
Submitted:
31 October 2024
Posted:
01 November 2024
You are already at the latest version
Abstract
The increasing global demand for food caused by a growing world population has resulted in environmental problems, such as the destruction of ecologically significant biomes and pollution of ecosystems. At the same time, the intensification of crop production in modern agriculture has led to the extensive use of synthetic fertilizers to achieve higher yields. Although chemical fertilizers provide essential nutrients and accelerate crop growth, they also pose significant health and environmental risks, including pollution of groundwater and and other bodies of water such as rivers and lakes. Soils that have been destabilized by indiscriminate clearing of vegetation undergo a desertification process that has profound effects on microbial ecological succession, impacting biogeochemical cycling, and thus the foundation of the ecosystem. Tropical countries have positive aspects that can be utilized to their advantage, such as warmer climates leading to increased primary productivity and, as a result, greater biodiversity. As an eco-friendly, cost-effective, and easy-to-apply alternative, biofertilizers have emerged as a solution to this issue. Biofertilizers consist of a diverse group of microorganisms able to promote plant growth and enhance soil health, even under challenging abiotic stress conditions. They can include plant growth-promoting rhizobacteria, arbuscular mycorrhizal fungi, and other beneficial microbial consortia. Bioremediators, on the other hand, are microorganisms that can reduce soil and water pollution or otherwise improve impacted environments. So, the use of microbial biotechnology relies on understanding the relationships between microorganisms and how they affect their environment, and, inversely, how abiotic factors influence microbial activity. The more recent introduction of genetically modified microorganisms into the gamut of biofertilizers and bioremediators requires further studies to assess potential adverse effects in various ecosystems. This article reviews and discusses these two soil correcting/improving processes with the aim of stimulating their use in developing tropical countries.
Keywords:
microorganisms
; biostimulation
; biogeochemical cycles
; extreme environments
; soil pollution
1. Introduction
Developing countries face a series of critical environmental processes due to the mismatch between their pace of development and their role in the global context [1]. Given their relatively short history, they have not had sufficient time to catch up with the global pace set by developed nations. Consequently, these countries are already showing signs of the impact of recent industrialization and urbanization [2]. Rapid development, population growth, inadequate responses to climate and environmental risks, and inefficiencies in governance and environmental management pose substantial environmental challenges for developing countries.
A significant number of these countries are located in warmer climates, allowing them to become major global food producers and leading to the excessive use of their land for agriculture. This can lead to biomes rich in biodiversity being destroyed to make way for monocultures of food plants, depleting ecosystems and their soils. Chemical fertilizers introduced into these systems to boost agricultural productivity disrupt the local soil geochemical balance and affect adjacent environments such as rivers, lakes, and coastal areas [3]. Ultimately, this process can lead to imbalances in water bodies, resulting in hyperproductivity and subsequent eutrophication [4].
In these sense, tropical soil ecosystems exhibit a rich tapestry of diversity and complexity, setting them apart from their temperate counterparts. Within the expansive realms of the tropics, a myriad of ecosystems serves as the canvas for diverse microbial niches and evolutionary processes. The relationship between latitude and bacterial diversity is not statistically significant, but communities can be aptly characterized by their ecotypes and functional attributes. Delving into desert soils, where extremes of solar radiation, minimal precipitation, and fluctuating temperatures prevail, one discovers the existence of intricately diverse bacterial communities, a subject briefly touched upon here. In the semi-arid tropical biomes dominated by grassy vegetation, a resilient and dry-adapted bacterial biodiversity coexists harmoniously with plants [5]. Venturing into water-rich tropical environments, encompassing rainforest soils and the sediments of mangroves, unveils an astonishing array of diversity, harboring untapped biotechnological potential. It’s noteworthy that tropical soils play a crucial role in providing sustenance for around 40% of the world’s population, with a significant portion residing in developing countries. In Brazil, the application of microbial inoculants derived from tropical soils has yielded substantial improvements in large-scale food production. This particular brand of biotechnology is now being shared by Embrapa with tropical nations in Africa. The utilization of tropical soil bacteria extends beyond agriculture, with applications in remediating polluted soils, promoting reforestation, and safeguarding water resources. However, the imperative lies in scaling up these efforts across all tropical regions. Preserving designated areas within the intricate tapestry of tropical ecosystems becomes paramount, ensuring that the invaluable soil microbial gene pool remains intact for the benefit of future generations [6].
Soil, plant and water-associated microorganisms play a vital role in ecosystem functioning, participating in biogeochemical cycles and degrading organic matter [7]. Over 80 coexisting beneficial soil microorganisms, including photosynthetic bacteria, lactic acid bacteria, yeasts, actinomycetes, and fermenting fungi had been identified up to 2014 [8]; with the greater use of DNA-based technologies, this number has increased substantially. These microorganisms can be employed as biofertilizers, to replace the harmful chemicals still being used. Ranipa et al. (2023) [9] point out the role of biofertilizers in sustainable agriculture and review the organisms and their applications. Zhang et al. (2021) [10] investigated the role of arbuscular mycorrhizal fungi in symbiosis with plants, revealing their catalytic effect on organic matter decomposition. This symbiosis also promoted soil aggregation and contributed to soil stabilization through mycelial growth, ultimately enhancing nutrient uptake, particularly phosphorus, by plants. Kheirfam et al. (2017) [11] demonstrated that soil crusts enriched with beneficial microorganisms improved soil properties; poor soils could be improved by adding cyanobacteria, for instance. The application of these bacteria proved to be a rapid, long-lasting, environmentally friendly, and cost-effective technique for enhancing soil quality [11]. Liu et al. (2022) [12] demonstrated that the application of biofertilizers resulted in the increased presence of beneficial bacteria within the sugarcane rhizosphere, while reducing the population of pathogenic bacteria (such as Leifsonia). This shift in microbial communities could help mitigate or hinder the development of diseases.
These examples underscore the significance of biofertilizers, microorganism-based fertilizers, in sustainable agriculture; they have a long-lasting impact on soil fertility [13,14]. In addition, the ability of microorganisms to balance soils and water bodies and to degrade toxic compounds provides them with another important use, commonly referred to as bioremediation.
Biofertilizers and bioremediators are formulations comprising living microbial cells, either a single strain or multiple strains (a consortium), that enhance plant growth by increasing nutrient availability and acquisition and reducing adverse conditions. The term “biofertilizer” has evolved over the past 30 years and has received various interpretations [15,16]. Macik et al. (2020) [16] point out that the most common misconception in nomenclature arises when microbial inoculants with other beneficial activities (e.g., biopesticides and phytostimulators) are included under the umbrella of biofertilizers. Plant growth-promoting bacteria and rhizobacteria (PGPB/PGPR) may also be distinguished, as not all PGPB/PGPR function as biofertilizers [17]. However, it is worth mentioning that biofertilizers can offer additional direct and indirect benefits for plant growth, including phytostimulation, enhanced tolerance to abiotic stress, and control of plant pathogens [18,19,20].
2. Biofertilizers and Bioremediators
One of the key elements of fertile soil is the presence of beneficial soil microbiota, which play a crucial role in enhancing the nutrient pool through biogeochemical processes in the soil. Consequently, it is essential for biofertilizers (microbial inoculants) to be compatible with the soil environment [21]. However, a significant proportion of arable soils worldwide do not offer ideal cultivation conditions due to various abiotic stressors, such as organic pollutants [22], heavy metals [23], drought [24], salinity [25], and extreme temperature variations [26]. Among these stressors, the release of metals from diverse human activities [27,28,29] poses a substantial threat to the sustainability of crop production systems.
Biofertilizers and bioremediators consist of organic or composite products containing living microorganisms with the ability to enhance biogeochemical cycles and, as a result, increase primary productivity and environmental health. The basic difference between the two lies in their purpose and consequent composition: Biofertilizers are used to enhance crop productivity, while bioremediators are employed for the removal of excessive or toxic compounds [30]. In the particular case of biofertilizers, they are mainly used in agriculture as an alternative to traditional chemical fertilizers, since they can play a significant role in providing essential plant nutrients, such as nitrogen, phosphorus, and potassium, in a more sustainable manner. Various biofertilizer types are available, including nitrogen-fixing varieties, mycorrhizae fostering symbiotic associations between fungi and plant roots to enhance nutrient absorption, phosphorus-solubilizing bacteria, and liquid fertilizers containing microorganisms that facilitate organic matter decomposition, thus enhancing soil structure and nutrient availability [31]. In comparison to chemical fertilizers, biofertilizers are deemed more sustainable, as they reduce the dependence on harsh chemicals, promote soil well-being, and, in many instances, encourage environmentally-friendly agricultural practices [32,33,34,35,36]. Bioremediators, on the other hand, contribute to the reduction of pollution and the associated environmental impact stemming from conventional agriculture, deforestation of forests with concomitant soil degradation, and the ultimate contamination of soil and water bodies.
2.1. Agricultural Fertilization
The global human population continues to expand at a concerning rate, giving rise to various issues, including food insecurity [37,38]. To ensure the supply of food, it is imperative to enhance crop production through fertile agrosystems. Soil fertility and crop productivity are often used interchangeably; however, they differ significantly. Soil fertility refers to the inherent capacity of the soil to provide essential plant nutrients in sufficient quantities [39,40], while crop productivity is defined as H/P x Y, where H is acres harvested, P is acres planted and Y is the yield per acre. Both, however, rely on the availability of essential plant nutrients such as N, P, K, Ca, Mg, S, Cu, Cl and Si. These nutrients are produced by the natural decomposition of soil organic matter and the addition of chemical fertilizers. Inoculating biofertilizers represents a promising approach for enhancing crop productivity while decreasing reliance on synthetic fertilizers, thereby promoting environmentally sustainable agriculture [40,41,42].
Microbial inoculants play a crucial role in accelerating the decomposition process, which results in the release of these essential nutrients. This, in turn, leads to an overall increase in crop productivity [43]. As the global population currently stands at around 8 billion people and is projected to reach 9.7 billion by 2050, there is a pressing need to produce 321 million tons of food to feed this growing populace [44]. However, relying solely on chemical fertilizers is no longer a sustainable solution due to their cost and detrimental effects on soil health. The emphasis is shifting toward cost-effective, environmentally friendly, and sustainable biofertilizers, which not only enhance the physical, chemical, and biological properties of the soil but also boost crop yield per unit area [45].
Biofertilizers are also gaining attention for their potential use in challenging environments such as elevated temperatures, saline soils, water scarcity, fluctuating pH levels, and the presence of environmental stressors like protein and heavy metal contaminants [46]. Their impact extends to the modulation of microbial communities within the rhizosphere, thus exerting an influence on the soil’s overall ecosystem [47].
The use of biofertilizers enhances both the function and structure of soil microorganisms and has implications for the physicochemical properties of the soil [48]. The effects of introducing plant growth-promoting rhizobacteria (PGPR) can be quite variable, impacting indigenous microbial populations in various ways. Some remain unaffected, while others may experience either stimulation or inhibition of their growth [49]. For instance, the probiotic strain Stenotrophomonas acidaminiphila BJ1 increased the bacterial population in the rhizosphere of Vicia faba in chlorothalonil-polluted soil [50]. Numerous studies have assessed the impact of microbial inoculants on soil microorganisms [51,52,53]. As early examples, we may cite Upadhyay et al. (2012a) [54], who reported positive effects on the growth and antioxidant status of wheat under saline conditions when inoculated with two PGPR strains, Bacillus subtilis SU47 and Arthrobacter sp. SU18, and Gangwar et al. (2013)[55], who observed enhanced root length, shoot length, root dry weight, and shoot dry weight of Mung Bean (Vigna radiata L.) following dual inoculation with plant growth-promoting Pseudomonas putida and Trichoderma viride.
2.2. Afforestation and Biostimulation
Afforestation involves the establishment of trees on land that has not been previously managed for forests or has been without forests for a minimum of 50 years (UNFCCC) [56]. It serves as a crucial restoration method for reclaiming abandoned agricultural and degraded land. A substantial portion of today’s agricultural land was once covered by forests. The ever-increasing human population and the rising need for food and fiber led to the expansion of agricultural activities, resulting in the conversion of forested areas into agricultural land [57]. The global agricultural land area increased from 15.84106 km2 in 1983 to 16.79106 km2 in 2003 [58]. As depicted in Figure 1, the escalating demand for agricultural land has caused the world to lose approximately one-third of its forests over the centuries.
The growing human and livestock population, uncontrolled extraction of forest resources, frequent forest fires, and mining operations have all contributed to soil erosion, diminished fertility, reduced moisture levels, and declining forest productivity. These issues create a multitude of challenges in ecosystem restoration. Consequently, soil-residing microbial inoculants play a vital and integral role in our soils, primarily by enhancing plant growth through nutrient accessibility, nitrogen fixation, the mobilization of otherwise unavailable nutrients, and the production of antifungal substances [59]. More specifically, deforestation for cattle production brings a series of impacts to the soil. Soil degradation can be classified as one of the most perilous human activities on the Earth’s surface, due to the fact that soil is not immediately renewable [60]. When deforestation reaches a stage that makes reoccupation by secondary forest impossible, the soil becomes exposed or occupied by low grasses, leading to changes in several abiotic factors that may result in desertification. Factors such as inappropriate temperature and salinity can make it impossible for the reestablishment of plant ecological succession, thus preventing the recolonization of deforested areas. Below, some impacts of deforestation on the abiotic parameters of the soil are listed, mentioning how the application of biofertilizers can contribute to the environmental restoration process. It should be taken into consideration, however, that the majority of studies on biostimulation are related to agriculture, with limited tests conducted on native species in tropical forests.
2.2.1. Temperature
Once soil is exposed by deforestation, the direct incidence of sunlight on the soil surface causes an increase in ambient temperature. This affects plant physiology by increasing respiration rates and leaf transpiration, altering photosynthate allocation [61,62]. The enzyme ribulose bisphosphate carboxylase/oxygenase (Rubisco), which produces organic carbon from the inorganic carbon dioxide in the air (Figure 2), is one of those affected.
At high temperatures, Rubisco’s affinity for carbon dioxide decreases, while its affinity for oxygen increases [64,65]. Carbon dioxide solubility decreases more than that of oxygen with rising temperatures, resulting in reduced carbon dioxide concentration in the chloroplast compared to oxygen [66]. Additionally, plants close their stomata to reduce water loss through evapotranspiration when temperatures rise. Stomatal closure leads to a rapid decline in carbon dioxide concentration, while oxygen concentration increases, limiting photosynthesis and increasing photorespiration [67].
Heat stress triggers complex molecular, biochemical, and physiological responses in plants [65], leading to the synthesis of heat shock proteins, reactive oxygen species (ROS), osmoprotectant compounds, amino acids, sugars, and sulfur compounds [68]. Consequently, heat stress stimulates oxidative stress and ROS production, which are detected by histidine kinases and heat stress factors (Hsfs). Redox-sensitive transcription factors downstream from these signals are activated through the mitogen–activated-protein kinases signaling pathway, subsequently turning on other transcription factors (e.g., BF1c and Rboh) to trigger the expression of genes involved in the synthesis of antioxidant enzymes [69]. However, although ROS accumulate during abiotic stresses, such as heat, ROS themselves are toxic and can react with cell components; the classical response to heat stress involves Hsfs, HSPs and various levels of ROS [70]. The heat shock response is a complex and finely tuned system.
Hormonal signaling plays a significant role in heat stress responses, with ethylene being one of the key players [71,72,73]. Ethylene is involved not only in processes like senescence, development, and plant physiology but also in plant responses to various abiotic stressors, including heat, salinity, and drought [72,73,74]. Protection of plants against heat stress can be enhanced by microbial biostimulants. For instance, the synthesis of enzymes that degrade ROS, such as superoxide, catalase, peroxidases, and dismutase, can improve heat stress tolerance in plants. This enhancement can be observed in plants colonized by beneficial bacteria like Pseudomonas and Bacillus, as well as mycorrhizal fungi like Septoglomus deserticola and Septoglomus constrictum [75]. Some available biostimulants contain P. fluorescens and P. aeruginosa, which contribute to improving soil quality and heat stress tolerance, working as bioremediators, phytostimulators, and enhancing soil fertility [65]. Similarly, Bacillus spp. have been developed not only as biopesticides but also in the form of biostimulant products. While these microorganisms are utilized in various commercial treatments, whether individually or in combination, their potential to mitigate heat stress is not always highlighted as one of their benefits [76].
The use of microorganisms capable of reducing ethylene emissions holds promise because decreasing ethylene levels during stressful situations could help plants avoid the detrimental effects of heat stress [77]. Bacteria that possess 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase activity appear particularly promising, including species like Bacillus subtilis BERA 7, Leclercia adecarboxylata MO1, Pseudomonas fluorescens YsS6, and Pseudomonas migulae 8R6 [78]. For example, the ACC deaminase-producing Paraburkholderia phytofirmans PsJN has been found to support normal tomato development under heat stress [79]; however, this strain has not yet been commercialized [65].
Despite the documented positive effects of biostimulants as mitigators of thermal stress, further research is needed to better understand their mechanisms of action and to develop efficient formulations that can mitigate the impact of heat stress in the recuperation of desertified areas.
2.2.2. Salinity
The soil serves as a reservoir of biodiversity, but the worldwide issue of soil salinization poses a significant threat to crop cultivation, affecting all living organisms and hindering the achievement of sustainable development goals, particularly in ensuring food security. Salinity in the soil can be attributed to a variety of natural and human activities. Reduced water availability and osmotic pressure in saline soil are indicated by poor seed germination, leaf wilting, and in extreme instances, the death of plants [80]. Mitsuchi et al. (1989) [81] conducted a review of both natural and human-induced factors contributing to soil salinization. Of the various human-induced factors, salinization resulting from deforestation was considered to be prevalent. In saline soil, the accumulation of solutes reduces the osmotic potential of the liquid phase, limiting water absorption by plant roots [82]. Soil salinization affects approximately 30% of total arable land in Mediterranean regions [83]. Drought stress and the resulting soil salinization are major contributors to desertification in overexploited regions, affecting soil biodiversity and composition, and leading to plant deterioration, reduced soil coverage, and subsequent soil erosion [84]. Drylands now cover 46% of the world’s land surface, impacting around 250 million people in developing countries [85].
Plant tolerance to drought and salinity can be enhanced by microbial biostimulants through various direct and plant-mediated processes. For example, microbial biostimulants can produce bacterial exopolysaccharides that improve soil structure by forming micro and macroaggregates [86], promoting plant growth under water stress conditions [65]. Additionally, exopolysaccharides form hydrophilic biofilms, creating a microenvironment that retains water by shielding microbes from drought stress [87] and binding Na+ ions to reduce their uptake by plants [88,89]. Examples of bacteria that produce protective exopolysaccharides include Pseudomonas spp. PF23 [88], Pseudomonas putida GAP-P45 [90] and Bacillus licheniformis [91]. Root capacity can also be strengthened by mycorrhizal fungi [92], leading to increased root biomass, improved soil structure, enhanced water retention, and reduced leaching of mineral nutrients [93,94]. For instance, the arbuscular mycorrhizal fungus, Glomus sp., produces a glycoprotein called glomalin, which enhances soil structure, plant growth, and drought tolerance [95]. Similarly, ascomycetes (P. glomerata LWL2, Exophiala sp. LHL08, P. formosus LHL10, and Penicillium sp. LWL3) colonize cucumber plants, enhancing leaf development and chlorophyll content under drought stress [92,96].
