Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
The Locus Coeruleus (LC) is a critical area in the brain that plays an important role in several neural pathways associated with a range of physiological and behavioral processes. Its activity can modulate both norepinephrine (NE) and dopamine neurotransmission, particularly in the prefrontal cortex (PFC). In the present study, we show that a chemogenetically induced increase in norepinephrine release from the LC to the PFC reduced hyperactive behavioral patterns in rats lacking the dopamine transporter (DAT-KO rats) with spontaneously elevated dopamine transmission. These manipulations in hyperdopaminergic mutants also caused amelioration of cognitive abnormalities in spatial learning task as evidenced by decreases in perseverative activity and the number of visits to the error zone. Furthermore, chemogenetic activation of NE neurotransmission in these animals significantly improved their performance in in this maze test. Thus, the results obtained of this study highlight an important modulatory role of NE on hyperactivity and cognitive dysfunctions of hyperdopaminergic DAT-KO rats lacking the dopamine transporter.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Animals
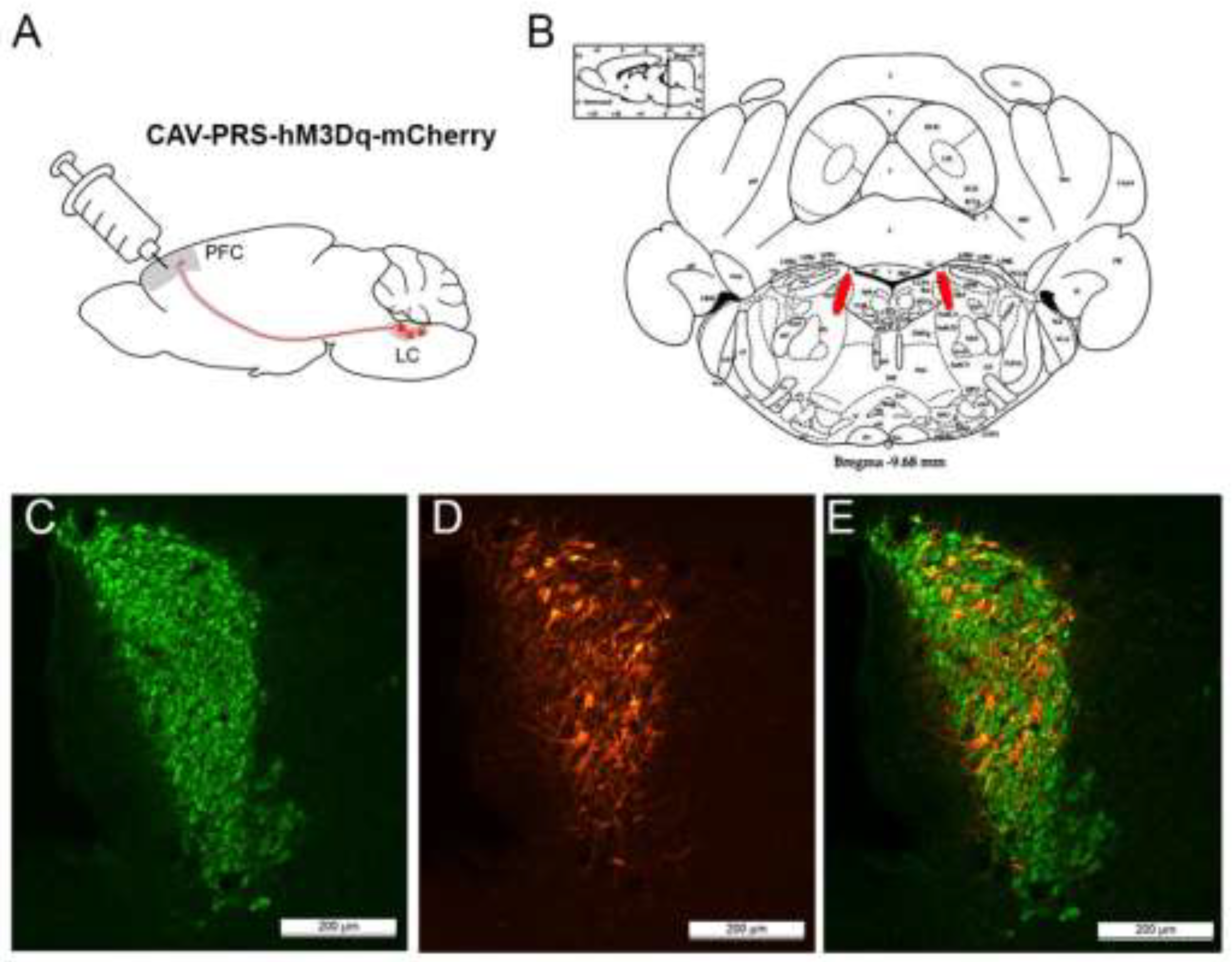
2.2. Viral Vector Microinjection
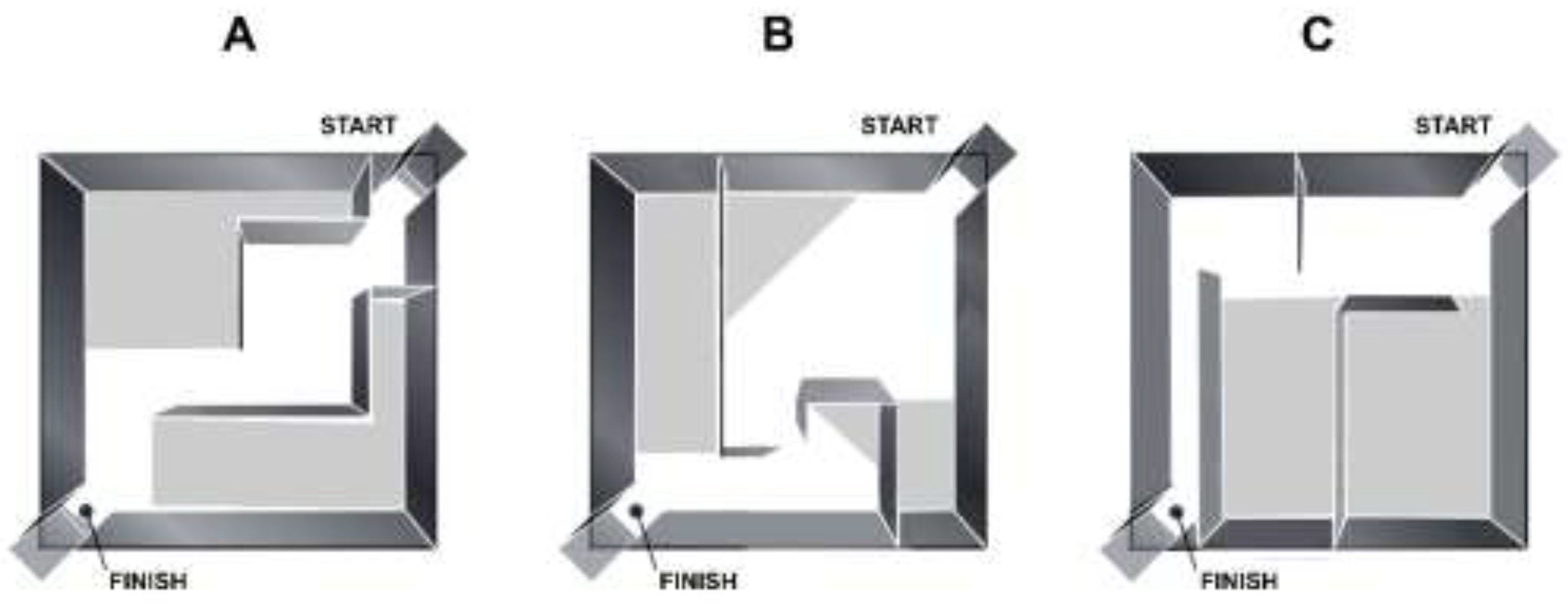
2.3. Hebb-Williams Maze Apparatus and Experimental Setup
2.4. Immunohistochemistry
2.5. Statistical Analysis
3. Results
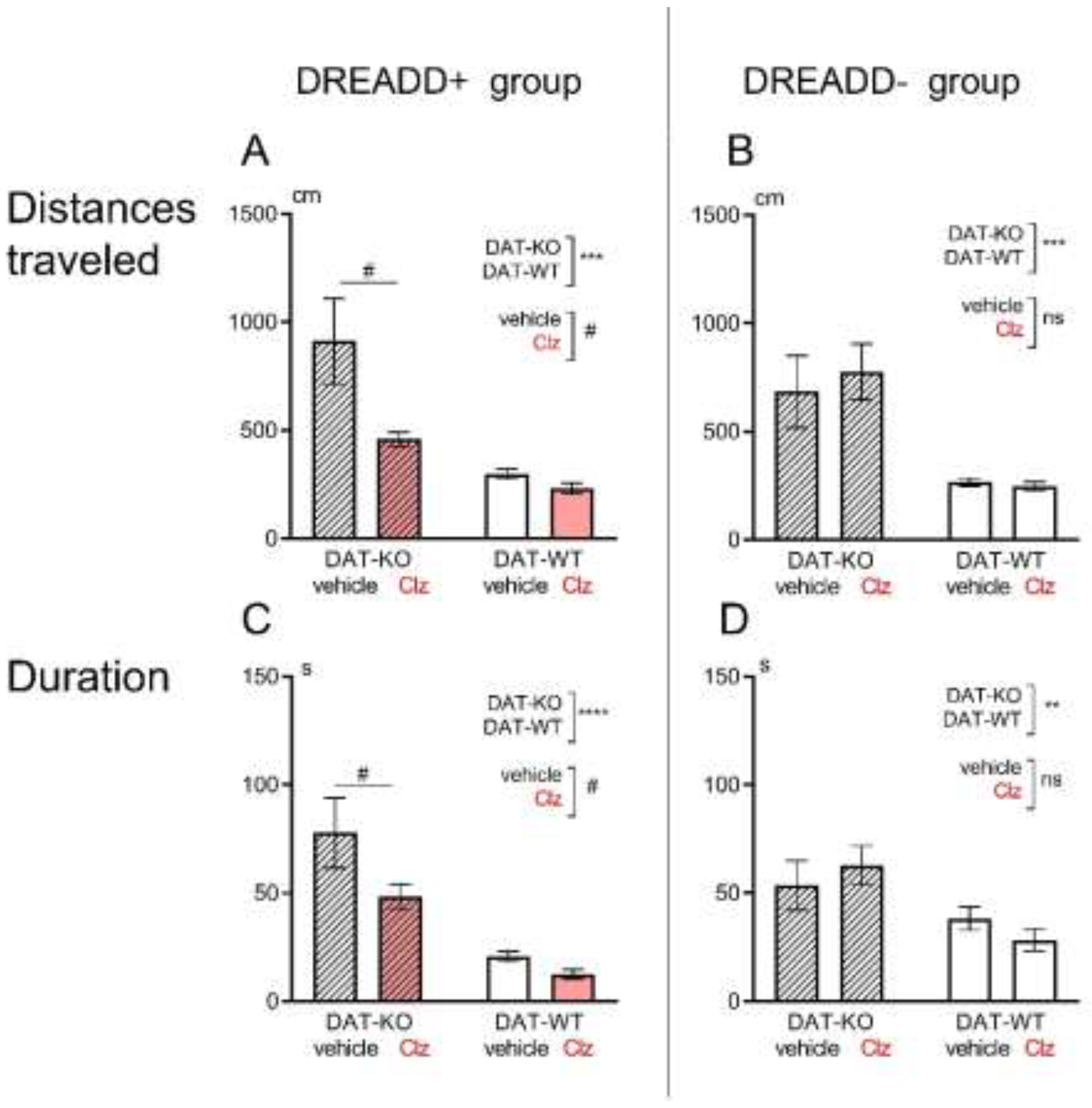
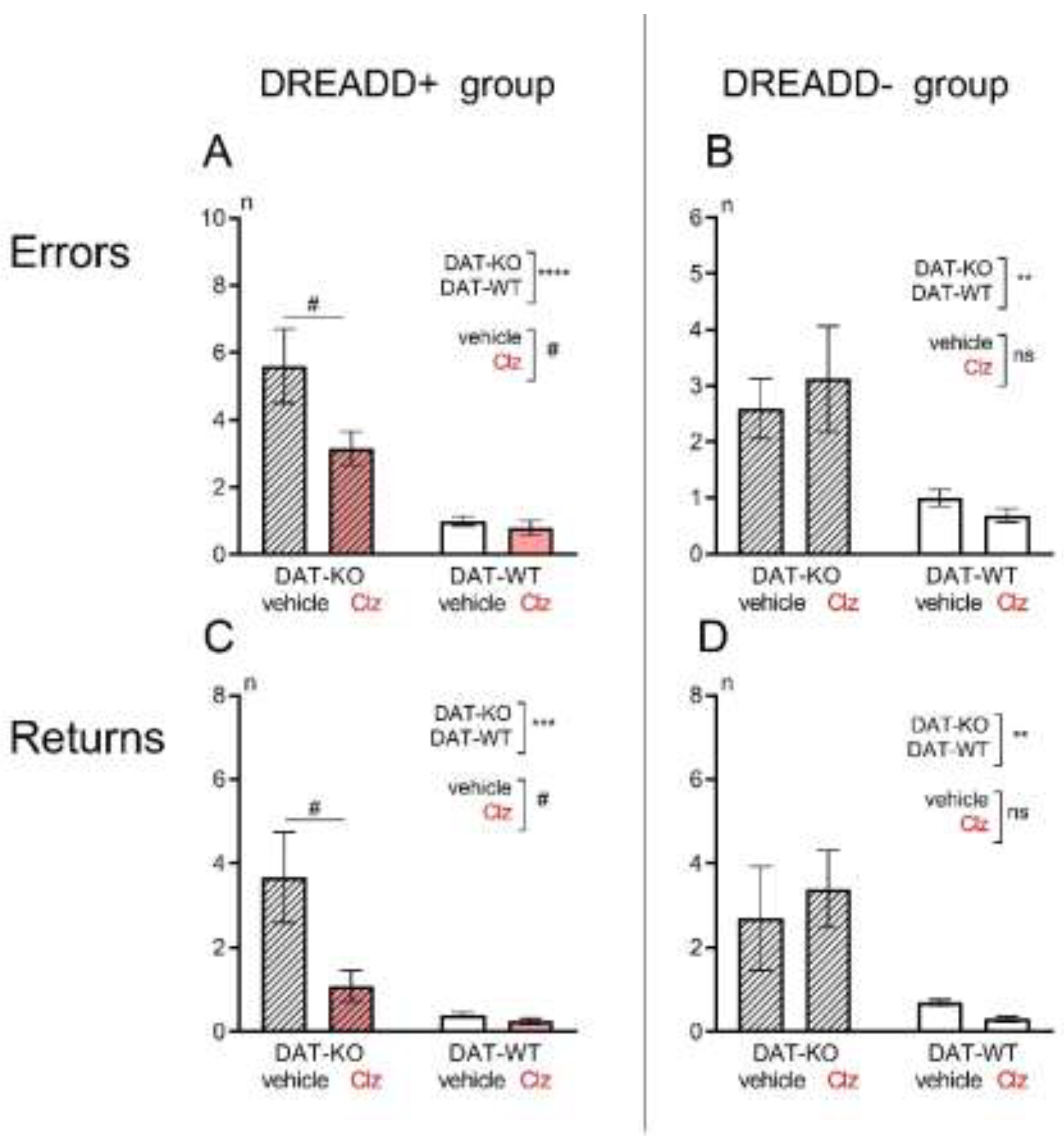
3.1. Hebb-Williams Maze
3.2. Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bari, B.A.; Chokshi, V.; Schmidt, K. Locus Coeruleus-Norepinephrine: Basic Functions and Insights into Parkinson’s Disease. Neural Regen. Res. 2020, 15, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Poe, G.R.; Foote, S.; Eschenko, O.; Johansen, J.P.; Bouret, S.; Aston-Jones, G.; Harley, C.W.; Manahan-Vaughan, D.; Weinshenker, D.; Valentino, R.; et al. Locus Coeruleus: A New Look at the Blue Spot. Nat. Rev. Neurosci. 