
Submitted:
12 November 2024
Posted:
14 November 2024
You are already at the latest version
Abstract
Phosphonates derivatives are compounds of interests and are applied as drugs of e.g. antibacterial antiviral activities, what is connected with their inhibitory activity towards different enzymes, what in turn is in the relation to the configuration of particular compounds izomers. Biological synthesis of such molecules is the method of choice and can be carried on with enzymes or whole cells of organisms. Photobiocatalysts employed in bioconversion of epoxy dimethyl phosphonate are able to convert this substrate into geometric isomers of the unsaturated product, which is classified as very rare and expensive compound of high added value. Six different strains were screened towards dimethyl epoxy phosphonate and in case of Synechococcus bigranulatus 99% of conversion degree was achieved. Product structure and yield of the reaction were confirmed with Mass Spectroscopy (MS); Nuclear Magnetic Resonance (NMR) of isotopes 1H, 13C, and 31P, 2D NMR; and Infrared Spectroscopy (IR).
Keywords:
cyanobacteria
; photobiocatalysts
; biotransformation
; substrates
; phosphonates
; high value-added compounds
1. Introduction
Organophosphorus compounds, especially phosphonates, are significant in a wide array of industrial, agricultural, and pharmaceutical applications due to their unique chemical and biological activities. The stable carbon-phosphorus (C–P) bond in phosphonates imparts excellent resistance to both chemical hydrolysis and enzymatic degradation, making these compounds highly valuable. This stability is important for improving biologically active molecules, including enzyme inhibitors (by imitating the transition state of phosphorylated intermediates), antimicrobial agents, and antiviral drugs [1]. The synthesis and application of phosphonates in pharmaceuticals influence their ability to mimic natural phosphate esters, thereby inhibiting a wide range of enzymes and initiating pathways for treating several ailments, including infectious diseases, cancer, and inflammatory disorders [2,3,4,5].
Epoxymethyl dimethyl phosphonate (EMDP) represents a crucial intermediate in organophosphorus chemistry as its unique structure, characterized by an epoxide group and a phosphonate moiety, provides extensive reactivity and versatility in synthetic applications. The epoxide group is exceptionally remarkable for its nucleophilic ring-opening reactions, while the phosphonate group contributes stability and bioactivity to the resulting molecules [6]. EMDP can be exploited as a precursor in the synthesis of several phosphonates, including hydroxyalkyl and vinyl phosphonates, which are fundamental for developing pharmaceuticals and agricultural chemicals [7,8,9]. The availability of the epoxide group in phosphonates can enhance the formation of new stereocenters, a feature that is vital for drug discovery and development [10]. Conventional chemical synthesis of phosphonates from EMDP often requires harsh conditions, such as strong acids or bases, elevated temperatures, and environmentally hazardous solvents. These conditions can lead to undesired side reactions and low selectivity [11,12], prompting the current research for greener, more feasible approaches. Hence, biocatalysis, employing enzymes or whole-cell systems, has emerged as a promising alternative to conventional chemical synthesis. Biocatalysis renders the benefits of ecofriendly reaction conditions, higher selectivity, and fewer by-products, aligning with the principles of green chemistry [13,14,15].
The final obtained product, dimethyl (1E)-3-hydroxyprop-1-enylphosphonate (Figure 1), synthesized from EMDP via photobiocatalytic processes, is a phosphonate of significant interest due to its biological and pharmacological prospectives, regarding the existed chemicals, which have been already applied. This compound presents a hydroxyl group adjacent to an unsaturated carbon-phosphorus bond which is relatively stable and resistant to hydrolysis, distinguishing it from more common carbon-oxygen-phosphorus (C-O-P) bonds available in phosphates, giving it a versatile intermediate for further functionalization and drug development [16,17]. The structural properties of dimethyl (1E)-3-hydroxyprop-1-enylphosphonate allow its application as a scaffold for designing enzyme inhibitors and other biologically active molecules, particularly in antiviral and anticancer research [3,5,18]. The availability of the hydroxyl group converses the compound with potential reactivity in condensation reactions, allowing the formation of hydrogen bonds, influencing its solubility and interaction with biological targets such as the active sites of enzymes, potentially inhibiting key biological processes. For instance, it can act as a transition-state analog in the inhibition of phosphatases or other enzymes that process phosphate esters [19,20]. The presence of a double bond between the second and third carbon atoms signals a degree of unsaturation, making the molecule more reactive in certain types of chemical reactions, including additions or oxidations. The phosphonate group imparts several vital properties, including acidity (due to the presence of acidic protons on the phosphonic acid moiety), coordination ability with metal ions, and potential bioactivity due to its imitation of phosphate esters, which are common in biological systems. Despite the high value of dimethyl (1E)-3-hydroxyprop-1-enylphosphonate, its large-scale production utilizing standard chemical methods is quite challenging. Problems associated with stereoselectivity, and yield make it difficult to achieve high purity and efficiency [21,22,23]. This challenge has drawn attention to biocatalysis as an alternative production method.
Cyanobacteria are photosynthetic microbes that have received attention for their potential in biotechnological applications as biocatalysts due to their ability to convert light energy into biochemical processes. The strains investigated in this study - Synechococcus bigranulatus, Nostoc cf-muscorum, Limnospira indica, Limnospira maxima, Leptolyngbya foveolarum, and Nodularia moravica - represent a diverse selection of cyanobacteria, each with unique metabolic characteristics influencing their usefulness as photobiocatalysts. Synechococcus species are well-known for their fast growth rates, effective photosynthetic systems, and metabolic flexibility. Synechococcus bigranulatus demonstrated a high potential for photobiocatalysis in this study [24]. This strain's photocatalytic potential can be ascribed to its efficient light-harvesting competences and robust metabolic pathways, which optimize energy transfer for enzymatic reactions and reinforce the transformation of several substrates [25,26]. In addition, genetic engineering research has improved the photosynthetic efficiency and biocatalytic capacities of Synechococcus strains, making them attractive microorganisms for industrial applications [27,28]. Nostoc cf-muscorum is a filamentous cyanobacterium known for its nitrogen-fixing abilities and flexibility under low light, nutrient availability and robust environmental adaptability [29]. The lower biocatalytic activity observed in this study may result from its slower growth rates and reduced photosynthetic efficiency under laboratory conditions [30,31]. Nostoc species often distribute significant metabolic resources to nitrogen fixation, which may constrain their capacity for other biocatalytic processes [29]. Thus, Nostoc strains might be appropriately suited for applications requiring stress tolerance and nitrogen fixation rather than high-rate chemical conversions. Limnospira species, earlier classified under the Spirulina genus, are identified for their high biomass productivity and adaptability. Both Limnospira indica and Limnospira maxima achieved significant conversion rates as Synechococcus bigranulatus in this study. Their high efficiency in photobiocatalysis is probably due to their effective photosynthetic machinery and ability to grow well under diverse controlled intensities of light [32,33,34,35]. Limnospira species are generally employed in industrial applications for biofuel production, nutraceuticals, and high-value chemicals due to their photosynthetic efficiency and scalability [36]. Leptolyngbya foveolarum is a filamentous cyanobacterium identified for its resistance in extreme environments, such as thermal springs and hypersaline conditions [37]. Although less studied in the context of biocatalysis, Leptolyngbya species have demonstrated promise in environmental applications, including the bioremediation of heavy metals and organic pollutants [38,39]. In this study, Leptolyngbya foveolarum demonstrated efficient conversion competence for EMDP, implying that its metabolic pathways assist organophosphorus transformations, although at a lower rate compared to Synechococcus and Limnospira strains. Future research could focus on optimizing reaction conditions and potentially engineering Leptolyngbya strains to boost their photobiocatalytic performance [40,41]. Nodularia species are primarily acknowledged for their nitrogen-fixing capacities and symbiotic relationships with plants in nutrient-deficient environments [42,43]. Nodularia sp., a filamentous strain, has been investigated for ecological applications, such as biofertilization and soil conditioning [44,45]. In the context of biocatalysis, Nodularia sp. exhibited reasonable conversion rates for EMDP, while not as efficient as Synechococcus strain or Limnospira strains in photobiocatalytic processes, Nodularia's unique metabolic attributes, including nitrogen fixation and tolerance to low-nutrient environments, could make it valuable for applications requiring multifunctional biocatalysis [46].
