Submitted:
16 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
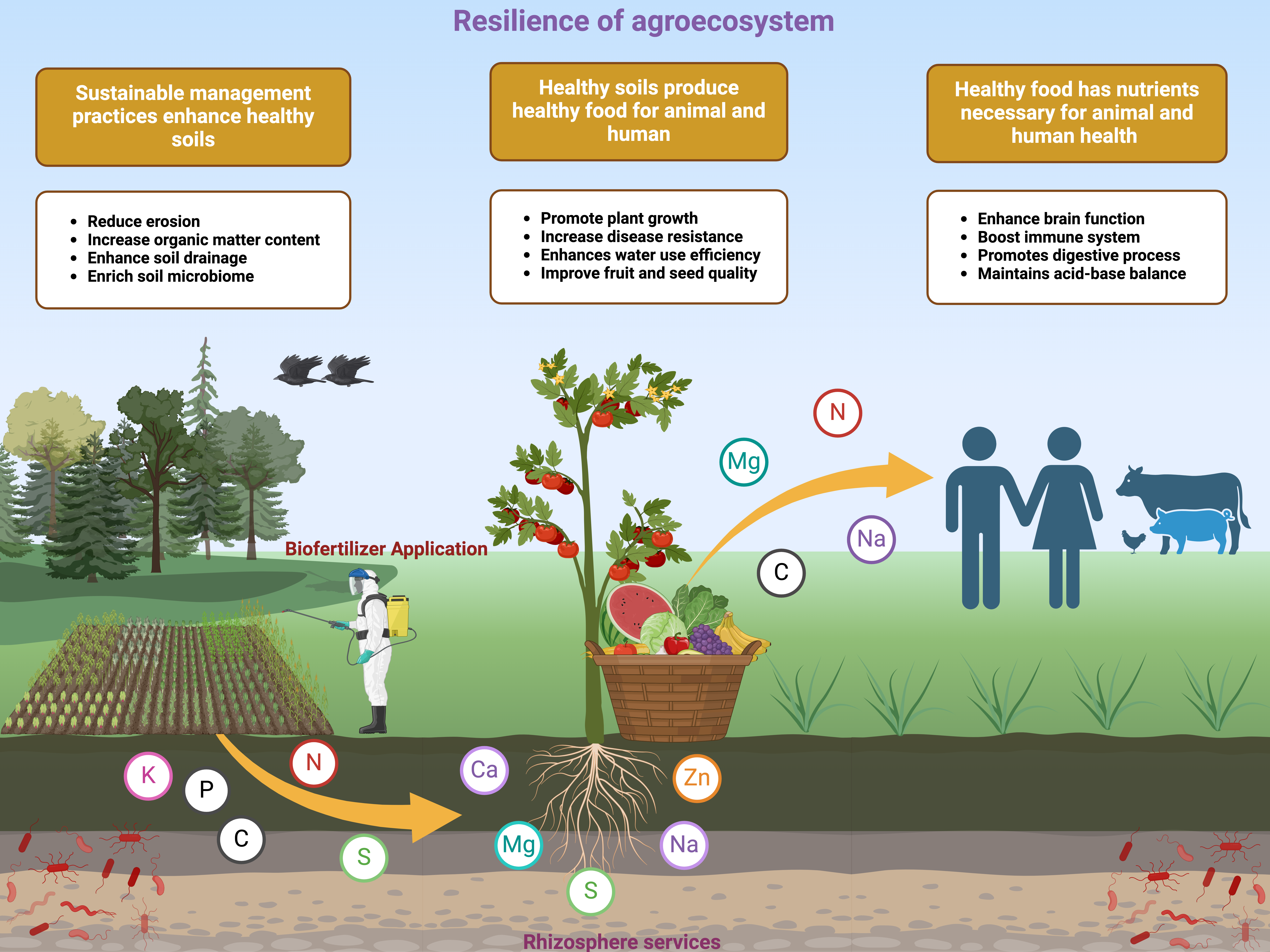
As farming techniques evolve and climate conditions shift, meeting the demands of a growing global population requires optimizing environmentally friendly practices and minimizing the ecological impacts of food production. Enhancing and manipulating rhizosphere processes have emerged as key strategies for promoting environmentally sustainable agriculture in the future. Considerate the relationships between soil micro-organisms and plant roots holds the possible to revolutionize agricultural production, increase crop yields, reduce dependence on synthetic fertilizers, and support sustainable farming practices through rhizosphere management. The rhizosphere, a region around plant roots, is a hotspot for microbial activity due to the exudates released by plant roots. These exudates, along with green manure cultivation and intercropping, change the structure of the soil’s microbial community. The plant provides essential nutrients to microorganisms, which in turn compete for these resources. Managing microbial com-petition in the rhizosphere, where microorganisms are abundant, is crucial. Beneficial plant-associated microbes, such as Trichoderma spp., Penicillium spp., Aspergillus spp., and Bacillus spp., are often employed; and these microorganisms have a one-to-one rela-tionship. However, the behavior of these inoculated microorganisms in the rhizosphere is still not fully understood. In addition, the use of green manures, intercropping, and growing multiple crops together in the same field offers various benefits. These practices promote soil health, enhance nutrient cycling, sequester carbon, reduce greenhouse gas emissions, decrease reliance on synthetic fertilizers, rebuild rhizosphere microbial communities, and provide economic advantages. These methods foster positive interactions between plants, microbes, and the rhizosphere, benefiting the entire ecosystem. The main aim of this review stands to critically explore in what way the resilience of agroe-cosystems can be strengthened through improved rhizosphere processes. By utilizing beneficial microbes, incorporating green manures, and practicing intercropping, we can support and promote sustainable cropping systems. This approach, combined with sound policy implementation, will help maintain healthy soils—an essential component of sustainable agroecosystems—and mitigate the predicted climate challenges of the near future.
Keywords:
Agroecosystems
; rhizosphere process
; soil microbes
; green manure
; intercropping
1. Introduction
The swift increase in the global population has markedly heightened the need for energy, food, and other essential goods, leading to the expansion of agricultural land use. However, poor farming techniques, urbanization, and inadequate soil management practices have had negative impacts on agricultural soil, such as reduced soil fertility, the loss of beneficial soil microbiota, and the migration of soil organisms [1]. The top layer of soil surrounding plant roots, known as the rhizosphere, is home to a diverse range of microbes that play vital roles in promoting plant growth, organic matter degradation, and maintain the soil sustainability. However, in the lack of proper soil management, coupled with inappropriate agricultural practices, the quality of rhizosphere soil deteriorates, and its microbial communities weaken [2,3].
The rhizosphere serves as a microbial hotspot owing to the exudates emitted by plant roots, which supply vital nutrients for microbial activities [4]. Dynamics for example root zones, plant growth phases, plant species, and environmental conditions change the diversity and composition of soil microbial communities (e.g., bacterial and fungi) in the rhizosphere [5]. Multiple species can compose microbial groups, demonstrating comparable mechanisms of action, yet differing in their tolerance to plant cultivars and various environmental stresses. The microorganisms found in the rhizosphere include fungi, bacteria, and nematodes [6]. Large numbers of this community have an impartial effect on soil and plants, but some microbes are directly or indirectly connected with the plant–soil systems and provide nutrients from the rhizosphere to the plant. Additionally, rhizosphere microbiota contains microorganisms with both positive and negative effects on plant growth and soil health. Harmful microbes include pathogenic fungi, nematodes, and bacteria, while beneficial microbes, such as plant growth-promoting microbes (PGPM), nitrogen-fixing bacteria, and mycorrhizal fungi, contribute to plant nutrition and soil health [6,7]. However, beneficial microbes provide essential plant nutrients and contribute to lowering greenhouse gas emissions, improving nutrient availability, suppressing soil-borne plant diseases, and balancing nutrient cycling [8].
Conversely, many authors have pointed out that use of various agricultural techniques, overuse of mineral fertilizers, agrochemicals and farming practices could reduce soil fertility and disturb the rhizosphere microbiota, resulting in decreased soil health and crop production [9,10]. In general, overuse of agrochemicals (i.e., fungicide, herbicides, nematicides, insecticides) is being used non-judiciously, which has a harmful effect on soil positive microbiota in the soil as well as the overall health and quality of the soil [11,12]. This impact is mostly related to changes in microbial parameters such as enzymatic activity and biomass, which are the most important features of soil health and are important tools in monitoring soil quality. Eighty to ninety percent of soil processes are controlled by soil microorganisms and are strongly linked with plant and soil health, in addition to the role of microorganisms in the suppression of soil-borne diseases, nutrient cycling, soil resilience, and maintenance of soil sustainability [13,14]. Nevertheless, several authors reported that green manure is an alternate way to sustain agriculture and maintain soil health. The integration of the entire plant, containing more nutrients than crop straw, into the soil enhances soil fertility by adding more organic matter. Green manure improved crop yield and growth; reduced pests insects, suppress plant diseases; and increased soil enzyme activities, which regulate nutrient fluxes and stimulate plant growth [15,16,17]. Therefore, a sustainable solution must be sought by utilizing all available resources to modulate the rhizospheric process for sustainably enhancing agroecosystems productivity.
It is important for long-term sustainability in agricultural systems to keep agroecosystems stable and improve rhizosphere processes that make soil more resilient. This includes keeping and expanding the diversity of microorganisms in the rhizosphere. The rhizosphere is crucial for facilitating nutrient cycling, enhancing soil structure, and promoting plant health. The primary goal of this review is to investigate several methods for improving agricultural ecosystem resilience through rhizosphere activity optimization. We can implement these practices by utilizing beneficial microbes, green manures, and intercropping. These strategies jointly enhance synergistic communications between soil microorganisms and plant roots, creating an environment favorable for soil health and plant growth. The findings of this review improve comprehension of sustainable agriculture techniques, highlighting the importance of natural and organic resources in boosting crop production while preserving ecological balance. Using these rhizosphere-driven processes, we could develop more sustainable and resilient agricultural systems that ensure food security and environmental preservation amidst changing climate situations.
2. The Rhizosphere: Its Mechanism and Roles
The rhizosphere refers to the area surrounding the root zone [18,19]. This zone is considered essential for biological activities because many exudates produced by plant roots provide nutrients for soil microorganisms. Thus, this region is vital for disease resistance, nutrient cycling, and plant growth [20]. The rhizosphere zone has a central role in the food chain of soil microorganisms. Microbial food chain mainly includes dead plant matter and living plant roots, which are the primary source of nutrients [21], and carbon from living plant tissues and root exudates. These are crucial for plant health and growth as these support disease suppression, protection, and nutrient uptake [22]. The activity of beneficial microorganisms in the rhizosphere not only provides essential nutrients to plants but also influences the quality and quantity of root exudates [22,23]. Furthermore, various metabolites exuded by plant roots into the soil primarily shape and attract the microbial community composition in the rhizosphere, facilitating interactions with both biotic and abiotic factors. Plants frequently adjust the diversity of these microbes according to the benefits they provide for growth and health [24]. In the rhizosphere, plants engage in a wide range of interactions with soil-dwelling organisms, including exploitation, competition, neutrality, mutualism, and commensalism. These diverse interactions are essential for plants to adapt to varying environmental conditions, which is crucial for their survival and agricultural productivity. In the rhizosphere, plants host a diverse collection of microbes, and they can inhabit the inner tissues (endosphere), and surface tissues (phylloplane and rhizoplane). Collectively, these microorganisms are referred to as the plant microbiome [25,26]. The plant and soil microbiome is critical for stimulating plant development, improving soil quality and fertility, and supporting agricultural sustainability by strengthening resilience to environmental stressors, nutrient cycling, and disease fighting [27,28]. The interplay between roots and their surrounding soil constitutes one of the most prolific and diverse ecosystems in nature. The rhizosphere, the tiny and thin soil layer around the roots, has a vast population of microbes, fueled by carbon-rich compounds exuded during photosynthesis, and provide essential nourishment for the recruited microorganisms [29]. Overall, the rhizosphere is a kind of microenvironment wherein plant root development and nutrient uptake, soil properties, and microbe activities interact in a corresponding manner (Figure 1).
2.1. Rhizosphere Processes: Nutrient Retention in Agroecosystems
The rhizosphere hosts diverse microbial groups that play critical roles in nutrient cycling, promoting plant growth, and protecting plants from pathogens, as well as from biotic and abiotic stresses [30]. Changes in rhizosphere processes directly contribute to nutrient retention in agroecosystems. Through plant-soil interactions, plants and their residues provide organic carbon (C) to the soil. The rate of soil carbon input depends primarily on the rate of plant growth, which is driven by photosynthesis. Root-derived carbon input, in particular, is a key regulatory factor in the plant-soil interaction within the rhizosphere, as it has a more direct impact on soil processes than carbon input from shoots [31]. This root-derived carbon plays a significant role in soil carbon sequestration. The amount of carbon that can be stored as soil organic matter (SOM) depends on the balance between carbon input and the rate at which plant materials decompose. Decomposition is the process by which microorganisms break down organic molecules into their inorganic components, such as nutrients and CO2. Carbon dioxide, produced during microbial respiration, is released from the soil back into the atmosphere. However, some decomposition products are protected from further breakdown, playing a critical role in forming stable SOM, which is essential for long-term carbon sequestration in agroecosystems [31]. As a result, the rhizosphere process is particularly crucial for the nutrients cycling, stabilizing SOM, making nutrients available for plant growth, and ultimately handling soil microorganisms. On the other hand, maintaining and increasing soil and it’s process through utilizing carbonized materials such as “Biochar” that have long-term stability [32]. Biochars can reduce the emission of N2O and CH4 from the soil [33]. Biochar comprises a stable C compound as a derivative of gasification and pyrolysis and is available either as a pelletized or powder form. Char is ubiquitous in natural soils, containing as much as 35% of total organic C (TOC) in fire-impacted soils. Laird [34] reported that the application of biochar in to the soil was found to enhance the soil fertility, significantly increase soil organic carbon (SOC) and nutrients, which leads to the improvement of soil health. SOC consists of a complex mix of partially decomposed substances, including polysaccharides, lignin, aliphatic biopolymers, tannins, lipids, proteins, and amino sugars derived from plant litter, as well as microbial and faunal biomass [35]. However, SOC has been classified into slow, active, and inert pools, depending upon the decomposition rate in soil [36]. Furthermore, biochar largely supports the SOC pool for stabilization while or green manure incorporation add to the more bio-accessible fraction of the soil C reserve. This is because the biochar decomposes typically slower than that of uncharred biomass, e.g., green manure incorporation, fresh crop residues [37]. However, green manure or fresh crop residues in agricultural fields, on the other hand, can be used as a SOC storage/sequestration strategy by supporting cover crops; conservation tillage provides benefits including nutrient cycling, control of surface run off of water, wind erosion, and crop production [38]. Yazdanpanah [39] reported that applying green manure or plat residues improved the stability and total porosity of soil aggregates and increased the soil C pool but depended on the type and application rate of soil amendments. Whereas Kenney, et al. [40] reported that removal of crop residues (e.g., corn stover, zea mays L.) decreased the soil C pool. Therefore, the following strategy should be considered to achieve higher soil C sequestration via soil amendments produced from green manure/crop residues or biochar. Indirect effects of using these amendments should be considered in the context of production/control of soil GHG, environmental health and economic cost benefits of their use. As per our knowledge, the combined effect of biochar and crop residues have rarely been investigated except one recently study Nguyen, et al. [41], and it should be considered in near future. Overall, biochar seems to have a greater positive impact on total soil SC while the incorporation of crop residues/green manures can increases the microbial biomass C of soil, enhances the rhizosphere process and reduce the carbon dioxide emission.
