
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
Bacteria of the genus Pseudomonas are the most studied microorganisms in the biodestruction of persistent perfluoroorganic pollutants, and the research on their application for the remediation of environmental sites using biotechnological methods remains relevant. The aim of this study was to investigate the ability of a known destructor of perfluorooctane sulfonic acid from the genus Pseudomonas to accelerate and enhance the depth of destruction of long-chain perfluorocarboxylic acids (PFCAs), specifically perfluorooctanoic acid and perfluorononanoic acid, in water and soil in association with the strain P. mosselii 5(3), which has previously confirmed genetic potential for degrading PFCAs. The complete genome (length 5.86 million base pairs) of the strain 2,4-D, a new species of Pseudomonas, was sequenced, assembled, and analyzed. The genomes of both strains contain genes involved in the defluorination of fluorinated compounds, including haloacetate dehalogenase H-1 (dehH1) and haloalkane dehalogenase (dhaA). The strain 2,4-D also has a multi-component enzyme system consisting of a dioxygenase component, an electron carrier, and 2-halobenzoate 1,2-dioxygenase (CbdA) with a preference for fluorides. The strain 2,4-D was able to completely decarboxylate and defluorinate PFCAs in an aqueous cultivation system within 7 days, using them as the sole source of carbon and energy, but it only converted them to perfluorheptanoic acid. However, it assisted strain 5(3) (which defluorinated them during the same period to perfluorohexanoic acid) in converting PFCAs to perfluoropentanoic acid, accelerating the process by 24 hours. The degradation and transformation of the anionic acids were assessed using LC-MS analysis, and the release of indicators for mineralization of fluoride ions was determined by ion chromatography. In a model experiment for bioaugmentation of microorganisms in artificially contaminated soil, the degradation of PFCAs by the association of pseudomonads also occurred faster and deeper than by the individual strains, achieving a degree of biodestruction of 75% over 60 days, with perfluoropentanoic acid being the main metabolite of destruction. Thus, it has been demonstrated that strains of Pseudomonas bacteria with genetic potential for the biodegradation of PFCAs are capable of realizing this potential in association, exhibiting an additive effect.
Keywords:
perfluorocarboxylic acids
; biodegradation
; association of bacteria
; Pseudomonas
; transformation
; genome
1. Introduction
Halogenated chemicals are continuously entering various environmental objects due to industrial pollution and persist there for decades [1]. Perfluorocarboxylic acids (PFCA) are industrial chemicals that, due to the characteristics of their chemical structure and their high chemical stability and resistance, have been widely used for over 50 years. The widespread use of PFCA, particularly as fire-fighting foams in the military and mining sectors, along with their recently identified toxicity, highlights the scale of soil and groundwater contamination by these substances worldwide [2,3,4].
Alongside the well-known perfluorooctanoic acid (PFOA), perfluorononanoic acid (PFNA) can also be highlighted. The area of application for PFNA includes its use as a polymerization additive in the production of fluoropolymers [5] in the commercial product Surflon S-111 (CAS 72968-3-88, 74% by weight). PFNA is very stable and is known not to degrade in the environment due to oxidative processes because of the strength of the carbon-fluorine bond. Like the eight-carbon PFOA, the nine-carbon PFNA is a toxic substance for development of the organism and the immune system [6], and it can disrupt the human thyroid hormone system, with possible negative consequences for pregnancy and the subsequent development of the fetus and child [7]. Their toxicity has been repeatedly confirmed for both animal and human health [8,9]. They not only move with water [10,11] but also accumulate in soils [12,13], thus entering cultivated food crops [14,15]. Due to their combined toxic properties concerning living organisms, they have been included in Annex B of the Stockholm Convention on Persistent Organic Pollutants [16].
Accordingly, the issues related to their degradation are relevant and require solutions. Initially, various physical methods (pyrolysis, membrane and adsorption separation, sonolysis) [17,18,19] and chemical methods (ozonation, plasma, photocatalytic and electrochemical reactions) [20,21,22] were used for this purpose (degradation, defluorination, destruction of PFCAs). However, considering their high costs for energy supply, multi-stage pre-treatment, and the formation of toxic by-products [23], microbial destruction, primarily bacterial, is a relevant alternative. Biocatalytic technologies for the degradation of perfluoroorganic compounds are promising, environmentally friendly, and economical in terms of energy use. Although two to three decades ago, PFCAs were referred to as 'forever chemicals,' and the inability of microorganisms to degrade them was attributed to a lack of time for their evolution and the absence of natural analogs to stimulate the evolution of the necessary enzymes [24]. But perhaps the time has come, and it is precisely 'here and now' that progress has been made in the cleavage of robust chemical bonds in persistent halogenated organic compounds, leading to their transformation into harmless substances. Initially, microorganisms learned to break carbon covalent bonds that do not contain fluorides (for example, CS, CO) during the biodegradation of perfluorocarboxylic (sulfonic) acids [25,26,27,28]. Not stopping there, they progressed to the effective cleavage of C-F bonds, thereby reducing the chemical stability of the organic molecule of perfluoroalkyl compounds and making it available for further degradation [29], while also expanding the range of perfluoroalkyl compounds used as substrates by microorganisms to C7 - C10, even confirming defluorination through the detection of released fluoride ions [30].
In recent years, it has been shown that bacteria and their communities are capable of metabolizing certain PFCAs, forming a range of metabolites. One of the proposed pathways for microbial destruction of fluorinated organic acids involves oxidation to perfluorohexanoic acid, which has been demonstrated for the strain Gordonia sp. NB4-1Y [31]. Prior to this, several strains of bacteria from the genus Pseudomonas with similar activity were known [32,33,34].
Recent reviews have touched upon the limited published materials regarding the enzymes and molecular tools of microorganisms involved in the destruction and transformation of PFCA through the release of fluoride ions and protection against its toxic effects [35,36]. So far, the number of microorganisms and enzymes demonstrating activity in this area remains limited. A list has been presented, including dehalogenases of halogenated acids, reductive dehalogenases, laccases, desulfonases, and mechanisms of microbial resistance to intracellular fluoride.
Enzymatic biocatalysts for breaking C-F bonds under mild conditions have been discovered relatively recently, and they generally do not require the use of cofactors [37]. For example, several related dehalogenases with defluorination potential were found in the genome of the strain Delftia acidovaerens D4B, isolated from soil contaminated with PFCAs, which were later identified as DeHa2 (haloacid dehalogenase) and DeHa4 (fluoroacetate dehalogenase) [38,39]. One of the latest reports in this cluster of studies concerned the discovery of the rdhA gene, which plays a key role in the defluorination of PFCAs by the strain Acidimicrobium sp. A6 [40].
We previously demonstrated that the strain Pseudomonas mosselii 5(3) is capable of genetically and effectively degrading C7-C10 PFCAs to C6 [30], while the strain Pseudomonas sp. 2,4-D also exhibited biodestructive activity towards perfluorooctane sulfonate, breaking it down to perfluoroenanthic acid [28]. Given the high potential for a possible additive effect in the destruction of PFCAs through the associative use of these strains, this study involved the sequencing of the genome of a representative new species of the genus Pseudomonas strain 2,4-D, examining its genetic features and biodegradation potential to increase the depth and rate of PFCA destruction when cultivated in an aqueous environment and through the bioaugmentation of microorganisms in artificially contaminated soil.
2. Materials and Methods
Bacterial strains and conditions for their cultivation in liquid medium.
Strain 5(3), identified by whole genome sequencing as Pseudomonas mosselii, capable of degrading C7-C10 PFCA [30], was isolated from pesticide-contaminated soil in the Yanaul district of the Republic of Bashkortostan. The project of its genome has been deposited in GenBank under the number JAUHUJ000000000 and the BioSample and BioProject numbers SAMN36271085 and PRJNA990579, respectively.