2020 2111 2020, 21, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.J. The Locus Coeruleus and Noradrenergic Modulation of Cognition. Nat. Rev. Neurosci. 2009 103 2009, 10, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Radley, J.J.; Williams, B.; Sawchenko, P.E. Noradrenergic Innervation of the Dorsal Medial Prefrontal Cortex Modulates Hypothalamo-Pituitary-Adrenal Responses to Acute Emotional Stress. J. Neurosci. 2008, 28, 5806. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.; Sara, S.J. Network Reset: A Simplified Overarching Theory of Locus Coeruleus Noradrenaline Function. Trends Neurosci. 2005, 28, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Berridge, C.W.; Waterhouse, B.D. The Locus Coeruleus–Noradrenergic System: Modulation of Behavioral State and State-Dependent Cognitive Processes. Brain Res. Rev. 2003, 42, 33–84. [Google Scholar] [CrossRef]
- Bouras, N.N.; Mack, N.R.; Gao, W.J. Prefrontal Modulation of Anxiety through a Lens of Noradrenergic Signaling. Front. Syst. Neurosci. 2023, 17, 1173326. [Google Scholar] [CrossRef]
- Agster, K.L.; Mejias-Aponte, C.A.; Clark, B.D.; Waterhouse, B.D. Evidence for a Regional Specificity in the Density and Distribution of Noradrenergic Varicosities in Rat Cortex. J. Comp. Neurol. 2013, 521, 2195–2207. [Google Scholar] [CrossRef]
- Ranjbar-Slamloo, Y.; Fazlali, Z. Dopamine and Noradrenaline in the Brain; Overlapping or Dissociate Functions? Front. Mol. Neurosci. 2020, 12, 504957. [Google Scholar] [CrossRef]
- Russell, V.A. Overview of Animal Models of Attention Deficit Hyperactivity Disorder (ADHD). Curr. Protoc. Neurosci. 2011. [CrossRef]
