
Submitted:
06 August 2025
Posted:
06 August 2025
You are already at the latest version
Abstract
Alterations and maladaptations of the immune system remain some of the most controversial concepts in autism spectrum disorder (ASD). Nonetheless, intensifying evidence confirms that much of what ASD involves is related not to fixed ´autistic states of being´ but rather to the consequences of environmental insult and complex psychological and physiological processes along a neuro-immune-microbiota-axis. This paper focuses on clinical research and is written to specialists and non-specialists, to provide access to multi- and interdisciplinary perspectives with wide-ranging cutting-edge implications for all people with ASD. Beginning with an epidemiological and etiological underpinning, we elaborate on the current role of the immune system in the pathophysiology of ASD. Theoretical and scientific discourse on the relationship of the immune system with the nervous system and host microbiota in homeostasis/allostasis, neurodevelopment, and psychological and physiological health and disease is also provided. This gives us a platform for not only examining the role of the immune system in the etiology, pathogenesis, and pathophysiology of ASD but also understanding social and higher-level processes of consciousness for individuals on the spectrum. Finally, taking a neuroimmunological perspective, we highlight the need for a multi-scale, holistic, and ´middle-out´ approach to understanding and developing future therapeutic modalities to address the core symptoms of ASD that go beyond the current reductionist and ´magic-bullet´ medical paradigm.
Keywords:
autism spectrum disorder
; immune system
; neuroimmunology
; gut-brain axis
; bioregulatory systems medicine
1. Introduction
In the 1910s, the Swiss psychiatrist Eugen Bleuler introduced the term autism to refer to what he identified as one of the symptoms of schizophrenia [61]. A fundamental description of autism spectrum symptoms was later developed in the 1920s by Sucharew [481], followed by systematic case reports by Kanner [253] and Asperger [29] in the 1940s. Kanner and Asperger laid the foundation to make the distinction between autism and childhood schizophrenia [535], later clarified in the 1980s with the 3rd edition of The Diagnostic and Statistical Manual of Mental Disorders (DSM-III) [20,440]. Over time, the apparent heterogeneous nature of autism in clinical presentation, etiology, underlying neurobiology, and degree of severity led to the adoption and common usage of the term autism spectrum disorder (ASD) [334]. This conveys a shift from categorical concepts of autistic syndromes to a dimensional concept, within the larger framework of neurodevelopmental disorders (NDD). The current consensus, according to the National Institute of Mental Health (NIMH), defines ASD as “a neurological and developmental disorder that affects how people interact with others, communicate, learn, and behave” [381]; while the current diagnostic criteria for ASD according to DSM-V [19], states that a child must have persistent deficits in areas of social communication and interaction plus restricted repetitive behaviors and interests. Other disorders that share features with ASD include Asperger’s disorder, Rett’s disorder, Landau-Kleffner Syndrome, Fragile X syndrome, childhood disintegrative disorder (Heller’s syndrome), and pervasive developmental disorder not otherwise specified [239].
ASD is broadly characterized as a neuropsychiatric disorder, while its manifestation is that of a psychiatric syndrome [449]. Its most brutal manifestation would indicate, as Eugen Bleuler first identified, “a neonate’s tendency to turn away from reality and retire into a subjective world” [62]. Current real-world evidence documents that, in a significant number of cases, parents witness the occurrence of ASD behavioral phenotypes in normal babies who have reached all previous developmental milestones [37,175,314,317,557]. While former studies suggested that regressive forms of onset were not common in ASD, more recent investigations indicate that the rates are quite high and may be under-reported. For example, Ozonoff et al. [391] report an 88% regressive ASD phenotype in a systematic investigation of infants with and without a family history of ASD. Changes in the postnatal or early developmental periods include dramatic abnormalities in social interaction and withdrawal (i.e., lack of pair-bonding, “avert eyes” [109], and lack of neonatal imitation [437]), impairments in verbal and non-verbal communication, and a restricted repertoire of interests and activities [40], as well as seeking and finding comfort in repetitive behaviors. Babies, adolescents, and adults with ASD are also reported to have difficulties in emotion processing, in particular, problems with recognizing and discriminating emotions in others [80].
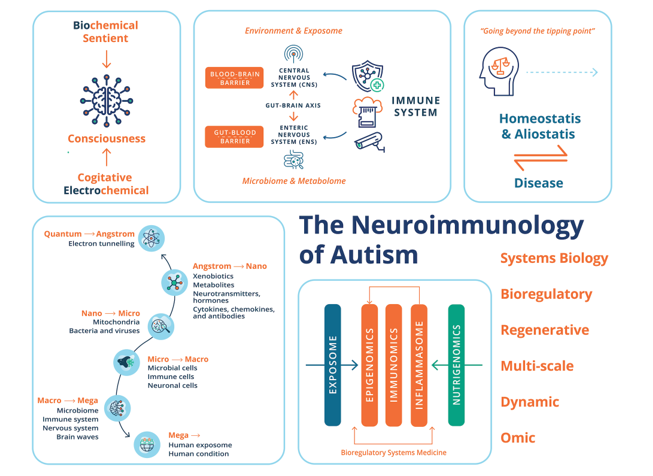
Growing evidence shows that much of what ASD involves is related not to fixed “autistic states of being” but rather to the consequences of an allostasis systems malfunction, principally mediated through environmental insult and deficiencies, leading to maladaptation and pathology of the immune system as well as metabolic processes. Response to acute or chronic stress promotes allostasis or adaptation and promotes survival by protecting the body from damage and adaptive responses for which it has immunologic memory [343]. However, when the allostatic systems are overworked beyond a “tipping point”, the capacity to respond acutely and appropriately functions poorly and, if the immunologic memory is for an inflammatory or autoimmune response within the nervous system, the shorter-term stress effect could exacerbate into a chronic disease process [344], generating heterogeneous ASD behavioral phenotypes with heterogeneity deriving from interindividual systems differences.
This review aims to move beyond genetic determinism to epigenetic regulation and its modulation by acute and chronic environmental toxic exposure and insult to the immune system in individuals with ASD. Historically, the documentation of psychological features largely preceded the documentation of physiological components of ASD. However, the pathological mechanisms on which we elaborate involve interacting sets of complex “whole-body” psychological and physiological processes along the immune-neuro-microbiota-axis, working through individuals with ASD at different rates, with different equilibria or set points when compared to neurotypical people. Real-world and clinical evidence of fever has been reported from the onset of ASD symptoms [346], which would indicate an inflammatory response in its etiology. Several immunological abnormalities have been detected both in the peripheral and central nervous system (CNS) for the pathogenesis of ASD [173]. The role of toxic compounds and their accumulation in the CNS and their role in neurodegenerative and NDDs has been acknowledged for many decades [257]. However, ASD was not thought to be associated with any loss or maladaptation of the function of the immune system, as a malfunction in the CNS, which was thought to be the source and locus of autism, was understood to be shielded from immune influences by barriers. Indeed, since the 1960’s the brain was almost axiomatically viewed as an immune-privileged site [458]. However, over the last decade, mounting evidence has uncovered the substantial role of the immune system in CNS health and functioning and disease.
If the critical role of environmental insult in the pathogenesis of ASD is taken a priori, and the contributions of such insults to immune dysfunction must be acknowledged and the role of the immune system becomes paramount in its pathophysiology. This has been reviewed from the 2000s by Pardo and coworkers [396,397], Anderson et al., [23], Patterson [400], and Ashwood and coworkers [28,389] among others. Nonetheless, alterations and maladaptation of the immune system, including both adaptive and innate systems, still remain some of the most controversial concepts in ASD [414]. Building on this pioneering work, this review provides an updated and current commentary on the roles of the immune system and neuroimmunology within a broader environmental “exposome” framework in the pathology and pathophysiology of ASD.
This paper focuses on clinical research and is written for specialists and non-specialists to provide access to multi- and interdisciplinary perspectives with wide-ranging cutting-edge implications for all individuals with ASD. This development is a result of an interplay between the expansion of tools for observation, investigation, experimentation, and frames of reference, as well as major changes in social and environmental life conditions, and a significant increase in autism prevalence. We highlight the current state-of-the-art, detailing the interconnection, interdependence, and interference with or subjugation (as would be the case for autoinflammatory and autoimmune conditions) of the nervous system and host-microbiota by the immune system and the role of these interactions in the pathogenesis of ASD. We begin with the epidemiological, genetic, and environmental underpinnings of vulnerability to ASD, and on this basis, turn our attention to the current scientific discourse regarding the relationship of the immune system with the nervous system and host microbiota in homeostasis/allostasis, neurodevelopment, and psychological and physiological health and disease. This gives us a platform for not only examining the role of the immune system in the etiology, pathogenesis, and pathophysiology of ASD but also understanding social and higher-level processes of consciousness in individuals on the autistic spectrum.
We also highlight the need for a multi-scale, holistic, and “middle-out” approach to understanding and developing future therapeutic modalities for ASD [212], which can be greatly facilitated through multidimensional omics technology and a bioregulatory systems (BrSYS) approach. Considering the heterogeneous nature of ASD and the vulnerable nature of individuals on the spectrum, we also endorse and highlight non-pharmacological, nutritional, botanical, and “mind-body” therapeutic modalities, particularly considering the growing body of scientific literature documenting the impacts of many of these approaches on neuroplasticity, immune function, and the gut-brain axis (GBA). Optimally, multiple interventions with low-risk profiles would be synergistically woven into an individualized dynamic strategy individually tailored to the specific, usually multisystem, set of vulnerabilities of each person, so as to not only provide exceptional safety profiles but also go beyond a reductionist pharmacological medical paradigm, with the capacity to not only ameliorate the symptoms of ASD but also reverse its core phenotypes.
2. Epidemiological Preambles of ASD
ASD has complex heterogeneous clinical manifestations [43], a strong bias toward males, and a broad crossover with numerous neuropsychiatric disorders. ASD phenotypes (social-communication impairments and restricted, repetitive patterns of behavior) are commonly associated with intellectual disabilities, anxiety, depression, epilepsy, attention deficit hyperactivity disorder (ADHD), sleep disorders, dyspraxia, and lack of verbal communication [38]. The significant occurrence of physiological comorbidities in ASD is also highlighted [267] by reports of increased rates of chronic physical health conditions across all organ systems in autistic children, adolescents, and adults [548]. This can be broadly sub-grouped as related to gastrointestinal [71] and metabolic disorders [169]; immune disorders (allergies, infections, primary immunodeficiency, and autoimmune) [232]; and motor/coordination disorders [354,474].
ASD is not strictly associated with mental retardation and some people on the ASD spectrum are exceedingly brilliant. Nonverbal people with ASD can also be articulate and expressive, through assistance and technologies that facilitate spelling and typing [242]. While a person unable to speak may be assumed to lack the capacity for symbolic thought that underlies language, verbal speech is only one means of expressive language, and literacy by non-verbal means has been extensively reported in nonspeaking people with ASD [242]. For example, some have learned to communicate by pointing to alphabet letters. This method remains controversial because it requires the assistance of another person who could theoretically cue them to point to letters [306]. A study published in 2020 by Jaswal et al. [243] used head-mounted eye-tracking to investigate communicative agency in a sample of nine non-speaking autistic individuals. Researchers measured the speed and accuracy with which individuals looked at and pointed to letters as they responded to novel questions. Participants pointed to about one letter per second, rarely made spelling errors, and visually fixated on most letters for about half a second before pointing to them. Additionally, their response times reflected planning and production processes characteristic of fluent spelling in non-autistic typists. These findings by Jaswal et al. would render a cueing account of participants’ performance unlikely and a blanket dismissal of assisted autistic communication unwarranted.
While ASD can involve exquisite gifts and unusual qualities of perception and thought, the condition can also involve a great deal of suffering for individuals on the spectrum as well as family and wider community. In a descriptive study, Weitlauf et al., [553] report that up to 25% of 726 participants with ASD have severe disabilities and require substantial support and 24-hour-a-day care. Siegel et al., [471] report that 11% of children with ASD are psychiatrically hospitalized in the USA before age twenty-one, and Lui et al. [307] report adolescents with ASD accessed emergency department services four times as often as adolescents without ASD. Depending on the degree of severity and intervention strategies accessed, some children with ASD may develop into independent and interdependent adults with full-time employment and self-sufficiency; however, this is seldom the case. Moreover, a clear need also becomes apparent, not only to address the core characteristics but also deeper existential suffering and loneliness for a growing and vulnerable adult population with ASD [219]. Such issues may relate to loss and change, freedom of choice versus loss of control, the dignity of the self, a sense of fundamental aloneness, altered quality of relationships, searching for meaning, mystery of what seems unknowable, and death anxiety [101,277].
ASD remained a relatively unknown disorder affecting less than 1 in 2,500 children up until the 1980s [374]. Current estimates by the CDC [321,469] and Dietz et al., [119] indicate that in the USA, up to 1 in 31 children and 1 in 45 adults have ASD, respectively. This apparent dramatic increase in ASD prevalence over the past five decades has provoked a heated debate in the academic and medical community. While the broad clinical manifestations of ASD have remained largely unchanged since Kanner’s and Asperger’s first descriptions, speculations on the increasing prevalence of ASD have emerged with better and more expanded diagnosis in observational studies [375], as well as differing external methodological factors used in epidemiological research estimating the prevalence of ASD. A rising prevalence of ASD also coincides with an epidemic of chronic childhood diseases in the USA, each of which has its own set of diagnostic criteria. At present, the CDC reports that up to 2 in 5 US students aged 6 to 17 have chronic health conditions such as asthma, diabetes, or epilepsy [81]. This again represents a dramatic increase from estimates of 18 in 1000 children reported to have such conditions sufficiently severely to interfere with usual daily activities in the 1960s [410] and estimates of 1 in 100 to 1 in 25 children under 16 to have a severe chronic illness in the 1980s [195]. In 2011, based on a 2007 National Survey of Children’s Health, Bethell et al. estimated that 54% of US children had at least 1 of 20 chronic health conditions and/or were at risk for developmental delays [53]. Based on surveys Ullah and Kaelber [528] conducted between 2016 and 2018, an estimated 40% of children and adolescents have at least one chronic disease, including obesity, eczema, asthma, food allergies, ADHD, and hypertension.
A rising, often medically complicated, and aging global population with ASD also has economic implications. For the individuals with ASD identified from 1990–2019, the lifetime social (medical, educational, productivity, and care) cost for the US was estimated to be more than $7 trillion in 2019 dollars [77]. As discussed by Cakir et al., [77] even if one assumes that the rate of increase in the prevalence of ASD is static for the next decade (2020–2029), the projected cost estimate for ASD in the US will increase to $11.5 trillion in 2019 dollars. These numbers are not unique to the USA. Autism is often regarded internationally as the most expensive disability [236]. In the United Kingdom, the total annual cost for children with ASD is $4.3 billion, while for adults it is reported to be $40.5 billion [279]; and the national cost of ASD in Australia is estimated to range from $4.5 to $7.2 billion [228].
3. Underpinnings of ASD: From Genetic to Environmental and Gene/Environment Etiologies
The ongoing debate on the rise and prevalence of ASD over nearly half a century can be associated with the central scientific dogma that ASD is a highly heritable genetic disease of the brain. Indeed, the current professional standard of care emanates from the genetic narrative’s static encephalopathy-based model of autism. Yet, although over $1 billion has been spent on genetic research in autism over the past 10 years by the NIH, Autism Speaks, and the Simons Foundation, unequivocal evidence that a genetic association is “hardwired” has not been established [211]. Moreover, genetic investigations have failed to yield a single therapy to treat the core symptoms. Concordance (shared diagnosis) of 90% of monozygotic (identical twins) and 10% of dizygotic (fraternal twins), published in the 1970s, were initially used to justify an intensive focus on genetics to the exclusion of environmental influence [146]. These findings have yet to be corroborated. For example, in 2011, Hallmayer et al. [188] reported on the largest twin study to date and reported a lower monozygotic concordance and higher dizygotic concordance. Their results yielded a smaller gap between autism rates in identical as compared with fraternal twins, with 55% of the variance for strict autism and 58% for ASD explained by shared environmental factors, with moderate genetic heritability of 37-38% [188]. In 2014, using an epidemiological sample from Sweden, Gaugler et al. [161] concluded that autism’s genetic architecture has a narrow-sense heritability of ≈52.4%, with most due to the common variations and rare de novo mutations.
A growing body of evidence implicates a strong interplay between environmental insults and epigenetics [116] in the pathophysiology of ASD [182,519], with chronic and acute multifactorial xenobiotic exposure occurring during peri- or postnatal or even preconception periods. Fuller et al. [154] report that up to nine million deaths per year (16% of all deaths worldwide) are attributed to air, water, and soil pollution alone. This is unsurprising when one considers that today’s typical contemporary diet and lifestyle are corrupted by ultra-processed and nutrient-poor food [299,348], and chemical exposure through cosmetics, home-care products, household construction materials, agrochemicals, aerosols, and pharmaceuticals in the home, school, and workplace. Scientific literature spanning many decades implicates exposure to non-metabolic heavy metals such as mercury [51,162,264] and aluminum [134,135,467]. Both have a long environmental legacy from mining in North America and have had widespread application as adjuvants in biologics recommended during pregnancy and to infants [51,60,97,163,227,264,265]. Other heavy metals implicated, again with no metabolic function in the human body, include cadmium, lead, and arsenic, all found in contaminated superfund sites. All these metals are highly neurotoxic and are known to cause neurodevelopmental deficits [583]. Energy, porphyrin, and neurotransmitter homeostasis are the key metabolic pathways affected by heavy metal exposure. Other pharmaceuticals implicated in the etiology of ASD include antibiotics [383], valproate [578], acetaminophen [247], antidepressants [347], anticonvulsants [571], and antiemetics taken by mothers during pregnancy [84]. Fetal and neonatal exposure to toxins can also occur from industrial chemicals such as polyfluorinated substances, dioxins, polychlorinated biphenyls, alkylphenols, and plasticizers like phthalates and bisphenol A [58]. All these chemicals are known to be endocrine-disrupting chemicals and can disrupt normal immune function in the brain, leading to chronic or excessive neuroinflammation. Organophosphorus (OP) compounds are a class of acetylcholinesterase inhibitors used in agrochemicals (pesticides, insecticides, herbicides, and fungicides) and have a long history in chemical warfare as neurotoxins [421]. Some chronic illnesses that manifest symptomatology overlapping with ASD, such as Gulf War Illness, have (at least in part) been attributed to OP exposure [240,517,576]. A proposed etiological role of glyphosate for ASD has recently been proposed by Seneff, Kyriakopoulos, and Nigh [460]. Glyphosate is a widely used active ingredient in agricultural herbicides, inhibiting the biosynthesis of aromatic amino acids in plants by targeting their shikimate pathway [446]. The shikimate pathway is not present in mammals per se; however, the pathway is present in gut bacteria [453]. Walsh, Hill, and Ross [545] further hypothesize and explore the potential of glyphosate to inhibit the growth or functionality of beneficial microbes in the gut. Moreover, glyphosate can substitute for glycine in protein synthesis, creating potential havoc [461]. Studies spanning many decades have shown that exposure to low-level, low-frequency electromagnetic radiation (EMF) can break DNA chains, damage proteins, even increase the blood-brain barrier permeability, disturb sleep, and cause fatigue, memory, and concentration (ADHD) problems [483]. Herbert and Sage [213,214] reviewed the pathophysiological damage to core cellular processes that are associated both with ASD and with the biological effects of EMF exposures that contribute to chronically disrupted homeostasis and human health [372]. Epidemiological studies have also shown a clear association between maternal immune activation (MIA) and schizophrenia or autism in the progeny [133]. Maternal autoimmune disorders, allergies, asthma, acute stress, and exposure to environmental pollutants have been linked to an enhanced risk of ASD and schizophrenia [133]. Emerging evidence suggests similar links for disorders like cerebral palsy and aging-associated neurodegenerative diseases, positioning MIA as a factor in the brain’s responsiveness to cumulative lifetime exposure to environmental insults [281].
4. The Immune & Nervous Systems: “Systems of Relations”
In an abstract sense, the current dogma states that the immune system is tasked to monitor and interpret insults and potential threats (i.e., toxins, pathogens, and wounds) from the external world and mount appropriate defensive actions. This would include a sophisticated innate and adaptive defense system to counter environmental insults successfully. At the same time, it also monitors the states of self of internal organs, including the nervous system, facilitates resistance to stress and maintenance of homeostasis, and has an essential role in allostasis and healing responses [424]. Of note, a general evolutionary trend indicates an inverse correlation between the ability to regenerate damaged/lost body parts and the development of an advanced immune system [3]. The same abstract framing of the immune system also applies to the nervous system, particularly the CNS, which processes information from the external and internal worlds and commands reactions to external and internal stimuli to maintain homeostasis, allostasis, and survival.
Homeostasis involves the maintenance and defense of vital physiological variables such as blood pressure and blood sugar. Walter Cannon first defined this concept in 1929 as the principle underlying physiological regulation [145]. Later, in the late 1980’s Sterling and Eyer coined the term allostasis to reflect the process whereby adaptive organisms must be able to change the defended levels of one or more regulated parameters as needed to adjust to new or changing environments [344]. Allostatic systems promote adaptation to stressful experiences and are generally most useful when rapidly mobilized and terminated. However, when they are prolonged without resolution, potentially irreversible diseased states can be reached. Allostatic processes can undermine mental and physical health, primarily because of their effects on brain plasticity, immune, microbiome, and metabolic pathophysiology [475]. Homeostasis in the nervous tissue is controlled by glial cells (astrocytes, microglia) and mast cells, resident and invasive immune cells (e.g., T-cell and B-cell lymphocytes, neutrophils, dendritic cells, macrophages, mast cells); and immune signaling and inflammatory regulators (e.g., chemokines and cytokines). The immune system is essential for normal healthy functioning and the neuroinflammatory response [1] and tumor immunosurveillance [480]. However, it can also act as a double-edged sword and exhibit complementary and inhibitory functions [575]. For example, the chronic activation of the immune system has been highlighted to trigger self-reinforcing disease processes through failed shut-off of stress-responsive hormone systems [132]. Immune dysfunction and neuroinflammatory components are also associated with several neurodegenerative diseases, including Alzheimer’s disease and Parkinson’s disease [207,291].
Following on and as reviewed by Hiller-Sturmhöfel and Bartke [224], the CNS and the glandular endocrine system are intimately connected, forming the neuroendocrine system. Hormones can be produced by endocrine glands in the hypothalamus, pituitary gland, adrenal glands, and gonads [278]. In this system, the “neural part” can recognize its environment and can memorize it, and influence the endocrine system, which acts as the “biochemical executor” [99]. In tandem with the neuroendocrine system, immune cells also synthesize, store, and secrete various hormones identical to those secreted by endocrine glands. These include the proopiomelanocortin hormones (e.g., endorphin), the thyroid system hormones, growth hormones, prolactin, melatonin, histamine, serotonin, and catecholamines [99]. In the immune-endocrine axis, immune cells can recognize and store the information (memory) and execute the commands provoked by the recognition. Immune cells are also mobile, meaning they can appear in any place in the organism, under local factors that attract them, e.g., in the case of inflammation. Blalock further theorizes that the immune system can be deemed a “sensory organ” that uses the same signals and receptors as the neuroendocrine system [59] and even as “the seventh sense” that informs the brain about its wider external and internal environment, including parasites and toxins. Indeed, neuroimmunology has established that the nervous and the immune systems are two functionally related physiological and complementary “systems of relations” that closely work together [103]. This is in distinction to systems of structure (skeletal, muscular) and maintenance (digestive, respiratory, renal, endocrine, cardiovascular, hemopoietic, and reproductive).
Finally, the nervous system and the immune system are found to be correspondingly controlled and engaged in controlling a complex, dynamic, and diversely composed microbiota with residence in various niches of the human body, which would include the oral cavity, nasal passages, lungs, skin, hair, bladder, and vagina [95]. Not only do immune cells and the microbiota help control infections and malignancy, but the extraordinary plasticity and motility of immune cells and their reliance on the highly dynamic microbiota also bridge virtually all physiological systems, making the immune system and microbiota central regulators of host homeostasis [26].
5. “The ASD Bone Is Connected to the Immune Bone, the Digestive Bone, & the Brain Bone”
The role of microorganisms in disease, also known as “germ theory”, was established in the 19th century [296], and the central role of microorganisms in childhood morbidity and mortality during the Industrial Revolution is well documented [223]. Infectious diseases in this period were principally mediated through poor sanitation, inadequate waste disposal systems and water supply, poverty, and deprivation [235,285]. In juxtaposition, more recent discoveries over the last three decades have revealed that the human body harbors a diverse ecosystem of symbiotic microorganisms, including bacteria, viruses, and fungi, collectively known as the microbiota or microbiome, that are vital for normal health and well-being and for the prevention of disease [117]. The ability of microorganisms to selectively colonize the human niche reflects their evolutionary adaptation and results in reciprocal interacting processes that form a single unified process [36] and superorganism (or “holobiont”) [64,327,486]. The most studied microbiome of the human body resides in the gastrointestinal (GI) tract. GI mucosal surfaces are intimately associated with the most abundant and diverse microbial communities in the human body [318]. The current consensus implicates four prominent bacterial phyla in the gut. I.e., Actinobacteria, Firmicutes, Proteobacteria, and Bacteroides [95]. Stress can sway the balance of the microbiome away from homeostasis. Of note, while many triggers to disease have been identified, far less is known about the triggers of a natural return to homeostasis [430]. The immune system learns to tolerate the dynamic evolution of commensal microbiota and respond appropriately to pathogens and antigens; in turn, the host microbiota is integral to educating the immune system to function correctly [329]. From this viewpoint, the immune system represents the most conspicuous set of anti-exploitation adaptations involved in human–microbial symbiosis [117]. Which mechanisms that underlie the acquisition of microbial communities are still debated and controversial. It remains unclear whether pioneer colonization starts during fetal life (i.e., “in utero colonization” hypothesis) or whether it occurs during birth and the early postnatal period (i.e., “sterile womb paradigm”) [409,487,488]. Furthermore, maternal antibodies are emerging as a key player in shaping initial interactions with the microbiota [26].
The GBA [98,292,337] provides an essential link between two important systems of the nervous system–namely, the CNS and the enteric nervous system (ENS), and the gut microbiome and metabolome [426]. The ENS represents an interdependent and extensive network of nerve cells within the gut. Its local complement component acts as a vigilant sentinel to a dynamic and evolving environment in the GI tract [568]. Even though it is now considered a third branch of the autonomic nervous system (ANS), the ENS has been referred to as the “second brain” [166], based on its size, complexity, and similarity in neurotransmitters and signaling molecules with the brain [166]. Since ENS neurons do not extend into the intestinal lumen, their ability to sense the microbiota is indirect, accomplished either by microbial molecules that have penetrated the epithelial barrier or by sensing through epithelial cells themselves [320]. The ENS is viewed as a peripheral extension of the limbic system into the gut, where it is exposed closely to our complex internal environment, including powerful mechanical, (bio)chemical, and microbial influences [566]. The GBA essentially represents the bidirectional communication between the CNS and the ENS, linking the emotional and cognitive centers of the brain with peripheral intestinal functions. The GBA has a critical role in the integration of external and internal changes in the environment and the maintenance of bodily homeostasis. Indeed, there is growing recognition that the GBA is a critical regulator of neurological functions and an intermediary to neurotoxicity by environmental stressors such as drugs, environmental contaminants, and dietary factors [113]. Interactions represent a system with bidirectional communication channels and multiple feedback loops. GBA crosstalk occurs via multiple channels, with rapid neuronal signaling primarily by the vagus nerve, as well as the gut connectome (via the semi-autonomous ENS), with more delayed feedback being achieved through neuroendocrine and neuroimmune signals into the circulation [338,490] in the form of gut-derived neurotransmitters (dopamine (DA), serotonin (5-HT), GABA and histamine), hormones and inflammatory mediators (i.e., cytokines). 90% of vagal fibers between the gut and brain are afferent, suggesting that the brain is more of a receiver than a transmitter concerning brain-gut communication [52].
Nonetheless, preclinical studies indicate the destabilization of microbial diversity and disturbed microbiome (called dysbiosis) after acute brain trauma and chronic physiological stress [255,261], suggesting the CNS is a critical player in this GBA interplay. The GBA is not only of importance for well-being but also a bridge to our understanding of homeostasis, adaptation, disease, and the body-mind connection [490]. Mounting evidence implicates the GBA in the pathogenesis of multiple chronic diseases, including inflammatory bowel disease, coeliac disease, allergies, asthma, metabolic syndrome, cardiovascular disease, and obesity [49], and the pathology of numerous neurological and psychiatric disorders. GI dysbiosis is also implicated in the etiology of ASD [423,543], and children with ASD often experience chronic GI symptoms (e.g., diarrhea, constipation, bloating, and gastroesophageal reflux [544]). Moreover, a strong positive correlation has been observed between the severity of GI symptoms and the severity of ASD symptoms [516]. Indeed, in 2010, an expert panel of the American Academy of Pediatrics strongly recommended further investigation into the role of GI abnormalities in the pathophysiology of ASD [71].
Two important boundaries in the CNS and GBA are the blood-brain barrier (BBB) and the gut-blood barrier (GBB) [320]. The integrity of both GBB and BBB has been reported to be impaired in ASD individuals [509]. The BBB constitutes the largest interface between the blood and the brain, separating the brain interstitial fluid from blood plasma [2]. In disease states, BBB breakdown and dysfunction lead to leakages of inappropriate as well as harmful blood components into the CNS, cellular infiltration, and aberrant transport and clearance of molecules. BBB breakdown can lead to neurotoxic accumulates of fibrin, thrombin, and plasmin, and red blood cell extravasation, the release of hemoglobin and ferrous iron, causing reactive oxygen species (ROS), which can all injure dopaminergic neurons [497]. Like any other organ, the brain is vascularized from the surrounding vascular plexus during embryogenesis, and the BBB is tempered by the body’s immune system from embryonic development [497]. Both the arteriolar and capillary vessel wall is covered by astrocytic end feet and glia limitans. Components of the neurovascular unit, including smooth muscle cells, pericytes (mural cells), and astrocytes, all have neuronal innervation. Sweeney et al. [497] extensively review the involvement of BBB breakdown and dysfunction in neurological deficits and other pathologies, including chronic conditions (Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis, and multiple sclerosis), as well as acute CNS disorders such as stroke, traumatic brain injury, spinal cord injury, and epilepsy. The GBB, i.e., the intestinal epithelium, is a highly dynamic structure, is estimated to completely self-renew every 4–7 days, and plays an important role in homeostasis [386]. Weakened intestinal epithelial barrier function has been suggested as a possible source of chronic inflammation in schizophrenia [386] and ASD [201] and the development of autoimmune diseases [22]. However, there are still major gaps in our understanding of how specific microbial compounds influence human conditions. The associations between circulating metabolite levels and brain function are difficult to interpret without knowledge of at least BBB permeability. In addition, many of the currently proposed mediators of microbiota-gut-brain signaling have no known receptors in the CNS.
6. The Pathophysiology of ASD
From a pathophysiological perspective, maladaptation of inflammation processes is a common feature of many acute and chronic neurodegenerative diseases [21,206] and NDDs [245]. Immune abnormalities were first described in individuals with ASD in 1977 by Stubbs et al. [493]. Since then, a growing body of research spanning nearly half a century has implicated oxidative stress, mitochondrial dysfunction, and inflammatory processes, as well as chronic and systemic immune dysregulation in the pathophysiology of ASD. These can be broadly characterized as neuroinflammation (encephalitis) and autoimmune encephalitis, involving the innate and adaptive immune systems respectively [136,345,389,452] (Figure 1). Alongside this, a growing body of evidence indicates the influence of the gut microbiota and microbial signaling molecules and metabolites (e.g., bile acids, short-chain fatty acids, and tryptophan metabolites) on neurodevelopment and behavior. Many studies have shown that early colonization, mode of delivery, and antibiotic usage significantly affect the development of the gut microbiome and the onset of ASD [500]. Many authors speculate that gut-microbiota dysbiosis may be central to the etiology and pathogenesis of ASD [71,200,226,233,403,422,491,500]. However, as discussed by White [560] and Beopoulos [50], an alternative (or perhaps complementary) hypothesis also explored here is that it may be a secondary consequence of immune pathology in the GI tract and ENS [70,380,385].