2.2. Rhizosphere Engineering: Turn into Sustainably Increases the Agroecosystem and Its Productivity
The rhizosphere is a hotspot of microbes because many exudates are produced by plant roots, which are the primary source of nutrients for essential microbial processes and sustain the agroecosystems and their functions. The rhizosphere contains diverse microbial groups that perform various functions and affect the nutrients cycling and plant growth promotion and this portion affected via different strategies such as crop rotation, green manuring, and sustainable use of PGPM and intercropping. However, addition of PGPM, Asghar and Kataoka [42] found that plant growth promoting fungi changed the fungal community composition, enhances the phosphatase and glycosidase enzymes activities in the soil and stimulated the lettuce and brassica plant growth. Further, Kataoka, et al. [43] reported that incorporating leguminous green manure (Hairy Vetch, Vicia villosa L.) enhances the soil fungal biomass and diversity and provides a healthier environment by producing soil phosphatase enzymes. Furthermore, these rhizosphere engineering and its processes are explained with different strategies. In short, these interventions improve the rhizospheric processes and sustainably increase the agroecosystem and its productivity. On the other side of the agroecosystems and maintaining the rhizosphere process, biochar has emerged as a crucial innovation in scientific research, offering multiple benefits for sustainable agriculture, rhizosphere management, and environmental health [44]. Biochar, a type of charcoal obtained after the combustion of feedstock under a minimal Biochar is a type of charcoal produced through the combustion of organic feedstock with limited oxygen. It is considered a valuable soil conditioner and an effective means of carbon sequestration, which helps mitigate global warming and climate change [45]. Biochar decomposes slowly, making it highly durable when applied to soil, where it can remain for the long term [46]. However, generally biochar is derived from waste materials such as animal manures, forest residues and agricultural byproducts [47]. Through scientific processes, these waste materials are transformed into valuable products that can directly or indirectly improve soil health and promote plant growth. Biochar enhances crop yield, improves soil quality, and maintains soil health by preserving biochemical properties [48]. In terms of soil biological properties, Uzoma, et al. [49] found that biochar positively influences soil microbes and their activity, leading to increased crop yields. However, the effect of biochar depends on the specific soil type and specific crops. Another study, Lu, et al. [50] highlighted that biochar porous structure creates a favorable niche for soil microbiota, boosting microbial populations. Conversely, biochar derived from rice straw has been shown to reduce certain microbial communities, such as Ascomycota fungi and Actinobacteria, though the abundance and diversity of soil microbes may vary with different types of biochar [51]. The application of biochar not only alters soil microbial communities but also affects nutrient cycling, which can have a direct impact on rhizospheric processes and agroecosystem productivity.
3. Role of Soil Microbes in the Rhizosphere Processes
The rhizosphere is home to various soil microbes, including fungi, bacteria, protozoa and nematodes, which play essential roles in nutrient cycling, plant growth, and maintaining agroecosystems. These microbes facilitate the breakdown of organic matter and nitrogen fixation, making nutrients available to plants [1]. In agriculture, two groups of soil microorganisms are particularly important—rhizosphere fungi and bacteria—due to their critical functions. Fungi and bacteria thrive in soil, adapting to a wide range of environments, including harsh and high-salt conditions [52,53]. Fungi in the rhizosphere include pathogenic, saprotrophic, and mutualistic species, with mycorrhizal and plant-growth-promoting fungi being especially important for increasing nutrient availability, promoting plant growth and stress tolerance, and improving soil structure and disease resistance [54]. In contrast, the main rhizosphere bacterial phyla are Firmicutes, Actinobacteria, Acidobacteria, and Proteobacteria [55]. Plant growth-promoting rhizobacteria (PGPR) are crucial for enhancing plant growth, improving disease resistance, responding to carbon inputs, nutrient uptake, and assisting in phytoremediation [56,57,58]. In recent decades, a substantial body of literature has emerged demonstrating the activities of microbial species in plant growth promotion, biocontrol of phytopathogens, and nutrient cycling, highlighting their potential for development as alternative or supplementary agrochemicals in sustainable agriculture.
3.1. The Role of Microbes in Agroecosystems
Rhizosphere microorganisms, such as fungi and bacteria, not only act as pathogens in agricultural plants but also facilitate plant growth, increase nutrient availability, and control plant diseases through their role as biocontrol agents [59]. Recent research has highlighted the utilization of rhizosphere microorganisms to enhance plant development and mitigate diseases, thereby reducing reliance on pesticides and herbicides [60]. These microorganisms establish symbiotic associations with plant roots, exemplified by mycorrhizal fungi, which augment water and nutrient absorption, especially phosphorus. Furthermore, specific bacterial and fungal species might inhibit detrimental infections, thereby reducing dependence on chemical pesticides. Furthermore, these microbes help in the degradation of pollutants, thereby improving soil quality and resilience [61,62]. In compare, the excessive application of chemical fertilizers and pesticides has detrimental environmental consequences, and growing reliance on these substances is concerning. Commercially available as biofertilizers and biocontrol agents, a wide range of microorganisms are utilized in agricultural production and the management of agroecosystems through rhizosphere processes those chemical fertilizers often disrupt. The diversity and activity of soil microbial communities are essential for preserving soil structure, enhancing plant growth, and sustaining ecosystem stability, rendering these microorganisms crucial for sustainable agricultural operations [63]. Therefore, enhancing the resilience of agroecosystems through rhizosphere microbes is crucial for promoting sustainable agriculture.
3.2. Plant Growth Promotion Via Microbes
The application of chemical fertilizers to meet the food demand for a growing human population has led to many unpredicted environmental concerns related to soil-plant health and agroecosystems [64]. To protect soil-plant health and the environment, there is a need to develop and adopt new biological techniques that are not harmful to agroecosystems, soil, and plant health. Many researchers and scientists have already reported that microbes related to biofertilizers are functional and could be an alternative to chemical fertilizers, provide maximum environmental benefits, and improve crop yield and soil microbial biomass. In this context, microbe-related biofertilizers are capable and could be used in an eco-friendly manner to enhance crop production and maintain agroecosystems [65]. Microbe-related biofertilizers are defined as biological inoculants of beneficial fungal and bacterial strains mobilizer that can increase crop productivity and sustain soil biodiversity [66]. Mazid and Khan [66] reported that these inoculants, applicable directly or indirectly to soil and plant roots, are cost-effective and supply important nutrients for plants, enhancing soil nutritional composition, structure, and fertility..
Moreover, biofertilizer-related microbes are commonly known as plant growth promoting microbes (PGPM). There are various fungal and bacterial strains that have already been reported and used commercially to improve plant growth, soil conditions, and the nutritional demand of plants. Such bacterial and fungal strains are Azotobacter, Pseudomonas, Azospirillum, Rhizobium, Streptomyces, Bacillus spp., Trichoderma, Penicillium, Phoma, Fusarium, and Aspergillus spp., and these strains have been gaining global attention due to their potential to control soil diseases, promote plant growth, and maintain agroecosystems [67,68,69]. The significant role and potential activities of PGPM are shown in Table 1.
3.3. Singling Between Plants and Microbes in the Rhizosphere
The rhizosphere contains four main components: soil, microorganisms, roots, and their interactions. These components collectively influence a range of physical, chemical, and biological processes that affect nutrient use, plant development, and overall plant health. The primary goal of rhizosphere signaling is the interaction between plants and the symbiotic microbes in their surrounding area, which promotes the growth of various rhizosphere microbial communities that have a beneficial impact on plant productivity [82]. A robust reciprocal influence marks the interaction between the plant and the rhizosphere microbiome (Figure 1), where they interact through the exchange of signaling molecules produced and recognized by both the plant and its related microbiosystem [19]. Disease-free and asymptomatic plants have complex relationships with their rhizosphere microbes, which improve plant performance and maintain the agroforestry system [23,83]. Plants change the pH level, the structure of the soil, the amount of oxygen that is available, and the energy source that comes from carbon-rich exudates. These changes have an effect on the microbiome around the plants’ roots. Many of the chemically varying primary and secondary metabolites found in plant root exudates have bioactive effects on microorganisms, influencing their composition and function [84]. The exudation of carbon from plant roots, which accounts for almost one-third of the carbon produced by photosynthesis, significantly influences the results of chemical interactions at both the individual and community levels [85]. Root exudates serve as a principal means of communication between plants and their living environment, enabling several reactions, including nutrient uptake, competition for resources, signaling across species, the attraction of microorganisms, and several other interactions [86]. Furthermore, sugars, amino acids, organic acids, phenolic compounds, and secondary metabolites like coumarins, glucosinolates, benzoxazinoids, camalexin, and triterpenes are the main organic parts that come from exudate [87,88]. Plant species cultivate a unique microbial community in their rhizosphere by creating a varied carbon-rich environment. This community provides many adaptive benefits to the plant host by influencing the composition of microorganisms and adjusting their advantageous characteristics [88]. Additionally, chemicals made from root exudates play a key role in creating a stress-resistant microbiome that helps plants deal with abiotic stresses like not getting enough food, being sick, and drought. The identification of stress-derived metabolomics and microbiota is a viable approach to addressing both abiotic and biotic limitations [89,90,91]. However, we have not thoroughly investigated the positive impacts of root-enriched microbial species, supported by specialized metabolites produced from root exudate [92]. This paragraph of the manuscript on a review of current research on the plant rhizosphere microbiota and the compounds produced by root exudates for nutrient acquisition as depicted in the conceptual (Figure 2).
4. The Role of Green Manures in Agroecosystems
Green manures represent a promising strategy to sustaining agroecosystems and improving the soil health and rhizosphere processes. Green manure can reduce chemical fertilizers in agroecosystems, ultimately improving soil health and quality [93]. Green manure provides various benefits to the agroecosystem and soil, such as soil covering, reducing soil temperature, organic matter improvement, increasing water infiltration, and reducing weed infestation [94]. Besides, green manure has environmental and economic benefits such as food production, maintaining biodiversity, soil carbon sequestration, and soil retention [95]. However, to enhance the resilience of agroecosystems and maintain soil health, planting the green manure is an essential and valuable practice and works to reduce soil-borne diseases through biofumigation. Green manure provides better soil health and biodiversity and promotes the beneficial fungal genera that break the soil-borne pathogen’s life cycle that is connected with the crop genotype [96]. Many types of green manures, including leguminous and non-leguminous, have multiple functions and benefits for soil and plant health such as incorporating leguminous green manure to enhance wheat yield and production due to the mineralization of inorganic nitrogen and stimulating biological fixation [97]. Furthermore, Kataoka, Nagasaka, Tanaka, Yamamura, Shinohara, Haramoto, Hayakawa and Sakamoto [43] also reported that incorporating leguminous green manure (Hairy Vetch, Vicia villosa L.) enhances the soil fungal biomass and diversity and provides a healthier environment by producing soil phosphatase enzymes. Therefore, it could be assumed that planting and incorporating different green manures could maintain the agroecosystems and rhizosphere processes by enhancing biological processes and increasing organic matter. Another point of view, green manure may alter the soil N and C availability such as hairy vetch (leguminous plant) fixes N2 whereas barley (Non-leguminous plant) has high biomass productivity, thus adds more C, which influences ecosystems functioning and microbial groups in soil [98]. Chavarria, Verdenelli, Serri, Restovich, Andriulo, Meriles and Vargas-Gil [98] reported that the green manure incorporation can improves agroecosystems services by improving nutrient retention capacity, accelerating soil organic matter, and reducing greenhouse gases in agricultural soils. However, applying alternative management such as green manuring practices stimulates the improvement of soil diverse microbial communities, accelerating the specific enzyme activities related to P and C [99]. Thus the relationship between microbial community composition and their activity stimulate the plant growth and soil quality.
4.1. Green Manure and Their Beneficial Effects on the Rhizosphere Processes
The application of green manure is an additional practice for enhancing the resilience of agroecosystems and rhizosphere processes and increasing soil’s productive capacity through nutrient availability. This application is geared towards maintaining agroecosystems and long-term environmental sustainability by enhancing processes such as nutrient cycling and preserving and maintaining soil biodiversity and the physical and chemical stability of the soil. Incorporating green manure can positively affect soil microbial communities by directly adding nutrients after incorporation and indirectly by changing plant and soil properties [100,101]. At the same time, Sun, Zhang, Guo, Wang and Chu [54] reported that the plant could affect soil microbial diversity and community through releasing root exudates. The decomposing residues of brassica green manure release glucosinolates, which assist in controlling parasitic nematodes [102]. Furthermore, during this phase, they can induce chemical and physical alterations in the soil, promote root penetration of subsequent crops, and serve as a kind of buffer protecting the soil. Furthermore, the exclusive use of grass species may improve the soil nutrient profile compared to monoculture. The advantages of green manure designate it as a sustainable practice capable of providing various agro-ecological services; however, effective management of green manure is essential to substantially enhance soil carbon stocks and mitigate climate change, particularly in semi-arid Mediterranean regions, by decreasing greenhouse gas emissions [103,104]. Furthermore, many studies have proven that planting and incorporating green manure have multiple beneficial effects on soil health and the rhizosphere process [93,105]. The different types of green manure and their beneficial effects on the rhizosphere are shown in Table 2.
4.2. Green Manure and Their Effects on Soil Health
Soil health is a primary requirement for agriculture and plant growth, but most of the agricultural soils are poor in organic nutrients and organic matters due to the high applications of inorganic or chemical-based fertilizers to achieve high crop yields. Different agricultural techniques should be adopted; otherwise, it can become very hazardous and lead to a lack of organic matter. Improving soil health through chemical, physical, and biological properties is attributed to the integrated application of green manure and organic application due to better nutrient uptake and preservation of soil health [115]. A variety of green manures, including grains, root crops, legumes, and oil crops, can serve as vegetative covers, each offering different benefits for enhancing soil health. Therefore, growing and incorporating green manure in agricultural lands help maintain and preserve soil health and soil nutrient availability and ultimately increase soil organic matter content. Incorporation of green manure provides soil nutrients and organic matter to the soil, and organic matter plays an important role in soil biodiversity, soil microbiota, and the arrangement of soil aggregates to improve the soil structure [116,117]. Therefore, incorporating green manure leads to an increased organic matter content, which has an important role in soil health and quality. Furthermore, green manure suppresses soil-borne diseases through biofumigation. Soil-borne diseases also affect soil health, reduce crop productivity, and negatively affect soil biodiversity. Many soil-borne pathogens have negative effects on plant growth and crop production, such as Phytophthora spp., Fusarium spp., Verticillium spp., and Rhizoctonia spp. Soil-borne diseases reduce agricultural yield by as much as 50–70%, including those of wheat, vegetables, and cotton [118,119]. However, the practices of green manure in agricultural lands produces significant benefits such as increased soil nutrient availability, enhanced soil organic carbon, reduced soil compaction, improved particle aggregation and soil structure, as well as strengthened microbe activity, diversity, abundance, and suppression of soil-borne diseases.
