
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
Metabolic reprogramming is a hallmark of cancer, with cancer cells acquiring many unique met-abolic traits to support malignant growth, and extensive intra- and inter-tumour metabolic het-erogeneity. Understanding these metabolic characteristics presents opportunities in precision medicine for both diagnosis and therapy. However, despite its potential, metabolic phenotyping has lagged behind genetic, transcriptomic and immunohistochemical profiling in clinical appli-cations. This is partly due to the lack of a single experimental technique capable of profiling the entire metabolome, necessitating the use of multiple technologies and approaches to capture the full range of cancer metabolic plasticity. This review examines the repertoire of tools available for profiling cancer metabolism, demonstrating their applications in preclinical and clinical settings. It also presents case studies illustrating how metabolomic profiling has been integrated with oth-er omics technologies to gain insights into tumour biology and guide treatment strategies. This information aims to assist researchers in selecting the most effective tools for their studies and highlights the importance of combining different metabolic profiling techniques to comprehen-sively understand tumour metabolism.
Keywords:
metabolomics
; cancer
; precision medicine
; therapy
; metabolome
1. Introduction
Metabolic reprogramming is undoubtedly one of the crucial hallmarks of cancer pathogenesis [1]. Tumour cells undergo several metabolic adaptations, often in a nutrient-deficient microenvironment, to sustain their increased rate of growth and proliferation [2].Perhaps the most well-known example of cancer metabolic reprogramming is the ‘Warburg effect’, which refers to the tumour cells’ preference for using glycolysis even in the presence of oxygen (aerobic glycolysis) [3]. However, the switch from aerobic cellular respiration to aerobic glycolysis represents just the tip of the iceberg, and cancer cells acquire many other unique metabolic traits to support their malignant growth, including enhanced generation of antioxidant and detoxification capacity, upregulation of lipid metabolism or addiction to glutamine metabolism [4]. Tumours further display extensive intra and inter-tumour metabolic heterogeneity which is manifested in both spatial and temporal dimension [5].
Understanding the metabolic characteristics and dependencies of cancer cells presents several opportunities in precision medicine from both a diagnostic and therapeutic perspective. Non-invasive metabolic profiling has been routinely applied to accurately detect and monitor tumour progression, whilst metabolic biomarkers show immense promise in identifying altered signalling pathways associated with the specific cancer being profiled [6]. Despite promising applications, the use of metabolic phenotyping in precision medicine has lagged as a diagnostic and therapeutic tool compared to the more routinely used genetic or immunohistochemical profiling [7]. More cancer cohorts are choosing to focus their efforts on profiling cancer genomes, transcriptomes and proteomes ignoring that the metabolome can often depict a more accurate and direct functional readout of the pathophysiological state of living systems [8].
This is also instigated by the fact that unlike other omics profiling technologies, there is no singular experimental technique that enables the entire metabolome to be profiled at once [9]. As a consequence, it is rather challenging to capture the full range of metabolic plasticity cancer cells possess, limiting the identification and application of specific metabolic biomarkers. Given the vast chemical space a cellular metabolome encompasses, metabolic profiling through multiple technologies and approaches is necessary to better understand and leverage tumour metabolic reprogramming. The purpose of this review is to examine the repertoire of tools available to researchers in tracking cancer metabolism to better aid them in decisions about which would be most effective for their studies. We also then proceed to demonstrate relevant examples of how these technologies have been applied both pre-clinically and in patient samples. Lastly, we demonstrate two case studies of how metabolomic profiling has been integrated with other omics technologies to gain better insights into tumour biology and guide treatment strategies in patients.
2. An Overview of Technologies and Approaches in Studying Tumour Metabolism
A wide range of technologies and approaches are being utilised in studying tumour metabolism, starting from metabolomics which refers to the set of analytical chemical techniques to characterise the composition of biochemical mixtures [7]. Metabolomic techniques can be used to profile a wide range of metabolites from a variety of sources, including cells, tissues, and bodily fluids [10]. Unlike with genomics, transcriptomics, and proteomics, metabolomics cannot be performed using a single instrument or technique because metabolites have a wide range of concentrations, sources, and chemical properties [9]. Another limitation of current metabolomic techniques is that it is often challenging to accurately capture the cell’s metabolic state by quenching all its metabolic pathways [11]. As a result, most metabolomic techniques focus on profiling and quantifying a smaller subset of pre-selected metabolites. Different metabolic profiling tools are often combined to take advantage of their strengths and compensate for their weaknesses. This section will focus on these tools and how they are combined to more comprehensively profile tumour metabolism.
Figure 1.
Overview of tools and technologies used to investigate metabolic phenotypes in cancer. Figure created using images adapted from various publications [12,13,14] and Biorender.
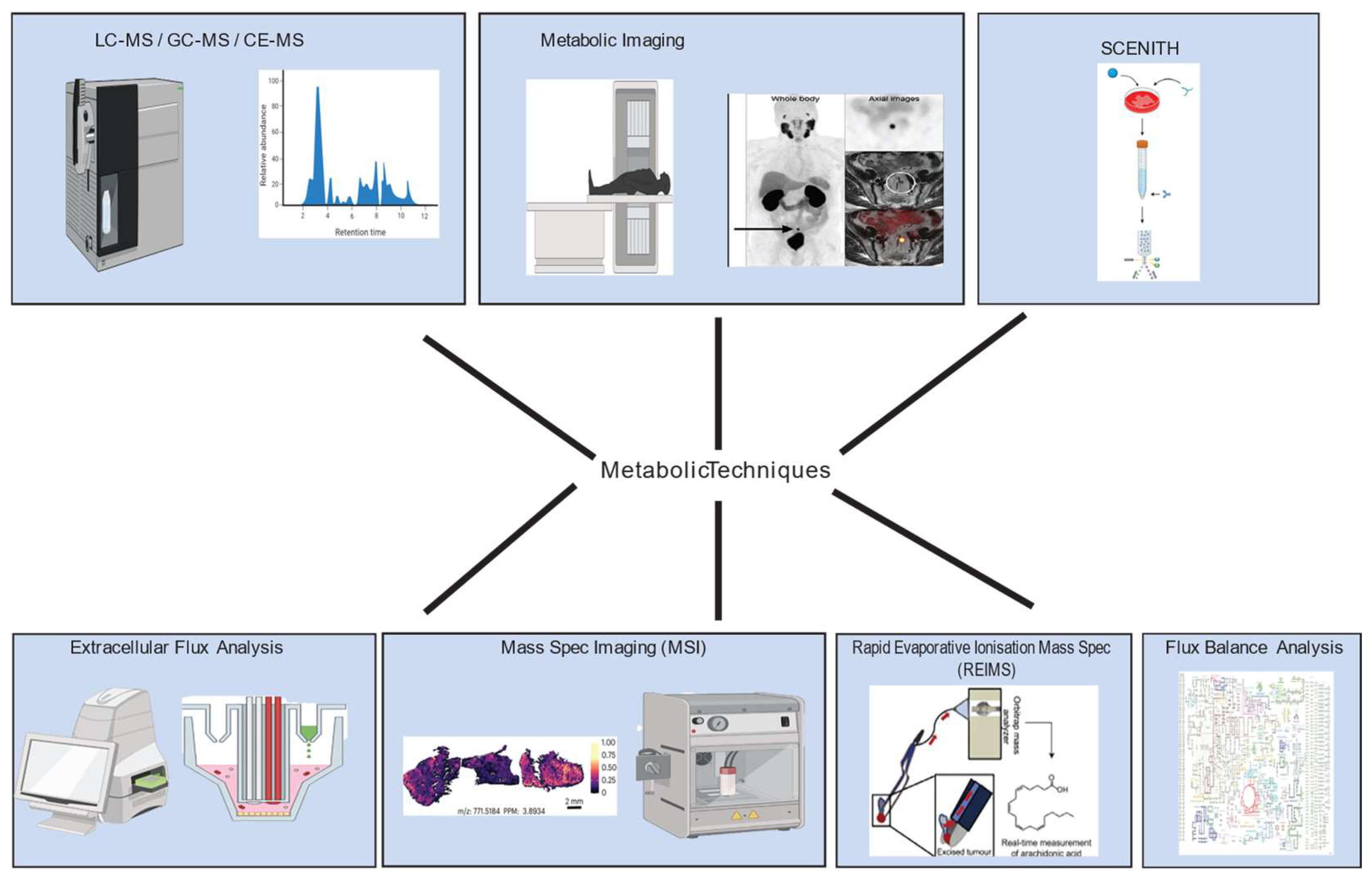
2.1. Nuclear Magnetic Resonance (NMR) Spectroscopy, and Mass Spectrometric Methods Remain the Cornerstone of Metabolomic Profiling in Cancer
Over the past few decades, nuclear magnetic resonance (NMR) spectroscopy and separation techniques coupled to mass spectrometry have become standard practice to be able to profile and quantify metabolites in biological samples. These methods have been extensively reviewed elsewhere [15,16], hence this section will only briefly address the workings of NMR, gas chromatography mass spectrometry (GC-MS), liquid chromatography mass spectrometry (LC-MS) and capillary electrophoresis mass spectrometry (CE-MS). NMR uses radiofrequency pulses and strong magnetic fields to analyse the environment of certain atomic nuclei, most commonly 1H and 13C [17]. In the strong magnetic field, these nuclei absorb and re-emit energy producing characteristic signals providing information about the chemical environment they are in. These spectra can then be used to estimate the relative concentration of specific metabolites or study the chemical environment of the analyte of interest. NMR can be applied in untargeted settings to quantify specific metabolic species most commonly from 1H spectra [18]. However, these spectra can often be complex and difficult to analyse; computational techniques involving machine learning can help facilitate analysis and quantification of the metabolites [19]. More commonly, 13C NMR is used more commonly in targeted metabolomics settings for direct measurement of labelled carbons. Different tracers can be used to study specific metabolic pathways - for example 13C1,2- glucose can be used to distinguish between oxidative and non-oxidative branches of the pentose phosphate pathway from differential labelling in downstream metabolites. 13C glutamine can be used to study glutaminolysis while 13C-palmitic acid is often used to study fatty acid oxidation [16]. Metabolites can often be shuttled into many different pathways, but NMR allows the elucidation and quantification of metabolic flux into specific pathways due to the chemical environment of the labelled carbon, making it an attractive and powerful technique to use in metabolomics settings.
Despite its ease of use, NMR is limited in its ability to detect low-abundance metabolites and has lower sensitivity compared to mass-spectrometric techniques [20]. Mass spectrometry is most commonly coupled with three different separation techniques: gas chromatography, liquid chromatography and capillary electrophoresis. The choice of separation technique often depends on the analyte and metabolic species to be determined [21]. GC-MS is best suited for detecting volatile compounds with low boiling points such as free fatty acids [22]. GC-MS separation takes place inside capillary columns consisting of two phases – a stationary phase coated on the inner surface of the column and a carrier gas with the vapourised metabolite sample or the mobile phase. Each metabolite in the mobile phase interacts differently with the stationary phase of the column depending on its chemical properties [23]. Metabolites are, therefore, separated based on their travel speed through the column. Gas chromatograms are often coupled to time-of-flight mass spectrometers (TOF-MS) that allow metabolite identification. Control of metabolite injection into the TOF analyser allows better detection of lower concentration metabolites and good resolution of the analytes in question [24]. GC-MS can detect certain volatile compounds with high precision, but is limited to compounds that can easily be vaporized and are thermally liable. This represents a small subset of the cellular metabolome limiting the applicability of GC-MS.
After the commercial introduction of atmospheric pressure ionisation sources, LC-MS gained more widespread use to profile metabolite mixtures [25]. Like GC-MS, LC-MS consists of a stationary solid phase and a mobile phase (which is a liquid in the case of LC-MS) inside a silica column. The mobile phase carries the metabolites to be analysed that interact and are eluted from the column depending on their chemical properties [25]. Advances in chromatographic separation techniques in LC-MS, such as Hydrophilic interaction liquid mass chromatography (HILIC), has further led to better separation of compounds and resolution of peaks for LC-MS [26]. As a result, many complex metabolite mixtures can now be analysed using LC-MS with high accuracy.
