
Submitted:
05 December 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
Three fungal strains were employed for stereoselective oxidation of the cheap and commercially available substrate 2-phenylethanol, what resulted in chiral building blocks receiving. Whole-cells biocatalysts were as follows: Beauveria bassiana DSM 1344, Beauveria brongniartii DSM 6651 and Rhizopus arrhizus DSM 1185. The main product of Beauveria bassiana bioconversion, was 1-phenylethane-1,2-diol obtained, depending on the form of the biocatalyst, as a R-enantiomer (e.g. 99.9%), with fresh biomass application or as racemic mixture in case of immobilization in agar- agar. The very best and pioneer results for the synthesis of R-enantiomer of diol were received under precisely defined conditions, and as a results of scaling study conducted in automatic bath reactor (Table 2.). This is pioneer result, since in the previous studies, fresh mycelium of Aspergillus niger allowed receiving this product as (S) enantiomer. Also the use of Rhizopus arrhizus DSM 1185 (immobilized in polyurethane foams), gave important results, since the bioconversion of phenyl ethanol led, indeed to the racemic mixture of 1-phenylethane-1,2-diol, but it was accompanied by the noticeable tyrosol synthesis, what is reported for the first time.
Keywords:
biotransformation
; 2-phenylethanol
; fungi
; Beauveria bassiana
; Beauveria brongniartii
; Rhizopus arrhizus
; (R)-1-phenylethane-1
; 2-diol
; tyrosol
1. Introduction
Whole-cell biocatalysis based on the use of free or immobilized fungal, bacterial or plant cells is an important method for obtaining compounds with broad applications in the chemical, biofuel, pharmaceutical and cosmetic industries [1,2,3,4]. The possibility of carrying out selective transformations under mild conditions with reduced waste generation means, that biocatalyzed reactions are an attractive alternative to classical chemical methods. Biotransformations of compounds of expand steric hindrance are of special interest, including polycyclic compounds, where the introduction of functional groups at the specific location or/and specific configuration receiving, is particularly difficult by classical methods. Fungal biocatalysts are valuable for this type of modification, especially, since it is possible to direct the selectivity of the reaction by simply manipulating the form of the biocatalyst or by engineering the reaction conditions [5].
Entomopathogenic fungi of the genus Beauveria sp. are known for their broad spectrum of enzymatic activities e.g. Beauveria bassiana is one of the most common fungal biocatalyst [6]. It is well-known in steroids [7,8,9,10] and flavonoids biotransformations [11,12,13]. Its hydroxylation, oxidation, glycosidation, Baeyer–Villiger oxidation and ester hydrolysis activities [6] allows the production of compounds, that are difficult to synthesize by traditional chemical methods.
Another example is Rhizopus arrhizus strain also widely used in biotransformation processes due to its hydroxylation and selective reduction activity. Its biocatalytic capabilities have been described in the literature, among others, in reduction of arylalkanones [14] and acetophenone [15], in synthesis of chiral secondary carbinols and halohydrins [16] and in biotransformation of betulonic acid [17]. Similarly to B. bassiana, it has found application in the biotransformation of steroids [18].
From a practical point of view, an important group of compounds are low molecular weight polyphenols such as hydroxyl derivatives of 2-phenylethanol. These compounds are applied as factors because of their biological activity. For example, tyrosol (4-(2-hydroxyethyl)phenol) is a very well-known cosmetic ingredient [19]. It is one of the main phenolic compounds naturally occurring in olive oil [20]. In biotransformation studies it serves as a substrate for obtaining hydroxytyrosol [21,22] and its derivatives have been reported as ones, which exhibit antioxidant, anticancer, antiviral or anti-inflammatory activity [23,24,25]. Another 2-phenylethanol derivative (R)-1-phenyl-1,2-ethanediol is applied as biomarker in the exposure of humans to styrene [26] and as an intermediate used in the production of pharmaceutical [27,28,29], agrochemical and chemical industries [28,29]. Biocatalytic preparation of this compound, without the burden of the environment is an alternative to organic synthesis. The substrates in (R)-1-phenyl-1,2-ethanediol synthesis via biocatalytic methods are mainly 2-hydroxyacetophenone, racemic styrene oxide and phenylglyoxal [30,31,32,33,34].
As an alternative biocatalytic pathway the biohydroxylations of cheap, commercially available substrate- 2-phenylethanol (Figure 1) [35,36,37] may be considered.
So far, the biotransformations of 2-phenylethanol have resulted in compounds such as hydroxytyrosol, tyrosol, (S)-1-phenylethane-1,2-diol and 4-hydroxyphenylacetic acid [36,37]. All these chemical compounds are widely used in various industries. Previous studies demonstrated that, among others, Aspergillus niger IAFB 2301 cells and Aspergillus niger (OPI) spores are capable of bioconverting 2-phenylethanol to its derivatives [36,37]. However, bioconversion of 2-phenylethanol into the opposite enantiomer - (R)-1-phenylethane-1,2-diol and tyrosol derivatives has not been achieved yet. This was an assumption to further research with next fungal strains.
In this study, three different fungal strains were applied for the biotransformation of 2-phenylethanol: Beauveria bassiana DSM 1344, Beauveria brongniartii DSM 6651 and Rhizopus arrhizus DSM 1185.
The biocatalytic capacity of biocatalyst cells is influenced by its preparation to the process. So, methods such as immobilization (e.g. polyurethane foams [36], agar-agar and alginate [38]) or pre-incubation under starvation conditions were checked [39]. Such modifications, usually affect the reaction efficiency and the optical purity of the obtained product [5]. This depends on the applied conditions as once altering the permeability of cellular envelopes (immobilization) or in case of nutrient deficiency, inducing the activity of the enzymes, which are knock out in the cells cultivated under optimal conditions.
The research includes a number of experiments, finally resulted in receiving of the optically pure (R)- 1-phenylethane-1,2-diol and tyrosol and also forming the racemic mixture of diol (Scheme 1).
2. Materials and Methods
2.1. Chemicals
Cultivation medium as recommended by DSMZ- German Collection of Microorganisms and Cell Cultures GmbH for B. brongniartii: 4 g yeast extract, 15 g soluble starch, 1 g K2HPO4, 0.5 g MgSO4 × 7 H2O per litre; B. bassiana: 30 g malt extract, 3 g soya peptone per litre. Medium for R. arrhizus: potato dextrose broth (PDB): 4 g potato extract, 20 g glucose per litre.
Commercially available standards of (R)-1-phenylethane-1,2-diol, (S)-1-phenylethane-1,2-diol, 1-phenylethane-1,2-diol, tyrosol (4-(2-hydroxyethyl)phenol), 2-phenylethanol, Triton X-100 and sodium alginate were purchased from Sigma-Aldrich.
Ethyl acetate, anhydrous magnesium sulphate, ethanol (99,8%), n-hexane (HPLC), 2-propanol (HPLC), acetonitrile (HPLC), formic acid, glucose, agar-agar, dipotassium phosphate were purchased from Avantor.
Agar-agar, soya peptone, malt extract, Potato Dextrose Broth (PDB) and Potato Dextrose Agar (PDA) were purchased from Biocorp, soluble starch from POCH and yeast extract from VWR Chemicals.
Calcium chloride, barium chloride and magnesium sulfate heptahydrate were purchased from Chempur.
