
Submitted:
05 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
The elimination of hydrocarbon pollution is a critical environmental challenge, often addressed using biotechnological approaches. The study of alkane oxidizing bacteria is an important task, as it will allow developing new approaches to oil pollution treatment and improving existing ones. Representatives of the genus Rhodococcus are well-known hydrocarbon-oxidizing bacteria with high biotechnological potential. A distinctive feature of some Rhodococcus species is the presence of multiple homologous alkB genes within their genomes. However, the reasons behind this redundancy and the functional diversity of the alkane monooxygenases encoded by these genes remain unclear. Furthermore, despite the significant diversity of alkB genes even within this single genus, a comprehensive classification system has yet to be established. In this study, we conducted a phylogenetic analysis of all AlkB family alkane monooxygenases identified in Rhodococcus genomes and examined the genomic context of their corresponding genes. Our findings reveal a remarkable diversity of AlkB enzymes, refine the classification of previously described AlkB types, and propose three novel types.For the AlkB0 type, we identified deviations from previously reported signature amino acid motifs. Our results highlight the key role of colocalization of alkane monooxygenase alkB genes with rubredoxin rubB genes in the further functional classification of alkane monooxygenases.
Keywords:
alkan monooxygenase
; alkane hydroxylase
; rubredoxin
; Rhodococcus
; alkane degradation
; bioremediation
1. Introduction
Microorganisms capable of utilizing hydrocarbon substrates as their sole source of carbon and energy are widespread in nature. They have been identified in various environments, including soil [1,2,3], coastal sediments [4,5], seawater [6,7], and rhizospheres [8,9,10]. These microorganisms are even found in extreme environments, such as the Arctic [11,12] and Antarctic [13,14] regions. Aliphatic alkanes are one of the most common hydrocarbons in the natural environment. Alkanes enter ecosystems through a variety of sources. They are produced as metabolites by cyanobacteria [15], form the primary components of waxy substances of plant origin [16,17], and arise during the decomposition of plant residues [18]. Natural seepage of crude oil [19,20] also contributes to alkane release.
Human activities significantly amplify the presence of alkanes in the environment. This occurs through the extraction, transportation, and refining of crude oil and its products. Alkanes constitute 20–40% of crude oil, making them the most abundant fraction [21]. Anthropogenic sources contribute to more than 50% of oil entering marine environments according to some estimates [22]. Pollution arises from various incidents, ranging from moderate spills [23] to extreme events, such as the Deepwater Horizon oil spill (10.1126/science.1195223), from regular, low-level routine leaks [24]) and as part of wastewater from oil refineries [25]. Both acute oil spills and chronic hydrocarbon pollution have severe impacts on ecosystems, harming biodiversity [26,27,28], human health, and public well-being [29,30,31]. Oil pollution thus remains a critical environmental issue, underscoring the need to study hydrocarbon-oxidizing bacteria and the mechanisms of alkane biodegradation. Such research facilitates the development and application of effective, environmentally friendly remediation techniques [32,33,34].
Under aerobic conditions, the predominant pathway for the microbial consumption of n-alkanes involves their sequential oxidation. This process converts n-alkanes into the corresponding alcohols, then into intermediate aldehydes, and finally into fatty acids, which subsequently undergo beta-oxidation [35]. From a chemical perspective, alkanes are relatively inert substrates, so the key step in their metabolism is the activation of the substrate molecule through terminal or subterminal hydroxylation to form primary or secondary alcohols, respectively [36].
Therefore, when studying alkane metabolism in bacteria, particular attention is given to the enzymes responsible for substrate hydroxylation: alkane monooxygenases (also known as alkane hydroxylases) [37].
Currently, several families of bacterial alkane monooxygenases are known. Short-chain alkanes (up to C9) are oxidized by copper-containing membrane-bound monooxygenases (CuMMO family), including butane monooxygenase (pBMO), soluble di-iron monooxygenases (SDIMO family) such as methane monooxygenase (sMMO), butane monooxygenase (sBMO), and propane monooxygenase family (PRM) [38,39,40,41]. Soluble monooxygenases CYP153 from the gem-thiolate cytochrome P450 superfamily are probably responsible for the oxidation of medium-length alkanes (C4-C16) [42]. These enzymes exhibit significant potential for biotechnological applications in the bioprocessing industry, including the transformation of various molecules and the production of valuable compounds [43,44].
Luciferase-like flavoprotein monooxygenase (LadA), belonging to the SsuD subfamily, has been identified in thermophilic Geobacillus species and is reported to degrade alkanes with carbon chains up to C36 (10.1016/j.jmb.2007.11.069). Similarly, the almaA gene in Acinetobacter sp. strain DSM 17874 encodes the flavin monooxygenase AlmA, which is presumed to degrade long-chain alkanes from C32 [45].
Among bacterial alkane monooxygenases, the most extensively studied are the non-heme diiron alkane monooxygenases of the AlkB family, which belong to the superfamily of membrane fatty acid desaturases [46]. The organization of the corresponding genes and the pathway for n-alkane metabolism under aerobic conditions have been described in detail, notably in studies of the OCT-plasmid in Pseudomonas putida strain GPo1 (P. oleovorans strain TF4-1L = ATCC 29347) [47]. In addition to alkane monooxygenase itself, two other proteins are essential for alkane hydroxylation: rubredoxin (RubA), an electron carrier, and rubredoxin reductase (RubB) responsible for reducing rubredoxin. During substrate oxidation, a two-electron transfer occurs, consuming one molecule of the reducing equivalent NADH and reducing molecular oxygen to water [37]. Usually, alkane monooxygenase is represented as a singe domaine proteine. However, fused forms, such as AlkB-RubA [48] and even three-domain proteins like AlkB-RubA-RubB, have also been identified [49]. Interestingly, the substrate specificity of AlkB is not confined to alkanes. Enzymes in this family can catalyze the hydroxylation of alkyl radicals of esters [50] and the epoxidation of 1,7-octadiene [51]. Moreover, competition between hydroxylation and desaturation reactions has been demonstrated using norcarane, cyclohexane, and methylphenylcyclopropane as examples [52].
The alkB genes are widely distributed in the environment [5,53,54]. The greatest diversity of alkB genes encoding alkane monooxygenases of the AlkB family has been observed in representatives of the phyla Pseudomonadota and Actinomycetota [55,56], which include typical oil-degrading genera such as Pseudomonas and Rhodococcus. Analysis of the alkB gene has been used to assess the community composition of hydrocarbon-oxidizing microorganisms [57,58], as a catabolic marker for alkane degraders [57,59], and even as a complementary tool for monitoring crude oil presence [60,61]. The abundance and detectable diversity of alkB genes positively correlate with the natural content of alkanes in ecotopes [17]. An increase in the copy number of alkB genes has been observed during acute hydrocarbon exposure in both laboratory experiments [54,61] and natural conditions, such as emergency oil spills, as exemplified by the Gulf of Mexico case [62]. Enrichment of the hydrocarbon degradation gene fraction, including alkB, is also evident following chronic hydrocarbon pollution [63]. However, many studies show results that are inconsistent with the above, demonstrating the absence of a clear and explainable relationship between the alkB gene profile and alkane content [5,64,65].
One of the reasons for such apparent discrepancies may be the incomplete understanding of the true diversity of alkB genes, and the results of recent studies indicate that the true picture of that diversity has yet to be revealed. Comprehensive analyses of genomic and metagenomic data have shown that, an entirely new clade of alkB genes absent in uncontaminated samples emerges in metagenomes in the presence of hydrocarbon contamination [66]. of Uncultured microorganisms reseaches have significantly expanded knowledge of the prevalence of degradation genes [67,68].
Nevertheless, much of this research focuses on specific communities, whether natural or laboratory-based, with only a few studies assessing the overall diversity of degradation genes deposited in public databases [55,69]. Systematic bioinformatics research on alkB genes, considering their genomic context, has enabled the characterization of a new, diverse clade of alkB within the phylum Pseudomonadota [70]. However, this approach has limitations too: when analyzing sequences at high taxonomic ranks, the resulting resolution is too low to adequately capture diversity at lower taxonomic levels. A more comprehensive understanding of the true diversity of alkB genes is essential for elucidating evolutionary relationships, characterizing the corresponding proteins, studying alkane-degrading communities, and advancing bioremediation applications.
Representatives of the genus Rhodococcus (phylum Actinomycetota) are particularly noteworthy in this regard. Rhodococcus is considered an environmentally significant genus with immense biotechnological potential due to its ability to degrade a wide range of pollutants [71,72]. These bacteria are typical hydrocarbon-oxidizing members of indigenous microbiomes and are actively used in bioremediation technologies for oil-contaminated environments [73].
A distinctive feature of rhodococci is the presence of multiple chromosomal alkB homologs in the genomes of certain species [74]. The reason for this redundancy remains unclear. Moreover, we observed that some studies do not account for this intrageneric diversity in their phylogenetic analyses [75,76].
A significant portion of studies on rhodococcal alkB genes relies on amplicon sequencing. The obvious disadvantages of this approach are the low sensitivity of degenerate primers even in the case of their specialized design, which will significantly underestimate the detectable gene diversity. Additionally, the obtained sequences had been partial, restricting the resolution of phylogenetic analyses. It can also be noted that the the detectable phylum-level diversity of alkB genes is typically much lower for Actinomycetota compared to Pseudomonadota [56,68,70,77], bioinformatic assessment is not consistent with this (10.1038/srep04968). It is challenging to determine whether this discrepancy reflects the actual state of diversity or is the result of methodological bias.
In our study, we used the approach of investigating AlkB diversity within the genus of Rhodococcus. Based on the performed phylogenetic analysis of amino acid sequences and examination of the genomic context of the corresponding genes, we propose a more consistent classification for AlkB. Our results allow us to generalize the known AlkB types and identify three novel ones. To the best of our knowledge, this is the first comprehensive analysis of AlkB family proteins that utilized the entire available array of amino acid sequences at the genus level. These findings enhance our understanding of alkane monooxygenase diversity and provide valuable insights to the further understanding of the mechanisms of bacterial alkane degradation systems.
2. Results
2.1. Defining a Rhodococcus (sensu lato) group
First of all, we defined the taxonomic boundaries of the group for which the AlkB search will be performed. There are discrepancies in the scientific community regarding the phylogenetic position of some Rhodococcus species, associated with their allocation to other genera: Rhodococcoides, Prescottella, Antrihabitans [92,93]. Taxonomy within Rhodococcus (sensu stricto) also faces difficulties, for example, due to the very close phylogenetic relationships of the species R. qingshengii and R. erythropolis, which complicates their resolution to species [94]. For these reasons, the same objects may have different taxonomic assignments in different databases (Table 1).
All four of the above mentioned genera are listed in GenBank as separate genera. The Rhodococcus fascians clade was separated from the genus Rhodococcus in a recent update. Since NCBI Taxonomy database is not an authoritative source for nomenclature or classification [84], we examined the taxonomy of Rhodococcus with the Genome Taxonomy database (GTDB) [85]. In the GTDB classification the genera Prescottella and Rhodococcoides are absent, and all their representatives are assigned to different Rhodococcus clusters. The genus Antrihabitans is present, but the only one available genome of Antrihabitans cavernicola is assigned to Rhodococcus cavernicola. The taxonomy of Rhodococcus in the Type (Strain) Genome Server database (10.1038/s41467-019-10210-3) is largely consistent with the GTDB. According to the List of Prokaryotic names with Standing in Nomenclature (LPSN) for the taxa Rhodococcoides, Prescottella soli, Prescottella subtropica and Antrihabitans cavernicola the correct genus name is Rhodococcus [95].
As a result, it was decided to cover the largest number of sequences for further work. We identified a provisional group Rhodococcus (sensu lato), which included all species classified as Rhodococcus in at least one database, namely Rhodococcus (sensu stricto) spp., Prescottella spp., Rhodococcoides spp., and Antrihabitans cavernicola. Hereafter, the names of taxa are given according to the NCBI Taxonomy.
2.2. Phylogenetic analysis of AlkB sequences and examination of the genomic context
A BLAST search of AlkB family alkane monooxygenases was performed among representatives of the Rhodococcus (sensu lato) group (Table S1). After filtering nd removing outliers (Table S2), 927 non-redundant translated amino acid sequences of length 349-447 were obtained, the minimum % identity was 44.99%.
