
Submitted:
07 November 2024
Posted:
08 November 2024
You are already at the latest version
Abstract
The increasing antibiotic resistance among bacteria challenges the biotech industry to search for new antibacterial molecules. Endolysin TP84_28 is a thermostable, lytic enzyme, encoded by the bacteriophage TP-84, and it effectively digests host bacteria cell wall. Biofilms, together with antibiotic resistance, are major problems in clinical medicine and industry. The primary challenge is to keep antibacterial molecules at the site of desired action, as their diffusion leads to loss of efficacy. The gene encoding TP84_28 endolysin was cloned into a custom expression-fusion vector, forming a fusion gene cbd_tp84_28_his with a cellulose binding domain from the cellulase enzyme. The CBD_TP84_28_His protein was biosynthesized in Escherichia coli and purified. Thermostability and enzymatic activity against various bacterial species were measured by turbidity reduction assay, spot assay, and biofilms removal. Cellulose binding properties were confirmed via interaction with microcellulose and cellulose paper-based immunoblotting. The high affinity of CBD allowed for high concentration of the fusion enzyme at desired target sites such as cellulose-based wound dressings, artificial heart valves and food packaging. CBD_TP84_28_His exhibits lytic effect against thermophilic bacteria Geobacillus stearothemophilus, Thermus aquaticus, Bacillus stearothermophilus and minor effects against mesophilic Bacillus cereus, Bacillus subtilis. CBD_TP84_28_His retains full activity after preincubation in the temperatures of 30-65°C and exhibits significant activity up to its melting point at 73°C. CBD_TP84_28_His effectively reduces biofilms. These findings suggest that integrating CBDs into thermostable endolysins could enable the development of targeted antibacterial recombinant proteins with diverse clinical and industrial applications.
Keywords:
thermophage
; bacteriophage
; TP-84
; endolysin
; cellulose-binding domain
; antimicrobial
1. Introduction
Antibiotic resistance is growing worldwide, and there are at least 2.8 million antimicrobial-resistant infections causing as many as 35,000 deaths yearly in the USA alone [1]. Therefore, innovative approaches employing bacteriophages (phage therapy) and their enzymes to fight bacterial infections are gaining increasing attention. Endolysins are bacteriophage-produced lytic proteins, vital for exiting the host cell. Many endolysins have been tested in vitro, providing promising results. To date, none of them have been registered as a drug, but clinical research on the safety and efficacy of endolysins is ongoing [2]. One of the endolysins, Staphefekt™, is available in Gladskin cosmetics, which are registered as medical devices Endolysins are promising candidates for replacing or complementing traditional antibiotics. Most endolysins target highly conserved bonds within peptidoglycan, making it very difficult for bacteria to evolve endolysin-resistance. Exposing bacteria to low endolysins concentrations shows that endolysin-resistance development is a very rare event [4,5].
Due to the significant variation in the chemical structure of peptidoglycan in gram-positive bacteria, the presence of peptidase catalytic and binding domains in gram-positive specific endolysins often leads to the relatively narrow host range. In contrast, endolysins from gram-negative specific bacteriophages are small globular proteins usually lacking a binding domain [6]. They have a broader activity range, which is consistent with a relatively conserved structure of peptidoglycan across gram-negative bacteria [7].
Four types of bacterial cell wall-lytic enzymes can be distinguished, based on the different types of peptidoglycans they target:
- (i)
- glycosidases, cleaving the bonds between N-acetylglucosamine and N-acetylmuramic acid, including two subgroups: N-acetylmuramidases and N-acetylglucosaminidases
- (ii)
- transglucosylases, cleaving bonds between N-acetylmuramic acid and N-acetylglucosamine (like N-acetylmuramidases), but in a different mechanism; they do not require water, thus are not considered hydrolyses
- (iii)
- amidases, cleaving the amide bond between N-acetylmuramic acid and L-alanine, the first amino acid in the cross-linking peptide
- (iv)
Bacteriophage TP-84 was discovered in 1952 when L. Leon Campbell isolated it from the greenhouse soil using Bacillus (B.) stearothermophilus strain 2184 [12]. This bacterium was recently reclassified as Geobacillus (G.) stearothermophilus. The closely related G. stearothermophilus strain 10 was further used as a host, since TP-84 exhibits a narrow host range, limited to several B. stearothermophilus and G. stearothermophilus strains [13,14]. We have also sequenced its genome, and we proposed to rename it G. thermoleovorans strain 10 [15]. However, before publication of the full genomic sequence, for clarity we use here the name G. stearothermophilus strain 10.
Bacteriophage TP-84 exhibits an extraordinarily wide temperature growth range of 30-80°C, covering both mesophilic and thermophilic range, with optimum at 55-65°C. TP-84 has the capability to proliferate within the entire temperature range, although with widely varying efficiency. We have sequenced the TP-84 genome and bioinformatically characterised its 81 ORFs, 73 of which were confirmed by proteomics analysis [16,17]. Three of those, TP84_26 glycosylase-depolymerase, TP84_27 holin and TP84_28 endolysin were assigned as hypothetical lytic proteins. TP-84 belongs to the family Siphoviridae, and we have shown that it is a genomic orphan with very little sequence similarity to any bacterial or viral genomes at the time of analysis in 2018 [16]. This makes TP-84 an interesting source of novel proteins, which may exhibit properties not observed in those derived from other species [11]. Bioinformatic analysis of the tp84_28 gene has revealed an enzymatic catalytic domain GH25_Lyc-like (21-206 aa) [cd06525 - protein blast alignment on-line 16.10.2024]. Furthermore, there are three LysM superfamily domains [cl21525 - protein BLAST alignment on-line 16.10.2024] responsible for peptidoglycan binding, which have also been seen in proteins exhibiting functions other than lysis [11].
We have cloned and expressed the tp84_28 gene and determined that TP84_28 is a thermostable protein with melting point at 77.6°C, which coincides with its loss of lytic activity. The protein is active in a wide range of pH: from 4.0 to 10.0, with maximum at 7.5-8.0. In this work, we have constructed and evaluated a bioactive fusion protein, consisting of TP84_28 endolysin and Cellulose Binding Domain (CBD). CBDs in general are substantially autonomous domains of cellulases, still capable of binding to cellulose if separated from cellulase’s catalytic domains [18]. This feature is retained in a number of fusion protein constructs, where the cellulase catalytic domain was replaced by another protein.
2. Results
2.1. Cloning, Overproduction, and Purification of Recombinant Fusion Endolysin CBD_TP84_28_His
The pET28_delSapI_CBD_His (Supplementary file S1) expression clones coding for recombinant fusion endolysin CBD_TP84_28_His were stably maintained in various E. coli strains (not shown). However, IPTG-induced cultures of E. coli BL21(DE3) pET28_CBD_TP84_28_His exhibited slower growth rate as compared to both uninduced culture and control E. coli BL21(DE3) culture, devoid of pET28_CBD_TP84_28_His plasmid (Supplementary file S2). This indicates moderate toxicity to the E. coli host, similar to what we have observed during expression of recombinant, but not-fused TP84_28_His endolysin [11]. The simple purification protocol, which included removal of nucleic acids and acidic protein by PEI precipitation, followed by metal affinity chromatography on a NiNTA HisTrap HP column, enabled obtaining nearly homogeneous CBD_TP84_28_His (over 95%) (Figure 1). The purified preparation was stored at 4℃ for more than 3 months, and at -80℃ with added glycerol (10%) for over 6 months, without loss of activity. The final yield was 2 mg (final concentration 0.5 mg/ml) of enzymatically active recombinant protein from 1 L of the expression culture.
2.2. Properties of the Recombinant Fusion Endolysin CBD_TP84_28_His
2.2.1. Lytic Activity of the Recombinant Fusion Endolysin CBD_TP84_28_His Against Test Bacteria
Two methods of testing lytic activity of CBD_TP84_28_His against bacteria were used: qualitative spot assay (diffusion test) and quantitative TRA. Spot assay results obtained for bacteria incubated at 42°C have shown no lytic effect of CBD_TP84_28_His against B. subtilis. Minor lytic effect was observed against E. coli and moderate lytic effect was observed in the case of B. cereus. Considerable lytic effect was visible for G. stearothermophilus strain 10, T. aquaticus, Geobacillus ICI, B. stearothermophilus (Figure 2). To exclude the influence of incubation temperature on the activity of CBD_TP84_28_His, its lytic potency was also assessed via TRA. After centrifugation and resuspension in buffer R, each tested bacterial strain was subjected to digestion by CBD_TP84_28_His at 55°C (Figure 3). Reduction of resuspended bacteria OD600 upon addition of CBD_TP84_28_His progressed rapidly for T. aquaticus, B. stearothermophilus, Geobacillus ICI and G. stearothermophilus strain 10. Minor lytic effect was observed in case of B. cereus and B. subtilis, and negligible lytic effect comparable to the control was observed for E. coli.
2.2.2. Activity of the Recombinant Fusion Endolysin CBD_TP84_28_His in Comparison to Recombinant Endolysin TP84_28_His
To evaluate the effect of fusing CBD to endolysin TP84_28_His on the lytic activity, equimolar amounts of fusion recombinant CBD_TP84_28_His and recombinant TP84_28_His were used to perform TRA. The results indicated that the endolysin TP84_28_His digests G. stearothermophilus, strain 10 2-3 times faster than the bimodular CBD_TP84_28_His in the linear activity range between 5 and 10 minutes (Figure 4).
2.2.3. Lytic Activity of the Recombinant Fusion Endolysin CBD_TP84_28_His Against Biofilm
- Inhibition of the biofilm formation by G. stearothermophilus strain 10
This assay was conducted in microtiter plate format for quantitative determination of required CBD_TP84_28_His amounts for efficient biofilm digestion. This was conducted by adding various amounts of CBD_TP84_28_His to biofilms formed in a microtiter plate. 0.05 µg of CBD_TP84_28_His reduced biofilm formation by G. stearothermophilus strain 10 by 10%, adding 0.25 µg reduced it by 59%, 0.5 µg by 79% and 5 µg by 96% and 50 µg by a 98% reduction in biofilm formation, respectively (Figure 5).
- Inhibition of biofilm formation by mesophilic pathogenic-related bacteria
The effect of endolysin CBD_TP84_28_His on the formation of mesophilic bacterial biofilms was investigated. For safety reasons, these were formed from bacterial strains closely related to real pathogenic bacteria, as the latter present serious risks. Endolysin CBD_TP84_28_His showed the greatest inhibition of biofilm formation by S. aureus (as much as 81%); for P. aeruginosa, it was 42%, for E. coli 39%, and for S. enteritidis. and B. cereus, the inhibition of biofilm formation was slightly above 30% (Table 1).
2.2.4. Thermostability of the Recombinant Fusion Endolysin CBD_TP84_28_His
Thermostability of CBD_TP84_28_His was evaluated through 13 temperature values in the range of 30-100°C and autoclaving for 30 min in the buffer R, followed by TRA assay. Similar rates of reaction were observed for samples preincubated at the temperature range 30-65.1°C. A significant reduction in reaction rate was noted for the sample preincubated at 70°C, and further reduction in reaction rate was observed for the sample incubated at the temperature of 74.8°C (Figure 6). A further increase in the preincubation temperature caused essentially the same results as for 74.8°C. Measurements on Prometheus Panta device confirmed those findings: the protein starts to unfold at 64.98°C, and the melting point is 73.06°C.