Comparative analysis of the cyanobacterial strains in this study features significant variations in their competences for the photobiocatalytic conversion of EMDP to dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. Synechococcus bigranulatus and Limnospira species exhibited great conversion rates and limited side reactions, attributed to their efficient photosynthetic systems and metabolic flexibility [47,48,49,50,51,52]. In contrast, Nostoc cf-muscorum, Leptolyngbya foveolarum, and Nodularia sp. exhibited moderate conversion efficiencies, possibly due to their metabolic focus on nitrogen fixation or resilience in extreme environments rather than high-rate biocatalysis of phosphonates [24,53,54,55]. In addition, previous research emphasizes the capabilities of Leptolyngbya foveolarum, Nostoc cf-muscorum and Nodularia sp. as promising biocatalysts particularly in the enantioselective synthesis of chiral phosphonates [24]. Nodularia species in previous investigation demonstrated a significant degree of conversion and optical purity in producing diethyl (S)-2-hydroxy-2-phenylethylphosphonate, indicating its potential in biocatalysis despite metabolic adaptations toward resilience in extreme conditions like nitrogen fixation [24]. Recent developments in synthetic biology and metabolic engineering can offer promising prospects for advancing the performance of cyanobacteria in biocatalytic applications, enabling the feasible production of high-value products like dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. For instance, research have revealed that engineered Synechococcus elongatus can efficiently produce C3 platform chemicals like glycerol, dihydroxyacetone, and 3-hydroxypropionic acid, which serve as precursors for biofuels and biodegradable materials [56]. This integration has the potential to minimize carbon footprints while generating valuable chemicals through photosynthesis [56,57,58]. In another development, integration of Green Chemistry Technologies (such as cyanobacterial systems) with other sustainable technologies, such as enzyme immobilization or continuous flow reactors will further advance process efficiency and scalability of high-value organophosphorus compounds in a sustainable means [58,59,60,61].
This study aims to harness photobiocatalytic processes to facilitate the complete conversion of epoxymethyl dimethyl phosphonate into dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. This is a rare compound with remarkable value due to its unique chemical structure and versatile potential in advanced pharmaceutical, agricultural, and industrial applications. By exploiting this innovative technique, future research could significantly advance the green synthesis of valuable phosphonates.
2. Materials and Methods
Target organic substrate employed in the photobiocatalysis technique was epoxymethyl dimethyl phosphonate, sourced from SPECIFIC POLYMERS, France. Growth media components were purchased from Sigma Aldrich, Chempur, Avantor Performance Materials. The biomass of cyanobacterial strains and their growth media used include: Synechococcus bigranulatus - CCALA 187 (BG-11 medium); Nostoc cf-muscorum - CCALA 129 (BG-10 medium); Limnospira indica - PCC 8005 (Spirulina medium); Limnospira maxima - CCALA 27 (Spirulina medium); Leptolyngbya foveolarum - CCALA 76 (BG-11 medium); Nodularia moravica - CCALA 114 (BG-11 medium). These strains were obtained from the Culture Collection of Autotrophic Organisms (CCALA) at the Institute of Botany, Czech Academy of Sciences. The analytical instruments used include Mass Spectrometry (MS), Infrared Spectroscopy (IR), High-Performance Liquid Chromatography (HPLC) and Nuclear Magnetic Resonance (NMR). NMR spectra were measured on a Bruker Avance™ 600 at 600.58 MHz for 1H and 243.12 MHz for 31P or on a Jeol ECZ 400 S at 161.92 MHz for 31P. Chemical shifts (δ) were reported in ppm and coupling constants (J) were given in Hz. MS spectra were obtained using high resolution mass spectrometer with analyzer time-of-flight (TOFMS) from LCT PremierTM XE.
Growth media were prepared according to standard laboratory protocols [62]. Each strain was inoculated into separate 250 mL Erlenmeyer flasks containing 100 mL of the appropriate growth medium. The growth of the strains was monitored under a controlled photobioreactor (growth chamber) conditions: continuous illumination at 15–22 μmol photons/m²/s (Power Glo fluorescent bulb, 30W, Hagen) at 29°C ± 1°C under sterile conditions for 21 days. The photobiocatalytic reaction was initiated by adding 15 µL of epoxymethyl dimethyl phosphonate to each 100 mL cyanobacterial biomass solution, followed by gentle mixing to achieve a uniform substrate concentration of 1 mM. The cultures were then maintained under consistent photobioreactor conditions for 1 to 7 days to optimize conversion rates. Each experiment was conducted in triplicate. After biotransformation, the reaction mixture from each strain was centrifuged at 4000 rpm for 10 minutes, and the supernatant was collected. The supernatant was subjected to extraction twice using 50 mL of ethyl acetate. The organic layer was dried over anhydrous MgSO4 for 24 hours, and then filtered. The ethyl acetate was evaporated under reduced pressure using a rotary evaporator at 60°C. The resulting oil residue (product) was purified using High Pressure Preparative Liquid Chromatography System (Teledyne ISCO ACCQPrep HP125) with reversed phase column RediSep Prep C18, 5 μm, 250 mm. General procedure of purification using HPLC: 10 min of isocratic flow of 10% of acetonitrile in water, 5 min of 15% of acetonitrile in water, 5 min from 15 to 30% of acetonitrile in water, 10 min of isocratic flow of 30% of acetonitrile in water; flow 5 mL/min, Rf1 = 23 min. The obtained products were analyzed by Mass Spectroscopy (MS), Infrared Spectroscopy (IR), Nuclear Magnetic Resonance Spectroscopy (NMR) – MS (TOF MS ES+) Calcd for C5H11O4P [M + H]+ 167.0473; found: 167.0481; IR: 3362 (OH), 1638 (CH=CH) cm-1; 31P NMR (CDCl3, δ, ppm): 22.64; 1H NMR (CDCl3, δ, ppm): 3.69 (d, J = 11 Hz, 6H, 2*P(O)OCH3), 4.28 (m, 2H, CH2OH), 5.96 (ddt, J= 2, 17, 21 Hz, 1H, P-CH=CH), 6.86 (ddt, J = 3, 17, 23 Hz, 1H, P-CH=CH); 13C NMR (CDCl3, δ, ppm): 52.58 (d, J = 5.7 Hz, 1C, POCH3), 62.36 (d, J = 21.8 Hz, 1C, HOCH2=CH), 113.22 (d, J = 190.7 Hz, 1C, P-CH=CH), 153.69 (d, J = 5.2 Hz, 1C, P-CH=CH).
