Submitted:
04 November 2024
Posted:
05 November 2024
You are already at the latest version
Abstract
The emergence of antibiotic-resistant Acinetobacter baumannii (A. baumannii) is a pressing threat in clinical settings. As the last line of defense, colistin is currently widely used to treat multidrug-resistant A. baumannii. Unfortunately colistin-resistant phenotypes in A.baumannii isolates have also been reported, highlighting the desperate need to develop alternative medications to address these serious bugs. To resist colistin, A. baumannii has developed several mechanisms. These include the loss of outer membrane lipopolysaccharide (LPS) due to mutation of LPS biosynthetic genes, modification of lipid A (a constituent of LPS) structure through the addition of phosphoethanolamine (PEtN) moieties to the lipid A component by overexpression of chromosomal pmrCAB operon genes and eptA gene, or acquisition plasmid-encoded mcr genes through horizontal gene transfer. Other resistance mechanisms involve alterations of outer membrane permeability through porins, expulsion of colistin by efflux pumps and heteroresistance. In response to the rising threat of colistin-resistant A. baumannii, researchers have developed various treatment strategies, including antibiotic combination therapy, adjuvants to potentiate antibiotic activity, repurposing existing drugs, antimicrobial peptides, nanotechnology, photodynamic therapy, CRISPR/Cas and phage therapy. While many of these strategies have shown promise in vitro and in vivo, further clinical trials are necessary to ensure their efficacy and widen their clinical applications. Ongoing research is crucial to identify optimal therapeutic strategies for managing colistin-resistant A. baumannii. This review explores the genetic mechanisms underlying colistin resistance and assesses potential treatment options for this challenging pathogen.
Keywords:
Colistin
; antibiotic resistant
; A. baumannii
; mutation
; lipid A
1. Introduction
Multidrug resistant Acinetobacter baumannii (A.baumannii) is a notorious pathogen, posing a great threat to human health, and is responsible for bacteremia, pneumonia, urinary tract, and skin and soft tissue infections, contributing to significant morbidity and mortality [1]. The World Health Organization (WHO and the Centers for Disease Control and Prevention (CDC) have declared it critical priority pathogen and urges the exploration of new treatment options to combat this pathogen [2]. The emergence of multidrug-resistant (MDR) and extensively drug-resistant (XDR) isolates of A. baumannii make it more challenging to address the associated infections. Several antibiotics are usually considered effective for the treatment of these infections, such as polymyxin E and B, sulbactam, tigecycline, piperacillin/tazobactam, tigecycline, cefiderocol and aminoglycosides used alone or in combination [3]. Nevertheless, the resistant strains are becoming more rampant, resulting in limited treatment options [4]
Polymyxin antibiotics, mainly colistin, are currently used as last-line defense against MDR A. baumannii infections. Colistin is a polycationic peptide that belongs to the class of polymyxin antibiotics, with only two members- polymyxin B and colistin- used in clinical settings [5]. It was first isolated in 1947 from the bacterium Paenibacillus polymyxa subspecies Colistinus and introduced into clinical use in the 1950s, but its use in human medicine was mainly limited to the treatment of pulmonary infections caused by MDR Gram-negative pathogens in patients with cystic fibrosis [6]. Initially limited to topical use in human medicine due to its nephrotoxic and neurotoxic risks when administered systemically, colistin has become increasingly relied upon as a last-resort antibiotic for treating challenging infections caused by multidrug-resistant (MDR) Gram-negative pathogens [7,8]. It is commonly used in critical cases such as bacteremia, sepsis, and ventilator-associated pneumonia (VAP) in intensive care units. Additionally, colistin is utilized as an alternative treatment for urinary tract infections, osteomyelitis, joint infections, meningitis, pneumonia, gastrointestinal infections, pyoderma, soft tissue infections, as well as eye and ear infections [9]. Colistin is still commonly used as an additive in livestock feed to promote growth and treat intestinal infections. It is primarily used in food-producing animals such as pigs and poultry to control these infections. In the sea food industries, colistin sulfate is also used to promote fish growth [10,11]. However, extensive reliance on colistin in animals and human health exerts high selective pressure.
Consequently, the growing clinical reliance on colistin for the treatment of infections has led to a surge of colistin resistance in clinical strains [12]. Colistin-resistance in A. baumannii can occur through various mechanisms. One significant mechanism involves the addition of 4-amino-4-deoxy-L-arabinose (L-Ara4N) or phosphoethanolamine (pEtN) to the lipid A component of lipopolysaccharide (LPS). This addition reduces the overall negative charge of the bacterial outer membrane (OM), making it more stable and hindering colistin penetration [13]. The PmrAB two-component system plays a role in regulating colistin resistance by controlling the expression of PmrC, an enzyme that aids in adding pEtN to lipid A. This modification decreases the negative charge of the outer membrane, making it harder for colistin to bind [14]. Furthermore, mutations in genes responsible for LPS biosynthesis, such as lpxC, lpxA, or lpxD, can lead to the loss of LPS and contribute to colistin resistance [15,16]. Besides, in A. baumannii, the plasmid carrying the mobile colistin resistance (mcr)-1 gene contributes to colistin resistance by encoding a phosphoethanolamine transferase. This enzyme catalyzes the addition of a phosphoethanolamine group to lipid A in the bacterial outer membrane, potentially altering its structure [17] . Interestingly, many A. baumannii isolates have been frequently reported to exhibit heteroresistance to colistin. This phenomenon involves apparently colistin-susceptible strains harboring a subpopulation of colistin-resistant cells. Under selective pressure, both in vitro and in vivo, heteroresistant A. baumannii strains can quickly evolve into strains with a high level of colistin resistance. Treating infections caused by heteroresistant isolates may lead to the selection of colistin-resistant subpopulations, potentially resulting in therapeutic failures [18].
In this review, we discuss the colistin-resistant mechanisms of A. baumannii and the alternative therapeutic drugs to address colistin-resistant A. baumanii (Col-RAB) infections.
2. Colistin Mechanism of Action
Colistin exerts its antibacterial activity on the outer membrane which harbors lipopolysaccharides that constitute the cell surface [19]. These LPS molecules limit the entry of hydrophobic components and antibiotics as well as confers stability and integrity to the outer membrane. Positively charged colistin binds to the negatively charged phosphate group of lipid A through electrostatic interaction. Lipid A is a hydrophobic constituent of the LPS of gram-negative bacteria, and plays a substantial role in bacterial permeability and exchange with the cell exterior. After the initial interaction, colistin competitively displaces the divalent calcium (Ca2+) and magnesium (Mg2+) cations, resulting in destabilizing the cytoplasmic membrane. This disruption weakens the outer membrane LPS and leads to the loss of inner cellular contents, ultimately causing bacterial cell death [20]. Colistin binds to lipid A (lipid A is an endotoxin) and exerts anti-endotoxin activity by neutralizing its effects. Thus, colistin inhibits the endotoxin functions of lipid A by binding to the LPS [21]. Colistin could also promote the intracellular accumulation of reactive oxygen species (ROS), which leads to the bactericidal effect on Col-RAB. ROS- generated during colistin traversing through the outer and inner membrane are converted into H2O2 by superoxide dismutase enzymes. H2O2 participates in the Fenton reaction oxidizing Fe2+into Fe3+, and concomitantly producing hydroxyl radicals (•OH). This reaction can induce oxidative damage to DNA and membrane lipids [22,23] (Figure 1).
3. Mechanism of Colistin Resistance in A. baumannii
A. baumannii can rapidly develop resistance to colistin through two main mechanisms, both of which involve complete loss or significant alterations of the lipid A component of LPS, the primary target of colistin activity. In both cases, the net negative charge on the cell surface decreases affecting the electrostatic interaction between positively charged colistin and the negatively charged lipid A. Loss of LPS is achieved through the inactivation of genes (lpxA, lpxC and lpxD genes) involved in the initial step of lipid A synthesis. The most common mechanism, however, involves the modification of lipid A through the addition of phosphoethanolamine (PEtN). This modification is genetically driven by the chromosomal pmrCAB operon and eptA gene, or through plasmid-borne mcr genes, all of which encode enzymes responsible for altering the structure of lipid A, thereby conferring resistance to colistin in A. baumannii (Figure 2).
The genes lpxA and lpxD on the A. baumannii chromosome encode acyl-transferases involved in lipid A biosynthesis [24]. Similarly, lpxB and lpxC, also involved in lipid A biosynthesis, are located in separate chromosomal regions [25,26]. These genes mainly regulate the first three steps of lipid A biosynthesis pathway, and the hydrophobic anchor of lipopolysaccharide [27]. Therefore, disruption or mutation of lpxA, lpxC, and lpxD genes in A. baumannii may have account for the observed LPS deficiency leading to colistin resistance. Moffat and colleagues were the first to report that a lack of lipopolysaccharide (LPS) results in colistin resistance in A. baumannii [28]. They worked with 13 colistin-resistant variants of the A. baumannii strain ATCC 19606 and found mutations in the lpxA, lpxC, or lpxD genes involved in lipid A biosynthesis. These mutations led to a complete absence of LPS production. The absence of LPS was confirmed in a clinical Col-RAB isolate. Carbohydrate silver staining of whole-cell lysates revealed the lack of LPS in the colistin-resistant strains. Mutations in the lpxA, lpxC, and lpxD genes were identified through PCR amplification and sequencing, with mutations ranging from single nucleotide changes to large deletions up to 445 base pairs. These mutations affected critical residues in the lipid A biosynthesis pathway, such as P30 in LpxC and H159, G68, and Q72 in LpxA, highlighting their importance for the proper function of LpxC and LpxA [28]. Further research with Col-RAB strains, has found that the insertion of ISAba1 or ISAba11 into the lpxC gene is a frequent occurrence, causing the inactivation of lpxC and lpxA genes. This inactivation leads to the loss of lipopolysaccharides (LPS). ISAba11 was found to be inserted between nucleotides 390-393 in four strains and at nucleotide 420 or 421 in three strains, indicating that these specific regions may serve as hot spots for insertion [29]. Besides, under-expression of lpxACD has also been reported in colistin-resistant A. baumannii, leading to the decreased production of lipid A [30].
Modification of lipid A moieties, such as the addition of molecules like 4-amino-4-deoxy-l-arabinose (L-Ara4N), phosphoethanolamine (PEtN), or galactosamine to lipid A, is another common mechanism of colistin resistance in A. baumannii. Unlike other gram-negative bacteria, A. baumannii lacks the biosynthesis machinery for L-Ara4N and instead utilizes PEtN or galactosamine to bind to lipid A [31,32]. The structural modification of lipid A results in a decrease in the net negative charge on the cell membrane, which interferes with the binding of positively charged colistin to lipid A, diminishing its efficacy. Similar to enteric bacteria, the PmrA/PmrB two component system (TCS) has been shown to play a role in mediating colistin resistance in this bacterium [33,34]. The pmrA/pmrB system consists of a two-component response regulator (PmrA) and sensor kinase (PmrB) that enable bacteria to sense and respond to environmental cues. This system also controls the pmrC gene, responsible for encoding a lipid A phosphoethanolamine (PEtN) transferase that alters lipid A in bacterial LPS. Upon activation of the PmrA/PmrB system, phosphorylated PmrA interacts with the pmrCAB operon promoter, leading to increased expression and the synthesis of the enzyme responsible for adding PEtN to lipid A [14,35,36,37]. Colistin resistance develops when phosphoethanolamine (pEtN) is added to lipid A by the phosphoethanolamine transferase PmrC, encoded by the pmrC gene. This process is controlled by the PmrA/PmrB two-component system (TCS). Mutations in the pmrA, pmrB, or pmrC genes can lead to overexpression of pmrC, resulting in resistance. The pmrB gene is most commonly mutated, causing overexpression of pmrAB and enabling PmrC to modify lipid A with pEtN, ultimately contributing to colistin resistance [38,39]. A study identified two novel mutations in pmrA (I13M) and pmrB (Q270P) that contribute to colistin resistance. They also found a mutation in miaA, a posttranscriptional regulator previously reported to affect cell growth and virulence. The I221V mutation enhanced the colistin resistance of pmrA (P102R) [40]. Mutations in the pmrB gene of A. baumannii, which upregulate pmr genes, are the most common cause of colistin resistance, with diverse amino acid changes in the PmrB protein are frequently observed [41,42,43]. Nurtop et al. discover highest number of mutations in pmrB genes [44]. A study identified three colistin-resistant clinical A. baumannii strains from different patients. Non-synonymous mutations were present in various domains of PmrB, with a significant number concentrated in the HisKA domain which is essential for autophosphorylation and transferring the phosphoryl group to the PmrA response regulator [42,45]. Mutations in the receiver domain of the response regulator PmrA were also identified in Col-RAB strains. Palethorpe et al. provided structural insights into the N-terminal domain of A. baumannii PmrA using X-ray crystallography and developed a full-length model through molecular modeling, which helped deduce the impact of two key PmrA mutations, PmrA::I13M and PmrA::P102R, both of which contribute to increased colistin resistance [46]. The PmrAB TCS also controls the naxD gene in colistin-resistant strains with activating mutations in PmrB. An activating mutation in PmrB can cause overexpression of naxD. NaxD deacetylates N-acetylgalactosamine to galactosamine, necessary for adding galactosamine to lipid A. This modification adds a positive charge to lipid A, enhancing resistance [47,48]. A study identified at least 20 amino acid substitutions in PmrC in colistin-susceptible isolates, which were associated with colistin MICs ranging from ≤0.125 µg/ml to 0.5 µg/ml [49] . In another study, colistin-susceptible A. baumannii strain also adopt PEtN [32]. These findings indicate that the resistance mechanism may vary by strain, and the presence of PEtN alone may not completely explain colistin resistance. According to Beceiro et al. the emergence of moderate colistin resistance in A. baumannii is linked to distinct genetic alterations. These alterations comprise a minimum of one (01) point mutation in the pmrB gene, heightened expression of the pmrAB operon, and the activation of pmrC, leading to the incorporation of phosphoethanolamine into lipid A [50].