2.2.5. Cellulose- Binding Properties of the Recombinant Fusion Endolysin CBD_TP84_28
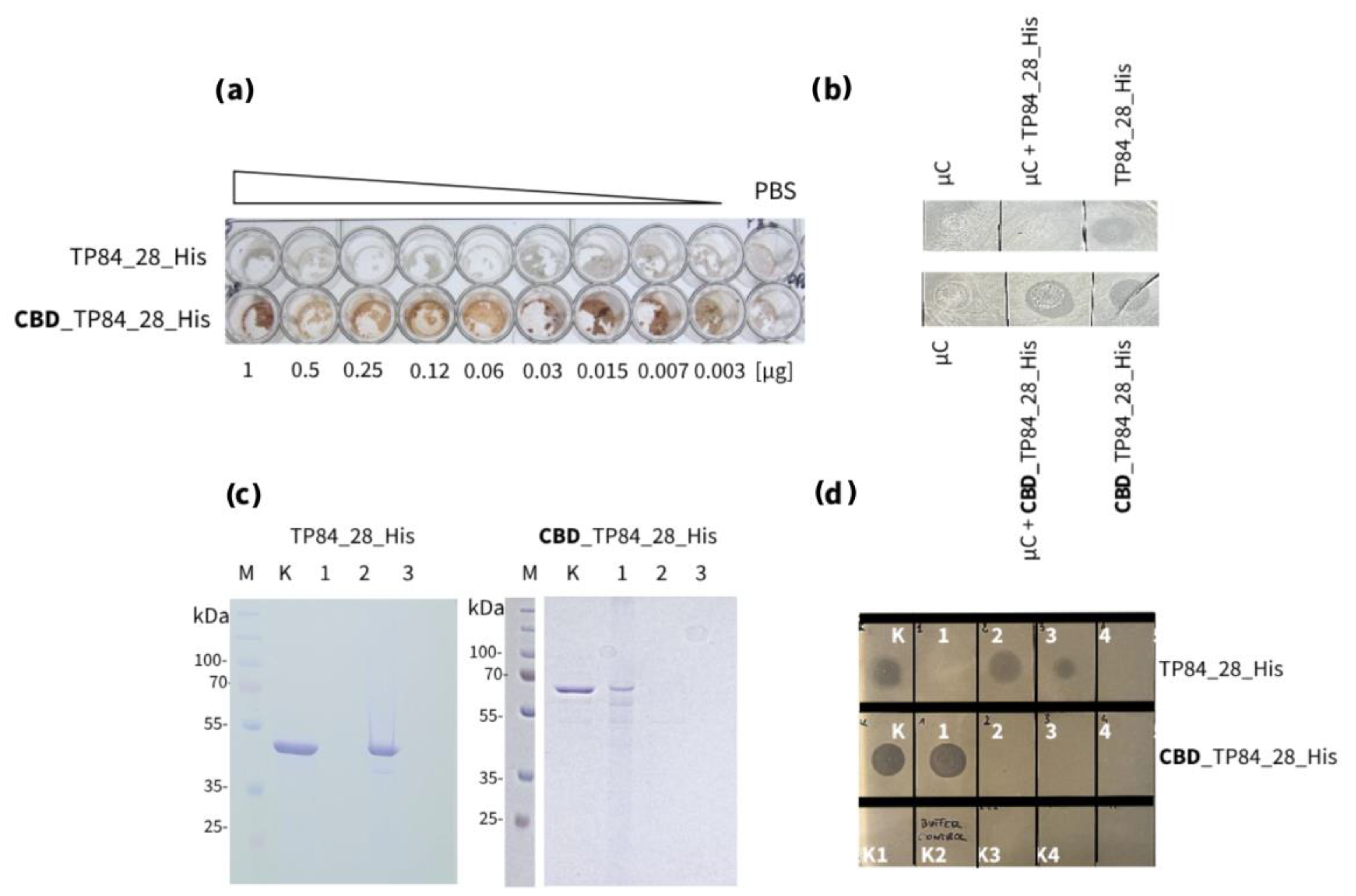
- Interaction with microcellulose (µC) assayFigure 7. Interactions comparison of recombinant endolysin TP84_28_His and recombinant fusion endolysin CBD_TP84_28_His with µC. Panel (a) Binding of TP84_28_His and CBD_TP84_28_His to µC in a 96-well plate using detection of His-tag for Western blots (Method 2.2.3.1). Both proteins have a His-tag that is located at the C-terminus, which allows for detection of the protein attached to the µC using anti-His antibodies. Panel (b) Dot blot assay of endolysin activity using the host G. stearothermophilus strain 10. A control - µC i 1 x PBS dot applied, µC complex formed with each of the enzymes: TP84_28_His and CBD_TP84_28_His were applied to the agar plate with spread G. stearothermophilus strain 10. Panel (c) SDS-PAGE comparative analysis of the formation of insoluble µC complexes with CBD_TP84_28 and with TP84_28_His. Lane M, PageRuler Plus Stained Protein Ladder; lane K, untreated TP84_28_His (left)/CBD_TP84_28 (right); lane 1, insoluble µC complex formed with TP84_28_His (left)/CBD_TP84_28_His (right); lane 2, supernatants of complexes formation – unbound protein; lane 3, washing supernatant of the complexes with 1 x PBS. Panel (d) µC complex formed with each of the enzymes: TP84_28_His and CBD_TP84_28_His were applied to the agar plate with spread G. stearothermophilus strain 10 to measure µC complexes activity; dot K, TP84_28_His/CBD_TP84_28_His; dot 1, formed and washed µC complexes with TP84_28_His/CBD_TP84_28_His; dot 2, supernatants of insoluble µC complexes formation; dot 3, washes of the complexes with 1 x PBS; dots 4, first washes of the complex with water.Figure 7. Interactions comparison of recombinant endolysin TP84_28_His and recombinant fusion endolysin CBD_TP84_28_His with µC. Panel (a) Binding of TP84_28_His and CBD_TP84_28_His to µC in a 96-well plate using detection of His-tag for Western blots (Method 2.2.3.1). Both proteins have a His-tag that is located at the C-terminus, which allows for detection of the protein attached to the µC using anti-His antibodies. Panel (b) Dot blot assay of endolysin activity using the host G. stearothermophilus strain 10. A control - µC i 1 x PBS dot applied, µC complex formed with each of the enzymes: TP84_28_His and CBD_TP84_28_His were applied to the agar plate with spread G. stearothermophilus strain 10. Panel (c) SDS-PAGE comparative analysis of the formation of insoluble µC complexes with CBD_TP84_28 and with TP84_28_His. Lane M, PageRuler Plus Stained Protein Ladder; lane K, untreated TP84_28_His (left)/CBD_TP84_28 (right); lane 1, insoluble µC complex formed with TP84_28_His (left)/CBD_TP84_28_His (right); lane 2, supernatants of complexes formation – unbound protein; lane 3, washing supernatant of the complexes with 1 x PBS. Panel (d) µC complex formed with each of the enzymes: TP84_28_His and CBD_TP84_28_His were applied to the agar plate with spread G. stearothermophilus strain 10 to measure µC complexes activity; dot K, TP84_28_His/CBD_TP84_28_His; dot 1, formed and washed µC complexes with TP84_28_His/CBD_TP84_28_His; dot 2, supernatants of insoluble µC complexes formation; dot 3, washes of the complexes with 1 x PBS; dots 4, first washes of the complex with water.
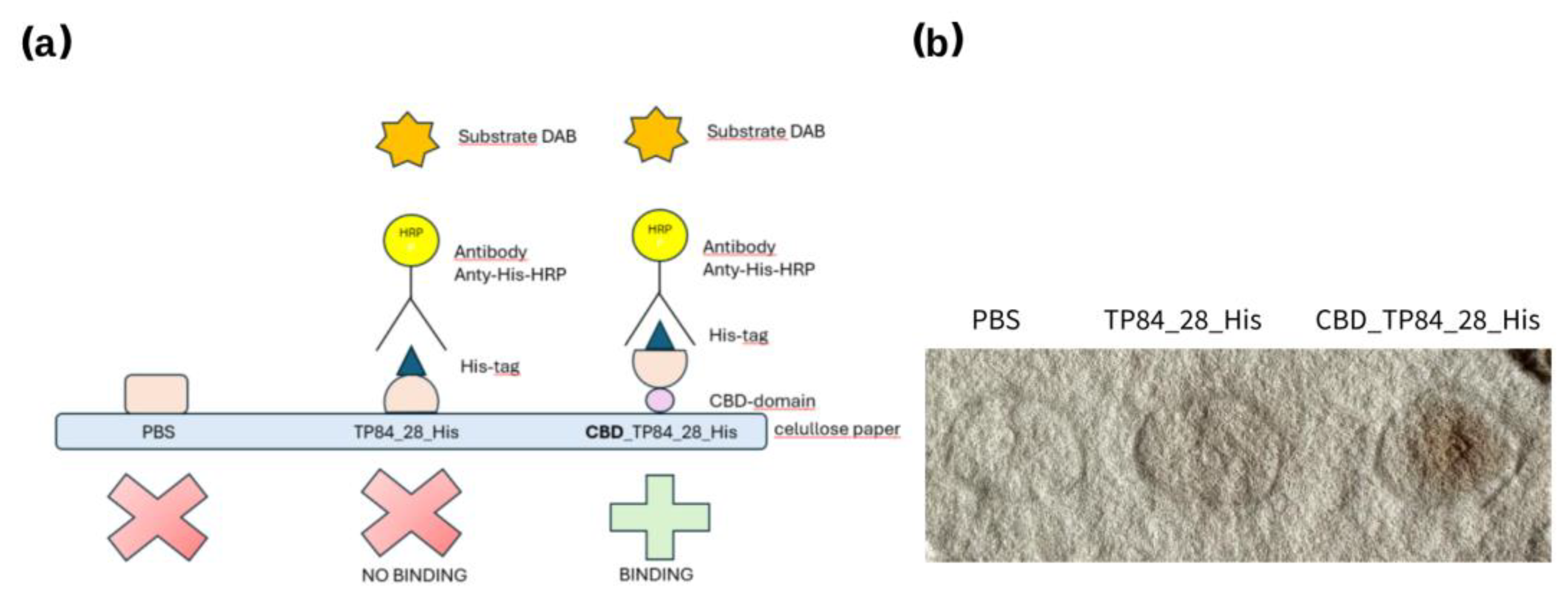
- Cellulose paper-based immunoblottingFigure 8. CBD_TP84_28_His interaction with cellulose paper: (a) scheme showing CBD-domain binding to cellulose filter paper and how His-Tag binds with anti-His-HRP antibody, which enables visualisation of the reaction after colour development using DAB. (b) cellulose filter paper with spots: PBS buffer (control), TP84_28_His and CBD_TP84_28_His.Figure 8. CBD_TP84_28_His interaction with cellulose paper: (a) scheme showing CBD-domain binding to cellulose filter paper and how His-Tag binds with anti-His-HRP antibody, which enables visualisation of the reaction after colour development using DAB. (b) cellulose filter paper with spots: PBS buffer (control), TP84_28_His and CBD_TP84_28_His.
