finland). A response was deemed elevated if the optical density exceeded the threshold, defined as the mean of the negative controls plus three standard deviations.
2.8. Computational Tools
The protein sequences of interest (Q44767, P11089, O51173, Q44849, and P0CL66) from the B. burgdorferi American strain were initially retrieved from the UniProt database [http://www.uniprot.org/] (Accessed on 11 August 2022). Similarity analysis with proteins from other organisms was conducted using BLASTP. To identify the epitope locations within the 3D structures of these proteins, we generated models using UCSF ChimeraX, Version 1.5 https// https://www.rbvi.ucsf.edu/chimerax/; Accessed on 12 August 2021). Structure generation of the five proteins was performed by employing the AlphaFold server [47] and the Protein Data Bank (PDB). Searches for B. burgdorferi peptides were conducted in the Protein Information Resource (PIR) database (https://research.bioinformatics.udel.edu/peptide match/index.jsp, Accessed on 12 April 2022) using previously identified sequences in other organisms.
2.9. Data Analysis
The ELISA test results were statistically analyzed using MedCalc software version 20.218 [48]. Each peptide's reactivity index (RI) was calculated as the ratio of the optical density (OD) of a specific sample to the cutoff OD values for each test. Sensitivity, specificity, and cross-reactivity with leptospirosis and syphilis samples were also assessed. RI values were classified as positive (>1.00) or negative (≤1.00), with a Gray zone defined as RI values within ±10% of the cutoff, where results may be ambiguous. Statistical analysis of cross-reactivity was performed using GraphPad Prism version 5.0. Differences between groups were assessed using the Kruskal-Wallis test, with statistical significance set at a p-value ≤ 0.05. A one-way ANOVA with the Kruskal-Wallis post-test was used to compare multiple groups, with p-values <0.005 considered statistically significant
3. Result
3.1. Detection of IgM Epitopes in Surface Proteins of B. burgdorferi
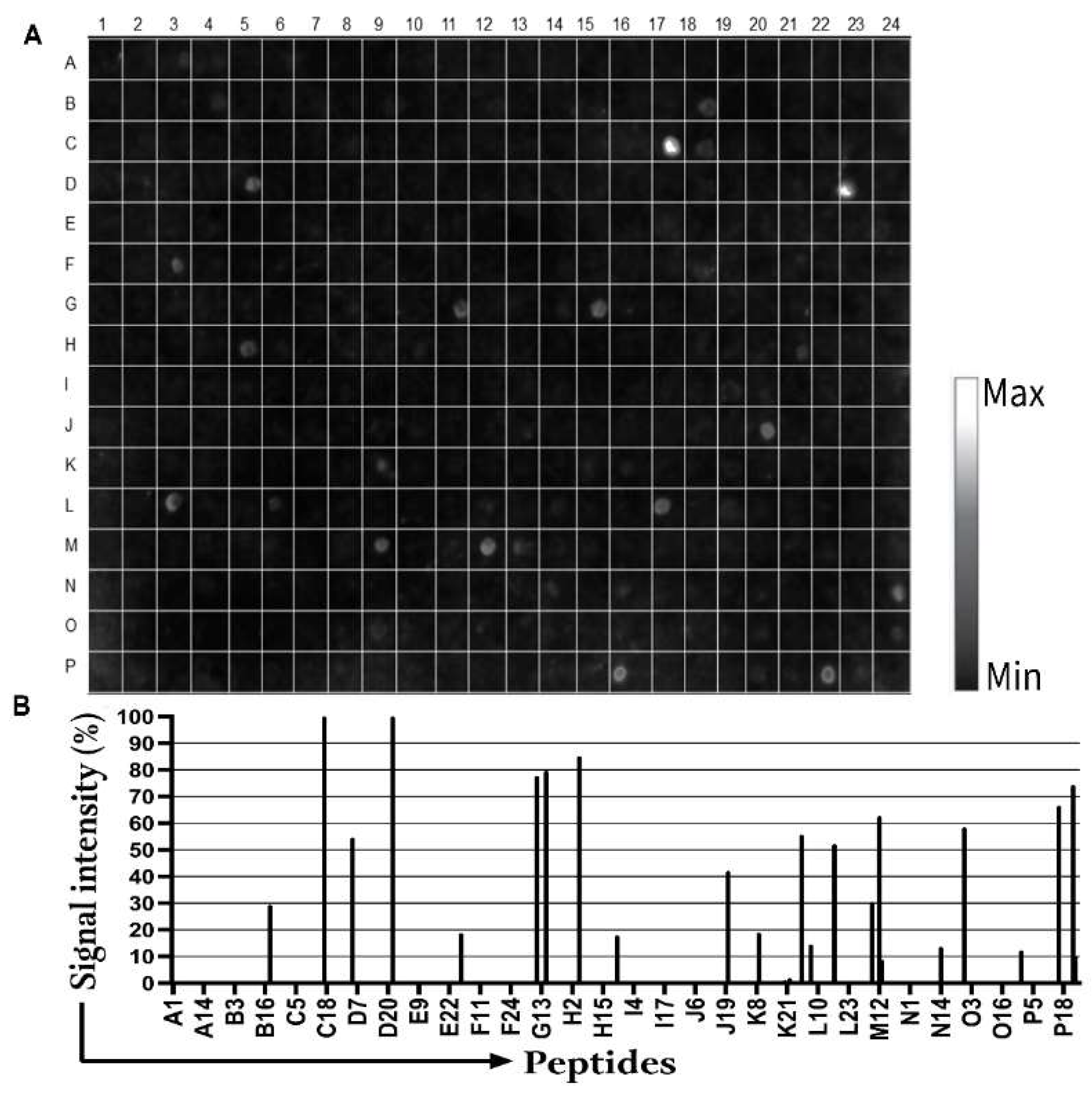
Proteins are known to contain a finite number of epitopes that are recognized by antibodies, but their molecular interactions remain largely undefined. In the case of Borreliosis, only a few proteins have been extensively studied. Therefore, this study analyzed five membrane proteins by synthesizing a library of representative peptides. These peptides were incubated with human seropositive samples and IgM secondary antibodies (see Materials and Methods). As shown in Figure 2A, the analysis of signal intensity (with a cutoff above 30%) of the primary sequences from the proteins containing B cell epitopes in the 384-peptide library (Figure 2B and Table S1) identified 14 peptides that were reactive to IgM from a pool of patient's sera. Table 1 summarizes the immunoreactivities of the identified peptides, which include two peptides from the flagellar hook protein (Bburg/01/huM and Bburg/02/huM), two peptides from the 41-kDa flagellin core protein (Bburg/03/huM and Bburg/04/huM), four peptides from the flagellar hook-associated protein 2 (Bburg/06/huM to Bburg/09/huM), two peptides from the putative outer membrane protein BBA03 (Bburg/10/huM and Bburg/11/huM), and two peptides from outer surface protein A (Bburg/12/huM and Bburg/13/huM).
Figure 1.
Mapping of linear B-cell epitopes in proteins (Q44767, P11089, O51173, Q44849, and P0CL66) from B. burgdorferi using a pool of sera (n=7) from Borreliosis: (A) Chemiluminescent assay image showing IgM-reactive peptides. (B) Graph depicting the signal intensity of the immunoreactive peptides. The x-axis represents the peptide distribution on the membrane, and the y-axis shows the relative signal intensity in percentage.
Figure 1.
Mapping of linear B-cell epitopes in proteins (Q44767, P11089, O51173, Q44849, and P0CL66) from B. burgdorferi using a pool of sera (n=7) from Borreliosis: (A) Chemiluminescent assay image showing IgM-reactive peptides. (B) Graph depicting the signal intensity of the immunoreactive peptides. The x-axis represents the peptide distribution on the membrane, and the y-axis shows the relative signal intensity in percentage.
3.2. Secondary Structure and Structural Mapping of the IgM Epitopes
The I-TASSER service predicted the protein's secondary structure components in detail. Molecular modeling generated five protein models, ranked according to their C-scores and TM-scores, with the lowest values appearing first. The models with the highest C-scores and TM-scores were chosen based on their secondary structures. Table 1 illustrates the secondary structures of the 14 peptides containing epitopes ranging from 4 to 14 amino acids and their positions within each protein. These peptides exhibited reactive epitopes in their secondary structures' coil, helix, and strand regions. PIR alignment showed that four peptides exhibited similarities to peptide sequences from various Borrelia species, while the remaining peptides shared homology with sequences from other animals (Table 1).

