
Submitted:
19 November 2024
Posted:
20 November 2024
You are already at the latest version
Abstract
This study investigates the diversity and distribution of intertidal Cystoseira sensu lato (s.l.) species across different protection zones within the "Capo Gallo-Isola delle Femmine" Marine Protected Area (MPA) in the central Mediterranean Sea. Five Cystoseira s.l. species (Cystoseira compressa, C. foeniculacea, Ericaria amentacea, E. brachycarpa and E. crinita) were observed on the intertidal rocky shores, with varied abundances across the MPA's protection zones. Ericaria amentacea was the only species found in all zones, with a much higher cover percentage in the most protected area (zone A). However, its morpho-functional traits showed an inverse trend, with the largest thalli found in the moderately protected zone B and C. The remaining Cystoseira s.l. species were only found in zone B and C probably due to their wider area and greater habitat diversity compared to zone A. The presence of two non-indigenous species, Asparagopsis taxiformis and Caulerpa cylindracea, was ob-served exclusively in the less protected zones B and C. Our findings highlight the complex inter-actions between MPA protection levels and Cystoseira s.l. species conservation, with protection having, in some respects, a positive influence on selected Cystoseira s.l. species, indicating the im-portance of careful consideration in MPA design and management. Furthermore, this study pro-vides a baseline for future monitoring of Cystoseira s.l. populations in light of ongoing environmental changes in the Mediterranean Sea.
Keywords:
biodiversity
; macroalgal distribution
; Marine Protected Areas
; Mediterranean Sea
; Cystoseira sensu lato
; Cystoseira
; Ericaria
; Gongolaria
1. Introduction
Marine Protected Areas (MPAs) have emerged as vital tools for biodiversity conservation and ecosystem management worldwide [1,2]. These protected zones serve as valuable resources for studying the effects of human activities on marine ecosystems and the potential for recovery when such pressures are reduced or eliminated [3]. Among the various indicators of ecosystem health in Mediterranean coastal waters, brown seaweeds of the genus Cystoseira sensu lato (s.l.) - including the genera Cystoseira, Ericaria, and Gongolaria [4,5] - have emerged as particularly significant.
In the Mediterranean rocky shores, seaweeds belonging to the genus Cystoseira s.l. play a valuable role as foundation species, forming complex three-dimensional habitats that support high biodiversity and provide numerous ecosystem services [6,7,8]. These seaweeds significantly enhance the structural complexity and productivity of coastal communities from the intertidal down to the upper circalittoral zone [9,10,11,12]. Additionally, they are considered useful indicators of water and ecosystem quality according to the Water Framework Directive (2000/60/EC) and the Marine Strategy Framework Directive (2008/56/EC) [13,14].
Some species of Cystoseira s.l. are recognized as highly impacted, particularly near urban areas, due to the interplay effects of local anthropogenic pressures (e.g., pollution, urbanization, the introduction of non-native species, overfishing, coastal aquaculture), and global climate change [7,15,16,17,18]. Consequently, the decline or loss of Cystoseira s.l. populations has been observed on many rocky coasts, leading to a shift from complex and productive benthic communities to less structured assemblages dominated by turf-forming algae, mussels, or sea urchin barrens [19,20], and a decrease of valuable ecosystem services [21].
While MPAs can play a critical role in the conservation of Cystoseira s.l. forests by providing protection from various anthropogenic impacts [22], their effectiveness in preserving these foundation seaweeds remains understudied. The protective environment within MPAs may facilitate higher reproduction rates of Cystoseira s.l. populations by reducing local anthropogenic stressors, facilitating conditions to successful reproduction and the establishment of new individuals [22].
We investigated the diversity and distribution of Cystoseira s.l. populations on the intertidal rocky shore within the “Capo Gallo-Isola delle Femmine” MPA in the central Mediterranean Sea and tested whether the diverse protection zones within the MPA may affect their conservation differently. For Ericaria amentacea (C.Agardh) Molinari & Guiry 2020, one of the main species characterizing the intertidal zone, we also explored changes in key morpho-functional traits across the protection zones. Additionally, we documented the presence of non-indigenous seaweeds in the investigated area, providing crucial information for understanding current challenges to conservation efforts. With this study we provide a baseline understanding of Cystoseira s.l. population dynamics within the “Capo Gallo-Isola delle Femmine” MPA (Figure 1) by updating a previous research [23]. This data will be essential for the MPA to plan future monitoring activities, which are mandatory for effective conservation strategies in the face of global change.
2. Results
A total of 5 Cystoseira s.l. species (Cystoseira compressa (Esper) Gerloff & Nizamuddin 1975, Cystoseira foeniculacea (Linnaeus) Greville 1830, E. amentacea, E. brachycarpa (J.Agardh) Molinari & Guiry 2020 and E. crinita (Duby) Molinari & Guiry 2020) were identified along the intertidal rocky shore of the MPA of Capo Gallo – Isola delle Femmine (Figure 2).
Ericaria amentacea was mainly found on the outer edge of the vermetid reef. It was the only species among that discovered in this study, present in all the MPA protection zones. In particular, the cover percentage of E. amentacea was significantly higher in zone A, while decreased in the other protection zones, with the lowest values in zone C (Figure 2a, Table S1-S2).
Cystoseira compressa was mainly discovered within intertidal pools along the vermetid reef, or close to the external edge of the vermetid reef, where substitutes E. amentacea. It was rarely associated with E. amentacea. Cystoseira compressa was found in B and in C zones with average cover of 2.6 % and 0.44%, respectively (Figure 2b).
