
Submitted:
19 November 2024
Posted:
21 November 2024
You are already at the latest version
Abstract
A growing body of evidence indicates a link between circulating neurotoxic lipids and the development of chronic neuroinflammatory diseases in the peripheral and central nervous systems. Therefore, strategies to modify circulating lipid profiles may complement the management of neuroinflammatory diseases, including neuropathic pain. In a previous study, we observed a sig-nificant shift in the metabolomic profile of patients' plasma with symptoms of painful diabetic neuropathy (pDN) following three months of docosahexaenoic acid (DHA)-rich supplementation, leading to improved pDN symptoms. However, it is important to identify the specific lipid mediators responsible for this therapeutic effect and elucidate potential mechanism(s). This study investigates whether DHA-rich supplementation reduces neurotoxic lipid mediators associated with pDN in individuals with type 2 diabetes mellitus (T2DM). Forty volunteers diagnosed with type 2 diabetes were enrolled in the "En Balance-PLUS" diabetes education study. The volunteers participated in weekly lifestyle/nutrition education and daily supplementation with 1000 mg DHA and 200 mg eicosapentaenoic acid. The Short-Form McGill Pain Questionnaire validated the clinical determination of baseline and post-intervention pain complaints. Untargeted Lipidomic analyses were conducted using blood serum collected at baseline and after three months of participation in the dietary regimen. The lipidomic data were analyzed using a non-parametric paired Wilcoxon rank-sum test and random forest analysis. ELISA further eval-uated participant serum samples to investigate associated biomarkers of necrosis (MLKL), autophagy (ATG5), and lipid chaperone protein (FABP5). Untargeted lipidomic analysis revealed that several neurotoxic-associated lipids significantly decreased after DHA-rich supplementation. Also, circulating levels of MLKL were reduced, while protein levels of ATG5 and FABP5 significantly increased. The reduction of circulating neurotoxic lipids and increase of neuroprotective lipids following DHA-rich supplementation is consistent with the reported roles of omega-3 polyunsaturated fatty acids (PUFAs) in reducing adverse symptoms associated with neuroinflammatory diseases and painful neuropathy.
Keywords:
Docosahexaenoic acid
; lipidomics
; autophagy
; painful diabetic neuropathy
1. Introduction
The treatment of painful diabetic neuropathy (pDN) remains inadequate, and it is considered a significant co-morbidity of this condition [1]. Pharmacological treatment only assists about a third of patients to achieve 50% pain relief, often complicated by side effects [2,3,4]. Until recently, there was a consensus that hyperglycemia was the primary driver of diabetic neuropathy (DN). However, growing evidence suggests that lipid substrate overload, commonly observed in T2DM, has been proposed as a significant contributing pathogenic factor to DN [1,5,6,7]. The need for novel targets in the treatment of DN stems from the 2012 Cochrane review, which indicated that intensive glycemic control only marginally improved neuropathy in multiple type 2 diabetes cohorts [8]. Furthermore, intensive glycemic control significantly increases the risk of severe hypoglycemic episodes and is associated with increased mortality in T2DM [9]. Since the publication of these findings, a directed focus on lipid mediators in the development of DN is intensifying [10,11,12].
Dorsal root ganglion (DRG) neurons and Schwann cells are the primary cells for intact sensory processing. DRG neurons depend on mitochondrial ATP production throughout the axon and rely on mitochondrial transport mechanisms to distribute mitochondria for normal nerve function . When exposing DRG neurons to elevated levels of C16:0 and C18:0 saturated fatty acids (SFAs), a marked decrease in mitochondrial trafficking is observed [14,15]. Ultimately, increased levels of SFA lead to mitochondrial depolarization and impaired mitochondrial bioenergetics, triggering DRG apoptosis [12,13]. In line with these findings, the neuronal model of nerve growth factor-differentiated pheochromocytoma cells (NGF-PC12) exposed to elevated palmitic acid levels demonstrated significant alterations in mitochondrial transmembrane potential, alongside increased mRNA expression of key genes regulating cell death and survival.[16,17]. Notably, our research showed that treating neuronal cells under lipotoxic stress with DHA reduced necroptosis activity, as evidenced by a marked decrease in Mixed Lineage Kinase Domain-Like protein (MLKL) levels, while simultaneously enhancing autophagy activity. Additionally, SFA substrate overload decreases β-oxidation, increasing available palmitoyl-CoA [18]. The increased availability of palmitoyl-CoA, in turn, raises the levels of ceramides to toxic levels, inhibiting axonal growth [19,20]. These studies indicate that an overabundance of certain SFAs impairs mitochondrial function and plays a critical role in peripheral neuropathy progression.
Omega-3 PUFAs have the potential to shield neuronal tissues from SFA-induced metabolic stress, as demonstrated in previous studies [16,21]. DHA, a well-known omega-3 PUFA, has been shown to protect and reverse palmitic acid-induced lipotoxicity in neuronal cell models by inhibiting mitochondrial membrane depolarization, a significant biochemical feature in the development of DN [16,19]. Furthermore, omega-3 PUFAs have been found to reduce nerve damage and alleviate neuropathic pain in various pain models [22,23,24,25]. Significantly, DHA effectively reduced ceramide production and associated oxidative stress [26]. These findings underscore the role of omega-3 PUFAs in counteracting key metabolic pathways associated with DN, suggesting their potential as a therapeutic intervention.
We have demonstrated that the neuroprotective effects of DHA are mediated through its interaction with FABP5, a lipid chaperone protein integral to neuronal survival and repair mechanisms [27]. FABP5 regulates the balance of bioactive lipids, mitigates oxidative stress, and enhances cellular repair following injury [28,29,30]. Specifically, it exerts antioxidant effects by reducing reactive oxygen species (ROS), which aids in nerve regeneration and functional recovery. Moreover, FABP5 exhibits ligand-dependent neuroprotective functions; for example, its interaction with DHA enhances endocannabinoid signaling and attenuates pro-inflammatory pathways, thereby reducing nociceptive sensitization and contributing to analgesic effects [27]. These findings underscore the synergistic relationship between FABP5 and DHA, highlighting a potential therapeutic mechanism for addressing neuropathic pain and neuroinflammatory conditions.
Recognizing lipid metabolism's pivotal role in DN's pathogenesis, our study takes a unique approach. We aim to identify changes in associated neurotoxic lipids in diabetic patients who report neuropathic pain symptoms before and after a dietary omega-3 PUFA supplementation. Furthermore, we sought to investigate proteins associated with DHA-induced neuroprotection (MLKL, ATG5, and FABP-5) in our human population. To our knowledge, no previous human study has investigated the global lipidomic profiles of T2DM patients reporting neuropathic pain or the effects of dietary omega-3 PUFAs supplementation on these profiles. Our approach employs a quantitative untargeted lipidomic analysis, which allows for an unbiased interpretation of the lipid mediators involved in DN and the impact of dietary omega-3 PUFA supplementation on the resolution of these neurotoxic lipids.
2. Materials and Methods
2.1. Study Design and Population
The En Balance-Plus study, a longitudinal single-arm study, was conducted in Loma Linda, California, to assess the effects of nutrition and diabetes education on type 2 diabetes in a Latino population, primarily Mexican Americans. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Loma Linda University Institutional Review Board (LLUIRB# 5110318) on 11/2014 [31,32]; informed consent was obtained from all participants prior to their involvement in the study.
2.2. Intervention
The En Balance-Plus study evaluated the effects of DHA-enriched dietary supplementation on participants' lipidomic profiles. Over a three-month period, participants received daily capsules containing 1000 mg of DHA and 200 mg of eicosapentaenoic acid (EPA) while attending weekly Spanish-language diabetes education sessions.
2.3. Data and Study Variables
We have previously described methods in detail [31,32]. Briefly, samples were collected for all participants at baseline and three months post-intervention, including fasting blood plasma and serum samples. Serum samples were sent to Metabolon for complex lipidomic analysis. Complex lipids were extracted from serum samples using a modified version of Lofgren's procedure [33].
Participants’ diets, medication use, and exercise habits were closely monitored throughout the three-month intervention to ensure that any changes identified in the untargeted lipidomic analysis could be attributed to the dietary supplementation. A paired statistical analysis method was utilized, comparing each participant’s post-intervention values to their baseline measurements [32], effectively minimizing the impact of external variables on the results.
2.4. Measurement of Neuropathic Pain Symptoms
Neuropathic pain symptoms were monitored as previously described [32]. At baseline, 26 participants reported neuropathic pain symptoms, with 14 classified as having low pain (sensory score < 7) and 12 as having moderate-to-high pain (sensory score > 7).