In addition to promoting root development, mycorrhizal fungi can improve water absorption through aquaporins [97], a family of integral membrane transporters that facilitate water movement through the cell membrane [98]. For example, Glomus intraradices colonizes common bean plants and mitigates water stress by affecting aquaporin activity and improving water conductivity in the roots [97,99]. G. intraradices, often found in commercial products, is typically formulated in combination with various beneficial bacterial and fungal strains or is available in single formulations [76].
Microbial biostimulants offer various supplementary benefits when it comes to alleviating the effects of drought stress in associated plants. These advantages encompass the enhancement of antioxidant defenses, the synthesis of protective osmolytes such as glycine betaine, phytohormones, and the emission of volatile organic compounds (VOCs) [100,101]. Drought stress often triggers increased ethylene production, which impedes plant growth. Microbial biostimulants, like Pseudomonas fluorescens, can address this issue by reducing aminocyclopropane-1-carboxylic acid (ACC) levels, consequently restricting ethylene production and mitigating ethylene-mediated inhibition [77]. Numerous microbial species, including Staphylococcus sp. Acb12, Staphylococcus sp. Acb13, Staphylococcus sp. Acb14, Bacillus, Achromobacter, Klebsiella, and Citrobacter spp., also synthesize ACC, offering the potential to alleviate the consequences of drought and salt stress [102,103]. Beneficial microorganisms that possess ACC deaminase activity, such as Achromobacter piechaudii ARV8 in pepper and tomato plants [104] and Pseudomonas fluorescens TDK1 in peanut seedlings [105], can assist in mitigating these unfavorable impacts. Furthermore, Achromobacter spp. and Pseudomonas spp. are recognized for their ability to stimulate plant growth and enhance soil quality [76].
According to Ouyang et al. (2017) [106], microbial biostimulants have the ability to biosynthesize indole acetic acid (IAA), which serves to stimulate the growth of plants and the proliferation of their root systems when they are exposed to drought stress. These authors suggested that this phenomenon is observed in various bacterial species, such as those falling within the genera Alcaligenes, Sinorhizobium, Serratia, Bacillus, and Arthrobacter. The bacterium Pseudomonas chlororaphis TSAU13, renowned for its ability to produce IAA, can bolster the resistance of cucumber and tomato plants to drought and salinity when introduced in salt-stressed conditions [107]. Similarly, the mycorrhizal fungus Funneliformis mosseae elevates IAA concentrations in the roots, fosters the development of root hairs, and stimulates the growth of orange plants experiencing drought stress [108]. Specific commercial products derived from Funneliformis mosseae can enhance plant nutrient and water uptake [65]. Microorganisms proficient in producing cytokinins and gibberellins can alleviate water stress by promoting stomatal opening and encouraging shoot growth in conditions of restricted water availability [109]. Plant growth-promoting rhizobacteria (PGPR) of the Acinetobacter, Pseudomonas, and Burkholderia genera can generate gibberellins, and can augment the growth of cucumber plants under salt and drought stress [110]. Despite their promising attributes, there are presently no commercially available products based on these species.
Following exposure to drought stress, the production of abscisic acid (ABA) increases, leading to stomatal closure. In soybean plants, the introduction of Pseudomonas putida H-2-3 decreases abscisic acid (ABA) levels, reducing the impact of drought stress [111]. P. putida is commonly used in conjunction with B. subtilis to enhance soil fertility, rather than specifically for alleviating drought stress. Similarly, the inoculation of lettuce with Glomus intraradices leads to a reduction in ABA concentration and increased resistance to salt [112]. ABA and water scarcity typically lead to increased ethylene production, which can inhibit plant development.
Water stress leads to the generation of reactive oxygen species (ROS) and subsequent oxidative damage to lipids, nucleic acids, and proteins. Many microorganisms can counteract the effects of elevated ROS by either producing antioxidant compounds or increasing the activity of antioxidant enzymes, such as peroxidases and catalase, in plants [113]. Pseudomonas species increase catalase activity in basil plants under water stress [113]. Similarly, Pseudomonas species, Bacillus lentus, and Azospirillum brasilense are microbial bioinoculants that have been employed individually or in consortia to mitigate drought stress in crops [65]. Ascorbate, peroxidase, and glutathione peroxidase in Pseudomonas species, Bacillus lentus, and Azospirillum brasilense have been utilized to alleviate drought stress [114].
The accumulation of osmocompatible solutes is a strategy employed by plants to combat water stress, allowing the buildup of inorganic and organic solutes in the vacuole and cytosol, respectively. This lowers the osmotic potential of the cell and maintains turgor pressure in water-stressed conditions [65]. Numerous bacteria can produce osmolytes [115], which often collaborate with osmolytes synthesized by the host plant, such as proline, to reduce osmotic potential and stabilize cell wall components [116]. The phosphate-solubilizing bacterium Bacillus polymyxa, when introduced to specific plants, produces proline, reducing the effects of water stress [117]. Betaine, produced by osmotolerant bacteria such as Streptomyces tendae F4 in the rhizosphere, can work in conjunction with the betaine produced by the host rice plant, increasing its tolerance to water stress [118]. Despite these promising results, Bacillus polymyxa is not yet available as a commercial product.
Some bacteria can interact with plants through volatile organic compounds (VOCs), which can induce stress adaptation responses related to mineral uptake, water conservation, and root growth [119]. Although the significance of hormone signaling pathways has been established [120], the underlying mechanisms of VOC-mediated interactions between plants and microbes under extreme conditions remain largely unexplored. Microbial VOCs can have positive effects on plants, such as the synthesis of osmoprotectants and regulation of stomatal closure [121] and increased plant fitness by 2,3-butanediol [65]. Compounds like 1-heptanol, 3-methyl-butanol, and 2-undecanone produced by Paraburkholderia phytofirmans enhance salinity tolerance [122], while butyrolactone and 1-butanol promote root growth and carbon exchange in the rhizosphere [123]. The production of VOCs by Bacillus thuringiensis AZP2 has been instrumental in mitigating drought stress in wheat [124]. The future development of VOCs for plant promotion will likely depend on the identification of stress-induced signaling pathways.
Although numerous microorganisms have demonstrated the ability to protect plants from water stress, only a limited number of commercial biofertilizers are available. Many of these are based on combinations of microorganisms, including Azospirillum brasilense, Bacillus altitudinis, Bacillus amyloliquefaciens, Bacillus licheniformis, Cellulomonas cellasea, Pseudomonas fluorescens, Pseudomonas putida, Pseudomonas stutzeri, Streptomyces albidoflavus, Glomus species, and Trichoderma species. These microorganisms exhibit not only the capacity to mitigate water stress but also the ability to enhance plant yields through the production of exopolysaccharides and the enrichment of nutrients and soil organic matter [76]. The development and production of more universally applicable commercial products, effective across various ecosystems, will greatly contribute to the mitigation of water stress in plants.
2.2.3. Flooding, Water Pooling, and Heavy Precipitation.
The root systems of trees help stabilize the soil, which in turn reduces the risk of flooding and erosion; trees also play a crucial role in soaking up excess water during the rainy season. However, when trees are removed from the environment, heavy rains, typical of tropical countries, can lead to severe consequences. Precipitation washes away essential topsoil and whatever nutrients remain in the soil. Flooding impacts approximately 13% of the Earth’s surface, and it is anticipated that the frequency and intensity of heavy rainfall events will increase globally in the future [65]. Heavy floods and rain result in water stagnation, causing root anoxia and hypoxia. When roots are subjected to flooding, they produce substantial amounts of the enzyme 1-aminocyclopropane-1-carboxylate (ACC) synthase, which is involved in ethylene production [65]. ACC oxidase, another enzyme required for the final step in ethylene biosynthesis, depends on oxygen for its catalytic activity. ACC is transported to the aerial parts of the plant through the xylem [125], where it is converted into ethylene, leading to symptoms such as wilting, chlorosis, leaf necrosis, fruit and flower loss, and reduced crop yields (Glick, 2014). Microbial biostimulants, through their ACC deaminase activity, can help to alleviate the stress caused by water stagnation by reducing endogenous ethylene levels [77,126]. Plants inoculated with ACC deaminase-producing strains of Pseudomonas spp. and Enterobacter spp. exhibit reduced anoxia stress and improved germination [65,127]. Pseudomonas and Enterobacter spp. can increase stress tolerance, although they do not specifically protect against waterlogging. Pseudomonas sp. [128] and Streptomyces sp. GMKU 336 [129] can increase chlorophyll content, plant growth, biomass, adventitious root formation, and leaf area, while reducing ethylene levels. S. lydicus WYEC 108 and Streptomyces K61 are beneficial bacteria often used against biotic stresses [130,131]. Future research should aim to develop more microbial biostimulants and comprehensively explore the potential of existing commercial products for mitigating water stress.
2.2.4. Imbalanced Nutrient Recycling
Forest degradation has profound effects on nutrient cycles. Along with its impact on ecosystem functions, it is exacerbated by changes in terrestrial environments. The primary anthropogenic influences on forests are logging and excessive pollution loads. Both processes result in detrimental effects on the soil, which subsequently hinder the natural regeneration of the forest [132]. The harm inflicted on forest soils jeopardizes the potential restoration of the original plant community because environmental fluctuations become deregulated. The soil’s ability to regulate environmental fluctuations relies on the continuous cycling of organic matter and sustained fertility. Disruption in forest nutrient cycles hampers effective land management, including sustainable landscape management [133].
An example is the Brazilian Amazon rainforest, which faces a significant threat from the expansion of cattle ranching activities [134,135]. During the process, farmers not only extract valuable timber, but also burn the remaining vegetation and replace it with fast-growing grasses [136]. This slash-and-burn technique alters the physical, chemical, and biological properties of the soil, facilitating the establishment of pastures in the Amazon region [137]. This practice is unsustainable, and pastures experience a rapid decline in productivity, ultimately becoming degraded and abandoned [138]. Under ideal conditions, ‘secondary forest’ is gradually formed [139], but studies suggest that deforestation is becoming increasingly common in the Brazilian Amazon region. Even though Carvalho et al. (2019) [140] state that these secondary forest areas are expanding, covering more than 100,000 km2, there is a need for more research on the re-establishment of secondary forests, especially with respect to soil microorganisms and nutrient cycling [141].
Phosphorus is considered an essential nutrient for plant growth and can limit primary productivity in various environments, including tropical and subtropical forests [142]. These regions typically have soils with high levels of iron and aluminum oxides due to intense weathering processes, which are exacerbated by the heavy rainfall and high temperatures in these areas. Fe and Al oxides can bind P, making it less available to plants [143,144]. Consequently, many plants rely on the recycling of P from litterfall and microbial P turnover [145].
Land-use changes negatively impact soil P dynamics in the Amazon region [144,147]. This is because the conversion of pristine forests to pasture using the slash-and-burn method alters P lability and increases P levels in more recalcitrant pools [144]. It can also release substantial amounts of nitrogen and P from forest biomass, which can be lost through leaching and runoff [148,149].
There is limited research and information available regarding the effects of secondary forest re-establishment on P dynamics. Existing studies suggest that secondary forest re-establishment has the potential to promote a gradual recovery of soil P [150,151,152]. According to these authors, the increase in plant diversity, the restoration of certain soil physicochemical properties (e.g., pH), and enzymatic activity (e.g., C, N, P, S enzymes) may favor specific bacterial and fungal groups such as P-mineralizing and P-solubilizing microorganisms. These groups play crucial roles in P transformation processes and availability. They are primarily involved in three key soil processes: (i) mineralization, (ii) solubilization, and (iii) immobilization [153] (Figure 4). These microorganisms produce enzymes that break down ester-phosphate bonds, releasing orthophosphate from recalcitrant organic P forms, as explained by Arenberg and Arai (2019) [154]. They possess genes encoding enzymes such as phytase (appA), alkaline phosphatase (phoD), acid phosphatase (olpA), phosphonatase (phnX), and C–P lyases (phn), which can mineralize organic-P compounds in soils [155,156].
P-solubilizing microorganisms can also produce and release organic acids like oxalic acid, malic acid, formic acid, citric acid, and gluconic acid. These acids can solubilize recalcitrant inorganic-P forms in soils [155,157]. They possess genes such as GCD (encoding glucose dehydrogenase) and the cofactor PQQ gene (encoding pyrroloquinoline quinone) that regulate the solubilization of unavailable inorganic P forms [158].
Microorganisms involved in P immobilization, on the other hand, can assimilate inorganic P into their biomass, competing with plants for available P [153]. They possess pst and pit transporter genes, which assist in the assimilation of inorganic P under P-limited and P-rich conditions.
The microbial groups involved in P transformations are ubiquitous in soils, but their population and activity are influenced by different environmental requirements [159]. They are also sensitive to disturbances, particularly changes in soil physicochemical properties [160,161]. This is a significant concern; previous studies have indicated that the conversion of pristine forests to pasture in the Amazon region alters soil physicochemical properties and the overall composition of the soil microbial community [162,163,164].
2.2.5. Soil Compaction by Cattle
Cattle trampling of agricultural soil can cause the collapse of larger soil pores, producing smaller pores. This leads to increased soil density and greater resistance to soil penetration, ultimately promoting more soil compaction. This, in turn, hinders the regrowth and renewal of pastures, resulting in reduced overall productivity [165]. Thus, soil compaction is closely linked to significant soil degradation processes, which manifest as reduced soil aeration, diminished water infiltration rates, and an increased risk of flooding and surface runoff [166,167,168,169]. These processes collectively have a negative impact on soil productive capacity [170]. Factors such as soil type, moisture content, and grazing management practices (like stocking rate, stocking density, and timing of activities) can exacerbate this soil compaction [165,171]. It is well-recognized that the risk is heightened when soil moisture content is increasing [172,173], as generally occurs in the wet summer seasons, which are thus the most vulnerable to soil compaction.
Currently, there are no reported studies that involve microorganisms and the impact of soil compaction resulting from cattle farming. However, some authors address the use of biochar to mitigate this problem. Gao and DeLuca (2022) [174] suggest that soil compaction and the consequent reduction in soil moisture and aeration may affect nutrient cycling, hindering natural repopulation with vegetation. There is thus a potential for enhancing nutrient cycling through the addition of specific microbial strains. The above authors noted an increase in soil organic matter and P content with biochar application. The use of microorganisms could further enhance this. Soil biofertilization could also stimulate primary production, leading to bioturbation by roots and detritivorous animals, thus promoting soil aeration. However, specific studies in tropical soils need to be carried out to substantiate this hypothesis. Porous materials can also play a crucial role in substrate enhancement [175,176,177]. Incorporating porous materials into the substrate augments soil porosity, elevates aeration rates [178], provides a conducive environment for microbial communities to thrive [179,180], and fosters the development of plant roots [181].
2.3. Treatment of Polluted Soils (Bioremediation)
The use of industrial effluents to irrigate agricultural crops is particularly prevalent in poor and less environmentally aware countries. This leads to the buildup of toxic substances in the soil with resulting effects on agricultural production and produce. Over the last decade, there has been a consistent growth in the number of publications discussing the application of microorganisms for the remediation of heavy metals and other contaminants in such polluted environments. While bacteria and fungi have been the predominant choices, yeasts and algae are also employed, the latter phylum mainly, but not exclusively, in aqueous environments [182]. Bioremediation usually relies on the metabolic activity of either a single organism or a consortium of microorganisms [183]. In this process, the transformation of contaminants may not provide significant benefits to the cell, when it is termed non-beneficial biotransformation [184,185]. Numerous studies have demonstrated that many organisms, both prokaryotes and eukaryotes, possess a natural capacity for biosorbing toxic heavy metal ions [186,187]; the biosorption sites may be located on or in the cells, or in the extracellular polymeric layer (EPS) of the microorganisms [188,189,190,191]. Alternative methods of bioremediating heavy metal contaminated soil include the use of microorganisms that change the toxicity of the metal by altering its oxidation state or by methylation [192,193,194,195] Examples of bacteria, fungi, and yeasts that have been studied and strategically employed in bioremediation applications for heavy metals in soil are shown in Table 1.
Rhizobacteria have been suggested to be particularly useful for the bioremediation of heavy metal-contaminated soils; the mechanisms of metal removal include chelation by siderophores, biotransformation (change to a less damaging oxidation number or methylation to reduce toxicity), biosorption and bioaccumulation in the cells, precipitation, ACC deaminase action, and biomineralization [223]. Biotechnological approaches by other microorganisms encompass bioleaching, bioextraction, and bioencapsulation [224], although not all of these are applicable to soil in situ.
When a contaminated site contains various pollutants, effective remediation requires a diverse array of microorganisms. Many microorganisms can degrade petroleum hydrocarbons, using them as a source of carbon and energy. The selection of these organisms for bioremediation varies based on the chemical nature of the pollutants and must be made carefully, as they can only thrive in the presence of a limited range of chemical contaminants. The efficiency of the degradation process is linked to the microbial capacity to introduce molecular oxygen into the hydrocarbon and generate intermediates that enter the cell’s energy-yielding metabolic pathways. Some bacteria have chemotactic responses, actively seeking out contaminants [225,226], while many produce powerful surface-active compounds that emulsify oil in water, aiding its uptake by the cells and hence removal from the soil [30,227]. Bacteria capable of degrading petroleum products include genera such as Pseudomonas, Aeromonas, Moraxella, Beijerinckia, Flavobacteria, Chromobacterium, Nocardia, Corynebacterium, Streptomyces, Bacillus, Arthrobacter, Aeromonas, as well as cyanobacteria [225,228], and certain yeasts [30]. One example is Pseudomonas putida MHF 7109 isolated from cow dung, which can be used for the biodegradation of selected petroleum hydrocarbon compounds, including benzene, toluene, and o-xylene (BTX) [186].
Genetically Modified Microorganisms for Bioremediation
Genetic engineering is a fundamental method to modify the metabolic pathways of microorganisms and improve their bioremediating activity, increasing efficiency and reducing the time required to produce the required effect. Genetic engineering can be applied to modify microorganisms to exhibit features like accelerated growth, tolerance to extreme environmental conditions and pH fluctuations, and cost-effective cultivation. Omics technologies, genomics, transcriptomics, proteomics and metabolomics, are being employed to increase our understanding of relevant microbial metabolisms, such as responses to heavy metals and high salt concentrations, and to indicate the appropriate engineering of potential microbial inoculants, with increased resistance and degrading activities for soil pollutants like heavy metals and hydrocarbons [229,230]. Perceived hazards in the release of genetically-modified organisms (GMOs), however, have led to problems in legislation for their placement on the market [231]. Similarly, care must be taken when releasing genetically-modified microorganisms (GMMs), defined as “microorganisms in which the genetic material has been altered in a way that does not occur naturally by mating and/or natural recombination” [232]. For this reason, genetic manipulations included in GMMs may include inbuilt senescence genes, so that unexpected negative interactions with native organisms will be limited [233]. Nevertheless, research on the use of GMMs in soil bioremediation in situ is now rather restricted.