Strain 2,4-D was isolated from a soil sample taken from a facility producing halogenated organic compounds (Ufa, Republic of Bashkortostan), previously identified as P. plecoglossicida 2,4-D and described as a destructor of perfluorooctane sulfonic acid [28].
Strains 2,4-D and 5(3) are deposited in the Microorganism Collection of the Ufa Institute of Biology, Subdivision of the Ufa Federal Research Centre of the Russian Academy of Sciences, under numbers UIB-53 and UIB-251.
The strains were cultivated at 28 °C in liquid Raymond mineral medium (per liter of distilled water): NH4NO3–2.0, MgSO4 × 7H2O–0.2, KH2PO4–2.0, Na2HPO4–3, CaCl2 × 6H2O–0.01, Na2CO3–0.1 [41], with PFOA or PFNA (250 mg/l) as the sole source of carbon and energy on an orbital shaker-incubator ES-20/60 (SIA BIOSAN, Riga, Latvia) at 180 rpm.
To obtain the inoculum, the strain was cultivated on Raymond's mineral medium with the addition of peptone (1 g/L) for 24 hours. The biomass was harvested and washed with sterile distilled water before inoculation into experimental flasks. The resulting pellet was resuspended in mineral medium and added to the experimental flasks until an OD600 of 0.1 was reached.
For the estimation of bacterial quantity and the isolation of individual colonies, LB medium [42] was used, consisting of (per liter of distilled water) 10 g of tryptone, 5 g of yeast extract, 5 g of NaCl, and 15 g of agar (PanReac, Barcelona, Spain).
To assess the effectiveness of PFCA degradation, the bacterium was cultured at 28 °C in a liquid mineral medium with individual C8-C9 PFCAs for 7 days. All experiments were conducted in three independent biological replicates.
Chemicals and reagents
Perfluorocarboxylic acids (PFCAs) - perfluorononanoic acid (PFNA) and perfluorooctanoic acid (PFOA) (both of high purity > 98%) were purchased from Sigma Aldrich (St. Louis, Missouri, USA). Acetonitrile and methanol (HPLC grade) were acquired from Merck (Darmstadt, Germany). All other reagents were of analytical grade.
Sequencing, assembly, and annotation of the genome.
The genomic DNA of strain 2,4-D was extracted from biomass using the phenol-chloroform method [43]. Sequencing was performed on DNBSEQ-G50 equipment (MGI, China), and paired-end reads of 100 bp were obtained.
Quality control of the reads was performed using 'HTQC' [44]. Low-quality (Q<25), short (<100 bp) reads and adapter sequences were removed using Trimomatic version 0.39 [45]. Genomes were assembled using SPAdes software version 3.15.4 [46]. Error correction was carried out using Pilon version 1.23 [47] and Bowtie2 version 2.3.5.1 [48]. To confirm the presence of the assembled circular replicon, we analyzed the presence of overlapping ends. For strain identification, average nucleotide identity (ANI) parameters (https://www.ezbiocloud.net/tools/ani [49]) and digital DNA-DNA hybridization (DDH) (https://ggdc.dsmz.de/ggdc.php [50]) were used with default settings.
Contigs shorter than 500 bp were removed. Annotation was performed using Prokka version 1.14.5 [51]. Analysis of target gene sequences was conducted using the NCBI BLAST service [52], utilizing the nr/nt and WGS databases.
The FASTME 2.1.6.1 program [53] was used to construct the minimum evolutionary tree based on the obtained intergenomic distances. The tree was rooted in the middle [54] and visualized using https://itol.embl.de/ (accessed October 2, 2024) [55]. Species and subspecies were grouped according to [56] and [57], respectively. The OrthoANI algorithm [58] was used to calculate the average nucleotide identity (ANI) between strains and related strains.
Model experiment on bioaugmentation of strains 2,4-D and 5(3) and their association in soil artificially contaminated with PFCAs.
For the model experiment, we used chernozem soil collected from an organic farming enterprise (Republic of Bashkortostan, Russia) that was not contaminated with chemical organic compounds. The experiment included four lines, each treated with PFOA or PFOS to a final concentration of 10 mg/kg of dry soil: 1) soil (Control); 2) soil augmented with the bacterial culture of strain 2,4-D (final culture concentration 2×106 CFU/g of soil) (2,4-D); 3) soil augmented with the bacterial culture of strain 5(3) (final culture concentration 2×106 CFU/g of soil) (5(3)); 4) soil augmented with an association of bacterial cultures of strains 2,4-D and 5(3) (final culture concentration 2×106 CFU/g of soil). The cultivation was conducted at 28±1°C, and the soil moisture was maintained at 40% of the soil's moisture capacity. The number of microorganisms was assessed using dilution method on Raymond's medium with the corresponding PFCA as a sole carbon source. The duration of the experiment was 60 days. Extraction of PFAS from soil samples was carried out using a modified method from Rankin et al. [59]. The prepared sample (3.5 g of dry soil) was transferred into polypropylene centrifuge tubes, previously washed with methanol. For PFAS extraction, a mixture of acetonitrile and water in a 9:1 ratio (8.5 ml) was used, with the addition of 400 µl of a 2 M NaOH solution. After adding the reagents, the samples were subjected to ultrasonic treatment in an ice bath for 15 minutes, followed by shaking on a shaker for 30 minutes and centrifugation for 10 minutes at 12,000 rpm. The resulting supernatants from the three extraction steps were combined in glass tubes. For sample preparation for analysis, the combined supernatant was evaporated to dryness under a stream of air. The dry residue was dissolved in two consecutive washes of acetonitrile and water in a 2:3 ratio, each with 0.5 ml. To assess the bacterial count, samples were inoculated onto Raymond's medium with the corresponding PFCA every 10 days of the experiment. All experiments were conducted in three independent biological replicates.
The efficiency of PFCA destruction was calculated using the formula
D(%) =100 – ((Сt×100)/ C0 ),
where D – is the destruction efficiency (%); Ct – is the concentration of PFCA after a certain period of time; C0 – is the concentration of PFCA at the initial moment in time.
Isolation and Identification of Biotransformation Products of PFCA
The content of PFCA in the medium was assessed and the products of their biotransformation were identified using a liquid tandem chromatography-mass spectrometer LCMS-IT-TOF (“Shimadzu”, Japan) at the Shared Equipment Center “AGIDEL” of the Ufa Scientific Center of the Russian Academy of Sciences, in ultrafiltrates (≤ 3 kDa) of culture media obtained by ultrafiltration using the “Vivaflow 50” system (Sartorius AG, Göttingen, Germany), as described in [28].
For chromatographic separation, a Shim-pack XR-ODS column (75 mm × 2.0 mm id, 2.2 μm) (Shimadzu, Kyoto, Japan) was used with a mobile phase consisting of 5 mM ammonium acetate in acetonitrile (A) and 0.1% acetic acid (v/v) in water (B). Elution was performed with a linear gradient as follows: from 0 min to 10 min, from 60% to 30% (B); from 10 to 20 min, from 30% to 60% (B). The separation was conducted at a flow rate of 0.25 ml/min, with an injection volume of 5 μl. Mass spectra were obtained using electrospray ionization (ESI) in negative ion mode with the following parameters: high voltage probe: -3.5 kV; nebulizer gas flow: 1.5 L/min; CDL temperature: 200 °C; heating block temperature: 50 °C; drying gas pressure: 150 kPa; TOF detector voltage: 1.57 kV. A trifluoroacetic acid solution was used as a standard sample for sensitivity and resolution calibration, as well as for mass calibration (ion trap and time-of-flight analyzer).
The structure of the obtained compounds was established based on a combination of mass spectrometry data, which focused on the fragmentation of the molecular ion, and comparisons with literature data. The release of fluoride ions was determined using ion chromatography with an LC-20 Prominence HPLC system equipped with a CDD-10Avp conductivity detector (Shimadzu, Japan). The analyte separation was performed on a Shodex column (Shodex, New York, NY, USA) at a flow rate of 1 mL/min with an eluent composed of 1.8 mM Na2CO3 + 1.7 mM NaHCO3, using the Xenoic® XAMS ASUREX-A100 suppressor (Diduco AB, Umeå, Sweden).
