
Submitted:
25 November 2024
Posted:
26 November 2024
You are already at the latest version
Abstract
The BBB is created by a special system of brain microvascular endothelial cells (BMECs), pericytes (PCs), the capillary basement membrane and the terminal branches (“end-feet”) of astrocytes. The key function of the BBB is to protect the central nervous system (CNS) from potentially harmful/toxic substances in the bloodstream by selectively controlling the entry of cells and molecules, including nutrients and components of the immune system. The loss of BBB integrity in response to neuroinflammation, as manifested by an increase in permeability (leakage), occurs with the dominant participation of proinflammatory cytokines. However, the pathomechanism of structural and functional changes in the BBB under the influence of individual cytokines is still poorly understood. This review summarizes the current state of knowledge on this topic, which is important from both pathophysiological and clinical–therapeutic points of view. The structures and functions of all components of the BBB are discussed, with particular attention given to the phenotypic differences between BMECs and the vascular endothelium in other locations of the circulatory system. The protein composition of the interendothelial tight junctions in the context of regulating BBB permeability is presented, as is the role of pericyte–BMEC interactions in the exchange of metabolites, ions and nucleic acids. Finally, the documented actions of proinflammatory cytokines within the BBB are summarized.
Keywords:
blood-brain barrier structure
; blood-brain barrier integrity
; proinflammatory cytokines
; neuroinflammation
; brain endothelial cells
; tight junctions
; pericyte-endothelial interaction
; astrocytes
; neurodegenerative processes
1. Introduction
The blood‒brain barrier (BBB), which is a specialized internal barrier, plays an important role in ensuring the normal functioning of the central nervous system (CNS), which is responsible for the vital functions of the body as a whole [1]. The BBB is a complex structure consisting of endothelial cells, pericytes, and astrocytes located between the brain parenchyma and the vascular system and is highly connected with surrounding neurons and microglia [2,3]. A loss of isolation between blood and nervous tissue can lead to damage to the CNS and changes in the composition of the cerebrospinal fluid (CSF). BBB disturbances commonly occur in response to neuronal dysfunction, neuroinflammation, neurodegeneration, and substance use disorders [4,5]. A precise understanding of the structure of the BBB and the pathomechanism by which proinflammatory cytokines influence the disruption (loss of integrity) of the BBB may contribute to improvements in existing preventive and therapeutic methods [6,7].
This study was challenging because existing research data regarding specific mechanisms that damage the BBB are far from sufficient. The majority of studies consider the disruption of this barrier structure in the context of the development of certain diseases (diabetes mellitus, multiple sclerosis, Alzheimer's disease, neuropsychiatric systemic lupus erythematosus, herpes simplex encephalitis, and cerebrovascular ischemia) or various pathological conditions and processes (stress, traumatic brain injury, subarachnoid hemorrhage, and sepsis) [8,9,10].
Watroba et al. [11] studied the impact of dysglycemia associated with diabetes mellitus on the BBB structure. According to these researchers, damage to the BBB structure caused by impaired blood sugar stability, especially when it exceeds the normal level (a hyperglycemic state), leads to damage to CNS cells, provoking neurodegeneration and dementia. In their review, the authors cited studies showing that hyperglycemia increases the expression of proinflammatory cytokines such as tumor necrosis factor alpha (TNF-α) and interleukins (IL-1, IL-4 and IL-6), but the detailed mechanism underlying their influence on the integrity and permeability of the BBB was not described [12].
Lesniak et al. [13] investigated how the loss of brain-derived neurotrophic factor due to blood‒brain barrier disruption caused by congenital or mild traumatic brain injury correlates with depression-like behavior. This study was conducted in a mouse model of BBB disruption due to congenital traumatic brain injury. Although the mechanism of damage to the barrier structure was not described in detail in this study, the behavioral changes caused by damage to the barrier structure were studied and the method used (the study of the BBB in a mouse model) can also be used to investigate the effects of cytokines on BBB disruption in vivo.
Małkiewicz et al. [14] studied the impact of substance abuse on the BBB. In reviewing scientific papers on the topic, the authors obtained reliable data on the disruption and increase in the permeability of the BBB owing to the activation of certain mechanisms caused by the use of psychoactive substances, while also citing the results of studies confirming the protective potential of other substances in maintaining and preserving BBB integrity.
A determination of the level of proteins associated with the BBB in patients with epilepsy was performed by Bronisz et al. [15]. The researchers assessed the serum levels of molecules associated with changes in BBB structure: matrix metalloproteinases 2 and 9 (MMP-2 and MMP-9), tissue inhibitors of metalloproteinases 1 and 2 (TIMP-1 and TIMP-2), the S100 calcium-binding protein beta (S100B), the chemokine CCL-2, intercellular adhesion molecule-1 (ICAM-1), P-selectin, and thrombospondin-2 (TSP-2). As a result, chronic processes of disruption and restoration of the BBB structure are accompanied by increased serum levels of MMP-9, MMP-2, TIMP-1, TIMP-2 and S100B, indicating their direct relationship with barrier permeability; however, this study did not provide a detailed description of the mechanism underlying this relationship.
Łach et al. [16] conducted a study on methods to create an experimental model to study the functions of the BBB. The discussion of methods for model development was based on data on the structure and functions of the BBB, as well as on the identification of parameters important for maintaining the barrier phenotype, which would allow an assessment of its state in vitro. The benefit of this study is that the implementation of theoretical developments of scientists allows an increase in the level of development of neuroscience and provides opportunities for studying the mechanisms underlying the effects of certain factors on the disruption of the BBB structure.
While systemic inflammation is a response to pathological changes associated with the development of diseases, some of which are the cause and some of which are the consequence of BBB disruption, pro-inflammatory cytokines may be directly related to damage to the BBB structure [17,18,19]. Therefore, this review aims to summarize the current state of knowledge concerning the effects of proinflammatory cytokines on the disruption of the integrity and permeability of the BBB based on the data on its structural features, which are important from both pathophysiological and clinical–therapeutic points of view. This approach has rational justification from both pathophysiological and clinical–therapeutic points of view [20,21].
2. Methodology of the Literature Search
The study of the structure of the BBB and the effects of proinflammatory cytokines on its disruption was performed via an analysis and synthesis of the research topic and generalization of the information obtained. In this manner, a harmonious formula of data presentation was built, which is helpful in the qualitative analysis of the research subject and thus in achieving the research goal.
The method of analyzing the subject of the study was applied to the peculiarities of the BBB structure and its functions, as this structure allows us to identify pathways and factors leading to BBB disorders, pathways by which proinflammatory cytokines pass through the BBB without disrupting its structure, and the effects of proinflammatory cytokines on BBB disorders. The materials for the analysis were selected from the most cited original scientific papers in the fields of anatomy, neurology, immunology and microbiology published in the electronic databases PubMed, Google Scholar and Scopus. The search for materials that included information on the structure and functions of the BBB was performed using the following keywords: “blood–brain barrier structure”, “capillary endothelium”, “basement membrane”, “astrocytes”, and “blood–brain barrier functions”. The search resulted in 33 sources that were relevant to the topic. Scientific papers that included information about the factors involved in BBB disorders and how this process is triggered were selected by queries containing the following keywords: “mechanisms of blood‒brain barrier damage”, “factors influencing blood‒brain barrier disorders”, and “diseases causing blood‒brain barrier disorders”. Priority was given to papers that contained observational or experimental data. Six publications that met the query criteria were selected for analysis. Sources for the review of proinflammatory cytokines that can cross the BBB without damaging its structure were selected from the list of studies containing both theoretical developments and experimental components using the keywords “proinflammatory cytokines and the blood‒brain barrier”, “types of proinflammatory cytokines that cross the blood‒brain barrier”, and “ways of crossing the blood‒brain barrier by proinflammatory cytokines”. Four sources corresponded to the given topic. The search for scientific papers reporting the analysis of data on the role of proinflammatory cytokines in the disruption of BBB integrity was performed using the keywords “the role of the inflammatory process in blood–brain barrier integrity disruption”, “negative impact of proinflammatory cytokines on the blood–brain barrier”, and “impact of proinflammatory cytokines on blood–brain barrier structure disruption”. Following the search, 32 sources were selected that contained theoretical information and the results of practical developments on this topic. A total of 75 scientific articles published between 1994 and 2023 were selected for the analysis. The data obtained during the analysis were synthesized to elucidate the mechanism by which proinflammatory cytokines affect the disruption of the BBB structure.
The synthesized information was summarized to determine the positive and negative effects of proinflammatory cytokines on the BBB structure and to identify methods that eliminate the damage caused by these effects, as well as areas that could use the principle of overcoming the barrier to solve certain specific problems.
3. Normal BBB Structure
The BBB is structurally designed to provide its main functions—protective and regulatory [22]. Any damage to the BBB can negatively affect the neurological system, as a loss of its integrity disrupts the isolation of blood from nervous tissue, abolishing the boundary between the central nervous system and dangerous substances in the blood and leading to changes in the stability of the cerebrospinal fluid [23,24]. The structure of the BBB must be determined in detail to understand how and what can disrupt or damage the BBB.
The BBB is not located in the entire CNS; it is absent in those parts of the CNS that function to analyze the chemical composition of the blood, regulate its main parameters, and detect the presence of toxins. These unprotected areas of the brain regulate the autonomic nervous system and endocrine glands. Due to their locations around the third and fourth ventricles, these structures are called circumventricular organs (CVOs). CVOs include the hypothalamus and other anatomically related structures, such as the pineal gland, median eminence, subcommisural organ, neurohypophysis, subfornical organ and organum vasculosum of the lamina terminalis, and vomiting center (anatomically corresponding to the area postrema) in the medulla oblongata [25,26].
The main elements of the BBB structure are the capillary endothelium, capillary basement membrane and perivascular membrane formed by glial cell processes (bulbous “legs” of astrocytes). The capillary endothelium, together with pericytes, is located on the basement membrane, on the other side of which are glial cell processes [1,3,20]. Multiprotein complexes between epithelial cells, known as tight junctions or zonulae occludentes, create a seal and prevent the leakage of solutes and water at the junction of endothelial cells [3,22].
The normal (undisturbed) structure of the BBB is composed of "layers" of biostructures, each of which acts as a level of protection for isolation of the blood and nervous tissue. At the first level, protection is provided by the capillary endothelium; at the second level, protection is provided by the basement membrane; and at the third level, protection is provided by the perivascular membrane formed from astrocyte processes [27]. The capillary endothelium restricts the access of bacteria and other microscopic objects and large hydrophilic molecules while ensuring the diffusion of small hydrophilic molecules and the transport of metabolic products. The tasks of the basement membrane are to preserve the structure of the barrier and ensure the exchange of signals between its components. Due to the contact between astrocyte processes and capillaries in the brain, the perivascular membrane maintains the normal BBB structure [28,29,30] (Figure 1).
3.1. Endothelial Cells
The capillary endothelium consists of firmly and tightly connected endothelial cells that are linked by interendothelial tight junctions. Phenotypically, the endothelial cells of the BBB differ from the endothelial cells localized in other parts of the body; they have a more flattened appearance, a greater number of mitochondria and a significantly smaller number of caveolae [32,33]. These features and the principle of communication between cells in the capillary endothelium affect the level of capillary permeability. Thus, it hinders the paracellular transport of hydrophilic molecules to nervous tissue and isolates the capillary lumen from the basolateral complex; however, it allows the passage of small lipophilic molecules through facilitated diffusion, as well as the transport of nutrients [glucose through the glucose transporter 1 (GLUT1) channel, essential amino acids, certain electrolytes, etc.], oxygen, carbon dioxide, lipids, anesthetics, ethanol, and nicotine. Through the mechanism of receptor-mediated endocytosis, it allows insulin, transferrin and leptin to cross the BBB [34,35,36].
In the lumen of the capillaries, the endothelium cells are covered with glycocalyx, which plays an essential role in maintaining homeostasis within the BBB. The endothelial glycocalyx extends the neurovascular unit, which influences the structure of the BBB. Specifically, the glycocalyx acts as a physical and ionic barrier; regulates mechanical transduction, vascular permeability, and the inflammatory response; and inhibits coagulation [37].
3.2. Tight Junctions
Tight contacts, which ensure the strength of the connections between endothelial cells, are formed from the main membrane proteins [claudin, occludin, junctional adhesion molecules (JAMs), and cadherins] and several additional cytoplasmic proteins (zonula occludens-1, -2, -3; cingulin; α-catenin; β-catenin; γ-catenin; vinculin and others). The mechanism of tight junctions involves auxiliary cytoplasmic proteins binding the main membrane proteins to the main cytoskeleton protein (actin), which allows the integrity and the required level of permeability of the capillary endothelium to be maintained [38,39] (Figure 2).