Polyurethane foams (porosity of 740-1040 µm or 1060-1600 µm or 2300-3300 µm) were purchased from Bogmar.
2.2. Microorganisms
Beauveria bassiana (DSM 1344), Beauveria brongniartii (DSM 6651), Rhizopus arrhizus (DSM 1185) were purchased from German Collection of Microorganisms and Cell Cultures.
2.3. Cultivation Conditions
2.3.1. Inoculum
Inoculum for microorganisms cultivation was consisted of their spores which were recovered from sporulated Petri dishes with solid medium (PDA). Spores were washed with a 10 mL of sterile Triton X-100 water solution (0,05%), suspended and transferred then to conical flasks. The concentration of the spore solution was adjusted to about 1,4·106 spores/mL for R. arrhizus, 3·109 spores/mL for B. bassiana and 4·107 spores/mL for B. brongniartii.
2.3.2. Cultivation Conditions - Fresh Biomas of Microorganisms
All cultures were grown in 250 mL flasks containing 100 mL of growth medium on a rotary shaker (135 rpm) at 24°C for 3 days (for R. arrhizus), 5 days (for B. bassiana), 6 days (for B. brongniartii) until the mid-log growth phase was reached. The average amount of biomass grown per 100 mL of culture medium was 10 g (R. arrhizus), 26 g (B. bassiana) and 16 g (B. brongniartii). Then these amounts were transferred each into 50 mL of biotransformation medium (distilled water).
2.3.3. Immobilization Procedure- Growth of the R. arrhizus and B. brongniartii in the Presence of Polyurethane Foams with Different Porosity
The polyurethane foams were cut into 1 cm cubes (porosity of 740-1040 µm or 1060-1600 µm or 2300-3300 µm), washed with sterile water and autoclaved in 250 mL Erlenmeyer’s flasks containing 100 mL of growth medium. Inoculum containing spores solution (500 µL) was transferred aseptically to sterile medium with polyurethane foams (20 cubes in each flask). The cultures were incubated at the same conditions as fresh biomass for 3 days (R. arrhizus) and 6 days (B. brongniartii).
2.3.4. Immobilization of B. bassiana Biomass in Agar-Agar
After incubation (6 days) fresh biomass was separated from growth medium by centrifugation (4000 rpm, 10 min, 4°C), then washed with distilled water, centrifuged again and then was used in immobilization process. Agar-agar solution (6.0%) was added to fresh biomass in a 1:1 ratio. Then solidifying fungal cells immobilized in agar-agar was divided into small pieces.
2.3.5. Immobilization of B. bassiana Biomass in Calcium Alginate
After incubation fresh fungal biomass (26 g) was separated from growth medium by centrifugation (4000 rpm, 10 min, 24°C), washed with distilled water, centrifuged again and then was used in immobilization process. Distilled water was added to fresh biomass (1:4.8 ratio) and mixed with equal portion of sodium alginate solution (2.0 %). The mixture was added dropwise to 1% CaCl2 water solution and left for 30 min. Fungal cells immobilized in calcium alginate beads were separate by filtration using Büchner funnel and transferred to 0,8% BaCl2 water solution for next 10 min. Immobilized fungal cells were separated again by filtration using Büchner funnel and then transferred to 250 ml conical flasks containing 50 ml of distilled water as a biotransformation medium.
2.3.6. Pre-Incubation of Fresh Biomass of R. arrhizus
Pre-incubation of mycelium of R. arrhizus under starvation conditions was performed in 250 mL conical flasks containing 50 mL of distilled water on a rotary shaker (135 rpm) at room temperature for 24 hours. After that, the biocatalyst was used for biotransformation reaction.
2.4. Procedures of Biotransformation
After cultivation, the biocatalyst (fresh biomass or immobilized ones) was transferred to conical flasks (250 mL) with biotransformation media (50 mL of distilled water) and the different amount (depending on the outcome) of the 2-phenylethanol was added as a substrate: 15 mg (2.5 mM) or 30 mg (5 mM) or 60 mg (10 mM). Flasks were shaken at 135 rpm at room temperature. Biotransformations were carried out for 3 hours and 1 to 5 days. Experiments were performed in triplicate. After each incubation period, biocatalyst was removed by filtration (R. arrhizus) or centrifugation 4000 rpm, 10 min, 4°C (B. bassiana, B. brongniartii) and the supernatant was extracted twice with ethyl acetate. Organic layers were dried over anhydrous MgSO4 and evaporated under reducing pressure. Final products mixture was dissolved in ethanol (99,8 %) or HPLC eluent mixture (acidic H2O (0.1% formic acid) and acetonitrile, 91:9) and analysed by HPLC method.
2.5. Semi-Preparative Biotransformation Procedures
2.5.1. Simplified Flow Bioreactor for R. arrhizus
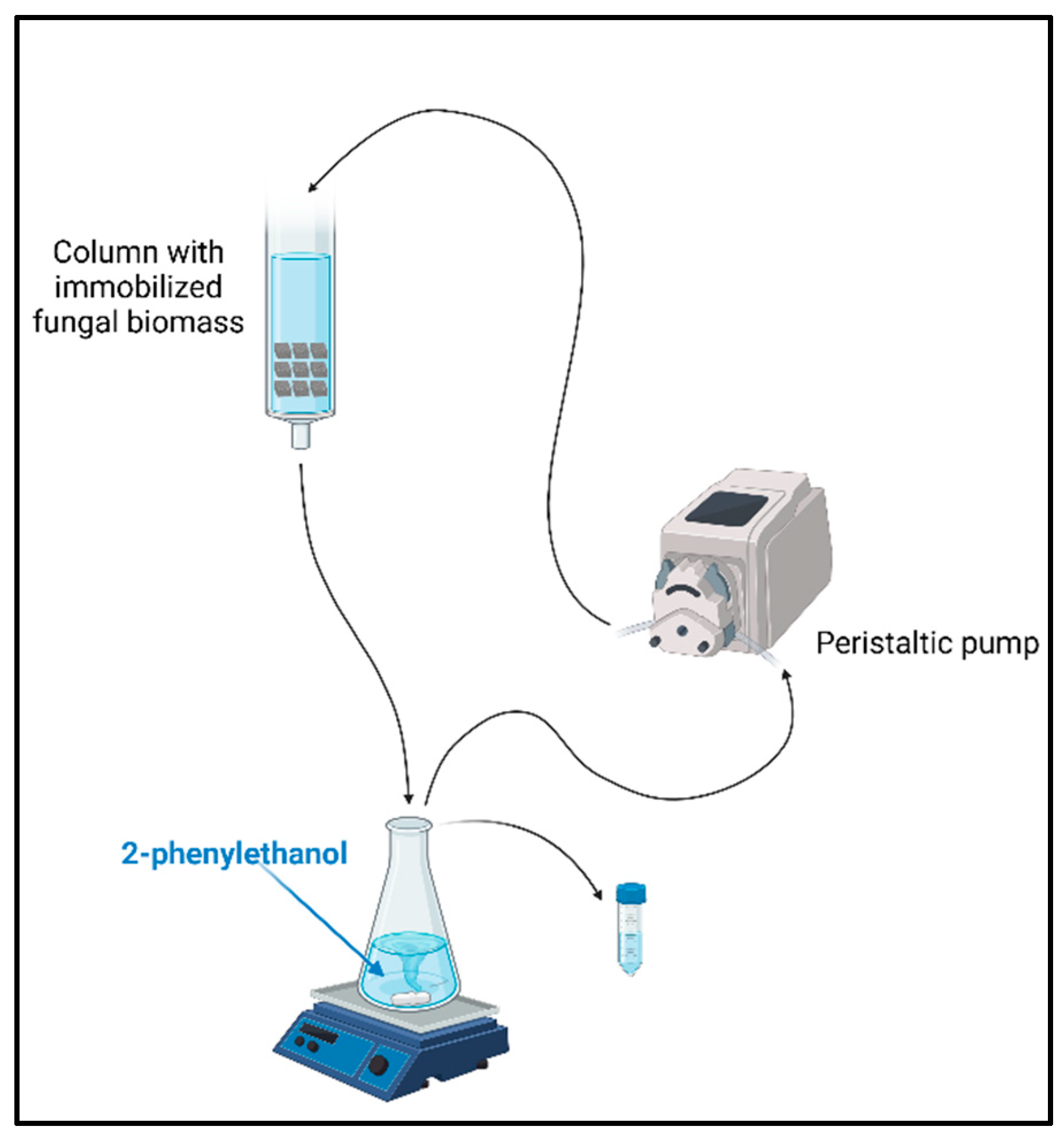
A simplified bioreactor model (Figure 2) was packed with immobilized cells (1060-1600 µm polyurethane foams, 20 cubes) and fill with biotransformation medium (150 mL of distilled water) containing substrate (5 mM) flowing continuously by the inside. The circulation of the medium through the column has been forced by peristaltic pump (3 mL min−1). Progress of bioconversion was monitored by HPLC analysis after 3 hours and after 1 to 5 days in the same time intervals (24h).