- Tripp, G.; Wickens, J. Reinforcement, Dopamine and Rodent Models in Drug Development for ADHD. Neurotherapeutics 2012, 9, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Ko, I.G.; Kim, S.E.; Kim, T.W.; Ji, E.S.; Shin, M.S.; Kim, C.J.; Hong, M.H.; Bahn, G.H. Swimming Exercise Alleviates the Symptoms of Attention-Deficit Hyperactivity Disorder in Spontaneous Hypertensive Rats. Mol. Med. Rep. 2013, 8, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Natsheh, J.Y.; Shiflett, M.W. Dopaminergic Modulation of Goal-Directed Behavior in a Rodent Model of Attention-Deficit/Hyperactivity Disorder. Front. Integr. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Ng, J.; Zhen, J.; Meyer, E.; Erreger, K.; Li, Y.; Kakar, N.; Ahmad, J.; Thiele, H.; Kubisch, C.; Rider, N.L.; et al. Dopamine Transporter Deficiency Syndrome: Phenotypic Spectrum from Infancy to Adulthood. Brain 2014, 137, 1107–1119. [Google Scholar] [CrossRef]
- Viggiano, D.; Vallone, D.; Sadile, A. Dysfunctions in Dopamine Systems and ADHD: Evidence from Animals and Modeling. Neural Plast. 2004, 11, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Chen, A.L.C.; Braverman, E.R.; Comings, D.E.; Chen, T.J.H.; Arcuri, V.; Blum, S.H.; Downs, B.W.; Waite, R.L.; Notaro, A.; et al. Attention-Deficit-Hyperactivity Disorder and Reward Deficiency Syndrome. Neuropsychiatr. Dis. Treat. 2008, 4, 893–917. [Google Scholar] [PubMed]
- Nikolaus, S.; Antke, C.; Beu, M.; Müller, H.W.; Nikolaus, S. Cortical GABA, Striatal Dopamine and Midbrain Serotonin as the Key Players in Compulsiveand Anxiety Disorders - Results from in Vivo Imaging Studies. Rev. Neurosci. 2010, 21, 119–140. [Google Scholar] [CrossRef]
- Del Campo, N.; Chamberlain, S.R.; Sahakian, B.J.; Robbins, T.W. The Roles of Dopamine and Noradrenaline in the Pathophysiology and Treatment of Attention-Deficit/ Hyperactivity Disorder. 2011. [CrossRef]
