
Submitted:
02 December 2024
Posted:
03 December 2024
You are already at the latest version
Abstract
HIV proviral reservoirs are cells that harbour integrated HIV proviral DNA within their nuclear genomes. These cells form a heterogeneous group, represented by peripheral blood mononuclear cells (PBMCs), tissue-resident lymphoid and monocytic cells and glial cells of the central nervous system. The importance of studying the properties of proviral reservoirs is connected with the inaccessibility of integrated HIV proviral DNA for modern therapies that block virus reproduction. If treatment is not effective enough or is interrupted, the proviral reservoir can reactivate. Early initiation of ART improves the prognosis of the course of HIV infection, which is explained by the reduction of the proviral reservoir pool in the early stages of the disease. HIV subtype provides differences in the number of latent reservoirs among different subtypes, as determined by structural and functional differences. Unique phenotypes of HIV-infected patients, such as elite controllers, have control over viral replication and can be said to have achieved functional cure of HIV infection. Uncovering the causes of this phenomenon will bring humanity closer to curing HIV infection, potential approaches to which are CRSPR/cas9, “Shock and kill” and “Block and lock”.
Keywords:
HIV-1
; Proviral reservoir
; HIV latency
; HIV cure
; ART
; HIV subtype
; Elite controllers
1. Introduction
At present, HIV remains a major global public health issue. As of the end of 2023, it was estimated that approximately 39.9 million people were living with HIV (with a range from 36.1 million to 44.6 million). In some regions, the trend of increasing new infections has resumed. Mortality due to HIV infection and associated opportunistic infections remains significant, amounting to approximately 630,000 deaths per year globally [1]. The development and implementation of antiretroviral therapy (ART) have contributed to a reduction in new HIV infections and deaths [2,3]. However, a complete cure for HIV remains currently unattainable [4]. Proviral reservoirs represent a major obstacle to the complete eradication of HIV from the human body [5,6]. These are cells in which HIV proviral DNA has integrated into the nuclear genome. For various reasons, some proviral reservoirs exist in a latent state, with either complete or partial cessation of viral gene transcription, creating a group of latent reservoirs [7]. In this state, infected cells can evade immune surveillance by preserving HIV DNA, until viral gene transcription is reactivated [8,9]. The existence of these latent reservoirs necessitates lifelong ART therapy and contributes to the development of chronic HIV infection [10].
Currently, the study of proviral reservoirs is one of the most pertinent areas in HIV research, as the integrity of integrated proviral DNA is critically important for the continued progression of the infection [11]. Studies have demonstrated the impact of the timing of ART initiation following diagnosis, the clinical characteristics of the patient, the HIV genetic variant, and a number of other factors on the characteristics of proviral reservoirs [12,13,14]. Based on this research, exposure to the HIV provirus may contribute to the development of new methods for treating HIV infection [15].
2. Establishment of Proviral Reservoirs and Their Role in HIV Pathogenesis
2.1. Definition of HIV Proviral Reservoirs and the Timing of Their Formation
Currently, there is no precise definition of proviral reservoirs, as there is no consensus on what should be included in this definition [16]. The most common interpretation in the literature defines proviral reservoirs as any cells capable of producing replication-competent virus in individuals who have been receiving ART for several years [17,18,19]. This definition accurately characterizes proviral reservoirs as a major challenge for HIV treatment. However, it may narrow the scope for future studies by excluding cells with HIV DNA that cannot produce replication-competent virus. The contribution of such defective proviral reservoirs to the potential cure of HIV may be significant and worth considering [20]. Additionally, this definition implies that proviral reservoirs are largely latent, as infected cells actively producing viral particles are often eliminated with long-term ART [21], although active release of virions may occur in distinct anatomical compartments (e.g., brain, lymph nodes) [22].
Therefore, in order to avoid restrictions for future research, it may be beneficial to define proviral reservoirs comprehensively as cells containing HIV proviral DNA integrated into the nuclear genome. Further division of these proviral reservoirs into latent/active and intact/defective categories can help identify important research targets within the context of proviral reservoirs, without limiting the overall scope of research. The proposed classification is illustrated in Figure 1.
HIV reservoirs begin to form at the earliest stages of HIV infection (stage I of acute HIV infection, according to Fiebieg, when HIV RNA can be detected [23]), as HIV DNA can already be detected in patients [13]. In HIV-infected individuals who do not receive ART during the acute phase, the levels of total and integrated HIV DNA, as well as 2-LTR circular forms, reach a peak within the first two weeks after infection. These levels remain relatively stable without treatment [24]. At this stage, peripheral blood mononuclear cells (PBMCs), and in particular CD4+ T lymphocytes, which transition into memory T cells after antigen stimulation ceases, are the primary cells that enter viral reservoirs [25].
2.2. Where Does the Provirus Hide?
Memory T cells have several subsets that may act as HIV proviral reservoirs: central memory T cells (Tcm), effector memory T cells (Tem), transient or transitional memory T cells (Ttm), stem memory T cells (Tscm), and naïve memory T cells (Tna) [26,27,28]. Other T cell subsets may also act as proviral reservoirs. For example, γδ T cells have been shown to produce replication-competent virus [29]. The level of CD4 T cell differentiation is not the only way to classify them, so some studies also consider the functional polarization of CD4 T cells [30]. Follicular T helper (Tfh) cells have been shown to be an important part of the proviral reservoir [31]. This information on the identification of cell populations containing HIV provirus is crucial, as these populations will be the targets for potential therapeutic methods aimed at eliminating proviral reservoirs. However, at present, obtaining an accurate distribution of proviral reservoirs among different T cell populations remains challenging due to their high heterogeneity and the technical difficulties associated with comprehensive analysis [25,32], necessitating further research on cell populations that may act as proviral reservoirs.
HIV reservoirs are also present in a number of other cell types, including cells of the monocyte-macrophage system and glial cells of the central nervous system [33]. These cells, like CD4 T lymphocytes, are among the earliest target cells of HIV, as they also express CD4 receptors and the CCR5 and CXCR4 co-receptors that allow the virus to bind to and enter the cell [34]. These reservoirs are significant as they are more resistant to apoptosis following HIV infection [35] and have the ability to migrate to various parts of the human body, evading immune surveillance, for example, to the lymph nodes and the central nervous system [36]. These characteristics suggest that cells of the monocyte-macrophage system may also form latent HIV reservoirs with distinctive features. Glial cells of the CNS also serve as a HIV reservoir, since the penetration of ART drugs into the brain is difficult due to the blood-brain barrier (BBB), which can lead to the ongoing production of HIV virions [37]. HIV replication in the brain causes neurocognitive disorders of varying degrees in most HIV-infected patients [38], as, with the advent of ART, the life expectancy of such patients has increased. However, HIV persists in the nervous system that leads to a cumulative cytotoxic effect mediated by viral infection and immune activity [39].
Astrocytes [40] and microglia [41] are the primary cellular reservoirs of HIV in the nervous system. Additionally, perivascular macrophages [42] have been shown to contribute to the HIV neuro reservoir. Viral proteins, such as Tat [43] and Vpr [44], induce neuronal death and provoke neuroinflammatory reactions. It is also known that HIV infection alters the levels of neurometabolites, with increased choline and myo-inositol and decreased N-acetylaspartate, which reflect an enhanced inflammatory background and neuronal dysfunction [45]. These findings suggest that neuroinflammation and neuronal dysfunction are integral components in the development of neurocognitive disorders associated with HIV infection. A link has been established between high choline and low N-acetylaspartate levels and the development of HIV-associated neurocognitive disorders (HANDs) [46].
2.3. Proviral Reservoirs and Latency
According to in vitro studies, approximately 65% of infected cells become latent proviral reservoirs after proviral integration [47]. HIV can directly infect resting cells, which also contributes to the replenishment of the latent reservoir [48]. Cells that were infected while in an active state and later entered a dormant state exhibit a higher activation frequency in vitro compared to those infected while at rest [49]. Infection of dormant cells more frequently results in the establishment of latent infection, while infection of activated cells typically leads to productive infection [50].
After the establishment of a latent reservoir, viral gene transcription is suppressed through several mechanisms. These mechanisms include transcriptional interference, epigenetic regulation of gene expression, and the interaction of transcription factors [7]. In combination with high mutability, evasion of apoptosis, and resistance to neutralizing antibodies, this suppression leads to the evasion of the cytotoxic immune response [51,52]. ART suppresses HIV replication and depletes proviral reservoirs to some extent, but this approach does not result in complete eradication [4]. A study of 30 HIV-positive participants who had been on ART for a prolonged period (7–12 years) showed that during the first year, there was a rapid depletion of reservoirs, with a sevenfold decrease in HIV DNA from a median value of 7319 copies/10^6 CD4 cells to 1054 copies/10^6 CD4 cells. However, further ART did not result in significant or rapid reservoir depletion [21]. This highlights the issue of the long-term persistence of the provirus within the human cell genome, as no effective therapeutic technique currently exists to target latent proviral reservoirs for the complete eradication of HIV.
2.4. Proviral Immortality
Maintaining a stable number of these reservoirs is one of the critical aspects of reservoir formation during the long-term course of HIV infection. Intact proviral reservoirs are believed to continue releasing replication-competent virus, even in individuals receiving effective ART, leading to a constant replenishment of the reservoir and the emergence of low-level viremia [53]. However, further research is needed to draw definitive conclusions about this phenomenon. Increasing evidence suggests that mechanisms of cellular proliferation play a significant role in the stability of these reservoirs [54]. These mechanisms include homeostatic proliferation, which refers to IL-7-dependent proliferation of CD4 cells when they are depleted [55]; antigen-dependent proliferation, especially in the intestine, where frequent contact with various antigens occurs [56]; and proliferation due to HIV integration into genomic regions that provide a survival advantage, such as the MCL2 and BACH2 genes, which are involved in cell growth and development, including that of T lymphocytes [57].
2.5. Intact and Defective Proviral Reservoirs: Which Are More Important?
The number of intact and defective proviral reservoirs is also significant, as intact reservoirs are believed to be the main pathogenic substrate for chronic HIV infection and to cause a large number of adverse events during ART, such as the emergence of drug-resistant mutant HIV and associated virologic failures [58]. Although this conclusion remains highly controversial, at present, it is impossible to provide a clear answer as to which part of the proviral reservoirs plays the most important role in achieving complete eradication of HIV infection. Studies have shown that the time it takes for viral load to resume after analytical treatment interruption (ATI) is strongly correlated with the levels of intact HIV DNA during the chronic phase of infection [59]. This underscores the need for further studies to gain a comprehensive understanding of the role of intact proviral reservoirs in viral load resumption. Such research is crucial for designing new eradication strategies.
Formation of defective proviruses begins early in HIV infection, and their number can reach up to 90% of the total reservoir size [60]. These defective proviruses do not undergo transcription and/or translation and do not contribute directly to HIV pathogenesis. However, some defective proviruses are transcriptionally and translationally competent and can contribute to chronic immune activation and, possibly, to the replication-competent portion of the reservoir [61]. It has been shown that defective proviral reservoirs can produce Gag and Nef proteins, which may contribute to immune activation even in patients with a suppressed viral load while taking ART [20]. It is also known that proviral reservoirs of this kind can transcribe unique HIV mRNA, in which the exons differ from the described spliced variants of HIV mRNA. Moreover, such mRNA contains translation-competent reading frames, which can provoke an immune response [62]. The persistence of defective proviral reservoirs eventually leads to the depletion of CD8 T cells and the disruption of their cytotoxic function [63], highlighting the need for their elimination to achieve complete HIV cure.
2.6. From Fundamental Research to Clinical Application
It is known that the number of proviral reservoirs serves as a predictor of HIV remission after ART cessation and the rate of HIV infection progression in the complete absence of ART [64,65,66,67]. One meta-analysis demonstrated a strong predictive ability of total HIV DNA regarding the progression of HIV infection to acquired immunodeficiency syndrome (AIDS) and fatal outcomes [68]. This finding is supported by other studies in which HIV DNA levels served as predictors of early HIV progression following suspected infection in naïve patients with confirmed HIV infection [66]. In another study, HIV DNA levels were shown to predict the progression of HIV infection, regardless of HIV RNA and CD4 cell levels in patients undergoing seroconversion [67]. Additionally, one study indicated that the total HIV DNA level at the time of analytical treatment interruption (ATI) was a prognostic factor for predicting the time until viral load recovery [65].