2.5. Measurements of MLKL, ATG5, and EFABP5 Serum Levels by ELISA
Fasting serum samples of all participants were collected at baseline and three months post-intervention. Specifically, Mixed lineage kinase domain-like (MLKL), Autophagy protein 5 (ATG5), and Fatty acid binding protein 5 (FABP5) serum levels were measured using a commercially available enzyme immunoassay according to manufacturers' instructions (MBS9300811, MBS2602759, and MBS2613697, MyBioSource Inc.).
2.6. Statistical Analysis
An occupational threshold of 70% was applied to lipidomic data, requiring complex lipids present in at least 70% of the participants to be considered for analysis. Statistical analyses were performed using Prism 9 (GraphPad Software, San Diego, CA, USA) and MetaboAnalyst 6.0 (accessed on April 19, 2024) [27]. A paired-sample t-test was used to analyze normally distributed continuous variables, while non-normally distributed continuous variables were evaluated using Wilcoxon signed-rank tests to determine significant differences between baseline and three months post-supplementation. Normality was assessed using the Kolmogorov–Smirnov and Shapiro–Wilk tests, and outliers were identified using Grubbs' test via GraphPad (www.graphpad.com, accessed on April 19, 2024). To control for false discovery rates, p-values were adjusted and reported as q-values. Data are presented as mean ± SD, with statistical significance set at α = 0.05 unless otherwise noted.
3. Results
3.1. Lipidomic Data Analysis
Metabolon TrueMass® complex lipid panel tests for several lipid species, concentrations, and compositions. For our analysis, we focused on fatty acid concentration [FA]. The panel consists of 283 known fatty acids. Of those, only 81 transformed [FA] significantly changed (p < 0.05) at three months on the matched pairs t-test (Figure 1). Specifically, the levels of 22 transformed [FA] were significantly increased (p < 0.05), while 59 were significantly decreased (p < 0.05) (Table 1). A summary of all significant [FA] is listed in Table 2. Interestingly, several known SFAs, MAG 16:0 and 18:0, LPC 16:0 and 18:0, LPE 18:0, and CER 16:0, considered to be neurotoxic and involved during neuropathy pathogenesis [10,34,35] significantly (p < 0.01) decrease post-DHA-rich dietary supplementation.
Next, to assess the significance of fatty acid (FA) features associated with DHA-rich supplementation, we conducted random forest (RF) classification using fatty acid concentrations to predict sample classification at baseline or post-intervention. The classification of serum samples before and after DHA-rich supplementation achieved approximately 96% accuracy, significantly higher than the 50% accuracy expected by random chance. Fatty acid importance in group classification was quantified as the mean decrease in accuracy, as shown in Figure 2. The top 15 fatty acids contributing to group differentiation were predominantly enriched with DHA lipid derivatives. Notably, palmitic and stearic acid derivatives, MAG[FA16:0] and MAG[FA18:0], were among the most important features for group separation.
3.2. Assessment of Associated Markers of Painful Diabetic Neuropathy Levels by ELISA
3.2.1. Measurement of MLKL Serum levels by ELISA
In a previous study, our group demonstrated that SFA overload reduces the viability of fully differentiated pheochromocytoma clone 12 (PC12) cells by necrosis, an established in vitro model for studying the cellular mechanisms associated with DN [36,37]. However, we rescued the cells in the presence of SFA overload by co-treating the cells with DHA [16,17,38]. To determine if this mechanism holds in humans, we measured the serum levels of MLKL before and after DHA-rich supplementation in a human cohort. Figure 3 shows a significant decrease in MLKL serum levels post DHA-rich supplementation in the overall group (regardless of pain status) and in the mod-high pain group, as defined previously [31].
3.2.2. Measurement of ATG5 Serum Levels by ELISA
Growing evidence suggests that critical pathways for cell maintenance, such as autophagy, play a crucial role in preventing and reversing neurodegenerative diseases, including DN [39,40,41,42]. Our previous research has shown that an overload of SFA can lead to neuronal cell toxicity, which can be alleviated by promoting autophagy [17]. For instance, we found that DHA can activate autophagy in the presence of SFA overload, and if autophagy is chemically inhibited, DHA's ability to protect against SFA toxicity is diminished. Based on our previous findings linking lipids, autophagy, and DN, we measured the serum levels of ATG5 before and after DHA-rich supplementation in a human cohort. Interestingly, we discovered that ATG5 levels only increased in the moderate-to-high pain group following DHA-rich supplementation, as depicted in Figure 4.
3.2.3. Measurement of FABP5 Serum Levels by ELISA
Our research group has shown that FABP5 plays an important role in neuronal development and neuronal injury and has antioxidant capacity [29,30,43,44]. We have also found that FABP5 is involved in locomotor recovery after spinal cord injury [27]. Additionally, FABP5 is essential for delivering neuroprotective fatty acids to the central nervous system, and a lack of FABP5 contributes to neuroinflammation [45,46]. Based on our experience with FABP5 and its neuroprotective effects in connection with DHA, we measured serum levels of FABP5 before and after DHA-rich supplementation in our cohort. As anticipated, we observed a significant increase in FABP5 serum levels after DHA-rich supplementation in the overall group (regardless of pain status) and the moderate to high pain group, as shown in Figure 5.
4. Discussion
4.1. Lipidomic implications of a DHA-rich supplementation in pDN
In addition to hyperglycemia, dyslipidemia is increasingly recognized as a significant contributing factor to neuropathy, especially in type 2 diabetes [1]. Moreover, many studies have demonstrated the harmful effects of SFA substrate overload on the nervous system [16,21,47,48]. Specifically, cellular studies showed that high concentrations of palmitic acid impaired sensory neuron mitochondrial trafficking and altered mitochondrial energy production [14]. Similarly, SFA overload in Schwann cells results in robust ER stress response, mitochondrial dysfunction, and increased oxidative stress [49], supporting the importance of dysregulated lipid metabolism in the pathogenesis of DN.
Specific palmitic and stearic acid derivatives are associated with the development of neuropathic pain. Long chain saturated fatty acid (LCSFA) containing LPC and LPE, when in excess, increases neuronal reactive oxygen species in the dorsal root ganglion, causing altered neuronal activity and signaling [34,35]. LCSFA containing LPC and LPE have also been identified as pain biomarkers [50]. Our study showed that participants with T2DM who experienced pDN had decreased levels of C16:0 neurotoxic derivatives containing MAG (C16:0, C18:0), LPC (C16:0, C18:0) and LPE (C18:0) after three months of dietary supplementation. Particularly noteworthy was the observation that the overall level of serum C16:0 did not significantly change, indicating that the supplementation had a targeted effect on C16:0-associated metabolites. Furthermore, the fact that the serum C16:0 remained at baseline levels aligns with the participant's report of not changing their diet aside from taking the provided DHA-rich supplementation. Lastly, our study is the first to report a significant decrease in associated neurotoxic LCSFA containing LPC and LPE after dietary omega-3 supplementation in patients with T2DM who reported neuropathic pain symptoms.
On the other hand, DHA-containing PC, LPE, and LPC have been reported to have neuroprotective properties [51]. Using RF analysis, our study demonstrates that DHA-PC is the most important complex lipid for distinguishing between baseline and post-intervention groups, indicating that DHA-PC levels could be a potential biomarker for pDN. Thus, the data suggest that individuals with T2DM and low levels of DHA-PC may be at a higher risk of developing pDN than those with higher levels of serum DHA-PC. Furthermore, we observed a significant increase in DHA-containing PC, LPC, and LPE serum levels after DHA-rich supplementation. Interestingly, DHA-containing PCs inhibit IL-6 signaling [51], an essential pro-inflammatory molecule involved in the abnormal activation of the NF-κB loop in DN [19].
Overall, we showed that complex lipid analysis identified key LCSFA derivatives of C16:0 and C18:0 involved in the pathogenesis of DN. More importantly, these findings indicate that dietary supplementation rich in DHA can reverse neurotoxic lipid profiles to a more neuro-regenerative composition. Specifically, this supplementation decreases LCSFA LPC and LPE levels while increasing DHA-containing PC, LPE, and LPC. As DHA supplementation is known to be safe for patients with T2DM, further studies are needed to define its potential role in the treatment of pDN.
4.2. Implicaions of Associated Markers of Neuronal Protection after DHA-Rich Dietary Intervention
Current pharmacotherapies fail to target the underlying nerve damage associated with pDN and expose the user to significant side effects [52]. This lack of effective therapies for pDN has prompted investigators to explore targeting central mechanisms of neuronal homeostasis to prevent/reverse the development of pDN. An emerging target is autophagy, an established pathway of critical importance for neuronal health [40,41,53,54]. Specifically, autophagy is an attractive target since it plays a critical role in regulating neuroinflammation and neuronal survival. Animal studies have recently demonstrated that upregulated autophagic activity can directly alleviate neuropathic pain [55,56].