- Vengeliene, V.; Bespalov, A.; Robmanith, M.; Horschitz, S.; Berger, S.; Relo, A.L.; Noori, H.R.; Schneider, P.; Enkel, T.; Bartsch, D.; et al. Towards Trans-Diagnostic Mechanisms in Psychiatry: Neurobehavioral Profile of Rats with a Loss-of-Function Point Mutation in the Dopamine Transporter Gene. DMM Dis. Model. Mech. 2017, 10, 451–461. [Google Scholar] [CrossRef]
- Sukhanov, I.; Leo, D.; Tur, M.A.; Belozertseva, I. V.; Savchenko, A.; Gainetdinov, R.R. Dopamine Transporter Knockout Rats as the New Preclinical Model of Hyper- and Hypo-Dopaminergic Disorders. V.M. BEKHTEREV Rev. PSYCHIATRY Med. Psychol. 2019, 0, 84–85. [Google Scholar] [CrossRef]
- Savchenko, A.; Targa, G.; Fesenko, Z.; Leo, D.; Gainetdinov, R.R.; Sukhanov, I. Dopamine Transporter Deficient Rodents: Perspectives and Limitations for Neuroscience. Biomol. 2023, Vol. 13, Page 806 2023, 13, 806. [Google Scholar] [CrossRef]
- Leo, D.; Sukhanov, I.; Zoratto, F.; Illiano, P.; Caffino, L.; Sanna, F.; Messa, G.; Emanuele, M.; Esposito, A.; Dorofeikova, M.; et al. Pronounced Hyperactivity, Cognitive Dysfunctions, and BDNF Dysregulation in Dopamine Transporter Knock-out Rats. J. Neurosci. 2018, 38, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.; Aristova, I.; Volnova, A.; Gainetdinov, R.R. Deficit in Working Memory and Abnormal Behavioral Tactics in Dopamine Transporter Knockout Rats during Training in the 8-Arm Maze. Behav. Brain Res. 2020, 390. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.; Belskaya, A.; Gromova, A.; Ignashchenkova, A.; Gainetdinov, R.R.; Volnova, A. Modulation of Spatial Memory Deficit and Hyperactivity in Dopamine Transporter Knockout Rats via A2A-Adrenoceptors. Front. Psychiatry 2022, 13, 503. [Google Scholar] [CrossRef] [PubMed]