The assignment of signals on 1H and 13C NMR has been confirmed by 1-1H COSY, 1H-13C HMQC and 1H-13C HMBC spectra. The product was also synthesized by chemical method [63]: Sodium methoxide in methanol (0.1 equiv) was added dropwise to epoxymethyl dimethyl phosphonate (2.57 mmol; 0,5 g) at 0 °C under stirring. After the addition, the reaction was stirred 1 h at room temperature. Then 2 g of Dowex 50W-X8 (H+) resin (200-400 mesh), previously washed with methanol, was added and the mixture was stirred for 15 min. Afterwards the resin was filtered off, washed with methanol, and the combined filtrate was evaporated to dryness to give the crude product (0.49 g). After synthesis, products were purified using Flash Chromatography System: Combi Flash® Rf 150 and reversed phase column PuriFlash C18-HP, 15 μm, 25 g. General procedure of purification included isocratic flow of pure water, flow 10 mL/min, Rf = 2.5 min. Product was analyzed by NMR, the assignment of signals on 1H and 13C NMR has been confirmed by 1-1H COSY, 1H-13C HMQC and 1H-13C HMBC spectra. The spectroscopic characterization was in agreement with the data reported earlier in the literature [63].
3. Results
Bioconversion results were thoroughly analyzed using Mass Spectroscopy (MS), Nuclear Magnetic Resonance (NMR) techniques for 1H, 13C, and 31P isotopes, two-dimensional NMR (2D NMR), and Infrared Spectroscopy (IR). Figure 3 highlights key findings from the 31P NMR and IR analyses.
3.1. Comparative Bioconversion Rates of Cyanobacterial Strains
Figure 2 shows the comparative bioconversion efficiency of six cyanobacterial strains - Synechococcus bigranulatus, Nostoc cf-muscorum, Limnospira indica, Limnospira maxima, Leptolyngbya foveolarum, and Nodularia moravica - in the conversion of epoxymethyl dimethyl phosphonate (EMDP) into dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. The figure reveals a time-dependent increase in bioconversion across all strains, with conversion degrees evaluated at intervals up to 7 days. The data indicate that Limnospira strains (Limnospira indica and Limnospira maxima) and Synechococcus bigranulatus all achieved the highest conversion efficiency, 99% after 3 days and maintaining this efficiency through subsequent days. However, Limnospira strains demonstrated rapid bioconversion rates, achieving 99% by after 24 hours and sustaining these levels. The high efficiency of these strains suggests a robust capability for photobiocatalysis, likely facilitated by their optimized photosynthetic pathways and compatibility with the experimental light conditions used.
In contrast, Leptolyngbya foveolarum and Nodularia moravica exhibited slower conversion rates, achieving 96% and 76% bioconversion, respectively, by day 7. Nostoc cf-muscorum showed the least conversion, with only 12.4% bioconversion by day 7, which may be attributed to inherent metabolic or structural limitations within this strain that affect substrate uptake or enzyme activity relevant to the transformation of EMDP. This variance among strains underscores the importance of strain-specific metabolic features in determining bioconversion efficiency.
3.2. Spectral Characterization of Biotransformation
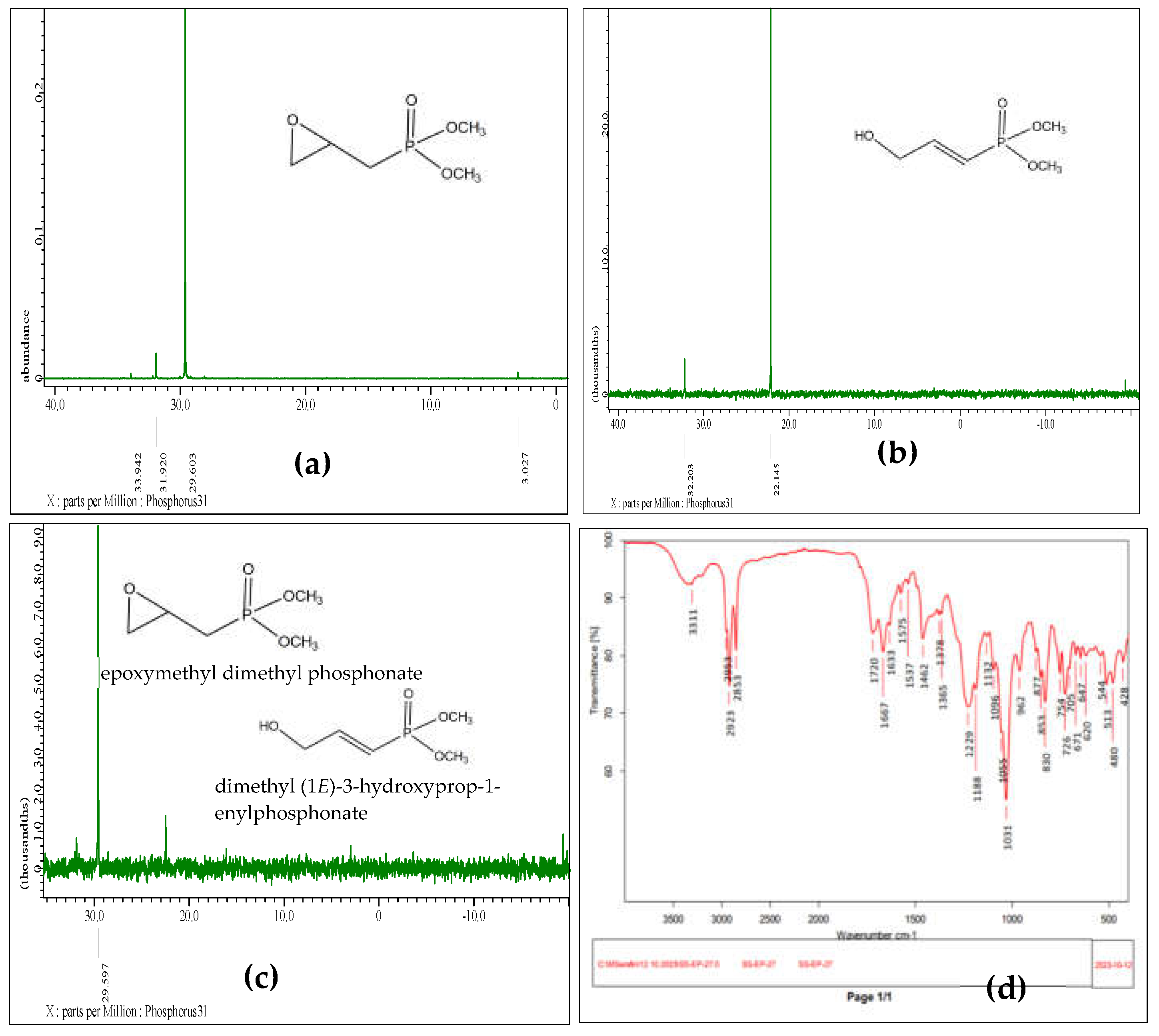
Thorough spectral characterization of the bioconversion process, using both 31P NMR and IR spectroscopy are presented in Figure 3. The spectral data provides insight into the structural changes occurring as EMDP is transformed into dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. Figure 3a presents the 31P NMR spectrum of the initial substrate, EMDP, with typical peaks that align with the characteristic phosphonate spectral signatures. Figure 3b displays the 31P NMR spectrum following complete bioconversion to the target product. Here, notable shifts in the 31P signal indicate successful transformation, corresponding to structural alterations involving the formation of an unsaturated C=C bond and the introduction of a hydroxyl group. Figure 3c shows the spectrum of a partially converted sample, where residual substrate peaks are visible alongside product signals, providing an intermediate perspective on the bioconversion process. This spectrum illustrates the sequential conversion stages and is useful in understanding the kinetics of the reaction. Figure 3d presents the IR spectrum of the fully converted product, indicating key vibrational modes that confirm the presence of both a hydroxyl group and an unsaturated C=C bond in the final product structure. The presence of these functional groups aligns with the proposed structure of dimethyl (1E)-3-hydroxyprop-1-enylphosphonate and corroborates findings from the NMR spectra [64,65,66]. Therefore, the data presented in Figure 3 confirms the structural integrity and reliability of the bioconversion process, with the spectral signatures validating the transition from EMDP to the targeted high value-added phosphonate product. These results provide a fundamental understanding of how cyanobacterial systems can be harnessed for efficient biocatalytic transformation of organophosphorus compounds, with each spectral shift marking a critical step in the pathway.