Cystoseira foeniculacea, E. crinita and E. brachycarpa were mostly found in the cuvettes of the vermetid platform. The three species, were absent in zone A, while are present in lower percentage in zones B and C. Specifically, C. foeniculacea is found in zone B with an average percentage of 0.25%, and in zone C with a percentage of 0.18%. Ericaria brachycarpa is present in zone B with an average percentage of 0.70%, and in zone C with an average coverage of 2% while E. crinita was found only in zone B with an average percentage of 0.48% (Figure 2c-e).
2.1. Morpho-Functional Parameters of Ericaria Amentacea
Ericaria amentacea presented bigger thalli in zone B, with a total length of about 10.4 ± 0.2 cm, compared to zone C and A, where thalli were 5 ± 0.2 cm, and 4 ± 0.3 cm, length respectively (Figure 3). The percentage of receptacles, epiphytes and burnt tissue were in general more present in zone B (3 ± 0.4, 4 ± 0.5, 6.4 ± 0.5 %, respectively) than in zone C (1.7 ± 0.2, 0.7 ± 0.2, 4.6 ± 0.3%, respectively), while were absent in zone A (Figure 3).
2.2. Non-Native Species
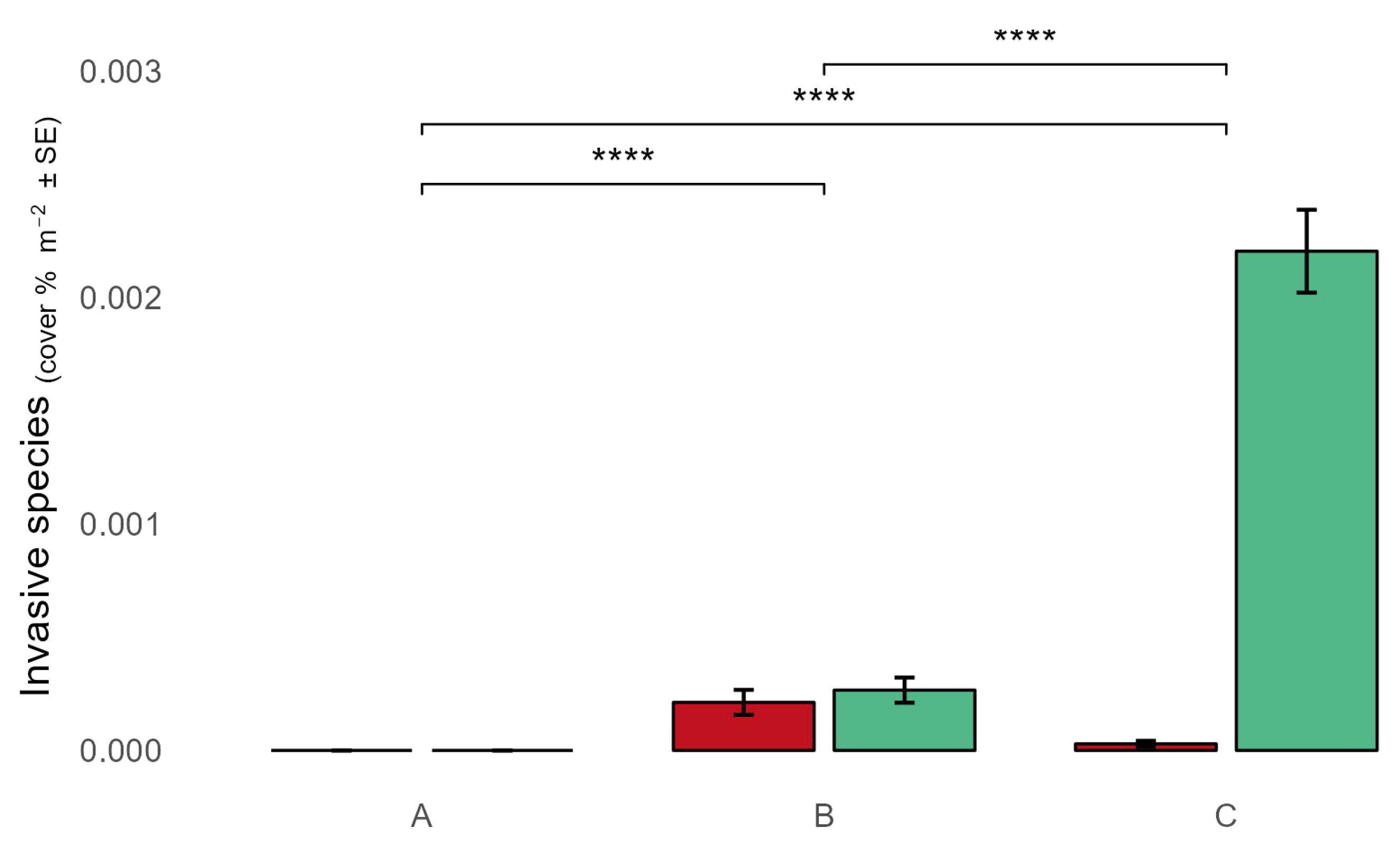
Two species, Asparagopsis taxiformis (Delile) Trevisan 1845 and Caulerpa cylindracea Sonder 1845, were discovered, with the last one resulting more abundant (Figure 4). These species were found only in zones B and C, with C. cylindracea being more abundant in zone C (Figure 4). Asparagopsis taxiformis was found in intertidal pools or in the outer part of the external edge of the vermetid reef, while C. cylindracea was mainly found in the cuvettes of the vermetid platform with thalli not longer than 1-2 cm.