- Belskaya, A.; Kurzina, N.; Savchenko, A.; Sukhanov, I.; Gromova, A.; Gainetdinov, R.R.; Volnova, A. Rats Lacking the Dopamine Transporter Display Inflexibility in Innate and Learned Behavior. Biomed. 2024, Vol. 12, Page 1270 2024, 12, 1270. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.S.; Bucci, D.J.; Luikart, B.W.; Mahler, S. V. Dreadds: Use and Application in Behavioral Neuroscience. Behav. Neurosci. 2021, 135, 89–107. [Google Scholar] [CrossRef]
- Roth, B.L. DREADDs for Neuroscientists. Neuron 2016, 89, 683–694. [Google Scholar] [CrossRef]
- Campbell, E.J.; Marchant, N.J. The Use of Chemogenetics in Behavioural Neuroscience: Receptor Variants, Targeting Approaches and Caveats. Br. J. Pharmacol. 2018, 175, 994–1003. [Google Scholar] [CrossRef]
- Kabanova, A.; Fedorov, L.; Eschenko, O. The Projection-Specific Noradrenergic Modulation of Perseverative Spatial Behavior in Adult Male Rats. eNeuro 2024, 11. [Google Scholar] [CrossRef]
- Kabanova, A.; Yang, M.; Logothetis, N.K.; Eschenko, O. Partial Chemogenetic Inhibition of the Locus Coeruleus Due to Heterogeneous Transduction of Noradrenergic Neurons Preserved Auditory Salience Processing in Wild-Type Rats. Eur. J. Neurosci. 2024. [CrossRef]
- Pritchett, K.; Mulder, G.B. Operant Conditioning. J. Am. Assoc. Lab. Anim. Sci. 2004, 43, 35–36. [Google Scholar]
- Falcy, B.A.; Mohr, M.A.; Micevych, P.E. Immunohistochemical Amplification of MCherry Fusion Protein Is Necessary for Proper Visualization. MethodsX 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012 97 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, A.; Zelli, S.; Leo, D.; Carbone, C.; Mus, L.; Illiano, P.; Alleva, E.; Gainetdinov, R.R.; Adriani, W. Behavioral Characterization of DAT-KO Rats and Evidence of Asocial-like Phenotypes in DAT-HET Rats: The Potential Involvement of Norepinephrine System. Behav. Brain Res. 2019, 359, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.; Volnova, A.; Aristova, I.; Gainetdinov, R.R. A New Paradigm for Training Hyperactive Dopamine Transporter Knockout Rats: Influence of Novel Stimuli on Object Recognition. Front. Behav. Neurosci. 2021, 15, 654469. [Google Scholar] [CrossRef]