4. Discussion
The research’s findings highlight the effectiveness of cyanobacterial strains as biocatalysts in transforming epoxymethyl dimethyl phosphonate into dimethyl (1E)-3-hydroxyprop-1-enylphosphonate (Figure 3a–d). Among the six strains tested, Synechococcus bigranulatus and Limnospira strains achieved an exceptional conversion rate, (Figure 2). This exceptional performance emphasizes the metabolic adaptability and photosynthetic competence of these strains, predominantly their capability to harvest light energy and optimize energy transfer for biotransformation processes. The performance of Synechococcus aligns with previous research which demonstrated that Synechococcus photoautotrophic abilities, coupled with genetic engineering, improve the synthesis of high value-added biochemicals under controlled light conditions [67]. For example, engineered Synechococcus elongatus strains have been advanced for effective generation of high-value compounds like succinic acid, highlighting the potential of Synechococcus in scalable chemical production [67,68]. Limnospira strains, specifically Limnospira indica and Limnospira maxima have been recorded for their biofuels and nutraceutical production, and their strong photosynthetic machinery and adaptability to diverse light intensities make them highly suitable for biocatalytic applications. In addition, the high biomass productivity of Limnospira sp make them cost-effective options for industrial-scale production and their applicability in photobiocatalysis [24].
Other strains such as Leptolyngbya foveolarum and Nodularia moravica exhibited moderate conversion efficiencies while Nostoc cf-muscorum exhibited the least conversion efficiency. Although previous investigation demonstrated that these strains show potential in enantioselective synthesis of chiral phosphonates [24,60,69], however their performance in this study was limited, possibly due to their slower growth rates and metabolic path toward nitrogen fixation. Nostoc species, for example, direct significant metabolic resources towards nitrogen fixation, that may constrain high-rate biocatalysis for phosphonates [24]. Previous studies have also highlighted Nostoc’s applications in nutrient-deficient environments where nitrogen fixation is prioritized over other metabolic functions [70,71]. Hence, while these strains may be less efficient for EMDP conversion, they could serve specialized roles in environments that require multifunctional biocatalysis or bioremediation [72,73,74]. Cyanobacterial species, like Nostoc and Nodularia, can synthesize chiral hydroxyphosphonates via enantioselective bioreduction, suggesting further investigation of these strains for diverse chiral synthesis applications [48].
Prospective research efforts could focus on advancing genetic engineering techniques to improve biocatalytic efficacy across various cyanobacterial strains. For instance, offering metabolic pathways for advanced phosphonate synthesis in strains such as Leptolyngbya and Nodularia could increase their applicability. In addressing challenges faced by conventional chemical synthesis of phosphonates in terms of selectivity and environmental impact, incorporating these cyanobacterial systems with green chemistry practices, like enzyme immobilization or continuous flow reactors, could significantly boost process scalability and efficiency in eco-friendly production of high value-added phosphonates [68,75,76,77].
5. Conclusions
This research presents a pioneering exploration of cyanobacteria as biocatalysts for the eco-efficient transformation of epoxymethyl dimethyl phosphonate into the valuable product dimethyl (1E)-3-hydroxyprop-1-enylphosphonate. Notably, Synechococcus bigranulatus and Limnospira strains emerged as highly effective agents, demonstrating the potential of cyanobacteria to revolutionize biocatalysis through natural photosynthetic mechanisms. These findings highlight a feasible approach to chemical production, with significant implications for the pharmaceutical and agricultural industries. The scalability and cost-effectiveness of cyanobacteria underscore their promise for industrial applications, offering an environmentally friendly alternative to traditional synthetic methods.
Future research should prioritize advanced genetic and metabolic engineering strategies to further enhance biocatalytic efficiency and expand the utility of cyanobacteria in complex synthetic pathways, including chiral phosphonate production. This innovative pathway could establish a greener, more resourceful framework for the synthesis of valuable compounds, positioning cyanobacteria as integral to the future of feasible biochemical products.