The metabolic pathways were drawn using ChemDraw Ultra v. 12.0.2.1076.
Statistical processing and data analysis
The statistical analysis was performed using Microsoft Office Excel 2021. All experiments were conducted in triplicate. Standard methods of parametric statistics were used to describe the research results: the arithmetic mean and standard deviation were calculated. The comparison of two groups was carried out using the two-tailed Student's t-test. The critical level of significance was set at 0.05 for this study.
3. Results
3.1. Identification of the Strain 2,4-D and Functional Annotation of Its Genome
Previously, based on analyses of the physiological-biochemical properties and the 16S rRNA gene, the strain was identified as Pseudomonas plecoglossicida [28]. The same study demonstrated its ability to transform perfluorooctanesulfonic acid. In the current work, we present the results of sequencing and obtaining a reliable draft assembly of the genome of the pure culture, which were used for identification and subsequent functional analysis. Sequencing and complete assembly of the genome of the strain Pseudomonas sp. 2,4-D revealed a circular chromosomal replicon of 5,861,101 bp (GC content: 61.84%), with other assembly characteristics presented in Figure 1A. The genome project for strain 2,4-D has been deposited in GenBank under the number JAUHUI000000000 and the BioSample and BioProject numbers SAMN36271038 and PRJNA990562, respectively. Based on whole-genome sequencing, an attempt was made to determine the phylogenetic relationship between the studied strain and its closest type strains on the tree (Figure 1B). Following the recommendations in [60], a comparison of the genome of the studied strain was conducted with the genomes of closely related type strains P. kurunegalensis RW1P2, P. putida NBRC14164, and P. monteilii DSM14164 using Average Nucleotide Identity (ANI) and Digital DNA–DNA Hybridization (DDH) parameters. The ANI values ranged from 89.34% to 89.83%, while the DDH values ranged from 56.40% to 60.30%. According to the criteria for describing new bacterial species, these values should not be lower than 95% and 70%, respectively, compared to the type strains of known species.
None of the parameters allow the studied strain to meet the species threshold for classification as a specific species, and it is likely that the strain Pseudomonas sp. 2,4-D represents a new species. As shown by the results of the functional annotation, the genome contains genes for all the metabolic pathways necessary for the existence of an autonomous culture (Figure 1C). Considering that detailed systematics of this genus is still lacking, and despite the long history of study and extensive databases, the genus is annually supplemented with new species.
Using the KEGG metabolic pathway and network analysis programs, it was found that the genome of the strain encodes several genes potentially involved in the biodegradation and metabolism of xenobiotics. This category contains 34 genes. The genome of the 2,4-D strain was analyzed for genetic elements potentially associated with the degradation of PFCA. It contains, similar to the genome of the PFCA-degrading strain 5(3) [30], genes potentially involved in the defluorination of fluorinated compounds, such as haloacetate dehalogenase H-1 (dehH1), haloalkane dehalogenase (dhaA), and alkane sulfonate monooxygenase (SsuE). However, unlike strain 5(3), the genome of the 2,4-D strain does not contain a fluoride ion transporter (CrcB), but it does have a multicomponent enzyme system consisting of a dioxygenase component, an electron transport protein, and a 2-halobenzoate 1,2-dioxygenase large subunit (CbdA) with a preference for fluorides, which has not been previously mentioned in the context of PFCA.
The presence of genes for haloacetate dehalogenase H-1 (dehH1), haloalkane dehalogenase (dhaA), or similar ones is also characteristic of other PFCA-degrading organisms, particularly Delftia acidovaerens D4B [38]. The A6 strain has been found to contain genes encoding a homolog of reductive dehalogenase (RdhA), a homolog of fluoroacetate dehalogenase (FceA), as well as two putative genes for halogen acid dehalogenases (dhl_1 and dhl_2). Molecular dynamics simulations have been performed, and the docking of the enzyme complex in the corresponding configuration for PFAS substrates has been investigated [40].
The absence of the fluoride ion transporter gene (CrcB) likely negatively impacts the viability of the culture during the transformation of PFOS and the inability to overcome the C7 barrier due to the toxic effects of the released fluoride. This is also supported by data indicating that effective defluorination activity will require a high level of tolerance to elevated intracellular concentrations of F- in the microbial host [61]. The discovery of the putative fluoride ion transporter, the CrcB gene, in the A6 genome highlights a significant problem posed by the toxicity of fluoride, a byproduct of the cleavage of the C–F bond during metabolism. The lack of a rapid and efficient mechanism for fluoride export would render this recently developed metabolic pathway detrimental to the organism, ultimately leading to its exclusion from the population. As noted in [40], this underscores the need for a reliable enzymatic apparatus, potentially involving ATP-consuming processes, to catalyze the cleavage of C–F bonds and mitigate the toxic effects of very high fluoride levels [62].
A multicomponent enzyme system (with hydroxylation, defluorination, and decarboxylation properties), consisting of a dioxygenase component, an electron carrier, and the 2-halobenzoate 1,2-dioxygenase large subunit (CbdA) originally from P. cepacia 2CBS [63], which catalyzed the double hydroxylation of 2-halobenzoates with the concomitant release of halide and carbon dioxide, but has broad substrate specificity and is likely involved in the transformation of PFCA by strain 2,4-D.
Among the associated genes, it is worth highlighting the presence of the alternative vanadium nitrogenase gene VnfA, which is capable of supplying microorganisms with nitrogen by converting it into ammonia under microaerobic conditions (and low temperatures) to sustain their vitality [64]. Their hosts are used in associations to enhance various biotechnological processes [65].
The genes identified in the genomes do not always function or do not function as researchers assume. Therefore, the actual activity of the enzymes they encode in defluorination processes still requires confirmation. Nevertheless, the process of biodegradation of 'forever chemicals' has been initiated, and it seems plausible that such degrading enzymes are involved in it.
3.2. Biodegradation and Defluorination of PFCA
Both studied Pseudomonas strains, as previously shown [28,30], actively grew in mineral media with perfluorinated organic acids as sole source of energy and carbon, reaching maximum microbial numbers in the culture liquid after 7 days, completely transforming the substrate during this time. The difference lay in the depth of transformation: strain 5(3) degraded them only to C6 PFCA, while strain 2,4-D degraded them only to perfluoroenanthic acid. Accordingly, an association of these microorganisms was formed to overcome this barrier. Associations or consortia of microorganisms are generally more effective than their individual counterparts, as they consist of different strains with broad and complementary enzymatic capabilities.
In the current study, individual strains required 7 days for the complete degradation of PFCA, but in association, they accomplished this 24 hour faster with both PFOA (Figure 2A) and PFNA (Figure 2C). This was confirmed by the release of fluoride ions into the medium (up to 30 mg/L), with the level of fluoride release in the case of the microbial association exceeding that of the individual strains by 1.6 to 2.2 times.
Considering the special non-traditional properties of PFCA, they are analyzed using liquid chromatography/tandem mass spectrometry (Figure 3A), the detection of fluoride ions released during the biodegradation process is carried out using conductometric HPLC (Figure 3B).
The conducted LCMS analysis also showed a difference in the depth of transformation of PFCAs by microorganisms and their association (Figure 2B, D). At the end of cultivation in a medium with PFOA (Figure 2B), for strain 2,4-D, we observed dissociated acid ions (with m/z 363) of PFHpA, for strain 5(3) – mostly PFHxA (with m/z 313), and for their association – mostly PFPeA (with m/z 263). Similarly, the degradation of PFNA (Figure 2D) occurs under the influence of bacteria and their association. The obtained products differ in mass by the amount of CF2- groups cleaved by bacterial dehalogenases, i.e., by m/z 50. Depending on the acid, the bacteria undergo a different number of cycles.