3. Discussion
Our findings reveal a diversified pattern of Cystoseira s.l. species distribution across different protection zones within the MPA of “Capo Gallo - Isola delle Femmine”, highlighting the potential effectiveness of MPAs in conserving these important foundation species. We identified five Cystoseira s.l. species (C. compressa, C. foeniculacea, E. amentacea, E. brachycarpa and E. crinita) on the intertidal rocky shores of the MPA. This diversity underscores the ecological importance of the MPA and aligns with previous studies documenting the richness of Cystoseira s.l. species in Mediterranean coastal ecosystems (Ballesteros et al., 2009; Thibaut et al., 2015, Mancuso et al., 2024). Compared to previous research carried out into the MPA, we observed changes in Cystoseira s.l. composition rather than in number of species. In fact, previous study highlighted the presence of Cystoseira humilis Schousboe ex Kützing 1860 [23]. We believe the absence of this species in our study may be due to its restricted range in the intertidal zone, as it is usually confined to intertidal rocky pools. It’s possible that during our survey, C. humilis may have been retracted or less visible due to high temperatures that intertidal pools can reach during the sampling season. However, this hypothesis requires further investigation to confirm. The discrepancy in our findings underscores the importance of considering temporal factors in surveys particularly for intertidal species that are highly subjected variation of environmental variables.
We found that the distribution of these species varied significantly across the MPA’s protection zones, suggesting that the level of protection influences their abundance and composition. Ericaria amentacea was the widely distributed and abundant species and was the only species present in all protection zones, aligning with previous observation [23]. However, compared to previous research E. amentacea presented a significantly higher cover percentages in zone A (no-take/no-access), suggesting an improvement in its status and supporting the hypothesis that stricter protection measures can benefit Cystoseira s.l. populations, possibly by reducing direct human stressors and maintaining favorable environmental conditions [22]. The variable abundances of Cystoseira s.l. species across protection zones demonstrate the complex relationship between conservation measures and species-specific ecological needs. Ericaria amentacea thrived in the highly protected zone A, whereas C. compressa, C. foeniculacea, E. brachycarpa, and E. crinita were more abundant in the less strictly protected Zones B and C. The observed distribution patterns of Cystoseira s.l. species across protected zones are most likely the result of a complex interaction of factors such as species-specific environmental tolerances, anthropogenic stressors, and interspecific competition. The geomorphological properties of the seabed have an important influence in shaping coastal marine ecosystems and, by extension, seaweeds distribution. In the intertidal zone, substrate composition, topographical features, and hydrodynamic conditions all have a significant influence on the formation and composition of macroalgal communities [24]. Substrate qualities have a direct impact on algal attachment and growth, with various species preferring specific substrate types [25,26].
Furthermore, hydrodynamic conditions influenced by wave exposure and water flow patterns promote the dispersion of reproductive propagules and nutrients, influencing the distribution and diversity of macroalgal assemblages [27]. While we did not specifically include geomorphological data in our analysis, we recognize its potential importance in understanding the observed differences in macroalgal distribution among protected zones. Future research incorporating extensive seabed geomorphology analysis will surely provide greater insights into the distribution patterns of Cystoseira s.l. populations, hence improving our understanding of these complex coastal ecosystems. The absence of some species (C. foeniculacea, E. brachycarpa and E. crinita) from zone A may be attributed to greater habitat variability (e.g., the presence of intertidal pools) and specific habitat requirements that are not met in the most strictly protected areas, rather than the level of protection.
The analysis of E. amentacea’s morpho-functional traits across protection zones reveals interesting patterns. Surprisingly, individuals in zone B exhibited the largest thalli, contrary to our expectation that the most protected area (zone A) would host the most well-developed specimens. This finding suggests that, while the higher cover percentage of E. amentacea in zone A may be facilitated by reducing human stressors such as trampling (especially during summer), which has been shown to have a negative impacts on its growth [28,29], the higher area covered by moderate levels of protection, combined with greater variation in geomorphological characteristics, may provide an optimal balance of environmental conditions for E. amentacea growth. The higher percentages of receptacles, epiphytes, and burnt tissue in zones B and C, compared to their absence in zone A, may indicate also in this case that greater variation in geomorphological characteristics of these less protected areas may experience greater environmental stress or disturbance. These factors could influence the reproductive strategies and overall health of E. amentacea populations. The presence of burnt tissue, in particular, may be an indicator of exposure to extreme temperatures or desiccation, which could be more pronounced in areas with higher human activity [18].
The detection of two non-native species, Asparagopsis taxiformis and Caulerpa cylindracea, exclusively in zones B and C may be concerning. However, their absence from zone A suggests that strict protection measures may help resist biological invasions, possibly by maintaining more intact and resilient native communities [30]. The higher abundance of C. cylindracea in zone C indicates that areas with lower protection levels may be more susceptible to invasion, potentially due to increased disturbance or altered competitive dynamics.
Our findings have important implications for the management of MPAs and the conservation of Cystoseira s.l. species. The effectiveness of strict protection (zone A) in supporting high cover of E. amentacea underscores the value of no-take/no-access areas in preserving key foundation species, especially for trampling related stressors. The variability in species responses to protection levels highlights the need for a diverse approach to MPA zoning to accommodate the requirements of different Cystoseira s.l. species, especially in terms of habitat suitability (e.g., intertidal pools). The presence of invasive species emphasizes the importance of monitoring and management strategies to prevent their spread.