To protect crops from soil-borne diseases, many fumigants and fungicides need to be used regularly. Later, it was noted that the use of fumigants and fungicides or any other chemical-based product causes ecological problems such as human health hazards and reduced beneficial microbes in the soil biodiversity, directly affecting soil health [120]. In this consequence, organically alternative strategies should be adopted to reduce soil-borne diseases, preserve soil health, and improve crop production. Alternatively, incorporation of green manure and cover crops are useful strategies for protecting soil health from soil-borne diseases and improving soil and plant health through biofumigation, providing nutrients and increasing beneficial microbial activity. Hao, et al. [121] reported that broccoli (B. oleracea) grown as green manure reduces the S. minor sclerotia in the rhizosphere. Whalen [122] also reported that Fusarium wilt is another soil-borne disease that reduces watermelon production in the southeastern United States. The soil-borne disease Fusarium wilts is due to Fusarium oxysporum that attacks the crop of watermelon and reduces the annual number of fruits, directly affecting the annual yield of watermelon production. For the management of Fusarium wilt disease, hairy vetch (Vicia villosa. L) green manure was incorporated. After incorporating hairy vetch, the occurrence of Fusarium wilt disease in the rhizosphere of the watermelon field was reduced, leading to an increased annual watermelon yield by 45% [123]. Papp, et al. [124] demonstrated that the cultivation of green manures influences soil microorganism environments, which are essential for sustaining soil functions and ecological system sustainability because they participate in nutrient cycling and organic matter turnover, providing multiple services. Researchers have observed that the incorporation of green manure modifies the composition and dynamics of soil fungus and bacterial communities, and encourage the beneficial microorganisms. Ntalli and Caboni [125] observed that certain cruciferous green manures, including canola (Brassica napus L.) and rape (Brassica rapa L.), upon termination, release isothiocyanates (ITCs) via the hydrolysis of glucosinolates (GSLs) during the decomposition of their tissues, serving as effective biofumigants against various soil-borne pathogens and pests. The utilization of cruciferous species as green manures may facilitate the natural management of possible soil-borne diseases. Furthermore, Waisen, et al. [126] demonstrated that brown mustard (Brassica juncea L.) and radish (Raphanus raphanistrum subsp. sativus) serve as excellent biofumigant crops that protect against plant-parasitic nematodes, while keeping the soil healthy and maintaining the strength of the nematode population structure strong. Aydınlı and Mennan [127] also found that planting arugula (Eruca sativa) and radish (R. sativus) as winter crops before planting plants that are easily damaged by the root-knot nematode Meloidogyne arenaria may lower the number of eggs laid, the gall index, and the damage that occurs, while also increasing crop yields. Leguminous green manure can substantially enrich the soil with nitrogen throughout its growth and can acidify the rhizosphere by enhancing the absorption of insoluble phosphorus into the soil [128,129]. Leguminous green manure can fix atmospheric nitrogen via symbiosis with rhizobia in root nodules [130]. Furthermore, the fixed nitrogen can be allocated to intercropped non-leguminous plants within mixed cropping systems, or it may succeed crops in rotational practices. Biological nitrogen-fixing (BNF) systems may decrease the reliance on commercial nitrogen fertilizers [131,132]. Further, Scavo, et al. [133] reported that the existence of Trifolium subterraneum for three straight years resulted in a significant rise in nitric nitrogen, ammoniacal nitrogen, and nitrogen cycle bacteria. Campiglia, et al. [134] observed similar wheat yields when they used underground clover as living mulch in intercropping systems. In addition, Guardia, et al. [135] showed that the mitigating effect of the legume green manure (vetch) mostly comes from less synthetic nitrogen being added to the next cash crop, less indirect N2O emissions from NO3– leaching, and more carbon being stored because photosynthesis is higher. Therefore, green manure and cover crops should be adopted to protect soil health, reduce soil-borne diseases as well as refining water use efficiency and many more beneficial impacts as depicted in the conceptual (Figure 3).
5. Role of Cropping Systems in Agroecosystems
Understanding the value of agroecosystems requires knowledge of soil, its different properties, and the processes that maintain its health. Soil is a complex ecosystem and is one of our most important resources due to their many vital functions, such as providing food, fuel, and fibre; recycling of nutrients that turn into beneficial nutrients for crop and plant necessities; organic matter decomposition, including plants and animal residues; regulation of water supply and quality; and provision of soil microorganisms, which are important for soil health and play vital roles in agroecosystems [136]. Unfortunately, the soil is currently being degraded rapidly due to extensive application of chemical-based fertilizers, lack of agricultural techniques, and irregular cropping are rapidly degrading the soil, but what is most concerning is that soil is a non-renewable resource at a human temporal scale [96]. Therefore, scientists and researchers have been trying to adopt alternative strategies to protect soil health, ultimately enhancing agroecosystems and rhizosphere processes. Cropping systems, such as intercropping and crop rotation practices, could serve this purpose in agricultural practices, thereby positively influencing soil health and agroecosystems [137]. Cropping systems are adopted to maximize the crop yields of agroecosystems as well as to maintain agroecosystems through soil health and quality preservation. In contrast, minimization of chemical-based products directly or indirectly affect agroecosystems and rhizosphere processes [138]. With the primary goal of soil health maintenance to ensure long-term stability of agroecosysemts and high production of crops, we need to adopt agronomic practices such as intercropping or crop rotations. Intercropping practices can improve agroecosystems by reducing the use of chemical-based fertilizers, soil pollution, and preserving soil microbial diversity.
5.1. Intercropping and Their Effects on Rhizosphere Processes and Soil Health
Intercropping, which involves the cultivation of two or more crops in a field concurrently, is a useful strategy in agronomic practice. Different crops such as cover and cash crops grow at the same time. The practice has gained global attention due to their beneficial effects for the enhancement of agroecosystems and soil health. In brief, intercropping practices can maintain agroecosystems and soil health by the reduced use of chemical-based fertilizers [139], enhancing soil-plant functions [140], suppressing the occurrence of soil-borne diseases [137], increasing soil nutrient and organic matter contents [141], and promoting the beneficial soil microorganisms, which support the rhizosphere processes [142]. Cong, et al. [143] reported that intercropping systems, including that of faba beans, corn, and wheat, had about 11%, 4%, and 23% higher belowground plant biomass as well as increased organic carbon (C) and nitrogen (N) contents than those in plant species rotations found in Gansu, China. Furthermore, de Medeiros, et al. [144] reported that intercropping with beans and pigeon pea considerably minimize black root rot (Scytalidium lignicola) in cassava by up to 50% when matched with cassava in monoculture systems, as well as enhanced the rhizosphere process, that is, increasing the soil enzymes activities, microbial biomass, and soil nutrients related to organic C and N. Increasing the use of intercropping with different plant species have been typically interconnected with the enhancement of rhizosphere processes, providing environmental benefits [145]. For example, leguminous species are advantageous for soil through providing nitrogen-fixing microorganisms and enhancing soil enzymes activities. On the other hand, non-leguminous species usually provide soil cover and nitrogen availability; such intercropping self-regulates soil nitrogen levels to improve soil nutrient use and reduce the C footprint [146]. Furthermore, Graf, et al. [147] also reported that compared to sole cropping, intercropping practices on Dactylis glomerata and Medicago sativa have led to increased shoot biomass and nitrous oxide (N2O) production rate, which suggests that understanding the intercropping strategies is helpful for the maintenance of agroecosystems and soil health. Intercropping is regaining agronomical and ecological importance because of its ability to self-regulate soil nutrients and preserve soil health. An overview of intercropping in agroecosystems and its beneficial interactions based on Brooker, et al. [148] and their outcomes are shown in Figure 4.
Furthermore, legume and cereal intercropping systems have been extensively employed to address limitations on natural resources, promote sustainable agricultural development, and guarantee food security due to their economic and ecological advantages [149]. Faba/wheat intercropping decreased nitrogen fertilizer application by 5–16% while simultaneously enhancing wheat production by 15–25% [150]. While, Luo, et al. [151] reported that a 40 percent reduction in synthetic nitrogen input, coupled with soybean intercropping, might sustain sugarcane productivity and promote sustainable soil management. Many research studies have demonstrated that belowground facilitative interactions among species can improve nutrient availability and utilization. [152] revealed that the root interactions between maize and faba beans, along with the root exudates from maize, significantly improved the symbiotic N2 fixation and nodulation in faba beans grown alongside maize. Intercropping can make crop roots release more organic acids and control the mineralization of organic phosphorus, the dissolution of inorganic phosphorus, or both. This makes more phosphorus available in the soil. Recent research shows that growing cereals and legumes together improves metabolic function and changes the structure of the soil’s microbial community through interactions between species [153]. According to Zhang, et al. [154], elevated carbon uptake by maize (Zea mays L.) promoted nutrient cycling by altering the abundance of functional groups in soil microbes and improving the stability and complexity of microbial networks in soybean/maize intercropping systems. Teshita, et al. [155] proposed that, particularly in low-nitrogen soils, intercropping maize with alfalfa (Medicago sativa L.) could enhance the structural complexity of the soil food web. Moreover, intercropping improves agricultural sustainability by utilizing crop diversity to exploit synergistic interactions across species [156]. This method improves crop yield and nitrogen utilization efficiency through similar and beneficial interactions. The rhizosphere of leguminous plants cultivates distinct microbial communities, including nitrogen-fixing bacterium (nifh) populations, via root exudates in legume-based intercropping systems, enhancing ecological benefits such as nitrogen fixation and phosphorus mobilization [157,158]. Within these approaches, maize/soybean intercropping is notable for its effective transfer of fixed nitrogen from soybean to maize, hence improving nitrogen efficiency and yields [159]. Multiple factors, such as cropping configurations and fertilization methods, affect how well maize and soybean intercropping systems work. Improving microbes is crucial for nutrient absorption and yield maximization [160,161]. Understanding how rhizosphere microbial communities react, especially in terms of their ability to fix nitrogen, would help improve intercropping patterns and fertilizer management, which would lead to more sustainable farming and food security.
5.2. Crop Rotation and Their Effects on Rhizosphere Processes and Soil
Crop rotation is a conventional and effective method for maintaining agroecosystems, managing biodiversity by improving soil health, suppressing disease and pest occurrences, and hence enhancing crop yields [162]. The efficiency and value of crop rotation are dependent upon various elements, including the types of crops utilized in the rotation, the sequence and frequency of specific crops, the duration of rotation, the agronomic history of the agricultural land, and the properties of the soil [163,164]. Recent studies reported that farmers had used crop rotation through traditional methods to manage crop pests and diseases and manage soil productivity [165,166]. Usually, these crop rotations involve growing three or four different types of crops in a sequence. However, with increasing food demand and agricultural production in recent years, many farmers grow just one or two crop species, with pesticides and mineral fertilizers contributing to compensate for the lack of crop rotations [167]. The crop rotations are not always helpful and also decline the soil quality and crop production have been reported with the number of crops grown linked with short rotation or continuous cropping, including soybean, sugarcane, maize, wheat and oilseed rape [168]. Interestingly in crop rotation, oilseed rape is very important for the rotation and even highly profitable when grown as a break crop for cereals. Crop rotations with oilseed rape and maize affect the dynamics and structure of soil plant-associated microbial communities, are known beneficial for soil health via suppression of soil-borne diseases and plant growth promotion [169]. Microbial species living nearby or associated with plants are directly influenced by the root’s architecture and the chemical characteristics of root exudates when we rotate with maize or oilseed rape crops [170]. The direct influence of plant roots around soil is known as the rhizosphere. The rhizosphere is the habitat of plant beneficial microorganisms and plants pathogens [171]. With the research evidence of Benitez, Osborne and Lehman [169], crop rotations promote beneficial plant growth-promoting microbes and affect the soil microbial community in the rhizosphere, and influences maize seedling growth characteristics.
Furthermore, rotating crops with grain legumes can significantly improve the yield and protein content of subsequent wheat crops, owing to the increased nitrogen availability in the soil from legume biological fixation [97]. Leguminous crops (such as peas and chickpeas) and various chickpea genotypes (cultivars) in rotation can alter soil functional microbial populations and affect the productivity of pulse crops and subsequent wheat crops. Different crops can produce root exudates and distinct residues that enhance soil microbial diversity and activity, as well as microbial biomass, nitrogen and carbon cycling [164,172,173]. Modifications in crop rotation duration and frequency of identical crops throughout time might influence the prevalence of root rot diseases and improve crop production and soil health. Short series rotations exhibit heightened sensitivity to host-specific diseases, resulting in poorer yields compared to lengthier series rotations [167,174]. While, Lupwayi, et al. [175] found that wheat phase of a five-year rotation exhibited greater microbial biomass in both rhizosphere and bulk soil compared to three-year rotation of wheat phase, likely due to increased carbon inputs from crop residues. In Canada particularly western part, two phases of pea in a four-year rotation increased soil nitrogen levels, whereas three legume phases markedly altered the function and composition of the rhizosphere bacterial community in comparison to continuous wheat cultivation and growth [176]. Increasing the rate of one crop in rotation can be detrimental for soil health. For example, Bainard, et al. [177] observed that a higher pulse phase in rotation caused host-specific fungal pathogens to build up in the soil, which could make rotation less beneficial for crop yield and soil health. Meanwhile, an additional crucial factor in crop rotation design is the ability of soil-borne diseases to utilize alternate crops as hosts or to remain dormant in the soil for extended periods, as well as the response of these crops to disease. Utilizing non-host plants in crop rotations to manage diseases transmitted by the soil is critical for mitigating yield losses, particularly given that many pathogens can persist in the soil for extended periods as spores or other latent forms in the absence of their preferred host plants [167,178]. For example, Nayyar, et al. [179] reported that Fusarium root rot in peas cultivated in rotation on the Canadian prairie was associated with a restricted soil microbial community and reduced populations of arbuscular mycorrhizal fungi and beneficial bacteria. In certain instances, continuous cropping with enhanced crop diversification elevated the population of antagonistic soil microorganisms, hence lowering the populations of soil pathogens and reducing the “take-all” effect in wheat. Typically, incorporating three or more crops within a cropping system can improves soil health and optimizes the productivity of crops [166,167]. However, the benefits of crop rotation are practical; understanding the specific effects of crop systems and how they affect subsequent crops will encourage and adopt a worthy crop rotation, which will promote soil health and increase crop productivity. These findings make it easier to create customized interventions that use microbial populations to maintain soil health, decrease the emissions of greenhouse gases (GHGs) and crop production, and the efficient use of resources in intercropping systems. According to references, we propose intercropping in agroecosystems, as depicted in the conceptual (Figure 5).
6. Indicators for Assessing Soil Health in Agroecosystems
Soil is a highly intricate and complex, multifunctional system including gaseous, liquid, and solid elements that interact through various chemical, physical, and biological processes. A healthy soil sustains agroecosystems and facilitates the provision of essential services to ecosystem [180]. Moreover, soil health typically refers to the capacity of soil to operate as a crucial living system that supports biological productivity, preserves both soil and plant health, enhances environmental quality, and maintains an agroecosystem [181]. In order to evaluate and sustain agroecosystems, it is essential to consider the biological, physical, and chemical functions and traits of the soil, particularly the biological ones that serve as sensitive indicators of soil health and quality [182]. Microbiological and biochemical markers show that the variety and activity of soil microbes are important for the long-term health of agroecosystems because they keep soil health functions going, like cycling carbon and nutrients [183]. Microbial indicators exhibit more sensitivity than physical traits to environmental alterations such as soil usage and management; hence, they enable early detection and forecasting of changes and disturbances in environmental sustainability [184]. Related studies recognize soil microbial biomass, which includes fungi, bacteria, and algae, as a primary biological indicator of soil health and as a crucial source of nutrient cycling and delivery relative to plant demand [185]. Additionally, through photosynthesis, plants fix and transfer carbon as carbohydrates into the food web, making it one of the most critical biological processes on Earth [183]. Both agricultural and forestry soils widely use soil respiration and microbial biomass as bio-indicators of soil health.
We expect biochemical indicators to effectively integrate the combined impact on soil’s chemical, physical, and biological processes and characteristics, making them suitable for diverse management and environmental conditions. This review aims to elucidate and augment our focus on soil quality and related research, particularly with biochemical soil quality indicators such as enzymatic and microbial activity [186]. Due to their role in nutrient cycling through microbial processes, soil enzyme activities frequently serve as sensitive indicators of soil biochemical quality and as measures of ecological quality. They can effectively respond to both anthropogenic and natural alterations in soil and are easily quantifiable [187]. However, soil enzymes are important in functioning of soil ecosystems, including nutrient and carbon cycling, and maintenance [188]. For example soil enzymes have an important role in C (β-glucosidase and galactosidase), N (urease), P (acid phosphatases and alkaline) cycling, and these enzymes play crucial roles in breakdown of OM and release nutrient, which are important for soil health and plant growth promotion. Dick and Tabatabai [189] reported that soil enzymes serve as reliable indicators of soil quality because of their close association with soil biological processes, their ease of measurement, and their swift response to alterations in soil management. In summary, soil enzymes are valuable tools for assessing both long and short-term changes in soil quality and management practices.