Author Contributions
E.Z.D., S.O.S., and M.S.L. conceptualized the idea and methodology; E.Z.D. and M.S.L. supervised the article; S.O.S., E.Z.D., M.S.L., M.B.R., and M.K.O. wrote the original drafted manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McPherson, J.C.; Runner, R.; Buxton, T.B.; Hartmann, J.F.; Farcasiu, D.; Bereczki, I.; Rőth, E.; Tollas, S.; Ostorházi, E.; Rozgonyi, F.; et al. Synthesis of osteotropic hydroxybisphosphonate derivatives of fluoroquinolone antibacterials. Eur. J. Med. Chem. 2012, 47, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Mikołajczyk M (2019) Phosphonate reagents and building blocks in the synthesis of bioactive compounds, natural products and medicines. Pure Appl Chem 91:811–838. [CrossRef]
- Krečmerová M, Majer P, Rais R, Slusher BS (2022) Phosphonates and Phosphonate Prodrugs in Medicinal Chemistry: Past Successes and Future Prospects. Front Chem 10:889737. [CrossRef]
- Sevrain, C.M.; Berchel, M.; Couthon, H.; Jaffrès, P.-A. Phosphonic acid: preparation and applications. Beilstein J. Org. Chem. 2017, 13, 2186–2213. [Google Scholar] [CrossRef] [PubMed]
- Kaboudin B, Daliri P, Faghih S, Esfandiari H (2022) Hydroxy- and Amino-Phosphonates and -Bisphosphonates: Synthetic Methods and Their Biological Applications. Front Chem 10:890696. [CrossRef]
- Alkenoic acids and their derivatives. In: Tetrahedron Organic Chemistry Series. pp 199–282.
- Turhanen PA, Demadis KD, Kafarski P (2021) Editorial: Phosphonate Chemistry in Drug Design and Development. Front Chem 9:695128. [CrossRef]
- Manghi, M.C.; Masiol, M.; Calzavara, R.; Graziano, P.L.; Peruzzi, E.; Pavoni, B. The use of phosphonates in agriculture. Chemical, biological properties and legislative issues. 2021, 283, 131187. [Google Scholar] [CrossRef]
- Xuan, C.; Zhu, Z.; Li, Z.; Shu, C. Recent developments in the synthesis of pharmacological alkyl phosphonates. Adv. Agrochem 2024. [Google Scholar] [CrossRef]
- Kaur, B.; Singh, P. Epoxides: Developability as active pharmaceutical ingredients and biochemical probes. Bioorganic Chem. 2022, 125, 105862. [Google Scholar] [CrossRef]
- Montchamp JL (2019) Challenges and solutions in phosphinate chemistry. Pure Appl Chem 91:113–120. [CrossRef]
- Cybulska P, Legrand YM, Babst-Kostecka A, Diliberto S, Leśniewicz A, Oliviero E, Bert V, Boulanger C, Grison C, Olszewski TK (2022) Green and Effective Preparation of α-Hydroxyphosphonates by Ecocatalysis. Molecules 27:3075. [CrossRef]
- Serafin-Lewańczuk M, Brzezińska-Rodak M, Lubiak-Kozłowska K, Majewska P, Klimek-Ochab M, Olszewski TK, Żymańczyk-Duda E (2021) Phosphonates enantiomers receiving with fungal enzymatic systems. Microb. Cell Fact. 20.
- Kolodiazhna, A.O.; Kolodiazhnyi, O.I. Catalytic Asymmetric Synthesis of C-Chiral Phosphonates. Symmetry 2022, 14, 1758. [Google Scholar] [CrossRef]
- Sobkowski M, Kraszewski A, Stawinski J (2014) Recent Advances in H-Phosphonate Chemistry. Part 2. Synthesis of C-Phosphonate Derivatives. Top Curr Chem 361:179–216. [CrossRef]
- Schrewe, M.; Julsing, M.K.; Bühler, B.; Schmid, A. Whole-cell biocatalysis for selective and productive C–O functional group introduction and modification. Chem. Soc. Rev. 2013, 42, 6346–6377. [Google Scholar] [CrossRef]
- Ran, N.; Zhao, L.; Chen, Z.; Tao, J. Recent applications of biocatalysis in developing green chemistry for chemical synthesis at the industrial scale. Green Chem. 2007, 10, 361–372. [Google Scholar] [CrossRef]
- Groaz E, De Jonghe S (2021) Overview of Biologically Active Nucleoside Phosphonates. Front Chem 8:616863. [CrossRef]
- Hilderbrand, R.L. Henderson TO (2018) Phosphonic acids in nature. Role Phosphonates Living Syst 5–30. [CrossRef]
- Hanson, J.E.; Kaplan, A.P.; Bartlett, P.A. Phosphonate analogs of carboxypeptidase A substrates are potent transition-state analog inhibitors. Biochemistry 1989, 28, 6294–6305. [Google Scholar] [CrossRef]
- Ruffolo, F.; Dinhof, T.; Murray, L.; Zangelmi, E.; Chin, J.P.; Pallitsch, K.; Peracchi, A. The Microbial Degradation of Natural and Anthropogenic Phosphonates. Molecules 2023, 28, 6863. [Google Scholar] [CrossRef]
- Harsági, N.; Keglevich, G. The Hydrolysis of Phosphinates and Phosphonates: A Review. Molecules 2021, 26, 2840. [Google Scholar] [CrossRef] [PubMed]
- Horsman, G.P.; Zechel, D.L. Phosphonate Biochemistry. Chem. Rev. 2016, 117, 5704–5783. [Google Scholar] [CrossRef] [PubMed]
- Górak, M.; Żymańczyk-Duda, E. Application of cyanobacteria for chiral phosphonate synthesis. Green Chem. 2015, 17, 4570–4578. [Google Scholar] [CrossRef]
- Tüllinghoff, A.; Djaya-Mbissam, H.; Toepel, J.; Bühler, B. Light-driven redox biocatalysis on gram-scale in Synechocystis sp. PCC 6803 via an in vivo cascade. Plant Biotechnol. J. 2023, 21, 2074–2083. [Google Scholar] [CrossRef]
- Włodarczyk, A.; Selão, T.T.; Norling, B.; Nixon, P.J. Newly discovered Synechococcus sp. PCC 11901 is a robust cyanobacterial strain for high biomass production. Commun. Biol. 2020, 3, 1–14. [Google Scholar] [CrossRef]
- Leu S, Boussiba S (2014) Advances in the Production of High-Value Products by Microalgae. Ind. Biotechnol. 1: 10.