3. Factors Affecting the Characteristics of Proviral Reservoirs.
3.1. ART in the Early Stages of HIV Infection Results in a Significant Reduction in the Number of Proviral Reservoirs and Alters their Distribution Among Different Subsets of T Cells
Many studies have shown that initiating ART in the first few weeks or months after infection can reduce the amount of HIV cell-associated DNA [69,70,71,72,73]. The effects of early ART on the amount of HIV cell-associated DNA in the PBMC population were examined, with patients being assigned standard ART regimens during the acute phase of HIV infection. The results showed a decrease in the amount of HIV cell-associated DNA in PBMCs in all patients one year after starting ART, with median values of 3.04 log copies/10^6 PBMC before ART initiation and 2.18 log copies/10^6 PBMC after one year of therapy [73]. These data are consistent with an earlier study that also investigated the effect of early ART on proviral reservoirs. This study found that, 48 weeks after starting treatment, patients in the acute phase of HIV infection showed a decrease in the median level of cell-associated HIV DNA from 2.8 log copies/10^6 PBMC to 1.6 log copies/10^6 PBMC. Additionally, 9.5% of the patients had undetectable HIV DNA levels in their PBMCs [70]. Later initiation of ART, starting from the Fiebig II stage, resulted in higher frequencies of PBMCs with cell-associated HIV DNA. In patients at the Fiebig I stage, the median value of HIV cell-associated DNA in PBMCs was 8 copies/10^6 PBMC, whereas in patients at the Fiebig II stage, this value was 96 copies/10^6 PBMC [69]. However, the decrease in total HIV DNA is primarily due to non-integrated forms of DNA. The role of these forms in viral reservoirs is still unclear and requires a more cautious interpretation of data from studies that examine only the level of total HIV DNA [71]. A study that included HIV-infected patients at different stages of the disease showed that, in patients with chronic HIV infection who started ART, the main form of HIV DNA that decreased during treatment was non-integrated DNA. However, in patients who started ART at an early stage of HIV infection (Fiebig II-III), integrated forms of HIV DNA also decreased significantly. This confirms the benefit of early ART initiation [72].
It is known that the kinetics of proviral reservoir depletion are most favorable for patients who start ART within a short period after infection (1-3 months), with a decrease of 0.07 log copies per year. In HIV-positive individuals who begin ART during the chronic phase of infection, the rate of reservoir depletion is slower (0.01 log copies per year) [27]. It is believed that short-lived effector CD4 memory T cells are more susceptible to HIV and are actively infected during the acute phase, while in the chronic phase, HIV primarily targets long-lived central memory CD4 T cells [74]. These cells have an increased likelihood of becoming latent reservoirs due to their long lifespan and prolonged dormancy. Early ART leads to the rapid elimination of proviral reservoirs from effector CD4 T cells, whereas late ART is less effective, as CD4 memory T cells are located in less accessible anatomical sites, are inactive, and do not express viral genes.
ART has different effects on the distribution of integrated HIV DNA among subsets of memory T cells at various stages of HIV infection. When ART is initiated at the Fiebig II-III stage, there is a decrease in integrated HIV DNA in Tcm, Ttm, and Tem cells. However, in patients who begin ART at the Fiebig IV-VI stages, there is no significant reduction in integrated HIV DNA levels. It has also been shown that the contribution of the Tcm subpopulation to the total number of proviral reservoirs after ART initiation is lower in patients at Fiebig stages II-III compared to those in the chronic phase [72]. This suggests that in the chronic phase, most proviral reservoirs are latent and inaccessible to eradication through ART, emphasizing the greater benefit of early ART initiation [27].
3.2. The ART Effect On The Number Of Intact And Defective Proviral Reservoirs
The use of next-generation sequencing (NGS) in a study on the distribution of intact and defective proviral reservoirs in patients with acute and chronic infection showed that ART has little effect on the ratio of intact to defective genomes at early stages [60]. However, another study found that the number of intact reservoirs decreased more quickly in patients already taking ART (the median time after ART initiation was 617 days). The median annual decay rate of intact proviral reservoirs was 15.7% for the first seven years after the start of the study, with a slower decline to 3.6% after that. The median decay rate of defective proviral reservoirs was 4% annually for the first seven years. After that, the decline slowed to 1.5% annually [75]. It can be assumed that these differences are due to lower pressure of the immune system on the defective proviral reservoirs. This is associated with ineffective transcription and translation in most defective clones. However, it is also possible that there is a different distribution of intact and defective proviral reservoirs among cell populations. Further research is needed to determine the exact cause of these differences.
3.3. Is it Necessary to Take Into Account the HIV Subtype when Analyzing Proviral Reservoirs?
In the genetic group M, which is the most prevalent worldwide, there are more than 10 subtypes and numerous recombinant forms of HIV. Some of these variants significantly impact the epidemiological situation in various regions [76]. Studying the differences between genetic variants of the virus in relation to the characteristics of proviral reservoirs is crucial for developing and implementing new, effective HIV treatment strategies aimed at controlling the global spread of the virus.
Several studies have established a link between the HIV gene and the progression of HIV infection. One early study conducted in Senegal examined the rate of AIDS progression among female sex workers infected with subtypes A, C, D, and G. The results revealed that patients infected with subtype A developed AIDS approximately eight times more slowly than those infected with other subtypes [77]. Further studies comparing subtypes C, A, and D confirmed similar findings: patients infected with subtype C experienced a more rapid decline in CD4 cells, an increase in viral load, and faster progression to AIDS. Similarly, patients infected with subtype D exhibited a quicker rise in viral load and progression to AIDS compared to those infected with subtype A [78]. Another study reinforced these differences between subtypes D, A, and C, showing that subtype D was associated with a faster disease progression and a lower CD4 cell count [79]. Since the number of proviral reservoirs is believed to predict the clinical progression of HIV infection [64], it is essential to consider the quantity of these reservoirs in relation to subtype-specific differences in HIV development.
One of the most informative studies compared cohorts of HIV-positive patients from Uganda and the United States. In Ugandan patients, subtype D was the most prevalent, while subtype A and the A/D recombinant form were less common. Subtype B, on the other hand, was predominantly found in US patients. This study demonstrated that patients with subtype B had a larger reservoir of infectious units (median value: 1.08 infectious units per million cells (IUPM)) in a latent state compared to Ugandan patients (median value: 0.36 IUPM) [14]. Another study focused on the number of proviral reservoirs in patients who began ART in the early stages of infection. The findings were consistent, revealing a higher number of proviral reservoirs in patients with subtype B compared to those with other subtypes. This difference was evident both before ART initiation (median value: 3.4 log10 copies/10^6 PBMC versus 2.8 log10 copies/10^6 PBMC) and after 48 weeks of ART (median value: 2.5 log10 copies/10^6 PBMC). Patients with subtype B also appeared to have more active viral protein Nef, which helps the virus evade immune surveillance by downregulating the expression of CD4 and MHC-I in infected cells. This may be linked to the larger number of proviral reservoirs observed in these patients. However, no direct correlation between CD4 expression and the number of proviral reservoirs was noted in this study [80]. A separate study, involving over 1,000 HIV-positive patients, revealed differences in total HIV DNA levels among patients infected with different subtypes. Patients with subtype B had higher levels of total HIV DNA compared to those with other subtypes, particularly in comparison to subtype C [81]. These findings suggest that the total number of reservoirs, and the number of latent HIV reservoirs, are influenced by the genetic variant of the virus responsible for the infection.
To confirm the above conclusions, it is important to take into account molecular differences among HIV genetic variants, since there is evidence that different genetic variants may establish latency in different ways and with different kinetic rates. Among the molecular mechanisms related to latent HIV proviral reservoirs, the LTR structure is usually associated with the promoter area, which have binding sites for cell transcription factors and, accordingly, are responsible for reactivating the latent reservoir [82]. Additionally, the functioning of viral proteins such as Nef [83], Vif [84], and Vpu [85] also contribute to the establishment of latency within proviral reservoirs.
3.4. The Unique Phenotypes of HIV-Infected Patients Have Features in the Characteristics of Proviral Reservoirs
Researchers are also interested in the unique phenotypes of HIV-infected patients who show the best indicators of HIV infection progression on clinical and laboratory tests due to a number of features. These phenotypes include elite controllers (EC), post-treatment controllers (PTCs), and the Low Viral Reservoir Treated (LoViReT) phenotype.
Elite controllers are a group of HIV-positive patients who have been monitored for viral replication without ART for at least 12 months. During this time, they may confirm HIV infection through serological tests, but viral RNA cannot be detected in their blood plasma using PCR [86]. A study compared the virological and immunological characteristics of elite controllers with those of patients at the chronic stage of HIV infection. Elite controllers had significantly lower levels of total and intact HIV DNA in their CD4 T cells compared to patients with chronic infection. Additionally, ECs retained actively infected cells that produce viral antigens, as they had high levels of activation markers on specific CD8 T cells. This suggests a potentially strong cytotoxic response in elite controllers [87]. In another study, elite controllers were found to have lower levels of total and intact HIV DNA in PBMCs compared to HIV-infected patients on ART at the chronic stage. The median values were 30.4 copies/10^6 PBMCs for elite controllers versus 602.52 copies/10^6 PBMCs for the chronically infected group. At the same time, the ratio of intact to total HIV DNA in ECs was similar to or even higher than that of chronically progressing patients, indicating that elite controllers do not have a large number of defective reservoirs [88]. This suggests that elite controllers may represent a model for a functional cure of HIV infection. They have a replication-competent provirus but are able to maintain complete remission due to fewer proviral reservoirs and a more effective HIV-specific cytotoxic immune response. Similar results have been observed in PTCs, although their mechanism of functional cure is more attributed to the early initiation of ART, which leads to a smaller number of proviral reservoirs, rather than the strong cytotoxic response typical of ECs [89].
Patients with the LoViReT phenotype are a cohort of HIV-infected individuals taking ART and having a relatively small reservoir size. A study conducted on this cohort found these patients had lower levels of starting proviral DNA in their CD4 cells, and that this DNA was less transcriptionally active. This phenotype differs in the distribution of proviral reservoirs. Thus, in LoViReT, most of the reservoir is located in short-lived Ttm and Tem cells. This is thought to be one of the reasons for the small reservoir size in this group of patients. Interestingly, the time at which ART was started does not correlate with reservoir size in patients with LoViReT. This finding suggests that there may be other factors involved in determining the size of the reservoirs in HIV-infected individuals [90]. A graphical representation of the above phenotypes and their characteristics are shown in Figure 2.
3.5. Modern Methods of Analyzing Proviral Reservoirs
Real-time PCR has been widely used to assess the amount of HIV DNA in laboratory samples. However, this method tends to overestimate the number of replication-competent viruses due to methodological limitations in quantifying total cell-associated HIV DNA [60]. To address these limitations, several alternative techniques have been developed, including droplet digital PCR (ddPCR) and digital PCR (dPCR). These methods focus on detecting intact viral genomes [91] and enable the quantification of viral genes, as well as the assessment of the proportion of genomes that include all key HIV genes (such as gag, pol, env, Nef, etc.). The interpretation of results from these techniques can vary depending on the platform and the specific primer/probe sets used, which differ across studies [92]. One advantage of these methods is that they provide a more accurate estimate of the absolute amount of HIV DNA in a sample compared to real-time PCR. However, some researchers have raised concerns about the potential for false positives, particularly in samples lacking the matrix or in individuals not infected with HIV [93,94].
Sequencing of proviral genomes is an important method for analyzing proviral reservoirs. Currently, there are many methods available that allow us to obtain genomes or sequences of almost full length, which allows us to better understand the characteristics of proviral reservoirs in different cell populations [11,28,60]. One promising method of sequencing-based analysis is mapping HIV infection sites [95]. The HIV provirus can integrate into genes responsible for cell growth and development [57], making it necessary to study the features of integration and its connection with the establishment of latency in proviral reservoirs.
Quantitative analysis of viral growth, also known as Quantitative Viral Outgrowth Assay (QOVA), is the most well-known and widely used method for quantitatively assessing replication-competent reservoirs. However, this analysis may underestimate the true number of replication-competent reservoirs, as some of them are in a deep latent state and are not activated even by repeated stimulation [96]. The aforementioned methods are shown in Figure 3.
4. Therapeutic Options for Treating HIV Infection Aimed at Controlling Proviral Reservoirs.
4.1. ART Cannot Eliminate Proviral Reservoirs on Its Own
The presence of latent proviral reservoirs in the HIV-infected body, which can be reactivated, is one of the most important challenges in achieving a complete cure for HIV infection. Therefore, it is essential to develop new strategies to treat HIV infection and eliminate proviral reservoirs. Currently, widely used ART is a successful method of converting HIV infection into a chronic, controlled condition, but it requires lifelong drug intake. Today, ART allows people to live with the disease, but it is not a complete cure. ART reduces the number of reservoirs, but does not eliminate them entirely. Even the intensification or initial use of more potent ART regimens, such as MegaART (which includes Raltegravir and Maraviroc), does not show better results in reducing the amount of proviral reservoirs compared to standard ART, encouraging the search for alternative treatments [99]. New HIV treatments are currently under active development. These treatments can be divided into several categories: genetic editing of the genome using CRISPR/Cas to eliminate the provirus from proviral reservoirs [100]; activation of latent proviral reservoirs and stimulation of the immune system to destroy all cells containing the provirus (Shock and Kill) [101]; and induction of a more dormant state of proviral reservoirs, preventing further activation (Block and Lock) [102].