Claudin proteins form a strong seal, which includes both bonds with other claudin proteins between the endothelial cells on which they are located and with zonula occludens proteins (ZO-1, ZO-2, and ZO-3). The strength of the bonds provided by this protein affects the regulation of BBB permeability during the development of inflammation, exposure to toxic substances, trauma, and other pathological processes. Thus, damage to claudin can adversely affect the permeability of the barrier, allowing the passage of monocytes in some inflammatory diseases [40,41]. The loss of this protein occurs during brain tumor development, stroke, and complex central nervous system inflammatory processes [42,43,44,45].
The occludin protein differs from claudin in its larger size (molecular weight of 65 kDa, compared with 20-27 kDa for claudin) and amino acid sequence (522 amino acids, compared with 102–150 amino acids for claudin, depending on the transmembrane domains). Together with claudin, occludin forms intramembrane filaments that promote the diffusion of hydrophilic molecules and ions and regulate paracellular permeability [46,47,48].
Junctional adhesion molecules (JAMs) are immunoglobulin proteins with a molecular weight of 40 kDa. JAMs increase the adhesion density by binding to actin via ZO-1, but data on their role in monocyte migration across the BBB have not been conclusively confirmed, and research in this area is still ongoing [49].
Cadherins are important components of a tight junction that bind to the cytoskeleton of a cell with the help of catenins to form tight adhesive contacts.
The main task of additional cytoplasmic proteins is to establish strong connections between membrane proteins and the cytoskeleton. The zonula occludens proteins ZO-1, ZO-2 and ZO-3 bind claudins, and ZO-1 binds occludins and JAMs. Through cingulin, ZO-1, ZO-2 and ZO-3 establish connections between claudin and occludin and between each other and intracellular actin [50]. The function of the β-catenin and γ-catenin proteins is to connect cadherins to the cytoskeleton via the α-catenin protein or its analog vinculin [51,52,53].
3.3. Pericytes
Pericytes are localized outside the capillary endothelium and share a basement membrane with it, and they are involved in the synthesis of elements of the basement membrane (see Figure 1) [54,55]. Pericytes are critical yet understudied cells that are a central component of the neurovascular unit [56]. Through processes that envelop the endothelium and the ability to contract, pericytes regulate the permeability of the interendothelial junction to fluids. Due to their close contact with the capillary endothelium, metabolites, ribonucleic acids (RNAs) and ions are exchanged between cells, and the safety of this exchange is ensured by the ability of pericytes to phagocytose them, which allows these cells to neutralize toxic metabolites [57,58,59,60,61]. Other important functions of pericytes are the expression of vasopressin, angiotensin, and endothelin receptors, which allows them to participate in cerebral autoregulation; the regulation of the expression of BBB-specific genes in endothelial cells; and the induction of the polarization of glial cell processes (astrocytes) that envelop the CNS capillaries [62,63]. Numerous studies have revealed the important role of pericytes in the activation of endothelial and astrocyte functions in the regulation of the BBB [54,58]. Damage to pericytes can occur during many neurological diseases, especially degenerative diseases (dementia, Alzheimer's disease, multiple sclerosis, etc.) [56,64,65,66,67]. A decrease in the resistance of endothelial cells to apoptosis can be caused by the loss of their connection with pericytes, which can lead to a loss of BBB integrity.
Although pericytes are increasingly recognized as important in the control of BBB permeability and vascular flow, pericyte heterogeneity and species-specific differences between animals (mouse) and humans have contributed to many discrepancies in research results [68]. The precise identification and definition of pericytes and pericyte subtypes in humans should facilitate an understanding of the biology of these cells and the development of new therapeutic approaches that specifically target disease-associated pericyte subtypes in individuals with neurodegenerative diseases [69].
3.4. Basement Membrane
The basement membrane of capillaries, a form of extracellular matrix, is located under endothelial and epithelial cells. Its main functions are the adhesion of cells on its surface, maintenance of the BBB structure, and transmission of signals between its elements [70]. The basement membrane consists of four proteins (collagen IV, laminin, nidogen, and perlecan) that form a plate of 50 to 100 nm thick [71]. These proteins are synthesized by the endothelial cells of blood vessels in the brain, pericytes and astrocytes. Collagen IV is the most abundant basement membrane protein. Its functional feature is that it does not participate in the formation of the membrane structure, but it maintains it. Collagen IV also plays an important role in maintaining the integrity of vessels [72]. Another element of the barrier is laminin. The laminin protein can be produced in different isoforms depending on the type of cell that synthesizes it. Laminin synthesized by astrocytes plays an important role in maintaining the BBB, which is indispensable for the integrity of the barrier [73,74]. Collagen IV and laminin are stabilized by the protein nidogen (entactin) [75]. The role of nidogen in maintaining the integrity of the BBB is not yet fully understood [28,76]. Perlecan, a large heparan sulfate proteoglycan (HSPG), is involved in the formation of the basement membrane starting in the embryonic period. The protein core of perlecan is 400 kDa in size and consists of five distinct structural domains that enable interactions with extracellular matrix proteins. The influence and importance of perlecan in maintaining the integrity of the BBB are not sufficiently understood [77,78,79].
3.5. Perivascular Membrane
The perivascular membrane formed by glial cell processes consists of astrocytes, which are the most common cells in the central nervous system and perform many important functions; in particular, astrocytes play an important role in the inflammatory process during the development of neurodegenerative diseases [80]. Given that astrocytes are formed in the postnatal period, i.e., after the BBB has formed and thickened, these cells may not be involved in the formation of the barrier itself. However, an important role in maintaining the BBB structure is played by the processes (“legs”) of astrocytes, which are in close contact with the brain capillaries [81]. Researchers have not yet been established whether astrocytes affect the level of BBB permeability, but some studies have confirmed the involvement of these cells in the formation of tight junctions by modulating the expression of vascular endothelial growth factor (VEGF) and angiotensin-1 [82,83].
Understanding the main structural units of the BBB as comprehensively as possible provides the basis for a detailed analysis of the impact of proinflammatory cytokines on their integrity.
4. Influence of Proinflammatory Cytokines on the Disruption of the BBB Structure
The effects of cytokines on the BBB structure depend on the mechanism of barrier penetration [84]. When cytokines enter the body exogenously, for example, through injection, they can quickly and easily cross the BBB without disrupting its structure. This process can occur through retrograde axonal transport, saturated influx transport (SIT), or simple diffusion in areas of the brain where the BBB is incomplete. These areas correspond to CVOs. The cytokines IL-1α, IL-6, and TNF-α can cross the barrier via these CVO-related pathways [85,86]. Usually, the exogenous route of cytokine transport into the body is controlled, and thus when the barrier is crossed, its structure is not disturbed and the disturbance is insignificant. However, this type of crossing, even without structural disruption, can compromise the integrity of the BBB by activating free calcium and potentially disrupting its homeostasis in the brain [87]. The endogenous pathway through which proinflammatory cytokines pass through the BBB is more dangerous because of the structural integrity of the barrier. Damage to the BBB structure and, accordingly, an increase in its permeability are most often associated with various neurological disorders, psychiatric disorders, and neurodegenerative and infectious diseases of the CNS, which increase the levels of proinflammatory cytokines [18,88] (Figure 3).
Increased levels of proinflammatory cytokines depend on the nature and characteristics of pathological processes and conditions of the nervous system [127].
The permeability of the BBB is indirectly affected by autoimmune diseases, which lead to an increase in cytokine levels. Thus, certain changes in the immune system disrupt BBB permeability, after which autoreactive T lymphocytes enter the central nervous system, increasing the levels of the proinflammatory cytokines IL-1, TNF-α, and γ-interferon. As a result, the process of autoimmune attack on the myelin sheath is triggered, leading to the development of multiple sclerosis [128,129].
The release of cytokines, along with oxidants and proteolytic enzymes, also occurs in ischemic stroke. The ingress of these elements into the brain tissue provokes the development of cytotoxic edema and impaired BBB permeability, which leads to the penetration of leukocytes through the capillary endothelium into the brain tissue and damage to healthy neurons [130].
The main role of the neuroinflammatory process in the pathogenesis of epilepsy involves the mobilization of the proinflammatory cytokines IL-1β, IL-6 and TNF-α in response to pathological changes in the nervous system caused by traumatic brain injury (TBI), brain tumors, stroke, drug exposure, etc. [131,132] Discussions about which process is primary—damage to the BBB or seizures—are ongoing. After considering one of the most common causes of epilepsy, TBI, investigations of how BBB damage is associated with seizures are possible. Depending on the consequences of TBI, BBB disruption occurs mechanically or as a result of proinflammatory cytokines that are activated in response to trauma. Blood components, one of which is thrombin, enter the cerebrospinal fluid through a hemorrhage, where they provoke neuronal excitability and, accordingly, seizures [133,134].
The tau (τ) protein, a neuronal protein that supports axonal transport and microtubule dynamics, plays an important role in the inflammatory response to TBI, which also affects the integrity of the BBB [135]. Tau preserves cellular integrity and shifts cellular signaling away from pro-cell death signaling cascades. Tauopathies represent a heterogeneous group of approximately 20 different neurodegenerative diseases characterized by the abnormal deposition of the microtubule-associated protein tau (MAPT) in cells of the nervous system [136]. The BBB damage observed in individuals with tauopathies without amyloid-β overproduction suggests a role for tau in BBB damage that is driven by chronic neuroinflammation initiated within the microglial compartment by extracellular tau aggregates (e-τ) released from injured neurons [137].
The increase in the levels of proinflammatory cytokines (IL-1β, IL-6, IL-12, TNF-α and IFN-γ) is also influenced by several mental disorders, including depression. Inflammatory activity can both provoke and maintain a specific (proinflammatory) cytokine profile, and so prolonged exposure of the nervous system to this state alter BBB permeability and, accordingly, provoke other neurological disorders and diseases [138].
The activation of astrocytes and increased levels of the cytokines IL-1β, IL-6, and TNF-α have been observed under the influence of neurotoxic drugs. Thus, damage to the BBB and an increase in its permeability caused by methamphetamine neurotoxicity are manifested by decreases in the levels of claudin and occludin, swelling of astrocytes and their processes, a decrease in pericyte coverage, and the loss of tight junctions of endothelial cells [139,140,141].
The factors that provoke increased production of proinflammatory cytokines (IL-6 and TNF-α) include the histamine H4 receptor, which indirectly activates mast cells, and along with cytokines, actively produce chemokines and histamine [142].
Chemokines are cytokines with chemotactic activity that are involved in the physiological and pathological processes of the CNS [143,144,145,146]. Under physiological conditions in the nervous system, chemokines contribute to cellular interactions, the activation of signaling pathways, and the maintenance of CNS homeostasis. For example, the chemokines CCL2, CCL19, CCL20, CCL21, and CXCL12 are involved in immunological surveillance by transmitting signals and activating immune cells; CX3CL1 (fractalkine) is involved in maintaining adult neurogenesis, ensuring the balance of homeostasis, and limits the expression of the proinflammatory cytokines IL-1β, IL-6, and TNF-α [147,148]. The chemokines CXCL12, CCL19, CCL20, and CCL21 are expressed in the vascular system of the BBB [149]. Under pathological conditions, chemokines act as mediators of cell migration and participate in the regulation of inflammatory and autoimmune (differentiation and growth of cells, including tumors) processes in the CNS [150]. The mechanism by which CX3CL1 affects BBB integrity depends on the cells involved in migration. For example, one mechanism is triggered by a significant accumulation of CD16+ monocytes on inflamed cerebral endothelial cells due to their transendothelial migration in response to CX3CL1 expression [151]. In addition, the migration of CD4+ T cells is ensured by the chemoattractant activity of CX3CL1 [152]. Changes in BBB permeability are affected by an increase in CXCL13 levels, which occurs as a result of the development of tumors with cerebral metastases. This process leads to an increase in the paracellular permeability of the capillary endothelium and a decrease in the expression and localization of the tight junction proteins claudin-5 and occludin [153].
Investigating and identifying the effect of histamine, a biogenic amine formed by the conversion of histidine via histidine decarboxylase (HDC), on BBB integrity are important [154]. The histamine molecule is not small enough to penetrate the capillary endothelium, and thus it cannot cross the BBB unhindered [155]. However, histamine H1 and H2 receptors are expressed on both luminal (blood-facing) and abluminal (brain-facing) plasma membranes of the brain endothelium and regulate several endothelial functions, such as blood coagulation and the permeability of the BBB [156,157]. Histamine is able to increase the permeability of the BBB by downregulating the major tight junction membrane proteins claudin-5 and occludin and the additional cytoplasmic protein ZO-1. Moreover, the expression of the histamine H2 receptor can be used as a predictor of the barrier permeability provoked by histamine [158]. Conversely, endothelial histamine H1 receptor signaling reduces BBB permeability and the susceptibility to autoimmune encephalomyelitis [159]. Brain tissue should also be considered a separate source of histamine, the abundance of which is determined by the availability of L-histidine and the local activity of HDCs [160]. Newly synthesized neuronal histamine is thought to be stored within nerve terminal vesicles. Both in vivo and in vitro studies have shown that the depolarization of nerve terminals activates the exocytotic release of histamine via a voltage- and calcium-dependent mechanism [161].