This study provides a baseline for future monitoring of Cystoseira s.l. intertidal populations in the “Capo Gallo-Isola delle Femmine” MPA. Long-term studies are needed to track changes in species distribution, abundance, and traits over time, particularly in the context of global climate change. Additionally, investigating the specific mechanisms driving the observed patterns, such as water quality, herbivory pressure, or competitive interactions, would provide valuable insights for conservation strategies.
4. Material and Methods
4.1. Study Area
Sampling was preformed on the intertidal rocky shore within the “Capo Gallo-Isola delle Femmine” MPA (Lat: 38.213961, Long: 13.277121) located in the northwestern coast of Sicily, Italy (Figure 1). The “Capo Gallo-Isola delle Femmine” MPA, established in 2002 by the Italian Ministry of Environment and Protection of Land and Sea, affects the sea stretch between the towns of Palermo and Isola delle Femmine. Covering approximately 22 km2 of sea area and a coastline of about 16 km, it is bounded to the east by the gulf of Mondello and to the west by the bay of Carini. An imposing calcareous dolomitic mountain crest (Capo Gallo, 562 m a.s.l.) defines the coastal strip, resulting in a steep and rocky coastal morphology. Due to the limestone nature, flowing waters generate karst phenomena, leading to caves of high naturalistic value (Grotta dell’Olio and Grotta della Mazzara). Only towards the western part, the rocky coast assumes a flat conformation, enlivened by the presence, about 300 meters from the mainland, of the Isola delle Femmine (also known as Isola di Fuori), an isolated vestige of the aforementioned calcareous ridge.
The MPA is divided into three main zones (A, B and C), each with its own level of environmental protection (Figure 1). There are two no-take/no-access zones (zone A, total area of 1 km2, Figure 1), one in the north sector of Isola delle Femmine and the other in the stretch of sea at the west of Capo Gallo promontory, between the Puntazza and the Capo Gallo lighthouse. Zones B and C are buffer zones where human use restrictions, including fishing, become progressively lower. In particular, there are three general protection zones (zone B, total area of 2 km2, Figure 1), while the remaining sea within the MPA’s border includes a partial protection area (zone C, total area of 19 km2, Figure 1). The MPA area is also identified as Site of Community Importance (SCI, ITA020047 – Fondali di Isola delle Femmine Capo Gallo).
4.2. Sampling
The MPA coastline was divided into sectors of 300 meters each, totaling 36 stations. Each station of 300 m was divided into 12 sectors of 25 meters where sampling was performed visually on 6 random replicates using a 20 x 20 cm quadrat, the minimum area recommended for sampling Mediterranean assemblages in the upper infralittoral zone [31]. In each quadrat the percentage coverage of Cystoseira s.l. species was estimated using the method proposed by Dethier et al. (1993), using a frame divided into 25 equal squares: we attributed a cover score from 0 to 4 to each square, and then summed up scores where the taxon was present. Organisms filling < 1⁄4 square were given the value of 0.5. For E. amentacea a series of morpho-functional characteristics (length of the cauloid, branch length, total length of the thallus, presence/absence and percentage of receptacles, and percentage of burned tissue) were also analysed. All surveys were carried out between May and June 2022, when the thalli of Cystoseira s.l. species in this area reach their maximum development [33].
4.3. Data Analysis
Differences on Cystoseira s.l. species cover percentage among the MPA protection zones (fixed factor with 3 levels: zone A, zone B, and zone C) was tested using one-way Analysis of Variance (ANOVA). Assumptions of normality and homogeneity of variances were tested using Shapiro-Wilk and Levene’s tests, respectively. As regards the morpho-functional parameters of E. amentacea, a Draftsman Plot was first created to check for high correlations among the morpho-functional parameters. Total thallus length, cauloid length, and branch length were found to be highly correlated (cor. > 0.90), as were the presence and percentage of receptacles (cor. = 0.95) (Figure S1, Table S1). Consequently, the following parameters were selected for investigating variations among the MPA’s zones: total thallus length, percentage of receptacles, burnt tissue, and presence of epiphytes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, F.P.M. and A.M.M.; methodology, F.P.M. and A.M.M.; data curation, F.P.M.; investigation, F.P.M. and A.M.M.; visualization, F.P.M.; laboratory analyses, F.P.M. and A.M.M.; writing—original draft, F.P.M., G.S. and A.M.M.; writing—review and editing, F.P.M., G.S. and A.M.M.; supervision, G.S. and A.M.M.; funding acquisition, G.