3. Discussion
The functionalization of the thermostable endolysin TP84_28 with CBD for antimicrobial applications, such as cellulose-based wound dressings, offers distinct advantages over alternatives like T4 lysozyme fused with a CBD. The TP84_28 endolysin, derived from the bacteriophage TP-84, exhibits high thermostability, retaining activity at temperatures up to 77.6°C. This makes it suitable for environments that involve high heat exposure, deactivating other mesophilic enzymes like T4 lysozyme and thus eliminating their potential applications at high temperatures [11]. Recombinant fusion endolysin CBD_TP84_28_His was expressed in the T7 promoter-based expression system in E. coli BL21(DE3) Gold. The yield of 2 mg / 1 L culture was rather low due to the expected and apparent toxicity of the endolysin part of the fusion protein against E. coli. Synthesis of the fusion endolysin slowed bacterial growth by 35%. In contrast, when the enzyme was applied to E. coli from outside in the diffusion assay, it was not able to effectively lyse E. coli (Figure 2). One possible explanation of this phenomenon is that endolysin CBD_TP84_28_His can penetrate cytoplasmic membrane of E. coli cell and digest the bacterial peptidoglycan cell wall from inside, whereas the outer membrane forms a non-permeable barrier for the enzyme. There are profound differences between endolysins derived from bacteriophages targeting Gram-positive and Gram-negative bacteria due to the architecture of the cell wall [6]. It was also observed that P. aeruginosa, a Gram-negative species, was marginally susceptible to CBD_TP84_28_His. This corroborates with the finding that CBD_TP84_28_His exhibits highest lytic activity towards thermophilic Gram-positive bacteria related to the host of TP-84 bacteriophage, G. stearothermophilus strain 10, Geobacillus ICI and B. stearothermophilus, similarly as unfused TP84_28_His endolysin [11].
T. aquaticus, even though not related to Geobacillus, is also highly sensitive to CBD_TP84_28_His as it is Gram-positive and therefore without an external membrane (Figure 2 and Figure 3). However, the fact that activity against T. aquaticus was comparable to activity against the reference species G. stearothermophilus strain 10 points to an interesting, putative difference in enzymatic specificity when compared to unfused TP84_28_His, for which activity against T. aquaticus was moderate [11]. A possible reason for this finding may lie in the way the presence of CBD domain affects folding of the endolysin itself. Slight changes in conformation might shift catalytic preference to other peptidoglycan structural variants. In general, TP84_28 displays strong lytic activity against thermophilic Gram-positive bacteria, which are major culprits in wound infections. Its ability to target resistant bacteria, including those that form biofilms, is particularly advantageous in clinical settings, where biofilm-associated infections are a challenge. This property is further enhanced when coupled with a CBD, as the enzyme can anchor onto cellulose-based wound dressings, providing localized and sustained antibacterial action. Prolonged retention at the wound site ensures continuous antimicrobial activity.
The activity of CBD_TP84_28_His against mesophilic Gram-positive bacteria related to G. stearothermophilus strain 10 remains low. In the diffusion test, this may be attributed to the conditions favouring cell wall regeneration/bacterial growth (bacterial medium availability, temperature of 42°C) rather than lysis by the applied CBD_TP84_28_His, which has higher temperature optimum range. In TRA, however, to slow down the regeneration process, the buffer optimal for the enzyme CBD_TP84_28_His was used instead of media promoting bacterial growth. Additionally, the temperature was optimal for the enzyme, not for the bacteria (55°C), yet the enzyme’s activity against the mesophiles remained low.
The poor activity of CBD_TP84_28_His against mesophilic Bacillus spp. is somewhat surprising, as these are Gram-positive bacteria related to the G. stearothermophilus strain 10. This points to the conclusions that:
- (i)
- the specificity of the CBD_TP84_28_His appears to depend both on the thermophilicity and phylogenetic relatedness of the bacteria
- (ii)
- the structure of peptidoglycan varies substantially across Bacillus bacterial species
- (iii)
- the structure of peptidoglycan of the tested thermophiles has common features, sensitive to the CBD_TP84_28_His
- (iv)
- differences in external polysaccharide envelopes may play an important role, preventing the enzyme access to the cell wall.
Given the limited activity of CBD_TP84_28_His against mesophilic bacteria, T4 bacteriophage lysozyme could serve as a complementary or alternative enzyme to be mixed with CBD_TP84_28_His. However, even though the T4 lysozyme fused with a CBD acquires cellulose-binding capacity [19], it lacks the thermostability advantages of CBD_TP84_28_His.
Lytic activity of CBD_TP84_28_His against two common pathogenic strains was tested and found to be low (Gram-negative P. aeruginosa and Gram-positive S. aureus; data not shown); this is consistent with the conclusions above. Endolysins derived from Gram-positive targeting bacteriophages can be supported in accessing the cell wall of Gram-negative species in multiple ways. Pretreatments with high hydrostatic pressure, EDTA, or organic acids all weaken the outer membrane of Gram-negative bacteria [20,21], but these methods are not applicable in clinical settings. More promising approaches include combining endolysins with antibiotics for synergistic effect; packing endolysins into liposomes to help them cross the outer membrane; or combining them with silver nanoparticles to disrupt the outer membrane [20,21]. Modular proteins fused with an additional domain disrupting the outer membrane have also been reported [4,22].
Artilysins®, a concept by Briers et al. (2014) are endolysins with an additional polycationic nanopeptide dedicated to destabilisation of LPSs, a component of the outer membrane. Artilysins® are an antimicrobial technology platform and Lysando, the biotechnology company that develops the concept, claims that “it is possible to design specific Artilysin® to target nearly every bacterial species” [23].
Innolysins are endolysins fused with proteins binding to the receptors in the outer membrane of Gram-negative bacteria and they were reported by Zampara et al. (2020) to have lytic activity against Gram-negative P. aeruginosa [24]. It seems that expanding functionality of endolysins by adding new moieties and creating a fusion protein is one way to pursue the desired effect of the potential drug.
Although recombinant fusion endolysin CBD_TP84_28_His has not shown high activity against the tested pathogen-related bacteria responsible for nosocomial infections, it has shown lytic activity towards biofilm formed by all the bacteria tested, including highly pathogenic S. aureus (Table 1). Biofilm is a mixture of microbial cells, exopolysaccharides, DNA and proteins which serves as a protective milieu for the bacteria and so poses a major clinical problem. Somewhat surprisingly, the highest reduction in biofilm was observed for the S. aureus (81%), secondly for P. aeruginosa (42%) – both of which were strains of very low sensitivity to CBD_TP84_28_His in the diffusion test and TRA (data not shown). The main component of S. aureus biofilm and the only exopolysaccharide is polysaccharide intercellular adhesin (PIA, PNAG) – a partially deacetylated polymer of β-1,6-N-acetylglucosamine (Nguyen et al. 2020), suspected to be the target for CBD_TP84_28_His. PIA is also known to be produced by species such as E. coli, Acinetobacter baumanii, Yersinia pestis, Bordatella spp. (Yakandavala et. al, 2020). Therefore, CBD_TP84_28_His could be expected to demonstrate lytic activity against those species, but this requires further testing. The biofilm of P. aeruginosa is known to consist of several types of exopolysaccharides [25].
Biofilm formation is a serious problem associated with the long-term use of various medical devices and implants. Biofilm forms a barrier which can be impossible to lyse even with the application of broad-spectrum antibiotics [26]. Additionally, due to the effort to reduce the plastic burden, immunologically neutral cellulose-based materials are going to become increasingly popular in the construction of such equipment.
An example of a potential advanced application of CBD_TP84_28_His is treating infectious endocarditis: this life-threatening condition is often not susceptible to antibiotics due to biofilm formation on artificial heart valves. If innovative cellulose-based medical devices and implants, such as the artificial heart valves just patented by Piotr Siondalski and his team [27] become popular, CBD_TP84_28_His has a high potential to become an effective treatment for associated infections. Cellulose would serve as a scaffold for CBD_TP84_28_His, keeping the antibacterial activity highly localised and therefore highly efficient. At the same time, this overcomes the issues posed by systemic application linked to proteinaceous nature of the enzyme (i.e. evoking allergic reactions, and degrading before reaching target site). The biofilm-disrupting properties of CBD_TP84_28_His could also facilitate the efficacious application of life-saving antibiotics.
There are also promising prospective applications for CBD_TP84_28_His in food production: the industry will inevitably be switching to alternatives to plastic for packaging solutions, many of which are cellulose derivatives. Target genera of the fusion endolysin, Bacillus and Geobacillus, are responsible for spoilage of various types of food [28,29]. The CBD domain allows effective binding to cellulose food packaging material, immobilising the protein and preventing it from mixing with the content inside the package. This property is especially important, as using a preservative that is not immobilised would allow it to mix with the food contents, and thus require extensive safety testing. In contrast to food additives, the proposed immobilised agent has a much lower potential to cause adverse reactions, since most of it will never be consumed. Ultimately, CBD_TP84_28_His is just a protein; even if ingested, it will simply be digested and broken down into its constituent amino acids.
On top of all the intriguing properties and potential applications, CBD_TP84_28_His is highly thermostable. It proves able to preserve its full activity in temperatures as high as 65°C (Fig. 6). Interestingly, spot assays, which are qualitative tests, have proven that some residual CBD_TP84_28_His activity is preserved even after autoclaving (121°C, 20 min). Measurements using nanoDSF technology confirmed that the melting point of the protein is 73.06°C, which is slightly less than the melting point of lone TP84_28_His at 77.6°C [11]. The change in melting point is most likely due to the presence of the additional cellulose-binding domain. Thermostability gives CBD_TP84_28_His a major advantage, as it can be stored or used without decrease in activity in variable conditions.