Phylogenetic analysis of the translated amino acid sequences demonstrates the formation of a large number of well-isolated clades with high representation (Figure 1). To identify patterns in AlkB clustering and to classify them, we studied the genomic context of the corresponding genes. All genomes of the Rhodococcus (sensu lato) group with an assembly level “complete” or “chromosome” as well as available genomes of type strains of all species were analyzed (excluding “Atypical genomes”, “Metagenome-assembled genomes”, and “Genomes from large multi-isolate projects”). Plasmids were not considered if present. A total of 221 assemblies were considered. For all strains with unspecified species designations we identified them using the Type (Strain) Genome Server and checked them in GTDB (Table S3). Based on these resutls, the following types can be distinguished.
The AlkB1 type is characterized by the localization of alkB gene in the so-called complete alkB-cluster. It is an operon that includes the alkB gene, two rubredoxin rubA genes, and the rubredoxin reductase rubB gene (Figure 2). In most of the cases considered, the co-directional gene of the TetR/AraC family regulatory protein alkU is located downstream. It can also be part of an operon, as demonstrated for the Rhodococcus jostii RHA1 strain (10.3390/microorganisms7110479). However, in the R. aetherivorans, R. indonesiensis, and R. ruber it is directed convergently, and in Mycobacterium tuberculosis H37Rv co-transcription of this gene together with the others was not observed (10.1128/spectrum.01969-22). A total of 110 alkB1-containing regions were found. Outside the complete alkB cluster, the genomic context is variable.
The genes we assigned to the AlkB2 subtype (a total of 115 regions were examined) are localized in a partial alkB-cluster that includes only two rubA and alkU genes, in the absence of rubB (Figure 2). A distinctive feature of the partial cluster is its localization in a fairly conservative manA-ahcY region. The downstream region is highly conservative and is preserved in all the genomes examined: after alkU, the genes adenosylhomocysteinase ahcY, thymidylate kinase, two-component sensor histidine kinase mtrA and mtrB are localized. The upstream region contains the genes amino acid permease, cation diffusion transporter fieF mannose-6-phosphate isomerase manA, phosphoglucose / phosphomannose isomerase tobH, phosphomannomutase / phosphoglucomutase manB and DUF3499 domain-containing protein. The upstream part is more variable: there are variants without transporter, or without transporter and permease, but the other genes are always present. It can be noted that all the listed genes are co-directional, although there is no data on their operon organization.
In 5 cases, not a partial but a complete alkB cluster was localized in the manA-ahcY region. i.e. together with the rubB gene. We searched for such regions and found that all corresponding AlkB sequences formed a clade designated as type AlkB1N. The opposite picture was shown for 9 regions: the partial operon was localized not in the manA-ahcY region. The genomic context was variable and did not coincide with any of the found variants of the alkB1 context. All corresponding proteins, designated AlkB2N, clustered on the tree together with AlkB1 but did not form a clade.
Neither AlkB1 nor AlkB2 types formed monophyletic groups. AlkB1 type forms two clades, AlkB2 — three (if we exclude the unique clade of two singleton sequences of AlkB2 Antrihabitans cavernicola). At the same time, one of the AlkB2 clades is formed exclusively by sequences of the genus Prescottella, in which, however, AlkB2 is the only homologue. A distinctive feature uniting these two types is the colocalization of the alkB gene with the rubA genes. The genomic context of all other alkB types did not contain rubA. Also, no cases were found when the rubB gene was present in the region, but the rubA gene was absent. The only exception was the strain Rhodococcoides yunnanense strain G21638-S1: the region included alkU, convergently located, alkB and rubB, and the corresponding protein was clustered together with the AlkB1 type.
A significant exception from the AlkB1-AlkB2 sequence cluster, which includes all cases of alkB and rubA colocalization, are the sequences we assigned to the AlkB8 type (Figure 1 and Figure 2). For them, only 4 regions were initially considered, 3 of which belonged to Rhodococcoides kroppenstedtii and 1 to Rhodococcoides corynebacterioides. Conserved determinants of the genomic context, which allowed us to isolate this type, were the alpha/beta fold hydrolase menH and glycosyltransferase mshA upstream genes from the region and the downstream ROK family transcriptional regulator. Sequences of this type clustered on a fairly distant branch, which was part of a common cluster with the AlkB1 and AlkB2. This branch itself is divided into two more clades, and the proteins of the regions we analyzed fall into only one of them. Checking the alignment of all proteins in the branch showed that the key difference between the sequences from the two clusters is the absence of the first 30 amino acids from the N-terminus in the clade that included proteins from regions that we did not analyze. It is likely that these are errors in determining the CDS during annotation: longer AlkB8 type proteins were completely aligned to the translated sequences of the corresponding nucleotide regions, which does not confirm the hypothetical deletion. Analysis of the genomic context of all other sequences localized in this branch confirmed the high conservation of the environment. This allowed us to combine all these high similarity sequences into one AlkB8 type.
A large distant clade is formed by sequences of the AlkB0 type, the genes of which are localized in the manA-ahcY region (22 regions), but all other elements of the alkB cluster, rubA, rubB, and alkU, are absent so that the downstream from alkB is immediately localized ahcY gene (Figure 2). The upstream region in this case is more conservative than for the alkB2 tytpe: the manA and permease genes are always located upstream. Since such a frequent localization of the alkB gene in the manA-ahcY region was found, we checked those region in genomes where alkB1A, alkB2, and alkB0 were not found. In these cases, alkU is always localized between the manA and the ahcY genes. As in the case of the alkB2 type, the permease gene, or both the permease and the transporter, can be also localized between manA and alkU.
Among the AlkB2 and AlkB0 types, the unique organization of the genomic context in Antrihabitans cavernicola is particularly noteworthy. Only one genome has been deposited in GenBank for this species (INSDC and RefSeq accessions, with slight differences in protein sequences, which explains the presence of two leaves on the phylogenetic tree). The genome of this strain contains the manA-ahcY region, in which a partial alkB-cluster is localized, as for the typical AlkB2 type, but another AlkB gene is localized between the operon and the ahcY gene. The AlkB protein corresponding to the first gene from the operon is clustered together with AlkB2, and the protein corresponding to the second gene with AlkB0. Both sequences are quite distant from the other representatives of the types. Interestingly, we found a similar colocalization of a partial alkB-cluster with alkB0 in Nocardia asteroides genomes. In this case, the downstream localised ahcY gene, and in the upstream there was a local rearrangement of the region with manA (Figure 3).
AlkB3 type was colocalized with downstream class F sortase gene, AlkB4 type — with downstream glutamate-tRNA ligase gltX, AlkB6 — with upstream ABC transporter substrate-binding protein (Figure 2). All sequences of such proteins are localized in one common clade. In this clade AlkB found in the sole genome of Rhodococcus globerulus was clustered relatively close to the AlkB3, although the genomic context coincided only by one downstream short hypothetical protein. We additionally analyzed all deposited Rhodococcus globerulus genomes (7 accessions) and checked the genomic context of two more proteins (for Rhodococcus sp. WS3 and Rhodococcus sp. MS16 strains) belonging tothe same clade. All 9 tested regions had a conservative organization, confirming that AlkB9 type can be proposed.
The AlkB5 type is represented in 66 regions and forms a large, distinct clade. In the genomic context, the upstream genes are aminotransferase and cold shock protein, and the downstream is either the AbiEi family antitoxin domain-containing protein gene or the non-ribosomal peptide synthase gene (Figure 2).
The AlkB7 type gene is located between alpha/beta hydrolase and ATP-binding protein. It is found in representatives of the genus Rhodococcoides (7 regions), some Rhodococcus species (3 regions), and Rhodococcus of unknown species (4 regions) (Figure 2).
Eight very similar AlkB sequences clustered in a distant clade basal to the AlkB5 clade. We tested all proteins of this clade and all corresponding genomic regions. All strains belonged only to the species Rhodococcoides kroppenstedtii, and the genomic context was different. We designated this clade as AlkBK, but according to our approach we cannot distinguish these alkane monooxygenases as a separate type.
Another 5 unique sequences, designated as AlkBX type, clustered separately from all other types and had no common elements of genomic context.
2.3. Distribution of AlkB types by species
The distribution of AlkB across Rhodococcus (sensu lato) species was degenerate: different species could contain one set of AlkB types, but representatives of a single species always carried the same set (Figure 4).
The most represented type from the taxonomic point of view is AlkB2 (25 species), followed by AlkB1 (19 species). It should be noted that each species has at least one of these types, with the exception of R. artemisiae with the only homologue of AlkB2 belongs to type 5. On the other hand, the simultaneous presence of genes of types 1 and 2 is observed only within one clade: “erythropolis, qingshengii, baikonurensis”. The AlkB1 sequences of these species cluster in the most distant AlkB1 clade from all other AlkB1s. The AlkB2N type is found in fairly distant taxa: in the Rhodococcoides cluster and in the “Rhodococcus phenolicus, Rhodococcus zopfii” clade. AlkB1A is present in the genomes of 4 species that fall into one cluster together with Rhodococcus spelaei, containing AlkB2. In all members of the genus Prescottella, the only type present was AlkB2.
If the distribution of AlkB1 and AlkB2 by species has a polyphyletic character, then the remaining types were distributed within monophyletic groups. If we do not consider the species Antrihabitans cavernicola, which is controversial in taxonomic position and unique in the organization of the region, then AlkB0 is found only within the clade containing all representatives of the genus Rhodococcoides (6 species) and three representatives of Rhodococcus. All representatives of this clade had the AlkB7 type, except for Rhodococcoides kroppenstedtii and Rhodococcoides corynebacterioides. At the same time, only these two phylogenetically closest species had the AlkB8 type, and the conditional AlkBK type in R. kroppenstedtii. Both AlkB8 and AlkBK were localized in isolated and remote clusters on the AlkB sequence tree.
Alkane monooxygenases AlkB3, AlkB4 and AlkB6 were found in a single clade with only four species. AlkB4 was found in Rhodococcus qingshengii, R. erythropolis, R. baikonurensis and R. globerulus; AlkB6 - in R. qingshengii, R. erythropolis and R. baikonurensis; AlkB3 - only in R. qingshengii and R. baikonurensis and in one representative of R. erythropolis. AlkB5 type is present in 15 species that make up a monophyletic group, which includes R. rhodochrous, R. aetherivorans, R. pyridinivorans, R. ruber.
Most genomes contain 1-2 homologous alkB genes. 3 homologs are found in almost all representatives of the Rhodococcoides cluster (7 out of 9). Also, 3 copies are contained in the genome of Rhodococcus globerulus. And the closest group of species R. erythropolis, R. qingshengii, and R. baikonurensis is unique: only its representatives carry 4 (erythropolis) and 5 (qingshengii, baikonurensis) copies of AlkB. Four species did not contain homologs of AlkB genes at all: Rhodococcus antarcticus, R. rhodnii, R. spongiicola and R. xishaensis. They are distributed along different branches. The species R. spongiicola and R. xishaensis formed a basal clade in relation to the Prescottella clade. R. rhodnii was included in a clade with triatomae, and R. antarcticus was the most distant species among all the Rhodococcus (sensu lato) species considered.
2.4. AlkB amino acid motifs
The search for amino acid motifs using the MEME tool was performed for the entire obtained set of alkane monooxygenase sequences among representatives of the Rhodococcus (sensu lato) group, as well as for individual types. The main signature histidine-containing motifs were HE[LM]GH[KR], EHN[RHF]GHH, NY[LVI]EHY[AG], LQRHSDHHA. All of them had high statistical significance (E-value close to zero) (Figure S1). To our knowledge, this is the first de novo search for motifs performed for a large set of alkane monooxygenase sequences. Comparison of motifs found in individual types showed that the main patterns in them coincide, with the exception of the AlkB0 type (see Discussion).
3. Discussion
The existence of several homologous alkane monooxygenases in representatives of the genus Rhodococcus was first demonstrated in the paper by van Beilen et al. [74]. The same group of authors performed a detailed characterization: the genomic context was described, and the possible role of alkane monooxygenases and rubredoxins in the oxidation of alkanes was assessed using heterologous expression [96]. In two strains of Rhodococcus erythropolis, the presence of four homologous copies of alkB was shown, according to which we identified the types AlkB1, AlkB2, AlkB3, and AlkB4 (Table 2).
In the work by Kim et al. [97], amplicons of alkB genes were obtained for several Rhodococcus strains, which were assigned by the authors to new types. Since only partial sequences are provided and the genomes of the studied strains are absent, the genomic context for these alkB genes is unavailable. However, clustering on a common tree allows us to assign AlkB6 and AlkB7 types, according to Kim et al., to the AlkB2 and AlkB1 types we proposed, respectively. The AlkB5 type clusters into a new clade.