2.5.2. Batch Bioreactor for B. bassiana
The microbiological bioreactor (Applikon Biotechnology my-Control, capacity of 1 L) was packed with fresh biomass (100 g/600 mL biotransformation medium- distilled water). Substrate was added (600 mg- 8.36 mM) and biotransformation proceeded for 5 days. The process was carried out with stirring at 400-700 rpm, temperature of 25°C and aeration (1 L min−1). Samples were collected within 24-hour time intervals and analysed by HPLC method.
2.6. Analytical Methods
1-phenylethane-1,2-diol and tyrosol were detected by HPLC (Beckman System Gold 126 Solvent Module, Supelcosil LC-18 column, 25 cm × 4.6 mm, 5 µm). Acidic H2O (0.1% formic acid) and acetonitrile were used as the mobile phase at a flow rate of 1 mL min−1. Percentage of eluent composition (% acidic H2O : % acetonitrile) was 91:9. Injections were done in full loop (20 µL). Products were detected with λ= 276 nm. Retention times were as follows: products - tyrosol (12.2 min) and 1-phenylethane-1,2-diol (13.2 min); substrate- 2-phenylethanol (26.6 min)
(R)-1-phenylethane-1,2-diol was detected by HPLC (AccQPrep HP125 UV-Vis, CHIRALCEL – OB column, particle size 10 μm, 4.6 mm × 250 mm). The analysis was carried out using n-hexane/2-propanol (96:4) as an eluent, at a flow rate 1.0 mL min−1, temperature: 24°C. Product was detected with λ= 254 nm. Retention times were as follows: product - (R)-1-phenylethane-1,2-diol (16.9 min); (S)-1-phenylethane-1,2-diol (20.9 min); substrate - 2-phenylethanol (13.3 min).
Results were confirmed by comparison to commercially available standards and the amount of the products was determined using calibration curves.
3. Results and Discussion
An unquestionable advantage of biotransformations is the ability to direct the bioconversion of particular substrate into desired product, by simply altering the reaction conditions and modifying the form/mode of the biocatalyst. Among fungal strains, even familiar ones, the diversity of the enzymes activities involved in metabolism is enormous. The differences are strain-specific or are related to external factors impacting the cells, e.g. temperature, pH or xenobiotics presence. Therefore, the change in the single parameter of the reaction can be a “game changer”, allowing achieving satisfactory results. The appropriate selection of the reaction conditions and immobilization method can have a significant impact on the biocatalyst's resistance to external factors [40] as well as on the reaction efficiency and enantioselectivity [5].
Three different fungal strains (Beauveria bassiana DSM 1344, Beauveria brongniartii DSM 6651 and Rhizopus arrhizus DSM 1185) were used for the biotransformation of 2-phenylethanol. Different conditions (pre-incubation under starvation conditions, different process duration and substrate concentration) and different immobilization methods (polyurethan foams, agar-agar and calcium alginate) were used to optimize the biocatalytic capabilities of these whole-cell biocatalysts.
Results collected in Table 1. show, that depending on the conditions, bioconversion with B. bassiana DSM 1344 and R. arrhizus DSM 1185, allowed forming (R)- 1-phenylethane-1,2-diol and as racemic mixture, also tyrosol was received (R. arrizus).
A key parameter that must be under consideration in biocatalytic process is the appropriate substrate concentration, which critically influence the yield of the reaction by impacting the biocatalyst cells. So, differed in 2-phenylethanol concentration (15, 30 and 60 mg/50 mL- flasks system) were checked to study biomass sensitivity.
These parameter is crucial one, what can be seen on Scheme 2, which correlated the mode of the biocatalysts, the duration of the process, the substrate concentration with the progress of biotransformation of 2-phenylethanol catalysed by R. arrhizus. This reaction leads to the formation of the mixture of the products, which is consisted of racemic 1-phenylethane-1,2-diol (main product) and tyrosol (up to 5% in the product mixture). In the initial phase of biotransformation (3 hours), the amount of substrate used has little effect on the reaction (Scheme 2). However, with time extending, biocatalytic process with 30 mg of substrate (in 50 mL of medium), resulted in better bioconversion progress in 3 day (1,23 mg). On the other hand, increasing amount of 2-phenylethanol to 60 mg resulted in more products mixture formation in two day (1,85 mg). Unfortunately, further extension of time causes a decrease in the amount of products formed, which may be the result of the toxic effect of the xenobiotic used [41,42], or the resulting derivatives undergo further metabolic changes.
In comparison to the above, in the case of the Beauveria bassiana strain, similarly, the optimal time of bioconversion of 2-phenylethanol used in the amount of 60 mg/ 50 mL was the third day. Although in this case the amount of the product lower (0.38 mg) than in the case of R. arrhizus, in exchange for that, the reaction was highly enantioselective and the only product formed was (R)-1-phenylethane-1,2-diol (e.e. 99.9%). It is worth noting that in this case, strict control of the biotransformation time was necessary because in the following days a decrease in the enantiomeric excess of the product was observed as a probable effect of the increase in the activity of enzymes with opposite enantioselectivity. This results were the assumption for further studies on B. bassiana applying as biocatalysts.
