
Submitted:
19 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
Chronic kidney disease (CKD) is increasingly common in older cats. The transforming growth factor-beta (TGF-β) pathway is associated with renal fibrosis. This study aimed to quantify the mRNA expression of TGFβ, MAPK, and Bcl2 genes and the protein expression of TGF-β and MAPK in feline kidney cells and tissues. Gene expression analysis was conducted using relative gene expression, while protein expression was assessed through western blot analysis. The immunohistochemistry staining of TGF-β and MAPK was performed on feline kidney tissues. The result reveals the significant upregulation of TGFβ (P = 0.001) and considerable downregulation of Bcl2 (P = 0.010) in doxorubicin-treated feline kidney cells. Protein expression level of TGF-β and MAPK also tended to increase in doxorubicin-induced feline kidney cells. The immunostaining levels of TGF-β and MAPK were higher in the kidney tissues of cats with CKD compared to the no lesions group. A deeper understanding of the TGF-β pathway could enable veterinarians to monitor disease progression and mitigate complications in feline CKD.
Keywords:
Bcl-2
; cat
; chronic kidney disease
; mitogen-activated protein kinase
; transforming growth factor-β
1. Introduction
Chronic kidney disease (CKD) is an important disease in cats. The definition of CKD is an abnormality in the anatomy or function of one or both kidneys that persist for at least three months [1]. The prevalences of feline CKD were different in each country, with rate of 3.6% in the United Kingdom [2], 2.37 % in Chiang Mai, Thailand [3], and 50% in the United States [4]. Moreover, the prevalence increased with age; 6 months-5 years was 37.5%, 5-10 years was 40.9%, 10-15 years was 42.1%, and 15-20 years was 80.9% [2]. The glomerular filtration rate (GFR) measurement is the gold standard of renal function assessment. Nevertheless, this method is complicated to perform in routine health checks. The guideline for the feline CKD diagnosis was suggested by the International Renal Interest Society (IRIS). They suggest routine investigation of serum creatinine, blood urea nitrogen (BUN), urinalysis, symmetric dimethylarginine (SDMA), and ultrasonography. Criteria for CKD consist of creatinine greater than 1.6 mg/dl and/or inappropriate urine (USG less than 1.035) for several months [5]. The most common lesion of feline CKD is a tubulointerstitial lesion, including interstitial fibrosis, interstitial inflammation, tubular mineralization, and hyperplastic arteriolosclerosis. Interstitial fibrosis score was found to be increased with the IRIS stage [6]. In the renal fibrosis process, there are four pro-fibrotic mediators, including TGF-β, transglutaminase 2 (TG2), endothelin 1 (ET1), and the renin-angiotensin-aldosterone system (RAAS) components. The most crucial pro-fibrotic mediator is TGF-β, which changes various types of cells to myofibroblasts [7].
Transforming growth factor-beta (TGF-β) plays a role in regulating cell proliferation as either an inhibitor or stimulator, depending on signaling pathways. In addition, TGF-β is involved in wound healing, differentiation of cells, and the immune system. It is also associated with pathological processes, including connective tissue disorder, fibrosis, and cancer [8]. The TGF- β signaling pathway consists of 2 pathways: smad-dependent/canonical pathway and smad-independent/non-canonical pathway [9]. TGF-β is associated with apoptosis through these two pathways: the smad-dependent pathway affects BCL-2 family expression, and the smad-independent pathway induces apoptosis by activating p38 and JNK pathways [10]. In cats, previous study demonstrated that the urinary TGF-β1: creatinine ratio was significantly enhanced in CKD cats compared to healthy cats and positively correlated with creatinine in serum [11]. The study of feline kidney tissue found that TGF-β immunohistochemistry staining in CKD cats was stronger than in healthy cats [12]. While TGF-β concentration in CKD cats’ blood was significantly less than in healthy cats. Moreover, they found that cats with lower circulating TGF-β had shorter life spans than those with higher circulating TGF-β [13]. The Mitogen-activated protein kinase (MAPK) is an enzyme that functions as a serine/threonine protein kinase. The MAPK was controlled by various extracellular stimuli, which then caused gene expression, cell division, and cell survival. The MAPK pathway contributes to acute kidney injury (AKI) and CKD [14]. Previous studies indicated the association between the MAPK pathway and renal fibrosis [15,30]. In mice, renal artery stenosis (RAS) surgery is a technique for inducing tubular atrophy and interstitial fibrosis. The result revealed increasing p38 MAPK signaling in mice undergoing RAS surgery compared to sham mice. Moreover, blocking p38 MAPK with the inhibitors in RAS mice led to a reduction in interstitial inflammation, interstitial fibrosis, and tubular atrophy compared to RAS without p38 MAPK inhibitors [15]. Furthermore, the MAPK pathway is involved in the apoptosis process, with the p38 MAPK and JNK pathways regulating the Bcl2 family [16]. The B-cell lymphoma 2 (Bcl-2) protein family plays a role in cell growth and apoptosis regulation. It is found to be associated with renal cell death during kidney injury [17]. There are three classes of the Bcl-2 protein family: anti-apoptosis (Bcl-XL, Bcl-2), pro-apoptosis (Bak, Bax), and BH3-only proteins (Bid, Bad) [18]. In cats, Bcl-2 expression in the blood of the CKD cats was significantly lower than in healthy cats. Bcl-2 immunoreactivity in feline renal tissues was found in glomerular and tubular epithelium cells. Previous studies suggested that Bcl-2 was an acceptable diagnostic marker for feline CKD. It was also indicated that higher CKD stages were associated with lower Bcl-2 concentration [19].
However, there were few studies of the TGF-β signaling pathway, which involves MAPK and Bcl-2, in cats with chronic kidney disease. This study aimed to evaluate TGFβ, MAPK, and Bcl2 gene expression in doxorubicin-induced feline kidney cells and kidney tissues of cats with naturally occurring CKD and to determine TGF-β and MAPK protein expression in doxorubicin-treated feline kidney cells.
2. Materials and Methods
2.1. Sample
Feline kidney cell line
The Cradle Rees Feline Kidney (CRFK) cell line was procured from ATCC® (CCL-94), LOT 60980362. These cells originate from the cortex epithelium of the kidney of 12-week-old female domestic cats. Cells were grown in Dulbecco’s Modified Eagle Medium (DMEM), 5% fetal bovine serum (FBS), 100 µg/ml streptomycin, 100 units/ml penicillin, and non-essential amino acids solution and incubated at 37 °C in 5% CO2 in a humidified incubator. The cells used in this study were in passages 125-132 passages.
Kidney tissues
Kidney tissues were collected from cats with no kidney lesions (n = 6) and cats with CKD (n = 6) that died at the Small Animal Hospital, Faculty of Veterinary Medicine, Chiang Mai University. The samples were preserved in 10% formalin or frozen at -20 °C to keep them fresh. Paraffin block samples were prepared from 10% formalin-preserving tissues, cut into slides, and stained with Hematoxylin and Eosin. A pathologist examined the histological morphologies and categorized them as no kidney lesions or CKD. The Ethics Committee approved the Human and Animal Experimentation protocol, Faculty of Veterinary Medicine, Chiang Mai University, under the reference number S1/2566.
2.2. Sample Collection
Cytotoxicity test of doxorubicin-induced CRFK
CRFK cells were incubated with different concentrations and durations of doxorubicin (DOX) (Catalogue no. 324380). The concentrations of DOX used were 0, 1, 2, 4, and 8 µM, and the cells were incubated for 48, 72, and 96 hours [20]. The CRFK cells were grown in a 96-wells plate and kept overnight in a 37 °C, 5% CO2 humidified incubator before adding DOX. Cell viability was determined using MTT colorimetric assay (Mosmann, 1983) [21]. The study applied the appropriate dose and duration that could decrease cell viability by 50%.
DOX-induced cytotoxicity test
CRFK cells were seeded into 6-well plates and then incubated at 37 °C in a humidified 5% CO2 incubator overnight. The appropriate dose and duration of DOX were 8 µM for 48 hours. Next, DOX 8 µM was added to CRFK cells and incubated for 48 hours. After that, feline kidney cells with DOX-induced cytotoxicity were evaluated for the mRNA expression of TGFβ, MAPK, and Bcl2.
RNA extraction
The RNA was harvested from CRFK cells and kidney tissues. CRFK cells were grown in a 6-well plate before harvesting the RNA. RNA was extracted using TRIzol® reagent. First, the medium was removed, and then TRIzol® reagent was added to the plate for 15 minutes. The extraction was then transferred to the Eppendorf tube and centrifuged at 4 °C 12,000 rpm for 3 minutes. The supernatant was removed, and then isopropanol was added at room temperature for 10 minutes. The mixture was centrifuged at 4 °C 12,000 rpm for 3 minutes. The supernatant was removed, and 75% ethanol was added and centrifuged at 4 °C 8,000 rpm for 3 minutes. The supernatant was removed, and the RNA pellet was mixed with RNase-free water and transferred to a new Eppendorf tube. The RNA samples were then stored at -80 °C. For the kidney tissues, the kidney tissues were ground in liquid nitrogen and homogenized with TRIzol® reagent. Liquid sample was mixed with TRIzol® reagent at room temperature for 15 minutes. The process of RNA extraction was the same as cell line. The RNA quantification was measured using a NanoDrop spectrophotometer.