3. Environmental Parameters Influencing Microbial Fertilization and Bioremediation in Soils
Bioremediation involves harnessing microorganisms, including bacteria, algae, and fungi, as well as plants, to expedite the breakdown, alteration, elimination, immobilization, or detoxification of diverse physical and chemical contaminants in the environment. The microorganisms can employ metabolic pathways to accelerate biochemical reactions that break down pollutants [239,240]. For bioremediation to be effective, microorganisms must be supplied with the necessary energy and nutrients to support their growth. Various factors, including physical, chemical, biological, soil type, carbon and nitrogen sources and the type of microorganisms (whether single or a consortium), influence the efficacy of bioremediation [241]. Microbial consortia often exhibit multifunctionality and resistance, and thus are more efficient than a single microorganism [242]. These may be natural microbial communities. For instance, carbon, one of the most crucial nutrients, was found to enhance in situ bioremediation by increasing natural consortium metabolic activity and expediting the breakdown of existing pollutants [243]. Soil type also impacts bioremediation, with sandy soils generally allowing higher levels of pollutant bioremediation than clay [244]. The factors influencing bioremediation and the methods employed for soil remediation, both in situ and ex situ, are discussed by Sales da Silva et al. (2020) [245]. These factors include the microbial population, contaminant accessibility, and the physicochemical characteristics of the environment (Figure 5). The same considerations apply to the use of biofertilizers. Extra activities involved may be excretion of phytohormones, suppression of phytopathogens and protection of plants from various types of stress [246,247].
3.1. Temperature
An important physical factor that significantly influences the survival of microorganisms as well as the degradation of pollutants is temperature. For example, in cold regions such as the Arctic, the natural degradation of oil is slow, creating a greater reliance on microbes to remediate oil spills. In these conditions, sub-zero temperatures can freeze the transport channels of microorganisms, hindering their ability to carry out metabolic processes. Temperature also has a direct impact on the turnover of enzymes involved in degradation, which may vary depending on the pollutant. The physiological properties of microorganisms are influenced by temperature, which can either accelerate or decelerate the bioremediation process. Higher temperatures generally promote increased microbial activity up to a certain limit, and this activity typically decreases if temperatures rise or fall abruptly, eventually ceasing altogether at temperature extremes [248,249]. However, extremophilic microorganisms, thermophiles and psychrophiles, are active in extreme environments such as the tropics and polar regions and can be used in bioremediation and biofertilization in such areas, as well as in other extreme environments [250,251,252,253,254].
3.2. Salinity
One of the factors that can limit bioremediation and that could be tackled by the use of extremophiles is salinity. Salt levels in soils used for agriculture are rising because of natural and anthropic activities [255]. Unsustainable soil management, such as irrigation with brackish water or overapplication of fertilizers, are responsible for some of this [256]. Hypersaline soils not only reduce plant growth, but also limit the possibilities of bioremediation. Non-saline soils have been shown to have approximately four times higher total petroleum hydrocarbon biodegradation compared to saline (1% NaCl) soils [257]. Salinity reduces the bioremediation of motor oil [258] and petroleum hydrocarbons [259,260] in soil. Nguyen et al. (2021) [261] suggest that salinity exposure may influence Cr(VI) bioremediation; Cr-resistant microorganisms may be able to deal with this problem [195]. There are several examples of the use, or potential use, of halophiles in bioremediation of polluted environments [195]. Although still undergoing considerable research effort, the use of halophilic microorganisms for bioremediation in, or of, saline environments appears to be a real possibility [267,268,269]
3.3. pH
The acidity, alkalinity, and basicity of a compound influence microbial metabolism and the bioremediation process. Soil pH, similarly, has an impact on nutrient availability as well as enzyme activity. Soil pH can be used to predict microbial growth since even slight pH variations have a notable effect on metabolic processes [270]. In the case of petroleum hydrocarbon biodegradation, a pH close to neutrality is optimal [271], but this will clearly vary depending on the pollutant and the microorganisms. For example, modeling the removal of the antibiotic, azithromycin, from contaminated soil by various fungi showed pH to be the most important physicochemical parameter and the optimum to be 5.5 [272], and acidophilic methanotrophs, growing optimally at pH 5 or below, have been suggested for the bioremediation of chlorinated volatile organic compounds in groundwater aquifers [273]. It should be noted, however, that biofertilizers themselves may be able to alter soil pH advantageously [274].
3.4. Microbiological Diversity
Quite apart from the microorganisms added to attain the desired effect, the resident microbiota will have a large part to play in the final outcome of a bioremediation or biofertilization treatment. There will be competition between the autochthonous and the added species, as well as between the indigenous microorganisms themselves [275,276]. For example, when petroleum degrading bacteria (PDB), Acinetobacter radioresistens KA5 and Enterobacter hormaechei KA6, were inoculated into soil containing indigenous microorganisms, the level of petroleum breakdown was considerably lower than that produced by the PDB alone [277]. When two different types of pollutants require removal, or when biofertilization is the aim, the choice of augmenting microorganisms becomes much more complex; antagonistic or highly competitive interactions must be closely monitored. The negative effects of one pollutant on the degradation of another are discussed and exemplified by Olaniran et al. (2013) [278], but double bioremediation is achievable, although perhaps not at maximum efficiency. In the case of metal-contaminated soils, one approach is to use metal-resistant microorganisms. Roane et al. (2001) [279] isolated 4 bacterial strains of different genera that reduced soluble Cd levels in the contaminated soil and also supported the degradation of 500-μg ml−1 2,4-D by the cadmium-sensitive 2,4-D degrader Ralstonia eutropha JMP134, while Wani et al. (2023) [280] reported that Brevibcillus parabrevis OZF5 removed both Cr and hydrocarbons from contaminated soil, enhancing growth of beans. Microbial surfactants, such as surfactin, rhamnolipid, and sophorolipids, may also be used to augment bioremediation of various pollutants, including DDT, atrazine, hexachlorocyclohexane and cyprodinil [281], as well as the obvious hydrocarbons [282].
Finally, it must be noted that any change in the structure of the microbial community can result in shifts not only in temperature adaptation at the community level but also in growth strategies: copiotrophs can promptly respond to favorable environmental conditions and proliferate rapidly when labile carbon sources are available. In contrast, oligotrophs display gradual, sustained growth under conditions of low carbon and nutrient availability, finally yielding a higher biomass per unit substrate [283,284]. As eloquently articulated by Neidhardt (1999) [285], when the primary objective is growth, delving exclusively into individual enzymes and genes, or even isolated pathways and mechanisms, is insufficient. There comes a point where one must return to the holistic perspective of the entire cell and consider the coordinated orchestration of various processes. When several microbial species are mixed, the community must work together and support each other to grow and become active in the bioremediation/biofertilization process in a symbiosis (Moqsud et al., 2015) [286].
3.5. Oxygen Levels
The biodegradation process is directly related to the available oxygen content in the environment. Most bioremediation systems run under aerobic conditions [287]. Often, however, in aquatic sediments, sludges or compacted soils, for example, there is limited oxygen available. With the impossibility to aerate the environment, the use of anaerobic decomposers or facultative organisms can be more effective [288]. Ji et al. (2014) [289] investigated how the activities of microorganisms were influenced by the presence of dissolved oxygen (DO) in polluted river water. Their study revealed that the removal efficiency of different substances, such as chemical oxygen demand, total phosphorus, and ammonia nitrogen, increased with higher levels of DO, showing improvements of 4.59%, 18.18%, and 34.34%, respectively, suggesting that this bioremediation works even under aerobic conditions.
Petroleum-contaminated sludge is a particular problem in coastal wetlands, where costly physicochemical treatment is currently the norm [290]. Anaerobic bioremediation, utilizing sulfate-reducing bacteria, denitrifying bacteria, iron and manganese reducing bacteria, and, particularly, methanogenic bacteria is more economical, although currently less efficient than aerobic bioremediation [290]. Intermittent oxygen supplies may also be useful in some cases, For example, Chen et al. (2023) [291] found that intermittent microoxygenation facilitated codegradation of trichloroethane and toluene; the main bacterial genera responsible for the dechlorination were Dehalogenimonas, Dehalococcoides, and, mainly, Dehalogenimonas. One possibility could be the use of cyanobacteria, which can grow anaerobically, under the correct conditions and produce oxygen by photosynthesis. Their potential as biofertilizers in green technology is discussed by Bhuyan et al. (2023) [292].
3.6. Soil Moisture
The soil’s moisture content is also an important factor in accelerating biogeochemical processes. Soil that is excessively waterlogged is not suitable for microbiological treatment [293]. The moisture level in the soil impacts the movement of water and soluble nutrients in and out of microorganisms and excessive moisture diminishes oxygen levels during the aerobic process [294] and hence bioremediation [295]. With this effect of soil moisture in mind, Naorem (2022) [296] considered the potential effects of climate change on biodeterioration efficiency. Bahmani et al. (2018) [297], using laboratory experiments and various moisture levels in hydrocarbon-contaminated soil, developed a mathematical model for the process. They defined a first-order rate equation for removal of total petroleum hydrocarbons (TPH); this equation held good between a specific moisture value at which the constant is zero and that at which the constant reaches its maximum.
ν dθ/dt=qin-qout (1) qin=0 (2) qout=APET (θ-θr /θsat-θr)
Where, ν=volume of beaker, θ=water content, q=volumetric flow rate of water, A=surface of beaker, PET=potential evaporation (constant during their experiment at 0.00495 m day-1), θr=residual water content, θsat=saturation water content.
In their study, the best TPH removal from a sandy clay soil was achieved by adjusting the water to 60% capacity and adding water every 2 days. This moisture level is higher than that quoted by other workers; for example, Kumari et al. (2023) [298] suggested 35%. However, Zeng et al. (2023) [299] and Norozpour et al. (2023) [300] suggest 60% and 70-80% respectively. These authors were all working with PAH-contamination.
3.7. Nutrient Availability
Nutrients play a crucial role in influencing the growth of microorganisms and the rate of biodegradation. Enhancing the soil C:N:P ratio can significantly enhance biodegradation efficiency, particularly when vital nutrients like nitrogen and phosphorus are provided. When these nutrients are present in low concentrations, the degradation of hydrocarbons is constrained. Addition of extraneous nutrients, such as glucose, can augment pollutant removal by stimulating microbial activity [301]. Introduction of nutrients is particularly important for bioremediation activities in cold environments; it can boost the metabolic activity and diversity of microorganisms and, consequently, the rate of biodegradation [80]. Microbes that consume oil rely on these essential nutrients to thrive, and they are typically present in limited quantities in natural settings [302,303].
Cavazolli et al. (2023) [304] used DNA sequencing techniques to investigate the bacterial populations of soil co-contaminated with heavy metals and hydrocarbons. Bioremediation was carried out using meat and bone meal (MBM) as nutrient addition and cyclodextrin (Cdx), as a surfactant to release previously unavailable nutrients. After 12 weeks, only Cdx had increased bacterial diversity more than the control soil; MBM accelerated the change in populations, but final diversity after 12 weeks was not altered. Oil-degrading groups of bacteria increased in prevalence during the procedure, but hydrocarbon and heavy metal levels were not determined. The effects of addition of any specific nutrient, or nutrients, to contaminated soil are difficult, perhaps impossible, to predict and should be carefully monitored.
3.8. Pollutant Complexity
The release of diverse chemical pollutants into the environment by industries can disrupt the delicate equilibrium of ecosystems. These anthropogenic pollutants, in particular, tend to exhibit greater complexity and heightened resistance to degradation, necessitating the use of specific bioremediating strains capable of withstanding the toxicity of substances such as metals and hydrocarbons [305]. It has been suggested that GMOs and mixed strains of microorganisms may be the preferred route for future treatment of complex pollutants [306,307]. While the potential of GMOs in bioremediation is substantial, its practical application is circumscribed in various ways [308] and the discussion above (Section 2.3.1) points out the problems of this approach.
Li et al. (2020) [309] suggest that fungi may be more appropriate for the degradation of complex pollutants. Fungi may oxidize pollutants extracellularly using laccases, manganese peroxidases and lignin peroxidases, or intracellularly using cytochrome P450, reductive dehalogenases, and nitroreductases [309]. Recently, innovative remediation techniques have been proposed, including the utilization of extracellular and/or cell-free enzymes, together with metabolomics and proteomics to study the response of the microorganisms to complex and mixed pollutants [310,311]. Using a single microbial species in a bioremediation treatment for complex molecules is unlikely to obtain success; microbial consortia are now being intensely studied to find species that can survive and work together. The engineering of microbial consortia is discussed by Li et al. (2021) [312]; they consider 2 main approaches, “bottom-up” and “top-down”. Interactions between microorganisms include antagonism and symbiosis; the bottom-up approach involves reconfiguring negative interactions to ensure co-survival and continued activity. The top-down approach involves modifying the environment to compel the organisms to perform the desired activities; it is not an easy option because of the complexity of the microbial interactions. The bottom-up approach requires a detailed knowledge of the metabolism and interactions of the consortium members. The new omics techniques have allowed this to become a less onerous and more achievable goal and will assist the development of consortia for the bioremediation of complex pollutants such as PAHs.
Cell-free systems have also been investigated for bioremediation of pollutants such as organophosphate pesticides [313], aromatic polymers like chlorophenols [314] and nitrile and amide-based xenobiotics [315]. Immobilization of the enzymes on a suitable, and economic, material can be advantageous to ensure their continued activity, but the support must be carefully chosen to prevent any reduction in efficacy [316]. Screening of microorganisms for enzyme activities, in addition to being used for the identification of potential enzymes to be extracted or engineered, could also be used in place of laboratory biodegradation tests for detection of efficient functional isolates for bioremediation procedures [317].
4. Risks to Ecosystem Health: Impact on the Local Biota
In theory, the biofertilization and bioremediation approaches should lead to minimal or well-controlled consequences concerning factors such as dispersal, persistence, alterations to microbial function and biogeochemical cycles, and shifts in the ecosystem communities [318]. A substantial area of concern lies in the potential impact of introduced microorganisms on the existing microbiome, which may result from direct ecological interactions, whether they are cooperative or antagonistic in nature. Invasive, alien species are bad for ecosystems, since they may reduce biodiversity and disrupt food chains, including our own. Species from other regions that establish self-sustaining populations in the invaded area are referred to as naturalized species. Among naturalized species, those that quickly spread beyond the site of their initial introduction in the invaded region are classified as invasive, as described by Richardson et al. (2000) [319], Pyšek et al. (2004) [320], and Blackburn et al. (2011)[321]. For instance, the chestnut blight fungus entered North America surreptitiously through contaminated nursery stock [322], leading to the demise of four billion trees over a span of 40 years. Especially in tropical regions, like South Asia and South America, plant invasions are one of several factors that threaten the local biodiversity. However, the economic and social challenges resulting from plant invasions are on the rise across all continents. The expenses directly associated with the harm caused by invasive species are thirteen times greater than the costs incurred for their management [323].
Finally, biological invasions rank as the fifth most significant driver of global environmental change, according to IPBES 2019 [324]. Approximately 14,000 taxa, constituting about 4% of the world’s flora, are recognized to have naturalized from the global plant species pool [325,326]. Among them, around 2,500 species are identified as invasive (Pagad et al. 2018), with the Asteraceae family contributing the highest number of naturalized taxa [327]. The escalating international trade has led to a global increase in the number of invasive species [328].
The phenomenon of horizontal gene transfer has also been identified as a relevant consideration [329,330,331]. Horizontal gene transfer involves the transfer of genetic information between organisms, facilitating the dissemination of antibiotic resistance genes among bacteria, excluding those passed from parent to offspring. This process contributes to the evolution of pathogens [332]. For instance, the simple use of manure as an organic fertilizer in agriculture introduces fecal bacteria, along with their plasmids and antimicrobial resistance genes, into the soil. The exchange of genetic material, which includes plasmids [333] and even entire genomic segments, is particularly prevalent in the rhizosphere—an area rich in microbial activity. Such exchanges have been observed even between species that are quite distantly related. There have, for example, been documented cases of gene transfer between Pseudomonas putida and E. coli facilitated by the TOL plasmid (pWW0) [334]. Macedo et al. (2022) [333], through a microcosm study involving E. coli, recorded the successful transference of a broad host range plasmid to soil and manure bacteria by conjugation. Similar instances of plasmid transfer have been witnessed within various strains of Azotobacter vinelandii. Martínez-Carranza et al. (2019) [335] suggest that A. vinelandii may have a polyphyletic origin, with a genomic resemblance to the pathogenic bacterium P. aeruginosa. This hypothesis opens up the possibility of horizontal transfer from P. aeruginosa to A. vinelandii of genetic information encoding pathogenicity [335].
It has been suggested that microorganisms may influence ecosystem biodiversity or induce infections in humans, animals, or plants, thereby creating an adverse environmental impact. As a response to this concern, the European authorities have tried to protect humans, plants, animals and the environment by developing European Regulation (EU) 2019/1009 [336] on biofertilizers.
Current understanding suggests that plants are not isolated entities; instead, they exist in association with a diverse range of microbes [337]. Introduced microorganisms can indirectly alter resident microbial communities by influencing plant physiology and morphology, via the acquisition of nutrients, reduction of plant stress and suppression of plant diseases. For instance, some plant growth-promoting microbes are known to modify root architecture and exudation [338], which can impact rhizosphere communities [339,340]. In mixed plant communities, indirect effects on the resident microbiome could also occur if the introduced microorganism induces changes in plant diversity and composition, as has been seen with some AMF inoculants [341,342].
Inoculation with rhizobia, commonly used in biofertilizers, has shown significant effects on soil and plant-associated microbial communities in soybean [343], alfalfa [344] and common bean crops [345]. These effects can extend beyond the rhizosphere; field inoculation of Phaseolus vulgaris with two native rhizobia strains (separately or combined) altered the structure and increased the phylotype richness of bacterial communities in the bulk soil [346]. Changes in microbial structure can result from both positive and negative interactions of rhizobia with the rhizosphere microbiome. The close relationship between root endophytes and root tissues facilitates the exchange of nutrients between plants and microbes [347]. For instance, Azospirillum, a bacterial genus known for its free-living organisms with nitrogen-fixing capabilities, among other plant growth-promoting traits, has shown variable effects on the resident microbiome [346]. Inoculation of maize with A. lipoferum induced a shift in the composition of rhizosphere bacterial communities lasting for at least one month [348]. However, variable results were observed when inoculating the same or other crops with A. brasilense [349,350,351], even though this bacterium can induce physiological and morphological changes in the root system.
According to Trabelsi and Mhamdi (2013) [346], the effects of Azospirillum inoculation on rhizosphere microorganisms are likely driven by nitrogen dynamics, although a wide array of factors are involved. While rhizobia and Azospirillum have been more widely studied regarding their effects on the resident microbiome, research on other taxa, such as Pseudomonas sp., remains limited, even though they could potentially modify both bacterial and fungal communities [352,353].
Arbuscular mycorrhizal fungi (AMF) biofertilizers are the most widely used and readily available, despite insufficient evaluation of ecological risks associated with their field application [341,354]. The introduction of non-native AMF can impact indigenous AMF communities in various ways, such as displacing them and reducing their diversity [355], or altering their composition [356], although these effects on the native AMF community may not always be evident [357]. In a study conducted by Jin et al. (2013b) [358], a diverse AMF inoculum had a less pronounced positive impact on plant fitness but had a milder influence on the composition of subsequent AMF communities. In addition to affecting native mycorrhizal fungi, introduced AMF can disrupt other soil microbial communities, especially those associated with their extraradical mycelium, known as the mycorrhizosphere [346].