4. Materials and Methods
4.1. Bacterial Strains, Media, Reagents, DNA, SOFTWARE and devices
Bacteriophage TP-84 (NC_041918.2), its host, G. stearothermophilus strain 10 (BGSC No. 9A21, NCBI ID 272567) and bacteria used to assess CBD_TP84_28_His lytic activity came originally from Epstein and Campbell, 1975. Escherichia. coli (E. coli) BL21(DE3) (B F_ ompT hsdS (rB_mB-) dcm+ Tetr galλ (DE3) endA Hte) (Agilent Technologies, Palo, Alto, CA, USA) was used for gene expression and E. coli DH5α (F- φ80lacZ∆M15 ∆(lacZYA-argF) U169 deoR recA1 endA1 hsdR17 (rk - , mk +) phoA supE44 λ- thi-1 gyrA96 relA1 Mrr -) from New England Biolabs (Ipswich, MA, USA) was used for plasmid propagation.For CBD_TP84_28_His lytic activity measure used bacteria: E. coli (KPD 168-BA), B. stearothermophilus (KPD 109-BA), B. cereus (KPD 110-BA), B. subtilis (KPD 112-BA) and Geobacillus ICI (KPD 1699). Thermus aquaticus DSM 625 (T. aquaticus) was obtained from the DSM collection. For culturing G. stearothermophilus strain 10, B. cereus, B. subtilis, E. coli, B. stearothermophilus LB medium (1% tryptone, 0.5% yeast extract, 1% NaCl) was used, for T. aquaticus TM medium (0.3% tryptone, 0.2% yeast extract, 1x Castenholz salts) was used, for Geobacillus ICI 2xYT medium (1.6% tryptone, 1% yeast extract, 0.5% NaCl) was used and their corresponding solid media were supplemented with 1.5% of agar. For bacterial biofilm disruption assessment, TSBg medium (1.7% peptone K; 0.3% peptone SP; 0.5% NaCl; 0.25% K2HPO4; 0.25% glucose, pH 7.3) was used. Bacterial strains used to assess activity of fusion protein CBD_TP84_28_His against biofilm included the host strain G. stearothermophilus strain 10 and bacteria known for their pathogenicity for humans: E. coli DSM 1103 and B. cereus DSM 31 from DSM collection, as well as Staphylococcus aureus ATCC 25923 (S. aureus), Pseudomonas aeruginosa ATCC 17503 (P. aeruginosa), Salmonella enteritidis ATCC 25928 (S. enteritidis) from the ATCC collection. The Plasmid Mini kit and Gel-Out ACX kit were from A&A Biotechnology (Gdańsk, Poland). SapI restriction endonuclease, Q5® High Fidelity DNA polymerase and Q5® High GC Enhancer and dNTPs were from New England Biolabs (Ipswich, MA, USA). PCR primers synthesis and DNA sequencing was conducted at Eurofins Genomics (Ebersberg, Germany). T4 DNA Ligase and low-melting point agarose were supplied by ThermoFisher Scientific Baltics UAB (Vilnus, Lithuania). Agarose was from Bioshop (Burlington, Canada). VivaSpin Turbo membranes came from Sartorius (Burlington, USA). Chromatographic column Ni-NTA HisTrap HP was obtained from GEHealthcare Bio-Sciences AB (Uppsala, Sweden). Clone ID kit was from Lucigen Corporation (Middleton, USA). Genetic constructs were designed with SnapGene 4.1 software. Microplate reader Infinite M200 PRO by Tecan Trading AG (Männedorf, Switzerland), Victor3 V multilabel plate reader (Waltham, USA) and spectrophotometer Jenway 7205 by Cole-Parmer Instrument Company, LCC (St Neots, UK) were used for spectrophotometric measurements. T100 Thermal Cycler and Next Generation Chromatography apparatus were supplied by Bio-Rad (Hercules, USA). The Prometheus Panta device by NanoTemper Technologies (Munich, Germany) was used for nano-Differential Scanning Fluorimetry (nanoDSF) to measure unfolding temperature of the fusion CBD_TP84_28_His. 6-well, flat bottom polystyrene microplates by Nest Scientific Biotechnology (Rahway, USA) were used to pour bacterial solution and then to assess biofilm formation inhibition. Corning® 96-well Clear Flat Bottom Polystyrene High Bind Microplate purchased from Corning (Corning, USA) was used to place microcellulose (µC) when assessing its interactions with CBD_TP84_28_His. To visualize interactions of CBD_TP84_28_His with cellulose materials using dedicated antibodies, EveryBlot Blocking Buffer from Bio-Rad (Hercules, USA), 3,3’- Diaminobenzidine (DAB) from Sigma-Aldrich (Saint Louis, USA) and anti-His HRP-conjugated antibodies from Merck (Darmstadt, Germany) were used. Microcellulose (µC) used to assess the affinity of the fusion CBD_TP84_28_His to cellulose was prepared as we described before [11]. All other chemical reagents were purchased from Merck (Darmstadt, Germany).
4.2. Construction, Expression and Purification of Fusion Endolysin - CBD_TP84_28_His
4.2.1. Cloning tp84_28 Gene into pET28_delSapI_CBD_His Vector
The fusion gene, coding for fusion protein CBD_TP84_28_His, was constructed by cloning using the pET28_delSapI_CBD_His custom-made expression-fusion vector (a derivative of the pET21d (+) vector) for cloning the PCR amplified tp84_28 gene. The vector contains the sequence coding for the CBD domain, derived from cellulase of Clostridium cellulovorans [18] in perfect fusion with the vector’s START codon. The enterokinase site was included between sequences coding for the CBD domain and TP84_28_His domain, and the His6-tag was included at the C-terminus of the recombinant protein. The resulting fusion ORF was under the control of the T7 promoter. The tp84_28 gene was amplified in a PCR using TP-84 genomic DNA as a template, Q5® High Fidelity DNA Polymerase, Q5® High GC Enhancer and primers: F - pET_TP28_SapI_F – gagctcttcacccgggatgcaagcaagat, R - pET_TP28_SapI_R – CATGCTCTTCTGGGCCCgtttggaattttgatcgt. The PCR temperature conditions were denaturing 98°C, 30 cycles: 98°C per 30 s, 62°C per 30 s, 72°C per 2 min, annealing 72°C per 4 min. The reaction product was phenol-chloroform purified and ethanol precipitated. The vector was SapI digested, dephosphorylated using E. coli bacterial alkaline phosphatase, separated along with SapI digested PCR product on agarose electrophoresis, both Gel-Out AX kit-purified and ligated using T4 ligase at the insert / vector molar ratio 6:1 at 16°C for 16 h. The mixture was transformed into E. coli DH5α, plated onto kanamycin-supplemented (30 μg/ml) LB plates and resulting clones PCR were analysed using Clone ID kit and primers pET-tert (TGCTAGTTATTGCTCAGCGG) and pET-up (GATGCGTCCGGCGTAGA). Reaction conditions included: 25 PCR cycles, denaturation at 98°C for 2 min, annealing at 52°C for 30 sec, elongation at 72°C for 1 min, final fill-in at 72°C for 3 min. Resulting pET28_delSapI_CBD_TP84_28_His clones were sequenced and isolated plasmid DNA transformed into expression strain E. coli BL21(DE3), previously used for overproduction of TP84_28 endolysin [11]. The sequence of pET28_delSapI_CBD_TP84_28_His was submitted to GeneBank under the accession number PP894717.
4.2.2. Gene Expression and Overproduction of CBD_TP84_28_His
A 100 ml overnight culture (16 h) of E. coli BL21(DE3) pET28_CBD_TP84_28_His was started from a single bacterial colony in LB medium, supplemented with kanamycin (30 µg/ml). It was used to inoculate 1 L of freshly prepared LB medium with kanamycin (30 µg/ml) to optical density at wavelength 600 nm (OD600) 0.1. The culture was grown until OD600=0.6, induced with 1 mM isopropyl-β-D-1-thiogalactopyranoside (IPTG), cultivated for 4 h at 30°C and cells were harvested by centrifugation (2739 x g, 15 min).
4.2.3. Recombinant Fusion Endolysin CBD_TP84_28_His Purification
Bacterial cells (1.6 g) were resuspended in a buffer A (50 mM Tris-HCl pH=7.0, 500 mM NaCl) in mass ratio 1:10. Phenylmethylsulphonyl fluoride (PMSF) was added to a final concentration of 1 mM, lysozyme to final concentration of 0.5 mg/ml, and the mixture was incubated for 30 min at 4°C. Lysis was completed by sonication, with ice-cooling of the lysate, which was subsequently centrifuged (16 000 x g, 20 min) and filtered through 0.22 µm syringe filter. Further purification of the recombinant CBD_TP84_28_His protein was performed by removing the thermolabile E. coli proteins by heating at 55°C for 10 min and centrifugation (12 000 x g, 30 min). The bulk of the other proteins contaminating the supernatant and fragmented nucleic acids were precipitated with 0.2% polyethyleneimine (PEI) buffered to pH 8.0. Precipitate was removed by centrifugation (centrifugation 12 000 x g, 30 min) and dialysed to the buffer A: 50 mM Tris-HCl, 500 mM NaCl (pH=7.0). The CBD_TP84_28 was further metal affinity purified using NGC chromatography and a 5 ml Ni-NTA HisTrap HP column. 15 ml of the solution containing the protein of interest was applied on the column; the column was washed with buffer A (10 column volume (CV)), and CBD_TP84_28_His was eluted with imidazole gradient (20 CV), from 0 mM to 500 mM. The recombinant CBD_TP84_28_His was eluted at 300-400 mM imidazole concentration. Further processing included sample concentration on 10 kDa polystyrene ultrafiltration units, while changing buffer R (50 mM Na/PO4, 150 mM NaCl, pH=7.0). SDS-PAGE analysis was performed to assess the purity of the protein.
4.3. Characterization of Recombinant Fusion Endolysin CBD_TP84_28_His
4.3.1. Evaluation of the Lytic Activity of CBD_TP84_28_His
- Spot assay (diffusion test)
Petri dishes with agar medium for the test bacteria were prepared: (i) LA for E. coli, B. subtilis, B. cereus, B. stearothermophilus, G. stearothermophillus strain 10; (ii) 2xYT solid medium for Geobacillus ICI; (iii) TM solid medium for T. aquaticus and 100 µl of fresh bacterial culture was rubbed into the plate. Bacteria were cultivated at their optimal temperatures (E. coli, B. subtilis, B. cereus at 37°C, G. stearothermophillus strain 10 at 55°C, B. stearothermophilus, T. aquaticus, Geobacillus ICI at 60°C) for variable periods of time, until they formed a uniform bacterial lawn. After that, 10 µl of PBS (control) or 10 µl of CBD_TP84_28_His preparation was spotted in the middle of the Petri dish and allowed to dry for 40 minutes. The plates were left for 3 h at the selected temperature for tested bacteria, and the lytic activity of CBD_TP84_28_His was documented by photographing.
- Turbidity reduction assay (spectrophotometer variant)
Spectrophotometer-quantified turbidity reduction assay (TRA) was used [11]. Overnight (16 h), cultures of test bacteria were grown with vigorous aeration at corresponding media and temperatures (sections “Bacterial strains, media, reagents, DNA, software and devices” and “Spot assay (diffusion test)”). Then, they were used to inoculate fresh medium of the same type, which was cultivated until the mid-exponential phase (OD600 = 0.6). The cultures were then harvested by centrifugation (4000 x g, 10 min, 4°C). The obtained bacterial pellets were resuspended in buffer R at room temperature to approximately the same initial OD600. 100 µl of the buffer R (controls) or 100 µl of the CBD_TP84_28_His solution (0.145 mg/m) was added to 10 ml of the freshly prepared bacterial suspensions to the final concentration of 1.43 µg/ml and shaken at 55°C. 1 ml samples were taken at time intervals and OD600 was measured.
- Turbidity reduction assay (Tecan microplate reader variant)
Overnight (16 h), culture of G. stearothermophilus strain 10 was grown in LB medium at 55 °C. It was used to inoculate 30 mL of fresh LB medium, which was cultivated until the mid-exponential phase (OD600 = 0.6). The culture was harvested by centrifugation (4000 x g, 10 min, 4°C). The resulting bacterial pellet was resuspended in the buffer R at room temperature to approximately the same optical density as the initial culture, according to Tecan 96-well plate reader measurement. 95 µl of the freshly prepared bacterial suspension was placed in the consecutive 14 wells. Then, 5µg of buffer R (control) or 5 µl of 0.5 mg/mL CBD_TP84_28_His in buffer R (preincubated at different temperatures, as described in section “Thermostability of recombinant fusion CBD_TP84_28_His”) was added to each well to final concentration of 25 µg/ml. Measurements of OD600 were performed in 5 min intervals simultaneously for all the wells.