5. Proinflammatory Cytokines Within the BBB: Clinical Approaches and Potential Therapies
An investigation of the structure of the BBB and the effects of proinflammatory cytokines on damage to the BBB revealed that the cytokines IL-1, IL-6 and TNF-α are most often involved in barrier disruption [149]. Therefore, counteracting the mechanism underlying their effect on the BBB should be addressed in the development of therapeutic approaches to preserve BBB integrity and prevent neurological disorders provoked by the ingress of blood elements into nervous tissue. Moreover, the ability of proinflammatory cytokines to increase BBB permeability can also be used to develop new therapeutic strategies, for example, creating drugs based on cytokine derivatives with good CNS penetration for use in anticancer therapy [162,163,164].
Under normal conditions, each system of the body functions and interacts safely with other systems, supporting the process of vital activity. The influence of pathological factors causes a corresponding increase in the immune response, the regulators of which, while performing their tasks at the highest level of intensity, can cause damage to other systems and structures [165,166]. Studies of the BBB revealed that its main functions (protection and regulation) are performed primarily through the structures of the combination of all elements, from the specific shape of endothelial cells and special tight connections between them to the placement of astrocyte processes in a tight ring around the basement membrane. In response to pathological processes in the nervous system, including autoimmune diseases (multiple sclerosis, Guillain–Barré syndrome, and myasthenia gravis), the immune response involves systemic inflammation mobilized by proinflammatory cytokines [167,168,169,170,171,172]. As reported in previous studies, a significant increase in the concentrations of these cytokines in the blood can damage the BBB structure and increase its permeability through a disruption of the tight junctions of capillary endothelial cells and other barrier components. The data obtained can be used as a foundation for the development of therapeutic methods to control the levels of proinflammatory cytokines to preserve the integrity of the BBB.
This issue was addressed by Takeshita et al. [173]. These scientists built static BBB models to study long-term barrier function and have shown that IL-6 blockade suppresses BBB disorders, preventing the onset of visual spectrum neuromyelitis. This study revealed that the inhibition of T-cell migration to the nervous tissue of the spinal cord was influenced by the blockade of IL-6 signal transduction, which prevented the increase in BBB permeability. IL-6 blockade was achieved by transferring neuromyelitis optica immunoglobulin G (NMO-IgG) autoantibodies and satralizumab through the BBB using a triple culture system, which mimics the close contact of endothelial cells, pericytes and astrocyte processes [173]. The conclusions of this study are convincing and promising because the results can serve as examples for the development of methods to block other proinflammatory cytokines, such as IL-1β and TNF-α, using the BBB model proposed by the authors.
As already mentioned, the ability of proinflammatory cytokines to overcome the BBB should be considered not only to find methods to limit it but also to use it as a tool to overcome the BBB as an obstacle to drug penetration into the nervous tissue. Corti et al. [174] investigated the use of TNF-α as a “conductor” of drugs to cross the BBB to deliver antitumor therapy directly to brain tumors. This team of researchers has studied the capabilities and properties of the drug NGR-TNF, a fusion of a peptide and the cytokine, which includes the NGR-TNF-α molecule combined with the peptide Cys-Asn-Gly-Arg-Cys-Gly (CNGRCG) (denoted as NGR in the drug's name) and the ligand of aminopeptidase N (CD13) of tumor blood vessel-positive (+) blood vessels. The results of preclinical and clinical trials have shown the effectiveness of this drug in changing the selectivity of the tumor BBB, which has improved the quality of chemotherapy and increased patient survival. However, the authors also noted the problems that could arise due to the instability and molecular heterogeneity of this drug. Given the therapeutic potential of drugs based on the combination of peptides and cytokines, the strategy of this nonimmunogenic approach for the safe treatment of the BBB at the tumor site seems promising, increasing the therapeutic index of cytokines in cancer therapy.
In addition to factors related to exogenous BBB disruption (brain injuries, infections, radiation, etc.), nervous system processes provoked by internal causes such as stress or sleep loss also affect BBB integrity. The regulatory effect of sleep on the BBB was studied by Hurtado-Alvarado et al. [175]. They reported that sleep loss (sleep deprivation) provoked a low-grade inflammatory state. According to the study, increased BBB permeability was observed in experimental mice with limited sleep time. The hippocampus of these mice presented increased expression of neuroinflammatory markers, such as TNF, IFN-γ and the adenosine receptor. Notably, BBB disruptions appeared after ten days of prolonged sleep loss or disruption, and the damaged BBB structure showed a limited ability to rebuild/heal, which could provoke complex nervous disorders and neurodegenerative diseases.
The pandemic of acute respiratory disease caused by COVID-19 aroused the interest of researchers in the effects of the highly transmissible, pathogenic severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) on various body systems. Zhang et al. [176] reported that SARS-CoV-2 overcomes the BBB, accompanied by the destruction of the basement membrane without affecting tight junctions. An in vivo experiment detected the SARS-CoV-2 RNA in the vascular wall, perivascular space, and microvascular endothelial cells of the brains of infected mice, and BBB breakdown was observed in infected hamsters. The peculiarity of BBB damage in animals is associated with the fact that the expression of the main proteins forming the tight junctions of endothelial cells (claudin, occludin, and JAMs) does not change, but the basement membrane is disrupted. This study reveals the transcellular pathway of BBB damage caused by the SARS-CoV-2 virus, which bypasses tight junctions and breaks the barrier of endothelial cells. In addition, this effect of SARS-CoV-2 on the BBB is hypothetically dangerous for the integrity of the barrier, since the virus damages one of its structural units and activates the immune response in the form of the mobilization of proinflammatory –cytokines; moreover, the subsequent recovery process may be significantly complicated or even impossible. However, verification of this statement requires additional research.
The impact of SARS-CoV-2 is also considered to cause elevated levels of proinflammatory cytokines, referred to as the cytokine storm [177]. When investigating the pathogenesis of the cytokine storm, Soy et al. [178] studied data from modern scientific articles on the topic and reported that the manifestation of such a strong and dangerous process may be associated with the genetic features of SARS-CoV-2, as well as gene mutations in the body. The consequences of the cytokine storm are a high mortality rate, and treatment, such as antiviral anti-inflammatory and antirheumatic therapy, is administered to target several processes at once. The effects of cytokine storms on the BBB are poorly understood [179]. Researchers have assumed that the mechanism underlying its effect on the barrier is standard but much more intense, but no studies have determined the level of BBB recovery after successful treatment of the cytokine storm.
Chen et al. [180] conducted a study that examined the role of IL-17 in neuroinflammation causing chronic migraine. The study revealed that, by crossing the BBB in the medulla oblongata, IL-17 causes the activation of previously injected nitroglycerin through microglia-mediated neuroinflammation. Thus, the mechanism of chronic migraine triggered by nitroglycerin was established, and a new therapeutic approach, which consists of blocking IL-17 as a factor contributing to neuroinflammation, was proposed. Given that the results were obtained in an in vivo study in rats, any proposed therapeutic approaches must involve full preclinical studies and clinical trials in humans.
In terms of its main functions, the BBB plays a very important role not only in the CNS but also in the vital activities of the whole organism. Knowledge regarding the effects of proinflammatory cytokines on the BBB structure reviewed in this paper may be useful in the development and implementation of therapeutic strategies that, depending on the task, are based on a blockade of these cytokines that is safe for the immune system or quickly and effectively restores the integrity of the damaged BBB structure [81].
6. Concluding Remarks
This review summarizes the current knowledge of the structure of the BBB and how proinflammatory cytokines act to damage it. A review of the literature was performed, which included information on the main components of the BBB and the mechanisms of action of proinflammatory cytokines that disrupt the integrity of the BBB.
The most important barrier element of the BBB is the capillary endothelium. The peculiarities of its structure lie in its specific structure, the location of endothelial cells and presence of tight junctions that ensure the reliability of communication between them, resulting in the selective permeability of the BBB. This task is ensured by the mechanism of building tight contacts, namely, the connections established by additional cytoplasmic proteins (zonula occludens-1, -2, -3; cingulin; α-catenin; β-catenin; γ-catenin; and vinculin) with the main membrane proteins (claudin, occludin, JAMs, and cadherins) and the main cytoskeletal protein (actin). Pericytes play important roles in regulating the permeability of the interendothelial junction to fluids and the exchange of metabolites, RNA, and ions with endothelial cells. The shape of the basement membrane (a structure consisting of four proteins: collagen IV, laminin, nidogen and perlecan) allows capillary endothelial cells and pericytes to adhere to it, thus ensuring the transmission of signals between them and maintaining the structure of the BBB. Due to the close contact of astrocytes with capillaries in the brain, the perivascular membrane formed from their processes ensures the integrity of the barrier.
The effects of proinflammatory cytokines on disruptions in the BBB structure and permeability are directly related to their main function—mobilization of the inflammatory response to pathological processes in the nervous system. Most often, this effect is associated with the actions of the cytokines IL-1β, IL-6, and TNF-α. The mechanism of their interaction with the structural elements of the barrier is as follows: disruption of the tight junctions of capillary endothelial cells due to the destruction of the structural protein claudin, a reduction in the level of the β-catenin protein, and damage to astrocytes.
In conclusion, determining the mechanism underlying the effects of proinflammatory cytokines on the BBB, in addition to theory, is of practical importance, as it can be used to develop therapeutic methods to neutralize these negative effects. However, exploring the possibility of using drugs that penetrate the BBB may be a key concept in the development of drugs for the treatment of pathological changes in nervous tissue, such as brain tumors.