In the context of disorders of the CNS, immune cell abnormalities have been reported in brain-resident innate and adaptive immune cells in children and adults diagnosed with ASD. This includes glial cells (i.e., microglia, astrocytes, and oligodendrocytes), monocytes, natural killer (NK) cells, thrombocytes, mast cells, and dendritic cells [23,28,396,397]; and T cells (e.g., CD4+ T-helper cells (Th) (major lineages including Th1, Th2, Th17, Treg) d[225] and CD8+ lymphocytes (Tc) (major lineages include Tc1, Tc2, Tc9, Tc17, Tc22) [284,504]), and regulatory B cell lymphocytes [368]. Of note, immune memory was long thought to be restricted to the adaptive immune system; however, increasing evidence suggests that concepts of a trained immune system also apply to the innate immune system. Netea et al. [373] review the mounting evidence that cells of the innate immune system, which lack the antigen specificity, clonality, and longevity of T cells and B cells, do have some capacity to remember. This property would allow monocytes, macrophages, and NK cells to maintain homeostasis/allostasis by enhanced responsiveness when they reencounter xenobiotic insult. Many researchers have also investigated myriad chemical messengers and signaling cascades that regulate pro- and anti-inflammatory processes for their role in the etiology of ASD and as biomarkers of CNS pathology. They include, for example, cytokines (such as interleukin (Il), interferon, tumor necrosis factor (TNF), and chemokines [579]), neurotrophic factors (e.g., nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF)) [309] and insulin-like growth factor-1 (IGF-1) [202]). In parallel, current research also highlights the role of neurotransmitters (e.g., serotonin, dopamine, norepinephrine, acetylcholine, oxytocin, endogenous opioids, cortisol, histamine, glutamate, and GABA) [293,328] in the pathophysiology of ASD. All these molecules are not restricted to modulating only static neurons; they also modulate mobile immune cells. Their role from a neuroimmunological perspective would be important and evidence is provided for their involvement in the pathophysiology of ASD. This would indicate an overall departure from brain biochemical homeostasis. That said, it is noteworthy that no universal or robust biological signature or biomarker for ASD has been found. This would suggest that there is no single specific “chemical imbalance” in the CNS of all autistic individuals. Finally, sex bias and the role of sex hormones in the pathogenesis of ASD (male bias) and autoimmune disorders (female bias) have been well-documented and also further explored as a possible nexus between these disorders.
6.1. ASD, Inflammation & Oxidative Stress
Oxidative stress can be defined as an imbalance between pro-oxidants and antioxidants, resulting in a damaging action toward the cell caused by reactive oxygen species (ROS) and reactive free radicals [476]. All these molecules are produced during the activity of peroxisomes, endoplasmic reticulum, proteasome, and mitochondria (PERM) [89]. Chirumbolo and Bjørklund [89] introduce the “PERM hypothesis” to describe the complex dynamical system as a single master tuner of cellular decision-making. The authors highlighted that the ability of this system to adapt to stressors, insults, and stimuli may lie in its chaotic behavior, mainly formed by synchronized oscillatory mechanisms, involving calcium signaling, ROS, and mitochondria polarization.
Recent evidence indicates that mitochondria lie at the heart of immunity and play a key role in innate and adaptive immune response [24,350,552]. I.e., mitochondrial signaling dictates macrophage polarization and function and is necessary for responses to activators of innate immune signaling. Mitochondrial ROS regulates Th (T helper) cell activation, differential metabolic pathways regulate CD4+ cell differentiation, and mitochondrial metabolism regulates CD8+ memory formation [552]. Hanaford and Johnson [192] review preclinical and clinical evidence for the immune system’s role in the pathogenesis of mitochondrial dysfunction and disease. Current knowledge highlights a disturbed mitochondrial function in immune cells for various immunological diseases [552]. Other psychiatric diseases associated with mitochondrial disorders include bipolar disorder, anxiety disorders, schizophrenia, and ASD [326].
Several studies have suggested that redox imbalance and oxidative stress are integral parts of ASD pathophysiology [23,57,442,443,529] and inflammatory responses [472]. Substantial percentages of ASD patients display peripheral markers of mitochondrial energy metabolism dysfunction [416], such as elevated lactate, pyruvate, and alanine levels in blood, urine, and cerebrospinal fluid [152]; serum carnitine deficiency [143,152]; and enhanced oxidative stress [152,393]. Recent evidence from post-mortem studies of autistic brains points toward abnormalities in mitochondrial function as possible downstream consequences of unreactive immunity and altered calcium signaling [393]. Children with ASD are considered more vulnerable to oxidative stress because of their imbalance in intracellular and extracellular glutathione levels and decreased glutathione reserve capacity. Ghanizadeh et al., [168] discuss the role of glutathione in the context of ASD, including its involvement in neuroprotection against oxidative stress and neuroinflammation.
6.2. Innate Immune Deregulation, Encephalitis & ASD
Current research indicates that innate neuroimmune reactions play a major pathogenic role in an undefined proportion of autistic patients [531]. Glial cells (including astrocytes, microglia [429], and oligodendrocytes [468]), natural killer (NK) cells [131], mast cells [88], dendritic cells [93], and platelets [108] play pivotal yet distinct roles at various developmental stages of the fetal and neonatal nervous system. Accordingly, the immune system would have explicit and important roles in the pathogenesis of any neurodevelopmental disease, especially those whose etiology is rooted in environmental xenobiotic insult. Moreover, in the developing brain, it is important to note that the energy requirement during the rapid postnatal growth period is due to a swift developmental progression, predominantly driven by the intricate maturation and refinement of existing neurons, as neurogenesis primarily occurs before birth. Cantando et al. [79] elaborate on the interplay between astrocytes, microglia, and metabolic dysregulation in the context of ASDs, revealing a complex landscape for their potential contribution to the pathogenesis of these neurodevelopmental conditions.
6.2.1. The Pathophysiology of Glial Cells in ASD
Microglia are the major brain-resident macrophages that act as the first line of defense against injury and infection in the CNS [335]. In addition, microglia are reported to be the nervous system’s “electricians” [179] and have an important function in forming a network of immune components within the CNS and ENS [178,203,450]. New controversies have also emerged, such as the question of whether microglia are active or reactive players in neurodegenerative disease conditions, or whether they may be “victims” themselves [178]. The current consensus indicates that microglia both play a prominent role in neurodevelopmental processes like synaptic pruning and neuronal network maturation [362] and are involved in neurodegenerative diseases [178]. Microglia induce the formation of both inhibitory (I) and excitatory (E) synapses in the hippocampus as regulators of physiological homeostasis [441]. Moreover, using in-vivo imaging in rodent models, Haruwaka et al. [197] demonstrate that microglia play a dual role in maintaining BBB integrity. Deng et al. [114] further elaborate on how gut metabolites can directly regulate the functional states of microglia, or indirectly regulate them via the GBA, and their role in the pathogenesis of neurodegenerative diseases and NDDs. Microglial dysfunction in ASD is reported to be attenuated or excessive synaptic pruning, impairment of the brain’s excitation vs. inhibition (E vs. I or E/I) balance [137,275,287,295,315,411,569]. Microglial abnormalities have been identified in ASD, including in density, function, and morphology in the brain [137,287], deficient microglia autophagy [275], and pathological mechanisms involving other brain resident glial cells (i.e., astrocytes and oligodendrocytes) [315,411,569].
Astrocytes compose at least one-half of human brain tissue volume, and up to a few decades ago, were assumed to be primarily giving structural, metabolic, and functional support for neurons [405]. More recent discoveries have identified multiple functionalities of astrocytes beyond primary supportive roles, including homeostatic (molecular, cellular & network, systemic, organ, metabolic) and defensive processes of the CNS [533,542]. In the brain, astrocytes are involved in the control of synapse formation, neurogenesis, and brain vascular tone [237]. They may have not only harmful effects of aggravating neuro-inflammation and hindering synaptic sprouting or axon growth, but also beneficial effects of anti-inflammation and neuroprotection [124]. Brain physiology and pharmacology research further indicate that wakefulness and sleep depend on astroglia calcium signaling [63,404]. Astrocytes are also deemed essential to the maturation, maintenance, and regulation of the BBB in the healthy brain [124]. However, as discussed by Pociūtė et al. [415], little is known about the effects of astrocyte-secreted factors on the integrity of the BBB under physiological conditions. Alterations in the neuron–astrocyte partnership have emerged in the literature and have been shown to underlie brain lesions in pathologies as varied as brain tumors, Alzheimer’s disease, and amyotrophic lateral sclerosis [399]. Astrocytic excitation is chemically encoded and is revealed not through electrophysiology, as for neurons, but by assays of intracellular calcium concentration transients and oscillations [478]. Astrocytes are seen as local communication elements within the CNS that can generate various signals, for example, through the regulated release of ‘gliotransmitters’ including glutamate [542]. Alterations in astrocytic processing (neurogenesis, synaptogenesis, inflammation, myelination, glutamate) and number have been deemed significant contributors to ASD pathophysiology [16,79,185,530].
Oligodendrocytes myelinate the brain and spinal cord to insulate axons electrically and provide neurons with trophic and metabolic factors [45]. Emerging evidence supports the notion that oligodendrogenesis and neural myelination may play a pivotal role in the pathophysiology of ASD and its clinical presentation [159]. Steinman and Mankuta [485] provide evidence of the putative role of IGF-1 in the genesis of ASD. IGF-1 directly affects the rate at which oligodendrocytes promote myelination in the CNS [484]. IGF-1 signaling pathways are crucial for adequate axonal myelination and oligodendrocyte differentiation, but, when disrupted, can lead to white matter alterations, learning challenges, ASD-like behaviors, and neurodevelopmental and neuropsychiatric disorders [432,435].
6.2.2. The Pathophysiology of NK Cells in ASD
NK cells were originally defined as effector lymphocytes of the innate immune system [538] that control several types of tumors and microbial infections by limiting their spread and subsequent tissue damage [208]. Recent research highlights that NK cells are also regulatory cells engaged in reciprocal interactions with dendritic cells, macrophages, T cells, and endothelial cells [539]. NK cells are potent effectors of immune homeostasis, with receptors allowing them to sense the “non–self” or “missing-self” status of target cells. There is increasing evidence that NK cells link innate and adaptive immunity and play an important role in the pathogenesis of autoimmune conditions [171,308]. NK cells have been implicated in the pathology of neurological and behavioral disorders, including Tourette syndrome, schizophrenia, multiple sclerosis, neuromyelitis optica spectrum disorders, autoimmune encephalitis, Guillain-Barré Syndrome (GBS), chronic inflammatory demyelinating polyneuropathy, myasthenia gravis, and idiopathic inflammatory myopathy [131].
Over 30 years ago, Warren, Foster, and Margerten [550] reported altered NK activity in adults and children with ASD. Recently, Ebrahimi, Rostam-Abadia, and Rezaei [127] reviewed and highlighted a growing body of research detailing NK cell dysfunction in children with ASD as well as their parents. The authors discuss changes in the frequency, gene expressions, cytotoxicity features, and receptors of NK cells in children and adults with ASD. Highlighted studies include those of López-Cacho et al. [312]. Their investigation indicated an increase in the percentages of CD8+ Tc cells and B-cells, a decrease in NK cells, and a trend toward increased apoptosis in monocytes in patients with ASD. Vojdani et al., [540] further explored NK cell activity in 1027 blood samples from autistic children obtained from 10 clinics in the USA and compared the results to 113 healthy controls. 45% of a subgroup of children with ASD suffered from low NK cell activity. The authors discuss the role of low intracellular levels of glutathione [540], as also underscored by Ghanizadeh et al. [168].
6.2.3. The Pathophysiology of Mast Cells & Dendritic Cells in ASD
Mast cells are critical for allergic reactions but are also important in immune response and inflammation cascades in the nervous system [286]. Together with dendritic cells, they are the first line of defense in the immune system against invading pathogens [370]. Mast cells are present in the brain and its meninges, including the area postrema, choroid plexus, and thalamic hypothalamic region [205]. Mast cell–neuron interactions can involve the ENS and regulation of the GBB permeability and GI pathophysiology [509]. For example, mast cell activation by allergic, infectious, environmental, and stress-related triggers, especially perinatally, can release pro-inflammatory and neurotoxic molecules and disrupt the GBB and BBB, contributing to ASD pathogenesis and phenotype [25,433].
The role of mast cells in the pathophysiology of ASD has been extensively explored and discussed by Theoharides and coworkers [507,508,510,511]. Researchers speculate that stress and environmental stimuli trigger cascades involving mast cells and microglia, leading to abnormal synaptic pruning and dysfunctional neuronal connectivity [511] in brain pathology. For example, processes at a cellular level in these systems could alter the “fear threshold” in the amygdala and lead to an exaggerated “fight-or-flight” response. As a further example, corticotropin-releasing factor is secreted from the hypothalamus under stress and, together with neurotensin, can stimulate brain mast cells to release inflammatory and neurotoxic mediators that disrupt the BBB, stimulate microglia, and cause focal inflammation [510,511]. Also noteworthy, in allergy/immunology practice, it is not unusual to find food allergies in children with ASD [570]. Angelidou et al. [25] further speculate that subjects with hypersensitive mast cells may represent a unique subgroup of patients who are more likely to respond to environmental and stress triggers, precipitating or worsening ASD. Jyonouchi highlights studies that indicate a high prevalence of non-IgE-mediated food allergies in young children with ASD and further speculates that food allergies may account for some but not all GI symptoms observed in children with ASD [249]. These findings also suggest that non-allergic autoimmune mechanisms involving mast cell activation, probably in response to environmental and stress triggers, could contribute to inflammation and the pathogenesis of autism.
Alongside mast cells, dendritic cells have important functions in the modulation of the immune system and the phagocytosis of pathogens or debris, antigen presentation, activation of naïve T cells, induction of tolerance, and cytokine/chemokine production [104]. Dendritic integration plays a fundamental role in sensory processing, cognition, and conscious perception processes [189] and is hypothesized to be impaired in individuals with ASD and NDDs. Impairments include problems with dendrite morphogenesis [93], integration [371], and frequency [332]. Breece et al., [66] conducted a study of the frequencies of dendritic cells and their association with behavioral assessment and MRI measurements of amygdala volume. 57 patients with ASD were enrolled and compared to 29 typically developing controls. Researchers reported a significant increase in the frequency of myeloid dendritic cells, which was also correlated with abnormal right and left amygdala enlargement, severity of GI symptoms, and increased repetitive behaviors. Alterations in dendritic cell frequency were corroborated in a study by Saad et al., [447] of 32 children with ASD vs. 30 healthy children. Researchers enumerated data from flow cytometry of peripheral blood samples and indicated higher percentages of myeloid dendritic cells and plasmacytoid dendritic cells in the ASD group. Basheer et al. [42] also reported elevated myeloid dendritic cells in a comprehensive evaluation of various peripheral immune cell subsets and associated serum cytokine levels in 30 children with ASD, compared to 30 typically developing children.
6.2.4. The Pathophysiology of Platelets in ASD
The classical role attributed to platelets is the maintenance of hemostasis; however, more recent evidence has highlighted a central role for platelets in the host inflammatory and immune responses [244], by virtue of their high prevalence and ability to rapidly release a broad spectrum of immunomodulatory cytokines, chemokines, and other mediators, as circulating sentinels [244]. Moreover, platelets share common biological and molecular characteristics with neurons (calcium-dependent activation and secretion mechanism, cell surface receptor, and secretory vesicles with neurotransmitters such as serotonin, dopamine, glutamine, and GABA transporters) [72]. They also contribute to brain homeostasis [298], mediate protective neuroinflammation, and promote neuronal plasticity at the site of neuronal injury [126]. In general, dysfunction, abnormal activation, and morphological alteration of platelets have been implicated in many complex neurological disorders such as schizophrenia, migraine, Parkinson’s disease, Alzheimer’s disease, and ASD [177]. Clinical research by Farmer et al. [138] suggests that elevation in BDNF may be partially explained by higher platelet counts in children with ASD. Although many studies reveal associations between platelet biomarkers and ASD, there is an important knowledge gap in linking these markers with ASD and explaining the altered platelet phenotypes detected in ASD patients [392].
6.3. Adaptive Immune Dysfunction, Autoimmune Conditions, & The Pathophysiology of ASD
Alongside a decline in the incidence of most infectious diseases since the 1900s, there has been a steady rise in the incidence in industrialized countries of chronic inflammatory disorders and autoimmune diseases [31]. Autoimmune diseases are caused by an inappropriate immune response against “self” antigens, resulting in local tissue-specific and systemic chronic cascades of inflammation and tissue damage. Autoimmune conditions are hypothesized to occur through exacerbation of the immunoregulatory deficiency. This deficiency is speculated to be due to the loss of microorganisms (dubbed the “hygiene hypothesis” or “Old Friends hypothesis”) from modern urban environments, with which humans have coevolved and tolerated and relied on for the development of immunoregulatory circuits [439]. Like other chronic diseases, autoimmune diseases exhibit shared alterations in the gut microbiota. Wang et al., [547] report microbial alterations among autoimmune diseases that were substantially more consistent compared with those of other diseases (cancer, metabolic disease, and neurological disease), with microbial signatures exhibiting notable discriminative power for disease prediction. Akdis [11] further discussed an epithelial (e.g., skin and gut) barrier hypothesis, which proposes that the increase in epithelial barrier-damaging agents linked to industrialization, urbanization, and modern life underlies the rise in allergic, autoimmune, and other chronic conditions, including ASD. One alternative hypothesis that ties infection with autoimmune disease is molecular mimicry [14]. Mechanisms of molecular mimicry essentially involve infectious agents containing an antigen cross-reactive to a host antigen in the body; a less specific activation of the innate immune response can also promote autoimmune disease [13,14]. No convincing or rigorous proof exists for the theory of molecular mimicry by pathogenic agents [13,14]. However, as reviewed by Segal and Shoenfeld [459], a growing body of research implicates mechanisms involving xenobiotics, including prophylactics, in the development of autoimmune conditions. As with ASD, autoimmune diseases are heterogeneous in prevalence, manifestations, and pathogenesis. The CDC estimates that as many as 50 million people in the USA have an autoimmune disease, making it the third most prevalent disease category, surpassed only by cancer and heart disease [379]. There are more than 80 known autoimmune diseases, including about 30 autoimmune disorders of the nervous system [54,506].
Oxidative stress plays an important role in the pathogenesis of autoimmune diseases, and many environmental agents can participate in and amplify the cascade mechanisms involved [268]. Autoimmune diseases are typically defined by the autoantibodies that are produced by autoreactive B-cells and autoantigen-reactive Th cells against their host [311]. Natural autoantibodies provide immediate protection against infection and prevent inflammation by facilitating the clearance of oxidized lipids, oxidized proteins, and apoptotic cells [310]. The role of autoantibodies in the development of autoimmunity is still unclear [311] and the initial trigger for autoantibody production in patients with CNS autoimmunity is still widely unknown. However, autoantibody detection to neuronal or glial targets has resulted in a better understanding of CNS autoimmunity, and reclassification of some diseases previously thought to result from infectious, ‘idiopathic,’ or psychogenic causes [420]. Pathogens and commensals stimulate pattern recognition receptors, including toll-like receptors (TLRs), to protect against autoimmunity [31]. Pathogenic mechanisms of autoantibodies in autoimmune diseases include interaction with cell surface receptors, cell surface binding and lysis, immune complex-mediated damage, binding to extracellular molecules, and autoantibody transfer across the placenta [311]. One theory speculates that faults of the CD4+ T cell line [581] and reduced microbial stimulation of the TLRs in early life could lead to a weaker Th1 response and a stronger Th2 response to allergens. Th1 cells mainly develop following infections by intracellular bacteria and some viruses, whereas Th2 cells predominate in response to infestations by GI nematodes and are associated with allergies [438]. Emerging evidence suggests Th2 responses also play a crucial role in CNS homeostasis and disease pathogenesis [325]. Th17 cells have also developed a reputation as a destructive element in several chronic diseases [333] and auto-inflammatory neurological disorders such as multiple sclerosis, Alzheimer’s disease, Parkinson’s disease, and schizophrenia [499]. Regulatory T (Treg) cells are a population of T cells that can functionally suppress an immune response and are fundamental in maintaining T cell tolerance to self-antigens and immune homeostasis in healthy individuals [259]. Mitochondrial-regulated Tregs are also hypothesized to be involved in the occurrence and progression of autoimmune diseases of the CNS [191].
Antibodies against self-antigens are also found in cancer and during massive tissue damage [47]. Cancer involves uncontrolled cell proliferation, whereas NDDs are connected to anomalies in the development of the nervous system. Wen and Herbert [555] discuss the overlap between ASD and cancer and speculate on possible common mechanisms regarding signaling pathways related to metabolic alterations. Wen et al., [554] also conducted pathway network analyses for ASD, revealing multisystem involvement, major overlaps with other diseases, and convergence upon MAPK and calcium signaling pathways. In 2023, Yavyz et al. [572] published a large-scale study of de novo mutations in approximately 8000 samples with NDDs and approximately 10,000 tumor samples from The Cancer Genome Atlas. Mutations in NDDs tend to have a weaker functional impact and are more likely to influence differentiation compared to those in cancer.
A myriad of publications implicate MIA in schizophrenia and ASD [133,230,281]. This would support a link between cellular immune dysregulation and ASD-related behaviors. For example, Braunschweig et al. report an association between the transfer of IgG autoantibodies during early neurodevelopment and the risk of developing autism in some children [65]. Informed by these studies, an alternative and novel hypothesis for the etiology of ASD implicates autoimmune pathology involving a maladaptation of the immune system in the CNS [129]. Money, Bobrow, and Clarke [358] provide the first reports that autoimmunity of the CNS may be etiologically important in ASD. In a case report in 1971 authors describe a child with multiple diagnoses with a strong family history of autoimmune disorders. 35 years later, Edmiston, Ashwood, and Van de Water provided the first review of investigations, implicating autoimmunity and autoantibodies in individuals with ASD [129]. Moreover, epidemiological studies document a significant association between ASD and autoimmune disorders such as celiac disease, type 1 diabetes, asthma, multiple sclerosis [139], epilepsy, and atopic disease [123,507].
In 2017, Ahmad et al., [8] reported that children with ASD have imbalances between the anti- and pro-inflammatory milieu in blood leukocytes. Based on an analysis of peripheral blood mononuclear cells of children comparing ASD with a typically developing control group, researchers reported increased pro-inflammatory cytokine production and decreased anti-inflammatory molecules. In a sequel paper, Ahmad et al. [9] indicated dysregulation of Th1, Th2, Th17, and Treg cell-related transcription factor signaling in children with ASD. In the final study published in 2019, Ahmad et al., [7] used RT-PCR and western blotting to report elevated expression and significant mRNA and protein induction of IL-16 in children with ASD compared with typically developing controls. IL-16 is a chemoattractant for various CD4+ cell lines associated with proinflammatory processes and activation of glial cells. It is also reported to be closely involved in the pathology of multiple sclerosis and other inflammatory diseases in the CNS [229]. Nie et al. [378] also investigated CD4+ T cell subsets in 82 children with ASD and 50 healthy typically developing children from the Medical University of Yunnan Province. Overall, their data suggested an imbalance in inflammatory and regulatory immune responses in ASD. A higher effective T cell (Teff) to Treg ratio was associated with more severe problematic behavioral symptoms. Researchers also report that proinflammatory cytokine levels were higher in the plasma of children with ASD compared to typical controls. Molloy et al., [355] report that children with ASD had increased activation of both Th2 and Th1 arms of the adaptive immune response, with a Th2 predominance. Li et al. [303] reported elevated immune responses in the brains of autistic patients with elevated proinflammatory cytokines. However, in disagreement with reports by Molloy et al., the Th1/Th2 ratio was significantly increased in ASD patients enrolled in this study. Basheer et al. [42] report activated Th17 cells on analysis of blood serum from children with ASD. The role of Th17 cells in auto-inflammatory neurological disorders has been highlighted [229]. Moreover, Th17 cells increase the migration of other immune cells, such as neutrophils, into the inflamed CNS through the BBB and trigger inflammatory reactions that occasionally lead to irreversible neuronal damage [418]. B cells play multiple roles, including the capacity to produce antibodies and cytokine milieu, and play fundamental roles in autoimmune diseases [336]. Nadeem et al., [368] report an imbalance in pro-inflammatory and anti-inflammatory cytokines in B cells of children with ASD. This study indicated that in ASD subjects, pro-inflammatory cytokines such as IL-6 and TNF-α were elevated in B cells while anti-inflammatory cytokine IL-10 was lowered. The elevation of TNF in individuals with ASD would indicate neuropathology involving neuronal signaling and homeostasis [360,398]. Cruz-Machado et al., [473] report higher nocturnal saliva levels of TNF and IL-6 in 20 individuals with ASD compared to 20 normally developing individuals. Cruz- Machado et al. [473] also speculate that the involvement of immune-pineal axis activation, with elevated TNF but not IL-6 level, is associated with disrupted pineal melatonin release and sleep dysfunction in ASD. Further discussion of the brain-immune crosstalk in sleep is provided in a review by Marshall and Born [330]. The limbic system, amygdala, and related structures have been extensively researched in the context of the pathophysiology of ASD [498]. Limbic encephalitis is characterized by adaptive autoimmune inflammation of the limbic system. It has recently been identified as a major cause of temporal lobe epilepsy accompanied by progressive memory disturbance and emotional and behavioral changes [56].
This is noteworthy as it is now established and widely recognized that children with ASD are at high risk for developing epilepsy [522,541]. Several researchers [522] provide further reflection on the autism-epilepsy phenotype and a growing body of evidence for shared neuronal networks that can account for both ASD and epilepsy. Epilepsy is a paroxysmal disorder characterized by abnormal electrical brain activity associated with a variety of behavioral manifestations. Autoimmune encephalitis and epilepsy have been linked to neural-specific autoantibodies targeting both intracellular and plasma membrane antigens [515]. The autism-epilepsy phenotype has also recently been shown to be associated with macrocephaly, a pathologic condition due to accelerated brain growth in early development, leading to ASD. Cerebellar alterations are often observed with epilepsy, including structural changes and modulation during seizures [492].
The role of cerebellar circuitry alterations in the pathology of sensorimotor and other ASD-related behaviors has been extensively highlighted [46,121,366,494]. Numerous studies have focused on the cell pathology of the cerebella involving Purkinje cells for ASD [140]. Purkinje neurons project into deep cerebellar nuclei and are the only output cells of the cerebellar cortex; they play an important role in motor coordination, sensory processes, and cognition [107]. They contain the inhibitory neurotransmitter GABA and the calcium-binding protein calbindin, and their output from the cerebellar cortex is wholly inhibitory [83]. Reports have indicated 35–95% fewer cerebellar Purkinje cells [46] and a reduction of cell size in ASD brains compared to controls [140]. The cerebellum can be a frequent target of autoimmune attacks. The underlying cause for this vulnerability is unclear. Hampe and Mitoma [190] speculate that region-specific differences in BBB permeability, the high concentration of neurons in the cerebellum, and the presence of autoantigens on Purkinje cells, as potential explanations. Wills et al. [564] examined plasma from children with ASD for antibodies directed against human cerebellar protein. Western blot analysis revealed that 13 of 63 subjects with ASD possessed autoantibodies that demonstrated specific reactivity to a cerebellar protein. Intense immunoreactivity to what was determined morphologically to be the Golgi cell of the cerebellum was noted for 7 of 34 subjects with ASD.
GBS is an autoimmune disease of the peripheral nervous system. Its annual incidence in children under 15 years old ranges from 0.34 to 1.34 per 100,000 [563]. TLR and cytokines have been reported to have a critical role in the pathogenesis of the disease [384]. Despite having great clinical value, the interconnection between GBS and ASD has not been established and remains a mystery. Hasib et al., [198] evaluated the differential expression pattern of genes from two RNA-seq datasets to discover potential common biomarkers for GBS and ASD. 17 common differentially expressed genes were identified for these two disorders. Common genes identified the protein-protein interaction (PPI) network and pathways associated with both disorders. PPI networks are distributed in complex diseases, including cancer and autoimmune disorders [448].
Taken together, the findings of autoimmunity in families and the plethora of anti-brain antibodies reported for individuals with ASD suggest that in some patients, autoimmune encephalitis and autoantibodies that target the CNS and ENS may be a pathological or exacerbating factor in neuronal and ASD development.
6.4. Autoimmune Encephalitis, N-Methyl-D-Aspartate (NMDAR) Encephalitis & ASD
Autoimmune encephalitis comprises a group of CNS inflammatory disorders. The presentation of autoimmune encephalitis can be like encephalitis secondary to an environmental insult. A study by the California Encephalitis Project, a center focused on the epidemiology and etiology of encephalitis, found that 63% of the patients remained without infectious etiology based on a battery of tests for 16 potential infectious agents. Of these patients, the most common etiology was immune-mediated [156]. Encephalitis is indicated by acute or subacute symptoms of decreased or altered level of consciousness, lethargy, short-term memory loss, and personality changes such as apathy, irritability, agitation, and, commonly, seizures [462]. In children, neurological manifestations such as seizure, movement disorders, and focal neurological deficits are more prominent at initial presentation than psychiatric or behavioral symptoms [194]. When psychiatric symptoms do occur, they often manifest as temper tantrums, aggression, agitation, and rarely psychosis. Speech disorders are also a common complication of encephalitis in children (estimated 15-38% [382]).
NMDAR (N-methyl-D-aspartate (NMDA) receptor) encephalitis is the most common form of antibody-mediated limbic encephalitis in children and adults [160]. NMDARs are voltage-dependent ionotropic glutamate receptors [395], and glutamate is a major excitatory neurotransmitter of the CNS. It is implicated in many basic neuronal functions and CNS processes, including learning, memory, and synaptic plasticity [377]. The anti-NMDAR autoantibody is a typical synaptic protein that can bind to synaptic NMDA glutamate receptors, leading to dysfunctional glutamate neurotransmission in the brain that manifests as psychiatric symptoms (psychosis, hallucinations, and personality changes) [258]. A growing body of case reports documents NMDAR antibodies in the serum and cerebrospinal fluid of children with developmental regression, particularly of social communication skills, mimicking an autistic regression [186,273]. Kern et al., [266] review the evidence for encephalitis and microglial and astrocytic activation, a unique and elevated proinflammatory profile of cytokines, and aberrant maladaptation of B cells. The authors speculate that at least 69% of individuals with an ASD diagnosis have encephalitis. Tzang et al., [523] correlated ASD with dysfunctional autoimmunity and anti-NMDAR encephalitis. Authors also report autoantibodies, cytokines, decreased lymphocytes, serum immunoglobulin level imbalance, and T cell lineage to mediate immune responses as important biomarkers for autoimmune pathologies in ASD. Tarabeux et al., [501] explored rare or de novo mutations in NDDs. Researchers sequenced the seven genes encoding for NMDARs in a large cohort of individuals affected with schizophrenia (n = 429) or ASD (n = 428) vs. typical controls (n = 568). Sequencing identified two de novo mutations in patients with sporadic schizophrenia and one de novo mutation in a patient with ASD. Data here further supports the hypothesis that rare de novo mutations could account for some cases of sporadic ASD. This study also further supports the importance of NMDARs in psychiatric disorders with neurodevelopmental origins. In NMDAR encephalitis, unique electroencephalogram (EEG) patterns of 20–30 Hz beta activity riding on rhythmic 1–3 Hz delta waves, called extreme delta brush, have been described [54].
It remains to be seen if the relation between encephalitis and ASD is uni- or bidirectional, that is, whether children with ASD have an epigenetic diathesis to developing encephalitis (such as those mediated by the NMDAR), or conversely, if deranged or inflamed neuroreceptor processes are implicated in its development, or on occasion, both.
6.5. GI Pathology, the Immune System & Pathophysiology of ASD
There is substantial evidence that provides not only a correlational but also a causal relationship between intestinal pathology, autoimmune [147,271], and neurodevelopmental diseases [4]. Notably, the largest component of the body’s immune system is gut-associated lymphoid tissue (GALT), comprising secretory lymphoid aggregates known as Peyer’s and caecal patches. GALT plays a central role in maintaining GI homeostasis, including baseline levels of inflammation, prevention of microbial overgrowth, and neuroimmune interactions. Microfold (M) cells found in Peyer’s patches actively transport luminal antigens to the underlying lymphoid follicles to initiate an immune response, leading to autoimmune diseases [283], and play a role at the interface between innate and adaptive immunity [248]. M cells can engulf molecules in the intestinal mucosa and pass on information to the antigen-presenting cells, such as macrophages and dendritic cells, with further crosstalk with B cells for antibody production [349].