4.2. CRISPR/Cas9
Genetic editing using the CRISPR/Cas9 system has shown great promise for curing HIV infection since its inception. This system consists of two components: the Cas restriction endonuclease and the CRISPR RNA guide targeting complementary DNA/RNA sequences [103]. The Cas protein, with its endonuclease activity, can introduce mutations into integrated HIV DNA or completely remove it from the nuclear genome. Relatively conserved genes, such as Gag [104,105], Pol [105,106], Rev [106] and Vpr [107] have been identified as potential targets in the HIV genome. Several studies have shown encouraging results in inhibiting viral transcription and translation [107,108,109]. However, in most cases, it is often preferable to apply a combined approach using multiple CRISPR RNA targets, as single genome modifications may induce the non-homologous end joining (NHEJ) repair mechanism, which can lead to mutations in the viral genome. This, in turn, can lead to resistance to the CRISPR/Cas system, as it can no longer recognize the sites for restriction, which further leads to the resumption of productive infection [111,112].
4.3. Shock and Kill
The Shock and Kill approach leads to the reactivation of HIV gene transcription through latent reservoir inhibitors (LARs), initiating viral protein translation and subsequent cell lysis via various immune mechanisms [113]. The main LARs include histone deacetylase inhibitors [114,115], histone methyltransferase inhibitors [116], bromodomain inhibitors [117], protein kinase C agonists [118], and Toll-like receptor (TLR) agonists [119]. Although these drugs have shown promising results in reversing HIV latency, infected cells activated through this process may survive despite the cytopathic effects of HIV and the action of cytotoxic CD8 T cells [120]. Bcl-2 antagonists [121], PI3K/Akt inhibitors [122], Smac mimetics [123] and RIG-I inducers [124] are proposed as potential drugs that could increase the efficiency of proviral reservoir lysis by working synergistically with the immune system.
4.4. Block and Lock.
Block and Lock refers to an approach in which HIV remains in the body but becomes completely unable to replicate, aiming for a functional cure of HIV infection [125]. This strategy is proposed to be implemented through a combination of drugs, including Didehydro-Cortistatin A, which inhibits the Tat protein [126]; LEDGINs, a group of drugs that block the interaction between HIV integrase and the cellular chromatin tethering factor LEDGF/p75 [127]; Curaxin CBL0100, an inhibitor of the chromatin-facilitating transcription complex (FACT) [128]; heat shock protein 90 inhibitors (HSP90) [129]; Jak-STAT inhibitors; kinase inhibitors [130], and other drug classes.
5. Conclusions
HIV proviral reservoirs remain the primary obstacle in the quest for a cure for HIV infection. For nearly 40 years, scientists and healthcare professionals worldwide have been working toward finding a cure, but no proven method for eliminating the disease has yet been discovered. Potential therapeutic approaches, such as CRISPR/Cas9, Block and Lock, and Shock and Kill, are still being explored and refined. However, none have demonstrated definitive success in clinical trials, underscoring the need for continued research into the properties of HIV proviral reservoirs.
ART has shown significant impact on the size of proviral reservoirs, with the timing of its initiation being a critical factor. Even at early stages of HIV (Fiebieg stages I and II), there is a 12-fold difference in cell-associated DNA. However, cell-associated DNA encompasses several forms of HIV DNA, not all of which lead to productive infection. Intact proviral reservoirs are likely a key component of the overall reservoir and should be prioritized for both analysis and treatment. Research findings in this area remain inconclusive, necessitating further confirmation. Currently, IPDA, NGS, and QOVA are considered the best methods for studying intact proviral reservoirs. Their varied applications enable researchers to explore different aspects of HIV molecular biology, including mapping integration sites, determining the proportion of defective and intact reservoirs, and assessing the inducibility of these reservoirs.
The genetic variant of HIV plays an important role in proviral reservoirs, with several studies showing that patients with subtype B have a larger number of reservoirs, both general and latent. These differences are linked to the structure of the long terminal repeat (LTR) region and the function of HIV proteins, which make them potential targets for intervention.
Elite and post-treatment controllers, who maintain an undetectable viral load without ART and do not progress in HIV infection, are valuable subjects for research and the identification of unique traits. Strong CD8 cytotoxic responses in elite controllers have already been shown to be a critical factor in achieving remission. Similarly, early ART initiation in post-treatment controllers highlights the benefits of this approach. Together, these findings open up promising new avenues for developing HIV treatment strategies, with the goal of achieving a stable and functional cure—a highly desirable outcome for HIV-positive individuals.
Author Contributions
Conceptualization, N. G.; Writing the original manuscript and drawing design, A. M.; writing-review and editing K. E., N. G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The figures in this manuscript were created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- «The urgency of now: AIDS at a crossroads. Geneva: Joint United Nations Programme on HIV/AIDS; 2024. Licence: CC BY-NC-SA 3.0 IGO..» 2023.
- R. A. Legarth, M. G. Ahlström, G. Kronborg, C. S. Larsen, C. Pedersen, G. Pedersen, R. Mohey, J. Gerstoft и N. Obel, «Long-Term Mortality in HIV-Infected Individuals 50 Years or Older: A Nationwide, Population-Based Cohort Study.» J Acquir Immune Defic Syndr. 2016, 71, 213–218. [CrossRef]
- H. Samji, A. Cescon, R. S. Hogg, S. P. Modur, K. N. Althoff, K. Buchacz, A. N. Burchell, M. Cohen, K. A. Gebo, M. J. Gill, A. Justice, G. Kirk, M. B. Klein, P. T. Korthuis, J. Martin, S. Napravnik, S. B. Rourke, T. R. Sterling, M. J. Silverberg, S. Deeks, L. P. Jacobson, R. J. Bosch, M. M. Kitahata, J. J. Goedert, R. Moore и S. J. Gange, «Closing the Gap: Increases in Life Expectancy among Treated HIV-Positive Individuals in the United States and Canada.» PLoS ONE, 2013, 8, e81355. [CrossRef]
- M. C. Pitman, J. S. Y. Lau, J. H. McMahon и S. R. Lewin, «Barriers and strategies to achieve a cure for HIV» Lancet HIV, 2018, 5, e317–e328. [CrossRef]
- T. -W. Chun, S. Moir и A. S. Fauci, «HIV reservoirs as obstacles and opportunities for an HIV cure.» Nat. Immunol., 2015, 16, 584–589. [CrossRef]
- M. S. Dahabieh, E. Battivelli и E. Verdin, «Understanding HIV Latency: The Road to an HIV Cure.» Annu. Rev. Med., 2015, 66, 407–421. [CrossRef]
- D. C. Cary, K. Fujinaga и B. M. Peterlin, «Molecular mechanisms of HIV latency.» J. Clin. Investig., 2016, 126, 448–454. [CrossRef]
- M. J. Pace, L. Agosto, E. H. Graf и U. O’Doherty, «HIV reservoirs and latency models.» Virology, 2011, 411, 344–354. [CrossRef]
- A. Shukla, N.-G. P. Ramirez и I. D’Orso, «HIV-1 Proviral Transcription and Latency in the New Era.» Viruses, 2020, 12, 555. [CrossRef]
- P. A. Volberding и S. G. Deeks, «Antiretroviral therapy and management of HIV infection.» Lancet, 2010, 376, 49–62. [CrossRef]
- Y. -C. Ho, L. Shan, N. Hosmane, J. Wang, S. Laskey, D. S. Rosenbloom, J. Lai, J. Blankson, J. Siliciano и R. Siliciano, «Replication-Competent Noninduced Proviruses in the Latent Reservoir Increase Barrier to HIV-1 Cure.» Cell, 2013, 155, 540–551. [CrossRef]
- J. Ananworanich, K. Dubé и N. Chomont, «How does the timing of antiretroviral therapy initiation in acute infection affect HIV reservoirs?.» Curr Opin in HIV AIDS, 2015, 10, 18–28. [CrossRef]
- J. Ananworanich, C. P. Sacdalan, S. Pinyakorn, N. Chomont, M. Souza, T. Luekasemsuk, A. Schuetz, S. J. Krebs, R. Dewar, L. Jagodzinski, S. Ubolyam, R. Trichavaroj, S. Tovanabutra, S. Spudich, V. Valcour, I. Sereti, N. Michael, M. Robb, P. Phanuphak, J. H. Kim и N. Phanuphak, «Virological and immunological characteristics of HIV-infected individuals at the earliest stage of infection.» J Virus Erad, 2016, 2, 43–48. [CrossRef]
- J. L. Prodger, J. Lai, S. J. Reynolds, J. C. Keruly, R. D. Moore, J. Kasule, T. Kityamuweesi, P. Buule, D. Serwadda, M. Nason, A. A. Capoferri, S. F. Porcella, R. F. Siliciano, A. D. Redd, J. D. Siliciano и T. C. Quinn, «Reduced Frequency of Cells Latently Infected With Replication-Competent Human Immunodeficiency Virus-1 in Virally Suppressed Individuals Living in Rakai, Uganda.» Clin Infect Dis, 2017, 65, 1308–1315. [CrossRef]
- L. B. Cohn, N. Chomont и S. G. Deeks, «The Biology of the HIV-1 Latent Reservoir and Implications for Cure Strategies.» Cell Host Microbe, 2020, 27, 519–530. [CrossRef]
- M. J. Churchill, S. G. Deeks, D. M. Margolis, R. F. Siliciano и R. Swanstrom, «HIV reservoirs: what, where and how to target them.» Nat Rev Microb, 2015, 14, 55–60. [CrossRef]
- E. Eisele и R. Siliciano, «Redefining the Viral Reservoirs that Prevent HIV-1 Eradication.» Immunity, 2012, 37, 377–388. [CrossRef]
- S. D. Falcinelli, C. S. D. Falcinelli, C. Ceriani, D. M. Margolis и N. M. Archin, «New Frontiers in Measuring and Characterizing the HIV Reservoir.» Front Microbiol, 2019, 10. [CrossRef]
- J. Vanhamel, A. Bruggemans и Z. Debyser, «Establishment of latent HIV-1 reservoirs: what do we really know?.» J Virus Erad, 2019, 5, 3–9. [CrossRef]
- H. Imamichi, M. Smith, J. W. Adelsberger, T. Izumi, F. Scrimieri, B. T. Sherman, C. A. Rehm, T. Imamichi, A. Pau, M. Catalfamo, A. S. Fauci и H. C. Lane, «Defective HIV-1 proviruses produce viral proteins.» Proc Natl Acad Sci U S A, 2020, 117, 3704–3710. [CrossRef]
- G. J. Besson, C. M. Lalama, R. J. Bosch, R. T. Gandhi, M. A. Bedison, E. Aga, S. A. Riddler, D. K. McMahon, F. Hong и J. W. Mellors, «HIV-1 DNA Decay Dynamics in Blood During More Than a Decade of Suppressive Antiretroviral Therapy.» Clin Infect Dis, 2014, 59, 1312–1321. [CrossRef]
- M. F. Oliveira, A. Pankow, T. Vollbrecht, N. M. Kumar, G. Cabalero, C. Ignacio, M. Zhao, A. Vitomirov, B. Gouaux, M. Nakawawa, B. Murrell, R. J. Ellis и S. Gianella, «Evaluation of Archival HIV DNA in Brain and Lymphoid Tissues.» J Virol, 2023, 97, e00543–23. [CrossRef]
