Submitted:
26 November 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
Aims Breast cancer is a heterogeneous disease and, according to Breast Cancer Statistics (CDC), ranks as the second leading cause of death among women. The intratumoral microbiota, a subject of recent research attention, plays a crucial role in the tumor microenvironment, contributing significantly to its repair and regeneration and serving as an effective therapeutic factor in cancers. Additionally, non-ionizing magnetic fields with extremely low frequencies (ELF-EFM) induce apoptosis, leading to the suppression of tumor cells. Studies have shown that ELF-EFM also plays a vital role in reducing bacterial cell counts. Therefore, by influencing the intratumoral microbiota population through ELF-EFM exposure, we can potentially address cancer and infectious diseases. Materials and methods ELF-EFM exposure for two hours per day with a frequency of 1Hz and an intensity of 100mT was investigated in triple-negative breast cancer (TNBC) mouse models and triple-positive breast cancer (TPBC) mouse models. After 28 days, the mice were euthanized, and the intratumoral microbiota population was examined using IHC tests (LPS/LTA) and real-time PCR (16SrRNA). Additionally, the mice's immune response was assessed by examining factors affecting tumor growth and metastasis: IHC (CD4/CD8, VEGF) and H&E (TILs). Key findings Significantly, TNBC mice in the exposure group did not show a significant change in tumor growth rate compared to the control group. However, TPBC mice in the exposure group exhibited a reduction in tumor growth rate compared to the control group. The number of Tumor-Infiltrating Lymphocytes (TILs) was lower in the TNBC exposure group compared to the TPBC group. In the intratumoral microbiota population, the number of Gram-negative bacteria, based on IHC test results (LPS/LTA) and examination of the expression of the 16SrRNA gene, was lower in TNBC mice compared to the TPBC group. Significance Our study results confirm the impact of ELF-EFM exposure on reducing the intratumoral microbiota population and inducing apoptosis, both of which are considered tumor cell suppressors.
Keywords:
ELF-MF
; anti-microbial
; TNBC
; TPBC
1. Introduction
The human microbiome, which is likely a vital factor in the initiation, progression, and prognosis of various cancer types, is rapidly expanding [1]. The collective genomes and by-products of all microorganisms living in the human body are referred to as the human microbiome, including bacteria, viruses, fungi, and more[2]. There exists a dynamic and complex relationship between the human host and the microbial composition of the human body (microbiota), and the interaction between the human microbiome and cancer is referred to as "oncobiome" [3]. Additionally, the human host can influence the microbiota and their mechanisms. These microbiota, present in various parts of the human body, including the mouth, skin, gut, and others, can impact food digestion, boost the immune system, and may even affect your mood and behavior [4]. In recent years, the microbiota has been found in various cancer tissues, including breast cancer, lung cancer, colorectal cancer, and prostate cancer. Nejman and colleagues conducted a study involving 1,526 tumors from seven different types of cancer and their adjacent normal tissues, and they found that each tumor type has a unique microbiome. This includes breast cancer, which is characterized by a diverse and rich microbiome. The breast cancer tissue contains its distinct microbiota [5].This includes breast cancer, which is characterized by a diverse and rich microbiome. The breast cancer tissue contains its distinct microbiota. Recent preclinical data suggests that changes in the breast microbiota may contribute to the initiation and progression of breast cancer.
Breast cancer (BC) is a heterogeneous disease arising from genetic and epigenetic factors in breast tissue. According to GLOBOCAN 2020, an estimated 2.3 million new cases of breast cancer occurred worldwide[6]. Triple-negative breast cancer (TNBC), constituting 10-20% of BC cases, is an aggressive subtype characterized by the lack of certain cell surface receptors, making current therapies less effective, and leading to metastasis or chemotherapy resistance [7]. Triple-positive breast cancer (TPBC), expressing higher than normal levels of all three cell surface receptors, responds to hormone and HER2-targeted therapies despite its high prevalence [8]. Treatment choices, including surgery, immunotherapy, chemotherapy, radiation, and hormone therapy, are tailored based on BC type, stage, and patient conditions [6,9,10] . As current treatments for BC patients are not universally effective, research is exploring potential efficacy through the tumor microbiome and non-ionizing electromagnetic fields with very low frequencies.
In recent years, non-ionizing, non-thermal extremely low-frequency electromagnetic fields (ELF-EMF) have been studied. Researchers have found that ELF-EMF can induce apoptosis in cancer cells through ROS oscillation and death signal communication. The effects of ELF-EMF depend on factors such as the type of cancerous/bacterial cell, exposure duration, field intensity, and frequency [11,12]. Due to the indiscriminate use of antibiotics, antibiotic resistance has become a challenging issue in infectious diseases. Researchers propose that exposing ELF-EMF, along with using bioelectrically effective antibiotics, could enhance the treatment of infectious diseases [13]. Additionally, it is hypothesized that utilizing the anti-tumor and antibacterial effects of ELF-EMF could be employed in cancer treatment by reducing or eliminating intratumoral microbiota.