Because of their ability to transport luminal antigens and bacteria, Peyer’s patches can be considered immune sensors in the intestine [248]. The essential role of Peyer’s patches includes their influence on gut microbiome composition, communication in the ENS, and influence on behavior as part of the GBA [4,155]. A high comorbidity of celiac disease, an immune-mediated, autoimmune reaction to gluten, and ASD has also been reported and extensively discussed in the context of ASD pathophysiology [92,118]. Of note, the pathophysiology of celiac disease has been speculated to be neurological, involving loss of Purkinje cells and atrophy and gliosis of the cerebellum [367]. A commonly used restrictive dietary intervention for ASD is the gluten-free casein-free diet (GFCF) [91], which is based on the opioid excess theory [502]. Panksepp first proposed this theory to explain the pathogenesis of ASD and hypothesized that opioid peptides released from gluten and casein could pass through the mucosa and cross the blood-brain barrier, reaching the CNS and affecting brain function [394].
With opioids, the neurotransmitter serotonin has also been extensively investigated for its role in the pathophysiology of ASD in the CNS [263,551] and its ability to modulate neural plasticity and networks [301]. Serotonin is detected in the first trimester and regulates neuronal growth, differentiation, migration, and survival [105]. Serotonin also shares a strong relationship with the pathological cases of tumors (migration, metastatic dissemination, and angiogenesis), inflammation, as well as pathogen infection, and is speculated to participate in the pathogenesis of autoimmune disease [466,546]. In a systematic review and meta-analysis, Gabriele, Sacco, and Persico [157] report that approximately 30% of children with ASD show higher levels of serotonin in the blood in comparison to typically developing control groups. Serotonin signaling is also ubiquitous in the GI tract, as a neurotransmitter in the ENS, and is involved in epithelial proliferation and decreased injury from intestinal inflammation [463]. Outside the CNS, serotonin is produced in the ENS and released predominantly by enterochromaffin cells (estimated to be up to 90% of the body’s serotonin production [164]) of the gut mucosa and immune cells, with influence not only on the ENS but also on the CNS via the GBA [274]. In addition, not only does the gut secrete serotonin, but it also expresses a kaleidoscopic abundance of serotonin receptors and serotonin transporters [167]. The impressive multi-functional nature of enteric serotonin has made the identification of physiological roles difficult and sometimes controversial. A novel hypothesis for documented GI pathology for ASD could be that immune-mediated insult damages the enteric innervation (enteric ganglionitis) with increased mucosal serotonin produced, predominantly by enterochromaffin cells of the GI tract [110].
6.6. Sex Hormones, the Pathogenesis of ASD & the Immune System
While sex hormones have long been recognized for their roles in reproductive functions, within the past two decades, scientists have found that sex hormones are integral signaling modulators of the human immune system. Sex hormones are implicated in the immune response, with estrogens as enhancers at least of the humoral immunity and androgens and progesterone (and glucocorticoids) as natural immune suppressors [100]. The sex hormonal status is also known to modulate the hypothalamic-pituitary-adrenal axis (HPA), with accumulating evidence indicating that estrogen has excitatory effects, while androgen has inhibitory effects on HPA function [340]. There is also compelling evidence to suggest that sex hormones are important in the maturation of the immune system during fetal life [342]. The inherent differences in sex-dependent neuroimmune development are predominantly orchestrated by a surge of gonadal hormones unique to males, specifically occurring within a critical perinatal window that significantly impacts brain development [27].
Fetal testosterone exposure is one of several hypotheses that attempt to explain the male preponderance of NDDs, especially in ASD [141]. Fetal testosterone is involved in many aspects of development and may interact with neurotransmitters, neuropeptides, or immune pathways to contribute to male vulnerability. Gonadal hormones may also impart male-biased behavioral vulnerabilities to immune activation via microglial mitochondrial function and directly influence neurons, as well as oligodendrocytes, endothelial cells, and astrocyte functioning. McCarthy and Wright [339] review the role of microglia and astrocytes in directing brain masculinization. These authors further speculate that the natural process of brain masculinization puts males at risk of ASD by moving them closer to a vulnerability threshold, breached by refractory inflammation during critical periods of brain development [556]. Alternative hypotheses are provided by El-Ansary, Bhat, and Zayad [130], who speculate that higher ASD phenotypes in males compared with females could be attributed to the protective effect of estrogen, the higher diversity and predominance of probiotics in females, and the lower liability of females to develop leaky gut, neuroinflammation, and excitotoxicity. Kushak and Winter [289] discuss the bidirectional relationship between sex hormones and intestinal microbiota and speculate that sex bias in males may be associated with more significant changes in the intestinal microbiome than in affected girls. The sexually dimorphic prevalence of autoimmune disease for females [5,100,115,280,365,376,577] and ASD for males [141,339,556] remains one of the most intriguing clinical observations among both groups of these disorders. This raises the question of whether sex hormones could provide a bridge to understanding common etiologies of these conditions, in some neonates, occurring through differing gender-dependent mechanisms of pathogenesis.
7. ASD: From Biochemical, Immune, & Electrophysiological Correlates of Cognition, Sentience & Consciousness to the Biology of the Self
Moving on from the underlying biology of ASD, consideration can be given to linking across the levels at which ASD can be distinguished from neurotypicality, including pathophysiology (e.g., genomic vulnerability, toxicant induced metabolic and microbiome dysfunction and immune dysregulation) to conditions such as brain disease (e.g., encephalopathy), to higher levels of coordination that generate neuromotor, perceptual, emotional, cognitive, social, consciousness capacities. High-level processes of consciousness, including self-consciousness and processing of emotion in autistic individuals, have been researched from standpoints and in the context of a “brain-centered” neurocentric psychiatric disorder. From this hierarchical perspective, the fundamental perturbations of human consciousness associated with the ASD syndrome that we perceive directly with our perceptual capacities (and which are heterogeneous across the ASD spectrum) need to be linked more clearly to sets of alterations in underlying biology. However, partly due to a reductionist structuring of most “brain disorder” research, the process of identifying these linkages is essentially in its infancy.
We briefly review central and current neuroanatomical, neurofunctional, and neurocognitive theories of ASD as an integrated complement to neuroimmunological standpoints provided thus far. Moreover, based on the mounting evidence of the role of the immune system in the pathophysiology of ASD, we would endorse an expansion from primarily psychiatric and neurocentric perspectives to a broader framework that also includes (neuro)immunocentric, microbiome, and environmental underpinnings. This is not only to understand the pathophysiology of ASD, but also to incorporate components of higher-level cognition and sentience for individuals on the spectrum into comprehensive multiscale/multisystem perspectives. Such perspectives should not only consider the interactions of the nervous and immune systems with the host microbiome, but also, their collective role as “systems of relations” with all other physiological systems of the body, at all biological scales (or different hierarchical levels of resolution) (Figure 2), for a “middle-out” explanation of the system functioning, regulation or maladaptation. This would, in addition, potentially provide a framework for understanding and potentially approving existing as well as novel therapeutic modalities able to address the core symptoms of ASD. Such perspectives would also provide for useful future directions to establish a comprehensive basis to achieve partial to full recovery of at least some individuals, especially children, on the spectrum.
7.1. ASD from Underpinnings of Homeostasis/Allostasis & Sentience
The fundamental interplay between the nervous and the immune systems with the host microbiome to dynamically control physiological and psychological processes of homeostasis and allostasis cannot be overstated. Physically, every immune organ is innervated, and bi-directional communication between neural and immune systems at a cellular level is established in numerous physiological systems [322]. In addition, as highlighted by this study, the gut microbiome is in a persistent state of dysbiosis for some individuals on the spectrum. As elucidated in this review, the GBA and ENS are fundamental drivers of CNS homeostasis and brain plasticity and are therefore fundamental inputs to the development and maintenance of higher-level processes of consciousness, not only for healthy individuals but especially for those on the spectrum or with other neurological challenges.
The capacity for–and qualities of–feeling (or sentience) can be perceived as closely related to the capacity to dynamically control the physiological processes of homeostasis and allostasis [427]. Pereira distinguishes between sentience and cognitive consciousness, with a specific emphasis and application to the medical sciences [407]. As part of this paradigm, sentience refers to feeling or sensation as distinguished from perception and thought or cognition. The physiological correlates of sentience are proposed to be the system underpinning the dynamic control of biochemical homeostasis and allostasis, interrelating with functions of emotions and the capacity to feel that are higher in the overall hierarchy of functions. In contrast, the correlates of cognitive consciousness are referred to as patterns of bioelectrical activity in networks manifesting in cognition and the capacity to think. Cognition as part of this paradigm is proposed to be principally correlated to E/I balance of the brain in the context of homeostatic synaptic plasticity [87] and “metaplasticity” [234]. Based on the paradigm presented by Pereira, cognitive consciousness depends on sentience, but not vice versa. Pereira’s dynamic of sentience is adaptive, not restricted to the central nervous system (CNS) but composed of “whole-body” [408] interoceptive cycles that involve elaborate innervation of vital organs (e.g., the heart) [30] with their inputs from and outputs to the CNS and active striated motor with kinesthetic sensors. Pereira’s main hypothesis is that cognitive consciousness depends on sentience, implying that more attention should be given to the neural correlates of sentience in therapy.
7.2. From Neuroanatomical to Neurofunctional Underpinnings of ASD
Through the examination of postmortem brain tissue specimens and in vivo analysis with advanced imaging techniques such as structural nuclear magnetic resonance imaging (MRI) [111]. In the vast majority of cases, an immature autistic brain will display global and regional structural differences when compared to comparably aged non-autistic controls. Brain regions investigated in the context of ASD include the brainstem, cerebellum, hippocampus, limbic system, prefrontal cerebral cortex, thalamus, and white matter volume [44,125,210,215,216,217,479]. For high-level processes of consciousness, sentience, and cognition, there is a common distinction in neuropsychology between the limbic system as an emotional center and the associative prefrontal cortex as the seat of reason and self-consciousness. Khetrapal highlights the role of the amygdala in ASD and further explores the implication of the interaction of emotion with consciousness, and provides a framework for disturbed affective consciousness in ASD [272]. Baron-Cohen also proposes an “amygdala theory of autism” and presents plausible abnormalities of the amygdala as a cause of ASD based on a review of MRI studies [41].
Many of these early studies assumed a priori that brain anatomy was fixed, as per early assumptions that ASD was a congenital disease that irreversibly determined the state of the mind. However, with the emergence of a body of data documenting differences in autistic brains that were most likely of physiological origin–particularly neuropathological identification of neural and glial inflammation associated with enlargement of the areas of the brain, including cerebrum, that continue to grow and mature postnatally–the assumption of congenital origin came under question, since the processes of generating such changes are often associated with environmental etiologies including oxidative stress and neuroinflammation generated by noxious environmental exposures, including activation of glial cells [44,262,305,363,397,505,531]. Due to its neurodevelopmental character and the large phenotypic heterogeneity among individuals on the autism spectrum, the neurobiology of ASD is inherently difficult to describe, and much is still to be learned about specific neurobiological mechanisms driving atypical brain development. However, as elucidated by Ecker [128], it is important to note that the neuroanatomical differences are neither unique to nor necessarily causal for ASD. Instead, there is considerable overlap between the set of brain regions that mediate symptoms and, as also highlighted in this study, the neural systems underlying similar symptoms in other psychiatric and neurological conditions. We would endorse, as do Donovan and Basson [125], the performance of large-scale, longitudinal studies to determine the scale and incidence of anatomical differences, particularly during the development of the cerebellum, amygdala, and cerebrum (including both grey and white matter). Moreover, based on the mounting evidence of an immune component in some individuals with ASD and the putative role of the GBA in its pathophysiology, anatomical (volume, weight, and uniformity), network, and functional alterations should be considered neither the principal cause nor necessarily fixed in ASD but rather a correlate of disease in the context of a multifactorial and multidimensional/multiscale aetiology and pathogenesis [111] along with multifactorial environmental insult in the setting of genetic and genomic vulnerability.
State-of-the-art functional and molecular imaging using nuclear radiotracers includes magnetic resonance spectroscopy imaging (MRS), and proton electron (PET) and single-photon emission computed (SPECT) tomography, further providing insight into dynamic cellular processes in the brain of autistic individuals and higher resolution to decipher the role of atypical pathophysiology and environmental insult in its etiology and pathogenesis. This includes mitochondrial dysfunction and metabolic and oxidative stress. Cagnin, Gerhard, and Banati [76] briefly outline the rationale for employing PET for imaging of activated microglia to detect in vivo inflammatory changes occurring in a variety of brain diseases and at different disease stages, and to monitor the progression of neuroinflammation as an in vivo marker of disease activity. Researchers note that while imaging of activated microglia with PET does not serve to override an a priori clinical classification, it does provide a generic measurement of disease location and progression in the clinical presentation of patients with neurodegenerative disorders. Giron and Mazzi [172] and Tillisch et al. [513] discuss PET imaging and agents that could be applied to study the dynamic interactions of the GBA. Researchers conclude that by combining the evaluation of symptoms, behavior, and brain responses to relevant stimuli, the use of neuroimaging provides a basis to move the focus of the study of brain–gut disorders to an emphasis on underlying neural and physiological networks, with more subjective outcomes playing, if anything, a secondary role.
Thomson et al. [512] review literature of MRS studies measuring brain metabolites, including those involved in neural and glial integrity (e.g., N-acetyl aspartate (NAA), choline, myo-inositol), and oxidative stress (e.g., glutathione) in autism cohorts. The authors report significantly lower concentrations of GABA and NAA in ASD and speculate that lower concentrations of NAA in the limbic and prefrontal regions may be indicative of mitochondrial dysfunction. Zürcher et al. [584] and Li et al. [304] provide a systematic review of molecular PET and SPECT for adults and children with ASD. A clear single indicator of specific “chemical imbalance” is unapparent since serotonergic, dopaminergic, glutamatergic, and GABAergic systems are all deemed critical in shaping the disturbed neural circuitry of ASD, with few studies providing for neuroinflammatory perspectives.
A notable study is provided by Suzuki et al. [495] using PET. In this study, researchers investigated microglial activation and distribution in high-functioning autistic adults. Study results indicated increased binding in brain regions such as the fusiform gyrus, the orbitofrontal cortex, the cingulate cortex, the midbrain, and the cerebellum. Suzuki et al. [495] also endorse the need for multi-center and multimodal investigations to individualize and elucidate unique signatures and biomarkers of neurophysiological underpinnings and delineate biologically and clinically meaningful subgroups of ASD. On the other hand, there are also risks and health effects associated with the use of nuclear magnetic resonance diagnostic devices and radiotracers. For example, current safety standards do not account for synergistic effects of ionizing radiation and magnetic and electromagnetic fields [69]. In addition, to avoid motion artifacts, the sedation of some young participants is necessary. A careful risk/benefit evaluation must therefore be performed when planning the enrolment of children, especially before widespread clinical application [304].
7.3. ASD at the Level of Brainwaves & Synchronized Neural Activity
Thus far, after the historical introduction about early conceptions of autism, this paper has discussed a wide range of biological processes found to be different from “normal” in at least some subclasses of ASD. The question then becomes: how do we bridge the gap between these atypical underlying immune, host microbiota, metabolic, and other processes and the features of the higher-level phenomenology of ASD? One model suggests that such underlying processes alter or compromise brain oscillatory activity or “brainwaves” and their syntax, which can arguably be considered a fulcrum between the underlying tissue physiology and the manifest phenomena and alterations in outputs at higher levels of integration, including neuroanatomy, neurofunctional coordination, and consciousness.
Neuronal networks in several anatomical locations support oscillatory E/I activities and related synchronization phenomena in association with a wide range of cognitive and executive processes, with oscillatory bands that span from approximately 0.05 Hz or even lower to 500 Hz [75,158] and categorized as delta (1-3 Hz), theta (4-7 Hz), alpha (8-12 Hz), beta (13-30 Hz), and gamma (30-200 Hz) [471]. John [246] highlights the modulation and neuronal synchronization created by gamma oscillation, principally correlated to neural oscillations in cortical structures, the hippocampus, and the basal ganglia. Gamma band oscillations are deemed a necessary condition for awareness of stimuli and large-scale integration, or ‘binding’ of local field potentials, which is crucial for perception, movement, memory, and emotion [183,246]. Gallotto et al. [158] review intensifying evidence that cortical oscillatory signatures, for example, activity in the alpha-band, may index or even causally support conscious perception. Ward [549] reviews the thalamic dynamic core theory of conscious experience and the central importance of synchronized neural activity in cortico-thalamic circuits.
Neurodegenerative diseases, including Alzheimer’s and Parkinson’s are associated with abnormal neural synchronization [526], and numerous publications report impaired gamma oscillations in schizophrenic patients in a variety of behavioral tasks [525,527]. Guan et al. [183] highlight the susceptibility of gamma-band wave oscillations to disrupt CNS homeostasis. Buskila, Bellot-Saez and Morley [74] and Oliveira and Araque [387] review and elaborate on mechanisms by which astrocytic Ca2+ signaling and glutamate clearance play an essential role in the regulation of the network activity and K+ homeostasis, which ultimately would affect the neuronal excitability underlying network oscillations, including cortical oscillations [48,234]. These studies have provided a basis to further mine EEG data for inference of the progression of any neuroinflammatory pathologies early into the diagnosis of ASD, especially in neonates.
In comparison to studies of schizophrenia, the data on brainwave activity in autism are scarce and controversial [455,525]. Uhlhaas et al. [525] review EEG studies detailing the potential role of neural synchrony in ASD and the relationship between neural synchrony and aberrant brain development in ASDs. Researchers highlight dysfunctional gamma- (e.g., [390]) and alpha- (e.g., [412]) band activity. In a recent study, published in 2023, Santarone et al. [455] retrospectively reviewed 292 routine polysomnographic EEG tracings of preschool children (age < 6 years) with ASD. In 78.0% of cases, the EEG recordings were found to be abnormal, particularly during sleep. During wakefulness, most of the children (96.0%) did not display EEG alterations, while a minor percentage (4.0%) showed paroxysmal slowing and interictal epileptiform discharges or abnormal slow waves. Using magnetoencephalography, Kenet and coworkers [269,270] report increased long-range feedforward connectivity and reduced long-range feedback as characteristic of functional connectivity abnormalities in ASD.
Alterations in abnormal neural synchronization, brain E/I homeostasis, and synaptic plasticity are vital for network stability and sophisticated processing of patterned inputs and cognitive functions [87]. In 2003, Rubenstein and Merzenich hypothesized that some forms of ASD might be caused by a reduction in signal-to-noise in key neural circuits (sensory, mnemonic, social, and emotional) [444]. However, the precise nature of such abnormalities is inconsistent across studies, and there is no consensus in peer-reviewed literature regarding how altered cortical structure relates to E/I balance or how altered neural oscillations relate to underlying pathophysiology [115]. Contributors to such phenomena could include altered brain homeostasis, oxidative stress, and neuroinflammatory cascades, which have been correlated by EEG data for NMDAR encephalitis [54] and myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) [297].
Routine EEGs are not recommended as a screen for epileptic discharges in current practice guidelines for children with ASD [496]. However, not only is there significant comorbidity between epilepsy and ASD [522,541], but it is also the case that there are non-epileptic electroencephalographic correlates of ASD [401]. Screening EEGs for children not only with ASD but also normally developing children [496] could provide a safe, high-resolution, low-cost, and non-invasive diagnostic tool for wide-scale clinical application. In addition, it can provide a reference and benchmark if faced with clinical changes or when the patient develops into adulthood [251]. That said, the use of EEG to detect non-epileptiform abnormalities requires a level of sophistication that is not common, and in addition, most EEG studies are performed in non-shielded environments, where the pervasive presence of Wi-Fi and other sources of electromagnetic fields adds noise to EEG data that complicates interpretation, which has unfortunately discouraged EEG’s use.
Further electrophysiological correlates of ASD are also provided by the auditory brainstem response (ABR) of developing neonates. ABRs are “far-field” (i.e., recorded from the scalp) reflections of neural potentials evoked by a brief auditory stimulus and represent the neuroelectric events in the cochlear end organ and brainstem pathways set in motion by the auditory response. Evidence from ABR studies suggests that prenatal maturation in the central auditory system far precedes other aspects of sensory-motor maturation [479,482,521], which in turn form the foundation of social interactions and communication. In a landmark study, Torres et al. [521] provide the basis for a digital screening biomarker to flag neurodevelopmental derailment in neonates, capturing the nonstationary and nonlinear nature of the neonatal data using ABR. In this pilot study, babies were followed over time, and researchers report that babies who go on to receive an ASD diagnosis show profound delays perinatally in the latencies of the ABR across all seven regions of the brainstem.
7.4. Neurocognitive Theories of ASD
Hill and Frith outline the three main neurocognitive theories of ASD as: (a) theory of mind (ToM) deficit, (b) weak central coherence, and (c) weak executive dysfunction [149]. ToM refers to the ability to understand the mental states and behavior of oneself and others [39]. Ploog discusses unresolved and critical issues surrounding ToM [413] and highlights the scientific “conundrum” of the “unobservable”, the logical fallacy of circular reasoning and reification, the lack of neuroscientific correlates of ToM, and the role of language as a prerequisite for ToM. Indeed, speech and language have historically been conflated, and loss of speech in ASD doesn’t always mean lack of language. For example, in a retrospective study of 31 adolescents and adults with ASD, with limited or no speech, Jaswal et al. [242] objectively developed and assessed literacy with an iPad game. As earlier discussed, researchers report that many autistic individuals who do not talk could learn how to spell or have already learned orthographic English conventions. Jaswal et al. further speculated that if they were given appropriate opportunities, individuals with ASD who lack verbal communication would be able to learn to communicate by typing.
A characteristic of neurotypical information processing is the ability to draw together diverse information to construct higher-level meaning in context, or “central coherence” [193]. Individuals with ASD have been reported not to show the normal bias towards processing certain types of information at a global level, i.e., inefficiencies in global processing in combination with a greater propensity for detail-focused processing [150,241]. Frith and Happe suggest that weak central coherence in autism may be most appropriately viewed as a preference for a particular cognitive style, rather than a form of deficit or impairment. “Executive function” is an umbrella term used to describe cognitive tasks such as planning, working memory, impulse control, inhibition, and shifting set, as well as the initiation and monitoring of action [221,425,574]. In an extensive review, Hill [222] acknowledges the complexity of investigating executive function in ASD. This would include the possible influence of IQ on executive performance in these groups, and the possibility of overlap between performance on tests of executive function in other NDDs that are likely to involve differences in the frontal lobes. Additionally, there are considerably contradictory findings of people with ASD and diminished executive function by laboratory measures, operational tasks, and ratings-based measures, especially in preschoolers with ASD [282].
As highlighted in this study, the leading causes of any cognitive impairment in ASD could involve neurological, immune, and gastrointestinal processes, which should account for environmental and nutritional influences pre- and perinatal. Given the critical importance of food to both basic survival and cultural interaction, Allen [15] introduced a ‘‘theory of food’’ (ToF) (analogous to ToM) to represent another complex network essential for normal cognition and in the context of neurocognitive adaptation. As part of this theory, Allen speculates that the evolved internal cognitive representation of our diets in our minds, like other complex cognitive abilities, relies on complex and overlapping dedicated neural networks that develop in childhood under familial and cultural influences. Implications of ToF, as highlighted by Allen, would be that cognitive activities related to food may be cognitive enhancers for maintaining healthy brain function in aging, with implications for neurodevelopmental disorders and ASD.
7.5. ASD the Biology of the Self
From psychiatric perspectives, the ASD syndrome can be integrally understood to involve fundamental perturbations of human consciousness and self-consciousness. The first perceptual awareness of the human organism is evident in the neural integration and prospective, primary sensorimotor intentions of the second-trimester fetus [112], with self-consciousness emerging around the third year of life. Self-consciousness is a multidimensional concept that involves as much the awareness of one’s own body and image as the recognition of oneself with awareness of one’s own identity. Disorders of self-consciousness have been extensively researched in devastating neuropsychiatric disorders such as schizophrenia [532]. Like ASD, schizophrenia is a heterogeneous condition with no known or single specific etiology and is characterized by delusions, hallucinations, passivity phenomena, disordered thought processes, disorganized behavior, and cognitive deficit. Tordjman et al. [518] discuss self-consciousness disorders in schizophrenia and ASD and how impairments of body-self can lead to impairments of self-other differentiation and its consequences on social communication, emotion, cognition, and empathy. Body-self impairments, for example, can be expressed through abnormal recognition of self-image in its physical dimension, and alterations of body experience are characterized by an experience of embodiment default. Authors highlight how ASD provides a model for exploring the nature of self-consciousness, requiring the ability to reflect on mental states and the awareness of one’s own body and image as the recognition of oneself with awareness of one’s identity. Frith and Happé highlight experimental studies of normal and abnormal development, suggesting that the abilities to attribute mental states to self and others are closely related [151].
From a philosophical standpoint, the characteristics of self/oneself recognition, specificity, and memory are shared only by the immune and nervous systems [238]. Although the mechanisms by which peripheral immunity may influence neuronal function are largely unknown, recent findings implicate its influence on cognitive capabilities, such as spatial learning and memory [582]. Filiano et al. [142] implicate adaptive immune dysfunction in social dysfunction and suggest a co-evolutionary link between social behaviour and an anti-pathogen immune response driven by interferon signalling. In 2002, Robertson elucidated an “astrocentric hypothesis” describing the essential role of astrocytes in consciousness and memory formation [436]. Pereira further elaborates on the potential connection between the immune system and conscience using a neuro-astroglia interaction model of mental activity [406]. Pereira also speculates on transitioning from a neurocentric to an astrocentric perspective for cognitive and sensory processing [405].
Costantini [94] provides perspectives on the biological link between bodily self-consciousness and the immune system [106], and Sánchez-Ramón and Faure [454] present a brain self-theory from an evolutionary biological perspective by analogy with the “immune self”. Researchers hypothesize that the distinction of self/non-self (internal/external) inputs by the brain has a cellular basis similar to the immune self. With further expansion of the analogy to outline the postulates for archetypical autophrenic diseases involving the “rupture of the self,” such as schizophrenia and some forms of epilepsy, by extrapolating from the postulates that define autoimmune diseases. Bhat et al. [55] question if false inference at an immunological level alters the message passing at a psychological level (or vice versa) through a principled exchange between the two systems. Researchers also introduce the novel concept of “neuroimmunological diaschisis” and the possibility that a diaschisis of threat-avoidance may contribute to the overlap between psychiatric disorders, immunological hypersensitivities, and environmental influences and etiologies.
8. ASD and Biological Influences: A Two-Way Street
Placing the underlying physiological mechanisms and contributory environmental exposures that have been identified alongside the soaring rates over time, it is hard to avoid the conclusion that from the biochemical/pathophysiological standpoint, ASD lies, at least for the most part, on the other side of various kinds of “tipping points”, some perhaps yet biophysiologically uncharacterized, in the development of function that would represent a departure from the usual modes of brain homeostasis, which has been assumed to be, for the most part, permanent. From the perspective of ASD, the concept of “tipping point” refers to a critical threshold at which perturbations can shift a system into a new and different type of self-organization [475]. In most of this discussion, “new and different self-organization” has been tacitly assumed to be “worse.” However, such an assumption paints an incomplete picture. First, many people with autism cherish their differences and have unusual or unique talents. However, there is no intrinsic reason to exclude either the possibility or the value of such eventualities. Secondly, environmental trauma can have both constructive and destructive roles. Nonetheless, there are many cases of partial to full recoveries from ASD, often achieved through extensive and independent effort on the part of the parents of autistic children [204]. Comparable to ecological restoration, they involve increasing healthy inputs, aggressively reducing the total load of destructive inputs, and seeking multiple levels of such interventions at which these hopefully constructive changes produce positive transformation, and regain physiological and psychological processes of homeostasis.
9. Future Directions & Therapeutic Modalities for ASD
In the 1940s, Leo Kanner attributed the etiology of ASD to the emotional unavailability of an affected child’s mother [196]. While erroneous, this misconception originally and lastingly branded autism as a “brain-centered” psychiatric disorder, requiring psychiatric therapy and medications [96]. In addition, there have been nearly eight decades of a common scientific consensus on a “static” genetic etiology. Only in the last decades have researchers taken “whole-body” pathologies for neonates and children diagnosed with ASD [209]. Moreover, much like the issue of regressive autism, the idea that ASD for some is not a lifelong condition is a relatively recent addition to the knowledge of ASD. Isolated reports talking about children “growing out of autism” peppered the research literature in the 1980s and 1990s following some earlier initial interest in this topic [561].
A further limitation to our understanding of the ASD phenotype includes the reductionist paradigm, which views the human organism as a compilation of single targets for intervention and symptom alleviation without investigating its networks of interconnection. This paradigm has been central to modern healthcare models for over 150 years [514]. Although it can offer a useful first step in understanding complex systems, this paradigm also tends to oversimplify non-communicable diseases and neglect the heterogeneity of their contributing factors. This makes it not only an ineffective tool for sufficiently understanding and treating the complex nature of heterogeneous disorders such as ASD, but also an obstacle to the aspiration for the restoration of normal psychological and physiological well-being, which requires a non-linear dynamical systems approach. It is also clear that the non-linear, dynamic nature of the developing mind and the nervous and immune systems implies that potential deviations from typical development cannot be meaningfully captured with static, discrete, and linear parametric scales imposed a priori [520].
The debate continues as to the role of the immune system in ASD and whether the immune abnormality in ASD children is hyperactive or hypoactive [70]. In addition, the heterogeneous nature of ASD is reflected not only on a cellular level but also in whole-brain anatomy [397,419] and disorders of the peripheral nervous systems [67]. The need to empower and accelerate precision medicine [199] and personalized medicine approaches has become apparent and essential for the efficacious treatment of ASD [419]. Moreover, modern therapeutic modalities usually take a reductionist approach and often use medication under the guise of a “magic bullet” that can purportedly eliminate a disease’s cause or symptom. This single-target perspective tends to neglect consideration of how these medications unintentionally impact the overall regulatory ability of the human organism from “whole-body” perspectives. A systems approach would instead suggest that medications should be designed to mimic, modulate, or promote the body’s natural resolution mechanisms instead of interfering with them. Given the current healthcare and medical challenges, it is evident that the single-molecule, single-target paradigm does not provide the specificity and sophistication that a multitargeting model has the potential to offer [176].
9.1. ASD: Viewpoints from Environmental Exposomes, Omics Technologies & BrSYS Medicine
Critical windows of vulnerability include early pregnancy and childhood, where the establishment of the immune system, host microbiome, nervous system, and endocrine system can proceed in a healthy or unhealthy direction depending on the exposome [324]. The window of vulnerability to fetal and neonatal neurodevelopment brings further specificity to the ASD phenotype. Recent research in various health disciplines demonstrates that deficiency and toxicity are common etiological determinants of ill health in the contemporary era [165]. This totality of this exposure (and deficiency) can be described as the “human exposome” and refers to the variety of external and internal sources, including chemical agents, biological agents, or radiation, from conception onward, over a complete lifetime [537]. The chemical exposome further captures the diversity and range of exposures to synthetic chemicals, dietary constituents, psychosocial stressors, physical factors, and their corresponding biological responses [534]. The exposome chemical space remains largely uncharted due to the sheer number of possible chemical structures, estimated at over 1060 unique forms [451]. Individual responses to current exposures and disease susceptibility are influenced by multiple factors, including genetics, genomic variants, epigenetics, physiology, and health status, which all involve changes in biological pathways; these factors may devolve from present or past exposures (i.e., from a legacy of prior exposures or even ancestral exposures) [288]. Krausová et al. [288] review the potential early-life impact of multi-factorial exposures that can influence the exposome during gestation or after birth and highlight the lack of large-scale studies covering a broad range of xenobiotics. Hertz-Picciotto et al., [218] also report that exposure levels in pregnant women and infants are largely unknown for most contaminants and that exposome-scale studies of large cohorts with a broader chemical space coverage are scarce. Recently, Tartaglione et al., [503] conducted a systematic review of 24 case-control studies and indicated several pollutants within the first three years of life to be significantly associated with the risk of NDDs. However, the authors acknowledge that further studies are also needed to more accurately characterize the risk of NDDs by applying an “exposome approach”.