- E. W. Fiebig, D. J. Wright, B. D. Rawal, P. E. Garrett, R. T. Schumacher, L. Peddada, C. Heldebrant, R. Smith, A. Conrad, S. H. Kleinman и M. P. Busch, «Dynamics of HIV viremia and antibody seroconversion in plasma donors: implications for diagnosis and staging of primary HIV infection.» AIDS, 2003, 17, 1871–1879. [CrossRef]
- J. Ananworanich, N. Chomont, L. A. Eller, E. Kroon, S. Tovanabutra, M. Bose, M. Nau, J. L. K. Fletcher, S. Tipsuk, C. Vandergeeten, R. J. O’Connell, S. Pinyakorn, N. Michael, N. Phanuphak и M. L. Robb, «HIV DNA Set Point is Rapidly Established in Acute HIV Infection and Dramatically Reduced by Early ART.» EBioMedicine, 2016, 11, 68–72. [CrossRef]
- K. J. Kwon, A. E. Timmons, S. Sengupta, F. R. Simonetti, H. Zhang, R. Hoh, S. G. Deeks, J. D. Siliciano и R. F. Siliciano, «Different human resting memory CD4 + T cell subsets show similar low inducibility of latent HIV-1 proviruses.» Sci Transl Med, 2020, 12, eaax6795. [CrossRef]
- C. Gálvez, J. Grau-Expósito, V. Urrea, B. Clotet, V. Falcó, M. J. Buzón и J. Martinez-Picado, «Atlas of the HIV-1 Reservoir in Peripheral CD4 T Cells of Individuals on Successful Antiretroviral Therapy.» mBio, 2021, 12, e03078-21. [CrossRef]
- M. J. Buzon, E. Martin-Gayo, F. Pereyra, Z. Ouyang, H. Sun, J. Z. Li, M. Piovoso, A. Shaw, J. Dalmau, N. Zangger, J. Martinez-Picado, R. Zurakowski, X. G. Yu, A. Telenti, B. D. Walker, E. S. Rosenberg и M. Lichterfeld, «Long-Term Antiretroviral Treatment Initiated at Primary HIV-1 Infection Affects the Size, Composition, and Decay Kinetics of the Reservoir of HIV-1-Infected CD4 T Cells.» J Virol, 2014, 88, 10056–10065. [CrossRef]
- B. Hiener, B. A. Horsburgh, J.-S. Eden, K. Barton, T. E. Schlub, E. Lee, S. von Stockenstrom, L. Odevall, J. M. Milush, T. Liegler, E. Sinclair, R. Hoh, E. A. Boritz, D. Douek, R. Fromentin, N. Chomont, S. G. Deeks, F. M. Hecht и S. Palmer, «Identification of Genetically Intact HIV-1 Proviruses in Specific CD4 + T Cells from Effectively Treated Participants.» Cell Rep, 2017, 21, 813–822. [CrossRef]
- N. Soriano-Sarabia, N. M. Archin, R. Bateson, N. P. Dahl, A. M. Crooks, J. D. Kuruc, C. Garrido и D. M. Margolis, «Peripheral Vγ9Vδ2 T Cells Are a Novel Reservoir of Latent HIV Infection.» PLOS Pathog, 2015, 11, e1005201. [CrossRef]
- H. Sun, D. Kim, X. Li, M. Kiselinova, Z. Ouyang, L. Vandekerckhove, H. Shang, E. S. Rosenberg, X. G. Yu и M. Lichterfeld, «Th1/17 Polarization of CD4 T Cells Supports HIV-1 Persistence during Antiretroviral Therapy.» J Virol, 2015, 89, 11284–11293. [CrossRef]
- M. Perreau, A.-L. Savoye, E. De Crignis, J.-M. Corpataux, R. Cubas, E. K. Haddad, L. De Leval, C. Graziosi и G. Pantaleo, «Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production.» J Exp Med, 2012, 210, 143–156. [CrossRef]
- V. H. Wu, J. M. L. Nordin, S. Nguyen, J. Joy, F. Mampe, P. M. del Rio Estrada, F. Torres-Ruiz, M. González-Navarro, Y. A. Luna-Villalobos, S. Ávila-Ríos, G. Reyes-Terán, P. Tebas, L. J. Montaner, K. J. Bar, L. A. Vella и M. R. Betts, «Profound phenotypic and epigenetic heterogeneity of the HIV-1-infected CD4+ T cell reservoir.» Nat Immunol, 2022, 24, 359–370. [CrossRef]
- A. Alexaki, Y. Liu и B. Wigdahl, «Cellular Reservoirs of HIV-1 and their Role in Viral Persistence.» Curr HIV Res, 2008, 6, 388–400. [CrossRef]
- R. T. Veenhuis, C. M. Abreu, P. A. G. Costa, E. A. Ferreira, J. Ratliff, L. Pohlenz, E. N. Shirk, L. H. Rubin, J. N. Blankson, L. Gama и J. E. Clements, «Monocyte-derived macrophages contain persistent latent HIV reservoirs.» Nat Microbiol, 2023, 8, 833–844. [CrossRef]
- S. Swingler, A. M. Mann, J. Zhou, C. Swingler и M. Stevenson, «Apoptotic Killing of HIV-1–Infected Macrophages Is Subverted by the Viral Envelope Glycoprotein.» PLoS Pathog, 2007, 3, e134. [CrossRef]
- C. Vérollet, S. Souriant, E. Bonnaud, P. Jolicoeur, B. Raynaud-Messina, C. Kinnaer, I. Fourquaux, A. Imle, S. Benichou, O. T. Fackler, R. Poincloux и I. Maridonneau-Parini, «HIV-1 reprograms the migration of macrophages.» Blood, 2015, 125, 1611–1622. [CrossRef]
- L. Ene, S. M. Ruta, «How much do antiretroviral drugs penetrate into the central nervous system?.», J Med Life., 2011, 4, 432–439.
- R. K. Heaton, D. B. Clifford, D. R. Franklin, S. P. Woods, C. Ake, F. Vaida, R. J. Ellis, S. L. Letendre, T. D. Marcotte, J. H. Atkinson, M. Rivera-Mindt, O. R. Vigil, M. J. Taylor, A. C. Collier, C. M. Marra, B. B. Gelman, J. C. McArthur, S. Morgello, D. M. Simpson, J. A. McCutchan, I. Abramson, A. Gamst, C. Fennema-Notestine, T. L. Jernigan, J. Wong и I. Grant, «HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study.» Neurology, 2010, 75, 2087–2096. [CrossRef]
- M. K. Ash, L. Al-Harthi и J. R. Schneider, «HIV in the Brain: Identifying Viral Reservoirs and Addressing the Challenges of an HIV Cure.» Vaccines (Basel), 2021, 9, 867. [CrossRef]
- V. Lutgen, S. D. Narasipura, H. J. Barbian, M. Richards, J. Wallace, R. Razmpour, T. Buzhdygan, S. H. Ramirez, L. Prevedel, E. A. Eugenin и L. Al-Harthi, «HIV infects astrocytes in vivo and egresses from the brain to the periphery.» PLOS Pathog, 2020, 16, e1008381. [CrossRef]
- Y. Tang, A. Chaillon, S. Gianella, L. M. Wong, D. Li, T. L. Simermeyer, M. Porrachia, C. Ignacio, B. Woodworth, D. Zhong, J. Du, E. de la Parra Polina, J. Kirchherr, B. Allard, M. L. Clohosey, M. Moeser, A. L. Sondgeroth, G. D. Whitehill, V. Singh, A. Dashti, D. M. Smith, J. J. Eron, K. J. Bar, A. Chahroudi, S. B. Joseph, N. M. Archin, D. M. Margolis и G. Jiang, «Brain microglia serve as a persistent HIV reservoir despite durable antiretroviral therapy.» Journal of Clinical Investigation, 2023, 133, e167417. [CrossRef]
- K. A. Thompson, C. L. Cherry, J. E. Bell и C. A. McLean, «Brain Cell Reservoirs of Latent Virus in Presymptomatic HIV-Infected Individuals.» Am J Pathol, 2011, 179, 1623–1629. [CrossRef]
- J. E. King, E. A. Eugenin, C. M. Buckner и J. W. Berman, «HIV tat and neurotoxicity.» Microbes Infect, 2006, 8, 1347–1357. [CrossRef]
- M. K. Mamik, E. Hui, W. G. Branton, B. A. McKenzie, J. Chisholm, E. A. Cohen и C. Power, «HIV-1 Viral Protein R Activates NLRP3 Inflammasome in Microglia: implications for HIV-1 Associated Neuroinflammation.» J Neuroimmune Pharmacol, 2016, 12, 233–248. [CrossRef]
- S. Dahmani, N. Kaliss, J. W. VanMeter, D. J. Moore, R. J. Ellis и X. Jiang, «Alterations of Brain Metabolites in Adults With HIV: A Systematic Meta-analysis of Magnetic Resonance Spectroscopy Studies.» Neurology, 2021, 97, e1085–e1096. [CrossRef]
- A. Alakkas, R. J. Ellis, C. W.-M. Watson, A. Umlauf, R. K. Heaton, S. Letendre, A. Collier, C. Marra, D. B. Clifford, B. Gelman, N. Sacktor, S. Morgello, D. Simpson, J. A. McCutchan, A. Kallianpur, S. Gianella, T. Marcotte, I. Grant и C. Fennema-Notestine, «White matter damage, neuroinflammation, and neuronal integrity in HAND.» J NeuroVirol, 2018, 25, 32–41. [CrossRef]
- M. S. Dahabieh, M. Ooms, V. Simon и I. Sadowski, «A Doubly Fluorescent HIV-1 Reporter Shows that the Majority of Integrated HIV-1 Is Latent Shortly after Infection.» J Virol, 2013, 87, 4716–4727. [CrossRef]
- W. J. Swiggard, C. Baytop, J. J. Yu, J. Dai, C. Li, R. Schretzenmair, T. Theodosopoulos и U. O’Doherty, «Human Immunodeficiency Virus Type 1 Can Establish Latent Infection in Resting CD4+T Cells in the Absence of Activating Stimuli.» J Virol, 2005, 79, 14179–14188. [CrossRef]
- S. D. Rezaei, H. K. Lu, J. J. Chang, A. Rhodes, S. R. Lewin и P. U. Cameron, «The Pathway To Establishing HIV Latency Is Critical to How Latency Is Maintained and Reversed.» J Virol, 2018, 92, e02225–17. [CrossRef]
- L. Chavez, V. Calvanese и E. Verdin, «HIV Latency Is Established Directly and Early in Both Resting and Activated Primary CD4 T Cells.» PLOS Pathog, 2015, 11, e1004955. [CrossRef]
- Y. Meng, J. Zhong, Y. Lv и W. Zou, «Research progress on HIV-1 immune escape mechanisms.» AIDS Rev, 2022, 24, 133–138. [CrossRef]
- A. O. Pasternak и B. Berkhout, «HIV persistence: silence or resistance?.» Curr Opin Virol, 2023, 59, 101301. [CrossRef]
- L. Sarmati, G. D’Ettorre, S. Parisi и M. Andreoni, «HIV Replication at Low Copy Number and its Correlation with the HIV Reservoir: A Clinical Perspective.» Curr HIV Res, 2015, 13, 250–257. [CrossRef]
- M. C. Virgilio и K. L. Collins, «The Impact of Cellular Proliferation on the HIV-1 Reservoir.» Viruses, 2020, 12, 127. [CrossRef]
- N. Chomont, M. El-Far, P. Ancuta, L. Trautmann, F. A. Procopio, B. Yassine-Diab, G. Boucher, M.-R. Boulassel, G. Ghattas, J. M. Brenchley, T. W. Schacker, B. J. Hill, D. C. Douek, J.-P. Routy, E. K. Haddad и R.-P. Sékaly, «HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation.» Nat Med, 2009, 15, 893–900. [CrossRef]
- P. Mendoza, J. R. Jackson, T. Y. Oliveira, C. Gaebler, V. Ramos, M. Caskey, M. Jankovic, M. C. Nussenzweig и L. B. Cohn, «Antigen-responsive CD4+ T cell clones contribute to the HIV-1 latent reservoir.» J Exp Med, 2020, 217, e20200051. [CrossRef]
- F. Maldarelli, X. Wu, L. Su, F. R. Simonetti, W. Shao, S. Hill, J. Spindler, A. L. Ferris, J. W. Mellors, M. F. Kearney, J. M. Coffin и S. H. Hughes, «Specific HIV integration sites are linked to clonal expansion and persistence of infected cells.» Science, 2014, 345, 179–183. [CrossRef]
- F. R. Simonetti, J. A. White, C. Tumiotto, K. D. Ritter, M. Cai, R. T. Gandhi, S. G. Deeks, B. J. Howell, L. J. Montaner, J. N. Blankson, A. Martin, G. M. Laird, R. F. Siliciano, J. W. Mellors и J. D. Siliciano, «Intact proviral DNA assay analysis of large cohorts of people with HIV provides a benchmark for the frequency and composition of persistent proviral DNA.» Proc Natl Acad Sci U S A, 2020, 117, 18692–18700. [CrossRef]
- J. Z. Li, M. Melberg, A. Kittilson, M. Abdel-Mohsen, Y. Li, E. Aga, R. J. Bosch, E. R. Wonderlich, J. Kinslow, L. B. Giron, C. Di Germanio, M. Pilkinton, L. MacLaren, M. Keefer, L. Fox, L. Barr, E. Acosta, J. Ananworanich, R. Coombs, J. Mellors, S. Deeks, R. T. Gandhi, M. Busch, A. Landay, B. Macatangay и D. M. Smith, «Predictors of HIV rebound differ by timing of antiretroviral therapy initiation.» JCI Insight, 2024, 9, e173864. [CrossRef]
- K. M. Bruner, A. J. Murray, R. A. Pollack, M. G. Soliman, S. B. Laskey, A. A. Capoferri, J. Lai, M. C. Strain, S. M. Lada, R. Hoh, Y.-C. Ho, D. D. Richman, S. G. Deeks, J. D. Siliciano и R. F. Siliciano, «Defective proviruses rapidly accumulate during acute HIV-1 infection.» Nat Med, 2016, 22, 1043–1049. [CrossRef]