- Volnova, A.; Kurzina, N.; Belskaya, A.; Gromova, A.; Pelevin, A.; Ptukha, M.; Fesenko, Z.; Ignashchenkova, A.; Gainetdinov, R.R. Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine. Biomedicines 2023, 11, 222. [Google Scholar] [CrossRef]
- Savchenko, A.; Müller, C.; Lubec, J.; Leo, D.; Korz, V.; Afjehi-Sadat, L.; Malikovic, J.; Sialana, F.J.; Lubec, G.; Sukhanov, I. The Lack of Dopamine Transporter Is Associated With Conditional Associative Learning Impairments and Striatal Proteomic Changes. Front. Psychiatry 2022, 13, 269. [Google Scholar] [CrossRef]
- Benarroch, E.E. Locus Coeruleus. Cell Tissue Res. 2017 3731 2017, 373, 221–232. [Google Scholar] [CrossRef]
- Breton-Provencher, V.; Drummond, G.T.; Sur, M. Locus Coeruleus Norepinephrine in Learned Behavior: Anatomical Modularity and Spatiotemporal Integration in Targets. Front. Neural Circuits 2021, 15, 638007. [Google Scholar] [CrossRef]
- Carter, M.E.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; De Lecea, L. Tuning Arousal with Optogenetic Modulation of Locus Coeruleus Neurons. Nat. Neurosci. 2010 1312 2010, 13, 1526–1533. [Google Scholar] [CrossRef]
- Naganuma, F.; Nakamura, T.; Kuroyanagi, H.; Tanaka, M.; Yoshikawa, T.; Yanai, K.; Okamura, N. Chemogenetic Modulation of Histaminergic Neurons in the Tuberomamillary Nucleus Alters Territorial Aggression and Wakefulness. Sci. Reports 2021 111 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Uematsu, A.; Tan, B.Z.; Ycu, E.A.; Cuevas, J.S.; Koivumaa, J.; Junyent, F.; Kremer, E.J.; Witten, I.B.; Deisseroth, K.; Johansen, J.P. Modular Organization of the Brainstem Noradrenaline System Coordinates Opposing Learning States. Nat. Neurosci. 