Protein extraction
The protein was extracted from CRFK cells in a 6-well plate. The growth medium was removed, and the cells were washed with PBS. Next, trypsin was applied to detach cells, and then growth medium was added. The samples were transferred to Eppendorf tubes and centrifuged at 4 °C at 1,200 rpm for 10 minutes. The supernatant was removed, and RIPA (lysis buffer) was added and left in an ice container for 30 minutes. The samples were centrifuged at 4 °C at 12,000 rpm for 10 minutes. The supernatant was collected and stored at -80 °C. Protein quantification was measured using the Bradford assay. The Bradford solution (Catalogue no. 500-0006) was diluted at 1:3 with PBS. The diluted-Bradford was added to the protein samples in 96-well plates. Then, protein lysates were added to each well and incubated at room temperature for 10 minutes. The protein quantification was assessed at 540 nm.
2.3. Sample Processing
Relative gene expression
The total RNA from CRFK cells and kidney tissues was extracted using TRIzol® reagent (Catalogue no. R4533). Real-time polymerase chain reaction was performed using iQ5 real-time PCR with IO SYBR green supermix (Catalogue no. 08-24-00001). The qPCR protocol involved an initial polymerase activation step to 95 °C for 12 min, followed by 40 cycles of denaturation at 95 °C for 15 sec and annealing/extension at 60°C for 30 sec. Data acquisition was performed at the end of the annealing/extension step. All reactions were conducted in triplicate using 96-well reaction plates (2 µl per reaction). In this study, the housekeeping gene was β-actin. The mRNA expression levels of TGFβ, MAPK, and Bcl2 were calculated as relative gene expression ratio with β-actin using 2˗∆CT. The primer sequences used in this study are detailed in Table 1.
Western blot analysis
The protein samples at a concentration of 30 µl were mixed with 2✕Laemmli sample buffer (Bio-Rad) containing β-mercaptoethanol and heated at 95 °C for 5 min. Next, proteins were separated on 12% SDS-PAGE gels and transferred onto a polyvinylidene fluoride (PVDF) membrane 0.45 µm pores (Bio-Rad), utilizing a Trans-Blot® SD Semi-Dry Transfer Cell (Bio-Rad). Next, the membrane was washed with Tris-buffered saline with 0.05% Tween (TBST) and blocked with BSA at room temperature for 45 minutes. The membrane was incubated for 2 hours with primary antibodies for TGF-β, MAPK, or β-actin (direct-blot HRP mouse monoclonal anti-β-actin, BIOL-643808, BioLegend) and washed with TBST. After that, it was incubated with a secondary antibody labeled as HRP-conjugated antibody for 45 minutes at room temperature. The primary and secondary antibodies are shown in Table 2. The signal detection was performed using a DAB substrate (Bio Basic). Protein quantifying densitometry was determined using Image StudioTM Lite (LI-COR, Lincoln, Nebraska, USA) and then calculated as the ratio of TGF-β or MAPK to β-actin as loading control.
Immunohistochemistry
The kidney tissues from dead cats were collected through necropsy. The tissues were fixed in 10% formalin, and embedded in paraffin, then sliced into sections of 2-7 µm thickness using a rotary microtome. Deparaffinization was conducted using xylene, followed by 100% ethanol and 95% ethanol before staining. For heat-induced epitope retrieval (HIER), microwave 800 W was used for 20 minutes with sodium citrate buffer at pH 6. The tissue area was outlined using an immunohistochemistry pen, and 0.3% hydrogen peroxidase (H2O2) was applied for 5 minutes. The slides were washed with PBST for 5 minutes, three times. Next, 2.5% BSA in PBS was added to the samples at room temperature for 5 minutes and washed with PBST for 5 minutes, three times. Immunohistochemical staining was performed by incubating the tissue section with the primary antibodies at 37 °C for 2 hours, followed by washing with PBS for 5 minutes, three times. The tissue section was incubated with a dilution of PBS with normal goat serum (1:5) for 30 minutes and washed with PBS for 5 minutes, three times. The secondary antibodies were incubated at room temperature in a dark room for 45 minutes and washed with PBS for 5 minutes, three times. DAB solution was used to develop the staining at room temperature for 5 minutes, followed by washing the slide with tap water and staining with Hematoxylin and Lithium. The primary and secondary antibodies are shown in Table 2. CaseViewer program was used to evaluate the quality of immunoreactivity.
2.1. Statistical Analysis
The data were presented as either mean ± standard deviation (SD) or median. We conducted a normality test using the Shapiro-Wilk test. The one-way analysis of variance (ANOVA) with the Bonferroni post hoc test was used to compare the mean between groups. An unpaired t-test or Welch two-sample t-test was performed to compare the mean between the two groups. Each experiment consisted of at least n = 5, where n was the number of monolayers or cat kidney tissues. A P-value less than 0.05 was considered significant.
3. Results
3.1. Cytotoxic Test of Doxorubicin-Induced Cytotoxicity in Feline Kidney Cells
The cell viability of CRFK was measured after being treated with DOX at concentration of 1, 2, 4, and 8 µM for 48, 72, and 96 hours. The results revealed that after 48 hours, the cell viability was 83.08%, 76.78%, 64.87%, and 53.83% for the respective DOX concentrations (Figure S1). After 72 hours, the cell viability percentages were 111.35%, 97.53%, 107.04%, and 59.37% for the corresponding DOX concentrations (Figure S2). The cell viability after 96 hours of treatment was 82.97%, 68.93%, 62.61%, and 61.04% for the respective DOX concentration (Figure S3). Our study focused on using DOX at a concentration of 8 µM for 48 hours.
3.2. TGFβ, MAPK, and Bcl2 Relative Gene Expression in Feline Kidney Cells
The results revealed that the mean fold-change of TGFβ gene expression in the DOX-treated group was 4.084. The difference in TGFβ gene expression fold change between the control and the DOX-treated group was 3.084 ± 0.668 (Figure 1 and Figure 2). TGFβ gene expression in the DOX-treated group was significantly higher compared to the control group. The mean fold-change of MAPK gene expression in the DOX-treated group was 1.072. The difference in MAPK gene expression fold-change between the control and the DOX-treated group was 0.072 ± 0.509 (Figure 1 and Figure 2). MAPK gene expression in the DOX-treated group did not differ from the control group. The mean fold-change of Bcl2 gene expression in the DOX group was 0.596. The difference in Bcl2 gene expression fold-change between the control and the DOX-treated group was -0.404 ± 0.125 (Figure 1 and Figure 2). Bcl2 gene expression in the DOX-treated group was significantly lower than in the control group. These results of TGFβ, MAPK, and Bcl2 mRNA expression are available in Table 3.
3.3. TGFβ, MAPK, and Bcl2 Relative Gene Expression in Kidney Tissues
The fold-change of TGFβ, MAPK, and Bcl2 gene expression were 0.563, 0.925, and 0.683, respectively (Figure 3). The difference of TGFβ, MAPK, and Bcl2 gene expression between cats with no kidney lesions and cats with CKD were -0.437 ± 0.222, -0.075 ± 0.373, and -0.317 ± 0.251, respectively (Figure 4). TGFβ and Bcl2 gene expressions in CKD cats were slightly lower than in cats with no kidney lesions. However, the MAPK gene expression in the CKD cats was not significantly different from that in the cats with no kidney lesions. The results of gene expression in kidney tissues are shown in Table 4.
3.4. Protein Expression of TGF-β and MAPK in Feline Kidney Cells
The protein expression of TGF-β was a trend to increase in the doxorubicin-treated group (0.06 ± 0.07) compared to the control group (0.04 ± 0.05) (Figure 5). MAPK protein expression of the doxorubicin group was 2.28 ± 2.64, and the control group was 0.44 ± 0.14. The protein expression of MAPK was a trend to increase in the doxorubicin-treated group (Figure 6). The results are shown in Table 5.
3.5. Immunohistochemistry of TGF-β and MAPK in Cat Kidney Tissues
4. Discussion
Doxorubicin can lead to oxidative stress in glomerular epithelial cells [22] and result in kidney damage [23]. An eight µM dose of DOX for 48 hours was determined to be the appropriate dosage and duration to induce cytotoxicity. This finding is consistent with a previous study in CRFK cells, which demonstrated a 50% decrease in cell viability of eight µM doxorubicin for 48 hours [24]. It is similar to the study of human kidney cells. The cell viability of HK-2 cells significantly decreased at a doxorubicin concentration of 8 µM for 24 hours, dropping to nearly 50% [20,25].