Bayir and colleagues conducted studies revealing that the dual exposure of S. aureus and E. coli bacteria to a low frequency with a field intensity of 4 mT is affected. When the field intensity changes to 2 mT, S. aureus is inhibited, while E. coli requires a higher field intensity. Their studies suggest that ELF-EMF has both positive and negative effects on the cellular growth and viability of bacteria, as well as their antibiotic sensitivities, depending on factors such as exposure duration, frequency, field density, and bacterial cell type. Moreover, ELF-EMF acts as a stressor on lactobacilli, reducing their colony-forming units (CFU) [11].
Fojt and colleagues investigated the impact of ELF-EMF on positive/negative gram bacteria and found that gram-negative bacteria are more affected than gram-positive bacteria under conditions of 10 mT, 50 Hz, and 24 minutes [14].
Segator and colleagues reported that E.coli and P.aeruginosa strains, incubated with sub-inhibitory concentrations, experienced a reduction in cell count after exposure to ELF-EMF for 4, 6, and 8 hours, while the percentage of cells increased during the 24-hour exposure. However, no significant differences were observed in antibiotic sensitivity and growth rate between the test and control groups [15].
Given the significance of the intratumoral microbiota in cancer progression and treatment, we aim to investigate the anticancer and antibacterial effects of ELF-MF on the intratumoral microbiota in TNBC and TNBC mouse models. This research will pave the way for future clinical trials to assess the effectiveness of ELF-MF in the treatment of TNBC patients, following the completion of studies and experiments.
2. Materials and Methods
2.1. Cell Line
Cell lines of breast cancer, 4T1, and MC4L2, were obtained from the Iranian Biological Resource Center and cultured in RPMI1640 and h-DMEM/DMEM/F1 media with 10% FBS, 1% penicillin, and streptomycin at 37℃ in 5% CO2. When cell Confluence reached 80-90%, the cells were counted and prepared for the subsequent experiments.
2.2. Animals
Adult female BALB/C mice (6-7 weeks old) were purchased from the Pasteur Institute of Iran and randomly assigned to groups of 4-5 in each cage. The mice were housed under standard temperature, humidity, and regular light/dark cycles of 12 hours. To induce breast tumors, 1 million cells in 100μL PBS buffer were subcutaneously injected into the left mammary fat pad. All animal procedures were conducted by approved protocols by the Ethics Committee of the Cancer Institute (ACER) in Iran.
2.3. Measurement of Tumor Size And Body Weight
Daily, the body weight and tumor size were measured using a digital scale and manual calipers, respectively. The tumor volume was calculated using the formula:
Tumor size (mm3) =1/2*length*(with) 2
When the tumor size reached 120-200 mm3, the mice were prepared for exposure to ELF. Throughout the exposure period, the tumor size and body weight of the mice were regularly measured.
2.4. ELF Exposure
The ELF electromagnetic device consists of two pairs of square-shaped wire coils at the two ends of a U-shaped iron core within a power transformer. It features a waveform generator connected to copper wire coils. This device provides magnetic fields with intensities ranging from 1 to 100 mT and frequencies from 1 to 60 Hz in the calibrated state. In the experimental setup, TNBC and TPBC mouse models, housed in square-shaped enclosures in groups of 4-5, are exposed to ELF with an intensity of 100 mT and a frequency of 1 Hz for 2 hours daily over 28 days. Control groups undergo the same conditions but without exposure to ELF. During ELF exposure, the mouse models are maintained at optimal temperatures with access to standard nutrition.
2.6 Hematoxylin and Eosin (H&E) Staining
After the tumors and organs underwent tissue processing, 5 μm tissue sections were prepared using a microtome, followed by deparaffinization with xylene to clear the tissue. Subsequently, dehydration was performed, and finally, the sections were stained with hematoxylin and eosin in sequence. The resulting images were obtained using an Olympus light microscope with magnifications of 10X and 40X [16,17].
2.7. Gram staining
After undergoing the stages of fixation, dehydration, embedding, and sectioning, the tumors were stained with Giemsa dye, and images were obtained using an Olympus light microscope with magnifications of 10X and 40X [18] .
2.8. Immunohistochemistry (IHC)
After deparaffinization of tumors in xylene and dehydration with varying alcohol concentrations, a set-up protocol was employed to remove formalin during two stages of antigen retrieval and neutralization of endogenous peroxidases. Throughout each step, washing was performed using DDW/PBS. Tumor sections were incubated with primary antibodies against proteins, including Anti-LPS, Anti-LTA, Anti-CD4, Anti-CD8, and Anti-VEGF, overnight at 4℃. Subsequently, they were washed three times with PBS and incubated for 1 hour at RT with secondary antibodies. Finally, images were captured using an Olympus light microscope with 10X and 40X magnifications [19].