2017 2011 2017, 20, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, S.; Li, Y.; Randall, A.; Kremer, E.J.; Pickering, A.E. Functional Dichotomy in Spinal-vs Prefrontal-Projecting Locus Coeruleus Modules Splits Descending Noradrenergic Analgesia from Ascending Aversion and Anxiety in Rats. Elife 2017, 6. [Google Scholar] [CrossRef]
- McCall, J.G.; Siuda, E.R.; Bhatti, D.L.; Lawson, L.A.; McElligott, Z.A.; Stuber, G.D.; Bruchas, M.R. Locus Coeruleus to Basolateral Amygdala Noradrenergic Projections Promote Anxiety-like Behavior. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Robbins, T.W.; Arnsten, A.F.T. The Neuropsychopharmacology of Fronto-Executive Function: Monoaminergic Modulation. Annu. Rev. Neurosci. 2009, 32, 267–287. [Google Scholar] [CrossRef]
- Howes, O.D.; McCutcheon, R.; Owen, M.J.; Murray, R.M. The Role of Genes, Stress, and Dopamine in the Development of Schizophrenia. Biol. Psychiatry 2017, 81, 9–20. [Google Scholar] [CrossRef]
- Winograd-Gurvich, C.; Fitzgerald, P.B.; Georgiou-Karistianis, N.; Bradshaw, J.L.; White, O.B. Negative Symptoms: A Review of Schizophrenia, Melancholic Depression and Parkinson’s Disease. Brain Res. Bull. 2006, 70, 312–321. [Google Scholar] [CrossRef]
- Nutt, D.J. Relationship of Neurotransmitters to the Symptoms of Major Depressive Disorder. J. Clin. Psychiatry 2008, 69, 17337. [Google Scholar]
- Kessi, M.; Duan, H.; Xiong, J.; Chen, B.; He, F.; Yang, L.; Ma, Y.; Bamgbade, O.A.; Peng, J.; Yin, F. Attention-Deficit/Hyperactive Disorder Updates. Front. Mol. Neurosci. 2022, 15, 925049. [Google Scholar] [CrossRef]
- Koirala, S.; Grimsrud, G.; Mooney, M.A.; Larsen, B.; Feczko, E.; Elison, J.T.; Nelson, S.M.; Nigg, J.T.; Tervo-Clemmens, B.; Fair, D.A. Neurobiology of Attention-Deficit Hyperactivity Disorder: Historical Challenges and Emerging Frontiers. Nat. Rev. Neurosci. 2024 2512 2024, 25, 759–775. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Wetsel, W.C.; Jones, S.R.; Levin, E.D.; Jaber, M.; Caron, M.G. Role of Serotonin in the Paradoxical Calming Effect of Psychostimulants on Hyperactivity. Science (80-. ). 1999, 283, 397–401. [Google Scholar] [CrossRef]