The significant lesion in CKD cats is tubulointerstitial fibrosis [6]. However, tubular atrophy is also markedly found in the kidneys of CKD cats [13]. Hence, fibrosis and apoptosis are associated with feline CKD development. The pro-fibrotic mediator associated with renal fibrosis is TGF-β [7]. The oxidative stress stimulates TGF-β production [26]. A TGF-β signaling pathway is composed of 2 cascades: smad-dependent (canonical) pathway and smad-independent (non-canonical) pathway [9]. Our study demonstrated the expression of the TGFβ gene significantly increased in the DOX-treated group compared to the control group. In feline kidney cells, a previous study found an increase in α-SMA, CTGF, TNC, TSP-1, and COL1 gene expression in the TGF-β-treated group [27]. In rat kidney cells, TGF-β treated group had higher α-SMA, COL1, and COL3 than the control group [28]. The previous studies in kidney cells demonstrated the pro-fibrotic mediator effect of TGF-β [27,28]. These results may explain that fibrosis was related to oxidative stress in feline kidney cytotoxicity.
The present study found that TGFβ gene expression in kidney tissues of CKD cats was lower than in cats with no kidney lesions. The gene expression of TGFβ revealed higher in the kidney tissues of mice with unilateral ureteral obstruction (UUO) when compared to sham mice [29]. In diabetic nephropathy mice, TGFβ mRNA expression in kidney tissue of diabetic mice with blood glucose fluctuation (BGF) was the highest compared to diabetic mice without BGF and control mice [30]. This difference in RNA stability in tissue can be attributed to temperature variations and collection procedures. It is important to note that the stability of RNA in the tissue is influenced by temperature [31]. A post-mortem study demonstrated that human kidney tissues were unsuitable for RNA evaluation [32]. Only one sample in our study was obtained via nephrectomy, while the other tissues were collected post-mortem. Urine TGF-β: creatinine in cats was correlated with fibrotic severity [33]. However, the kidney tissue in the immunohistochemistry part was derived from a cat with CKD stage II that may exhibit mild fibrosis. The CKD stage III tissue sample was from a cat with polycystic kidney disease (PKD) and severe subcapsular effusion. These kidney tissues in the CKD stage III group were compressed by fluid, possibly leading to reduced TGFβ gene expression.
Our study demonstrated that TGF-β protein expression in feline kidney cells tended to increase in the DOX-treated group compared to the control group. Immunohistochemistry of TGF-β in kidney tissue also indicated positive immunostaining in the tubular area of CKD cats. In agreement with a previous study, TGF-β immunostaining was predominantly located in the distal tubules and collecting ducts of kidney tissue of cats with CKD [13]. In contrast, normal feline kidney tissue showed weak immunostaining of TGF-β [12]. Additionally, kidneys tissues from patients experiencing renal fibrosis showed higher levels of TGF-β and α-SMA immunostaining compared to negative control [29]. The previous study reveals that the urine TGF-β to creatinine ratio was significantly higher in CKD cats than in healthy ones [11]. Moreover, a moderate correlation was identified between the urine TGF-β to creatinine ratio and interstitial fibrosis [33]. Analysis of TGF-β levels in the blood of cats indicated that the TGF-β concentration in those with CKD was significantly lower than in healthy cats. Furthermore, cats with low circulating TGF-β concentrations had shorter survival times [13]. Therefore, targeting the TGF-β pathway is important for developing therapies aimed at preventing the progression of fibrosis in the kidney [34]. In a previous study, IN-1130 showed potential in reducing renal fibrosis in rats subjected to UUO by inhibiting the TGF-β pathway [35].
Our study demonstrated that the MAPK gene expression response to DOX-induced cytotoxicity. MAPK protein expression in the DOX-treated cells was a trend to increase compared to the control cells. MAPK plays a role in signaling transmission of extracellular stimuli to intracellular response and apoptosis [16] and oxidative stress [36]. In cardiac cells, phosphorylated p38 MAPK protein expression was significantly increased in DOX-induced H9c2 cardiac cells. However, total p38 MAPK in DOX-induced cardiac cells was not different from control cells [37]. Moreover, the MAPK signaling pathway can be stimulated by TGF-β through TAK-1/MKK6/p38MAPK pathway [38], which was categorized as a TGF-β smad-independent pathway [39]. In a previous study, mice undergoing unilateral ischemia-reperfusion injury had higher TGFβ and p38MAPK mRNA expression than sham mice [40]. Our study also indicated that MAPK immunoreactivity in the kidney tissue of CKD cats was higher than in cats with no kidney lesions. Several previous studies revealed a high level of MAPK immunoreactivity in other species with kidney problems [41,42]. In patients with IgA nephropathy, p-p38MAPK immunostaining was predominantly in high-grade renal fibrosis [41]. In mice models, p-p38MAPK immunostaining was eminently in UUO mice [40]. In experimental nephrotic syndrome, p-p38MAPK staining was increased in podocyte, parietal epithelial cells, and glomeruli [42]. Therefore, MAPK may be associated with feline kidney disease through hypoxia and renal fibrosis, especially the TGF-β pathway.
The present study revealed that Bcl2 gene expression in the DOX-treated group was significantly lower than in the control group. Moreover, the gene expression of Bcl2 in the kidney tissue of CKD cats was a trend to decrease compared to cats with no kidney lesions. In the experiment-induced apoptosis, there was downregulation of Bcl-2 and upregulation of p38MAPK and Bax expression [43,44]. Additionally, the level of Bcl-2 in cats with CKD was lower than in healthy cats [19]. The study in leukemia cells found that treating cells with TGF-β promoted cell death, increased Bax, and deceased Bcl-2 expression [45]. In patients with glomerulonephritis, TGF-β had a negative correlation with Bcl-2 but a positive correlation with Bax [46]. These results may explain that apoptosis was related to feline kidney cytotoxicity and CKD development in cats. Bcl2 was regulated by the TGF-β smad-dependent pathway [39], and the downregulation of Bcl-2 may regulate apoptosis in feline CKD [19].
Oxidative stress, renal fibrosis, and apoptosis play crucial roles in the progression of feline CKD [7,29,47]. The present study provided information on the TGF-β mediator that may be associated with renal fibrosis and apoptosis in feline CKD. Our study demonstrated the TGF-β mediator gene and protein expression in cats with naturally occurring CKD. This study may lead to further investigation into therapeutic medication that can delay renal fibrosis and apoptosis in CKD progression and benefit in prolonging the life span of cats with CKD. However, this study has limitations, including a small sample size of kidney tissue and incomplete blood profile data. Obtaining more blood samples and increasing the number of tissue samples could provide valuable information about these mediators.
5. Conclusions
The current study revealed that TGFβ was upregulated in feline kidney cells but downregulated in feline kidney tissues. Additionally, Bcl2 was downregulated in both feline kidney cells and tissues. However, there was no significant change in MAPK gene expression in feline kidney cells and tissues. Protein expression of TGF-β and MAPK in the DOX-treated cells showed a trend to increase compared to control. The immunoreactivity of TGF-β and MAPK in kidney tissues of cats with CKD was highly stained compared to healthy tissue. This result indicated that TGF-β, MAPK, and Bcl-2 may regulate renal fibrosis and the progression of apoptosis in feline kidney cells (Figure 8). There have been studies on TGF-β inhibitors in organ fibrosis. However, knowledge regarding TGF-β inhibitors in renal fibrosis among animals is limited. TGF-β inhibitors have not been studied in cats with chronic kidney disease (CKD). Further research on these mediators could lead to the development of therapeutic medications that delay the progression of CKD.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Supplementary Materials
The following supporting information can be downloaded at: the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.P. and P.C.; methodology, C.I., K.P., N.M. and P.C.; software, C.I., K.P. and P.C.; validation, C.I., K.P., N.M. and P.C.; formal analysis, C.I., K.P. and P.C.; investigation, C.I., K.P., N.M. and P.C.; resources, C.I., B.S., K.P., N.M. and P.C.; data curation, K.P., P.C. and P.P.; writing—original draft preparation, C.I.; writing—review and editing, K.P and P.C.; visualization, C.I. and K.P.; supervision, K.P. and P.C.; project administration, K.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee approved the Human and Animal Experimentation protocol, Faculty of Veterinary Medicine, Chiang Mai University (protocol code S1/2566 and February 15, 2023).
Informed Consent Statement
Not applicable
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Polzin, D.J. Chronic kidney disease in small animals. Veterinary Clinics: Small Animal Practice 2011, 41, 15–30. [Google Scholar] [CrossRef] [PubMed]
- O'neill, D.; Church, D.; McGreevy, P.; Thomson, P.; Brodbelt, D. Prevalence of disorders recorded in cats attending primary-care veterinary practices in England. The Veterinary Journal 2014, 202, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Piyarungsri, K.; Tangtrongsup, S.; Thongtharb, A.; Sodarat, C.; Bussayapalakorn, K. The risk factors of having infected feline leukemia virus or feline immunodeficiency virus for feline naturally occurring chronic kidney disease. Veterinary Integrative Sciences 2020, 18, 119–131. [Google Scholar]
- Marino, C.L.; Lascelles, B.D.X.; Vaden, S.L.; Gruen, M.E.; Marks, S.L. Prevalence and classification of chronic kidney disease in cats randomly selected from four age groups and in cats recruited for degenerative joint disease studies. Journal of feline medicine and surgery 2014, 16, 465–472. [Google Scholar] [CrossRef]
- Sparkes, A.H.; Caney, S.; Chalhoub, S.; Elliott, J.; Finch, N.; Gajanayake, I.; Langston, C.; Lefebvre, H.P.; White, J.; Quimby, J. ISFM consensus guidelines on the diagnosis and management of feline chronic kidney disease. Journal of feline medicine and surgery 2016, 18, 219–239. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Syme, H.; Brown, C.; Elliott, J. Histomorphometry of feline chronic kidney disease and correlation with markers of renal dysfunction. Veterinary pathology 2013, 50, 147–155. [Google Scholar] [CrossRef]
- Lawson, J.; Elliott, J.; Wheeler-Jones, C.; Syme, H.; Jepson, R. Renal fibrosis in feline chronic kidney disease: known mediators and mechanisms of injury. The Veterinary Journal 2015, 203, 18–26. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β family: context-dependent roles in cell and tissue physiology. Cold Spring Harbor perspectives in biology 2016, 8, a021873. [Google Scholar] [CrossRef]
- Gordeeva, O. TGFβ family signaling pathways in pluripotent and teratocarcinoma stem cells’ fate decisions: Balancing between self-renewal, differentiation, and cancer. Cells 2019, 8, 1500. [Google Scholar] [CrossRef]
- Bartholin, L.; Vincent, D.F.; Valcourt, U. TGF-β as tumor suppressor: in vitro mechanistic aspects of growth inhibition. TGF-β in Human Disease 2013, 113-138.