Table 1.
List of B-cell linear IgM epitopes with signal intensity ≥ 30%, Peptide Match, secondary structure (C, coil; H, helix; S, strand) predicted by I-TASSER (accessed on 20 November 2022).
Table 1.
List of B-cell linear IgM epitopes with signal intensity ≥ 30%, Peptide Match, secondary structure (C, coil; H, helix; S, strand) predicted by I-TASSER (accessed on 20 November 2022).
| Protein | Epitope | 2nd structure* | a number | Peptide match** |
|---|---|---|---|---|
| FlgE | Bburg/01/huM | C | 209 SLYDSFGN 216 | Various Borrelia sp |
| Bburg/02/huM | C | 321 GYGMGYME 328 | Various Borrelia sp | |
| Bburg/03/huM | C+S | 381 VRIGETGLAGLGDIR 395 | Another organism | |
| Flg41 kDa | Bburg/04/huM | H+C | 121 ANLSKTQEKLSSGYR 135 | Another organism |
| Bburg/05/huM | H+C | 322 AQANQVPQYVLSLLR 336 | Another organism. | |
| Flg hook2 | Bburg/06/huM | C | 08 PGLESKYN 15 | Various Borrelia sp |
| Bburg/07/huM | C+S | 76 SGNSSNSEVLTLSTR 90 | Another organism | |
| Bburg/08/huM | C+S | 391 AENAKIKFDGVDVER 405 | Another organism | |
| Bburg/09/huM | H+C+S | 546 RYLRLDEKKFDESIR 560 | Another organism | |
| Bburg/10/huM | H | 616 QKNKVEDYKKKYEDR 630 | Another organism | |
| BBA03 | Bburg/11/huM | C | 31 DEKSQAKSNLVD 42 | Another organism |
| Bburg/12/huM | H+C | 46 IEFSKATPLEKLVSR 60 | Another organism | |
| OSP A | Bburg/13/huM | C | 56 ATVDKLELKGTSDKN 70 | Various Borrelia sp |
| Bburg/14/huM | C | 266 EGSAVEITKL 275 | Another organism |
C, coil; H, helix; S, strand; * based on an AlphaFold analysis; ** PIR (Protein Information Resource).
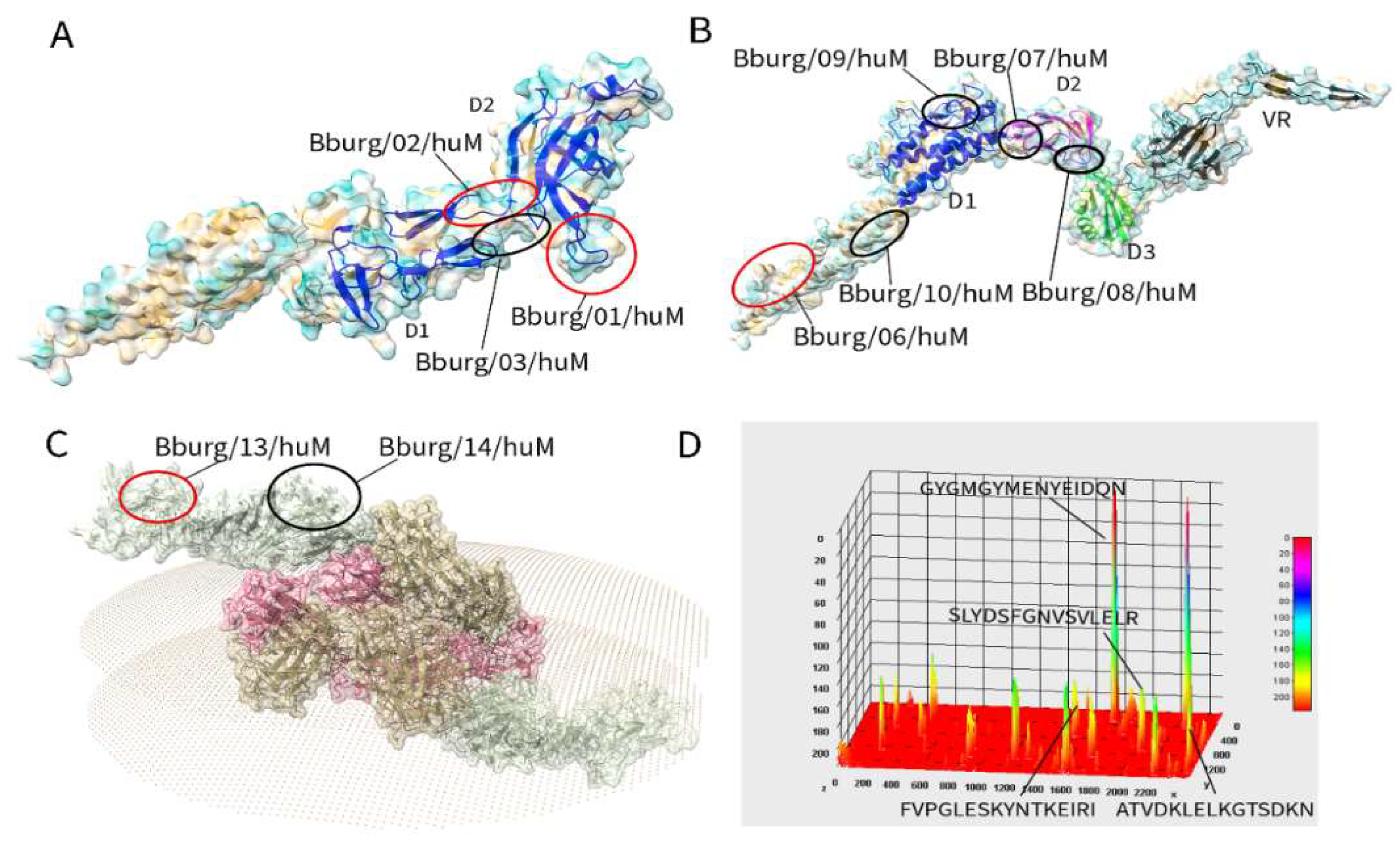
The spatial localization of linear B-cell IgM epitopes was identified and mapped onto 3D protein structures, as illustrated in Figure 2. Figure 2A,B represent flagellar proteins, while Figure 2C displays a surface protein. Red-circled epitopes were selected for validation, highlighting their exposed surface positioning, which enhances accessibility for antibody binding. The spatial arrangement of coil within the outer membrane protein was examined via 3D structural analysis using ChimeraX, which revealed that these epitopes reside within the coiled-coil region of the flagellar protein, located between domain 1 (D1) and domain 2 (D2) and extend near the N-terminal.
Figure 2D shows the surface cellulose presenting the sequence and increasing signal intensity, providing a detailed visual representation of the reactivity with IgM antibodies and correlates with greater serum levels of the target, suggesting differences in epitope immunogenicity.
Figure 2.
3D structural model protein fromBorrelia burgdorferi and localization of the immunodominant IgM epitopes. The picture represents the location of the epitopes on the ribbon diagram of FlgE (Figure 2A), (Osp A) outer surface protein (Figure 2B), (Flg 41 kDa) flagellar filament protein (Figure 2C) and (Flg hook 2) flagellar protein (Figure 2D). The molecular modelling of the proteins was based on homology using AlphaFold v2.0 scrip and crystal structure of OSPA (PDB:1fj1). The fourteen highly immunogenic IgM epitopes mapped are highlighted in blue, and the four epitopes used for the peptide-ELISA assay are depicted.
Figure 2.
3D structural model protein fromBorrelia burgdorferi and localization of the immunodominant IgM epitopes. The picture represents the location of the epitopes on the ribbon diagram of FlgE (Figure 2A), (Osp A) outer surface protein (Figure 2B), (Flg 41 kDa) flagellar filament protein (Figure 2C) and (Flg hook 2) flagellar protein (Figure 2D). The molecular modelling of the proteins was based on homology using AlphaFold v2.0 scrip and crystal structure of OSPA (PDB:1fj1). The fourteen highly immunogenic IgM epitopes mapped are highlighted in blue, and the four epitopes used for the peptide-ELISA assay are depicted.