The exposome can be seen as a complement to the genome [562] and encompasses all external and internal factors that influence functional proteins, metabolites, and gene expression [34,431]. It would be vital to consider individual exposomes as a basis for precision medicine [256]. Our understanding of the role of chemical exposomes in chronic non-communicable diseases has advanced considerably with high-throughput molecular-omics techniques (metagenomics, proteomics, metabolomics) [220], which provide tools for a new emphasis on interdisciplinary research and personalized medicine for ASD. Maitre et al. [323] developed a blueprint of how multi-omics technologies can capture individual exposomes and establish signatures of disease. In a multi-center cohort of 1301 mother-child pairs, researchers associated individual exposomes of >100 chemical, outdoor, social, and lifestyle exposures during pregnancy and childhood. Of note, pregnancy exposures were predominantly associated with child DNA methylation changes. In contrast, childhood exposures are associated with features across all omics layers. A review by Higdon et al. [220] elaborates on how distinct disease subtypes can be identified in ASD, through the integration and analysis of clinical and multi-omics data.
The science of omics technologies and the immune system in particular can be described as immunomics [73]. Immunomics represents a bridge to elucidating the complex interactions between genes, environment, disease pathology, and the immune system. It also provides a new dimension to understanding immunopharmacology and immunotoxicology, with huge potential to impact disease etiology and pathogenesis, and for the development of novel and personalized therapeutics. Inflammasomes are multimeric regulatory proteins that form in the process of cytosis, in response to danger signals that perturb cellular homeostasis and are involved in inflammatory responses, including the production of proinflammatory cytokines and programmed pyroptosis and autophagy [457]. Inflammasome activity is closely associated with numerous human disorders, including autoinflammation, cardiovascular diseases, neurodegeneration, and cancer [153]. Saresella et al. [456] report that multiple inflammasome complexes are activated in ASD and suggest that therapeutic strategies targeting inflammasome activation could be useful in ASD. Considering the role of environmental insults in the etiology and pathogenesis of ASD, these early investigations provide a basis for a broad and multifactorial analysis of potential individual root causes of ASD, and strategies for precision and personalized therapy. Further direction to benchmark individual exposomes on a diagnosis of ASD in children would provide high-resolution inference and be essential to diminish any further escalation of disease states in the first instance.
Complex systems are now recognized as demonstrating “emergent” behavior with functions arising from the totality of system interactions, which are often nonlinear and stochastic. With the application of omics technologies, a systems biology approach [212] further provides a “middle-out”, dynamic, multi-scaled, and “whole-body” approach to understanding ASD and the development of personalized therapeutics. This approach defines the human body as a complex biological network of interconnected components (molecules, cells, tissues, organs) [428]. This includes not only the immune system and the nervous system but also integrated systems of structure and maintenance. Goldman et al. [176] further extend the systems biology paradigm by merging it with regulatory and relevant empirical and treatment models, which they designate as “bioregulatory systems medicine”. BrSM is rooted in the idea that robust and effective solutions for complex diseases should optimize an individual’s homeostatic and allostatic systems and their interactions across all biological systems, and emerges in part as a response to the limitations of the reductionist perspective model [514]. A goal of BrSM would be to eventually develop biomarkers tailored to individual pathophysiological disease states that predict the health-disease continuum and allow for therapeutic interventions at the specific stage of a patient’s disease. A BrSM approach also incorporates a multi-scale approach and would also emphasize the simultaneous and complex dynamic interactions of activities across all biological scales [181].
9.2. ASD Therapeutic Modalities: Standard of Care & State-Of-The-Art
The common phrase “children are not little adults” is even more true in drug development, with infants, toddlers, and children placed in the special “vulnerable population” category [180]. Clinical development of therapeutics for ASD poses challenges regarding the identification of suitable endpoints, lack of biomarkers for progression, and regulatory considerations [170,341]. Moreover, in nonverbal children or children with communication difficulties, side effects can be challenging to recognize, since children may exhibit different behaviors as side effects to the same therapeutic intervention [559]. Adverse events may also be camouflaged by comorbidities in individuals with ASD. On this basis, one must acknowledge the explicit need for an authentic human connection between practitioner and patient with ASD.
Doctors often use psychiatric drugs to address symptoms associated with ASDs. Risperidone and aripiprazole are both antipsychotics and are the only medications currently approved by the US FDA for the management of irritability in children with ASD [90]. Other classes of pharmaceuticals commonly prescribed off-label include stimulants (e.g., amphetamines and methylphenidates), selective serotonin reuptake inhibitors (SSRIs), α2-adrenergic agonist inhibitors (e.g., clonidine and guanfacine), and selective norepinephrine reuptake inhibitors [184,290,417]. West et al., [559] found that current pharmacological interventions used for ASD are only moderately useful in treating symptoms associated with ASD, such as attention, aggression, or repetitive movements, and they are only minimally effective in addressing the core deficits of ASD. All these classes of pharmaceuticals carry specific side effects and require professional long-term monitoring and management of adverse effects [18,331,524,558]. As an example, King et al. [276] conducted a randomized clinical trial of 149 individuals with ASD who received either citalopram (n = 76) or placebo (n = 76) over the 12-week therapeutic period. Clinical results indicated no significant difference in the rate of positive response. Moreover, citalopram use was significantly more likely to be associated with adverse events, particularly increased energy level, impulsiveness, decreased concentration, hyperactivity, stereotypy, diarrhea, insomnia, and dry skin or pruritus. Additionally, the vast majority of psychiatric drugs display a high potential for physiological dependence [356,357] which would make testing and use in pediatric populations challenging and potentially risky, if not redundant.
Biologics and immunosuppressive drugs are the primary treatment tools for autoimmune disorders, along with non-steroidal anti-inflammatory drugs and SSRIs [174,364]. Immunomodulating agents are designed to resemble antibodies, receptors, and other immunological factors. Emerging immunomodulating agents, including biologics, many of which were developed only recently, may provide additional treatment options for ASD subjects. However, published data on clinical trials in ASD subjects are scant [250]. The use of anti-inflammatory drugs in ASD has been reviewed by Hafizi, Tabatabaei, and Lai [187]. This included amantadine an antiviral and anti-Parkinson drug; celecoxib, a nonsteroidal anti-inflammatory drug; the corticosteroid, galantamine, an acetylcholinesterase inhibitor; intravenous immunoglobulin; lenalidomide, a derivative of thalidomide; memantine, an NMDA receptor blocker; minocycline, a tetracycline-class antibiotic with anti-inflammatory effect; the antioxidant N-acetylcysteine; pioglitazone, an antidiabetic medication; riluzole, an inhibitor of presynaptic release of glutamate; palmitoylethanolamide, a fatty acid amide with neuroprotective, anti-inflammatory, and anti-nociceptive properties; pentoxifylline, a xanthine derivative that has an inhibitory effect on inflammatory responses; spironolactone, an antagonist for aldosterone receptor; and topiramate, an anti-epilepsy medication. The authors concluded that for all pharmaceuticals reviewed, there was a lack of rigorous clinical trials and very few successful replications that did address the core characteristics or associated symptoms of ASD. Moreover, all therapeutic agents exhibited low safety profiles, including serious systemic side effects.
First-line treatments for autoimmune encephalitis include aggressive pharmaceuticals, including intravenous steroids and immunoglobulin, plasma exchange, and immunoadsorption [470]. Efficacy studies with these drugs have yielded contradictory results, and each of these treatments can result in an immune-compromised state, putting the affected population at risk for serious infections or immune-related illnesses, malignancy, and cardiovascular conditions [319]. Clinical investigations using stem cell products are currently being applied for a wide spectrum of conditions using a variety of stem cell types [120]. Siniscalco, Bradstreet, and Antonucci [477] review the therapeutic potential of hematopoietic stem cells (HSC) in ASD-related inflammation. Researchers conclude that although HSCs are attractive candidates for the potential restoration of ASD-related immune-mediated pathologies, concerns about several life-threatening risks were raised. These include abnormal trafficking of stem cells by the BBB into the CNS, resultant unintended consequences, the bidirectional nature of stem cells and potential contribution to the inflammatory state, and the risk of graft-versus-host disease [302].
Other novel interventions for ASD include microbial transplant therapy (MTT). Indeed, in 2019, the FDA recognized microbial transplant therapy with a “fast-track” designation for ASD treatment [6]. MTT is a novel method to modify the microbial ecosystem in the GI tract of the hosts. MTT has indicated success in clinical trials with significant improvements in GI symptoms, autism-related symptoms, and gut microbiota ecology. Kang et al. [252] report important changes in gut microbiota at the end of treatment that remained at follow-up, including significant increases in bacterial diversity and relative abundances of Bifidobacteria and Prevotella. However, such interventions are intrusive, can have adverse effects, and come with several limitations regarding the source and uniformity of donor feces. In addition, the use of adult fecal donors may correlate with the development of aging physiological function and unwanted immune reactions [86]. Besides, there is a lack of clinical data in children regarding many facets of MTT, such as the uniformity of transplant protocols used and the route of administration (oral, endoscopy, or enema).
9.3. ASD Therapeutics: Personalized & Precision Nutrition and Mind-Body Modalities
The maxims of “let thy food be thy medicine” attributed to Hippocrates (circa 400 BC) and ‘you are what you eat’ by Jean Anthelme Brillat-Savarin (in 1826) remain as pertinent today as they were then. In the contemporary era, research into these bioactive natural products originating from medicinal plants, fungi, and microorganisms is also based on ancient ethnopharmacological (including entheogenic) human practices [353,573]. Real-world evidence has provided ample evidence from parents of children with ASD and clinicians, of the utility of food and diet for the treatment of symptoms and sporadic reversal of the core symptoms of ASD [82]. These approaches principally involve an adjustment in dieting habits that begins with the removal of all ultra-processed foods and the encouragement of organic and whole-food diets. Indeed, the essential role of nutrition in optimizing, promoting, and recovering mental health is only now becoming recognized within medical practice and the scope of practice in psychiatry [445]. Peper and Shufor [402] coined the term “grandmother therapy assessment”, in a recommendation that diet and nutritional deficiency be the first line of assessment of ASD and ADHD. GI disorders in ASD and their treatment through nutritional therapy may also provide paths for modulating the expression of social and behavioral symptoms through the GBA. This includes not only the application of foods rich in probiotics [465], but also direct administration of prebiotics, as well as postbiotics [580]. The latter provides strategies to support natural gut healing and a shift in GI homeostasis from potentially pathogenic to beneficial and congenital microbial ecologies or, at least, to restore gut eubiosis. Indeed, GI microbiome restoration would be a prerequisite to the successful amendment of inflammation processes identified in any chronic disease [313,316]. van den Brink et al. [68] further elaborate on strategies to (re)gain inflammatory flexibility by regular mild metabolic triggers through perspectives of trained immunity, hormesis, and pro-resolution.
Modulating the neuro-immune-microbiome axis with “full-spectrum” functional foods and botanical plant-based medicines further provides a multitargeting paradigm and allows for a synergistic or “entourage effect”. The “entourage effect” essentially refers to the interaction of not only primary and secondary therapeutic agents (e.g., alkaloids, pre- and postbiotics, antioxidants, and microbial communities living in fermented foods) shown to have therapeutic potential, but also myriad secondary metabolites in the “nutritional dark matter” in the “foodome” [434,464,573]. Animals generally avoid these potentially toxic molecules, but our hominid ancestors elected to selectively consume foods rich in secondary metabolites [300] (i.e., polyphenols, flavonoids, carotenoids, phytosterols, polysaccharides, triterpenoids, tannins, saponins, and ash) as means of complementing their nutrition and which later formed the basis of natural and omnipresent archaic medicine practices and folklore. Wellness and therapeutic metabolites found in food and botanicals when taken in their natural matrix are generally safe even for chronic consumption, with extremely low toxicity, present ease in uptake or breakdown by the body, have low retention, and present low potential for physiological dependency [565].
Despite considerable interest and real-world evidence for the utility of dietary interventions for ASD, no consensus exists regarding optimal nutritional therapy, while there is much controversy regarding its effectiveness [122,148,254,260,294,359]. As for pharmaceutical interventions, the lack of consensus is unsurprising, considering the heterogeneous nature of ASD. This further emphasizes that there is no “one-size-fits-all” and that the need for a precision and personalized medicine paradigm that goes beyond the modern reductionist approach, through the application of multi-dimensional, multi-disciplinary nutritional therapy, is strong. One paradigm to address current pitfalls for successful nutritional intervention in ASD could be through the convergence of nutrition science with omics technology, or “nutrigenomics” [388] (Figure 3). The integration of nutritional systems with gene and protein expression profiles and metabolic fingerprints allows a more extensive analysis of the individual response to any nutritional intervention [32]. “Nutrigenomics” provides a more global, comprehensive understanding of how food components influence health status [32] and new opportunities oriented toward personalized interventions [32]. A few studies of transcriptomic, proteomic, and metabolomic studies have demonstrated the ability of functional foods and their bioactive components to fight against oxidative damage [361] and to modulate the underlying dynamics of low-grade chronic, systemic inflammation, impaired metabolic flexibility, and dysregulation [78]. Indeed, although the concept of combined “omics” (Figure 3) is interesting conceptually, it remains theoretical, and much work will be needed to develop such a platform as a standard of care in the clinic.
Moving along from “body-mind” modalities, “mind-body” modalities have also been endorsed by health practitioners active in the field and there is modest clinical evidence supporting their efficacy for outcomes such as social communication and cognitive functions, and anxiety [12,231]. Non-pharmacological interventions that are standard of care include cognitive and behavioral therapies (CBT) to address issues of anxiety for children with ASD [489,567] and promote autonomy and emotional well-being [351]. CBT helps strengthen cognitive skills, such as mental flexibility and problem-solving, which are essential for an independent and fulfilling life [351]. Also of note, it has been speculated that approximately 90% of vagal fibers between the gut and brain are afferent, suggesting that the brain is more of a receiver than a transmitter concerning brain-gut communication [52]. Nonetheless, this still establishes the role of non-pharmacological “mind-body” interventions for the restoration of normal neuro-immuno-microbiota functioning for individuals with ASD. At the very state-of-the-art, Baniel et al., [35] provide a theoretical foundation for the application of the Feldenkrais method and Baniel Method® NeuroMovement® intervention modalities with children and adults experiencing diverse motoric, cognitive, and social challenges. These methods leverage neuroplasticity through the utilization of movement, through a dance-like dyadic process of self-discovery that mimics the spontaneous, organic way typically developing children play, learn, and grow. This therapeutic modality would also exemplify the application of sentience [408] to influence and remedy higher-level processes of self-consciousness and cognition, and improve social interactions for individuals with ASD.
Other cutting-edge non-pharmacological technologies include breathwork training [10,352]. The use of conscious breathing practices for physical, psychological, emotional, and spiritual healing has a long and varied history [144]. However, there has been little work to bring these practices into a coherent and unified form that contributes to the field of body psychotherapy [536]. Overall, research findings indicate that breathwork may be efficacious for treating anxiety, depression, and PTSD [10]. Physical benefits of breathwork can be summarized as improving immune function; regulating arousal; decreasing sinus problems; balancing hormones, enzymes, neurotransmitters and brainwaves; stabilizing blood gasses; promoting digestion, circulation, and proper organ function; facilitating waste metabolism, aligning posture, decreasing muscle tension; and increasing motility, mobility and movement efficiency [17,33,85,536].. Despite preliminary evidence for breathwork’s efficacy in treating common psychological distress, more research is needed to evaluate its utility for treating symptoms and core deficits of ASD.
10. Concluding Remarks
The explicit role of environmental (xenobiotic) insult in the etiology and pathogenesis of ASD has been speculated upon for over half a century and highlighted here. Chronic or acute xenobiotic activation of the immune system would trigger self-reinforcing disease processes through failed shut-off of stress-responsiveness. ASD, therefore, is an example of a situation that emerges as things “go beyond the tipping point” at various vulnerable states of neurological and immunological development. As highlighted by this review, the complex pathophysiology and heterogeneous ASD phenotype are hypothesized, for some individuals with ASD, to be related to the deregulation of the immune system. This raises the question for some individuals diagnosed with ASD of whether the manifestation of immune cell pathology, and immune system dysregulation, is truly involved in the pathogenesis, or whether they are epiphenomena of the disorder; and whether their brain is truly and intrinsically electrochemically “defective” or instead biochemically “obstructed”? In support of the latter model recently D’Adamo et al., [102] provided a case report of the reversal of autism symptoms in two dizygotic twins through a personalized lifestyle and environmental modification approach. These findings and other published cases of ASD reversal are encouraging. We would endorse more dialogue between parents, clinicians, and academics, with further documentation of regression and recovery. In addition, we would also endorse more research to take a “bottom-up/middle-out/top-down” approach to address the problem of how to explain the transmission of immune/biochemical/metabolic/bioenergetic perturbations (bottom) into neurological synchronization (middle) and thence into neurocognition/behavior/social functioning (top) for individuals with ASD. A crossover of ASD pathophysiology with autoinflammatory and autoimmune conditions further provides a novel hypothesis for understanding the pathogenesis of ASD. Moreover, omics and immunomics could provide game-changing platforms to elucidate the complex interactions between genes, environment, disease pathology, and the immune system, leading to ASD phenotypes and the development of safe and effective personalized therapeutic solutions that address the core symptoms of ASD. Indeed, a key takeaway of this study would be to highlight the primary need for a multiscale, multisystem, individualized intervention approach to helping people with ASD. In addition, as such a program begins to mature, we would also endorse the design and implementation of collaborative research infrastructures to capture cognitive/emotional/social/consciousness outcomes and account for the full range of hierarchical levels of biological resolution, in the people receiving these integrated treatments.
In 2009, Plotkin, Gerber, and Offit made the bold statement that: “Autism is not an immune-mediated disease. Unlike autoimmune diseases such as multiple sclerosis, there is no evidence of immune activation or inflammatory lesions in the CNS of people with autism” [414]. Perspectives, theories, and scientific and clinical research provided in this study are in stark contradiction to the statement expressed above. In addition, after reviewing the incredibly complex neuro-immuno-microbiota interactions implicated in clinical research, we believe that only once we understand that ASD is not genetically inevitable or a genetic tragedy but an environmental and physiological catastrophe, will we truly be able to grasp and address the root causes of the dramatic rise in its prevalence. Indeed, scientific evidence presented here would suggest that people with ASD are the “canaries in the coal mine” – that is, they are the most susceptible among us who are affected first by problems that might eventually reach us all [369]. Moreover, it is quite plausible that this catastrophe goes beyond ASD. The rise of ASD may well to a significant extent evolve, or emerge, from overlapping vulnerability pathways associated with systemic problems of other chronic diseases [554]. The point henceforward becomes not just to support and seek full recovery for those diagnosed with ASD, but also to forthrightly address how we as individuals, families, communities, and society in the contemporary era can most effectively protect future generations.
References
- Abbaoui, A.; Fatoba, O.; Yamashita, T. Meningeal T cells function in the central nervous system homeostasis and neurodegenerative diseases. Front. Cell. Neurosci. 2023, 17, 1181071. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Abnave, P.; Ghigo, E. Role of the immune system in regeneration and its dynamic interplay with adult stem cells. Semin. Cell Dev. Biol. 2019, 87, 160–168. [Google Scholar] [CrossRef]
- Abo-Shaban, T.; Sharna, S.S.; Hosie, S.; Lee, C.Y.Q.; Balasuriya, G.K.; McKeown, S.J.; Franks, A.E.; Hill-Yardin, E.L. Issues for patchy tissues: defining roles for gut-associated lymphoid tissue in neurodevelopment and disease. J. Neural Transm. 2022, 130, 269–280. [Google Scholar] [CrossRef]
- Ackerman, L.S. Sex Hormones and the Genesis of Autoimmunity. Arch. Dermatol. 2006, 142, 371–376. [Google Scholar] [CrossRef]
- Adams, J.B.; Borody, T.J.; Kang, D.-W.; Khoruts, A.; Krajmalnik-Brown, R.; Sadowsky, M.J. Microbiota transplant therapy and autism: lessons for the clinic. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 1033–1037. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Al-Ayadhi, L.Y.; Attia, S.M. Elevated IL-16 expression is associated with development of immune dysfunction in children with autism. Psychopharmacology 2018, 236, 831–838. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Nadeem, A.; Ansari, M.A.; Bakheet, S.A.; Attia, S.M.; Zoheir, K.M.; Al-Ayadhi, L.Y.; Alzahrani, M.Z.; Alsaad, A.M.; Alotaibi, M.R.; et al. Imbalance between the anti- and pro-inflammatory milieu in blood leukocytes of autistic children. Mol. Immunol. 2017, 82, 57–65. [Google Scholar] [CrossRef]
- Ahmad, S.F.; A Zoheir, K.M.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Al-Ayadhi, L.Y.; Alzahrani, M.Z.; Al-Shabanah, O.A.; Al-Harbi, M.M.; Attia, S.M. Dysregulation of Th1, Th2, Th17, and T regulatory cell-related transcription factor signaling in children with autism. Mol. Neurobiol. 2016, 54, 4390–4400. [Google Scholar] [CrossRef]
- Aideyan, B.; Martin, G.C.; Beeson, E.T. A Practitioner’s Guide to Breathwork in Clinical Mental Health Counseling. J. Ment. Heal. Couns. 2020, 42, 78–94. [Google Scholar] [CrossRef]
- Akdis, C.A. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat. Rev. Immunol. 2021, 21, 739–751. [Google Scholar] [CrossRef]
- Akers, J.S.; Davis, T.N.; Gerow, S.; Avery, S. Decreasing motor stereotypy in individuals with autism spectrum disorder: A systematic review. Res. Autism Spectr. Disord. 2020, 77. [Google Scholar] [CrossRef]
- Albert, L.J.; Inman, R.D. Gram-negative pathogens and molecular mimicry: is there a case for mistaken identity? Trends Microbiol. 2000, 8, 444–445. [Google Scholar] [CrossRef]
- Albert, L.J.; Inman, R.D.; Epstein, F.H. Molecular Mimicry and Autoimmunity. New Engl. J. Med. 1999, 341, 2068–2074. [Google Scholar] [CrossRef]
- Allen, J.S. “Theory of food” as a neurocognitive adaptation. Am. J. Hum. Biol. 2012, 24, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.; Huang, B.S.; Notaras, M.J.; Lodhi, A.; Barrio-Alonso, E.; Lituma, P.J.; Wolujewicz, P.; Witztum, J.; Longo, F.; Chen, M.; et al. Astrocytes derived from ASD individuals alter behavior and destabilize neuronal activity through aberrant Ca2+ signaling. Mol. Psychiatry 2022, 27, 2470–2484. [Google Scholar] [CrossRef]
- Almahayni, O.; Hammond, L.; Hamasaki, H. Does the Wim Hof Method have a beneficial impact on physiological and psychological outcomes in healthy and non-healthy participants? A systematic review. PLOS ONE 2024, 19, e0286933. [Google Scholar] [CrossRef]
- Alsayouf, H.A. Growing evidence of pharmacotherapy effectiveness in managing attention-deficit/hyperactivity disorder in young children with or without autism spectrum disorder: a minireview. Front. Psychiatry 2024, 15, 1408876. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association, D. , & American Psychiatric Association, D. (2013). Diagnostic & statistical manual of mental disorders: DSM-5 (Vol. 5). American Psychiatric Association Washington, DC.
- American Psychiatric Association Staff, & Spitzer, R. L. (1980). Diagnostic & Statistical Manual of Mental Disorders (DSM-III). American Psychiatric Publishing.
- Amor, S.; Puentes, F.; Baker, D.; Van Der Valk, P. Inflammation in neurodegenerative diseases. Immunology 2010, 129, 154–169. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Liu, Y.; Wang, Y.; Fan, R.; Hu, X.; Zhang, F.; Yang, J.; Chen, J. The Role of Intestinal Mucosal Barrier in Autoimmune Disease: A Potential Target. Front. Immunol. 2022, 13, 871713. [Google Scholar] [CrossRef]
- Anderson, W.F.; Dien, B.S.; Brandon, S.K.; Peterson, J.D. Assessment of Bermudagrass and Bunch Grasses as Feedstock for Conversion to Ethanol. Appl. Biochem. Biotechnol. 2007, 145, 13–21. [Google Scholar] [CrossRef]
- Angajala, A.; Lim, S.; Phillips, J.B.; Kim, J.-H.; Yates, C.; You, Z.; Tan, M. Diverse Roles of Mitochondria in Immune Responses: Novel Insights Into Immuno-Metabolism. Front. Immunol. 2018, 9, 1605. [Google Scholar] [CrossRef]
- Angelidou, A.; Alysandratos, K.-D.; Asadi, S.; Zhang, B.; Francis, K.; Vasiadi, M.; Kalogeromitros, D.; Theoharides, T.C. Brief Report: “Allergic Symptoms” in Children with Autism Spectrum Disorders. More than Meets the Eye? J. Autism Dev. Disord. 2011, 41, 1579–1585. [Google Scholar] [CrossRef]
- Ansaldo, E.; Farley, T.K.; Belkaid, Y. Control of Immunity by the Microbiota. Annu. Rev. Immunol. 2021, 39, 449–479. [Google Scholar] [CrossRef]
- Ardalan, M.; Mallard, C. From hormones to behavior through microglial mitochondrial function. Brain, Behav. Immun. 2024, 117, 471–472. [Google Scholar] [CrossRef] [PubMed]
- Ashwood, P. , & Van de Water, J. (2004). A review of autism and the immune response. Clinical & Developmental Immunology, 11(2), 165. [CrossRef]
- Asperger, H. Die „Autistischen Psychopathen” im Kindesalter. Eur. Arch. Psychiatry Clin. Neurosci. 1944, 117, 76–136. [Google Scholar] [CrossRef]
- Azzalini, D.; Rebollo, I.; Tallon-Baudry, C. Visceral Signals Shape Brain Dynamics and Cognition. Trends Cogn. Sci. 2019, 23, 488–509. [Google Scholar] [CrossRef]
- Bach, J.-F. The hygiene hypothesis in autoimmunity: the role of pathogens and commensals. Nat. Rev. Immunol. 2017, 18, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G.; Padro, T. Systems biology approaches to understand the effects of nutrition and promote health. Br. J. Clin. Pharmacol. 2016, 83, 38–45. [Google Scholar] [CrossRef]
- Bahi, C.; Irrmischer, M.; Franken, K.; Fejer, G.; Schlenker, A.; Deijen, J.B.; Engelbregt, H. Effects of conscious connected breathing on cortical brain activity, mood and state of consciousness in healthy adults. Curr. Psychol. 2023, 43, 10578–10589. [Google Scholar] [CrossRef]
- Balcells, C.; Xu, Y.; Gil-Solsona, R.; Maitre, L.; Gago-Ferrero, P.; Keun, H.C. Blurred lines: Crossing the boundaries between the chemical exposome and the metabolome. Curr. Opin. Chem. Biol. 2023, 78, 102407. [Google Scholar] [CrossRef] [PubMed]
- Baniel, A.; Almagor, E.; Sharp, N.; Kolumbus, O.; Herbert, M.R. From fixing to connecting—developing mutual empathy guided through movement as a novel path for the discovery of better outcomes in autism. Front. Integr. Neurosci. 2025, 18, 1489345. [Google Scholar] [CrossRef]
- Baquero, F.; Coque, T.M.; Galán, J.C.; Martinez, J.L. The Origin of Niches and Species in the Bacterial World. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Barbaro, J.; Dissanayake, C. Autism Spectrum Disorders in Infancy and Toddlerhood: A Review of the Evidence on Early Signs, Early Identification Tools, and Early Diagnosis. J. Dev. Behav. Pediatr. 2009, 30, 447–459. [Google Scholar] [CrossRef]
- Barlattani, T. , D’Amelio, C., Cavatassi, A., De Luca, D., Di Stefano, R., Di Berardo, A., et al. (2023). Autism spectrum disorders and psychiatric comorbidities: a narrative review. Journal of Psychopathology. [CrossRef]
- Baron-Cohen, S. (2000). Theory of mind and autism: A review. In International Review of Research in Mental Retardation (Vol. 23, pp. 169–184). Academic Press. https://doi.org/10.1016/S0074-7750(00)80010-5.
- Baron-Cohen, S. The cognitive neuroscience of autism. J. Neurol. Neurosurg. Psychiatry 2004, 75, 945–948. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Ring, H.; Bullmore, E.; Wheelwright, S.; Ashwin, C.; Williams, S. The amygdala theory of autism. Neurosci. Biobehav. Rev. 2000, 24, 355–364. [Google Scholar] [CrossRef]
- Basheer, S.; Venkataswamy, M.M.; Christopher, R.; Van Amelsvoort, T.; Srinath, S.; Girimaji, S.C.; Ravi, V. Immune aberrations in children with Autism Spectrum Disorder: a case-control study from a tertiary care neuropsychiatric hospital in India. Psychoneuroendocrinology 2018, 94, 162–167. [Google Scholar] [CrossRef]
- Bauman, M.L. Medical Comorbidities in Autism: Challenges to Diagnosis and Treatment. Neurotherapeutics 2010, 7, 320–327. [Google Scholar] [CrossRef]
- Bauman, M.L.; Kemper, T.L. Neuroanatomic observations of the brain in autism: a review and future directions. Int. J. Dev. Neurosci. 2005, 23, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Baumann, N.; Pham-Dinh, D. Biology of Oligodendrocyte and Myelin in the Mammalian Central Nervous System. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef] [PubMed]
- Becker, E. B. E., & Stoodley, C. J. (2013). Chapter One - Autism Spectrum Disorder and the Cerebellum. In G. Konopka (Ed.), International Review of Neurobiology (Vol. 113, pp. 1–34). Academic Press. https://doi.org/10.1016/B978-0-12-418700-9.00001-0.
- Bei, R.; Masuelli, L.; Palumbo, C.; Modesti, M.; Modesti, A. A common repertoire of autoantibodies is shared by cancer and autoimmune disease patients: Inflammation in their induction and impact on tumor growth. Cancer Lett. 2009, 281, 8–23. [Google Scholar] [CrossRef]
- Bellot-Saez, A.; Cohen, G.; van Schaik, A.; Ooi, L.; Morley, J.W.; Buskila, Y. Astrocytic modulation of cortical oscillations. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Benakis, C.; Martin-Gallausiaux, C.; Trezzi, J.-P.; Melton, P.; Liesz, A.; Wilmes, P. The microbiome-gut-brain axis in acute and chronic brain diseases. Curr. Opin. Neurobiol. 2020, 61, 1–9. [Google Scholar] [CrossRef]
- Beopoulos, A.; Gea, M.; Fasano, A.; Iris, F. Autonomic Nervous System Neuroanatomical Alterations Could Provoke and Maintain Gastrointestinal Dysbiosis in Autism Spectrum Disorder (ASD): A Novel Microbiome–Host Interaction Mechanistic Hypothesis. Nutrients 2021, 14, 65. [Google Scholar] [CrossRef]
- Bernard, S.; Enayati, A.; Redwood, L.; Roger, H.; Binstock, T. Autism: a novel form of mercury poisoning. Med Hypotheses 2001, 56, 462–471. [Google Scholar] [CrossRef]
- Berthoud, H.-R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Bethell, C.D.; Kogan, M.D.; Strickland, B.B.; Schor, E.L.; Robertson, J.; Newacheck, P.W. A National and State Profile of Leading Health Problems and Health Care Quality for US Children: Key Insurance Disparities and Across-State Variations. Acad. Pediatr. 2011, 11, S22–S33. [Google Scholar] [CrossRef] [PubMed]
- Bhagavati, S. Autoimmune Disorders of the Nervous System: Pathophysiology, Clinical Features, and Therapy. Front. Neurol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Parr, T.; Ramstead, M.; Friston, K. Immunoceptive inference: why are psychiatric disorders and immune responses intertwined? Biol. Philos. 2021, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Bien, C.G.; Elger, C.E. Limbic encephalitis: A cause of temporal lobe epilepsy with onset in adult life. Epilepsy Behav. 2007, 10, 529–538. [Google Scholar] [CrossRef]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; et al. Oxidative Stress in Autism Spectrum Disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef]
- Bjørklund, G.; Mkhitaryan, M.; Sahakyan, E.; Fereshetyan, K.; A Meguid, N.; Hemimi, M.; Nashaat, N.H.; Yenkoyan, K. Linking Environmental Chemicals to Neuroinflammation and Autism Spectrum Disorder: Mechanisms and Implications for Prevention. Mol. Neurobiol. 2024, 61, 6328–6340. [Google Scholar] [CrossRef]
- E Blalock, J. The immune system as a sensory organ. J. Immunol. 1984, 132, 1067–1070. [Google Scholar] [CrossRef]
- Blaxill, M.F.; Redwood, L.; Bernard, S. Thimerosal and autism? A plausible hypothesis that should not be dismissed. Med Hypotheses 2004, 62, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Bleuler, E. (1911). Dementia praecox: oder Gruppe der Schizophrenien. F. Deuticke.