Further cytokine research in this area may include the study of the effects of anti-inflammatory cytokines and a group of cytokines (e.g., chemokines) regulating cellular and humoral immunity on the BBB structure, which will allow us to obtain a holistic picture of the types of interactions of these proteins with this barrier.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing in not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BBB | blood‒brain barrier |
| BMECs | brain microvascular endothelial cells |
| CCL2 | C-C motif chemokine ligand 2, also known as monocyte chemotactic protein 1 (MCP1) |
| CCL19 | C-C motif chemokine ligand 19, also known as macrophage inflammatory protein-3 beta (MIP-3β) |
| CCL20 | C-C motif chemokine ligand 20 |
| CCL21 | C-C motif chemokine ligand 21 |
| CD13 | cluster of differentiation 13, also known as aminopeptidase N (APN) or alanyl aminopeptidase |
| CNS | central nervous system |
| COVID-19 | coronavirus disease 2019 |
| CSF | cerebrospinal fluid |
| CVOs | circumventricular organs |
| CX3CL1 | C-X3-C motif chemokine ligand 1, also known as fractalkine |
| CXCL12 | C-X-C motif chemokine ligand 12, also known as stromal cell-derived factor 1 (SDF-1) |
| e-τ | extracellular tau aggregates |
| GBM | glioblastoma multiforme |
| GLUT1 | glucose transporter 1 |
| HDC | histidine decarboxylase |
| HSPG | heparan sulfate proteoglycan |
| ICAM-1 | intercellular adhesion molecule-1, also known as cluster of differentiation 54 (CD54) |
| IFN-γ | Interferon gamma |
| IL-1, IL-1α, IL-1β, IL-2, IL-4, IL-6, IL-10, IL-12, IL-15, IL-17, IL-18 | interleukins: interleukin 1, interleukin 1-alpha, interleukin 1-beta, interleukin 2, interleukin 4, interleukin 6; interleukin 10, interleukin 12, interleukin 15, interleukin 17, interleukin 18, respectively |
| JAMs | junctional adhesion molecules |
| kDa | kilodalton (a non-SI unit of mass) |
| MAPT | microtubule-associated protein tau |
| MMP-2, MMP-3, MMP-9 | matrix metalloproteinases 2, 3 and 9, respectively |
| NGR-TNF | a drug created by fusing a peptide Cys-Asn-Gly-Arg-Cys-Gly (CNGRCG), denoted as NGR, with a TNF-α molecule |
| NK cells | natural killer cells |
| NMO-IgG | neuromyelitis optica immunoglobulin G |
| PCs | pericytes |
| P/E selectins | platelet/endothelial selectins, also known as CD62P/CD62E, respectively |
| RNAs | ribonucleic acids |
| ROS | reactive oxygen species |
| S100B | S100 calcium-binding protein beta |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| SHH | sonic hedgehog homolog (protein) |
| SIT | saturated influx transport |
| TBI | traumatic brain injury |
| TIMP-1, TIMP-2 | tissue inhibitors of metalloproteinases 1 and 2, respectively |
| TNF-α | tumor necrosis factor alpha |
| TSP-2 | thrombospondin-2 |
| VCAM-1 | vascular cell adhesion molecule 1, also known as cluster of differentiation 106 (CD106) |
| VEGF | vascular endothelial growth factor |
| ZO-1, ZO-2 and ZO-3 | zonula occludens proteins 1, 2, and 3, respectively |
References
- Dyrna F, Hanske S, Krueger M, Bechmann I. The blood-brain barrier. J Neuroimmune Pharmacol. 2013; 8(4):763-73. [CrossRef]
- Khan E. An examination of the blood-brain barrier in health and disease. Br J Nurs. 2005;14(9):509-13. [CrossRef]
- Benz F, Liebner S. Structure and Function of the Blood-Brain Barrier (BBB). Handb Exp Pharmacol. 2022;273:3-31. [CrossRef]
- Kim JH, Byun HM, Chung EC, Chung HY, Bae ON. Loss of Integrity: Impairment of the Blood-brain Barrier in Heavy Metal-associated Ischemic Stroke. Toxicol Res. 2013;29(3):157-64. [CrossRef]
- Yang C, Hawkins KE, Doré S, Candelario-Jalil E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am J Physiol Cell Physiol. 2019;316(2):C135-C153. [CrossRef]
- Takata F, Nakagawa S, Matsumoto J, Dohgu S. Blood-Brain Barrier Dysfunction Amplifies the Development of Neuroinflammation: Understanding of Cellular Events in Brain Microvascular Endothelial Cells for Prevention and Treatment of BBB Dysfunction. Front Cell Neurosci. 2021; 15:661838. [CrossRef]
- Zhao B, Yin Q, Fei Y, Zhu J, Qiu Y, Fang W, Li Y. Research progress of mechanisms for tight junction damage on blood-brain barrier inflammation. Arch Physiol Biochem. 2022;128(6):1579-1590. [CrossRef]
- Li X, Cai Y, Zhang Z, Zhou J. Glial and Vascular Cell Regulation of the Blood-Brain Barrier in Diabetes. Diabetes Metab J. 2022;46(2):222-238. [CrossRef]
- Cai Z, Qiao PF, Wan CQ, Cai M, Zhou NK, Li Q. Role of Blood-Brain Barrier in Alzheimer's Disease. J Alzheimers Dis. 2018;63(4):1223-1234. [CrossRef]
- Nikolopoulos D, Manolakou T, Polissidis A, Filia A, Bertsias G, Koutmani Y, Boumpas DT. Microglia activation in the presence of intact blood-brain barrier and disruption of hippocampal neurogenesis via IL-6 and IL-18 mediate early diffuse neuropsychiatric lupus. Ann Rheum Dis. 2023;82(5):646-657. [CrossRef]
- Wątroba M, Grabowska AD, Szukiewicz D. Effects of Diabetes Mellitus-Related Dysglycemia on the Functions of Blood-Brain Barrier and the Risk of Dementia. Int J Mol Sci. 2023;24(12):10069. [CrossRef]
- Wang J, Li G, Wang Z, Zhang X, Yao L, Wang F, Liu S, Yin J, Ling EA, Wang L, Hao A. High glucose-induced expression of inflammatory cytokines and reactive oxygen species in cultured astrocytes. Neuroscience. 2012;202:58-68. [CrossRef]
- Lesniak A, Poznański P, Religa P, Nawrocka A, Bujalska-Zadrozny M, Sacharczuk M. Loss of Brain-Derived Neurotrophic Factor (BDNF) Resulting From Congenital- Or Mild Traumatic Brain Injury-Induced Blood-Brain Barrier Disruption Correlates With Depressive-Like Behaviour. Neuroscience. 2021;458:1-10. [CrossRef]
- Małkiewicz MA, Małecki A, Toborek M, Szarmach A, Winklewski PJ. Substances of abuse and the blood brain barrier: Interactions with physical exercise. Neurosci Biobehav Rev. 2020;119:204-216. [CrossRef]
- Bronisz E, Cudna A, Wierzbicka A, Kurkowska-Jastrzębska I. Blood-Brain Barrier-Associated Proteins Are Elevated in Serum of Epilepsy Patients. Cells. 2023;12(3):368. [CrossRef]
- Łach A, Wnuk A, Wójtowicz AK. Experimental Models to Study the Functions of the Blood-Brain Barrier. Bioengineering (Basel). 2023;10(5):519. [CrossRef]
- Patel JP, Frey BN. Disruption in the Blood-Brain Barrier: The Missing Link between Brain and Body Inflammation in Bipolar Disorder? Neural Plast. 2015;2015:708306. [CrossRef]
- Yarlagadda A, Alfson E, Clayton AH. The blood brain barrier and the role of cytokines in neuropsychiatry. Psychiatry (Edgmont). 2009;6(11):18-22. https://www.ncbi.nlm.nih. 2801.
- Haley MJ, Lawrence CB. The blood-brain barrier after stroke: Structural studies and the role of transcytotic vesicles. J Cereb Blood Flow Metab. 2017;37(2):456-470. [CrossRef]
- Mayer MG, Fischer T. Microglia at the blood brain barrier in health and disease. Front Cell Neurosci. 2024;18:1360195. [CrossRef]
- Lim SH, Yee GT, Khang D. Nanoparticle-Based Combinational Strategies for Overcoming the Blood-Brain Barrier and Blood-Tumor Barrier. Int J Nanomedicine. 2024;19:2529-2552. [CrossRef]
- Wu D, Chen Q, Chen X, Han F, Chen Z, Wang Y. The blood-brain barrier: structure, regulation, and drug delivery. Signal Transduct Target Ther. 2023;8(1):217. [CrossRef]
- McCabe SM, Zhao N. The Potential Roles of Blood-Brain Barrier and Blood-Cerebrospinal Fluid Barrier in Maintaining Brain Manganese Homeostasis. Nutrients. 2021;13(6):1833. [CrossRef]
- Patabendige A, Janigro D. The role of the blood-brain barrier during neurological disease and infection. Biochem Soc Trans. 2023;51(2):613-626. [CrossRef]
- Dotiwala AK, McCausland C, Samra NS. Anatomy, Head and Neck: Blood Brain Barrier. [Updated 2023 Apr 4]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-. Available from: https://www.ncbi.nlm.nih. 5195.
- Ganong WF. Circumventricular organs: definition and role in the regulation of endocrine and autonomic function. Clin Exp Pharmacol Physiol. 2000;27(5-6):422-7. [CrossRef]
- Verkhratsky A, Pivoriūnas A. Astroglia support, regulate and reinforce brain barriers. Neurobiol Dis. 2023;179:106054. [CrossRef]
- Xu L, Nirwane A, Yao Y. Basement membrane and blood-brain barrier. Stroke Vasc Neurol. 2018; 4(2):78-82. [CrossRef]
- Erickson MA, Banks WA. Transcellular routes of blood-brain barrier disruption. Exp Biol Med (Maywood). 2022;247(9):788-796. [CrossRef]
- Persidsky Y, Ramirez SH, Haorah J, Kanmogne GD. Blood-brain barrier: structural components and function under physiologic and pathologic conditions. J Neuroimmune Pharmacol. 2006;1(3):223-36. [CrossRef]
- Weiss N, Miller F, Cazaubon S, Couraud PO. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim Biophys Acta. 2009 Apr;1788(4):842-57. [CrossRef]
- Xie Y, He L, Lugano R, Zhang Y, Cao H, He Q, Chao M, Liu B, Cao Q, Wang J, Jiao Y, Hu Y, Han L, Zhang Y, Huang H, Uhrbom L, Betsholtz C, Wang L, Dimberg A, Zhang L. Key molecular alterations in endothelial cells in human glioblastoma uncovered through single-cell RNA sequencing. JCI Insight. 2021;6(15):e150861. [CrossRef]
- Langen UH, Ayloo S, Gu C. Development and Cell Biology of the Blood-Brain Barrier. Annu Rev Cell Dev Biol. 2019;35:591-613. [CrossRef]
- Ozgür B, Helms HCC, Tornabene E, Brodin B. Hypoxia increases expression of selected blood-brain barrier transporters GLUT-1, P-gp, SLC7A5 and TFRC, while maintaining barrier integrity, in brain capillary endothelial monolayers. Fluids Barriers CNS. 2022;19(1):1. [CrossRef]
- Bhowmick S, D'Mello V, Caruso D, Wallerstein A, Abdul-Muneer PM. Impairment of pericyte-endothelium crosstalk leads to blood-brain barrier dysfunction following traumatic brain injury. Exp Neurol. 2019;317:260-270. [CrossRef]
- Kakava S, Schlumpf E, Panteloglou G, Tellenbach F, von Eckardstein A, Robert J. Brain Endothelial Cells in Contrary to the Aortic Do Not Transport but Degrade Low-Density Lipoproteins via Both LDLR and ALK1. Cells. 2022;11(19):3044. [CrossRef]
- Zhao F, Zhong L, Luo Y. Endothelial glycocalyx as an important factor in composition of blood-brain barrier. CNS Neurosci Ther. 2021;27(1):26-35. Erratum in: CNS Neurosci Ther. 2021;27(7):862. [CrossRef]
- Kadry H, Noorani B, Cucullo L. A blood-brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS. 2020;17(1):69. [CrossRef]