3.3. ELISA Screening
The reactivity index (IR) and gray zone (GZ) were obtained by ELISA assay using peptides Bburg/01/huM, Bburg/02/huM, Bburg/06/huM and Bburg/13/huM. The result obtained is shown in Figure 3. The selection of four peptides reactive to IgM antibody was based on signal strength, secondary structure, and peptide match. The peptide-ELISA was evaluated on a panel of 19 seropositive, 39 seronegative, and suspected case samples obtained for three months (n=11) and less than three months (n=13). The grey zone (GZ) to RI value was 1.0% ± 10%.
Figure 3.
Reactivity Index (RI) of synthetic peptide reactive in individuals with anti-Borrelia IgM (n= 19), non-infected (n=39), Infected SC < 3 months (n= 13), SC > 3 months (n=11). The dashed line represents the reactivity index (RI) cutoff value. The area delimited by a gray rectangle indicates the indeterminate zone (RI ±10%). Significance level P: (ns) P > 0.05; (**) P ≤ 0.01; (******) P ≤ 0.0001. SC, Suspected case.
Figure 3.
Reactivity Index (RI) of synthetic peptide reactive in individuals with anti-Borrelia IgM (n= 19), non-infected (n=39), Infected SC < 3 months (n= 13), SC > 3 months (n=11). The dashed line represents the reactivity index (RI) cutoff value. The area delimited by a gray rectangle indicates the indeterminate zone (RI ±10%). Significance level P: (ns) P > 0.05; (**) P ≤ 0.01; (******) P ≤ 0.0001. SC, Suspected case.
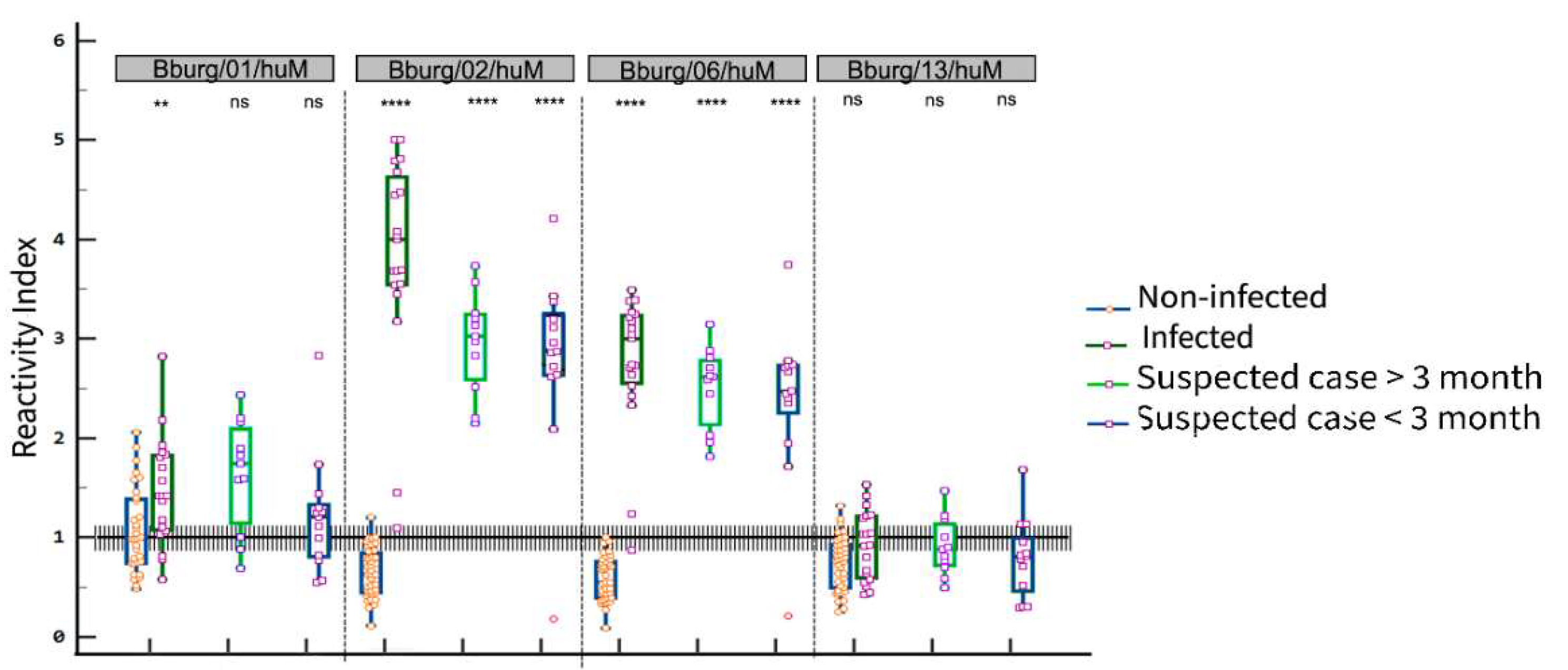
Table 2 presents the performance of peptides in an ELISA assay with samples from patients with Borreliosis. The cutoff was determined using the ROC curve, and the peptides (Bburg/02/huM and Bburg/06/huM) demonstrated a potential capacity to detect positive cases with a sensitivity of ≥ 94.74%. Specificity was ≥ 97.44%, as seen in Table 2, suggesting strong specificity, which means they could properly identify negative cases.
On the other hand, peptides (Bburg/01/huM and Bburg/13/huM) showed an antagonistic profile concerning sensitivity and specificity. According to Table 2, peptide Bburg/01/huM presented high sensitivity. However, the specificity was slightly lower, equivalent to 56.2%. The performance of peptide Bburg/13/huM revealed a sensitivity of 47.37%, suggesting a relatively low capacity to identify the infected individuals. However, the specificity, which is the ability to identify the non-infected, was high.
Furthermore, the (Area under the curve) AUC was determined for each peptide, and the findings showed that peptides Bburg/02/huM and Bburg/06/huM 162 were highly accurate. The accuracy of peptide Bburg/01/huM was good, suggesting a good performance in identifying between infected and non-infected patients. Peptide Bburg/13/huM, on the other hand, had a lower AUC value, indicating a less accurate performance. According to the analysis, the peptides were evaluated and found to have a significant discriminatory ability with a P-value or significance level of (P< 0,0001) to peptides (Bburg/02/huM and Bburg/06/huM), Bburg/01/huM (P= 0,0052), and Bburg/13/huM (P= 0,0703).
We saw that the peptides Bburg/02/huM and Bburg/06/huM used in the ELISA assays with samples from suspected cases exhibited a sensitivity of ≥ 92.3% and a specificity of 100.0% (Table 2). Additionally, the sensitivity and specificity for the peptides Bburg/01/huM and Bburg/13/huM were 72.7% and 56.2%, respectively, with no significant variation.
Table 2.
The diagnostic assessment of the ELISA-peptide and Receiver operating characteristic curve (ROC) with 95% confidence intervals.
Table 2.
The diagnostic assessment of the ELISA-peptide and Receiver operating characteristic curve (ROC) with 95% confidence intervals.
| Suspected case | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Infected | > 3 months | < 3 months | |||||||||||
| Peptide | Se (%) | Sp (%) | AUC | Ac (%) | Se (%) | Sp (%) | AUC | Se (%) | Sp (%) | AUC | |||
| Bburg/01/huM | 82.4 | 56.2 | 0.721 | 66.6 | 72.7 | 56.2 | 0.557 | 76.9 | 59.4 | 0.671 | |||
| Bburg/02/huM | 100.0 | 97.44 | 0.999 | 96.2 | 100.0 | 100.0 | 1 | 92.3 | 100.0 | 0.925 | |||
| Bburg/06/huM | 94.74 | 100.0 | 0.996 | 98.2 | 100.0 | 100.0 | 1 | 92.3 | 100.0 | 0.925 | |||
| Bburg/13/huM | 47.37 | 87.18 | 0.652 | 74.1 | 72.7 | 59.0 | 0.622 | 76.9 | 59.4 | 0.694 | |||
The area under the curve (AUC), Accuracy (Ac), sensitivity (Se), and specificity (Sp)
3.4. Potential cross-reactivity of the epitopes and validation of bi-specific peptides as antigens
The BLASTP analysis of B. burgdorferi proteins [Flagellar E (FlgE, Q44849) and outer surface protein A (OspA, P0CL66)] examined in this study revealed a significant alignment with proteins from other Borrelia species, indicating that the identified Borrelia-specific epitopes are universal. In contrast, the flagellar filament 41 kDa protein (Flg 41 kDa, Q4767), flagellar hook-associated protein 22 (Flg hook 2, PI1089), and Omp BBA03 (Omp BBA03, O51173) showed sequence similarities ranging from 39.29% to 57.4% with Treponema pallidum and Leptospira interrogans (Table S2).