- Vijay, D.; Akhtar, M.K.; Hess, W.R. (2019) Genetic and metabolic advances in the engineering of cyanobacteria. Curr Opin Biotechnol 59:150–156. [CrossRef]
- El-Fayoumy, E.A.; Shanab, S.M.; Hassan, O.M.A.; Shalaby, E.A. Enhancement of active ingredients and biological activities of Nostoc linckia biomass cultivated under modified BG-110 medium composition. Biomass- Convers. Biorefinery 2021, 13, 6049–6066. [Google Scholar] [CrossRef]
- Flores, E.; López-Lozano, A.; Herrero, A. (2015) Nitrogen fixation in the oxygenic phototrophic prokaryotes (cyanobacteria): the fight against oxygen. Biol Nitrogen Fixat 2–2:879–890. [CrossRef]
- Kulasooriya SA, Magana-Arachchi DN (2016) Nitrogen fixing cyanobacteria: Their diversity, ecology and utilization with special reference to rice cultivation. J Natl Sci Found Sri Lanka 44:111–128. [CrossRef]
- Pineda-Rodríguez YY, Herazo-Cárdenas DS, Vallejo-Isaza A, Pompelli MF, Jarma-Orozco A, Jaraba-Navas J de D, Cordero-Ocampo JD, González-Berrio M, Arrieta DV, Pico-González A, Ariza-González A, Aviña-Padilla K, Rodríguez-Páez LA (2023) Optimal Laboratory Cultivation Conditions of Limnospira maxima for Large-Scale Production. Biology (Basel) 12:1462. [CrossRef]
- Segers C, Mysara M, Coolkens A, Wouters S, Baatout S, Leys N, Lebeer S, Verslegers M, Mastroleo F (2023) Limnospira indica PCC 8005 Supplementation Prevents Pelvic Irradiation-Induced Dysbiosis but Not Acute Inflammation in Mice. Antioxidants 12:. [CrossRef]
- Poughon L, Laroche C, Creuly C, Dussap C-G, Paille C, Lasseur C, Monsieurs P, Heylen W, Coninx I, Mastroleo F (2020) limnospira indica pcc8005 growth in photobioreactor: model and simulation of the iss and ground experiments. Life Sci Sp Res 25. [CrossRef]
- López-Rodríguez A, Mayorga J, Flaig D, Fuentes G, Hernández V, Gómez PI (2023) Genetic characterization and assessment of the biotechnological potential of strains belonging to the genus Arthrospira/Limnospira (Cyanophyceae) deposited in different culture collections. Algal Res 73:103164. [CrossRef]
- Ali, S.S.; El-Sheekh, M.; Manni, A.; Ruiz, H.A.; Elsamahy, T.; Sun, J.; Schagerl, M. Microalgae-mediated wastewater treatment for biofuels production: A comprehensive review. Microbiol. Res. 2022, 265, 127187. [Google Scholar] [CrossRef]
- Keshari N, Zhao Y, Das SK, Zhu T, Lu X (2022) Cyanobacterial Community Structure and Isolates From Representative Hot Springs of Yunnan Province, China Using an Integrative Approach. Front Microbiol 13:. [CrossRef]
- Negi, R.; Nigam, M.; Singh, R.K. Harnessing Leptolyngbya for antiproliferative and antimicrobial metabolites through lens of modern techniques: A review. Algal Res. 2024, 83. [Google Scholar] [CrossRef]
- Kumar, V.R.; Narendrakumar, G.; Thyagarajan, R.; Melchias, G. A comparative analysis of biodiesel production and its properties from Leptolyngbya sp. BI-107 and Chlorella vulgaris under heat shock stress. Biocatal. Agric. Biotechnol. 2018, 16, 502–506. [Google Scholar] [CrossRef]
- Cui, J.; Morita, M.; Ohno, O.; Kimura, T.; Teruya, T.; Watanabe, T.; Suenaga, K.; Shibasaki, M. Leptolyngbyolides, Cytotoxic Macrolides from the Marine Cyanobacterium Leptolyngbya sp.: Isolation, Biological Activity, and Catalytic Asymmetric Total Synthesis. Chem. Eur. J. 2017, 23, 8500–8509. [Google Scholar] [CrossRef]
- Żymańczyk-Duda, E.; Głąb, A.; Górak, M.; Klimek-Ochab, M.; Brzezińska-Rodak, M.; Strub, D.; Śliżewska, A. Reductive capabilities of different cyanobacterial strains towards acetophenone as a model substrate – Prospect of applications for chiral building blocks synthesis. Bioorganic Chem. 2019, 93, 102810. [Google Scholar] [CrossRef] [PubMed]
- Jodlbauer, J.; Rohr, T.; Spadiut, O.; Mihovilovic, M.D.; Rudroff, F. Biocatalysis in Green and Blue: Cyanobacteria. Trends Biotechnol. 2021, 39, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Sun, H.; Chen, R.; Sun, J.; Mo, G.; Luan, G.; Lu, X. Multiple routes toward engineering efficient cyanobacterial photosynthetic biomanufacturing technologies. Green Carbon 2023, 1, 210–226. [Google Scholar] [CrossRef]
- Baunach, M.; Guljamow, A.; Miguel-Gordo, M.; Dittmann, E. Harnessing the potential: advances in cyanobacterial natural product research and biotechnology. Nat. Prod. Rep. 2023, 41, 347–369. [Google Scholar] [CrossRef]
- Prasanna, R.; Saxena, A.K.; Jaiswal, P.; Nayak, S. Development of alternative support system for viable count of cyanobacteria by most probable number method. Folia Microbiol. 2006, 51, 455–458. [Google Scholar] [CrossRef]
- Parte, S.; Sirisha, V.L.; D’Souza, J.S. Biotechnological Applications of Marine Enzymes From Algae, Bacteria, Fungi, and Sponges. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 80, pp. 75–106. [Google Scholar]
- Knoop, H.; Gründel, M.; Zilliges, Y.; Lehmann, R.; Hoffmann, S.; Lockau, W.; Steuer, R. Flux Balance Analysis of Cyanobacterial Metabolism: The Metabolic Network of Synechocystis sp. PCC 6803. PLOS Comput. Biol. 2013, 9, e1003081. [Google Scholar] [CrossRef]
- Zhao L, Lin LZ, Zeng Y, Teng WK, Chen MY, Brand JJ, Zheng LL, Gan NQ, Gong YH, Li XY, Lv J, Chen T, Han BP, Song LR, Shu WS (2023) The facilitating role of phycospheric heterotrophic bacteria in cyanobacterial phosphonate availability and Microcystis bloom maintenance. Microbiome 11:1–16. [CrossRef]
- González-Morales SI, Pacheco-Gutiérrez NB, Ramírez-Rodríguez CA, Brito-Bello AA, Estrella-Hernández P, Herrera-Estrella L, López-Arredondo DL (2020) Metabolic engineering of phosphite metabolism in Synechococcus elongatus PCC 7942 as an effective measure to control biological contaminants in outdoor raceway ponds. Biotechnol Biofuels 13:1–19. [CrossRef]
- Dexter, J.; Fu, P. Metabolic engineering of cyanobacteria for ethanol production. Energy Environ. Sci. 2009, 2, 857–864. [Google Scholar] [CrossRef]
- Pattharaprachayakul, N.; Choi, J.-I.; Incharoensakdi, A.; Woo, H.M. Metabolic Engineering and Synthetic Biology of Cyanobacteria for Carbon Capture and Utilization. Biotechnol. Bioprocess Eng. 2020, 25, 829–847. [Google Scholar] [CrossRef]
- Gomaa, M.A.; Al-Haj, L.; Abed, R.M.M. Metabolic engineering of Cyanobacteria and microalgae for enhanced production of biofuels and high-value products. J. Appl. Microbiol. 2016, 121, 919–931. [Google Scholar] [CrossRef]
- Bhati R, Mallick N (2012) Production and characterization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) co-polymer by a N 2-fixing cyanobacterium, Nostoc muscorum Agardh. J Chem Technol Biotechnol 87:505–512. [CrossRef]
- Dodds WK, Gudder DA, Mollenhauer D (1995) THE ECOLOGY OF NOSTOC. J Phycol 31:2–18. [CrossRef]
- Nostoc muscorum - microbewiki. https://microbewiki.kenyon.edu/index.php/Nostoc_muscorum. 2024.