Such bacterial activity of biodestruction of perfluorinated substances confirmed both genetically and through the mineralization of fluorine, has not been reported by the authors in the literature.
To date, the closest result achieved involved the use of the bacterial strain Acidimicrobium sp. A6 for the removal of PFOA and PFOS in aerobic conditions, where the degree of biodegradation did not exceed 63% for a concentration of 100 mg/L over 100 days [19]. The removal of PFOA and PFOS has also been documented in systems with microbial consortia, showing a 16% reduction with chemoorganotrophic bacteria and a 36% reduction with a consortium of yeast and mold over 28 days. Their LC/MS analysis shows the production of monofluorinated fatty acids [66]. Additionally, data on the degradation of various perfluoroalkyl substances by a consortium of microorganisms is presented in study [67], which detected PFOA, PFHxA, and PFHpA as metabolites in aqueous solutions after 10 months of incubation.
For the defluorination of PFCA, bacteria need to transport it into the cell and produce an active enzyme that catalyzes the cleavage of the C−F bond. The destructive capabilities of the studied strains are also supported by partially published data [68] regarding the extraction from their culture supernatants, partial purification, and molecular characterization of extracellular dehalogenases (with molecular weights of 72 and 80 kDa) that vary in structure and possibly in mechanisms of action.
Considering that strain 5(3) specializes in the degradation of PFCA, while strain 2,4-D focuses on the destruction of PFOS, we conclude that the latter enhances the depth and speed of PFCAs degradation. The genetically determined presence of a multienzyme system with broad substrate specificity, exhibiting decarboxylation, hydroxylation, and defluorination properties (the presence of the CbdA gene), further supports this conclusion. Additionally, the overcoming of the physiological barrier of fluoride toxicity in these bacteria during the degradation of PFCA is explained by the presence of a fluoride ion transporter (CrcB), which allows them to bypass this barrier and conduct defluorination at a new level in association.
3.3. Model Experiment on Bioaugmentation of PFCA-Contaminated Soil
Bioremediation requires various microorganisms to break down and transform pollutants into energy, cellular biomass, and harmless byproducts. The effectiveness of bioremediation strategies can be enhanced through bioaugmentation—introducing specific biodegraders to accelerate the degradation of contaminants [69]. This method is also used in contexts involving halogenated organics [70,71] and PFAS [72].
In the present study, model soil systems were developed to evaluate the potential application of a bacterial consortium as a bioremediation agent. It was demonstrated that the introduction of bacteria reduced the concentration of PFCA in the soil by 22-25% (strain 2,4-D) and 38-40% (strain 5 (3)) over a period of 60 days (Figure 4 A, D). The microbial titers in the soil increased linearly during this time, reaching values at the end of the experiment that differed from the initial levels by slightly more than an order of magnitude (Figures 4 B, F). The use of the bacterial consortium resulted in a significant enhancement of PFCA degradation efficiency in the soil, achieving levels of 75-78%, which was corroborated by improved microbial survival in the soil (the titer increased by two orders of magnitude over the course of the experiment). It should be noted that in model systems lacking the introduced microorganisms, there was virtually no degradation of PFCA. According to LCMS analysis, it can be observed (Figures 4 C, E) that the degradation of PFCA in the soil differs in metabolite profiles from biodegradation in aquatic environments. In all augmented soil experiments, a dissociated acid ion (with m/z 263) corresponding to perfluoropentanoic acid was detected, and in the trials with the consortia, it was the predominant species, indicating accelerated transformation of the parent PFCA specifically in their presence.
Similar studies demonstrating comparable effectiveness in the biodegradation of PFCA in soil have not been identified by the authors. The further degradation of PFCA and their complete bacterial mineralization is likely a question that will be answered in the near future through continued investigation into the catabolic capabilities of the microorganisms studied.
In response to the question posed in reference [72]: "Rehabilitation of soils contaminated with poly- and perfluoroalkyl substances (PFAS): to mobilize, immobilize, or decompose?" we provide a clear answer: "Decomposition through microorganisms, even considering that the removal of PFAS in soil using bacteria will be complicated in practice due to the complex mixture of PFAS compounds in the contaminated environment and limited biodegradation.
4. Conclusions
The capabilities of the well-known destructor of perfluorooctanesulfonic acid from the genus Pseudomonas strain 2,4-D to accelerate and enhance the degradation depth of long-chain perfluorocarboxylic acids (PFCAs), specifically perfluorooctanoic acid and perfluorononanoic acid, in water and soil in association with strain P. mosselii 5(3)—which has previously been genetically confirmed to possess significant potential for degrading PFCA—were studied. The complete genome (length 5.86 million base pairs) of the destructor Pseudomonas sp. strain 2,4-D, isolated from soil contaminated with halogenated organic compounds (previously identified as P. plecoglossicida 2,4-D), was sequenced, assembled, and analyzed. Based on the average nucleotide identity and digital DNA-DNA hybridization compared to type strains of pseudomonads, the studied microorganism likely belongs to a new species. Both strains genomes contain genes potentially involved in the defluorination of fluorinated organic compounds, including haloacetate dehalogenase H-1 (dehH1), haloalkane dehalogenase (dhaA), and alkane sulfonate monooxygenase (SsuE); however, unlike strain 5(3), the genome of strain 2,4-D does not contain the fluoride ion transporter (CrcB) but features a multi-component enzyme system (with hydroxylation, defluorination, and decarboxylation properties) consisting of a dioxygenase component, an electron transfer component, and the large subunit of 2-halobenzoate 1,2-dioxygenase (CbdA) with a preference for fluorides. Strain 2,4-D was capable of completely decarboxylating and defluorinating PFCAs in a cultivation medium within 7 days, using them as the sole carbon and energy source, but only to perfluorheptanoic acid; it facilitated strain 5(3) (which defluorinated them to perfluorohexanoic acid in the same time frame) in converting PFCAs to perfluoropentanoic acid, accelerating the process by 24 hours. The degradation and transformation of the anionic acids were assessed using HPLC-MS analysis, while the release of mineralization indicators for fluoride ions was determined by ion chromatography. In a model experiment on the bioaugmentation of microorganisms in artificially contaminated soil, the degradation of PFCAs by the association of pseudomonads occurred more rapidly and deeply than by the strains individually, achieving a biodegradation rate of 75-78% over 60 days, with perfluoropentanoic acid being the primary degradation metabolite. Thus, it was demonstrated that bacterial strains of the genus Pseudomonas with genetic potential for PFCA biodegradation can effectively realize this potential in association, exhibiting an additive effect.