Since, the experimental with this strain on laboratory scale (50 mL of biotransformation medium), resulted in pure (R)-1-phenylethane-1,2-diol formation, scaling attempts were made. It allows carrying the reaction under controlled conditions in a 1 L bioreactor, and resulted in a 12-fold scale increasing. As on the laboratory scale, during the biotransformation in the bioreactor, the third day turned out to be optimal for (R)-1-phenylethane-1,2-diol (28.8 mg / 600 mL) production (Table 2). It is worth emphasizing that despite the decrease in the amount of the product formed in the subsequent days of biotransformation (probable further metabolic changes), no decrease in the enantioselectivity of the reaction was observed, which may indicate a dominant enantioselectivity of R-selective enzymes. So, controlled conditions in the bioreactor are crucial for the enantioselectivity and final good results. In case of flasks system applying, it is difficult to assure the constant level of aeration, which is crucial for the activity of enzymes involved in substrate oxy-functionalization, what can imply in creating the unfavourable conditions. As a consequence, the secondary metabolic pathways can be triggered on and the dropping in optical purity of the product can be observed with the activation of other enzymes groups.
Although both tested fungal strains (R. arrhizus and B. bassiana) are active towards the substrate, the reactions proceed with different enantioselectivity. In both cases, the main or the only product formed was 1-phenylethane-1,2-diol. The formation of this derivative may be the result of a unspecific peroxygenases (EC 1.11.2.1) secreted by some fungal cells [43], or multi-stage reaction proceeding according to the sequence of transformations: dehydrogenation (formation of a double bond), epoxidation (result of monooxygenase activity), epoxide hydrolysis and the formation of a diol [44]. It seems that in the case of the B. bassiana strain, the spatial structure of the active/binding site of the enzyme allows particular arrangement of 2-phenylethanol and/or the intermediate product of the multi-stage transformation reaction, that finally favours oxirane ring opening with R-diol formation. On the other hand, it is known from the literature data [43] that some fungi can produce peroxygenases capable of, through hydroxylation, epoxidation, dealkylation, heteroatom oxidation etc., functionalizing a wide spectrum of substrates including alkyl benzenes with a longer (C2-C6) side chain. For example, in the case of ethylbenzene and propylbenzene as the end reaction product, the corresponding R-1-phenylalcohol is formed with a high yield and an enantiomeric excess of above 99 % e.e. [45]. However, in the case of biotransformation involving the Rhizopus arrhizus strain, biocatalysis may rely on enzymes physiologically specific to substrates with expand steric hindrance (e.g. steroids), therefore the binding of the substrate or oxidized (epoxide) intermediate may proceed in a statistically random manner, hence the obtained product is racemic. Additionally, other groups of enzymes (e.g. monooxygenases, epoxide hydrolases of different enantioselectivity etc.) may be involved in the reaction, as evidenced by the presence of small amounts of tyrosol in the post-reaction mixture.
The next approaches to improve the results were focused on the other parameters strongly influencing the progress and selectivity of biotransformations, such as the deficiency of nutrients. This is a very well-known strategy introduced in the biotechnological processes relying on the activities of secondary pathways. However in presented studies such approach resulted in the activation of the pathways (Beauveria strains), which enabled the fast and effective degradation of the substrate. In the research, 2-phenylethanol was considered as xenobiotic, which can be converted by enzymatic systems involved in the secondary metabolism, switched on by starvation conditions. Obviously, environmental stress factors can be different as deficiency of carbon or nitrogen source, changes of temperature, light, and pH [46,47,48]. In case of discuss processes, the pre-incubation of the biocatalysts under starvation conditions was implemented, to force the bioconversion of the substrate [49,50]. However, studies conducted with R. arrhizus cells, showed that the starvation step did not affect the substrate bioconversion. In this case, the activity or synthesis of enzymes involved in substrate bioconversion is inhibited by stress conditions. An extremely different situation was observed in the case of Beauveria strains, where the use of pre-incubation under starvation conditions resulted in the activation of degradative pathways (data not shown) and the gradual utilization of 2-phenylethanol for cellular purposes within a maximum of 72 hours.
These conclusions were the base for the modifications of the form of the biocatalyst, which is another common method applied in biotransformations, as a solution allowed protecting the viable microbes. On the other hand, different immobilization methods may affect the mechanisms of transport through cell membranes and change the activity of enzymes associated with the biocatalyst cell envelope, which may lead to a change in selectivity. The selection of the appropriate immobilization method is a process that also depends on the type of growth of the microorganism tested and must be determined individually.
This approach was very effective in the case of R. arrhizus strain, since it leads to improve the bioconversion results. So, this fungus was immobilised on polyurethane foams. Effective overgrow of the support material by the fungal mycelium was tested for two types of foams (740-1040 µm and 1060-1600 µm porosity) and in both cases was observed. Biotransformation of 2-phenylethanol catalysed by immobilized R. arrhizus cells leads to the formation of the same mixture of products (tyrosol and mainly racemic 1-phenylethane-1,2-diol), but the amount of desired products (in 50 mL of medium) was much higher using biocatalyst immobilized on/in foams with 740-1040 µm porosity in comparison to the fresh mycelium. Formation of products (1-phenylethane-1,2-diol and tyrosol) under such conditions, just after the first day of biotransformation allowed to obtain a much larger amount of product mixture (1.55 mg/ 50 mL) than in the case of biotransformations carried out with the fresh biomass, the maximum was reached after three days of bioprocess (about 2 mg, of which 1-phenylethane-1,2-diol mainly formed was 1.85 mg and tyrosol 0.15 mg, Scheme 2). This susceptibility to immobilization of this fungus, can be explained by the characteristic of R. arrhizus grow. Usually it grows as a filamentous form, so it can overgrown the foams without any obstacles it resistant to negative effects of external factors can be increased [51,52,53]. Confirmation of his facts can be seen by noting, that the immobilized form of the biocatalyst allowed obtaining the comparable (4th day) or higher (3, 5th day) amount of products during the transformation of 30 mg of substrate comparing to the initial conditions with the fresh mycelium applied for transformation of 60 mg of substrate (Scheme 2- green and blue series).
Such immobilized mycelium, which overgrown the polyurethane foams was also subjected to further to scaling experimental with the simplified continuous flow bioreactor (Figure 2.). However, in this case the partial washing out of the catalyst from the polyurethane foams was observed, so the results were irrelevant for scaling approaches.
The same method of immobilization was also applied in case of the B. brongniartii DSM 6651 strain (B. bassiana mycelium did not match the pores of the foams and immobilization failed). The results obtained indicate that, both in the case of the biotransformation experiment carried out by fresh and immobilised in foams biomass, that the degradation of 2-phenylethanol is occurred. (Table 1). The only differences were noticed in the rate of degradation. Fresh biomass carried out this process faster (48 h), than immobilized ones on polyurethane foams, due to the mass exchange problems, so complete degradation was occurred within 72 h. It seems, that B. brongniartii consumes 2-phenylethanol as an energy and carbon source [54,55]. Degradative activity is also useful in industry. Pharmaceutical wastes disposal is today a challenge for environmental protection, especially in aquatic ecosystems. Therefore, fungi of such potential are significant in bioremediations [56].