...2.9Real-Time PCR
The frozen tissues were initially homogenized into small pieces using a sterile mortar and pestle and then thoroughly homogenized in 1 ml Rnx Plus (SinaClon BioScience). Total RNA was extracted using a pre-determined protocol. The concentration of total RNA was determined using a NanoDrop device. cDNA was synthesized following the manufacturer's protocol using a kit. Primers for the 16srRNA gene were designed for gene amplification. Real-time PCR was performed using Hot-Start SYBR Green qPCR Master Mix (Amplicon). DNA was denatured at 95°C for 7 seconds, followed by an annealing step at 56°C for 20 seconds and an extension step at 72°C for 30 seconds. The relative expression of the 16srRNA gene was calculated using the formula: change =2- [20,21].

Table 1.
Primers used for PCR validation of 16srRNA.:
| Micro-organism | Primers | Sequence (5′-3′) | Annealing temp and time | Extension temp and time | Amplicon size (bp) | Detail |
|---|---|---|---|---|---|---|
| E.coli | 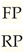 |
56°C for 20 s | 72°C for 30 s | 40 | Self-designed |
2.10. Statistical Analysis
All data were analyzed using GraphPad Prism software (Version 8) with t-Student and ANOVA tests. A significance level of p < 0.05 was considered statistically significant.
3. Results
3.1. Tumor Size
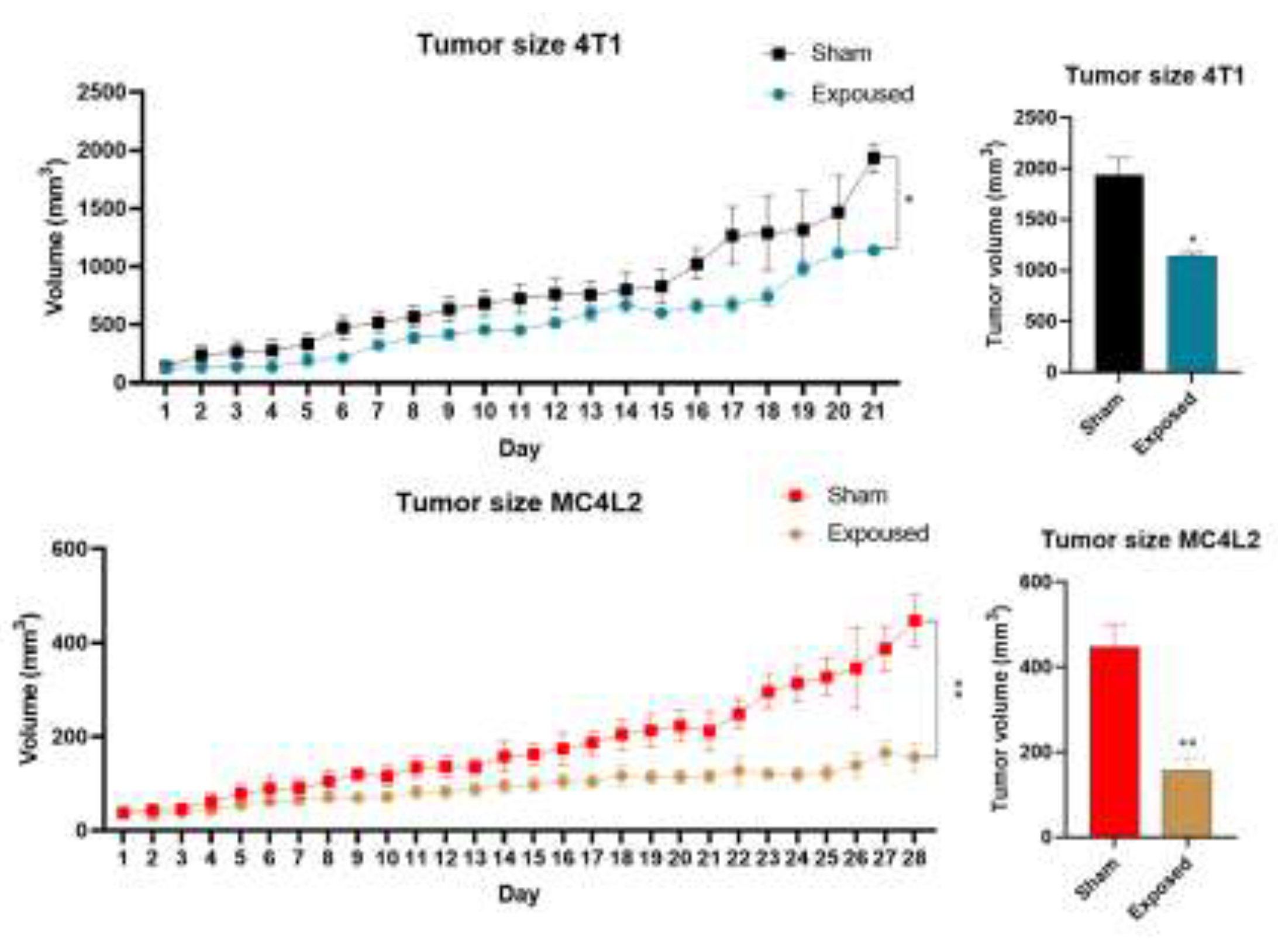
As depicted in Figure 1, tumor size changes in the experimental group, after exposure to ELF-EMF for two hours per day, significantly reduced tumor growth rate (final volume) compared to the control group in TPBC mouse models from day 16 onwards. However, in TNBC mouse models, no significant difference in tumor growth rate was observed after exposure to ELF-EMF.