In addition, the overexpression of another pmrC homolog, named eptA (ethanolamine phosphotransferase A) is found in Col-RAB isolates. The mere presence of eptA gene in bacteria does not inherently confers resistance. However, when the insertion sequences ISAba1 integrate into the upstream of the eptA gene, they can enhance the expression of this enzyme, thus contributing to colistin resistance [51]. The gene eptA is frequently found in clinical strains of A. baumannii belonging to International Clone 2 (IC2) and can be present in multiple copies within a single isolate (three or more). It is located distantly from the pmrCAB operon in the genome, which enhances resistance. In order for these strains to display colistin resistance, the integration of the ISAbaI element upstream of eptA is essential [51,52,53,54]. Lesho et al. discovered that colistin resistance was associated with point mutations in the pmrA1 and/or pmrB genes. They also identified homologs of pmrC, named eptA-1 and eptA-2, which were located distantly from the operon. Colistin-resistant isolates exhibited significantly increased expression of pmrC1A1B, eptA-1, and eptA-2 compared to colistin-susceptible isolates [55]. In a study, two A. baumannii isolates from a patient showed increasing colistin resistance before and after failed treatment. Genome sequencing identified an extra insertion sequence (ISAba125) in a transcriptional regulator gene in the highly resistant strain. Deleting hns in the less resistant strain increased colistin resistance, while restoring hns in the highly resistant strain reversed resistance. Transcriptomic analysis revealed over 150 differentially expressed genes in the resistant strain, including increased expression of eptA, a gene associated with colistin resistance [56].
Plasmid-mediated transferable colistin resistance is mainly caused by mobile colistin resistance (mcr) genes, which produce phosphoethanolamine transferases (MCR enzymes). These enzymes alter lipid A of the lipopolysaccharide (LPS) by adding phosphoethanolamine, a crucial mechanism of colistin resistance in Gram-negative bacteria [57,58,59]. So far, different versions of the mcr-1 gene, from mcr-1 to mcr-10, and their variants have been detected worldwide in various Gram-negative bacteria, primarily found in food-producing animals [60,61,62,63,64]. In A. baumannii, two variants of mcr, mcr-1 and mcr-4.3, have been reported. In 2019, mcr-1 was first reported in clinical isolates from Pakistan [17]. Subsequently, it has been isolated from India [65], Egypt [66], China [67], and Iraq [68]. In 2008, mcr-4.3 positive isolates of A. baumannii from Brazil were found to carry a novel plasmid (pAb-MCR4.3) containing mcr-4.3 within a Tn3-like transposon [69]. The mcr-4.3 gene was subsequently found in pig feces at a slaughterhouse in China, where it was carried by the plasmid pAB18PR065 through horizontal gene transfer [70]. It was also detected in isolates from the Czech Republic, hypothesizing that food imports can disseminate this gene in Europe [71].
A. baumannii can develop resistance to colistin through mechanisms other than modifications or loss of LPS structure, such as the involvement of efflux pumps. Lin et al. [72] suggested that EmrAB efflux pumps play a role in colistin resistance in A. baumannii. Studies have shown that the overexpression of genes like adeI, adeC, emrB, mexB, and macAB, HlyD family —, which encode efflux pump proteins, is associated with colistin resistance, as indicated by transcriptomic (RNAseq) analysis of resistant strains [73,74]. The contribution of efflux pumps is further supported by the reversal of resistance using efflux pump inhibitors (EPIs) and cyanide-3-chlorophenylhydrazone (CCCP) in various bacteria, including A. baumannii, P. aeruginosa, K. pneumoniae, and S. maltophilia [75,76]. Moreover, mutations in vacJ, pldA, ttg2C, pheS, and a conserved hypothetical protein have been linked to decreased colistin susceptibility through novel resistance mechanisms [77]. In another study, Paul et al. found notable changes in the transcription of genes encoding sugE, ydhE, ydgE, mdfA, ynfA, and tolC in Col-RAB when exposed to subinhibitory concentrations of colistin. This indicates that A. baumannii efficiently removes colistin using efflux pumps from the MATE and SMR families [78]. Outer membrane proteins (OMPs) are implicated in the development of colistin resistance in A. baumannii. Specifically, OmpW plays a role in colistin binding and iron homeostasis regulation in the bacterium [79]. The OMP inhibitor AOA-2 has been found to increase the susceptibility of both reference and clinical colistin-susceptible and colistin-resistant strains to colistin in vitro, underscoring the role of OMPs in colistin resistance. In vivo studies have shown that combining AOA-2 with colistin significantly reduces bacterial loads in tissues and blood, and enhances mouse survival rates compared to colistin treatment alone [80]. Colistin resistance in A. baumannii may also be associated with non-Lpx proteins that support the structure and integrity of the outer membrane. One such protein, LpsB, a glycosyltransferase essential for LPS core formation, is crucial for the bacterium's virulence. Deleting LpsB enhances susceptibility to colistin and cationic antimicrobial peptides, underscoring its role in maintaining outer membrane stability [81]. Heteroresistance is a progressive stage in the development of antimicrobial resistance, leading to the failure of anti-infective treatments [82]. It occurs when a small subset of bacterial cells in a genetically uniform group shows resistance to an antibiotic, while the majority of the population remains susceptible [83]. Li et al. first observed heteroresistance to colistin in 15 out of 16 clinical A. baumannii isolates. Subpopulations (<0.1%) of ATCC 19606 and most clinical isolates were able to grow in the presence of 3 to 10 μg/ml colistin. Following four consecutive passages in colistin-containing broth (up to 200 μg/ml), the resistant subpopulation in ATCC 19606 increased from 0.000023% to 100%. Even after 16 passages in colistin-free broth, 2.1% of the population remained resistant [84].
4. Biological Cost of Acquiring Colistin-Resistant Trait
When bacteria become resistant to antibiotics due to genetic mutations, they often experience a "biological cost." This cost refers to a decrease in their overall fitness, pathogenicity, and virulence compared to non-resistant bacteria [85]. In vivo and in vitro studies have found lower fitness and reduced virulence potential in Col-RAB compared to their colistin-susceptible parental strains [86,87,88]. LPS is considered a key factor in the virulence of A. baumannii [89]. As a crucial component of the outer membrane in Gram-negative bacteria, LPS plays a vital role in creating a barrier that controls the passage of substances into the cell and helps maintain the structural integrity of bacteria [90]. Previous research has shown that the absence of LPS in A. baumannii makes it more susceptible to antibiotics like azithromycin, rifampicin, and vancomycin [91]. A study examined various virulence characteristics in colistin-resistant, LPS-deficient versions of the A. baumannii ATCC 19606 strain and five multidrug-resistant clinical isolates, as well as their LPS-deficient, colistin-resistant counterparts. The results indicated that the loss of LPS caused growth deficiencies compared to the original strain both in laboratory conditions and in human serum. Additionally, the LPS-deficient strains showed reduced ability to grow and spread in live organisms. In a mouse model of widespread sepsis, infection with LPS-deficient strains resulted in lower levels of pro-inflammatory cytokines TNF-α and IL-6 in the bloodstream compared to infections with the parent strain, suggesting decreased virulence. The absence of LPS also affected biofilm formation, surface motility, growth in low-iron environments, and increased susceptibility to various disinfectants used in healthcare settings. These findings highlight the significant impact of LPS loss on different virulence factors, shedding light on the relatively infrequent occurrence of LPS-deficient colistin-resistant strains in clinical practice [92]. Col-RAB strains that lack LPS due to mutations in lpx genes only weakly activate neutrophils, leading to decreased production of reactive oxygen species and cytokines. Despite this, neutrophils exhibit a preference for killing these LPS-deficient strains, which are more susceptible to lysozyme and lactoferrin [93].
Mutations in the pmrAB two-component system result in colistin resistance in A. baumannii, which is correlated to impaired fitness, diminished virulence, lower biofilm formation, and decreased infectivity [94,95]. In a study, two strains of A. baumannii were isolated from a ventilator-associated pneumonia patient's respiratory tract. One strain was colistin-susceptible (ABCS), and the other was colistin-resistant (ABCR). Whole-genome sequencing confirmed that ABCR evolved from parental strain due to mutations in the pmrA and rpoB genes and the absence of a prophage. The colistin resistance in ABCR was attributed to a pmrA mutation (E8D), indicating a close relationship between reduced virulence and colistin resistance. The loss of the prophage in ABCR may have also contributed to its decreased in vivo virulence [96]. In contrast, another Col-RAB isolate ( CR17) remains sporadic, although colistin resistance was attained through pmrAB mutation [97]. While strain CR17 remained infectious, it showed decreased virulence and fitness in a murine sepsis model compared to the original susceptible strain, colistin-susceptible 01 (CS01). CS01 was more virulent than CR17, resulting in higher mortality rates and a shorter time to death [98]. Pournaras et al. discovered that the two Colistin-resistant isolates (Ab249 and Ab347, with colistin MICs of 128 and 32 μg/ml, respectively), which acquired resistant through pmrAB mutation, exhibited slower growth compared to their Colistin-susceptible clinical counterparts (Ab248 and Ab299, both having a colistin MIC of 0.5 μg/ml), indicating reduced fitness. Furthermore, one Colistin-resistant isolate demonstrated reduced expression of the Csu system, which is crucial for biofilm formation, along with the outer membrane protein CarO and antioxidant proteins that defend against reactive oxygen species (ROS) generated by macrophages [23]. Further studies by the same researchers showed that A. baumannii strains, Ab249 and Ab347, had a notable decrease in biofilm formation in both static and dynamic assays (P < 0.001) [99]. Nonetheless, a study revealed that LPS-deficient (lpxD mutant) Colistin resistant strains exhibited reduced growth rate, biofilm formation, and biofilm-associated gene expression compared to their Colistin susceptible counterparts. These strains also showed increased susceptibility to azithromycin, vancomycin, and rifampin. In contrast, LPS-modified (pmrB mutant) strains did not display significant differences in these characteristics. The loss of LPS also hindered surface motility, while the expression of type IV pili remained unaffected [100]. LPS plays a critical role in biofilm formation in A. baumannii. The absence of LPS can have a negative impact on biofilm development as it is crucial for surface adhesion and structural stability. However, if LPS undergoes modifications instead of complete loss, biofilm formation may still occur as these modifications may not disrupt the essential processes involved in biofilm. Anyway, LPS loss mutants incur higher fitness costs than pmrB mutants in A. baumannii [101].