The influence of AMF on soil microbial communities appears to be variable and is influenced by multiple factors [359,360,361]. For instance, among the principal factors influencing microbial activity lies the microbial capability to reduce organic materials, utilizing them as sources of energy. The average oxidation state of carbon in a contaminant stands as an effective energy source for aerobic heterotrophic organisms. A heightened oxidation state, consequently, correlates with a diminished energy yield, thereby offering a reduced energy output for microbial degradation. The outcome of any biodegradation undertaking is contingent upon various factors, encompassing microbial dynamics (population diversity, enzyme activities, and biomass concentration), substrate attributes (physical and chemical properties, molecular concentration, and structure), and an assortment of environmental determinants (pH, moisture content, temperature, electrical conductivity, availability of electron acceptors and carbon, and energy sources). These parameters exert influence on the adaptation period of microorganisms to the given substrate. The concentration and molecular structure of contaminants wield substantial influence over the feasibility of bioremediation practices and the nature of microbial metamorphosis, determining whether the compound will function as a primary, secondary, or co-metabolic substrate [362].
Notably, bacteria residing in the mycorrhizosphere have demonstrated plant growth-promoting activity and are believed to work in synergy with AMF [363]. The impact of non-AMF fungal inoculants has received less attention, but there is evidence that endophytic fungi [364,365] and Trichoderma spp. [293] can induce changes in local microbial communities.
The impact of introduced microorganisms on the existing microbial communities hinges, to some extent, on their abundance, survival, and persistence within the ecosystem [366]. This intricate relationship implies that the very traits that contribute to successful plant growth promotion also elevate the risk of ecological invasion. Surprisingly, there is a dearth of comprehensive studies that delve into the consequences of large-scale, repeated inoculations, as well as the enduring effects, both in the long term and across subsequent crops [330,346]. Hart et al. (2018) [341] emphasize the significance of long-term repercussions, particularly concerning AMF inoculants. The assessment of the establishment and endurance of AMF inoculants is compounded by their complex genetic makeup. These concerns become all the more alarming when considering that the effects of inoculation may linger even as the population of introduced microorganisms diminishes [346]. One of the underlying mechanisms responsible for these enduring “legacy” effects is the phenomenon of plant-soil feedback. This is more likely to occur when the introduced microorganism has a symbiotic relationship with or a strong affinity for a specific plant, potentially affecting non-target plants within the agroecosystem [342,366].
The diverse outcomes witnessed when introducing microorganisms through inoculation into the resident microbiome imply a multifaceted interplay of influencing factors, notably including the host plant [359]. Correa et al. (2007) [351] found that the plant’s genetic makeup exerts control over the genetic and physiological responses of phyllosphere and rhizoplane bacterial communities to A. brasilense seed inoculation. Rhizosphere bacterial communities were found to respond differently to A. brasilense at distinct growth stages of the crop, displaying more pronounced effects during the jointing stage than the grain-filling stage [367].
The resident microbiome susceptibility and buffering capacity can vary considerably depending on its diversity [346,368] and the presence of specific antagonistic or synergistic taxa [369,370]. Environmental conditions, such as soil moisture [361,371] and light intensity [359], may emerge as pivotal factors in regulating the influence of introduced microorganisms.
In general, it appears that the majority of studies investigating the effects of biofertilizers tend to place emphasis on structural attributes rather than functional aspects [346]. Recognizing the functional implications of these alterations is of paramount importance, as they exert a direct influence on ecosystem operation and well-being. To tackle the complexities of these ongoing dynamics and foresee microbial responses in natural conditions, multidimensional omics and advanced data analysis serve as indispensable tools [366,372]. The question remains as to the lasting environmental impact of introduced microorganisms, encompassing their potential for long-term and legacy effects. Additionally, there is a need for more in-depth exploration regarding genetically modified microorganisms, encompassing considerations of metabolic load [329] and the associated risks of horizontal gene transfer and dispersion [273,274]. Diverse inoculants may emerge as a more environmentally benign alternative, as implied by studies on AMF or Bacillus spp [76].
Bioremediation as a method presents its own set of constraints. The microbial breakdown of specific chemicals may lead to the creation of intermediary substances that not only prove more toxic but also exhibit greater mobility than the original compound. For example, the reductive dehalogenation of TCE can result in the accumulation of vinyl chloride, a highly toxic and carcinogenic byproduct. Hence, bioremediation is a technology demanding extensive research and a comprehensive understanding of microbial processes prior to its implementation. Failure to do so could yield even more severe repercussions for the ecosystem than the initial contaminant itself [375].
5. Conclusions and Perspectives
Currently, developing countries are going through particular historical phases compared to those experienced by developed nations. This is due to both globalization, which defines their role in the global context, and their geographical characteristics, which directly influence their ecological, cultural, and productive dynamics. Consequently, they are currently grappling with issues stemming from their rapid industrialization, lacking the necessary infrastructure and cultural maturity to address the adverse effects of development. On the other hand, their natural attributes bestow upon them advantages that have not yet been fully harnessed. The warm climate, coupled with increased sunlight and more intense weathering, results in higher primary productivity and, thus, greater biodiversity, which could serve as a key to their sustainable development. Throughout this review, we briefly discussed characteristics related to the communities of microorganisms in nature, as well as the effects of microbial coexistence. It becomes evident that there is a direct relationship between environmental health and the microbial balance. The targeted use of these microorganisms can lead to beneficial effects on ecosystems. Introducing microbial strains into low-quality soils, which are typically unfavorable for plant growth, has a dual benefit. It not only enhances soil fertility, boosts nutrient cycling, and mitigates soil salinity, but also enhances the profitability of cash crops and fosters the sustainability of farming practices. Thus, the key to sustainable development lies in the proper management of these biological tools. Understanding the product of the interaction between microorganisms and environmental factors is essential for the development of cost-effective sustainable environmental management techniques.
Bioremediation is a burgeoning technology that can be employed in conjunction with other physical and chemical treatments to comprehensively address a wide range of environmental pollutants, in addition to helping to solve a range of other environmental issues, further in-depth studies on its other uses should be undertaken to fully explore the potential of microbial biotechnology, which appears to offer a sustainable approach to environmental pollution management. It is imperative to establish a synergistic relationship between the environmental influences on the fate and behavior of environmental contaminants and the selection and efficacy of the most suitable bioremediation technique, as well as other pertinent methods, to ensure the efficient and successful execution and monitoring of bioremediation processes.
The vast majority of articles published on the effects of using microorganisms in bioremediation, particularly studies on the use of microbial strains in the remediation of natural environments, reveal a range of methodological limitations, like the use of known and secure microbiota species, the use of the host species with a short-life cycles, the control and manipulation of mineral nutrients level in soil and the appropriate combination of micobiota species and host plant. It is essential to bear in mind that the majority of this research has been conducted in temperate climates. This article proposes that the environmental conditions of tropical regions could offer an advantage for the utilization of this more ecologically sustainable technology, which could usefully be employed to tackle problems in developing countries.
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare that they have no competing interests.
Author Contributions
Christine C. Gaylarde and Estefan M. da Fonseca: conception; development of the theory; discussion of results and contribution to final manuscript.
Acknowledgments
This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
- Benzerrouk, Z.; Abid, M.; Sekrafi, H. Pollution haven or halo effect? A comparative analysis of developing and developed countries. Energy Rep. 2021, 7, 4862–4871. [Google Scholar] [CrossRef]
- Ameen, R.F.M.; Mourshed, M. Urban environmental challenges in developing countries—A stakeholder perspective. Habitat Int. 2017, 64, 1–10. [Google Scholar] [CrossRef]
- Akhtar, N.; Ishak, M.I.S.; Bhawani, S.A.; Umar, K. Various Natural and Anthropogenic Factors Responsible for Water Quality Degradation: A Review. Water 2021, 13, 2660. [Google Scholar] [CrossRef]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and Eutrophication: Where Do We Go from Here? Sustainability 2014, 6, 5853–5875. [Google Scholar] [CrossRef]
- Parkinson, D.; Coleman, D.C. Microbial communities, activity and biomass. Agriculture. Ecosystems & Environment 1991, 24, 3–33. [Google Scholar]
- Macrae, A., Coelho. Tropical Soil Microbial Communities. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin, Heidelberg, 2013. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology and Biochemistry; Elsevier: Amsterdam, NX, Netherlands, 2015. [Google Scholar]
- Mukherjee, T.; Kao, N. PLA Based Biopolymer Reinforced with Natural Fibre: A Review. J. Polym. Environ. 2011, 19, 714–725. [Google Scholar] [CrossRef]
- Sharma, A.K. Biofertilizer – A Key Player in Enhancing Soil Fertility and Agricultural Sustainability. Int. J. Agric. Environ. Biotechnol. 2023, 16. [Google Scholar] [CrossRef]
- Cui, K.; Xu, L.; Tao, T.; Huang, L.; Li, J.; Hong, J.; Li, H.; Chi, Y. Mechanical behavior of multiscale hybrid fiber reinforced recycled aggregate concrete subject to uniaxial compression. Journal of Building Engineering 2023, 71, 106504. [Google Scholar] [CrossRef]
- Kheirfam, H.; Sadeghi, S.H.; Homaee, M.; Darki, B.Z. Quality improvement of an erosion-prone soil through microbial enrichment. Soil Tillage Res. 2017, 165, 230–238. [Google Scholar] [CrossRef]
- Liu, Q.; Pang, Z.; Yang, Z.; Nyumah, F.; Hu, C.; Lin, W.; Yuan, Z. Bio-fertilizer Affects Structural Dynamics, Function, and Network Patterns of the Sugarcane Rhizospheric Microbiota. Microb. Ecol. 2021, 84, 1195–1211. [Google Scholar] [CrossRef]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil Microbial Resources for Improving Fertilizers Efficiency in an Integrated Plant Nutrient Management System. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, D.; Gupta, A.; Pandey, K.D.; Singh, P.; Kumar, A. Plant Growth Promoting Rhizobacteria. In PGPR Amelioration in Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2019; pp. 41–66. [Google Scholar] [CrossRef]
- El-Ramady, H.; El-Ghamry, A.; Mosa, A.A.; Alshaal, T. Nanofertilizers vs. Biofertilizers: New Insights. Environ. Biodivers. Soil Secur. 2018, 2, 40–50. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Riaz, U.; Mehdi, S.M.; Iqbal, S.; Khalid, H.I.; Qadir, A.A.; Anum, W.; et al. Bio-fertilizers: eco-friendly approach for plant and soil environment,” in Bioremediation and Biotechnology: Sustainable Approaches to Pollution Degradation, eds K. R. Hakeem, R. A. Bhat, and H. Qadri (Cham: Springer) 2020.
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-Producing Rhizobacteria as a Promising Tool for Empowering Plants to Cope with Iron Limitation in Saline Soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Liu, N.; Shao, C.; Sun, H.; Liu, Z.; Guan, Y.; Wu, L.; Zhang, L.; Pan, X.; Zhang, Z.; Zhang, Y.; et al. Arbuscular mycorrhizal fungi biofertilizer improves American ginseng (Panax quinquefolius L.) growth under the continuous cropping regime. Geoderma 2020, 363, 114155. [Google Scholar] [CrossRef]
- Shirmohammadi, E.; Alikhani, H.A.; Pourbabaei, A.A.; Etesami, H. Improved Phosphorus (P) Uptake and Yield of Rainfed Wheat Fed with P Fertilizer by Drought-Tolerant Phosphate-Solubilizing Fluorescent Pseudomonads Strains: a Field Study in Drylands. J. Soil Sci. Plant Nutr. 2020, 20, 2195–2211. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Abrantes, I.; Caracciolo, A.B.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proença, D.N. Belowground Microbiota and the Health of Tree Crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef]
- Tomar, O.; Akarca, G.; Çağlar, A.; Beykaya, M.; Gök, V. The effects of kefir grain and starter culture on kefir produced from cow and buffalo milk during storage periods. Food Sci. Technol. 2020, 40, 238–244. [Google Scholar] [CrossRef]
- Rizvi, A.; Zaidi, A.; Ameen, F.; Ahmed, B.; AlKahtani, M.D.F.; Khan, M.S. Heavy metal induced stress on wheat: phytotoxicity and microbiological management. RSC Adv. 2020, 10, 38379–38403. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Song, S.; Rao, P.; Wang, R.; Liu, S. Preparation and properties of soil conditioner microspheres based on self-assembled potassium alginate and chitosan. Int. J. Biol. Macromol. 2019, 147, 877–889. [Google Scholar] [CrossRef]
- Zahra, N.; Raza, Z.A.; Mahmood, S. Effect of Salinity Stress on Various Growth and Physiological Attributes of Two Contrasting Maize Genotypes. Braz. Arch. Biol. Technol. 2020, 63. [Google Scholar] [CrossRef]
- Narayanan, S. Effects of high temperature stress and traits associated with tolerance in wheat. Open Access J. Sci. 2018, 2, 1. [Google Scholar] [CrossRef]
- Bankole, M.T.; Abdulkareem, A.S.; Mohammed, I.A.; Ochigbo, S.S.; Tijani, J.O.; Abubakre, O.K.; Roos, W.D. Selected Heavy Metals Removal From Electroplating Wastewater by Purified and Polyhydroxylbutyrate Functionalized Carbon Nanotubes Adsorbents. Sci. Rep. 2019, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Adamiec, E.; Jarosz-Krzemińska, E.; Wieszała, R. Heavy metals from non-exhaust vehicle emissions in urban and motorway road dusts. Environ. Monit. Assess. 2016, 188, 1–11. [Google Scholar] [CrossRef]
- Luo, X.; Wu, C.; Lin, Y.; Li, W.; Deng, M.; Tan, J.; Xue, S. Soil heavy metal pollution from Pb/Zn smelting regions in China and the remediation potential of biomineralization. J. Environ. Sci. 2022, 125, 662–677. [Google Scholar] [CrossRef]
- Kumar, S.; Diksha; Sindhu, S.S.; Kumar, R. Biofertilizers: An ecofriendly technology for nutrient recycling and environmental sustainability. Curr. Res. Microb. Sci. 2021, 3, 100094. [Google Scholar] [CrossRef]
- Kumar, R.; Chaudhry, V.; Prakash, O. Editorial: Multi-omics profiling of unique niches to reveal the microbial and metabolite composition. Front. Microbiol. 2022, 13, 997191. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant Growth-Promoting Rhizobacteria Allow Reduced Application Rates of Chemical Fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef]
- Bertola, M.; Mattarozzi, M.; Sanangelantoni, A.M.; Careri, M.; Visioli, G. PGPB Colonizing Three-Year Biochar-Amended Soil: Towards Biochar-Mediated Biofertilization. J. Soil Sci. Plant Nutr. 2019, 19, 841–850. [Google Scholar] [CrossRef]
- Ullah, N.; Ditta, A.; Khalid, A.; Mehmood, S.; Rizwan, M.S.; Ashraf, M.; Mubeen, F.; Imtiaz, M.; Iqbal, M.M. Integrated Effect of Algal Biochar and Plant Growth Promoting Rhizobacteria on Physiology and Growth of Maize Under Deficit Irrigations. J. Soil Sci. Plant Nutr. 2019, 20, 346–356. [Google Scholar] [CrossRef]
- Murgese, P.; Santamaria, P.; Leoni, B.; Crecchio, C. Ameliorative Effects of PGPB on Yield, Physiological Parameters, and Nutrient Transporter Genes Expression in Barattiere (Cucumis melo L.). J. Soil Sci. Plant Nutr. 2020, 20, 784–793. [Google Scholar] [CrossRef]
- Fasusi, O.A.; Cruz, C.; Babalola, O.O. Agricultural Sustainability: Microbial Biofertilizers in Rhizosphere Management. Agriculture 2021, 11, 163. [Google Scholar] [CrossRef]
- Igiri, B.E.; Okoduwa, S.I.; Idoko, G.O.; Akabuogu, E.P.; Adeyi, A.O.; Ejiogu, I.K. Toxicity and Bioremediation of Heavy Metals Contaminated Ecosystem from Tannery Wastewater: A Review. J. Toxicol. 2018, 2018, 2568038. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rodríguez, A.M.; Gastelum, L.A.S.; Pablos, C.M.F.; Parra-Cota, F.I.; Santoyo, G.; Puente, M.L.; Bhattacharya, D.; Mukherjee, J.; Santos-Villalobos, S.d.L. The Current and Future Role of Microbial Culture Collections in Food Security Worldwide. Front. Sustain. Food Syst. 2021, 4. [Google Scholar] [CrossRef]
- Panda, S.C. Organic Farming for Sustainable Agriculture, third ed.; Kalyani publisher: New Delhi, 2018; 9327216334. [Google Scholar]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Chakraborty, T.; Akhtar, N. Biofertilizers: Prospects and challenges for future. Biofertilizers 2021, 575–590. [Google Scholar] [CrossRef]
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight Into the Role of PGPR in Sustainable Agriculture and Environment. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Conway, G. One Billion Hungry: Can we Feed the World? Comstock Publishing Associates, 2012. [Google Scholar]
- Kumar, R.; Kumawat, N.; Sahu, Y.K. Role of biofertilizers in agriculture. Popular Kheti 2017, 5(4), 63–66. [Google Scholar]
- Odoh, C.K., Sam. Microbial consortium as biofertilizers for crops growing under extreme habitats. In Plant Microbiomes for Sustainable Agriculture; Yadav, A.N., Singh, J., Rastegari, A.A., Yadav, N., Eds.; Springer: Cham, 2020; pp. 381–424. [Google Scholar]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Berg, G. Plant–microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Brossi, M.J.d.L.; Kuramae, E.E.; Tsai, S.M. Land-use system shapes soil bacterial communities in Southeastern Amazon region. Appl. Soil Ecol. 2015, 95, 151–160. [Google Scholar] [CrossRef]
- Zhang, Q.; Saleem, M.; Wang, C. Probiotic strain Stenotrophomonas acidaminiphila BJ1 degrades and reduces chlorothalonil toxicity to soil enzymes, microbial communities and plant roots. AMB Express 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects A review. J. soil Sci. Plant Nutr. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, H.; Zhang, B.; Geng, M.; Cai, X.; Wang, J.; Wang, Y. Investigating the effect of microbial inoculants Frankia F1 on growth-promotion, rhizosphere soil physicochemical properties, and bacterial community of ginseng. Appl. Soil Ecol. 2021, 172, 104369. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, Y.; Yang, Y.; Zhang, M.; Mao, X.; Guo, Y.; Li, X.; Tao, B.; Qi, Y.; Ma, L.; et al. Co-application of biochar and microbial inoculants increases soil phosphorus and potassium fertility and improves soil health and tomato growth. J. Soils Sediments 2022, 23, 947–957. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 2011, 14, 605–611. [Google Scholar] [CrossRef]
- Gangwar, R.K., Bhushan. Combined effects of plant growth promoting rhizobacteria and fungi on mung bean (Vigna radiata l.). International Journal of Pharmaceutical Sciences and Research 2013, 4, 4422–4426. [Google Scholar] [CrossRef]
- Niu, X.; Duiker, S.W. Carbon sequestration potential by afforestation of marginal agricultural land in the Midwestern U.S. For. Ecol. Manag. 2006, 223, 415–427. [Google Scholar] [CrossRef]
- Rudel, T.K.; Coomes, O.T.; Moran, E.; Achard, F.; Angelsen, A.; Xu, J.; Lambin, E. Forest transitions: towards a global understanding of land use change. Glob. Environ. Chang. 2005, 15, 23–31. [Google Scholar] [CrossRef]
- Benayas, J.R.; Martins, A.; Nicolau, J.M.; Schulz, J.J. Abandonment of agricultural land: An overview of drivers and consequences. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 1–14. [Google Scholar] [CrossRef]
- Asif, M.; Mughal, A.H.; Bisma, R.; Mehdi, Z.; Saima, S.; Ajaz, M.; Malik, M.A.; Masood, A.; Sidique, S. Application of Different Strains of Biofertilizers for Raising Quality Forest Nursery. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 3680–3686. [Google Scholar] [CrossRef]
- Samec, P.; Kučera, A.; Tomášová, G. Soil Degradation Processes Linked to Long-Term Forest-Type Damage. Intech Open 2023. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant, Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Malhotra, S.K. Horticultural crops and climate change: A review. Indian J. Agric. Sci. 2017, 87, 12–22. [Google Scholar] [CrossRef]
- Gupta, R.; Kim, S.T. Depletion of RuBisCO protein using the protamine sulfate precipitation method. Methods in Molecular Biology 2017, 1295, 225–233. [Google Scholar] [CrossRef]
- Jordan, D.B.; Ogren, W.L. The CO2/O2 specificity of ribulose 1,5-bisphosphate carboxylase/oxygenase. Planta 1984, 161, 308–313. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Donati, I.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing Climate Change: Application of Microbial Biostimulants to Mitigate Stress in Horticultural Crops. Agronomy 2020, 10, 794. [Google Scholar] [CrossRef]
- Ku, S.-B.; Edwards, G.E. Oxygen inhibition of photosynthesis. Planta 1978, 140, 1–6. [Google Scholar] [CrossRef]
- Bhattacharya, A. Effect of High Temperature on Crop Productivity and Metabolism of Macro Molecules; Elsevier: Amsterdam, NX, Netherlands, 2019. [Google Scholar]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Qu, A.-L.; Ding, Y.-F.; Jiang, Q.; Zhu, C. Molecular mechanisms of the plant heat stress response. Biochem. Biophys. Res. Commun. 2013, 432, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Driedonks, N.; Xu, J.; Peters, J.L.; Park, S.; Rieu, I. Multi-Level Interactions Between Heat Shock Factors, Heat Shock Proteins, and the Redox System Regulate Acclimation to Heat. Front. Plant Sci. 2015, 6, 999. [Google Scholar] [CrossRef] [PubMed]
- Byerlee, D.; Stevenson, J.; Villoria, N. Does intensification slow crop land expansion or encourage deforestation? Glob. Food Secur. 2014, 3, 92–98. [Google Scholar] [CrossRef]
- Husain, T.; Fatima, A.; Suhel, M.; Singh, S.; Sharma, A.; Prasad, S.M.; Singh, V.P. A brief appraisal of ethylene signaling under abiotic stress in plants. Plant Signal. Behav. 2020, 15. [Google Scholar] [CrossRef]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge. Plants 2022, 11, 2211. [Google Scholar] [CrossRef]
- Dubois, M.; Van den Broeck, L.; Inzé, D. The Pivotal Role of Ethylene in Plant Growth. Trends Plant Sci. 2018, 23, 311–323. [Google Scholar] [CrossRef]
- Duc, N.H.; Csintalan, Z.; Posta, K. Arbuscular mycorrhizal fungi mitigate negative effects of combined drought and heat stress on tomato plants. Plant Physiol. Biochem. 2018, 132, 297–307. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Babalola, O.O.; Santoyo, G.; Perazzolli, M. The Potential Role of Microbial Biostimulants in the Amelioration of Climate Change-Associated Abiotic Stresses on Crops. Front. Microbiol. 2022, 12, 829099. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- del Carmen Orozco-Mosqueda, M.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Esmaeel, Q.; Miotto, L.; Rondeau, M.; Leclère, V.; Clément, C.; Jacquard, C.; Sanchez, L.; Barka, E.A. Paraburkholderia phytofirmans PsJN-Plants Interaction: From Perception to the Induced Mechanisms. Front. Microbiol. 2018, 9, 2093. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Perinjelil, S.J.; Thakur, N.D. Chapter 11 - Soil salinization and bioremediation using halophiles and halotolerant microorganisms. Gustavo Santoyo, Ajay Kumar, Mohd Aamir, Ed.; Mitigation of Plant Abiotic Stress by Microorganisms, Academic Press, 2022. [Google Scholar] [CrossRef]
- Mitsuchi, M., Wichaidit, P., Jeungnijnirund, S. Soils of the Northeast Plateau, Thailand. Technical bulletin of the Tropical Agriculture Research Center. Japan, No. 25. 1989. https://www.jircas.go.jp/en/publication/techtarc/25/1.