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fondo europeo per gli affari marittimi e la pesca Misura: “1.40—Protezione e ripristino della biodiversità e degli ecosistemi marini e dei regimi di compensazione nell’ambito di attività di pesca sostenibili””, and by the European Union—NextGenerationEU; award number: Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, Project title: “National Biodiversity Future Center—NBFC”. the Ministero della transizione ecologica (MiTE) through the projects “Realizzazione di misure di conservazione del sito di interesse comunitario ITA 020047 “Fondali Isola delle Femmine-Capo Gallo”” and “Valutazione dello stato di conservazione della vegetazione algale nell’AMP di Capo Gallo-Isola delle Femmine and Ministero delle politiche agricole alimentari, forestali e del turismo (Mipaaft) FEAMP PO 2014– 2020
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We thank Gabriella Lo Cicero for the assistance during the sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sala, E.; Mayorga, J.; Bradley, D.; Cabral, R.B.; Atwood, T.B.; Auber, A.; Cheung, W.; Costello, C.; Ferretti, F.; Friedlander, A.M.; et al. Protecting the Global Ocean for Biodiversity, Food and Climate. Nature 2021, 592, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Giakoumi, S.; Scianna, C.; Plass-Johnson, J.; Micheli, F.; Grorud-Colvert, K.; Thiriet, P.; Claudet, J.; Di Carlo, G.; Di Franco, A.; Gaines, S.D.; et al. Ecological Effects of Full and Partial Protection in the Crowded Mediterranean Sea: A Regional Meta-Analysis. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, P.; Baiata, P.; Ballesteros, E.; Di Franco, A.; Hereu, B.; Macpherson, E.; Micheli, F.; Pais, A.; Panzalis, P.; Rosenberg, A.A.; et al. Large-Scale Assessment of Mediterranean Marine Protected Areas Effects on Fish Assemblages. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira Sensu Lato (Fucales, Phaeophyceae) in the Eastern Atlantic and Mediterranean Based on Morphological and DNA Evidence, Including Carpodesmia Gen. Emend. and Treptacantha Gen. Emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Molinari Novoa, E.A.; Guiry, M.D. Reinstatement of the Genera Gongolaria Boehmer and Ericaria Stackhouse (Sargassaceae, Phaeophyceae). Not. Algarum 2020, 172, 1–10. [Google Scholar]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.M.; Francour, P. Nursery Value of Cystoseira Forests for Mediterranean Rocky Reef Fishes. J. Exp. Mar. Bio. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Mineur, F.; Arenas, F.; Assis, J.; Davies, A.J.; Engelen, A.H.; Fernandes, F.; Malta, E.; Thibaut, T.; Van Nguyen, T.; Vaz-Pinto, F.; et al. European Seaweeds under Pressure: Consequences for Communities and Ecosystem Functioning. J. Sea Res. 2015, 98, 91–108. [Google Scholar] [CrossRef]
- Schiel, D.R.; Foster, M.S. The Population Biology of Large Brown Seaweeds: Ecological Consequences of Multiphase Life Histories in Dynamic Coastal Environments. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 343–372. [Google Scholar] [CrossRef]
- Giaccone, G.; Alongi, G.; Pizzuto, F.; Cossu, A.V.L. La Vegetazione Marina Bentonica Fotofila Del Mediterraneo: 2. Infralitorale e Circalitorale: Proposte Di Aggiornamento. Boll. dell’Accademia Gioenia di Sci. Nat. 1994, 27, 111–157. [Google Scholar]
- Falace, A.; Bressan, G. Seasonal Variations of Cystoseira Barbata (Stackhouse) C. Agardh Frond Architecture. Hydrobiologia 2006, 555, 193–206. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Acunto, S.; Cinelli, F.; Hawkins, S.J. The Influence of Canopy Algae on Vertical Patterns of Distribution of Low-Shore Assemblages on Rocky Coasts in the Northwest Mediterranean. J. Exp. Mar. Bio. Ecol. 2002, 267, 89–106. [Google Scholar] [CrossRef]
- Ballesteros, E.; Garrabou, J.; Hereu, B.; Zabala, M.; Cebrian, E.; Sala, E. Deep-Water Stands of Cystoseira Zosteroides C. Agardh (Fucales, Ochrophyta) in the Northwestern Mediterranean: Insights into Assemblage Structure and Population Dynamics. Estuar. Coast. Shelf Sci. 2009, 82, 477–484. [Google Scholar] [CrossRef]
- European Commission Water Framework Directive (2000/60/EC); 2000; Vol. L 269, pp. 1–15.
- Commission, E. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive). Off. J. Eur. Union 2008, 164, 19–40. [Google Scholar]
- Strain, E.M.A.; Thomson, R.J.; Micheli, F.; Mancuso, F.P.; Airoldi, L. Identifying the Interacting Roles of Stressors in Driving the Global Loss of Canopy-Forming to Mat-Forming Algae in Marine Ecosystems. Glob. Chang. Biol. 2014, 20, 3300–3312. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Airoldi, L. Loss, Status and Trends for Coastal Marine Habitats of Europe. In Oceanography and Marine Biology: An Annual Review; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; CRC Press, 2007; Vol. 45, pp. 345–405.