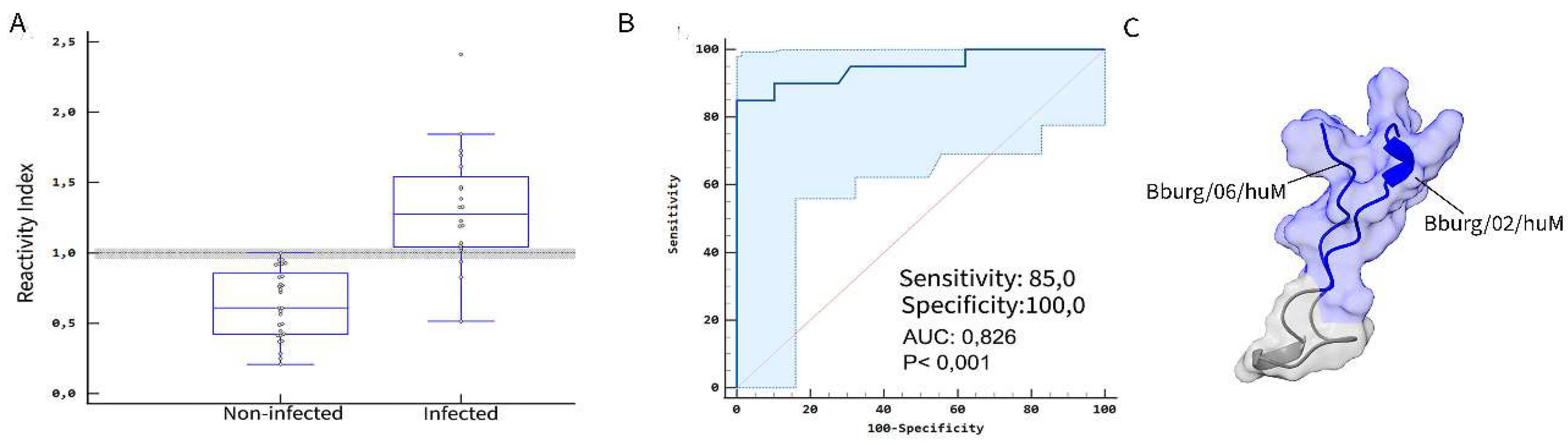
To validate the specificity of the selected epitopes, we employed MAPs4 bi-specific antigen (FlgE /Flg Hook 2) peptides (Bburg/02/huM and Bburg/06/huM) as antigens and the patients' sera. ELISA test results showed a similar sensitivity (85.0%) and specificity (100.0%) with a p-value of 0.001 (Figure 4), like the ELISA with single peptides (Table 3).
Figure 4.
Performance parameters of the bi-specific antigen peptides for diagnostic confirmation. (A) The Scatter plots show the reactivity of the polypeptide and IgM by peptide-based ELISA of the serum sample negative (n=29) and serum sample B. burgdorferi-infected individuals (n=20). The dashed line represents the gray zone, which was defined as positive (> 1.1), indeterminate (1.1- 09), or negative (< 0.9). (B) ROC (receiver operating characteristic) curve for the ELISA-polypeptide technique [cutoff, sensitivity, specificity, and AUC (area under curve) are depicted]. (C) bi-epitope peptide structure prediction.
Figure 4.
Performance parameters of the bi-specific antigen peptides for diagnostic confirmation. (A) The Scatter plots show the reactivity of the polypeptide and IgM by peptide-based ELISA of the serum sample negative (n=29) and serum sample B. burgdorferi-infected individuals (n=20). The dashed line represents the gray zone, which was defined as positive (> 1.1), indeterminate (1.1- 09), or negative (< 0.9). (B) ROC (receiver operating characteristic) curve for the ELISA-polypeptide technique [cutoff, sensitivity, specificity, and AUC (area under curve) are depicted]. (C) bi-epitope peptide structure prediction.
An ELISA assay assessed cross-reactivity with serum samples from leptospirosis and syphilis cases. Analysis of polypeptide-specific anti-Borreliosis IgM antibodies with the Kruskal-Walli's test revealed a significant difference in IgM antibody reactivity across leptospirosis and syphilis (p < 0.0001), as shown in Figure 5.
The polypeptide demonstrated significantly higher reactivity to anti-Borreliosis IgM antibodies than other diseases, as indicated by the median values in Figure 5 and Table 3. Additionally, Dunn's multiple comparisons test, conducted as a post-hoc analysis to pinpoint specific pairwise differences, confirmed that the polypeptide's IgM antibody reactivity was significant. Pairwise comparisons using LB as a reference showed a substantial difference between leptospirosis (p < 0.0424) and syphilis (p < 0.0001). This indicates a significant difference between the disease groups, as supported by the median and P values (Figure 5 and Table 3).
Figure 5.
Box and whisker plot comparing ELISA-polypeptide IgM cross-reactivity using sera samples from borreliosis (LB, n=20), leptospirosis (LEP, n=20), and syphilis (SIF, n=20) individuals. The Kruskal-Walli’s test result graph displaying the p-value (P<0.0001 (****); P= 0,0424 (*)).
Figure 5.
Box and whisker plot comparing ELISA-polypeptide IgM cross-reactivity using sera samples from borreliosis (LB, n=20), leptospirosis (LEP, n=20), and syphilis (SIF, n=20) individuals. The Kruskal-Walli’s test result graph displaying the p-value (P<0.0001 (****); P= 0,0424 (*)).
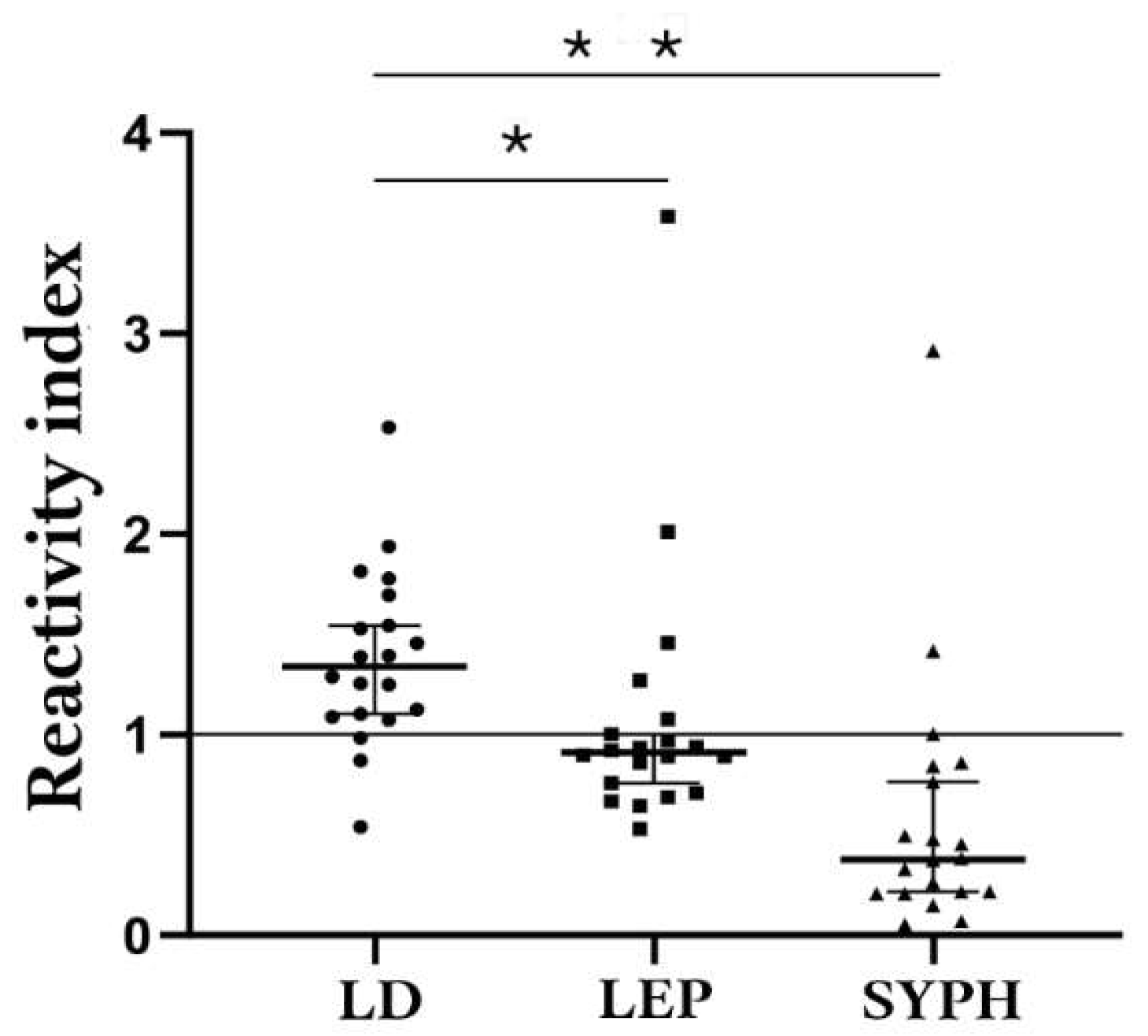
Table 3.