- Inhibition of the biofilm formation by the recombinant fusion endolysin CBD_TP84_28_His
The ability of the fusion protein CBD_TP84_28_His to prevent or disrupt biofilm formation by E. coli, S. aureus, P. aeruginosa, S. enteritidis., B. cereus and G. stearothermophilus strain 10 was evaluated using a quantitative spectrophotometric microtiter plate assay as described before [11]. The selected bacterial culture was diluted with TSBg medium in the ratio 1:150 and left 24 h at a temperature optimal for a given species (E. coli, S. aureus, P. aeruginosa, S. enteritidis., B. cereus - at 37°C and G. stearothermophilus strain 10 at 55°C). Suspensions (4 mL each) were poured into consecutive wells of a 6-well flat-bottom polystyrene microplate. CBD_TP84_28_His was then added at amount ranging from 0.05 to 50 µg (which corresponds to the final concentration of 0.0125 12.5 µg/mL), when efficacy against biofilm formation by G. stearothermophilus strain 10 was analysed. For other bacterial substrates, 2 µg (0.5 µg/ml) was used. The plates were incubated for 24 h at either 55°C (G. stearothermophilus strain 10) or 24 h at 37°C (E. coli, S. aureus, P. aeruginosa, S. enteritidis, B. cereus). After the initial incubation, the plate containing mesophilic bacteria was further incubated for 1 h at 55°C to elevate the CBD_TP84_28_His to its optimal activity temperature. The plates were washed with 4 mL of 1x PBS buffer (137 mM NaCl; 10 mM Na2HPO4; 1,8 mM KH2PO4; 2,7 mM KCl, pH=7.4) and stained with 0.1% crystal violet in 33% (vol/vol) acetic acid for 15 min, then washed twice with 4 mL of 1 x PBS. OD600 was measured with a Spectra Wallac microplate reader. TSBg and bacteria not treated with CBD_TP84_28_His were used as a negative control.
4.3.2. Evaluation of the Thermostability of Recombinant Fusion CBD_TP84_28_His
To assess the thermostability of the CBD_TP84_28_His, 100 µl samples were incubated for 30 min in a thermocycler at selected temperatures (37.7°C , 45.7°C, 50.0°C, 57.5°C, 65.1°C, 70°C, 74.8°C, 81.2°C, 86.2°C, 90.0°C, 95.8°C, 100°C for 30 min; and in an autoclave at 121°C for 20 min). Then, TRA was performed as described in section “Turbidity reduction assay (spectrophotometer variant)”, and data on lytic activity was collected for each of the preincubated samples. Additionally, nanoDSF, an advanced differential scanning fluorimetry, was used to measure the unfolding temperature of the fusion CBD_TP84_28_His.
4.3.3. Cellulose- Binding Properties of Recombinant Fusion Endolysin CBD_TP84_28_His
- Interaction with microcellulose assay
We have devised methods for assessing cellulose-binding properties of recombinant fusion endolysin CBD_TP84_28_His in a format of interaction with microcellulose (µC) [30], patent application (WIPO ST 10/C PL446913)]. A high-binding, flat 96-well plate was coated with a suspension of µC (100 mg in 100 µl 1 x PBS) and incubated for 18 h at 4°C. The plate was then centrifuged at 4000 x g for 10 min. Serial dilutions of enzymes, starting at 1 µg per well, were applied to row A (TP84_28_His) and row B- (CBD_TP84_28_His) in a volume of 100 µl and incubated at 4°C for 18 h. The plate was then centrifuged, and the supernatant extracted. The wells were washed 3 times with 1 x PBS by adding the solution and centrifuging the plate and incubated with a 100 µl blocking buffer per 5 minutes. Then, 100 µl of 1 x TBST (0.05 M Tris-HCl pH=7.5, 0.15 M NaCl, 0.1% Tween 20) was added, and the plate was shaken for 5 min at room temperature. The wells were again vortexed and washed 3 times with TBST buffer by vortexing the plate. The complexes formed were resuspended in 100 µl of a 1:2000 solution of anti-His HRP-conjugated antibodies and incubated for 1 h at room temperature. They were centrifuged again, and the wells were washed 3 times with a TBST buffer. A DAB solution was then added to the washed wells to visualise the formed µC-CBD_TP84_28_His complexes by HRP-DAB chemical reaction (qualitative method).
- Cellulose paper-based immunoblotting assay
Cellulose-binding properties of CBD_TP84_28_His were also assessed using cellulose paper-based immunoblotting, a previously described method [31]. The procedure was run in parallel for CBD_TP84_28_His and controls: recombinant TP84_28_His [11] and PBS. The solution with CBD_TP84_28_His was spotted onto cellulose paper and allowed to adsorb for 2 min at room temperature, which was followed by 10 min of incubation of the sample with 5 mL of blocking buffer (BSA solution). The sample was incubated for 1 h at 30°C with anti-His HRP-conjugated, diluted at 1:2000 in 3mL of 1 x TBST buffer. The sample was washed with 5 mL of 1 x TBST for 5 min. This step was repeated 3 times and premixed DAB solution was spotted onto the paper. The appearance of dark spots indicated the successful detection of the His-tag.
5. Patents
Related patent applications of our research group: (i) microcellulose synthesis method- WIPO ST 10/C PL446913; (ii) fusion recombinant protein CBD_TP84_28_His; the method of obtaining it and its application as published the following work: WIPO ST 10/C PL449273.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Plasmid pET28_CBD_TP84_28_His; Figure S2: Growth curve of E. coli BL(DE3) Gold with recombinant plasmid pET28delSapI_CBD_TP84_28_His after the induction of fusion gene cbd_tp84_28 expression and controls.
Author Contributions
All authors contributed to the study conception and design. JZ designed all experiments in the manuscript, purified the enzyme and studied its biofilm degradation properties, as well as the interaction of the protein with cellulose. MP contributed to the design of the experiments and purification of the enzyme, characterised the enzyme's properties, activity and thermostability and participated in cellulose binding experiments. All the experiments and data collection and analysis were performed by MP and JŻ. The first draft of the manuscript was written by MP and JŻ. PS came up with the concept of CBD fusions for attachment to micro- and nanocellulose, reviewed the manuscript and secured the funding for the project. All authors read and approved the final manuscript.
Funding
This research was funded by a grant: National Center for Research and Development NCBR TECHMATSTRATEG2/410747/11/NCBR/2019.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data generated or analysed during this study are included in this published article. The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request. Supplementary Materials:Figure S1: Plasmid pET28_CBD_TP84_28_His; Figure S2: Growth curve of E. coli BL(DE3) Gold with recombinant plasmid pET28delSapI_CBD_TP84_28_His after induction of fusion gene cbd_tp84_28 expression and controls.
Acknowledgments
Prof. Piotr Mucha is appreciated for preparing the microcellulose interactions available for the study. Thanks to Anna Pałgan and Paul Gavin for help with graphical design and language corrections.
Conflicts of Interest
MP, JZ and PS declare no competing interests. See the related patent applications of our research group - see section 5. Patents.
References
- Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases, Division of Healthcare Quality Promotion. About Antimicrobial Resistance. Available online: https://www.cdc.gov/drugresistance/about.html (accessed on 24 January 2023).
- Liu, H.; Hu, Z.; Li, M.; Yang, Y.; Lu, S.; Rao, X. Therapeutic potential of bacteriophage endolysins for infections caused by Gram-positive bacteria. J. Biomed. Sci. 2023, 30, 29. [Google Scholar] [CrossRef] [PubMed]
- Gladskin, EU. Microbiome-balancing skincare | endolysin science | Gladskin. Available online: https://www.gladskin.eu/collections/staphefekt (accessed on 28 March 2024).
- Briers, Y.; Walmagh, M.; Grymonprez, B.; Biebl, M.; Pirnay, J.P.; Defraine, V.; Michiels, J.; Cenens, W.; Aertsen, A.; Miller, S.; Lavigne, R. Art-175 is a highly efficient antibacterial against multidrug-resistant strains and persisters of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 3774–3784. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Karyagina, A.S.; Vasina, D.V.; Vasina, I.V.; Gushchin, V.A.; Lunin, V.G. Resistance to peptidoglycan-degrading enzymes. Crit. Rev. Microbiol. 2020, 703–726. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.U.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a promising solution against antimicrobial resistance. Antibiotics (Basel) 2021, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Walter, A.; Mayer, C. Peptidoglycan structure, biosynthesis, and dynamics during bacterial growth. In Extracellular Sugar-Based Biopolymers Matrices. Biologically-Inspired Systems, vol 12; Cohen, E., Merzendorfer, H., Eds.; Springer: Cham, 2019. [Google Scholar] [CrossRef]
- Fenton, M.; Ross, P.; McAuliffe, O.; O'Mahony, J.; Coffey, A. Recombinant bacteriophage lysins as antibacterials. Bioeng. Bugs 2010, 1, 9–16. [Google Scholar] [CrossRef]
- Liu, B.; Guo, Q.; Li, Z.; Guo, X.; Liu, X. Bacteriophage endolysin: A powerful weapon to control bacterial biofilms. Protein J. 2023, 42, 463–476. [Google Scholar] [CrossRef]
- Lee, C.; Kim, H.; Ryu, S. Bacteriophage and endolysin engineering for biocontrol of food pathogens: recent advances and future trends. Crit. Rev. Food Sci. Nutr. 2023, 63, 8919–8938. [Google Scholar] [CrossRef]
- Żebrowska, J.; Żołnierkiewicz, O.; Ponikowska, M.; Puchalski, M.; Krawczun, N.; Makowska, J.; Skowron, P.M. Cloning and characterization of a thermostable endolysin of bacteriophage TP-84 as a potential disinfectant and biofilm-removing biological agent. Int. J. Mol. Sci. 2022, 23, 7612. [Google Scholar] [CrossRef]
- Saunders, G.F.; Campbell, L.L. Characterization of a thermophilic bacteriophage for Bacillus stearothermophilus. J. Bacteriol. 1966, 91, 340–348. [Google Scholar] [CrossRef]
- Epstein, I.; Campbell, L.L. Production and purification of the thermophilic bacteriophage TP-84. Appl. Microbiol. 1975, 29, 219–223. [Google Scholar] [CrossRef]
- Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sobolewski, I.; Skowron, P.M. Bacteriophages of thermophilic Bacillus group bacteria - a review. Microorganisms 2021, 9, 1522. [Google Scholar] [CrossRef] [PubMed]
- Żebrowska, J.; Witkowska, M.; Struck, A.; Laszuk, P.E.; Raczuk, E.; Ponikowska, M.; Skowron, P.M.; Zylicz-Stachula, A. Antimicrobial potential of the genera Geobacillus and Parageobacillus, as well as endolysins biosynthesized by their bacteriophages. Antibiotics (Basel) 2022, 11, 242. [Google Scholar] [CrossRef] [PubMed]
- Skowron, P.M.; Kropinski, A.M.; Żebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; Zylicz-Stachula, A. Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84 virus genus. PLoS ONE 2018, 13, 1–23. [Google Scholar] [CrossRef]
- Skowron, P.M.; Łubkowska, B.; Sobolewski, I.; Zylicz-Stachula, A.; Šimoliūnienė, M.; Šimoliūnas, E. Bacteriophages of thermophilic 'Bacillus Group' bacteria: A systematic review, 2023 update. Int. J. Mol. Sci. 2024, 25, 3125. [Google Scholar] [CrossRef]
- Shoseyov, O.; Takagi, M.; Goldstein, M.A.; Doi, R.H. Primary sequence analysis of Clostridium cellulovorans cellulose binding protein A. Proc. Natl. Acad. Sci. USA 1992, 89, 3483–3487. [Google Scholar] [CrossRef]
- Abouhmad, A.; Mamo, G.; Dishisha, T.; Amin, M. A.; Hatti-Kaul, R. . T4 lysozyme fused with cellulose-binding module for antimicrobial cellulosic wound dressing materials. Journal of applied microbiology 2016, 121, 115–125. [Google Scholar] [CrossRef]
- Murray, E.; Draper, L.A.; Ross, R.P.; Hill, C. The advantages and challenges of using endolysins in a clinical setting. Viruses 2021, 13, 680. [Google Scholar] [CrossRef]
- Gontijo, M.T.P.; Jorge, G.P.; Brocchi, M. Current status of endolysin-based treatments against Gram-negative bacteria. Antibiotics (Basel) 2021, 10, 1143. [Google Scholar] [CrossRef]
- Wang, S.; Gu, J.; Lv, M.; Guo, Z.; Yan, G.; Yu, L.; Du, C.; Feng, X.; Han, W.; Sun, C.; Lei, L. The antibacterial activity of E. coli bacteriophage lysin Lysep3 is enhanced by fusing the Bacillus amyloliquefaciens bacteriophage endolysin binding domain D8 to the C-terminal region. J. Microbiol. 2017, 55, 403–408. [Google Scholar] [CrossRef]
- Lysando. Lysando-Brochure-Digital. Available online: https://www.lysando.com/wp-content/uploads/2023/08/Lysando-Brochure-Digital.pdf (accessed on 29 March 2024).