- Bleuler, E. (1951). Autistic thinking. In Organization & pathology of thought: Selected sources (pp. 399–437). Columbia University Press.
- Bojarskaite, L.; Bjørnstad, D.M.; Pettersen, K.H.; Cunen, C.; Hermansen, G.H.; Åbjørsbråten, K.S.; Chambers, A.R.; Sprengel, R.; Vervaeke, K.; Tang, W.; et al. Astrocytic Ca2+ signaling is reduced during sleep and is involved in the regulation of slow wave sleep. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Theis, K.R.; Waldor, M.K. Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. PLOS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [PubMed]
- Braunschweig, D.; Ashwood, P.; Krakowiak, P.; Hertzpicciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Vandewater, J.; Van de Water, J. Autism: Maternally derived antibodies specific for fetal brain proteins. NeuroToxicology 2007, 29, 226–231. [Google Scholar] [CrossRef]
- Breece, E.; Paciotti, B.; Nordahl, C.W.; Ozonoff, S.; Van de Water, J.A.; Rogers, S.J.; Amaral, D.; Ashwood, P. Myeloid dendritic cells frequencies are increased in children with autism spectrum disorder and associated with amygdala volume and repetitive behaviors. Brain, Behav. Immun. 2013, 31, 69–75. [Google Scholar] [CrossRef]
- Brincker, M.; Torres, E.B. Noise from the periphery in autism. Front. Integr. Neurosci. 2013, 7, 52681. [Google Scholar] [CrossRef]
- Brink, W.v.D.; van Bilsen, J.; Salic, K.; Hoevenaars, F.P.M.; Verschuren, L.; Kleemann, R.; Bouwman, J.; Ronnett, G.V.; van Ommen, B.; Wopereis, S. Current and Future Nutritional Strategies to Modulate Inflammatory Dynamics in Metabolic Disorders. Front. Nutr. 2019, 6, 129. [Google Scholar] [CrossRef]
- Brix, G.; Nekolla, E.A.; Nosske, D.; Griebel, J. Risks and safety aspects related to PET/MR examinations. Eur. J. Nucl. Med. 2008, 36, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Mehl-Madrona, L. Autoimmune and gastrointestinal dysfunctions: does a subset of children with autism reveal a broader connection? Expert Rev. Gastroenterol. Hepatol. 2011, 5, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Buie, T.; Campbell, D.B.; Fuchs, G.J., 3rd; Furuta, G.T.; Levy, J.; Vandewater, J.; Whitaker, A.H.; Atkins, D.; Bauman, M.L.; Beaudet, A.L.; et al. Evaluation, Diagnosis, and Treatment of Gastrointestinal Disorders in Individuals With ASDs: A Consensus Report. Pediatrics 2010, 125 (Suppl. 1), S1–S18. [Google Scholar] [CrossRef] [PubMed]
- Burnouf, T.; Walker, T.L. The multifaceted role of platelets in mediating brain function. Blood 2022, 140, 815–827. [Google Scholar] [CrossRef]
- Burns-Naas, L.A.; Dearman, R.J.; Germolec, D.R.; Kaminski, N.E.; Kimber, I.; Ladics, G.S.; Luebke, R.W.; Pfau, J.C.; Pruett, S.B. “Omics” Technologies and the Immune System. Toxicol. Mech. Methods 2006, 16, 101–119. [Google Scholar] [CrossRef]
- Buskila, Y.; Bellot-Saez, A.; Morley, J.W. Generating Brain Waves, the Power of Astrocytes. Front. Neurosci. 2019, 13, 1125. [Google Scholar] [CrossRef]
- Buzsáki, G.; Watson, B.O. Brain rhythms and neural syntax: implications for efficient coding of cognitive content and neuropsychiatric disease. Dialog- Clin. Neurosci. 2012, 14, 345–367. [Google Scholar] [CrossRef]
- Cagnin, A. In vivo imaging of neuroinflammation. Eur. Neuropsychopharmacol. 2002, 12, 581–586. [Google Scholar] [CrossRef]
- Cakir, J.; Frye, R.E.; Walker, S.J. The lifetime social cost of autism: 1990–2029. Res. Autism Spectr. Disord. 2020, 72. [Google Scholar] [CrossRef]
- Calçada, D.; Vianello, D.; Giampieri, E.; Sala, C.; Castellani, G.; de Graaf, A.; Kremer, B.; van Ommen, B.; Feskens, E.; Santoro, A.; et al. The role of low-grade inflammation and metabolic flexibility in aging and nutritional modulation thereof: A systems biology approach. Mech. Ageing Dev. 2014, 136-137, 138–147. [Google Scholar] [CrossRef]
- Cantando, I.; Centofanti, C.; D’aLessandro, G.; Limatola, C.; Bezzi, P. Metabolic dynamics in astrocytes and microglia during post-natal development and their implications for autism spectrum disorders. Front. Cell. Neurosci. 2024, 18, 1354259. [Google Scholar] [CrossRef]
- Carré, A.; Chevallier, C.; Robel, L.; Barry, C.; Maria, A.-S.; Pouga, L.; Philippe, A.; Pinabel, F.; Berthoz, S. Tracking Social Motivation Systems Deficits: The Affective Neuroscience View of Autism. J. Autism Dev. Disord. 2015, 45, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- CDC. (2021). Managing Chronic Health Conditions. CDC Healthy Schools. https://www.cdc.gov/healthyschools/chronicconditions.htm. 1 June.
- Cekici, H.; Sanlier, N. Current nutritional approaches in managing autism spectrum disorder: A review. Nutr. Neurosci. 2017, 22, 145–155. [Google Scholar] [CrossRef]
- Cellot, G.; Cherubini, E. GABAergic Signaling as Therapeutic Target for Autism Spectrum Disorders. Front. Pediatr. 2014, 2, 70. [Google Scholar] [CrossRef]
- Chambers, C., & Schaefer, C. (2015). 2.10 - Epilepsy and antiepileptic medications. In C. Schaefer, P. Peters, & R. K. Miller (Eds.), Drugs During Pregnancy and Lactation (Third Edition) (pp. 251–291). San Diego: Academic Press. https://doi.org/10.1016/B978-0-12-408078-2.00011-1.
- Chavez, J.A.; Zappaterra, M. Can Wim Hof Method breathing induce conscious metabolic waste clearance of the brain? Med Hypotheses 2023, 177. [Google Scholar] [CrossRef]
- Chen, C.-C.; Chiu, C.-H. Current and future applications of fecal microbiota transplantation for children. Biomed. J. 2021, 45, 11–18. [Google Scholar] [CrossRef]
- Chen, L.; Li, X.; Tjia, M.; Thapliyal, S. Homeostatic plasticity and excitation-inhibition balance: The good, the bad, and the ugly. Curr. Opin. Neurobiol. 2022, 75, 102553–102553. [Google Scholar] [CrossRef]
- Chia, S.L.; Kapoor, S.; Carvalho, C.; Bajénoff, M.; Gentek, R. Mast cell ontogeny: From fetal development to life-long health and disease. Immunol. Rev. 2023, 315, 31–53. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S.; Bjørklund, G. PERM Hypothesis: The Fundamental Machinery Able to Elucidate the Role of Xenobiotics and Hormesis in Cell Survival and Homeostasis. Int. J. Mol. Sci. 2017, 18, 165. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kim, J.H.; Yang, H.S.; Kim, J.Y.; Cortese, S.; Smith, L.; Koyanagi, A.; Dragioti, E.; Radua, J.; Fusar-Poli, P.; et al. Pharmacological and non-pharmacological interventions for irritability in autism spectrum disorder: a systematic review and meta-analysis with the GRADE assessment. Mol. Autism 2024, 15, 1–14. [Google Scholar] [CrossRef]
- Cieślińska, A.; Kostyra, E.; Savelkoul, H.F. ; Wageningen University & Research Treating Autism Spectrum Disorder with Gluten-Free and Casein-Free Diet: The Underlying Microbiota-Gut-Brain Axis Mechanisms. Clin. Immunolgy Immunother. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Clappison, E.; Hadjivassiliou, M.; Zis, P. Psychiatric Manifestations of Coeliac Disease, a Systematic Review and Meta-Analysis. Nutrients 2020, 12, 142. [Google Scholar] [CrossRef]
- Copf, T. Impairments in dendrite morphogenesis as etiology for neurodevelopmental disorders and implications for therapeutic treatments. Neurosci. Biobehav. Rev. 2016, 68, 946–978. [Google Scholar] [CrossRef]
- Costantini, M. Bodily self and immune self: is there a link? Front. Hum. Neurosci. 2014, 8, 138. [Google Scholar] [CrossRef]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial Community Variation in Human Body Habitats Across Space and Time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [PubMed]
- Coury, D.L.; Anagnostou, E.; Manning-Courtney, P.; Reynolds, A.; Cole, L.; McCoy, R.; Whitaker, A.; Perrin, J.M. Use of Psychotropic Medication in Children and Adolescents With Autism Spectrum Disorders. Pediatrics 2012, 130, S69–S76. [Google Scholar] [CrossRef]
- Crépeaux, G.; Authier, F.-J.; Exley, C.; Luján, L.; Gherardi, R.K. The role of aluminum adjuvants in vaccines raises issues that deserve independent, rigorous and honest science. J. Trace Elements Med. Biol. 2020, 62, 126632. [Google Scholar] [CrossRef]
- Cryan, J.F.; O'RIordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Csaba, G. Hormones in the immune system and their possible role. A critical review. Acta Microbiol. Et Immunol. Hung. 2014, 61, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Cutolo, M.; Capellino, S.; Sulli, A.; Serioli, B.; Secchi, M.E.; Villaggio, B.; Straub, R.H. Estrogens and Autoimmune Diseases. Ann. New York Acad. Sci. 2006, 1089, 538–547. [Google Scholar] [CrossRef]
- Cwik, J. C. (2021). Spiritual needs of people with autism spectrum disorder. In Spiritual Needs in Research and Practice: The Spiritual Needs Questionnaire as a Global Resource for Health and Social Care (pp. 265–280). Springer.
- D’adamo, C.R.; Nelson, J.L.; Miller, S.N.; Hong, M.R.; Lambert, E.; Ruhm, H.T. Reversal of Autism Symptoms among Dizygotic Twins through a Personalized Lifestyle and Environmental Modification Approach: A Case Report and Review of the Literature. J. Pers. Med. 2024, 14, 641. [Google Scholar] [CrossRef] [PubMed]
- Daëron, M. The immune system as a system of relations. Front. Immunol. 2022, 13, 984678. [Google Scholar] [CrossRef] [PubMed]
- D’aGostino, P.M.; Gottfried-Blackmore, A.; Anandasabapathy, N.; Bulloch, K. Brain dendritic cells: biology and pathology. Acta Neuropathol. 2012, 124, 599–614. [Google Scholar] [CrossRef]
- Daly, E., D. Tricklebank, M., & Wichers, R. (2019). Chapter Two - Neurodevelopmental roles and the serotonin hypothesis of autism spectrum disorder. In M. D. Tricklebank & E. Daly (Eds.), The Serotonin System (pp. 23–44). Academic Press. https://doi.org/10.1016/B978-0-12-813323-1.00002-5.
- Damasio, A. Mental self: The person within. Nature 2003, 423, 227–227. [Google Scholar] [CrossRef]
- D’Angelo, E. (2018). Physiology of the cerebellum. Handbook of Clinical Neurology, 154, 85–108. https://doi.org/10.1016/B978-0-444-63956-1.00006-0.
- Davenport, P.; Sola-Visner, M. Platelets in the neonate: Not just a small adult. Res. Pr. Thromb. Haemost. 2022, 6, e12719. [Google Scholar] [CrossRef]
- Davies, S.; Bishop, D.; Manstead, A.S.R.; Tantam, D. Face Perception in Children with Autism and Asperger's Syndrome. J. Child Psychol. Psychiatry 1994, 35, 1033–1057. [Google Scholar] [CrossRef]
- De Giorgio, R.; Guerrini, S.; Barbara, G.; Stanghellini, V.; De Ponti, F.; Corinaldesi, R.; Moses, P.L.; A Sharkey, K.; Mawe, G.M. Inflammatory neuropathies of the enteric nervous system☆. Gastroenterology 2004, 126, 1872–1883. [Google Scholar] [CrossRef]
- Del Casale, A.; Ferracuti, S.; Alcibiade, A.; Simone, S.; Modesti, M.N.; Pompili, M. Neuroanatomical correlates of autism spectrum disorders: A meta-analysis of structural magnetic resonance imaging (MRI) studies. Psychiatry Res. Neuroimaging 2022, 325, 111516. [Google Scholar] [CrossRef]
- Delafield-Butt, J.; Ciaunica, A. Sensorimotor foundations of self-consciousness in utero. Curr. Opin. Behav. Sci. 2024, 59. [Google Scholar] [CrossRef]
- Dempsey, J.L.; Little, M.; Cui, J.Y. Gut microbiome: An intermediary to neurotoxicity. NeuroToxicology 2019, 75, 41–69. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Yi, P.; Xiong, Y.; Ying, J.; Lin, Y.; Dong, Y.; Wei, G.; Wang, X.; Hua, F. Gut Metabolites Acting on the Gut-Brain Axis: Regulating the Functional State of Microglia. Aging Dis. 2024, 15, 480–502. [Google Scholar] [CrossRef]
- Denman, A.M. Sex hormones, autoimmune diseases, and immune responses. BMJ 1991, 303, 2–3. [Google Scholar] [CrossRef]
- Deth, R.C. Autism: A Redox/Methylation Disorder. Glob. Adv. Heal. Med. 2013, 2, 68–73. [Google Scholar] [CrossRef]
- Dethlefsen, L.; McFall-Ngai, M.; Relman, D.A. An ecological and evolutionary perspective on human–microbe mutualism and disease. Nature 2007, 449, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Di Liberto, D.; D’anneo, A.; Carlisi, D.; Emanuele, S.; De Blasio, A.; Calvaruso, G.; Giuliano, M.; Lauricella, M. Brain Opioid Activity and Oxidative Injury: Different Molecular Scenarios Connecting Celiac Disease and Autistic Spectrum Disorder. Brain Sci. 2020, 10, 437. [Google Scholar] [CrossRef]
- Dietz, P.M.; Rose, C.E.; McArthur, D.; Maenner, M. National and State Estimates of Adults with Autism Spectrum Disorder. J. Autism Dev. Disord. 2020, 50, 4258–4266. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Ding, S.; A Rando, T.; Trounson, A. Translational strategies and challenges in regenerative medicine. Nat. Med. 2014, 20, 814–821. [Google Scholar] [CrossRef]
- D'MEllo, A.M.; Stoodley, C.J. Cerebro-cerebellar circuits in autism spectrum disorder. Front. Neurosci. 2015, 9, 408. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.; Foley, K.-R.; Schloss, J. Complementary and Alternative Medicine for Autism – A Systematic Review. J. Autism Dev. Disord. 2024, 1–11. [Google Scholar] [CrossRef]
- Donald, K.; Finlay, B.B. Early-life interactions between the microbiota and immune system: impact on immune system development and atopic disease. Nat. Rev. Immunol. 2023, 23, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Donovan, A.P.A.; Basson, M.A. The neuroanatomy of autism – a developmental perspective. Am. J. Anat. 2016, 230, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Dukhinova, M.; Kuznetsova, I.; Kopeikina, E.; Veniaminova, E.; Yung, A.W.; Veremeyko, T.; Levchuk, K.; Barteneva, N.S.; Wing-Ho, K.K.; Yung, W.-H.; et al. Platelets mediate protective neuroinflammation and promote neuronal plasticity at the site of neuronal injury. Brain, Behav. Immun. 2018, 74, 7–27. [Google Scholar] [CrossRef]
- Meimand, S.E.; Rostam-Abadi, Y.; Rezaei, N. Autism spectrum disorders and natural killer cells: a review on pathogenesis and treatment. Expert Rev. Clin. Immunol. 2020, 17, 27–35. [Google Scholar] [CrossRef]
- Ecker, C. The neuroanatomy of autism spectrum disorder: An overview of structural neuroimaging findings and their translatability to the clinical setting. Autism 2016, 21, 18–28. [Google Scholar] [CrossRef]
- Edmiston, E.; Ashwood, P.; Van de Water, J. Autoimmunity, Autoantibodies, and Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 383–390. [Google Scholar] [CrossRef]
- El-Ansary, A.; Bhat, R.S.; Zayed, N. Gut Microbiome and Sex Bias in Autism Spectrum Disorders. Curr. Behav. Neurosci. Rep. 2020, 7, 22–31. [Google Scholar] [CrossRef]
- Enstrom, A.M.; Lit, L.; Onore, C.E.; Gregg, J.P.; Hansen, R.L.; Pessah, I.N.; Hertz-Picciotto, I.; Van de Water, J.A.; Sharp, F.R.; Ashwood, P. Altered gene expression and function of peripheral blood natural killer cells in children with autism. Brain, Behav. Immun. 2008, 23, 124–133. [Google Scholar] [CrossRef]
- Epel, E.S.; McEwen, B.S.; Ickovics, J.R. Embodying Psychological Thriving: Physical Thriving in Response to Stress. J. Soc. Issues 1998, 54, 301–322. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef]
- Exley, C. Human exposure to aluminium. Environ. Sci. Process. Impacts 2013, 15, 1807–1816. [Google Scholar] [CrossRef]
- Exley, C.; Clarkson, E. Aluminium in human brain tissue from donors without neurodegenerative disease: A comparison with Alzheimer’s disease, multiple sclerosis and autism. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Famitafreshi, H.; Karimian, M. Overview of the Recent Advances in Pathophysiology and Treatment for Autism. CNS Neurol. Disord. - Drug Targets 2018, 17, 590–594. [Google Scholar] [CrossRef]
- Fan, G.; Ma, J.; Ma, R.; Suo, M.; Chen, Y.; Zhang, S.; Zeng, Y.; Chen, Y. Microglia Modulate Neurodevelopment in Autism Spectrum Disorder and Schizophrenia. Int. J. Mol. Sci. 2023, 24, 17297. [Google Scholar] [CrossRef]
- Farmer, C.A.; Thurm, A.E.; Honnekeri, B.; Kim, P.; Swedo, S.E.; Han, J.C. The contribution of platelets to peripheral BDNF elevation in children with autism spectrum disorder. Sci. Rep. 2021, 11, 1–6. [Google Scholar] [CrossRef]
- Fasano, A. Leaky Gut and Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2011, 42, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, S.H.; Halt, A.R.; Realmuto, G.; Earle, J.; Kist, D.A.; Thuras, P.; Merz, A. Purkinje Cell Size Is Reduced in Cerebellum of Patients with Autism. Cell. Mol. Neurobiol. 2002, 22, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Ferri, S.L.; Abel, T.; Brodkin, E.S. Sex Differences in Autism Spectrum Disorder: a Review. Curr. Psychiatry Rep. 2018, 20, 1–17. [Google Scholar] [CrossRef]
- Filiano, A.J.; Xu, Y.; Tustison, N.J.; Marsh, R.L.; Baker, W.; Smirnov, I.; Overall, C.C.; Gadani, S.P.; Turner, S.D.; Weng, Z.; et al. Unexpected role of interferon-γ in regulating neuronal connectivity and social behaviour. Nature 2016, 535, 425–429. [Google Scholar] [CrossRef]
- Filipek, P.A.; Juranek, J.; Nguyen, M.T.; Cummings, C.; Gargus, J.J. Relative Carnitine Deficiency in Autism. J. Autism Dev. Disord. 2004, 34, 615–623. [Google Scholar] [CrossRef]
- Fincham, G.W.; Strauss, C.; Montero-Marin, J.; Cavanagh, K. Effect of breathwork on stress and mental health: A meta-analysis of randomised-controlled trials. Sci. Rep. 2023, 13, 1–14. [Google Scholar] [CrossRef]
- Fleming, D. Walter B. Cannon and homeostasis.. 1984, 51, 609–40. [Google Scholar]
- Folstein, S.; Rutter, M. Genetic influences and infantile autism. Nature 1977, 265, 726–728. [Google Scholar] [CrossRef]
- Forbes, J.D.; Van Domselaar, G.; Bernstein, C.N. The Gut Microbiota in Immune-Mediated Inflammatory Diseases. Front. Microbiol. 2016, 7, 1081. [Google Scholar] [CrossRef] [PubMed]
- Fraguas, D.; Díaz-Caneja, C.M.; Pina-Camacho, L.; Moreno, C.; Durán-Cutilla, M.; Ayora, M.; González-Vioque, E.; de Matteis, M.; Hendren, R.L.; Arango, C.; et al. Dietary Interventions for Autism Spectrum Disorder: A Meta-analysis. Pediatrics 2019, 144, e20183218. [Google Scholar] [CrossRef] [PubMed]
- Frith, U. Autism and theory of mind in everyday life. Soc. Dev. 1994, 3, 108–124. [Google Scholar] [CrossRef]
- Frith, U.; Happé, F. Autism: beyond “theory of mind”. Cognition 1994, 50, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Frith, U.; Happé, F. Theory of Mind and Self-Consciousness: What Is It Like to Be Autistic? Mind Lang. 1999, 14, 82–89. [Google Scholar] [CrossRef]
- Frye, R.E.; Rincon, N.; McCarty, P.J.; Brister, D.; Scheck, A.C.; Rossignol, D.A. Biomarkers of mitochondrial dysfunction in autism spectrum disorder: A systematic review and meta-analysis. Neurobiol. Dis. 2024, 197, 106520. [Google Scholar] [CrossRef]
- Fu, J.; Schroder, K.; Wu, H. Mechanistic insights from inflammasome structures. Nat. Rev. Immunol. 2024, 24, 518–535. [Google Scholar] [CrossRef]
- Fuller, R.; Rahona, E.; Fisher, S.; Caravanos, J.; Webb, D.; Kass, D.; Matte, T.; Landrigan, P.J. Pollution and non-communicable disease: time to end the neglect. Lancet Planet. Heal. 2018, 2, e96–e98. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The Enteric Nervous System and Gastrointestinal Innervation: Integrated Local and Central Control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin, Germany, 2014; Volume 817, pp. 39–71. [Google Scholar] [CrossRef]
- Gable, M.S.; Sheriff, H.; Dalmau, J.; Tilley, D.H.; Glaser, C.A. The Frequency of Autoimmune N-Methyl-D-Aspartate Receptor Encephalitis Surpasses That of Individual Viral Etiologies in Young Individuals Enrolled in the California Encephalitis Project. Clin. Infect. Dis. 2012, 54, 899–904. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Persico, A.M. Blood serotonin levels in autism spectrum disorder: A systematic review and meta-analysis. Eur. Neuropsychopharmacol. 2014, 24, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Gallotto, S.; Sack, A.T.; Schuhmann, T.; de Graaf, T.A. Oscillatory Correlates of Visual Consciousness. Front. Psychol. 2017, 8, 1147–1147. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Contreras, A.Y.; Zarate-Lopez, D.; Torres-Chavez, A.L.; Gonzalez-Perez, O. Role of Oligodendrocytes and Myelin in the Pathophysiology of Autism Spectrum Disorder. Brain Sci. 2020, 10, 951. [Google Scholar] [CrossRef]
- Garg, D.; Mohammad, S.S.; Sharma, S. Autoimmune Encephalitis in Children: An Update. Indian Pediatr. 2020, 57, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Gaugler, T.; Klei, L.; Sanders, S.J.; A Bodea, C.; Goldberg, A.P.; Lee, A.B.; Mahajan, M.; Manaa, D.; Pawitan, Y.; Reichert, J.; et al. Most genetic risk for autism resides with common variation. Nat. Genet. 2014, 46, 881–885. [Google Scholar] [CrossRef]
- D. A., G.; J.K., K.; M.R., G. The biological basis of autism spectrum disorders: Understanding causation and treatment by clinical geneticists. Acta Neurobiol. Exp. 2010, 70, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Geier, D.A.; King, P.G.; Hooker, B.S.; Dórea, J.G.; Kern, J.K.; Sykes, L.K.; Geier, M.R. Thimerosal: Clinical, epidemiologic and biochemical studies. Clin. Chim. Acta 2015, 444, 212–220. [Google Scholar] [CrossRef]
- Geng, Z.-H.; Zhu, Y.; Li, Q.-L.; Zhao, C.; Zhou, P.-H. Enteric Nervous System: The Bridge Between the Gut Microbiota and Neurological Disorders. Front. Aging Neurosci. 2022, 14, 810483. [Google Scholar] [CrossRef]
- Genuis, S.J. What's Out There Making Us Sick? J. Environ. Public Heal. 2011, 2012, 1–10. [Google Scholar] [CrossRef]
- Gershon, M.D. The Enteric Nervous System: A Second Brain. Hosp. Pr. 1999, 34, 31–52. [Google Scholar] [CrossRef]
- Gershon, M. D. (2022). The Shaggy Dog Story of Enteric Signaling: Serotonin, a Molecular Megillah. In N. J. Spencer, M. Costa, & S. M. Brierley (Eds.), The Enteric Nervous System II (pp. 307–318). Cham: Springer International Publishing.
- Ghanizadeh, A.; Akhondzadeh, S.; Hormozi, M.; Makarem, A.; Abotorabi-Zarchi, M.; Firoozabadi, A. Glutathione-Related Factors and Oxidative Stress in Autism, A Review. Curr. Med. Chem. 2012, 19, 4000–4005. [Google Scholar] [CrossRef] [PubMed]
- Ghaziuddin, M.; Al-Owain, M. Autism Spectrum Disorders and Inborn Errors of Metabolism: An Update. Pediatr. Neurol. 2013, 49, 232–236. [Google Scholar] [CrossRef]
- Ghosh, A.; Michalon, A.; Lindemann, L.; Fontoura, P.; Santarelli, L. Drug discovery for autism spectrum disorder: challenges and opportunities. Nat. Rev. Drug Discov. 2013, 12, 777–790. [Google Scholar] [CrossRef]
- Gianchecchi, E.; Delfino, D.V.; Fierabracci, A. NK cells in autoimmune diseases: Linking innate and adaptive immune responses. Autoimmun. Rev. 2018, 17, 142–154. [Google Scholar] [CrossRef]
- Giron, M.C.; Mazzi, U. Molecular imaging of microbiota-gut-brain axis: searching for the right targeted probe for the right target and disease. Nucl. Med. Biol. 2021, 92, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Gładysz, D.; Krzywdzińska, A.; Hozyasz, K.K. Immune Abnormalities in Autism Spectrum Disorder—Could They Hold Promise for Causative Treatment? Mol. Neurobiol. 2018, 55, 6387–6435. [Google Scholar] [CrossRef] [PubMed]
- Gobin, V.; Van Steendam, K.; Denys, D.; Deforce, D. Selective serotonin reuptake inhibitors as a novel class of immunosuppressants. Int. Immunopharmacol. 2014, 20, 148–156. [Google Scholar] [CrossRef]
- Goldberg, W.A.; Osann, K.; Filipek, P.A.; Laulhere, T.; Jarvis, K.; Modahl, C.; Flodman, P.; Spence, M.A. Language and Other Regression: Assessment and Timing. J. Autism Dev. Disord. 2003, 33, 607–616. [Google Scholar] [CrossRef]
- Goldman, A.W.; Burmeister, Y.; Cesnulevicius, K.; Herbert, M.; Kane, M.; Lescheid, D.; McCaffrey, T.; Schultz, M.; Seilheimer, B.; Smit, A.; et al. Bioregulatory systems medicine: an innovative approach to integrating the science of molecular networks, inflammation, and systems biology with the patient's autoregulatory capacity? Front. Physiol. 2015, 6, 225. [Google Scholar] [CrossRef] [PubMed]
- Goubau, C.; Buyse, G.M.; Van Geet, C.; Freson, K. The contribution of platelet studies to the understanding of disease mechanisms in complex and monogenetic neurological disorders. Dev. Med. Child Neurol. 2014, 56, 724–731. [Google Scholar] [CrossRef]
- Graeber, M.B.; Streit, W.J. Microglia: biology and pathology. Acta Neuropathol. 2009, 119, 89–105. [Google Scholar] [CrossRef]
- Graeber, M.B.; Stre'RT, W.J. Microglia: Immune Network in the CNS. Brain Pathol. 1990, 1, 2–5. [Google Scholar] [CrossRef]
- Green, D.J.; Park, K.; Bhatt-Mehta, V.; Snyder, D.; Burckart, G.J. Regulatory Considerations for the Mother, Fetus and Neonate in Fetal Pharmacology Modeling. Front. Pediatr. 2021, 9. [Google Scholar] [CrossRef]
- Greenberg, G. , & Tobach, E. (2014). Behavioral Evolution & Integrative Levels: The T.C. Schneirla Conferences Series, Volume 1. Taylor & Francis.
- Gropman, A., & Sadle, C. J. (2024). Epigenetics of autism spectrum disorder. In Neuropsychiatric Disorders & Epigenetics (pp. 81–102). Elsevier. https://doi.org/10.1016/B978-0-443-18516-8.00017-X.
- Guan, A.; Wang, S.; Huang, A.; Qiu, C.; Li, Y.; Li, X.; Wang, J.; Wang, Q.; Deng, B. The role of gamma oscillations in central nervous system diseases: Mechanism and treatment. Front. Cell. Neurosci. 2022, 16, 962957. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gupta, M. Off-label psychopharmacological interventions for autism spectrum disorders: strategic pathways for clinicians. CNS Spectrums 2023, 29, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Gzielo, K.; Nikiforuk, A. Astroglia in Autism Spectrum Disorder. Int. J. Mol. Sci. 2021, 22, 11544. [Google Scholar] [CrossRef]
- Hacohen, Y.; Wright, S.; Gadian, J.; Vincent, A.; Lim, M.; Wassmer, E.; Lin, J. N-methyl-d-aspartate (NMDA) receptor antibodies encephalitis mimicking an autistic regression. Dev. Med. Child Neurol. 2016, 58, 1092–1094. [Google Scholar] [CrossRef]
- Hafizi, S.; Tabatabaei, D.; Lai, M.-C. Review of Clinical Studies Targeting Inflammatory Pathways for Individuals With Autism. Front. Psychiatry 2019, 10, 849. [Google Scholar] [CrossRef]
- Hallmayer, J.; Cleveland, S.; Torres, A.; Phillips, J.; Cohen, B.; Torigoe, T.; Miller, J.; Fedele, A.; Collins, J.; Smith, K.; et al. Genetic Heritability and Shared Environmental Factors Among Twin Pairs With Autism. Arch. Gen. Psychiatry 2011, 68, 1095–1102. [Google Scholar] [CrossRef]
- Hameroff, S. The “conscious pilot”—dendritic synchrony moves through the brain to mediate consciousness. J. Biol. Phys. 2009, 36, 71–93. [Google Scholar] [CrossRef]
- Hampe, C.S.; Mitoma, H. A Breakdown of Immune Tolerance in the Cerebellum. Brain Sci. 2022, 12, 328. [Google Scholar] [CrossRef]
- Han, A.; Peng, T.; Xie, Y.; Zhang, W.; Sun, W.; Xie, Y.; Ma, Y.; Wang, C.; Xie, N. Mitochondrial-regulated Tregs: potential therapeutic targets for autoimmune diseases of the central nervous system. Front. Immunol. 2023, 14, 1301074. [Google Scholar] [CrossRef]
- Hanaford, A.; Johnson, S.C. The immune system as a driver of mitochondrial disease pathogenesis: a review of evidence. Orphanet J. Rare Dis. 2022, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Happé, F.G.E. Wechsler IQ Profile and Theory of Mind in Autism: A Research Note. J. Child Psychol. Psychiatry 1994, 35, 1461–1471. [Google Scholar] [CrossRef]
- Hardy, D. Autoimmune Encephalitis in Children. Pediatr. Neurol. 2022, 132, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Harkey, J. The Epidemiology of Selected Chronic Childhood Health Conditions. Child. Heal. Care 1983, 12, 62–71. [Google Scholar] [CrossRef]
- Harris, J. Leo Kanner and autism: a 75-year perspective. Int. Rev. Psychiatry 2018, 30, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef]
- Al Hasib, R.; Ali, C.; Rahman, H.; Ahmed, S.; Sultana, S.; Summa, S.Z.; Shimu, M.S.S.; Afrin, Z.; Jamal, M.A.H.M. Integrated gene expression profiling and functional enrichment analyses to discover biomarkers and pathways associated with Guillain-Barré syndrome and autism spectrum disorder to identify new therapeutic targets. J. Biomol. Struct. Dyn. 2023, 42, 11299–11321. [Google Scholar] [CrossRef] [PubMed]
- Hawgood, S.; Hook-Barnard, I.G.; O’bRien, T.C.; Yamamoto, K.R. Precision medicine: Beyond the inflection point. Sci. Transl. Med. 2015, 7, 300ps17–300ps17. [Google Scholar] [CrossRef]
- Hazan, S.; Spradling-Reeves, K.D.; Papoutsis, A.; Walker, S.J. Shotgun Metagenomic Sequencing Identifies Dysbiosis in Triplet Sibling with Gastrointestinal Symptoms and ASD. Children 2020, 7, 255. [Google Scholar] [CrossRef]
- Heberling, C.A.; Dhurjati, P.S.; Sasser, M. Hypothesis for a systems connectivity model of autism spectrum disorder pathogenesis: Links to gut bacteria, oxidative stress, and intestinal permeability. Med Hypotheses 2013, 80, 264–270. [Google Scholar] [CrossRef]
- Heemskerk, V.H.; Daemen, M.A.; A Buurman, W. Insulin-like growth factor-1 (IGF-1) and growth hormone (GH) in immunity and inflammation. Cytokine Growth Factor Rev. 1999, 10, 5–14. [Google Scholar] [CrossRef]
- Heiss, C.N.; Olofsson, L.E. The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocr. 2019, 31, e12684. [Google Scholar] [CrossRef]
- Helt, M.; Kelley, E.; Kinsbourne, M.; Pandey, J.; Boorstein, H.; Herbert, M.; Fein, D. Can Children with Autism Recover? If So, How? Neuropsychol. Rev. 2008, 18, 339–366. [Google Scholar] [CrossRef]
- Hendriksen, E.; van Bergeijk, D.; Oosting, R.S.; Redegeld, F.A. Mast cells in neuroinflammation and brain disorders. Neurosci. Biobehav. Rev. 2017, 79, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: the role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Herberman, R.B.; Ortaldo, J.R. Natural killer cells: their roles in defenses against disease. Science 1981, 214, 24–30. [Google Scholar] [CrossRef]
- Herbert, M. , & Weintraub, K. (2013). The Autism Revolution: Whole-Body Strategies for Making Life All It Can Be. Random House Publishing Group.