Author Contributions
Conceptualization, S.C.; methodology, S.C., S.S. and D.S.; software, G.H.; validation, S.C. and S.S.; formal analysis; investigation, D.S., S.S.; resources, S.C.; data curation, S.C.; writing—original draft preparation, S.C.; writing—review and editing, S.C. and G.H.; visualization, G.H., D.S. and S.S.; supervision, S.C.; project administration, S.C.; funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Russian Science Foundation, grant number 23-24-00154.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The project of its genome has been deposited in GenBank under the number JAUHUJ000000000 and the BioSample and BioProject numbers SAMN36271085 and PRJNA990579, respectively.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Baggi, G.; Bernasconi, S.; Zangrossi, M.; Cavalca, L.; Andreoni, V. Co-metabolism of di- and trichlorobenzoates in a 2-chlorobenzoate-degrading bacterial culture: Effect of the position and number of halo-substituents. Int. Biodeterior. Biodegrad. 2008, 62, 57–64. [CrossRef]
- Groffen, T.; Wepener, V.; Malherbe, W.; Bervoets, L. Distribution of perfluorinated compounds (PFASs) in the aquatic environment of the industrially polluted Vaal River, South Africa. Sci. Total Environ. 2018, 627, 1334–1344. [CrossRef]
- Jian, J.-M.; Guo, Y.; Zeng, L.; Liang-Ying, L.; Lu, X.; Wang, F.; Zeng, E.Y. Global distribution of perfluorochemicals (PFCs) in potential human exposure source–A review. Environ. Int. 2017, 108, 51–62. [CrossRef]
- Toms, L.M.L.; Br€aunig, J.; Vijayasarathy, S.; Phillips, S.; Hobson, P.; Aylward, L.L.; Kirk, M.D.; Mueller, J.F. Per- and polyfluoroalkyl substances (PFAS) in Australia: current levels and estimated population reference values for selected compounds. Int. J. Hyg Environ. Health. 2019, 222, 387–394. [CrossRef]
- Prevedouros, K.; Cousins, I.T.; Buck, R.C.; Korzeniowski, S.H. Sources, fate and transport of perfluorocarboxylates. Environ Sci Technol. 2006, 40 (1), 32–44. [CrossRef]
- Fang, X.; Zhang, L.; Feng, Y.; Zhao, Y.; Dai, J. Immunotoxic effects of perfluorononanoic acid on BALB/c mice. Toxicol. Sci. 2008, 105(2), 312–21. [CrossRef]
- Coperchini, F.; Croce, L.; Ricci, G.; Magri, F.; Rotondi, M.; Imbriani, M.; Chiovato, L. Thyroid Disrupting Effects of Old and New Generation PFAS. Front. Endocrinol. 2021, 11, 612320. [CrossRef]
- Domingo, J.L.; Nadal, M. Human exposure to per- and polyfluoroalkyl substances (PFAS) through drinking water: A review of the recent scientific literature. Environ Res. 2019, 177, 108648. [CrossRef]
- Peritore, A.F.; Gugliandolo, E.; Cuzzocrea, S.; Crupi, R.; Britti, D. Current Review of Increasing Animal Health Threat of Per- and Polyfluoroalkyl Substances (PFAS): Harms, Limitations, and Alternatives to Manage Their Toxicity. Int J Mol Sci. 2023, 24(14), 11707. [CrossRef]
- Post, G.B.; Cohn, P.D.; Cooper, K.R. Perfluorooctanoic acid (PFOA), an emerging drinking water contaminant: a critical review of recent literature. Environ Res. 2012, 116, 93-117. [CrossRef]
- Pan, Y.; Zhang, H.; Cui, Q.; Sheng, N.; Yeung, L.W.Y.; Sun, Y.; Guo, Y.; Dai, J. Worldwide Distribution of Novel Perfluoroether Carboxylic and Sulfonic Acids in Surface Water. Environ Sci Technol. 2018, 52(14), 7621-7629. [CrossRef]
- Zhu, Q.; Qian, J.; Huang, S.; Li, Q.; Guo, L.; Zeng, J.; Zhang, W.; Cao, X.; Yang, J. Occurrence, distribution, and input pathways of per- and polyfluoroalkyl substances in soils near different sources in Shanghai. Environ Pollut. 2022, 308, 119620. [CrossRef]
- Sim, W.; Park, H.; Yoon, J.K.; Kim, J.I.; Oh, J.E. Characteristic distribution patterns of perfluoroalkyl substances in soils according to land-use types. Chemosphere. 2021, 276, 130167. [CrossRef]
- Li, J.; Sun, J.; Li, P. Exposure routes, bioaccumulation and toxic effects of per- and polyfluoroalkyl substances (PFASs) on plants: A critical review. Environ Int. 2022, 158, 106891. [CrossRef]
- Xu, K.; Huang, J.; Zhang, Y.; Wu, X.; Cai, D.; Hu, G.; Li, Y.; Ni, Z.; Lin, Q.; Wang, S.; Qiu, R. Crop Contamination and Human Exposure to Per- and Polyfluoroalkyl Substances around a Fluorochemical Industrial Park in China. Toxics. 2024, 12(4), 269. [CrossRef]
- Report of the Conference of the Parties of the Stockholm Convention on Persistent Organic Pollutants on the Work of Its Fourth Meeting, 4–8 May//UNEP/POPS/COP.4/38; Stockholm Convention Secretariat: Geneva, Switzerland, 2009; pp. 66–69.
- Cheng, J.; Vecitis, C.D.; Park, H.; Mader, B.T.; Hoffmann, M.R. Sonochemical degradation of perfluorooctane sulfonate (PFOS) and perfluorooctanoate (PFOA) in landfill groundwater: environmental matrix effects. Env. Sci. Technol. 2008, 42, 8057–8063. [CrossRef]
- Wang, F.; Shih, K.; Lu, X.; Liu, C. Mineralization behavior of fluorine in perfluorooctanesulfonate (PFOS) during thermal treatment of lime-conditioned sludge. Env. Sci. Technol. 2013, 47, 2621–2627. [CrossRef]
- Kabiri, S.; Navarro, D.A.; Hamad, S.A.; Grimison, C.; Higgins, C.P.; Mueller, J.F.; Kookana, R.S.; McLaughlin, M.J. Physical and chemical properties of carbon-based sorbents that affect the removal of per- and polyfluoroalkyl substances from solution and soil. Sci Total Environ. 2023, 875, 162653. [CrossRef]
- Cao, C.S.; Wang, J.; Yang, L.; Wang, J.; Zhang, Y.; Zhu, L. A review on the advancement in photocatalytic degradation of poly/perfluoroalkyl substances in water: Insights into the mechanisms and structure-function relationship. Sci Total Environ. 2024, 946, 174137. [CrossRef]
- Liu X, He S, Yang Y, Yao B, Tang Y, Luo L, Zhi D, Wan Z, Wang L, Zhou Y. A review on percarbonate-based advanced oxidation processes for remediation of organic compounds in water. Environ Res. 2021, 200, 111371. [CrossRef]
- Yang, L.; He, L.; Xue, J.; Ma, Y.; Xie, Z.; Wu, L.; Huang, M.; Zhang, Z. Persulfate-based degradation of perfluorooctanoic acid (PFOA) and perfluorooctane sulfonate (PFOS) in aqueous solution: Review on influences, mechanisms and prospective. J Hazard Mater. 2020, 393, 122405. [CrossRef]
- Marquínez-Marquínez, A.N.; Loor-Molina, N.S.; Quiroz-Fernández, L.S.; Maddela, N.R.; Luque, R.; Rodríguez-Díaz, J.M. Recent advances in the remediation of perfluoroalkylated and polyfluoroalkylated contaminated sites. Environmental Research. 2023, 219, 115152. [CrossRef]
- Wackett, L.P.; McMahon, K. Why Is the biodegradation of polyfluorinated compounds so rare? mSphere. 2021, 6(5): Article e00721–21. [CrossRef]
- Ruiz-Urigüen, M.; Shuai, W.; Huang, S.; Jaffé, P.R. Biodegradation of PFOA in microbial electrolysis cells by Acidimicrobiaceae sp. strain A6. Chem. 2022, 292, 133506. [CrossRef]
- Huang, S.; Jaffé, P.R. Defluorination of perfluorooctanoic acid (PFOA) and perfluorooctane sulfonate (PFOS) by Acidimicrobium sp. Strain A6. Environ. Sci. Technol. 2019, 53(19), 11410–11419. [CrossRef]