CE-MS is a relatively new technique that is especially suited for analysis of polar and charged metabolites in biological samples such as nucleic acids, amino acids, peptides and charged lipids [27]. Charged analytes migrate through a capillary that’s placed in an electric field produced from electrodes. Analytes migrate according to their electrophoretic ability and are subsequently detected by a spectrometer. CE-MS has excellent separation for polar metabolites that may be structurally similar, but possess different mass:charge ratios and require very little sample preparation. However, due to the relative low use of CE-MS, challenges remain in its reproducibility and optimisation for specific metabolite classes [28].
Similar to NMR, GC-, LC- and CE-MS can also be used in fluxomics applications to measure metabolic flux through specific reactions or pathways. This is achieved through radioisotope labelling of certain substrates such as 13C-labelled glucose, or 15N-labelled glutamine, and incorporation analysis of the labelled isotope into other compounds to track metabolic rewiring [29]. While mass spectrometry methods have been widely used, GC-, LC- and CE-MS are constrained in how many different species it can profile simultaneously and has difficulty detecting certain metabolites present at very low concentrations. Often to profile many different metabolites, multiple different columns with different properties must be used which can be quite expensive [10]. While NMR and mass-spectrometry methods may not provide a truly global metabolomic profile, they still nonetheless remain indispensable for detecting and quantifying metabolites. Many metabolomic strategies may use other techniques to profile cancer metabolism, but still rely on the aforementioned technologies to validate findings.
2.2. Metabolic Imaging Enables the Examination of Metabolism In Vivo
As mentioned before, tumour metabolism can differ quite significantly from the metabolism of the surrounding normal tissue given tumours’ high metabolic demands. This can be exploited through metabolic imaging that offers insights into altered biochemical processes characteristic of malignant growth [30]. While multiple imaging techniques exist, this section will address the two most widely used: positron emission tomography (PET) and magnetic resonance spectroscopy (MRS). PET provides detailed information about the metabolic activity of tissues in the body. It makes use of a radiotracer, most commonly 18F-fluorodeoxyglucose (FDG) that mimics glucose and is taken up in cells in a manner proportional to their metabolic activity. Once 18F-FDG is taken up, it is phosphorylated by hexokinase to FDG-6 phosphate, which unlike glucose, cannot be metabolised further, so it accumulates in tissues. Once inside tissues, the tracer emits positrons, which upon encountering electrons, emits photons that move in opposite directions. PET detectors that surround the body can detect these photons and use this information to infer the three-dimensional distribution of the tracer in the body [31]. This allows a visualisation of metabolically active regions of the tissue such as tumours and can aid diagnosis and staging of tumours. PET is quite sensitive to changes in metabolism and can thus aid early cancer detection. It is also used to sensitively monitor the response of tumours to treatment, and because it can be used to scan the entire body, can enable the detection and monitoring of metastases [32]. The biggest drawback of PET is its low spatial resolution to precisely locate tumours or fine details surrounding the tumour. PET is also limited in the types of species that can be used in radiotracing and the metabolites it can image in vivo [33]. Nonetheless, PET still has important applications in cancer diagnostics and detection with multiple Food and Drug Administration (FDA)-approved radiotracers used in the clinic.
MRS borrows from the workings of nuclear magnetic resonance to non-invasively profile tissues at the molecular level (see NMR section for further details). MRS is a versatile technique that provides quantitative data on metabolite concentrations in many different tissue types. However, it is limited by its spatial resolution and sensitivity as it requires metabolite concentrations to be on the order of millimolar (mM) [34]. MRS’s applications, however, have massively improved with the arrival of dissolution dynamic nuclear polarization (DNP) that can enhance the sensitivity of 13C labelling by over 10,000 times. DNP hyperpolarizes nuclear spins which means DNP 13C-labelled metabolites can be detected much more sensitively and at much lower concentrations than when using their non-hyperpolarized counterparts [35]. For full details on the DNP MRS methodology, readers are referred to other relevant reviews [36].
While PET and MRS can’t be used to truly profile the metabolome of cancer, their importance is underscored by their application to profile metabolism in vivo which can have important applications in precision medicine. These techniques are particularly useful when combined with other metabolomic methods such as stable-isotope assisted LC-MS or MS imaging that can highlight important metabolites to examine in vivo.
2.3. Matrix-Assisted Laser Desorption/Ionisation (MALDI) and DESI Preserve Spatial Data During Metabolomic Profiling
While NMR, GC-MS and LC-MS can provide quantitative measurements for a variety of metabolites, they utilise cell extracts from many cells and therefore, sacrifice spatial information for the corresponding metabolomics data. MALDI and DESI are both mass-spectrometry imaging (MSI) techniques that aim to profile many metabolites while allowing for spatial information preservation [37]. MALDI makes use of a matrix-like compound typically added onto a tissue segment or a biological extract [38]. The matrix is then irradiated with a laser and transfers the energy to the metabolites leading to the desorption and ionisation of metabolites from the matrix. These ions are then accelerated into the mass analyser of a mass spectrometer for identification and relative quantification [39]. The laser desorption and ionisation process can be focused on a particular area of a slide, allowing for spatial information preservation [38]. MALDI is well suited for the analysis of small molecules, as well as those with low abundance. MALDI is also less harsh on the metabolite samples as they are not directly irradiated with the laser compared to some ionisation methods used in LC-MS [39]. Thus, MALDI is often adept for analysis of labile compounds that can undergo fragmentation under harsher ionisation procedures.
DESI, on the other hand, can analyse the molecular composition of tissues directly without the need for external sample preparation. As it operates under atmospheric conditions, tissues in their native state can be analysed [40]. Instead of a matrix, DESI utilises a charged solvent, which contains ions used for desorption and ionization, that is continuously sprayed onto the tissue surface. This spray forms small droplets upon hitting the sample surface, that interact with metabolites on the surface. This leads to a transfer of energy to the analytes on the surface causing their desorption into the gas phase [41]. These desorbed molecules enter the gaseous phase and can then be ionized and analysed by a mass spectrometer. By moving the sample or DESI probe, spatial information can be generated for the tissue of interest [41]. DESI and MALDI excel at providing metabolomic profiling with spatial context, which is difficult to achieve with most other metabolomic techniques [42]. Advancements in technology have also allowed for their use with minimal sample preparation increasing their applicability [43].
However, MALDI and DESI can only profile limited dynamic ranges of metabolites and struggle with the identification of isobaric compounds, limiting their more widespread use as a metabolomic profiling technology [44]. LC-MS, on the other hand, makes use of chromatographic separation and so can handle broader ranges of metabolites [45]. Moreover, MALDI and DESI provide semi-quantitative metabolomics data, and given the lack of internal standards that are present in other mass-spectrometry based techniques, they often face issues in reliably quantifying metabolites [46]. Technical issues with each technique such as ion suppression effects (the presence of one analyte affects the ionisation of another) in MALDI, and polarity bias (where ionisation of certain compounds is favoured over others) in DESI, further limit the suitability of their use in metabolomic profiling [47]. Despite all these limitations, MALDI and DESI are incredibly powerful and versatile technologies as they provide spatially resolved metabolomics data for samples. With regards to precision medicine, this has important implications in studying metabolic heterogeneity of individual tumour samples, the metabolic crosstalk between the tumour microenvironment and cancer cells, as well as identification of drivers for metabolic phenotypes [44]. It is possible that MALDI and DESI could be used in clinical applications to identify metabolic phenotypes associated with drug response and identify tumour areas that may be more resistant to specific targeted therapies.
2.4. Extracellular Flux Analysis (EFA) Facilitates Investigation into Metabolic Phenotypes
Studying tumour metabolism involves examining not just metabolite levels and pathways, but also the way cancer cells manage their energy sources via metabolic phenotypes. The Agilent Seahorse Extracellular Flux (XF) analyser, is used to quantify “real-time” bioenergetic profiles of cells upon treatment with pharmacological agents [48]. It can simultaneously measure oxygen consumption rate (OCR) as a readout for mitochondrial oxidative phosphorylation (OXPHOS) activity, and extracellular acidification rate (ECAR) that acts as an indicator of lactate secretion or glycolysis [49]. Importantly, the Seahorse instrument allows administration of up to four drugs enabling the study of cellular metabolic adaptation under different conditions. An example application of EFA to fully characterise mitochondrial activity is through the common Cell mito stress test, which starts out with a basal measurement of OCR for evaluating the basal respiration of cells [50]. The cells are then exposed to oligomycin that greatly reduces mitochondrial respiration by inhibiting ATP synthase causing a decrease in OCR. The second injection involves adding carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP) to cells, which simulates maximal electron flow through the electron transport chain so that the respiratory chain operates at its maximum capacity. The difference between the maximal and the basal oxygen consumption is known as the spare respiratory capacity which gives an indication of cells’ fitness and ability to respond to cellular stresses and different energetic demands. Lastly, rotenone and antimycin are injected in the media – these inhibit complex I and III respectively and shut down mitochondrial respiration, which also allows to determine non-mitochondrial respiration driven by processes outside the mitochondria. Different combinations of drugs can be used to test for various metabolic capacities and phenotypes of tumour cells in analogous ways [50].
EFA assays have enabled a more thorough investigation into overall metabolic phenotypes. For example, when tumours exhibit gross phenotypic shifts due for example the acquisition of specific driver mutations or development of resistance to certain targeted therapies, they undergo major metabolic shifts, such as a change in a preferred carbon source or dependence on a particular nutrient. EFA assays are easy to perform and give a quantitative readout of various bioenergetic parameters; the ability to inject custom drugs also enables metabolic vulnerabilities under different conditions to be tested [6]. However, care must be taken while performing these experiments and interpreting the results as factors like media composition, cell density and inhibitor concentration can all greatly affect the observed phenotypes. In addition, while OCR and ECAR give some measure of oxidative and glycolytic activity, many pathways contribute to these readings and the contribution of secondary bioenergetic pathways cannot be ignored [51]. EFA assays offer a coarse interpretation of cellular metabolic shifts and are often used to accompany other signalling or metabolic findings but, nonetheless, represent useful accompaniments when exploring metabolic shifts and novel vulnerabilities [52].
2.5. SCENITH Can Be Used for Immuno-Metabolic Profiling Applications
In vitro EFA assays typically use many cells in bulk and specific growth media to perform the assays. They’re unable to profile heterogeneous populations of cells, classify rare sub-populations of cells and the change in media can lead to artefacts. Single Cell ENergetIc metabolism by profiling Translation inHibition (SCENITH) is a technique that allows single cell energetic profiling of multiple cell types ex vivo and allows shifts in metabolism in certain sub-populations of cells to be detected upon certain events [53]. SCENITH allows the study of metabolic responses by measuring protein synthesis rates as a proxy for cellular metabolism. Protein synthesis rates can be reliably measured using incorporation of puromycin and show a strong correlation with metabolic rates. Samples will typically be incubated with puromycin for a short time to allow labelling. Using a novel anti-puromycin monoclonal antibody, SCENITH then leverages cell sorting technologies such as fluorescence-activated cell sorting (FACS) to measure incorporation of puromycin at a single cell resolution. Similar to EFA assays, SCENITH uses a combination of different metabolic inhibitors along with puromycin to study the contribution of different energetic pathways for specific cell types. An example of this may be incubating the cells with puromycin and 2-deoxy-d-glucose to study the energetic contribution of glycolysis [53].
SCENITH’s primary use has been for immuno-metabolic profiling as it allows for metabolic profiling of different cell types in the tumour micro-environment at single cell resolution [53]. Identification of immune cells has been heavily optimised in FACS experiments and so SCENITH complements existing technologies to better understand the tumour micro-environment. This has especially important applications in improving the efficacy of immunotherapy as it can help in the understanding of why certain patients respond and potential ways of metabolically reprogramming the microenvironment to improve response to immunotherapies [6]. SCENITH also can be performed ex vivo and so can be directly applied to patient serum samples bypassing artefacts induced by culturing in non-physiological media that are common in vitro. Despite this, SCENITH has some drawbacks in energetically profiling cells. SCENITH relies on measuring protein synthesis which is an indirect measurement of cellular metabolism as opposed to the more direct measurements of OCR and ECAR used in EFA assays. SCENITH also relies on cellular fixation and cannot analyse energetic profiles in real time in response to certain inhibitors like EFA assays can [53]. Nonetheless, the primary draw of SCENITH that most other technologies fail to provide is single cell energetic profiling. Given how heterogenous the immune cell population in the tumour micro-environment is, SCENITH is emerging as a valuable tool to understand the numerous interactions tumour cells have with immune cells and how energetic profiles of immune cells govern their phenotypic behaviour. When combined with single cell RNA-seq (scRNA-seq), SCENITH holds immense promise for improving the efficacy of immunotherapies [52].