- Habenicht, L.M.; Webb, T.L.; Clauss, L.A.; Dow, S.W.; Quimby, J.M. Urinary cytokine levels in apparently healthy cats and cats with chronic kidney disease. Journal of feline medicine and surgery 2013, 15, 99–104. [Google Scholar] [CrossRef]
- Uehara, Y.; Furusawa, Y.; Islam, M.S.; Yamato, O.; Hatai, H.; Ichii, O.; Yabuki, A. Immunohistochemical Expression of TGF-β1 in Kidneys of Cats with Chronic Kidney Disease. Veterinary Sciences 2022, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Piyarungsri, K.; Chuammitri, P.; Pringproa, K.; Pila, P.; Srivorakul, S.; Sornpet, B.; Pusoonthornthum, R. Decreased circulating transforming growth factor-beta (TGF-β) and kidney TGF-β immunoreactivity predict renal disease in cats with naturally occurring chronic kidney disease. Journal of feline medicine and surgery 2023, 25, 1098612X231208937. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, H.; Radford, R.; Slyne, J.; O’Connell, S.; Slattery, C.; Ryan, M.P.; McMorrow, T. The role of MAPK in drug-induced kidney injury. Journal of signal transduction 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Warner, G.M.; Yin, P.; Knudsen, B.E.; Cheng, J.; Butters, K.A.; Lien, K.R.; Gray, C.E.; Garovic, V.D.; Lerman, L.O. Inhibition of p38 MAPK attenuates renal atrophy and fibrosis in a murine renal artery stenosis model. American Journal of Physiology-Renal Physiology 2013, 304, F938–F947. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK signaling pathways in apoptosis. International journal of molecular sciences 2020, 21, 2346. [Google Scholar] [CrossRef]
- Borkan, S.C. The role of BCL-2 family members in acute kidney injury. Seminars in nephrology 2016. [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: opposing activities that mediate cell death. Nature reviews Molecular cell biology 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Pila, P.; Chuammitri, P.; Patchanee, P.; Pringproa, K.; Piyarungsri, K. Evaluation of Bcl-2 as a marker for chronic kidney disease prediction in cats. Frontiers in Veterinary Science 2023, 9, 1043848. [Google Scholar] [CrossRef]
- Chaotham, C.; De-Eknamkul, W.; Chanvorachote, P. Protective effect of plaunotol against doxorubicin-induced renal cell death. Journal of natural medicines 2013, 67, 311–319. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of immunological methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Ghiggeri, G.M.; Bertelli, R.; Ginevri, F.; Oleggini, R.; Altieri, P.; Trivelli, A.; Gusmano, R. Multiple mechanisms for doxorubicin cytotoxicity on glomerular epithelial cells ‘in vitro’. European Journal of Pharmacology: Environmental Toxicology and Pharmacology 1992, 228, 77–83. [Google Scholar] [CrossRef]
- Wu, Q.; Li, W.; Zhao, J.; Sun, W.; Yang, Q.; Chen, C.; Xia, P.; Zhu, J.; Zhou, Y.; Huang, G. Apigenin ameliorates doxorubicin-induced renal injury via inhibition of oxidative stress and inflammation. Biomedicine Pharmacotherapy 2021, 137, 111308. [Google Scholar] [CrossRef] [PubMed]
- Piyarungsri, K.; Pusoonthornthum, R.; Rungsipipat, A.; Sritularak, B. (2014). INVESTIGATION OF RISK FACTORS INVOLVING IN FELINE CHRONIC KIDNEY DISEASE, OXI DATIVE STRESS AND STUDY THE EFFECT OF ANTIDESMA ACIDUM CRUDE EXTRACT IN F ELINE KIDNEY CELL LINE Doctoral thesis, Chulalongkorn University].
- Park, E.J.; Kwon, H.K.; Choi, Y.M.; Shin, H.J.; Choi, S. Doxorubicin induces cytotoxicity through upregulation of pERK-dependent ATF3. PLoS One 2012, 7, e44990. [Google Scholar] [CrossRef]
- Shin, D.-M.; Jeon, J.-H.; Kim, C.-W.; Cho, S.-Y.; Lee, H.-J.; Jang, G.-Y.; Jeong, E.M.; Lee, D.-S.; Kang, J.-H.; Melino, G. TGFβ mediates activation of transglutaminase 2 in response to oxidative stress that leads to protein aggregation. The FASEB Journal 2008, 22, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- van Beusekom, C.D.; Zimmering, T.M. Profibrotic effects of angiotensin II and transforming growth factor beta on feline kidney epithelial cells. Journal of feline medicine and surgery 2019, 21, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wu, G.; Dai, T.; Lang, Y.; Chi, Z.; Yang, S.; Dong, D. Naringin attenuates renal interstitial fibrosis by regulating the TGF-β/Smad signaling pathway and inflammation. Experimental and Therapeutic Medicine 2021, 21, 1–1. [Google Scholar] [CrossRef]
- Yao, Z.; Yang, S.; He, W.; Li, L.; Xu, R.; Zhang, X.; Li, H.; Zhan, R.; Sun, W.; Tan, J. P311 promotes renal fibrosis via TGFβ1/Smad signaling. Scientific reports 2015, 5, 17032. [Google Scholar] [CrossRef]
- Cheng, X.; Gao, W.; Dang, Y.; Liu, X.; Li, Y.; Peng, X.; Ye, X. (2013). Both ERK/MAPK and TGF-Beta/Smad signaling pathways play a role in the kidney fibrosis of diabetic mice accelerated by blood glucose fluctuation. Journal of diabetes research, 2013.
- Lv, Y. h.; Ma, K. j.; Zhang, H.; He, M.; Zhang, P.; Shen, Y. w.; Jiang, N.; Ma, D.; Chen, L. A Time Course Study Demonstrating m RNA, micro RNA, 18 S r RNA, and U 6 sn RNA Changes to Estimate PMI in Deceased Rat's Spleen. Journal of forensic sciences 2014, 59, 1286–1294. [Google Scholar] [CrossRef]
- Thakral, S.; Purohit, P.; Mishra, R.; Gupta, V.; Setia, P. (2023). The impact of RNA stability and degradation in different tissues to the determination of post-mortem interval: A systematic review. Forensic Sci Int, 349, 111772. [CrossRef]
- Lawson, J.; Syme, H.; Wheeler-Jones, C.; Elliott, J. (2016). Urinary active transforming growth factor β in feline chronic kidney disease. The Veterinary Journal, 214, 1-.
- Meng, X.-m.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: the master regulator of fibrosis. Nature Reviews Nephrology 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Moon, J.-A.; Kim, H.-T.; Cho, I.-S.; Sheen, Y.; Kim, D.-K. IN-1130, a novel transforming growth factor-β type I receptor kinase (ALK5) inhibitor, suppresses renal fibrosis in obstructive nephropathy. Kidney international 2006, 70, 1234–1243. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.M.; Xu, W.M.; Lin, J.C.; Mo, L.Q.; Hua, X.X.; Chen, P.X.; Wu, K.; Zheng, D.D.; Feng, J.Q. Activation of the p38 MAPK/NF-kappaB pathway contributes to doxorubicin-induced inflammation and cytotoxicity in H9c2 cardiac cells. Mol Med Rep 2013, 8, 603–608. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell research 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Fan, T.; Xiao, C.; Tian, H.; Zheng, Y.; Li, C.; He, J. TGF-beta signaling in health, disease, and therapeutics. Signal Transduct Target Ther 2024, 9, 61. [Google Scholar] [CrossRef]
- An, J.N.; Yang, S.H.; Kim, Y.C.; Hwang, J.H.; Park, J.Y.; Kim, D.K.; Kim, J.H.; Kim, D.W.; Hur, D.G.; Oh, Y.K. Periostin induces kidney fibrosis after acute kidney injury via the p38 MAPK pathway. American Journal of Physiology-Renal Physiology 2019, 316, F426–F437. [Google Scholar] [CrossRef]
- Lee, J.; An, J.N.; Hwang, J.H.; Lee, H.; Lee, J.P.; Kim, S.G. p38 MAPK activity is associated with the histological degree of interstitial fibrosis in IgA nephropathy patients. PLoS One 2019, 14, e0213981. [Google Scholar] [CrossRef] [PubMed]
- Koshikawa, M.; Mukoyama, M.; Mori, K.; Suganami, T.; Sawai, K.; Yoshioka, T.; Nagae, T.; Yokoi, H.; Kawachi, H.; Shimizu, F. Role of p38 mitogen-activated protein kinase activation in podocyte injury and proteinuria in experimental nephrotic syndrome. Journal of the American Society of Nephrology 2005, 16, 2690–2701. [Google Scholar] [CrossRef]
- Hui, K.; Yang, Y.; Shi, K.; Luo, H.; Duan, J.; An, J.; Wu, P.; Ci, Y.; Shi, L.; Xu, C. The p38 MAPK-regulated PKD1/CREB/Bcl-2 pathway contributes to selenite-induced colorectal cancer cell apoptosis in vitro and in vivo. Cancer letters 2014, 354, 189–199. [Google Scholar] [CrossRef]
- Gholami, M.; Harchegani, A.; Saeedian, S.; Owrang, M.; Parvizi, M. (2021). Effect of N-acetyl cysteine on oxidative stress and Bax and Bcl2 expression in the kidney tissue of rats exposed to lead. Ukrainian Biochemical Journal.