Another new AlkB type was proposed based on the analysis of partial sequences of the Rhodococcus sp. TMP2 strain [98]. The authors designated it as AlkB5 but did not take into account the results of the previous work, so we chronologically designate the clade in which this sequence clusters as the AlkB6 type. At the same time, the described strain also contained the AlkB1, AlkB2, AlkB3, and AlkB4 types, i.e., a total of five copies of the alkB genes, which is completely consistent with our results for the Rhodococcus qingshengii species.
Amouric et al. [99] used the AlkB types from Whyte et al. [96] and Kim et al. [97] to classify alkB amplicons obtained for the strains Rhodococcus sp. (ruber) SP2B and R. ruber DSM 43338. Two copies of alkB were found in each strain, one pair of which the authors attributed to AlkB1. For one of these genes, a genomic context was described that included two rubA genes and a rubB gene, which confirms our classification. Another pair of homologues was proposed to be classified either as a new type or as AlkB5. Our classification supports the second assumption. Thus, the AlkB composition in both strains completely corresponds to the composition for R. ruber.
An alkane monooxygenase found in Rhodococcus sp. CH91 [100] was proposed to be classified as a new phylum. Based on the sequence clustering on the tree and the corresponding genomic context, this protein can be assigned to the AlkB5 type. The strain genome contained a second homologue, which belongs to the AlkB2 type. This AlkB composition is typical for R. rhodochrous and a number of closely related species. Analysis of the Rhodococcus sp. CH91 genome in TSGS showed that it is probably a separate species, but the closest one is R. rhodochrous, which has a similar AlkB composition.
Analysis of the genomic context of rhodococcal alkane monooxygenases allowed us to identify the AlkB7 type, which contains the upstream gene alpha/beta hydrolase [101]. The same work showed the presence of single alkB genes in Rhodococcus fascians, localized between the permease and ahcY genes. We attributed these to the AlkB0 type.
Thus, among the types we proposed, AlkB1N, AlkB8, and AlkB9 were introduced for the first time (Figure 5).
As can be seen, in a number of the above-described works, alkane monooxygenases were classified as a new type solely based on clustering their sequences at a sufficient distance from those described earlier. The results of our phylogenetic analysis show that many types are characterized by significant internal heterogeneity of clusters. Using a sufficiently high threshold of amino acid sequence similarity as the sole criterion, it would be possible to propose additional types. However, such an approach to classifying AlkB prroteins would break the phylogenetic tree into many small clades, creating an enormous number of groups, which would only complicate the overall picture of diversity.
The alkB sequences have previously been proposed as species markers [105], although this approach does not always provide the desired taxonomic resolution [106]. Our results suggest that the phylogenetic analysis of alkane monooxygenases may have limited use as an additional marker for species identification.
We have shown for the first time the existence of species in the genus Rhodococcus that do not contain alkB homologues, where the absence of this gene may itself serve as an indirect indicator of species affiliation. Unfortunately, most of the taxa considered are represented by only a small number of genomes or even a single genome, which limits the formulation of general conclusions.
An interesting case is the possibility of distinguishing the species Rhodococcus qingshengii and Rhodococcus erythropolis, which is a topic of ongoing discussion [94]. The applicability of molecular genetic analysis of alkB genes for identifying representatives of these two species has already been demonstrated [107]. Our results confirm these conclusions. Moreover, species separation can be preliminarily achieved simply by determining the copy number of alkB genes.
For six strains, we noticed a discrepancy between the GenBank species and the species-specific AlkB composition. We identified these strains using the Type (Strain) Genome Server and checked them in GTDB (Table S4). Clarified species matched the composition of AlkB types in five cases, and the only representative of Rhodococcus erythropolis containing five copies of alkB as Rhodococcus qingshengii was an exception.
In all known alkane monooxygenases, four highly conserved histidine-containing motifs are identified: HEXXHK, EHXXGHH, NYXEHYG, and LQRHXDHHA [96]. The first, second, and fourth motifs belong to the fatty acid desaturase superfamily [102], while the third (HYG motif) is specific to alkane monooxygenases [96]. Based on these patterns, sequence annotations are performed, and primers are designed for amplicon sequencing and hybridization [103,104].
Our search for motifs confirms their presence in all alkane monooxygenases of the Rhodococcus (sensu lato) group with high confidence. However, when comparing motifs among individual types, a difference was observed in the AlkB0 type: the first and third motifs are HEXXHR and NYXEHY[AG], respectively. In the fourth motif, a small fraction of sequences also shows the replacement of the last alanine by threonine (Figure 6). Since these motifs are localized in the sites forming the enzyme’s active center, substitutions in them may influence the functional role of this type of alkane monooxygenases compared to others.
Colocalization of alkane monooxygenase genes with rubredoxin genes was first demonstrated in the OCT plasmid of Pseudomonas putida (oleovorans) GPo1 [108]. Subsequent studies confirmed the widespread occurrence of this gene arrangement [35,70]. Fusion proteins combining alkane monooxygenase and rubredoxin domains are also charac [109,110]. Given the biochemical role of rubredoxin in the hydroxylation of alkanes, this organization appears functionally advantageous.
The stability and prevalence of such gene arrangements are directly linked to the functional role of rubredoxin in alkane hydroxylation. Interestingly, the rubB gene, which encodes rubredoxin reductase and is also essential for alkane oxidation, is less commonly found in a single operon with alkB and rubA. This single operon arrangement seems characteristic of certain members of the order Mycobacteriales. However, the clustering of the AlkB1 and AlkB2 types into one clade suggests evolutionary proximity and functional similarity, regardless of the presence of the rubB gene.
This hypothesis is further supported by the relatively rare co-occurrence of AlkB1 and AlkB2 types within the same genome, indicating potential duplication of function. The near-universal presence of at least one of these types in almost all Rhodococcus sensu lato species indicates strong positive selection for the colocalization of alkB and rubA. Studies on the biochemistry of alkane monooxygenases and their functional roles predominantly focus on enzymes encoded by genes colocalized with rubA ([111,112,113]). Such cases also include the studies of fusion proteins [109,110].
Conversely, our results reveal that alkane monooxygenases whose genes are localized without a neighboring rubredoxin gene form a separate, large, and diverse cluster. Even the AlkB8 type, which clusters with AlkB1-AlkB2, exhibits significant evolutionary distance. Based on this observation, we propose using the colocalization of alkB and rubA genes as a primary parameter for classifying AlkB family alkane monooxygenases. We suggest two generalized types: AlkBR (R for “rubredoxin”) and AlkBS (S for “single”) (Figure 6).
Considering all factors, we speculate that these two types of alkane monooxygenases may have distinct functional roles. This hypothesis warrants further investigation and provides a rationale for focusing future research efforts on the AlkBS type of alkane monooxygenases and offers new insights into the evolutionary and functional diversity of these enzymes.
4. Materials and Methods
4.1. Phylogenetic analysis of sequences of alkane monooxygenases of the AlkB family
To obtain translated amino acid sequences of alkane monooxygenases of the AlkB family for representatives the Rhodococcus (sensu lato) group, a blastp algorithm (blast.ncbi.nlm.nih.gov/Blast.cgi) was used to search for the sequence of alkane 1-monooxygenase Rhodococcus rhodochrous EP4 (AYA27164.1) in the GenBank database “non-redundant protein sequences (nr)” with a restriction on the taxa Rhodococcus, Rhodococcoides, Prescottella and Antrihabitans cavernicola [78]. The obtained sequences were filtered by length (>300), sequences marked as “partial” and “uncultured” were removed, and outliers were removed.
Sequence alignment was performed using the ClustalO v1.2.4 algorithm (ebi.ac.uk/Tools/msa/clustalo) [79].
The phylogenetic tree of amino acid sequences was constructed using the maximum likelihood method in the IQ-TREE v1.6.12 program (iqtree.cibiv.univie.ac.at) [80] with the search for the best substitution model by ModelFinder [81] and Ultrafast bootstrap [82].
Visualization and annotation of phylogenetic trees were performed in the online service iTOL v7 (itol.embl.de/) [83].
4.2. Assessment of the taxonomy classification
The taxonomic status of microorganisms was checked in the NCBI Taxonomy database[84], The Genome Taxonomy Database (GTDB, release 220) (gtdb.ecogenomic.org) [85].
The taxonomic position of strains was clarified using the Type (Strain) Genome Server service (tygs.dsmz.de) [86].
The phylogenetic tree of Rhodococcus (sensu lato) group and related taxa was constructed based
Genebank type strains genomes using the GTDB-Tk v2.4.0 program without including GTDB reference genomes in multiple sequence alignment (“--skip_gtdb_refs” option for “align” command) [87].
4.3. Characterisation of the genomic context
Genomic context was investigated using NCBI’s graphical GenBank and the online service SyntTax (archaea.i2bc.paris-saclay.fr/SyntTax) [88].
Gene clusters were searched using cblaster (cagecat.bioinformatics.nl/tools/search) [89].
Genomic regions were visualised using clinker (cagecat.bioinformatics.nl/tools/clinker) [90].
4.4. Characterisation of amino acid motifs
Motifs discovery was performed in MEME Version 5.5.7 tool (meme-suite.org/meme/tools/meme) wih “One Occurrence Per Sequence (oops)” and motif width 6-15 parameters [91].
Sequence logos were obtained from the WebLogo v2.8.2 online service (weblogo.berkeley.edu/logo.cgi).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Motifs discovered by MEME tool in AlkB protens. Table S1: BLAST results description table. Table S2: BLAST results outliers excluded from the final analysis. Table S3: Taxonomy assignment of the strains with unspecified species designations. Table S4: Taxonomy checking of the strains with unspecified species designations.