- Kniesel U, Wolburg H. Tight junctions of the blood-brain barrier. Cell Mol Neurobiol. 2000;20(1): 57-76. [CrossRef]
- Feng S, Zou L, Wang H, He R, Liu K, Zhu H. RhoA/ROCK-2 Pathway Inhibition and Tight Junction Protein Upregulation by Catalpol Suppresses Lipopolysaccaride-Induced Disruption of Blood-Brain Barrier Permeability. Molecules. 2018;23(9):2371. [CrossRef]
- Haseloff RF, Dithmer S, Winkler L, Wolburg H, Blasig IE. Transmembrane proteins of the tight junctions at the blood-brain barrier: structural and functional aspects. Semin Cell Dev Biol. 2015; 38:16-25. [CrossRef]
- Koumangoye R, Penny P, Delpire E. Loss of NKCC1 function increases epithelial tight junction permeability by upregulating claudin-2 expression. Am J Physiol Cell Physiol. 2022;323(4):C1251-C1263. [CrossRef]
- Yamamoto TM, Webb PG, Davis DM, Baumgartner HK, Woodruff ER, Guntupalli SR, Neville M, Behbakht K, Bitler BG. Loss of Claudin-4 Reduces DNA Damage Repair and Increases Sensitivity to PARP Inhibitors. Mol Cancer Ther. 2022;21(4):647-657. [CrossRef]
- Kim NY, Pyo JS, Kang DW, Yoo SM. Loss of claudin-1 expression induces epithelial-mesenchymal transition through nuclear factor-κB activation in colorectal cancer. Pathol Res Pract. 2019;215(3):580-585. [CrossRef]
- Greene C, Hanley N, Campbell M. Claudin-5: gatekeeper of neurological function. Fluids Barriers CNS. 2019;16(1):3. [CrossRef]
- Günzel D, Fromm M. Claudins and other tight junction proteins. Compr Physiol. 2012;2(3):1819-52. [CrossRef]
- Komarova YA, Kruse K, Mehta D, Malik AB. Protein Interactions at Endothelial Junctions and Signaling Mechanisms Regulating Endothelial Permeability. Circ Res. 2017;120(1):179-206. [CrossRef]
- Liu WY, Wang ZB, Zhang LC, Wei X, Li L. Tight junction in blood-brain barrier: an overview of structure, regulation, and regulator substances. CNS Neurosci Ther. 2012;18(8):609-15. [CrossRef]
- Bergmann S, Lawler SE, Qu Y, Fadzen CM, Wolfe JM, Regan MS, Pentelute BL, Agar NYR, Cho CF. Blood-brain-barrier organoids for investigating the permeability of CNS therapeutics. Nat Protoc. 2018;13(12):2827-2843. [CrossRef]
- Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta. 2008;1778(3):660-9. [CrossRef]
- Kuo WT, Odenwald MA, Turner JR, Zuo L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann N Y Acad Sci. 2022;1514(1):21-33. [CrossRef]
- Wang Q, Huang X, Su Y, Yin G, Wang S, Yu B, Li H, Qi J, Chen H, Zeng W, Zhang K, Verkhratsky A, Niu J, Yi C. Activation of Wnt/β-catenin pathway mitigates blood-brain barrier dysfunction in Alzheimer's disease. Brain. 2022;145(12):4474-4488. [CrossRef]
- Liebner S, Gerhardt H, Wolburg H. Differential expression of endothelial beta-catenin and plakoglobin during development and maturation of the blood-brain and blood-retina barrier in the chicken. Dev Dyn. 2000;217(1):86-98. [CrossRef]
- Alarcon-Martinez L, Yemisci M, Dalkara T. Pericyte morphology and function. Histol Histopathol. 2021;36(6):633-643. [CrossRef]
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97(6):512-23. [CrossRef]
- Whitehead B, Karelina K, Weil ZM. Pericyte dysfunction is a key mediator of the risk of cerebral ischemia. J Neurosci Res. 2023;101(12):1840-1848. [CrossRef]
- Kamouchi M, Ago T, Kitazono T. Brain pericytes: emerging concepts and functional roles in brain homeostasis. Cell Mol Neurobiol. 2011;31(2):175-93. [CrossRef]
- Sá-Pereira I, Brites D, Brito MA. Neurovascular unit: a focus on pericytes. Mol Neurobiol. 2012; 45(2):327-47. [CrossRef]
- Armulik A, Genové G, Mäe M, Nisancioglu MH, Wallgard E, Niaudet C, He L, Norlin J, Lindblom P, Strittmatter K, Johansson BR, Betsholtz C. Pericytes regulate the blood-brain barrier. Nature. 2010; 468(7323):557-61. [CrossRef]
- Dudvarski Stankovic N, Teodorczyk M, Ploen R, Zipp F, Schmidt MHH. Microglia-blood vessel interactions: a double-edged sword in brain pathologies. Acta Neuropathol. 2016;131(3):347-63. [CrossRef]
- Fu J, Liang H, Yuan P, Wei Z, Zhong P. Brain pericyte biology: from physiopathological mechanisms to potential therapeutic applications in ischemic stroke. Front Cell Neurosci. 2023;17:1267785. [CrossRef]
- Su X, Huang L, Qu Y, Xiao D, Mu D. Pericytes in Cerebrovascular Diseases: An Emerging Therapeutic Target. Front Cell Neurosci. 2019;13:519. [CrossRef]
- Brown LS, Foster CG, Courtney JM, King NE, Howells DW, Sutherland BA. Pericytes and Neurovascular Function in the Healthy and Diseased Brain. Front Cell Neurosci. 2019;13:282. [CrossRef]
- Liu Q, Yang Y, Fan X. Microvascular pericytes in brain-associated vascular disease. Biomed Pharmacother. 2020;121:109633. [CrossRef]
- Anwar MM, Özkan E, Gürsoy-Özdemir Y. The role of extracellular matrix alterations in mediating astrocyte damage and pericyte dysfunction in Alzheimer's disease: A comprehensive review. Eur J Neurosci. 2022;56(9):5453-5475. [CrossRef]
- Rivera FJ, Hinrichsen B, Silva ME. Pericytes in Multiple Sclerosis. Adv Exp Med Biol. 2019;1147:167-187. [CrossRef]
- Benarroch E. What Are the Roles of Pericytes in the Neurovascular Unit and Its Disorders? Neurology. 2023;100(20):970-977. [CrossRef]
- Bohannon DG, Long D, Kim WK. Understanding the Heterogeneity of Human Pericyte Subsets in Blood-Brain Barrier Homeostasis and Neurological Diseases. Cells. 2021;10(4):890. [CrossRef]
- Nwadozi E, Rudnicki M, Haas TL. Metabolic Coordination of Pericyte Phenotypes: Therapeutic Implications. Front Cell Dev Biol. 2020;8:77. [CrossRef]
- Khalilgharibi N, Mao Y. To form and function: on the role of basement membrane mechanics in tissue development, homeostasis and disease. Open Biol. 2021;11(2):200360. [CrossRef]
- Halder SK, Sapkota A, Milner R. The importance of laminin at the blood-brain barrier. Neural Regen Res. 2023;18(12):2557-2563. [CrossRef]
- Boudko SP, Danylevych N, Hudson BG, Pedchenko VK. Basement membrane collagen IV: Isolation of functional domains. Methods Cell Biol. 2018;143:171-185. [CrossRef]
- Biswas S, Bachay G, Chu J, Hunter DD, Brunken WJ. Laminin-Dependent Interaction between Astrocytes and Microglia: A Role in Retinal Angiogenesis. Am J Pathol. 2017;187(9):2112-2127. [CrossRef]
- Gautam J, Zhang X, Yao Y. The role of pericytic laminin in blood brain barrier integrity maintenance. Sci Rep. 2016;6:36450. [CrossRef]
- Töpfer U, Holz A. Nidogen in development and disease. Front Cell Dev Biol. 2024;12:1380542. [CrossRef]
- Thomsen MS, Routhe LJ, Moos T. The vascular basement membrane in the healthy and pathological brain. J Cereb Blood Flow Metab. 2017;37(10):3300-3317. [CrossRef]
- Keaney J, Campbell M. The dynamic blood-brain barrier. FEBS J. 2015;282(21):4067-79. [CrossRef]
- Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood-brain barrier. Neurobiol Dis. 2010;37(1):13-25. [CrossRef]
- Töpfer, U. Basement membrane dynamics and mechanics in tissue morphogenesis. Biol Open. 2023;12(8):bio059980. [CrossRef]
- Clarke LE, Barres BA. Emerging roles of astrocytes in neural circuit development. Nat Rev Neurosci. 2013;14(5):311-21. [CrossRef]
- Fong H, Zhou B, Feng H, Luo C, Bai B, Zhang J, Wang Y. Recapitulation of Structure-Function-Regulation of Blood-Brain Barrier under (Patho)Physiological Conditions. Cells. 2024;13(3):260. [CrossRef]
- Manu DR, Slevin M, Barcutean L, Forro T, Boghitoiu T, Balasa R. Astrocyte Involvement in Blood-Brain Barrier Function: A Critical Update Highlighting Novel, Complex, Neurovascular Interactions. Int J Mol Sci. 2023;24(24):17146. [CrossRef]
- Schiera G, Di Liegro CM, Schirò G, Sorbello G, Di Liegro I. Involvement of Astrocytes in the Formation, Maintenance, and Function of the Blood-Brain Barrier. Cells. 2024;13(2):150. [CrossRef]
- Naveed M, Zhou QG, Han F. Cerebrovascular inflammation: A critical trigger for neurovascular injury? Neurochem Int. 2019;126:165-177. [CrossRef]
- Quan N. Immune-to-brain signaling: how important are the blood-brain barrier-independent pathways? Mol Neurobiol. 2008;37(2-3):142-52. [CrossRef]
- Miyata S. Glial functions in the blood-brain communication at the circumventricular organs. Front Neurosci. 2022;16:991779. [CrossRef]
- Varatharaj A, Galea I. The blood-brain barrier in systemic inflammation. Brain Behav Immun. 2017; 60:1-12. [CrossRef]
- Yang J, Ran M, Li H, Lin Y, Ma K, Yang Y, Fu X, Yang S. New insight into neurological degeneration: Inflammatory cytokines and blood-brain barrier. Front Mol Neurosci. 2022;15:1013933. [CrossRef]
- Ransohoff RM, Schafer D, Vincent A, Blachère NE, Bar-Or A. Neuroinflammation: Ways in Which the Immune System Affects the Brain. Neurotherapeutics. 2015;12(4):896-909. [CrossRef]
- Ramesh G, MacLean AG, Philipp MT. Cytokines and chemokines at the crossroads of neuroinflammation, neurodegeneration, and neuropathic pain. Mediators Inflamm. 2013;2013:480739. [CrossRef]
- Liu LR, Liu JC, Bao JS, Bai QQ, Wang GQ. Interaction of Microglia and Astrocytes in the Neurovascular Unit. Front Immunol. 2020;11:1024. [CrossRef]
- Rawat M, Nighot M, Al-Sadi R, Gupta Y, Viszwapriya D, Yochum G, Koltun W, Ma TY. IL1B Increases Intestinal Tight Junction Permeability by Up-regulation of MIR200C-3p, Which Degrades Occludin mRNA. Gastroenterology. 2020;159(4):1375-1389. [CrossRef]
- Versele R, Sevin E, Gosselet F, Fenart L, Candela P. TNF-α and IL-1β Modulate Blood-Brain Barrier Permeability and Decrease Amyloid-β Peptide Efflux in a Human Blood-Brain Barrier Model. Int J Mol Sci. 2022;23(18):10235. [CrossRef]
- Kaneko N, Kurata M, Yamamoto T, Morikawa S, Masumoto J. The role of interleukin-1 in general pathology. Inflamm Regen. 2019;39:12. [CrossRef]
- Wang Y, Jin S, Sonobe Y, Cheng Y, Horiuchi H, Parajuli B, Kawanokuchi J, Mizuno T, Takeuchi H, Suzumura A. Interleukin-1β induces blood-brain barrier disruption by downregulating Sonic hedgehog in astrocytes. PLoS One. 2014;9(10):e110024. [CrossRef]
- Gajtkó A, Bakk E, Hegedűs K, Ducza L, Holló K. IL-1β Induced Cytokine Expression by Spinal Astrocytes Can Play a Role in the Maintenance of Chronic Inflammatory Pain. Front Physiol. 2020; 11:543331. [CrossRef]
- Sjöström EO, Culot M, Leickt L, Åstrand M, Nordling E, Gosselet F, Kaiser C. Transport study of interleukin-1 inhibitors using a human in vitro model of the blood-brain barrier. Brain Behav Immun Health. 2021;16:100307. [CrossRef]