- Zampara, A.; Sørensen, M.C.H.; Grimon, D.; Antenucci, F.; Vitt, A.R.; Bortolaia, V.; Briers, Y.; Brøndsted, L.
- Nguyen, H.T.T.; Nguyen, T.H.; Otto, M. The staphylococcal exopolysaccharide PIA - biosynthesis and role in biofilm formation, colonization, and infection. Comput. Struct. Biotechnol. J. 2020, 18, 3324–3334. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Siondalski, P.; Kołaczkowska, M.; Bieńkowski, M.; Pęksa, R.; Kowalik, M.M.; Dawidowska, K.; Vandendriessche, K.; Meuris, B. Bacterial cellulose as a promising material for pulmonary valve prostheses: In vivo study in a sheep model. J. Biomed. Mater. Res. Part B: Appl. Biomater. 2024, 112, e35355. [Google Scholar] [CrossRef] [PubMed]
- Marangoz, B.; Kahraman, S.; Bostan, K. Bacillus spp. responsible for spoilage of dairy products. Int. J. Food Eng. 2018, 4, 43–46. [Google Scholar]
- André, S.; Vallaeys, T.; Planchon, S. Spore-forming bacteria responsible for food spoilage. Res. Microbiol. 2017, 168, 379–387. [Google Scholar] [CrossRef]
- Żebrowska, J.; Mucha, P.; Prusinowski, M.; Krefft, D.; Żylicz-Stachula, A.; Deptuła, M.; Skoniecka, A.; Tymińska, A.; Zawrzykraj, M.; Zieliński, J.; Pikuła, M.; Skowron, P.M. Development of hybrid biomicroparticles: cellulose-exposing functionalized fusion proteins. Microb. Cell Fact. 2024, 23, 81. [Google Scholar] [CrossRef]
- Sun, X.; Yang, S.; Al-Dossary, A.A.; Broitman, S.; Ni, Y.; Guan, M.; Yang, M.; Li, J.; et al. Nanobody-functionalized cellulose for capturing SARS-CoV-2. Appl. Environ. Microbiol. 2022, 88, e0230321. [Google Scholar] [CrossRef]
Figure 1.
SDS-PAGE analysis of the purified recombinant fusion endolysin CBD_TP84_28_His. M, PageRuler Plus Stained Protein Ladder; E, endolysin CBD_TP84_28_His. Arrow points at a band corresponding in size to the endolysin CBD_TP84_28_His (64.2 kDa).
Figure 1.
SDS-PAGE analysis of the purified recombinant fusion endolysin CBD_TP84_28_His. M, PageRuler Plus Stained Protein Ladder; E, endolysin CBD_TP84_28_His. Arrow points at a band corresponding in size to the endolysin CBD_TP84_28_His (64.2 kDa).
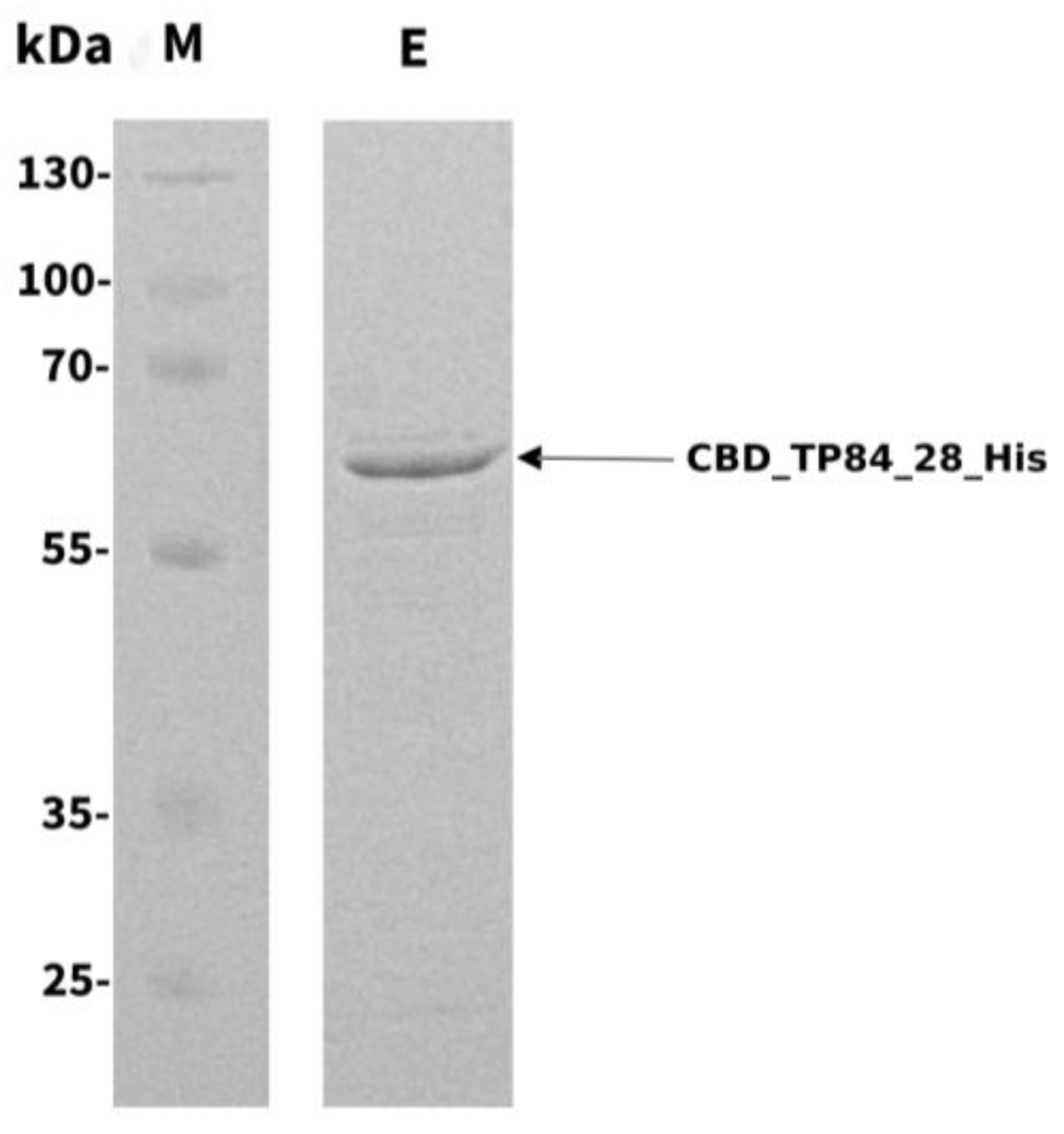
Figure 2.
Recombinant fusion endolysin CBD_TP84_28_His activity evaluation - spot assay Top row (a) shows activity of CBD_TP84_28_His against mesophilic bacteria: B. cereus, B. subtilis, E. coli. Bottom row (b) shows activity of CBD_TP_84_28_His against thermophilic bacteria: G. stearothermophilus strain 10, T. aquaticus, Geobacillus ICI, B. stearothermophilus. Arrows indicate where CBD_TP84_28_His solution was spotted and transparent circles on the bacterial lawn indicate lytic effect of the enzyme.
Figure 2.
Recombinant fusion endolysin CBD_TP84_28_His activity evaluation - spot assay Top row (a) shows activity of CBD_TP84_28_His against mesophilic bacteria: B. cereus, B. subtilis, E. coli. Bottom row (b) shows activity of CBD_TP_84_28_His against thermophilic bacteria: G. stearothermophilus strain 10, T. aquaticus, Geobacillus ICI, B. stearothermophilus. Arrows indicate where CBD_TP84_28_His solution was spotted and transparent circles on the bacterial lawn indicate lytic effect of the enzyme.