2.6. Rapid Evaporative Ionisation Mass Spectrometry (REIMS) Permits Real-Time Analysis of Surgical Samples
A key feature that distinguishes metabolism from other cellular processes, is its ability to be analysed immediately. Once a sample is ready to be analysed by a mass spectrometer, the instrument can generate a spectrum and identify certain metabolites within a short duration. This is in direct contrast, for instance, to profiling tumour genomics as even after a tedious process of DNA extraction, it takes significantly longer to sequence and identify mutations. The entire process from sampling to mass detection is shortened by recent technological advancements coupling surgical techniques to metabolomic profiling through rapid evaporative ionisation mass spectrometry (REIMS) [54]. REIMS allows near-instantaneous metabolic profiling of tissues, enabling surgeons to identify boundaries between tumours and surrounding non-cancerous tissue. REIMS is typically used alongside an electrosurgical knife that cauterizes tissue during surgery. As the knife cauterizes the tissue, it generates an aerosol known as the electrosurgical plume which contains small particles of the tissue. The electrosurgical plume is directed into the REIMS ionization source in which the tissue particles are subject to an electric field and heated vaporization chamber. The high temperature in the chamber causes the particles to rapidly evaporate leading to the lipids in the tissue being ionized allowing for them to be analysed by mass spectrometry [55]. Given that cancerous and non-cancerous tissues have distinct metabolic profiles, REIMS allows for differentiation between the two. Importantly, it has also been shown that distinct metabolic signatures can be linked to specific oncogenic drivers – for example PIK3CA mutant tumours display unique metabolic profiles characterised by enhanced arachidonic acid levels [56]. In addition to identifying tumour boundaries, this allows for REIMS to provide near real-time diagnosis of tumours driven by specific oncogenic drivers [56], opening therapeutic opportunities during surgery even for regions of the tumour that can’t be fully surgically excised.
2.7. Flux Balance Analysis (FBA) Allows a Near Genome Wide View of Cancer Metabolism
While it is difficult to globally profile the metabolome with current experimental methods, computational methods can aid in predicting cellular metabolomes. One such method is flux balance analysis (FBA) that analyses the flow of metabolites through metabolic networks [57]. FBA calculates metabolite flow using genome scale metabolic network reconstructions curated over the past few decades. It operates under the assumption that cellular metabolic processes seek a steady state with fluxes through metabolic reactions optimised to reach certain cellular objectives such as maximisation of growth, maintenance of energy levels represented through ATP:ADP ratios or maximizing a cell’s reductive potential to protect against harmful reactive oxygen species. FBA utilises linear programming to predict optimal flux distributions through minimising or maximizing objective functions subject to constraints of a predefined stoichiometric model of the metabolic network. These flux distributions can then be interpreted as a surrogate for metabolic activity through a particular reaction or pathway.
FBA’s real value lies in being able to set appropriate constraints on the metabolic network from available transcriptomics or proteomics data – this allows unique solutions for flux vectors to be generated depending on the sample of interest. In this way FBA can model, for example, different cancer cell lines or patients and identify key metabolic pathways influencing metabolic phenotypes. In addition, FBA can also be used to identify potential metabolic vulnerabilities that upon knockout in silico, lead to a decrease in the value of the objective function and serve as a readout for reduction or loss in viability [58]. FBA has been used routinely in microbial settings particularly in industrial applications involving metabolic engineering [59]. While FBA is not commonplace in studying human metabolism, advances in model simulation methods and curation are improving the accuracy of its use in studying cancer [58]. FBA holds great promise as a companion technique in metabolomics offering a quantitative framework to reduce the metabolic space to study. This can allow focus on a few select reactions or pathways through experimental methods such as LC-MS-based metabolomics.
2.8. Comparison of Different Metabolomics Techniques

Table 1.
Overview of the advantages and disadvantages of different metabolomics techniques.
| Technique | Advantage | Disadvantage |
|---|---|---|
| GC-MS |
|
|
| LC-MS |
|
|
| Metabolic imaging - PET |
|
|
| Metabolic Imaging – MRS |
|
|
| Mass Spectrometry Imaging |
|
|
| Extracellular Flux Analysis |
|
|
3. The Application of Metabolomics in the Diagnosis and Treatment of Cancer
This section covers some key applications of the aforementioned metabolomics techniques in cancer research. In particular, how metabolomics can aid in tumour detection, understanding tumourigenesis, monitoring response to therapies, identifying novel metabolic vulnerabilities and improving the outcome of existing therapies.
3.1. Non-Invasive Metabolic Tracking for Tumour Detection and Response to Therapy
Tumour metabolic reprogramming often occurs downstream of signalling pathways perturbations or other important regulators. Metabolomics can detect specific metabolites whose levels are altered throughout neoplastic progression, providing effective biomarkers for cancer diagnosis and disease monitoring [6]. In particular, non-invasive metabolic tracking has attracted significant attention due to its simplicity and increased patience compliance compared to obtaining biopsies to profile cancers. Non-invasive metabolic tracking tools in clinical settings usually comprise of metabolic imaging of selective biomarkers or direct metabolomics analysis of biofluids [60]. While many different types of samples can be used for the latter analysis, such as urine, saliva, faecal matter and breath, this review will examine a few brief examples using serum and plasma, and the reader is referred to other reviews for more comprehensive coverage of this topic in different cancer subtypes [61,62,63,64,65,66].
Imaging techniques including magnetic resonance imaging and PET are widely used clinically for the detection and diagnosis of cancer. As mentioned in the previous section, one of the most common and first tracers used for PET imaging was 18F-FDG-PET that takes advantage of tumour’s enhanced metabolic activity and glucose uptake. Its application extends to initial diagnoses including estimation of staging, lymph node involvement and presence of metastases. It is also used after treatment to assess response to therapy and follow up recurrent cases [67]. Due to its high retention within cells and high rate of clearance from the body, 18F-FDG-PET is used to diagnose many different types of cancers including lung, head and neck, ovarian and breast cancers amongst others [68]. 18F-FDG-PET is used in guiding treatment strategies for pancreatic cancer where it has been applied to aid in choosing between curative and palliative approaches to treatment [69]. However, 18F-FDG may not always be the most appropriate metabolite for PET imaging. Some tumours, such as prostate cancers, have low glucose avidity, and instead rely more on fatty acid oxidation [70], while in other tumours, such as gliomas, the high background of the surrounding neural tissue can interfere with detection [71]. Other tracers that examine different metabolic pathways or substrates can be used instead. For example, L-[5-11C]-glutamine [72], [18F]-(2S,4R)-4-fluoroglutamine, and [18F]-(2S,4S)-4-(3-fluoropropyl)glutamine [73] have been used to study glutaminolysis which has been shown to be significantly altered in some cancers. [18F]-(2S,4R)-4-fluoroglutamine has been applied to a recent glioma clinical trial as gliomas show higher rates of glutaminolysis [74]. Other tumours rely more on lipid metabolism and show increased lipid species synthesis; this idea has been exploited to develop [18F]-fluoromethylcholine and [18F]-fluoroethylcholine which label the phospholipid head group choline and can be used as a proxy for membrane synthesis [75,76] . These tracers are currently used in the clinic to image prostate cancer. The use of different metabolic species makes PET a very versatile and useful clinical tool for cancer diagnosis.
Magnetic resonance spectroscopy imaging (MRSI) is another invaluable imaging modality that allows for the detection of metabolites spatially resolved in vivo, and unlike PET, does not involve exposure to ionizing radiation. Early studies using 1H and 13C MRSI had some limitations in their application due to low spatial resolution, but rapid advances have been made with the advent on DNP [77]. One of the first, and most widely used molecules to be hyperpolarised for MRSI is [1-13C] pyruvate [78]. The hyperpolarised 13C from [1-13C] pyruvate can shift its environment if it’s converted to lactate, catalysed by lactate dehydrogenase, and serves as a proxy for measuring pyruvate’s entry into the mitochondria and the TCA cycle. Due to its sensitive detection, hyperpolarised [1-13C] pyruvate has been used in preclinical studies to show that lactate labelling can be used to assess tumour grade and early treatment response [79,80]. It has been applied to clinical studies of breast, prostate, brain, renal and pancreatic cancers and has been used to assess response to immune checkpoint inhibitors in a prostate cancer patient [81]. MRSI of lactate labelling from hyperpolarised [1-13C] pyruvate can also identify tumour metabolic subtypes and gain insights into likely responses to therapy. For example, in a study conducted on orthotopically implanted patient-derived glioblastoma xenografts, levels of lactate labelling corresponded to transcriptomic derived subtypes [82]. High levels of lactate labelling resembled the mesenchymal subtype that was more radioresistant [83] compared to tumours that showed lower levels of lactate labelling and resembled more oxidative neural progenitor like glioblastomas [82]. While hyperpolarised [1-13C] pyruvate has been more widely used clinically, other emerging hyperpolarised metabolites such as [1,4-13C2] fumarate for imaging necrotic cell death [84] are also increasingly being used in various applications.
While PET and MRSI have been applied quite extensively in clinical contexts, they are not without drawbacks. FDG-PET for example is often useful for diagnosing later, but not early-stage cancers. This is because 18F-FDG is a substrate for facilitated glucose transporters (GLUTs), but not sodium-dependent glucose transporters (SGLTs), the former of which are often more highly expressed in later stage tumours [85]. Moreover, whilst FDG-PET has been widely applied to monitor cancer therapy response, it has shown low accuracy in the context of some targeted therapies. Inflammatory phenotypes, in particular, have been well documented to compromise early assessment of response. An example that exemplifies this are melanomas that when treated with anti-CTLA4 or anti-PDL1 immunotherapies can often show heightened immune responses which would be detected on a PET scan as high FDG uptake. This suggests it would be harder to differentiate tumour relapse from response to immunotherapy [86]. FDG-PET also has limited applicability in studying certain tumours and metastases that are surrounded by tissue with high glucose uptake. Continuing on from the melanoma example, a common site of metastases is to the brain that shows the highest uptake of glucose in the body making it difficult for detection using FDG-PET. MRSI has the potential to improve detection and evaluate treatment response in brain metastases through evaluation of metabolite concentrations such as choline, creatine and N-acetylaspartate that change in tumours due to treatment [87]. While MRSI can aid in overcoming some of PET’s limitations, the lack of standard procedures and difficulty in the analysis of spectra, as well as the fact that hyperpolarised metabolites can only retain their polarised state for 20-30 seconds, limit its applicability in the clinic [87]. Both PET and MRSI are also limited in the species they can measure and fail to capture the whole tumour metabolome.
Compared to imaging techniques, analysis of biofluids allows convenient and sensitive profiling of a greater diversity of metabolic species. As a result, there have been efforts to use serum and plasma-derived metabolic biomarkers to detect cancer and monitor response. For specific tissue types of cancer, specific serum markers have been associated with tumour detection. Examples include L-octanoylcarnitine, 5-oxoproline, hypoxanthine, and docosahexaenoic acid for breast cancer [88] and glutamate, choline, 1,5-anhydro-D-glucitol, betaine, and methylguanidine amongst others. Another study focused on the presence of free amino acids in plasma and highlighted consistent biomarkers amongst many different types of cancer including lung, gastric, breast, colorectal and prostate cancers compared to the matched, healthy control specimens [89].