- Cinque, S.; Zoratto, F.; Poleggi, A.; Leo, D.; Cerniglia, L.; Cimino, S.; Tambelli, R.; Alleva, E.; Gainetdinov, R.R.; Laviola, G.; et al. Behavioral Phenotyping of Dopamine Transporter Knockout Rats: Compulsive Traits, Motor Stereotypies, and Anhedonia. Front. Psychiatry 2018, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Pogorelov, V.M.; Rodriguiz, R.M.; Insco, M.L.; Caron, M.G.; Wetsel, W.C. Novelty Seeking and Stereotypic Activation of Behavior in Mice with Disruption of the Dat1 Gene. Neuropsychopharmacology 2005, 30, 1818–1831. [Google Scholar] [CrossRef] [PubMed]
- Arnsten, A.F.T. Guanfacine’s Mechanism of Action in Treating Prefrontal Cortical Disorders: Successful Translation across Species. Neurobiol. Learn. Mem. 2020, 176, 107327. [Google Scholar] [CrossRef] [PubMed]
- Ota, T.; Yamamuro, K.; Okazaki, K.; Kishimoto, T. Evaluating Guanfacine Hydrochloride in the Treatment of Attention Deficit Hyperactivity Disorder (Adhd) in Adult Patients: Design, Development and Place in Therapy. Drug Des. Devel. Ther. 2021, 15, 1965–1969. [Google Scholar] [CrossRef]
- Zerbi, V.; Floriou-Servou, A.; Markicevic, M.; Vermeiren, Y.; Sturman, O.; Privitera, M.; von Ziegler, L.; Ferrari, K.D.; Weber, B.; De Deyn, P.P.; et al. Rapid Reconfiguration of the Functional Connectome after Chemogenetic Locus Coeruleus Activation. Neuron 2019, 103, 702–718.e5. [Google Scholar] [CrossRef]
- Rossetti, Z.L.; Carboni, S. Noradrenaline and Dopamine Elevations in the Rat Prefrontal Cortex in Spatial Working Memory. J. Neurosci. 2005, 25, 2322–2329. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