The whole genome sequencing (WGS) data of four clinical A. baumannii strains isolated from patients treated with colistin were analyzed to study the evolution and regression of resistance. Colistin resistance developed in all four patients due to mutations in the pmr locus. However, in three cases, susceptible strains became dominant after colistin treatment was stopped. In one case, resistance was lost because of a compensatory mutation that reduced the chances of regaining resistance. Despite initial indications of fitness costs, genomic analysis revealed stable resistance that was not detected by standard clinical tests. Transcriptional studies confirmed increased pmr expression, and adjustments in environmental conditions restored the resistance phenotype [102]. In another study, an immunocompromised patient developed a Col-RAB infection while on long-term colistin therapy. The study examined phenotypic and genotypic characteristics, with a focus on colistin resistance mechanisms and strain fitness. Both colistin-sensitive and colistin-resistant strains were extensively drug-resistant (XDR) and belonged to the same ST78 genotype. Unlike previous findings, the colistin resistance, attributed to a P233S mutation in the PmrB sensor kinase, did not lead to decreased fitness, growth, or virulence [103]. Moreover, whole-genome sequencing of five isolates showed that C440 and C428 were colistin-resistant and genetically identical to their susceptible counterparts. Colistin resistance in C440 was associated with a known P233S mutation in pmrB, while C428 had a novel ΔI19 mutation in pmrB. There were no discernible differences in virulence among isolates C080, C314, and C428 in vitro [104]. Further research is needed to explore the potential impact of compensatory mutations, post-translational modifications, or physiological alterations.
5. Available Therapies and Future Prospects for Col-RAB Infections
The increase in Col-RAB is a major global concern due to the rise of multidrug-resistant strains resulting from the overuse of colistin. This worrisome development threatens to regress clinicians and patients to a time with limited treatment options akin to the pre-antibiotic era [105]. Therefore, there is an urgent need for innovative therapeutic approaches to address A. baumannii infections. The management of Col-RAB is similar to that of carbapenem-resistant strains, but colistin alone is not recommended as a standalone treatment and may no longer be the primary component of combination therapy [106,107,108]. To tackle this issue, potential solutions include the development of new drugs, repurposing existing medications, and exploring combination therapies involving colistin with other agents or adjuvants. Promising strategies such as nanotechnology-based therapies, antimicrobial peptides (AMP), photodynamic therapy, vaccines, CRISPR interference (CRISPRi), and phage therapy offer hope for effectively combating Col-RAB.
Combination of colistin with other drugs has been proposed as an effective strategy to fight against MDR A. baumannii persister cells, which are known as combination therapy. When tobramycin is combined with colistin or ciprofloxacin, it can effectively eliminate persister cells of A. baumannii during the late exponential and stationary growth phases. Colistin helps tobramycin enter the cells by boosting membrane permeability and inducing hyperpolarization of the inner membrane, leading to higher levels of reactive oxygen species (ROS) production [109]. Other antibiotics, including econazole, tigecycline, meropenem, rifampin, fosfomycin, amikacin, ampicillin/sulbactam, polymyxin B, minocycline, eravacycline, ceftazidime/avibactam, trimethoprim/sulfamethoxazole, rifabutin, and azithromycin, are promising combinations with colistin to combat Col-RAB [110,111,112,113,114,115,116,117,118,119,120,121]. In a study on colistin-based combinations, the synergistic effects of two-drug combinations using eight commercially available antibiotics were assessed using the checkerboard approach. Among the combinations tested, vancomycin, aztreonam, ceftazidime, and imipenem showed the highest potency, demonstrating synergistic activity against the strains studied [122]. The combination of fosfomycin or fusidic acid to colistin led to a significantly better microbiological response, as well as a potential improvement in clinical outcomes and lower mortality rates compared to using colistin alone. Fusidic acid was also effective in preventing the development of colistin resistance, a phenomenon that occurred more frequently when colistin was used as a standalone treatment [123,124]. Pregnadiene-11-hydroxy-16,17-epoxy-3,20-dione-1 (PYED-1), a steroid, has shown notable antimicrobial, antibiofilm, and antivirulence properties against both Gram-positive and Gram-negative bacteria, as well as Candida spp. When combined with colistin, PYED-1 can synergistically enhance its antimicrobial efficacy against multidrug-resistant strains of A. baumannii [125].
Adjuvants are non-antibiotic molecules that boost the effectiveness of antibiotics such as colistin. They help lower the necessary treatment dose and reduce its harmful effects. These compounds function by enhancing the antibiotic's penetration into bacterial cells or blocking its expulsion, thus restoring susceptibility in extensively drug-resistant (XDR) strains. This method can be applied to address infections resistant to conventional therapies, presenting a hopeful tactic to tackle drug-resistant bacteria by enhancing the efficiency of current antibiotics. Table 1 lists successful adjuvants used with colistin against Col-RAB.
Drug repurposing is a potential approach in drug discovery that identifies new therapeutic uses for existing drugs to treat resistant infections. This strategy boosts the discovery process by leveraging the known pharmacological properties of these drugs, leading to faster and more cost-effective development compared to developing new drugs. By skipping preclinical trials and moving directly to Phase 2 to test drug efficacy, this approach offers economic advantages and rapidly expands the range of available infection treatments [126,127,128]. Drugs used for anthelmintic purposes, such as Niclosamide, closantel and oxyclozanide restore the colistin activity against Col-RAB and other gram-negative bacteria when used in combination with colistin [129,130,131,132]. Tavaborole was approved by the FDA in July 2014 as an antifungal agent. A recent study by Borges et al. showed that tavaborole has strong antimicrobial activity, with a minimum inhibitory concentration (MIC) value of 2 μg/ml. It also demonstrated potent activity against both the standard ATCC strain and multidrug-resistant (MDR) clinical strains of A. baumannii, as well as effective efficacy against biofilms from multidrug-resistant strains at a concentration of 16 μg/ml [133]. Mitomycin C and 5-fluorouracil are anticancer agents, fluspirilene is an antipsychotic drug, and Bay 11-7082, an inhibitor of κB kinase, is a broad-spectrum drug, that has been shown to be effective against MDR A. baumannii [134,135]. In experimental pneumonia models in mice, the efficacy of N-desmethyltamoxifen, a metabolite of the anticancer agent tamoxifen, was assessed in combination with colistimethate sodium (CMS) or tigecycline. The results demonstrated significant effectiveness against A. baumannii and E. coli strains. Furthermore, combining N-desmethyltamoxifen with antibiotics led to a reduction in bacterial concentrations in the lungs and blood for both A. baumannii strains [136]. A study has identified A. baumannii dihydroorotate dehydrogenase (DHODH) as a potential target for treating drug-resistant infections. Researchers repurposed compounds from a malaria DHODH program and discovered potent inhibitors, such as DSM186, that exhibited strong activity against various A. baumannii strains. Another compound, DSM161, demonstrated protective effects in mouse infection models without any observed resistance. The structure of AbDHODH bound to DSM186 was resolved, confirming its potential as a target for developing new antimicrobial agents [137].
Antimicrobial peptides (AMPs) are an important substitute for antibiotics that affect a wide range of microbes. AMPs offer a potential avenue to fight against MDR, XDR, and Col-RAB through a variety of mechanisms [138,139]. Numerous studies on AMPs suggest that AMPs, either alonee or in combination with colistin, show effectiveness against MDR, XDR and Col-RAB. Peptides such as Esc (1-21), melittin, indolicidin, mastoparan, Ω76, NuriPep 1653, Cec4, 2K4L, LS-sarcotoxin and LS-stomoxyn have demonstrated effectiveness against A. baumannii [140,141,142,143,144,145,146].
Nanotechnology provides a promising solution to combat drug resistance, offering new treatment options and helping to preserve our antibiotic resources [147]. Nanoparticles in combination with colistin exert synergistic effects against Col-RAB. For example, the combination of colistin and silver nanoparticles significantly reduced bacterial growth and viability; this , however, was increased in colistin monotherapy [148]. In addition, colistin-silver nanoparticle synergism led to a reduction in the MICs of colistin by more than four times [149]. A study by Usjak et al. [150] revealed that a combination of low concentrations of colistin (0.5 or 1 μg/mL) and selenium nanoparticles (0.5 μg/mL) significantly decreased the initial bacterial load within the first 4 hours of incubation. This effect was not observed with colistin alone at concentrations of 0.5, 1, or 2 μg/mL. The synergistic action of these agents at low doses implies that these concentrations could be feasibly achieved through systemic administration in vivo.
Antimicrobial photodynamic therapy (APDT) has emerged as a cutting-edge technique in modern health care systems, offering a promising alternative to traditional antibiotic therapy for photo inactivating a wide range of bacterial pathogens [151]. Boluki et al. evaluated photodynamic therapy (PDT) using toluidine blue O (TBO) and a light-emitting diode (LED) as a photosensitizer and radiation source on pandrug-resistant A. baumannii isolated from a burn patient. In this study, PDT was able to eradicate the bacterial infection regardless of resistance by directly modulating the pmrA/pmrB two-component regulatory system activity [152]. A subsequent study has shown that after exposing PDT (TBO and LED), the outer membrane protein A degrades due to the overexpression of the ompA gene in Col-RAB, which could assist in promoting antibiotic penetration [153]. In addition, PDT resulted in an 83.7% decrease in bacterial count compared to the control group. When PDT was used in combination with colistin, a robust synergistic effect was seen against A. baumannii, achieving a 100% kill rate and a 9-log reduction in bacterial load at all tested colistin concentrations. Besides, PDT significantly lowered the minimal inhibitory concentration (MIC) of colistin against Col-RAB by over 11-fold [154].
CRISPR-associated gene editing could offer a new solution to combat PDR A. baumannii. A research has revealed that the clinical isolate AB43, equipped with a full I-Fb CRISPR-Cas system, utilizes the Cas3 nuclease to modulate quorum sensing and influence drug resistance. Through the downregulation of the quorum-sensing synthase AbaI, CRISPR-Cas activity resulted in decreased efflux pump activity, reduced biofilm formation, heightened ROS production, and diminished antibiotic resistance [155]. A study by Wang et al. used CRISPR/Cas9 to remove plasmids in E. coli isolates, making them more susceptible to antibiotics. Plasmids were successfully removed from isolate 14EC033, and multiple plasmids were eliminated in 14EC007. A single sgRNA deleted the colistin-resistant mcr-1 gene in one plasmid, but unintended recombination occurred due to the presence of IS5 upstream of mcr-1 in p14EC033a. While the method was effective for plasmid elimination and gene deletion, caution should be taken to avoid unintended genetic alterations [156].
Phage therapy is one of the most potent therapeutic approaches gaining momentum to address Col-RAB infections. A lysin from bacteriophage PMK34 in combination with colistin resulted in up to a 32-fold reduction of the MIC of colistin and reverted colistin-resistant strains to susceptible in both Mueller-Hinton broth and 50% human serum [157]. The phage effectively reduced biofilms and prevented new ones. When combined, phage vWU2001 and colistin showed decreased bacterial growth. In Galleria mellonella, the combined therapy improved survival and bacterial clearance compared to individual treatments, suggesting a synergistic effect against CRAB [158]. Another lytic phage, IsfAB78, isolated from wastewater was able to reduce the biofilm of Col-RAB by up to 87% [159]. A 68-year-old diabetic patient with necrotizing pancreatitis developed a multidrug-resistant A. baumannii infection that was unresponsive to antibiotics, including colistin and tigecycline. After 4 months of worsening condition, bacteriophage therapy was initiated using nine phages that targeted the bacteria. The treatment was administered intravenously and into abscesses, leading to a reversal of the patient's clinical deterioration and clearance of the infection. Five days into bacteriophage therapy, minocycline was introduced, showing an additional effect in vitro against phage-resistant A. baumannii. This case highlights the potential of combining bacteriophages with antibiotics like colistin and minocycline to treat resistant infections [160].