Metabolic states such as T2DM are often marked by lipid substrate overload. This metabolic imbalance is significant, as excessive lipid accumulation is linked to reduced autophagic turnover [57,58]. Our previous research in preclinical models demonstrated that DHA protects SFA overload, with autophagy playing a central role in DHA-mediated neuronal protection from lipotoxicity [17,29]. In the current study, we observed that participants with moderate to high pain had significantly increased serum ATG5 levels after three months of DHA-rich supplementation. These findings reinforce the potential of targeting central mechanisms of neuronal homeostasis, such as autophagy, as an effective strategy to improve clinical outcomes in patients with pDN.
In our intervention, we focused on increasing omega-3 PUFAs in our participants' diets; therefore, we measured the levels of FABP5, a fatty acid-binding protein involved in neuronal protection and fatty acid transport. Previously, our group and others showed that FABP5 protects neuronal models from lipotoxicity, increases after omega-3 dietary supplementation and deficiency in FABP5 is associated with neuronal fragility [16,27,29,46,59,60]. In the present study, FABP5 serum levels increased in the participants after the dietary DHA-rich supplementation. This finding, along with our previous reports [31], supports the role of FABP5 in protecting the nervous system from lipotoxicity, oxidative stress, and inflammatory damage, pathways associated with neurodegeneration and pain [27,29,32]. As FABP5 and autophagy processes appear to be central to DHA action, further study is needed on the potential FABP5 involvement in autophagic activity during DHA regulation.
4.3. Study Limitations and Strengths
We have previously addressed the limitations of this study in detail [31,32]. In summary, the analysis lacks a traditional control group, and participants engaged in an interactive, supportive environment, which may have introduced a Hawthorne or placebo effect on the reported SF-MPQ pain scores.
Confounding factors often complicate the evaluation of outcomes in community-based interventions, particularly when reliable quantitative measures are limited or the inclusion of a control group conflicts with the study's objectives [61]. To address these challenges, we employed a paired analysis approach, using each participant as their own control and assessing changes relative to baseline values [62]. A key strength of this study lies in its use of lipidomic profiling, which offered detailed insights into the impact of DHA-rich supplementation. The untargeted lipidomic analysis identified significant changes in several lipids associated with DHA metabolism, further highlighting the intervention's biochemical effects.
5. Conclusion
In conclusion, this study highlights the potential of docosahexaenoic acid (DHA)-rich supplementation to modify circulating lipid profiles and reduce neurotoxic lipid mediators associated with painful diabetic neuropathy (pDN) in individuals with type 2 diabetes mellitus (T2DM). The intervention not only significantly decreased neurotoxic lipids but also increased levels of neuroprotective markers, including ATG5 and FABP5, while reducing MLKL, a biomarker of necrosis. These findings align with the established roles of omega-3 polyunsaturated fatty acids in mitigating neuroinflammatory processes and alleviating symptoms of neuropathic pain. This research provides valuable insights into the mechanisms underlying DHA's therapeutic effects and underscores the potential of lipid-targeted interventions as complementary strategies for managing chronic neuroinflammatory diseases and neuropathic pain.
Author Contributions
Conceptualization, M.D.L.; methodology, A.M.D., and M.D.L.; software, A.M.D.; validation, A.M.D., F.Z., and M.D.L.; formal analysis, A.M.D.; investigation, A.M.D., and M.D.L.; resources, M.D.L.; data curation, A.M.D.; writing—original draft preparation, A.M.D., F.Z., and M.D.L.; writing—review and editing, A.M.D., F.Z, and M.D.L.; visualization, A.M.D.; supervision, M.D.L.; project administration, M.D.L.; funding acquisition, M.D.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by NIH grants 5P20MD006988 and 5R25GM060507.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of Loma Linda University.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study (LLU-IRB# 5110318).
Data Availability Statement
Data supporting reported results can be found in a dataset generated during the study. Additional information is available upon request.
Acknowledgments
The authors thank all investigators, study teams, and volunteers for participating in this study. We also would like to acknowledge the contributions of late Dr. Zaida Cordero-MacIntyre, Dr. Anthony Firek, and Dr. Larry Beeson, PhD for original contributions in in starting the En Balance-Plus diabetic educational program.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Callaghan, B. C.; Gallagher, G.; Fridman, V.; Feldman, E. L. , Diabetic neuropathy: what does the future hold? Diabetologia 2020, 63(5), 891–897. [Google Scholar] [CrossRef] [PubMed]
- Sloan, G.; Shillo, P.; Selvarajah, D.; Wu, J.; Wilkinson, I. D.; Tracey, I.; Anand, P.; Tesfaye, S. , A new look at painful diabetic neuropathy. Diabetes Res Clin Pract 2018, 144, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Tang, H. Y.; Jiang, A. J.; Ma, J. L.; Wang, F. J.; Shen, G. M. , Understanding the Signaling Pathways Related to the Mechanism and Treatment of Diabetic Peripheral Neuropathy. Endocrinology 2019, 160(9), 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T. S.; Karlsson, P.; Gylfadottir, S. S.; Andersen, S. T.; Bennett, D. L.; Tankisi, H.; Finnerup, N. B.; Terkelsen, A. J.; Khan, K.; Themistocleous, A. C.; Kristensen, A. G.; Itani, M.; Sindrup, S. H.; Andersen, H.; Charles, M.; Feldman, E. L.; Callaghan, B. C. , Painful and non-painful diabetic neuropathy, diagnostic challenges and implications for future management. Brain 2021, 144(6), 1632–1645. [Google Scholar] [CrossRef] [PubMed]
- Yorek, M. A. , The Potential Role of Fatty Acids in Treating Diabetic Neuropathy. Curr Diab Rep 2018, 18(10), 86. [Google Scholar] [CrossRef]
- Feldman, E. L.; Callaghan, B. C.; Pop-Busui, R.; Zochodne, D. W.; Wright, D. E.; Bennett, D. L.; Bril, V.; Russell, J. W.; Viswanathan, V. , Diabetic neuropathy. Nat Rev Dis Primers 2019, 5(1), 41. [Google Scholar] [CrossRef]
- Callaghan, B. C.; Gao, L.; Li, Y.; Zhou, X.; Reynolds, E.; Banerjee, M.; Pop-Busui, R.; Feldman, E. L.; Ji, L. , Diabetes and obesity are the main metabolic drivers of peripheral neuropathy. Ann Clin Transl Neurol 2018, 5(4), 397–405. [Google Scholar] [CrossRef]
- Callaghan, B. C.; Little, A. A.; Feldman, E. L.; Hughes, R. A. C. , Enhanced glucose control for preventing and treating diabetic neuropathy. Cochrane Database of Systematic Reviews 2012, (6). [Google Scholar] [CrossRef]
- Rodriguez-Gutierrez, R.; Gonzalez-Gonzalez, J. G.; Zuniga-Hernandez, J. A.; McCoy, R. G. , Benefits and harms of intensive glycemic control in patients with type 2 diabetes. BMJ 2019, 367, l5887. [Google Scholar] [CrossRef]
- Afshinnia, F.; Reynolds, E. L.; Rajendiran, T. M.; Soni, T.; Byun, J.; Savelieff, M. G.; Looker, H. C.; Nelson, R. G.; Michailidis, G.; Callaghan, B. C.; Pennathur, S.; Feldman, E. L. , Serum lipidomic determinants of human diabetic neuropathy in type 2 diabetes. Ann Clin Transl Neurol 2022, 9(9), 1392–1404. [Google Scholar] [CrossRef]