Furthermore, indicators of soil health for sustaining agroecosystems need to be associated with soil processes and responsive to alterations in management and environmental conditions. Soil biological traits, such as microbial biomass and its activities, serve as sensitive and rapid indicators that capture responses and data from diverse environmental conditions [190,191]. We use soil biological metrics, such as microbial density, activity, and biomass, as indicators of soil health [192]. Furthermore, a specific drawback of employing soil microbial characteristics as soil health indicators is the technological limitations encountered in the research of soil microbial populations. Molecular techniques that are more advanced, especially next-generation sequencing (NGS) methods (such as shotgun sequencing for structural and amplicon sequencing and functional microbial diversity investigations), have made it possible to study the part of soil microbial communities that can’t be grown in a lab. This is because most soil microorganisms don’t grow on media or in a lab setting. However, while analyzing the data, we must acknowledge the limitations of these novel methodologies. In conclusion, we can use soil to focus on agroecosystem maintenance and enhance rhizosphere processes; soil health indicators are used to inform management techniques, optimize fertilizer application, and promote sustainable crop productivity.
7. Integrated Agricultural Practices for Improving Rhizospheric Processes
Integrated agricultural management established soil properties (biological, chemical, and physical properties) that influence nutrients cycling, soil microbial community composition and structure, and enhance the rhizospheric process [193]. The disturbance of rhizosphere processes and depletion of organic matter are restraining agricultural productivity around the globe via improper agricultural practices. To reduce adverse effects, integrative biofertilizers, crop rotation, green manuring, intercropping, and mulching practices should be promoted and tested worldwide to improve the rhizosphere processes and agroecosystems resilience. In short about integrated agricultural practices (IAP), are based on the integration of different crops, livestock wastes, and fertilizers into production systems that, through valuable management practices, maintain a high level of soil fertility, productivity, and quality and pursue to reduce external inputs of agrochemical and fertilizers, and enhance the functions of soil inside biological cycles and its processes. Mixed crop-livestock and other available resources such as seed primed, the basis of integrated agricultural practices, allow the most efficient and effective use of natural resources and biological cycles to improve belowground activities [194]. Sarkar, et al. [195] reported that microbial-assisted nutrient management is a sustainable, eco-friendly, and cost-effective option under integrated agricultural systems. The usage of less amount of agrochemicals but provides support organically or naturally to protect the environment from nutrients denitrification or runoff, land degradation, and soil pollution. Duarah, et al. [196] stated that reducing the amount of synthetic NPK fertilizer application and seed priming (with bacteria) can significantly improve the nitrogen use efficiency. While, Entesari, et al. [197] witnessed that bio-priming seeds of soybean with Trichoderma sp., prior to planting enhanced the nutrient status of crop and improve the crop yield. Phosphorus (P), an essential nutrient, is often fixed in the soil and forms complexes with metals, limiting its availability to plants. However, certain fungal and bacterial communities have a trait called P solubilization, which allows them to convert unavailable or fixed phosphorus into forms that plants can absorb. Kim, et al. [198] reported that incorporating green manure can help re- mobilize phosphorus and make non-exchangeable potassium available, circulating nutrients more efficiently. This integrated approach also enhances phosphorus availability, which is crucial for both plant growth and the activity of soil microorganisms. However, advantages of IAP are the selection of crops and available resources into creative integrated agricultural systems that can effectively achieve following points: minimize the disease and insect problems, minimize the outputs and reduces the energy requirements, reduces the need for agrochemical and pesticides, and also reduces the climate and economic fluctuations risks. Improved rhizosphere processes promote healthy soil, allowing agroecosystems to continue providing ecosystem services that benefit humans and animals. Thus, optimizing the rhizosphere is essential for the agroecosystem’s resilience and future food and fiber production. To achieve these benefits are significant for the improvement of the rhizospheric processes and agricultural systems.
8. Concluding Remarks and Future Directions
Understanding the significance of the rhizosphere and its interactions with microorganisms holds tremendous potential for fostering sustainable agroecosystems. The rhizosphere provides an ideal environment for a variety of microbes that enhance plant growth by improving nutrient availability, controlling pathogens, facilitating hormone signaling, and altering soil structure. Exudates from plant roots, green manure cultivation, and intercropping of various plant species are key drivers influencing microbial community dynamics in the rhizosphere. Emerging research underscores the importance of rhizosphere engineering, coupled with green manures and intercropping, in delivering essential life-support functions for host plants, including nutrient uptake, growth promotion, tolerance to abiotic stress, disease resistance, and the overall maintenance of agroecosystems. The cultivation of green manures offers multifaceted benefits: improving soil health, enhancing nutrient cycling, increasing carbon sequestration, reducing greenhouse gas emissions, decreasing reliance on synthetic fertilizers, and rebuilding rhizosphere microbial communities. These practices also contribute to economic advantages by stabilizing crop yields and enhancing soil fertility through organic matter accumulation. Additionally, green manures improve water conservation, soil structure, and pest and weed management, all of which are critical for supporting rhizosphere functions and maintaining agroecosystem stability.
A thriving agroecosystem upholds soil fertility, prompting the production of food and fiber, which is crucial for both human and animal health. Optimizing rhizosphere activities such as nutrient cycling and beneficial microbial interactions improves soil structure, nutrient availability, and plant growth. These enhancements increase tolerance to environmental impacts and reduce synthetic inputs, ensuring agricultural system sustainability. Improved rhizosphere processes promote healthy soil, allowing agroecosystems to continue providing ecosystem services that benefit humans and animals. Thus, optimizing the rhizosphere is essential for the agroecosystem’s resilience and future food and fiber production.
There is growing interest in utilizing microbial inoculants, microbial-derived products, green manures, and intercropping systems to bolster rhizosphere activity, enhance crop productivity, and sustain soil quality. These methods have the potential to enhance the long-term sustainability of low-input agroecosystems, contributing to food security and environmental resilience. The findings from this review provide valuable insights to agronomists, soil scientists, and crop managers on optimizing rhizosphere processes to improve agroecosystem resilience. In closing, the rhizosphere will be central to future ecological innovations in agriculture, driving both resilience and sustainability. However, further experimental research is required to fully unlock the prospective of the rhizosphere. Future studies should focus on integrating advanced technologies, such as microbial genomics and precision agriculture, to optimize rhizosphere management for more resilient and sustainable agroecosystems. The development of smart agricultural strategies, coupled with policy support, will be vital for addressing the growing challenges posed by climate change and soil degradation.
Author Contributions
W.A. and R.K. designed and wrote the manuscript. K.D. R.K. reviewed and supervision. A.M., J.S., and J.F., edited and wrote the manuscript.
Funding
This work has not received any funding and grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data was used for the research described in the article.
Acknowledgments
Figures for the review manuscript were created by BioRender (https://biorender.com).
Conflicts of Interest
The authors declare no conflict of interest. The authors declare that this article is original research and has not been submitted previously to any journal. We hereby transfer the copyright ownership to the International Journal of Molecular Sciences for due process of review and editing. Besides, the authors listed below declare no conflict of interest and competing interests.
References
- Omotayo, O.P.; Babalola, O.O. Resident rhizosphere microbiome’s ecological dynamics and conservation: Towards achieving the envisioned Sustainable Development Goals, a review. International Soil and Water Conservation Research 2020. [Google Scholar] [CrossRef]
- Liang, H.; Wang, X.; Yan, J.; Luo, L. Characterizing the intra-vineyard variation of soil bacterial and fungal communities. Front microbiol 2019, 10, 1239. [Google Scholar]
- Schmidt, J.E.; Vannette, R.L.; Igwe, A.; Blundell, R.; Casteel, C.L.; Gaudin, A.C. Effects of agricultural management on rhizosphere microbial structure and function in processing tomato plants. AEM 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Asghar, W.; Kataoka, R. Co-application of Green Manure and Trichoderma spp. Induced Plant Growth Promotion by Nutrient Improvement and Increased Fungal Biomass in Soil. Agric. Res 2024, 1–10. [Google Scholar]
- Asghar, W.; Craven, K.D.; Kataoka, R.; Mahmood, A.; Asghar, N.; Raza, T.; Iftikhar, F. The application of Trichoderma spp., an old but new useful fungus, in sustainable soil health intensification: A comprehensive strategy for addressing challenges. Plant Stress 2024, 100455. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Sturz, A.; Carter, M.; Johnston, H. A review of plant disease, pathogen interactions and microbial antagonism under conservation tillage in temperate humid agriculture. Soil Till Res 1997, 41, 169–189. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, J.; Zhang, D.; Cheng, H.; Hao, B.; Cao, A.; Yan, D.; Wang, Q.; Li, Y. Beneficial effect on the soil microenvironment of Trichoderma applied after fumigation for cucumber production. PLoS One 2022, 17, e0266347. [Google Scholar] [CrossRef]
- Jiang, S.-Q.; Yu, Y.-N.; Gao, R.-W.; Wang, H.; Zhang, J.; Li, R.; Long, X.-H.; Shen, Q.-R.; Chen, W.; Cai, F. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial community in a tomato cultivated coastal saline soil. Science of the total environment 2019, 687, 601–609. [Google Scholar] [CrossRef]
- Bajsa, N.; Morel, M.A.; Braña, V.; Castro-Sowinski, S. The effect of agricultural practices on resident soil microbial communities: focus on biocontrol and biofertilization. Mol Microb Ecol Rhizosphere 2013, 2, 687–700. [Google Scholar]
- Meena, H.; Meena, R.S.; Rajput, B.S.; Kumar, S. Response of bio-regulators to morphology and yield of clusterbean [Cyamopsis tetragonoloba (L.) Taub.] under different sowing environments. Journal of Applied and Natural Science 2016, 8, 715–718. [Google Scholar] [CrossRef]
- Dubey, R.K.; Dubey, P.K.; Chaurasia, R.; Singh, H.B.; Abhilash, P.C. Sustainable agronomic practices for enhancing the soil quality and yield of Cicer arietinum L. under diverse agroecosystems. Journal of environmental management 2020, 262, 110284. [Google Scholar] [PubMed]
- Bruinsma, M.; Kowalchuk, G.; Van Veen, J. Effects of genetically modified plants on microbial communities and processes in soil. Biol Fertil Soils 2003, 37, 329–337. [Google Scholar] [CrossRef]
- Sharma, S.K.; Ramesh, A.; Sharma, M.P.; Joshi, O.P.; Govaerts, B.; Steenwerth, K.L.; Karlen, D.L. Microbial community structure and diversity as indicators for evaluating soil quality. In Biodiversity, biofuels, agroforestry and conservation agriculture; Springer, 2010; pp. 317–358. [Google Scholar]
- Asghar, W.; Kataoka, R. Green manure incorporation accelerates enzyme activity, plant growth, and changes in the fungal community of soil. Arch Microbiol 2022, 204, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, X.; Liao, Y.; Lu, Y.; Nie, J.; Cao, W. Co-incorporation of rice straw and green manure benefits rice yield and nutrient uptake. Crop Sci. 2019, 59, 749–759. [Google Scholar] [CrossRef]
- Khan, M.I.; Gwon, H.S.; Alam, M.A.; Song, H.J.; Das, S.; Kim, P.J. Short term effects of different green manure amendments on the composition of main microbial groups and microbial activity of a submerged rice cropping system. Appl Soil Ecol 2020, 147, 103400. [Google Scholar] [CrossRef]
- York, L.M.; Carminati, A.; Mooney, S.J.; Ritz, K.; Bennett, M.J. The holistic rhizosphere: integrating zones, processes, and semantics in the soil influenced by roots. Journal of experimental botany 2016, 67, 3629–3643. [Google Scholar] [CrossRef]
- Solomon, W.; Janda, T.; Molnár, Z. Unveiling the significance of rhizosphere: implications for plant growth, stress response, and sustainable agriculture. Plant Physiology and Biochemistry 2023, 108290. [Google Scholar] [CrossRef]
- De Luna, L.Z.; Stubbs, T.L.; Kennedy, A.C.; Kremer, R.J. Deleterious bacteria in the rhizosphere. roots and soil management: interactions between roots and the soil 2005, 48, 233-261.
- RUEss, L.; Ferris, H. Decomposition pathways and successional changes. Nematology Monographs and Perspectives 2004, 2, 547–556. [Google Scholar]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sc 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Asghar, W.; Kondo, S.; Iguchi, R.; Mahmood, A.; Kataoka, R. Agricultural utilization of unused resources: liquid food waste material as a new source of plant growth-promoting microbes. Agronomy 2020, 10, 954. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; Tuinen, D.v.; Berg, G. Plant-driven selection of microbes; Springer, 2009. [Google Scholar]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front Plant Sci 2017, 8, 1617. [Google Scholar] [CrossRef] [PubMed]
- Barea, J. Future challenges and perspectives for applying microbial biotechnology in sustainable agriculture based on a better understanding of plant-microbiome interactions. J. Soil Sci. Plant Nutr 2015, 15, 261–282. [Google Scholar] [CrossRef]
- He, Z.; Shang, X.; Zhang, T.; Yun, J. Ca and mg stimulate protein synthesis in maize kernel through the action of endogenous hormones and defense enzymes. Plant Physiology and Biochemistry 2024, 206, 108280. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hossain, M.S.; Akter, M. Challenges faced by plant growth-promoting bacteria in field-level applications and suggestions to overcome the barriers. Physiological and Molecular Plant Pathology 2023, 126, 102029. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering With Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Frontiers in Sustainable Food Systems 2021, 5, 16. [Google Scholar] [CrossRef]
- de Graaff, M.-A. Interactions between plants and soil nutrient cycling under elevated CO2; 2007.
- Joseph, S.; Downie, A.; Munroe, P.; Crosky, A.; Lehmann, J. Biochar for carbon sequestration, reduction of greenhouse gas emissions and enhancement of soil fertility; a review of the materials science. In Proceedings of Proceedings of the Australian combustion symposium; pp. 130–133.