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-Term Decline of the Populations of Fucales (Cystoseira Spp. and Sargassum Spp.) in the Albères Coast (France, North-Western Mediterranean). Mar. Pollut. Bull. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of Vulnerable Cystoseira Populations along the Italian Infralittoral Fringe, and Relationships with Environmental and Anthropogenic Variables. Mar. Pollut. Bull. 2018, 129, 762–771. [Google Scholar] [CrossRef]
- Ling, S.D.; Scheibling, R.E.; Rassweiler, A.; Johnson, C.R.; Shears, N.; Connell, S.D.; Salomon, A.K.; Norderhaug, K.M.; Pérez-Matus, A.; Hernández, J.C.; et al. Global Regime Shift Dynamics of Catastrophic Sea Urchin Overgrazing. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 1–10. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Scheibling, R.E. Sea Urchin Barrens as Alternative Stable States of Collapsed Kelp Ecosystems. Mar. Ecol. Prog. Ser. 2014, 495, 1–25. [Google Scholar] [CrossRef]
- Teagle, H.; Hawkins, S.J.; Moore, P.J.; Smale, D.A. The Role of Kelp Species as Biogenic Habitat Formers in Coastal Marine Ecosystems. J. Exp. Mar. Bio. Ecol. 2017, 492, 81–98. [Google Scholar] [CrossRef]
- Gianni, F.; Bartolini, F.; Airoldi, L.; Ballesteros, E.; Francour, P.; Guidetti, P.; Meinesz, A.; Thibaut, T.; Mangialajo, L. Conservation and Restoration of Marine Forests in the Mediterranean Sea and the Potential Role of Marine Protected Areas. Adv. Oceanogr. Limnol. 2013, 4, 83–101. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Sarà, G.; Mannino, A.M. Conserving Marine Forests: Assessing the Effectiveness of a Marine Protected Area for Cystoseira Sensu Lato Populations in the Central Mediterranean Sea. Plants 2024, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Ramos, E.; Díaz de Terán, J.R.; Puente, A.; Juanes, J.A. The Role of Geomorphology in the Distribution of Intertidal Rocky Macroalgae in the NE Atlantic Region. Estuar. Coast. Shelf Sci. 2016, 179, 90–98. [Google Scholar] [CrossRef]
- Tsiamis, K.; Salomidi, M.; Gerakaris, V.; Mogg, A.O.M.; Porter, E.S.; Sayer, M.D.J.; Küpper, F.C. Macroalgal Vegetation on a North European Artificial Reef (Loch Linnhe, Scotland): Biodiversity, Community Types and Role of Abiotic Factors. J. Appl. Phycol. 2020, 32, 1353–1363. [Google Scholar] [CrossRef]
- Muth, A.F. Effects of Zoospore Aggregation and Substrate Rugosity on Kelp Recruitment Success. J. Phycol. 2012, 48, 1374–1379. [Google Scholar] [CrossRef] [PubMed]
- Gaylord, B.; Reed, D.C.; Washburn, L.; Raimondi, P.T. Physical–Biological Coupling in Spore Dispersal of Kelp Forest Macroalgae. J. Mar. Syst. 2004, 49, 19–39. [Google Scholar] [CrossRef]
- Milazzo, M.; Chemello, R.; Badalamenti, F.; Riggio, S. Short-Term Effect of Human Trampling on the Upper Infralittoral Macroalgae of Ustica Island MPA (Western Mediterranean, Italy). J. Mar. Biol. Assoc. United Kingdom 2002, 82, 745–748. [Google Scholar] [CrossRef]
- Micheli, F.; Heiman, K.W.; Kappel, C. V.; Martone, R.G.; Sethi, S.A.; Osio, G.C.; Fraschetti, S.; Shelton, A.O.; Tanner, J.M. Combined Impacts of Natural and Human Disturbances on Rocky Shore Communities. Ocean Coast. Manag. 2016, 126, 42–50. [Google Scholar] [CrossRef]
- Giakoumi, S.; Pey, A. Assessing the Effects of Marine Protected Areas on Biological Invasions: A Global Review. Front. Mar. Sci. 2017, 4, 1–6. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti-Cecchi, L.; Morri, C.; Pasini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Hard Bottoms. In: GAMBI M.C., DAPPIANO M. (Eds), Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study. In Biol. Mar. Medit.; 2004; Vol. 11 (Suppl., pp. 185–215.
- Dethier, M.; Graham, E.; Cohen, S.; Tear, L. Visual versus Random-Point Percent Cover Estimations: “objective” Is Not Always Better. Mar. Ecol. Prog. Ser. 1993, 96, 93–100. [Google Scholar] [CrossRef]
- Mancuso, F.P.P.; Milazzo, M.; Chemello, R. Decreasing in Patch-Size of Cystoseira Forests Reduces the Diversity of Their Associated Molluscan Assemblage in Mediterranean Rocky Reefs. Estuar. Coast. Shelf Sci. 2021, 250, 107163. [Google Scholar] [CrossRef]
Figure 1.
The MPA of Capo Gallo – Isola delle Femmine. Red, yellow, and blue areas denote no-take (zone A), general (zone B), and limited (zone C) protection zones, respectively.
Figure 1.
The MPA of Capo Gallo – Isola delle Femmine. Red, yellow, and blue areas denote no-take (zone A), general (zone B), and limited (zone C) protection zones, respectively.