6. Concluding Remarks
Acinetobacter baumannii employs a complex strategy to resist colistin, with a key mechanism involving the complete loss of lipopolysaccharide (LPS) due to mutations in the lpxA, lpxB, and lpxC genes. These mutations disrupt LPS biosynthesis, making the bacterium less susceptible to the antibiotic. Additionally, modifications to Lipid A structure, such as the addition of phosphoethanolamine (PEtN) or galactosamine, decrease colistin's binding affinity. Other factors contributing to resistance include efflux pumps, changes in outer membrane proteins, and mutations in non-lpx genes. The acquisition of mcr genes through horizontal gene transfer is particularly worrisome as its further boosts colistin resistance in A. baumannii. Additionally, colistin heteroresistance can result in antibiotic treatment failure and may lead to the emergence of colistin-resistant strains if not addressed properly. Taken together, Col-RAB poses a significant challenge to healthcare systems. To address this threat effectively, a comprehensive alternative strategy is needed. Col-RAB strains often exhibit a fitness cost, resulting in reduced virulence due to growth defects and impaired infection capability. Understanding these fitness costs could provide insights into managing antibiotic resistance by exploiting the increased susceptibility to other antibiotics through combination therapies [161,162]. While no single treatment has shown clear superiority, there are various non-polymyxin-based regimens available, with colistin still playing a role in synergistic combination therapies. The recommended treatment approach for carbapenem- and Col-RAB is sulbactam, with sulbactam/durlobactam as the preferred option if possible, or ampicillin/sulbactam as an alternative. Moreover, adjuvants, repurposing of existing drugs, AMPs, nanotechnology, photodynamic therapy, CRISPR/Cas, and phage therapy may serve as alternative treatment options in the future.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (Grant Nos. 2022R1F1A1071415 and NRF-RS-2023-00275307) and Basic Science Research Capacity Enhancement Project through Korea Basic Science Institute (National research Facilities and Equipment Center) grant funded by the Ministry of Education (Grant No. 2019R1A6C1010033).
Authors Contribution
M.M.I: Writing original draft, illustrated figures, designed the review and performed in searching works of literature. W.S.S: Conceptualized the review, editing the manuscript & fund acquisition. M.H.O: Conceptualized, supervised the study, fund acquisition, review and editing the manuscript.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no competing interests.
References
- Jiang M, Chen X, Liu S, Zhang Z, Li N, Dong C, et al. Epidemiological Analysis of Multidrug-Resistant Acinetobacter baumannii Isolates in a Tertiary Hospital Over a 12-Year Period in China. Frontiers in public health. 2021, 9:707435. [CrossRef]
- WHO WHO: WHO Bacterial Priority Pathogens List, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. In.; 2024.
- Serapide F, Guastalegname M, Gullì SP, Lionello R, Bruni A, Garofalo E, et al. Antibiotic Treatment of Carbapenem-Resistant Acinetobacter baumannii Infections in View of the Newly Developed β-Lactams: A Narrative Review of the Existing Evidence. 2024, 13, 506.
- Cain AK, Hamidian M. Portrait of a killer: Uncovering resistance mechanisms and global spread of Acinetobacter baumannii. PLOS Pathogens. 2023, 19, e1011520. [CrossRef]
- Yang S, Wang H, Zhao D, Zhang S, Hu C. Polymyxins: recent advances and challenges. 2024, 15. [CrossRef]
- El-Sayed Ahmed MAE, Zhong LL, Shen C, Yang Y, Doi Y, Tian GB. Colistin and its role in the Era of antibiotic resistance: an extended review (2000-2019). Emerging microbes & infections. 2020, 9, 868–885. [CrossRef]
- Rabi R, Enaya A, Sweileh MW, Aiesh BM, Namrouti A, Hamdan ZI, et al. Comprehensive Assessment of Colistin Induced Nephrotoxicity: Incidence, Risk Factors and Time Course. Infection and drug resistance. 2023, 16:3007-17. [CrossRef]
- Torres DA, Seth-Smith HMB, Joosse N, Lang C, Dubuis O, Nüesch-Inderbinen M, et al. Colistin resistance in Gram-negative bacteria analysed by five phenotypic assays and inference of the underlying genomic mechanisms. BMC Microbiology. 2021, 21, 321. [CrossRef]
- Sharma J, Sharma D, Singh A, Sunita K. Colistin Resistance and Management of Drug Resistant Infections. The Canadian journal of infectious diseases & medical microbiology = Journal canadien des maladies infectieuses et de la microbiologie medicale. 2022, 2022:4315030. [CrossRef]
- Lencina FA, Bertona M, Stegmayer MA, Olivero CR, Frizzo LS, Zimmermann JA, et al. Prevalence of colistin-resistant Escherichia coli in foods and food-producing animals through the food chain: A worldwide systematic review and meta-analysis. Heliyon. 2024, 10, e26579. [CrossRef]
- Kumar H, Chen BH, Kuca K, Nepovimova E, Kaushal A, Nagraik R, et al. Understanding of Colistin Usage in Food Animals and Available Detection Techniques: A Review. Animals : an open access journal from MDPI. 2020, 10(10). [CrossRef]
- Mondal AH, Khare K, Saxena P, Debnath P, Mukhopadhyay K, Yadav D. A Review on Colistin Resistance: An Antibiotic of Last Resort. Microorganisms. 2024, 12(4). [CrossRef]
- Jiang X, Yang K, Han ML, Yuan B, Li J, Gong B, et al. Outer Membranes of Polymyxin-Resistant Acinetobacter baumannii with Phosphoethanolamine-Modified Lipid A and Lipopolysaccharide Loss Display Different Atomic-Scale Interactions with Polymyxins. ACS Infect Dis. 2020, 6, 2698–2708. [CrossRef]
- Ouyang Z, He W, Jiao M, Yu Q, Guo Y, Refat M, et al. Mechanistic and biophysical characterization of polymyxin resistance response regulator PmrA in Acinetobacter baumannii. 2024, 15. [CrossRef]
- Zhang W, Aurosree B, Gopalakrishnan B, Balada-Llasat J-M, Pancholi V, Pancholi P. The role of LpxA/C/D and pmrA/B gene systems in colistin-resistant clinical strains of Acinetobacter baumannii. Frontiers in Laboratory Medicine. 2017, 1, 86–91. [CrossRef]
- Moffatt JH, Harper M, Harrison P, Hale JDF, Vinogradov E, Seemann T, et al. Colistin Resistance in <i>Acinetobacter baumannii</i> Is Mediated by Complete Loss of Lipopolysaccharide Production. 2010, 54, 4971–4977. [CrossRef]
- Hameed F, Khan MA, Muhammad H, Sarwar T, Bilal H, Rehman TU. Plasmid-mediated mcr-1 gene in Acinetobacter baumannii and Pseudomonas aeruginosa: first report from Pakistan. Revista da Sociedade Brasileira de Medicina Tropical. 2019, 52:e20190237. [CrossRef]
- Novović K, Jovčić B. Colistin Resistance in Acinetobacter baumannii: Molecular Mechanisms and Epidemiology. 2023, 12, 516.
- Sabnis A, Hagart KLH, Klöckner A, Becce M, Evans LE, Furniss RCD, et al. Colistin kills bacteria by targeting lipopolysaccharide in the cytoplasmic membrane. eLife. 2021, 10:e65836. [CrossRef]
- Papazachariou A, Tziolos R-N, Karakonstantis S, Ioannou P, Samonis G, Kofteridis DP. Treatment Strategies of Colistin Resistance Acinetobacter baumannii Infections. 2024, 13, 423.
- Roberts JL, Cattoz B, Schweins R, Beck K, Thomas DW, Griffiths PC, et al. In Vitro Evaluation of the Interaction of Dextrin-Colistin Conjugates with Bacterial Lipopolysaccharide. J Med Chem. 2016, 59, 647–654. [CrossRef]
- Gadar K, de Dios R, Kadeřábková N, Prescott TAK, Mavridou DAI, McCarthy RR. Disrupting iron homeostasis can potentiate colistin activity and overcome colistin resistance mechanisms in Gram-Negative Bacteria. Communications biology. 2023, 6, 937. [CrossRef]
- Pournaras S, Poulou A, Dafopoulou K, Chabane YN, Kristo I, Makris D, et al. Growth retardation, reduced invasiveness, and impaired colistin-mediated cell death associated with colistin resistance development in Acinetobacter baumannii. Antimicrobial agents and chemotherapy. 2014, 58, 828–832. [CrossRef]
- Saleh NM, Hesham MS, Amin MA, Samir Mohamed R. Acquisition of Colistin Resistance Links Cell Membrane Thickness Alteration with a Point Mutation in the lpxD Gene in Acinetobacter baumannii. Antibiotics (Basel, Switzerland). 2020, 9(4). [CrossRef]
- Möller A-M, Vázquez-Hernández M, Kutscher B, Brysch R, Brückner S, Marino EC, et al. Common and varied molecular responses of Escherichia coli to five different inhibitors of the lipopolysaccharide biosynthetic enzyme LpxC. Journal of Biological Chemistry. 2024, 300(4). [CrossRef]
- Martínez-Guitián M, Vázquez-Ucha JC, Álvarez-Fraga L, Conde-Pérez K, Bou G, Poza M, et al. Antisense inhibition of lpxB gene expression in Acinetobacter baumannii by peptide-PNA conjugates and synergy with colistin. The Journal of antimicrobial chemotherapy. 2020, 75, 51–59. [CrossRef]
- Wang X, Quinn PJ, Yan A. Kdo2 -lipid A: structural diversity and impact on immunopharmacology. Biological reviews of the Cambridge Philosophical Society. 2015, 90, 408–427. [CrossRef]
- Moffatt JH, Harper M, Harrison P, Hale JD, Vinogradov E, Seemann T, et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrobial agents and chemotherapy. 2010, 54, 4971–4977. [CrossRef]
- Moffatt JH, Harper M, Adler B, Nation RL, Li J, Boyce JD. Insertion sequence ISAba11 is involved in colistin resistance and loss of lipopolysaccharide in Acinetobacter baumannii. Antimicrobial agents and chemotherapy. 2011, 55, 3022–3024. [CrossRef]
- Cafiso V, Stracquadanio S, Lo Verde F, Gabriele G, Mezzatesta ML, Caio C, et al. Colistin Resistant A. baumannii: Genomic and Transcriptomic Traits Acquired Under Colistin Therapy. 2019, 9. [CrossRef]
- Kim S-H, Yun S, Park W. Constitutive Phenotypic Modification of Lipid A in Clinical Acinetobacter baumannii Isolates. 2022, 10, e01295–22. [CrossRef]
- Gerson S, Betts JW, Lucaßen K, Nodari CS, Wille J, Josten M, et al. Investigation of Novel pmrB and eptA Mutations in Isogenic Acinetobacter baumannii Isolates Associated with Colistin Resistance and Increased Virulence In Vivo. Antimicrobial agents and chemotherapy. 2019, 63(3). [CrossRef]
- Sun B, Liu H, Jiang Y, Shao L, Yang S, Chen D, et al. New Mutations Involved in Colistin Resistance in Acinetobacter baumannii. mSphere. 2020, 5(2). [CrossRef]
- Yamada N, Kamoshida G, Shiraishi T, Yamaguchi D, Matsuoka M, Yamauchi R, et al. PmrAB, the two-component system of <i>Acinetobacter baumannii</i>, controls the phosphoethanolamine modification of lipooligosaccharide in response to metal ions. 2024, 206, e00435–23. [CrossRef]
- Hua J, Jia X, Zhang L, Li Y. The Characterization of Two-Component System PmrA/PmrB in Cronobacter sakazakii. 2020, 11. [CrossRef]
- Kato A, Chen HD, Latifi T, Groisman EA. Reciprocal control between a bacterium's regulatory system and the modification status of its lipopolysaccharide. Molecular cell. 2012, 47, 897–908. [CrossRef]
- Adams MD, Nickel GC, Bajaksouzian S, Lavender H, Murthy AR, Jacobs MR, et al. Resistance to Colistin in <i>Acinetobacter baumannii</i> Associated with Mutations in the PmrAB Two-Component System. 2009, 53, 3628–3634. [CrossRef]
- Trebosc V, Gartenmann S, Tötzl M, Lucchini V, Schellhorn B, Pieren M, et al. Dissecting Colistin Resistance Mechanisms in Extensively Drug-Resistant Acinetobacter baumannii Clinical Isolates. mBio. 2019, 10(4). [CrossRef]