3.2. H&E Staining Results on Vital Organs
The results from H&E staining in TNBC mice, in both the exposure and control groups exposed to ELF-EMF for two hours per day, did not show a significant change in the number of tumor-infiltrating lymphocytes (TILs). However, in TPBC mice exposed to ELF-EMF under the same conditions, the number of TILs increased.
- The H&E results: The control group of TNBC mice showed high nuclear polymorphism and mitotic figures. However, there was no significant difference in the number of TILs between both the exposure and control groups. Therefore, there was no observed response to the treatment. Nevertheless, in the exposure group of TPBC mice, a reduction in mitotic figures was observed, indicating that exposure to ELF-EMF for two hours per day resulted in decreased tumor growth and a treatment response.
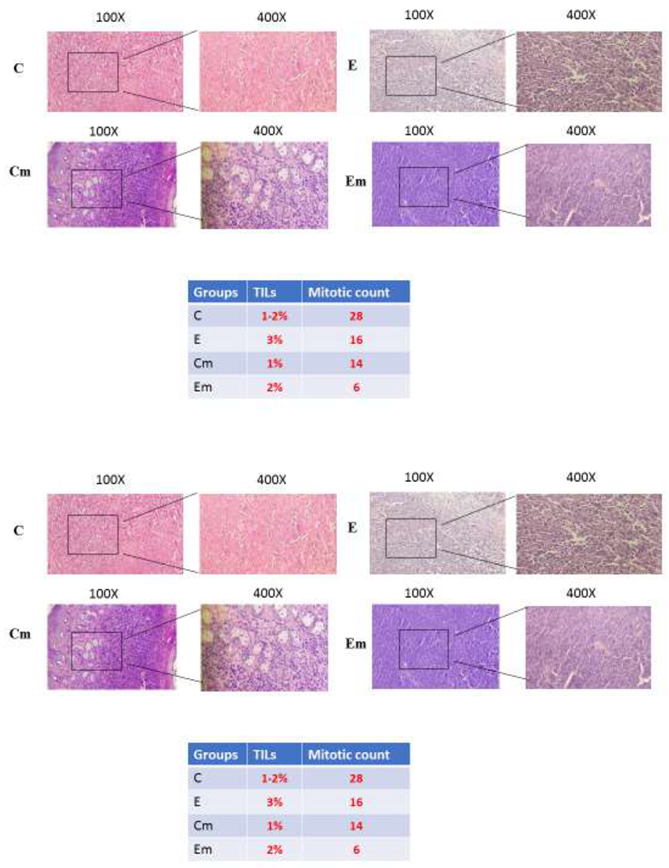
3.3. Gram Staining Results on Tumors
The population of intratumoral microbiota in the exposure and control groups of TNBC and TPBC mice was examined using Gram staining. The presence of intratumoral microbiota was confirmed in the mouse models, and it was observed that gram-negative bacteria were more prevalent extracellularly. However, no significant difference in the population of intratumoral microbiota was observed between the exposure and control groups of TNBC and TPBC mice.
- The presence of rod-shaped bacterial entities with distinct nuclei around the tumor cells of TNBC and TPBC mice is observable. However, no significant change in the population of the microbiota was observed in both exposure and control groups in both mouse models.