Kruskal-Walli’s test analysis of the reactivity of the Borrelia-polypeptide with sera from leptospirosis and syphilis individuals.
Table 3.
Kruskal-Walli’s test analysis of the reactivity of the Borrelia-polypeptide with sera from leptospirosis and syphilis individuals.
| Comparison Group | Median | 75% Percentile | 25% Percentile |
|---|---|---|---|
| Lyme-Borreliosis | 0,2613 | 0,3235 | 0,2131 |
| Leptospirosis | 0,0755 | 0,08788 | 0,060 |
| Syphilis | 0,02175 | 0,04738 | 0,01213 |
4. Discussion
In this study, we investigate the antigenic properties of five surface proteins of B. burgdorferi to find Borrelia-specific epitopes. Through this approach, we successfully identified several linear IgM epitopes, including sequences derived from FlgE, the 41 kDa flagellin core, and the flagellar hook-associated protein FliD, also known as Hook-associated protein 2.
Notably, identifying epitopes in conserved regions of these proteins underscores their potential to elicit a robust and specific antibody response. This finding highlights their relevance as potential targets for neutralizing antibodies, as conserved regions are less likely to undergo antigenic variation, enhancing diagnostic and therapeutic utility [49,50,51,52,53]. These results provide a strong foundation for further exploration of these epitopes in developing diagnostic assays or immune-based interventions for Borrelia infections.
The localization of the Bburg/12/huM epitope was found in the α1 region Bburg/13/huM and Bburg/14/huM were located at the N-terminus and C-terminal regions, respectively, which are considered conserved regions. [50,51]. Information regarding the location of the final exposed epitope on the surface was obtained from previous research that detailed the N-terminal region (residues 16–27) of OspA, which is a structurally flexible tethering domain of only 12 residues [54,55].
Synthetic peptides have been gaining prominence in diagnostic tests because of their low cost, rapid production (without worrying about cell culture and protein purification), and high sensitivity and specificity [45,56,57].
The FliD protein, also known as flagellar hook-associated protein 2, plays a key role in the assembly and stability of the B. burgdorferi flagellum [58]. In this study, the mapping of epitopes recognized by sera from patients with Lyme disease revealed the sequence 8PGLESKYN15 in the N-terminal region of FliD [58]. The C- and N-terminus regions, as well as D1, are highly conserved [58]; the presence of an epitope in this region suggests that in addition to generating a robust production of specific antibodies in response to infection, it may be a relevant target for neutralizing antibodies. In this study, we identified FlgE epitopes that have great potential as targets for diagnosing Borreliosis in patients [5]. Among them, the epitopes 321GYGMGYME328 and 381VRIGETGLAGLGDIR395 located between the D1 and D2 domains of the FlgE protein stand out as a possible diagnostic marker, given their location in the structure and accessibility. The structure of the Borrelia FlgE protein is composed of three main domains: D0, which forms the inner core of the tubular structure, and the D1 and D2 domains, which integrate the outer layers of the structure [59]. The specific intermolecular cross-linking of lysin-alanine (Lal) between the D1 and D2 domains plays a central role in the structural stability of the flagellar protein. The structural relevance of Lal in FlgE was highlighted by the study with Treponema denticola FlgE, deficient in Lal cross-linking, which presented normal hook assembly and cell morphology but with impaired motility, as described by Lynch et al. [54]. This data indicates that Lal cross-linking may play a key role not only in the structure but also in the function of the flagellum, highlighting Lal as a possible antimicrobial target to inhibit pathogenic spirochetes. The detection of the flagellar hook gene (flgE) in blood samples from individuals diagnosed with Borrelia infection corroborates the applicability of this epitope in diagnostic strategies.
We also identified two peptides belonging to the putative outer membrane protein BBA03 and two peptides belonging to outer surface protein A. A previous study showed that the localization of these highly immunogenic and diverse proteins on the membrane allows their exposure [50]. The epitopes identified in the BBA03 protein in the coil and α1 region in this study corroborate the understanding of the functional role of this protein in B. burgdorferi [55]. Its conformation presents seven α-helices and regions conserved among different genera of Borrelia, which may confer protection against multiple strains. The function of BBA03 becomes even more prominent in mixed infections, where B. burgdorferi coexists with other spirochetes [55], suggesting its importance in competitive environments.
Epitope 56ATVDKLELKGTSDKN70 e 266EGSAVEITKL275 mapped in our study corresponds with previously identified antibody binding. The first epitope is found in the conserved region, mainly among the six OspA serotypes [56,57]. Of the two identified, EGSAVEITKL is found in a poorly conserved region [50,51]. OspA has been widely studied as an immunogen, only providing 76-92% protection [58-60]. Although it is a surface protein, it has variations among different serotypes of B. burgdorferi; the epitopes may vary depending on the prevalent serotype in the region studied [61-63].
Synthetic peptides have been gaining prominence in diagnostic tests because of their low cost, rapid production (without worrying about cell culture and protein purification), and high sensitivity and specificity [64,65]. Therefore, to assess the performance of the mapped epitopes, we synthesized four Borrelia epitopes and evaluated their immunoreactivity in anti-IgM testing using ELISA.
The peptide Bburg/01/huM demonstrated a sensitivity of 82.4% and specificity of 56.2%, whereas the peptide Bburg/13/huM had a sensitivity of 47.37% and specificity of 87.18%. These two peptides showed antagonistic profiles, indicating a limited ability to identify infected individuals but a remarkable capacity to identify non-infected individuals. The latter result is consistent with studies that showed low sensitivity for early acute infections [66,67]. In contrast, peptides Bburg/02/huM and Bburg/06/huM demonstrated a high sensitivity of ≥94.74% and specificity of ≥97.44%c (Table 2).
The existence of different serotypes could generate a variation in the reactivity of the patient's serum, thus influencing the identification of epitopes. However, a blast performed with the main pathogenic strains showed that the selected peptides are genus-specific.
Though commercial MTTT tests show consistency comparable to the standard method, previous studies showed that the combination of selected epitopes can result in higher sensitivity (76.0%) and specificity (97.4%) [68,69]. Therefore, combining single epitopes is expected to result in tests with better sensitivity and specificity [70]. In fact, recently, a point-of-care serological assay for LBD leveraging IgM-specific synthetic peptides derived from different proteins was used in a paper-based platform for rapid and cost-effective diagnosis has been developed [48].
We produced bi-specific peptides, but the optical density obtained was inferior to the single peptides. In our study, the ELISA results using the chimeritope revealed a sensitivity of 85.0%, specificity of 100.0%, and p-value of 0.001. However, previous studies have revealed cross-reactivity between serological tests for syphilis [71], viral infections [72], and IgG anti-Borrelia antibodies in Yersinia. Given the above, we proceeded with an assay to evaluate polypeptides specific to IgM antibodies in borreliosis using serum samples obtained from individuals diagnosed with leptospirosis and syphilis. The results revealed a substantial difference between the disease groups, as evidenced by the median and p-values.