- Yang Y-S, Jeng W-Y, Lee Y-T, Hsu C-J, Chou Y-C, Kuo S-C, et al. Ser253Leu substitution in PmrB contributes to colistin resistance in clinical Acinetobacter nosocomialis. Emerging microbes & infections. 2021, 10, 1873–1880. [CrossRef]
- Sun B, Liu H, Jiang Y, Shao L, Yang S, Chen D. New Mutations Involved in Colistin Resistance in Acinetobacter baumannii. mSphere. 2020, 5(2). [CrossRef]
- Karakonstantis, S. A systematic review of implications, mechanisms, and stability of in vivo emergent resistance to colistin and tigecycline in Acinetobacter baumannii. Journal of Chemotherapy. 2020, 33, 1–11. [Google Scholar] [CrossRef]
- Marano V, Marascio N, Pavia G, Lamberti AG, Quirino A, Musarella R, et al. Identification of pmrB mutations as putative mechanism for colistin resistance in A. baumannii strains isolated after in vivo colistin exposure. Microbial pathogenesis. 2020, 142:104058. [CrossRef]
- Adams MD, Nickel GC, Bajaksouzian S, Lavender H, Murthy AR, Jacobs MR, et al. Resistance to colistin in Acinetobacter baumannii associated with mutations in the PmrAB two-component system. Antimicrobial agents and chemotherapy. 2009, 53, 3628–3634. [CrossRef]
- Nurtop E, Bayındır Bilman F, Menekse S, Kurt Azap O, Gönen M, Ergonul O, et al. Promoters of Colistin Resistance in Acinetobacter baumannii Infections. Microbial drug resistance (Larchmont, NY). 2019, 25, 997–1002. [CrossRef]
- Haeili M, Kafshdouz M, Feizabadi MM. Molecular Mechanisms of Colistin Resistance Among Pandrug-Resistant Isolates of Acinetobacter baumannii with High Case-Fatality Rate in Intensive Care Unit Patients. Microbial drug resistance (Larchmont, NY). 2018, 24, 1271–1276. [CrossRef]
- Palethorpe S, Milton ME, Pesci EC, Cavanagh J. Structure of the Acinetobacter baumannii PmrA receiver domain and insights into clinical mutants affecting DNA binding and promoting colistin resistance. Journal of biochemistry. 2022, 170, 787–800. [CrossRef]
- Chin C-Y, Gregg KA, Napier BA, Ernst RK, Weiss DS. A PmrB-Regulated Deacetylase Required for Lipid A Modification and Polymyxin Resistance in Acinetobacter baumannii. 2015, 59, 7911–7914. [CrossRef]
- Pelletier MR, Casella LG, Jones JW, Adams MD, Zurawski DV, Hazlett KR, et al. Unique structural modifications are present in the lipopolysaccharide from colistin-resistant strains of Acinetobacter baumannii. Antimicrobial agents and chemotherapy. 2013, 57, 4831–4840. [CrossRef]
- Zafer MM, Hussein AFA, Al-Agamy MH, Radwan HH, Hamed SM. Retained colistin susceptibility in clinical Acinetobacter baumannii isolates with multiple mutations in pmrCAB and lpxACD operons. Frontiers in cellular and infection microbiology. 2023, 13:1229473. [CrossRef]
- Beceiro A, Llobet E, Aranda J, Bengoechea JA, Doumith M, Hornsey M, et al. Phosphoethanolamine modification of lipid A in colistin-resistant variants of Acinetobacter baumannii mediated by the pmrAB two-component regulatory system. Antimicrobial agents and chemotherapy. 2011, 55, 3370–3379. [CrossRef]
- Trebosc V, Gartenmann S, Tötzl M, Lucchini V, Schellhorn B, Pieren M, et al. Dissecting Colistin Resistance Mechanisms in Extensively Drug-Resistant Acinetobacter baumannii Clinical Isolates. 2019, 10. [CrossRef]
- Potron A, Vuillemenot J-B, Puja H, Triponney P, Bour M, Valot B, et al. ISAba1-dependent overexpression of eptA in clinical strains of Acinetobacter baumannii resistant to colistin. Journal of Antimicrobial Chemotherapy. 2019, 74, 2544–2550. [CrossRef]
- Vijayakumar S, Swetha RG, Bakthavatchalam YD, Vasudevan K, Abirami Shankar B, Kirubananthan A, et al. Genomic investigation unveils colistin resistance mechanism in carbapenem-resistant Acinetobacter baumannii clinical isolates. Microbiology Spectrum. 2024, 12(2). [CrossRef]
- Palmieri M, D’Andrea MM, Pelegrin AC, Perrot N, Mirande C, Blanc B, et al. Abundance of Colistin-Resistant, OXA-23- and ArmA-Producing Acinetobacter baumannii Belonging to International Clone 2 in Greece. 2020, 11. [CrossRef]
- Lesho E, Yoon EJ, McGann P, Snesrud E, Kwak Y, Milillo M, et al. Emergence of colistin-resistance in extremely drug-resistant Acinetobacter baumannii containing a novel pmrCAB operon during colistin therapy of wound infections. The Journal of infectious diseases. 2013, 208, 1142–1151. [CrossRef]
- Deveson Lucas D, Crane B, Wright A, Han ML, Moffatt J, Bulach D, et al. Emergence of High-Level Colistin Resistance in an Acinetobacter baumannii Clinical Isolate Mediated by Inactivation of the Global Regulator H-NS. Antimicrobial agents and chemotherapy. 2018, 62(7). [CrossRef]
- Gaballa A, Wiedmann M, Carroll LM. More than mcr: canonical plasmid- and transposon-encoded mobilized colistin resistance genes represent a subset of phosphoethanolamine transferases. 2023, 13. [CrossRef]
- Stojanoski V, Sankaran B, Prasad BVV, Poirel L, Nordmann P, Palzkill T. Structure of the catalytic domain of the colistin resistance enzyme MCR-1. BMC Biology. 2016, 14, 81. [CrossRef]
- Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. The Lancet Infectious diseases. 2016, 16, 161–168. [CrossRef]
- Hussein NH, Al-Kadmy IMS, Taha BM, Hussein JD. Mobilized colistin resistance (mcr) genes from 1 to 10: a comprehensive review. Molecular biology reports. 2021, 48, 2897–2907. [CrossRef]
- Calero-Cáceres W, Rodríguez K, Medina A, Medina J, Ortuño-Gutiérrez N, Sunyoto T, et al. Genomic insights of mcr-1 harboring Escherichia coli by geographical region and a One-Health perspective. Frontiers in microbiology. 2022, 13:1032753. [CrossRef]
- Lei C-W, Zhang Y, Wang Y-T, Wang H-N. Detection of Mobile Colistin Resistance Gene <i>mcr-10.1</i> in a Conjugative Plasmid from <i>Enterobacter roggenkampii</i> of Chicken Origin in China. 2020, 64. [CrossRef]
- Göpel L, Prenger-Berninghoff E, Wolf SA, Semmler T, Bauerfeind R, Ewers C. Occurrence of Mobile Colistin Resistance Genes mcr-1–mcr-10 including Novel mcr Gene Variants in Different Pathotypes of Porcine Escherichia coli Isolates Collected in Germany from 2000 to 2021. 2024, 4, 70–84.
- Zhang J, Chen L, Wang J, Yassin AK, Butaye P, Kelly P, et al. Molecular detection of colistin resistance genes (mcr-1, mcr-2 and mcr-3) in nasal/oropharyngeal and anal/cloacal swabs from pigs and poultry. Scientific Reports. 2018, 8, 3705. [CrossRef]
- Rahman M, Ahmed S. Prevalence of colistin resistance gene mcr-1 in clinical Isolates Acinetobacter Baumannii from India. International Journal of Infectious Diseases. 2020, 101:81. [CrossRef]
- Seleim SM, Mostafa MS, Ouda NH, Shash RY. The role of pmrCAB genes in colistin-resistant Acinetobacter baumannii. Sci Rep. 2022, 12, 20951. [CrossRef]
- Fan R, Li C, Duan R, Qin S, Liang J, Xiao M, et al. Retrospective Screening and Analysis of mcr-1 and blaNDM in Gram-Negative Bacteria in China, 2010–2019. 2020, 11. [CrossRef]
- Kareem, SM. Emergence of mcr- and fosA3-mediated colistin and fosfomycin resistance among carbapenem-resistant Acinetobacter baumannii in Iraq. Meta Gene. 2020, 25:100708. [CrossRef]
- Martins-Sorenson N, Snesrud E, Xavier DE, Cacci LC, Iavarone AT, McGann P, et al. A novel plasmid-encoded mcr-4.3 gene in a colistin-resistant Acinetobacter baumannii clinical strain. The Journal of antimicrobial chemotherapy. 2020, 75, 60–64. [CrossRef]
- Ma F, Shen C, Zheng X, Liu Y, Chen H, Zhong L, et al. Identification of a Novel Plasmid Carrying <i>mcr-4.3</i> in an <i>Acinetobacter baumannii</i> Strain in China. 2019, 63. [CrossRef]
- Bitar I, Medvecky M, Gelbicova T, Jakubu V, Hrabak J, Zemlickova H, et al. Complete Nucleotide Sequences of mcr-4.3-Carrying Plasmids in Acinetobacter baumannii Sequence Type 345 of Human and Food Origin from the Czech Republic, the First Case in Europe. Antimicrobial agents and chemotherapy. 2019, 63. [CrossRef]
- Lin MF, Lin YY, Lan CY. Contribution of EmrAB efflux pumps to colistin resistance in Acinetobacter baumannii. Journal of microbiology (Seoul, Korea). 2017, 55, 130–136. [CrossRef]
- Boinett CJ, Cain AK, Hawkey J, Do Hoang NT, Khanh NNT, Thanh DP, et al. Clinical and laboratory-induced colistin-resistance mechanisms in Acinetobacter baumannii. Microbial genomics. 2019, 5(2). [CrossRef]
- Cheah SE, Johnson MD, Zhu Y, Tsuji BT, Forrest A, Bulitta JB, et al. Polymyxin Resistance in Acinetobacter baumannii: Genetic Mutations and Transcriptomic Changes in Response to Clinically Relevant Dosage Regimens. Sci Rep. 2016, 6, 26233. [CrossRef]
- Ni W, Li Y, Guan J, Zhao J, Cui J, Wang R, et al. Effects of Efflux Pump Inhibitors on Colistin Resistance in Multidrug-Resistant Gram-Negative Bacteria. Antimicrobial agents and chemotherapy. 2016, 60, 3215–3218. [CrossRef]
- Osei Sekyere J, Amoako DG. Carbonyl Cyanide m-Chlorophenylhydrazine (CCCP) Reverses Resistance to Colistin, but Not to Carbapenems and Tigecycline in Multidrug-Resistant Enterobacteriaceae. 2017, 8. [CrossRef]
- Thi Khanh Nhu N, Riordan DW, Do Hoang Nhu T, Thanh DP, Thwaites G, Huong Lan NP, et al. The induction and identification of novel Colistin resistance mutations in Acinetobacter baumannii and their implications. Scientific Reports. 2016, 6, 28291. [CrossRef]
- Paul D, Mallick S, Das S, Saha S, Ghosh AK, Mandal SM. Colistin Induced Assortment of Antimicrobial Resistance in a Clinical Isolate of Acinetobacter baumannii SD01. Infectious disorders drug targets. 2020, 20, 501–505. [CrossRef]
- Catel-Ferreira M, Marti S, Guillon L, Jara L, Coadou G, Molle V, et al. The outer membrane porin OmpW of Acinetobacter baumannii is involved in iron uptake and colistin binding. 2016, 590, 224–231. [CrossRef]
- Parra-Millán R, Vila-Farrés X, Ayerbe-Algaba R, Varese M, Sánchez-Encinales V, Bayó N, et al. Synergistic activity of an OmpA inhibitor and colistin against colistin-resistant Acinetobacter baumannii: mechanistic analysis and in vivo efficacy. Journal of Antimicrobial Chemotherapy. 2018, 73, 3405–3412. [CrossRef]
- Hood MI, Becker KW, Roux CM, Dunman PM, Skaar EP. genetic determinants of intrinsic colistin tolerance in Acinetobacter baumannii. Infection and immunity. 2013, 81, 542–551. [CrossRef]
- Levin BR, Berryhill BA, Gil-Gil T, Manuel JA, Smith AP, Choby JE, et al. Theoretical considerations and empirical predictions of the pharmaco- and population dynamics of heteroresistance. 2024, 121, e2318600121. [CrossRef]
- Band VI, Weiss DS. Heteroresistance to beta-lactam antibiotics may often be a stage in the progression to antibiotic resistance. PLoS biology. 2021, 19, e3001346. [CrossRef]
- Li J, Rayner CR, Nation RL, Owen RJ, Spelman D, Tan KE, et al. Heteroresistance to colistin in multidrug-resistant Acinetobacter baumannii. Antimicrobial agents and chemotherapy. 2006, 50, 2946–2950. [CrossRef]
- Andersson, DI. The biological cost of mutational antibiotic resistance: any practical conclusions? Current opinion in microbiology. 2006, 9, 461–465. [Google Scholar] [CrossRef]
- Beceiro A, Moreno A, Fernández N, Vallejo JA, Aranda J, Adler B, et al. Biological cost of different mechanisms of colistin resistance and their impact on virulence in Acinetobacter baumannii. Antimicrobial agents and chemotherapy. 2014, 58, 518–526. [CrossRef]
- Da Silva GJ, Domingues S. Interplay between Colistin Resistance, Virulence and Fitness in Acinetobacter baumannii. 2017, 6, 28.