- Wang, Y.; Tao, F.; Ni, J.; Li, C.; Xu, P. Production of C3 platform chemicals from CO2 by genetically engineered cyanobacteria. Green Chem. 2015, 17, 3100–3110. [Google Scholar] [CrossRef]
- Pembroke JT, Ryan MP (2020) Cyanobacterial Biofuel Production: Current Development, Challenges and Future Needs. 35–62. [CrossRef]
- Gu, F.; Li, C.; Zheng, H.; Ni, J. Engineering cyanobacteria for the production of aromatic natural products. Blue Biotechnol. 2024, 1, 1–17. [Google Scholar] [CrossRef]
- Satta, A.; Esquirol, L.; Ebert, B.E. Current Metabolic Engineering Strategies for Photosynthetic Bioproduction in Cyanobacteria. Microorganisms 2023, 11, 455. [Google Scholar] [CrossRef]
- Lai, M.C.; Lan, E.I. Advances in Metabolic Engineering of Cyanobacteria for Photosynthetic Biochemical Production. Metabolites 2015, 5, 636–658. [Google Scholar] [CrossRef]
- Salehi, S.; Abdollahi, K.; Panahi, R.; Rahmanian, N.; Shakeri, M.; Mokhtarani, B. Applications of Biocatalysts for Sustainable Oxidation of Phenolic Pollutants: A Review. Sustainability 2021, 13, 8620. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111. [Google Scholar] [CrossRef]
- Just G, Potvin P, Hakimelahi GH 3-Diethylphosphonoacrolein diethylthioacetal anion (6*), a reagent for the conversion of aldehydes to u,p-unsaturated ketene dithioacetals and three-carbon homologated u ,p-unsaturated aldehydes.
- Galezowska, J.; Gumienna-Kontecka, E. Phosphonates, their complexes and bio-applications: A spectrum of surprising diversity. Co-ord. Chem. Rev. 2012, 256, 105–124. [Google Scholar] [CrossRef]
- Thornburg, C.C.; Cowley, E.S.; Sikorska, J.; Shaala, L.A.; Ishmael, J.E.; Youssef, D.T.A.; McPhail, K.L. Apratoxin H and Apratoxin A Sulfoxide from the Red Sea Cyanobacterium Moorea producens. J. Nat. Prod. 2013, 76, 1781–1788. [Google Scholar] [CrossRef]
- H:\chm_51\website\chapters\carey-13.PDF | Enhanced Reader.
- Sengupta S, Jaiswal D, Sengupta A, Shah S, Gadagkar S, Wangikar PP (2020) Metabolic engineering of a fast-growing cyanobacterium Synechococcus elongatus PCC 11801 for photoautotrophic production of succinic acid. Biotechnol Biofuels 13:1–18. [CrossRef]
- Zhang, S.; Sun, J.; Feng, D.; Sun, H.; Cui, J.; Zeng, X.; Wu, Y.; Luan, G.; Lu, X. Unlocking the potentials of cyanobacterial photosynthesis for directly converting carbon dioxide into glucose. Nat. Commun. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- Ciebiada, M.; Kubiak, K.; Daroch, M. Modifying the Cyanobacterial Metabolism as a Key to Efficient Biopolymer Production in Photosynthetic Microorganisms. Int. J. Mol. Sci. 2020, 21, 7204. [Google Scholar] [CrossRef]
- Mouga T, Pereira J, Moreira V, Afonso C (2024) Unveiling the Cultivation of Nostoc sp. under Controlled Laboratory Conditions. Biology (Basel) 13:306. [CrossRef]
- Brenes-Álvarez, M.; Olmedo-Verd, E.; Vioque, A.; Muro-Pastor, A.M. A nitrogen stress-inducible small RNA regulates CO2 fixation in Nostoc. Plant Physiol. 2021, 187, 787–798. [Google Scholar] [CrossRef]
- Ciani M, Adessi A (2023) Cyanoremediation and phyconanotechnology: cyanobacteria for metal biosorption toward a circular economy. Front Microbiol 14:1166612. [CrossRef]
- Cepoi, L.; Zinicovscaia, I.; Valuta, A.; Codreanu, L.; Rudi, L.; Chiriac, T.; Yushin, N.; Grozdov, D.; Peshkova, A. Bioremediation Capacity of Edaphic Cyanobacteria Nostoc linckia for Chromium in Association with Other Heavy-Metals-Contaminated Soils. Environments 2021, 9, 1. [Google Scholar] [CrossRef]
- Lourembam J, Haobam B, Singh KB, Verma S, Rajan JP (2024) The molecular insights of cyanobacterial bioremediations of heavy metals: the current and the future challenges. Front Microbiol 15:1450992. [CrossRef]
- Zhu, Y.; Chen, Q.; Shao, L.; Jia, Y.; Zhang, X. Microfluidic immobilized enzyme reactors for continuous biocatalysis. React. Chem. Eng. 2019, 5, 9–32. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Miyazaki, M. Bioremediation of Hazardous Pollutants Using Enzyme-Immobilized Reactors. Molecules 2024, 29, 2021. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef]
Figure 1.
Cascade of photobiocatalysis.