- Sharifi, R.; Ryu, C.-M. Sniffing bacterial volatile compounds for healthier plants. Curr. Opin. Plant Biol. 2018, 44, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Rehman, M.U.; Siddiqi, T.O.; Ahmad, P. Salt Stress: Causes, Types and Responses of Plants. In Ecophysiology and Responses of Plants under Salt Stress; Springer, 2012; pp. 1–24. [Google Scholar]
- Alonso-Español, A.; Bravo, E.; Ribeiro-Vidal, H.; Virto, L.; Herrera, D.; Alonso, B.; Sanz, M. The Antimicrobial Activity of Curcumin and Xanthohumol on Bacterial Biofilms Developed over Dental Implant Surfaces. Int. J. Mol. Sci. 2023, 24, 2335. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, G.; Zhang, Y.; Guan, X.; Wei, Y.; Guo, R. Global desertification vulnerability to climate change and human activities. Land Degrad. Dev. 2020, 31, 1380–1391. [Google Scholar] [CrossRef]
- Grover, M.; Ali, S.Z.; Sandhya, V.; Rasul, A.; Venkateswarlu, B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 2010, 27, 1231–1240. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial Exopolysaccharides: Functionality and Prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Multifunctional Exopolysaccharides from Pseudomonas aeruginosa PF23 Involved in Plant Growth Stimulation, Biocontrol and Stress Amelioration in Sunflower Under Saline Conditions. Curr. Microbiol. 2014, 69, 484–494. [Google Scholar] [CrossRef]
- El-Ghany, M.F.A.; Attia, M. Effect of Exopolysaccharide-Producing Bacteria and Melatonin on Faba Bean Production in Saline and Non-Saline Soil. Agronomy 2020, 10, 316. [Google Scholar] [CrossRef]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. A Halotolerant Bacterium Bacillus licheniformis HSW-16 Augments Induced Systemic Tolerance to Salt Stress in Wheat Plant (Triticum aestivum). Front. Plant Sci. 2016, 7, 1890. [Google Scholar] [CrossRef]
- Khan, A.L.; Hussain, J.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Endophytic fungi: resource for gibberellins and crop abiotic stress resistance. Crit. Rev. Biotechnol. 2013, 35, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; van der Heijden, M.G. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-Bacterial Amelioration of Plant Abiotic and Biotic Stress. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Gong, M.; Tang, M.; Chen, H.; Zhang, Q.; Feng, X. Effects of two Glomus species on the growth and physiological performance of Sophora davidii seedlings under water stress. New For. 2012, 44, 399–408. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Ayangbenro, A.S.; Babalola, O.O. Organic Farming Enhances the Diversity and Community Structure of Endophytic Archaea and Fungi in Maize Plant: a Shotgun Approach. J. Soil Sci. Plant Nutr. 2020, 20, 2587–2599. [Google Scholar] [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozan, J.M. How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporins in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytol. 2007, 173, 808–816. [Google Scholar] [CrossRef]
- Bray, E. A “Response to abiotic stress,” in Biochemistry and Molecular Biology of Plants. (2000). eds B. B. Buchanan, W. Gruissem, and R. L. Jones (Rockville, MD: ASPP), 1158–1203.
- Omomowo, I.O.; Fadiji, A.E.; Omomowo, O.I. Assessment of bio-efficacy of Glomus versiforme and Trichoderma harzianum in inhibiting powdery mildew disease and enhancing the growth of cowpea. Ann. Agric. Sci. 2018, 63, 9–17. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Plant-growth-promoting rhizobacteria: drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 2015, 66, 35–42. [Google Scholar] [CrossRef]
- Sharifi, R.; Ryu, C.-M. Sniffing bacterial volatile compounds for healthier plants. Curr. Opin. Plant Biol. 2018, 44, 88–97. [Google Scholar] [CrossRef]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; et al. A Drought Resistance-Promoting Microbiome Is Selected by Root System under Desert Farming. PLOS ONE 2012, 7, e48479. [Google Scholar] [CrossRef]
- Jayakumar, A.; Padmakumar, P.; Nair, I.C.; Radhakrishnan, E.K. Drought tolerant bacterial endophytes with potential plant probiotic effects from Ananas comosus. Biologia 2020, 75, 1769–1778. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria that confer resistance to water stress in tomatoes and peppers. Plant Sci. 2003, 166, 525–530. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Samiyappan, R. ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 2006, 102, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Pei, H.; Xu, Z. Low nitrogen stress stimulating the indole-3-acetic acid biosynthesis of Serratia sp. ZM is vital for the survival of the bacterium and its plant growth-promoting characteristic. Arch. Microbiol. 2016, 199, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D. Survival of Pseudomonas extremorientalis TSAU20 and P. chlororaphis TSAU13 in the rhizosphere of common bean (Phaseolus vulgaris) under saline conditions. Plant, Soil Environ. 2011, 57, 122–127. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Zhang, F.; Zhang, D.-J.; Srivastava, A.; Wu, Q.-S.; Zou, Y.-N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Liu, F.; Xing, S.; Ma, H.; Du, Z.; Ma, B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Kang, S.-M.; Radhakrishnan, R.; You, Y.-H.; Khan, A.L.; Park, J.-M.; Lee, S.-M.; Lee, I.-J. Cucumber performance is improved by inoculation with plant growth-promoting microorganisms. Acta Agric. Scand. Sect. B — Soil Plant Sci. 2014, 65, 36–44. [Google Scholar] [CrossRef]
- Shaffique, S.; Hussain, S.; Kang, S.-M.; Imran, M.; Hoque, I.U.; Khan, M.A.; Lee, I.-J. Phytohormonal modulation of the drought stress in soybean: outlook, research progress, and cross-talk. Front. Plant Sci. 2023, 14, 1237295. [Google Scholar] [CrossRef]
- Jahromi, F.; Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Influence of Salinity on the In Vitro Development of Glomus intraradices and on the In Vivo Physiological and Molecular Responses of Mycorrhizal Lettuce Plants. Microb. Ecol. 2007, 55, 45–53. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Golpayegani, A. Effects of water stress and inoculation with plant growth promoting rhizobacteria (PGPR) on antioxidant status and photosynthetic pigments in basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 2011, 11, 57–61. [Google Scholar] [CrossRef]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular signature of hypersaline adaptation: insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70–R70. [Google Scholar] [CrossRef] [PubMed]
- Sanders, G.J.; Arndt, S.K. Osmotic Adjustment Under Drought Conditions. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 199–229. [Google Scholar] [CrossRef]
- Shintu, P. , Jayaram, K. Phosphate solubilising bacteria (Bacillus polymyxa)-An effective approach to mitigate drought in tomato (Lycopersicon esculentum Mill.). Tropical Plant Research 2015, 2, 17–22. [Google Scholar]
- Dimkpa, C.O.; Merten, D.; Svatos, A.; Büchel, G.; Kothe, E. Siderophores mediate reduced and increased uptake of cadmium by Streptomyces tendae F4 and sunflower (Helianthus annuus), respectively. J. Appl. Microbiol. 2009, 107, 1687–1696. [Google Scholar] [CrossRef]
- Bailly, A.; Groenhagen, U.; Schulz, S.; Geisler, M.; Eberl, L.; Weisskopf, L. The inter-kingdom volatile signal indole promotes root development by interfering with auxin signalling. Plant J. 2014, 80, 758–771. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Garladinne, M.; Lee, Y.H. Volatile Indole Produced by Rhizobacterium Proteus vulgaris JBLS202 Stimulates Growth of Arabidopsis thaliana Through Auxin, Cytokinin, and Brassinosteroid Pathways. J. Plant Growth Regul. 2014, 34, 158–168. [Google Scholar] [CrossRef]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.-S.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Choline and Osmotic-Stress Tolerance Induced in Arabidopsis by the Soil Microbe Bacillus subtilis (GB03). Mol. Plant-Microbe Interactions® 2010, 23, 1097–1104. [Google Scholar] [CrossRef]
- Ledger, T.; Rojas, S.; Timmermann, T.; Pinedo, I.; Poupin, M.J.; Garrido, T.; Richter, P.; Tamayo, J.; Donoso, R. Volatile-Mediated Effects Predominate in Paraburkholderia phytofirmans Growth Promotion and Salt Stress Tolerance of Arabidopsis thaliana. Front. Microbiol. 2016, 7, 1838. [Google Scholar] [CrossRef]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; de la Cruz, H.R.; Macías-Rodríguez, L. Plant growth-promoting rhizobacteria modulate root-system architecture in Arabidopsis thaliana through volatile organic compound emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.-M.; Lee, I.-J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- Else, M.A.; Jackson, M.B. Transport of 1-aminocyclopropane-1-carboxylic acid (ACC) in the transpiration stream of tomato (Lycopersicon esculentum) in relation to foliar ethylene production and petiole epinasty. Funct. Plant Biol. 1998, 25, 453–458. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.-C. Plant Growth Promotion Under Water: Decrease of Waterlogging-Induced ACC and Ethylene Levels by ACC Deaminase-Producing Bacteria. Front. Microbiol. 2018, 9, 1096. [Google Scholar] [CrossRef] [PubMed]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containingplant growth-promoting bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.-J.; Hao, Z.-P.; Li, H.; Chen, B.-D. Aquaporin genesGintAQPF1andGintAQPF2fromGlomus intraradicescontribute to plant drought tolerance. Plant Signal. Behav. 2013, 8, e24030. [Google Scholar] [CrossRef]
- Jaemsaeng, R.; Jantasuriyarat, C.; Thamchaipenet, A. Positive role of 1-aminocyclopropane-1-carboxylate deaminase-producing endophytic Streptomyces sp. GMKU 336 on flooding resistance of mung bean. Agric. Nat. Resour. 2018, 52, 330–334. [Google Scholar] [CrossRef]
- Fravel, D. Commercialization and Implementation of Biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Manikandan, A., Anandham, R., Johnson, I., Krishnamoorthy, R., Senthilkumar, M., Raghu, R., et al. Bacillus and Streptomyces for Management of Biotic Stresses in Plants for Sustainable Agriculture. In S. Chhabra, R. Prasad, N. R. Maddela, & N. Tuteja (Eds.), Plant Microbiome for Plant Productivity and Sustainable Agriculture (pp. 263-288). Singapore: Springer Nature Singapore. 2023. [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef]
- Bernal, S.; Hedin, L.O.; Likens, G.E.; Gerber, S.; Buso, D.C. Complex response of the forest nitrogen cycle to climate change. Proc. Natl. Acad. Sci. 2012, 109, 3406–3411. [Google Scholar] [CrossRef]
- Vale, P.; Gibbs, H.; Vale, R.; Christie, M.; Florence, E.; Munger, J.; Sabaini, D. The Expansion of Intensive Beef Farming to the Brazilian Amazon. Glob. Environ. Chang. 2019, 57, 101922. [Google Scholar] [CrossRef]
- INPE, 2021. National Institute for Space Research. http://www.inpe.br/.
- Navarrete, A.A.; Tsai, S.M.; Mendes, L.W.; Faust, K.; de Hollander, M.; Cassman, N.A.; Raes, J.; van Veen, J.A.; Kuramae, E.E. Soil microbiome responses to the short-term effects of Amazonian deforestation. Mol. Ecol. 2015, 24, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Melo, V.F.; Orrutéa, A.G.; Motta, A.C.V.; Testoni, S.A. Land Use and Changes in Soil Morphology and Physical-Chemical Properties in Southern Amazon. Rev. Bras. De Cienc. Do Solo 2017, 41. [Google Scholar] [CrossRef]
- Braz, A.M.d.S.; Fernandes, A.R.; Alleoni, L.R.F. SOIL ATTRIBUTES AFTER THE CONVERSION FROM FOREST TO PASTURE IN AMAZON. Land Degrad. Dev. 2011, 24, 33–38. [Google Scholar] [CrossRef]
- Cenciani, K.; Lambais, M.R.; Cerri, C.C.; de Azevedo, L.C.B.; Feigl, B.J. Bacteria diversity and microbial biomass in forest, pasture and fallow soils in the southwestern Amazon basin. Rev. Bras. De Cienc. Do Solo 2009, 33, 907–916. [Google Scholar] [CrossRef]
- Carvalho, R.; Adami, M.; Amaral, S.; Bezerra, F.G.; de Aguiar, A.P.D. Changes in secondary vegetation dynamics in a context of decreasing deforestation rates in Pará, Brazilian Amazon. Appl. Geogr. 2019, 106, 40–49. [Google Scholar] [CrossRef]
- Pedrinho, A.; Mendes, L.W.; Merloti, L.F.; Andreote, F.D.; Tsai, S.M. The natural recovery of soil microbial community and nitrogen functions after pasture abandonment in the Amazon region. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef]
- Porder, S.; Vitousek, P.M.; Chadwick, O.A.; Chamberlain, C.P.; Hilley, G.E. Uplift, Erosion, and Phosphorus Limitation in Terrestrial Ecosystems. Ecosystems 2007, 10, 159–171. [Google Scholar] [CrossRef]
- Gama-Rodrigues, A.C.; Sales, M.V.S.; Silva, P.S.D.; Comerford, N.B.; Cropper, W.P.; Gama-Rodrigues, E.F. An exploratory analysis of phosphorus transformations in tropical soils using structural equation modeling. Biogeochemistry 2014, 118, 453–469. [Google Scholar] [CrossRef]
- Alonso-Español, A.; Bravo, E.; Ribeiro-Vidal, H.; Virto, L.; Herrera, D.; Alonso, B.; Sanz, M. The Antimicrobial Activity of Curcumin and Xanthohumol on Bacterial Biofilms Developed over Dental Implant Surfaces. Int. J. Mol. Sci. 2023, 24, 2335. [Google Scholar] [CrossRef]
- Johnson, A.H.; Frizano, J.; Vann, D.R. Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure. Oecologia 2003, 135, 487–499. [Google Scholar] [CrossRef]
- Shrivastava, M., Srivastava, P., D’Souza, S. Phosphate-Solubilizing Microbes: Diversity and Phosphates Solubilization Mechanism. In: Meena, V. (eds) Role of Rhizospheric Microbes in Soil. Springer, Singapore. 2018. [CrossRef]
- Chavarro-Bermeo, J.P.; Arruda, B.; Mora-Motta, D.A.; Bejarano-Herrera, W.; Ortiz-Morea, F.A.; Somenahally, A.; Silva-Olaya, A.M. Responses of Soil Phosphorus Fractions to Land-Use Change in Colombian Amazon. Sustainability 2022, 14, 2285. [Google Scholar] [CrossRef]
- Hamer, U.; Potthast, K.; Burneo, J.I.; Makeschin, F. Nutrient stocks and phosphorus fractions in mountain soils of Southern Ecuador after conversion of forest to pasture. Biogeochemistry 2012, 112, 495–510. [Google Scholar] [CrossRef]
- Garcia-Montiel, D.C.; Neill, C.; Melillo, J.; Thomas, S.; Steudler, P.A.; Cerri, C.C. Soil Phosphorus Transformations Following Forest Clearing for Pasture in the Brazilian Amazon. Soil Sci. Soc. Am. J. 2000, 64, 1792–1804. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology and Biochemistry, fourth ed. Academic Press, London. 2014.