As it was mentioned above, Beauveria (B. bassiana) mycelium was unable to efficiently overgrow the pore network of the foam, so alternative methods were implemented, such as entrapment in calcium alginate and in agar-agar matrix. Immobilization in agar-agar, resulted in a slightly higher resistance to the toxic effects of the substrate or products, which resulted in a higher amount of obtained product (0.43 mg in 50 mL) compared to the fresh mycelium of this strain (0.38 mg). However, immobilization had a negative impact on the enantioselectivity of the reaction and racemic 1-phenylethane-1,2-diol was formed. Increasing the amount of substrate to 60 mg (10 mM) implies in the lack of the bioconversion, probably, as a result of enhanced toxicity. The crucial factor lowering the reaction effectiveness in this case, seems to be the limitation in the contact between the mycelium and the substrate. Also, the encapsulation of the mycelium in the agar network can be a reason of partial deactivation of some enzymes (involved in bioconversion), especially those associated with the cell membrane, which in turn allowed manifesting the activity of enzymes of lower or opposite selectivity. Additionally, immobilization in agar-agar can affect the enantioselectivity by moderating the selectivity of transport across the cell envelope, which is in direct contact with the entrapped material [5]. In some studies the use of two different immobilization methods completely changed the enantioselectivity of the biocatalyst and, depending on the selected immobilization material (agar-agar or alginate), the S or R enantiomer of the resulted product was obtained [38].
The important prove of the blocking of the contact between the substrate and enzymatic system of fungal mycelium is the research on B. bassiana also entrapped in an alginate matrix. So, calcium alginate beads hindered the contact of the substrate with the biocatalyst and no biotransformation of 2-phenylethanol or its degradation was observed. The same results were obtained regardless the substrate concentration (5 mM or 10 mM). This type of carrier tightly seals the biocatalyst and can prevent the contact of fungal cells with the substrate, which results in a lack of biotransformation.
In our earlier studies, biotransformation of 2-phenylethanol on a semi-preparative scale with A. niger resulted in the optically pure (S)-1-phenylethane-1,2-diol synthesis [36]. In this studies, the R enantiomer of the same product was formed during the bioprocess with B. bassiana. These are meaningful results, since both optical isomers are formed after bioconversions starting from cheap, common reagent as phenylethanol.
Author Contributions
A.R., B.S-M., M.B-R, E.Ż-D. conceptualized the idea and methodology; A.R., M.B-R., E.Ż-D. wrote the original drafted manuscript; M.K-O., E.Ż-D., M.B-R. editing and revising the manuscript; E.Ż-D., M.B-R. supervision of experiments and writing. All authors have read and agreed to the published version of the manuscript
Funding
The work was supported in part by the project Minigrants for doctoral students of the Wroclaw University of Science and Technology, number 50SD/0068/24.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Choudhary, M.; Gupta, S.; Dhar, M.K.; Kaul, S. Endophytic Fungi-Mediated Biocatalysis and Biotransformations Paving the Way Toward Green Chemistry. Front Bioeng Biotechnol 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Schrewe, M.; Julsing, M.K.; Bühler, B.; Schmid, A. Whole-Cell Biocatalysis for Selective and Productive C–O Functional Group Introduction and Modification. Chem Soc Rev 2013, 42. [Google Scholar] [CrossRef] [PubMed]
- Madavi, T.B.; Chauhan, S.; Keshri, A.; Alavilli, H.; Choi, K.Y.; Pamidimarri, S.D.V.N. Whole-Cell Biocatalysis: Advancements toward the Biosynthesis of Fuels. Biofuels, Bioproducts and Biorefining.
- Schwarz, F.M.; Müller, V. Whole-Cell Biocatalysis for Hydrogen Storage and Syngas Conversion to Formate Using a Thermophilic Acetogen. Biotechnol Biofuels 2020, 13. [Google Scholar] [CrossRef]
- Raczyńska, A.; Jadczyk, J.; Brzezińska-Rodak, M. Altering the Stereoselectivity of Whole-Cell Biotransformations via the Physicochemical Parameters Impacting the Processes. Catalysts 2021, 11. [Google Scholar] [CrossRef]
- Grogan, G.J.; Holland, H.L. The Biocatalytic Reactions of Beauveria Spp. J Mol Catal B Enzym 2000, 9. [Google Scholar] [CrossRef]
- Kozłowska, E.; Urbaniak, M.; Hoc, N.; Grzeszczuk, J.; Dymarska, M.; Stępień, Ł.; Pląskowska, E.; Kostrzewa-Susłow, E.; Janeczko, T. Cascade Biotransformation of Dehydroepiandrosterone (DHEA) by Beauveria Species. Sci Rep 2018, 8. [Google Scholar] [CrossRef]
- Huszcza, E.; Dmochowska-Gładysz, J.; Bartmańska, A. Transformations of Steroids by Beauveria Bassiana. Zeitschrift fur Naturforschung - Section C Journal of Biosciences 2005, 60. [Google Scholar] [CrossRef]
- Kozłowska, E.; Matera, A.; Sycz, J.; Kancelista, A.; Kostrzewa-Susłow, E.; Janeczko, T. New 6,19-Oxidoandrostan Derivatives Obtained by Biotransformation in Environmental Filamentous Fungi Cultures. Microb Cell Fact 2020, 19. [Google Scholar] [CrossRef]
- Świzdor, A.; Kołek, T.; Panek, A.; Białońska, A. Microbial Baeyer-Villiger Oxidation of Steroidal Ketones Using Beauveria Bassiana: Presence of an 11α-Hydroxyl Group Essential to Generation of D-Homo Lactones. Biochim Biophys Acta Mol Cell Biol Lipids 2011, 1811. [Google Scholar] [CrossRef]