- Yu, Y.; Zhang, K.; Li, Z.; Ren, C.; Chen, J.; Lin, Y.-H.; Liu, J.; Men, Y. Microbial cleavage of C-F bonds in two C6 Per- and polyfluorinated compounds via reductive defluorination. Environ. Sci. Technol. 2020, 54(22), 14393–14402. [CrossRef]
- Chetverikov, S.P.; Sharipov, D.A.; Korshunova, T.Y.; Loginov, O.N. Degradation of perfluorooctanyl sulfonate by strain Pseudomonas plecoglossicida 2.4-D. Appl. Biochem. Microbiol. 2017, 53 (5), 533–538. [CrossRef]
- Zhang, Z.; Sarkar, D.; Biswas, J.K.; Datta, R. Biodegradation of per- and polyfluoroalkyl substances (PFAS): a review. Bioresour. Technol. 2022, 344, 126223. [CrossRef]
- Chetverikov, S.; Hkudaygulov, G.; Sharipov, D.; Starikov, S.; Chetverikova, D. Biodegradation Potential of C7-C10 Perfluorocarboxylic Acids and Data from the Genome of a New Strain of Pseudomonas mosselii 5(3). Toxics. 2023, 11(12), 1001. [CrossRef]
- Mothersole, R.G.; Mothersole, M.K, Goddard, H.G.; Liu, J.; Van Hamme, J.D. Enzyme Catalyzed Formation of CoA Adducts of Fluorinated Hexanoic Acid Analogues using a Long-Chain acyl-CoA Synthetase from Gordonia sp. Strain NB4-1Y. Biochemistry. 2024, 63(17), 2153–2165. [CrossRef]
- Kwon, B.G.; Lim, H-J.; Na, S-H.; Choi, B-I.; Shin, D-S.; Chung, S-Y. Biodegradation of perfluorooctanesulfonate (PFOS) as an emerging contaminant. Chemosphere. 2014, 109, 221–225. [CrossRef]
- Yi, L.; Tang, C.; Peng, Q.; Peng, Q.; Chai, L. Draft genome sequence of perfluorooctane acid-degrading bacterium Pseudomonas parafulva YAB-1. Genome Announc. 2015, 3, e00935–e00915. [CrossRef]
- Harris, J.; Gross, M.; Kemball, J.; Farajollahi, S.; Dennis, P.; Sitko, J.; Steel, J.J.; Almand, E.; Kelley-Loughnane, N.; Varaljay, V.A. Draft Genome Sequence of the Bacterium Delftia acidovorans Strain D4B, Isolated from Soil. Microbiol Resour Announc. 2021, 10(44), e0063521. [CrossRef]
- Hu, M.; Scott, C. Toward the development of a molecular toolkit for the microbial remediation of per-and polyfluoroalkyl substances. Appl Environ Microbiol. 2024, 90(4), e0015724. [CrossRef]
- Berhanu, A.; Mutanda, I.; Taolin, J.; Qaria, M.A.; Yang, B.; Zhu, D. A review of microbial degradation of per- and polyfluoroalkyl substances (PFAS): Biotransformation routes and enzymes. Sci Total Environ. 2023, 859(Pt 1), 160010. [CrossRef]
- Khusnutdinova, A.N.; Batyrova, K.A.; Brown, G.; Fedorchuk, T.; Chai, Y.S.; Skarina, T.; Flick, R.; Petit, A.P.; Savchenko, A.; Stogios, P.; Yakunin, A.F. Structural insights into hydrolytic defluorination of difluoroacetate by microbial fluoroacetate dehalogenases. FEBS J. 2023, 290(20), 4966–4983. [CrossRef]
- Farajollahi, S.; Lombardo, N.V.; Crenshaw, M.D.; Guo, H.B.; Doherty, M.E.; Davison, T.R.; Steel, J.J.; Almand, E.A.; Varaljay, V.A.; Suei-Hung, C.; Mirau, P.A.; Berry, R.J.; Kelley-Loughnane, N.; Dennis, P.B. Defluorination of Organofluorine Compounds Using Dehalogenase Enzymes from Delftia acidovorans (D4B). ACS Omega. 2024, 9(26), 28546–28555. [CrossRef]
- Harris, J.D.; Coon, C.M.; Doherty, M.E.; McHugh, E.A.; Warner, M.C.; Walters, C.L.; Orahood, O.M.; Loesch, A.E.; Hatfield, D.C.; Sitko, J.C.; Almand, E.A.; Steel, J.J. Engineering and characterization of dehalogenase enzymes from Delftia acidovorans in bioremediation of perfluorinated compounds. Synth Syst Biotechnol. 2022, 7(2), 671–676. [CrossRef]
- Jaffé, P.R.; Huang, S.; Park, J.; Ruiz-Urigüen, M.; Shuai, W.; Sima, M. Defluorination of PFAS by Acidimicrobium sp. strain A6 and potential applications for remediation. Methods Enzymol. 2024, 696, 287–320. [CrossRef]
- Raymond, R.L. Microbial oxidation of n-paraffinic hydrocarbons. Dev. Ind. Microbiol. 1961, 2, 23–54.
- Bertani, G. Studies on lysogenesis I. J. Bacteriol. 1951, 62, 293–300. [CrossRef]
- Sambrook, J.; Russell, D.W. Purification of nucleic acids by extraction with phenol:chloroform. Cold Spring Harb Protoc. 2006, 2006(1), pdb.prot4455. [CrossRef]
- Yang, X.; Liu, D.; Liu, F.; Wu, J.; Zou, J.; Xiao, X.; Zhao, F.; Zhu, B. HTQC: A fast quality control toolkit for Illumina sequencing data. BMC Bioinform. 2013, 14, 33. [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics. 2014, 30(15), 2114–20. [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.; Nikolenko, S.; Pham, S.; Prjibelski, A.; Pyshkin, A.; Sirotkin, A.; Vyahhi, N.; Tesler, G.; Alekseyev, M.A.; Pevzner, P.A. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012, 19, 455–477. [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; Earl, A.M. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE. 2014, 9, e112963. eCollection 2014. [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods. 2012, 9, 357–359. [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.M.; Kwon, S.J.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek. 2017, 110(10), 1281–1286. [CrossRef]
- Meier-Kolthoff, J.P.; Sardà Carbasse, J.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: a database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acid Res. 2022, 50(D1), D801–D807. [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics. 2014, 30, 2068–2069. [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015; 32: 2798–2800. DOI:.
- Farris, J.S. Estimating phylogenetic trees from distance matrices. Am. Nat. 1972, 6, 645–667.
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [CrossRef]
- Meier-Kolthoff, J.P.; Goker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [CrossRef]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V.; Fiebig, A.; Rohde, C.; Rohde, M.; Fartmann, B.; Goodwin, L.A.; Chertkov, O.; Reddy, T.; Pati, A.; Ivanova, N.N.; Markowitz, V.; Kyrpides, N.C.; Woyke, T.; Göker, M.; Klenk, H-P. Complete genome sequence of DSM 30083(T), the type strain (U5/41(T)) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genom. Sci. 2014, 9, 2. eCollection 2014. [CrossRef]
- Lee, I.; Ouk Kim, Y.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [CrossRef]
- Rankin, K.; Mabury, S.A.; Jenkins, T.M.; Washington, J.W. A North American and global survey of perfluoroalkyl substances in surface soils: Distribution patterns and mode of occurrence. Chemosphere. 2016, 161, 333-341. [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; Trujillo, M.E. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [CrossRef]
- Wackett, L.P. Pseudomonas: versatile biocatalysts for PFAS. Environ Microbiol. 2022, 24 (7), 2882-2889. [CrossRef]
- Calero, P.; Gurdo, N.; Nikel, P.I. Role of the CrcB transporter of Pseudomonas putida in the multi-level stress response elicited by mineral fluoride. Env Microbiol. 2022, 24(11), 5082–5104. [CrossRef]
- Haak, B.; Fetzner, S.; Lingens, F. Cloning, nucleotide sequence, and expression of the plasmid-encoded genes for the two-component 2-halobenzoate 1,2-dioxygenase from Pseudomonas cepacia 2CBS. J Bacteriol. 1995, 177(3), 667-75. [CrossRef]
- Eady, R.R. The vanadium-containing nitrogenase of Azotobacter. Biofactors. 1988, 1(2), 111-6.