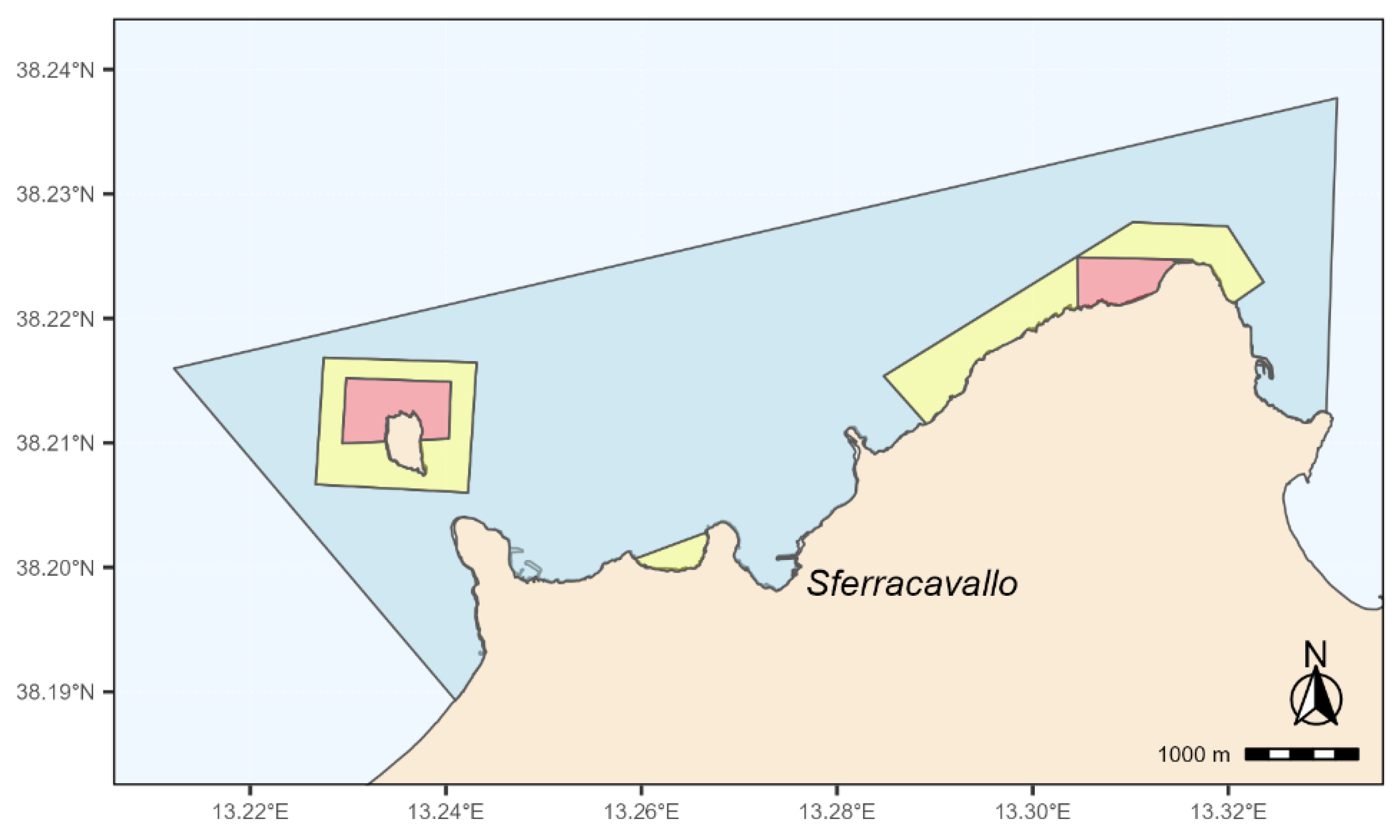
Figure 2.
Cover percentage of Ericaria amentacea (a), Cystoseira compressa (b), Cystoseira foeniculacea (c), Ericaria brachycarpa (d), and Ericaria crinita (e) among the AMP protection zones.
Figure 2.
Cover percentage of Ericaria amentacea (a), Cystoseira compressa (b), Cystoseira foeniculacea (c), Ericaria brachycarpa (d), and Ericaria crinita (e) among the AMP protection zones.
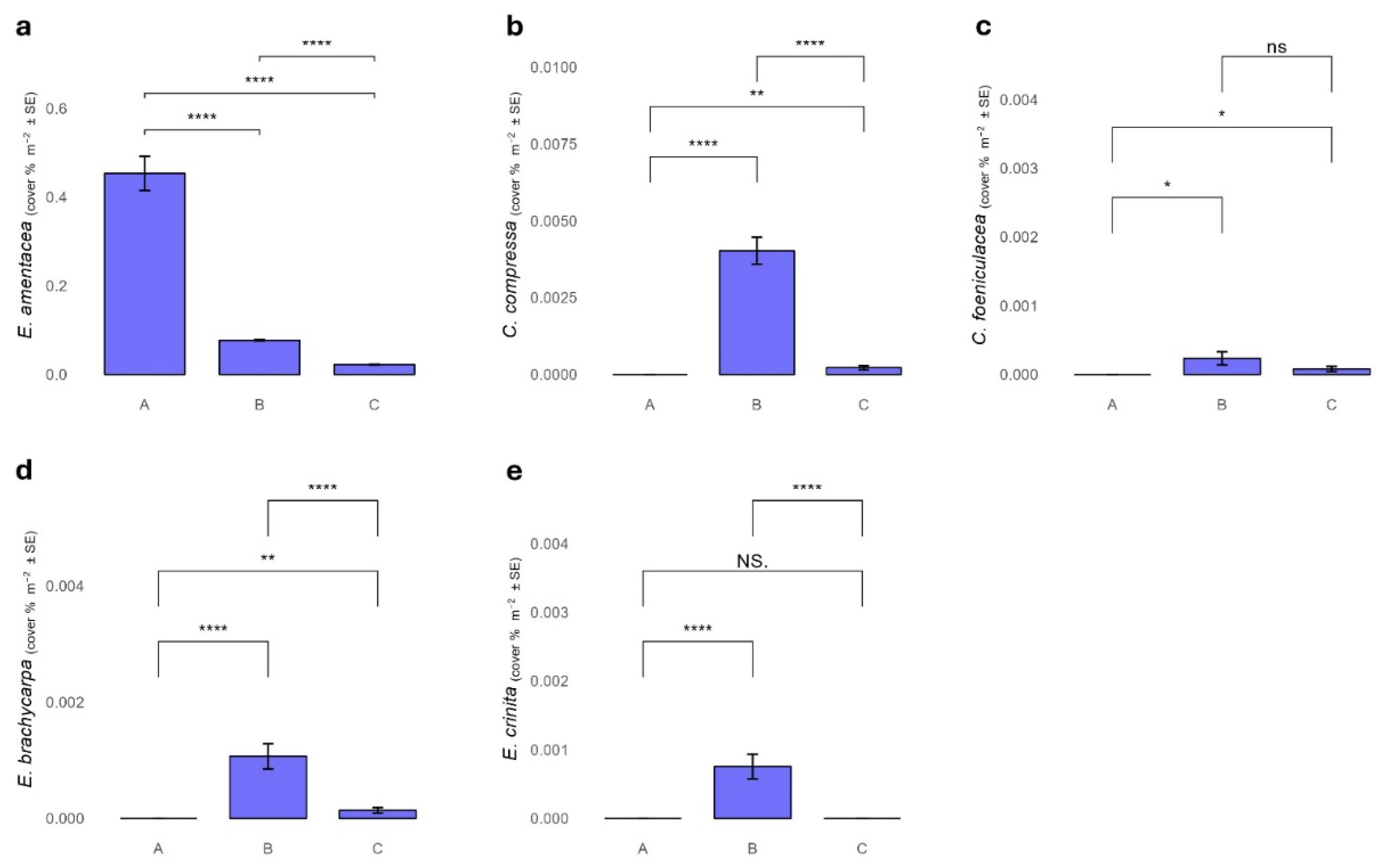
Figure 3.