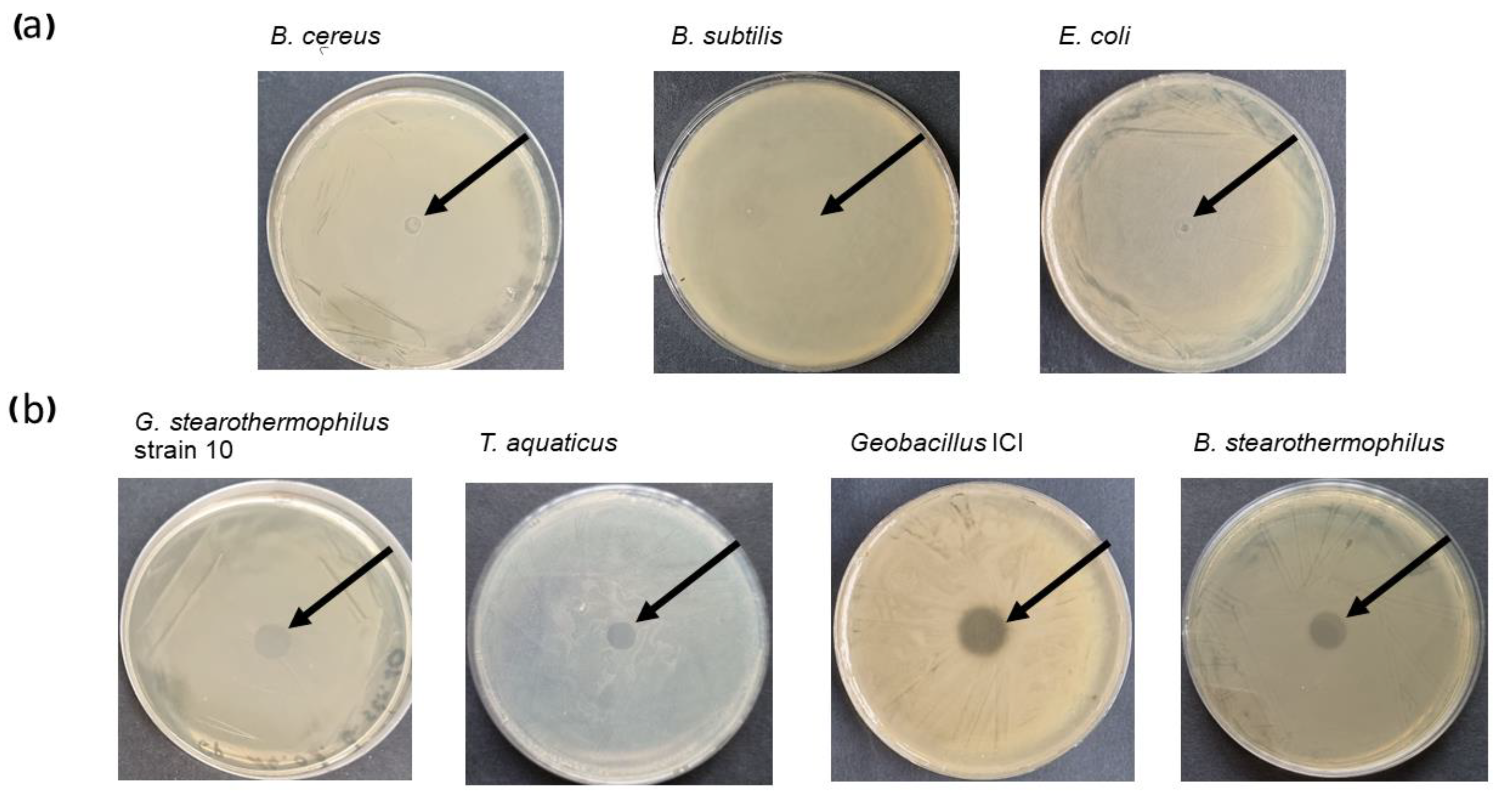
Figure 3.
Recombinant fusion endolysin CBD_TP84_28_His activity evaluation – TRA assay on various bacterial strains. The graph shows the reduction of relative OD600 (ratio of OD600 of the sample treated with CBD_TP84_28_His to the OD600 of the control) in bacterial substrates suspensions (E. coli, Geobacillus ICI, G. stearothermophilus, strain 10, B. stearothermophilus, B. cereus, T. aquaticus, B. subtilis) upon addition of purified CBD_TP84_28_His to the final concentration of 1.43 µg/ml at 55°C.
Figure 3.
Recombinant fusion endolysin CBD_TP84_28_His activity evaluation – TRA assay on various bacterial strains. The graph shows the reduction of relative OD600 (ratio of OD600 of the sample treated with CBD_TP84_28_His to the OD600 of the control) in bacterial substrates suspensions (E. coli, Geobacillus ICI, G. stearothermophilus, strain 10, B. stearothermophilus, B. cereus, T. aquaticus, B. subtilis) upon addition of purified CBD_TP84_28_His to the final concentration of 1.43 µg/ml at 55°C.
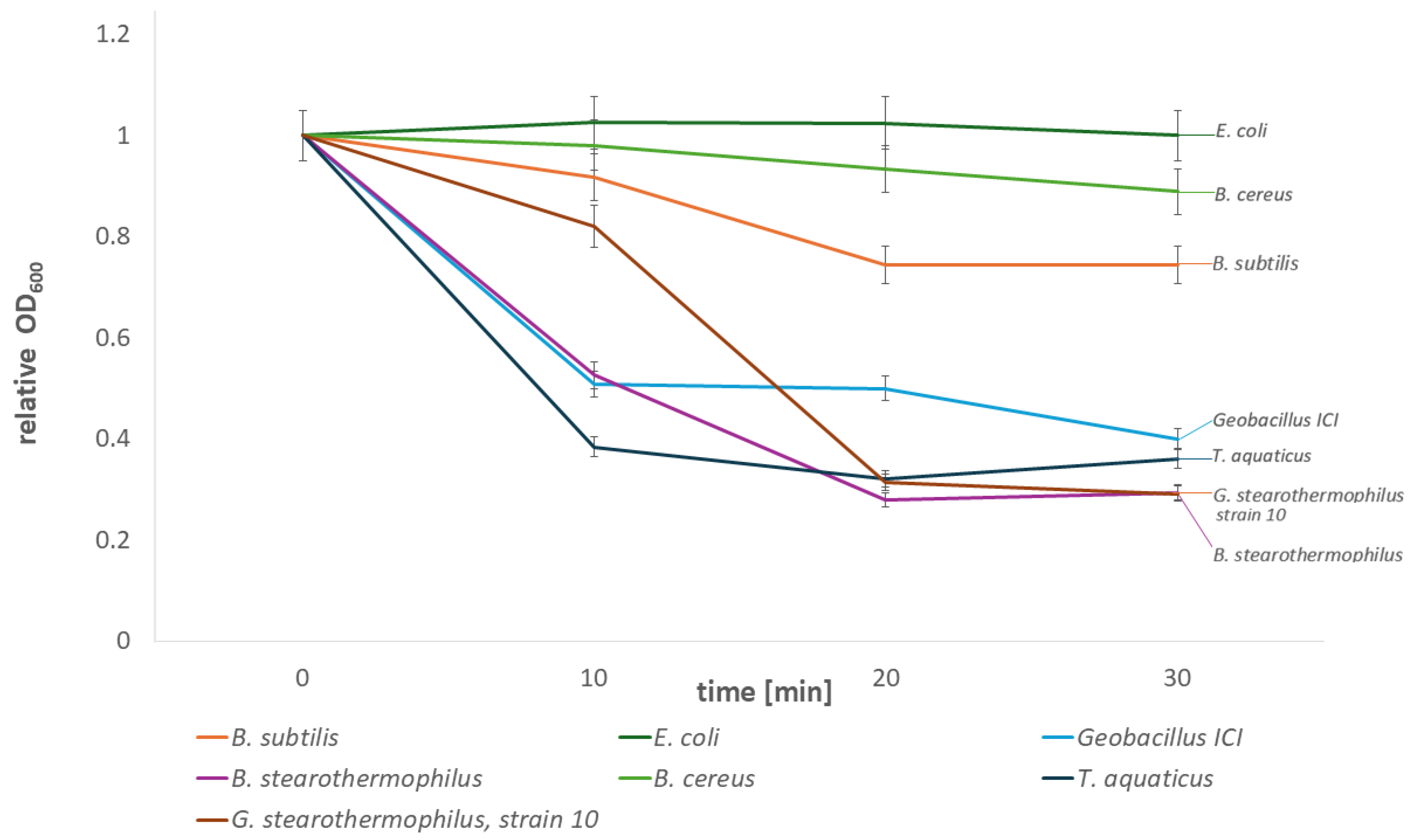
Figure 4.
Comparison of activity of the recombinant fusion endolysin CBD_TP84_28_His and recombinant TP84_28_His. The graph shows reduction in OD600 in G. stearothermophilus strain 10, resuspended in buffer R, after addition of equimolar amounts of CBD_TP84_28_His, TP84_28_His or the reaction buffer alone [control (-)].
Figure 4.
Comparison of activity of the recombinant fusion endolysin CBD_TP84_28_His and recombinant TP84_28_His. The graph shows reduction in OD600 in G. stearothermophilus strain 10, resuspended in buffer R, after addition of equimolar amounts of CBD_TP84_28_His, TP84_28_His or the reaction buffer alone [control (-)].
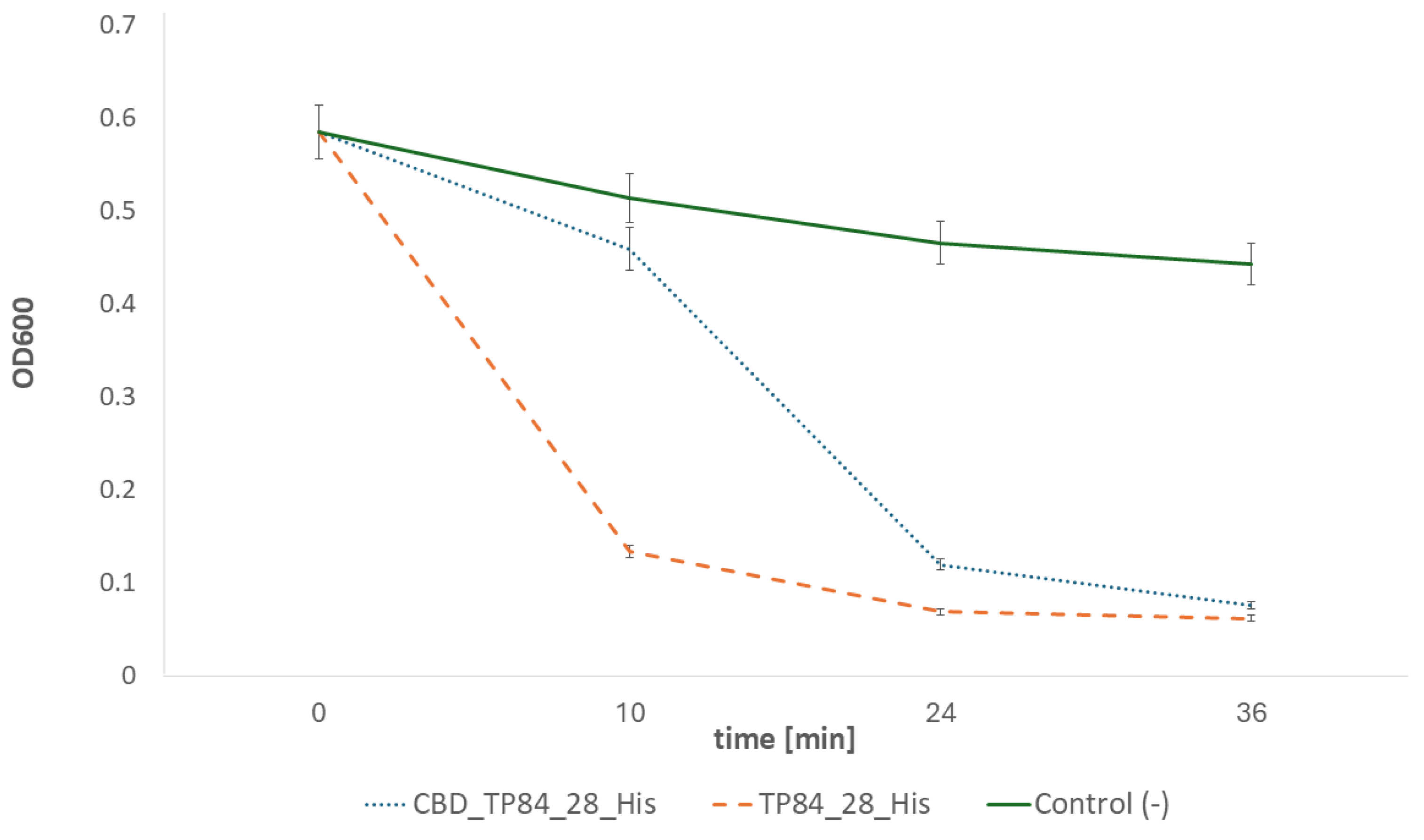
Figure 5.