Although these studies have revealed potential biomarkers for diagnosis, they are quite limited in their cohort size, lack of independent cohort for validation and limited reproducibility due to differences in how metabolites are extracted and profiled between studies. To address these issues and identify metabolic biomarkers in an unbiased way, recent studies have utilised metabolomics that measures levels of many metabolites coupled with machine learning methods to identify subsets of metabolites whose levels can have diagnostic value. One recent example comes from Chen et. al, that analysed plasma metabolite levels of 147 metabolites from 702 patients comprising of three distinct cohorts from a multi-centre study [90]. Using LASSO regression to select the most relevant metabolites coupled with random forest classifiers, the authors were able to develop a diagnostic model based on 10 metabolites that accurately differentiated gastric cancer samples from healthy patients for all stages including stage 1A. Using a similar method, they also developed a prognostic model based on 28 metabolites that could predict prognosis and survival at different stages. Importantly, these models were able to outperform models that use existing clinical characteristics highlighting the usefulness of plasma metabolomics as a diagnostic and prognostic tool.
3.2. Metabolomic Profiling of Clinical Specimens to Understand Tumour Progression
Non-invasive profiling of tumours has great diagnostic and prognostic value, but has limited value in being able to understand tumour metabolic heterogeneity, the relationship between the tumour’s genome, transcriptome and metabolome, and to discover metabolic vulnerabilities that can aid therapeutic targeting. When surgical specimens become available, metabolomic analysis of tumours and adjacent normal tissues allows a more thorough understanding of metabolic reprogramming and metabolic interactions that occur between tumours and the tumour microenvironment. Mass spectrometry-based metabolomics have been applied widely for this purpose and relevant examples are reviewed in Section 4; this section will briefly examine applications of MSI.
MSI’s major power comes from preserving spatial metabolomic information and the ability to integrate other spatial multi-omic data, such as spatial genomics and transcriptomics. The latter has been exemplified by studies that investigated Myc metabolic reprogramming in breast cancer. Using DESI MSI analysis of tumours derived from inducible models of Myc, Kreuzaler et al. were able to demonstrate that upregulation of Myc led to higher levels of pantothenic acid, a precursor to coenzyme A, which is involved in a myriad of metabolic processes [14]. Importantly, they were able to demonstrate in tumours with heterogeneous Myc expression that pantothenic acid levels followed a distribution similar to the expression patterns of Myc. Of note, this Myc-associated metabolic biomarker was missed looking at the bulk metabolic analysis of Myc heterogeneous tumours, highlighting the spatial information preserved by DESI. In a separate study [86], DESI was also able to show spatiotemporal changes in metabolism associated with Myc amplification that led to lipogenesis induction through SREBP cooperation. This led to increases in phosphatidylglycerol (PG) species altering membrane and mitochondrial function.
Perhaps the most relevant application of DESI in recent times is its ability to study spatial metabolic heterogeneity and interaction of tumour cells with the TME as shown in a recent study on gastric cancer. Sun et al. revealed significant intratumor heterogeneity, identifying distinct tissue regions with varied metabolite, lipid, and gene expression signatures [91]. Notably, they observed stepwise metabolic alterations along the progression from normal epithelium to a serrated lesion to a tumour, shedding light on the underlying metabolic shifts driving gastric cancer development. Furthermore, the study highlighted the immunometabolic dynamics at the tumour interface region, where immune and inflammation-related signatures are prominent. By mapping the spatial transcriptome, the researchers identified a distinct “interface” cluster rich in immune cell populations, particularly plasma B cells and follicular B cells. Tumour cells at this interface were characterized by alterations in glutamine metabolism and increased levels of long-chain unsaturated fatty acids. The upregulation of genes involved in FA synthesis and arachidonic acid metabolism in PLT indicated an enhanced inflammatory response, potentially influencing immune cell functions and tumour cell proliferation. Overall, this study shows how MSI can provide comprehensive insights into the spatial heterogeneity and immunometabolic dynamics of cancer, unveiling novel molecular signatures and potential therapeutic targets.
3.3. Metabolomics Applied to Therapeutic Settings
Metabolic rewiring by cancer cells has been extensively studied and exploited to develop therapeutics to better treat tumours. This section briefly focuses on applications of metabolomics techniques to discover metabolic vulnerabilities or targets that can be used to improve existing therapies. For a more thorough review on metabolic therapies in cancer, readers are referred to the following excellent reviews [60,92,93].
LC-MS and GC-MS based methods have predominantly been used to identify potential metabolic targets for therapy and have been successfully applied to the clinic. Perhaps the most classical example is the development of IDH-1/2 mutant inhibitors used to treat acute myeloid leukaemia (AML) and gliomas. In IDH-1/2 mutant AML tumours, LC-MS revealed that there was accumulation of the D-2HG oncometabolite, supporting malignant transformation [94]. Targeting the mutant IDH-1/2 enzyme through Ivosidenib and Enasidenib greatly limited proliferation in these tumours and was subsequently approved by the FDA [95]. Since then, LC-MS has been used to discover various metabolic targets for therapy including enzymes involved in nucleotide synthesis, redox metabolism and amino acid metabolism. Further examples are examined in Section 4.
EFA methods can also be used to uncover relevant metabolic therapies pertaining to energy metabolism. Metabolic pathways related to the generation of ATP have been shown to be important not just for cancer cell’s energetic needs, but also for their signalling, differentiation, invasion and ability to metastasize. Accordingly, inhibitors of glycolysis [96], fatty acid metabolism, glutaminolysis, the TCA cycle and oxidative phosphorylation (OXPHOS) have been developed [97] which can be used to directly target tumours or sensitise them to existing therapies as part of combination therapy strategies [98]. EFA methods, such as the Seahorse assay, can unveil the metabolic pathways tumours primarily rely on and expose relevant metabolic vulnerabilities. An example of this was applied in studying NRAS-mutated melanoma which lacks any clinically approved targeted therapy [99]. The authors were able to use the Seahorse assay to show that NRAS-mutated melanoma cells were dependent on glucose and showed low fuel flexibility, even under glucose deprivation. This was followed up by showing that NRAS mutant tumours activated BRAF signalling that led to the phosphorylation of 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-2/3 (PFKFB2/PFKFB3) and allosteric activation of PFKM1. This led to a positive feedback cycle coupling glycolytic flux to RAS signalling. Subsequently, inhibition of glucose metabolism through a combination of 2-deoxy-D-glucose (2-DG) and sorafenib disrupted this cycle and inhibited tumour growth in patient derived xenografts. These studies illustrate how energy metabolism through EFA can be used to inform on appropriate metabolic therapies to treat certain tumour subsets.
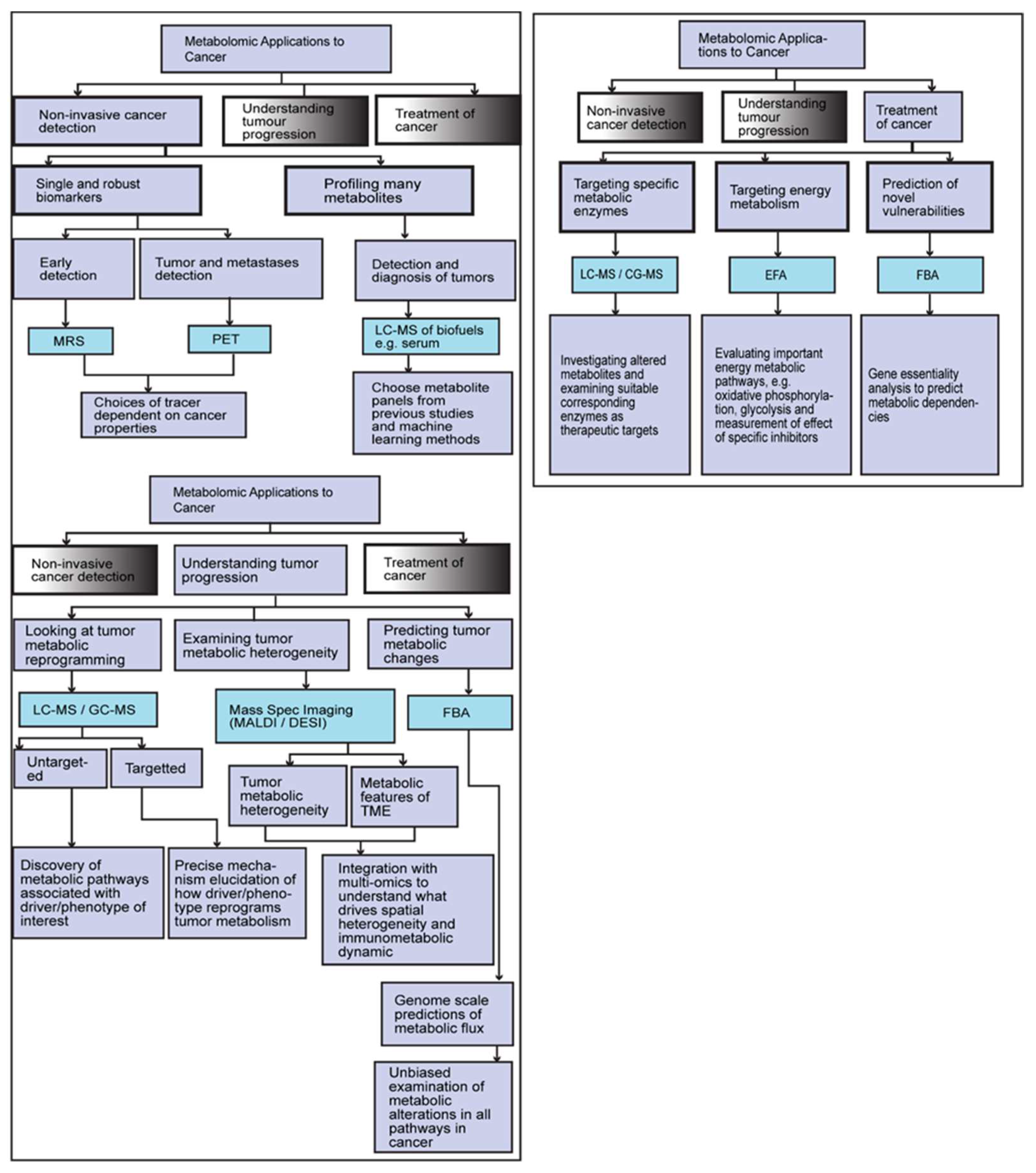
Figure 2.
Describing how different metabolic tracking tools and methodologies fit in with different applications for detecting, understanding and treating cancer.
Figure 2.
Describing how different metabolic tracking tools and methodologies fit in with different applications for detecting, understanding and treating cancer.
4. Metabolomics in Multimodal Profiling of Clinical Cohorts
Advancements in metabolomics technologies have now allowed better profiling of patients’ metabolomes. However, many clinical trials still omit profiling cancer metabolomes often choosing to focus on other cellular profiles such as the genome, transcriptome or the proteome [8]. Nevertheless, recent studies have shown that multimodal approaches that utilise metabolomics carries great diagnostic and prognostic potential in understanding tumour progression and identifying key metabolic vulnerabilities in patients. While it is not possible to review all these studies, this review will focus on two specific case studies and how metabolomics has been applied to patient cohorts to better understand the underlying cancer biology. The first concerns triple negative breast cancer in the Fudan cohort and the second studies a glioblastoma (GBM) cohort.
4.1. Fudan Cohort
Breast cancers are histologically characterised by the expression of receptor subtypes. Tumours not expressing the estrogen receptor (ER) or progesterone receptor (PR) and have no human epidermal growth factor receptor 2 (HER2; referred also to by its gene name ERBB2) amplification are classified as triple negative breast cancer (TNBC) [100]. TNBCs typically make up 10-20% of most breast cancer cases and are biologically more aggressive, often associated with the worst five-year prognosis with higher rates of early distant recurrence [101]. They are among the most heterogeneous subtypes of cancers, with different cases presenting various drivers and lacking more obvious targets such as the ER or HER2 receptors [102]. Because of the challenges in targeting TNBCs, there are limited treatment options. While there are some exceptions, such as the use of Olaparib [100] and an immune checkpoint inhibitor, Atezolizumab [103], for patients with germline mutations in BRCA genes and PD-L1+ tumours, respectively, there are relatively few approved targeted therapeutics for the treatment of TNBCs, with chemotherapy still remaining the standard of care. Therefore, there is an urgent need to identify targetable pathways for TNBCs.