- Jones CL, Singh SS, Alamneh Y, Casella LG, Ernst RK, Lesho EP, et al. <i>In Vivo</i> Fitness Adaptations of Colistin-Resistant Acinetobacter baumannii Isolates to Oxidative Stress. 2017, 61, 10–1128. [CrossRef]
- Oh MH, Shin WS, Mahbub NU, Islam MM. Phage-encoded depolymerases as a strategy for combating multidrug-resistant Acinetobacter baumannii. 2024, 14. [CrossRef]
- Bertani B, Ruiz N. Function and Biogenesis of Lipopolysaccharides. EcoSal Plus. 2018, 8. [CrossRef]
- García-Quintanilla M, Carretero-Ledesma M, Moreno-Martínez P, Martín-Peña R, Pachón J, McConnell MJ. Lipopolysaccharide loss produces partial colistin dependence and collateral sensitivity to azithromycin, rifampicin and vancomycin in Acinetobacter baumannii. International journal of antimicrobial agents. 2015, 46, 696–702. [CrossRef]
- Carretero-Ledesma M, García-Quintanilla M, Martín-Peña R, Pulido MR, Pachón J, McConnell MJ. Phenotypic changes associated with Colistin resistance due to Lipopolysaccharide loss in Acinetobacter baumannii. Virulence. 2018, 9, 930–942. [CrossRef]
- Kamoshida G, Akaji T, Takemoto N, Suzuki Y, Sato Y, Kai D, et al. Lipopolysaccharide-Deficient Acinetobacter baumannii Due to Colistin Resistance Is Killed by Neutrophil-Produced Lysozyme. Frontiers in microbiology. 2020, 11:573. [CrossRef]
- Rolain J-M, Roch A, Castanier M, Papazian L, Raoult D. Acinetobacter baumannii Resistant to Colistin With Impaired Virulence: A Case Report From France. The Journal of infectious diseases. 2011, 204, 1146–1147. [CrossRef]
- Li J, Nation RL, Owen RJ, Wong S, Spelman D, Franklin C. Antibiograms of Multidrug-Resistant Clinical Acinetobacter baumannii: Promising Therapeutic Options for Treatment of Infection with Colistin-Resistant Strains. Clinical Infectious Diseases. 2007, 45, 594–598. [CrossRef]
- Hraiech S, Roch A, Lepidi H, Atieh T, Audoly G, Rolain JM, et al. Impaired virulence and fitness of a colistin-resistant clinical isolate of Acinetobacter baumannii in a rat model of pneumonia. Antimicrobial agents and chemotherapy. 2013, 57, 5120–5121. [CrossRef]
- López-Rojas R, Jiménez-Mejías ME, Lepe JA, Pachón J. Acinetobacter baumannii Resistant to Colistin Alters Its Antibiotic Resistance Profile: A Case Report From Spain. The Journal of infectious diseases. 2011, 204, 1147–1148. [CrossRef]
- López-Rojas R, McConnell MJ, Jiménez-Mejías ME, Domínguez-Herrera J, Fernández-Cuenca F, Pachón J. Colistin resistance in a clinical Acinetobacter baumannii strain appearing after colistin treatment: effect on virulence and bacterial fitness. Antimicrobial agents and chemotherapy. 2013, 57, 4587–4589. [CrossRef]
- Dafopoulou K, Xavier BB, Hotterbeekx A, Janssens L, Lammens C, Dé E, et al. Colistin-Resistant Acinetobacter baumannii Clinical Strains with Deficient Biofilm Formation. Antimicrobial agents and chemotherapy. 2015, 60, 1892–1895. [CrossRef]
- Farshadzadeh Z, Taheri B, Rahimi S, Shoja S, Pourhajibagher M, Haghighi MA, et al. Growth Rate and Biofilm Formation Ability of Clinical and Laboratory-Evolved Colistin-Resistant Strains of Acinetobacter baumannii. Frontiers in microbiology. 2018, 9:153. [CrossRef]
- Mu X, Wang N, Li X, Shi K, Zhou Z, Yu Y, et al. The Effect of Colistin Resistance-Associated Mutations on the Fitness of Acinetobacter baumannii. 2016, 7. [CrossRef]
- Snitkin ES, Zelazny AM, Gupta J, Palmore TN, Murray PR, Segre JA. Genomic insights into the fate of colistin resistance and Acinetobacter baumannii during patient treatment. Genome research. 2013, 23, 1155–1162. [CrossRef]
- Durante-Mangoni E, Del Franco M, Andini R, Bernardo M, Giannouli M, Zarrilli R. Emergence of colistin resistance without loss of fitness and virulence after prolonged colistin administration in a patient with extensively drug-resistant Acinetobacter baumannii. Diagnostic Microbiology and Infectious Disease. 2015, 82, 222–226. [CrossRef]
- Dahdouh E, Gómez-Gil R, Sanz S, González-Zorn B, Daoud Z, Mingorance J, et al. A novel mutation in pmrB mediates colistin resistance during therapy of Acinetobacter baumannii. International journal of antimicrobial agents. 2017, 49, 727–733. [CrossRef]
- Gogry FA, Siddiqui MT, Sultan I, Haq QMR. Current Update on Intrinsic and Acquired Colistin Resistance Mechanisms in Bacteria. 2021, 8. [CrossRef]
- Thadtapong N, Chaturongakul S, Soodvilai S, Dubbs P. Colistin and Carbapenem-Resistant Acinetobacter baumannii Aci46 in Thailand: Genome Analysis and Antibiotic Resistance Profiling. Antibiotics (Basel, Switzerland). 2021, 10(9). [CrossRef]
- Tamma PD, Aitken SL, Bonomo RA, Mathers AJ, van Duin D, Clancy CJ. Infectious Diseases Society of America 2023 Guidance on the Treatment of Antimicrobial Resistant Gram-Negative Infections. Clinical Infectious Diseases. 2023. [CrossRef]
- Saelim W, Changpradub D, Thunyaharn S, Juntanawiwat P, Nulsopapon P, Santimaleeworagun W. Colistin plus Sulbactam or Fosfomycin against Carbapenem-Resistant Acinetobacter baumannii: Improved Efficacy or Decreased Risk of Nephrotoxicity? Infect Chemother. 2021, 53, 128–140.
- Kashyap S, Kaur S, Sharma P, Capalash N. Combination of colistin and tobramycin inhibits persistence of Acinetobacter baumannii by membrane hyperpolarization and down-regulation of efflux pumps. Microbes and Infection. 2021, 23, 104795. [CrossRef]
- Xie M, Chen K, Chan EW, Chen S. Synergistic Antimicrobial Effect of Colistin in Combination with Econazole against Multidrug-Resistant Acinetobacter baumannii and Its Persisters. Microbiol Spectr. 2022, 10, e0093722. [CrossRef]
- Wu H, Feng H, He L, Zhang H, Xu P. In Vitro Activities of Tigecycline in Combination with Amikacin or Colistin Against Carbapenem-Resistant Acinetobacter baumannii. Applied Biochemistry and Biotechnology. 2021, 193, 3867–3876. [CrossRef]
- Abdul-Mutakabbir JC, Yim J, Nguyen L, Maassen PT, Stamper K, Shiekh Z, et al. In Vitro Synergy of Colistin in Combination with Meropenem or Tigecycline against Carbapenem-Resistant Acinetobacter baumannii. 2021, 10, 880.
- Müderris T, Dursun Manyaslı G, Sezak N, Kaya S, Demirdal T, Gül Yurtsever S. In-vitro evaluation of different antimicrobial combinations with and without colistin against carbapenem-resistant Acinetobacter baumannii clinical isolates. European Journal of Medical Research. 2024, 29, 331. [CrossRef]
- Ju YG, Lee HJ, Yim HS, Lee M-G, Sohn JW, Yoon YK. In vitro synergistic antimicrobial activity of a combination of meropenem, colistin, tigecycline, rifampin, and ceftolozane/tazobactam against carbapenem-resistant Acinetobacter baumannii. Scientific Reports. 2022, 12, 7541. [CrossRef]
- Zhu S, Song C, Zhang J, Diao S, Heinrichs TM, Martins FS, et al. Effects of amikacin, polymyxin-B, and sulbactam combination on the pharmacodynamic indices of mutant selection against multi-drug resistant Acinetobacter baumannii. 2022, 13. [CrossRef]
- Shafiee F, Naji Esfahani SS, Hakamifard A, Soltani R. In vitro synergistic effect of colistin and ampicillin/sulbactam with several antibiotics against clinical strains of multi-drug resistant Acinetobacter baumannii. Indian Journal of Medical Microbiology. 2021, 39, 358–362. [CrossRef]
- Ungthammakhun C, Vasikasin V, Changpradub D. A Randomized Controlled Trial of Colistin Combined with Sulbactam: 9 g per Day versus 12 g per Day in the Treatment of Extensively Drug-Resistant Acinetobacter baumannii Pneumonia: An Interim Analysis. 2022, 11, 1112.