- Llamas, A.; Leon-Miranda, E.; Tejada-Jimenez, M. Microalgal and Nitrogen-Fixing Bacterial Consortia: From Interaction to Biotechnological Potential. Plants. 2023, 12(13), 2476. [CrossRef]
- Beškoski, V.P., Yamamoto, A., Nakano, T., Yamamoto, K., Matsumura, C., Motegi, M., Beškoski, L.S., Inui, H. Defluorination of perfluoroalkyl acids is followed by production of monofluorinated fatty acids. Sci Total Environ. 2018, 636, 355-359. [CrossRef]
- Tang, Z., Vogel, T.M., Wang, Q., Wei, C., Ali, M., Song, X. Microbial defluorination of TFA, PFOA, and HFPO-DA by a native microbial consortium under anoxic conditions. J Hazard Mater. 2024, 465, 133217. [CrossRef]
- Starikov, S.N., Hkudaygulov, G.G., Chetverikov, S.P. Isolation of perfluorocarboxylic acid dehalogenases from Pseudomonas plecoglossicida 2,4-D and Pseudomonas mosselii 5(3). Èkobioteh. 2024, 7 (3), 204-210. [CrossRef]
- Tyagi, M., da Fonseca, M.M., de Carvalho, C.C. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation. 2011, 22(2), 231-41. [CrossRef]
- Olaniran, A.O., Pillay, D., Pillay, B. Biostimulation and bioaugmentation enhances aerobic biodegradation of dichloroethenes. Chemosphere. 2006, 63(4), 600-8. [CrossRef]
- Willmann, A., Trautmann, A.L., Kushmaro, A., Tiehm, A. Intrinsic and bioaugmented aerobic trichloroethene degradation at seven sites. Heliyon. 2023, 9(2), e13485. [CrossRef]
- Bolan, N., Sarkar, B., Yan, Y., Li, Q., Wijesekara, H., Kannan, K., Tsang, D.C.W., Schauerte, M., Bosch, J., Noll, H., Ok, Y.S., Scheckel, K., Kumpiene, J., Gobindlal, K., Kah, M., Sperry, J., Kirkham, M.B., Wang, H., Tsang. Y.F., Hou, D., Rinklebe, J. Remediation of poly- and perfluoroalkyl substances (PFAS) contaminated soils - To mobilize or to immobilize or to degrade? J Hazard Mater. 2021, 401, 123892. [CrossRef]
Figure 1.
Assembly metrics of the strain 2,4-D (A), whole-genome phylogenetic tree within the genus Pseudomonas (B), and the number of genes associated with common functional categories in its genome according to KEGG classification (C).
Figure 1.
Assembly metrics of the strain 2,4-D (A), whole-genome phylogenetic tree within the genus Pseudomonas (B), and the number of genes associated with common functional categories in its genome according to KEGG classification (C).
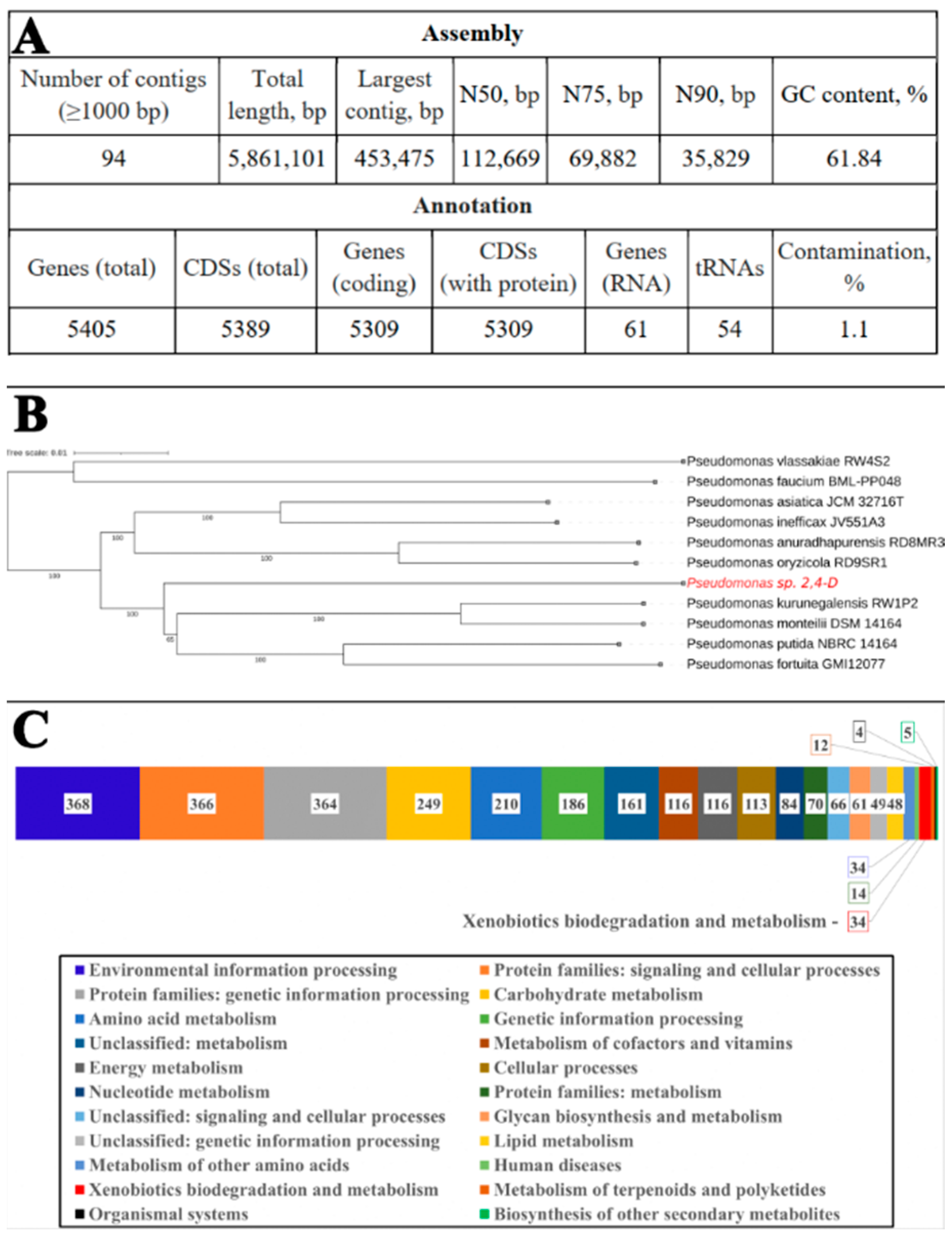
Figure 2.
Dynamics of fluoride ion release (x10 mg/L, continuous line), changes in PFCAs concentrations (mg/L, dashed line) during cultivation in liquid mineral medium ((A) with PFOA, (C) with PFNA) of Pseudomonas sp. 2,4-D strains (blue), P. mosselii 5(3) (black), their association (red), and depth of biodegradation potential based on component ratios (%) in the mixture on day 7 in variants (B) with PFOA, (D) with PFNA.
Figure 2.
Dynamics of fluoride ion release (x10 mg/L, continuous line), changes in PFCAs concentrations (mg/L, dashed line) during cultivation in liquid mineral medium ((A) with PFOA, (C) with PFNA) of Pseudomonas sp. 2,4-D strains (blue), P. mosselii 5(3) (black), their association (red), and depth of biodegradation potential based on component ratios (%) in the mixture on day 7 in variants (B) with PFOA, (D) with PFNA.