To better understand the heterogeneity of the TNBC landscape and to find actionable targets, recent efforts have focused on multi-omic profiling of patients. One such example comes from the Fudan University Shanghai Cancer Center (FUSCC) that profiled over 465 Chinese TNBC patients [104]. Initially, researchers identified numerous subgroups within the cohort and demonstrated that their transcriptomic subtyping best explained clinical features. Accordingly, the FUSCC-TNBC cohort was classified into four subtypes: Luminal Androgen Receptor (LAR), immunomodulatory (IM), basal-like immune-suppressed (BLIS) and mesenchymal-like (MES) [104]. This was subsequently followed up with the FUTURE trial that used the enrichment of certain transcriptomic signatures and genomic mutations within each subgroup to inform on which targeted therapies patients were most likely to respond to. The IM and MES subtypes showed satisfactory objective response rates (ORR) to targeted therapies the latter of which achieved over 50% ORR. However, the two other subgroups LAR and BLIS did not meet expectations and showed ORRs under 20% [105].
Given that the initial analysis did not lead to good response rates for some TNBC subtypes, the authors expanded their study by integrating their transcriptomic subtyping with metabolomics data derived from LC-MS. In particular, they focused on finding metabolic vulnerabilities in the LAR and BLIS subtypes. To this end, 594 polar metabolites and 1944 lipids were profiled for 330 TNBC samples and 149 paired normal breast tissues making this one of the biggest metabolic atlases created for TNBCs [106]. Using the similarity fusion network method, the authors divided samples into three clusters: C1 that possessed upregulations in sphingolipid and fatty acid metabolism and predicted to have an increased dependence on fatty acids for energetic requirements; C2 that possessed an upregulation of glutamate pathways and hence likely to rely more on glutamate metabolism; and C3 that possessed only small metabolomic differences compared to normal tissue.
Subsequently, the authors investigated the correlation between the metabolic subtypes and their previous transcriptomics classification. In particular, the C1 subtype showed a strong correlation with the transcriptomic LAR subtype. The LAR TNBC subtype showed increased levels of ceramides; this was validated using tracing experiments that highlighted the increased de novo synthesis and degradation of ceramides consistent with the C1 subtype’s increased reliance on fatty acid metabolism. To reveal potential metabolic vulnerabilities, the authors used inhibitors of every step in the ceramide biosynthesis pathway and demonstrated that SPHK1 inhibition was most effective in reducing the viability of LAR patient derived organoids compared to non-LAR models. This approach highlights how metabolomics can complement other omics techniques to highlight specific vulnerabilities.
Association of the other TNBC transcriptomics subtypes with the metabolomics subtypes was less straightforward. However, using computational methods, the authors were able to establish that BLIS tumours that overlapped with the C2 metabolic subtype (BLIS-C2) showed a worse relapse-free survival compared to BLIS-C3 tumours. Using LASSO regression followed by a support vector machine classifier, the authors were able to distinguish BLIS-C2 from BLIS-C3 tumours based on the abundance of as few as 6 metabolites highlighting a potential role for metabolomics in predicting patient outcome. Using a similar approach as for the LAR tumours and focusing on BLIS-C2 with worse outcomes, the authors were able to identify NAAG as a metabolite of interest. RIMKLB, an enzyme that produces NAAG, was shown to be highly expressed in BLIS tumours and positively correlate with NAAG levels in the TNBC cohort. shRNA-mediated inhibition of RIMKLB led to a significant reduction in cancer cell proliferation, migration and invasion both in vitro, and in vivo that could be rescued by NAAG supplementation. RIMKLB, therefore, presents a novel target that can be used to treat the BLIS subtype which is otherwise refractory to most therapies.
To further investigate metabolic vulnerabilities in TNBC subtypes, the authors integrated the multi-omics data of their cohort to develop a ferroptosis atlas. Ferroptosis is an iron-dependent form of cell death driven by an overload of lipid peroxides on cellular membranes and has attracted considerable attention as it may be used to treat tumours resistant to chemotherapy or other targeted therapies. A critical feature of ferroptosis is its execution via phospholipid peroxidation which can be catalysed by lipoxygenases relying on iron, reactive oxygen species (ROS) and phospholipids containing primarily polyunsaturated fatty acid chains. On the other hand, cells possess three primary defence mechanisms that allows them to escape ferroptosis through detoxification of lipid hyperperoxides via the action of the glutathione peroxidase 4 (Gpx4), lipophilic antioxidants like coenzyme Q10 (CoQ10) and thioredoxin reductase which directly reduces lipid hydroperoxides by NADPH and selenocystine. Targeting these escape mechanisms could, therefore, be used to treat tumours that have high rates of lipid peroxides.
Using an integrated transcriptomic and metabolomic approach, the authors were able to demonstrate the LAR subtype had multiple pathways shown to be upregulated that are related to ferroptosis, including fatty acid synthesis, ROS accumulation and glutathione (GSH) metabolism. LAR tumours had higher rates of lipid peroxidation and were shown to upregulate the SLC7A11/GSH/GPX4 axis to escape ferroptosis. Using various pharmacological inhibitors for ferroptosis suppressors, the authors were able to show that targeting GPX4, that catalyses the oxidation of GSH to GSSG, through the inhibitor RSL3 led to much greater cell death in LAR tumours compared to other TNBC subtypes. Interestingly, the authors showed that GPX4’s expression was dependent on androgen receptor (AR) signalling but treating the tumours with AR inhibitors did not prime LAR tumours for ferroptosis. This is likely due to the plural role of AR in ferroptosis, with high AR expression leading to 2,4-Dienoyl-CoA Reductase 1 (DECR1) downregulation, whose knockdown selectively inhibits β-oxidation of PUFAs that serve as substrates for lipid peroxidation. As a result, GPX4, but not AR, inhibition led to ferroptosis in LAR tumours highlighting the importance of this specific metabolic target. The authors further showed that GPX4 inhibition in vivo led to reprogramming of the tumour microenvironment and increased infiltration of CD8+ T cells and M1 macrophages that exert anti-tumour effects. The authors thus suggested that a novel combination therapy of ferroptosis inducers and immune checkpoint inhibitors (ICIs) could show synergy to treat LAR tumours and significantly improve patient outcomes.
Overall, these studies underscore the value of performing metabolomics alongside other omics profiling technologies. While transcriptomics has led to better stratification of TNBCs, there is still an unmet clinical need in being able to predict therapies that these tumours would respond to. Using metabolomics, the authors were able to uncover new metabolic vulnerabilities that showed promising pre-clinical applications and demonstrated that metabolic therapies can also be used to potentiate other forms of treatment such as ICIs. This case study demonstrates the value of multi-modal pipeline for treatment refractory TNBCs.
4.2. Glioblastomas
Glioblastoma multiforme (GBM) is one of the most lethal and most common subtypes of central nervous system tumours and is often associated with very poor survival rates [107]. Much work has gone into studying GBM’s genomes and transcriptomes and yielded better understanding of molecular alterations that drive GBMs, including alterations in the core RTK/RAS/PI3K, p53 and RB signalling pathways [108]. Previous profiling have led to GBM’s stratification into four transcriptomic subtypes namely classical, mesenchymal, neural and proneural [109]. GBMs are also classified based on their IDH1/2 mutation status [110]. While studies have led to better knowledge of GBM drivers, GBMs display extensive genetic and transcriptomic heterogeneity that limits the efficacy of specific targeted therapies [111]. This has led to the majority of glioblastomas being incurable with current treatment options, highlighting an urgent clinical need to identify more effective therapeutic strategies for these tumours [112].
Similar to the work done for TNBC, multi-omics cohorts for GBMs have leveraged metabolomics to better understand GBM progression and discover metabolic vulnerabilities that can be used for treatment. One such study comes from Minami et al. that integrated genomics, transcriptomics and lipidomics from a diverse GBM cohort consisting of patient samples, orthotopic xenografts (PDXs) and gliomaspheres (GS) cell cultures [113]. Using shotgun lipidomics, the authors were able to identify 1020 different lipid species from 15 lipid subgroups making this one of the largest lipidomics studies performed for GBMs. They were able to separate the metabolic species into four lipid groups that contained species representing specific subclasses and FA saturation: L1 that mostly consisted of triacylglycerols (TAGs) with monounsaturated fatty acids (MUFAs), L2 which consisted of TAGs with polyunsaturated fatty acids (PUFAs), L3 that contained various phospholipid species including phosphatidylcholines (PCs), phosphatidylethanolamine (PEs), free fatty acids (FFAs) and diacylglycerols (DAGs), and L4 which primarily consisted of ether lipids and ceramides. These lipid clusters had strong correspondence with gene related metabolic signatures based on gene ontology enrichment showing a strong transcriptomic-metabolic relationship.
The authors were then able to leverage this relationship to identify cell intrinsic regulators of GBM metabolism. They identified that CDKN2A, which is deleted in up to 60% of GBMs, had an important role in tail length and saturation state of a significant subset of lipids in all the aforementioned lipid subgroups. More specifically, the authors showed that CDKN2A governed the partitioning of oxidizable PUFAs into TAGs where they were protected from lipid peroxidation. As CDKN2A null tumours lacked this ability, they had highly desaturated TAGs with shorter fatty acid tails. This led to an increase in proportion of oxidizable PUFAs in phospholipids in the cellular membrane and as a result, higher rates of lipid peroxidation. This primed CDKN2A null GBMs for ferroptosis and inhibition of GPX4 through RSL3 led to significant cell death in CDKN2A null tumours but not CDK2NA WT tumours. This vulnerability was recapitulated in vivo through mouse models that showed that GPX4 inhibition led to prolonged survival in CDK2NA null GBMs, but not CDK2NA WT tumours. Importantly, the authors were able to show that this trend was captured in primary tumours, but not cell lines, providing further evidence for the importance of conducting metabolomics in patient cohorts.
Overall, these studies highlight the importance of performing metabolomics studies in large patient cohorts. In the case of GBMs, metabolomics allowed for a better understanding of tumour progression and helped discover previously unknown roles of key cancer drivers such as CDK2NA’s role in PUFA partitioning, whilst in TNBCs, it was shown that it can be used to better predict survival in patients such as the BLIS transcriptomic subtype. Importantly, it also helped to unveil novel metabolic vulnerabilities that can be used to treat tumours refractory to therapies, as is the case for GPX4 inhibition in tumours with high rates of lipid peroxidation. These case studies thus highlight the multi-faceted role of metabolomics in patient cohorts.
5. Future Directions
Metabolomics has shown extensive applications for diagnosing and treating tumours but is most often used in isolation and not always integrated with other profiling modalities such as spatial omics or proteomics. Further advances in metabolomics and computational techniques will allow a more wholistic understanding of tumour biology. This section briefly looks at some analytic techniques that show promise in their application to cancer precision medicine.
5.1. Using Multi-Omics Tools and Machine Learning to Examine Tumour Metabolism and Predict Therapy Response
Multi-omics methods have been applied pre-clinically through cell line and mouse models to derive many insights about tumour metabolism and identify novel metabolic vulnerabilities. Pan-cancer analyses in cell lines have linked tumour metabolic changes to specific genetic alterations, epigenetic features or transcriptional dependencies. This study unveiled genomic factors driving tumour metabolic reprogramming including amplification of the malic enzyme 2 and proposed targeting of asparaginase metabolism in gastric cancer. Another study integrated metabolomics in cell lines with proteomics and transcriptomics to uncover transcriptional regulators of metabolism and crosstalk between the metabolic and transcriptional levels. In particular, they uncovered a relationship between glucose and one-carbon metabolism suggesting tumours that have lower glucose uptake rates could be sensitive to anti-folate drugs. This computational framework can exploit tumour metabolic profiles to find appropriate transcriptional regulators that underly targetable metabolic dysregulations. While these studies have shed light on novel metabolic targets, as evidenced in the glioblastoma study in Section 4, there is still discord between the applicability of tumour metabolic targets pre-clinically and in patients. Accordingly, multi-omics studies such as those outlined in Section 4 must be performed to gain more meaningful and physiologically relevant insights.