- Yanai, Y.; Toyota, K.; Okazaki, M. Effects of charcoal addition on N2O emissions from soil resulting from rewetting air-dried soil in short-term laboratory experiments. Soil Sci Plant Nutr 2007, 53, 181–188. [Google Scholar] [CrossRef]
- Laird, D.A. The charcoal vision: a win–win–win scenario for simultaneously producing bioenergy, permanently sequestering carbon, while improving soil and water quality. Agronomy journal 2008, 100, 178–181. [Google Scholar] [CrossRef]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; De Courcelles, V.d.R.; Singh, K. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric Ecosyst Environ 2013, 164, 80–99. [Google Scholar] [CrossRef]
- Yuan, P.; Wang, J.; Pan, Y.; Shen, B.; Wu, C. Review of biochar for the management of contaminated soil: Preparation, application and prospect. Science of the Total Environment 2019, 659, 473–490. [Google Scholar] [CrossRef] [PubMed]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Neogi, S.; Dutta, T.; Powel, M.A.; Banik, P. The impact of biochar on soil carbon sequestration: meta-analytical approach to evaluating environmental and economic advantages. Journal of environmental management 2019, 250, 109466. [Google Scholar] [CrossRef] [PubMed]
- Yazdanpanah, N. CO 2 emission and structural characteristics of two calcareous soils amended with municipal solid waste and plant residue. Solid Earth 2016, 7, 105–114. [Google Scholar] [CrossRef]
- Kenney, I.; Blanco-Canqui, H.; Presley, D.R.; Rice, C.W.; Janssen, K.; Olson, B. Soil and crop response to stover removal from rainfed and irrigated corn. Gcb Bioenergy 2015, 7, 219–230. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Scheer, C.; Rowlings, D.W.; Grace, P.R. Rice husk biochar and crop residue amendment in subtropical cropping soils: effect on biomass production, nitrogen use efficiency and greenhouse gas emissions. Biol Fertil Soils 2016, 52, 261–270. [Google Scholar] [CrossRef]
- Asghar, W.; Kataoka, R. Effect of co-application of Trichoderma spp. with organic composts on plant growth enhancement, soil enzymes and fungal community in soil. Arch Microbiol 2021, 10.1007/s00203-021-02413-4. [Google Scholar] [CrossRef]
- Kataoka, R.; Nagasaka, K.; Tanaka, Y.; Yamamura, H.; Shinohara, S.; Haramoto, E.; Hayakawa, M.; Sakamoto, Y. Hairy vetch (Vicia villosa), as a green manure, increases fungal biomass, fungal community composition, and phosphatase activity in soil. Appl Soil Ecol 2017, 117, 16–20. [Google Scholar] [CrossRef]
- Ayaz, M.; Feizienė, D.; Tilvikienė, V.; Akhtar, K.; Stulpinaitė, U.; Iqbal, R. Biochar Role in the Sustainability of Agriculture and Environment. Sustainability 2021, 13, 1330. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for environmental management: an introduction; Routledge, 2015. [Google Scholar]
- Lu, L.; Yu, W.; Wang, Y.; Zhang, K.; Zhu, X.; Zhang, Y.; Wu, Y.; Ullah, H.; Xiao, X.; Chen, B. Application of biochar-based materials in environmental remediation: from multi-level structures to specific devices. Biochar 2020, 2, 1–31. [Google Scholar] [CrossRef]
- Brewer, C.E.; Chuang, V.J.; Masiello, C.A.; Gonnermann, H.; Gao, X.; Dugan, B.; Driver, L.E.; Panzacchi, P.; Zygourakis, K.; Davies, C.A. New approaches to measuring biochar density and porosity. Biomass and bioenergy 2014, 66, 176–185. [Google Scholar] [CrossRef]
- Arif, M.; Ali, S.; Ilyas, M.; Riaz, M.; Akhtar, K.; Ali, K.; Adnan, M.; Fahad, S.; Khan, I.; Shah, S. Enhancing phosphorus availability, soil organic carbon, maize productivity and farm profitability through biochar and organic–inorganic fertilizers in an irrigated maize agroecosystem under semi-arid climate. Soil Use Manag 2021, 37, 104–119. [Google Scholar] [CrossRef]
- Uzoma, K.; Inoue, M.; Andry, H.; Fujimaki, H.; Zahoor, A.; Nishihara, E. Effect of cow manure biochar on maize productivity under sandy soil condition. Soil Use Manag 2011, 27, 205–212. [Google Scholar] [CrossRef]
- Lu, H.; Yan, M.; Wong, M.H.; Mo, W.Y.; Wang, Y.; Chen, X.W.; Wang, J.-J. Effects of biochar on soil microbial community and functional genes of a landfill cover three years after ecological restoration. Science of The Total Environment 2020, 717, 137133. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.; Kuhn, T.K.; Macdonald, L.M.; Maddern, T.M.; Murphy, D.V.; Hall, P.A.; Singh, B.P.; Baumann, K.; Krull, E.S.; Baldock, J.A. Microbial utilisation of biochar-derived carbon. Science of the Total Environment 2013, 465, 288–297. [Google Scholar] [CrossRef]
- Marschner, P.; Crowley, D.; Rengel, Z. Rhizosphere interactions between microorganisms and plants govern iron and phosphorus acquisition along the root axis–model and research methods. Soil Biol Biochem 2011, 43, 883–894. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Sun, R.; Zhang, X.-X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol Biochem 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Lagos, L.; Maruyama, F.; Nannipieri, P.; Mora, M.; Ogram, A.; Jorquera, M. Current overview on the study of bacteria in the rhizosphere by modern molecular techniques: a mini‒review. J. Soil Sci. Plant Nutr 2015, 15, 504–523. [Google Scholar]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. PNAS 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Mahmood, A.; Kataoka, R.; Takagi, K. Dichlorodiphenyltrichloroethane (DDT) degradation by Streptomyces sp. isolated from DDT contaminated soil. Bioremediation Journal 2020, 1–14. [Google Scholar]
- Asghar, W.; Kataoka, R. Fungal volatiles from green manure-incorporated soils promote the growth of lettuce (Lactuca sativa) and mediate antifungal activity against Fusarium oxysporum in vitro. Plant Soil 2023, 1–12. [Google Scholar] [CrossRef]
- Selvasekaran, P.; Chidambaram, R. Agriculturally Important Fungi for Crop Protection. In Agriculturally Important Fungi for Sustainable Agriculture; Springer, 2020; pp. 1–53. [Google Scholar]
- Ito, K.; Mahmood, A.; Kataoka, R.; Takagi, K. Dichlorodiphenyltrichloroethane (DDT) degradation by Streptomyces sp. isolated from DDT contaminated soil. Bioremediation Journal 2021, 25, 148–158. [Google Scholar]
- Mahmood, A.; Kataoka, R. Metabolite profiling reveals a complex response of plants to application of plant growth-promoting endophytic bacteria. Microbiol Res 2020, 234, 126421. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Iguchi, R.; Kataoka, R. Multifunctional food waste fertilizer having the capability of Fusarium-growth inhibition and phosphate solubility: A new horizon of food waste recycle using microorganisms. Waste Manage 2019, 94, 77–84. [Google Scholar] [CrossRef]
- Da Costa, P.B.; Beneduzi, A.; de Souza, R.; Schoenfeld, R.; Vargas, L.K.; Passaglia, L.M. The effects of different fertilization conditions on bacterial plant growth promoting traits: guidelines for directed bacterial prospection and testing. Plant Soil 2013, 368, 267–280. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatalysis and Agricultural Biotechnology 2020, 23, 101487. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.A. Future of bio-fertilizers in Indian agriculture: an overview. International Journal of Agricultural and Food Research 2015, 3. [Google Scholar] [CrossRef]
- Murali, M.; Amruthesh, K.; Sudisha, J.; Niranjana, S.; Shetty, H. Screening for plant growth promoting fungi and their ability for growth promotion and induction of resistance in pearl millet against downy mildew disease. Journal of Phytology 2012, 4, 30–36. [Google Scholar]
- Murali, M.; Amruthesh, K.N. Plant Growth-promoting Fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J Phytopathol 2015, 163, 743–754. [Google Scholar] [CrossRef]
- Sayyed, R.; Chincholkar, S.; Reddy, M.; Gangurde, N.; Patel, P. Siderophore producing PGPR for crop nutrition and phytopathogen suppression. In Bacteria in agrobiology: disease management; Springer, 2013; pp. 449–471. [Google Scholar]
- Hossain, M.M.; Sultana, F.; Kubota, M.; Koyama, H.; Hyakumachi, M. The plant growth-promoting fungus Penicillium simplicissimum GP17-2 induces resistance in Arabidopsis thaliana by activation of multiple defense signals. Plant and cell physiology 2007, 48, 1724–1736. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Sultana, F.; Miyazawa, M.; Hyakumachi, M. The plant growth-promoting fungus Penicillium spp. GP15-1 enhances growth and confers protection against damping-off and anthracnose in the cucumber. Journal of oleo science 2014, 63, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Abdel-lateif, K.S. Trichoderma as biological control weapon against soil borne plant pathogens. Afr. J. Biotechnol 2017, 16, 2299–2306. [Google Scholar]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Cervantes-Badillo, M.G.; Rosales-Saavedra, M.T.; Islas-Osuna, M.A.; Casas-Flores, S. The plant growth-promoting fungus Aspergillus ustus promotes growth and induces resistance against different lifestyle pathogens in Arabidopsis thaliana. Journal of microbiology and biotechnology 2011, 21, 686–696. [Google Scholar] [CrossRef]
- Cavello, I.A.; Crespo, J.M.; García, S.S.; Zapiola, J.M.; Luna, M.F.; Cavalitto, S.F. Plant growth promotion activity of keratinolytic fungi growing on a recalcitrant waste known as “Hair Waste”. Biotechnology research international 2015. [Google Scholar] [CrossRef]
- Yamagiwa, Y.; Inagaki, Y.; Ichinose, Y.; Toyoda, K.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 emits β-caryphyllene, which promotes plant growth and induces resistance. J. Gen. Plant Pathol 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Kumar, V.; Behl, R.K.; Narula, N. Establishment of phosphate-solubilizing strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat cultivars under green house conditions. Microbiol Res 2001, 156, 87–93. [Google Scholar] [CrossRef]
- Joseph, B.; Ranjan Patra, R.; Lawrence, R. Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). International Journal of Plant Production 2012, 1, 141–152. [Google Scholar]
- Ahemad, M.; Khan, M.S. Alleviation of fungicide-induced phytotoxicity in greengram [Vigna radiata (L.) Wilczek] using fungicide-tolerant and plant growth promoting Pseudomonas strain. Saudi journal of biological sciences 2012, 19, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S. Plant growth promoting rhizobacteria. Resonance 2013, 18, 275–281. [Google Scholar] [CrossRef]
- Sunpapao, A.; Chairin, T.; Ito, S.-i. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings. Biocontrol 2018, 123, 36–42. [Google Scholar] [CrossRef]
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere signaling: Insights into plant–rhizomicrobiome interactions for sustainable agronomy. Microorganisms 2022, 10, 899. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. Journal of Experimental Botany 2021, 72, 57–69. [Google Scholar] [CrossRef]
- O’Banion, B.S.; O’Neal, L.; Alexandre, G.; Lebeis, S.L. Bridging the gap between single-strain and community-level plant-microbe chemical interactions. MPMI 2020, 33, 124–134. [Google Scholar] [CrossRef]
- Rizaludin, M.S.; Stopnisek, N.; Raaijmakers, J.M.; Garbeva, P. The chemistry of stress: understanding the ‘cry for help’of plant roots. Metabolites 2021, 11, 357. [Google Scholar] [CrossRef]
- Jacoby, R.P.; Chen, L.; Schwier, M.; Koprivova, A.; Kopriva, S. Recent advances in the role of plant metabolites in shaping the root microbiome. F1000Research 2020, 9. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: from community assembly to plant health. Nature reviews microbiology 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Mönchgesang, S.; Strehmel, N.; Schmidt, S.; Westphal, L.; Taruttis, F.; Müller, E.; Herklotz, S.; Neumann, S.; Scheel, D. Natural variation of root exudates in Arabidopsis thaliana-linking metabolomic and genomic data. Sci Rep 2016, 6, 29033. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, S.; Kumar, P. Rhizosphere microbiome: revisiting the synergy of plant-microbe interactions. Annals of Microbiology 2019, 69, 307–320. [Google Scholar] [CrossRef]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Science 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zhou, Q.; Hao, Y.; Huang, A.C. Crafting the plant root metabolome for improved microbe-assisted stress resilience. New Phytologist 2022, 234, 1945–1950. [Google Scholar] [CrossRef]
- Toom, M.; Tamm, S.; Talgre, L.; Tamm, I.; Tamm, Ü.; Narits, L.; Hiiesalu, I.; Mäe, A.; Lauringson, E. The effect of cover crops on the yield of spring barley in Estonia. Agriculture 2019, 9, 172. [Google Scholar] [CrossRef]
- Florentín, M.A.; Peñalva, M.; Calegari, A.; Derpsch, R.; McDonald, M. Green manure/cover crops and crop rotation in conservation agriculture on small farms. Integrated Crop Management 2010, 12. [Google Scholar]
- Ntakirutimana, L.; Li, F.; Huang, X.; Wang, S.; Yin, C. Green manure planting incentive measures of local authorities and farmers’ perceptions of the utilization of rotation fallow for sustainable agriculture in Guangxi, China. Sustainability 2019, 11, 2723. [Google Scholar] [CrossRef]
- Yang, T.; Siddique, K.H.; Liu, K. Cropping systems in agriculture and their impact on soil health-A review. Global Ecology and Conservation 2020, e01118. [Google Scholar] [CrossRef]
- Gan, Y.; Miller, P.; McConkey, B.; Zentner, R.; Stevenson, F.; McDonald, C. Influence of diverse cropping sequences on durum wheat yield and protein in the semiarid northern Great Plains. Agronomy Journal 2003, 95, 245–252. [Google Scholar] [CrossRef]
- Chavarria, D.N.; Verdenelli, R.A.; Serri, D.L.; Restovich, S.B.; Andriulo, A.E.; Meriles, J.M.; Vargas-Gil, S. Effect of cover crops on microbial community structure and related enzyme activities and macronutrient availability. Eur J Soil Biol 2016, 76, 74–82. [Google Scholar] [CrossRef]
- Balota, E.L.; Calegari, A.; Nakatani, A.S.; Coyne, M.S. Benefits of winter cover crops and no-tillage for microbial parameters in a Brazilian Oxisol: A long-term study. Agric Ecosyst Environ 2014, 197, 31–40. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol Biochem 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Khan, M.I.; Hwang, H.Y.; Kim, G.W.; Kim, P.J.; Das, S. Microbial responses to temperature sensitivity of soil respiration in a dry fallow cover cropping and submerged rice mono-cropping system. Appl Soil Ecol 2018, 128, 98–108. [Google Scholar] [CrossRef]
- Quintarelli, V.; Radicetti, E.; Allevato, E.; Stazi, S.R.; Haider, G.; Abideen, Z.; Bibi, S.; Jamal, A.; Mancinelli, R. Cover crops for sustainable cropping systems: a review. Agriculture 2022, 12, 2076. [Google Scholar] [CrossRef]
- Lazcano, C.; Gonzalez-Maldonado, N.; Yao, E.H.; Wong, C.T.; Merrilees, J.J.; Falcone, M.; Peterson, J.D.; Casassa, L.F.; Decock, C. Sheep grazing as a strategy to manage cover crops in Mediterranean vineyards: Short-term effects on soil C, N and greenhouse gas (N2O, CH4, CO2) emissions. Agric Ecosyst Environ 2022, 327, 107825. [Google Scholar] [CrossRef]
- Fiorini, A.; Maris, S.C.; Abalos, D.; Amaducci, S.; Tabaglio, V. Combining no-till with rye (Secale cereale L.) cover crop mitigates nitrous oxide emissions without decreasing yield. Soil Till Res 2020, 196, 104442. [Google Scholar] [CrossRef]
- Doltra, J.; Olesen, J.E. The role of catch crops in the ecological intensification of spring cereals in organic farming under Nordic climate. Eur J Agron 2013, 44, 98–108. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, A.; Kahlon, C.; Brar, A.; Grover, K.; Dia, M.; Steiner, R. The Role of Cover Crops towards Sustainable Soil Health and Agriculture—A Review Paper. American Journal of Plant Sciences 2018, 09, 1935–1951. [Google Scholar] [CrossRef]
- Brennan, R.J.B.; Glaze-Corcoran, S.; Robert, W.; HASHeMi, M. Biofumigation: An alternative strategy for the control of plant parasitic nematodes. Journal of Integrative Agriculture 2020, 19, 1680–1690. [Google Scholar] [CrossRef]
- Srivastava, J.; Ghatak, A. Biofumigation: a control method for the soil-borne diseases. International Journal of Plant Protection 2017, 10, 453–460. [Google Scholar] [CrossRef]
- Dubey, R.K.; Dubey, P.K.; Abhilash, P. Sustainable soil amendments for improving the soil quality, yield and nutrient content of Brassica juncea (L.) grown in different agroecological zones of eastern Uttar Pradesh, India. Soil Till Res 2019, 195, 104418. [Google Scholar] [CrossRef]
- Clark, A. Managing cover crops profitably; Diane Publishing, 2008. [Google Scholar]
- Thapa, V.R.; Ghimire, R.; Acosta-Martínez, V.; Marsalis, M.A.; Schipanski, M.E. Cover crop biomass and species composition affect soil microbial community structure and enzyme activities in semiarid cropping systems. Appl Soil Ecol 2021, 157, 103735. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Ncube, B.; Mulidzi, R.; Lewu, F.B. Management impact and benefit of cover crops on soil quality: A review. Soil Till Res 2020, 204, 104717. [Google Scholar] [CrossRef]
- Magdoff, F.; Van Es, H. Building soils for better crops; Sustainable Agriculture Network Beltsville, 2000; Volume 4. [Google Scholar]
- Boselli, R.; Fiorini, A.; Santelli, S.; Ardenti, F.; Capra, F.; Maris, S.C.; Tabaglio, V. Cover crops during transition to no-till maintain yield and enhance soil fertility in intensive agro-ecosystems. Field Crops Res 2020, 255, 107871. [Google Scholar] [CrossRef]
- Patra, D.; Anwar, M.; Chand, S. Integrated nutrient management and waste recycling for restoring soil fertility and productivity in Japanese mint and mustard sequence in Uttar Pradesh, India. Agric Ecosyst Environ 2000, 80, 267–275. [Google Scholar] [CrossRef]
- Annabi, M.; Houot, S.; Francou, C.; Poitrenaud, M.; Bissonnais, Y.L. Soil aggregate stability improvement with urban composts of different maturities. Soil Science Society of America Journal 2007, 71, 413–423. [Google Scholar] [CrossRef]
- Belachew, T.; Abera, Y. Effect of green manuring in combination with nitrogen on soil fertility and yield of bread wheat (Triticum aestivum) under double cropping system of Sinanadinsho, Southeast Ethiopia. Journal of Biodiversity and Environmental Sciences 2011, 1, 1–11. [Google Scholar]
- Baysal-Gurel, F.; Gardener, B.; Miller, S.A. Soil Borne Disease Management in Organic Vegetable Production. Organic Agri 2012. [Google Scholar]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Mokhtar, M.; El-Mougy, N. Bio-compost application for controlling soil-borne plant pathogens–a review. International Journal of Engineering and Innovative Technology 2014, 4, 2277–3754. [Google Scholar]
- Hao, J.; Subbarao, K.V.; Koike, S.T. Effects of broccoli rotation on lettuce drop caused by Sclerotinia minor and on the population density of sclerotia in soil. Plant Disease 2003, 87, 159–166. [Google Scholar] [CrossRef]
- Whalen, J. Crop profile for watermelon in Maryland. Online. Natl. Info. System for the Regional IPM Centers, North Carolina State Univ., Raleigh, NC 1999.