- Sordon, S.; Popłoński, J.; Tronina, T.; Huszcza, E. Regioselective O-Glycosylation of Flavonoids by Fungi Beauveria Bassiana, Absidia Coerulea and Absidia Glauca. Bioorg Chem 2019, 93. [Google Scholar] [CrossRef]
- Perz, M.; Krawczyk-Łebek, A.; Dymarska, M.; Janeczko, T.; Kostrzewa-Susłow, E. Biotransformation of Flavonoids with -NO2, -CH3 Groups and -Br, -Cl Atoms by Entomopathogenic Filamentous Fungi. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Łużny, M.; Tronina, T.; Kozłowska, E.; Dymarska, M.; Popłoński, J.; Łyczko, J.; Kostrzewa-Susłow, E.; Janeczko, T. Biotransformation of Methoxyflavones by Selected Entomopathogenic Filamentous Fungi. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Salvi, N.A.; Chattopadhyay, S. Studies on Rhizopus Arrhizus Mediated Enantioselective Reduction of Arylalkanones. Tetrahedron 2001, 57. [Google Scholar] [CrossRef]
- Salvi, N.A.; Patil, P.N.; Udupa, S.R.; Banerji, A. Biotransformations with Rhizopus Arrhizus: Preparation of the Enantiomers of 1-Phenylethanol and 1-(Fo-, m- and p-Methoxyphenyl)Ethanols. Tetrahedron Asymmetry 1995, 6. [Google Scholar] [CrossRef]
- Salokhe, P.R.; Salunkhe, R.S. Rhizopus Arrhizus Mediated SAR Studies in Chemoselective Biotransformation of Haloketones at Ambient Temperature. Biocatal Biotransformation 2022, 40. [Google Scholar] [CrossRef]
- Song, K.N.; Lu, Y.J.; Chu, C.J.; Wu, Y.N.; Huang, H.L.; Fan, B.Y.; Chen, G.T. Biotransformation of Betulonic Acid by the Fungus Rhizopus Arrhizus CGMCC 3.868 and Antineuroinflammatory Activity of the Biotransformation Products. J Nat Prod 2021, 84, 2664–2674. [Google Scholar] [CrossRef]
- Holland, H.L. Biotransformations of Δ4-3-Ketosteroids by the Fungus Rhizopus Arrhizus. Acc Chem Res 1984, 17. [Google Scholar] [CrossRef]
- Madureira, J.; Margaça, F.M.A.; Santos-Buelga, C.; Ferreira, I.C.F.R.; Verde, S.C.; Barros, L. Applications of Bioactive Compounds Extracted from Olive Industry Wastes: A Review. Compr Rev Food Sci Food Saf 2022, 21. [Google Scholar] [CrossRef]
- Heath, R.S.; Ruscoe, R.E.; Turner, N.J. The Beauty of Biocatalysis: Sustainable Synthesis of Ingredients in Cosmetics. Nat Prod Rep 2022, 39. [Google Scholar] [CrossRef]
- Brooks, S.J.; Doyle, E.M.; O’Connor, K.E. Tyrosol to Hydroxytyrosol Biotransformation by Immobilised Cell Extracts of Pseudomonas Putida F6. Enzyme Microb Technol 2006, 39. [Google Scholar] [CrossRef]
- Anissi, J.; Sendide, K.; Ouardaoui, A.; Benlemlih, M.; El Hassouni, M. Production of Hydroxytyrosol from Hydroxylation of Tyrosol by Rhodococcus Pyridinivorans 3HYL DSM109178. Biocatal Biotransformation 2021, 39. [Google Scholar] [CrossRef]
- Grech-Baran, M.; Sykłowska-Baranek, K.; Pietrosiuk, A. Biotechnological Approaches to Enhance Salidroside, Rosin and Its Derivatives Production in Selected Rhodiola Spp. in Vitro Cultures. in Vitro Cultures. Phytochemistry Reviews 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Han, T. Effects of Salidroside Pretreatment on Expression of Tumor Necrosis Factor-Alpha and Permeability of Blood Brain Barrier in Rat Model of Focal Cerebralischemia-Reperfusion Injury. Asian Pac J Trop Med 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Han, J.; Zhang, J.; Xiao, Q.; Hu, J.; Chen, L. Pharmacological Activities, Mechanisms of Action, and Safety of Salidroside in the Central Nervous System. Drug Des Devel Ther 2018, 12. [Google Scholar] [CrossRef]
- Lima, J.J.; Aguilar, A.; Sánchez, F.G.; Díaz, A.N. Enantiomeric Fraction of Styrene Glycol as a Biomarker of Occupational Risk Exposure to Styrene. Chemosphere 2017, 168. [Google Scholar] [CrossRef]
- Wang, S.; Xu, Y.; Zhang, R.; Zhang, B.; Xiao, R. Improvement of (R)-Carbonyl Reductase-Mediated Biosynthesis of (R)-1-Phenyl-1,2-Ethanediol by a Novel Dual-Cosubstrate-Coupled System for NADH Recycling. Process Biochemistry 2012, 47. [Google Scholar] [CrossRef]
- Nie, Y.; Xu, Y.; Mu, X.Q. Highly Enantioselective Conversion of Racemic 1-Phenyl-1,2-Ethanediol by Stereoinversion Involving a Novel Cofactor-Dependent Oxidoreduction System of Candida Parapsilosis CCTCC M203011. Org Process Res Dev 2004, 8. [Google Scholar] [CrossRef]
- Pandey, R.K.; Fernandes, R.A.; Kumar, P. An Asymmetric Dihydroxylation Route to Enantiomerically Pure Norfluoxetine and Fluoxetine. Tetrahedron Lett 2002, 43. [Google Scholar] [CrossRef]
- Cui, Z.M.; Zhang, J.D.; Fan, X.J.; Zheng, G.W.; Chang, H.H.; Wei, W.L. Highly Efficient Bioreduction of 2-Hydroxyacetophenone to (S)- and (R)-1-Phenyl-1,2-Ethanediol by Two Substrate Tolerance Carbonyl Reductases with Cofactor Regeneration. J Biotechnol 2017, 243. [Google Scholar] [CrossRef]
- Cao, L.; Lee, J.; Chen, W.; Wood, T.K. Enantioconvergent Production of (R)-1-Phenyl-1,2-Ethanediol from Styrene Oxide by Combining the Solanum Tuberosum and an Evolved Agrobacterium Radiobacter AD1 Epoxide Hydrolases. Biotechnol Bioeng 2006, 94. [Google Scholar] [CrossRef]
- Peng, F.; Su, H.H.; Ou, X.Y.; Ni, Z.F.; Zong, M.H.; Lou, W.Y. Immobilization of Cofactor Self-Sufficient Recombinant Escherichia Coli for Enantioselective Biosynthesis of (R)-1-Phenyl-1,2-Ethanediol. Front Bioeng Biotechnol 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Chen, Q.S.; Li, F.Z.; Ou, X.Y.; Zong, M.H.; Lou, W.Y. Using Deep Eutectic Solvents to Improve the Biocatalytic Reduction of 2-Hydroxyacetophenone to (R)-1-Phenyl-1,2-Ethanediol by Kurthia Gibsonii SC0312. Molecular Catalysis 2020, 484. [Google Scholar] [CrossRef]