- A. E. Baxter, U. O’Doherty и D. E. Kaufmann, «Beyond the replication-competent HIV reservoir: transcription and translation-competent reservoirs.» Retrovirology, 2018, 15, 18. [CrossRef]
- H. Imamichi, R. L. Dewar, J. W. Adelsberger, C. A. Rehm, U. O’Doherty, E. E. Paxinos, A. S. Fauci и H. C. Lane, «Defective HIV-1 proviruses produce novel protein-coding RNA species in HIV-infected patients on combination antiretroviral therapy.» Proc Natl Acad Sci U S A, 2016, 113, 8783–8788. [CrossRef]
- M. El-Far, R. Halwani, E. Said, L. Trautmann, M. Doroudchi, L. Janbazian, S. Fonseca, J. van Grevenynghe, B. Yassine-Diab, R.-P. Sékaly и E. K. Haddad, «T-cell exhaustion in HIV infection.» Curr HIV/AIDS Rep, 2008, 5, 13–19. [CrossRef]
- V. Avettand-Fènoël, L. Hocqueloux, J. Ghosn, A. Cheret, P. Frange, A. Melard, J.-P. Viard и C. Rouzioux, «Total HIV-1 DNA, a Marker of Viral Reservoir Dynamics with Clinical Implications.» Clin Microbiol Rev, 2016, 29, 859–880. [CrossRef]
- J. P. Williams, J. Hurst, W. Stöhr, N. Robinson, H. Brown, M. Fisher, S. Kinloch, D. Cooper, M. Schechter, G. Tambussi, S. Fidler, M. Carrington, A. Babiker, J. Weber, K. K. Koelsch, A. D. Kelleher, R. E. Phillips и J. Frater, «HIV-1 DNA predicts disease progression and post-treatment virological control.» eLife, 2014, 3, e03821. [CrossRef]
- C. Goujard, M. Bonarek, L. Meyer, F. Bonnet, M.-L. Chaix, C. Deveau, M. Sinet, J. Galimand, J.-F. Delfraissy, A. Venet, C. Rouzioux и P. Morlat, «CD4 Cell Count and HIV DNA Level Are Independent Predictors of Disease Progression after Primary HIV Type 1 Infection in Untreated Patients.» Clin Infect Dis, 2006, 42, 709–715. [CrossRef]
- C. Rouzioux, J. Hubert, M. Burgard, C. Deveau, C. Goujard, M. Bary, D. Séréni, J. Viard, J. Delfraissy и L. Meyer, «Early Levels of HIV-1 DNA in Peripheral Blood Mononuclear Cells Are Predictive of Disease Progression Independently of HIV-1 RNA Levels and CD4+T Cell Counts.» J Infect Dis, 2005, 192, 46–55. [CrossRef]
- C. G. Tsiara, G. K. Nikolopoulos, P. G. Bagos, C. Goujard, T. L. Katzenstein, A. K. Minga, C. Rouzioux и A. Hatzakis, «Impact of HIV Type 1 DNA Levels on Spontaneous Disease Progression: A Meta-Analysis.» AIDS Res Hum Retroviruses, 2012, 28, 366–373. [CrossRef]
- J. Ananworanich, A. Schuetz, C. Vandergeeten, I. Sereti, M. de Souza, R. Rerknimitr, R. Dewar, M. Marovich, F. van Griensven, R. Sekaly, S. Pinyakorn, N. Phanuphak, R. Trichavaroj, W. Rutvisuttinunt, N. Chomchey, R. Paris, S. Peel, V. Valcour, F. Maldarelli, N. Chomont, N. Michael, P. Phanuphak и J. H. Kim, «Impact of Multi-Targeted Antiretroviral Treatment on Gut T Cell Depletion and HIV Reservoir Seeding during Acute HIV Infection.» PLoS ONE, 2012, 7, e33948. [CrossRef]
- B. Hoen, D. A. Cooper, F. C. Lampe, L. Perrin, N. Clumeck, A. N. Phillips, L.-E. Goh, S. Lindback, D. Sereni, B. Gazzard, J. Montaner, H.-J. Stellbrink, A. Lazzarin, D. Ponscarme, S. Staszewski, L. Mathiesen, D. Smith, R. Finlayson, R. Weber, L. Wegmann, G. Janossy и S. Kinloch-de Loes, «Predictors of Virological Outcome and Safety in Primary HIV Type 1-Infected Patients Initiating Quadruple Antiretroviral Therapy: QUEST GW PROB3005.» Clin Infect Dis, 2007, 45, 381–390. [CrossRef]
- K. Koelsch, L. Liu, R. Haubrich, S. May, D. Havlir, H. Günthard, C. Ignacio, P. Campos-Soto, S. Little, R. Shafer, G. Robbins, R. D’Aquila, Y. Kawano, K. Young, P. Dao, C. Spina, D. Richman и J. Wong, «Dynamics of Total, Linear Nonintegrated, and Integrated HIV-1 DNA In Vivo and In Vitro.» J Infect Dis, 2008, 197, 411–419. [CrossRef]
- L. Leyre, E. Kroon, C. Vandergeeten, C. Sacdalan, D. J. Colby, S. Buranapraditkun, A. Schuetz, N. Chomchey, M. de Souza, W. Bakeman, R. Fromentin, S. Pinyakorn, S. Akapirat, R. Trichavaroj, S. Chottanapund, S. Manasnayakorn, R. Rerknimitr, P. Wattanaboonyoungcharoen, J. H. Kim, S. Tovanabutra, T. W. Schacker, R. O’Connell, V. G. Valcour, P. Phanuphak, M. L. Robb, N. Michael, L. Trautmann, N. Phanuphak, J. Ananworanich и N. Chomont, «Abundant HIV-infected cells in blood and tissues are rapidly cleared upon ART initiation during acute HIV infection.» Sci Transl Med, 2020, 12, eaav3491. [CrossRef]
- T. F. Leite, E. Delatorre, F. H. Côrtes, A. C. G. Ferreira, S. W. Cardoso, B. Grinsztejn, M. M. de Andrade, V. G. Veloso, M. G. Morgado и M. L. Guimarães, «Reduction of HIV-1 Reservoir Size and Diversity After 1 Year of cART Among Brazilian Individuals Starting Treatment During Early Stages of Acute Infection» Front Microbiol, 2019, 10, 145. [CrossRef]
- F. Groot, T. M. M. van Capel, J. H. N. Schuitemaker, B. Berkhout и E. C. de Jong, «Differential susceptibility of naïve, central memory and effector memory T cells to dendritic cell-mediated HIV-1 transmission.» Retrovirology, 2006, 3, 52. [CrossRef]
- M. J. Peluso, P. Bacchetti, K. D. Ritter, S. Beg, J. Lai, J. N. Martin, P. W. Hunt, T. J. Henrich, J. D. Siliciano, R. F. Siliciano, G. M. Laird и S. G. Deeks, «Differential decay of intact and defective proviral DNA in HIV-1–infected individuals on suppressive antiretroviral therapy.» JCI Insight, 2020, 5, e132997. [CrossRef]
- B. S. Taylor, M. E. Sobieszczyk, F. E. McCutchan и S. M. Hammer, «The Challenge of HIV-1 Subtype Diversity.» N Engl J Med, 2008, 358, 1590–1602. [CrossRef]
- P. Kanki, D. Hamel, J. Sankalé, C. Hsieh, I. Thior, F. Barin, S. Woodcock, A. Guèye-Ndiaye, E. Zhang, M. Montano, T. Siby, R. Marlink, I. NDoye, M. Essex и S. MBoup, «Human Immunodeficiency Virus Type 1 Subtypes Differ in Disease Progression.» J Infect Dis, 1999, 179, 68–73. [CrossRef]
- P. N. Amornkul, E. Karita, A. Kamali, W. N. Rida, E. J. Sanders, S. Lakhi, M. A. Price, W. Kilembe, E. Cormier, O. Anzala, M. H. Latka, L.-G. Bekker, S. A. Allen, J. Gilmour и P. E. Fast, «Disease progression by infecting HIV-1 subtype in a seroconverter cohort in sub-Saharan Africa.» AIDS, 2013, 27, 2775–2786. [CrossRef]
- D. Ssemwanga, R. N. Nsubuga, B. N. Mayanja, F. Lyagoba, B. Magambo, D. Yirrell, L. Van der Paal, H. Grosskurth и P. Kaleebu, «Effect of HIV-1 Subtypes on Disease Progression in Rural Uganda: A Prospective Clinical Cohort Study.» PLoS ONE, 2013, 8, e71768. [CrossRef]
- F. H. Omondi, S. Chandrarathna, S. Mujib, C. J. Brumme, S. W. Jin, H. Sudderuddin, R. L. Miller, A. Rahimi, O. Laeyendecker, P. Bonner, F. Y. Yue, E. Benko, C. M. Kovacs, M. A. Brockman, M. Ostrowski и Z. L. Brumme, «HIV Subtype and Nef-Mediated Immune Evasion Function Correlate with Viral Reservoir Size in Early-Treated Individuals.» J Virol, 2019, 93, e01832–18. [CrossRef]
- N. Bachmann, C. von Siebenthal, V. Vongrad, T. Turk, K. Neumann, N. Beerenwinkel, J. Bogojeska, J. Fellay, V. Roth, Y. L. Kok, C. W. Thorball, A. Borghesi, S. Parbhoo, M. Wieser, J. Böni, M. Perreau, T. Klimkait, S. Yerly, M. Battegay, A. Rauch, M. Hoffmann, E. Bernasconi, M. Cavassini, R. D. Kouyos, H. F. Günthard, K. J. Metzner, A. Anagnostopoulos, M. Battegay, E. Bernasconi, J. Böni, D. L. Braun, H. C. Bucher, A. Calmy, M. Cavassini, A. Ciuffi, G. Dollenmaier, M. Egger, L. Elzi, J. Fehr, J. Fellay, H. Furrer, C. A. Fux, H. F. Günthard, D. Haerry, B. Hasse, H. H. Hirsch, M. Hoffmann, I. Hösli, M. Huber, C. Kahlert, L. Kaiser, O. Keiser, T. Klimkait, R. D. Kouyos, H. Kovari, B. Ledergerber, G. Martinetti, B. M. d. Tejada, C. Marzolini, K. J. Metzner, N. Müller, D. Nicca, P. Paioni, G. Pantaleo, M. Perreau, A. Rauch, C. Rudin, A. U. Scherrer, P. Schmid, R. Speck, M. Stöckle, P. Tarr, A. Trkola, P. Vernazza, G. Wandeler, R. Weber и S. Yerly, «Determinants of HIV-1 reservoir size and long-term dynamics during suppressive ART.» Nat Commun, 2019, 10, 3193. [CrossRef]
- R. E. Jeeninga, M. Hoogenkamp, M. Armand-Ugon, M. de Baar, K. Verhoef и B. Berkhout, «Functional Differences between the Long Terminal Repeat Transcriptional Promoters of Human Immunodeficiency Virus Type 1 Subtypes A through G.» J Virol, 2000, 74, 3740–3751. [CrossRef]
- J. K. Mann, H. Byakwaga, X. T. Kuang, A. Q. Le, C. J. Brumme, P. Mwimanzi, S. Omarjee, E. Martin, G. Q. Lee, B. Baraki, R. Danroth, R. McCloskey, C. Muzoora, D. R. Bangsberg, P. W. Hunt, P. J. R. Goulder, B. D. Walker, P. R. Harrigan, J. N. Martin, T. Ndung’u, M. A. Brockman и Z. L. Brumme, «Ability of HIV-1 Nef to downregulate CD4 and HLA class I differs among viral subtypes.» Retrovirology, 2013, 10, 100. [CrossRef]
- I. Lisovsky, S. M. Schader, R. D. Sloan, M. Oliveira, D. Coutsinos, N. F. Bernard и M. A. Wainberg, «HIV-1 Subtype Variability in Vif Derived from Molecular Clones Affects APOBEC3G-Mediated Host Restriction.» Intervirology, 2013, 56, 258–264. [CrossRef]
- S. Langer, C. Hammer, K. Hopfensperger, L. Klein, D. Hotter, P. D. De Jesus, K. M. Herbert, L. Pache, N. Smith, J. A. van der Merwe, S. K. Chanda, J. Fellay, F. Kirchhoff и D. Sauter, «HIV-1 Vpu is a potent transcriptional suppressor of NF-κB-elicited antiviral immune responses.» eLife, 2019, 8, e41930. [CrossRef]
- S. G. Deeks и B. D. Walker, «Human Immunodeficiency Virus Controllers: Mechanisms of Durable Virus Control in the Absence of Antiretroviral Therapy.» Immunity, 2007, 27, 406–416. [CrossRef]
- B. D. Kennedy, J. Blazkova, J. S. Justement, V. Shi, M. A. Rai, M. R. Manning, L. Praiss, K. Gittens, P. A. Wender, S. Patro, X. Wu, S. Moir и T.-W. Chun, «Comprehensive analysis of HIV reservoirs in elite controllers.» J Clin Invest, 2023, 133, e165446. [CrossRef]
- A. K. R. Kwaa, C. C. Garliss, K. D. Ritter, G. M. Laird и J. N. Blankson, «Elite suppressors have low frequencies of intact HIV-1 proviral DNA.» AIDS, 2020, 34, 641–643. [CrossRef]
- A. Sáez-Cirión, C. Bacchus, L. Hocqueloux, V. Avettand-Fenoel, I. Girault, C. Lecuroux, V. Potard, P. Versmisse, A. Melard, T. Prazuck, B. Descours, J. Guergnon, J.-P. Viard, F. Boufassa, O. Lambotte, C. Goujard, L. Meyer, D. Costagliola, A. Venet, G. Pancino, B. Autran и C. Rouzioux, «Post-Treatment HIV-1 Controllers with a Long-Term Virological Remission after the Interruption of Early Initiated Antiretroviral Therapy ANRS VISCONTI Study.» PLoS Pathog, 2013, 9, e1003211. [CrossRef]
- A. Gálvez, V. Urrea, M. d. C. Garcia-Guerrero, S. Bernal, S. Benet, B. Mothe, L. Bailón, J. Dalmau, A. Martinez, A. Nieto, L. Leal, F. García, B. Clotet, J. Martinez-Picado и M. Salgado, «Altered T-cell subset distribution in the viral reservoir in HIV-1-infected individuals with extremely low proviral DNA (LoViReTs).» J Intern Med, 2022, 292, 308–320. [CrossRef]