Author Contributions
Conceptualization, I. Yu. P.-F.; methodology, A. A. V., I. Yu. P.-F.; investigation, K. V. P., A. A. I.; writing—original draft preparation, K. V. P., A. A. I.; writing—review and editing, A. A. V., O. I. S. and Yu. P.-F.; visualization, K. V. P., O. I. S.; supervision, Yu. P.-F.; project administration, K. V. P.; funding acquisition, K. V. P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, project no. 23-24-00192, https://rscf.ru/project/23-24-00192/.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data obtained in the work are included in the article and and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Muthukumar, B.; Parthipan, P.; AlSalhi, M.S.; Prabhu, N.S.; Rao, T.N.; Devanesan, S.; Maruthamuthu, M.K.; Rajasekar, A. Characterization of Bacterial Community in Oil-Contaminated Soil and Its Biodegradation Efficiency of High Molecular Weight (>C40) Hydrocarbon. Chemosphere 2022, 289, 133168. [Google Scholar] [CrossRef] [PubMed]
- Brzeszcz, J.; Steliga, T.; Ryszka, P.; Kaszycki, P.; Kapusta, P. Bacteria Degrading Both N-Alkanes and Aromatic Hydrocarbons Are Prevalent in Soils. Environ. Sci. Pollut. Res. 2023, 31, 5668–5683. [Google Scholar] [CrossRef]
- Ehiosun, K.I.; Godin, S.; Urios, L.; Lobinski, R.; Grimaud, R. Degradation of Long-Chain Alkanes through Biofilm Formation by Bacteria Isolated from Oil-Polluted Soil. Int. Biodeterior. Biodegradation 2022, 175, 105508. [Google Scholar] [CrossRef]
- Gosai, H.B.; Panseriya, H.Z.; Patel, P.G.; Patel, A.C.; Shankar, A.; Varjani, S.; Dave, B.P. Exploring Bacterial Communities through Metagenomics during Bioremediation of Polycyclic Aromatic Hydrocarbons from Contaminated Sediments. Sci. Total Environ. 2022, 842, 156794. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Smith, A.F.; Rattray, J.E.; England, W.E.; Hubert, C.R.J. Potential for Natural Attenuation of Crude Oil Hydrocarbons in Benthic Microbiomes near Coastal Communities in Kivalliq, Nunavut, Canada. Mar. Pollut. Bull. 2023, 196, 115557. [Google Scholar] [CrossRef]
- Goveas, L.C.; Nayak, S.; Selvaraj, R. Concise Review on Bacterial Degradation of Petroleum Hydrocarbons: Emphasis on Indian Marine Environment. Bioresour. Technol. Reports 2022, 19, 101136. [Google Scholar] [CrossRef]
- Mohanram, R.; Jagtap, C.; Kumar, P. Isolation, Screening, and Characterization of Surface-Active Agent-Producing, Oil-Degrading Marine Bacteria of Mumbai Harbor. Mar. Pollut. Bull. 2016, 105. [Google Scholar] [CrossRef]
- Cheng, X.; Jiang, L.; Zhao, X.; Wang, S.; Li, J.; Luo, C.; Zhang, G. Synergism of Endophytic Microbiota and Plants Promotes the Removal of Polycyclic Aromatic Hydrocarbons from the Alfalfa Rhizosphere. J. Hazard. Mater. 2024, 478, 135513. [Google Scholar] [CrossRef]
- Khan, A.L.; Numan, M.; Bilal, S.; Asaf, S.; Crafword, K.; Imran, M.; Al-Harrasi, A.; Al-Sabahi, J.N.; Rehman, N. ur; A-Rawahi, A.; et al. Mangrove’s Rhizospheric Engineering with Bacterial Inoculation Improve Degradation of Diesel Contamination. J. Hazard. Mater. 2022, 423, 127046. [Google Scholar] [CrossRef]
- Lü, H.; Tang, G.-X.; Huang, Y.-H.; Mo, C.-H.; Zhao, H.-M.; Xiang, L.; Li, Y.-W.; Li, H.; Cai, Q.-Y.; Li, Q.X. Response and Adaptation of Rhizosphere Microbiome to Organic Pollutants with Enriching Pollutant-Degraders and Genes for Bioremediation: A Critical Review. Sci. Total Environ. 2024, 912, 169425. [Google Scholar] [CrossRef]
- Góngora, E.; Chen, Y.-J.; Ellis, M.; Okshevsky, M.; Whyte, L. Hydrocarbon Bioremediation on Arctic Shorelines: Historic Perspective and Roadway to the Future. Environ. Pollut. 2022, 119247. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.K.; Li, C.; Jiang, C.; Chakraborty, A.; Grasby, S.E.; Hubert, C.R.J. Natural Attenuation of Spilled Crude Oil by Cold-Adapted Soil Bacterial Communities at a Decommissioned High Arctic Oil Well Site. Sci. Total Environ. 2020, 722, 137258. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, S.; Monien, P.; Pepino Minetti, R.; Jürgens, J.; Curtosi, A.; Villalba Primitz, J.; Frickenhaus, S.; Abele, D.; Mac Cormack, W.; Helmke, E. Bacterial Communities and Chemical Parameters in Soils and Coastal Sediments in Response to Diesel Spills at Carlini Station, Antarctica. Sci. Total Environ. 2017, 605–606, 26–37. [Google Scholar] [CrossRef] [PubMed]
- van Dorst, J.; Wilkins, D.; Crane, S.; Montgomery, K.; Zhang, E.; Spedding, T.; Hince, G.; Ferrari, B. Microbial Community Analysis of Biopiles in Antarctica Provides Evidence of Successful Hydrocarbon Biodegradation and Initial Soil Ecosystem Recovery. Environ. Pollut. 2021, 290, 117977. [Google Scholar] [CrossRef]
- Lea-Smith, D.J.; Biller, S.J.; Davey, M.P.; Cotton, C.A.R.; Perez Sepulveda, B.M.; Turchyn, A.V.; Scanlan, D.J.; Smith, A.G.; Chisholm, S.W.; Howe, C.J. Contribution of Cyanobacterial Alkane Production to the Ocean Hydrocarbon Cycle. Proc. Natl. Acad. Sci. 2015, 112, 13591–13596. [Google Scholar] [CrossRef]
- Schulz, S.; Giebler, J.; Chatzinotas, A.; Wick, L.Y.; Fetzer, I.; Welzl, G.; Harms, H.; Schloter, M. Plant Litter and Soil Type Drive Abundance, Activity and Community Structure of AlkB Harbouring Microbes in Different Soil Compartments. ISME J. 2012, 6, 1763–1774. [Google Scholar] [CrossRef]
- Hussain, B.; Chen, J.-S.; Hsu, B.-M.; Chao, W.-C.; Fan, C.-W. Niche-Specific Modulation of Long-Chain n-Alkanes Degrading Bacterial Community and Their Functionality in Forest Habitats across the Leaf Litter-Soil Compartments. Appl. Soil Ecol. 2024, 195, 105248. [Google Scholar] [CrossRef]
- Wiesenberg, G.L.B.; Lehndorff, E.; Schwark, L. Thermal Degradation of Rye and Maize Straw: Lipid Pattern Changes as a Function of Temperature. Org. Geochem. 2009, 40, 167–174. [Google Scholar] [CrossRef]
- Jatiault, R.; Henry, P.; Loncke, L.; Sadaoui, M.; Sakellariou, D. Natural Oil Seep Systems in the Aegean Sea. Mar. Pet. Geol. 2024, 163, 106754. [Google Scholar] [CrossRef]
- Kvenvolden, K.A.; Cooper, C.K. Natural Seepage of Crude Oil into the Marine Environment. Geo-Marine Lett. 2003, 23, 140–146. [Google Scholar] [CrossRef]
- Petrov, A.A. General Characteristics of Petroleum Hydrocarbons Molecular and Group-Type Methods of Analysis and Classification. In Petroleum Hydrocarbons; Springer Berlin Heidelberg: Berlin, Heidelberg, 1987; pp. 4–26. [Google Scholar]
- Burgherr, P. In-Depth Analysis of Accidental Oil Spills from Tankers in the Context of Global Spill Trends from All Sources. J. Hazard. Mater. 2007, 140, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Abou Samra, R.M.; Ali, R.R. Tracking the Behavior of an Accidental Oil Spill and Its Impacts on the Marine Environment in the Eastern Mediterranean. Mar. Pollut. Bull. 2024, 198, 115887. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Elkin, A.A.; Makarov, S.O.; Cunningham, C.J.; Peshkur, T.A.; Atlas, R.M.; Philp, J.C. Oil Spill Problems and Sustainable Response Strategies through New Technologies. Environ. Sci. Process. Impacts 2015, 17, 1201–1219. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yulisa, A.; Kim, S.; Hwang, S. Monitoring Microbial Community Structure and Variations in a Full-Scale Petroleum Refinery Wastewater Treatment Plant. Bioresour. Technol. 2020, 306, 123178. [Google Scholar] [CrossRef]
- Haider, F.U.; Ejaz, M.; Cheema, S.A.; Khan, M.I.; Zhao, B.; Liqun, C.; Salim, M.A.; Naveed, M.; Khan, N.; Núñez-Delgado, A.; et al. Phytotoxicity of Petroleum Hydrocarbons: Sources, Impacts and Remediation Strategies. Environ. Res. 2021, 197, 111031. [Google Scholar] [CrossRef]
- Barron, M.G.; Vivian, D.N.; Heintz, R.A.; Yim, U.H. Long-Term Ecological Impacts from Oil Spills: Comparison of Exxon Valdez, Hebei Spirit, and Deepwater Horizon. Environ. Sci. Technol. 2020, 54, 6456–6467. [Google Scholar] [CrossRef]
- Lassalle, G.; Scafutto, R.D.M.; Lourenço, R.A.; Mazzafera, P.; de Souza Filho, C.R. Remote Sensing Reveals Unprecedented Sublethal Impacts of a 40-Year-Old Oil Spill on Mangroves. Environ. Pollut. 2023, 331, 121859. [Google Scholar] [CrossRef]
- Stroope, S.; Slack, T.; Kroeger, R.A.; Keating, K.S.; Beedasy, J.; Sury, J.J.; Brooks, J.; Chandler, T. Deepwater Horizon Oil Spill Exposures and Long-Term Self-Rated Health Effects among Parents in Coastal Louisiana. Disaster Med. Public Health Prep. 2023, 17, e329. [Google Scholar] [CrossRef]
- Hemati, S.; Heidari, M.; Momenbeik, F.; Khodabakhshi, A.; Fadaei, A.; Farhadkhani, M.; Mohammadi-Moghadam, F. Co-Occurrence of Polycyclic Aromatic Hydrocarbons and Heavy Metals in Various Environmental Matrices of a Chronic Petroleum Polluted Region in Iran; Pollution Characterization, and Assessment of Ecological and Human Health Risks. J. Hazard. Mater. 2024, 478, 135504. [Google Scholar] [CrossRef]
- Ezeh, C.C.; Onyema, V.O.; Obi, C.J.; Moneke, A.N. A Systematic Review of the Impacts of Oil Spillage on Residents of Oil-Producing Communities in Nigeria. Environ. Sci. Pollut. Res. 2024, 31, 34761–34786. [Google Scholar] [CrossRef]
- Zhong, M.; Yang, C.; Su, L.; Sun, Z.; Xu, J.; Zhang, J.; Li, Q.; Hao, Y.; Ma, H.; Chen, H.; et al. Interactions between Plants and Bacterial Communities for Phytoremediation of Petroleum-Contaminated Soil. Environ. Sci. Pollut. Res. 2024, 31, 37564–37573. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, Q.; Cai, X.; Zhao, X.; Luo, C.; Zhang, G. Metabolic Characterization and Geochemical Drivers of Active Hydrocarbon-Degrading Microorganisms. J. Geophys. Res. Biogeosciences 2024, 129. [Google Scholar] [CrossRef]
- Chen, J.; Zhuang, J.; Dai, T.; Zhang, R.; Zeng, Y.; Jiang, B.; Guo, H.; Guo, X.; Yang, Y. Enhancing Soil Petrochemical Contaminant Remediation through Nutrient Addition and Exogenous Bacterial Introduction. J. Hazard. Mater. 2024, 476, 135078. [Google Scholar] [CrossRef]
- Rojo, F. Degradation of Alkanes by Bacteria. Environ. Microbiol. 2009, 11, 2477–2490. [Google Scholar] [CrossRef]