- Liang W, Liu XF, Huang J, Zhu DM, Li J, Zhang J. Activities of colistin- and minocycline-based combinations against extensive drug resistant Acinetobacter baumannii isolates from intensive care unit patients. BMC infectious diseases. 2011, 11:109. [CrossRef]
- Ozger HS, Cuhadar T, Yildiz SS, Demirbas Gulmez Z, Dizbay M, Guzel Tunccan O, et al. In vitro activity of eravacycline in combination with colistin against carbapenem-resistant A. baumannii isolates. The Journal of Antibiotics. 2019, 72, 600–604. [CrossRef]
- Zheng Z, Shao Z, Lu L, Tang S, Shi K, Gong F, et al. Ceftazidime/avibactam combined with colistin: a novel attempt to treat carbapenem-resistant Gram-negative bacilli infection. BMC infectious diseases. 2023, 23, 709. [CrossRef]
- Nepka M, Perivolioti E, Kraniotaki E, Politi L, Tsakris A, Pournaras S. In Vitro Bactericidal Activity of Trimethoprim-Sulfamethoxazole Alone and in Combination with Colistin against Carbapenem-Resistant Acinetobacter baumannii Clinical Isolates. Antimicrobial agents and chemotherapy. 2016, 60, 6903–6906. [CrossRef]
- Almutairi, MM. Synergistic activities of colistin combined with other antimicrobial agents against colistin-resistant Acinetobacter baumannii clinical isolates. PLoS One. 2022, 17, e0270908. [Google Scholar] [CrossRef]
- Sirijatuphat R, Thamlikitkul V. Preliminary Study of Colistin versus Colistin plus Fosfomycin for Treatment of Carbapenem-Resistant Acinetobacter baumannii Infections. 2014, 58, 5598–5601. [CrossRef]
- Phee LM, Betts JW, Bharathan B, Wareham DW. Colistin and Fusidic Acid, a Novel Potent Synergistic Combination for Treatment of Multidrug-Resistant Acinetobacter baumannii Infections. 2015, 59, 4544–4550. [CrossRef]
- Stabile M, Esposito A, Iula VD, Guaragna A, De Gregorio E. PYED-1 Overcomes Colistin Resistance in Acinetobacter baumannii. Pathogens (Basel, Switzerland). 2023, 12(11). [CrossRef]
- Gontijo AVL, Pereira SL, de Lacerda Bonfante H. Can Drug Repurposing be Effective Against Carbapenem-Resistant Acinetobacter baumannii? Curr Microbiol. 2021, 79, 13. [CrossRef]
- Peyclit L, Baron SA, Rolain JM. Drug Repurposing to Fight Colistin and Carbapenem-Resistant Bacteria. Frontiers in cellular and infection microbiology. 2019, 9:193. [CrossRef]
- Singh, A. Artificial intelligence for drug repurposing against infectious diseases. Artificial Intelligence Chemistry. 2024, 2, 100071. [Google Scholar] [CrossRef]
- Ayerbe-Algaba R, Gil-Marqués ML, Jiménez-Mejías ME, Sánchez-Encinales V, Parra-Millán R, Pachón-Ibáñez ME, et al. Synergistic Activity of Niclosamide in Combination With Colistin Against Colistin-Susceptible and Colistin-Resistant Acinetobacter baumannii and Klebsiella pneumoniae. 2018, 8. [CrossRef]
- Domalaon R, De Silva PM, Kumar A, Zhanel GG, Schweizer F. The Anthelmintic Drug Niclosamide Synergizes with Colistin and Reverses Colistin Resistance in Gram-Negative Bacilli. Antimicrobial agents and chemotherapy. 2019, 63. [CrossRef]
- Tran TB, Cheah S-E, Yu HH, Bergen PJ, Nation RL, Creek DJ, et al. Anthelmintic closantel enhances bacterial killing of polymyxin B against multidrug-resistant Acinetobacter baumannii. The Journal of Antibiotics. 2016, 69, 415–421. [CrossRef]
- Ayerbe-Algaba R, Gil-Marqués ML, Miró-Canturri A, Parra-Millán R, Pachón-Ibáñez ME, Jiménez-Mejías ME, et al. The anthelmintic oxyclozanide restores the activity of colistin against colistin-resistant Gram-negative bacilli. International journal of antimicrobial agents. 2019, 54, 507–512. [CrossRef]
- Borges KCM, Costa VAF, Neves BJ, Kipnis A, Junqueira-Kipnis AP. New antibacterial candidates against Acinetobacter baumannii discovered by in silico-driven chemogenomics repurposing. PLOS ONE. 2024, 19, e0307913. [CrossRef]
- Cruz-Muñiz MY, López-Jacome LE, Hernández-Durán M, Franco-Cendejas R, Licona-Limón P, Ramos-Balderas JL, et al. Repurposing the anticancer drug mitomycin C for the treatment of persistent Acinetobacter baumannii infections. International journal of antimicrobial agents. 2017, 49, 88–92. [CrossRef]
- Cheng Y-S, Sun W, Xu M, Shen M, Khraiwesh M, Sciotti RJ, et al. Repurposing Screen Identifies Unconventional Drugs With Activity Against Multidrug Resistant Acinetobacter baumannii. 2019, 8. [CrossRef]
- Herrera-Espejo S, Vila-Domínguez A, Cebrero-Cangueiro T, Smani Y, Pachón J, Jiménez-Mejías ME, et al. Efficacy of Tamoxifen Metabolites in Combination with Colistin and Tigecycline in Experimental Murine Models of Escherichia coli and Acinetobacter baumannii. Antibiotics (Basel, Switzerland). 2024, 13. [CrossRef]
- Russo TA, Umland TC, Deng X, El Mazouni F, Kokkonda S, Olson R, et al. Repurposed dihydroorotate dehydrogenase inhibitors with efficacy against drug-resistant <i>Acinetobacter baumannii</i>. 2022, 119, e2213116119. [CrossRef]
- Moghadam MT, Mojtahedi A, Moghaddam MM, Fasihi-Ramandi M, Mirnejad R. Rescuing humanity by antimicrobial peptides against colistin-resistant bacteria. Appl Microbiol Biotechnol. 2022, 106, 3879–3893. [CrossRef]
- Mwangi J, Hao X, Lai R, Zhang ZY. Antimicrobial peptides: new hope in the war against multidrug resistance. Zoological research. 2019, 40, 488–505. [CrossRef]
- Sacco F, Bitossi C, Casciaro B, Loffredo MR, Fabiano G, Torrini L, et al. The Antimicrobial Peptide Esc(1-21) Synergizes with Colistin in Inhibiting the Growth and in Killing Multidrug Resistant Acinetobacter baumannii Strains. Antibiotics (Basel, Switzerland). 2022, 11(2). [CrossRef]
- Vila-Farres X, Garcia de la Maria C, López-Rojas R, Pachón J, Giralt E, Vila J. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clinical Microbiology and Infection. 2012, 18, 383–387. [CrossRef]
- Nagarajan D, Roy N, Kulkarni O, Nanajkar N, Datey A, Ravichandran S, et al. Ω76: A designed antimicrobial peptide to combat carbapenem- and tigecycline-resistant <i>Acinetobacter baumannii</i>. 2019, 5, eaax1946. [CrossRef]
- Mohan NM, Zorgani A, Jalowicki G, Kerr A, Khaldi N, Martins M. Unlocking NuriPep 1653 From Common Pea Protein: A Potent Antimicrobial Peptide to Tackle a Pan-Drug Resistant Acinetobacter baumannii. 2019, 10. [CrossRef]
- Peng J, Wang Y, Wu Z, Mao C, Li L, Cao H, et al. Antimicrobial Peptide Cec4 Eradicates Multidrug-Resistant Acinetobacter baumannii in vitro and in vivo. Drug Design, Development and Therapy. 2023, 17, 977–992. [CrossRef]
- Ji F, Tian G, Shang D, Jiang F. Antimicrobial peptide 2K4L disrupts the membrane of multidrug-resistant Acinetobacter baumannii and protects mice against sepsis. 2023, 14. [CrossRef]
- Hirsch R, Wiesner J, Marker A, Pfeifer Y, Bauer A, Hammann PE, et al. Profiling antimicrobial peptides from the medical maggot Lucilia sericata as potential antibiotics for MDR Gram-negative bacteria. Journal of Antimicrobial Chemotherapy. 2018, 74, 96–107. [CrossRef]
- Hetta HF, Ramadan YN, Al-Harbi AI, E AA, Battah B, Abd Ellah NH, et al. Nanotechnology as a Promising Approach to Combat Multidrug Resistant Bacteria: A Comprehensive Review and Future Perspectives. Biomedicines. 2023, 11. [CrossRef]
- Kumar D, Singhal C, Yadav M, Joshi P, Patra P, Tanwar S, et al. Colistin potentiation in multidrug-resistant Acinetobacter baumannii by a non-cytotoxic guanidine derivative of silver. 2023, 13. [CrossRef]
- Khaled JM, Alharbi NS, Siddiqi MZ, Alobaidi AS, Nauman K, Alahmedi S, et al. A synergic action of colistin, imipenem, and silver nanoparticles against pandrug-resistant Acinetobacter baumannii isolated from patients. Journal of Infection and Public Health. 2021, 14, 1679–1685. [CrossRef]
- Ušjak D, Novović K, Filipić B, Kojić M, Filipović N, Stevanović MM, et al. In vitro colistin susceptibility of pandrug-resistant Ac. baumannii is restored in the presence of selenium nanoparticles. J Appl Microbiol. 2022, 133, 1197–1206. [CrossRef]
- Youf R, Müller M, Balasini A, Thétiot F, Müller M, Hascoët A, et al. Antimicrobial Photodynamic Therapy: Latest Developments with a Focus on Combinatory Strategies. 2021, 13, 1995.
- Boluki E, Kazemian H, Peeridogaheh H, Alikhani MY, Shahabi S, Beytollahi L, et al. Antimicrobial activity of photodynamic therapy in combination with colistin against a pan-drug resistant Acinetobacter baumannii isolated from burn patient. Photodiagnosis and Photodynamic Therapy. 2017, 18:1-5. [CrossRef]
- Boluki E, Moradi M, Azar PS, Fekrazad R, Pourhajibagher M, Bahador A. The effect of antimicrobial photodynamic therapy against virulence genes expression in colistin-resistance Acinetobacter baumannii. Laser therapy. 2019, 28, 27–33. [CrossRef]
- Pourhajibagher M, Kazemian H, Chiniforush N, Bahador A. Evaluation of photodynamic therapy effect along with colistin on pandrug-resistant Acinetobacter baumannii. Laser therapy. 2017, 26, 97–103. [CrossRef]
- Wang Y, Yang J, Sun X, Li M, Zhang P, Zhu Z, et al. CRISPR-Cas in Acinetobacter baumannii Contributes to Antibiotic Susceptibility by Targeting Endogenous AbaI. Microbiol Spectr. 2022, 10, e0082922. [CrossRef]
- Wang P, He D, Li B, Guo Y, Wang W, Luo X, et al. Eliminating mcr-1-harbouring plasmids in clinical isolates using the CRISPR/Cas9 system. The Journal of antimicrobial chemotherapy. 2019, 74, 2559–2565. [CrossRef]
- Abdelkader K, Gutiérrez D, Grimon D, Ruas-Madiedo P, Lood C, Lavigne R, et al. Lysin LysMK34 of <i>Acinetobacter baumannii</i> Bacteriophage PMK34 Has a Turgor Pressure-Dependent Intrinsic Antibacterial Activity and Reverts Colistin Resistance. 2020, 86, e01311–20. [CrossRef]
- Wintachai P, Phaonakrop N, Roytrakul S, Naknaen A, Pomwised R, Voravuthikunchai SP, et al. Enhanced antibacterial effect of a novel Friunavirus phage vWU2001 in combination with colistin against carbapenem-resistant Acinetobacter baumannii. Scientific Reports. 2022, 12, 2633. [CrossRef]
- Ebrahimi S, Sisakhtpour B, Mirzaei A, Karbasizadeh V, Moghim S. Efficacy of isolated bacteriophage against biofilm embedded colistin-resistant Acinetobacter baumannii. Gene Reports. 2021, 22, 100984. [CrossRef]
- Schooley RT, Biswas B, Gill JJ, Hernandez-Morales A, Lancaster J, Lessor L, et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter baumannii Infection. 2017, 61. [CrossRef]
- Helekal D, Keeling M, Grad YH, Didelot X. Estimating the fitness cost and benefit of antimicrobial resistance from pathogen genomic data. 2023, 20, 20230074. [CrossRef]
- Oyejobi GK, Olaniyan SO, Yusuf N-A, Ojewande DA, Awopetu MJ, Oyeniran GO, et al. Acinetobacter baumannii: More ways to die. Microbiological Research. 2022, 261, 127069. [CrossRef]
- Xu M, Yao Z, Kong J, Tang M, Liu Q, Zhang X, et al. Antiparasitic nitazoxanide potentiates colistin against colistin-resistant Acinetobacter baumannii and Escherichia coli in vitro and in vivo. Microbiol Spectr. 2024, 12, e0229523. [CrossRef]
- Minrovic BM, Jung D, Melander RJ, Melander C. New Class of Adjuvants Enables Lower Dosing of Colistin Against Acinetobacter baumannii. ACS Infect Dis. 2018, 4, 1368–1376. [CrossRef]
- Zhao Y, Liu Y, Feng L, Xu M, Wen H, Yao Z, et al. In vitro and in vivo synergistic effect of chrysin in combination with colistin against Acinetobacter baumannii. 2022, 13. [CrossRef]
- Kaur A, Sharma P, Capalash N. Curcumin alleviates persistence of Acinetobacter baumannii against colistin. Scientific Reports. 2018, 8, 11029. [CrossRef]
- Thadtapong N, Chaturongakul S, Napaswad C, Dubbs P, Soodvilai S. Enhancing effect of natural adjuvant, panduratin A, on antibacterial activity of colistin against multidrug-resistant Acinetobacter baumannii. Scientific Reports. 2024, 14, 9863. [CrossRef]
- Koller BH, Jania LA, Li H, Barker WT, Melander RJ, Melander C. Adjuvants restore colistin sensitivity in mouse models of highly colistin-resistant isolates, limiting bacterial proliferation and dissemination. 2024, 68, e00671–24. [CrossRef]
- Feng X, Liu S, Wang Y, Zhang Y, Sun L, Li H, et al. Synergistic Activity of Colistin Combined With Auranofin Against Colistin-Resistant Gram-Negative Bacteria. 2021, 12. [CrossRef]
- Vázquez-Ucha JC, Martínez-Guitián M, Lasarte-Monterrubio C, Conde-Pérez K, Arca-Suárez J, Álvarez-Fraga L, et al. Syzygium aromaticum (clove) and Thymus zygis (thyme) essential oils increase susceptibility to colistin in the nosocomial pathogens Acinetobacter baumannii and Klebsiella pneumoniae. Biomedicine & Pharmacotherapy. 2020, 130:110606. [CrossRef]
- Chen C, Cai J, Shi J, Wang Z, Liu Y. Resensitizing multidrug-resistant Gram-negative bacteria to carbapenems and colistin using disulfiram. Communications biology. 2023, 6, 810. [CrossRef]
- Guo T, Li M, Sun X, Wang Y, Yang L, Jiao H, et al. Synergistic Activity of Capsaicin and Colistin Against Colistin-Resistant Acinetobacter baumannii : In Vitro/Vivo Efficacy and Mode of Action. Front Pharmacol. 2021. [CrossRef]
Figure 1.