- I. M. Bruner, Z. Wang, F. R. Simonetti, A. M. Bender, K. J. Kwon, S. Sengupta, E. J. Fray, S. A. Beg, A. A. R. Antar, K. M. Jenike, L. N. Bertagnolli, A. A. Capoferri, J. T. Kufera, A. Timmons, C. Nobles, J. Gregg, N. Wada, Y.-C. Ho, H. Zhang, J. B. Margolick, J. N. Blankson, S. G. Deeks, F. D. Bushman, J. D. Siliciano, G. M. Laird и R. F. Siliciano, «A quantitative approach for measuring the reservoir of latent HIV-1 proviruses.» Nature, 2019, 566, 120–125. [CrossRef]
- M. Delporte, W. M. Delporte, W. van Snippenberg, E. E. Blomme, S. Rutsaert, M. Verschoore, E. De Smet, M.-A. De Scheerder, S. Gerlo, L. Vandekerckhove и W. Trypsteen, «Integrative assessment of total and intact HIV-1 reservoir by a five-region multiplexed Rainbow digital PCR assay.» bioRxiv, 2023, 553846. [CrossRef]
- S. Rutsaert, K. Bosman, W. Trypsteen, M. Nijhuis и L. Vandekerckhove, «Digital PCR as a tool to measure HIV persistence.» Retrovirology, 2018, 15, 16. [CrossRef]
- I. J. Bosman, M. Nijhuis, P. M. van Ham, A. M. J. Wensing, K. Vervisch, L. Vandekerckhove и W. De Spiegelaere, «Comparison of digital PCR platforms and semi-nested qPCR as a tool to determine the size of the HIV reservoir.» Sci Rep, 2015, 5, 1381. [CrossRef]
- I. B. Einkauf, G. Q. Lee, C. Gao, R. Sharaf, X. Sun, S. Hua, S. M. Y. Chen, C. Jiang, X. Lian, F. Z. Chowdhury, E. S. Rosenberg, T.-W. Chun, J. Z. Li, X. G. Yu и M. Lichterfeld, «Intact HIV-1 proviruses accumulate at distinct chromosomal positions during prolonged antiretroviral therapy.» J Clin Invest, 2019, 129, 988–998. [CrossRef]
- P. N. Enick, J. P. Brooker, C. M. Tumiotto, B. T. Staines, J. J. Eron, D. K. McMahon, R. T. Gandhi, J. W. Mellors и M. D. Sobolewski, «Comparison of methods to quantify inducible HIV-1 outgrowth.» J Virus Erad, 2021, 7, 100043. [CrossRef]
- Wagner TA, McLaughlin S, Garg K, Cheung CY, Larsen BB, Styrchak S, Huang HC, Edlefsen PT, Mullins JI, Frenkel LM. «Proliferation of cells with HIV integrated into cancer genes contributes to persistent infection.» Science, 2014, 345, 570–573(2014). [CrossRef]
- Paruzynski, A. , Arens, A. , Gabriel, R., Bartholomae, C. C., Scholz, S., Wang, W., Wolf, S., Glimm, H., Schmidt, M., & von Kalle, C. «Genome-wide high-throughput integrome analyses by nrLAM-PCR and next-generation sequencing». Nat Protoc, 2010, 5, 1379–1395. [Google Scholar] [CrossRef]
- I. Markowitz, T. H. Evering, D. Garmon, M. Caskey, M. La Mar, K. Rodriguez, V. Sahi, S. Palmer, N. Prada и H. Mohri, «A Randomized Open-Label Study of 3- Versus 5-Drug Combination Antiretroviral Therapy in Newly HIV-1–Infected Individuals.» J Acquir Immune Defic Syndr, 2014, 66, 140–147. [CrossRef]
- G. Wang, N. Zhao, B. Berkhout и A. T. Das, «CRISPR-Cas based antiviral strategies against HIV-1.» Virus Res, 2018, 244, 321–332. [CrossRef]
- Y. Kim, J. L. Anderson и S. R. Lewin, «Getting the “Kill” into “Shock and Kill”: Strategies to Eliminate Latent HIV.» Cell Host Microbe, 2018, 23, 14–26. [CrossRef]
- G. Vansant, A. Bruggemans, J. Janssens и Z. Debyser, «Block-And-Lock Strategies to Cure HIV Infection.» Viruses, 2020, 12, 84. [CrossRef]
- R. Barrangou и L. Marraffini, «CRISPR-Cas Systems: Prokaryotes Upgrade to Adaptive Immunity.» Mol Cell, 2014, 54, 234–244. [CrossRef]
- I. Fan, B. Berkhout и E. Herrera-Carrillo, «A combinatorial CRISPR-Cas12a attack on HIV DNA.» Mol Ther Methods Clin Dev, 2022, 25, 43–51. [CrossRef]
- H. Nguyen, H. Wilson, S. Jayakumar, V. Kulkarni и S. Kulkarni, «Efficient Inhibition of HIV Using CRISPR/Cas13d Nuclease System.» Viruses, 2021, 13, 1850. [CrossRef]
- W. Zhu, R. Lei, Y. Le Duff, J. Li, F. Guo, M. A. Wainberg и C. Liang, «The CRISPR/Cas9 system inactivates latent HIV-1 proviral DNA.» Retrovirology, 2015, 12, 22. [CrossRef]
- Z. Gao, M. Fan, A. T. Das, E. Herrera-Carrillo и B. Berkhout, «Extinction of all infectious HIV in cell culture by the CRISPR-Cas12a system with only a single crRNA.» Nucleic Acids Res, 2020, 48, 5527–5539. [CrossRef]
- G. Wang, N. Zhao, B. Berkhout и A. T. Das, «A Combinatorial CRISPR-Cas9 Attack on HIV-1 DNA Extinguishes All Infectious Provirus in Infected T Cell Cultures.» Cell Rep, 2016, 17, 2819–2826. [CrossRef]
- Y. Ophinni, M. Inoue, T. Kotaki и M. Kameoka, «CRISPR/Cas9 system targeting regulatory genes of HIV-1 inhibits viral replication in infected T-cell cultures.» Sci Rep, 2018, 8, 7784. [CrossRef]
- Q. Xiao, D. Guo и S. Chen, «Application of CRISPR/Cas9-Based Gene Editing in HIV-1/AIDS Therapy.» Front Cell Infect Microbiol, 2019, 9, 69. [CrossRef]
- Z. Wang, Q. Pan, P. Gendron, W. Zhu, F. Guo, S. Cen, M. Wainberg и C. Liang, «CRISPR/Cas9-Derived Mutations Both Inhibit HIV-1 Replication and Accelerate Viral Escape.» Cell Rep, 2016, 15, 481–489. [CrossRef]
- G. Wang, N. Zhao, B. Berkhout и A. T. Das, «CRISPR-Cas9 Can Inhibit HIV-1 Replication but NHEJ Repair Facilitates Virus Escape.» Mol Ther, 2016, 24, 522–526. [CrossRef]
- S. G. Deeks, «Shock and kill.» Nature, 2012, 487, 439–440. [CrossRef]
- I. M. Archin, J. L. Kirchherr, J. A. M. Sung, G. Clutton, K. Sholtis, Y. Xu, B. Allard, E. Stuelke, A. D. Kashuba, J. D. Kuruc, J. Eron, C. L. Gay, N. Goonetilleke и D. M. Margolis, «Interval dosing with the HDAC inhibitor vorinostat effectively reverses HIV latency.» J Clin Invest, 2017, 127, 3126–3135. [CrossRef]
- J. H. Elliott, F. Wightman, A. Solomon, K. Ghneim, J. Ahlers, M. J. Cameron, M. Z. Smith, T. Spelman, J. McMahon, P. Velayudham, G. Brown, J. Roney, J. Watson, M. H. Prince, J. F. Hoy, N. Chomont, R. Fromentin, F. A. Procopio, J. Zeidan, S. Palmer, L. Odevall, R. W. Johnstone, B. P. Martin, E. Sinclair, S. G. Deeks, D. J. Hazuda, P. U. Cameron, R.-P. Sékaly и S. R. Lewin, «Activation of HIV Transcription with Short-Course Vorinostat in HIV-Infected Patients on Suppressive Antiretroviral Therapy.» PLoS Pathog, 2014, 10, e1004473. [CrossRef]
- S. Bouchat, J.-S. Gatot, K. Kabeya, C. Cardona, L. Colin, G. Herbein, S. De Wit, N. Clumeck, O. Lambotte, C. Rouzioux, O. Rohr и C. Van Lint, «Histone methyltransferase inhibitors induce HIV-1 recovery in resting CD4+ T cells from HIV-1-infected HAART-treated patients.» AIDS, 2012, 26, 1473–1482. [CrossRef]
- G. Li, Z. Zhang, N. Reszka-Blanco, F. Li, L. Chi, J. Ma, J. Jeffrey, L. Cheng и L. Su, «Specific ActivationIn Vivoof HIV-1 by a Bromodomain Inhibitor from Monocytic Cells in Humanized Mice under Antiretroviral Therapy.» J Virol, 2019, 93, e00233–19. [CrossRef]
- A. Gutiérrez, S. Serrano-Villar, N. Madrid-Elena, M. J. Pérez-Elías, M. E. Martín, C. Barbas, J. Ruipérez, E. Muñoz, M. A. Muñoz-Fernández, T. Castor и S. Moreno, «Bryostatin-1 for latent virus reactivation in HIV-infected patients on antiretroviral therapy.» AIDS, 2019, 30, 1385–1392. [CrossRef]
- L. Vibholm, M. H. Schleimann, J. F. Højen, T. Benfield, R. Offersen, K. Rasmussen, R. Olesen, A. Dige, J. Agnholt, J. Grau, M. Buzon, B. Wittig, M. Lichterfeld, A. M. Petersen, X. Deng, M. Abdel-Mohsen, S. K. Pillai, S. Rutsaert, W. Trypsteen, W. De Spiegelaere, L. Vandekerchove, L. Østergaard, T. A. Rasmussen, P. W. Denton, M. Tolstrup и O. S. Søgaard, «Short-Course Toll-Like Receptor 9 Agonist Treatment Impacts Innate Immunity and Plasma Viremia in Individuals With Human Immunodeficiency Virus Infection.» Clin Infect Dis, 2017, 64, 1686–1695. [CrossRef]
- L. Shan, K. Deng, N. Shroff, C. Durand, S. Rabi, H.-C. Yang, H. Zhang, J. Margolick, J. Blankson и R. Siliciano, «Stimulation of HIV-1-Specific Cytolytic T Lymphocytes Facilitates Elimination of Latent Viral Reservoir after Virus Reactivation.» Immunity, 2012, 36, 491–501. [CrossRef]
- A. P. Chandrasekar, N. W. Cummins, S. Natesampillai, A. Misra, A. Alto, G. Laird и A. D. Badley, «The BCL-2 Inhibitor Venetoclax Augments Immune Effector Function Mediated by Fas Ligand, TRAIL, and Perforin/Granzyme B, Resulting in Reduced Plasma Viremia and Decreased HIV Reservoir Size during Acute HIV Infection in a Humanized Mouse Model.» J Virol, 2022, 96, e0173022. [CrossRef]
- I. Chugh, B. Bradel-Tretheway, C. M. R. Monteiro-Filho, V. Planelles, S. B. Maggirwar, S. Dewhurst и B. Kim, «Akt inhibitors as an HIV-1 infected macrophage-specific anti-viral therapy.» Retrovirology, 2008, 5, 11. [CrossRef]
- G. R. Campbell и S. A. Spector, «DIABLO/SMAC mimetics selectively kill HIV-1-infected resting memory CD4+ T cells: a potential role in a cure strategy for HIV-1 infection.» Autophagy, 2019, 15, 744–746. [CrossRef]
- I. Li, P. Kaiser, H. W. Lampiris, P. Kim, S. A. Yukl, D. V. Havlir, W. C. Greene и J. K. Wong, «Stimulating the RIG-I pathway to kill cells in the latent HIV reservoir following viral reactivation.» Nat Med, 2016, 22, 807–811. [CrossRef]
- G. Darcis, B. Van Driessche и C. Van Lint, «HIV Latency: Should We Shock or Lock?.» Trends Immunol, 2017, 38, 217–228. [CrossRef]
- A. F. Kessing, C. C. Nixon, C. Li, P. Tsai, H. Takata, G. Mousseau, P. T. Ho, J. B. Honeycutt, M. Fallahi, L. Trautmann, J. V. Garcia и S. T. Valente, «In Vivo Suppression of HIV Rebound by Didehydro-Cortistatin A, a “Block-and-Lock” Strategy for HIV-1 Treatment.» Cell Rep, 2017, 21, 600–611. [CrossRef]
- A. Bruggemans, G. Vansant, M. Balakrishnan, M. L. Mitchell, R. Cai, F. Christ и Z. Debyser, «GS-9822, a Preclinical LEDGIN Candidate, Displays a Block-and-Lock Phenotype in Cell Culture.» Antimicrob Agents Chemother, 2021, 65, e02328–20. [CrossRef]
- M. J. Jean, T. Hayashi, H. Huang, J. Brennan, S. Simpson, A. Purmal, K. Gurova, M. C. Keefer, J. J. Kobie, N. G. Santoso и J. Zhu, «Curaxin CBL0100 Blocks HIV-1 Replication and Reactivation through Inhibition of Viral Transcriptional Elongation.» Front Microbiol, 2017, 8, 2007. [CrossRef]
- I. Anderson, J. S. Low, S. Weston, M. Weinberger, A. Zhyvoloup, A. A. Labokha, G. Corazza, R. A. Kitson, C. J. Moody, A. Marcello и A. Fassati, «Heat shock protein 90 controls HIV-1 reactivation from latency.» Proc Natl Acad Sci U S A, 2014, 111, E1528–37. [CrossRef]
- C. Gavegnano, J. H. Brehm, F. P. Dupuy, A. Talla, S. P. Ribeiro, D. A. Kulpa, C. Cameron, S. Santos, S. J. Hurwitz, V. C. Marconi, J.-P. Routy, L. Sabbagh, R. F. Schinazi и R. P. Sékaly, «Novel mechanisms to inhibit HIV reservoir seeding using Jak inhibitors.» PLOS Pathog, 2017, 13, e1006740. [CrossRef]
- Research Toward a Cure Trials – Treatment Action Group. Available online: https://www.treatmentactiongroup.org/cure/trials/ (23.10.2024).