- Handzlik, M. K.; Gengatharan, J. M.; Frizzi, K. E.; McGregor, G. H.; Martino, C.; Rahman, G.; Gonzalez, A.; Moreno, A. M.; Green, C. R.; Guernsey, L. S.; Lin, T.; Tseng, P.; Ideguchi, Y.; Fallon, R. J.; Chaix, A.; Panda, S.; Mali, P.; Wallace, M.; Knight, R.; Gantner, M. L.; Calcutt, N. A.; Metallo, C. M. , Insulin-regulated serine and lipid metabolism drive peripheral neuropathy. Nature 2023, 614(7946), 118–124. [Google Scholar] [CrossRef] [PubMed]
- Hammad, S. M.; Baker, N. L.; El Abiad, J. M.; Spassieva, S. D.; Pierce, J. S.; Rembiesa, B.; Bielawski, J.; Lopes-Virella, M. F.; Klein, R. L. , Increased Plasma Levels of Select Deoxy-ceramide and Ceramide Species are Associated with Increased Odds of Diabetic Neuropathy in Type 1 Diabetes: A Pilot Study. Neuromolecular Med 2017, 19(1), 46–56. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X. T.; Huang, N.; Sheng, Z. H. , Programming axonal mitochondrial maintenance and bioenergetics in neurodegeneration and regeneration. Neuron 2022, 110(12), 1899–1923. [Google Scholar] [CrossRef] [PubMed]
- Rumora, A. E.; Lentz, S. I.; Hinder, L. M.; Jackson, S. W.; Valesano, A.; Levinson, G. E.; Feldman, E. L. , Dyslipidemia impairs mitochondrial trafficking and function in sensory neurons. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2018, 32(1), 195–207. [Google Scholar] [CrossRef] [PubMed]
- Rumora, A. E.; LoGrasso, G.; Haidar, J. A.; Dolkowski, J. J.; Lentz, S. I.; Feldman, E. L. , Chain length of saturated fatty acids regulates mitochondrial trafficking and function in sensory neurons. J Lipid Res 2019, 60(1), 58–70. [Google Scholar] [CrossRef] [PubMed]
- Almaguel, F. G.; Liu, J. W.; Pacheco, F. J.; Casiano, C. A.; De Leon, M. , Activation and reversal of lipotoxicity in PC12 and rat cortical cells following exposure to palmitic acid. Journal of neuroscience research 2009, 87(5), 1207–18. [Google Scholar] [CrossRef]
- Montero, M. L.; Liu, J. W.; Orozco, J.; Casiano, C. A.; De Leon, M. , Docosahexaenoic acid protection against palmitic acid-induced lipotoxicity in NGF-differentiated PC12 cells involves enhancement of autophagy and inhibition of apoptosis and necroptosis. J Neurochem 2020, 155(5), 559–576. [Google Scholar] [CrossRef]
- Guerbette, T.; Rioux, V.; Bostoen, M.; Ciesielski, V.; Coppens-Exandier, H.; Buraud, M.; Lan, A.; Boudry, G. , Saturated fatty acids differently affect mitochondrial function and the intestinal epithelial barrier depending on their chain length in the in vitro model of IPEC-J2 enterocytes. Front Cell Dev Biol 2024, 12, 1266842. [Google Scholar] [CrossRef]
- Feldman, E. L.; Nave, K. A.; Jensen, T. S.; Bennett, D. L. H. , New Horizons in Diabetic Neuropathy: Mechanisms, Bioenergetics, and Pain. Neuron 2017, 93(6), 1296–1313. [Google Scholar] [CrossRef]
- Mignard, V.; Dubois, N.; Lanoe, D.; Joalland, M. P.; Oliver, L.; Pecqueur, C.; Heymann, D.; Paris, F.; Vallette, F. M.; Lalier, L. , Sphingolipid distribution at mitochondria-associated membranes (MAMs) upon induction of apoptosis. J Lipid Res 2020, 61(7), 1025–1037. [Google Scholar] [CrossRef]
- Descorbeth, M.; Figueroa, K.; Serrano-Illan, M.; De Leon, M. , Protective effect of docosahexaenoic acid on lipotoxicity-mediated cell death in Schwann cells: Implication of PI3K/AKT and mTORC2 pathways. Brain Behav 2018, 8(11), e01123. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J. D.; Cordero, K.; Serrano-Illan, M.; Almeyda, A.; Baldeosingh, K.; Almaguel, F. G.; De Leon, M. , Metabolomics uncovers dietary omega-3 fatty acid-derived metabolites implicated in anti-nociceptive responses after experimental spinal cord injury. Neuroscience 2013, 255, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Coppey, L.; Davidson, E.; Shevalye, H.; Torres, M. E.; Yorek, M. A. , Effect of dietary oils on peripheral neuropathy-related endpoints in dietary obese rats. Diabetes Metab Syndr Obes 2018, 11, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Veigas, J. M.; Williams, P. J.; Halade, G.; Rahman, M. M.; Yoneda, T.; Fernandes, G. , Fish oil concentrate delays sensitivity to thermal nociception in mice. Pharmacol Res 2011, 63(5), 377–82. [Google Scholar] [CrossRef] [PubMed]
- Huang, C. T.; Tsai, Y. J. , Docosahexaenoic acid confers analgesic effects after median nerve injury via inhibition of c-Jun N-terminal kinase activation in microglia. J Nutr Biochem 2016, 29, 97–106. [Google Scholar] [CrossRef]
- Pacheco, F. J.; Almaguel, F. G.; Evans, W.; Rios-Colon, L.; Filippov, V.; Leoh, L. S.; Rook-Arena, E.; Mediavilla-Varela, M.; De Leon, M.; Casiano, C. A. , Docosahexanoic acid antagonizes TNF-α-induced necroptosis by attenuating oxidative stress, ceramide production, lysosomal dysfunction, and autophagic features. Inflamm Res 2014, 63(10), 859–71. [Google Scholar] [CrossRef]
- Figueroa, J. D.; Serrano-Illan, M.; Licero, J.; Cordero, K.; Miranda, J. D.; De Leon, M. , Fatty Acid Binding Protein 5 Modulates Docosahexaenoic Acid-Induced Recovery in Rats Undergoing Spinal Cord Injury. J Neurotrauma 2016, 33(15), 1436–49. [Google Scholar] [CrossRef]
- Liu, J. W.; Almaguel, F. G.; Bu, L.; De Leon, D. D.; De Leon, M. , Expression of E-FABP in PC12 cells increases neurite extension during differentiation: involvement of n-3 and n-6 fatty acids. J Neurochem 2008, 106(5), 2015–29. [Google Scholar] [CrossRef]
- Liu, J. W.; Montero, M.; Bu, L.; De Leon, M. , Epidermal fatty acid-binding protein protects nerve growth factor-differentiated PC12 cells from lipotoxic injury. J Neurochem 2015, 132(1), 85–98. [Google Scholar] [CrossRef]
- Liu, Y.; Longo, L. D.; De León, M. , In situ and immunocytochemical localization of E-FABP mRNA and protein during neuronal migration and differentiation in the rat brain. Brain Res 2000, 852(1), 16–27. [Google Scholar] [CrossRef]
- Durán, A. M.; Beeson, W. L.; Firek, A.; Cordero-MacIntyre, Z.; De León, M. , Dietary Omega-3 Polyunsaturated Fatty-Acid Supplementation Upregulates Protective Cellular Pathways in Patients with Type 2 Diabetes Exhibiting Improvement in Painful Diabetic Neuropathy. Nutrients 2022, 14(4). [Google Scholar] [CrossRef] [PubMed]
- Durán, A. M.; Salto, L. M.; Câmara, J.; Basu, A.; Paquien, I.; Beeson, W. L.; Firek, A.; Cordero-MacIntyre, Z.; De León, M. , Effects of omega-3 polyunsaturated fatty-acid supplementation on neuropathic pain symptoms and sphingosine levels in Mexican-Americans with type 2 diabetes. Diabetes Metab Syndr Obes 2019, 12, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Löfgren, L.; Ståhlman, M.; Forsberg, G. B.; Saarinen, S.; Nilsson, R.; Hansson, G. I. , The BUME method: a novel automated chloroform-free 96-well total lipid extraction method for blood plasma. J Lipid Res 2012, 53(8), 1690–700. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Y.; Tsai, Y. J.; Chen, S. H.; Lin, C. T.; Lue, J. H. , Lysophosphatidylcholine causes neuropathic pain via the increase of neuronal nitric oxide synthase in the dorsal root ganglion and cuneate nucleus. Pharmacol Biochem Behav 2013, 106, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Lin, J.; Yu, L.; Yan, M. , Lysophosphatidylcholine: Potential Target for the Treatment of Chronic Pain. Int J Mol Sci 2022, 23(15). [Google Scholar] [CrossRef]
- Saberi Firouzi, S.; Namazi Sarvestani, N.; Bakhtiarian, A.; Ghazi Khansari, M.; Karimi, M. Y.; Ranjbar, A.; Safa, M.; Hosseini, A. , Sildenafil protective effects on high glucose-induced neurotoxicity in PC12 cells: the role of oxidative stress, apoptosis, and inflammation pathways in an in vitro cellular model for diabetic neuropathy. Neurol Res 2018, 40(8), 624–636. [Google Scholar] [CrossRef]