Inhibition of biofilm formation by G. stearothermophilus strain 10. The graph shows OD600 measurements of G. stearothermophilus strain 10, cultivated on microtiter plate with addition of 0.05 – 50 µg of CBD_TP84_28_His for 24 h at 55°C. The picture below the chart shows biofilm stained with crystal violet: control on the left and samples treated with increasing amounts of the recombinant fusion endolysin CBD_TP84_28_His.
Figure 5.
Inhibition of biofilm formation by G. stearothermophilus strain 10. The graph shows OD600 measurements of G. stearothermophilus strain 10, cultivated on microtiter plate with addition of 0.05 – 50 µg of CBD_TP84_28_His for 24 h at 55°C. The picture below the chart shows biofilm stained with crystal violet: control on the left and samples treated with increasing amounts of the recombinant fusion endolysin CBD_TP84_28_His.
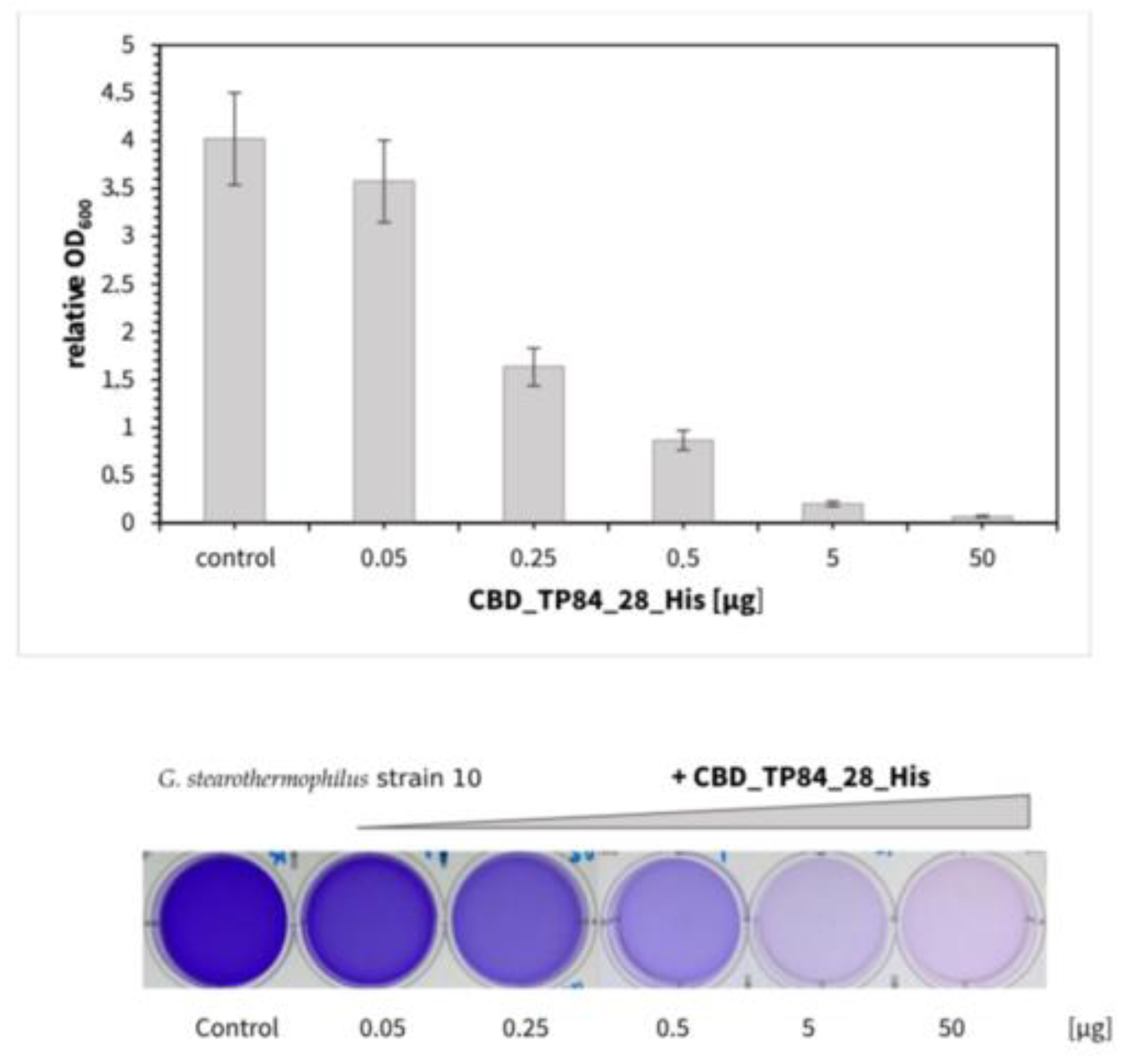
Figure 6.
Recombinant fusion CBD_TP84_28_His thermal stability evaluation. Lytic activity of CBD_TP84_28_His was assayed in optimal conditions after preincubation at various temperatures in a gradient thermocycler. The graph shows reduction in OD600 in G. stearothermophilus strain 10 suspension in buffer R after the addition of CBD_TP84_28_His, preincubated for 30 mins at temperatures of 37.7 – 95.8°C and autoclaving conditions (121°C, 20 min). As a negative control, the buffer was added instead of CBD_TP84_28_His. Measurements were taken using the Tecan microplate reader.
Figure 6.
Recombinant fusion CBD_TP84_28_His thermal stability evaluation. Lytic activity of CBD_TP84_28_His was assayed in optimal conditions after preincubation at various temperatures in a gradient thermocycler. The graph shows reduction in OD600 in G. stearothermophilus strain 10 suspension in buffer R after the addition of CBD_TP84_28_His, preincubated for 30 mins at temperatures of 37.7 – 95.8°C and autoclaving conditions (121°C, 20 min). As a negative control, the buffer was added instead of CBD_TP84_28_His. Measurements were taken using the Tecan microplate reader.
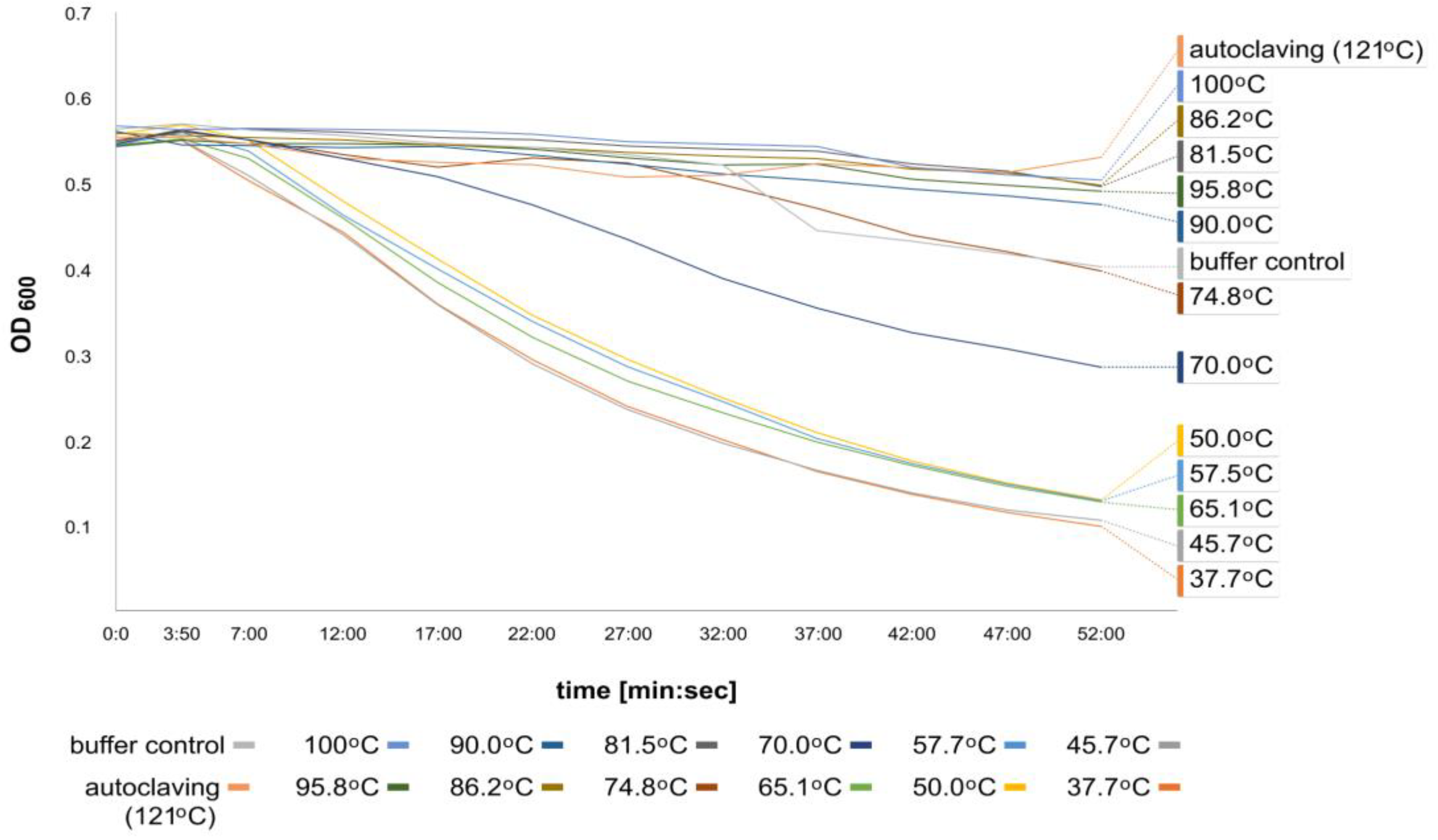

Table 1.
Inhibition of the biofilm formation by E. coli, S. aureus, P. aeruginosa, S. enteritidis., B. cereus. The table shows the relative OD600 value for a bacteria biofilm control process, as compared to a control reaction devoid of CBD_TP84_28_His. Biofilm of each bacterium was treated with 2 µg of CBD_TP84_28_His at 37°C for 24 h and at 55°C for 1 h.
Table 1.
Inhibition of the biofilm formation by E. coli, S. aureus, P. aeruginosa, S. enteritidis., B. cereus. The table shows the relative OD600 value for a bacteria biofilm control process, as compared to a control reaction devoid of CBD_TP84_28_His. Biofilm of each bacterium was treated with 2 µg of CBD_TP84_28_His at 37°C for 24 h and at 55°C for 1 h.
| Bacteria | Control [OD600] | Treatment [OD600] | Reduction [%] |
| E. coli (DSM 1103) | 5.4 | 3.3 | 39 |
| S. aureus (ATCC 25923) | 1.6 | 0.3 | 81 |
| P. aeruginosa (ATCC 17503) | 14.9 | 8.6 | 42 |
| S. enteritidis (ATCC 25928) | 0.14 | 0.09 | 34 |
| B. cereus (DSM 31) | 11.9 | 8.0 | 33 |
| Control | 0.0 | 0.0 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