- Keinath, A.P.; Hassell, R.L.; Everts, K.L.; Zhou, X.-G. Cover crops of hybrid common vetch reduce Fusarium wilt of seedless watermelon in the eastern United States. Plant Health Progress 2010, 11, 8. [Google Scholar] [CrossRef]
- Papp, R.; Marinari, S.; Moscatelli, M.C.; van der Heijden, M.; Wittwer, R.; Campiglia, E.; Radicetti, E.; Mancinelli, R.; Fradgley, N.; Pearce, B. Short-term changes in soil biochemical properties as affected by subsidiary crop cultivation in four European pedo-climatic zones. Soil Till Res 2018, 180, 126–136. [Google Scholar] [CrossRef]
- Ntalli, N.; Caboni, P. A review of isothiocyanates biofumigation activity on plant parasitic nematodes. Phytochemistry Reviews 2017, 16, 827–834. [Google Scholar] [CrossRef]
- Waisen, P.; Cheng, Z.; Sipes, B.S.; Koon-Hui, W. Biofumigation effects of brassicaceous cover crops on soil health in cucurbit agroecosystems in Hawaii, USA. Pedosphere 2022, 32, 521–531. [Google Scholar] [CrossRef]
- Aydınlı, G.; Mennan, S. Biofumigation studies by using Raphanus sativus and Eruca sativa as a winter cycle crops to control root-knot nematodes. Brazilian Archives of Biology and Technology 2018, 61, e18180249. [Google Scholar] [CrossRef]
- Gabriel, J.; Quemada, M. Replacing bare fallow with cover crops in a maize cropping system: yield, N uptake and fertiliser fate. Eur J Agron 2011, 34, 133–143. [Google Scholar] [CrossRef]
- Mukherjee, R.; Sen, S. Agricultural sustainability through nitrogen fixation: approaches and techniques. Harvest 2021, 6, 48–55. [Google Scholar]
- Wang, Y.; Lambers, H. Root-released organic anions in response to low phosphorus availability: recent progress, challenges and future perspectives. Plant Soil 2020, 447, 135–156. [Google Scholar] [CrossRef]
- Meena, R.S.; Vijayakumar, V.; Yadav, G.S.; Mitran, T. Response and interaction of Bradyrhizobium japonicum and arbuscular mycorrhizal fungi in the soybean rhizosphere. Plant Growth Regul 2018, 84, 207–223. [Google Scholar] [CrossRef]
- Lai, H.; Gao, F.; Su, H.; Zheng, P.; Li, Y.; Yao, H. Nitrogen distribution and soil microbial community characteristics in a legume–cereal intercropping system: A review. Agronomy 2022, 12, 1900. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Lombardo, S.; Fontanazza, S.; Abbate, C.; Pandino, G.; Anastasi, U.; Onofri, A.; Mauromicale, G. Improving soil health, weed management and nitrogen dynamics by Trifolium subterraneum cover cropping. Agron Sustain Dev 2020, 40, 1–12. [Google Scholar] [CrossRef]
- Campiglia, E.; Mancinelli, R.; Radicetti, E.; Baresel, J.P. Evaluating spatial arrangement for durum wheat (Triticum durum Desf.) and subclover (Trifolium subterraneum L.) intercropping systems. Field Crops Res 2014, 169, 49–57. [Google Scholar] [CrossRef]
- Guardia, G.; Aguilera, E.; Vallejo, A.; Sanz-Cobena, A.; Alonso-Ayuso, M.; Quemada, M. Effective climate change mitigation through cover cropping and integrated fertilization: A global warming potential assessment from a 10-year field experiment. Journal of Cleaner Production 2019, 241, 118307. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Sauer, T.J.; Cruse, R.M. Chapter one—soil: the forgotten piece of the water, food, energy nexus. Advances in agronomy 2017, 143, 1–46. [Google Scholar]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron Sustain Dev 2016, 36, 1–14. [Google Scholar] [CrossRef]
- Fargione, J.E.; Bassett, S.; Boucher, T.; Bridgham, S.D.; Conant, R.T.; Cook-Patton, S.C.; Ellis, P.W.; Falcucci, A.; Fourqurean, J.W.; Gopalakrishna, T. Natural climate solutions for the United States. Science Advances 2018, 4, eaat1869. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.; de Faccio Carvalho, P.C.; Dedieu, B. Integrated crop–livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric Ecosyst Environ 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Bukovsky-Reyes, S.; Isaac, M.E.; Blesh, J. Effects of intercropping and soil properties on root functional traits of cover crops. Agric Ecosyst Environ 2019, 285, 106614. [Google Scholar] [CrossRef]
- Hinsinger, P.; Betencourt, E.; Bernard, L.; Brauman, A.; Plassard, C.; Shen, J.; Tang, X.; Zhang, F. P for two, sharing a scarce resource: soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol 2011, 156, 1078–1086. [Google Scholar] [CrossRef]
- Sun, F.; Pan, K.; Olatunji, O.A.; Li, Z.; Chen, W.; Zhang, A.; Song, D.; Sun, X.; Huang, D.; Tan, X. Specific legumes allay drought effects on soil microbial food web activities of the focal species in agroecosystem. Plant Soil 2019, 437, 455–471. [Google Scholar] [CrossRef]
- Cong, W.F.; Hoffland, E.; Li, L.; Six, J.; Sun, J.H.; Bao, X.G.; Zhang, F.S.; Van Der Werf, W. Intercropping enhances soil carbon and nitrogen. Global change biology 2015, 21, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, E.V.; Notaro, K.d.A.; de Barros, J.A.; Duda, G.P.; Moraes, M.d.C.H.d.S.; Ambrósio, M.M.d.Q.; Negreiros, A.M.P.; Sales Júnior, R. Soils from intercropped fields have a higher capacity to suppress black root rot in cassava, caused by Scytalidium lignicola. J Phytopathol 2019, 167, 209–217. [Google Scholar] [CrossRef]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [PubMed]
- de Araújo Santos, G.A.; Moitinho, M.R.; de Oliveira Silva, B.; Xavier, C.V.; Teixeira, D.D.B.; Corá, J.E.; Júnior, N.L.S. Effects of long-term no-tillage systems with different succession cropping strategies on the variation of soil CO2 emission. Science of the total environment 2019, 686, 413–424. [Google Scholar] [CrossRef]
- Graf, D.R.; Saghaï, A.; Zhao, M.; Carlsson, G.; Jones, C.M.; Hallin, S. Lucerne (Medicago sativa) alters N2O-reducing communities associated with cocksfoot (Dactylis glomerata) roots and promotes N2O production in intercropping in a greenhouse experiment. Soil Biol Biochem 2019, 137, 107547. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.; Jones, H.G.; Karley, A.J. Improving intercropping: a synthesis of research in agronomy, plant physiology and ecology. New Phytologist 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Chai, Q.; Nemecek, T.; Liang, C.; Zhao, C.; Yu, A.; Coulter, J.A.; Wang, Y.; Hu, F.; Wang, L.; Siddique, K.H. Integrated farming with intercropping increases food production while reducing environmental footprint. PNAS 2021, 118, e2106382118. [Google Scholar] [CrossRef]
- Xiao, J.; Yin, X.; Ren, J.; Zhang, M.; Tang, L.; Zheng, Y. Complementation drives higher growth rate and yield of wheat and saves nitrogen fertilizer in wheat and faba bean intercropping. Field Crops Res 2018, 221, 119–129. [Google Scholar] [CrossRef]
- Luo, S.; Yu, L.; Liu, Y.; Zhang, Y.; Yang, W.; Li, Z.; Wang, J. Effects of reduced nitrogen input on productivity and N2O emissions in a sugarcane/soybean intercropping system. Eur J Agron 2016, 81, 78–85. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.-Y.; Wu, H.-M.; Zhang, F.-F.; Li, C.-J.; Li, X.-X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N2 fixation. PNAS 2016, 113, 6496–6501. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Dang, K.; Lv, S.; Zhao, G.; Wang, H.; Feng, B. Interspecific competition and nitrogen application alter soil ecoenzymatic stoichiometry, microbial nutrient status, and improve grain yield in broomcorn millet/mung bean intercropping systems. Field Crops Res 2021, 270, 108227. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, H.; Zhang, W.; Bezemer, T.M.; Liang, W.; Li, Q.; Li, L. Interspecific interactions between crops influence soil functional groups and networks in a maize/soybean intercropping system. Agric Ecosyst Environ 2023, 355, 108595. [Google Scholar] [CrossRef]
- Teshita, A.; Feng, Y.; Qian, R.; Wang, X.; Khan, W.; Gao, Y. Alfalfa and maize intercropping enhances soil nematode structure and food web complexity in low-nitrogen soils. Appl Soil Ecol 2023, 186, 104809. [Google Scholar] [CrossRef]
- Ma, H.; Zhou, J.; Ge, J.; Nie, J.; Zhao, J.; Xue, Z.; Hu, Y.; Yang, Y.; Peixoto, L.; Zang, H. Intercropping improves soil ecosystem multifunctionality through enhanced available nutrients but depends on regional factors. Plant Soil 2022, 480, 71–84. [Google Scholar] [CrossRef]
- Benitez-Alfonso, Y.; Soanes, B.K.; Zimba, S.; Sinanaj, B.; German, L.; Sharma, V.; Bohra, A.; Kolesnikova, A.; Dunn, J.A.; Martin, A.C. Enhancing climate change resilience in agricultural crops. Current Biology 2023, 33, R1246–R1261. [Google Scholar] [CrossRef]
- Mugi-Ngenga, E.; Bastiaans, L.; Zingore, S.; Anten, N.; Giller, K. The role of nitrogen fixation and crop N dynamics on performance and legacy effects of maize-grain legumes intercrops on smallholder farms in Tanzania. Eur J Agron 2022, 141, 126617. [Google Scholar] [CrossRef]
- Du, Q.; Zhou, L.; Chen, P.; Liu, X.; Song, C.; Yang, F.; Wang, X.; Liu, W.; Sun, X.; Du, J. Relay-intercropping soybean with maize maintains soil fertility and increases nitrogen recovery efficiency by reducing nitrogen input. The Crop Journal 2020, 8, 140–152. [Google Scholar] [CrossRef]
- Li, C.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.; Li, H.; Zhang, F.; van der Werf, W. Syndromes of production in intercropping impact yield gains. Nature Plants 2020, 6, 653–660. [Google Scholar] [CrossRef]
- Wang, N.; Wang, T.; Chen, Y.; Wang, M.; Lu, Q.; Wang, K.; Dou, Z.; Chi, Z.; Qiu, W.; Dai, J. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. Nature Communications 2024, 15, 839. [Google Scholar] [CrossRef]
- Barbieri, P.; Pellerin, S.; Seufert, V.; Nesme, T. Changes in crop rotations would impact food production in an organically farmed world. Nature Sustainability 2019, 2, 378–385. [Google Scholar] [CrossRef]
- Tiemann, L.; Grandy, A.S.; Atkinson, E.; Marin-Spiotta, E.; McDaniel, M. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecology letters 2015, 18, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jousset, A.; de Boer, W.; Carrión, V.J.; Zhang, T.; Wang, X.; Kuramae, E.E. Legacy of land use history determines reprogramming of plant physiology by soil microbiome. The ISME journal 2019, 13, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Hilton, S.; Bennett, A.J.; Keane, G.; Bending, G.D.; Chandler, D.; Stobart, R.; Mills, P. Impact of shortened crop rotation of oilseed rape on soil and rhizosphere microbial diversity in relation to yield decline. PLOS one 2013, 8, e59859. [Google Scholar] [CrossRef]
- Yang, T.; Siddique, K.H.; Liu, K. Cropping systems in agriculture and their impact on soil health-A review. Global Ecology and Conservation 2020, 23, e01118. [Google Scholar] [CrossRef]
- Bennett, A.J.; Bending, G.D.; Chandler, D.; Hilton, S.; Mills, P. Meeting the demand for crop production: the challenge of yield decline in crops grown in short rotations. Biological reviews 2012, 87, 52–71. [Google Scholar] [CrossRef]
- Crookston, R.; Kurle, J.; Copeland, P.; Ford, J.; Lueschen, W. Rotational cropping sequence affects yield of corn and soybean. Agronomy Journal 1991, 83, 108–113. [Google Scholar] [CrossRef]
- Benitez, M.-S.; Osborne, S.L.; Lehman, R.M. Previous crop and rotation history effects on maize seedling health and associated rhizosphere microbiome. Sci Rep 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS microbiology ecology 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: the microbial ecology of the rhizosphere. Nature Reviews Microbiology 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Yang, C.; Hamel, C.; Gan, Y.; Vujanovic, V. Pyrosequencing reveals how pulses influence rhizobacterial communities with feedback on wheat growth in the semiarid Prairie. Plant Soil 2013, 367, 493–505. [Google Scholar] [CrossRef]
- Gurr, G.M.; Lu, Z.; Zheng, X.; Xu, H.; Zhu, P.; Chen, G.; Yao, X.; Cheng, J.; Zhu, Z.; Catindig, J.L. Multi-country evidence that crop diversification promotes ecological intensification of agriculture. Nature plants 2016, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Vilich, V. Crop rotation with pure stands and mixtures of barley and wheat to control stem and root rot diseases. Crop Prot. 1993, 12, 373–379. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Larney, F.J.; Blackshaw, R.E.; Pearson, D.C.; Eastman, A.H. Soil microbial biomass and its relationship with yields of irrigated wheat under long-term conservation management. Soil Sci. 2018, 183, 179–187. [Google Scholar] [CrossRef]
- Hamel, C.; Gan, Y.; Sokolski, S.; Bainard, L.D. High frequency cropping of pulses modifies soil nitrogen level and the rhizosphere bacterial microbiome in 4-year rotation systems of the semiarid prairie. Appl Soil Ecol 2018, 126, 47–56. [Google Scholar] [CrossRef]
- Bainard, L.D.; Navarro-Borrell, A.; Hamel, C.; Braun, K.; Hanson, K.; Gan, Y. Increasing the frequency of pulses in crop rotations reduces soil fungal diversity and increases the proportion of fungal pathotrophs in a semiarid agroecosystem. Agric Ecosyst Environ 2017, 240, 206–214. [Google Scholar] [CrossRef]
- Merz, U.; Falloon, R.E. powdery scab of potato—increased knowledge of pathogen biology and disease epidemiology for effective disease management. Potato Research 2009, 52, 17–37. [Google Scholar] [CrossRef]
- Nayyar, A.; Hamel, C.; Lafond, G.; Gossen, B.D.; Hanson, K.; Germida, J. Soil microbial quality associated with yield reduction in continuous-pea. Appl Soil Ecol 2009, 43, 115–121. [Google Scholar] [CrossRef]
- Qaswar, M.; Ahmed, W.; Jing, H.; Hongzhu, F.; Xiaojun, S.; Xianjun, J.; Kailou, L.; Yongmei, X.; Zhongqun, H.; Asghar, W. Soil carbon (C), nitrogen (N) and phosphorus (P) stoichiometry drives phosphorus lability in paddy soil under long-term fertilization: A fractionation and path analysis study. PLoS One 2019, 14, e0218195. [Google Scholar] [CrossRef]
- Urra, J.; Alkorta, I.; Garbisu, C. Potential benefits and risks for soil health derived from the use of organic amendments in agriculture. Agronomy 2019, 9, 542. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P. Soil quality–A critical review. Soil Biol Biochem 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Cardoso, E.J.B.N.; Vasconcellos, R.L.F.; Bini, D.; Miyauchi, M.Y.H.; Santos, C.A.d.; Alves, P.R.L.; Paula, A.M.d.; Nakatani, A.S.; Pereira, J.d.M.; Nogueira, M.A. Soil health: looking for suitable indicators. What should be considered to assess the effects of use and management on soil health? Scientia Agricola 2013, 70, 274–289. [Google Scholar]
- Marchiori Junior, M.; Melo, W. Carbon, microbial biomass carbon and enzyme activity of a soil under natural forest, grassland and cotton culture. Revista Brasileira de Ciencia do Solo (Brazil) 1999. [Google Scholar]
- Sicardi, M.; Garcı́a-Préchac, F.; Frioni, L. Soil microbial indicators sensitive to land use conversion from pastures to commercial Eucalyptus grandis (Hill ex Maiden) plantations in Uruguay. Appl Soil Ecol 2004, 27, 125–133. [Google Scholar] [CrossRef]
- Karaca, A.; Cetin, S.C.; Turgay, O.C.; Kizilkaya, R. Soil enzymes as indication of soil quality. In Soil enzymology, Springer: 2010; pp. 119-148.