- Fu, D.; Wu, X.; Duan, C.; Chadwick, D.R.; Jones, D.L. Response of soil phosphorus fractions and fluxes to different vegetation restoration types in a subtropical mountain ecosystem. CATENA 2020, 193, 104663. [Google Scholar] [CrossRef]
- Teixeira, H.M.; Cardoso, I.M.; Bianchi, F.J.; Silva, A.d.C.; Jamme, D.; Peña-Claros, M. Linking vegetation and soil functions during secondary forest succession in the Atlantic forest. For. Ecol. Manag. 2019, 457, 117696. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability Update on Microbial Phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef]
- Arenberg, M.R.; Arai, Y. Uncertainties in soil physicochemical factors controlling phosphorus mineralization and immobilization processes, first ed. Elsevier, London. 2019. [CrossRef]
- Rodríguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, W.; Liu, G.; Liang, G.; He, P.; Liu, Z. Soil C/N and pH together as a comprehensive indicator for evaluating the effects of organic substitution management in subtropical paddy fields after application of high-quality amendments. Geoderma 2018, 337, 1116–1125. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Khan, S.M.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganism in sustainable agriculture: a review. Agronomy for Sustainable Development 2018, 27, 29–43. [Google Scholar] [CrossRef]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Ahmad, E.; Zaidi, A.; Oves, M. Functional aspect of phosphate solubilizing bacteria: importance in crop production. In Bacteria in Agrobiology: Crop Productivity; Maheshwari, D.K., Ed.; Springer: Berlin, 2013. [Google Scholar] [CrossRef]
- Bi, Q.-F.; Li, K.-J.; Zheng, B.-X.; Liu, X.-P.; Li, H.-Z.; Jin, B.-J.; Ding, K.; Yang, X.-R.; Lin, X.-Y.; Zhu, Y.-G. Partial replacement of inorganic phosphorus (P) by organic manure reshapes phosphate mobilizing bacterial community and promotes P bioavailability in a paddy soil. Sci. Total. Environ. 2019, 703, 134977. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.W.; Tsai, S.M.; Navarrete, A.A.; de Hollander, M.; van Veen, J.A.; Kuramae, E.E. Soil-Borne Microbiome: Linking Diversity to Function. Microb. Ecol. 2015, 70, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.-M.; Lee, I.-J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- Pedrinho, A.; Mendes, L.W.; Merloti, L.F.; Fonseca, M.d.C.d.; Cannavan, F.d.S.; Tsai, S.M. Forest-to-pasture conversion and recovery based on assessment of microbial communities in Eastern Amazon rainforest. FEMS Microbiol. Ecol. 2018, 95. [Google Scholar] [CrossRef]
- Batista, P.H.D.; de Almeida, G.L.P.; da Silva, J.L.B.; Pandorfi, H.; Da Silva, M.V.; da Silva, R.A.B.; de Melo, M.V.N.; Lins, F.A.C.; Junior, J.J.F.C. Short-term grazing and its impacts on soil and pasture degradation. DYNA 2020, 87, 123–128. [Google Scholar] [CrossRef]
- Sharrow, S.H. Soil compaction by grazing livestock in silvopastures as evidenced by changes in soil physical properties. Agrofor. Syst. 2007, 71, 215–223. [Google Scholar] [CrossRef]
- Pentoś, K.; Pieczarka, K.; Serwata, K. The Relationship between Soil Electrical Parameters and Compaction of Sandy Clay Loam Soil. Agriculture 2021, 11, 114. [Google Scholar] [CrossRef]
- de Moraes, M.T.; da Silva, V.R.; Zwirtes, A.L.; Carlesso, R. Use of penetrometers in agriculture: a review. Eng. Agricola 2014, 34, 179–193. [Google Scholar] [CrossRef]
- Krajco, J. Detection of soil compaction using soil electrical conductivity. Master’s Thesis, Cranfield University, Silsoe, UK, 2007.
- Nawaz, M.F.; Bourrié, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2012, 33, 291–309. [Google Scholar] [CrossRef]
- Mayerfeld, D.; Kruger, E.; Gildersleeve, R.; Rickenbach, M. Impacts of different grazing approaches on woodland ecosystem properties. Agrofor. Syst. 2021, 96, 527–540. [Google Scholar] [CrossRef]
- M., C.M. Effects of soil compaction by trampling of animals in soil productivity. Remediations. Rev. Colomb. de Cienc. Anim. - RECIA 2016, 8, 88–93. [Google Scholar] [CrossRef]
- Donkor, N.T.; Gedir, J.V.; Hudson, R.J.; Bork, E.W.; Chanasyk, D.S.; Naeth, M.A. Impacts of grazing systems on soil compaction and pasture production in Alberta. Can. J. Soil Sci. 2002, 82, 1–8. [Google Scholar] [CrossRef]
- Gao, S.; DeLuca, T.H. Rangeland application of biochar and rotational grazing interact to influence soil and plant nutrient dynamics. Geoderma 2022, 408. [Google Scholar] [CrossRef]
- An, N.; Zhang, L.; Liu, Y.; Shen, S.; Li, N.; Wu, Z.; Yang, J.; Han, W.; Han, X. Biochar application with reduced chemical fertilizers improves soil pore structure and rice productivity. Chemosphere 2022, 298, 134304. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Song, S.; Rao, P.; Wang, R.; Liu, S. Preparation and properties of soil conditioner microspheres based on self-assembled potassium alginate and chitosan. Int. J. Biol. Macromol. 2019, 147, 877–889. [Google Scholar] [CrossRef]
- Cui, K.; Xu, L.; Tao, T.; Huang, L.; Li, J.; Hong, J.; Li, H.; Chi, Y. Mechanical behavior of multiscale hybrid fiber reinforced recycled aggregate concrete subject to uniaxial compression. Journal of Building Engineering 2023, 71, 106504. [Google Scholar] [CrossRef]
- Cui, K.; Xu, L.; Tao, T.; Huang, L.; Li, J.; Hong, J.; Li, H.; Chi, Y. Mechanical behavior of multiscale hybrid fiber reinforced recycled aggregate concrete subject to uniaxial compression. Journal of Building Engineering 2023, 71, 106504. [Google Scholar] [CrossRef]
- Cui, Q.; Xia, J.; Yang, H.; Liu, J.; Shao, P. Biochar and effective microorganisms promote Sesbania cannabina growth and soil quality in the coastal saline-alkali soil of the Yellow River Delta, China. Sci. Total Environ. 2021, 756, 143801. [Google Scholar] [CrossRef]
- Cui, K.; Xu, L.; Tao, T.; Huang, L.; Li, J.; Hong, J.; Li, H.; Chi, Y. Mechanical behavior of multiscale hybrid fiber reinforced recycled aggregate concrete subject to uniaxial compression. Journal of Building Engineering 2023, 71, 106504. [Google Scholar] [CrossRef]
- Chander, M.P.; Kartick, C.; Gangadhar, J.; Vijayachari, P. Ethno medicine and healthcare practices among Nicobarese of Car Nicobar - an indigenous tribe of Andaman and Nicobar Islands. J. Ethnopharmacol. 2014, 158, 18–14. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L. M., Rezende, H.C., Coelho, L., M., de Sousa, P.A.R., Melo, D.F.O., Coelho, N.M.M. Bioremediation of Polluted Waters Using Microorganisms 2015. InTech. [CrossRef]
- Nicolaou, S.A.; Gaida, S.M.; Papoutsakis, E.T. A comparative view of metabolite and substrate stress and tolerance in microbial bioprocessing: From biofuels and chemicals, to biocatalysis and bioremediation. Metab. Eng. 2010, 12, 307–331. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A., Bisht. Review on bioremediation of polluted environment: a management tool. International Journal of Environmental Sciences 2011, 1, 1079–1093. [Google Scholar]
- Wasilkowski, D., Swedziol. The applicability of genetically modified microorganisms in bioremediation of contaminated environments. Chemik 2012, 66, 822–826. [Google Scholar]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef]
- Mushtaq, S.; e Bareen, F.; Tayyeb, A. Equilibrium kinetics and thermodynamic studies on biosorption of heavy metals by metal-resistant strains of Trichoderma isolated from tannery solid waste. Environ. Sci. Pollut. Res. 2022, 30, 10925–10954. [Google Scholar] [CrossRef]
- Atuchin, V.V.; Asyakina, L.K.; Serazetdinova, Y.R.; Frolova, A.S.; Velichkovich, N.S.; Prosekov, A.Y. Microorganisms for Bioremediation of Soils Contaminated with Heavy Metals. Microorganisms 2023, 11, 864. [Google Scholar] [CrossRef]
- Effendi, A.J.; Mastroiani, L.J.; Suhardi, S.H.; Ramadan, B.S. Potential use of extracellular polymeric substances (EPS) of Bacillus subtilis for biosorption of mercury produced from soil-washing effluent. Bioresour. Technol. Rep. 2023, 22. [Google Scholar] [CrossRef]
- Mathivanan, K.; Chandirika, J.U.; Srinivasan, R.; Charles, P.E.; Rajaram, R.; Zhang, R. Exopolymeric substances production by Bacillus cereus KMS3-1 enhanced its biosorption efficiency in removing Cd2+ and Pb2+ in single and binary metal mixtures. Environ. Res. 2023, 228, 115917. [Google Scholar] [CrossRef]
- Bioremediation of Heavy Metals Using Microorganisms. In Genetically Engineered Organisms in Bioremediation; Nagmote, M.S., Rai, A.R., Sharma, R., Desimone, M.F., Chaudhary, R.G., Singh, N.B., Eds.; CRC Press, 2024; pp. 168–190. [Google Scholar]
- Rahman, M.A.; Hassler, C. Is arsenic biotransformation a detoxification mechanism for microorganisms? Aquat. Toxicol. 2014, 146, 212–219. [Google Scholar] [CrossRef]
- Mala, J.G.S.; Sujatha, D.; Rose, C. Inducible chromate reductase exhibiting extracellular activity in Bacillus methylotrophicus for chromium bioremediation. Microbiol. Res. 2015, 170, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Barkay, T.; Gu, B. Demethylation─The Other Side of the Mercury Methylation Coin: A Critical Review. ACS Environ. Au 2021, 2, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.; Thomas, L.; Chetri, S.P.K.; Bodhankar, S.; Kumar, V.; Naidu, R. A comprehensive review on chromium (Cr) contamination and Cr(VI)-resistant extremophiles in diverse extreme environments. Environ. Sci. Pollut. Res. 2023, 30, 59163–59193. [Google Scholar] [CrossRef] [PubMed]
- Elnabi, M.K.A.; Elkaliny, N.E.; Elyazied, M.M.; Azab, S.H.; Elkhalifa, S.A.; Elmasry, S.; Mouhamed, M.S.; Shalamesh, E.M.; Alhorieny, N.A.; Elaty, A.E.A.; et al. Toxicity of Heavy Metals and Recent Advances in Their Removal: A Review. Toxics 2023, 11, 580. [Google Scholar] [CrossRef]
- Roane, T.M.; Josephson, K.L.; Pepper, I.L. Dual-Bioaugmentation Strategy To Enhance Remediation of Cocontaminated Soil. Appl. Environ. Microbiol. 2001, 67, 3208–3215. [Google Scholar] [CrossRef]
- Vullo, D.L.; Ceretti, H.M.; Daniel, M.A.; Ramírez, S.A.; Zalts, A. Cadmium, zinc and copper biosorption mediated by Pseudomonas veronii 2E. Bioresour. Technol. 2007, 99, 5574–5581. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z.; Chai, L.; Wang, Y.; Liu, Y.; Xiao, R. Bioleaching remediation of heavy metal-contaminated soils using Burkholderia sp. Z-90. J. Hazard. Mater. 2016, 301, 145–152. [Google Scholar] [CrossRef]
- Achal, V.; Pan, X.; Zhang, D. Remediation of copper-contaminated soil by Kocuria flava CR1, based on microbially induced calcite precipitation. Ecol. Eng. 2011, 37, 1601–1605. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. Genomic analysis of Bacillus cereus NWUAB01 and its heavy metal removal from polluted soil. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Achal, V.; Pan, X.; Fu, Q.; Zhang, D. Biomineralization based remediation of As(III) contaminated soil by Sporosarcina ginsengisoli. J. Hazard. Mater. 2011, 201-202, 178–184. [Google Scholar] [CrossRef]
- Li, X.; Dong, S.; Yao, Y.; Shi, W.; Wu, M.; Xu, H. Inoculation of bacteria for the bioremediation of heavy metals contaminated soil by Agrocybe aegerita. RSC Adv. 2016, 6, 65816–65824. [Google Scholar] [CrossRef]
- Kang, C.-H.; Kwon, Y.-J.; So, J.-S. Bioremediation of heavy metals by using bacterial mixtures. Ecol. Eng. 2016, 89, 64–69. [Google Scholar] [CrossRef]
- Rahman, A.; Nahar, N.; Nawani, N.N.; Jass, J.; Hossain, K.; Alam Saud, Z.; Saha, A.K.; Ghosh, S.; Olsson, B.; Mandal, A. Bioremediation of hexavalent chromium (VI) by a soil-borne bacterium,Enterobacter cloacaeB2-DHA. J. Environ. Sci. Heal. Part A 2015, 50, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Wani, P.A.; Rafi, N.; Wani, U.; A, H.B.; Khan, M.S.A. Simultaneous bioremediation of heavy metals and biodegradation of hydrocarbons by metal resistantBrevibacillus parabrevisOZF5 improves plant growth promotion. Bioremediation J. 2021, 27, 20–31. [Google Scholar] [CrossRef]
- Ridene, S.; Werfelli, N.; Mansouri, A.; Landoulsi, A.; Abbes, C. Bioremediation potential of consortium Pseudomonas Stutzeri LBR and Cupriavidus Metallidurans LBJ in soil polluted by lead. PLOS ONE 2023, 18, e0284120. [Google Scholar] [CrossRef]
- Vishwakarma, S.K.; Patil, A.; Pandey, A.; Arya, M. Biosorption of Heavy Metal (Mn2+) by Thermophilic Bacterial Strains Isolated from Surya Kund Hot Spring, Yamunotri, Uttarakhand. Appl. Biochem. Biotechnol. 2023, 196, 2518–2533. [Google Scholar] [CrossRef]
- Cam, N.; Benzerara, K.; Georgelin, T.; Jaber, M.; Lambert, J.-F.; Poinsot, M.; Skouri-Panet, F.; Cordier, L. Selective Uptake of Alkaline Earth Metals by Cyanobacteria Forming Intracellular Carbonates. Environ. Sci. Technol. 2016, 50, 11654–11662. [Google Scholar] [CrossRef]
- Al-Sherif, A.; Abd El-Hameed, M.S.; Mahmoud, M.A.; Ahmed, H.S. Use of cyanobacteria and organic fertilizer mixture as soil bioremediation. American-Eurasian Journal of Agricultural & Environmental Sciences 2015, 15, 794–799. [Google Scholar] [CrossRef]
- Jin, Z.; Deng, S.; Wen, Y.; Jin, Y.; Pan, L.; Zhang, Y.; Black, T.; Jones, K.C.; Zhang, H.; Zhang, D. Application of Simplicillium chinense for Cd and Pb biosorption and enhancing heavy metal phytoremediation of soils. Sci. Total. Environ. 2019, 697, 134148. [Google Scholar] [CrossRef]
- Chang, J.; Shi, Y.; Si, G.; Yang, Q.; Dong, J.; Chen, J. The bioremediation potentials and mercury(II)-resistant mechanisms of a novel fungus Penicillium spp. DC-F11 isolated from contaminated soil. J. Hazard. Mater. 2020, 396, 122638. [Google Scholar] [CrossRef]
- Din, G.; Hassan, A.; Dunlap, J.; Ripp, S.; Shah, A.A. Cadmium tolerance and bioremediation potential of filamentous fungus Penicillium chrysogenum FMS2 isolated from soil. Int. J. Environ. Sci. Technol. 2021, 19, 2761–2770. [Google Scholar] [CrossRef]
- Taştan, B.E.; Ertuğrul, S.; Dönmez, G. Effective bioremoval of reactive dye and heavy metals by Aspergillus versicolor. Bioresour. Technol. 2009, 101, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.K.; Congeevaram, S.; Thamaraiselvi, K. Evaluation of isolated fungal strain from e-waste recycling facility for effective sorption of toxic heavy metal Pb (II) ions and fungal protein molecular characterization-a Mycoremediation approach. Asian Journal of Experimental Biological Sciences 2011, 2, 342–347. [Google Scholar]
- Hasgul, E.; Malkoç, S.; Güven, A.; Dede, A.; Güven, K. Biosorption of cadmium and copper by Aspergillus spp. isolated from industrial ceramic waste sludge. Biol. Divers. Conserv. 2019, 12, 44–56. [Google Scholar] [CrossRef]
- Hassan, A.; Pariatamby, A.; Ahmed, A.; Auta, H.S.; Hamid, F.S. Enhanced Bioremediation of Heavy Metal Contaminated Landfill Soil Using Filamentous Fungi Consortia: a Demonstration of Bioaugmentation Potential. Water, Air, Soil Pollut. 2019, 230, 215. [Google Scholar] [CrossRef]
- Bahafid, W.; Tahri Joutey, N.; Sayel, H.; Boularab, I.; EL Ghachtouli, N. Bioaugmentation of chromium-polluted soil microcosms with Candida tropicalis diminishes phytoavailable chromium. J. Appl. Microbiol. 2013, 115, 727–734. [Google Scholar] [CrossRef]
- Damodaran, D.; Suresh, G.; Mohan, R. Bioremediation of soil by removing heavy metals using Saccharomyces cerevisiae. In 2nd international conference on environmental science and technology; Singapore, 2011. [Google Scholar]
- Imam, S.S.A. Comparative Study of Heavy Metal Bioremediation in Soil by Bacillus Subtilis and Saccharomyces Cerevisiae. Indian J. Sci. Technol. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Mao, Q.; Xie, X.; Pinzon-Nuñez, D.A.; Xie, Z.; Liu, T.; Irshad, S. Native microalgae and Bacillus XZM remediate arsenic-contaminated soil by forming biological soil crusts. J. Environ. Manag. 2023, 345, 118858. [Google Scholar] [CrossRef]
- Ri, C.; Tao, Y.; Tu, J.; Li, X.; She, S.; Hou, L.; Fu, Y.; Chen, L. Improvement of the Cd removal efficiency of a filamentous cyanobacterium Leptolyngbya sp. XZ1 through co-culture with Bacillus sp. S1. J. Appl. Phycol. 2023, 35, 2935–2944. [Google Scholar] [CrossRef]
- Gupta, R.; Khan, F.; Alqahtani, F.M.; Hashem, M.; Ahmad, F. Plant Growth–Promoting Rhizobacteria (PGPR) Assisted Bioremediation of Heavy Metal Toxicity. Appl. Biochem. Biotechnol. 2023, 196, 2928–2956. [Google Scholar] [CrossRef]
- Ledin, M. Accumulation of metals by microorganisms — processes and importance for soil systems. Earth-Science Rev. 2000, 51, 1–31. [Google Scholar] [CrossRef]
- Thapa, B.; Kc, A.K.; Ghimire, A. A Review On Bioremediation Of Petroleum Hydrocarbon Contaminants In Soil. 1970, 8, 164–170. [Google Scholar] [CrossRef]
- Gao, B.; Wang, X.; Ford, R.M. Chemotaxis along local chemical gradients enhanced bacteria dispersion and PAH bioavailability in a heterogenous porous medium. Sci. Total. Environ. 2022, 859, 160004. [Google Scholar] [CrossRef] [PubMed]
- Purnomo, A.S.; Putra, S.R.; Putro, H.S.; Hamzah, A.; Rohma, N.A.; Rohmah, A.A.; Rizqi, H.D.; Asranudin; Tangahu, B.V.; Warmadewanthi, I.D.A.A. The application of biosurfactant-producing bacteria immobilized in PVA/SA/bentonite bio-composite for hydrocarbon-contaminated soil bioremediation. RSC Adv. 2023, 13, 21163–21170. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Hou, J.; Du, M.; Zhang, Y.; Liu, W.; Christie, P.; Luo, Y. Surfactant-enhanced bioremediation of petroleum-contaminated soil and microbial community response: A field study. Chemosphere 2023, 322, 138225. [Google Scholar] [CrossRef]
- Malik, G.; Arora, R.; Chaturvedi, R.; Paul, M.S. Implementation of Genetic Engineering and Novel Omics Approaches to Enhance Bioremediation: A Focused Review. Bull. Environ. Contam. Toxicol. 2021, 108, 443–450. [Google Scholar] [CrossRef]
- Shyam, K.; Kumar, N.; Chandel, H.; Dogra, A.S.; Sharma, G.; Saxena, G. Omics Technologies in Environmental Microbiology and Microbial Ecology: Insightful Applications in Bioremediation Research. Wiley Online Library, Genomics Approach to Bioremediation: Principles, Tools, and Emerging Technologies 2023, 433–454. [Google Scholar] [CrossRef]
- Wesseler, J.; Kleter, G.; Meulenbroek, M.; Purnhagen, K.P. EU regulation of genetically modified microorganisms in light of new policy developments: Possible implications for EU bioeconomy investments. Appl. Econ. Perspect. Policy 2022, 45, 839–859. [Google Scholar] [CrossRef]
- Directive 2009/41/EC, European Parliament 2009. Source: https://www.fao.org/faolex/results/details/en/c/LEX-FAOC138068/.