In cases where metabolomics data is available for patients, machine learning (ML) can aid in using the data to classify patients based on response to therapies or specific phenotypes of interest. LASSO regression has been used extensively to reduce dimensional space and reduce the number of metabolites associated with predictions of phenotypes such as therapy response or cancer staging. This allows for more targeted metabolic panels to be applied to ML algorithms such as support vector machines, random forest or deep learning classifiers. ML and multi-omics further can converge through building boosted classifiers. Boosted classifiers are composed of two or more sub-classifiers that make predictions based on their respective information modalities. Predictions from each classifier can then be integrated to make better predictions. An example of this was applied recently to predicting radiation response in tumours that used a boosted classifier. One classifier was based on targeted metabolite panel developed earlier mostly composing of redox metabolites and another was based on a transcriptomics-based classifier. Integrating both classifiers significantly improved the performance of the classifier which was able to achieve a sensitivity of over 90% underscoring the power ML and multi-omics possess when integrated.
5.2. Analytic Techniques Can Aid in Discovering Metabolic Biomarkers or Vulnerabilities
It may not always be possible to perform untargeted metabolomics due to cost or the inability to profile desirable subsets of the metabolome. In this case, analytic techniques such as FBA can be used to study metabolic reprogramming to home in on specific pathways or metabolic targets. FBA’s major advantage is its ability to generate metabolic fluxomic data from just transcriptomic data. With the advent of more complete reference genome scale metabolic models such as Human1, FBA’s accuracy in predicting human metabolic reprogramming has greatly improved over the past few years. A recent example of the application of FBA to study tumour metabolic reprogramming was to study metabolism of TNBC metastases. In particular, breast cancer metastases upregulated both oxidative phosphorylation and glycolysis compared to the matched primary tumours to increase metabolic flexibility in harsher nutrient conditions. FBA also revealed an increase in transport reaction flux indicating metastases take up more nutrients from the microenvironment rather than synthesize de novo owing to the scarcity of some nutrients in the microenvironment. Further applications of FBA can be used to bridge the gap between metabolomics and other profiling technologies.
5.3. Single Cell and Spatial Metabolomics Technologies
Single-cell and spatial technologies have enabled unprecedented studies into understanding tumour heterogeneity and the role of the tumour microenvironment. However, in the past few years, single cell omics has primarily focused on genomics, transcriptomics and proteomics while metabolomics has lagged. However, recent technological developments hold promise in enabling single cell metabolomics to be performed. One such framework, ScSpaMet, has recently been developed to allow a highly multiplex untargeted metabolomic imaging on tumour tissue. In this framework, cells are labelled with metal-isotope conjugated antibodies and metabolic profiling is performed by 3D-SMF [114] Combining this with spatial proteomics approaches, the authors characterized the tumour metabolic environment of lung tumour microarrays. Accordingly, they were able to identify differences in metabolism between tumour and stromal associated regions including differences in glycolysis and cholesterol metabolism. While this approach is limited to profiling a few metabolic fragments, further advances in metabolite annotation in mass spectra will allow a more detailed profiling of tumour metabolic heterogeneity to complement current MSI approaches.
6. Conclusions
Metabolic reprogramming in cancer has only recently been acknowledged as a significant hallmark and is often overlooked in favour of other hallmarks such as altered signalling pathways or the importance of escape from the immune system. This is in large part due to the lack of a singular technology to profile whole cell metabolomics unlike for other omics technologies such as genomics and transcriptomics. However, through combining different technologies, it is possible to examine a large subset of the tumour metabolome and focus on relevant pathways. When metabolomic techniques are applied to study tumour biology, they can unveil biomarkers and targeted therapies not achievable by other profiling technologies. Metabolic imaging can be used to sensitively detect and monitor response to therapy non-invasively while plasma and serum metabolomics allows detailed diagnostic and prognostic information to be gathered easily from patients. Metabolic profiling of tumour samples allows a detailed understanding of tumour heterogeneity and can reveal new clinical targets that can better treat subtypes of tumours that are often intractable to therapy, such as glioblastomas or TNBCs. Successful integration of metabolomics with other omics technologies will allow clinicians to further advance personalised metabolic biomarkers and identify effective therapies for patients with cancer.
Funding
Work in the GP lab was supported by UK Research and Innovation (MR/W012030/1 and MC_PC_ MR/X013715/1).
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discovery 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, R.; Maeda, K.; Aki, S.; Osawa, T. Metabolic adaptations of cancer in extreme tumor microenvironments. Cancer Science 2023, 114, 1200–1200. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Kreuzaler, P.; Panina, Y.; Segal, J.; Yuneva, M. Adapt and conquer: Metabolic flexibility in cancer growth, invasion and evasion. Molecular Metabolism 2020, 33, 83–83. [Google Scholar] [CrossRef] [PubMed]
- Demicco, M.; Liu, X.Z.; Leithner, K.; Fendt, S.M. Metabolic heterogeneity in cancer. Nature Metabolism 2024 6:1 2024, 6, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Danzi, F.; Pacchiana, R.; Mafficini, A.; Scupoli, M.T.; Scarpa, A.; Donadelli, M.; Fiore, A. To metabolomics and beyond: a technological portfolio to investigate cancer metabolism. Signal Transduction and Targeted Therapy 2023 8:1 2023, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Clish, C.B. Metabolomics: an emerging but powerful tool for precision medicine. Cold Spring Harbor Molecular Case Studies 2015, 1, a000588–a000588. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, E.; Liu, E.M.; Tang, C.; Kuo, F.; Buyukozkan, M.; Park, T.; Park, J.; Correa, F.; Hakimi, A.A.; Intlekofer, A.M.; et al. A multimodal atlas of tumour metabolism reveals the architecture of gene–metabolite covariation. Nature Metabolism 2023 5:6 2023, 5, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Trifonova, O.P.; Maslov, D.L.; Balashova, E.E.; Lokhov, P.G. Current State and Future Perspectives on Personalized Metabolomics. Metabolites 2023, 13. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. MASS SPECTROMETRY-BASED METABOLOMICS. Mass spectrometry reviews 2007, 26, 51–51. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; Julià, A. Analytical methods in untargeted metabolomics: State of the art in 2015. Frontiers in Bioengineering and Biotechnology 2015, 3, 130318–130318. [Google Scholar] [CrossRef] [PubMed]
- Mitochondrial Respiration XF Cell Mito Stress Test | Agilent.
- Gatidis, S.; Hepp, T.; Früh, M.; La Fougère, C.; Nikolaou, K.; Pfannenberg, C.; Schölkopf, B.; Küstner, T.; Cyran, C.; Rubin, D. A whole-body FDG-PET/CT Dataset with manually annotated Tumor Lesions. Scientific Data 2022 9:1 2022, 9, 1–7. [Google Scholar] [CrossRef]
- Kreuzaler, P.; Inglese, P.; Ghanate, A.; Gjelaj, E.; Wu, V.; Panina, Y.; Mendez-Lucas, A.; MacLachlan, C.; Patani, N.; Hubert, C.B.; et al. Vitamin B5 supports MYC oncogenic metabolism and tumor progression in breast cancer. Nature Metabolism 2023, 5, 1870–1870. [Google Scholar] [CrossRef] [PubMed]
- Liesenfeld, D.B.; Habermann, N.; Owen, R.W.; Scalbert, A.; Ulrich, C.M. Review of mass spectrometry-based metabolomics in cancer research. Cancer epidemiology, biomarkers & prevention : a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology 2013, 22, 2182–2182. [Google Scholar] [CrossRef]
- Moco, S. Studying Metabolism by NMR-Based Metabolomics. Frontiers in Molecular Biosciences 2022, 9, 882487–882487. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Nagana Gowda, G.A.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9. [Google Scholar] [CrossRef]
- Raja, G.; Jung, Y.; Jung, S.H.; Kim, T.J. 1H-NMR-based metabolomics for cancer targeting and metabolic engineering –A review. Process Biochemistry 2020, 99, 112–122. [Google Scholar] [CrossRef]
- Cortés, I.; Cuadrado, C.; Hernández Daranas, A.; Sarotti, A.M. Machine learning in computational NMR-aided structural elucidation. Frontiers in Natural Products 2023, 2, 1122426–1122426. [Google Scholar] [CrossRef]
- Emwas, A.H.M. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods in molecular biology (Clifton, N.J.) 2015, 1277, 161–193. [Google Scholar] [CrossRef] [PubMed]
- Bouziani, A.; Yahya, M.; Bouziani, A.; Yahya, M. Mass Spectrometry Coupled with Chromatography toward Separation and Identification of Organic Mixtures. Biodegradation Technology of Organic and Inorganic Pollutants 2021. [Google Scholar] [CrossRef]
- Bajo-Fernández, M.; Souza-Silva, É.A.; Barbas, C.; Rey-Stolle, M.F.; García, A. GC-MS-based metabolomics of volatile organic compounds in exhaled breath: applications in health and disease. A review. Frontiers in Molecular Biosciences 2023, 10, 1295955–1295955. [Google Scholar] [CrossRef] [PubMed]
- Krone, N.; Hughes, B.A.; Lavery, G.G.; Stewart, P.M.; Arlt, W.; Shackleton, C.H.L. Gas chromatography/mass spectrometry (GC/MS) remains a pre-eminent discovery tool in clinical steroid investigations even in the era of fast liquid chromatography tandem mass spectrometry (LC/MS/MS). The Journal of Steroid Biochemistry and Molecular Biology 2010, 121, 496–496. [Google Scholar] [CrossRef] [PubMed]
- Komal Pramod, S.; Amol Navnath, K.; Mahesh Pramod, S.; Shri Babanrao, H.; Komal Pramod Hon Shri Babanrao Pachpute Vichardhara, S. A REVIEW ON GAS CHROMATOGRAPHY-MASS SPECTROMETRY (GC-MS). Certified Journal │ 741 World Journal of Pharmaceutical Research SJIF Impact Factor 2021, 10, 741–763. [Google Scholar] [CrossRef]
- Pitt, J.J. Principles and Applications of Liquid Chromatography-Mass Spectrometry in Clinical Biochemistry. The Clinical Biochemist Reviews 2009, 30, 19–19. [Google Scholar]
- Buszewski, B.; Noga, S. Hydrophilic interaction liquid chromatography (HILIC)—a powerful separation technique. Analytical and Bioanalytical Chemistry 2012, 402, 231–231. [Google Scholar] [CrossRef]