Although B. burgdorferi antigens contain private antigenic determinants, studies have shown that some common epitopes can cross-react with other pathogens and antibodies associated with autoimmune diseases [65, 73]. To address this, we analyzed the seroreactivity of populations with a high prevalence of these co-occurring infections, such as leptospirosis and syphilis patients, to the B. burgdorferi bi-epitope antigen. The results revealed significant differences between the disease groups, as evidenced by median and p-values. Therefore, the findings suggest that the chimeritope antigen can effectively distinguish between sera from patients with borreliosis and leptospirosis, syphilis, and healthy individuals (Figure 4). However, a low immune response was observed in some patients. This highlights the potential influence of genetic factors or time of sera collection on serological reactivity. This hypothesis warrants further investigation in larger cohorts to better understand the variability in immune responses and refine diagnostic accuracy
5. Conclusions
By integrating previously established procedures, including Spot-synthesis and BLAST homology searches, this study successfully identified IgM target epitopes with significant potential for use in a specific early diagnostic assay for Borreliosis. The method demonstrated high sensitivity and specificity, underscoring its effectiveness in detecting private key epitopes. The tested bi-epitope polypeptides yielded promising results, notably eliminating false-positive rates.
The study highlights the reliability of these epitopes as diagnostic markers for early Borrelia infections, attributed to their lack of cross-reactivity with other relevant conditions and their conservation across Borrelia genospecies. These findings pave the way for developing enhanced diagnostic tools for Borrelia infections, enabling more accurate diagnoses and improved treatment strategies. Moreover, combining multiple peptide epitopes in a chimeric protein or with a machine learning-based diagnostic model further enhances sensitivity without compromising specificity, showcasing a synergistic approach to advanced diagnostic solutions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. List of peptides synthesized for the analysis of IgM epitopes from five proteins of B. burgdorferi. [UniProt protein code 44767-Flagelar hoot open (spot A1-D15)]; [UniProt code P11089-Filament flagellar 41 kDa (spot D18-G11)]; (UniProt code O51173-Flagelar hoot protein 2 (spot G14-L24); [UniProt code Q4489-Putative Omp BURGA03 (spot M3-N10)]; [UniProt code P0CL66-Omp A (spot N13-P17)]. Table S2. List of B. burgdorferi proteins used in this study and percentage sequence identity to other organism’s proteins.
Author Contributions
Conceptualization and investigation: S.G.D.-S.; methodology: R.M.R.P., M.E.T.A.-C., V.L.N.B., G.S.G., A.L.A.S., P.N.-P., A.M.D., and J.P.R.S.C.; funding: S.G.D.-S. computational analysis: M.E.T.A.-C.; writing—original draft: M.E.T.A.-C. and S.G.D.-S. review and editing: S.G.D.-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Carlos Chagas Filho Foundation of Research Support of the State of Rio de Janeiro (FAPERJ #010.101.029/2018) and the Brazilian Council for Scientific Research (CNPq #301744/2019-0).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
The study was conducted according to the Declaration of Helsinki and approved by the Ethics Committee of Oswaldo Cruz Foundation (CAAE: 25836019.0.0000.5243) study center ethics committee.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
P.N-P.: G.C.L., and A.M.D. are postdoctoral fellows from the INOVA-FIOCRUZ Program. M.E.T.A.-C and J.P.R.S.C. are DSc fellows from the Federal Fluminense University, Post-Graduation Program on Science and Biotechnology. To Dr Virgínia L.N. Bonaldi and Dr Rosa M.R. Pereira (in memoriam) for easy access to patient serums.
Conflicts of Interest
The authors declare no disputes involving the research reported. The funding agencies had no role in the study design, data collection, data analysis, publication decision, or manuscript preparation.
References
- Mead, P.; Hinckley, A.; Kugeler, K. Lyme disease surveillance and epidemiology in the United States: A historical perspective. J Infect Dis 2024, 230, S11–S17. [Google Scholar] [CrossRef]
- Burn, L.; Vyse, A.; Pilz, A.; Tran, T.M.P.; Fletcher, M.A.; Angulo, F.J.; Gessner, B.D.; Moïsi, J.C.; Stark, J.H. Incidence of Lyme Borreliosis in Europe: A systematic review (2005-2020). Vector Borne Zoonotic Dis 2023, 23, 172–194. [Google Scholar] [CrossRef] [PubMed]
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J Public Health 2016, 39, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, N.H.; Bonoldi, V.L.N.; Bonin, S.; Falkingham, E.; Trevisan, G. The current state of knowledge on Baggio-Yoshinari syndrome (Brazilian Lyme Disease-like Illness): Chronological presentation of historical and scientific events observed over the last 30 Years. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Bonoldi, V.L.N.; Yoshinari, N.H.; Trevisan, G.; Bonin, S. Baggio-Yoshinari syndrome: A report of five cases. Microorganisms. 2024, 12, 2108. [Google Scholar] [CrossRef] [PubMed]
- Jorge, F.R.; Muñoz-Leal, S.; de Oliveira, G.M.B.; Serpa, M.C.A.; Magalhães, M.M.L.; de Oliveira, L.M.B.; Moura, F.B.P.; Teixeira, B.M.; Labruna, M.B. Novel Borrelia genotypes from Brazil indicate a new group of Borrelia spp. associated with South American Bats. J Med Entomol 2023, 60, 213–217. [Google Scholar] [CrossRef]
- Ohnishi, J.; Piesman, J.; de Silva, A.M. Antigenic and genetic heterogeneity of Borrelia burgdorferi populations transmitted by ticks. Proc Natl Acad Sci U S A 2001, 98, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Miziara, C.S.M.G.; Gelmeti Serrano, V.A.; Yoshinari, N. Passage of Borrelia burgdorferi through diverse Ixodid hard ticks causes distinct diseases: Lyme borreliosis and Baggio-Yoshinari syndrome. Clinics (Sao Paulo) 2018, 73, e394. [Google Scholar] [CrossRef]
- Wodecka, B.; Kolomiiets, V. Genetic diversity of Borreliaceae species detected in natural populations of Ixodes ricinus ticks in northern Poland. Life 2023, 13, 972. [Google Scholar] [CrossRef]
- Karageorgou, I.; Koutantou, M.; Papadogiannaki, I.; Voulgari-Kokota, A.; Makka, S.; Angelakis, E. Serological evidence of possible Borrelia afzelii Lyme disease in Greece. New Microbes New Infect 2022, 46, 100978. [Google Scholar] [CrossRef]
- Mancilla-Agrono, L.Y.; Banguero-Micolta, L.F.; Ossa-Lopez, P.A.; Ramirez-Chaves, H.E.; Castano-Villa, G.J.; Rivera-Paez, F.A. Is Borrelia burgdorferi sensu stricto in South America? First molecular evidence of its presence in Colombia. Trop Med Infect Dis 2022, 7, 428. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Horak, A.; Grubhoffer, L.; Mongodin, E.F.; Fraser, C.M.; Qiu, W.; Luft, B.J.; Morgan, R.G.; Casjens, S.R.; et al. , Genomic confirmation of Borrelia garinii. United States Emerg Infect Dis 2023, 29, 64–69. [Google Scholar] [CrossRef]
- Hansmann, Y.; Leyer, C.; Lefebvre, N.; Revest, M.; Rabaud, C.; Alfandari, S.; Christmann, D.; Tattevin, P. Feedback on difficulties raised by interpreting serological tests for Lyme disease diagnosis. Med Mal Infect 2014, 44, 199–205. [Google Scholar] [CrossRef]
- Kodym, P.; Kurzová, Z.; Berenová, D.; Pícha, D.; Smíšková, D.; Moravcová, L.; Malý, M. Serological diagnostics of Lyme Borreliosis: Comparison of universal and Borrelia species-specific tests based on whole-cell and recombinant antigens. J Clin Microbiol 2018, 56, e00601–18. [Google Scholar] [CrossRef] [PubMed]