- Fahey E, Doyle SL. IL-1 Family Cytokine Regulation of Vascular Permeability and Angiogenesis. Front Immunol. 2019;10:1426. [CrossRef]
- Souza PS, Gonçalves ED, Pedroso GS, Farias HR, Junqueira SC, Marcon R, Tuon T, Cola M, Silveira PCL, Santos AR, Calixto JB, Souza CT, de Pinho RA, Dutra RC. Physical Exercise Attenuates Experimental Autoimmune Encephalomyelitis by Inhibiting Peripheral Immune Response and Blood-Brain Barrier Disruption. Mol Neurobiol. 2017;54(6):4723-4737. [CrossRef]
- Balzano T, Dadsetan S, Forteza J, Cabrera-Pastor A, Taoro-Gonzalez L, Malaguarnera M, Gil-Perotin S, Cubas-Nuñez L, Casanova B, Castro-Quintas A, Ponce-Mora A, Arenas YM, Leone P, Erceg S, Llansola M, Felipo V. Chronic hyperammonemia induces peripheral inflammation that leads to cognitive impairment in rats: Reversed by anti-TNF-α treatment. J Hepatol. 2020;73(3):582-592. [CrossRef]
- Wylezinski LS, Hawiger J. Interleukin 2 Activates Brain Microvascular Endothelial Cells Resulting in Destabilization of Adherens Junctions. J Biol Chem. 2016;291(44):22913-22923. [CrossRef]
- Waguespack PJ, Banks WA, Kastin AJ. Interleukin-2 does not cross the blood-brain barrier by a saturable transport system. Brain Res Bull. 1994;34(2):103-9. [CrossRef]
- Gao W, Li F, Zhou Z, Xu X, Wu Y, Zhou S, Yin D, Sun D, Xiong J, Jiang R, Zhang J. IL-2/Anti-IL-2 Complex Attenuates Inflammation and BBB Disruption in Mice Subjected to Traumatic Brain Injury. Front Neurol. 2017;8:281. [CrossRef]
- Yshii L, Pasciuto E, Bielefeld P, Mascali L, Lemaitre P, Marino M, Dooley J, Kouser L, Verschoren S, Lagou V, Kemps H, Gervois P, de Boer A, Burton OT, Wahis J, Verhaert J, Tareen SHK, Roca CP, Singh K, Whyte CE, Kerstens A, Callaerts-Vegh Z, Poovathingal S, Prezzemolo T, Wierda K, Dashwood A, Xie J, Van Wonterghem E, Creemers E, Aloulou M, Gsell W, Abiega O, Munck S, Vandenbroucke RE, Bronckaers A, Lemmens R, De Strooper B, Van Den Bosch L, Himmelreich U, Fitzsimons CP, Holt MG, Liston A. Astrocyte-targeted gene delivery of interleukin 2 specifically increases brain-resident regulatory T cell numbers and protects against pathological neuroinflammation. Nat Immunol. 2022; 23(6):878-891. [CrossRef]
- Furutama D, Matsuda S, Yamawaki Y, Hatano S, Okanobu A, Memida T, Oue H, Fujita T, Ouhara K, Kajiya M, Mizuno N, Kanematsu T, Tsuga K, Kurihara H. IL-6 Induced by Periodontal Inflammation Causes Neuroinflammation and Disrupts the Blood-Brain Barrier. Brain Sci. 2020;10(10):679. [CrossRef]
- Scheller J, Chalaris A, Schmidt-Arras D, Rose-John S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta. 2011;1813(5):878-88. [CrossRef]
- Banks WA, Kastin AJ, Broadwell RD. Passage of cytokines across the blood-brain barrier. Neuroimmunomodulation. 1995;2(4):241-8. [CrossRef]
- Natesh K, Bhosale D, Desai A, Chandrika G, Pujari R, Jagtap J, Chugh A, Ranade D, Shastry P. Oncostatin-M differentially regulates mesenchymal and proneural signature genes in gliomas via STAT3 signaling. Neoplasia. 2015;17(2):225-37. [CrossRef]
- Serna-Rodríguez MF, Bernal-Vega S, de la Barquera JAO, Camacho-Morales A, Pérez-Maya AA. The role of damage associated molecular pattern molecules (DAMPs) and permeability of the blood-brain barrier in depression and neuroinflammation. J Neuroimmunol. 2022;371:577951. [CrossRef]
- Zhang Y, Tian Y, Wei J, Xiang Y. Relationship of Serum IL-12 to Inflammation, Hematoma Volume, and Prognosis in Patients With Intracerebral Hemorrhage. Emerg Med Int. 2022;2022:8688413. [CrossRef]
- Andreadou M, Ingelfinger F, De Feo D, Cramer TLM, Tuzlak S, Friebel E, Schreiner B, Eede P, Schneeberger S, Geesdorf M, Ridder F, Welsh CA, Power L, Kirschenbaum D, Tyagarajan SK, Greter M, Heppner FL, Mundt S, Becher B. IL-12 sensing in neurons induces neuroprotective CNS tissue adaptation and attenuates neuroinflammation in mice. Nat Neurosci. 2023;26(10):1701-1712. [CrossRef]
- Pan W, Wu X, He Y, Hsuchou H, Huang EY, Mishra PK, Kastin AJ. Brain interleukin-15 in neuroinflammation and behavior. Neurosci Biobehav Rev. 2013;37(2):184-92. [CrossRef]
- Pan W, Hsuchou H, Yu C, Kastin AJ. Permeation of blood-borne IL15 across the blood-brain barrier and the effect of LPS. J Neurochem. 2008;106(1):313-9. [CrossRef]
- Li Z, Han J, Ren H, Ma CG, Shi FD, Liu Q, Li M. Astrocytic Interleukin-15 Reduces Pathology of Neuromyelitis Optica in Mice. Front Immunol. 2018;9:523. [CrossRef]
- Burrack KS, Huggins MA, Taras E, Dougherty P, Henzler CM, Yang R, Alter S, Jeng EK, Wong HC, Felices M, Cichocki F, Miller JS, Hart GT, Johnson AJ, Jameson SC, Hamilton SE. Interleukin-15 Complex Treatment Protects Mice from Cerebral Malaria by Inducing Interleukin-10-Producing Natural Killer Cells. Immunity. 2018;48(4):760-772.e4. [CrossRef]
- Huppert J, Closhen D, Croxford A, White R, Kulig P, Pietrowski E, Bechmann I, Becher B, Luhmann HJ, Waisman A, Kuhlmann CR. Cellular mechanisms of IL-17-induced blood-brain barrier disruption. FASEB J. 2010;24(4):1023-34. [CrossRef]
- Jung HK, Ryu HJ, Kim MJ, Kim WI, Choi HK, Choi HC, Song HK, Jo SM, Kang TC. Interleukin-18 attenuates disruption of brain-blood barrier induced by status epilepticus within the rat piriform cortex in interferon-γ independent pathway. Brain Res. 2012;1447:126-34. [CrossRef]
- Clark PR, Kim RK, Pober JS, Kluger MS. Tumor necrosis factor disrupts claudin-5 endothelial tight junction barriers in two distinct NF-κB-dependent phases. PLoS One. 2015;10(3):e0120075. [CrossRef]
- Abd-El-Basset EM, Rao MS, Alshawaf SM, Ashkanani HK, Kabli AH. Tumor necrosis factor (TNF) induces astrogliosis, microgliosis and promotes survival of cortical neurons. AIMS Neurosci. 2021;8(4):558-584. [CrossRef]
- Muhammad M. Tumor Necrosis Factor Alpha: A Major Cytokine of Brain Neuroinflammation [Internet]. Cytokines. IntechOpen; 2020. Available from:. [CrossRef]
- Chen AQ, Fang Z, Chen XL, Yang S, Zhou YF, Mao L, Xia YP, Jin HJ, Li YN, You MF, Wang XX, Lei H, He QW, Hu B. Microglia-derived TNF-α mediates endothelial necroptosis aggravating blood brain-barrier disruption after ischemic stroke. Cell Death Dis. 2019;10(7):487. [CrossRef]
- Han EC, Choi SY, Lee Y, Park JW, Hong SH, Lee HJ. Extracellular RNAs in periodontopathogenic outer membrane vesicles promote TNF-α production in human macrophages and cross the blood-brain barrier in mice. FASEB J. 2019;33(12):13412-13422. [CrossRef]
- Ng CT, Fong LY, Abdullah MNH. Interferon-gamma (IFN-γ): Reviewing its mechanisms and signaling pathways on the regulation of endothelial barrier function. Cytokine. 2023;166:156208. [CrossRef]
- Sonar SA, Shaikh S, Joshi N, Atre AN, Lal G. IFN-γ promotes transendothelial migration of CD4+ T cells across the blood-brain barrier. Immunol Cell Biol. 2017;95(9):843-853. [CrossRef]
- Youakim A, Ahdieh M. Interferon-gamma decreases barrier function in T84 cells by reducing ZO-1 levels and disrupting apical actin. Am J Physiol. 1999;276(5):G1279-88. [CrossRef]
- Rahman MT, Ghosh C, Hossain M, Linfield D, Rezaee F, Janigro D, Marchi N, van Boxel-Dezaire AHH. IFN-γ, IL-17A, or zonulin rapidly increase the permeability of the blood-brain and small intestinal epithelial barriers: Relevance for neuro-inflammatory diseases. Biochem Biophys Res Commun. 2018; 507(1-4):274-279. [CrossRef]
- Zhang W, Xiao D, Mao Q, Xia H. Role of neuroinflammation in neurodegeneration development. Signal Transduct Target Ther. 2023;8(1):267. [CrossRef]
- Rochfort KD, Cummins PM. The blood-brain barrier endothelium: a target for pro-inflammatory cytokines. Biochem Soc Trans. 2015;43(4):702-6. [CrossRef]
- Lee JI, Choi JH, Kwon TW, Jo HS, Kim DG, Ko SG, Song GJ, Cho IH. Neuroprotective effects of bornyl acetate on experimental autoimmune encephalomyelitis via anti-inflammatory effects and maintaining blood-brain-barrier integrity. Phytomedicine. 2023;112:154569. [CrossRef]
- Pawluk H, Woźniak A, Grześk G, Kołodziejska R, Kozakiewicz M, Kopkowska E, Grzechowiak E, Kozera G. The Role of Selected Pro-Inflammatory Cytokines in Pathogenesis of Ischemic Stroke. Clin Interv Aging. 2020;15:469-484. [CrossRef]
- Soltani Khaboushan A, Yazdanpanah N, Rezaei N. Neuroinflammation and Proinflammatory Cytokines in Epileptogenesis. Mol Neurobiol. 2022;59(3):1724-1743. [CrossRef]
- Granata T, Fusco L, Matricardi S, Tozzo A, Janigro D, Nabbout R. Inflammation in pediatric epilepsies: Update on clinical features and treatment options. Epilepsy Behav. 2022;131(Pt B):107959. [CrossRef]
- Savotchenko А. В. Blood-brain barrier disfunction and development of epileptic seizures: According to the materials of scientific report at the meeting of the Presidium of the NAS of Ukraine, December 23, 2020. Visn. Nac. Akad. Nauk Ukr. 2021;1:53–61. [CrossRef]
- Kamali AN, Zian Z, Bautista JM, Hamedifar H, Hossein-Khannazer N, Hosseinzadeh R, Yazdani R, Azizi G. The Potential Role of Pro-Inflammatory and Anti-Inflammatory Cytokines in Epilepsy Pathogenesis. Endocr Metab Immune Disord Drug Targets. 2021;21(10):1760-1774. [CrossRef]
- Martin SP, Leeman-Markowski BA. Proposed mechanisms of tau: relationships to traumatic brain injury, Alzheimer's disease, and epilepsy. Front Neurol. 2024;14:1287545. [CrossRef]
- Langerscheidt F, Wied T, Al Kabbani MA, van Eimeren T, Wunderlich G, Zempel H. Genetic forms of tauopathies: inherited causes and implications of Alzheimer's disease-like TAU pathology in primary and secondary tauopathies. J Neurol. 2024 Mar 30. [CrossRef]
- Michalicova A, Majerova P, Kovac A. Tau Protein and Its Role in Blood-Brain Barrier Dysfunction. Front Mol Neurosci. 2020;13:570045. [CrossRef]
- Diener HC, Gaul C, Holle-Lee D, Jürgens TP, Kraya T, Kurth T, Nägel S, Neeb L, Straube A. Kopfschmerzen – Update 2018 [Headache - an Update 2018]. Laryngorhinootologie. 2019;98(3):192-217. German. [CrossRef]
- Huang J, Ding J, Wang X, Gu C, He Y, Li Y, Fan H, Xie Q, Qi X, Wang Z, Qiu P. Transfer of neuron-derived α-synuclein to astrocytes induces neuroinflammation and blood-brain barrier damage after methamphetamine exposure: Involving the regulation of nuclear receptor-associated protein 1. Brain Behav Immun. 2022;106:247-261. [CrossRef]