- Perera, I.C.; Hemamali, E.H. Genetically Modified Organisms for Bioremediation: Current Research and Advancements. In Bioremediation of Environmental Pollutants; Suyal, D.C., Soni, R., Eds.; Springer: Cham, 2022. [Google Scholar] [CrossRef]
- Sharma, B.; Shukla, P. A comparative analysis of heavy metal bioaccumulation and functional gene annotation towards multiple metal resistant potential by Ochrobactrum intermedium BPS-20 and Ochrobactrum ciceri BPS-26. Bioresour. Technol. 2020, 320, 124330. [Google Scholar] [CrossRef]
- Rafeeq, H.; Afsheen, N.; Rafique, S.; Arshad, A.; Intisar, M.; Hussain, A.; Bilal, M.; Iqbal, H.M. Genetically engineered microorganisms for environmental remediation. Chemosphere 2022, 310, 136751. [Google Scholar] [CrossRef]
- Tirkey, S.R.; Ram, S.; Chandrashekhar, P.; Mishra, S. Bioremediation of Soils Polluted with Hexavalent Chromium Using Bacteria. In Industrial Wastewater Reuse; Shah, M.P., Ed.; Springer: Singapore, 2023. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Han, F.; Liu, Z.; Zhao, C.; Lei, J.; Zhou, W. Enhanced degradation of petroleum in saline soil by nitrogen stimulation and halophilic emulsifying bacteria Bacillus sp. Z-13. J. Hazard. Mater. 2023, 459, 132102. [Google Scholar] [CrossRef] [PubMed]
- Elmore, J.R.; Dexter, G.N.; Baldino, H.; Huenemann, J.D.; Francis, R.; Peabody, G.L.; Martinez-Baird, J.; Riley, L.A.; Simmons, T.; Coleman-Derr, D.; et al. High-throughput genetic engineering of nonmodel and undomesticated bacteria via iterative site-specific genome integration. Sci. Adv. 2023, 9, eade1285. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P., Kandeler. Enzyme activities and microbiological and biochemical processes in soil. Enzymes in the Environment. CRC Press: Boca Raton, FL, USA, 2002; pp. 1–33. [Google Scholar]
- Khalid, F.; Hashmi, M.Z.; Jamil, N.; Qadir, A.; Ali, M.I. Microbial and enzymatic degradation of PCBs from e-waste-contaminated sites: a review. Environ. Sci. Pollut. Res. 2021, 28, 10474–10487. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Tripathi, M.; Srinath, T. Strategies for chromium bioremediation of tannery effluent. Reviews of Environmental Contamination and Toxicology 2012, 217, 75–140. [Google Scholar] [CrossRef] [PubMed]
- Abatenh, E.; Gizaw, B.; Tsegaye, Z.; Wassie, M. The Role of Microorganisms in Bioremediation- A Review. 2017, 2, 38–46. [Google Scholar] [CrossRef]
- Alvarez, A.; Saez, J.M.; Costa, J.S.D.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Haghollahi, A.; Fazaelipoor, M.H.; Schaffie, M. The effect of soil type on the bioremediation of petroleum contaminated soils. J. Environ. Manag. 2016, 180, 197–201. [Google Scholar] [CrossRef]
- da Silva, I.G.S.; de Almeida, F.C.G.; Silva, N.M.P.d.R.e.; Casazza, A.A.; Converti, A.; Sarubbo, L.A. Soil Bioremediation: Overview of Technologies and Trends. Energies 2020, 13, 4664. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Ng, Z.Y.; Ajeng, A.A.; Cheah, W.Y.; Ng, E.-P.; Abdullah, R.; Ling, T.C. Towards circular economy: Potential of microalgae – bacterial-based biofertilizer on plants. J. Environ. Manag. 2023, 349, 119445. [Google Scholar] [CrossRef]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wang, J.; Deng, Y.; Liu, Y.; Peng, B. The potential impact on the biodegradation of organic pollutants from composting technology for soil remediation. Waste Manag. 2017, 72, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Singh, S.P.; Parakh, S.K.; Tong, Y.W. Health hazards of hexavalent chromium (Cr (VI)) and its microbial reduction. Bioengineered 2022, 13, 4923–4938. [Google Scholar] [CrossRef] [PubMed]
- Giovanella, P.; Vieira, G.A.; Otero, I.V.R.; Pellizzer, E.P.; Fontes, B.d.J.; Sette, L.D. Metal and organic pollutants bioremediation by extremophile microorganisms. J. Hazard. Mater. 2019, 382, 121024. [Google Scholar] [CrossRef]
- Hualpa-Cutipa E., Acosta R. A. S., Cariga O. J. M., Espinoza-Medina M. A., Hansen-Reyes M., Medina-Cerna D., et al. “Omics Insights into Cold Environments: Cold-Tolerant Microorganisms and their Potential Use in Bioremediation” in Omics Insights in Environmental Bioremediation. eds. Kumar V., Thakur I. S. (Singapore: Springer Nature Singapore;) 2022, 437–453.
- Chaturvedi, R. Extremophiles: Biofactories for Bioremediation. In Integrative Approaches to Biotechnology; CRC Press, 2024; pp. 171–185. ISBN 9781003324706. [Google Scholar]
- Abdelaziz, S.; Belal, E.E.; Al-Quwaie, D.A.; Ashkan, M.F.; Alqahtani, F.S.; El-Tarabily, K.A.; El-Mageed, T.A.A.; Shami, A.; Nader, M.M.; Hemeda, N.F. Extremophilic bacterial strains as plant growth promoters and biocontrol agents against Pythium ultimum and Rhizocotnia solani. J. Plant Pathol. 2023, 105, 1347–1369. [Google Scholar] [CrossRef]
- Santos, A.P.; Belfiore, C.; Úrbez, C.; Ferrando, A.; Blázquez, M.A.; Farías, M.E. Extremophiles as Plant Probiotics to Promote Germination and Alleviate Salt Stress in Soybean. J. Plant Growth Regul. 2022, 42, 946–959. [Google Scholar] [CrossRef]
- Stavi, I.; Thevs, N.; Priori, S. Soil Salinity and Sodicity in Drylands: A Review of Causes, Effects, Monitoring, and Restoration Measures. Front. Environ. Sci. 2021, 9. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Garcia-Caparrós, P.; Nogales, A.; Abreu, M.M.; Santos, E.; Cortinhas, A.L.; Caperta, A.D. Sustainable agricultural management of saline soils in arid and semi-arid Mediterranean regions through halophytes, microbial and soil-based technologies. Environ. Exp. Bot. 2023, 212. [Google Scholar] [CrossRef]
- Atai, E.; Jumbo, R.B.; Cowley, T.; Azuazu, I.; Coulon, F.; Pawlett, M. Efficacy of bioadmendments in reducing the influence of salinity on the bioremediation of oil-contaminated soil. Sci. Total. Environ. 2023, 892, 164720. [Google Scholar] [CrossRef]
- Rhykerd, R.L.; Weaver, R.W.; McInnes, K.J. Influence of salinity on bioremediation of oil in soil. Environ. Pollut. 1995, 90, 127–130. [Google Scholar] [CrossRef]
- Qin, X.; Tang, J.; Li, D.; Zhang, Q. Effect of salinity on the bioremediation of petroleum hydrocarbons in a saline-alkaline soil. Lett. Appl. Microbiol. 2012, 55, 210–217. [Google Scholar] [CrossRef]
- Kansour, M.K.; Al-Mailem, D.M. Bioremediation of two oil-contaminated Kuwaiti hyper-saline soils by cross bioaugmentation and the role of indigenous halophilic/halotolerant hydrocarbonoclastic bacteria. Environ. Technol. Innov. 2023, 32. [Google Scholar] [CrossRef]
- Quoc, K.N.; Le Vinh, T.; Le Thanh, Q.; Ngoc, H.T.; Thi, X.D.; Huu, D.H.; Thanh, X.L.N.; My, T.L.T. Effects of biofertilizer supplementation, Rhodopseudomonas spp., on nitrogen and phosphorus uptakes, growth, and yield of sesame ( Sesamum indicum L.) on salt-affected soil. J. Plant Nutr. 2023, 47, 1–17. [Google Scholar] [CrossRef]
- Hadibarata, T.; Kristanti, R.A.; Bilal, M.; Yilmaz, M.; Sathishkumar, P. Biodegradation mechanism of chlorpyrifos by halophilic bacterium Hortaea sp. B15. Chemosphere 2022, 312, 137260. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Sivaperumal, P. Marine halophilic archaeal isolate Halococcus sp. with emphasis on bioremediation of radionuclides through extracellular biomolecules. J. Chem. Technol. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Iqbal, S., Hasni, M.S., Malik, Y., Baloch, S. and Aamir, R. Halophilic Fungi: Versatile Microorganisms for Biotechnological Advancements 2023. [CrossRef]
- Martínez-Espinosa, R.M.; Kumar, S.; Upadhyay, S.K.; Orhan, F. Editorial: Adaptation of halophilic/halotolerant microorganisms and their applications. Front. Microbiol. 2023, 14, 1252921. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Ma, Y.; Wang, X.; Zhang, B.; Zhang, G.; Bahadur, A.; Chen, T.; Liu, G.; Zhang, W.; et al. Research progress regarding the role of halophilic and halotolerant microorganisms in the eco-environmental sustainability and conservation. J. Clean. Prod. 2023, 418. [Google Scholar] [CrossRef]
- Hualpa-Cutipa, E., Acosta, R.A.S., Cariga, O.J.M., Espinoza-Medina, M.A., Hansen-Reyes, M., Medina-Cerna, D., Olanda, M.C., Cortez-Lázaro, A.A. Omics Insights into Cold Environments: Cold-Tolerant Microorganisms and their Potential Use in Bioremediation. In Omics Insights in Environmental Bioremediation 2022. (pp. 437-453). Singapore: Springer Nature Singapore. [CrossRef]
- Barghoth, M.G.; Desouky, S.E.; Radwan, A.A.; Shah, M.P.; Salem, S.S. Characterizations of highly efficient moderately halophilic toluene degrading exiguobacterium mexicanum M7 strain isolated from Egyptian saline sediments. Biotechnol. Genet. Eng. Rev. 2023, 1–19. [Google Scholar] [CrossRef]
- Kansour, M.K.; Al-Mailem, D.M. Bioremediation of two oil-contaminated Kuwaiti hyper-saline soils by cross bioaugmentation and the role of indigenous halophilic/halotolerant hydrocarbonoclastic bacteria. Environ. Technol. Innov. 2023, 32. [Google Scholar] [CrossRef]
- Rajkumar, R., Kurinjimalar, C. Microbiological Activity for Soil and Plant Health Management. Springer; Singapore. Microbes and Plant Mineral Nutrition 2021, 111–132. [CrossRef]
- Radhakrishnan, A.; Balaganesh, P.; Vasudevan, M.; Natarajan, N.; Chauhan, A.; Arora, J.; Ranjan, A.; Rajput, V.D.; Sushkova, S.; Minkina, T.; et al. Bioremediation of Hydrocarbon Pollutants: Recent Promising Sustainable Approaches, Scope, and Challenges. Sustainability 2023, 15, 5847. [Google Scholar] [CrossRef]
- Jalali, F.M.; Chahkandi, B.; Gheibi, M.; Eftekhari, M.; Behzadian, K.; Campos, L.C. Developing a smart and clean technology for bioremediation of antibiotic contamination in arable lands. Sustain. Chem. Pharm. 2023, 33. [Google Scholar] [CrossRef]
- Hwangbo, M.; Shao, Y.; Hatzinger, P.B.; Chu, K. Acidophilic methanotrophs: Occurrence, diversity, and possible bioremediation applications. Environ. Microbiol. Rep. 2023, 15, 265–281. [Google Scholar] [CrossRef] [PubMed]
- Quoc, B.N.; Armenta, M.; Carter, J.A.; Bucher, R.; Sukapanpotharam, P.; Bryson, S.J.; Stahl, D.A.; Stensel, H.D.; Winkler, M.-K.H. An investigation into the optimal granular sludge size for simultaneous nitrogen and phosphate removal. Water Res. 2021, 198, 117119. [Google Scholar] [CrossRef] [PubMed]
- Alegbeleye, O.O.; Opeolu, B.O.; Jackson, V.A. Polycyclic Aromatic Hydrocarbons: A Critical Review of Environmental Occurrence and Bioremediation. Environ. Manag. 2017, 60, 758–783. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.K.; Kumar, M.; Singh, D.K. Insight into the amoxicillin resistance, ecotoxicity, and remediation strategies. J. Water Process. Eng. 2020, 39, 101858. [Google Scholar] [CrossRef]
- Abtahi, H.; Parhamfar, M.; Saeedi, R.; Villaseñor, J.; Sartaj, M.; Kumar, V.; Coulon, F.; Parhamfar, M.; Didehdar, M.; Seifi, H.; et al. Effect of competition between petroleum-degrading bacteria and indigenous compost microorganisms on the efficiency of petroleum sludge bioremediation: Field application of mineral-based culture in the composting process. J. Environ. Manag. 2020, 258, 110013. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of Heavy Metals in Soil: Impact on Microbial Biodegradation of Organic Compounds and Possible Improvement Strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef]
- Roane, T.M.; Josephson, K.L.; Pepper, I.L. Dual-Bioaugmentation Strategy To Enhance Remediation of Cocontaminated Soil. Appl. Environ. Microbiol. 2001, 67, 3208–3215. [Google Scholar] [CrossRef]
- Wani, P.A.; Rafi, N.; Wani, U.; A, H.B.; Khan, M.S.A. Simultaneous bioremediation of heavy metals and biodegradation of hydrocarbons by metal resistantBrevibacillus parabrevisOZF5 improves plant growth promotion. Bioremediation J. 2021, 27, 20–31. [Google Scholar] [CrossRef]
- Sar, P.; Kundu, S.; Ghosh, A.; Saha, B. Natural surfactant mediated bioremediation approaches for contaminated soil. RSC Adv. 2023, 13, 30586–30605. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of Heavy Metals in Soil: Impact on Microbial Biodegradation of Organic Compounds and Possible Improvement Strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef]
- Roller, B.R.K.; Schmidt, T.M. The physiology and ecological implications of efficient growth. ISME J. 2015, 9, 1481–1487. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Neidhardt, F.C. Bacterial Growth: Constant Obsession with dN/dt. J. Bacteriol. 1999, 181, 7405–8. [Google Scholar] [CrossRef] [PubMed]
- Moqsud, M.; Yoshitake, J.; Bushra, Q.; Hyodo, M.; Omine, K.; Strik, D. Compost in plant microbial fuel cell for bioelectricity generation. Waste Manag. 2015, 36, 63–69. [Google Scholar] [CrossRef]
- Irfan, S.; Ranjha, M.M.A.N.; Shafique, B.; Ullah, M.I.; Siddiqui, A.R.; Wang, L. Bioremediation of Soil: An Overview. In Advances in Bioremediation and Phytoremediation for Sustainable Soil Management; Malik, J.A., Ed.; Springer: Cham, 2022. [Google Scholar] [CrossRef]
- Narancic, T.; O'Connor, K.E. Microbial biotechnology addressing the plastic waste disaster. Microb. Biotechnol. 2017, 10, 1232–1235. [Google Scholar] [CrossRef]
- Ji, M.; Zhu, J.; Zhu, R.; Chen, J.; Wang, C. Experiments on the Effects of Dissolved Oxygen on the Free and Immobilization Effective Microorganisms (EM) in Treating Polluted River Water. International Conference on Material and Environmental Engineering (ICMAEE 2014). LOCATION OF CONFERENCE, ChinaDATE OF CONFERENCE;
- Wang, B.; Kuang, S.; Shao, H.; Wang, L.; Wang, H. Anaerobic-petroleum degrading bacteria: Diversity and biotechnological applications for improving coastal soil. Ecotoxicol. Environ. Saf. 2021, 224, 112646. [Google Scholar] [CrossRef]
- Chen, W.-Y.; Wu, J.-H.; Wang, B.N. Intermittent Oxygen Supply Facilitates Codegradation of Trichloroethene and Toluene by Anaerobic Consortia. Environ. Sci. Technol. 2023, 57, 10252–10262. [Google Scholar] [CrossRef]
- Bhuyan, P.P.; Nayak, R.; Jena, M.; Pradhan, B. Convoluted role of cyanobacteria as biofertilizer: an insight of sustainable agriculture. Vegetos 2022, 36, 309–321. [Google Scholar] [CrossRef]
- Jangir, C.K.; Kumar, S.; Meena, R.S. Sustainable agriculture. Scientific Publisher; Jodhpur, India. Significance of soil organic matter to soil quality and evaluation of sustainability, 2019, 357–381.