- Ramautar, R. Capillary Electrophoresis-Mass Spectrometry for Clinical Metabolomics. Advances in clinical chemistry 2016, 74, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Ramautar, R.; Somsen, G.W.; de Jong, G.J. CE-MS for metabolomics: Developments and applications in the period 2016–2018. Electrophoresis 2019, 40, 165–165. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H.; Szczepski, K.; Al-Younis, I.; Lachowicz, J.I.; Jaremko, M. Fluxomics - New Metabolomics Approaches to Monitor Metabolic Pathways. Frontiers in Pharmacology 2022, 13, 805782–805782. [Google Scholar] [CrossRef]
- Pantel, A.R.; Ackerman, D.; Lee, S.C.; Mankoff, D.A.; Gade, T.P. Imaging Cancer Metabolism: Underlying Biology and Emerging Strategies. Journal of Nuclear Medicine 2018, 59, 1340–1340. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.Y.; Soloviev, D.; Brindle, K.M. Imaging Tumor Metabolism Using Positron Emission Tomography. Cancer journal (Sudbury, Mass.) 2015, 21, 129–129. [Google Scholar] [CrossRef]
- Kasem, J.; Wazir, U.; Mokbel, K. Sensitivity, Specificity and the Diagnostic Accuracy of PET/CT for Axillary Staging in Patients With Stage I-III Cancer: A Systematic Review of The Literature. In Vivo 2021, 35, 23–23. [Google Scholar] [CrossRef]
- Moses, W.W. Fundamental Limits of Spatial Resolution in PET. Nuclear instruments & methods in physics research. Section A, Accelerators, spectrometers, detectors and associated equipment 2011, 648 Supplement 1, S236. [Google Scholar] [CrossRef]
- Keshari, K.R.; Wilson, D.M. Chemistry and biochemistry of 13C hyperpolarized magnetic resonance using dynamic nuclear polarization. Chemical Society Reviews 2014, 43, 1627–1659. [Google Scholar] [CrossRef]
- Ardenkjær-Larsen, J.H.; Fridlund, B.; Gram, A.; Hansson, G.; Hansson, L.; Lerche, M.H.; Servin, R.; Thaning, M.; Golman, K. Increase in signal-to-noise ratio of >10,000 times in liquid-state NMR. Proceedings of the National Academy of Sciences of the United States of America 2003, 100, 10158–10163. [Google Scholar] [CrossRef]
- Brindle, K.M. Imaging cancer metabolism using magnetic resonance. npj Imaging 2024 2:1 2024, 2, 1–5. [Google Scholar] [CrossRef]
- Buchberger, A.R.; DeLaney, K.; Johnson, J.; Li, L. Mass Spectrometry Imaging: A Review of Emerging Advancements and Future Insights. Analytical chemistry 2018, 90, 240–240. [Google Scholar] [CrossRef] [PubMed]
- Aichler, M.; Walch, A. MALDI Imaging mass spectrometry: current frontiers and perspectives in pathology research and practice. Laboratory Investigation 2015 95:4 2015, 95, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Delgado, A.; Zhang, Z.; Bontempi, E.; Coccia, M.; Race, M.; Zhou, Y.; Wu, J.; Rong, Z.; Xiao, P.; Li, Y. Imaging Method by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry (MALDI-MS) for Tissue or Tumor: A Mini Review. Processes 2022, 10, 388–388. [Google Scholar] [CrossRef]
- Takáts, Z.; Wiseman, J.M.; Gologan, B.; Cooks, R.G. Mass spectrometry sampling under ambient conditions with desorption electrospray ionization. Science (New York, N.Y.) 2004, 306, 471–473. [Google Scholar] [CrossRef]
- Calligaris, D.; Caragacianu, D.; Liu, X.; Norton, I.; Thompson, C.J.; Richardson, A.L.; Golshan, M.; Easterling, M.L.; Santagata, S.; Dillon, D.A.; et al. Application of desorption electrospray ionization mass spectrometry imaging in breast cancer margin analysis. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 15184–15189. [Google Scholar] [CrossRef] [PubMed]
- Schwamborn, K.; Caprioli, R.M. Molecular imaging by mass spectrometry — looking beyond classical histology. Nature Reviews Cancer 2010 10:9 2010, 10, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Eberlin, L.S.; Ferreira, C.R.; Dill, A.L.; Ifa, D.R.; Cooks, R.G. Desorption electrospray ionization mass spectrometry for lipid characterization and biological tissue imaging. Biochimica et biophysica acta 2011, 1811, 946–960. [Google Scholar] [CrossRef] [PubMed]
- He, M.J.; Pu, W.; Wang, X.; Zhang, W.; Tang, D.; Dai, Y. Comparing DESI-MSI and MALDI-MSI Mediated Spatial Metabolomics and Their Applications in Cancer Studies. Frontiers in Oncology 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Schwaiger-Haber, M.; Stancliffe, E.; Anbukumar, D.S.; Sells, B.; Yi, J.; Cho, K.; Adkins-Travis, K.; Chheda, M.G.; Shriver, L.P.; Patti, G.J. Using mass spectrometry imaging to map fluxes quantitatively in the tumor ecosystem. Nature Communications 2023 14:1 2023, 14, 1–17. [Google Scholar] [CrossRef]
- Dueñas, M.E.; Larson, E.A.; Lee, Y.J. Toward mass spectrometry imaging in the metabolomics scale: Increasing metabolic coverage through multiple on-tissue chemical modifications. Frontiers in Plant Science 2019, 10, 449752–449752. [Google Scholar] [CrossRef]
- Ren, J.L.; Zhang, A.H.; Kong, L.; Wang, X.J. Advances in mass spectrometry-based metabolomics for investigation of metabolites. RSC Advances 2018, 8, 22335–22350. [Google Scholar] [CrossRef] [PubMed]
- Caines, J.K.; Barnes, D.A.; Berry, M.D. The Use of Seahorse XF Assays to Interrogate Real-Time Energy Metabolism in Cancer Cell Lines. Methods in molecular biology (Clifton, N.J.) 2022, 2508, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Salabei, J.K.; Gibb, A.A.; Hill, B.G. Comprehensive measurement of respiratory activity in permeabilized cells using extracellular flux analysis. Nature Protocols 2014 9:2 2014, 9, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Mercier-Letondal, P.; Marton, C.; Godet, Y.; Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine 2021, 19, 21–21. [Google Scholar] [CrossRef]
- Targeting Energy Metabolism for Cancer Therapeutic Discovery using Agilent Seahorse XF Technology - Drug Discovery World (DDW).
- Purohit, V.; Wagner, A.; Yosef, N.; Kuchroo, V.K. Systems-based approaches to study immunometabolism. Cellular & Molecular Immunology 2022 19:3 2022, 19, 409–420. [Google Scholar] [CrossRef]
- Argüello, R.J.; Combes, A.J.; Char, R.; Gigan, J.P.; Baaziz, A.I.; Bousiquot, E.; Camosseto, V.; Samad, B.; Tsui, J.; Yan, P.; et al. SCENITH: A flow cytometry based method to functionally profile energy metabolism with single cell resolution. Cell metabolism 2020, 32, 1063–1063. [Google Scholar] [CrossRef] [PubMed]
- Vaysse, P.M.; Demers, I.; van den Hout, M.F.C.M.; van de Worp, W.; Anthony, I.G.M.; Baijens, L.W.J.; Tan, B.I.; Lacko, M.; Vaassen, L.A.A.; van Mierlo, A.; et al. Evaluation of the Sensitivity of Metabolic Profiling by Rapid Evaporative Ionization Mass Spectrometry: Toward More Radical Oral Cavity Cancer Resections. Analytical Chemistry 2022, 94, 6939–6947. [Google Scholar] [CrossRef] [PubMed]
- Barlow, R.S.; Fitzgerald, A.G.; Hughes, J.M.; McMillan, K.E.; Moore, S.C.; Sikes, A.L.; Tobin, A.B.; Watkins, P.J. Rapid Evaporative Ionization Mass Spectrometry: A Review on Its Application to the Red Meat Industry with an Australian Context. Metabolites 2021, 11, 171–171. [Google Scholar] [CrossRef] [PubMed]
- Koundouros, N.; Karali, E.; Tripp, A.; Valle, A.; Inglese, P.; Perry, N.J.S.; Magee, D.J.; Anjomani Virmouni, S.; Elder, G.A.; Tyson, A.L.; et al. Metabolic Fingerprinting Links Oncogenic PIK3CA with Enhanced Arachidonic Acid-Derived Eicosanoids. Cell 2020, 181, 1596–1611.e27. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.D.; Thiele, I.; Palsson, B.O. What is flux balance analysis? Nature Biotechnology 2010 28:3 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.L.; Kocabaş, P.; Wang, H.; Cholley, P.E.; Cook, D.; Nilsson, A.; Anton, M.; Ferreira, R.; Domenzain, I.; Billa, V.; et al. An atlas of human metabolism. Science Signaling 2020, 13, 1482–1482. [Google Scholar] [CrossRef]
- Colarusso, A.V.; Goodchild-Michelman, I.; Rayle, M.; Zomorrodi, A.R. Computational modeling of metabolism in microbial communities on a genome-scale. Current Opinion in Systems Biology 2021, 26, 46–57. [Google Scholar] [CrossRef]
- Schmidt, D.R.; Patel, R.; Kirsch, D.G.; Lewis, C.A.; Heiden, M.G.V.; Locasale, J.W. Metabolomics in cancer research and emerging applications in clinical oncology. CA: A Cancer Journal for Clinicians 2021, 71, 333–358. [Google Scholar] [CrossRef] [PubMed]
- Armitage, E.G.; Barbas, C. Metabolomics in cancer biomarker discovery: Current trends and future perspectives. Journal of Pharmaceutical and Biomedical Analysis 2014, 87, 1–11. [Google Scholar] [CrossRef]
- Kdadra, M.; Höckner, S.; Leung, H.; Kremer, W.; Schiffer, E. Metabolomics Biomarkers of Prostate Cancer: A Systematic Review. Diagnostics 2019, Vol. 9, Page 21 2019, 9, 21–21. [Google Scholar] [CrossRef]
- Long, N.P.; Yoon, S.J.; Anh, N.H.; Nghi, T.D.; Lim, D.K.; Hong, Y.J.; Hong, S.S.; Kwon, S.W. A systematic review on metabolomics-based diagnostic biomarker discovery and validation in pancreatic cancer. Metabolomics 2018 14:8 2018, 14, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Poli, D.; Carbognani, P.; Corradi, M.; Goldoni, M.; Acampa, O.; Balbi, B.; Bianchi, L.; Rusca, M.; Mutti, A. Exhaled volatile organic compounds in patients with non-small cell lung cancer: Cross sectional and nested short-term follow-up study. Respiratory Research 2005, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, B.M.; Stirdivant, S.M.; Mitchell, M.W.; Wulff, J.E.; McDunn, J.E.; Li, Z.; Dennis-Barrie, A.; Neri, B.P.; Milburn, M.V.; Lotan, Y.; et al. Bladder Cancer Biomarker Discovery Using Global Metabolomic Profiling of Urine. PLOS ONE 2014, 9, e115870–e115870. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Y.; Zhao, W.; Deng, K.; Wang, Z.; Yang, C.; Ma, L.; Openkova, M.S.; Hou, Y.; Li, K.; et al. Metabolomics for biomarker discovery in the diagnosis, prognosis, survival and recurrence of colorectal cancer: a systematic review. Oncotarget 2017, 8, 35460–35472. [Google Scholar] [CrossRef] [PubMed]
- Use of PET for monitoring cancer therapy and for predicting outcome - PubMed.
- Almuhaideb, A.; Papathanasiou, N.; Bomanji, J. 18F-FDG PET/CT Imaging In Oncology. Annals of Saudi Medicine 2011, 31, 3–3. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Wang, C.; Zhao, S.; Xie, R.; Zhao, L.; Li, K.; Yang, C.; Zhang, R.; Tian, Y.; Tan, L.; et al. The clinical application of 18F-FDG PET/CT in pancreatic cancer: a narrative review. Translational Cancer Research 2021, 10, 3560–3560. [Google Scholar] [CrossRef] [PubMed]
- Griffeth, L.K. Use of PET/CT scanning in cancer patients: technical and practical considerations. Proceedings (Baylor University. Medical Center) 2005, 18, 321–321. [Google Scholar] [CrossRef]
- Verger, A.; Kas, A.; Darcourt, J.; Guedj, E. PET Imaging in Neuro-Oncology: An Update and Overview of a Rapidly Growing Area. Cancers 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.S.; Grudzinski, J.; Smith, G.T.; Peterson, T.E.; Whisenant, J.G.; Hickman, T.L.; Ciombor, K.K.; Cardin, D.; Eng, C.; Goff, L.W.; et al. First-in-Human PET Imaging and Estimated Radiation Dosimetry of l-[5-11C]-Glutamine in Patients with Metastatic Colorectal Cancer. Journal of nuclear medicine : official publication, Society of Nuclear Medicine 2022, 63, 36–43. [Google Scholar] [CrossRef]
- Role and therapeutic targeting of glutamine metabolism in non-small cell lung cancer (Review).