- Herbert, M.R. Large Brains in Autism: The Challenge of Pervasive Abnormality. Neurosci. 2005, 11, 417–440. [Google Scholar] [CrossRef]
- Herbert, M. S. 01.03 Autism: from static genetic brain defect to dynamic gene-environment modulated pathophysiology. Eur. Neuropsychopharmacol. 2013, 23. [Google Scholar] [CrossRef]
- Herbert, M. R. (2014). Translational implications of a whole-body approach to brain health in autism: how transduction between metabolism and electrophysiology points to mechanisms for neuroplasticity. In Frontiers in Autism Research: New Horizons for Diagnosis & Treatment, 515–556. http://dx.doi.org/10.1142/9789814602167_0021.
- Herbert, M.R.; Sage, C. Autism and EMF? Plausibility of a pathophysiological link – Part I. Pathophysiology 2013, 20, 191–209. [Google Scholar] [CrossRef]
- Herbert, M.R.; Sage, C. Autism and EMF? Plausibility of a pathophysiological link part II. Pathophysiology 2013, 20, 211–234. [Google Scholar] [CrossRef]
- Herbert, M.R.; Ziegler, D.A.; Deutsch, C.K.; O'Brien, L.M.; Kennedy, D.N.; Filipek, P.A.; Bakardjiev, A.I.; Hodgson, J.; Takeoka, M.; Makris, N.; et al. Brain asymmetries in autism and developmental language disorder: a nested whole-brain analysis. Brain 2004, 128, 213–226. [Google Scholar] [CrossRef]
- Herbert, M.R.; Ziegler, D.A.; Deutsch, C.K.; O'BRien, L.M.; Lange, N.; Bakardjiev, A.; Hodgson, J.; Adrien, K.T.; Steele, S.; Makris, N.; et al. Dissociations of cerebral cortex, subcortical and cerebral white matter volumes in autistic boys. Brain 2003, 126, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.R.; Ziegler, D.A.; Makris, N.; Filipek, P.A.; Kemper, T.L.; Normandin, J.J.; Sanders, H.A.; Kennedy, D.N.; Caviness, V.S. Localization of white matter volume increase in autism and developmental language disorder. Ann. Neurol. 2004, 55, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Hertz-Picciotto, I.; Schmidt, R.J.; Krakowiak, P. Understanding environmental contributions to autism: Causal concepts and the state of science. Autism Res. 2018, 11, 554–586. [Google Scholar] [CrossRef]
- Hickey, A.; Crabtree, J.; Stott, J. ‘Suddenly the first fifty years of my life made sense’: Experiences of older people with autism. Autism 2017, 22, 357–367. [Google Scholar] [CrossRef]
- Higdon, R.; Earl, R.K.; Stanberry, L.; Hudac, C.M.; Montague, E.; Stewart, E.; Janko, I.; Choiniere, J.; Broomall, W.; Kolker, N.; et al. The Promise of Multi-Omics and Clinical Data Integration to Identify and Target Personalized Healthcare Approaches in Autism Spectrum Disorders. OMICS: A J. Integr. Biol. 2015, 19, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.L. Executive dysfunction in autism☆. Trends Cogn. Sci. 2004, 8, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.L. Evaluating the theory of executive dysfunction in autism. Dev. Rev. 2004, 24, 189–233. [Google Scholar] [CrossRef]
- Hill, K. (1990). The decline of childhood mortality.
- Hiller-Sturmhöfel, S. , & Bartke, A. (1998). The endocrine system - An overview. Alcohol health & Research World, 22, 153–64. https://pubmed.ncbi.nlm.nih. 1570. [Google Scholar]
- Hirahara, K.; Nakayama, T. CD4 + T-cell subsets in inflammatory diseases: beyond the T h 1/T h 2 paradigm. Int. Immunol. 2016, 28, 163–171. [Google Scholar] [CrossRef]
- Ho, P.; Ross, D.A. More Than a Gut Feeling: The Implications of the Gut Microbiota in Psychiatry. Biol. Psychiatry 2017, 81, e35–e37. [Google Scholar] [CrossRef]
- Hooker, B.; Kern, J.; Geier, D.; Haley, B.; Sykes, L.; King, P.; Geier, M. Methodological Issues and Evidence of Malfeasance in Research Purporting to Show Thimerosal in Vaccines Is Safe. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Horlin, C.; Falkmer, M.; Parsons, R.; Albrecht, M.A.; Falkmer, T.; Mulle, J.G. The Cost of Autism Spectrum Disorders. PLOS ONE 2014, 9, e106552. [Google Scholar] [CrossRef] [PubMed]
- Hridi, S.U.; Barbour, M.; Wilson, C.; Franssen, A.J.; Harte, T.; Bushell, T.J.; Jiang, H.-R. Increased Levels of IL-16 in the Central Nervous System during Neuroinflammation Are Associated with Infiltrating Immune Cells and Resident Glial Cells. Biology 2021, 10, 472. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Chow, J.; Mazmanian, S.K.; Patterson, P.H. Modeling an autism risk factor in mice leads to permanent immune dysregulation. Proc. Natl. Acad. Sci. 2012, 109, 12776–12781. [Google Scholar] [CrossRef]
- Huang, J.; Li, X.; Chen, Z.; Zou, L.; Healy, S.; Tse, C.Y.A.; Li, C. Effects of Mind-Body Exercises on Health-related Outcomes in Children and Adolescents with Autism Spectrum Disorder: A Systematic Review. Rev. J. Autism Dev. Disord. 2023, 1–15. [Google Scholar] [CrossRef]
- Hughes, H.K.; Ko, E.M.; Rose, D.; Ashwood, P. Immune Dysfunction and Autoimmunity as Pathological Mechanisms in Autism Spectrum Disorders. Front. Cell. Neurosci. 2018, 12, 405. [Google Scholar] [CrossRef]
- Hughes, H.K.; Rose, D.; Ashwood, P. The Gut Microbiota and Dysbiosis in Autism Spectrum Disorders. Curr. Neurol. Neurosci. Rep. 2018, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Hulme, S.R.; Jones, O.D.; Raymond, C.R.; Sah, P.; Abraham, W.C. Mechanisms of heterosynaptic metaplasticity. Philos. Trans. R. Soc. B: Biol. Sci. 2014, 369, 20130148. [Google Scholar] [CrossRef]
- Humphries S, Bystrianyk R (2013) Dissolving illusions: Disease, vaccines and the forgotten history. CreateSpace Independent Publishing Platform.
- Hurley-Hanson, A. E., Giannantonio, C. M., Griffiths, A. J., Hurley-Hanson, A. E., Giannantonio, C. M., & Griffiths, A. J. (2020). The costs of autism. Springer. https://doi.org/10.1007/978-3-030-29049-8_3.
- Hussaini, S.M.Q.; Jang, M.H. New Roles for Old Glue: Astrocyte Function in Synaptic Plasticity and Neurological Disorders. Int. Neurourol. J. 2018, 22, S106–114. [Google Scholar] [CrossRef] [PubMed]
- E Ichim, T.; Solano, F.; Glenn, E.; Morales, F.; Smith, L.; Zabrecky, G.; Riordan, N.H. Stem Cell Therapy for Autism. J. Transl. Med. 2007, 5, 30–30. [Google Scholar] [CrossRef]
- Irwin, J. K. , MacSween, J., & Kerns, K. A. (2011). History and evolution of the autism spectrum disorders. In International Handbook of Autism & Pervasive Developmental disorders (pp. 3–16). Springer.
- Jaga, K.; Dharmani, C. The interrelation between organophosphate toxicity and the epidemiology of depression and suicide. Rev. Environ. Heal. 2007, 22, 57–74. [Google Scholar] [CrossRef]
- Jarrold, C.; Russell, J. Counting Abilities in Autism: Possible Implications for Central Coherence Theory. J. Autism Dev. Disord. 1997, 27, 25–37. [Google Scholar] [CrossRef]
- Jaswal, V.K.; Wayne, A.; Golino, H. Eye-tracking reveals agency in assisted autistic communication. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Jaswal, V.K.; Lampi, A.J.; Stockwell, K.M. Literacy in nonspeaking autistic people. Autism 2024, 28, 2503–2514. [Google Scholar] [CrossRef]
- Jenne, C.N.; Urrutia, R.; Kubes, P. Platelets: bridging hemostasis, inflammation, and immunity. Int. J. Lab. Hematol. 2013, 35, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.M.; Cowan, M.; Moonah, S.N.; Petri, W.A. The Impact of Systemic Inflammation on Neurodevelopment. Trends Mol. Med. 2018, 24, 794–804. [Google Scholar] [CrossRef]
- John, E. The neurophysics of consciousness. Brain Res. Rev. 2002, 39, 1–28. [Google Scholar] [CrossRef]
- Jones, J.P.; Williamson, L.; Konsoula, Z.; Anderson, R.; Reissner, K.J.; Parker, W. Evaluating the Role of Susceptibility Inducing Cofactors and of Acetaminophen in the Etiology of Autism Spectrum Disorder. Life 2024, 14, 918. [Google Scholar] [CrossRef]
- Jung, C.; Hugot, J.-P.; Barreau, F. Peyer's Patches: The Immune Sensors of the Intestine. Int. J. Inflamm. 2010, 2010, 823710. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H. Food allergy and autism spectrum disorders: Is there a link? Curr. Allergy Asthma Rep. 2009, 9, 194–201. [Google Scholar] [CrossRef]
- Jyonouchi, H. Autism spectrum disorder and a possible role of anti-inflammatory treatments: experience in the pediatric allergy/immunology clinic. Front. Psychiatry 2024, 15, 1333717. [Google Scholar] [CrossRef]
- Kaminska, A.; Cheliout-Heraut, F.; Eisermann, M.; de Villepin, A.T.; Lamblin, M.D. EEG in children, in the laboratory or at the patient's bedside. Neurophysiol. Clin. 2015, 45, 65–74. [Google Scholar] [CrossRef]
- Kang, D.-W.; Adams, J.B.; Coleman, D.M.; Pollard, E.L.; Maldonado, J.; McDonough-Means, S.; Caporaso, J.G.; Krajmalnik-Brown, R. Long-term benefit of Microbiota Transfer Therapy on autism symptoms and gut microbiota. Sci. Rep. 2019, 9, 5821. [Google Scholar] [CrossRef]
- Kanner, L. Autistic disturbances of affective contact. Nerv. Child 1943, 2, 217–250. [Google Scholar]
- Karhu, E.; Zukerman, R.; Eshraghi, R.S.; Mittal, J.; Deth, R.C.; Castejon, A.M.; Trivedi, M.; Mittal, R.; A Eshraghi, A. Nutritional interventions for autism spectrum disorder. Nutr. Rev. 2019, 78, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018, 9, 2013. [Google Scholar] [CrossRef]
- Karlsson, O. Chemical safety and the exposome. Emerg. Contam. 2023, 9. [Google Scholar] [CrossRef]
- Kaur, I.; Behl, T.; Aleya, L.; Rahman, H.; Kumar, A.; Arora, S.; Akter, R. Role of metallic pollutants in neurodegeneration: effects of aluminum, lead, mercury, and arsenic in mediating brain impairment events and autism spectrum disorder. Environ. Sci. Pollut. Res. 2021, 28, 8989–9001. [Google Scholar] [CrossRef]
- Kayser, M.S.; Dalmau, J. Anti-NMDA receptor encephalitis, autoimmunity, and psychosis. Schizophr. Res. 2016, 176, 36–40. [Google Scholar] [CrossRef]
- Keating, B. A., Lees, J. G., & Moalem-Taylor, G. (2019). The Roles of Regulatory T Cells in Central Nervous System Autoimmunity. In H. Mitoma & M. Manto (Eds.), Neuroimmune Diseases: From Cells to the Living Brain (pp. 167–193). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-030-19515-1_6.
- Keller, A.; Rimestad, M.L.; Rohde, J.F.; Petersen, B.H.; Korfitsen, C.B.; Tarp, S.; Lauritsen, M.B.; Händel, M.N. The Effect of a Combined Gluten- and Casein-Free Diet on Children and Adolescents with Autism Spectrum Disorders: A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 470. [Google Scholar] [CrossRef]
- Kelly, L.S.; Apple, C.G.; Gharaibeh, R.; Pons, E.E.B.; Thompson, C.W.B.; Kannan, K.B.; Darden, D.B.; Efron, P.A.; Thomas, R.M.; Mohr, A.M. Stress-related changes in the gut microbiome after trauma. J. Trauma Acute Care Surg. 2021, 91, 192–199. [Google Scholar] [CrossRef]
- Kemper, T.L.; Bauman, M. Neuropathology of Infantile Autism. J. Neuropathol. Exp. Neurol. 1998, 57, 645–652. [Google Scholar] [CrossRef]
- Kepser, L.-J.; Homberg, J.R. The neurodevelopmental effects of serotonin: A behavioural perspective. Behav. Brain Res. 2015, 277, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.; Geier, D.; Audhya, T.; King, P.; Sykes, L.; Geier, M. Evidence of parallels between mercury intoxication and the brain pathology in autism. Acta Neurobiol. Exp. 2012, 72, 113–153. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.K.; Geier, D.A.; Deth, R.C.; Sykes, L.K.; Hooker, B.S.; Love, J.M.; Bjørklund, G.; Chaigneau, C.G.; Haley, B.E.; Geier, M.R. Systematic Assessment of Research on Autism Spectrum Disorder (ASD) and Mercury Reveals Conflicts of Interest and the Need for Transparency in Autism Research. Sci. Eng. Ethic- 2017, 23, 1691–1718. [Google Scholar] [CrossRef]
- Kern, J.K.; Geier, D.A.; Sykes, L.K.; Geier, M.R. Relevance of Neuroinflammation and Encephalitis in Autism. Front. Cell. Neurosci. 2016, 9, 519. [Google Scholar] [CrossRef]
- Khachadourian, V.; Mahjani, B.; Sandin, S.; Kolevzon, A.; Buxbaum, J.D.; Reichenberg, A.; Janecka, M. Comorbidities in autism spectrum disorder and their etiologies. Transl. Psychiatry 2023, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Wang, G. Environmental agents, oxidative stress and autoimmunity. Curr. Opin. Toxicol. 2018, 7, 22–27. [Google Scholar] [CrossRef]
- Khan, S.; Gramfort, A.; Shetty, N.R.; Kitzbichler, M.G.; Ganesan, S.; Moran, J.M.; Lee, S.M.; Gabrieli, J.D.E.; Tager-Flusberg, H.B.; Joseph, R.M.; et al. Local and long-range functional connectivity is reduced in concert in autism spectrum disorders. Proc. Natl. Acad. Sci. 2013, 110, 3107–3112. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Michmizos, K.; Tommerdahl, M.; Ganesan, S.; Kitzbichler, M.G.; Zetino, M.; Garel, K.-L.A.; Herbert, M.R.; Hämäläinen, M.S.; Kenet, T. Somatosensory cortex functional connectivity abnormalities in autism show opposite trends, depending on direction and spatial scale. Brain 2015, 138, 1394–1409. [Google Scholar] [CrossRef]
- Kharrazian, D.; Herbert, M.; Lambert, J. The Relationships between Intestinal Permeability and Target Antibodies for a Spectrum of Autoimmune Diseases. Int. J. Mol. Sci. 2023, 24, 16352. [Google Scholar] [CrossRef] [PubMed]
- Khetrapal, N. The framework for disturbed affective consciousness in autism. Neuropsychiatr. Dis. Treat. 2008, 4, 531–533. [Google Scholar] [CrossRef]
- Khundakji, Y.; Masri, A.; Khuri-Bulos, N. Anti-NMDA receptor encephalitis in a toddler. Int. J. Pediatr. Adolesc. Med. 2018, 5, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-Y.; Camilleri, M. Serotonin: A Mediator of The Brain–Gut Connection. Am. J. Gastroenterol. 2000, 95, 2698–2709. [Google Scholar] [CrossRef]
- Kim, H.-J.; Cho, M.-H.; Shim, W.H.; Kim, J.K.; Jeon, E.-Y.; Kim, D.-H.; Yoon, S.-Y. Deficient autophagy in microglia impairs synaptic pruning and causes social behavioral defects. Mol. Psychiatry 2016, 22, 1576–1584. [Google Scholar] [CrossRef]
- King, B.H.; Hollander, E.; Sikich, L.; McCracken, J.T.; Scahill, L.; Bregman, J.D.; Donnelly, C.L.; Anagnostou, E.; Dukes, K.; Sullivan, L.; et al. Lack of Efficacy of Citalopram in Children With Autism Spectrum Disorders and High Levels of Repetitive Behavior. Arch. Gen. Psychiatry 2009, 66, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Kissane, D.W. The Relief of Existential Suffering. Arch. Intern. Med. 2012, 172, 1501–1505. [Google Scholar] [CrossRef]
- Kleine, B.; Rossmanith, W.G. Hormones and the Endocrine System; Springer Nature: Dordrecht, GX, Netherlands, 2016. [Google Scholar]
- Knapp, M.; Romeo, R.; Beecham, J. Economic cost of autism in the UK. Autism 2009, 13, 317–336. [Google Scholar] [CrossRef]
- Knudsen, G.P. Gender bias in autoimmune diseases: X chromosome inactivation in women with multiple sclerosis. J. Neurol. Sci. 2009, 286, 43–46. [Google Scholar] [CrossRef]
- Knuesel, I.; Chicha, L.; Britschgi, M.; Schobel, S.A.; Bodmer, M.; Hellings, J.A.; Toovey, S.; Prinssen, E.P. Maternal immune activation and abnormal brain development across CNS disorders. Nat. Rev. Neurol. 2014, 10, 643–660. [Google Scholar] [CrossRef]
- Ko, C.-L.; Lin, C.-K.; Lin, C.-L. Relationship between executive function and autism symptoms in preschoolers with autism spectrum disorder. Res. Dev. Disabil. 2024, 147, 104692. [Google Scholar] [CrossRef]
- Kobayashi, N.; Takahashi, D.; Takano, S.; Kimura, S.; Hase, K. The Roles of Peyer's Patches and Microfold Cells in the Gut Immune System: Relevance to Autoimmune Diseases. Front. Immunol. 2019, 10, 2345. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.-H.; Lee, S.; Kwak, M.; Kim, B.-S.; Chung, Y. CD8 T-cell subsets: heterogeneity, functions, and therapeutic potential. Exp. Mol. Med. 2023, 55, 2287–2299. [Google Scholar] [CrossRef]
- Konteh, F.H. Urban sanitation and health in the developing world: Reminiscing the nineteenth century industrial nations. Heal. Place 2009, 15, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Kovacheva, E.; Gevezova, M.; Maes, M.; Sarafian, V. Mast Cells in Autism Spectrum Disorder—The Enigma to Be Solved? Int. J. Mol. Sci. 2024, 25, 2651. [Google Scholar] [CrossRef]
- Koyama, R.; Ikegaya, Y. Microglia in the pathogenesis of autism spectrum disorders. Neurosci. Res. 2015, 100, 1–5. [Google Scholar] [CrossRef]
- Krausová, M.; Braun, D.; Buerki-Thurnherr, T.; Gundacker, C.; Schernhammer, E.; Wisgrill, L.; Warth, B. Understanding the Chemical Exposome During Fetal Development and Early Childhood: A Review. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 517–540. [Google Scholar] [CrossRef]
- Kushak, R.I.; Winter, H.S. Gut Microbiota and Gender in Autism Spectrum Disorders. Curr. Pediatr. Rev. 2020, 16, 249–254. [Google Scholar] [CrossRef]
- Lacivita, E.; Perrone, R.; Margari, L.; Leopoldo, M. Targets for Drug Therapy for Autism Spectrum Disorder: Challenges and Future Directions. J. Med. Chem. 2017, 60, 9114–9141. [Google Scholar] [CrossRef]
- Lahiri, D.K.; Maloney, B.; Wang, R.; Sokol, D.K.; Rogers, J.T.; Westmark, C.J. How autism and Alzheimer’s disease are TrAPPed. Mol. Psychiatry 2020, 26, 26–29. [Google Scholar] [CrossRef]
- Lai, Y.; Dhingra, R.; Zhang, Z.; Ball, L.M.; Zylka, M.J.; Lu, K. Toward Elucidating the Human Gut Microbiota–Brain Axis: Molecules, Biochemistry, and Implications for Health and Diseases. Biochemistry 2021, 61, 2806–2821. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.S.; Aman, M.G.; Arnold, L.E. Neurochemical correlates of autistic disorder: A review of the literature. Res. Dev. Disabil. 2006, 27, 254–289. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.W.; Hauser, J.; Reissmann, A. Gluten-free and casein-free diets in the therapy of autism. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 572–575. [Google Scholar] [CrossRef]
- Lebow, M., Kuperman, Y., & Chen, A. (2024). Prenatal-induced psychopathologies: All roads lead to microglia. In Stress: Immunology & Inflammation (pp. 199–214). Elsevier.
- Lederberg, J. Infectious History. Science 2000, 288, 287–293. [Google Scholar] [CrossRef]
- Lee, J.-S.; Sato, W.; Son, C.-G. Brain-regional characteristics and neuroinflammation in ME/CFS patients from neuroimaging: A systematic review and meta-analysis. Autoimmun. Rev. 2023, 23, 103484. [Google Scholar] [CrossRef] [PubMed]
- Leiter, O.; Walker, T.L. Platelets: The missing link between the blood and brain? Prog. Neurobiol. 2019, 183, 101695. [Google Scholar] [CrossRef]
- Leo, E.E.M.; Campos, M.R.S. Effect of ultra-processed diet on gut microbiota and thus its role in neurodegenerative diseases. Nutrients 2020, 71, 110609. [Google Scholar] [CrossRef]
- Lerer, L.; Varia, J. A long trip into the universe: Psychedelics and space travel. Front. Space Technol. 2022, 3. [Google Scholar] [CrossRef]
- Lesch, K.-P.; Waider, J. Serotonin in the Modulation of Neural Plasticity and Networks: Implications for Neurodevelopmental Disorders. Neuron 2012, 76, 175–191. [Google Scholar] [CrossRef]
- Leventhal, J.; Miller, J.; Abecassis, M.; Tollerud, D.J.; Ildstad, S.T. Evolving Approaches of Hematopoietic Stem Cell–Based Therapies to Induce Tolerance to Organ Transplants: The Long Road to Tolerance. Clin. Pharmacol. Ther. 2012, 93, 36–45. [Google Scholar] [CrossRef]
- Li, X.; Chauhan, A.; Sheikh, A.M.; Patil, S.; Chauhan, V.; Li, X.-M.; Ji, L.; Brown, T.; Malik, M. Elevated immune response in the brain of autistic patients. J. Neuroimmunol. 2009, 207, 111–116. [Google Scholar] [CrossRef]
- Li, X.; Zhang, K.; He, X.; Zhou, J.; Jin, C.; Shen, L.; Gao, Y.; Tian, M.; Zhang, H. Structural, Functional, and Molecular Imaging of Autism Spectrum Disorder. Neurosci. Bull. 2021, 37, 1051–1071. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Liu, Y.; Fu, X.; Li, Y. Postmortem Studies of Neuroinflammation in Autism Spectrum Disorder: a Systematic Review. Mol. Neurobiol. 2020, 57, 3424–3438. [Google Scholar] [CrossRef] [PubMed]
- Lilienfeld, S.O.; Marshall, J.; Todd, J.T.; Shane, H.C. The persistence of fad interventions in the face of negative scientific evidence: Facilitated communication for autism as a case example. Evidence-Based Commun. Assess. Interv. 2014, 8, 62–101. [Google Scholar] [CrossRef]
- Liu, G.; Pearl, A.M.; Kong, L.; Leslie, D.L.; Murray, M.J. A Profile on Emergency Department Utilization in Adolescents and Young Adults with Autism Spectrum Disorders. J. Autism Dev. Disord. 2016, 47, 347–358. [Google Scholar] [CrossRef]
- Liu, M.; Liang, S.; Zhang, C. NK Cells in Autoimmune Diseases: Protective or Pathogenic? Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Liu, S.-H.; Shi, X.-J.; Fan, F.-C.; Cheng, Y. Peripheral blood neurotrophic factor levels in children with autism spectrum disorder: a meta-analysis. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Lleo, A. (2014). Chapter 2 - What Is an Autoantibody? In Y. Shoenfeld, P. L. Meroni, & M. E. Gershwin (Eds.), Autoantibodies (Third Edition) (pp. 13–20). San Diego: Elsevier. https://doi.org/10.1016/B978-0-444-56378-1.00002-2.
- Lleo, A.; Invernizzi, P.; Gao, B.; Podda, M.; Gershwin, M.E. Definition of human autoimmunity — autoantibodies versus autoimmune disease. Autoimmun. Rev. 2010, 9, A259–A266. [Google Scholar] [CrossRef] [PubMed]
- López-Cacho, J.M.; Gallardo, S.; Posada, M.; Aguerri, M.; Calzada, D.; Mayayo, T.; Lahoz, C.; Cárdaba, B. Characterization of Immune Cell Phenotypes in Adults with Autism Spectrum Disorders. J. Investig. Med. 2016, 64, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- López-Varela, S.; González-Gross, M.; Marcos, A. Functional foods and the immune system: a review. Eur. J. Clin. Nutr. 2002, 56, S29–S33. [Google Scholar] [CrossRef]
- Lord, C.; Shulman, C.; DiLavore, P. Regression and word loss in autistic spectrum disorders. J. Child Psychol. Psychiatry 2004, 45, 936–955. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, Z. The Impact of Microglia on Neurodevelopment and Brain Function in Autism. Biomedicines 2024, 12, 210. [Google Scholar] [CrossRef] [PubMed]
- Luvián-Morales, J.; Varela-Castillo, F.O.; Flores-Cisneros, L.; Cetina-Pérez, L.; Castro-Eguiluz, D. Functional foods modulating inflammation and metabolism in chronic diseases: a systematic review. Crit. Rev. Food Sci. Nutr. 2021, 62, 4371–4392. [Google Scholar] [CrossRef]
- Luyster, R.; Richler, J.; Risi, S.; Hsu, W.-L.; Dawson, G.; Bernier, R.; Dunn, M.; Hepburn, S.; Hyman, S.L.; McMahon, W.M.; et al. Early Regression in Social Communication in Autism Spectrum Disorders: A CPEA Study. Dev. Neuropsychol. 2005, 27, 311–336. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Machado, A.P.; Ratliff, H.; Abdelwahab, A.; Vohra, M.H.; Kuang, A.; Shatila, M.; Khan, M.A.; Shafi, M.A.; Thomas, A.S.; Philpott, J.; et al. The Safety of Immunosuppressants Used in the Treatment of Immune-Related Adverse Events due to Immune Checkpoint Inhibitors: a Systematic Review. J. Cancer 2023, 14, 2956–2963. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Pachnis, V.; Prinz, M. Boundaries and integration between microbiota, the nervous system, and immunity. Immunity 2023, 56, 1712–1726. [Google Scholar] [CrossRef] [PubMed]
- Maenner, M.J. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years — Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2018. MMWR. Surveill. Summ. 2021, 70, 1–16. [Google Scholar] [CrossRef]
- Maier, S.F.; Watkins, L.R. Bidirectional communication between the brain and the immune system: implications for behaviour. Anim. Behav. 1999, 57, 741–751. [Google Scholar] [CrossRef]
- Maitre, L.; Bustamante, M.; Hernández-Ferrer, C.; Thiel, D.; Lau, C.-H.E.; Siskos, A.P.; Vives-Usano, M.; Ruiz-Arenas, C.; Pelegrí-Sisó, D.; Robinson, O.; et al. Multi-omics signatures of the human early life exposome. Nat. Commun. 2022, 13, 1–18. [Google Scholar] [CrossRef]
- Mallozzi, M.; Bordi, G.; Garo, C.; Caserta, D. The effect of maternal exposure to endocrine disrupting chemicals on fetal and neonatal development: A review on the major concerns. Birth Defects Res. Part C: Embryo Today: Rev. 2016, 108, 224–242. [Google Scholar] [CrossRef]
- Mamuladze, T.; Kipnis, J. Type 2 immunity in the brain and brain borders. Cell. Mol. Immunol. 2023, 20, 1290–1299. [Google Scholar] [CrossRef]
- Manji, H.; Kato, T.; Di Prospero, N.A.; Ness, S.; Beal, M.F.; Krams, M.; Chen, G. Impaired mitochondrial function in psychiatric disorders. Nat. Rev. Neurosci. 2012, 13, 293–307. [Google Scholar] [CrossRef]
- Margulis, L. , & Fester, R. (1991). Symbiosis as a Source of Evolutionary Innovation: Speciation & Morphogenesis. MIT Press.
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The Neurochemistry of Autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.E.; Marques, P.E.; Guabiraba, R.; Teixeira, M.M. Exploring the Homeostatic and Sensory Roles of the Immune System. Front. Immunol. 2016, 7, 125–125. [Google Scholar] [CrossRef] [PubMed]
- Marshall, L., & Born, J. (2002). Brain-Immune interactions in sleep. In International Review of Neurobiology (Vol. 52, pp. 93–131). Academic Press. https://doi.org/10.1016/S0074-7742(02)52007-9.