Mechanisms of action of colistin against A. baumannii. SOD (Superoxide dismutase). (Created with Biorender.com).
Figure 1.
Mechanisms of action of colistin against A. baumannii. SOD (Superoxide dismutase). (Created with Biorender.com).
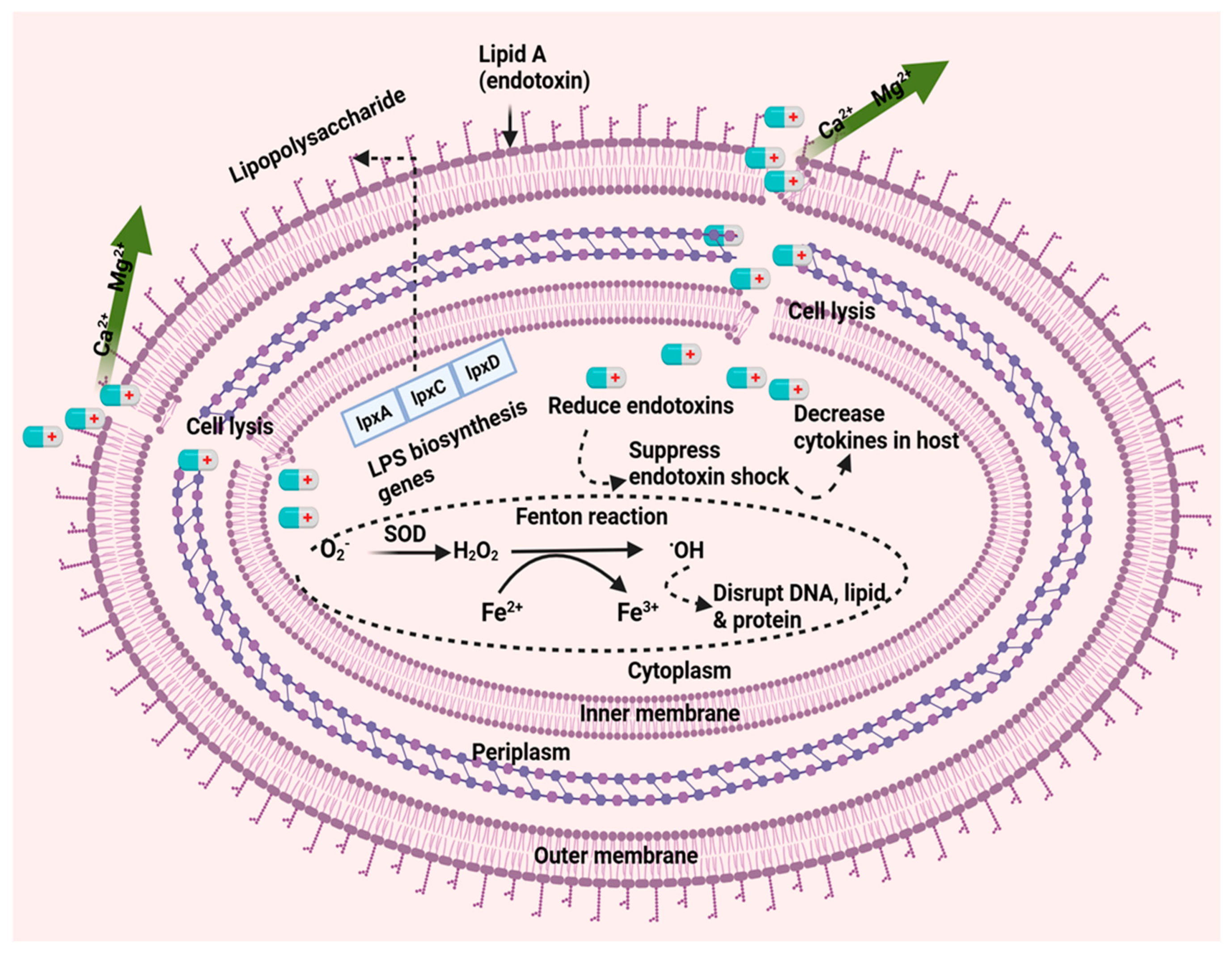
Figure 2.
A schematic illustration showing the general mechanisms of colistin resistance in A. baumannii. LPS (Lipopolysaccharide), GalN (Galactosamine), PEtN ( Phosphoethanolamine). (Created with Biorender.com).
Figure 2.
A schematic illustration showing the general mechanisms of colistin resistance in A. baumannii. LPS (Lipopolysaccharide), GalN (Galactosamine), PEtN ( Phosphoethanolamine). (Created with Biorender.com).
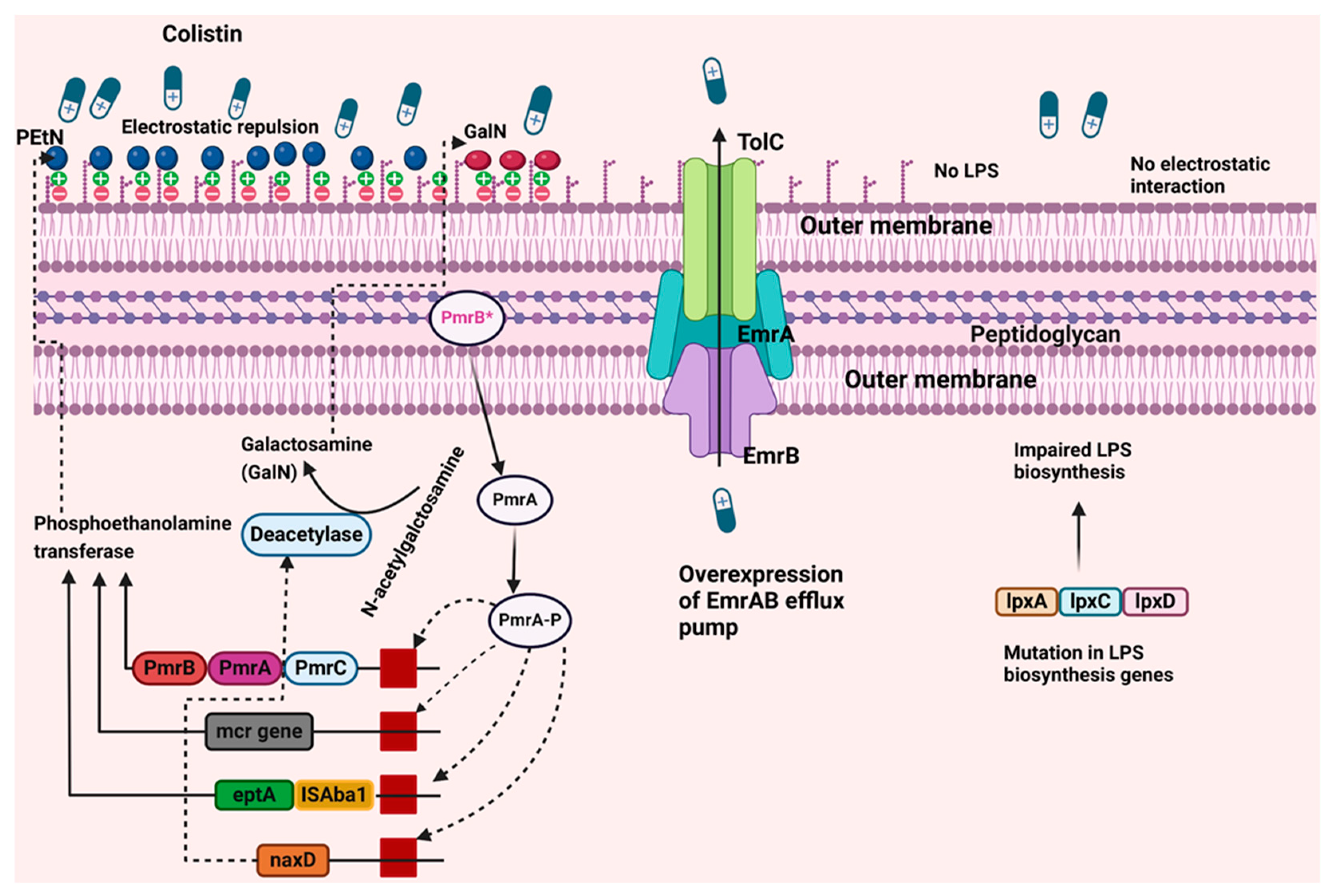

Table 1.
Compounds combine with colistin as adjuvants to fight against colistin-resistant A. baumannii infections.
Table 1.
Compounds combine with colistin as adjuvants to fight against colistin-resistant A. baumannii infections.
| Compounds | Subjects | Mechanisms | References |
|---|---|---|---|
| Kaempferol | Galleria mellonella | Disruption of iron homeostasis and fenton reaction leading to the generation of reactive oxygen species. | [22] |
| Nitazoxanide | Galleria mellonella | Inhibit biofilm | [163] |
| 2-aminoimidazole | Galleria mellonella | [164] | |
| Chrysin | Galleria mellonella/ Mouse | Damage extracellular membrane and modify bacterial membrane potential | [165] |
| Curcumin | In-vitro | Increase ROS and function as efflux pump inhibitor | [166] |
| AOA-2 | Mouse | OmpA inhibitor, significantly decrease the bacterial cell load in tissues and blood, and increased mouse survival rate | [80] |
| Panduratin A | In vitro | Inhibit biofilm formation | [167] |
| IMD-0354 | Mouse | Inhibit lipid A modification | [168] |
| NMD-27 | Mouse | [168] | |
| Auranofin | Mouse | Inhibition of MCR | [169] |
| Essential oils of clove and thyme | In vitro | Increase cell membrane permeability | [170] |
| Disulfiram | Mouse | Damage bacterial membrane and disrupt metabolism | [171] |
| Capsaicin | Mouse | Inhibit the biofilm formation, Disrupt outer membrane, Inhibition of protein synthesis and efflux pumps activity | [172] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