- Lelkes, E.; Unsworth, B. R.; Lelkes, P. I. , Reactive oxygen species, apoptosis and alte1red NGF-induced signaling in PC12 pheochromocytoma cells cultured in elevated glucose: AnIn Vitro cellular model for diabetic neuropathy. Neurotoxicity Research 2001, 3(2), 189–203. [Google Scholar] [CrossRef]
- Ulloth, J. E.; Almaguel, F. G.; Padilla, A.; Bu, L.; Liu, J. W.; De Leon, M. , Characterization of methyl-beta-cyclodextrin toxicity in NGF-differentiated PC12 cell death. Neurotoxicology 2007, 28(3), 613–21. [Google Scholar] [CrossRef]
- Huang, H. C.; Chen, L.; Zhang, H. X.; Li, S. F.; Liu, P.; Zhao, T. Y.; Li, C. X. , Autophagy Promotes Peripheral Nerve Regeneration and Motor Recovery Following Sciatic Nerve Crush Injury in Rats. J Mol Neurosci 2016, 58(4), 416–23. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Wang, L.; Wang, P.; Xue, Y.; Li, X.; Qiao, X.; Zhang, X.; Xu, T.; Liu, G.; Li, P.; Chen, C. , Autophagy impairment mediated by S-nitrosation of ATG4B leads to neurotoxicity in response to hyperglycemia. Autophagy 2017, 13(7), 1145–1160. [Google Scholar] [CrossRef]
- Mohseni, S.; Badii, M.; Kylhammar, A.; Thomsen, N. O. B.; Eriksson, K. F.; Malik, R. A.; Rosen, I.; Dahlin, L. B. , Longitudinal study of neuropathy, microangiopathy, and autophagy in sural nerve: Implications for diabetic neuropathy. Brain Behav 2017, 7(8), e00763. [Google Scholar] [CrossRef] [PubMed]
- Valencia, M.; Kim, S. R.; Jang, Y.; Lee, S. H. , Neuronal Autophagy: Characteristic Features and Roles in Neuronal Pathophysiology. Biomol Ther (Seoul) 2021, 29(6), 605–614. [Google Scholar] [CrossRef] [PubMed]
- de Le�n, M.; Welcher, A. A.; Nahin, R. H.; Liu, Y.; Ruda, M. A.; Shooter, E. M.; Molina, C. A. , Fatty acid binding protein is induced in neurons of the dorsal root ganglia after peripheral nerve injury. Journal of Neuroscience Research 1996, 44(3), 283–292. [Google Scholar] [CrossRef]
- Liu, Y.; Molina, C. A.; Welcher, A. A.; Longo, L. D.; De Leon, M. , Expression of DA11, a neuronal-injury-induced fatty acid binding protein, coincides with axon growth and neuronal differentiation during central nervous system development. J Neurosci Res 1997, 48(6), 551–62. [Google Scholar] [CrossRef]
- Kagawa, Y.; Jin, L.; Sun, J.; Nicolazzo, J.; Owada, Y.; Pan, Y. , Microglia with fatty acid-binding protein 5 deficiency exhibit proinflammatory phenotype contributing to neuroinflammation. Alzheimer's & Dementia 2022, 18(S4). [Google Scholar]
- Pan, Y.; Short, J. L.; Choy, K. H.; Zeng, A. X.; Marriott, P. J.; Owada, Y.; Scanlon, M. J.; Porter, C. J.; Nicolazzo, J. A. , Fatty Acid-Binding Protein 5 at the Blood-Brain Barrier Regulates Endogenous Brain Docosahexaenoic Acid Levels and Cognitive Function. J Neurosci 2016, 36(46), 11755–11767. [Google Scholar] [CrossRef]
- Bertea, M.; Rütti, M. F.; Othman, A.; Marti-Jaun, J.; Hersberger, M.; von Eckardstein, A.; Hornemann, T. , Deoxysphingoid bases as plasma markers in Diabetes mellitus. Lipids in health and disease 2010, 9(1), 84. [Google Scholar] [CrossRef]
- Eid, S.; Sas, K. M.; Abcouwer, S. F.; Feldman, E. L.; Gardner, T. W.; Pennathur, S.; Fort, P. E. , New insights into the mechanisms of diabetic complications: role of lipids and lipid metabolism. Diabetologia 2019, 62(9), 1539–1549. [Google Scholar] [CrossRef]
- Padilla, A.; Descorbeth, M.; Almeyda, A. L.; Payne, K.; De Leon, M. , Hyperglycemia magnifies Schwann cell dysfunction and cell death triggered by PA-induced lipotoxicity. Brain Res 2011, 1370, 64–79. [Google Scholar] [CrossRef]
- Ren, C.; Liu, J.; Zhou, J.; Liang, H.; Wang, Y.; Sun, Y.; Ma, B.; Yin, Y. , Lipidomic analysis of serum samples from migraine patients. Lipids in health and disease 2018, 17(1), 22. [Google Scholar] [CrossRef]
- Arima, H.; Hanada, M.; Hayasaka, T.; Masaki, N.; Omura, T.; Xu, D.; Hasegawa, T.; Togawa, D.; Yamato, Y.; Kobayashi, S.; Yasuda, T.; Matsuyama, Y.; Setou, M. , Blockade of IL-6 signaling by MR16-1 inhibits reduction of docosahexaenoic acid-containing phosphatidylcholine levels in a mouse model of spinal cord injury. Neuroscience 2014, 269, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pop-Busui, R.; Boulton, A. J.; Feldman, E. L.; Bril, V.; Freeman, R.; Malik, R. A.; Sosenko, J. M.; Ziegler, D. , Diabetic Neuropathy: A Position Statement by the American Diabetes Association. Diabetes Care 2017, 40(1), 136–154. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Nakamura, K.; Matsui, M.; Yamamoto, A.; Nakahara, Y.; Suzuki-Migishima, R.; Yokoyama, M.; Mishima, K.; Saito, I.; Okano, H.; Mizushima, N. , Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 2006, 441(7095), 885–9. [Google Scholar] [CrossRef] [PubMed]
- Yerra, V. G.; Kalvala, A. K.; Kumar, A. , Isoliquiritigenin reduces oxidative damage and alleviates mitochondrial impairment by SIRT1 activation in experimental diabetic neuropathy. J Nutr Biochem 2017, 47, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Y.; Zhou, F.; Xiao, Y.; Shi, L. , Inhibition of PI3K/AKT/mTOR signaling pathway promotes autophagy and relieves hyperalgesia in diabetic rats. Neuroreport 2020, 31(9), 644–649. [Google Scholar] [CrossRef]
- Li, J.; Tian, M.; Hua, T.; Wang, H.; Yang, M.; Li, W.; Zhang, X.; Yuan, H. , Combination of autophagy and NFE2L2/NRF2 activation as a treatment approach for neuropathic pain. Autophagy 2021, 17(12), 4062–4082. [Google Scholar] [CrossRef]
- Hernández-Cáceres, M. P.; Cereceda, K.; Hernández, S.; Li, Y.; Narro, C.; Rivera, P.; Silva, P.; Ávalos, Y.; Jara, C.; Burgos, P.; Toledo-Valenzuela, L.; Lagos, P.; Cifuentes Araneda, F.; Perez-Leighton, C.; Bertocchi, C.; Clegg, D. J.; Criollo, A.; Tapia-Rojas, C.; Burgos, P. V.; Morselli, E. , Palmitic acid reduces the autophagic flux in hypothalamic neurons by impairing autophagosome-lysosome fusion and endolysosomal dynamics. Mol Cell Oncol 2020, 7(5), 1789418. [Google Scholar] [CrossRef]
- Ortiz-Rodriguez, A.; Acaz-Fonseca, E.; Boya, P.; Arevalo, M. A.; Garcia-Segura, L. M. , Lipotoxic Effects of Palmitic Acid on Astrocytes Are Associated with Autophagy Impairment. Mol Neurobiol 2019, 56(3), 1665–1680. [Google Scholar] [CrossRef]
- Chouinard-Watkins, R.; Lacombe, R. J. S.; Bazinet, R. P. , Mechanisms regulating brain docosahexaenoic acid uptake: what is the recent evidence? Curr Opin Clin Nutr Metab Care 2018, 21(2), 71–77. [Google Scholar] [CrossRef]
- Pan, Y.; Morris, E. R.; Scanlon, M. J.; Marriott, P. J.; Porter, C. J. H.; Nicolazzo, J. A. , Dietary docosahexaenoic acid supplementation enhances expression of fatty acid-binding protein 5 at the blood-brain barrier and brain docosahexaenoic acid levels. J Neurochem 2018, 146(2), 186–197. [Google Scholar] [CrossRef]
- Wagner, E. F.; Swenson, C. C.; Henggeler, S. W. , Practical and Methodological Challenges in Validating Community-Based Interventions. Children's Services 2000, 3(4), 211–231. [Google Scholar] [CrossRef]
- Stevens, J. R.; Herrick, J. S.; Wolff, R. K.; Slattery, M. L. , Power in pairs: assessing the statistical value of paired samples in tests for differential expression. BMC Genomics 2018, 19(1), 953. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Matched pairs t-test with adjusted P-value (FDR) cutoff of 0.05. Eighty-one (81) lipid features were significantly different (p<0.05) at three months.