Figure 1.
Classification of proviral reservoirs. As the main classification criteria, we propose the ability to express viral mRNA and the potential to continue the viral life cycle, which determine the reservoir status in terms of latency. The ability to produce replication-competent virus defines proviral reservoirs as intact or defective, with defective reservoirs further subdivided based on the molecules they produce. Completely incompetent defective proviral reservoirs cannot produce any viral mRNA. Transcriptionally competent defective proviral reservoirs are capable of expressing viral mRNA, but their ability to translate it is limited. Translationally competent defective proviral reservoirs can synthesize HIV proteins, although the assembly of viable virions remains impossible.
Figure 1.
Classification of proviral reservoirs. As the main classification criteria, we propose the ability to express viral mRNA and the potential to continue the viral life cycle, which determine the reservoir status in terms of latency. The ability to produce replication-competent virus defines proviral reservoirs as intact or defective, with defective reservoirs further subdivided based on the molecules they produce. Completely incompetent defective proviral reservoirs cannot produce any viral mRNA. Transcriptionally competent defective proviral reservoirs are capable of expressing viral mRNA, but their ability to translate it is limited. Translationally competent defective proviral reservoirs can synthesize HIV proteins, although the assembly of viable virions remains impossible.
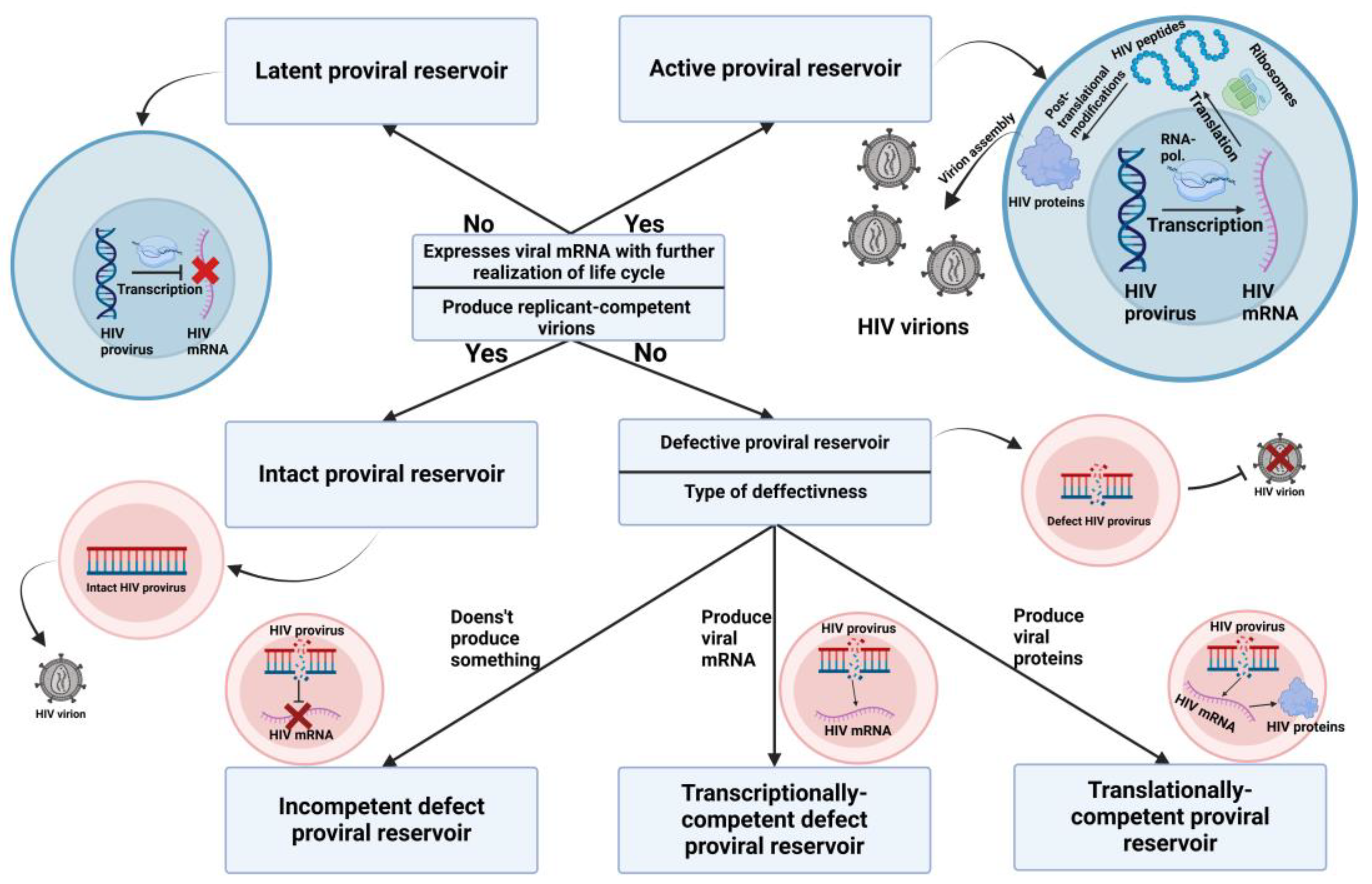
Figure 2.
Mechanisms responsible for the small size of the proviral reservoir. (A) In elite controllers, a strong HIV-specific CD8+ cytotoxic T cell response plays a major role in effectively controlling the size of the proviral reservoir. (B) Post-treatment controllers maintain a small proviral reservoir size through the early initiation of ART. This early treatment facilitates the clearance of active proviral reservoirs and prevents the colonization of large numbers of target cells. (C) The LoViReT phenotype is associated with a smaller proviral reservoir due to the alternative distribution of reservoirs towards short-lived Ttm and Tem populations. These subpopulations have a shorter lifespan and higher activation frequency, leading to immune clearance of the reservoirs. It is also believed that the HIV proviral DNA in this phenotype is less transcriptionally active, preventing the production of HIV virions and the active replenishment of the proviral reservoirs.
Figure 2.
Mechanisms responsible for the small size of the proviral reservoir. (A) In elite controllers, a strong HIV-specific CD8+ cytotoxic T cell response plays a major role in effectively controlling the size of the proviral reservoir. (B) Post-treatment controllers maintain a small proviral reservoir size through the early initiation of ART. This early treatment facilitates the clearance of active proviral reservoirs and prevents the colonization of large numbers of target cells. (C) The LoViReT phenotype is associated with a smaller proviral reservoir due to the alternative distribution of reservoirs towards short-lived Ttm and Tem populations. These subpopulations have a shorter lifespan and higher activation frequency, leading to immune clearance of the reservoirs. It is also believed that the HIV proviral DNA in this phenotype is less transcriptionally active, preventing the production of HIV virions and the active replenishment of the proviral reservoirs.
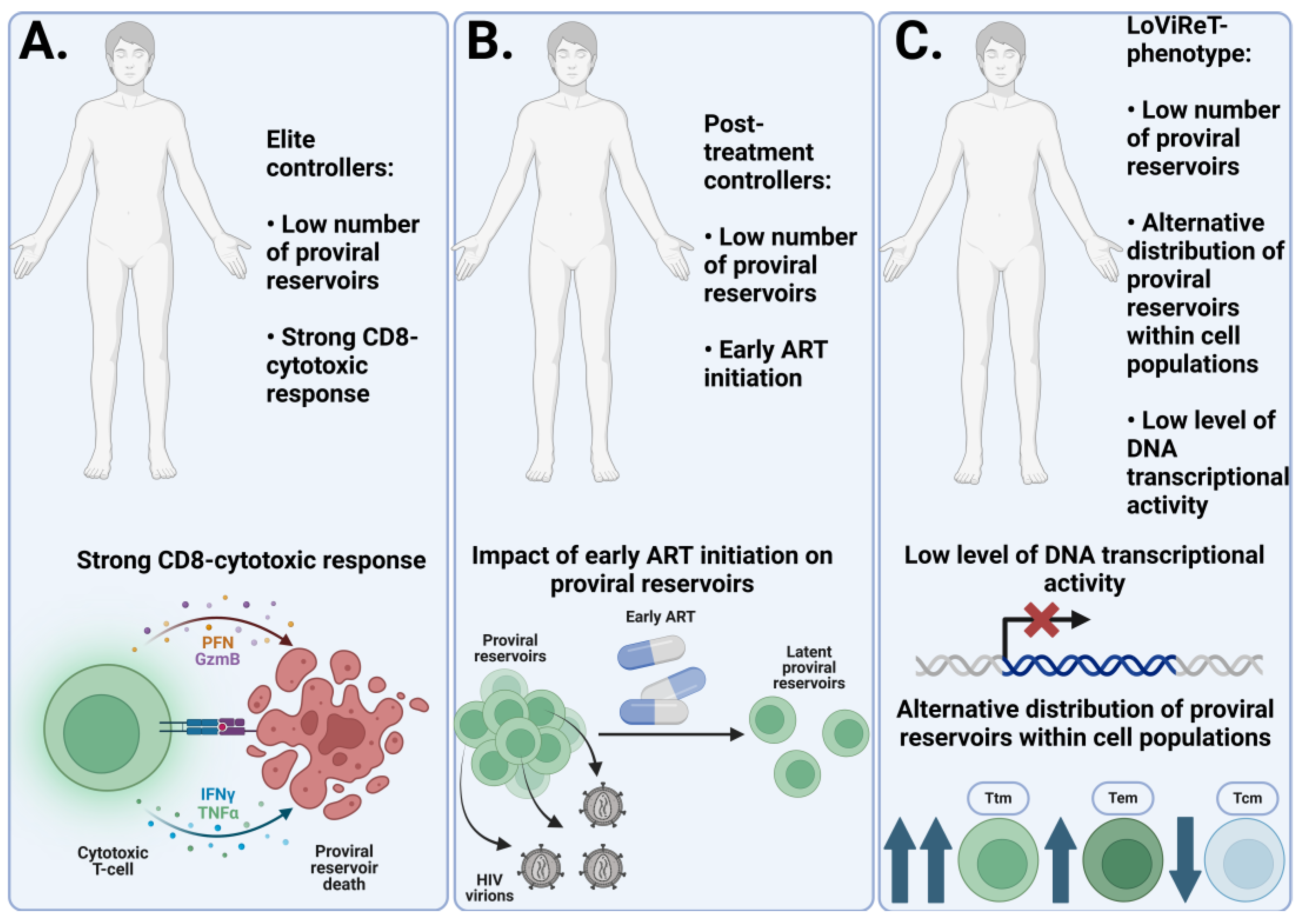
Figure 3.