- Jayanthi S, Daiwile AP, Cadet JL. Neurotoxicity of methamphetamine: Main effects and mechanisms. Exp Neurol. 2021;344:113795. [CrossRef]
- Zareifopoulos N, Skaltsa M, Dimitriou A, Karveli M, Efthimiou P, Lagadinou M, Tsigkou A, Velissaris D. Converging dopaminergic neurotoxicity mechanisms of antipsychotics, methamphetamine and levodopa. Eur Rev Med Pharmacol Sci. 2021;25(13):4514-4519. [CrossRef]
- Thangam EB, Jemima EA, Singh H, Baig MS, Khan M, Mathias CB, Church MK, Saluja R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front Immunol. 2018;9:1873. [CrossRef]
- van der Vorst EP, Döring Y, Weber C. Chemokines. Arterioscler Thromb Vasc Biol. 2015;35(11):e52-6. [CrossRef]
- Miller MC, Mayo KH. Chemokines from a Structural Perspective. Int J Mol Sci. 2017;18(10):2088. [CrossRef]
- Liu W, Jiang L, Bian C, Liang Y, Xing R, Yishakea M, Dong J. Role of CX3CL1 in Diseases. Arch Immunol Ther Exp (Warsz). 2016;64(5):371-83. [CrossRef]
- Vérité J, Janet T, Chassaing D, Fauconneau B, Rabeony H, Page G. Longitudinal chemokine profile expression in a blood-brain barrier model from Alzheimer transgenic versus wild-type mice. J Neuroinflammation. 2018;15(1):182. [CrossRef]
- Bachstetter AD, Morganti JM, Jernberg J, Schlunk A, Mitchell SH, Brewster KW, Hudson CE, Cole MJ, Harrison JK, Bickford PC, Gemma C. Fractalkine and CX 3 CR1 regulate hippocampal neurogenesis in adult and aged rats. Neurobiol Aging. 2011;32(11):2030-44. [CrossRef]
- Rogers JT, Morganti JM, Bachstetter AD, Hudson CE, Peters MM, Grimmig BA, Weeber EJ, Bickford PC, Gemma C. CX3CR1 deficiency leads to impairment of hippocampal cognitive function and synaptic plasticity. J Neurosci. 2011;31(45):16241-50. [CrossRef]
- Williams JL, Holman DW, Klein RS. Chemokines in the balance: maintenance of homeostasis and protection at CNS barriers. Front Cell Neurosci. 2014;8:154. [CrossRef]
- Legler DF, Thelen M. Chemokines: Chemistry, Biochemistry and Biological Function. Chimia (Aarau). 2016;70(12):856-859. [CrossRef]
- Ancuta P, Moses A, Gabuzda D. Transendothelial migration of CD16+ monocytes in response to fractalkine under constitutive and inflammatory conditions. Immunobiology. 2004;209(1-2):11-20. [CrossRef]
- Bertin J, Jalaguier P, Barat C, Roy MA, Tremblay MJ. Exposure of human astrocytes to leukotriene C4 promotes a CX3CL1/fractalkine-mediated transmigration of HIV-1-infected CD4⁺ T cells across an in vitro blood-brain barrier model. Virology. 2014;454-455:128-38. [CrossRef]
- Curtaz CJ, Schmitt C, Herbert SL, Feldheim J, Schlegel N, Gosselet F, Hagemann C, Roewer N, Meybohm P, Wöckel A, Burek M. Serum-derived factors of breast cancer patients with brain metastases alter permeability of a human blood-brain barrier model. Fluids Barriers CNS. 2020;17(1): 31. [CrossRef]
- Bañuelos-Cabrera I, Valle-Dorado MG, Aldana BI, Orozco-Suárez SA, Rocha L. Role of histaminergic system in blood-brain barrier dysfunction associated with neurological disorders. Arch Med Res. 2014;45(8):677-86. [CrossRef]
- Alstadhaug KB. Histamine in migraine and brain. Headache. 2014;54(2):246-59. [CrossRef]
- Abbott NJ. Inflammatory mediators and modulation of blood-brain barrier permeability. Cell Mol Neurobiol. 2000;20(2):131-47. [CrossRef]
- Sharma HS, Vannemreddy P, Patnaik R, Patnaik S, Mohanty S. Histamine receptors influence blood-spinal cord barrier permeability, edema formation, and spinal cord blood flow following trauma to the rat spinal cord. Acta Neurochir Suppl. 2006;96:316-21. [CrossRef]
- Wang Z, Cai XJ, Qin J, Xie FJ, Han N, Lu HY. The role of histamine in opening blood-tumor barrier. Oncotarget. 2016;7(21):31299-310. [CrossRef]
- Lu C, Diehl SA, Noubade R, Ledoux J, Nelson MT, Spach K, Zachary JF, Blankenhorn EP, Teuscher C. Endothelial histamine H1 receptor signaling reduces blood-brain barrier permeability and susceptibility to autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. 2010;107(44):18967-72. [CrossRef]
- Scammell TE, Jackson AC, Franks NP, Wisden W, Dauvilliers Y. Histamine: neural circuits and new medications. Sleep. 2019;42(1):zsy183. [CrossRef]
- Brown RE, Stevens DR, Haas HL. The physiology of brain histamine. Prog Neurobiol. 2001;63(6): 637-72. [CrossRef]
- Saxton RA, Glassman CR, Garcia KC. Emerging principles of cytokine pharmacology and therapeutics. Nat Rev Drug Discov. 2023;22(1):21-37. [CrossRef]
- Holder PG, Lim SA, Huang CS, Sharma P, Dagdas YS, Bulutoglu B, Sockolosky JT. Engineering interferons and interleukins for cancer immunotherapy. Adv Drug Deliv Rev. 2022;182:114112. [CrossRef]
- Pignataro G, Cataldi M, Taglialatela M. Neurological risks and benefits of cytokine-based treatments in coronavirus disease 2019: from preclinical to clinical evidence. Br J Pharmacol. 2022; 179(10):2149-2174. [CrossRef]
- Ge Y, Wu J, Zhang L, Huang N, Luo Y. A New Strategy for the Regulation of Neuroinflammation: Exosomes Derived from Mesenchymal Stem Cells. Cell Mol Neurobiol. 2024;44(1):24. [CrossRef]
- Klein RS, Hunter CA. Protective and Pathological Immunity during Central Nervous System Infections. Immunity. 2017;46(6):891-909. [CrossRef]
- Dobson R, Giovannoni G. Multiple sclerosis - a review. Eur J Neurol. 2019;26(1):27-40. [CrossRef]
- Shahrizaila N, Lehmann HC, Kuwabara S. Guillain-Barré syndrome. Lancet. 2021;397(10280):1214-1228. [CrossRef]
- Gilhus NE, Verschuuren JJ. Myasthenia gravis: subgroup classification and therapeutic strategies. Lancet Neurol. 2015;14(10):1023-36. [CrossRef]
- Younger DS. The Blood-Brain Barrier: Implications for Vasculitis. Neurol Clin. 2019;37(2):235-248. [CrossRef]
- Ortiz GG, Pacheco-Moisés FP, Macías-Islas MÁ, Flores-Alvarado LJ, Mireles-Ramírez MA, González-Renovato ED, Hernández-Navarro VE, Sánchez-López AL, Alatorre-Jiménez MA. Role of the blood-brain barrier in multiple sclerosis. Arch Med Res. 2014;45(8):687-97. [CrossRef]
- Khan AW, Farooq M, Hwang MJ, Haseeb M, Choi S. Autoimmune Neuroinflammatory Diseases: Role of Interleukins. Int J Mol Sci. 2023;24(9):7960. [CrossRef]
- Takeshita Y, Fujikawa S, Serizawa K, Fujisawa M, Matsuo K, Nemoto J, Shimizu F, Sano Y, Tomizawa-Shinohara H, Miyake S, Ransohoff RM, Kanda T. New BBB Model Reveals That IL-6 Blockade Suppressed the BBB Disorder, Preventing Onset of NMOSD. Neurol Neuroimmunol Neuroinflamm. 2021;8(6):e1076. [CrossRef]
- Corti A, Calimeri T, Curnis F, Ferreri AJM. Targeting the Blood-Brain Tumor Barrier with Tumor Necrosis Factor-α. Pharmaceutics. 2022;14(7):1414. [CrossRef]
- Hurtado-Alvarado G, Becerril-Villanueva E, Contis-Montes de Oca A, Domínguez-Salazar E, Salinas-Jazmín N, Pérez-Tapia SM, Pavon L, Velázquez-Moctezuma J, Gómez-González B. The yin/yang of inflammatory status: Blood-brain barrier regulation during sleep. Brain Behav Immun. 2018;69: 154-166. [CrossRef]
- Zhang L, Zhou L, Bao L, Liu J, Zhu H, Lv Q, Liu R, Chen W, Tong W, Wei Q, Xu Y, Deng W, Gao H, Xue J, Song Z, Yu P, Han Y, Zhang Y, Sun X, Yu X, Qin C. SARS-CoV-2 crosses the blood-brain barrier accompanied with basement membrane disruption without tight junctions alteration. Signal Transduct Target Ther. 2021;6(1):337. [CrossRef]
- Montazersaheb S, Hosseiniyan Khatibi SM, Hejazi MS, Tarhriz V, Farjami A, Ghasemian Sorbeni F, Farahzadi R, Ghasemnejad T. COVID-19 infection: an overview on cytokine storm and related interventions. Virol J. 2022;19(1):92. [CrossRef]
- Soy M, Keser G, Atagündüz P, Tabak F, Atagündüz I, Kayhan S. Cytokine storm in COVID-19: pathogenesis and overview of anti-inflammatory agents used in treatment. Clin Rheumatol. 2020; 39(7):2085-2094. [CrossRef]
- Hernández-Parra H, Reyes-Hernández OD, Figueroa-González G, González-Del Carmen M, González-Torres M, Peña-Corona SI, Florán B, Cortés H, Leyva-Gómez G. Alteration of the blood-brain barrier by COVID-19 and its implication in the permeation of drugs into the brain. Front Cell Neurosci. 2023;17:1125109. [CrossRef]
- Chen H, Tang X, Li J, Hu B, Yang W, Zhan M, Ma T, Xu S. IL-17 crosses the blood-brain barrier to trigger neuroinflammation: a novel mechanism in nitroglycerin-induced chronic migraine. J Headache Pain. 2022;23(1):1. [CrossRef]
Figure 1.
Schematic representation of the blood-brain barrier (BBB) in cross section illustrating the BBB's cellular structures [28,31]. From the capillary lumen: brain microvascular endothelial cells (BMECs) surrounded by pericyte located within the parenchymal basement membrane. The presence of tight junctions between endothelial cells maintains the integrity and required level of permeability of the BBB. The surface of the basement membrane is covered by the astrocytic end-feets. Microglia and neuronal synapse endings are located in the extracellular matrix surrounding the BBB from the side of.
Figure 1.
Schematic representation of the blood-brain barrier (BBB) in cross section illustrating the BBB's cellular structures [28,31]. From the capillary lumen: brain microvascular endothelial cells (BMECs) surrounded by pericyte located within the parenchymal basement membrane. The presence of tight junctions between endothelial cells maintains the integrity and required level of permeability of the BBB. The surface of the basement membrane is covered by the astrocytic end-feets. Microglia and neuronal synapse endings are located in the extracellular matrix surrounding the BBB from the side of.
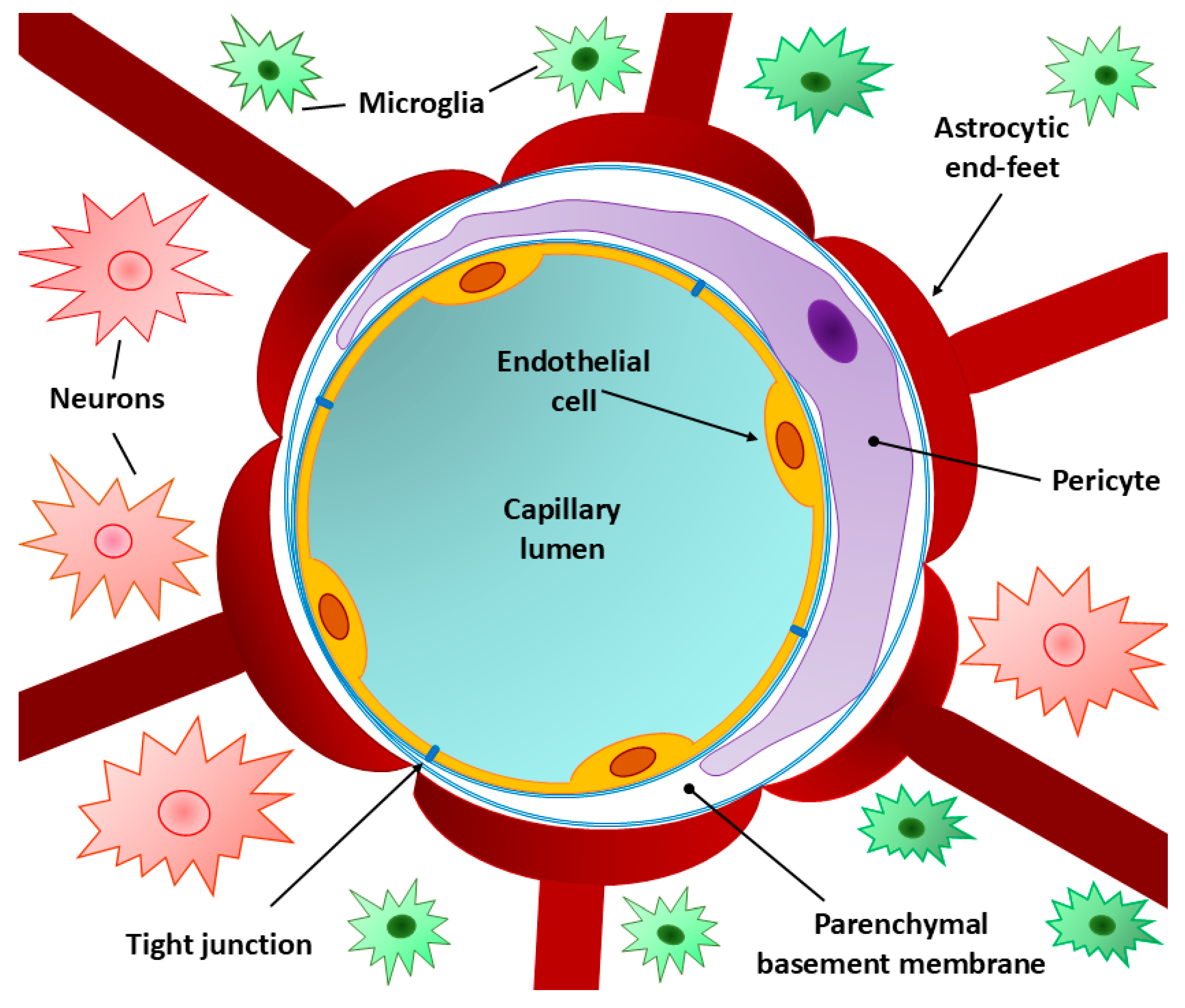
Figure 2.
Schematic representation of the tight contact between adjacent endothelial cells [38]. Tight junction (TJ) is formed by joining of major cellular membrane (CM) proteins of contacting cells, such as claudins, occludins and junctional adhesion molecules (JAMs). From the side of the cytoplasm of cells forming TJs, the participation of several additional proteins is necessary, which stabilize the connection by binding to elements of the cytoskeleton (actin) of endotheliocytes. These cytoplasmic proteins include zonula occludens-1, -2, and -3 (ZO-1, ZO-2, and ZO-3, respectively), cingulin, vinculin, catenins (α,β, and γ). The figure also shows a much weaker adherent junction (AJ) located below the TJ, which connects the adjoning plasma membranes with the electron-dense fibrillary molecules known as vascular endothelial (VE)-cadherins.
Figure 2.