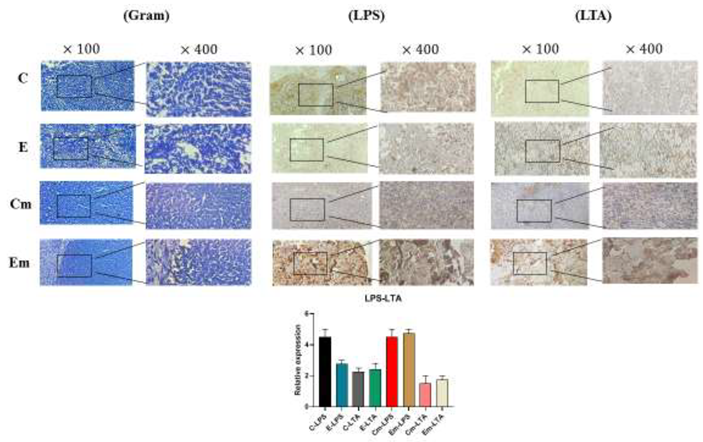
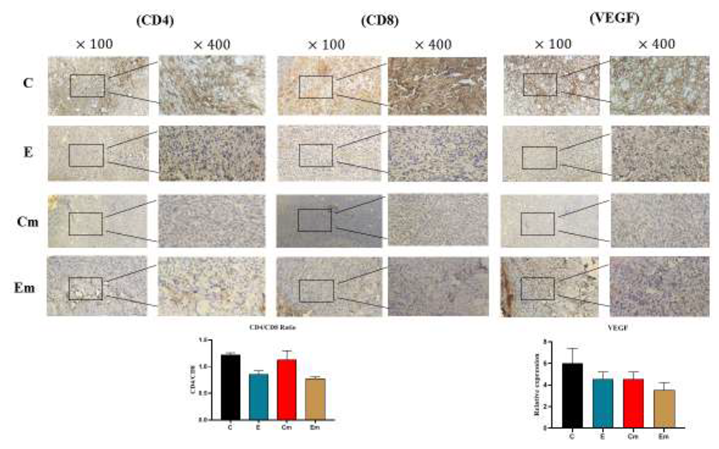
3.4. IHC Staining Results on Tumors
Immunohistochemical staining was performed to investigate the expression of cell surface biomarkers (LPS/LTA, CD4/CD8, VEGF) in TNBC and TPBC mouse models. No significant differences were observed in the CD4/CD8 ratios in the control and exposure groups of both TNBC and TPBC mouse models. On the other hand, the control groups in both mouse models showed high expression of VEGF protein, indicating angiogenesis in these groups. Furthermore, to explore the more specific presence of gram-positive/negative bacteria in the tumor microbiota, the expression of LPS/LTA proteins was examined. The level of LPS expression was higher in the exposure group, suggesting a higher presence of gram-negative bacteria in the tumor microbiota of both TNBC and TPBC mice.
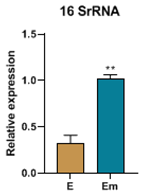
3.5. Real-time PCR results on tumors
In this study, the expression of the 16SrRNA gene, a marker for bacterial presence, was examined. The findings revealed that exposure to ELF-EMF led to a decrease in the expression of this gene in TNBC mice and an increase in its expression in TPBC mice.
4. Discussion
According to the global statistics provided by GLOBOCAN, breast cancer (BC) stands out as the most prevalent malignancy worldwide. Triple-negative breast cancer (TNBC), characterized by the absence of cell surface receptors, and high-prevalence triple-positive breast cancer (TPBC) command special attention. The evolving landscape of knowledge in Intratumoral Microbiota (ITM) has unveiled its pivotal role as a determinant in cancer progression and development, particularly in the metastatic setting, immune surveillance inhibition, and compromised therapeutic efficacy [7,22,23,24,25]. Researchers led by Jine et al. elucidated the profound impact of ITM on heightened tumor cell proliferation through the orchestration of chronic inflammation and intrinsic immune suppression [26]. The distinctive presence of microbiota in various tissue milieus was underscored by Nejman and associates [5]. Additionally, investigations by Geller et al. corroborated the influence of ITM on chemotherapeutic resistance [27]. Cancer Atlas reports that 15-20% of global cancer incidence can be ascribed to infectious etiologies. The indiscriminate use of antibiotics has engendered a concerning rise in antibiotic resistance, presenting a contemporary challenge in the management of infectious maladies. A promising therapeutic alternative lies in the strategic application of Extremely Low-Frequency Electromagnetic Fields (ELF-EMF), whose documented anti-cancer properties have come to the fore in recent research endeavors. In the course of these investigations, researchers delineated that ELF-EMF exposure precipitates apoptosis via the intricate Oscillation ROS mechanism. Notably, Moori et al. demonstrated that breast cancer cells subjected to (100mT) ELF-EMF experienced alterations in the expression profile of Bax and Bcl2, resulting in heightened Bax/Bcl2 ratios conducive to apoptotic induction. The ensuing apoptotic cascade in breast cancer cells underscores the potential therapeutic efficacy of ELF-EMF in the clinical management of breast cancer patients [28]. Furthermore, Aoshu et al. communicated that the exposure of breast cancer cells to four distinct frequencies of ELF-EMF, with particular emphasis on the 200Hz frequency, yielded superior outcomes by selectively suppressing the viability of breast cancer cells while exerting minimal impact on normal cellular counterparts [29]. An additional beneficial facet of ELF-EMF lies in its demonstrated antibacterial properties, rendering it a promising modality for the treatment of infectious maladies and cancers arising from infectious causation. Noteworthy studies have unveiled its modulating effect on the viability of both Gram-positive and Gram-negative bacterial strains, with a pronounced efficacy against the latter. Concurrent exposure to ELF-EMF and subinhibitory concentrations manifested a reduction in bacterial cell populations, with no discernible compromise in antibiotic susceptibility and growth kinetics observed in both Gram-positive and Gram-negative bacterial cohorts [14,15]. In the context of this study, which takes into consideration the nuanced conditions of patients afflicted with TNBC, and TPBC, and the extant challenges in contemporary therapeutic approaches, the investigation of ELF-EMF on the backdrop of ITM application was undertaken to elucidate its impact on breast cancer progression under in vivo conditions. This inquiry provides invaluable insights aimed at delineating efficacious therapeutic interventions for the advancement of breast cancer management.