Figure 1.
Matched pairs t-test with adjusted P-value (FDR) cutoff of 0.05. Eighty-one (81) lipid features were significantly different (p<0.05) at three months.
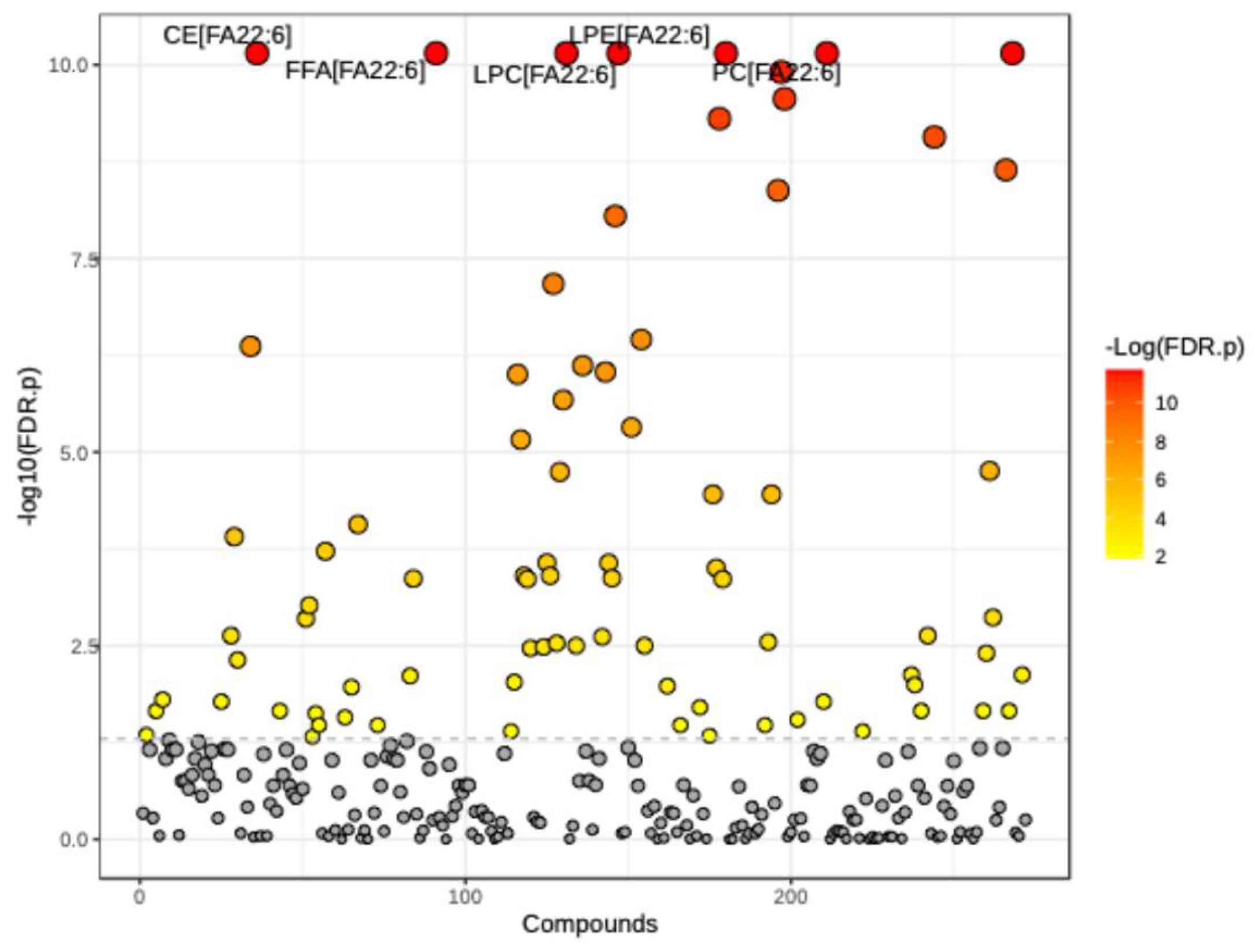
Figure 2.
RF classification of serum samples collected at baseline (0) and three (3) months after DHA-rich supplementation (1). Classification was ∽96% accurate for samples when a value of 50% would be expected by random chance. The plot for lipid features is important because it shows the top factors contributing to group separation.
Figure 2.
RF classification of serum samples collected at baseline (0) and three (3) months after DHA-rich supplementation (1). Classification was ∽96% accurate for samples when a value of 50% would be expected by random chance. The plot for lipid features is important because it shows the top factors contributing to group separation.
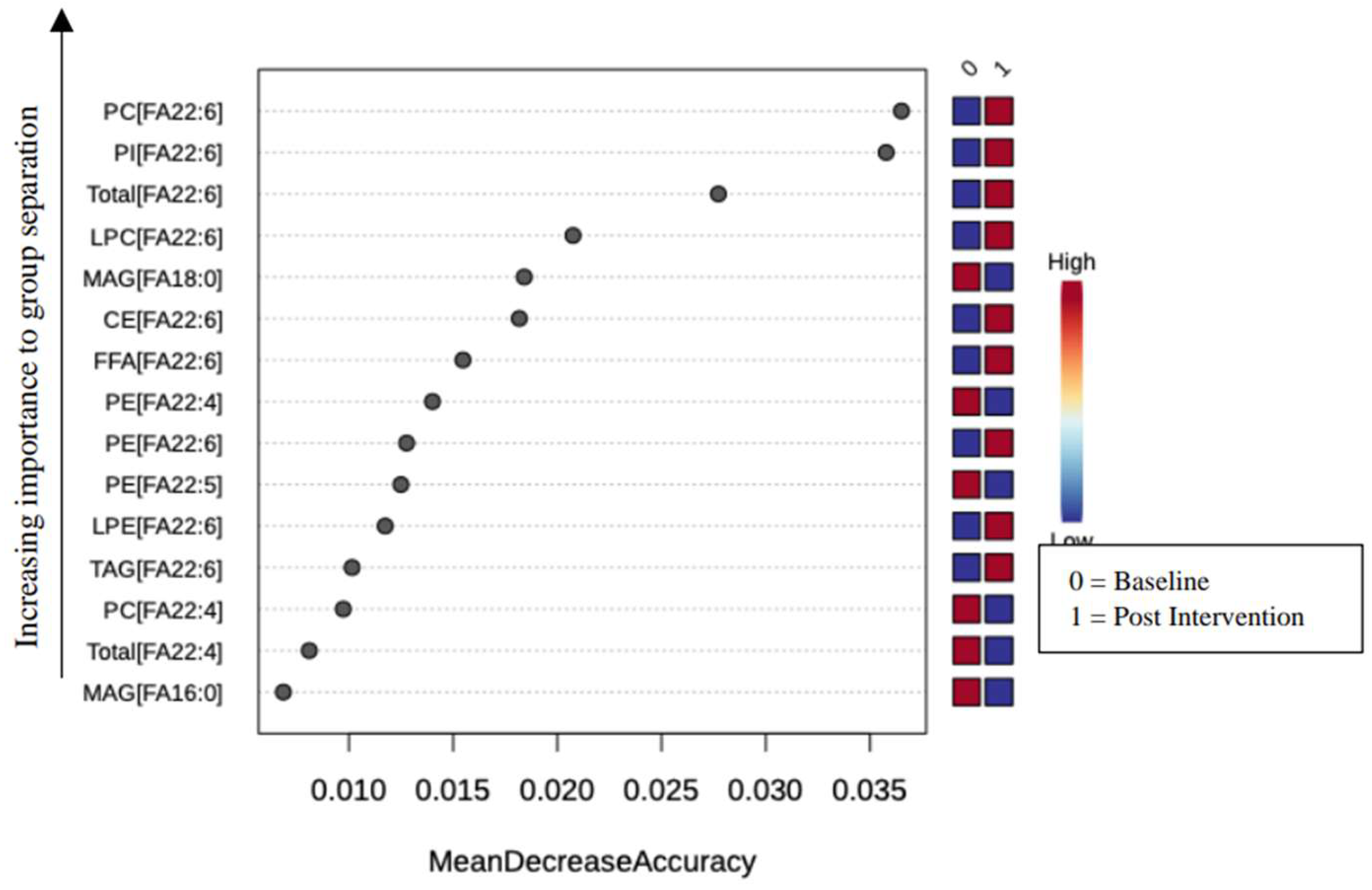
Figure 3.