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. Enzymes in the Environment 2002, 1–33. [Google Scholar]
- Bandick, A.K.; Dick, R.P. Field management effects on soil enzyme activities. Soil Biol Biochem 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Dick, W.; Tabatabai, M.A. Significance and potential uses of soil enzymes. Soil microbial ecology: applications in agricultural and environmental management. 1992, 95-127.
- Barrutia, O.; Garbisu, C.; Epelde, L.; Sampedro, M.; Goicolea, M.; Becerril, J. Plant tolerance to diesel minimizes its impact on soil microbial characteristics during rhizoremediation of diesel-contaminated soils. Science of the total environment 2011, 409, 4087–4093. [Google Scholar] [CrossRef]
- Galende, M.A.; Becerril, J.M.; Barrutia, O.; Artetxe, U.; Garbisu, C.; Hernández, A. Field assessment of the effectiveness of organic amendments for aided phytostabilization of a Pb–Zn contaminated mine soil. Journal of Geochemical Exploration 2014, 145, 181–189. [Google Scholar] [CrossRef]
- Pardo, T.; Clemente, R.; Epelde, L.; Garbisu, C.; Bernal, M. Evaluation of the phytostabilisation efficiency in a trace elements contaminated soil using soil health indicators. Journal of hazardous materials 2014, 268, 68–76. [Google Scholar] [CrossRef]
- Dubey, R.K.; Dubey, P.K.; Chaurasia, R.; Rao, C.S.; Abhilash, P.C. Impact of Integrated Agronomic Practices on Soil Fertility and Respiration on the Indo-Gangetic Plain of North India. Agronomy 2021, 11, 402. [Google Scholar] [CrossRef]
- Viaux, P.; Rieu, C. Integrated farming systems and sustainable agriculture in France. MONOGRAPHS-BRITISH CROP PROTECTION COUNCIL 1995, 297-297.
- Sarkar, D.; Rakshit, A.; Al-Turki, A.I.; Sayyed, R.; Datta, R. Connecting bio-priming approach with integrated nutrient management for improved nutrient use efficiency in crop species. Agriculture 2021, 11, 372. [Google Scholar] [CrossRef]
- Duarah, I.; Deka, M.; Saikia, N.; Boruah, H.D. Phosphate solubilizers enhance NPK fertilizer use efficiency in rice and legume cultivation. 3 Biotech 2011, 1, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Entesari, M.; Sharifzadeh, F.; Ahmadzadeh, M.; Farhangfar, M. Seed biopriming with Trichoderma species and Pseudomonas fluorescent on growth parameters, enzymes activity and nutritional status of soybean. International Journal of Agronomy and Plant Production 2013, 4, 610–619. [Google Scholar]
- Kim, P.J.; Chung, D.Y.; Malo, D. Characteristics of phosphorus accumulation in soils under organic and conventional farming in plastic film houses in Korea. Soil Sci Plant Nutr 2001, 47, 281–289. [Google Scholar] [CrossRef]
Figure 1.
The rhizosphere and the interactions between plant and soil microbes within this environment.
Figure 1.
The rhizosphere and the interactions between plant and soil microbes within this environment.
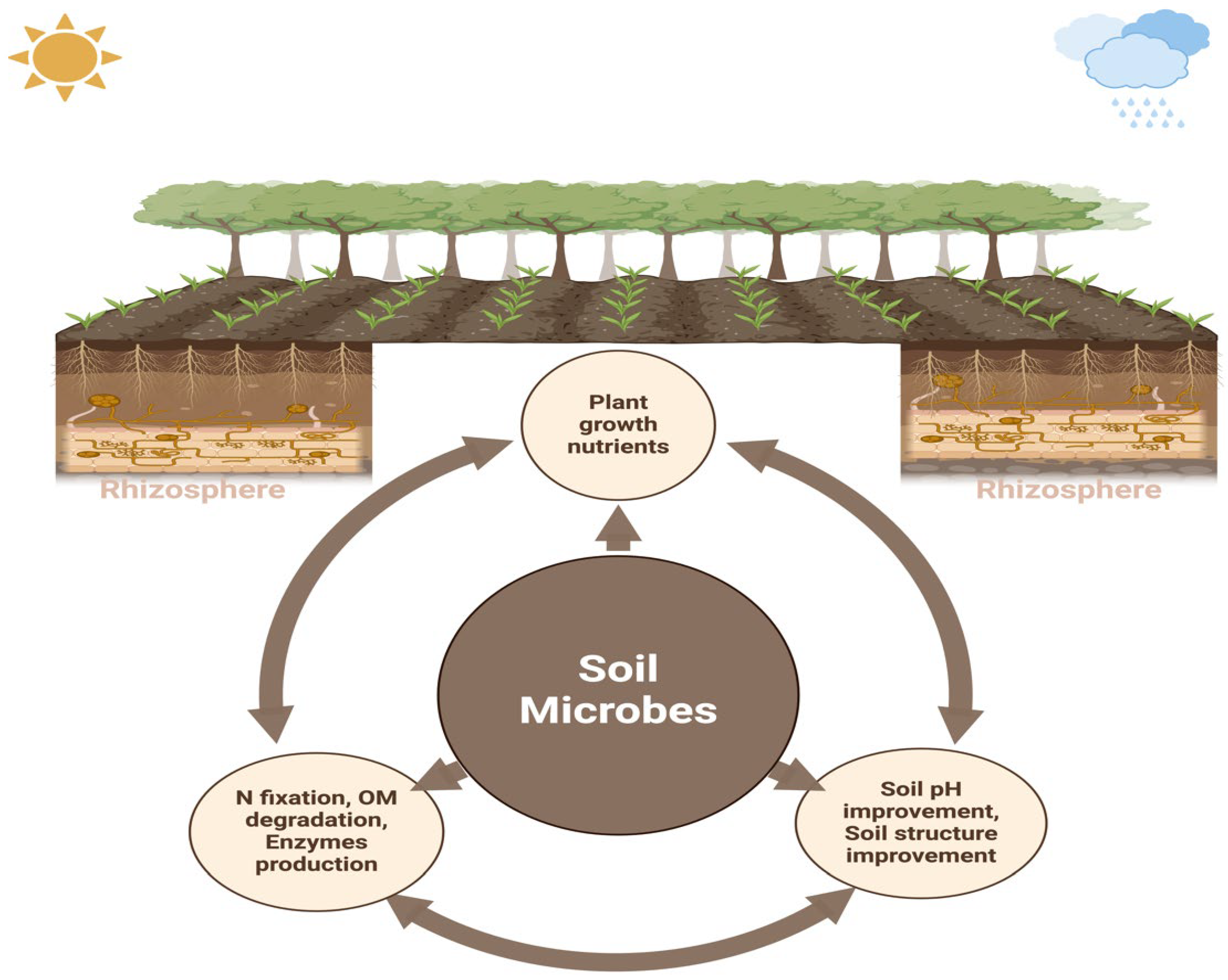
Figure 2.
Soils in agroecosystems serve as a primary nutrient reservoir for crops, supplying necessary nutrients and water while also functioning as the most significant microbial reproductive bank for the rhizosphere microbiome. The plant offers many microhabitats, including both the endosphere and rhizosphere, for the development of the rhizosphere and plant microbiome. Beneficial plant-microbe interactions facilitate the rhizosphere microbiome’s services. Microbiome services improve plant growth, stress tolerance, and disease resistance, as well as host resilience to abiotic challenges such as cold, salinity, and drought, resulting in biomass production increasing up to fourfold and enhancing rhizosphere processes through their direct and indirect mechanisms. We created the conceptual diagram to provide an overview of rhizosphere microbiome services.
Figure 2.
Soils in agroecosystems serve as a primary nutrient reservoir for crops, supplying necessary nutrients and water while also functioning as the most significant microbial reproductive bank for the rhizosphere microbiome. The plant offers many microhabitats, including both the endosphere and rhizosphere, for the development of the rhizosphere and plant microbiome. Beneficial plant-microbe interactions facilitate the rhizosphere microbiome’s services. Microbiome services improve plant growth, stress tolerance, and disease resistance, as well as host resilience to abiotic challenges such as cold, salinity, and drought, resulting in biomass production increasing up to fourfold and enhancing rhizosphere processes through their direct and indirect mechanisms. We created the conceptual diagram to provide an overview of rhizosphere microbiome services.
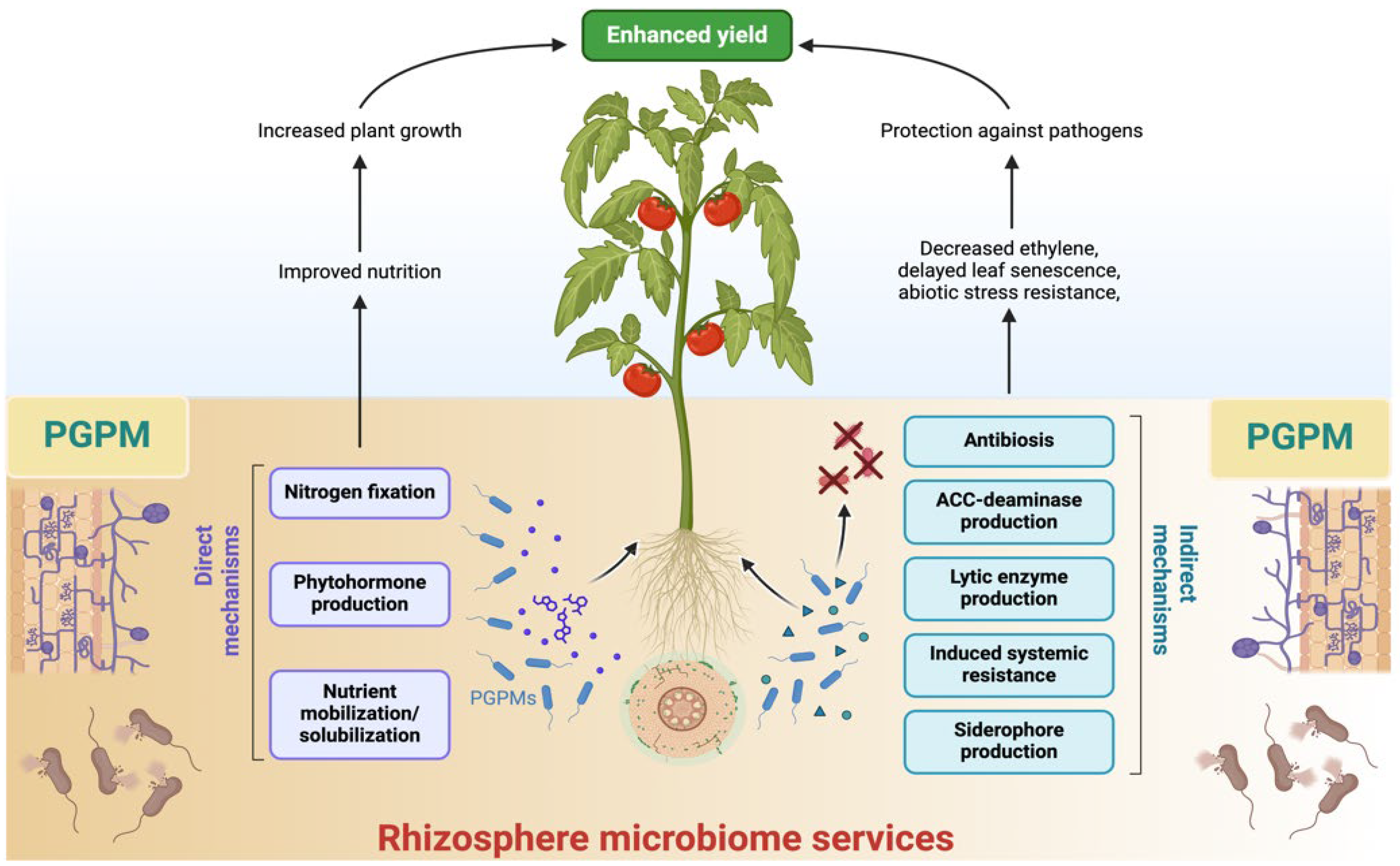
Figure 3.
Conceptual diagram that illustrates the functions of various types of green manures, including non-leguminous, leguminous, and Brassicaceous plants, within agroecosystems and their key ecological services.
Figure 3.
Conceptual diagram that illustrates the functions of various types of green manures, including non-leguminous, leguminous, and Brassicaceous plants, within agroecosystems and their key ecological services.