- Williams, S.C.; Austin, R.N. An Overview of the Electron-Transfer Proteins That Activate Alkane Monooxygenase (AlkB). Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Moreno, R.; Rojo, F. Enzymes for Aerobic Degradation of Alkanes in Bacteria. In Aerobic Utilization of Hydrocarbons, Oils, and Lipids. Handbook of Hydrocarbon and Lipid Microbiology; Rojo, F., Ed.; Springer International Publishing: Cham, 2017; pp. 1–25. [Google Scholar]
- Hamamura, N.; Storfa, R.T.; Semprini, L.; Arp, D.J. Diversity in Butane Monooxygenases among Butane-Grown Bacteria. Appl. Environ. Microbiol. 1999, 65, 4586–4593. [Google Scholar] [CrossRef]
- Chen, Y.; Ren, H.; Kong, X.; Wu, H.; Lu, Z. A Multicomponent Propane Monooxygenase Catalyzes the Initial Degradation of Methyl Tert -Butyl Ether in <i>Mycobacterium Vaccae</i? JOB5. Appl. Environ. Microbiol. 2023, 89. [Google Scholar] [CrossRef]
- Ferrari, E.; Di Benedetto, G.; Firrincieli, A.; Presentato, A.; Frascari, D.; Cappelletti, M. Unravelling the Role of the Group 6 Soluble Di-Iron Monooxygenase (SDIMO) SmoABCD in Alkane Metabolism and Chlorinated Alkane Degradation. Microb. Biotechnol. 2024, 17. [Google Scholar] [CrossRef]
- Zou, B.; Huang, Y.; Zhang, P.-P.; Ding, X.-M.; Op den Camp, H.J.M.; Quan, Z.-X. Horizontal Gene Transfer of Genes Encoding Copper-Containing Membrane-Bound Monooxygenase (CuMMO) and Soluble Di-Iron Monooxygenase (SDIMO) in Ethane- and Propane-Oxidizing Rhodococcus Bacteria. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef]
- Funhoff, E.G.; Bauer, U.; García-Rubio, I.; Witholt, B.; van Beilen, J.B. CYP153A6, a Soluble P450 Oxygenase Catalyzing Terminal-Alkane Hydroxylation. J. Bacteriol. 2006, 188, 5220–5227. [Google Scholar] [CrossRef]
- Cornelissen, S.; Julsing, M.K.; Volmer, J.; Riechert, O.; Schmid, A.; Bühler, B. Whole-Cell-Based CYP153A6-Catalyzed (S)-Limonene Hydroxylation Efficiency Depends on Host Background and Profits from Monoterpene Uptake via AlkL. Biotechnol. Bioeng. 2013, 110, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.; Park, B.G.; Yoo, H.-W.; Kim, J.; Choi, K.-Y.; Kim, B.-G. Semi-Rational Engineering of CYP153A35 to Enhance ω-Hydroxylation Activity toward Palmitic Acid. Appl. Microbiol. Biotechnol. 2018, 102, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Throne-Holst, M.; Wentzel, A.; Ellingsen, T.E.; Kotlar, H.-K.; Zotchev, S.B. Identification of Novel Genes Involved in Long-Chain n-Alkane Degradation by Acinetobacter Sp. Strain DSM 17874. Appl. Environ. Microbiol. 2007, 73, 3327–3332. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, J.; Han, L.; Lee, J.; Williams, S.C.; Forsberg, A.; Xu, Y.; Austin, R.N.; Feng, L. Structure and Mechanism of the Alkane-Oxidizing Enzyme AlkB. Nat. Commun. 2023, 14, 2180. [Google Scholar] [CrossRef]
- van Beilen, J.B.; Panke, S.; Lucchini, S.; Franchini, A.G.; Röthlisberger, M.; Witholt, B. Analysis of Pseudomonas Putida Alkane-Degradation Gene Clusters and Flanking Insertion Sequences: Evolution and Regulation of the Alk Genes. Microbiology 2001, 147, 1621–1630. [Google Scholar] [CrossRef]
- Williams, S.C.; Forsberg, A.P.; Lee, J.; Vizcarra, C.L.; Lopatkin, A.J.; Austin, R.N. Investigation of the Prevalence and Catalytic Activity of Rubredoxin-Fused Alkane Monooxygenases (AlkBs). J. Inorg. Biochem. 2021, 219, 111409. [Google Scholar] [CrossRef]
- Williams, S.C.; Luongo, D.; Orman, M.; Vizcarra, C.L.; Austin, R.N. An Alkane Monooxygenase (AlkB) Family in Which All Electron Transfer Partners Are Covalently Bound to the Oxygen-Activating Hydroxylase. J. Inorg. Biochem. 2022, 228, 111707. [Google Scholar] [CrossRef]
- Yoo, H.-W.; Kim, J.; Patil, M.D.; Park, B.G.; Joo, S.; Yun, H.; Kim, B.-G. Production of 12-Hydroxy Dodecanoic Acid Methyl Ester Using a Signal Peptide Sequence-Optimized Transporter AlkL and a Novel Monooxygenase. Bioresour. Technol. 2019, 291, 121812. [Google Scholar] [CrossRef]
- Shanklin, J.; Achim, C.; Schmidt, H.; Fox, B.G.; Munck, E. Mössbauer Studies of Alkane-Hydroxylase: Evidence for a Diiron Cluster in an Integral-Membrane Enzyme. Proc. Natl. Acad. Sci. 1997, 94, 2981–2986. [Google Scholar] [CrossRef]
- Naing, S.-H.; Parvez, S.; Pender-Cudlip, M.; Groves, J.T.; Austin, R.N. Substrate Specificity and Reaction Mechanism of Purified Alkane Hydroxylase from the Hydrocarbonoclastic Bacterium Alcanivorax Borkumensis (<i>Ab<i/>AlkB). J. Inorg. Biochem. 2013, 121, 46–52. [Google Scholar] [CrossRef]
- Heiss-Blanquet, S.; Benoit, Y.; Marechaux, C.; Monot, F. Assessing the Role of Alkane Hydroxylase Genotypes in Environmental Samples by Competitive PCR. J. Appl. Microbiol. 2005, 99, 1392–1403. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Wang, Y.; Chang, S.; Liu, G.; Chen, T.; Huo, G.; Zhang, W.; Wu, X.; Tai, X.; Sun, L.; et al. Diversity of Crude Oil-Degrading Bacteria and Alkane Hydroxylase (AlkB) Genes from the Qinghai-Tibet Plateau. Environ. Monit. Assess. 2017, 189, 116. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Chi, C.-Q.; Fang, H.; Liang, J.-L.; Lu, S.-L.; Lai, G.-L.; Tang, Y.-Q.; Wu, X.-L. Diverse Alkane Hydroxylase Genes in Microorganisms and Environments. Sci. Rep. 2014, 4, 4968. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Peng, Y.; Liao, J.; Liu, X.; Peng, J.; Wang, J.-H.; Shao, Z. Broad-Spectrum Hydrocarbon-Degrading Microbes in the Global Ocean Metagenomes. Sci. Total Environ. 2024, 926, 171746. [Google Scholar] [CrossRef]
- Révész, F.; Figueroa-Gonzalez, P.A.; Probst, A.J.; Kriszt, B.; Banerjee, S.; Szoboszlay, S.; Maróti, G.; Táncsics, A. Microaerobic Conditions Caused the Overwhelming Dominance of Acinetobacter Spp. and the Marginalization of Rhodococcus Spp. in Diesel Fuel/Crude Oil Mixture-Amended Enrichment Cultures. Arch. Microbiol. 2020, 202, 329–342. [Google Scholar] [CrossRef]
- Guibert, L.M.; Loviso, C.L.; Marcos, M.S.; Commendatore, M.G.; Dionisi, H.M.; Lozada, M. Alkane Biodegradation Genes from Chronically Polluted Subantarctic Coastal Sediments and Their Shifts in Response to Oil Exposure. Microb. Ecol. 2012, 64, 605–616. [Google Scholar] [CrossRef]
- Mukherjee, S.; Sipilä, T.; Pulkkinen, P.; Yrjälä, K. Secondary Successional Trajectories of Structural and Catabolic Bacterial Communities in Oil-Polluted Soil Planted with Hybrid Poplar. Mol. Ecol. 2015, 24, 628–642. [Google Scholar] [CrossRef]
- Wasmund, K.; Burns, K.A.; Kurtböke, D.I.; Bourne, D.G. Novel Alkane Hydroxylase Gene (AlkB) Diversity in Sediments Associated with Hydrocarbon Seeps in the Timor Sea, Australia. Appl. Environ. Microbiol. 2009, 75, 7391–7398. [Google Scholar] [CrossRef]
- Bagi, A.; Knapik, K.; Baussant, T. Abundance and Diversity of n-Alkane and PAH-Degrading Bacteria and Their Functional Genes – Potential for Use in Detection of Marine Oil Pollution. Sci. Total Environ. 2022, 810, 152238. [Google Scholar] [CrossRef]
- Hu, P.; Dubinsky, E.A.; Probst, A.J.; Wang, J.; Sieber, C.M.K.; Tom, L.M.; Gardinali, P.R.; Banfield, J.F.; Atlas, R.M.; Andersen, G.L. Simulation of Deepwater Horizon Oil Plume Reveals Substrate Specialization within a Complex Community of Hydrocarbon Degraders. Proc. Natl. Acad. Sci. 2017, 114, 7432–7437. [Google Scholar] [CrossRef]
- Lu, Z.; Deng, Y.; Van Nostrand, J.D.; He, Z.; Voordeckers, J.; Zhou, A.; Lee, Y.-J.; Mason, O.U.; Dubinsky, E.A.; Chavarria, K.L.; et al. Microbial Gene Functions Enriched in the Deepwater Horizon Deep-Sea Oil Plume. ISME J. 2012, 6, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Christian, W.C.; Butler, T.M.; Ghannam, R.B.; Webb, P.N.; Techtmann, S.M. Phylogeny and Diversity of Alkane-Degrading Enzyme Gene Variants in the Laurentian Great Lakes and Western Atlantic. FEMS Microbiol. Lett. 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.B.; Tolar, B.B.; Hollibaugh, J.T.; King, G.M. Alkane Hydroxylase Gene (AlkB) Phylotype Composition and Diversity in Northern Gulf of Mexico Bacterioplankton. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Hatt, J.K.; Kim, M.; Spain, J.C.; Huettel, M.; Kostka, J.E.; Konstantinidis, K.T. A Novel, Divergent Alkane Monooxygenase (AlkB Clade) Involved in Crude Oil Biodegradation. Environ. Microbiol. Rep. 2021, 13, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.L.; Zaugg, J.; Mason, O.U. Novel, Active, and Uncultured Hydrocarbon-Degrading Microbes in the Ocean. Appl. Environ. Microbiol. 2024. [Google Scholar] [CrossRef]
- Vázquez Rosas Landa, M.; De Anda, V.; Rohwer, R.R.; Angelova, A.; Waldram, G.; Gutierrez, T.; Baker, B.J. Exploring Novel Alkane-Degradation Pathways in Uncultured Bacteria from the North Atlantic Ocean. mSystems 2023, 8. [Google Scholar] [CrossRef]
- Rojas-Vargas, J.; Castelán-Sánchez, H.G.; Pardo-López, L. HADEG: A Curated Hydrocarbon Aerobic Degradation Enzymes and Genes Database. Comput. Biol. Chem. 2023, 107, 107966. [Google Scholar] [CrossRef]
- Wang, S.; Li, G.; Liao, Z.; Liu, T.; Ma, T. A Novel Alkane Monooxygenase (AlkB) Clade Revealed by Massive Genomic Survey and Its Dissemination Association with IS Elements. PeerJ 2022, 10, e14147. [Google Scholar] [CrossRef]
- Zampolli, J.; Zeaiter, Z.; Di Canito, A.; Di Gennaro, P. Genome Analysis and -Omics Approaches Provide New Insights into the Biodegradation Potential of Rhodococcus. Appl. Microbiol. Biotechnol. 2019, 103, 1069–1080. [Google Scholar] [CrossRef]
- Nazari, M.T.; Simon, V.; Machado, B.S.; Crestani, L.; Marchezi, G.; Concolato, G.; Ferrari, V.; Colla, L.M.; Piccin, J.S. Rhodococcus: A Promising Genus of Actinomycetes for the Bioremediation of Organic and Inorganic Contaminants. J. Environ. Manage. 2022, 323, 116220. [Google Scholar] [CrossRef]
- Vetrova, A.A.; Trofimov, S.Y.; Kinzhaev, R.R.; Avetov, N.A.; Arzamazova, A.V.; Puntus, I.F.; Sazonova, O.I.; Sokolov, S.L.; Streletskii, R.A.; Petrikov, K.V.; et al. Development of Microbial Consortium for Bioremediation of Oil-Contaminated Soils in the Middle Ob Region. Eurasian Soil Sci. 2022, 55, 651–662. [Google Scholar] [CrossRef]