Mixed lineage kinase domain-like (MLKL) decreases after three (3) months of DHA-rich supplementation in participants with type II diabetes and Neuropathic Pain. Match pairs Wilcoxon test A) ELISA measured all participants' serum levels of MLKL at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of MLKL, measured by ELISA, at baseline and three months. * p < 0.05, **** p < 0.0001.
Figure 3.
Mixed lineage kinase domain-like (MLKL) decreases after three (3) months of DHA-rich supplementation in participants with type II diabetes and Neuropathic Pain. Match pairs Wilcoxon test A) ELISA measured all participants' serum levels of MLKL at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of MLKL, measured by ELISA, at baseline and three months. * p < 0.05, **** p < 0.0001.
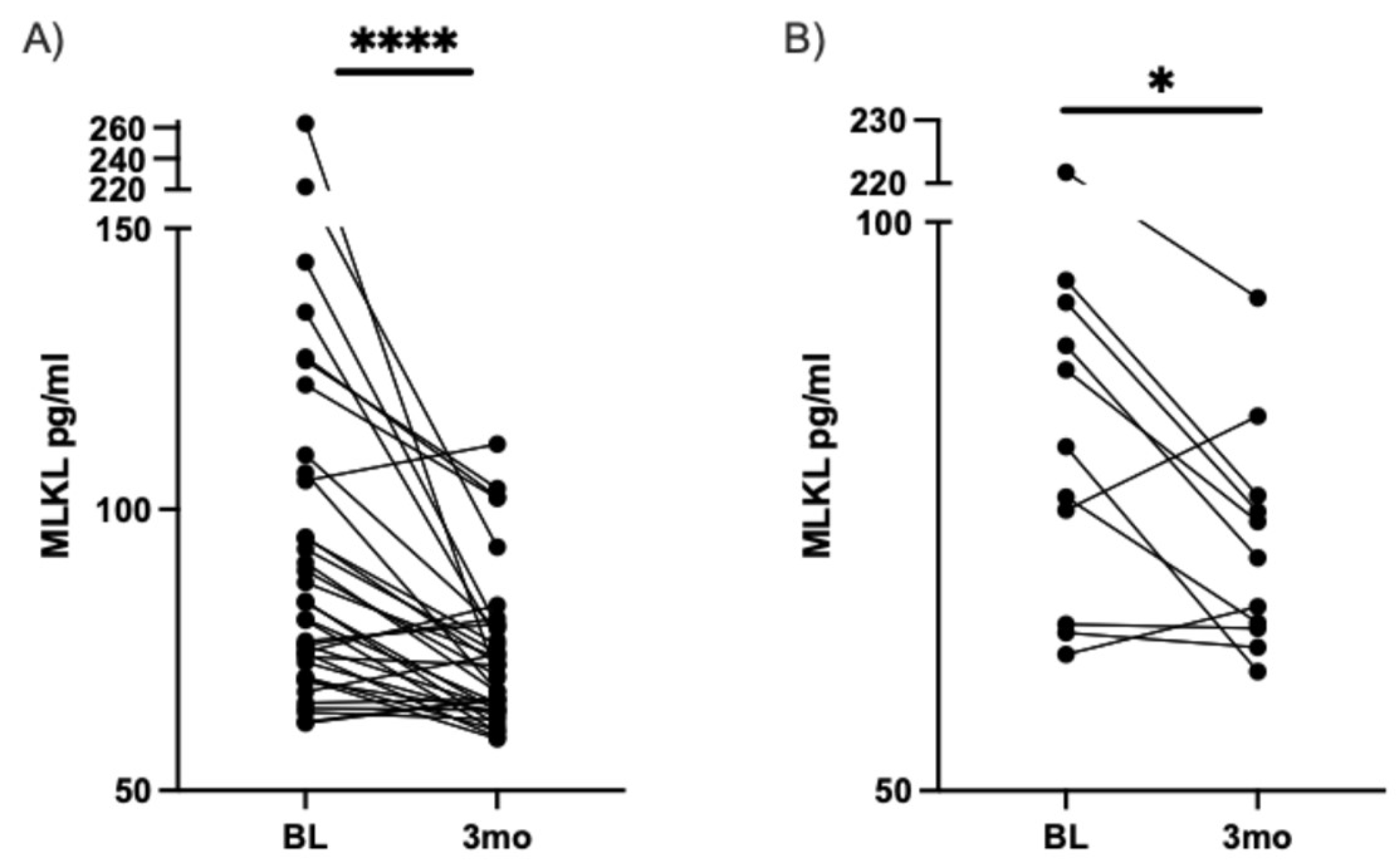
Figure 4.
ATG5 increases after three months of DHA-rich supplementation in participants with type II diabetes and with painful DN. Match pairs Wilcoxon test A) All participants' serum levels of ATG5, measured by ELISA, at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of ATG5, measured by ELISA, at baseline and for three months. * p < 0.05.
Figure 4.
ATG5 increases after three months of DHA-rich supplementation in participants with type II diabetes and with painful DN. Match pairs Wilcoxon test A) All participants' serum levels of ATG5, measured by ELISA, at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of ATG5, measured by ELISA, at baseline and for three months. * p < 0.05.
Figure 5.
FABP5 increases after three months of DHA-rich supplementation in participants with type II diabetes and Neuropathic Pain. Match pairs Wilcoxon test A) ELISA measured all participants' serum levels of FABP5 at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of FABP5, measured by ELISA, at baseline and for three months. ** p < 0.01, **** p < 0.0001.
Figure 5.
FABP5 increases after three months of DHA-rich supplementation in participants with type II diabetes and Neuropathic Pain. Match pairs Wilcoxon test A) ELISA measured all participants' serum levels of FABP5 at baseline and three months. B) Participants with moderately high pain at baseline and serum levels of FABP5, measured by ELISA, at baseline and for three months. ** p < 0.01, **** p < 0.0001.
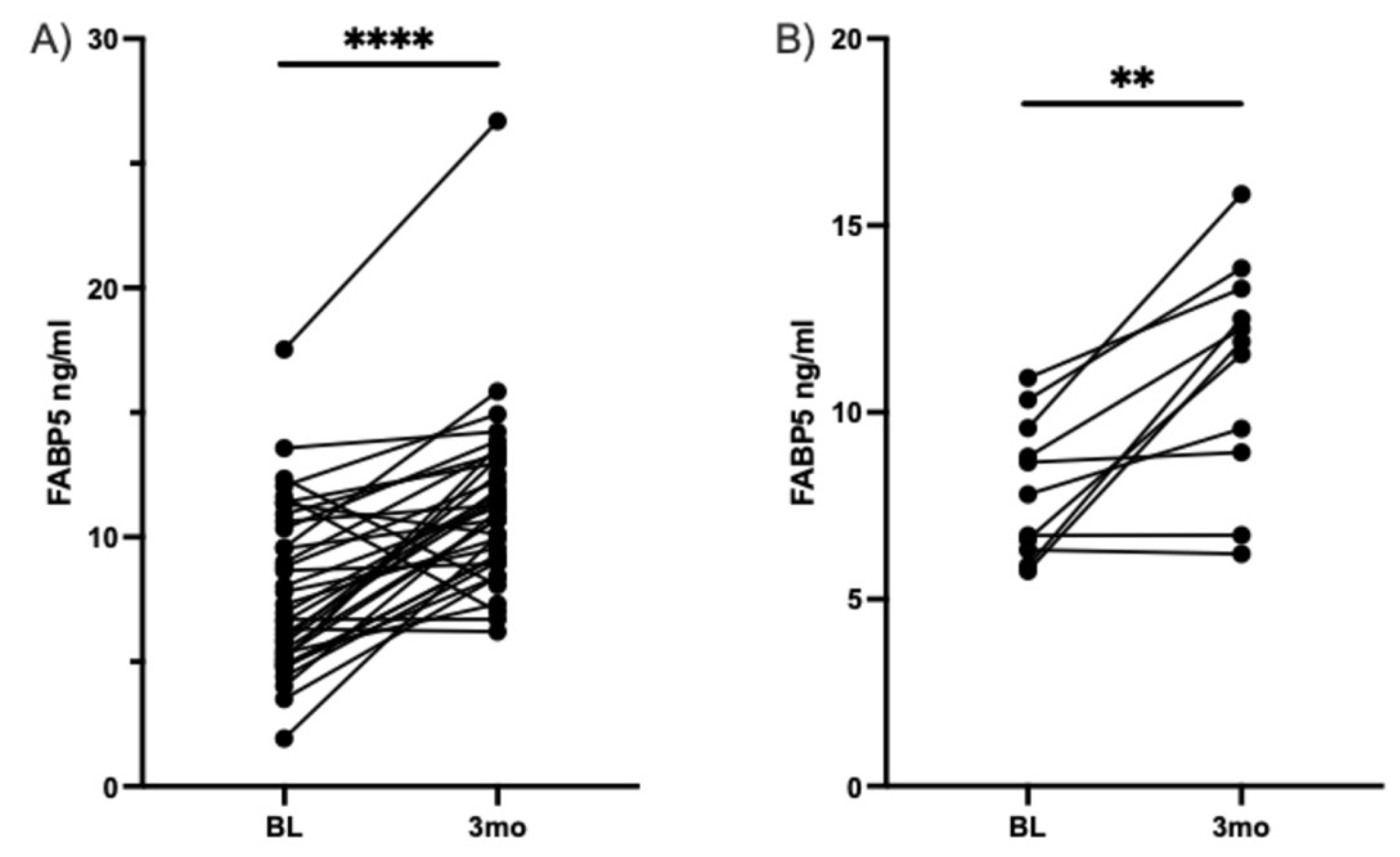

Table 1.