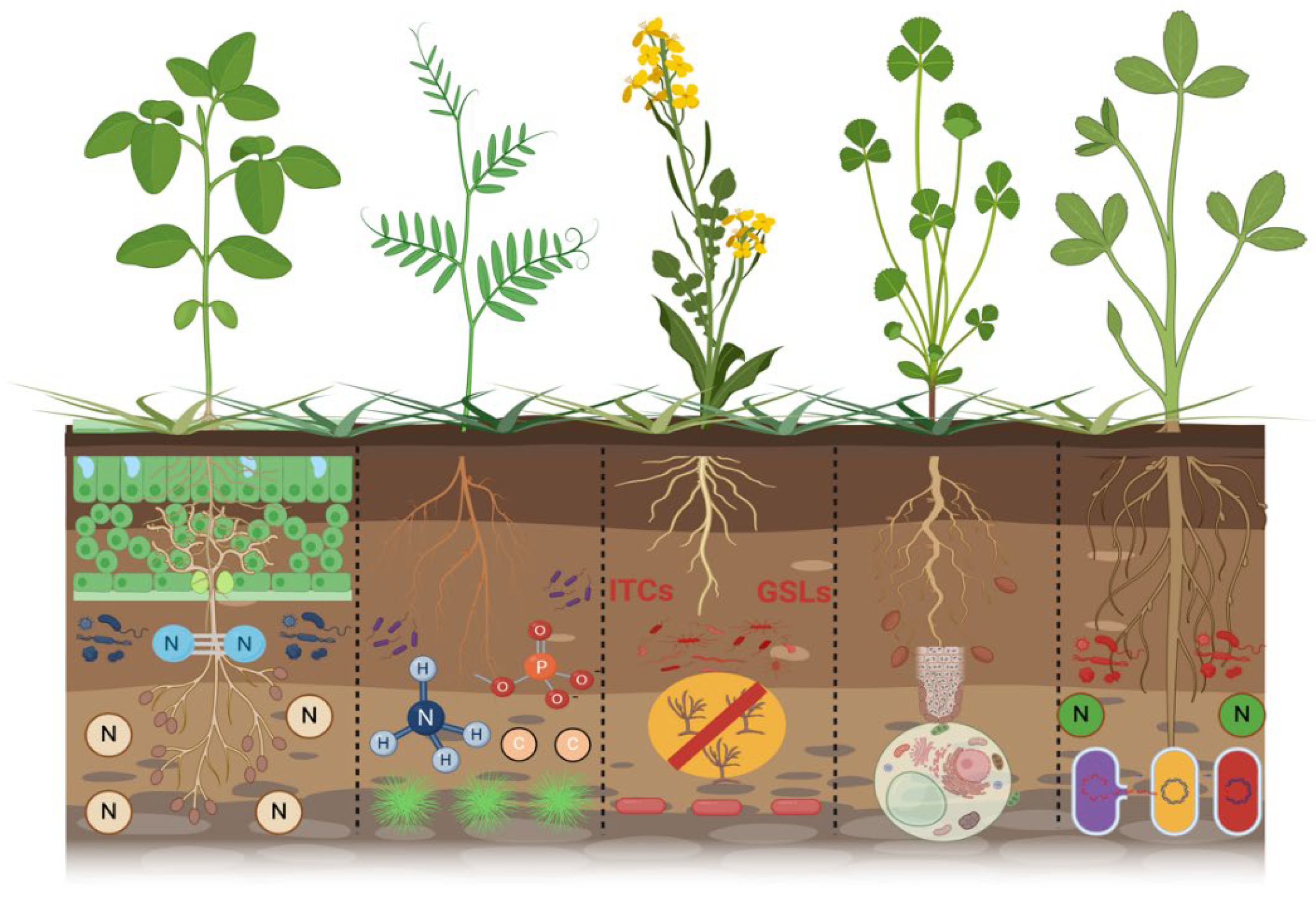
Figure 4.
An overview of intercropping systems and their beneficial impacts on cash crops and sustain the long-term agroecosystem.
Figure 4.
An overview of intercropping systems and their beneficial impacts on cash crops and sustain the long-term agroecosystem.
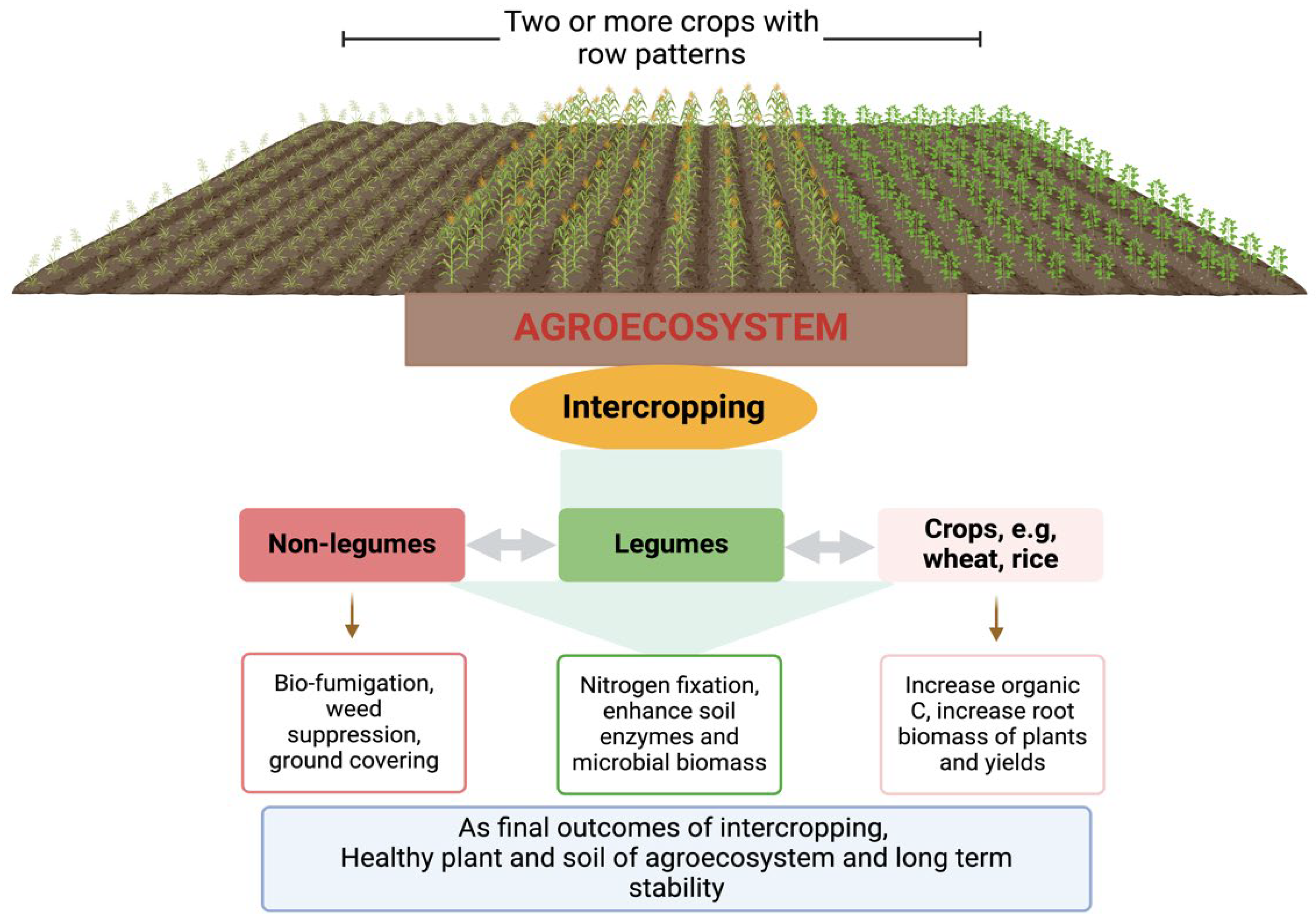
Figure 5.
We present a conceptual depiction of the integration of crop rotation systems, tracing their journey from challenges to outcomes. Crop rotation systems that incorporate cash and legume crops can sustain agricultural yields, increase farmers’ income, and minimize greenhouse gas emissions through legumes’ biological nitrogen fixation, which partially replaces synthetic nitrogen inputs. Incorporating legumes into crop rotations can improve soil health by promoting microbial and enzyme activity, optimizing nutrient cycling, and enhancing carbon sequestration.
Figure 5.
We present a conceptual depiction of the integration of crop rotation systems, tracing their journey from challenges to outcomes. Crop rotation systems that incorporate cash and legume crops can sustain agricultural yields, increase farmers’ income, and minimize greenhouse gas emissions through legumes’ biological nitrogen fixation, which partially replaces synthetic nitrogen inputs. Incorporating legumes into crop rotations can improve soil health by promoting microbial and enzyme activity, optimizing nutrient cycling, and enhancing carbon sequestration.
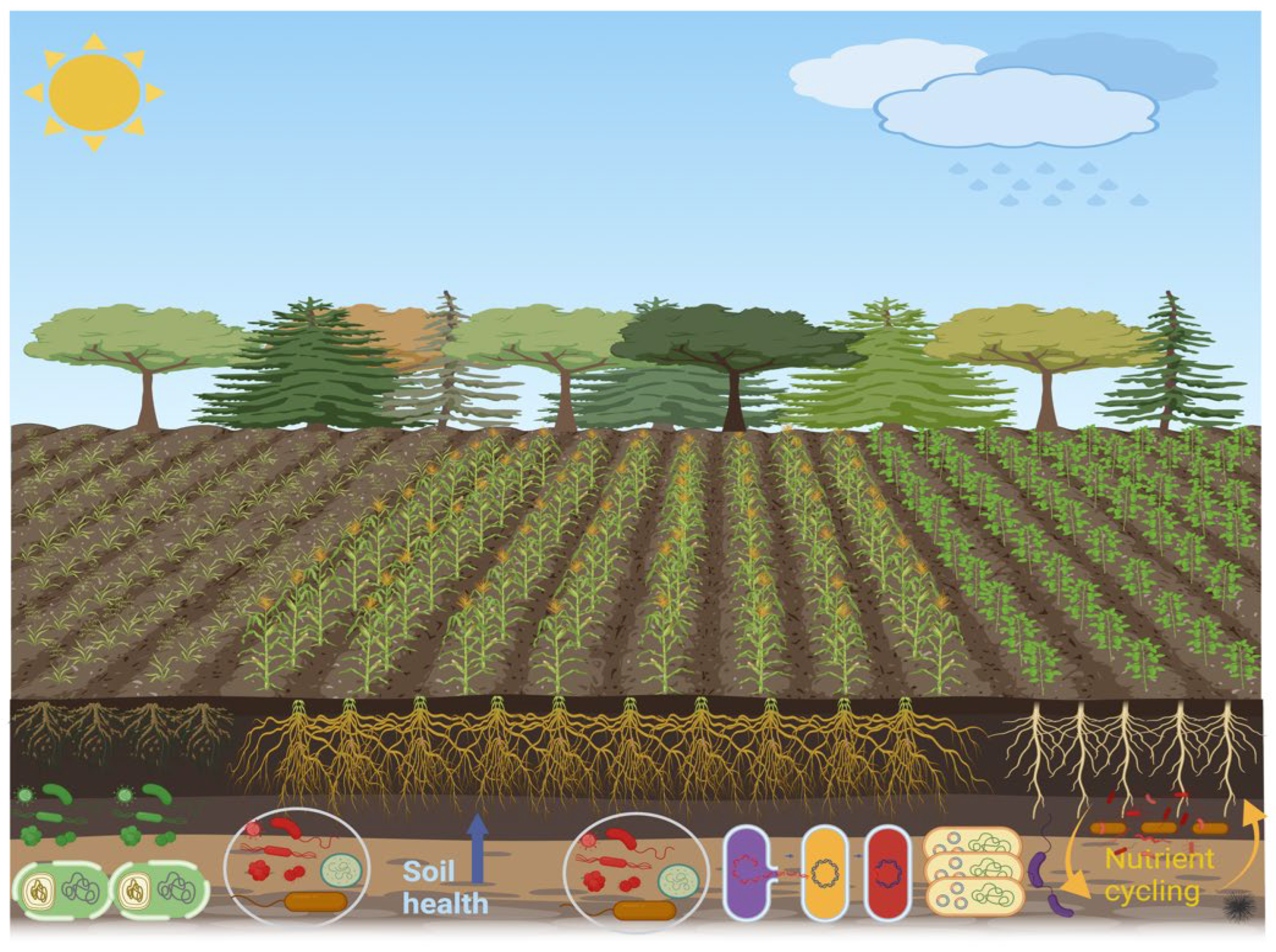

Table 1.
Potential role and activities of PGPM in the rhizosphere for sustainable agriculture.
| PGPM | Function and role | References |
|---|---|---|
| Penicillium spp. | Promotion of plant growth, increase systemic resistance, and confrontation to Pseudomonas syringae pv.Tomato | Hossain, et al. [70] Hossain, et al. [71] |
| Trichoderma spp. | Plant growth promotion, production of indole-3-acetic acid IAA, production of siderophores, resistance against soil-borne disease | Contreras-Cornejo, et al. [72] Abdel-lateif [73] Asghar and Kataoka [42] |
| Aspergillus spp. | Induces systemic resistance against necrotrophic fungus, plant growth promotion and phytohormones production | Salas-Marina, et al. [74] |
| Purpureocillium spp. | Plant growth promotion and siderophores and enzymes production | Cavello, et al. [75] |
| Talaromyces spp. | Promotion of plant growth, phosphate solubilization, siderophores production | Yamagiwa, et al. [76] |
| Bacillus spp. | Promotion of plant growth, Indole-3-acetic acid (IAA) and phosphate solubilization, siderophore production | Mahmood and Kataoka [62] |
| Azotobacter spp. | Ammonia production, Indole-3-acetic acid, growth enhancement of canola and lettuce plant, Phosphorus solubilization | Kumar, et al. [77] Joseph, et al. [78] |
| Pseudomonas spp. | Growth improvement of Canola and tomato plants, hormones production such as IAA, production of siderophore | Ahemad and Khan [79] Singh [80] |
| Azospirillum spp. | Indole-3-acetic acid (IAA) production, siderophore and exo-polysaccharides, and promote canola plant growth | Singh [80] Ahemad and Khan [79] |
| Streptomyces spp. | Streptomyces inhibit Curvularia oryzae and volatile organic compounds (VOCs) releasing, promote plant growth as well as activities of defense related enzymes | Sunpapao, et al. [81] |
Table 2.
Green manure and its beneficial effects.
| Green manure | Their beneficial effects | References |
|---|---|---|
| Oats, Hairy vetch | Nutrients availability, soil organic matter, enhancing of soil enzymes and fungal biomass and soil inorganic N and P, reducing soil erosion | Toom, Tamm, Talgre, Tamm, Tamm, Narits, Hiiesalu, Mäe and Lauringson [93] Kataoka, Nagasaka, Tanaka, Yamamura, Shinohara, Haramoto, Hayakawa and Sakamoto [43] Sharma, et al. [106] |
| Winter rye, Brassica | Biofumigation, reduction of soil-borne diseases, enhancement of soil enzymes and nitrogen fixation as well as improvement of soil biodiversity and structure | Sharma, Singh, Kahlon, Brar, Grover, Dia and Steiner [106] Brennan, et al. [107] Srivastava and Ghatak [108] Dubey, et al. [109] |
| Mustard, Barley | Soil and ground covering, nutrient cycling and increasing microbial biomass, as well as weed suppression, Increasing fungi and bacteria ration in soil | Khan, Gwon, Alam, Song, Das and Kim [17] Clark [110] |
| Red clover, Peas, Oat | Enhancement of biological activity in rhizosphere, nitrogen fixation and reduction of soil erosion, improve biological health, increase fungal abundance and soil enzymes activity | Clark [110] Thapa, et al. [111] |
| Cereal rye, Annual ryegrass | Enhancement of SOM, control weed growth, control N2O emissions, NO3- reduction, C sequestration and sustain cash crops yields | Adetunji, et al. [112] Magdoff and Van Es [113] Boselli, et al. [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