Regarding the tumor size, we observed that TPBC mice exposed to ELF-EMF for two hours daily exhibited a reduction in tumor size compared to the control group. This decrease in tumor growth rate is attributed to the induction of apoptosis in tumor cells, tumor necrosis, and the prevention of tumor tissue vascularization. As reported in studies by Tatarov et al., ELF-EMF induces apoptosis, leading to the demise of tumor cells, as confirmed by the TUNEL assay [30]. Furthermore, H&E staining results also indicate an increase in the number of Tumor-Infiltrating Lymphocytes (TILs) in the ELF-EMF-exposed group, reflecting enhanced immune response against tumor cells and impeding tumor cell proliferation [31,32]. Moreover, the results obtained from immunohistochemistry (IHC) revealed a reduction in the CD4/CD8 ratio at the tumor site, indicating an enhancement in cellular immunity in mice exposed to ELF-EMF. As reported in previous studies, the decrease in cytotoxic CD8 lymphocytes (CTLs) favors acquired immunity, while the reduction in CD4 lymphocytes also plays a crucial role in the suppression of tumor cells[33,34,35]. Additionally, we observed a decrease in the expression of Vascular Endothelial Growth Factor (VEGF) protein in mice exposed to ELF-EMF, which is a significant factor in tumor angiogenesis [36,37]. All results suggest immune enhancement in the ELF-EMF-exposed groups and the suppression of tumor cells.
In our study, we investigated the impact of ELF-EMF exposure on the intratumoral microbiota population by examining the expression level of the 16SrRNA gene. We observed an increase in the expression of this gene in TPBC mice in the exposure group and a decrease in its expression in TNBC mice in the exposure group [19,38,39]. Additionally, to further assess the specific presence of positive/negative bacteria, we utilized the LPS/LTA test (IHC), indicating a higher presence of Gram-negative bacteria in both TNBC and TPBC exposure groups [40,41].
-
*Banerjee et al. used a microarray-based approach called the “PathoChip” to determine the microbial signature for different breast cancer subtypes [42].Banerjee et al. [42] identified the unique microbial signatures linked with triple-negative breast cancer.Polyoma viruses, herpesviruses, papilloma viruses, poxviruses, Arcanobacterium haemolyticum, Prevotella nigrescens, Pleistophora mulleris, Piedraia hortae, Trichuris trichura, Leishmania in triple-negative breast tissues are much more frequently than normal tissues* plasmodium-derived molecules have been associated with antitumor properties in adult mice in vivo and in vitro, and may be used as targets for tumor immunotherapy in triple-negative breast cancer [43] .
مقالهTumor-related Microbiome in the Breast Microenvironment and Breast Cancer
- The results of the study suggest that ELF-EMF exposure can be employed to reduce the intratumoral microbiota population in breast cancer patients. This approach could serve as an alternative treatment for infectious diseases or patients with TNBC characterized by the absence of cell surface receptors. In this study, we were limited in utilizing additional tests to examine the intratumoral microbiota population. Furthermore, the findings from this study can be extrapolated to human cancers along with in vivo studies.
5. Conclusions
Exposure to ELF-EMF against breast cancer may possess anticancer properties due to the induction of apoptosis, tumor necrosis, and angiogenesis suppression at the tumor site. Furthermore, it leads to a reduction in the intratumoral microbiota within the tumor, indicating its antibacterial properties. The current study can be extended to various types of intratumoral microbiota in different cancer types. Additionally, the impact of ELF-EMF exposure on both Gram-negative and Gram-positive bacteria in other cancers was investigated. The results of this study may have potential utility in alternative treatments for infectious and cancerous diseases.