- Martin, A.; Scahill, L.; Anderson, G.M.; Aman, M.; Arnold, L.E.; McCracken, J.; McDougle, C.J.; Tierney, E.; Chuang, S.; Vitiello, B.; et al. Weight and Leptin Changes Among Risperidone-Treated Youths With Autism: 6-Month Prospective Data. Am. J. Psychiatry 2004, 161, 1125–1127. [Google Scholar] [CrossRef]
- Martínez-Cerdeño, V. Dendrite and spine modifications in autism and related neurodevelopmental disorders in patients and animal models. Dev. Neurobiol. 2016, 77, 393–404. [Google Scholar] [CrossRef]
- Marwaha, A.K.; Leung, N.J.; McMurchy, A.N.; Levings, M.K. TH17 Cells in Autoimmunity and Immunodeficiency: Protective or Pathogenic? Front. Immunol. 2012, 3, 129. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.; DeMayo, M.M.; Glozier, N.; Guastella, A.J. An Overview of Autism Spectrum Disorder, Heterogeneity and Treatment Options. Neurosci. Bull. 2017, 33, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Matcovitch-Natan, O.; Winter, D.R.; Giladi, A.; Aguilar, S.V.; Spinrad, A.; Sarrazin, S.; Ben-Yehuda, H.; David, E.; González, F.Z.; Perrin, P.; et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science 2016, 353, aad8670. [Google Scholar] [CrossRef]
- Mauri, C.; Bosma, A. Immune Regulatory Function of B Cells. Annu. Rev. Immunol. 2012, 30, 221–241. [Google Scholar] [CrossRef]
- Mayer, E.A. Gut feelings: the emerging biology of gut–brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut–Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Wright, C.L. Convergence of Sex Differences and the Neuroimmune System in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 402–410. [Google Scholar] [CrossRef]
- McCormick, C.M.; Furey, B.F.; Child, M.; Sawyer, M.J.; Donohue, S.M. Neonatal sex hormones have `organizational' effects on the hypothalamic-pituitary-adrenal axis of male rats. Dev. Brain Res. 1998, 105, 295–307. [Google Scholar] [CrossRef]
- McCracken, J.T.; Anagnostou, E.; Arango, C.; Dawson, G.; Farchione, T.; Mantua, V.; McPartland, J.; Murphy, D.; Pandina, G.; Veenstra-VanderWeele, J. Drug development for Autism Spectrum Disorder (ASD): Progress, challenges, and future directions. Eur. Neuropsychopharmacol. 2021, 48, 3–31. [Google Scholar] [CrossRef]
- McCruden, A. B., & Stimson, W. H. (1991). Sex Hormones and Immune Function. In R. ADER, D. L. FELTEN, & N. COHEN (Eds.), Psychoneuroimmunology (pp. 475–493). Academic Press. https://doi.org/10.1016/B978-0-12-043780-1.50021-X.
- McEwen, B.S. Protective and Damaging Effects of Stress Mediators. N. Engl. J. Med. 1998, 338, 171–179. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress, Adaptation, and Disease: Allostasis and Allostatic Load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef]
- Mead, J.; Ashwood, P. Evidence supporting an altered immune response in ASD. Immunol. Lett. 2015, 163, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Megremi, A.S. Is fever a predictive factor in the autism spectrum disorders? Med Hypotheses 2013, 80, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Mezzacappa, A.; Lasica, P.-A.; Gianfagna, F.; Cazas, O.; Hardy, P.; Falissard, B.; Sutter-Dallay, A.-L.; Gressier, F. Risk for Autism Spectrum Disorders According to Period of Prenatal Antidepressant Exposure. JAMA Pediatr. 2017, 171, 555–563. [Google Scholar] [CrossRef]
- Miclotte, L.; Van de Wiele, T. Food processing, gut microbiota and the globesity problem. Crit. Rev. Food Sci. Nutr. 2019, 60, 1769–1782. [Google Scholar] [CrossRef]
- Miller, H. , Zhang, J., Kuolee, R., Patel, G., & Chen, W. (2007). Intestinal M cells: The fallible sentinels? World Journal of Gastroenterology : WJG, 13, 1477–86. [CrossRef]
- Mills, E.L.; Kelly, B.; O’Neill, L.A.J. Mitochondria are the powerhouses of immunity. Nat. Immunol. 2017, 18, 488–498. [Google Scholar] [CrossRef]
- Misquita, A.G.; da Silva, C.N.F.; Souza, D.R.O.; dos Santos, D.N.B.; Brito, I.D.S.M.; Ribeiro, J.D.O.S.; Almeida, J.F.L.; Silva, J.A.G.; Barreto, N.P.M.; Rodrigues, S.C.P.; et al. Cognitive behavioral therapy: A valuable intervention in the autistic universe. Int. Seven- J. Heal. Res. 2024, 3, 754–760. [Google Scholar] [CrossRef]
- Mitsea, E.; Drigas, A.; Skianis, C. Cutting-Edge Technologies in Breathwork for Learning Disabilities in Special Education. Tech. Soc. Sci. J. 2022, 34, 136–157. [Google Scholar] [CrossRef]
- Modafferi, S.; Lupo, G.; Tomasello, M.; Rampulla, F.; Ontario, M.; Scuto, M.; Salinaro, A.T.; Arcidiacono, A.; Anfuso, C.D.; Legmouz, M.; et al. Antioxidants, Hormetic Nutrition, and Autism. Curr. Neuropharmacol. 2024, 22, 1156–1168. [Google Scholar] [CrossRef]
- Nordin, A.M.; Ismail, J.; Nor, N.K. Motor Development in Children With Autism Spectrum Disorder. Front. Pediatr. 2021, 9. [Google Scholar] [CrossRef]
- Molloy, C.; Morrow, A.; Meinzenderr, J.; Schleifer, K.; Dienger, K.; Manningcourtney, P.; Altaye, M.; Willskarp, M. Elevated cytokine levels in children with autism spectrum disorder. J. Neuroimmunol. 2006, 172, 198–205. [Google Scholar] [CrossRef]
- Moncrieff, J. Why is it so difficult to stop psychiatric drug treatment? Med Hypotheses 2006, 67, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Moncrieff, J.; Cohen, D.; Porter, S. The Psychoactive Effects of Psychiatric Medication: The Elephant in the Room. J. Psychoact. Drugs 2013, 45, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Money, J.; Bobrow, N.A.; Clarke, F.C. Autism and autoimmune disease: A family study. J. Autism Dev. Disord. 1971, 1, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.A.; dos Santos, A.A.A.; Gomes, L.M.M.; Rito, R.V.V.F. AUTISM SPECTRUM DISORDER: A SYSTEMATIC REVIEW ABOUT NUTRITIONAL INTERVENTIONS. Rev. Paul. de Pediatr. 2020, 38, e2018262. [Google Scholar] [CrossRef]
- Montgomery, S.L.; Bowers, W.J. Tumor Necrosis Factor-alpha and the Roles it Plays in Homeostatic and Degenerative Processes Within the Central Nervous System. J. Neuroimmune Pharmacol. 2011, 7, 42–59. [Google Scholar] [CrossRef]
- Moore, J.B.; Weeks, M.E. Proteomics and Systems Biology: Current and Future Applications in the Nutritional Sciences. Adv. Nutr. Int. Rev. J. 2011, 2, 355–364. [Google Scholar] [CrossRef]
- Mordelt, A.; de Witte, L.D. Microglia-mediated synaptic pruning as a key deficit in neurodevelopmental disorders: Hype or hope? Curr. Opin. Neurobiol. 2023, 79, 102674. [Google Scholar] [CrossRef]
- Morgan, J.T.; Chana, G.; Pardo, C.A.; Achim, C.; Semendeferi, K.; Buckwalter, J.; Courchesne, E.; Everall, I.P. Microglial Activation and Increased Microglial Density Observed in the Dorsolateral Prefrontal Cortex in Autism. Biol. Psychiatry 2010, 68, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Moroncini, G.; Calogera, G.; Benfaremo, D.; Gabrielli, A. Biologics in Inflammatory Immune-mediated Systemic Diseases. Curr. Pharm. Biotechnol. 2018, 18, 1008–1016. [Google Scholar] [CrossRef]
- Moroni, L.; Bianchi, I.; Lleo, A. Geoepidemiology, gender and autoimmune disease. Autoimmun. Rev. 2011, 11, A386–A392. [Google Scholar] [CrossRef]
- Mosconi, M.W.; Wang, Z.; Schmitt, L.M.; Tsai, P.; Sweeney, J.A. The role of cerebellar circuitry alterations in the pathophysiology of autism spectrum disorders. Front. Neurosci. 2015, 9, 296. [Google Scholar] [CrossRef] [PubMed]
- Mulder, S.J.; Mulder-Bos, G.C. Most probable origin of coeliac disease is low immune globulin A in the intestine caused by malfunction of Peyer’s patches. Med Hypotheses 2006, 66, 757–762. [Google Scholar] [CrossRef]
- Nadeem, A.; Ahmad, S.F.; Al-Harbi, N.O.; Al-Ayadhi, L.Y.; Sarawi, W.; Attia, S.M.; Bakheet, S.A.; Alqarni, S.A.; Ali, N.; AsSobeai, H.M. Imbalance in pro-inflammatory and anti-inflammatory cytokines milieu in B cells of children with autism. Mol. Immunol. 2022, 141, 297–304. [Google Scholar] [CrossRef]
- Naviaux, R.K. Perspective: Cell danger response Biology—The new science that connects environmental health with mitochondria and the rising tide of chronic illness. Mitochondrion 2020, 51, 40–45. [Google Scholar] [CrossRef]
- Nelissen, S.; Lemmens, E.; Geurts, N.; Kramer, P.; Maurer, M.; Hendriks, J.; Hendrix, S. The role of mast cells in neuroinflammation. Acta Neuropathol. 2013, 125, 637–650. [Google Scholar] [CrossRef]
- Nelson, A.D.; Bender, K.J. Dendritic Integration Dysfunction in Neurodevelopmental Disorders. Dev. Neurosci. 2021, 43, 201–221. [Google Scholar] [CrossRef]
- Nelson, I. Exploring the influence of Schumann resonance and electromagnetic fields on bioelectricity and human health. Electromagn. Biol. Med. 2025, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A.B.; Latz, E.; Mills, K.H.G.; Natoli, G.; Stunnenberg, H.G.; O’nEill, L.A.J.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098–aaf1098. [Google Scholar] [CrossRef] [PubMed]
- Nevison, C.; Blaxill, M.; Zahorodny, W. California Autism Prevalence Trends from 1931 to 2014 and Comparison to National ASD Data from IDEA and ADDM. J. Autism Dev. Disord. 2018, 48, 4103–4117. [Google Scholar] [CrossRef]
- Nevison, C.D.; Blaxill, M. Diagnostic Substitution for Intellectual Disability: A Flawed Explanation for the Rise in Autism. J. Autism Dev. Disord. 2017, 47, 2733–2742. [Google Scholar] [CrossRef]
- Ngo, S.T.; Steyn, F.J.; McCombe, P.A. Gender differences in autoimmune disease. Front. Neuroendocrinol. 2014, 35, 347–369. [Google Scholar] [CrossRef]
- Niciu, M.J.; Kelmendi, B.; Sanacora, G. Overview of glutamatergic neurotransmission in the nervous system. Pharmacol. Biochem. Behav. 2012, 100, 656–664. [Google Scholar] [CrossRef]
- Nie, Z.-Q.; Han, D.; Zhang, K.; Li, M.; Kwon, H.-K.; Im, S.-H.; Xu, L.; Yang, J.-C.; Li, Z.-W.; Huang, X.-W.; et al. TH1/Treg ratio may be a marker of autism in children with immune dysfunction. Res. Autism Spectr. Disord. 2022, 101. [Google Scholar] [CrossRef]
- NIEHS. (2024). Autoimmune Diseases. https://www.niehs.nih.gov/health/topics/conditions/autoimmune. 1 May.
- Niesler, B.; Kuerten, S.; Demir, I.E.; Schäfer, K.-H. Disorders of the enteric nervous system — a holistic view. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 393–410. [Google Scholar] [CrossRef]
- NIMH. (2024). Autism Spectrum Disorder. https://www.nimh.nih.gov/health/topics/autism-spectrum-disorders-asd. 16 July.
- Complication of encephalitis in children, an innovative approach for the treatment. Int. J. Pharm. Res. 2020, 12. [CrossRef]
- Njotto, L.L.; Simin, J.; Fornes, R.; Odsbu, I.; Mussche, I.; Callens, S.; Engstrand, L.; Bruyndonckx, R.; Brusselaers, N. Maternal and Early-Life Exposure to Antibiotics and the Risk of Autism and Attention-Deficit Hyperactivity Disorder in Childhood: a Swedish Population-Based Cohort Study. Drug Saf. 2023, 46, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Nyati, K.K.; Prasad, K.N. Role of Cytokines and Toll-Like Receptors in the Immunopathogenesis of Guillain-Barré Syndrome. Mediat. Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Pachnis, V. The Effect of Microbiota and the Immune System on the Development and Organization of the Enteric Nervous System. Gastroenterology 2016, 151, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: a therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2016, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Nelson, I. Exploring the influence of Schumann resonance and electromagnetic fields on bioelectricity and human health. Electromagn. Biol. Med. 2025, 1–11. [Google Scholar] [CrossRef]
- van Ommen, B.; Stierum, R. Nutrigenomics: exploiting systems biology in the nutrition and health arena. Curr. Opin. Biotechnol. 2002, 13, 517–521. [Google Scholar] [CrossRef]
- Onore, C.; Careaga, M.; Ashwood, P. The role of immune dysfunction in the pathophysiology of autism. Brain, Behav. Immun. 2012, 26, 383–392. [Google Scholar] [CrossRef]
- Orekhova, E.V.; Stroganova, T.A.; Nygren, G.; Tsetlin, M.M.; Posikera, I.N.; Gillberg, C.; Elam, M. Excess of High Frequency Electroencephalogram Oscillations in Boys with Autism. Biol. Psychiatry 2007, 62, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Ozonoff, S.; Iosif, A.-M.; Baguio, F.; Cook, I.C.; Hill, M.M.; Hutman, T.; Rogers, S.J.; Rozga, A.; Sangha, S.; Sigman, M.; et al. A Prospective Study of the Emergence of Early Behavioral Signs of Autism. J. Am. Acad. Child Adolesc. Psychiatry 2010, 49, 256–266.e2. [Google Scholar] [CrossRef]
- Padmakumar, M.; Van Raes, E.; Van Geet, C.; Freson, K. Blood platelet research in autism spectrum disorders: In search of biomarkers. Res. Pr. Thromb. Haemost. 2019, 3, 566–577. [Google Scholar] [CrossRef]
- Palmieri, L.; Persico, A.M. Mitochondrial dysfunction in autism spectrum disorders: Cause or effect? Biochim. et Biophys. Acta (BBA) - Bioenerg. 2010, 1797, 1130–1137. [Google Scholar] [CrossRef]
- Panksepp, J. A neurochemical theory of autism. Trends Neurosci. 1979, 2, 174–177. [Google Scholar] [CrossRef]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Pardo, C.A.; Eberhart, C.G. The Neurobiology of Autism. Brain Pathol. 2007, 17, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Pardo, C.A.; Vargas, D.L.; Zimmerman, A.W. Immunity, neuroglia and neuroinflammation in autism. Int. Rev. Psychiatry 2005, 17, 485–495. [Google Scholar] [CrossRef]
- Park, K.M.; Bowers, W.J. Tumor necrosis factor-alpha mediated signaling in neuronal homeostasis and dysfunction. Cell. Signal. 2010, 22, 977–983. [Google Scholar] [CrossRef]
- Pathak, D.; Sriram, K. Neuron-astrocyte omnidirectional signaling in neurological health and disease. Front. Mol. Neurosci. 2023, 16, 1169320. [Google Scholar] [CrossRef]
- Patterson, P.H. Immune involvement in schizophrenia and autism: Etiology, pathology and animal models. Behav. Brain Res. 2009, 204, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Paveenakiattikhun, S.; Likhitweerawong, N.; Sanguansermsri, C. EEG findings and clinical severity and quality of life in non-epileptic patients with autism spectrum disorders. Child Neuropsychol. 2024, 31, 255–265. [Google Scholar] [CrossRef]
- Peper, E.; Shufor, J. Reflections on the Increase in Autism, ADHD, Anxiety, and Depression: Part 2 – Exposure to Neurotoxins and Ultraprocessed Foods. NeuroRegulation 2024, 11, 219–228. [Google Scholar] [CrossRef]
- Peralta-Marzal, L.N.; Rojas-Velazquez, D.; Rigters, D.; Prince, N.; Garssen, J.; Kraneveld, A.D.; Perez-Pardo, P.; Lopez-Rincon, A. A robust microbiome signature for autism spectrum disorder across different studies using machine learning. Sci. Rep. 2024, 14, 1–11. [Google Scholar] [CrossRef]
- Pereira, A. (2020). Chapter Seven - Classical-quantum interfaces in living neural tissue supporting conscious functions. In R. R. Poznański & E. J. Brändas (Eds.), Advances in Quantum Chemistry (Vol. 82, pp. 213–252). Academic Press. https://doi.org/10.1016/bs.aiq.2020.08.002.
- Pereira, A.; Furlan, F.A. On the role of synchrony for neuron–astrocyte interactions and perceptual conscious processing. J. Biol. Phys. 2009, 35, 465–480. [Google Scholar] [CrossRef]
- Pereira, A.; Furlan, F.A. Astrocytes and human cognition: Modeling information integration and modulation of neuronal activity. Prog. Neurobiol. 2010, 92, 405–420. [Google Scholar] [CrossRef]
- Pereira, J. A. (2020). The Role of Sentience in the Theory of Consciousness and Medical Practice. [CrossRef]
- Pereira, A.; Wang, F. Neuromodulation, Emotional Feelings and Affective Disorders. Mens Sana Monogr. 2016, 14, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Perrin, J.M.; Anderson, L.E.; Van Cleave, J. The Rise In Chronic Conditions Among Infants, Children, And Youth Can Be Met With Continued Health System Innovations. Heal. Aff. 2014, 33, 2099–2105. [Google Scholar] [CrossRef]
- Petrelli, F.; Pucci, L.; Bezzi, P. Astrocytes and Microglia and Their Potential Link with Autism Spectrum Disorders. Front. Cell. Neurosci. 2016, 10, 21. [Google Scholar] [CrossRef]
- Pierce, S.; Kadlaskar, G.; Edmondson, D.A.; Keehn, R.M.; Dydak, U.; Keehn, B. Associations between sensory processing and electrophysiological and neurochemical measures in children with ASD: an EEG-MRS study. J. Neurodev. Disord. 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ploog, B. O. (2023). Theory of Mind in Autism. In A. El Idrissi & D. McCloskey (Eds.), Neurobiology of Autism Spectrum Disorders (pp. 23–35). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-031-42383-3_2.
- Gerber, J.S.; Offit, P.A. Vaccines and Autism: A Tale of Shifting Hypotheses. Clin. Infect. Dis. 2009, 48, 456–461. [Google Scholar] [CrossRef]
- Pociūtė, A.; Pivoriūnas, A.; Verkhratsky, A. Astrocytes dynamically regulate the blood-brain barrier in the healthy brain. Neural Regen. Res. 2023, 19, 709–710. [Google Scholar] [CrossRef]
- Poling, J.S.; Frye, R.E.; Shoffner, J.; Zimmerman, A.W. Developmental Regression and Mitochondrial Dysfunction in a Child With Autism. J. Child Neurol. 2006, 21, 170–172. [Google Scholar] [CrossRef]
- Politte, L.C.; Howe, Y.; Nowinski, L.; Palumbo, M.; McDougle, C.J. Evidence-Based Treatments for Autism Spectrum Disorder. Curr. Treat. Options Psychiatry 2015, 2, 38–56. [Google Scholar] [CrossRef]
- Pourgholaminejad, A., & Tahmasebinia, F. (2019). The Role of Th17 Cells in Immunopathogenesis of Neuroinflammatory Disorders. In H. Mitoma & M. Manto (Eds.), Neuroimmune Diseases: From Cells to the Living Brain (pp. 83–107). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-030-19515-1_3.
- Pretzsch, C.M.; Schäfer, T.; Lombardo, M.V.; Warrier, V.; Mann, C.; Bletsch, A.; Chatham, C.H.; Floris, D.L.; Tillmann, J.; Yousaf, A.; et al. Neurobiological Correlates of Change in Adaptive Behavior in Autism. Am. J. Psychiatry 2022, 179, 336–349. [Google Scholar] [CrossRef]
- Prüss, H. Autoantibodies in neurological disease. Nat. Rev. Immunol. 2021, 21, 798–813. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Yang, J.; Chang, L.; Qu, Y.; Wang, S.; Zhang, K.; Xiong, Z.; Zhang, J.; Tan, Y.; Wang, X.; et al. Maternal glyphosate exposure causes autism-like behaviors in offspring through increased expression of soluble epoxide hydrolase. Proc. Natl. Acad. Sci. 2020, 117, 11753–11759. [Google Scholar] [CrossRef]
- Pulikkan, J., Mazumder, A., & Grace, T. (2019). Role of the gut microbiome in autism spectrum disorders. Reviews on Biomarker Studies in Psychiatric & Neurodegenerative disorders, 253–269. https://doi.org/10.1007/978-3-030-05542-4_13.
- Puricelli, C.; Rolla, R.; Gigliotti, L.; Boggio, E.; Beltrami, E.; Dianzani, U.; Keller, R. The Gut-Brain-Immune Axis in Autism Spectrum Disorders: A State-of-Art Report. Front. Psychiatry 2022, 12, 755171. [Google Scholar] [CrossRef]
- Qi, H. Neuroimmunology: reviews and perspectives on recent advances. Cell. Mol. Immunol. 2023, 20, 1257–1258. [Google Scholar] [CrossRef]
- Rabinovici, G.D.; Stephens, M.L.; Possin, K.L. Executive Dysfunction. Contin. Lifelong Learn. Neurol. 2015, 21, 646–659. [Google Scholar] [CrossRef]
- Ramautar, R.; Berger, R.; van der Greef, J.; Hankemeier, T. Human metabolomics: strategies to understand biology. Curr. Opin. Chem. Biol. 2013, 17, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, D.S.; Woods, S.C. Clarifying the roles of homeostasis and allostasis in physiological regulation. Psychol. Rev. 2014, 121, 225–247. [Google Scholar] [CrossRef] [PubMed]
- Randolph-Gips, M.; Srinivasan, P. Modeling autism: a systems biology approach. J. Clin. Bioinform. 2012, 2, 17–17. [Google Scholar] [CrossRef]
- Reemst, K.; Noctor, S.C.; Lucassen, P.J.; Hol, E.M. The Indispensable Roles of Microglia and Astrocytes during Brain Development. Front. Hum. Neurosci. 2016, 10, 566. [Google Scholar] [CrossRef]
- Reid, G.; Younes, J.A.; Van Der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: natural and supplemented recovery of human microbial communities. Nat. Rev. Genet. 2011, 9, 27–38. [Google Scholar] [CrossRef]
- Renz, H.; Holt, P.G.; Inouye, M.; Logan, A.C.; Prescott, S.L.; Sly, P.D. An exposome perspective: Early-life events and immune development in a changing world. J. Allergy Clin. Immunol. 2017, 140, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Réthelyi, J.M.; Vincze, K.; Schall, D.; Glennon, J.; Berkel, S. The role of insulin/IGF1 signalling in neurodevelopmental and neuropsychiatric disorders – Evidence from human neuronal cell models. Neurosci. Biobehav. Rev. 2023, 153, 105330. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The crucial role of mast cells in blood–brain barrier alterations. Exp. Cell Res. 2015, 338, 119–125. [Google Scholar] [CrossRef]
- Ribeiro, S. Whole Organisms or Pure Compounds? Entourage Effect Versus Drug Specificity. In Plant Medicines, Healing and Psychedelic Science; Labate, B., Cavnar, C., Eds.; Springer: Cham, Switzerland, 2018; pp. 133–149. [Google Scholar] [CrossRef]
- Riikonen, R. Insulin-Like Growth Factors in the Pathogenesis of Neurological Diseases in Children. Int. J. Mol. Sci. 2017, 18, 2056. [Google Scholar] [CrossRef]
- Robertson, J.M. The Astrocentric Hypothesis: proposed role of astrocytes in consciousness and memory formation. J. Physiol. 2002, 96, 251–255. [Google Scholar] [CrossRef]
- Rogers, S. J. , Cook, I., & Meryl, A. (2005). Imitation and play in autism. In Handbook of Autism a& Pervasive Developmental Disorders (Vol. 1, pp. 382–405). Wiley Online Library.
- Romagnani, S. Th1/Th2 Cells. Inflamm. Bowel Dis. 1999, 5, 285–294. [Google Scholar] [CrossRef]
- Rook, G.A.W. Hygiene Hypothesis and Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2011, 42, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Rosen, N.E.; Lord, C.; Volkmar, F.R. The Diagnosis of Autism: From Kanner to DSM-III to DSM-5 and Beyond. J. Autism Dev. Disord. 2021, 51, 4253–4270. [Google Scholar] [CrossRef]
- Rosin, J.M.; Kurrasch, D.M. Emerging roles for hypothalamic microglia as regulators of physiological homeostasis. Front. Neuroendocr. 2019, 54, 100748. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Frye, R.E. A review of research trends in physiological abnormalities in autism spectrum disorders: immune dysregulation, inflammation, oxidative stress, mitochondrial dysfunction and environmental toxicant exposures. Mol. Psychiatry 2012, 17, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, D.A.; Frye, R.E. Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism. Front. Physiol. 2014, 5, 150. [Google Scholar] [CrossRef]
- Rubenstein, J.L.R.; Merzenich, M.M. Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes, Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef]
- Rucklidge, J.J.; Johnstone, J.M.; Kaplan, B.J. Nutrition Provides the Essential Foundation for Optimizing Mental Health. Evidence-Based Pr. Child Adolesc. Ment. Heal. 2021, 6, 131–154. [Google Scholar] [CrossRef]
- Rueda-Ruzafa, L.; Cruz, F.; Roman, P.; Cardona, D. Gut microbiota and neurological effects of glyphosate. NeuroToxicology 2019, 75, 1–8. [Google Scholar] [CrossRef]
- Saad, K.; Zahran, A.M.; Elsayh, K.I.; Abdel-Rahman, A.A.; Al-Atram, A.A.; Hussein, A.; El-Gendy, Y.G. Frequency of Dendritic Cells and Their Expression of Costimulatory Molecules in Children with Autism Spectrum Disorders. J. Autism Dev. Disord. 2017, 47, 2671–2678. [Google Scholar] [CrossRef]
- Safari-Alighiarloo, N. , Taghizadeh, M., Rezaei tavirani, M., Goliaei, B., & Peyvandi, A. (2014). Protein-Protein Interaction Networks (PPI) and complex diseases. Gastroenterology & Hepatology From Bed to Bench, 7, 17–31. https://pubmed.ncbi.nlm.nih. 2543. [Google Scholar]
- Salmond, C.H.; de Haan, M.; Friston, K.J.; Gadian, D.G.; Vargha-Khadem, F.; Frith, U.; Hill, E.L. Investigating individual differences in brain abnormalities in autism. Philos. Trans. R. Soc. B: Biol. Sci. 2003, 358, 405–413. [Google Scholar] [CrossRef]
- Salter, M.W.; Stevens, B. Microglia emerge as central players in brain disease. Nat. Med. 2017, 23, 1018–1027. [Google Scholar] [CrossRef]
- Samanipour, S.; Barron, L.P.; van Herwerden, D.; Praetorius, A.; Thomas, K.V.; O’bRien, J.W. Exploring the Chemical Space of the Exposome: How Far Have We Gone? JACS Au 2024, 4, 2412–2425. [Google Scholar] [CrossRef] [PubMed]
- Samsam, M.; Ahangari, R.; Naser, S.A. Pathophysiology of autism spectrum disorders: Revisiting gastrointestinal involvement and immune imbalance. World J. Gastroenterol. 2014, 20, 9942–51. [Google Scholar] [CrossRef] [PubMed]
- Samsel, A.; Seneff, S. Glyphosate’s Suppression of Cytochrome P450 Enzymes and Amino Acid Biosynthesis by the Gut Microbiome: Pathways to Modern Diseases. Entropy 2013, 15, 1416–1463. [Google Scholar] [CrossRef]
- Sánchez-Ramón, S.; Faure, F. Self and the Brain. The Immune Metaphor. Front. Psychiatry 2020, 11, 540676. [Google Scholar] [CrossRef]
- Santarone, M.E.; Zambrano, S.; Zanotta, N.; Mani, E.; Minghetti, S.; Pozzi, M.; Villa, L.; Molteni, M.; Zucca, C. EEG Features in Autism Spectrum Disorder: A Retrospective Analysis in a Cohort of Preschool Children. Brain Sci. 2023, 13, 345. [Google Scholar] [CrossRef]
- Saresella, M.; Piancone, F.; Marventano, I.; Zoppis, M.; Hernis, A.; Zanette, M.; Trabattoni, D.; Chiappedi, M.; Ghezzo, A.; Canevini, M.P.; et al. Multiple inflammasome complexes are activated in autistic spectrum disorders. Brain, Behav. Immun. 2016, 57, 125–133. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef]
- Schwartz, M.; Cahalon, L. The vicious cycle governing the brain–immune system relationship in neurodegenerative diseases. Curr. Opin. Immunol. 2022, 76, 102182. [Google Scholar] [CrossRef] [PubMed]
- Segal, Y.; Shoenfeld, Y. Vaccine-induced autoimmunity: the role of molecular mimicry and immune crossreaction. Cell. Mol. Immunol. 2018, 15, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Seneff, S.; Kyriakopoulos, A.M.; Nigh, G. Is autism a PIN1 deficiency syndrome? A proposed etiological role for glyphosate. J. Neurochem. 2024, 168, 2124–2146. [Google Scholar] [CrossRef]
- Seneff, S. , & Orlando, L. (2018). ( 8(1), 541. [CrossRef]
- Serrano-Pozo, A. (2018). Encephalopathy. In The Wiley Handbook on the Aging Mind and Brain (pp. 553–590). https://doi.org/10.1002/9781118772034.ch25.
- Shah, P.A.; Park, C.J.; Shaughnessy, M.P.; Cowles, R.A. Serotonin as a Mitogen in the Gastrointestinal Tract: Revisiting a Familiar Molecule in a New Role. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1093–1104. [Google Scholar] [CrossRef]
- Shahar, O.; Botvinnik, A.; Shwartz, A.; Lerer, E.; Golding, P.; Buko, A.; Hamid, E.; Kahn, D.; Guralnick, M.; Blakolmer, K.; et al. Effect of chemically synthesized psilocybin and psychedelic mushroom extract on molecular and metabolic profiles in mouse brain. Mol. Psychiatry 2024, 29, 2059–2073. [Google Scholar] [CrossRef]
- Shahbazi, R.; Sharifzad, F.; Bagheri, R.; Alsadi, N.; Yasavoli-Sharahi, H.; Matar, C. Anti-Inflammatory and Immunomodulatory Properties of Fermented Plant Foods. Nutrients 2021, 13, 1516. [Google Scholar] [CrossRef] [PubMed]
- Shajib, M.S.; Baranov, A.; Khan, W.I. Diverse Effects of Gut-Derived Serotonin in Intestinal Inflammation. ACS Chem. Neurosci. 2017, 8, 920–931. [Google Scholar] [CrossRef]
- Shaw, C.A.; Seneff, S.; Kette, S.D.; Tomljenovic, L.; Oller, J.W.; Davidson, R.M. Aluminum-Induced Entropy in Biological Systems: Implications for Neurological Disease. J. Toxicol. 2014, 2014, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.C.; Crombie, G.K.; Palliser, H.K.; Hirst, J.J. Impaired Oligodendrocyte Development Following Preterm Birth: Promoting GABAergic Action to Improve Outcomes. Front. Pediatr. 2021, 9. [Google Scholar] [CrossRef]
- Shaw, K.A. Prevalence and Early Identification of Autism Spectrum Disorder Among Children Aged 4 and 8 Years — Autism and Developmental Disabilities Monitoring Network, 16 Sites, United States, 2022. MMWR. Surveill. Summ. 2025, 74, 1–22. [Google Scholar] [CrossRef]
- Shin, Y.-W.; Lee, S.-T.; Park, K.-I.; Jung, K.-H.; Jung, K.-Y.; Lee, S.K.; Chu, K. Treatment strategies for autoimmune encephalitis. Ther. Adv. Neurol. Disord. 2017, 11. [Google Scholar] [CrossRef]
- Siegel, M.; Milligan, B.; Chemelski, B.; Payne, D.; Ellsworth, B.; Harmon, J.; Teer, O.; Smith, K.A. Specialized Inpatient Psychiatry for Serious Behavioral Disturbance in Autism and Intellectual Disability. J. Autism Dev. Disord. 2014, 44, 3026–3032. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: a concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Cruz-Machado, S.d.S.; Campos, L.M.G.; Fadini, C.C.; Anderson, G.; Markus, R.P.; Pinato, L. Disrupted nocturnal melatonin in autism: Association with tumor necrosis factor and sleep disturbances. J. Pineal Res. 2021, 70, e12715. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.S.; Ni, G.; Lam, V.; Demopoulos, C. Beyond words: an investigation of fine motor skills and the verbal communication spectrum in autism. Front. Psychiatry 2024, 15, 1379307. [Google Scholar] [CrossRef]
- Simons, M.; Levin, J.; Dichgans, M. Tipping points in neurodegeneration. Neuron 2023, 111, 2954–2968. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Siniscalco, D.; Bradstreet, J.J.; Antonucci, N. Therapeutic Role of Hematopoietic Stem Cells in Autism Spectrum Disorder-Related Inflammation. Front. Immunol. 2013, 4, 140. [Google Scholar] [CrossRef]
- Sloan, S.A.; Barres, B.A. Mechanisms of astrocyte development and their contributions to neurodevelopmental disorders. Curr. Opin. Neurobiol. 2014, 27, 75–81. [Google Scholar] [CrossRef]
- Smith, A.; Storti, S.; Lukose, R.; Jr, R.J.K. Structural and Functional Aberrations of the Auditory Brainstem in Autism Spectrum Disorder. J. Am. Osteopat. Assoc. 2019, 119, 41–50. [Google Scholar] [CrossRef]
- Smyth, M.J.; Godfrey, D.I.; Trapani, J.A. A fresh look at tumor immunosurveillance and immunotherapy. Nat. Immunol. 2001, 2, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Ssucharewa, G. Die schizoiden Psychopathien im Kindesalter. (Part 1 of 2). Eur. Neurol. 1926, 60, 235–247. [Google Scholar] [CrossRef]
- Starr, A.; Amlie, R.N.; Martin, W.H.; Sanders, S. Development of Auditory Function in Newborn Infants Revealed by Auditory Brainstem Potentials. Pediatrics 1977, 60, 831–839. [Google Scholar] [CrossRef]
- Stein, Y., Hänninen, O., Huttunen, P., Ahonen, M., & Ekman, R. (2015). Electromagnetic Radiation and Health: Human Indicators. In R. H. Armon & O. Hänninen (Eds.), Environmental Indicators (pp. 1025–1046). Dordrecht: Springer Netherlands. https://doi.org/10.1007/978-94-017-9499-2_57.