- Motyl, T.; Grzelkowska, K.; Zimowska, W.; Skierski, J.; Warȩski, P.; Płoszaj, T.; Trzeciak, L. Expression of bcl-2 and bax in TGF-β1-induced apoptosis of L1210 leukemic cells. European journal of cell biology 1998, 75, 367–374. [Google Scholar] [CrossRef]
- Goumenos, D.S.; Tsamandas, A.C.; Kalliakmani, P.; Tsakas, S.; Sotsiou, F.; Bonikos, D.S.; Vlachojannis, J.G. Expression of Apoptosis-Related Proteins Bcl-2 and Bax Along with Transforming Growth Factor (TGF-β1) in the Kidney of Patients with Glomerulonephritides. Renal failure 2004, 26, 361–367. [Google Scholar] [CrossRef]
- Piyarungsri, K.; Pusoonthornthum, R. Changes in reduced glutathione, oxidized glutathione, and glutathione peroxidase in cats with naturally occurring chronic kidney disease. Comparative Clinical Pathology 2016, 25, 655–662. [Google Scholar] [CrossRef]
Figure 1.
Fold-change of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
Figure 1.
Fold-change of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
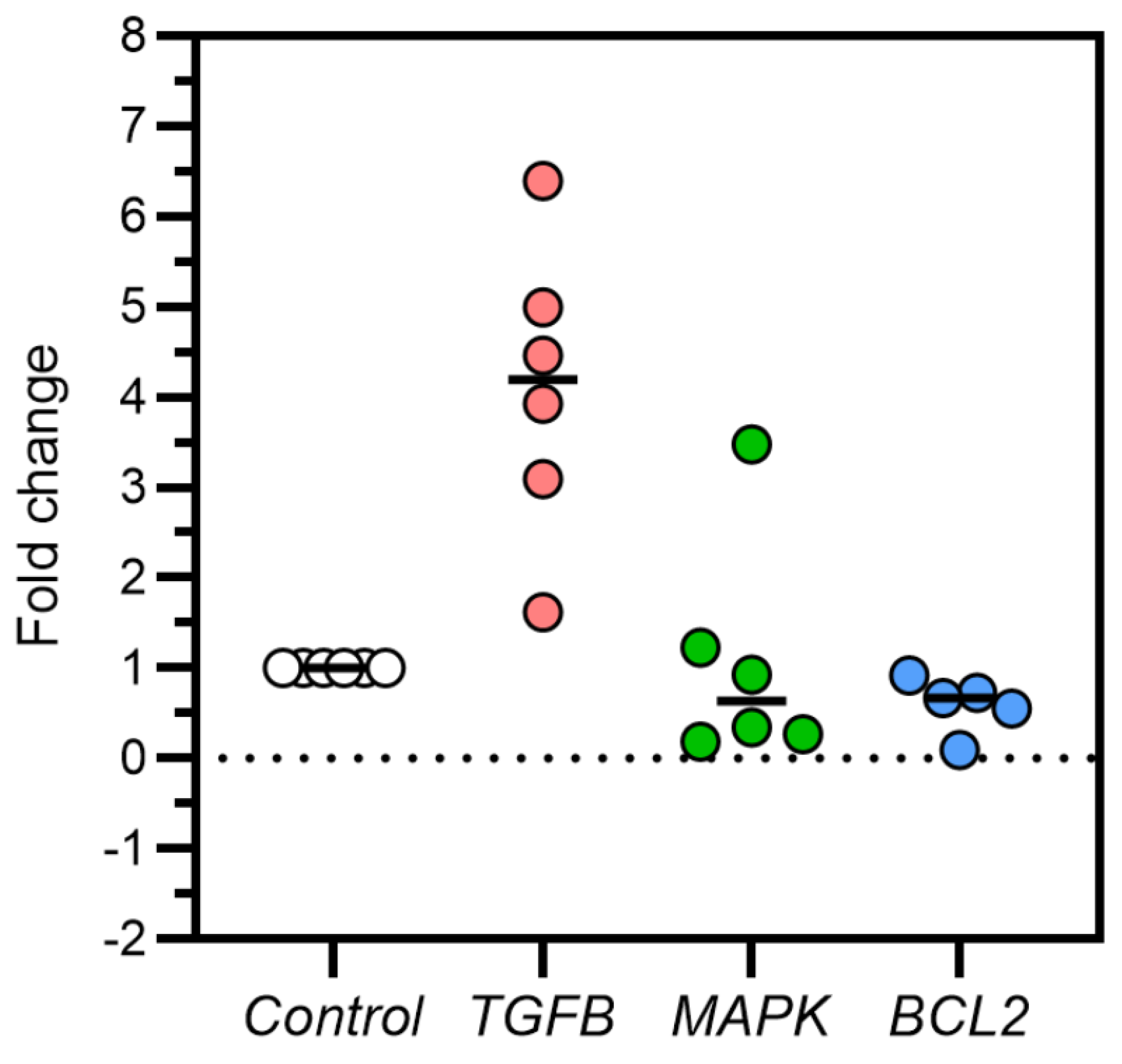
Figure 2.
Difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
Figure 2.
Difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
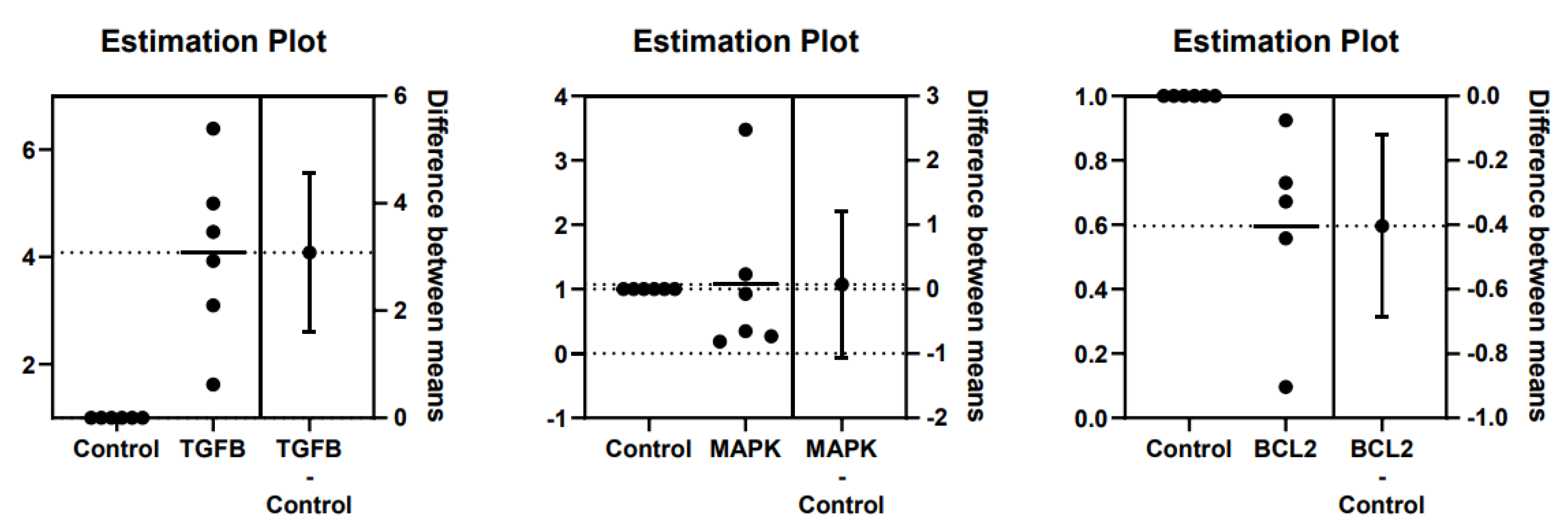
Figure 3.