- Ružić-Sabljić, E.; Cerar, T. Progress in the molecular diagnosis of Lyme disease. Expert Rev Mol Diagn. 2017, 17, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Eldin, C.; Parola, P.; Raoult., D. Limitations of diagnostic tests for bacterial infections. Med Mal Infect. 2019, 49, 98–101. [Google Scholar] [CrossRef]
- Toumanios C, Prisco L, Dattwyler RJ, Arnaboldi PM. Linear B cell epitopes derived from the multifunctional surface lipoprotein BBK32 as targets for the serodiagnosis of Lyme disease. mSphere 2019, 4, e00111-19. [Google Scholar] [CrossRef]
- Tokarz R, Guo C, Sanchez-Vicente S, Horn E, Eschman A, Turk SP, Lipkin WI, Marques A. Identification of reactive Borrelia burgdorferi peptides associated with Lyme disease. mBio 2024, 15, e0236024. [Google Scholar] [CrossRef]
- Durans AM, Napoleão-Pêgo P, Reis FCG, Dias ER, Machado LESF, Lechuga GC, Junqueira ACV, De-Simone SG, Provance-Jr DW. Chagas disease diagnosis with Trypanosoma cruzi exclusive epitopes in GFP. Vaccines (Basel) 2024, 12, 1029. [Google Scholar]
- Guérin, M.; Shawky, M.; Zedan, A.; Octave, S.; Avalle, B.; Maffucci, I.; Padiolleau-Lefèvre, S. Lyme borreliosis diagnosis: state of the art of improvements and innovations. BMC Microbiol. 2023, 23, 204. [Google Scholar] [CrossRef] [PubMed]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; Vanraden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein is required exclusively in a crucial early stage of mammalian infection. Infect Immun 2006, 74, 3554–3564. [Google Scholar] [CrossRef]
- Battisti, J.M.; Bono, J.L.; Rosa, P.A.; Schrumpf, M.E.; Schwan, T.G.; Policastro, P.F. Outer surface protein A protects Lyme disease spirochetes from acquired host immunity in the tick vector. Infect Immun 2008, 76, 5228–5237. [Google Scholar] [CrossRef]
- Bykowski, T.; Woodman, M.E.; Cooley, A.E.; Brissette, C.A.; Wallich, R.; Brade, V.; Kraiczy, P.; Stevenson, B. Borrelia burgdorferi complement regulator-acquiring surface proteins (BbCRASPs): Expression patterns during the mammal-tick infection cycle. Int J Med Microbiol 2008, 298, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Brissette, C.A.; Haupt, K.; Barthel, D.; Cooley, A.E.; Bowman, A.; Skerka, C.; Wallich, R.; Zipfel, P.F.; Kraiczy, P.; Stevenson, B. Borrelia burgdorferi infection-associated surface proteins ErpP, ErpA, and ErpC bind human plasminogen. Infect Immun 2009, 77, 300–306. [Google Scholar] [CrossRef]
- Xu, H.; Sultan, S.; Yerke, A.; Moon, K.H.; Wooten, R.M.; Motaleb, M.A. Borrelia burgdorferi CheY2 is dispensable for chemotaxis or motility but crucial for the infectious life cycle of the spirochete. Infect Immun 2017, 85. [Google Scholar] [CrossRef]
- Moon, K.H.; Hobbs, G.; Motaleb, M.A. Borrelia burgdorferi CheD promotes various functions in chemotaxis and the pathogenic life cycle of the spirochete. Infect Immun 2016, 84, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- Pulzova, L.; Bhide, M. Outer surface proteins of Borrelia: peerless immune evasion tools. Curr Protein Pept Sci 2014, 15, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Carroll, B.L.; Liu, J. Structural basis of bacterial flagellar motor rotation and switching. Trends Microbiol 2021, 29, 1024–1033. [Google Scholar] [CrossRef]
- Zhang, K.; Qin, Z.; Chang, Y.; Liu, J.; Malkowski, M.G.; Shipa, S.; Li, L.; Qiu, W.; Zhang, J.R.; Li, C. Analysis of a flagellar filament cap mutant reveals that HtrA serine protease degrades unfolded flagellin protein in the periplasm of Borrelia burgdorferi. Mol Microbiol 2019, 111, 1652–1670. [Google Scholar] [CrossRef]
- Rutemark, C.; Alicot, E.; Bergman, A.; Ma, M.; Getahun, A.; Ellmerich, S.; Carroll, M.C.; Heyman, B. Requirement for complement in antibody responses is not explained by the classic pathway activator IgM. Proc Natl Acad Sci U S A 2011, 108, E934–942. [Google Scholar] [CrossRef] [PubMed]
- Markowicz, M.; Reiter, M.; Gamper, J.; Stanek, G.; Stockinger, H. Persistent anti-Borrelia IgM antibodies without Lyme Borreliosis in the clinical and immunological context. Microbiol Spectr 2021, 9, e0102021. [Google Scholar] [CrossRef]
- Stavnezer J, Amemiya CT. Evolution of isotype switching. Semin. Immunol. 2004, 16, 257–275. [Google Scholar] [CrossRef]
- Chino, M.E.T.A.; Bonoldi, V.L.N.; Pereira, R.M.R.; Gazeta, G.S.; Carvalho, J.P.R.S.; Napoleão-Pêgo, P.; Durans, A.M.; Souza, A.L.A.; De-Simone, S.G. New epitopes for the serodiagnosis of human Borreliosis. Microorganisms 2024, 12, 2212. [Google Scholar] [CrossRef]
- Connolly, S.E.; Thanassi, D.G.; Benach, J.L. Generation of a complement-independent bactericidal IgM against a relapsing fever Borrelia. J Immunol 2004, 172, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Markowicz, M.; Reiter, M.; Gamper, J.; Stanek, G.; Stockinger, H. Persistent anti-Borrelia IgM antibodies without Lyme Borreliosis in the clinical and immunological context. Microbiol Spectr 2021, 9, e0102021. [Google Scholar] [CrossRef] [PubMed]
- Kalish, R.A.; McHugh, G.; Granquist, J.; Shea, B.; Ruthazer, R.; Steere, A.C. Persistence of immunoglobulin M or immunoglobulin G antibody responses to Borrelia burgdorferi 10-20 years after active Lyme disease. Clin Infect Dis 2001, 33, 780–785. [Google Scholar] [CrossRef]
- Bockenstedt LK, Wooten RM, Baumgarth N. Immune response to Borrelia: Lessons from Lyme disease spirochetes. Curr Issues Mol Biol. 2021, 42, 145–190. [Google Scholar] [CrossRef]
- Moore, A.; Nelson, C.; Molins, C.; Mead, P.; Schriefer, M. Current guidelines, common clinical pitfalls, and future directions for laboratory diagnosis of Lyme disease, United States. Emerg Infect Dis 2016, 22, 1169–1177. [Google Scholar] [CrossRef]
- Yoshinari, N.H.; Mantovani, E.; Bonoldi, V.L.; Marangoni, R.G.; Gauditano, G. Brazilian lyme-like disease or Baggio-Yoshinari syndrome: exotic and emerging Brazilian tick-borne zoonosis. Rev Assoc Med Bras 2010, 56, 363–369. [Google Scholar] [CrossRef]
- Haselbeck, A.H.; Im, J.; Prifti, K.; Marks, F.; Holm, M.; Zellweger, R.M. Serology as a tool to assess infectious disease landscapes and guide public health policy. Pathogens 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Signorino, G.; Arnaboldi, P.M.; Petzke, M.M.; Dattwyler, R.J. Identification of OppA2 linear epitopes as serodiagnostic markers for Lyme disease. Clin Vaccine Immunol 2014, 21, 704–711. [Google Scholar] [CrossRef]
- Arumugam, S.; Nayak, S.; Williams, T.; di Santa Maria, F.S.; Guedes, M.S.; Chaves, R.C.; Linder, V.; Marques, A.R.; Horn, E.J.; Wong, S.J.; et al. A multiplexed serologic test for diagnosis of Lyme disease for point-of-care use. J Clin Microbiol 2019, 57, e01142-19. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Morrissey, J.J.; Liu, L.; Wang, Y.; Zhou, Q.; Naik, R.R.; Singamaneni, S. Plasmonically enhanced ultrasensitive epitope-specific serologic assay for COVID-19. Anal Chem 2022, 94, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.S. Measuring molecular biomarkers in epidemiologic studies: Laboratory techniques and biospecimen considerations. Stat Med 2012, 31, 2400–2413. [Google Scholar] [CrossRef]
- De-Simone, S.G.; Napoleão-Pego, P.; Teixeira-Pinto, L.A.; Santos, J.D.; De-Simone, T.S.; Melgarejo, A.R.; Aguiar, A.S.; Marchi-Salvador, D.P. Linear B-cell epitopes in BthTX-1, BthTX-II and BthA-1, phospholipase A₂'s from Bothrops jararacussu snake venom, recognized by therapeutically neutralizing commercial horse antivenom. Toxicon 2013, 72, 90–101. [Google Scholar] [CrossRef]