- van Beilen, J.B.; Smits, T.H.M.; Whyte, L.G.; Schorcht, S.; Röthlisberger, M.; Plaggemeier, T.; Engesser, K.-H.; Witholt, B. Alkane Hydroxylase Homologues in Gram-Positive Strains. Environ. Microbiol. 2002, 4, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shan, G.; Shen, J.; Zhang, F.; Liu, Y.; Cui, C. In Situ Bioremediation of Petroleum Hydrocarbon–Contaminated Soil: Isolation and Application of a Rhodococcus Strain. Int. Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Sengupta, K. Computational-Based Insights into the Phylogeny, Structure, and Function of Rhodococcus Alkane-1-Monooxygenase. 3 Biotech 2020, 10, 391. [Google Scholar] [CrossRef]
- Wang, W.; Wang, L.; Shao, Z. Diversity and Abundance of Oil-Degrading Bacteria and Alkane Hydroxylase (AlkB) Genes in the Subtropical Seawater of Xiamen Island. Microb. Ecol. 2010, 60, 429–439. [Google Scholar] [CrossRef]
- Altschul, S. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Madeira, F.; Madhusoodanan, N.; Lee, J.; Eusebi, A.; Niewielska, A.; Tivey, A.R.N.; Lopez, R.; Butcher, S. The EMBL-EBI Job Dispatcher Sequence Analysis Tools Framework in 2024. Nucleic Acids Res. 2024, 52, W521–W525. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v6: Recent Updates to the Phylogenetic Tree Display and Annotation Tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A Comprehensive Update on Curation, Resources and Tools. Database 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Chuvochina, M.; Rinke, C.; Mussig, A.J.; Chaumeil, P.-A.; Hugenholtz, P. GTDB: An Ongoing Census of Bacterial and Archaeal Diversity through a Phylogenetically Consistent, Rank Normalized and Complete Genome-Based Taxonomy. Nucleic Acids Res. 2022, 50, D785–D794. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk v2: Memory Friendly Classification with the Genome Taxonomy Database. Bioinformatics 2022, 38, 5315–5316. [Google Scholar] [CrossRef]
- Oberto, J. SyntTax: A Web Server Linking Synteny to Prokaryotic Taxonomy. BMC Bioinformatics 2013, 14, 4. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Booth, T.J.; van Wersch, B.; van Grieken, L.; Medema, M.H.; Chooi, Y.-H. Cblaster: A Remote Search Tool for Rapid Identification and Visualization of Homologous Gene Clusters. Bioinforma. Adv. 2021, 1. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap.Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Val-Calvo, J.; Vázquez-Boland, J.A. Mycobacteriales Taxonomy Using Network Analysis-Aided, Context-Uniform Phylogenomic Approach for Non-Subjective Genus Demarcation. MBio 2023, 14. [Google Scholar] [CrossRef]
- Sangal, V.; Goodfellow, M.; Jones, A.L.; Schwalbe, E.C.; Blom, J.; Hoskisson, P.A.; Sutcliffe, I.C. Next-Generation Systematics: An Innovative Approach to Resolve the Structure of Complex Prokaryotic Taxa. Sci. Rep. 2016, 6, 38392. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Cognat, V.; Goodfellow, M.; Koechler, S.; Heintz, D.; Carapito, C.; Van Dorsselaer, A.; Mahmoud, H.; Sangal, V.; Ismail, W. Phylogenomic Classification and Biosynthetic Potential of the Fossil Fuel-Biodesulfurizing Rhodococcus Strain IGTS8. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Whyte, L.G.; Smits, T.H.M.; Labbé, D.; Witholt, B.; Greer, C.W.; van Beilen, J.B. Gene Cloning and Characterization of Multiple Alkane Hydroxylase Systems in Rhodococcus Strains Q15 and NRRL B-16531. Appl. Environ. Microbiol. 2002, 68, 5933–5942. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Engesser, K.-H.; Kim, S.-J. Physiological, Numerical and Molecular Characterization of Alkyl Ether-Utilizing Rhodococci. Environ. Microbiol. 2007, 9, 1497–1510. [Google Scholar] [CrossRef]
- Takei, D.; Washio, K.; Morikawa, M. Identification of Alkane Hydroxylase Genes in Rhodococcus Sp. Strain TMP2 That Degrades a Branched Alkane. Biotechnol. Lett. 2008, 30, 1447–1452. [Google Scholar] [CrossRef]
- Amouric, A.; Quéméneur, M.; Grossi, V.; Liebgott, P.-P.; Auria, R.; Casalot, L. Identification of Different Alkane Hydroxylase Systems in Rhodococcus Ruber Strain SP2B, an Hexane-Degrading Actinomycete. J. Appl. Microbiol. 2010, 108, 1903–1916. [Google Scholar] [CrossRef]
- Xiang, W.; Liang, Y.; Hong, S.; Wang, G.; You, J.; Xue, Y.; Ma, Y. Degradation of Long-Chain n-Alkanes by a Novel Thermal-Tolerant Rhodococcus Strain. Arch. Microbiol. 2022, 204, 259. [Google Scholar] [CrossRef]
- Bukliarevich, H.A.; Gurinovich, A.S.; Filonov, A.E.; Titok, M.A. Molecular Genetic and Functional Analysis of the Genes Encoding Alkane 1-Monooxygenase Synthesis in Members of the Genus Rhodococcus. Microbiology 2023, 92, 242–255. [Google Scholar] [CrossRef]
- Shanklin, J.; Whittle, E.; Fox, B.G. Eight Histidine Residues Are Catalytically Essential in a Membrane-Associated Iron Enzyme, Stearoyl-CoA Desaturase, and Are Conserved in Alkane Hydroxylase and Xylene Monooxygenase. Biochemistry 1994, 33, 12787–12794. [Google Scholar] [CrossRef]
- Kloos, K.; Munch, J.C.; Schloter, M. A New Method for the Detection of Alkane-Monooxygenase Homologous Genes (AlkB) in Soils Based on PCR-Hybridization. J. Microbiol. Methods 2006, 66, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Fenibo, E.O.; Selvarajan, R.; Abia, A.L.K.; Matambo, T. Medium-Chain Alkane Biodegradation and Its Link to Some Unifying Attributes of alkB Genes Diversity. Sci. Total Environ. 2023, 877, 162951. [Google Scholar] [CrossRef] [PubMed]
- Táncsics, A.; Benedek, T.; Szoboszlay, S.; Veres, P.G.; Farkas, M.; Máthé, I.; Márialigeti, K.; Kukolya, J.; Lányi, S.; Kriszt, B. The Detection and Phylogenetic Analysis of the Alkane 1-Monooxygenase Gene of Members of the Genus Rhodococcus. Syst. Appl. Microbiol. 2015, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Mougou, A.H.; Wu, C.J.; Gleason, M.L.; Rhouma, A. Sequence Analysis of 16S RDNA, GyrB and AlkB Genes of Plant-Associated Rhodococcus Species from Tunisia. Int. J. Syst. Evol. Microbiol. 2020, 70, 6491–6507. [Google Scholar] [CrossRef] [PubMed]
- Ratnikova, M.S.; Titok, M.A. Molecular Genetic Markers for Identification of Rhodococcus erythropolis and Rhodococcus qingshengii. Microbiology 2020, 89, 435–442. [Google Scholar] [CrossRef]
- van Beilen, J.B.; Wubbolts, M.G.; Witholt, B. Genetics of Alkane Oxidation by Pseudomonas Oleovorans. Biodegradation 1994, 5, 161–174. [Google Scholar] [CrossRef]
- Liang, J.-L.; Nie, Y.; Wang, M.; Xiong, G.; Wang, Y.-P.; Maser, E.; Wu, X.-L. Regulation of Alkane Degradation Pathway by a TetR Family Repressor via an Autoregulation Positive Feedback Mechanism in a Gram-Positive Dietzia Bacterium. Mol. Microbiol. 2016, 99, 338–359. [Google Scholar] [CrossRef]
- Chai, J.; Guo, G.; McSweeney, S.M.; Shanklin, J.; Liu, Q. Structural Basis for Enzymatic Terminal C–H Bond Functionalization of Alkanes. Nat. Struct. Mol. Biol. 2023. [Google Scholar] [CrossRef]
- van Beilen, J.B.; Smits, T.H.M.; Roos, F.F.; Brunner, T.; Balada, S.B.; Röthlisberger, M.; Witholt, B. Identification of an Amino Acid Position That Determines the Substrate Range of Integral Membrane Alkane Hydroxylases. J. Bacteriol. 2005, 187, 85–91. [Google Scholar] [CrossRef]
- Zampolli, J.; Collina, E.; Lasagni, M.; Di Gennaro, P. Biodegradation of Variable-Chain-Length n-Alkanes in Rhodococcus Opacus R7 and the Involvement of an Alkane Hydroxylase System in the Metabolism. AMB Express 2014, 4, 73. [Google Scholar] [CrossRef]
- Xiang, W.; Hong, S.; Xue, Y.; Ma, Y. Functional Analysis of Novel AalkB Genes Encoding Long-Chain n-Alkane Hydroxylases in Rhodococcus Sp. Strain CH91. Microorganisms 2023, 11, 1537. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Midpoint rooted maximum likelihood phylogenetic tree of AlkB family alkane monooxygenases of Rhodococcus (sensu lato) group. The tree was constructed using IQ-TREE, automatically determined best-fit substitution model was JTT+F+I+G4. Branches with ultrafast bootstrap support values ≥ 95% (1000 replicates) are marked with white circles. The outer ranges and branches colors marks the proposed AlkB types. The inner ranges and branches colors marks the genus Prescottella ssp. (light blue), the species Antrihabitans cavernicola (blue, purple), and AlkBK type of the species Rhodococcoides kroppenstedtii. The colored squares on the branches marks the AlkB2N (blue) and AlkBX (red) types.
Figure 1.
Midpoint rooted maximum likelihood phylogenetic tree of AlkB family alkane monooxygenases of Rhodococcus (sensu lato) group. The tree was constructed using IQ-TREE, automatically determined best-fit substitution model was JTT+F+I+G4. Branches with ultrafast bootstrap support values ≥ 95% (1000 replicates) are marked with white circles. The outer ranges and branches colors marks the proposed AlkB types. The inner ranges and branches colors marks the genus Prescottella ssp. (light blue), the species Antrihabitans cavernicola (blue, purple), and AlkBK type of the species Rhodococcoides kroppenstedtii. The colored squares on the branches marks the AlkB2N (blue) and AlkBX (red) types.
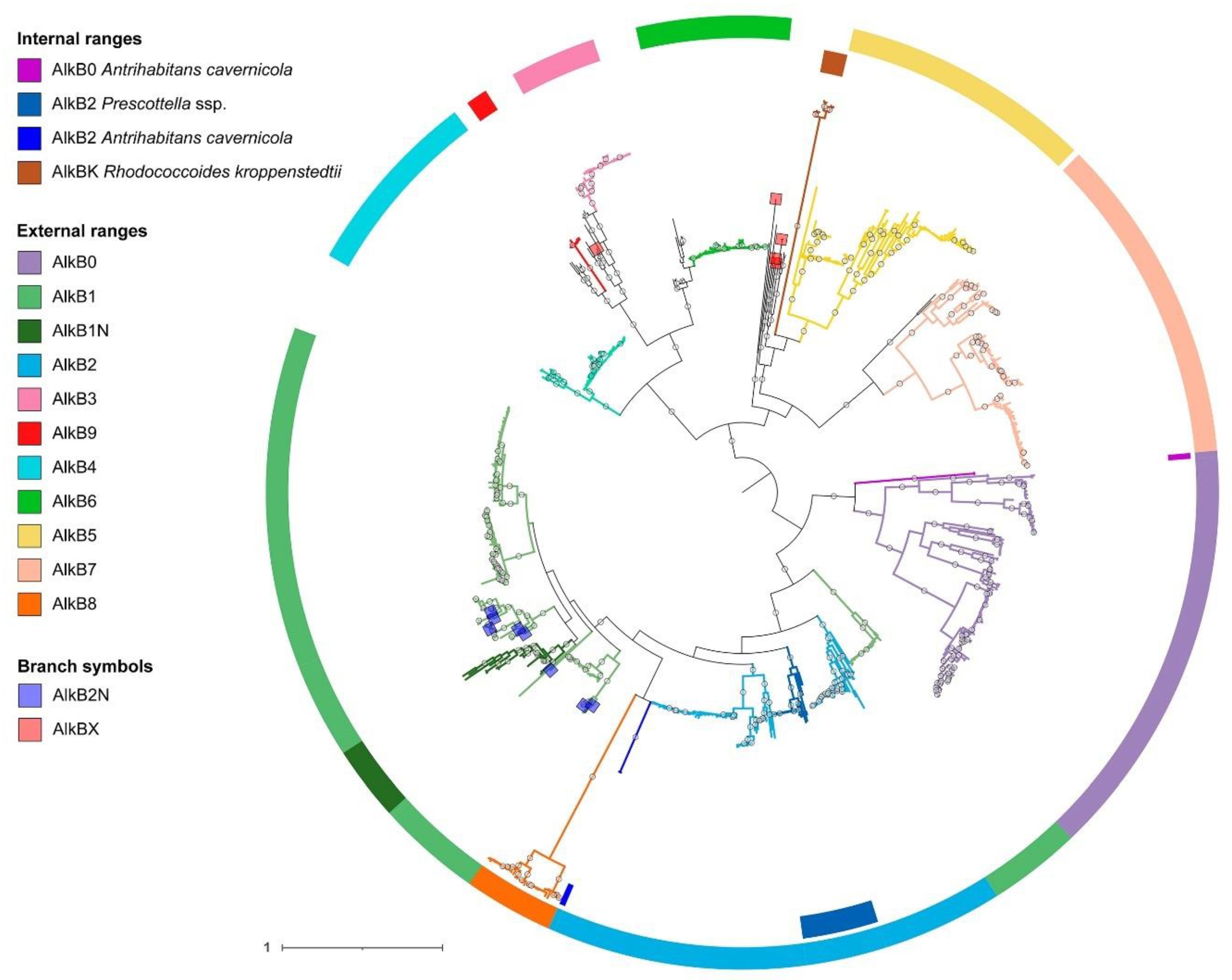
Figure 2.
Composition of the conserved genomic regions for AlkB of different types. merR — MerR family transcriptional regulator; CDGSH — CDGSH iron-sulfur domain-containing protein; GAF/ANTAR — GAF and ANTAR domain-containing protein, ycgM — fumarylacetoacetate hydrolase family protein, gltX — glutamate-tRNA ligase, abiEi — AbiEi family antitoxin domain-containing protein, glpC — iron-sulfur binding reductase, araC — AraC family transcriptional regulator, troA — TroA-like ABC transporter substrate-binding protein, ywnB — NAD(P)H-binding protein, rssA — patatin-like phospholipase family protein, ytpA — phospholipase YtpA (alpha/beta hydrolase), AAA — ATP-binding protein (AAA+ ATPase superfamily), leuA — 2-isopropylmalate synthase, mshA — inositol-3-phosphate glycosyltransferase, menH — alpha/beta fold hydrolase, ROK — ROK family transcriptional regulator.