Fold-change of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
Figure 3.
Fold-change of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
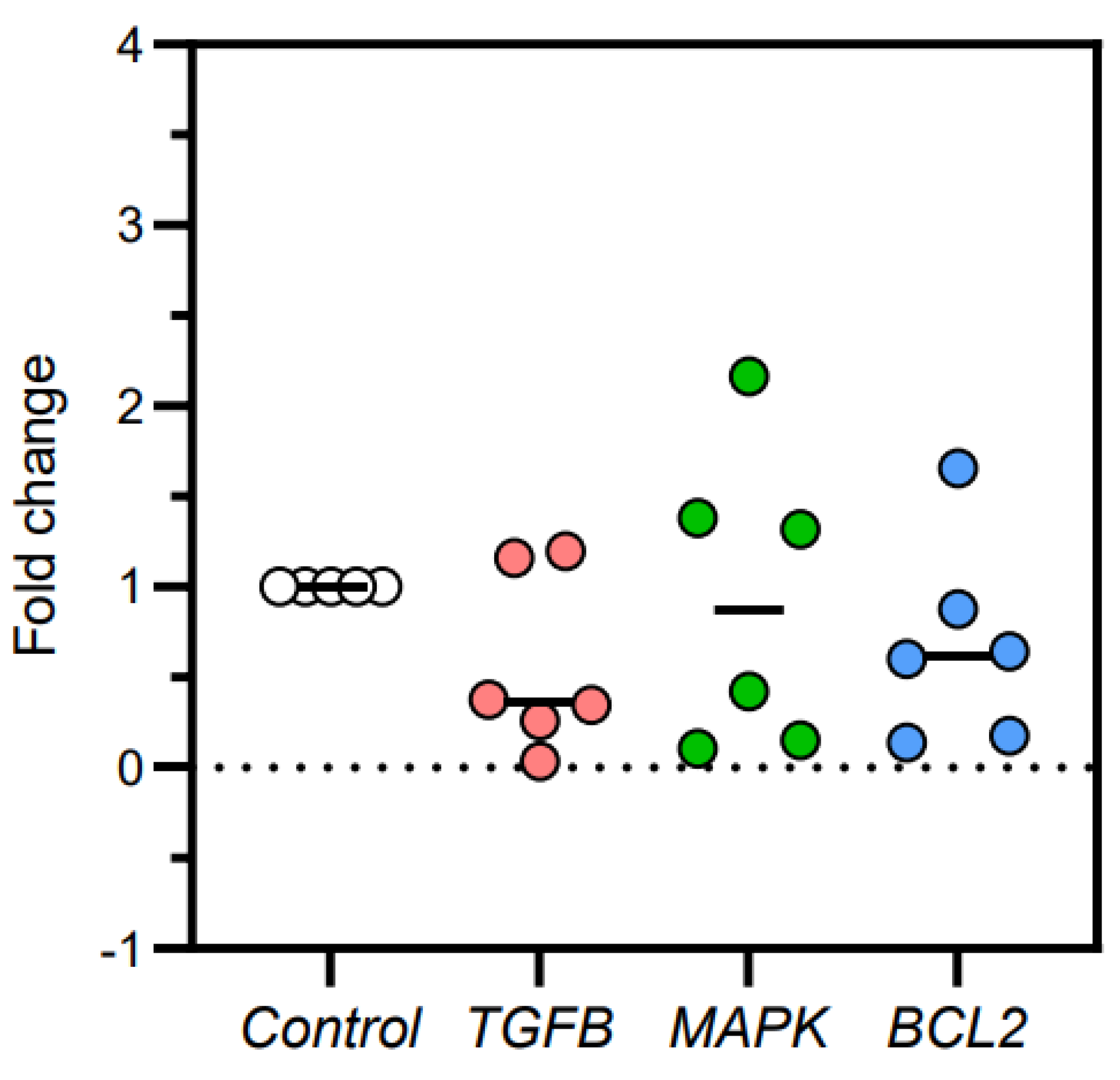
Figure 4.
Difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
Figure 4.
Difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
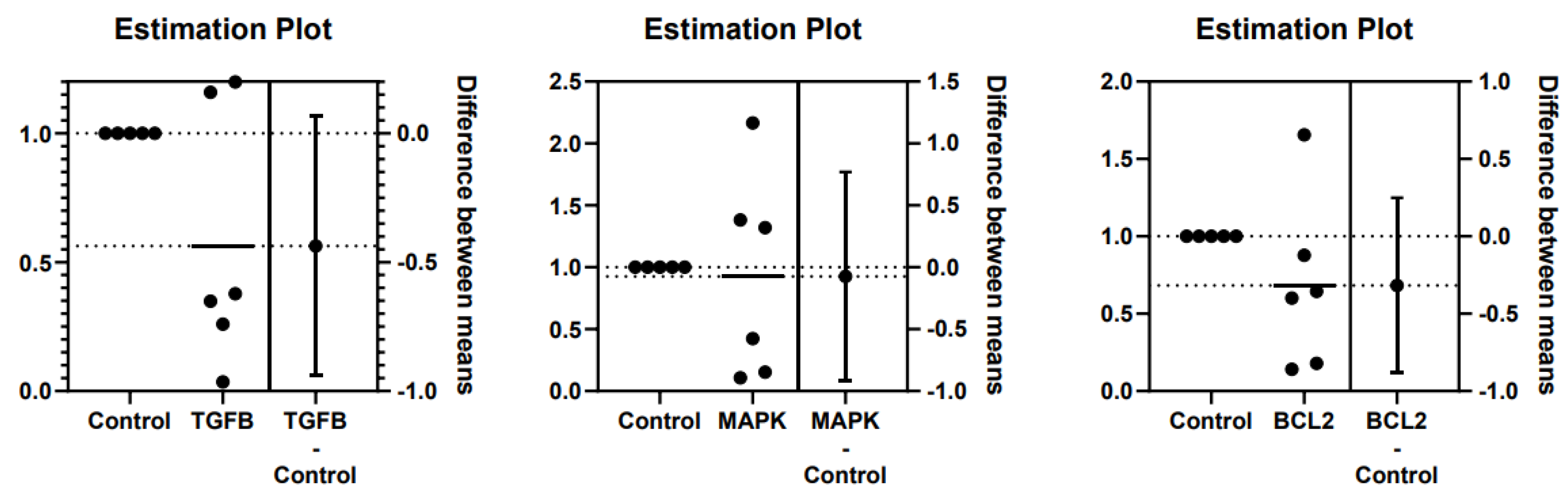
Figure 5.
TGF-β protein expressions in control and doxorubicin-induced feline kidney cell toxicity.
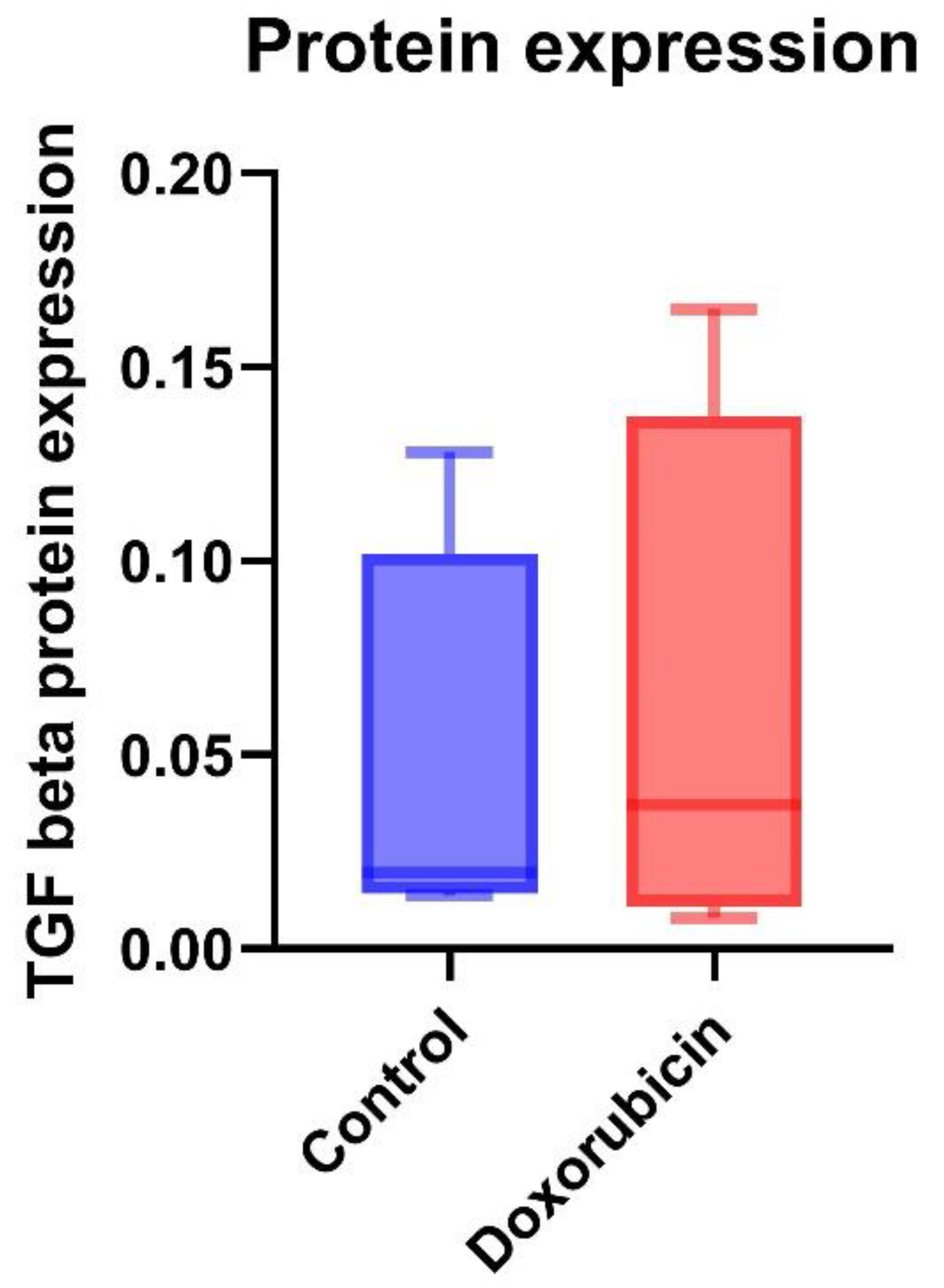
Figure 6.