References
- Knippel, R.J., J.L. Drewes, and C.L. Sears, The cancer microbiome: recent highlights and knowledge gaps. Cancer discovery, 2021. 11(10): p. 2378-2395. [CrossRef]
- Oliva, M., et al., Tumor-associated microbiome: where do we stand? International Journal of Molecular Sciences, 2021. 22(3): p. 1446.
- Thomas, R.M. and C. Jobin, The microbiome and cancer: is the ‘oncobiome’mirage real? Trends in cancer, 2015. 1(1): p. 24-35.
- Belkaid, Y. and S. Naik, Compartmentalized and systemic control of tissue immunity by commensals. Nature immunology, 2013. 14(7): p. 646-653. [CrossRef]
- Nejman, D., et al., The human tumor microbiome is composed of tumor type–specific intracellular bacteria. Science, 2020. 368(6494): p. 973-980. [CrossRef]
- Jayachandran, P., et al., Breast cancer and neurotransmitters: emerging insights on mechanisms and therapeutic directions. Oncogene, 2023. 42(9): p. 627-637. [CrossRef]
- Loizides, S. and A. Constantinidou, Triple negative breast cancer: Immunogenicity, tumor microenvironment, and immunotherapy. Frontiers in Genetics, 2023. 13: p. 1095839. [CrossRef]
- Schedin, T.B., V.F. Borges, and E. Shagisultanova, Overcoming therapeutic resistance of triple positive breast cancer with CDK4/6 inhibition. International journal of breast cancer, 2018. 2018.
- Thompson, K.J., et al., A comprehensive analysis of breast cancer microbiota and host gene expression. PloS one, 2017. 12(11): p. e0188873.
- Smrekar, K., A. Belyakov, and K. Jin, Crosstalk between triple negative breast cancer and microenvironment. Oncotarget, 2023. 14: p. 284. [CrossRef]
- Bayır, E., et al., The effects of different intensities, frequencies and exposure times of extremely low-frequency electromagnetic fields on the growth of Staphylococcus aureus and Escherichia coli O157: H7. Electromagnetic biology and medicine, 2015. 34(1): p. 14-18.
- Barati, M., et al., Cellular stress response to extremely low-frequency electromagnetic fields (ELF-EMF): An explanation for controversial effects of ELF-EMF on apoptosis. Cell Proliferation, 2021. 54(12): p. e13154. [CrossRef]
- Gualdi, G., et al., Wound repair and extremely low frequency-electromagnetic field: insight from in vitro study and potential clinical application. International Journal of Molecular Sciences, 2021. 22(9): p. 5037. [CrossRef]
- Fojt, L., et al., Comparison of the low-frequency magnetic field effects on bacteria Escherichia coli, Leclercia adecarboxylata and Staphylococcus aureus. Bioelectrochemistry, 2004. 63(1-2): p. 337-341. [CrossRef]
- Segatore, B., et al., Evaluations of the effects of extremely low-frequency electromagnetic fields on growth and antibiotic susceptibility of Escherichia coli and Pseudomonas aeruginosa. International Journal of Microbiology, 2012. 2012.
- Cahill, L.C., et al., Rapid virtual hematoxylin and eosin histology of breast tissue specimens using a compact fluorescence nonlinear microscope. Laboratory investigation, 2018. 98(1): p. 150-160. [CrossRef]
- Hong, S.D., et al., Hematoxylin and eosin staining for detecting biofilms: practical and cost-effective methods for predicting worse outcomes after endoscopic sinus surgery. Clinical and Experimental Otorhinolaryngology, 2014. 7(3): p. 193-197. [CrossRef]
- Becerra, S.C., et al., An optimized staining technique for the detection of Gram positive and Gram negative bacteria within tissue. BMC research notes, 2016. 9(1): p. 1-10. [CrossRef]
- Heymann, C.J., et al., The intratumoral microbiome: characterization methods and functional impact. Cancer letters, 2021. 522: p. 63-79. [CrossRef]
- Eisenhofer, R., et al., Contamination in low microbial biomass microbiome studies: issues and recommendations. Trends in microbiology, 2019. 27(2): p. 105-117. [CrossRef]
- Elie, C., et al., Comparison of DNA extraction methods for 16S rRNA gene sequencing in the analysis of the human gut microbiome. Scientific Reports, 2023. 13(1): p. 10279. [CrossRef]
- Yang, L., et al., Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy. Signal Transduction and Targeted Therapy, 2023. 8(1): p. 35. [CrossRef]
- Fu, A., et al., Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell, 2022. 185(8): p. 1356-1372. e26. [CrossRef]
- Gluz, O., et al., Triple-negative breast cancer—current status and future directions. Annals of Oncology, 2009. 20(12): p. 1913-1927. [CrossRef]
- Kumar, H., et al., A review of biological targets and therapeutic approaches in the management of triple-negative breast cancer. Journal of Advanced Research, 2023. [CrossRef]
- Jin, C., et al., Commensal microbiota promote lung cancer development via γδ T cells. Cell, 2019. 176(5): p. 998-1013. e16.