Current methods of analysis of proviral reservoirs. (A) Digital PCR, in its version for detecting defective and intact proviral reservoirs (Intact Proviral DNA Assay, IPDA), allows for an accurate estimation of the number of defective and intact reservoirs in a sample. This method involves the selection of primers and probes for key genes essential for the HIV life cycle (Gag, Pol, Env, Nef, etc.). The presence of a signal in one channel indicates the presence of defective proviral reservoirs for a particular target, while the presence of two signals from different channels indicates intact proviral reservoirs. The absence of signals indicates either defective reservoirs for both targets or the absence of HIV DNA in the nanowell. (B) Quantitative analysis of viral replication, presented in its various forms, allows us to estimate the number of replication-competent reservoirs. The main idea of the method is to incubate PBMC from an HIV-infected person with mitogens and feeder cells to activate them. After that, coculture with lymphoblasts or MOLT-4/CCR5 cell lines allows the virus to spread within this culture. At a certain point (7, 14, or 21 days later), the number of proviral reservoirs is counted using RT-qPCR or p24 ELISA techniques. (C) Genome-wide sequencing provides data that can be used to study various properties of proviral reservoirs. One promising area of research is the mapping of HIV integration sites. ISLA [97] and nrLAM-PCR [98] protocols have been developed to amplify and sequence HIV integration sites. Further data processing allows localization of HIV integration sites.
Figure 3.
Current methods of analysis of proviral reservoirs. (A) Digital PCR, in its version for detecting defective and intact proviral reservoirs (Intact Proviral DNA Assay, IPDA), allows for an accurate estimation of the number of defective and intact reservoirs in a sample. This method involves the selection of primers and probes for key genes essential for the HIV life cycle (Gag, Pol, Env, Nef, etc.). The presence of a signal in one channel indicates the presence of defective proviral reservoirs for a particular target, while the presence of two signals from different channels indicates intact proviral reservoirs. The absence of signals indicates either defective reservoirs for both targets or the absence of HIV DNA in the nanowell. (B) Quantitative analysis of viral replication, presented in its various forms, allows us to estimate the number of replication-competent reservoirs. The main idea of the method is to incubate PBMC from an HIV-infected person with mitogens and feeder cells to activate them. After that, coculture with lymphoblasts or MOLT-4/CCR5 cell lines allows the virus to spread within this culture. At a certain point (7, 14, or 21 days later), the number of proviral reservoirs is counted using RT-qPCR or p24 ELISA techniques. (C) Genome-wide sequencing provides data that can be used to study various properties of proviral reservoirs. One promising area of research is the mapping of HIV integration sites. ISLA [97] and nrLAM-PCR [98] protocols have been developed to amplify and sequence HIV integration sites. Further data processing allows localization of HIV integration sites.
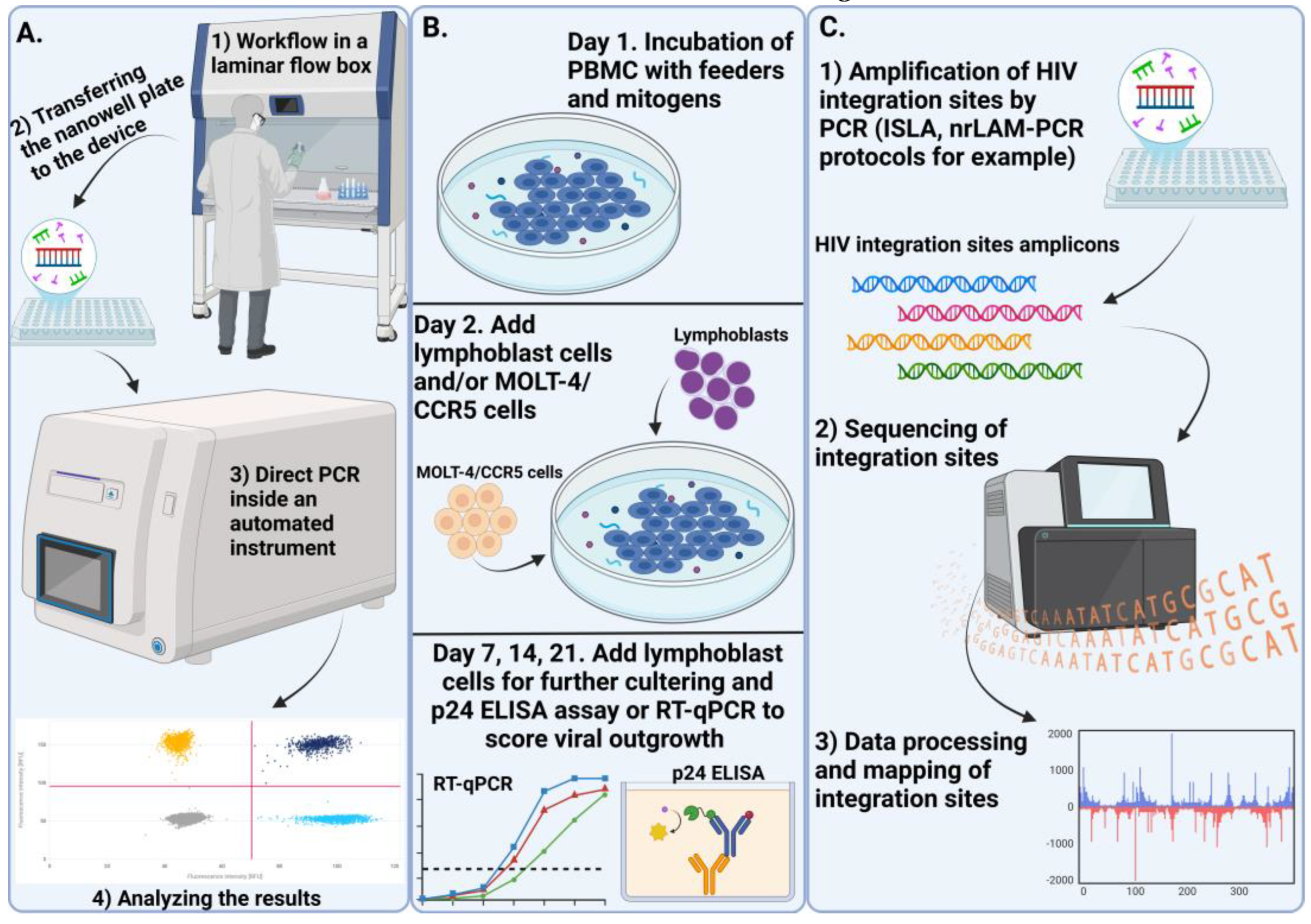
Figure 4.
Approaches to targeting proviral reservoirs for eradication. (A) In the absence of immune system intervention, the HIV life cycle proceeds normally. (B) The Block and Lock approach involves the use of molecules, known as latency agents (LAs), that induce a deep latent state. Didehydro-Cortistatin A is one of the most studied and promising LAs, which prevents the interaction between the viral Tat protein and the TAR site in the proviral genome, thereby halting the transcription of other proviral genes. (C) The use of CRISPR/Cas9 to excise the entire provirus or its individual components from the nuclear genome. (D) The Shock and Lock strategy involves initially reactivating the proviral gene transcription with latency reversal agents (LRAs). Histone deacetylase inhibitors (HDACi) and histone methyltransferase inhibitors (HMTi) can reactivate the HIV provirus by interfering with epigenetic modifications, enabling the immune system to detect and target proviral reservoirs with cytotoxic T lymphocytes (CTLs). When combined with cytolytic agents that enhance cell lysis, this approach facilitates effective eradication of proviral reservoirs.
Figure 4.
Approaches to targeting proviral reservoirs for eradication. (A) In the absence of immune system intervention, the HIV life cycle proceeds normally. (B) The Block and Lock approach involves the use of molecules, known as latency agents (LAs), that induce a deep latent state. Didehydro-Cortistatin A is one of the most studied and promising LAs, which prevents the interaction between the viral Tat protein and the TAR site in the proviral genome, thereby halting the transcription of other proviral genes. (C) The use of CRISPR/Cas9 to excise the entire provirus or its individual components from the nuclear genome. (D) The Shock and Lock strategy involves initially reactivating the proviral gene transcription with latency reversal agents (LRAs). Histone deacetylase inhibitors (HDACi) and histone methyltransferase inhibitors (HMTi) can reactivate the HIV provirus by interfering with epigenetic modifications, enabling the immune system to detect and target proviral reservoirs with cytotoxic T lymphocytes (CTLs). When combined with cytolytic agents that enhance cell lysis, this approach facilitates effective eradication of proviral reservoirs.
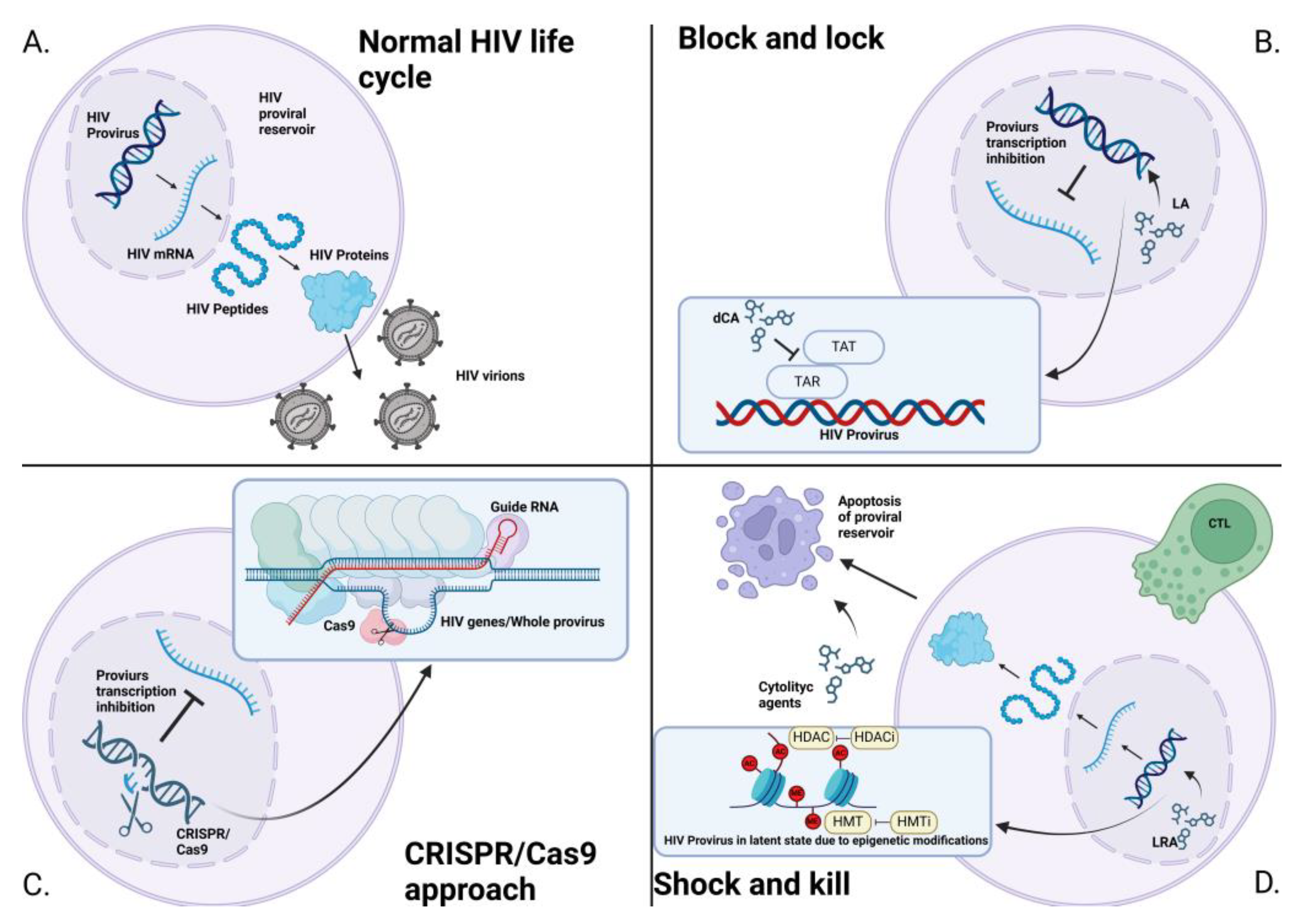
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