MAPK protein expressions in control and doxorubicin-induced feline kidney cell toxicity.
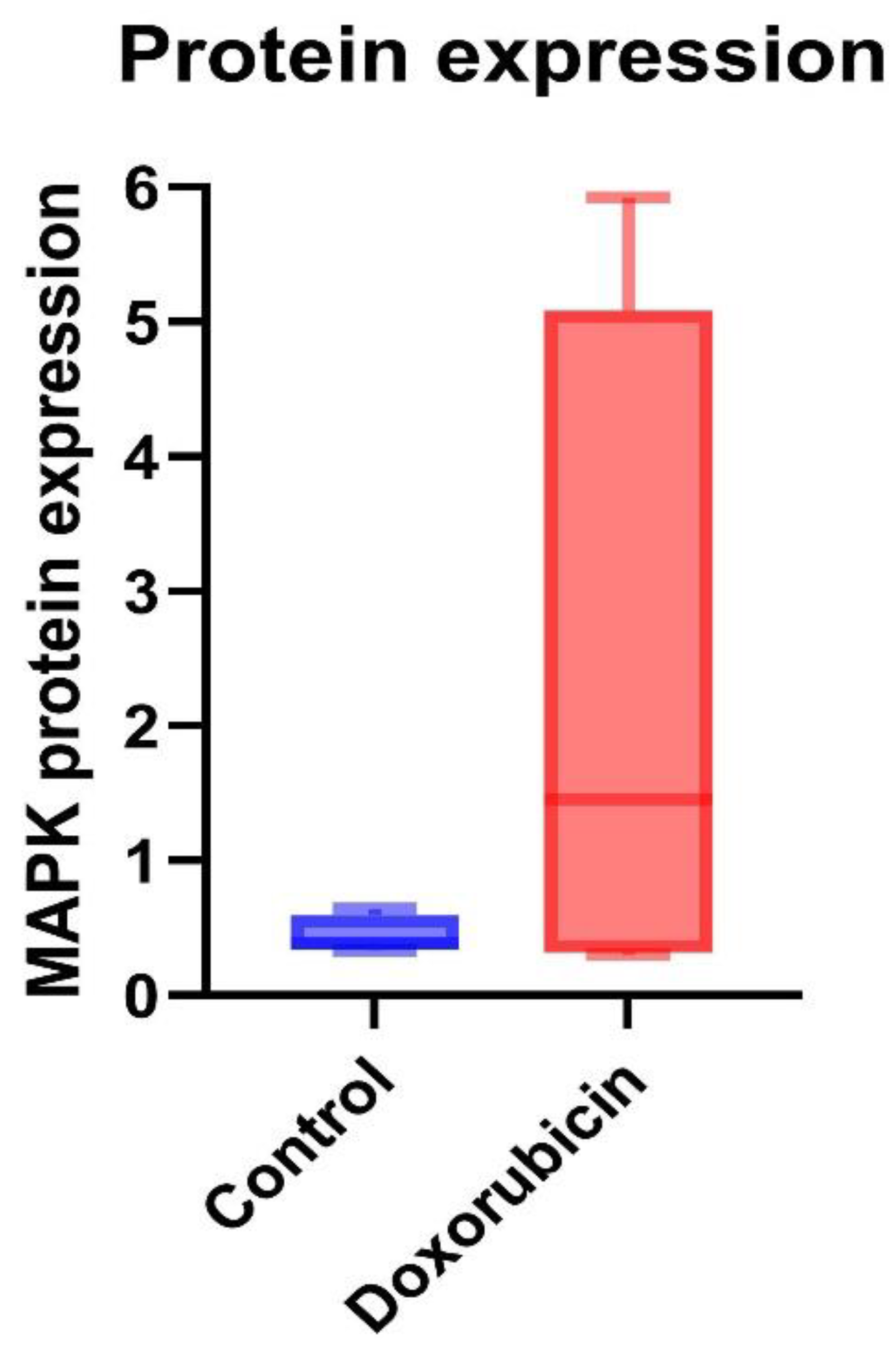
Figure 7.
The immunohistochemistry staining from cats’ kidney tissues. TGF–β immunohistochemistry staining in a cat with no lesion in kidney tissue (A) and a cat with CKD (B). MAPK immunohistochemistry staining in a cat with no lesion in kidney tissue (C) and a cat with CKD (D). Original magnification 400×. Scale bar = 20 µm.
Figure 7.
The immunohistochemistry staining from cats’ kidney tissues. TGF–β immunohistochemistry staining in a cat with no lesion in kidney tissue (A) and a cat with CKD (B). MAPK immunohistochemistry staining in a cat with no lesion in kidney tissue (C) and a cat with CKD (D). Original magnification 400×. Scale bar = 20 µm.
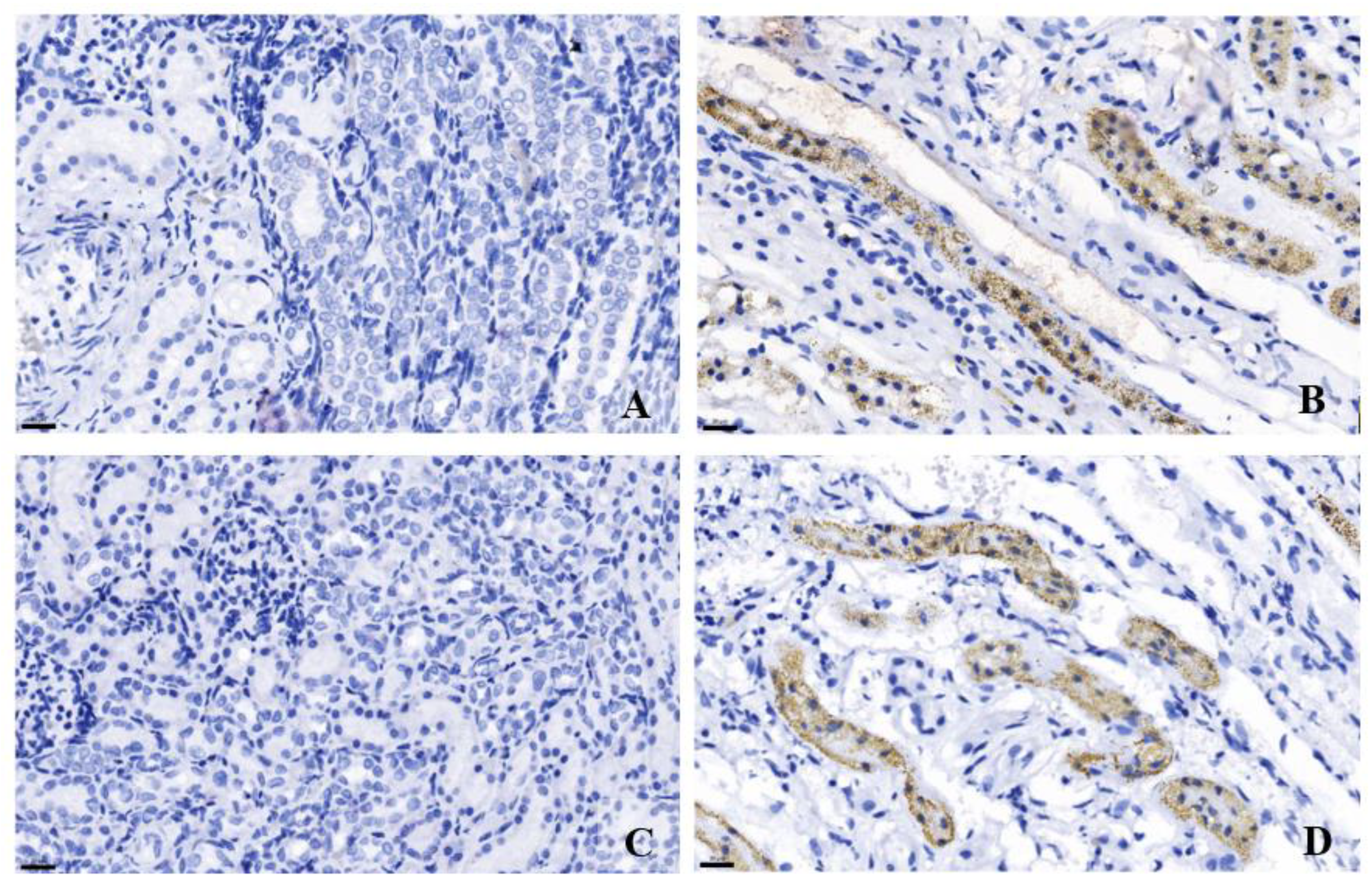
Figure 8.