- Geller, L.T. and R. Straussman, Intratumoral bacteria may elicit chemoresistance by metabolizing anticancer agents. Molecular & Cellular Oncology, 2018. 5(1): p. e1405139. [CrossRef]
- Moori, M., et al., Effects of 1Hz 100mT electromagnetic field on apoptosis induction and Bax/Bcl-2 expression ratio in breast cancer cells. Multidisciplinary Cancer Investigation, 2022. 6(1): p. 1-7. [CrossRef]
- Xu, A., Q. Wang, and T. Lin, Low-frequency magnetic fields (LF-MFs) inhibit proliferation by triggering apoptosis and altering cell cycle distribution in breast cancer cells. International Journal of Molecular Sciences, 2020. 21(8): p. 2952. [CrossRef]
- Tatarov, I., et al., Effect of magnetic fields on tumor growth and viability. Comparative medicine, 2011. 61(4): p. 339-345.
- Raskov, H., et al., Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. British journal of cancer, 2021. 124(2): p. 359-367. [CrossRef]
- Hwang, C., et al., Stromal tumor-infiltrating lymphocytes evaluated on H&E-stained slides are an independent prognostic factor in epithelial ovarian cancer and ovarian serous carcinoma. Oncology letters, 2019. 17(5): p. 4557-4565. [CrossRef]
- Lakritz, J.R., et al., Beneficial bacteria stimulate host immune cells to counteract dietary and genetic predisposition to mammary cancer in mice. International journal of cancer, 2014. 135(3): p. 529-540. [CrossRef]
- Farhood, B., M. Najafi, and K. Mortezaee, CD8+ cytotoxic T lymphocytes in cancer immunotherapy: A review. Journal of cellular physiology, 2019. 234(6): p. 8509-8521. [CrossRef]
- Umegaki, S., et al., Distinct role of CD8 cells and CD4 cells in antitumor immunity triggered by cell apoptosis using a Herpes simplex virus thymidine kinase/ganciclovir system. Cancer Science, 2023. [CrossRef]
- Inai, T., et al., Inhibition of vascular endothelial growth factor (VEGF) signaling in cancer causes loss of endothelial fenestrations, regression of tumor vessels, and appearance of basement membrane ghosts. The American journal of pathology, 2004. 165(1): p. 35-52. [CrossRef]
- Goel, H.L. and A.M. Mercurio, VEGF targets the tumour cell. Nature Reviews Cancer, 2013. 13(12): p. 871-882. [CrossRef]
- Chen, Y., et al., Influence of gut and intratumoral microbiota on the immune microenvironment and anti-cancer therapy. Pharmacological Research, 2021. 174: p. 105966. [CrossRef]
- Soto-Pantoja, D.R., et al., Diet alters entero-mammary signaling to regulate the breast microbiome and tumorigenesis. Cancer research, 2021. 81(14): p. 3890-3904. [CrossRef]
- Kong, F., et al., Abundance and metabolism disruptions of intratumoral microbiota by chemical and physical actions unfreeze tumor treatment resistance. Advanced Science, 2022. 9(7): p. 2105523. [CrossRef]
- Matson, V., C.S. Chervin, and T.F. Gajewski, Cancer and the microbiome—influence of the commensal microbiota on cancer, immune responses, and immunotherapy. Gastroenterology, 2021. 160(2): p. 600-613.
- Banerjee, S., et al., Distinct microbiological signatures associated with triple negative breast cancer. Scientific reports, 2015. 5(1): p. 15162.
- Junqueira, C., et al., Trypanosoma cruzi adjuvants potentiate T cell-mediated immunity induced by a NY-ESO-1 based antitumor vaccine. PloS one, 2012. 7(5): p. e36245. [CrossRef]
Figure 1.
ELF-EMF had no significant impact on tumor growth rate in TNBC mouse models, and the tumor growth rate in both the exposure and control groups was increasing. On the other hand, ELF-EMF led to a reduction in tumor growth rate in TPBC mouse models, and from day 9 onwards, the tumor growth rate in the exposure group was lower than in the control group.
Figure 1.
ELF-EMF had no significant impact on tumor growth rate in TNBC mouse models, and the tumor growth rate in both the exposure and control groups was increasing. On the other hand, ELF-EMF led to a reduction in tumor growth rate in TPBC mouse models, and from day 9 onwards, the tumor growth rate in the exposure group was lower than in the control group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