Untargeted Fatty Acid compound identification and statistical comparison.
| Total Biochemicals Identified | 283 | |||
| Total Biochemicals p ≤ 0.05, adjusted P-value (FDR) cutoff, matched paired t-test | 81 | |||
| Biochemicals (↑|↓) | 22 |59 |
Table 2.
Significantly different fatty acids post DHA-rich supplementation.
| Group | Lipid Class | Name | p-value | FDR |
| Neutral Lipids | Cholesteryl ester (CE) | CE[FA22:6] | 1.819E-12 | 7.0681E-11 |
| CE[FA22:4] | 2.6917E-08 | 4.3068E-07 | ||
| CE[FA20:4] | 1.3218E-05 | 0.00012397 | ||
| CE[FA20:3] | 0.0003791 | 0.0023435 | ||
| CE[FA20:5] | 0.00094117 | 0.0048302 | ||
| CE[FA20:0] | 0.0038675 | 0.016698 | ||
| Monoacylglycerol (MAG) | MAG[FA18:0] | 2.07E-08 | 3.519E-07 | |
| MAG[FA16:0] | 3.8752E-07 | 4.7911E-06 | ||
| MAG[FA18:1] | 0.00056988 | 0.0031634 | ||
| Triacylglycerol (TAG) | TAG[FA22:6] | 3.4561E-11 | 8.5459E-10 | |
| TAG[FA22:4] | 0.0003791 | 0.0023435 | ||
| TAG[FA20:2] | 0.0015176 | 0.0075053 | ||
| TAG[FA20:3] | 0.0021663 | 0.010159 | ||
| TAG[FA20:5] | 0.0053322 | 0.021975 | ||
| Diacylglycerol (DAG) | DAG[FA22:6] | 2.1048E-05 | 0.00019083 | |
| DAG[FA20:3] | 0.00014039 | 0.00095468 | ||
| DAG[FA20:2] | 0.0002192 | 0.0014196 | ||
| DAG[FA16:0] | 0.0053322 | 0.021975 | ||
| DAG[FA20:5] | 0.0060965 | 0.023689 | ||
| DAG[FA22:4] | 0.0093813 | 0.033575 | ||
| DAG[FA20:4] | 0.014108 | 0.047375 | ||
| Free fatty acid (FFA) | FFA[FA22:6] | 1.819E-12 | 7.0681E-11 | |
| FFA[FA20:4] | 5.8459E-05 | 0.00042975 | ||
| FFA[FA20:3] | 0.0015981 | 0.0077621 | ||
| FFA[FA16:0] | 0.0093813 | 0.033575 | ||
| Phospholipids | Phosphatidylcholine (PC) | PC[FA22:6] | 1.819E-12 | 7.0681E-11 |
| PC[FA22:4] | 1.819E-11 | 4.9477E-10 | ||
| PC[FA20:4] | 3.4888E-06 | 3.5146E-05 | ||
| PC[FA20:5] | 3.813E-05 | 0.00031428 | ||
| PC[FA22:5] | 6.2683E-05 | 0.00043717 | ||
| PC[FA14:1] | 0.0022767 | 0.010496 | ||
| PC[FA20:0] | 0.0046535 | 0.019777 | ||
| PC[FA17:0] | 0.0093813 | 0.033575 | ||
| PC[FA20:3] | 0.013558 | 0.046096 | ||
| Phosphatidylethanolamine (PE) | PE[FA22:5] | 3.638E-12 | 1.2369E-10 | |
| PE[FA22:6] | 9.0949E-12 | 2.7487E-10 | ||
| PE[FA22:4] | 2.0009E-10 | 4.1865E-09 | ||
| PE[FA20:5] | 3.4888E-06 | 3.5146E-05 | ||
| PE[FA20:4] | 0.00047945 | 0.002835 | ||
| PE[FA20:3] | 0.008995 | 0.033516 | ||
| Phosphatidylinositol (PI) | PI[FA22:6] | 1.819E-12 | 7.0681E-11 | |
| PI[FA22:5] | 0.0038675 | 0.016698 | ||
| PI[FA18:1] | 0.0075849 | 0.028654 | ||
| Lysophosphatidylcholine (LPC) | LPC[FA22:6] | 1.819E-12 | 7.0681E-11 | |
| LPC[FA20:4] | 3.7016E-09 | 6.7123E-08 | ||
| LPC[FA16:0] | 7.2841E-08 | 9.9064E-07 | ||
| LPC[FA22:5] | 1.6344E-07 | 2.1169E-06 | ||
| LPC[FA16:1] | 5.8416E-07 | 6.9084E-06 | ||
| LPC[FA22:4] | 1.6604E-06 | 1.8065E-05 | ||
| LPC[FA20:2] | 3.0617E-05 | 0.00026864 | ||
| LPC[FA17:0] | 5.0783E-05 | 0.00039466 | ||
| LPC[FA20:3] | 5.0783E-05 | 0.00039466 | ||
| LPC[FA18:0] | 6.2683E-05 | 0.00043717 | ||
| LPC[FA20:5] | 0.00050804 | 0.0029401 | ||
| LPC[FA20:1] | 0.00060329 | 0.0032819 | ||
| LPC[FA18:1] | 0.00063846 | 0.0034051 | ||
| LPC[FA15:0] | 0.0019595 | 0.0093506 | ||
| LPC[FA14:0] | 0.011537 | 0.040231 | ||
| Lysophosphatidylethanolamine (LPE) | LPE[FA22:6] | 1.819E-12 | 7.0681E-11 | |
| LPE[FA22:5] | 4.602E-10 | 8.9411E-09 | ||
| LPE[FA18:0] | 5.0639E-08 | 7.6521E-07 | ||
| LPE[FA20:4] | 6.4607E-08 | 9.249E-07 | ||
| LPE[FA20:5] | 3.1753E-05 | 0.0002699 | ||
| LPE[FA22:4] | 5.623E-05 | 0.00042485 | ||
| LPE[FA20:3] | 0.00040222 | 0.0024312 | ||
| LPE[FA16:1] | 0.00056988 | 0.0031634 | ||
| Sphingolipids | Sphingomyelin (SM) | SM[FA26:0] | 0.011537 | 0.040231 |
| Ceramide (CER) | CER[FA22:0] | 0.0035205 | 0.015698 | |
| CER[FA20:0] | 0.005577 | 0.021985 | ||
| CER[FA16:0] | 0.013026 | 0.044848 | ||
| Dihydroceramide (DCER) | DCER[FA26:0] | 8.848E-06 | 8.5952E-05 | |
| DCER[FA24:0] | 0.0023922 | 0.010845 | ||
| DCER[FA22:0] | 0.0069552 | 0.026645 | ||
| Total | N/A | Total [FA22:6] | 1.819E-12 | 7.0681E-11 |
| Total [FA22:4] | 1.0004E-10 | 2.2677E-09 | ||
| Total [FA20:4] | 1.5435E-06 | 1.7493E-05 | ||
| Total [FA20:5] | 0.0002059 | 0.001366 | ||
| Total [FA20:3] | 0.00075535 | 0.0039511 | ||
| Total [FA26:0] | 0.0015176 | 0.0075053 | ||
| Total [FA20:2] | 0.005577 | 0.021985 | ||
| Total [FA22:5] | 0.005577 | 0.021985 |
Significant fatty acid increases and decreases are highlighted in red and blue.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