Figure 2.
Composition of the conserved genomic regions for AlkB of different types. merR — MerR family transcriptional regulator; CDGSH — CDGSH iron-sulfur domain-containing protein; GAF/ANTAR — GAF and ANTAR domain-containing protein, ycgM — fumarylacetoacetate hydrolase family protein, gltX — glutamate-tRNA ligase, abiEi — AbiEi family antitoxin domain-containing protein, glpC — iron-sulfur binding reductase, araC — AraC family transcriptional regulator, troA — TroA-like ABC transporter substrate-binding protein, ywnB — NAD(P)H-binding protein, rssA — patatin-like phospholipase family protein, ytpA — phospholipase YtpA (alpha/beta hydrolase), AAA — ATP-binding protein (AAA+ ATPase superfamily), leuA — 2-isopropylmalate synthase, mshA — inositol-3-phosphate glycosyltransferase, menH — alpha/beta fold hydrolase, ROK — ROK family transcriptional regulator.
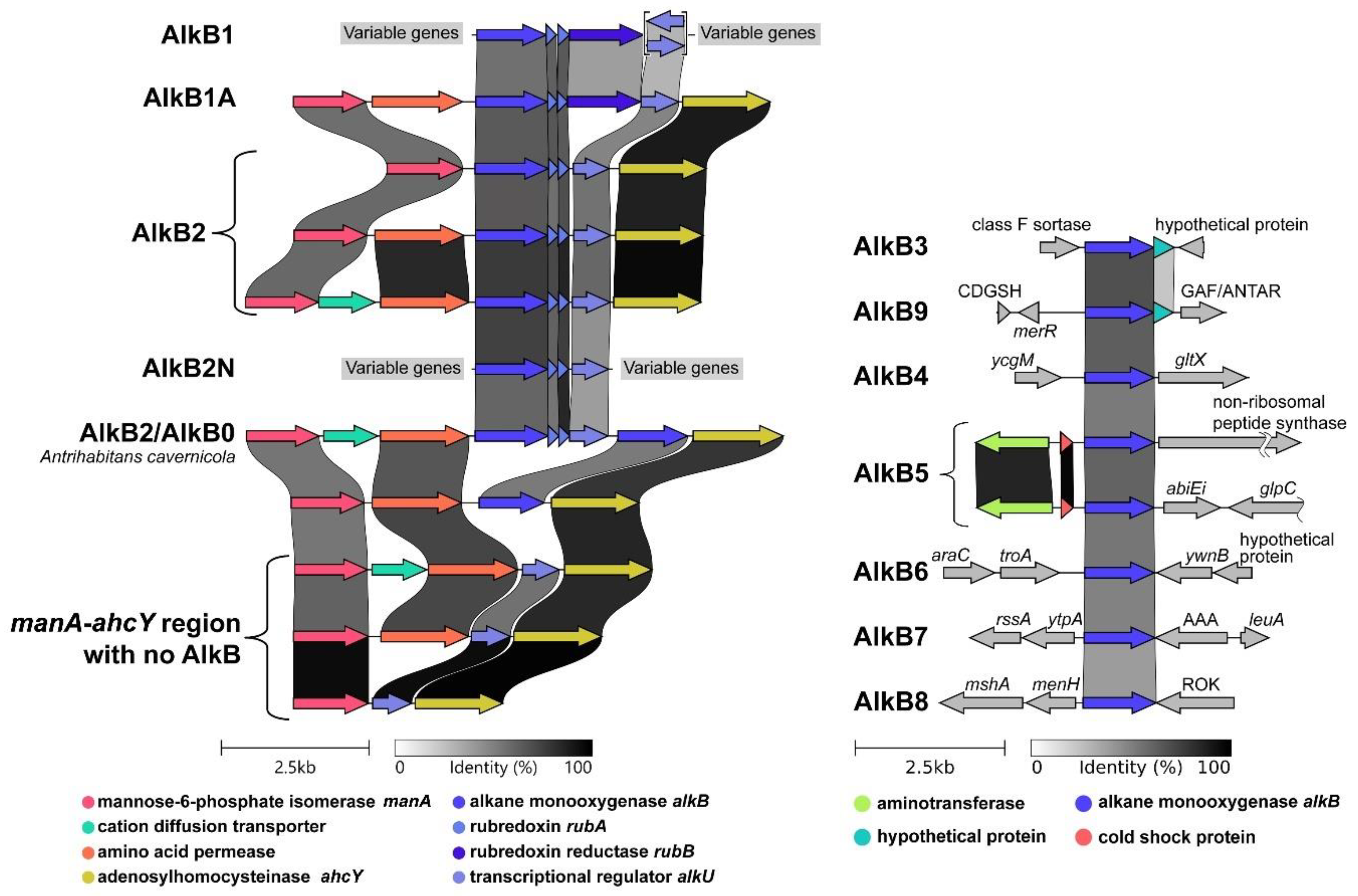
Figure 3.
Comparison of different gene organization variants of the manA-ahcY region.
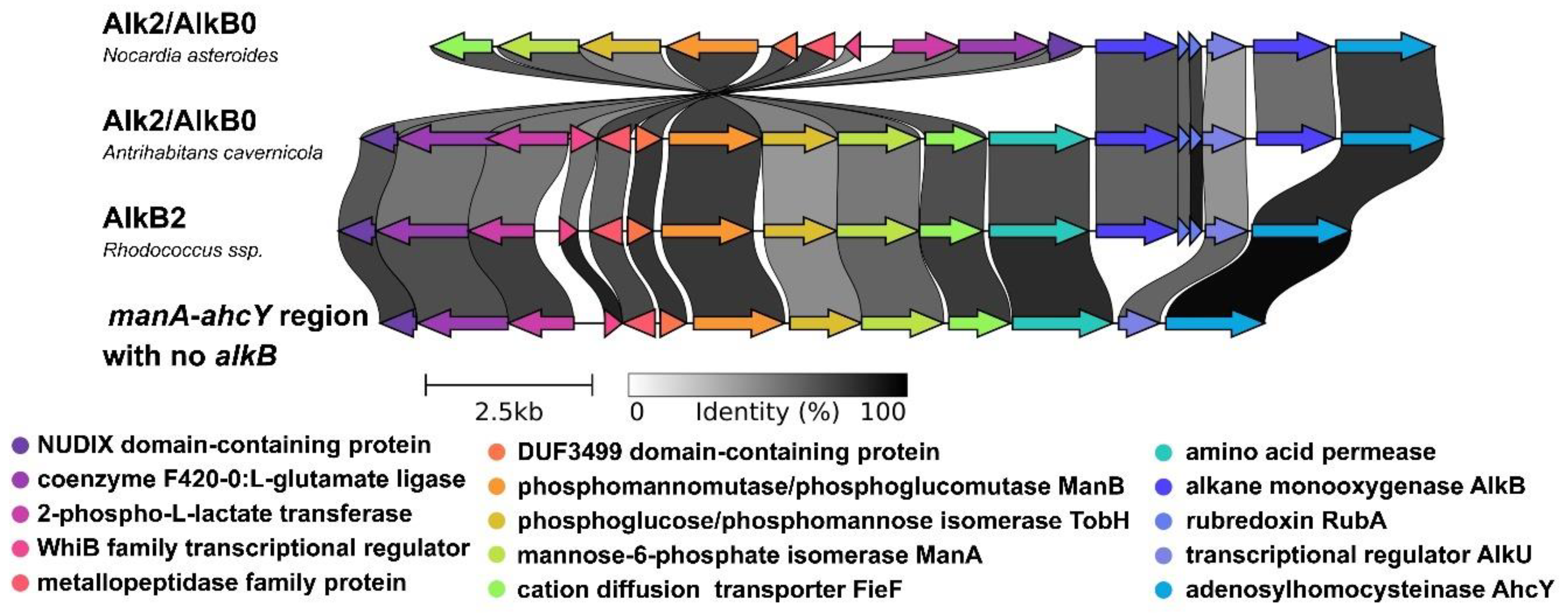
Figure 4.
Midpoint rooted phylogenetic tree of type strains of Rhodococcus (sensu lato) group and related species constructed using GTDB-tk. Branches with bootstrap support values ≥ 80% are marked with blue circles. “AlkB copy numbers” column shows the number of AlkB types in the genomes of species representatives. The size of the labels correspond to the the number of genomic regions considered in the paper for representatives of this species. Cluster containing all representatives of the genus Rhodococcoides and the clade consisting of all Prescotella ssp. is highlighted in colour.
Figure 4.
Midpoint rooted phylogenetic tree of type strains of Rhodococcus (sensu lato) group and related species constructed using GTDB-tk. Branches with bootstrap support values ≥ 80% are marked with blue circles. “AlkB copy numbers” column shows the number of AlkB types in the genomes of species representatives. The size of the labels correspond to the the number of genomic regions considered in the paper for representatives of this species. Cluster containing all representatives of the genus Rhodococcoides and the clade consisting of all Prescotella ssp. is highlighted in colour.
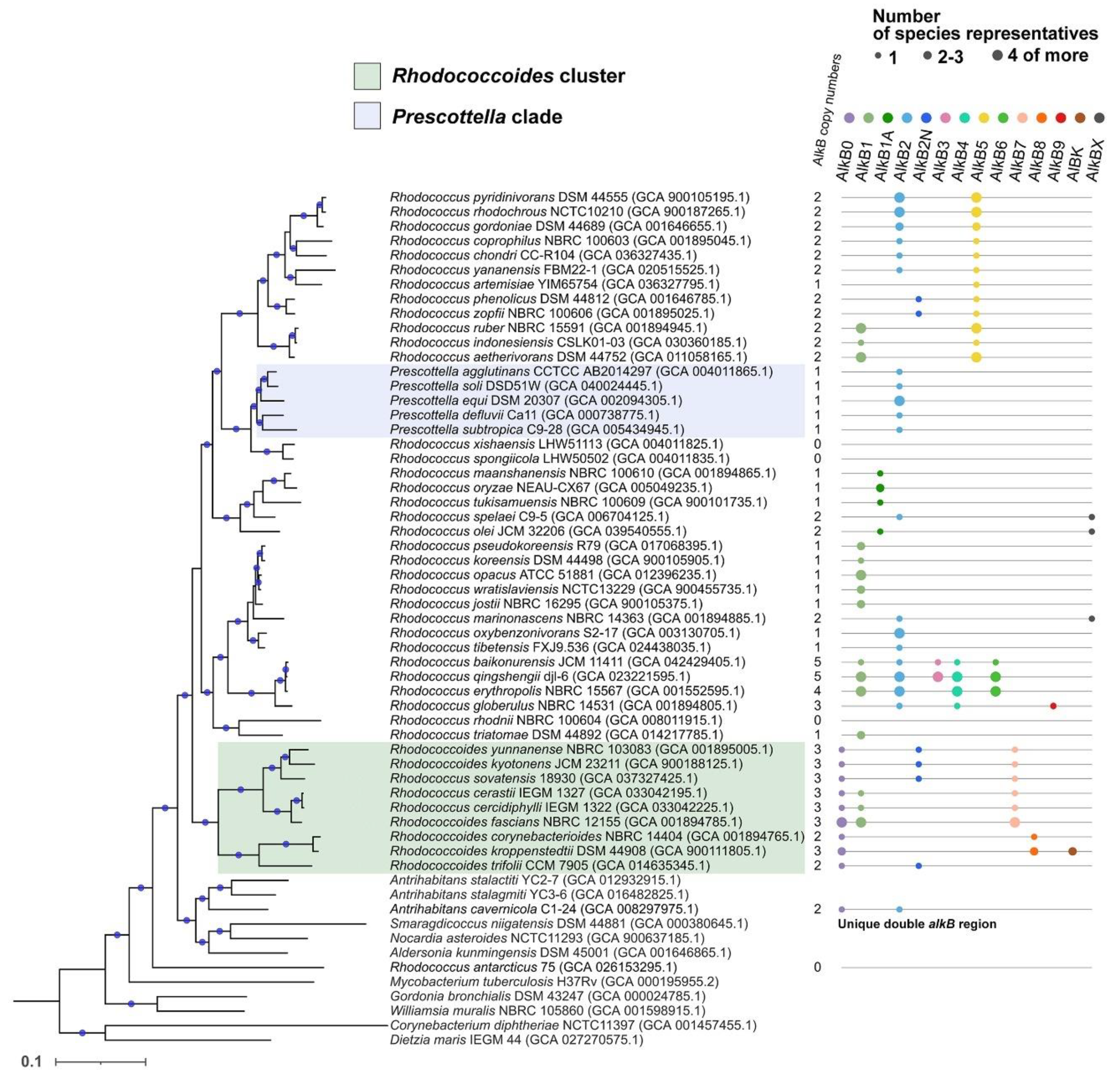
Figure 5.