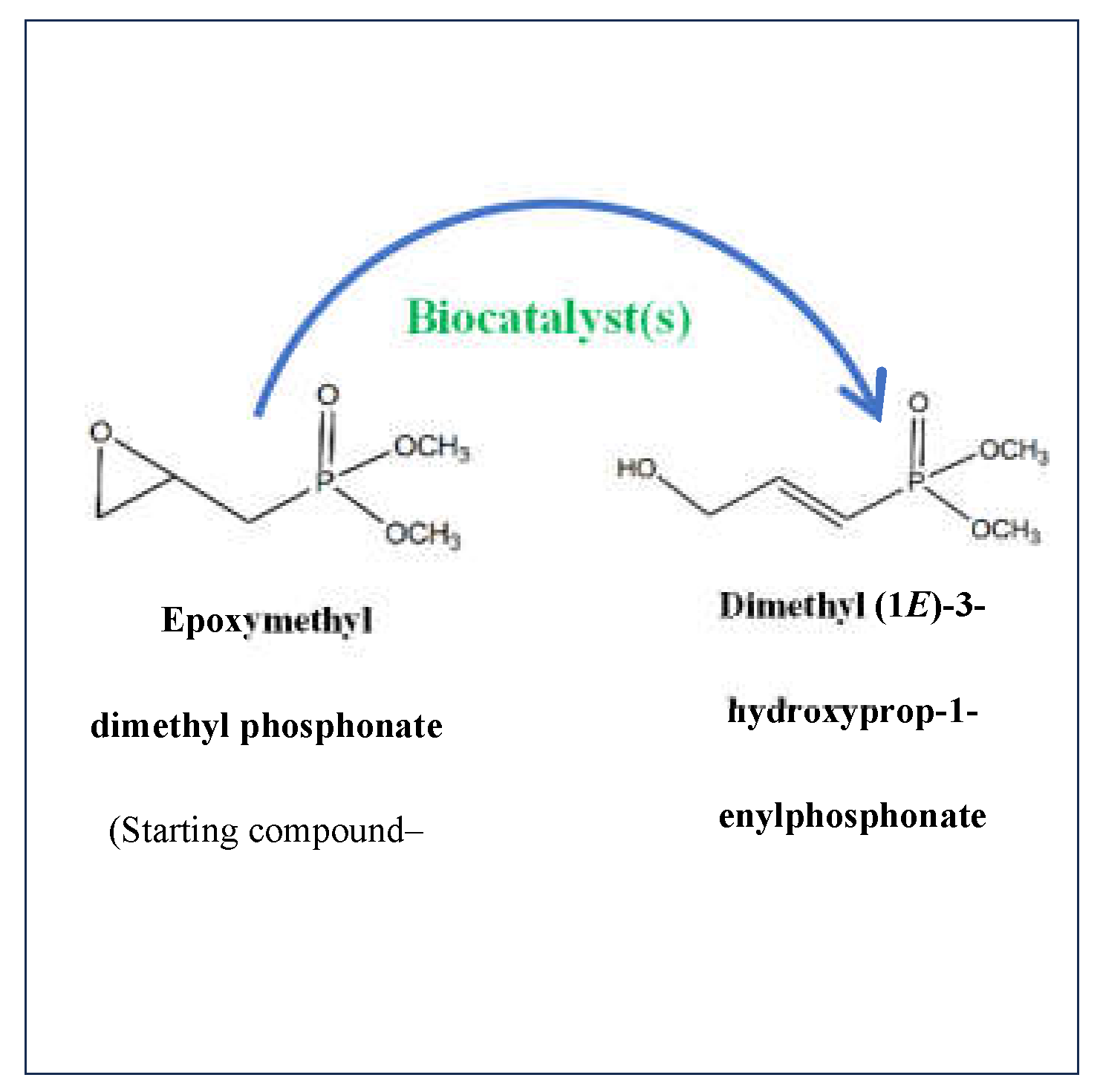
Figure 2.
Comparative analysis of the bioconversion degrees by cyanobacterial biocatalysts.
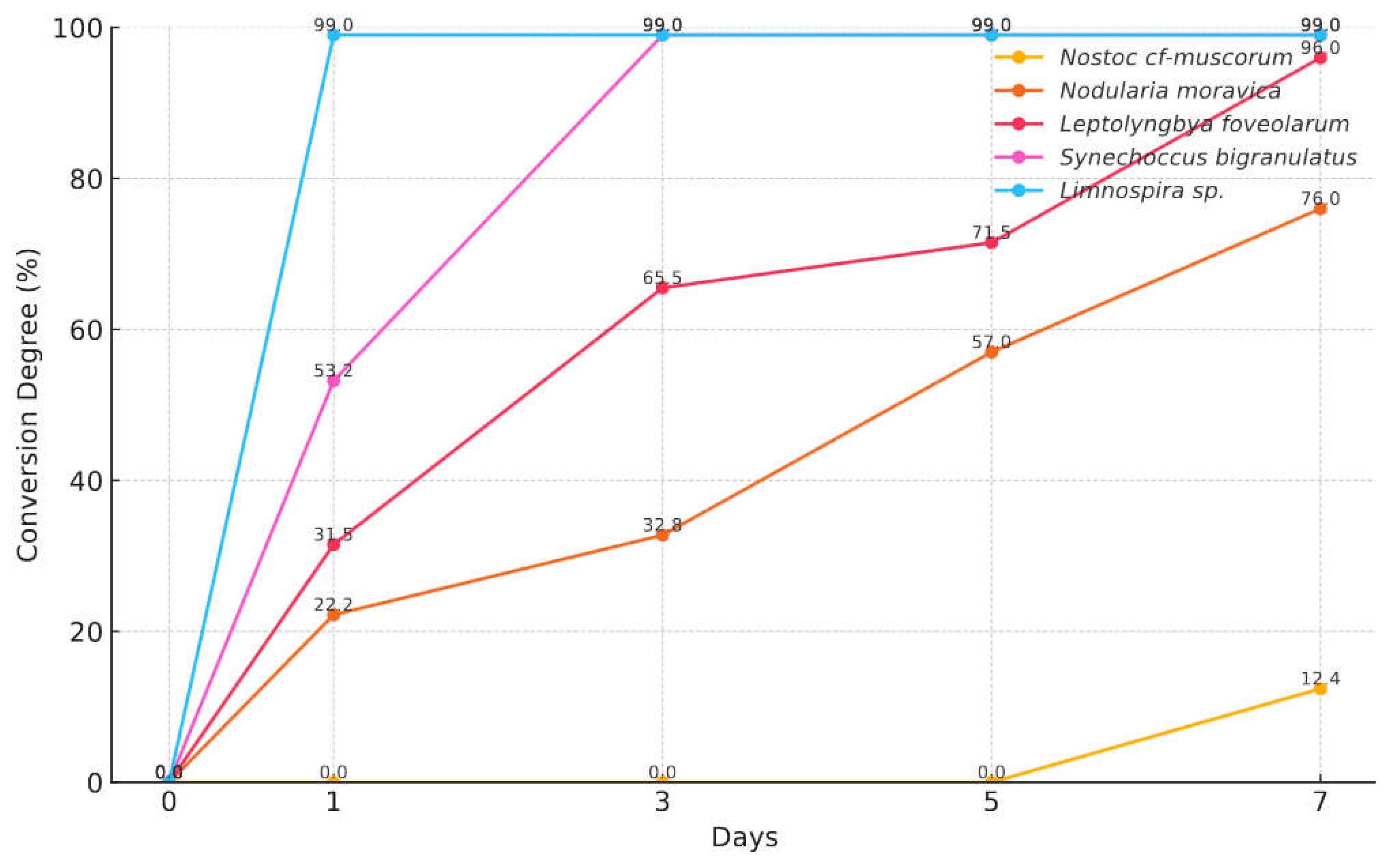
Figure 3.
Spectral analysis of the bioconversion process: (a) 31P NMR spectrum of the substrate, epoxymethyl dimethyl phosphonate; (b) 31P NMR spectrum of the fully converted product, dimethyl (1E)-3-hydroxyprop-1-enylphosphonate; (c) 31P NMR spectrum depicting partial substrate conversion; (d) IR spectrum of the fully converted bioconversion product.
Figure 3.
Spectral analysis of the bioconversion process: (a) 31P NMR spectrum of the substrate, epoxymethyl dimethyl phosphonate; (b) 31P NMR spectrum of the fully converted product, dimethyl (1E)-3-hydroxyprop-1-enylphosphonate; (c) 31P NMR spectrum depicting partial substrate conversion; (d) IR spectrum of the fully converted bioconversion product.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