- Miner, M.W.; Liljenbäck, H.; Virta, J.; Merisaari, J.; Oikonen, V.; Westermarck, J.; Li, X.G.; Roivainen, A. (2S, 4R)-4-[18F]Fluoroglutamine for In vivo PET Imaging of Glioma Xenografts in Mice: an Evaluation of Multiple Pharmacokinetic Models. Molecular Imaging and Biology 2020, 22, 969–969. [Google Scholar] [CrossRef]
- Umbehr, M.H.; Müntener, M.; Hany, T.; Sulser, T.; Bachmann, L.M. The Role of 11C-Choline and 18F-Fluorocholine Positron Emission Tomography (PET) and PET/CT in Prostate Cancer: A Systematic Review and Meta-analysis. European Urology 2013, 64, 106–117. [Google Scholar] [CrossRef]
- Wu, Z.; Zha, Z.; Li, G.; Lieberman, B.P.; Choi, S.R.; Ploessl, K.; Kung, H.F. [18F](2S,4S)-4-(3-Fluoropropyl)glutamine as a tumor imaging agent. Molecular Pharmaceutics 2014, 11, 3852–3866. [Google Scholar] [CrossRef] [PubMed]
- Chaumeil, M.M.; Najac, C.; Ronen, S.M. Studies of Metabolism Using (13)C MRS of Hyperpolarized Probes. Methods in enzymology 2015, 561, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.J.; Kurhanewicz, J.; Vigneron, D.B.; Larson, P.E.Z.; Harzstark, A.L.; Ferrone, M.; Van Criekinge, M.; Chang, J.W.; Bok, R.; Park, I.; et al. Metabolic imaging of patients with prostate cancer using hyperpolarized [1-13C]pyruvate. Science Translational Medicine 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Albers, M.J.; Bok, R.; Chen, A.P.; Cunningham, C.H.; Zierhut, M.L.; Zhang, V.Y.; Kohler, S.J.; Tropp, J.; Hurd, R.E.; Yen, Y.F.; et al. Hyperpolarized 13C Lactate, Pyruvate, and Alanine: Noninvasive Biomarkers for Prostate Cancer Detection and Grading. Cancer Research 2008, 68, 8607–8615. [Google Scholar] [CrossRef]
- Day, S.E.; Kettunen, M.I.; Gallagher, F.A.; Hu, D.E.; Lerche, M.; Wolber, J.; Golman, K.; Ardenkjaer-Larsen, J.H.; Brindle, K.M. Detecting tumor response to treatment using hyperpolarized 13C magnetic resonance imaging and spectroscopy. Nature Medicine 2007 13:11 2007, 13, 1382–1387. [Google Scholar] [CrossRef]
- de Kouchkovsky, I.; Chen, H.Y.; Ohliger, M.A.; Wang, Z.J.; Bok, R.A.; Gordon, J.W.; Larson, P.E.Z.; Frost, M.; Okamoto, K.; Cooperberg, M.R.; et al. Hyperpolarized 1-[13C]-Pyruvate Magnetic Resonance Imaging Detects an Early Metabolic Response to Immune Checkpoint Inhibitor Therapy in Prostate Cancer. European Urology 2022, 81, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Mair, R.; Wright, A.J.; Ros, S.; Hu, D.e.; Booth, T.; Kreis, F.; Rao, J.; Watts, C.; Brindle, K.M. Metabolic imaging detects low levels of glycolytic activity that vary with levels of C-MyC expression in patient-derived xenograft models of glioblastoma. Cancer Research 2018, 78, 5408–5418. [Google Scholar] [CrossRef]
- Fala, M.; Ros, S.; Sawle, A.; Rao, J.U.; Tsyben, A.; Tronci, L.; Frezza, C.; Mair, R.; Brindle, K.M. The role of branched-chain aminotransferase 1 in driving glioblastoma cell proliferation and invasion varies with tumor subtype. Neuro-Oncology Advances 2023, 5, 1–16. [Google Scholar] [CrossRef]
- Gallagher, F.A.; Kettunen, M.I.; Hu, D.E.; Jensen, P.R.; Zandt, R.I.t.; Karlsson, M.; Gisselsson, A.; Nelson, S.K.; Witney, T.H.; Bohndiek, S.E.; et al. Production of hyperpolarized [1,4-13C2]malate from [1,4-13C2]fumarate is a marker of cell necrosis and treatment response in tumors. Proceedings of the National Academy of Sciences of the United States of America 2009, 106, 19801–19806. [Google Scholar] [CrossRef]
- Barrio, J.R.; Huang, S.C.; Satyamurthy, N.; Scafoglio, C.S.; Yu, A.S.; Alavi, A.; Krohn, K.A. Does 2-FDG PET Accurately Reflect Quantitative In Vivo Glucose Utilization? Journal of Nuclear Medicine 2020, 61, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Gouw, A.M.; Margulis, K.; Liu, N.S.; Raman, S.J.; Mancuso, A.; Toal, G.G.; Tong, L.; Mosley, A.; Hsieh, A.L.; Sullivan, D.K.; et al. The MYC Oncogene Cooperates with Sterol-Regulated Element-Binding Protein to Regulate Lipogenesis Essential for Neoplastic Growth. Cell metabolism 2019, 30, 556–572.e5. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Andronesi, O.; Bogner, W.; Choi, I.Y.; Coello, E.; Cudalbu, C.; Juchem, C.; Kemp, G.J.; Kreis, R.; Krššák, M.; et al. Minimum Reporting Standards for in vivo Magnetic Resonance Spectroscopy (MRSinMRS): Experts' consensus recommendations. Nmr in Biomedicine 2021, 34. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Shin, Y.; Kim, T.H.; Kim, D.H.; Lee, A. Plasma metabolites as possible biomarkers for diagnosis of breast cancer. PLOS ONE 2019, 14, e0225129–e0225129. [Google Scholar] [CrossRef]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Miura, T.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma Free Amino Acid Profiling of Five Types of Cancer Patients and Its Application for Early Detection. PLOS ONE 2011, 6, e24143–e24143. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, B.; Zhao, Y.; Shao, X.; Wang, M.; Ma, F.; Yang, L.; Nie, M.; Jin, P.; Yao, K.; et al. Metabolomic machine learning predictor for diagnosis and prognosis of gastric cancer. Nature Communications 2024 15:1 2024, 15, 1–13. [Google Scholar] [CrossRef]
- Sun, C.; Wang, A.; Zhou, Y.; Chen, P.; Wang, X.; Huang, J.; Gao, J.; Wang, X.; Shu, L.; Lu, J.; et al. Spatially resolved multi-omics highlights cell-specific metabolic remodeling and interactions in gastric cancer. Nature Communications 2023 14:1 2023, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Halma, M.T.J.; Tuszynski, J.A.; Marik, P.E. Cancer Metabolism as a Therapeutic Target and Review of Interventions. Nutrients 2023, 15, 4245–4245. [Google Scholar] [CrossRef]
- Xiao, Y.; Yu, T.J.; Xu, Y.; Ding, R.; Wang, Y.P.; Jiang, Y.Z.; Shao, Z.M. Emerging therapies in cancer metabolism. Cell metabolism 2023, 35, 1283–1303. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.J.; Metallo, C.M. Metabolic consequences of oncogenic IDH mutations. Pharmacology & Therapeutics 2015, 152, 54–62. [Google Scholar] [CrossRef]
- Rohde, J.M.; Karavadhi, S.; Pragani, R.; Liu, L.; Fang, Y.; Zhang, W.; McIver, A.; Zheng, H.; Liu, Q.; Davis, M.I.; et al. Discovery and optimization of 2H-1λ2-pyridin-2-one inhibitors of mutant isocitrate dehydrogenase 1 for the treatment of cancer. Journal of medicinal chemistry 2021, 64, 4913–4913. [Google Scholar] [CrossRef] [PubMed]
- Gill, K.S.; Fernandes, P.; O'Donovan, T.R.; McKenna, S.L.; Doddakula, K.K.; Power, D.G.; Soden, D.M.; Forde, P.F. Glycolysis inhibition as a cancer treatment and its role in an anti-tumour immune response. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2016, 1866, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Phan, L.M.; Yeung, S.C.J.; Lee, M.H. Cancer metabolic reprogramming: importance, main features, and potentials for precise targeted anti-cancer therapies. Cancer Biology & Medicine 2014, 11, 1–1. [Google Scholar] [CrossRef]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting cancer metabolism in the era of precision oncology. Nature Reviews Drug Discovery 2021 21:2 2021, 21, 141–162. [Google Scholar] [CrossRef] [PubMed]
- McGrail, K.; Granado-Martínez, P.; Esteve-Puig, R.; García-Ortega, S.; Ding, Y.; Sánchez-Redondo, S.; Ferrer, B.; Hernandez-Losa, J.; Canals, F.; Manzano, A.; et al. BRAF activation by metabolic stress promotes glycolysis sensitizing NRASQ61-mutated melanomas to targeted therapy. Nature Communications 2022, 13, 1–22. [Google Scholar] [CrossRef]
- Zagami, P.; Carey, L.A. Triple negative breast cancer: Pitfalls and progress. npj Breast Cancer 2022, 8, 1–10. [Google Scholar] [CrossRef]
- Herrera Juarez, M.; Tolosa Ortega, P.; Sanchez De Torre, A.; Ciruelos Gil, E. Biology of the Triple-Negative Breast Cancer: Immunohistochemical, RNA, and DNA Features. Breast Care 2020, 15, 208–208. [Google Scholar] [CrossRef] [PubMed]
- Irshad, S.; Ellis, P.; Tutt, A. Molecular heterogeneity of triple-negative breast cancer and its clinical implications. Current opinion in oncology 2011, 23, 566–577. [Google Scholar] [CrossRef]
- Sharmni Vishnu, K.; Win, T.T.; Aye, S.N.; Basavaraj, A.K. Combined atezolizumab and nab-paclitaxel in the treatment of triple negative breast cancer: a meta-analysis on their efficacy and safety. BMC Cancer 2022, 22, 1139–1139. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.D.; Liu, Y.R.; Yu, Y.; et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer Cell 2019, 35, 428–440.e5. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xiao, Y.; Ding, J.H.; Jin, X.; Ma, D.; Li, D.Q.; Shi, J.X.; Huang, W.; Wang, Y.P.; Jiang, Y.Z.; et al. Ferroptosis heterogeneity in triple-negative breast cancer reveals an innovative immunotherapy combination strategy. Cell metabolism 2023, 35, 84–100.e8. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, D.; Yang, Y.S.; Yang, F.; Ding, J.H.; Gong, Y.; Jiang, L.; Ge, L.P.; Wu, S.Y.; Yu, Q.; et al. Comprehensive metabolomics expands precision medicine for triple-negative breast cancer. Cell research 2022, 32, 477–490. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pacific Journal of Cancer Prevention : APJCP 2017, 18, 3–3. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, M.Y.; Assem, M. Glioblastoma Genomics: A Very Complicated Story. Glioblastoma 2017, 3–25. [Google Scholar] [CrossRef]
- Sidaway, P. Glioblastoma subtypes revisited. Nature Reviews Clinical Oncology 2017 14:10 2017, 14, 587–587. [Google Scholar] [CrossRef]
- Cohen, A.L.; Holmen, S.L.; Colman, H. IDH1 and IDH2 Mutations in Gliomas. Current neurology and neuroscience reports 2013, 13, 345–345. [Google Scholar] [CrossRef]
- García-Montaño, L.A.; Licón-Muñoz, Y.; Martinez, F.J.; Keddari, Y.R.; Ziemke, M.K.; Chohan, M.O.; Piccirillo, S.G.M. Dissecting Intra-tumor Heterogeneity in the Glioblastoma Microenvironment Using Fluorescence-Guided Multiple Sampling. Molecular Cancer Research 2023, 21, 755–755. [Google Scholar] [CrossRef] [PubMed]
- Huse, J.T.; Holland, E.; DeAngelis, L.M. Glioblastoma: molecular analysis and clinical implications. Annual review of medicine 2013, 64, 59–70. [Google Scholar] [CrossRef]
- Minami, J.K.; Morrow, D.; Bayley, N.A.; Fernandez, E.G.; Salinas, J.J.; Tse, C.; Zhu, H.; Su, B.; Plawat, R.; Jones, A.; et al. CDKN2A deletion remodels lipid metabolism to prime glioblastoma for ferroptosis. Cancer Cell 2023, 41, 1048–1060.e9. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Allam, M.; Cai, S.; Henderson, W.; Yueh, B.; Garipcan, A.; Ievlev, A.V.; Afkarian, M.; Beyaz, S.; Coskun, A.F. Single-cell spatial metabolomics with cell-type specific protein profiling for tissue systems biology. Nature Communications 2023, 14, 1–20. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