Previously proposed “new” types of AlkB and generalized AlkB groups. Midpoint rooted maximum likelihood phylogenetic tree of AlkB family alkane monooxygenases of Rhodococcus (sensu lato) group with the addition of partial sequences of previously described alkane monooxygenases. The tree was constructed using IQ-TREE, automatically determined best-fit substitution model was JTT+F+I+G4. Branches with ultrafast bootstrap support values ≥ 95% (1000 replicates) are marked with white circles. The colored squares on the branches marks all previously described alkane monooxygenases (Table 2). The colored ranges corresponde proposed generalized AlkB types.
Figure 5.
Previously proposed “new” types of AlkB and generalized AlkB groups. Midpoint rooted maximum likelihood phylogenetic tree of AlkB family alkane monooxygenases of Rhodococcus (sensu lato) group with the addition of partial sequences of previously described alkane monooxygenases. The tree was constructed using IQ-TREE, automatically determined best-fit substitution model was JTT+F+I+G4. Branches with ultrafast bootstrap support values ≥ 95% (1000 replicates) are marked with white circles. The colored squares on the branches marks all previously described alkane monooxygenases (Table 2). The colored ranges corresponde proposed generalized AlkB types.
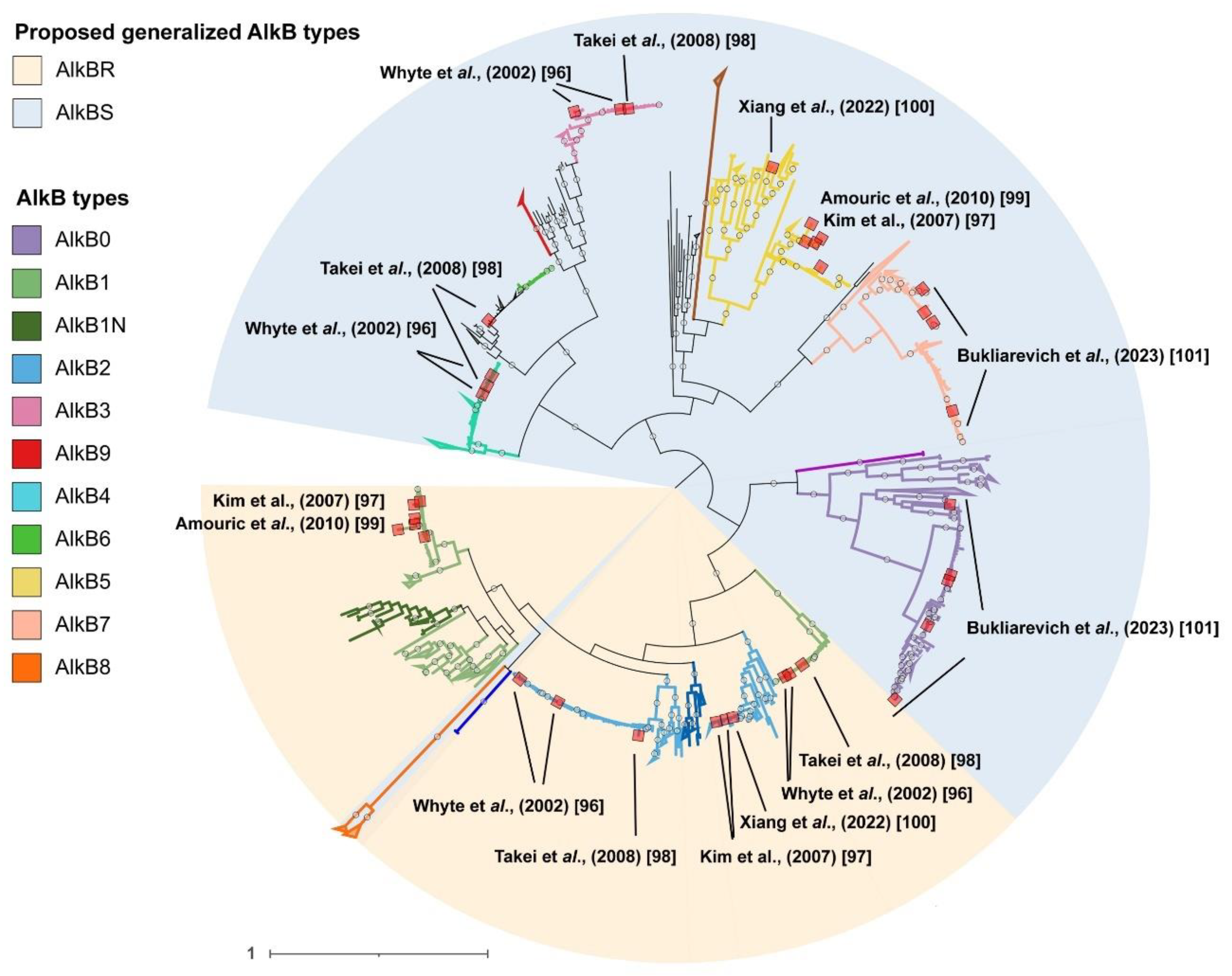
Figure 6.
Sequence logos of different types of alkane monooxygenases for regions of signature amino acid motifs.
Figure 6.
Sequence logos of different types of alkane monooxygenases for regions of signature amino acid motifs.


Table 1.
List of species belonging to the genus Rhodococcus and shows inconsistency of taxonomic assignment in different databases.
Table 1.
List of species belonging to the genus Rhodococcus and shows inconsistency of taxonomic assignment in different databases.
| NCBI Taxonomy | LPSN status of NCBI Taxonomy | GTDB Taxonomy | TSGS Taxonomy |
| Antrihabitans cavernicola | correct name, synonym: Rhodococcus cavernicola |
Rhodococcus_E cavernicola | Rhodococcus cavernicola |
| Prescottella agglutinans | correct name | Rhodococcus agglutinans | Rhodococcus agglutinans |
| Prescottella defluvii | correct name |
Rhodococcus defluvii, Rhodococcus defluvii_A |
Rhodococcus defluvii |
| Prescottella equi | correct name |
Rhodococcus equi, Rhodococcus equi_A |
Rhodococcus equi |
| Prescottella soli | synonym, correct name: Rhodococcus soli | absent | Rhodococcus soli |
| Prescottella subtropica | synonym, correct name: Rhodococcus subtropicus | Rhodococcus subtropicus | Rhodococcus subtropicus |
| Rhodococcoides corynebacterioides | synonym, correct name: Rhodococcus corynebacterioides |
Rhodococcus corynebacterioides, Rhodococcus corynebacterioides_A |
Rhodococcus corynebacterioides |
| Rhodococcoides fascians | synonym, correct name: Rhodococcus fascians |
Rhodococcus fascians, Rhodococcus fascians_E |
Rhodococcus fascians |
| Rhodococcoides kroppenstedtii | synonym, correct name: Rhodococcus kroppenstedtii | Rhodococcus kroppenstedtii | Rhodococcus kroppenstedtii |
| Rhodococcoides kyotonense | synonym, correct name: Rhodococcus kyotonensis |
Rhodococcus kyotonensis, Rhodococcus kyotonensis_B |
Rhodococcus kyotonensis |
| Rhodococcoides trifolii | synonym, correct name: Rhodococcus trifolii |
Rhodococcus trifolii | Rhodococcus trifolii |
| Rhodococcoides yunnanense | synonym, correct name: Rhodococcus yunnanensis |
Rhodococcus yunnanensis | Rhodococcus yunnanensis |
| Rhodococcus antarcticus | correct name | Rhodococcus_D antarcticus | Rhodococcus antarcticus |
| Rhodococcus baikonurensis | synonym, correct name: Rhodococcus erythropolis |
Rhodococcus qingshengii | Rhodococcus baikonurensis |
| Rhodococcus chondri | correct name | absent | Rhodococcus chondri |
| Rhodococcus indonesiensis | correct name | Rhodococcus sp030360185 | Rhodococcus indonesiensis |
| Rhodococcus olei | correct name | absent | Rhodococcus olei |
| Rhodococcus qingshengii | synonym, correct name: Rhodococcus erythropolis |
Rhodococcus qingshengii, Rhodococcus qingshengii_B |
Rhodococcus qingshengii |
| Rhodococcus sovatensis | correct name | absent | absent |
| Rhodococcus tibetensis | not validly published | Rhodococcus sp024438035 | Rhodococcus tibetensis |
Table 2.
Previously proposed “new” types of AlkB.
| Suggested namea | Genomic context if available | Protein GenBank ID | Strain | Source |
|---|---|---|---|---|
| AlkB1 | AlkB, RubA, RubA, RubB, AlkU | CAB51053.2 | R. erythropolis NRRL B-16531 | [96] |
| AlkB2 | cationic transporter†, AlkB, RubA, RubA, AlkU† | CAC37038.1 | ||
| AlkB3 | (putative) exported protein†, alkB | CAC40953.1 | ||
| AlkB4 | AlkB, glutamil t-RNA synthetase | CAC40954.1 | ||
| AlkB1 | AlkB, AubA, RubA, RubB, AlkU | AAK97448.1 | Rhodococcus sp. Q15 | |
| AlkB2 | cationic transporter†, AlkB, RubA, RubA, AlkU | AAK97454.1 | ||
| AlkB3 | — | AAK97446.1 | ||
| AlkB4 | — | AAK97447.1 | ||
| AlkB5 | — | ABI13998.1† | R. rhodochrous 116 | [97] |
| AlkB7 (AlkB1) | — | ABI13999.1† | Rhodococcus sp. DEE5151 | |
| AlkB5 | — | ABI14000.1† | Rhodococcus sp. DEE5301 | |
| AlkB7 (AlkB1) | — | ABI14001.1† | Rhodococcus sp. DEE5311 | |
| AlkB5 | — | ABI14002.1† | Rhodococcus sp. DEE5316 | |
| AlkB6 (AlkB2) | — | ABI14003.1† | Rhodococcus sp. DEOB100 | |
| AlkB7 (AlkB1) | — | ABI14004.1† | Rhodococcus sp. MOB100 | |
| AlkB6 (AlkB2) | — | ABI14005.1† | Rhodococcus sp. MOP100 | |
| AlkB7 (AlkB1) | — | ABI14006.1† | Rhodococcus sp. THF100 | |
| AlkB1 | — | BAG06232.1† | Rhodococcus sp. TMP2 | [98] |
| AlkB2 | — | BAG06233.1† | ||
| AlkB3 | — | BAG06234.1† | ||
| AlkB4 | — | BAG06235.1† | ||
| AlkB5b (AlkB6) | — | BAG06236.1† | ||
| AlkBa (AlkB1) | RubA, RubA, RubB† | ACX30747.1 | R. ruber SP2B | [99] |
| AlkBb (AlkB5) | — | ACX30751.1† | ||
| AlkBa (AlkB1) | — | ACX30755.1 | R. ruber DSM 43338 | |
| AlkBb (AlkB5) | — | ACX30752.1† | ||
| AlkB2 | ManA, AlkB, RubA, RubA, TetR, AhcY | WP_241385946.1 | Rhodococcus sp. CH91 | [100] |
| AlkB new (AlkB5) | aminotransferase, cold-shock protein, AlkB, AbiEi family protein | WP_241384812.1 | ||
| AlkB7 | alpha/beta hydrolase | AMY23060 | Rhodococcus fascians PBTS 2 | [101] |
| AJW41446 | Rhodococcus sp. B7740 | |||
| QII08422 | Rhodococcus fascians A25f | |||
| QIH99153 | Rhodococcus fascians A2ld2 | |||
| AlkB8 (AlkB0) | ManA, AlkB, AhcY | AYJ49258 | Rhodococcus sp. PI Y | |
| AMY51400 | Rhodococcus fascians D188 | |||
| AMY24665 | Rhodococcus fascians PBTS 2 | |||
| AJW39705 | Rhodococcus sp. B7740 | |||
| QII00755, QII06794c | Rhodococcus fascians A2ld2, Rhodococcus fascians A25f |
a The AlkB type we proposed is indicated in brackets if different. b Does not take into account the results of the Kim et al., 2007. c Identiclal sequences. † Partial sequence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