The summary of TGF-β pathway.
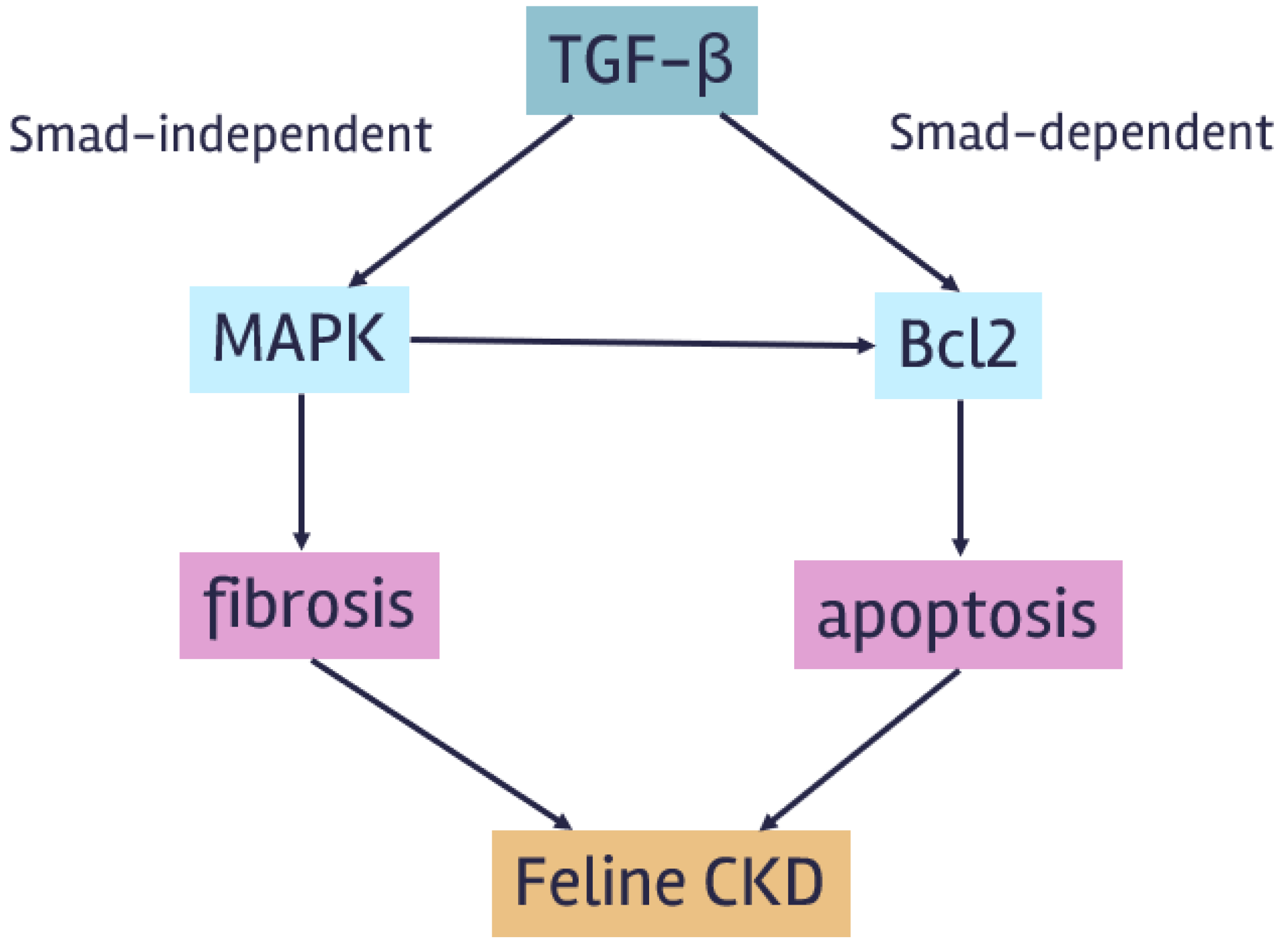

Table 1.
The primer sequences.
| Gene name | Accession | Direction | Sequence | Annealing Temp. (°C) | Size (bp) |
|---|---|---|---|---|---|
| TGFβ | M38449.1 | Forward | CCCTGGACACCAACTATTGC | 60 60 |
163 |
| Reverse | TCCAGGCTCCAAATGTAGGG | ||||
| MAPK | XM_003994973.5 | Forward | ACTGCTGAGCTAAGACCATGAG | 60 60 |
119 |
| Reverse | AAGTCAATGCCACAGTGTGC | ||||
| Bcl2 | NM_001009340.1 | Forward | CCTATCTGGGCCACAAGTGA | 60 60 |
123 |
| Reverse | TAAGAGACCACGGCTTCGTT | ||||
| β-actin | AB051104.1 | Forward | CCATCGAACACGGCATTGT | 60 60 |
147 |
| Reverse | TCTTCTCACGGTTGGCCTTG |
Bcl2 = B-cell lymphoma 2, MAPK = Mitogen-activated protein kinase, TGFβ = Transforming growth factor-beta.
Table 2.
Primary and secondary antibodies.
| Protein name | Antibodies | Dilution | |
|---|---|---|---|
| WB | IHC | ||
| TGF-β | Primary mouse monoclonal TGF-beta 1 Secondary goat anti-mouse |
1:1000 1:5000 |
1:200 |
| MAPK | Primary mouse polyclonal p38 MAPK Secondary goat anti-mouse |
1:1000 1:5000 |
1:200 |
| β-actin | Direct-Blot HRP mouse monoclonal anti-β-actin | 1:1000 | |
IHC = Immunohistochemistry, MAPK = Mitogen-activated protein kinase, TGF-β = Transforming growth factor-beta, WB = western blot analysis,.
Table 3.
Fold-change and difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
Table 3.
Fold-change and difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in controls and DOX-induced feline kidney cell toxicity.
| mRNA expression | Fold-change | Difference between mean ± SD | ANOVA P-value |
Unpaired t-test P-value |
|
|---|---|---|---|---|---|
| Control (n = 6) |
DOX-treated (n = 6) |
||||
| TGFβ | 1 | 4.084 | 3.084 ± 0.668 0.072 ± 0.509 -0.404 ± 0.125 |
0.002** | 0.001** |
| MAPK | 1 | 1.072 | 0.890 | ||
| Bcl2 | 1 | 0.596 | 0.010* | ||
ANOVA = Analysis of variance, Bcl2 = B-cell lymphoma 2, DOX = Doxorubicin, MAPK = Mitogen-activated protein kinase, SD = Standard deviation, TGFβ = Transforming growth factor-beta, * P < 0.05, ** P < 0.01.
Table 4.
Fold-change and difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
Table 4.
Fold-change and difference between mean ± SD of TGFβ, MAPK, and Bcl2 gene expressions in kidney tissues of cats with no kidney lesions and cats with chronic kidney disease (CKD).
| mRNA expression | Fold-change | Difference between mean ± SD | ANOVA P-value |
Unpaired t-test P-value |
|
|---|---|---|---|---|---|
| No kidney lesions (n = 6) |
CKD (n = 6) |
||||
| TGFβ | 1 | 0.563 | -0.437 ± 0.222 -0.075 ± 0.373 -0.317 ± 0.251 |
0.549 | 0.081 |
| MAPK | 1 | 0.925 | 0.846 | ||
| Bcl2 | 1 | 0.683 | 0.238 | ||
ANOVA = Analysis of variance, Bcl2 = B-cell lymphoma 2, CKD = Chronic kidney disease, DOX = Doxorubicin, MAPK = Mitogen-activated protein kinase, SD = Standard deviation, TGFβ = Transforming growth factor-beta.
Table 5.
Mean ± SD of TGF-β and MAPK protein expressions in control and DOX-induced feline kidney cell toxicity.
Table 5.
Mean ± SD of TGF-β and MAPK protein expressions in control and DOX-induced feline kidney cell toxicity.
| Protein expression | Control (n = 3) |
DOX-treated (n = 3) |
P-value |
|---|---|---|---|
| TGF-β | 0.04 ± 0.05 | 0.06 ± 0.07 | 0.73 |
| MAPK | 0.44 ± 0.14 | 2.28 ± 2.64 | 0.68 |
DOX = Doxorubicin, MAPK = Mitogen-activated protein kinase, TGF-β = Transforming growth factor-beta.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