- Li, Z.; Liu, W.; Chen, X.; Jia, S.; Wu, Q.; Zhu, D.; Ma, Y. Highly Enantioselective Double Reduction of Phenylglyoxal to (R)-1-Phenyl-1,2-Ethanediol by One NADPH-Dependent Yeast Carbonyl Reductase with a Broad Substrate Profile. Tetrahedron 2013, 69. [Google Scholar] [CrossRef]
- Żymańczyk-Duda, E.; Szmigiel-Merena, B.; Brzezińska-Rodak, M.; Klimek-Ochab, M. Natural Antioxidants–Properties and Possible Applications. Journal of Applied Biotechnology & Bioengineering 2018, 5. [Google Scholar] [CrossRef]
- Szmigiel-Merena, B.; Brzezińska-Rodak, M.; Klimek-Ochab, M.; Majewska, P.; Zymanćzyk-Duda, E. Half-Preparative Scale Synthesis of (S)-1-Phenylethane-1,2-Diol as a Result of 2-Phenylethanol Hydroxylation with Aspergillus Niger (IAFB 2301) Assistance. Symmetry (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Głąb, A.; Szmigiel-Merena, B.; Brzezińska-Rodak, M.; Żymańczyk-Duda, E. Biotransformation of 1- and 2-Phenylethanol to Products of High Value via Redox Reactions. Biotechnologia 2016, 97. [Google Scholar] [CrossRef]
- Lubiak-Kozłowska, K.; Brzezińska-Rodak, M.; Klimek-Ochab, M.; Olszewski, T.K.; Serafin-Lewańczuk, M.; Żymańczyk-Duda, E. (S)-Thienyl and (R)-Pirydyl Phosphonate Derivatives Synthesized by Stereoselective Resolution of Their Racemic Mixtures With Rhodotorula Mucilaginosa (DSM 70403) - Scaling Approaches. Front Chem 2020, 8. [Google Scholar] [CrossRef]
- Serafin-Lewańczuk, M.; Brzezińska-Rodak, M.; Lubiak-Kozłowska, K.; Majewska, P.; Klimek-Ochab, M.; Olszewski, T.K.; Żymańczyk-Duda, E. Phosphonates Enantiomers Receiving with Fungal Enzymatic Systems. Microb Cell Fact 2021, 20. [Google Scholar] [CrossRef]
- Lapponi, M.J.; Méndez, M.B.; Trelles, J.A.; Rivero, C.W. Cell Immobilization Strategies for Biotransformations. Curr Opin Green Sustain Chem 2022, 33. [Google Scholar] [CrossRef]
- Zingaro, K.A.; Nicolaou, S.A.; Papoutsakis, E.T. Dissecting the Assays to Assess Microbial Tolerance to Toxic Chemicals in Bioprocessing. Trends Biotechnol 2013, 31. [Google Scholar] [CrossRef]
- Nicolaou, S.A.; Gaida, S.M.; Papoutsakis, E.T. A Comparative View of Metabolite and Substrate Stress and Tolerance in Microbial Bioprocessing: From Biofuels and Chemicals, to Biocatalysis and Bioremediation. Metab Eng 2010, 12. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R. Oxidations Catalyzed by Fungal Peroxygenases. Curr Opin Chem Biol 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- VELASCO B., R.; GIL G., J. H.; GARCÍA P., C.M.; DURANGO R., D.L. PRODUCTION OF 2-PHENYLETHANOL IN THE BIOTRANSFORMATION OF CINNAMYL ALCOHOL BY THE PLANT PATHOGENIC FUNGUS <I>Colletotrichum Acutatum</I>. Vitae 2010, 17. [Google Scholar] [CrossRef]
- Kluge, M.; Ullrich, R.; Scheibner, K.; Hofrichter, M. Stereoselective Benzylic Hydroxylation of Alkylbenzenes and Epoxidation of Styrene Derivatives Catalyzed by the Peroxygenase of Agrocybe Aegerita. Green Chemistry 2012, 14. [Google Scholar] [CrossRef]
- Fox, E.M.; Howlett, B.J. Secondary Metabolism: Regulation and Role in Fungal Biology. Curr Opin Microbiol 2008, 11. [Google Scholar] [CrossRef]
- Yu, J.H.; Keller, N. Regulation of Secondary Metabolism in Filamentous Fungi. Annu Rev Phytopathol 2005, 43. [Google Scholar] [CrossRef]
- Medina, A.; Schmidt-Heydt, M.; Rodríguez, A.; Parra, R.; Geisen, R.; Magan, N. Impacts of Environmental Stress on Growth, Secondary Metabolite Biosynthetic Gene Clusters and Metabolite Production of Xerotolerant/Xerophilic Fungi. Curr Genet 2015, 61. [Google Scholar] [CrossRef]
- Kozyra, K.; Brzezińska-Rodak, M.; Klimek-Ochab, M.; Zymańczyk-Duda, E. Biocatalyzed Kinetic Resolution of Racemic Mixtures of Chiral α-Aminophosphonic Acids. J Mol Catal B Enzym 2013, 91. [Google Scholar] [CrossRef]
- Krohn, N.G.; Brown, N.A.; Colabardini, A.C.; Reis, T.; Savoldi, M.; Dinamarco, T.M.; Goldman, M.H.S.; Goldman, G.H. The Aspergillus Nidulans ATM Kinase Regulates Mitochondrial Function, Glucose Uptake and the Carbon Starvation Response. G3: Genes, Genomes, Genetics 2014, 4. [Google Scholar] [CrossRef]
- Faber, K. 2011.
- Chmiel, A. 1998.
- Bonin, S. Mikroorganizmy Immobilizowane. Agro Przemysł 2008, 8, 20–23. [Google Scholar]
- Hirano, J.I.; Miyamoto, K.; Ohta, H. Purification and Characterization of Aldehyde Dehydrogenase with a Broad Substrate Specificity Originated from 2-Phenylethanol-Assimilating Brevibacterium Sp. KU1309. Appl Microbiol Biotechnol 2007, 76. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Hirano, J.I.; Ohta, H. Efficient Oxidation of Alcohols by a 2-Phenylethanol-Degrading Brevibacterium Sp. Biotechnol Lett 2004, 26. [Google Scholar] [CrossRef] [PubMed]
- Arun, K.B.; Madhavan, A.; Tarafdar, A.; Sirohi, R.; Anoopkumar, A.N.; Kuriakose, L.L.; Awasthi, M.K.; Binod, P.; Varjani, S.; Sindhu, R. Filamentous Fungi for Pharmaceutical Compounds Degradation in the Environment: A Sustainable Approach. Environ Technol Innov 2023, 31. [Google Scholar] [CrossRef]
Figure 1.
2-Phenylethanol.
Scheme 1.
Biotransformations of 2-phenylethanol carried out by Beauveria bassiana DSM 1344 and Rhizopus arrhizus DSM 1185.
Scheme 1.
Biotransformations of 2-phenylethanol carried out by Beauveria bassiana DSM 1344 and Rhizopus arrhizus DSM 1185.
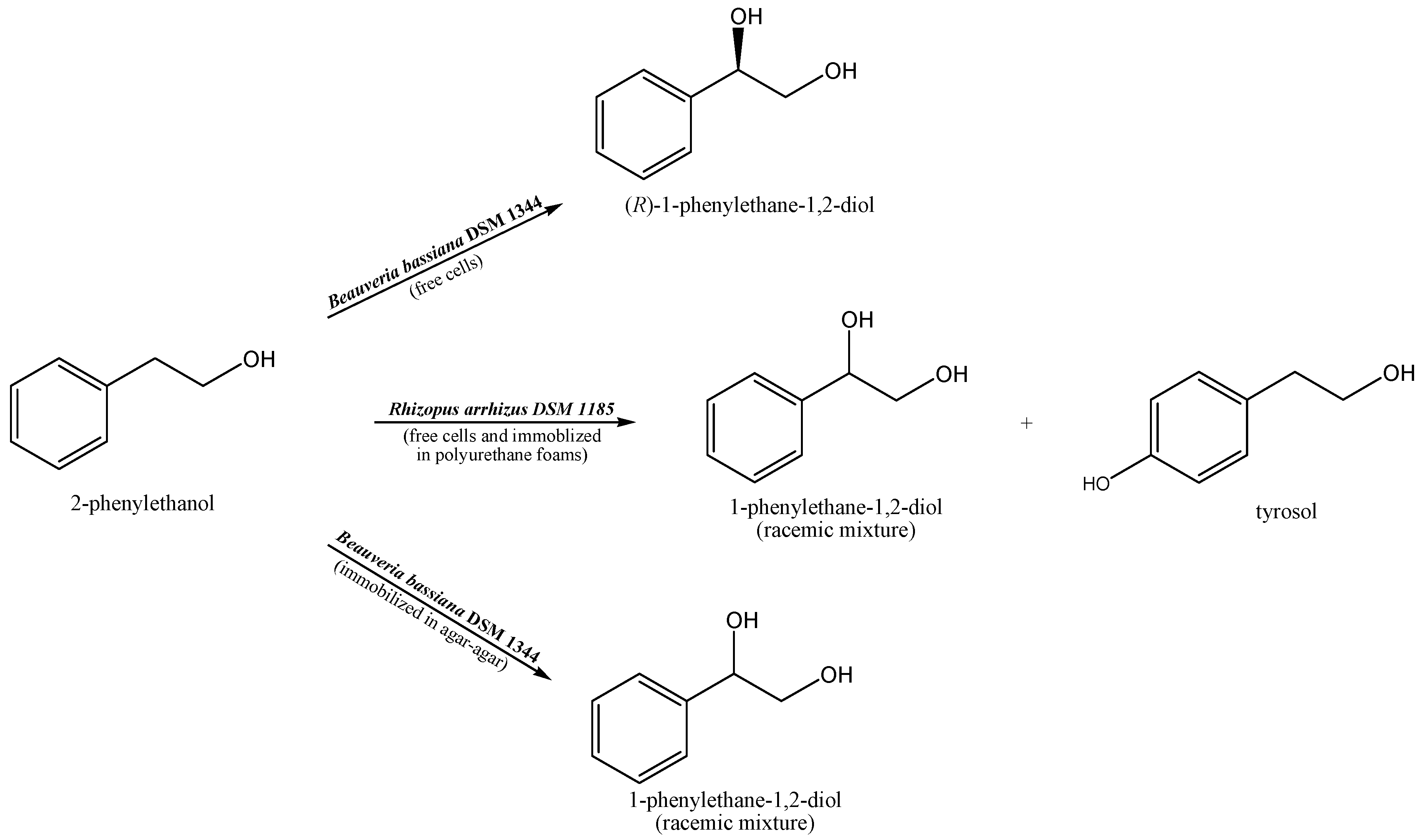
Figure 2.