- Steinman, G. Plausible etiology of brain dysconnectivity in autism – Review and prospectus. Med Hypotheses 2015, 85, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Steinman, G.; Mankuta, D. Insulin-like growth factor and the etiology of autism. Med Hypotheses 2013, 80, 475–480. [Google Scholar] [CrossRef]
- Stilling, R.M.; Bordenstein, S.R.; Dinan, T.G.; Cryan, J.F. Friends with social benefits: host-microbe interactions as a driver of brain evolution and development? Front. Cell. Infect. Microbiol. 2014, 4, 147–147. [Google Scholar] [CrossRef]
- Stinson, L.F.; Boyce, M.C.; Payne, M.S.; Keelan, J.A. The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Front. Microbiol. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Stinson, L.F.; Payne, M.S.; Keelan, J.A. Planting the seed: Origins, composition, and postnatal health significance of the fetal gastrointestinal microbiota. Crit. Rev. Microbiol. 2016, 43, 352–369. [Google Scholar] [CrossRef]
- Storch, E.A.; Arnold, E.B.; Lewin, A.B.; Nadeau, J.M.; Jones, A.M.; De Nadai, A.S.; Mutch, P.J.; Selles, R.R.; Ung, D.; Murphy, T.K. The Effect of Cognitive-Behavioral Therapy Versus Treatment as Usual for Anxiety in Children With Autism Spectrum Disorders: A Randomized, Controlled Trial. J. Am. Acad. Child Adolesc. Psychiatry 2013, 52, 132–142.e2. [Google Scholar] [CrossRef]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 1–11. [Google Scholar] [CrossRef]
- Streng, M.L.; Krook-Magnuson, E. The cerebellum and epilepsy. Epilepsy Behav. 2021, 121, 106909–106909. [Google Scholar] [CrossRef]
- Stubbs, E.G.; Crawford, M.L.; Burger, D.R.; Vandenbark, A.A. Depressed lymphocyte responsiveness in autistic children. J. Autism Child. Schizophr. 1977, 7, 49–55. [Google Scholar] [CrossRef]
- Su, L.-D.; Xu, F.-X.; Wang, X.-T.; Cai, X.-Y.; Shen, Y. Cerebellar Dysfunction, Cerebro-cerebellar Connectivity and Autism Spectrum Disorders. Neuroscience 2021, 462, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Sugihara, G.; Ouchi, Y.; Nakamura, K.; Futatsubashi, M.; Takebayashi, K.; Yoshihara, Y.; Omata, K.; Matsumoto, K.; Tsuchiya, K.J.; et al. Microglial Activation in Young Adults With Autism Spectrum Disorder. JAMA Psychiatry 2013, 70, 49–58. [Google Scholar] [CrossRef]
- Swatzyna, R.J.; Boutros, N.N.; Genovese, A.C.; MacInerney, E.K.; Roark, A.J.; Kozlowski, G.P. Electroencephalogram (EEG) for children with autism spectrum disorder: evidential considerations for routine screening. Eur. Child Adolesc. Psychiatry 2018, 28, 615–624. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Sweeten, T.L.; Posey, D.J.; Shekhar, A.; McDougle, C.J. The amygdala and related structures in the pathophysiology of autism. Pharmacol. Biochem. Behav. 2002, 71, 449–455. [Google Scholar] [CrossRef]
- Tahmasebinia, F.; Pourgholaminejad, A. The role of Th17 cells in auto-inflammatory neurological disorders. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2017, 79, 408–416. [Google Scholar] [CrossRef]
- Taniya, M.A.; Chung, H.-J.; Al Mamun, A.; Alam, S.; Aziz, A.; Emon, N.U.; Islam, M.; Hong, S.-T.S.; Podder, B.R.; Mimi, A.A.; et al. Role of Gut Microbiome in Autism Spectrum Disorder and Its Therapeutic Regulation. Front. Cell. Infect. Microbiol. 2022, 12, 915701. [Google Scholar] [CrossRef]
- Tarabeux, J.; Kebir, O.; Gauthier, J.; Hamdan, F.F.; Xiong, L.; Piton, A.; Spiegelman, D.; Henrion, É.; Millet, B.; Fathalli, F.; et al. Rare mutations in N-methyl-D-aspartate glutamate receptors in autism spectrum disorders and schizophrenia. Transl. Psychiatry 2011, 1, e55–e55. [Google Scholar] [CrossRef]
- Tarnowska, K.; Gruczyńska–Sękowska, E.; Kowalska, D.; Majewska, E.; Kozłowska, M.; Winkler, R. The opioid excess theory in autism spectrum disorders - is it worth investigating further? Crit. Rev. Food Sci. Nutr. 2021, 63, 3980–3993. [Google Scholar] [CrossRef]
- Tartaglione, A.; Camoni, L.; Calamandrei, G.; Chiarotti, F.; Venerosi, A. The contribution of environmental pollutants to the risk of autism and other neurodevelopmental disorders: A systematic review of case-control studies. Neurosci. Biobehav. Rev. 2024, 164, 105815. [Google Scholar] [CrossRef]
- Terrabuio, E.; Zenaro, E.; Constantin, G. The role of the CD8+ T cell compartment in ageing and neurodegenerative disorders. Front. Immunol. 2023, 14, 1233870. [Google Scholar] [CrossRef]
- Tetreault, N.A.; Hakeem, A.Y.; Jiang, S.; Williams, B.A.; Allman, E.; Wold, B.J.; Allman, J.M. Microglia in the Cerebral Cortex in Autism. J. Autism Dev. Disord. 2012, 42, 2569–2584. [Google Scholar] [CrossRef]
- Theofilopoulos, A.N.; Kono, D.H.; Baccala, R. The multiple pathways to autoimmunity. Nat. Immunol. 2017, 18, 716–724. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Tsilioni, I.; Patel, A.B.; Doyle, R. Atopic diseases and inflammation of the brain in the pathogenesis of autism spectrum disorders. Transl. Psychiatry 2016, 6, e844–e844. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Angelidou, A.; Alysandratos, K.-D.; Zhang, B.; Asadi, S.; Francis, K.; Toniato, E.; Kalogeromitros, D. Mast cell activation and autism. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2012, 1822, 34–41. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Doyle, R. Autism, Gut-Blood-Brain Barrier, and Mast Cells. J. Clin. Psychopharmacol. 2008, 28, 479–483. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Kavalioti, M.; Tsilioni, I. Mast Cells, Stress, Fear and Autism Spectrum Disorder. Int. J. Mol. Sci. 2019, 20, 3611. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Stewart, J.M.; Panagiotidou, S.; Melamed, I. Mast cells, brain inflammation and autism. Eur. J. Pharmacol. 2016, 778, 96–102. [Google Scholar] [CrossRef]
- Thomson, A.R.; Pasanta, D.; Arichi, T.; Puts, N.A. Neurometabolite differences in Autism as assessed with Magnetic Resonance Spectroscopy: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2024, 162, 105728. [Google Scholar] [CrossRef]
- Tillisch, K.; Wang, Z.; Kilpatrick, L.; Holschneider, D.P.; Mayer, E.A. Studying the Brain–Gut Axis with Pharmacological Imaging. Ann. New York Acad. Sci. 2008, 1144, 256–264. [Google Scholar] [CrossRef]
- Tillmann, T.; Gibson, A.R.; Scott, G.; Harrison, O.; Dominiczak, A.; Hanlon, P. Systems Medicine 2.0: Potential Benefits of Combining Electronic Health Care Records With Systems Science Models. J. Med Internet Res. 2015, 17, e64–e64. [Google Scholar] [CrossRef]
- Toledano, M.; Pittock, S.J. Autoimmune Epilepsy. Semin. Neurol. 2015, 35, 245–258. [Google Scholar] [CrossRef]
- Tomova, A.; Husarova, V.; Lakatosova, S.; Bakos, J.; Vlkova, B.; Babinska, K.; Ostatnikova, D. Gastrointestinal microbiota in children with autism in Slovakia. Physiol. Behav. 2015, 138, 179–187. [Google Scholar] [CrossRef]
- Toomey, R.; Kang, H.K.; Karlinsky, J.; Baker, D.G.; Vasterling, J.J.; Alpern, R.; Reda, D.J.; Henderson, W.G.; Murphy, F.M.; Eisen, S.A. Mental health of US Gulf War veterans 10 years after the war. Br. J. Psychiatry 2007, 190, 385–393. [Google Scholar] [CrossRef]
- Tordjman, S.; Celume, M.; Denis, L.; Motillon, T.; Keromnes, G. Reframing schizophrenia and autism as bodily self-consciousness disorders leading to a deficit of theory of mind and empathy with social communication impairments. Neurosci. Biobehav. Rev. 2019, 103, 401–413. [Google Scholar] [CrossRef]
- Tordjman, S.; Somogyi, E.; Coulon, N.; Kermarrec, S.; Cohen, D.; Bronsard, G.; Bonnot, O.; Weismann-Arcache, C.; Botbol, M.; Lauth, B.; et al. Gene-Environment Interactions in Autism Spectrum Disorders: Role of Epigenetic Mechanisms. Front. Psychiatry 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.B.; Denisova, K. Motor noise is rich signal in autism research and pharmacological treatments. Sci. Rep. 2016, 6, 37422. [Google Scholar] [CrossRef]
- Torres, E.B.; Varkey, H.; Vero, J.; London, E.; Phan, H.; Kittler, P.; Gordon, A.; E Delgado, R.; Delgado, C.F.; A Simpson, E.; et al. Sensing echoes: temporal misalignment in auditory brainstem responses as the earliest marker of neurodevelopmental derailment. PNAS Nexus 2023, 2, pgac315. [Google Scholar] [CrossRef]
- Tuchman, R.; Moshé, S.L.; Rapin, I. Convulsing toward the pathophysiology of autism. Brain Dev. 2008, 31, 95–103. [Google Scholar] [CrossRef]
- Tzang, R.-F.; Chang, C.-H.; Chang, Y.-C.; Lane, H.-Y. Autism Associated With Anti-NMDAR Encephalitis: Glutamate-Related Therapy. Front. Psychiatry 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Üçok, A.; Gaebel, W. Side effects of atypical antipsychotics: a brief overview. World Psychiatry 2008, 7, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P. Neural synchrony in cortical networks: history, concept and current status. Front. Integr. Neurosci. 2009, 3, 17. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Neural Synchrony in Brain Disorders: Relevance for Cognitive Dysfunctions and Pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Abnormal neural oscillations and synchrony in schizophrenia. Nat. Rev. Neurosci. 2010, 11, 100–113. [Google Scholar] [CrossRef]
- Ullah, F.; Kaelber, D.C. Using Large Aggregated De-Identified Electronic Health Record Data to Determine the Prevalence of Common Chronic Diseases in Pediatric Patients Who Visited Primary Care Clinics. Acad. Pediatr. 2021, 21, 1084–1093. [Google Scholar] [CrossRef]
- Usui, N.; Kobayashi, H.; Shimada, S. Neuroinflammation and Oxidative Stress in the Pathogenesis of Autism Spectrum Disorder. Int. J. Mol. Sci. 2023, 24, 5487. [Google Scholar] [CrossRef]
- Vakilzadeh, G.; Martinez-Cerdeño, V. Pathology and Astrocytes in Autism. Neuropsychiatr. Dis. Treat. 2023, ume 19, 841–850. [Google Scholar] [CrossRef]
- Vargas, D.L.; Nascimbene, C.; Krishnan, C.; Zimmerman, A.W.; Pardo, C.A. Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann. Neurol. 2005, 57, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubramanian, G. Understanding Schizophrenia as a Disorder of Consciousness: Biological Correlates and Translational Implications from Quantum Theory Perspectives. Clin. Psychopharmacol. Neurosci. 2015, 13, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, R.; Schymanski, E.L.; Barabási, A.-L.; Miller, G.W. The exposome and health: Where chemistry meets biology. Science 2020, 367, 392–396. [Google Scholar] [CrossRef]
- Vicedo, M. Moving beyond the search for the first discoverer of autism. Front. Psychiatry 2024, 15, 1266486. [Google Scholar] [CrossRef]
- Victoria, H.K.; Caldwell, C. Breathwork in body psychotherapy: Clinical applications. Body, Mov. Dance Psychother. 2013, 8, 216–228. [Google Scholar] [CrossRef]
- Vineis, P.; Robinson, O.; Chadeau-Hyam, M.; Dehghan, A.; Mudway, I.; Dagnino, S. What is new in the exposome? Environ. Int. 2020, 143, 105887. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or Adaptive Immunity? The Example of Natural Killer Cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Vojdani, A.; Mumper, E.; Granpeesheh, D.; Mielke, L.; Traver, D.; Bock, K.; Hirani, K.; Neubrander, J.; Woeller, K.N.; O'HAra, N.; et al. Low natural killer cell cytotoxic activity in autism: The role of glutathione, IL-2 and IL-15. J. Neuroimmunol. 2008, 205, 148–154. [Google Scholar] [CrossRef]
- Volkmar, F.R.; Nelson, D.S. Seizure Disorders in Autism. J. Am. Acad. Child Adolesc. Psychiatry 1990, 29, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Volterra, A.; Meldolesi, J. Astrocytes, from brain glue to communication elements: the revolution continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Wakefield, A.J. The Gut–Brain Axis in Childhood Developmental Disorders. J. Pediatr. Gastroenterol. Nutr. 2002, 34, S14–S17. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, A.J.; Puleston, J.M.; Montgomery, S.M.; Anthony, A.; O'LEary, J.J.; Murch, S.H. The concept of entero-colonic encephalopathy, autism and opioid receptor ligands. Aliment. Pharmacol. Ther. 2002, 16, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Walsh, L.; Hill, C.; Ross, R.P. Impact of glyphosate (Roundup TM ) on the composition and functionality of the gut microbiome. Gut Microbes 2023, 15, 2263935. [Google Scholar] [CrossRef]
- Wan, M.; Ding, L.; Wang, D.; Han, J.; Gao, P. Serotonin: A Potent Immune Cell Modulator in Autoimmune Diseases. Front. Immunol. 2020, 11, 186. [Google Scholar] [CrossRef]
- Wang, T.; Sternes, P.R.; Guo, X.-K.; Zhao, H.; Xu, C.; Xu, H. Autoimmune diseases exhibit shared alterations in the gut microbiota. Rheumatology 2023, 63, 856–865. [Google Scholar] [CrossRef]
- Ward, J.H.; Weir, E.; Allison, C.; Baron-Cohen, S. Increased rates of chronic physical health conditions across all organ systems in autistic adolescents and adults. Mol. Autism 2023, 14, 1–20. [Google Scholar] [CrossRef]
- Ward, L.M. The thalamic dynamic core theory of conscious experience. Conscious. Cogn. 2011, 20, 464–486. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.P.; Foster, A.; Margaretten, N.C. Reduced Natural Killer Cell Activity in Autism. J. Am. Acad. Child Adolesc. Psychiatry 1987, 26, 333–335. [Google Scholar] [CrossRef]
- Wegiel, J.; Chadman, K.; London, E.; Wisniewski, T.; Wegiel, J. Contribution of the serotonergic system to developmental brain abnormalities in autism spectrum disorder. Autism Res. 2024, 17, 1300–1321. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the Regulation of Innate and Adaptive Immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Weitlauf, A.S.; Gotham, K.O.; Vehorn, A.C.; Warren, Z.E. Brief Report: DSM-5 “Levels of Support:” A Comment on Discrepant Conceptualizations of Severity in ASD. J. Autism Dev. Disord. 2013, 44, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Alshikho, M.J.; Herbert, M.R.; Hu, V.W. Pathway Network Analyses for Autism Reveal Multisystem Involvement, Major Overlaps with Other Diseases and Convergence upon MAPK and Calcium Signaling. PLOS ONE 2016, 11, e0153329–e0153329. [Google Scholar] [CrossRef]
- Wen, Y.; Herbert, M.R. Connecting the dots: Overlaps between autism and cancer suggest possible common mechanisms regarding signaling pathways related to metabolic alterations. Med Hypotheses 2017, 103, 118–123. [Google Scholar] [CrossRef]
- Werling, D.M. The role of sex-differential biology in risk for autism spectrum disorder. Biol. Sex Differ. 2016, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.; Dawson, G. Validation of the Phenomenon of Autistic Regression Using Home Videotapes. Arch. Gen. Psychiatry 2005, 62, 889–895. [Google Scholar] [CrossRef]
- West, L.; Brunssen, S.H.; Waldrop, J. Review of the Evidence for Treatment of Children with Autism with Selective Serotonin Reuptake Inhibitors. J. Spéc. Pediatr. Nurs. 2009, 14, 183–191. [Google Scholar] [CrossRef]
- West, L.; Waldrop, J.; Brunssen, S. Pharmacologic Treatment for the Core Deficits and Associated Symptoms of Autism in Children. J. Pediatr. Heal. Care 2009, 23, 75–89. [Google Scholar] [CrossRef]
- White, J.F. Intestinal Pathophysiology in Autism. Exp. Biol. Med. 2003, 228, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, P.; Carr, K.; Shattock, P. Is Autism Inborn And Lifelong For Everyone? Neuropsychiatr. Dis. Treat. 2019, ume 15, 2885–2891. [Google Scholar] [CrossRef]
- Wild, C.P. Complementing the Genome with an “Exposome”: The Outstanding Challenge of Environmental Exposure Measurement in Molecular Epidemiology. Cancer Epidemiology Biomarkers Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef]
- Willison, H.J.; Jacobs, B.C.; A van Doorn, P. Guillain-Barré syndrome. Lancet 2016, 388, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Wills, S.; Cabanlit, M.; Bennett, J.; Ashwood, P.; Amaral, D.G.; Van de Water, J. Detection of autoantibodies to neural cells of the cerebellum in the plasma of subjects with autism spectrum disorders. Brain, Behav. Immun. 2008, 23, 64–74. [Google Scholar] [CrossRef]
- Witkamp, R.F.; van Norren, K. Let thy food be thy medicine….when possible. Eur. J. Pharmacol. 2018, 836, 102–114. [Google Scholar] [CrossRef]
- Wood, J.D.; Alpers, D.H.; Andrews, P.L.R. Fundamentals of neurogastroenterology. Gut 1999, 45, ii6–ii16. [Google Scholar] [CrossRef]
- Wood, J.J.; Kendall, P.C.; Wood, K.S.; Kerns, C.M.; Seltzer, M.; Small, B.J.; Lewin, A.B.; Storch, E.A. Cognitive Behavioral Treatments for Anxiety in Children With Autism Spectrum Disorder. JAMA Psychiatry 2020, 77, 474–483. [Google Scholar] [CrossRef]
- Wu, M.; Zheng, W.; Song, X.; Bao, B.; Wang, Y.; Ramanan, D.; Yang, D.; Liu, R.; Macbeth, J.C.; Do, E.A.; et al. Gut complement induced by the microbiota combats pathogens and spares commensals. Cell 2024, 187, 897–913.e18. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, J.; Li, Y. Microglia and astrocytes underlie neuroinflammation and synaptic susceptibility in autism spectrum disorder. Front. Neurosci. 2023, 17, 1125428. [Google Scholar] [CrossRef]
- Xu, G.; Snetselaar, L.G.; Jing, J.; Liu, B.; Strathearn, L.; Bao, W. Association of Food Allergy and Other Allergic Conditions With Autism Spectrum Disorder in Children. JAMA Netw. Open 2018, 1, e180279–e180279. [Google Scholar] [CrossRef]
- Xu, S.-C.; Zhong, Y.; Jiang, H.-Y.; Tang, J. Exposure to anti-seizure medication during pregnancy and the risk of autism and ADHD in offspring: a systematic review and meta-analysis. Front. Neurol. 2024, 15, 1440145. [Google Scholar] [CrossRef] [PubMed]
- Yavuz, B.R.; Arici, M.K.; Demirel, H.C.; Tsai, C.-J.; Jang, H.; Nussinov, R.; Tuncbag, N. Neurodevelopmental disorders and cancer networks share pathways, but differ in mechanisms, signaling strength, and outcome. npj Genom. Med. 2023, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.W.K.; Heinrich, M.; Kijjoa, A.; Tzvetkov, N.T.; Atanasov, A.G. The ethnopharmacological literature: An analysis of the scientific landscape. J. Ethnopharmacol. 2020, 250, 112414. [Google Scholar] [CrossRef]
- Yngve, M.; Lidström, H. Implementation of information and communication technology to facilitate participation in high school occupations for students with neurodevelopmental disorders. Disabil. Rehabilitation: Assist. Technol. 2023, 19, 2017–2025. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W.; Rivest, S. Taking Advantage of the Systemic Immune System to Cure Brain Diseases. Neuron 2009, 64, 55–60. [Google Scholar] [CrossRef]
- Zanchi, M.M.; Marins, K.; Zamoner, A. Could pesticide exposure be implicated in the high incidence rates of depression, anxiety and suicide in farmers? A systematic review. Environ. Pollut. 2023, 331, 121888. [Google Scholar] [CrossRef]
- Zandman-Goddard, G.; Peeva, E.; Shoenfeld, Y. Gender and autoimmunity. Autoimmun. Rev. 2007, 6, 366–372. [Google Scholar] [CrossRef]
- Zarate-Lopez, D.; Torres-Chávez, A.L.; Gálvez-Contreras, A.Y.; Gonzalez-Perez, O. Three Decades of Valproate: A Current Model for Studying Autism Spectrum Disorder. Curr. Neuropharmacol. 2024, 22, 260–289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M. , & An, J. ( 45(2), 27–37. [CrossRef]
- Zhang, S.; Han, F.; Wang, Q.; Fan, F. Probiotics and Prebiotics in the Treatment of Autism Spectrum Disorder: A Narrative Review. J. Integr. Neurosci. 2024, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. CD4 T cells: fates, functions, and faults. Blood 2008, 112, 1557–1569. [Google Scholar] [CrossRef]
- Ziv, Y.; Schwartz, M. Immune-based regulation of adult neurogenesis: Implications for learning and memory. Brain, Behav. Immun. 2008, 22, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Zota, A.R.; Riederer, A.M.; Ettinger, A.S.; A Schaider, L.; Shine, J.P.; Amarasiriwardena, C.J.; O Wright, R.; Spengler, J.D. Associations between metals in residential environmental media and exposure biomarkers over time in infants living near a mining-impacted site. J. Expo. Sci. Environ. Epidemiology 2015, 26, 510–519. [Google Scholar] [CrossRef]
- Zürcher, N.R.; Bhanot, A.; McDougle, C.J.; Hooker, J.M. A systematic review of molecular imaging (PET and SPECT) in autism spectrum disorder: Current state and future research opportunities. Neurosci. Biobehav. Rev. 2015, 52, 56–73. [Google Scholar] [CrossRef]
Figure 1.
Disorders of the Neuroimmune System & ASD.
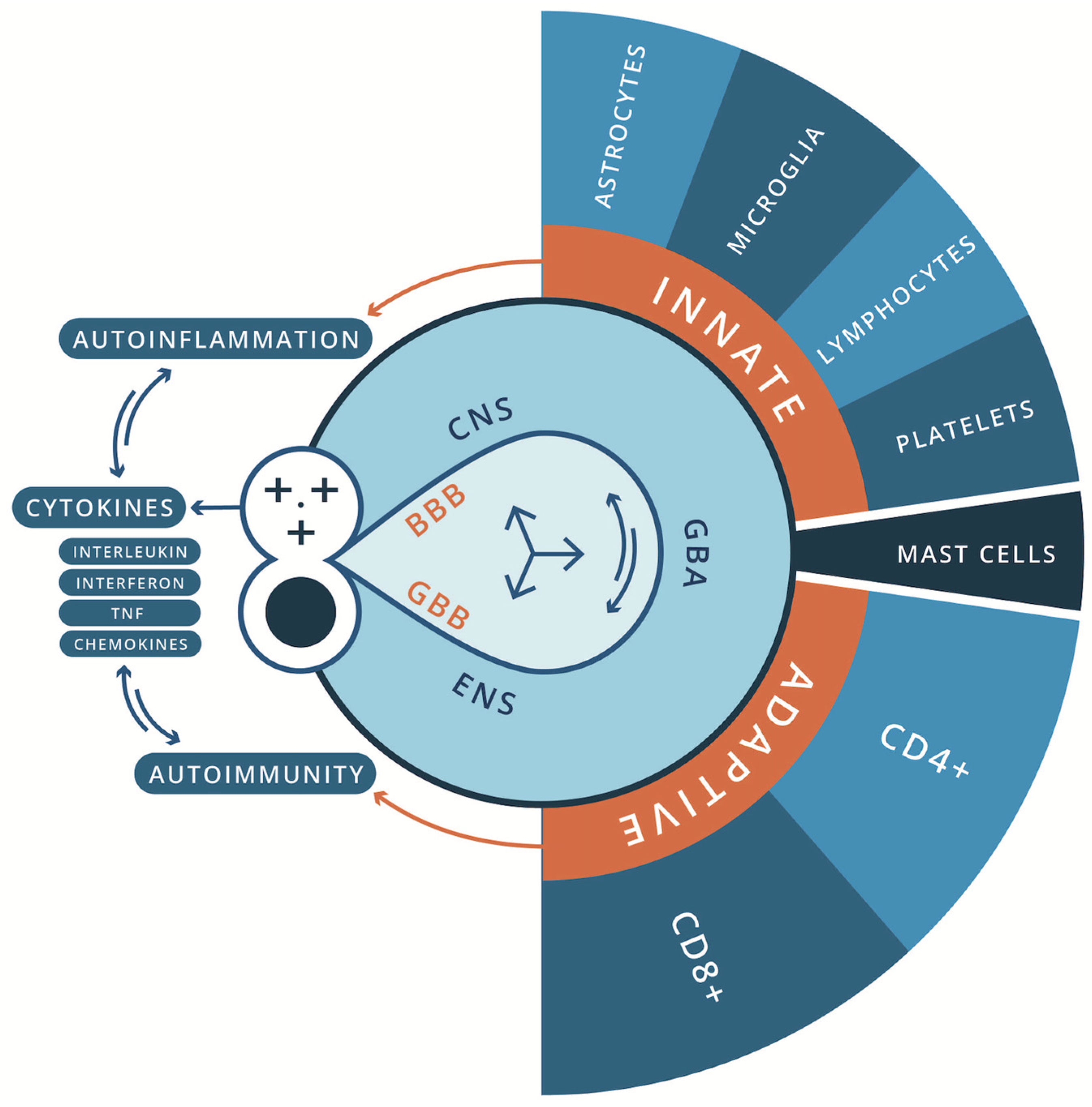
Figure 2.
ASD from Different Hierarchical Levels of Biological Resolution.

Figure 3.
Omics; Nutrigenomics & Bioregulatory Systems Medicine for ASD.
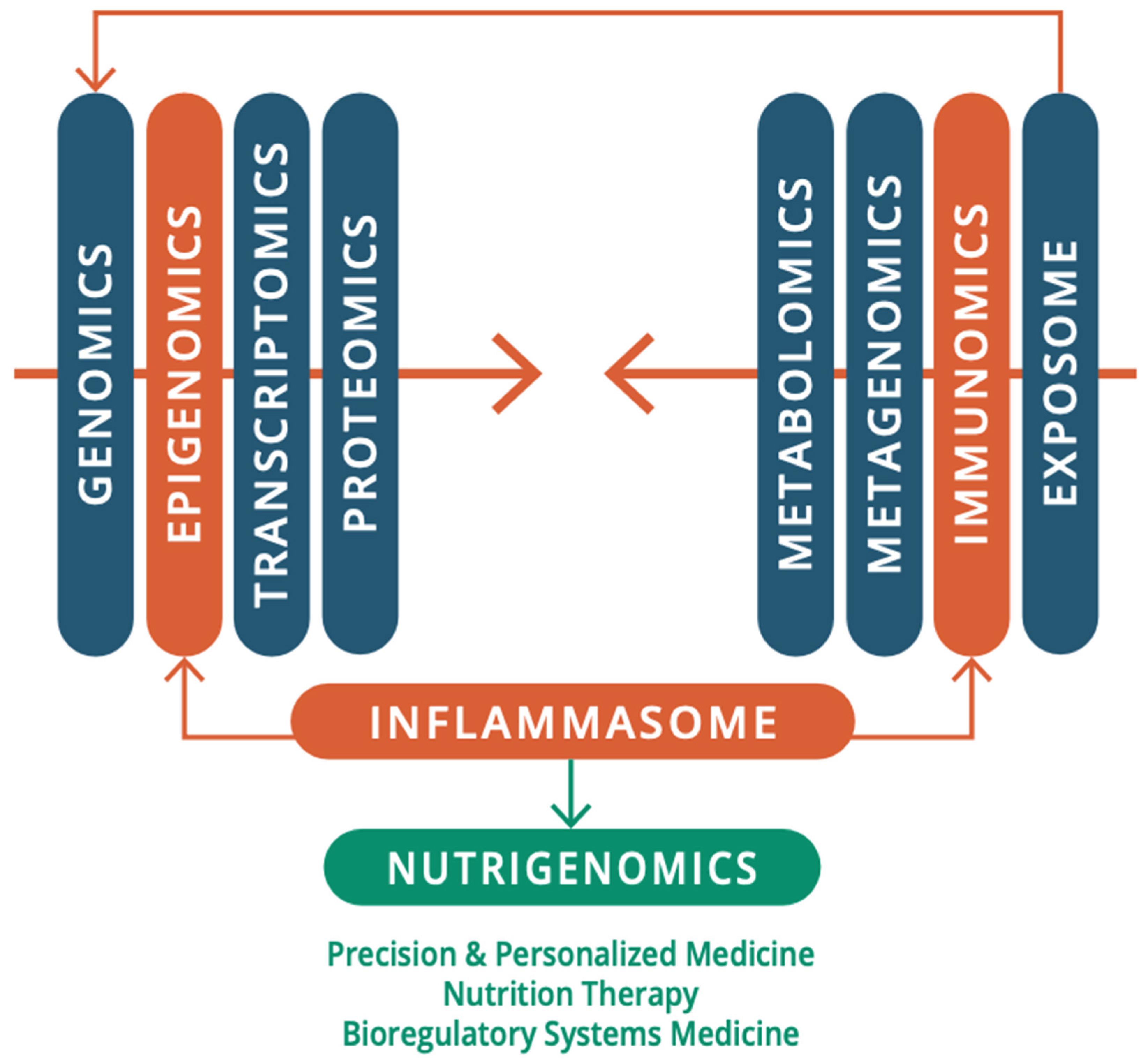
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