Variation of the morpho-functional parameters of Ericaria amentacea among the AMP protection zones.
Figure 3.
Variation of the morpho-functional parameters of Ericaria amentacea among the AMP protection zones.
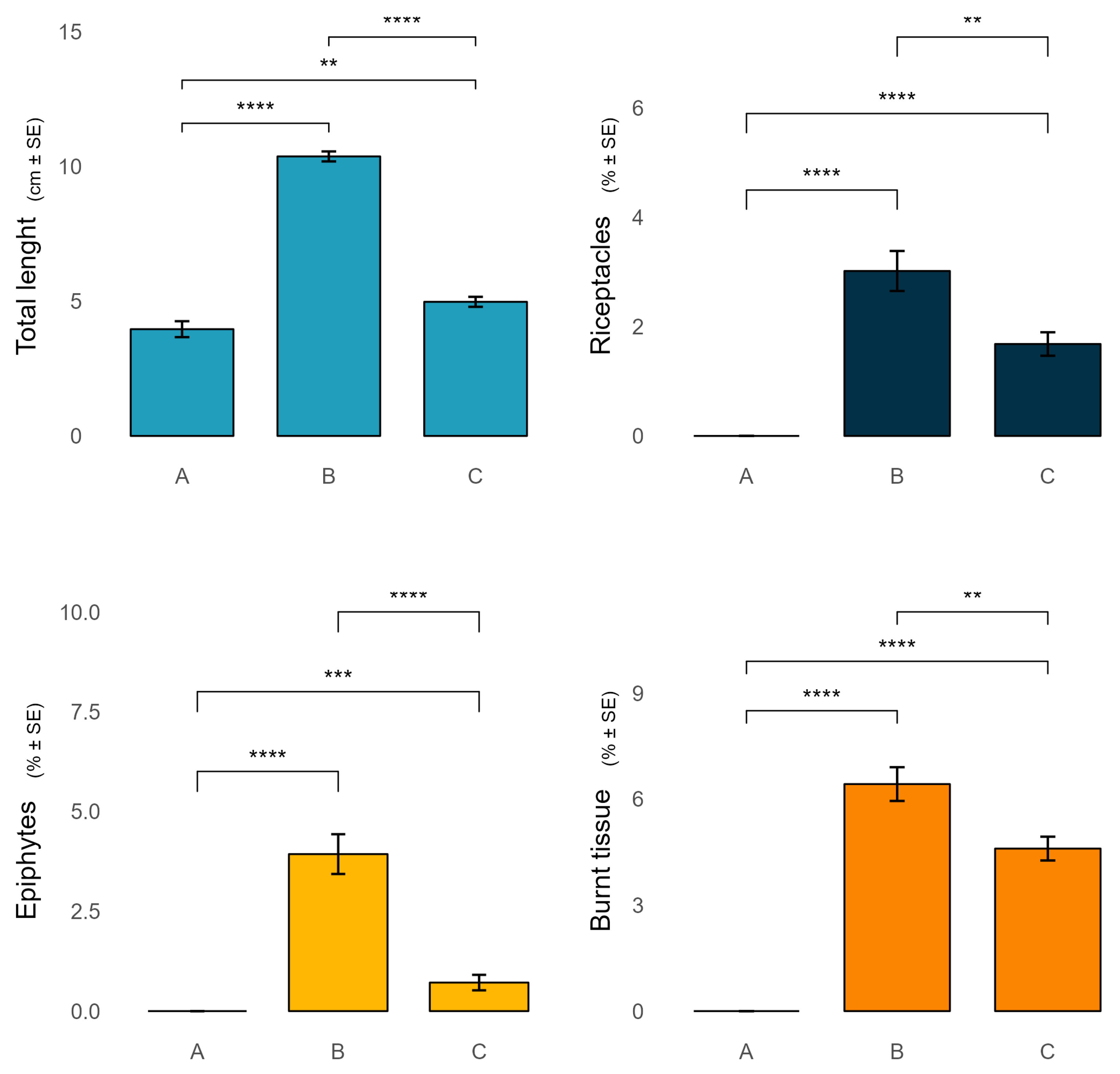
Figure 4.
Cover percentage of Asparagopsis taxiformis (red), and Caulerpa cylindracea (green), among the AMP protection zones.
Figure 4.
Cover percentage of Asparagopsis taxiformis (red), and Caulerpa cylindracea (green), among the AMP protection zones.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