General work scheme in semi-preparative experiment – simplified flow bioreactor model.
Scheme 2.
The amount of the mixture of products received after biotransformation of 2-phenylethanol with Rhizopus arrhizus DSM 1185 presented as function of substrate concentration (mg/50 mL) and form of biocatalyst.
Scheme 2.
The amount of the mixture of products received after biotransformation of 2-phenylethanol with Rhizopus arrhizus DSM 1185 presented as function of substrate concentration (mg/50 mL) and form of biocatalyst.
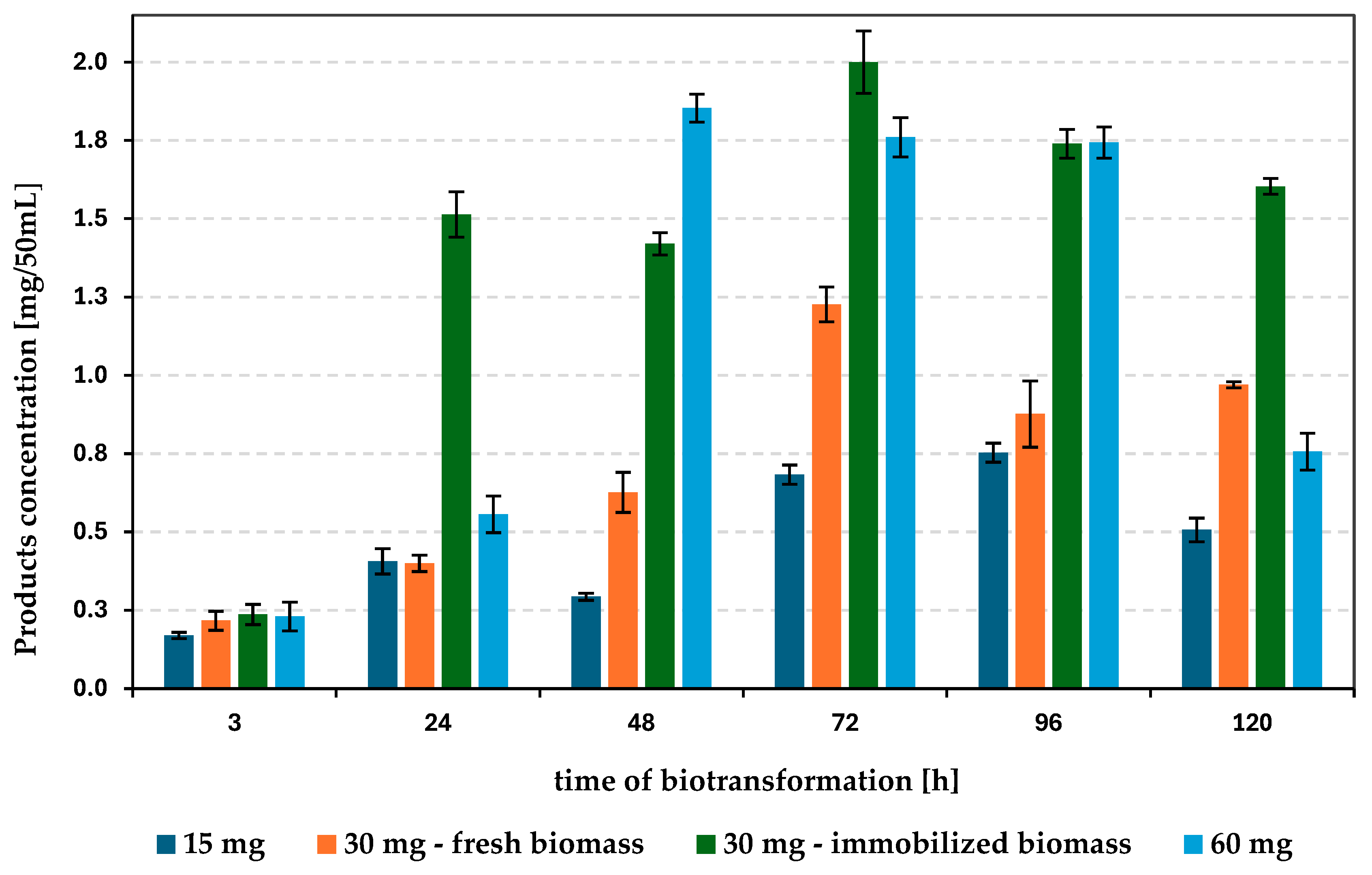

Table 1.
Biotransformation products depending on the selected strain and form of biocatalyst.
| Product |
(R)-1-phenylethane-1,2-diol | (R,S)-1-Phenylethane-1,2-diol | Tyrosol | |
|---|---|---|---|---|
| Biocatalyst | ||||
|
B. bassiana DSM 1344 |
fresh biomass in flasks | immobilized biomass (agar-agar) |
||
| fresh biomass in bioreactor | ||||
|
R. arrhizus DSM 1185 |
fresh biomass | fresh biomass | ||
| immobilized biomass (polyurethan foams) |
immobilized biomass (polyurethan foams) |
|||
|
B. brongniartii DSM 6651 |
2-phenylethanol degradation - no desired product formation | |||
Table 2.
Amount of (R)-1-fenyloetan-1,2-diol [mg] produced during the biotransformation of 2-phenylethanol (600 mg) in a batch bioreactor (600 mL medium).
Table 2.
Amount of (R)-1-fenyloetan-1,2-diol [mg] produced during the biotransformation of 2-phenylethanol (600 mg) in a batch bioreactor (600 mL medium).
| (R)-1-phenylethane-1,2-diol (e.e. 99.9%) [mg / 600 mL] | ||
|---|---|---|
| Strain | Biotransformation Time [Days] | Product [mg] |
| B. bassiana (fresh cells) | 1 | 12.9 (±0.095) |
| 2 | 17.6 (±0.110) | |
| 3 | 28.8 (±0.078) | |
| 4 | 10.0 (±0.130) | |
| 5 | 5.1(±0.075) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