- da Silva, F.R.; Napoleão-Pego, P.; De-Simone, S.G. Identification of linear B epitope of pertactin of Bordetela pertussis induced by immunization with whole and acellular vaccine. Vaccine 2014, 32, 6251–6258. [Google Scholar] [CrossRef]
- De-Simone, S.G.; Gomes, L.R.; Napoleão-Pêgo, P.; Lechuga, G.C.; de Pina, J.S.; da Silva, F.R. Epitope Mapping of the diphtheria toxin and development of an ELISA-specific diagnostic assay. Vaccines (Basel) 2021, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Ghosh, R.; Joung, H.A.; Goncharov, A.; Palanisamy, B.; Ngo, K.; Pejcinovic, K.; Krockenberger, N.; Horn, E.J.; Garner, O.B.; Ghazal, E.; et al. , Rapid single-tier serodiagnosis of Lyme disease. Nat Commun. 2024, 15, 7124. [Google Scholar] [CrossRef] [PubMed]
- Kenedy, M.R.; Lenhart, T.R.; Akins, D.R. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunol Med Microbiol 2012, 66, 1–19. [Google Scholar] [CrossRef]
- Wilske, B.; Luft, B.; Schubach, W.H.; Zumstein, G.; Jauris, S.; Preac-Mursic, V.; Kramer, M.D. Molecular analysis of the outer surface protein A (OspA) of Borrelia burgdorferi for conserved and variable antibody binding domains. Med Microbiol Immunol 1992, 181, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Wilske, B.; Luft, B.; Schubach, W.H.; Zumstein, G.; Jauris, S.; Preac-Mursic, V.; Kramer, M.D. Molecular analysis of the outer surface protein A (OspA) of Borrelia burgdorferi for conserved and variable antibody binding domains. Med Microbiol Immunol 1992, 181, 191–207. [Google Scholar] [CrossRef]
- Jiang, W.; Luft, B.J.; Schubach, W.; Dattwyler, R.J.; Gorevic, P.D. Mapping the major antigenic domains of the native flagellar antigen of Borrelia burgdorferi. J Clin Microbiol 1992, 30, 1535–1540. [Google Scholar] [CrossRef]
- Du, W.; Ma, X.; Nyman, D.; Povlsen, K.; Akguen, N.; Schneider, E.M. Antigen biochips verify and extend the scope of antibody detection in Lyme borreliosis. Diagn Microbiol Infect Dis 2007, 59, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.J.; Miller, M.; James, M.; Zhang, S.; Zhang, K.; Li, C.; Charon, N.W.; Crane, B.R. Structure and chemistry of lysinoalanine cross-linking in the spirochaete flagella hook. Nat Chem Biol 2019, 15, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Fridmanis, J.; Bobrovs, R.; Brangulis, K.; Tārs, K.; Jaudzems, K. Structural and Functional Analysis of BBA03, Borrelia burgdorferi competitive advantage promoting outer surface lipoprotein. Pathogens 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Mondino, S.; San Martin, F.; Buschiazzo, A. 3D cryo-EM imaging of bacterial flagella: Novel structural and mechanistic insights into cell motility. J Biol Chem 2022, 298, 102105. [Google Scholar] [CrossRef]
- Izac, J.R.; Oliver, L.D., Jr.; Earnhart, C.G.; Marconi, R.T. Identification of a defined linear epitope in the OspA protein of the Lyme disease spirochetes that elicits bactericidal antibody responses: Implications for vaccine development. Vaccine 2017, 35, 3178–3185. [Google Scholar] [CrossRef]
- Steere, A.; Sikand, V.K.; Meurice, F.; Parenti, D.L.; Fikrig, E.; Schoen, R.T.; Nowakowski, J.; Schmid, C.H.; Laukamp, S.; Buscarino, C.; et al. , Vaccination against Lyme disease with recombinant Borrelia burgdorferi outer-surface lipoprotein A with adjuvant. Lyme disease vaccine study group. N Engl J Med 1998, 339, 209–215. [Google Scholar] [CrossRef]
- Sigal, L.H.; Zahradnik, J.M.; Lavin, P.; Patella, S.J.; Bryant, G.; Haselby, R.; Hilton, E.; Kunkel, M.; Adler-Klein, D.; Doherty, T.; et al. , A vaccine consisting of recombinant Borrelia burgdorferi outer-surface protein A to prevent Lyme disease. Recombinant outer-surface protein A Lyme Disease Vaccine Study Consortium. N Engl J Med. Erratum in: N Engl J Med 1998, 339, 571. [CrossRef]
- Gingerich, M.C.; Nair, N.; Azevedo, J.F.; Samanta, K.; Kundu, S.; He, B.; Gomes-Solecki, M. Intranasal vaccine for Lyme disease provides protection against tick transmitted Borrelia burgdorferi beyond one year. NPJ Vaccines 2024, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Oeemig, J.S.; Kolodziejczyk, R.; Meri, T.; Kajander, T.; Lehtinen, M.J.; Iwaï, H.; Jokiranta, T.S.; Goldman, A. Structural basis for complement evasion by Lyme disease pathogen Borrelia burgdorferi. J Biol Chem 2013, 288, 18685–18695. [Google Scholar] [CrossRef] [PubMed]
- Makabe, K.; Tereshko, V.; Gawlak, G.; Yan, S.; Koide, S. Atomic-resolution crystal structure of Borrelia burgdorferi outer surface protein A via surface engineering. Protein Sci 2006, 15, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberger, M.A.B.; Sobe, R.C.; Sauder, A.B.; Chatterjee, S.; Peña, A.; Wang, F.; Giron, J.A.; Kiessling, V.; Costa, T.R.D.; Conticello, V.P.; et al. Flagellin outer domain dimerization modulates motility in pathogenic and soil bacteria from viscous environments. Nat Commun 2022, 13, 1422. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: current applications and future directions. Signal Transduct Target Ther 2022, 7, 48. [Google Scholar] [CrossRef]
- De-Simone, S.G.; Napoleão-Pêgo, P.; Lechuga, G.C.; Carvalho, J.P.R.S.; Gomes, L.R.; Cardozo, S.V.; Morel, C.M.; Provance, D.W.; Silva, F.R.d. High-throughput IgG epitope mapping of tetanus neurotoxin: Implications for immunotherapy and vaccine design. Toxins 2023, 15, 239. [Google Scholar] [CrossRef]
- Waddell, L.A.; Greig, J.; Mascarenhas, M.; Harding, S.; Lindsay, R.; Ogden, N. The accuracy of diagnostic tests for Lyme disease in humans, A systematic review and meta-analysis of North American research. PLoS One 2016, 11, e0168613. [Google Scholar] [CrossRef]
- Marques, A.R. Laboratory diagnosis of Lyme disease: advances and challenges. Infect Dis Clin North Am 2015, 29, 295–307. [Google Scholar] [CrossRef]
- Lagatie, O.; Verheyen, A.; Nijs, E.; Batsa Debrah, L.; Debrah, Y.A.; Stuyver, L.J. Performance evaluation of 3 serodiagnostic peptide epitopes and the derived multiepitope peptide OvNMP-48 for detection of Onchocerca volvulus infection. Parasitol Res 2019, 118, 2263–2270. [Google Scholar] [CrossRef]
- Lewandrowski, E.L. , Turbett S.E., Nigrovic L.E., Klontz E.H., Branda J.A. Comparative evaluation of commercial test kits cleared for use in modified two-tiered testing algorithms for serodiagnosis of Lyme disease. J Infect Dis 2024, 14, 230. [Google Scholar] [CrossRef]
- O’Bier, N.S.; Camire, A.C.; Patel, D.T.; Billingsley, J.S.; Hodges, K.R.; Marconi, R.T. Development of novel multi-protein chimeric immunogens. that protect against infection with the Lyme disease agent, Borreliella burgdorferi. mBio 2024, 7, e0215924. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Anderson, J.F.; Johnson, R.C. Cross-reactivity in serological tests for Lyme disease and other spirochetal infections. J Infect Dis 1987, 156, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska-Koszko, I.; Kwiatkowski, P.; Sienkiewicz, M.; Kowalczyk, M.; Kowalczyk, E.; Dołęgowska, B. Cross-reactive results in serological tests for Borreliosis in patients with active viral infections. Pathogens 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Doskaliuk, B.; Zimba, O. Borrelia burgdorferi and autoimmune mechanisms: implications for mimicry, misdiagnosis, and mismanagement in Lyme disease and autoimmune disorders. Rheumatol Int 2024, 44, 2265–2271. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