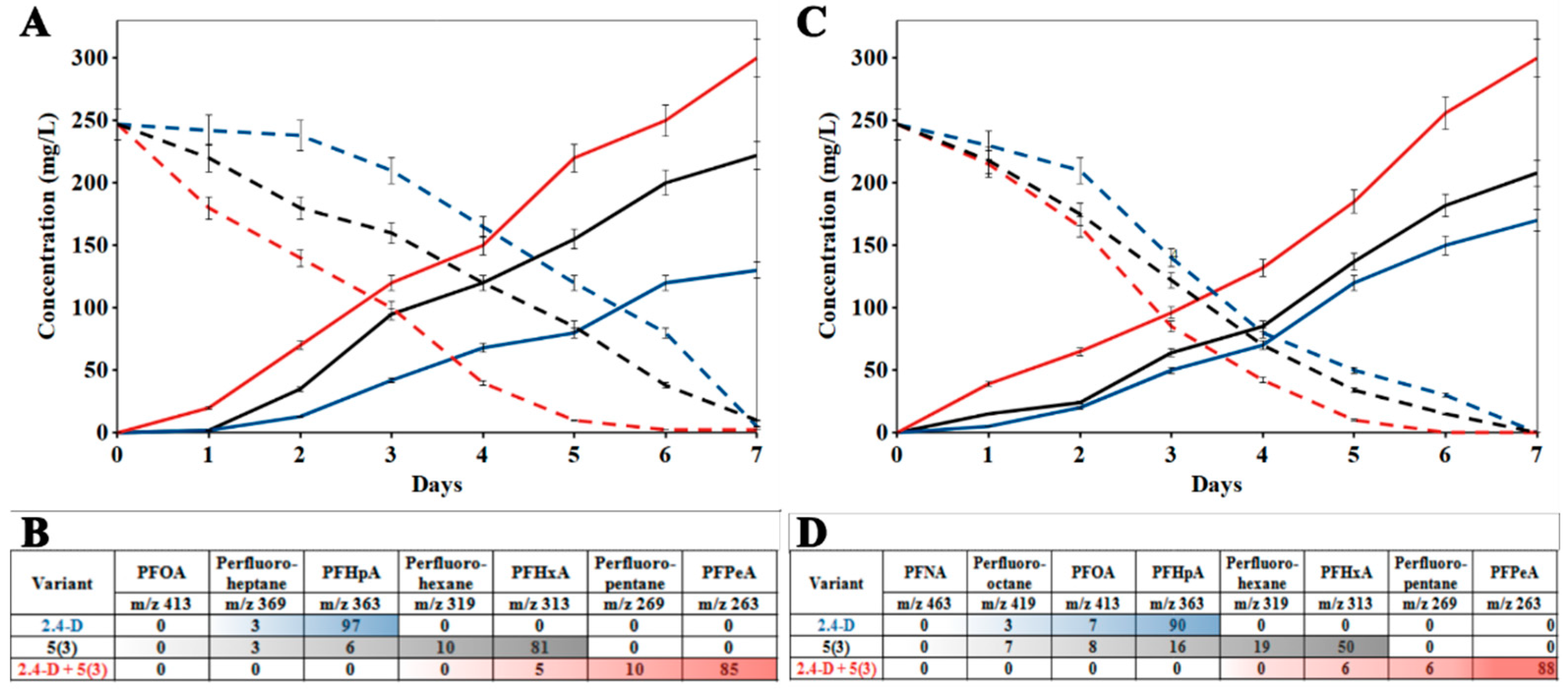
Figure 3.
Temporal characteristics and chromatograms of PFCA separation on a chromatograph-mass spectrometer (A) and detection of released fluoride ions by ion chromatography (B).
Figure 3.
Temporal characteristics and chromatograms of PFCA separation on a chromatograph-mass spectrometer (A) and detection of released fluoride ions by ion chromatography (B).
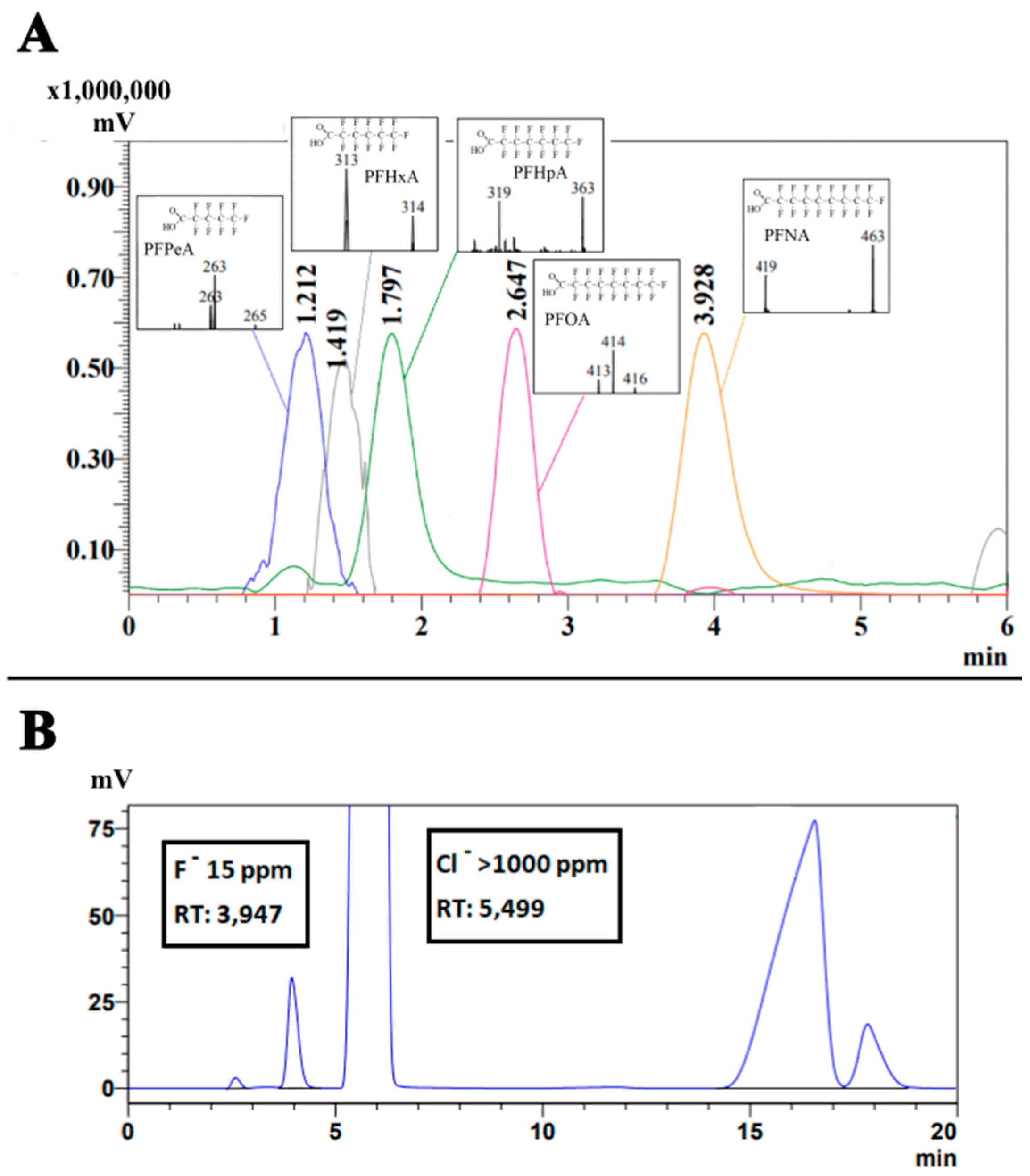
Figure 4.
Dynamics of changes during the augmentation of Pseudomonas sp. 2,4-D (blue), P. mosselii 5(3) (black), and their association (red) in the concentration of PFCAs (mg/kg), the number of destructive microorganisms (CFU/g), and the percentage ratio of PFCAs to their degradation products at the end of the model experiment in soil, respectively: A, B, C - with PFOA, D, E, F - with PFNA.
Figure 4.
Dynamics of changes during the augmentation of Pseudomonas sp. 2,4-D (blue), P. mosselii 5(3) (black), and their association (red) in the concentration of PFCAs (mg/kg), the number of destructive microorganisms (CFU/g), and the percentage ratio of PFCAs to their degradation products at the end of the model experiment in soil, respectively: A, B, C - with PFOA, D, E, F - with PFNA.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