Schematic representation of the tight contact between adjacent endothelial cells [38]. Tight junction (TJ) is formed by joining of major cellular membrane (CM) proteins of contacting cells, such as claudins, occludins and junctional adhesion molecules (JAMs). From the side of the cytoplasm of cells forming TJs, the participation of several additional proteins is necessary, which stabilize the connection by binding to elements of the cytoskeleton (actin) of endotheliocytes. These cytoplasmic proteins include zonula occludens-1, -2, and -3 (ZO-1, ZO-2, and ZO-3, respectively), cingulin, vinculin, catenins (α,β, and γ). The figure also shows a much weaker adherent junction (AJ) located below the TJ, which connects the adjoning plasma membranes with the electron-dense fibrillary molecules known as vascular endothelial (VE)-cadherins.
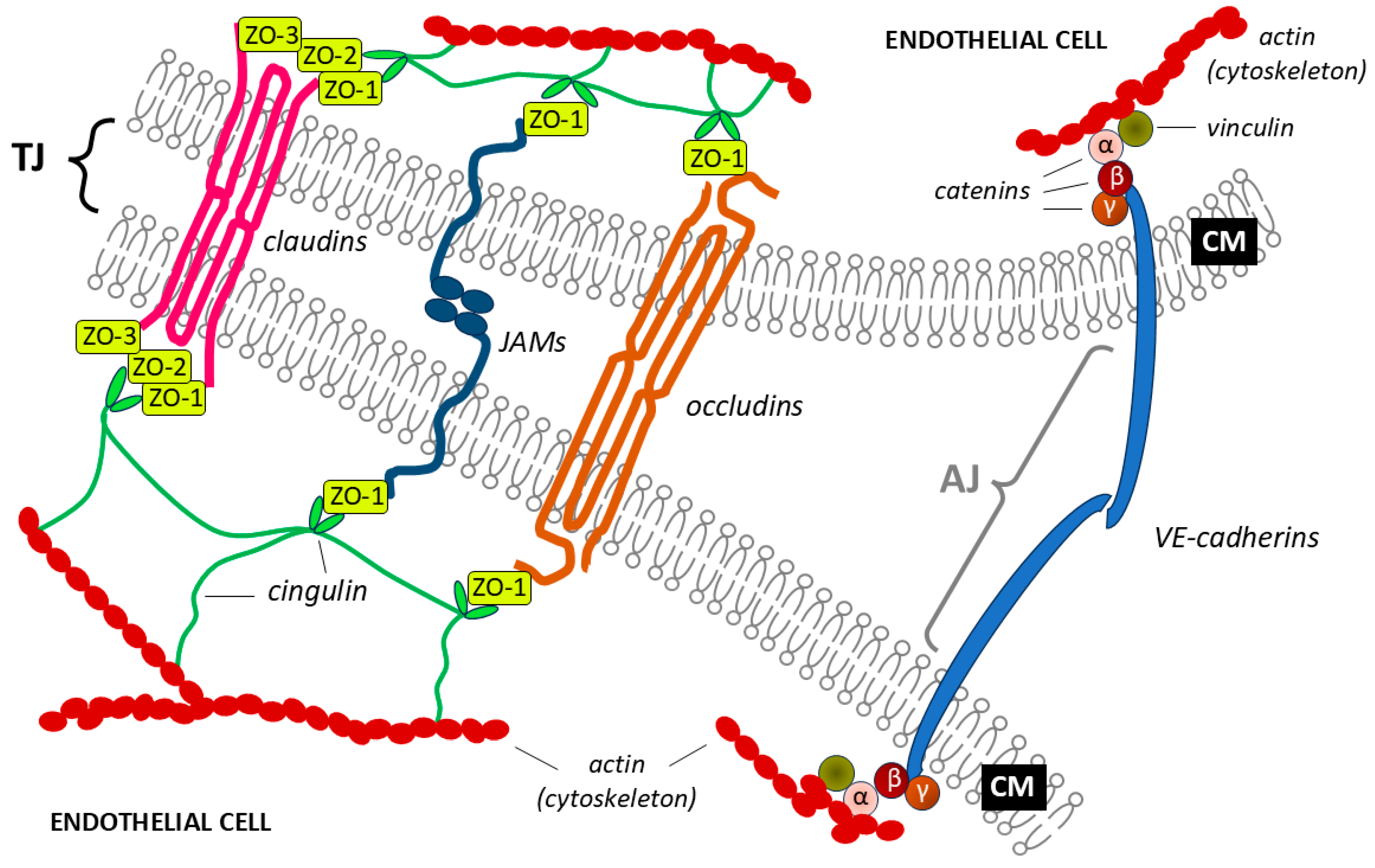
Figure 3.
The endogenous pathway of proinflammatory cytokines through the blood-brain barrier (BBB) [11,17]. Neuroinflammation may lead to loss of BBB integrity due to activation of brain-resident immune cells (marked with red asterisks), which then leads to damage of surrounding neurons and glial cells (marked with blue asterisks). The cellular component of neuroinflammation (summarized in the box on the left) is accompanied by a soluble component associated with the overproduction of pro-inflammatory cytokines and other inflammatory mediators (listed in the box on the right) in inflammation-activated cells [e.g., in polarized towards a pro-inflammatory (M1) phenotype microglia]. Decreased expression of membrane proteins (e.g., claudin-5) and cytoplasmic proteins [e.g., zonula occludens-1 (ZO-1)] is accompanied by impaired function of tight junctions. Combined with an increase in the expression of adhesion molecules on the surface of the brain microvascular endothelial cells (BMECs), an increase in BBB permeability may result in the influx of T cells from the bloodstream into the central nervous system (CNS). Other abbreviations: ROS – reactive oxygen species, MMPs – matrix metalloproteases.In response to pathological conditions in the CNS, the immune system uses inflammation as one of its tools to combat the pathology [89]. The inflammatory response is mobilized by proinflammatory cytokines produced by macrophages, leukocytes, neutrophils or other cells, depending on the pathological process [90]. For example, in response to stroke or lipopolysaccharide, the proinflammatory cytokines IL-1 and TNF-α are produced by microglia, which are activated in a proinflammatory phenotype during neuroinflammation to induce the formation of reactive astrocytes that acquire the same proinflammatory phenotype [91]. The effects of proinflammatory cytokines on BBB disorders are shown in Table 1.
Figure 3.
The endogenous pathway of proinflammatory cytokines through the blood-brain barrier (BBB) [11,17]. Neuroinflammation may lead to loss of BBB integrity due to activation of brain-resident immune cells (marked with red asterisks), which then leads to damage of surrounding neurons and glial cells (marked with blue asterisks). The cellular component of neuroinflammation (summarized in the box on the left) is accompanied by a soluble component associated with the overproduction of pro-inflammatory cytokines and other inflammatory mediators (listed in the box on the right) in inflammation-activated cells [e.g., in polarized towards a pro-inflammatory (M1) phenotype microglia]. Decreased expression of membrane proteins (e.g., claudin-5) and cytoplasmic proteins [e.g., zonula occludens-1 (ZO-1)] is accompanied by impaired function of tight junctions. Combined with an increase in the expression of adhesion molecules on the surface of the brain microvascular endothelial cells (BMECs), an increase in BBB permeability may result in the influx of T cells from the bloodstream into the central nervous system (CNS). Other abbreviations: ROS – reactive oxygen species, MMPs – matrix metalloproteases.In response to pathological conditions in the CNS, the immune system uses inflammation as one of its tools to combat the pathology [89]. The inflammatory response is mobilized by proinflammatory cytokines produced by macrophages, leukocytes, neutrophils or other cells, depending on the pathological process [90]. For example, in response to stroke or lipopolysaccharide, the proinflammatory cytokines IL-1 and TNF-α are produced by microglia, which are activated in a proinflammatory phenotype during neuroinflammation to induce the formation of reactive astrocytes that acquire the same proinflammatory phenotype [91]. The effects of proinflammatory cytokines on BBB disorders are shown in Table 1.
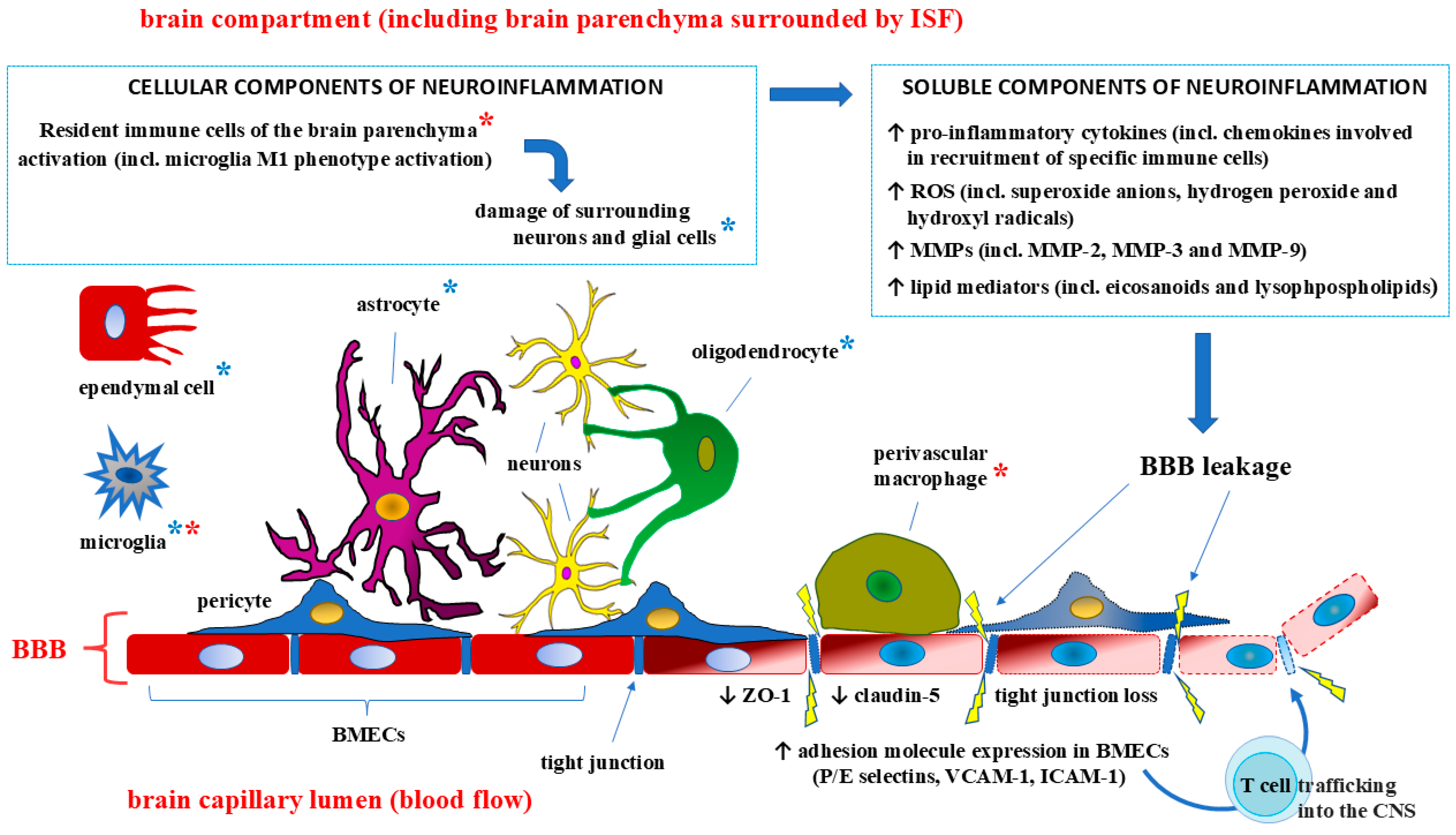

Table 1.
Influence of proinflammatory cytokines on the blood-brain barrier (BBB) structure.
| Pro-inflammatory cytokine | Impact on the structural elements of the BBB | References |
|---|---|---|
| IL-1 |
|
[92,93] [94] [95] [96] [93,97,98] |
| IL-1β |
|
[95,99] [95] [96,100] |
| IL-2 |
|
[101] [102] [103] [104] |
| IL-6 |
|
[105] [88] [106] [107,108] |
| IL-12 |
|
[109,110] [111] |
| IL-15 |
|
[112,113] [114] [115] |
| IL-17 |
|
[116] |
| IL-18 |
|
[117] |
| TNF-α |
|
[118] [119] [120] [121,122] |
| IFN-γ |
|
[123] [124] [125,126] |
Abbreviations: GBM – glioblastoma multiforme; IFN-γ – interferon gamma; IL-1 – interleukin 1; IL-2 – interleukin 2; IL-6 – interleukin 6; IL-12 – interleukin 12; IL-15 – interleukin 15; IL-17 – interleukin 17; IL-18 – interleukin 18; TNF-α – tumor necrosis factor alpha; ZO-1 – zonula occludens-1. Source: compiled by the authors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