- Bauder, D. L. Bioremediation of Contaminated Soil. Retrieved February 2006, 7, 2014 https://bioremediationofcontaminatedsoilwordpresscom/. [Google Scholar]
- Gao, H.; Wu, M.; Liu, H.; Ou, Y.; Zhang, T.; Duan, X. Unraveling the Positive Effect of Soil Moisture on the Bioaugmentation of Petroleum-Contaminated Soil Using Bioinformatics. Microb. Ecol. 2023, 1–11. [Google Scholar] [CrossRef]
- Naorem, A. Will Climate Change Alter the Efficiency of Bioremediation?. In: Malik, J.A. (eds) Advances in Bioremediation and Phytoremediation for Sustainable Soil Management. Springer, Cham. 2022. [CrossRef]
- Bahmani, F. , Ataei, S. A., Mikaili, M.A. The Effect of Moisture Content Variation on the Bioremediation of Hydrocarbon Contaminated Soils: Modeling and Experimental Investigation. Journal of Environmental Analytical Chemistry 2018, 5, 236. [Google Scholar] [CrossRef]
- Kumari, S.; Gautam, K.; Seth, M.; Anbumani, S.; Manickam, N. Bioremediation of polycyclic aromatic hydrocarbons in crude oil by bacterial consortium in soil amended with Eisenia fetida and rhamnolipid. Environ. Sci. Pollut. Res. 2023, 30, 82517–82531. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Wu, R.; Peng, T.; Li, Q.; Wang, Q.; Wu, Y.; Song, X.; Lin, X. Low-temperature thermally enhanced bioremediation of polycyclic aromatic hydrocarbon-contaminated soil: Effects on fate, toxicity and bacterial communities. Environ. Pollut. 2023, 335, 122247. [Google Scholar] [CrossRef]
- Norozpour, M.; Sarikhani, M.R.; Aliasgharzad, N. Monitoring of soil respiration changes in a heavy naphtha-contaminated sandy loam soil under different bioremediation treatments. Water and Soil Science 2023, 33, 53–72. [Google Scholar] [CrossRef]
- Peekate, L.P.; Diepiriye, A.D.; Amadi, M.C. Effect of glucose on biodegradation of hydrocarbons in crude-oil polluted soil undergoing bioremediation. Asia J. Appl. Microbiol. 2023, 10, 61–69. [Google Scholar] [CrossRef]
- Mupambwa, H.A.; Mnkeni, P.N.S. Optimizing the vermicomposting of organic wastes amended with inorganic materials for production of nutrient-rich organic fertilizers: a review. Environ. Sci. Pollut. Res. 2018, 25, 10577–10595. [Google Scholar] [CrossRef]
- Kebede, G.; Tafese, T.; Abda, E.M.; Kamaraj, M.; Assefa, F. Factors Influencing the Bacterial Bioremediation of Hydrocarbon Contaminants in the Soil: Mechanisms and Impacts. J. Chem. 2021, 2021, 1–17. [Google Scholar] [CrossRef]
- Cavazzoli, S.; Squartini, A.; Sinkkonen, A.; Romantschuk, M.; Rantalainen, A.-L.; Selonen, V.; Roslund, M.I. Nutritional additives dominance in driving the bacterial communities succession and bioremediation of hydrocarbon and heavy metal contaminated soil microcosms. Microbiol. Res. 2023, 270, 127343. [Google Scholar] [CrossRef]
- Bala, S.; Garg, D.; Thirumalesh, B.V.; Sharma, M.; Sridhar, K.; Inbaraj, B.S.; Tripathi, M. Recent Strategies for Bioremediation of Emerging Pollutants: A Review for a Green and Sustainable Environment. Toxics 2022, 10, 484. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M. Microbial bioremediation as a robust process to mitigate pollutants of environmental concern. Case Stud. Chem. Environ. Eng. 2020, 2, 100011. [Google Scholar] [CrossRef]
- Sharma, B.; Shukla, P. Futuristic avenues of metabolic engineering techniques in bioremediation. Biotechnol. Appl. Biochem. 2020, 69, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Basak, B.; Dey, A. Bioremediation Approaches for Recalcitrant Pollutants: Potentiality, Successes and Limitation. 2015. [CrossRef]
- Li, Q.; Liu, J.; Gadd, G.M. Fungal bioremediation of soil co-contaminated with petroleum hydrocarbons and toxic metals. Appl. Microbiol. Biotechnol. 2020, 104, 8999–9008. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Biswas, R.; Sarkar, A. Advancement of Omics: Prospects for Bioremediation of Contaminated Soils. In Microbial Bioremediation & Biodegradation; Shah, M., Ed.; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Rathore, S.; Varshney, A.; Mohan, S.; Dahiya, P. An innovative approach of bioremediation in enzymatic degradation of xenobiotics. Biotechnol. Genet. Eng. Rev. 2022, 38, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, S.; Dong, Y.; Fan, H.; Bai, Z.; Zhuang, X. Engineering Microbial Consortia towards Bioremediation. Water 2021, 13, 2928. [Google Scholar] [CrossRef]
- Mali, H.; Shah, C.; Raghunandan, B.; Prajapati, A.S.; Patel, D.H.; Trivedi, U.; Subramanian, R. Organophosphate pesticides an emerging environmental contaminant: Pollution, toxicity, bioremediation progress, and remaining challenges. J. Environ. Sci. 2022, 127, 234–250. [Google Scholar] [CrossRef]
- Khatoon, N.; Jamal, A.; Ali, M.I. Polymeric pollutant biodegradation through microbial oxidoreductase: A better strategy to safe environment. Int. J. Biol. Macromol. 2017, 105, 9–16. [Google Scholar] [CrossRef]
- Peter, J.K.; Singh, R.; Yadav, A.K.; Kothari, R.; Mehta, P.K. Toxicity of nitriles/amides-based products in the environment and their enzymatic bioremediation. J. Hazard. Mater. Adv. 2023, 13. [Google Scholar] [CrossRef]
- Wang, L.; Du, X.; Li, Y.; Bai, Y.; Tang, T.; Wu, J.; Liang, H.; Gao, D. Enzyme immobilization as a sustainable approach toward ecological remediation of organic-contaminated soils: Advances, issues, and future perspectives. Crit. Rev. Environ. Sci. Technol. 2023, 53, 1684–1708. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, F.; Guo, S. Highly efficient screening and optimal combination of functional isolates for bioremediation of hydrocarbon-polluted soil. Environ. Res. 2022, 219, 115064. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.-P. Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Newhouse, J.R. Blight C Sci. Am. 1990, 263, 106–111. Available online: http://www.jstor.org/stable/24996872. [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- IPBES Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. IPBES secretariat 2019, 1–1148. [CrossRef]
- van Kleunen, M.; Dawson, W.; Essl, F.; Pergl, J.; Winter, M.; Weber, E.; Kreft, H.; Weigelt, P.; Kartesz, J.; Nishino, M.; et al. Global exchange and accumulation of non-native plants. Nature 2015, 525, 100–103. [Google Scholar] [CrossRef]
- van Kleunen, M.; Pyšek, P.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Weigelt, P.; Stein, A.; Dullinger, S.; König, C.; et al. The Global Naturalized Alien Flora (GloNAF) database. Ecology 2018, 100, e02542. [Google Scholar] [CrossRef]
- Pyšek, P.; Pergl, J.; Essl, F.; Lenzner, B.; Dawson, W.; Kreft, H.; Weigelt, P.; Winter, M.; Kartesz, J.; Nishino, M.; et al. Naturalized alien flora of the world. Preslia 2017, 89, 203–274. [Google Scholar] [CrossRef]
- Seebens, H.; Essl, F.; Dawson, W.; Fuentes, N.; Moser, D.; Pergl, J.; Pyšek, P.; van Kleunen, M.; Weber, E.; Winter, M.; et al. Global trade will accelerate plant invasions in emerging economies under climate change. Glob. Chang. Biol. 2015, 21, 4128–4140. [Google Scholar] [CrossRef]
- Glick, B.R. Beneficial Plant-Bacterial Interactions. Springer Science and Business Media LLC, 2020; p. 383. [Google Scholar]
- Mawarda, P.C.; Le Roux, X.; van Elsas, J.D.; Salles, J.F. Deliberate introduction of invisible invaders: A critical appraisal of the impact of microbial inoculants on soil microbial communities. Soil Biol. Biochem. 2020, 148, 107874. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.-T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2021, 20, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.R. Horizontal Gene Transfer: Figure 1. Evol. Med. Public Heal. 2015, 2015, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.; Olesen, A.K.; Maccario, L.; Leal, L.H.; Maas, P.V.D.; Heederik, D.; Mevius, D.; Sørensen, S.J.; Schmitt, H. Horizontal Gene Transfer of an IncP1 Plasmid to Soil Bacterial Community Introduced by Escherichia coli through Manure Amendment in Soil Microcosms. Environ. Sci. Technol. 2022, 56, 11398–11408. [Google Scholar] [CrossRef]
- Ronchel, M.C.; Ramos-Díaz, M.A.; Ramos, J.L. Retrotransfer of DNA in the rhizosphere. Environ. Microbiol. 2000, 2, 319–323. [Google Scholar] [CrossRef]
- Martínez-Carranza, E.; García-Reyes, S.; González-Valdez, A.; Soberón-Chávez, G. Tracking the genome of four Pseudomonas aeruginosa isolates that have a defective Las quorum-sensing system, but are still virulent. Access Microbiol. 2020, 2, e000132. [Google Scholar] [CrossRef]
- European Regulation (EU) 2019/1009 soyce: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=PI_COM:Ares(2021)898281.
- Dastogeer, K.M.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant microbiome–an account of the factors that shape community composition and diversity. Curr. Plant Biol. 2020, 23, 100161. [Google Scholar] [CrossRef]
- Vacheron, J., Renoud, S., Muller, D., Babalola, O. O., Prigent-Combaret, C “Handbook for Azospirillum. Technical Issues and Protocols. Everything practical you always wanted to know about Azospirillum sp. but were afraid to ask,” in Alleviation of Abiotic and Biotic Stresses in Plants by Azospirillum, eds F. D. Cassan, Y. Okon, M. Cecilia, and M. C. Creus (Cham: Springer) 2015. 333–365.
- Jones, D.L. Organic acids in the rhizosphere – a critical review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Hart, M.M.; Antunes, P.M.; Chaudhary, V.B.; Abbott, L.K. Fungal inoculants in the field: Is the reward greater than the risk? Funct. Ecol. 2018, 32, 126–135. [Google Scholar] [CrossRef]
- Keswani, C.; Prakash, O.; Bharti, N.; Vílchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total. Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, Y.; Liu, P.; Xu, R.; Rensing, C.; Fu, X.; Liao, H. Genotype and rhizobium inoculation modulate the assembly of soybean rhizobacterial communities. Plant, Cell Environ. 2019, 42, 2028–2044. [Google Scholar] [CrossRef] [PubMed]
- Schwieger, F.; Tebbe, C.C. Effect of Field Inoculation with Sinorhizobium meliloti L33 on the Composition of Bacterial Communities in Rhizospheres of a Target Plant ( Medicago sativa ) and a Non-Target Plant ( Chenopodium album )—Linking of 16S rRNA Gene-Based Single-Strand Conformation Polymorphism Community Profiles to the Diversity of Cultivated Bacteria. Appl. Environ. Microbiol. 2000, 66, 3556–3565. [Google Scholar] [CrossRef] [PubMed]
- Horácio, E.H.; Zucareli, C.; Gavilanes, F.Z.; Yunes, J.S.; dos Santos Sanzovo, A.W.; Andrade, D.S. Co-inoculation of rhizobia, azospirilla and cyanobacteria for increasing common bean production. Londrina 2020, 41, 2015–2028. [Google Scholar] [CrossRef]
- Trabelsi, D.; Mhamdi, R. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Res. Int. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Harman, G.E.; Uphoff, N. Symbiotic Root-Endophytic Soil Microbes Improve Crop Productivity and Provide Environmental Benefits. Scientifica 2019, 2019, 1–25. [Google Scholar] [CrossRef]
- Baudoin, E.; Nazaret, S.; Mougel, C.; Ranjard, L.; Moënne-Loccoz, Y. Impact of inoculation with the phytostimulatory PGPR Azospirillum lipoferum CRT1 on the genetic structure of the rhizobacterial community of field-grown maize. Soil Biol. Biochem. 2009, 41, 409–413. [Google Scholar] [CrossRef]
- Herschkovitz, Y.; Lerner, A.; Davidov, Y.; Okon, Y.; Jurkevitch, E. Azospirillum brasilense does not affect population structure of specific rhizobacterial communities of inoculated maize (Zea mays). Environ. Microbiol. 2005, 7, 1847–1852. [Google Scholar] [CrossRef]
- Lerner, A.; Herschkovitz, Y.; Baudoin, E.; Nazaret, S.; Moenne-Loccoz, Y.; Okon, Y.; Jurkevitch, E. Effect of Azospirillum brasilense inoculation on rhizobacterial communities analyzed by denaturing gradient gel electrophoresis and automated ribosomal intergenic spacer analysis. Soil Biol. Biochem. 2006, 38, 1212–1218. [Google Scholar] [CrossRef]
- Correa, O.; Romero, A.; Montecchia, M.; Soria, M. Tomato genotype and Azospirillum inoculation modulate the changes in bacterial communities associated with roots and leaves. J. Appl. Microbiol. 2007, 102, 781–786. [Google Scholar] [CrossRef]
- Andreote, F.D.; de Araújo, W.L.; de Azevedo, J.L.; van Elsas, J.D.; da Rocha, U.N.; van Overbeek, L.S. Endophytic Colonization of Potato ( Solanum tuberosum L.) by a Novel Competent Bacterial Endophyte, Pseudomonas putida Strain P9, and Its Effect on Associated Bacterial Communities. Appl. Environ. Microbiol. 2009, 75, 3396–3406. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Yin, D.; Chen, S.; Xia, F.; Yang, J.; Li, Q.; Wang, W. Effect of Biocontrol Agent Pseudomonas fluorescens 2P24 on Soil Fungal Community in Cucumber Rhizosphere Using T-RFLP and DGGE. PLOS ONE 2012, 7, e31806. [Google Scholar] [CrossRef] [PubMed]
- Basiru, S.; Mwanza, H.P.; Hijri, M. Analysis of Arbuscular Mycorrhizal Fungal Inoculant Benchmarks. Microorganisms 2020, 9, 81. [Google Scholar] [CrossRef]
- Koch, A.M.; Antunes, P.M.; Barto, E.K.; Cipollini, D.; Mummey, D.L.; Klironomos, J.N. The effects of arbuscular mycorrhizal (AM) fungal and garlic mustard introductions on native AM fungal diversity. Biol. Invasions 2010, 13, 1627–1639. [Google Scholar] [CrossRef]
- Jin, H.; Germida, J.J.; Walley, F.L. Impact of arbuscular mycorrhizal fungal inoculants on subsequent arbuscular mycorrhizal fungi colonization in pot-cultured field pea (Pisum sativum L.). Mycorrhiza 2012, 23, 45–59. [Google Scholar] [CrossRef]
- Antunes, P.M.; Koch, A.M.; Dunfield, K.E.; Hart, M.M.; Downing, A.; Rillig, M.C.; Klironomos, J.N. Influence of commercial inoculation with Glomus intraradices on the structure and functioning of an AM fungal community from an agricultural site. Plant Soil 2008, 317, 257–266. [Google Scholar] [CrossRef]
- Jin, H.; Germida, J.J.; Walley, F.L. Impact of arbuscular mycorrhizal fungal inoculants on subsequent arbuscular mycorrhizal fungi colonization in pot-cultured field pea (Pisum sativum L.). Mycorrhiza 2012, 23, 45–59. [Google Scholar] [CrossRef]
- Marschner, P.; Timonen, S. Interactions between plant species and mycorrhizal colonization on the bacterial community composition in the rhizosphere. Appl. Soil Ecol. 2005, 28, 23–36. [Google Scholar] [CrossRef]
- Cavagnaro, T.R.; Jackson, L.E.; Six, J.; Ferris, H.; Goyal, S.; Asami, D.; Scow, K.M. Arbuscular Mycorrhizas, Microbial Communities, Nutrient Availability, and Soil Aggregates in Organic Tomato Production. Plant Soil 2006, 282, 209–225. [Google Scholar] [CrossRef]
- Monokrousos, N.; Papatheodorou, E.M.; Orfanoudakis, M.; Jones, D.G.; Scullion, J.; Stamou, G.P. The effects of plant type, AMF inoculation and water regime on rhizosphere microbial communities. Eur. J. Soil Sci. 2019, 71, 265–278. [Google Scholar] [CrossRef]
- Salilh, F.Z., Tarekegn, M.M. Cleaning polluted environments by bioremediation: Review. IJESC 2020, 10(8),27112–21. Available at: https://ijesc.org/upload/36172711a56684478405d1ee977ff36a.Cleaning%20Polluted%20Environments%20by%20Bioremediation-Review.pdf.
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria Associated With a Commercial Mycorrhizal Inoculum: Community Composition and Multifunctional Activity as Assessed by Illumina Sequencing and Culture-Dependent Tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef] [PubMed]
- Casas, C.; Omacini, M.; Montecchia, M.S.; Correa, O.S. Soil microbial community responses to the fungal endophyte Neotyphodium in Italian ryegrass. Plant Soil 2010, 340, 347–355. [Google Scholar] [CrossRef]
- Rojas, X.; Guo, J.; Leff, J.W.; McNear, D.H.; Fierer, N.; McCulley, R.L. Infection with a Shoot-Specific Fungal Endophyte (Epichloë) Alters Tall Fescue Soil Microbial Communities. Microb. Ecol. 2016, 72, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, A.; de Souza, R.; Passaglia, L.M.P. Ecological role of bacterial inoculants and their potential impact on soil microbial diversity. Plant Soil 2015, 400, 193–207. [Google Scholar] [CrossRef]
- Di Salvo, L.P.; Ferrando, L.; Fernández-Scavino, A.; de Salamone, I.E.G. Microorganisms reveal what plants do not: wheat growth and rhizosphere microbial communities after Azospirillum brasilense inoculation and nitrogen fertilization under field conditions. Plant Soil 2018, 424, 405–417. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Schulz, W.; Scheu, S.; Jousset, A. Niche dimensionality links biodiversity and invasibility of microbial communities. Funct. Ecol. 2012, 27, 282–288. [Google Scholar] [CrossRef]
- Vázquez, M.M.; César, S.; Azcón, R.; Barea, J.M. Interactions between arbuscular mycorrhizal fungi and other microbial inoculants (Azospirillum, Pseudomonas, Trichoderma) and their effects on microbial population and enzyme activities in the rhizosphere of maize plants. Appl. Soil Ecol. 2000, 15, 261–272. [Google Scholar] [CrossRef]
- Asiloglu, R.; Shiroishi, K.; Suzuki, K.; Turgay, O.C.; Murase, J.; Harada, N. Protist-enhanced survival of a plant growth promoting rhizobacteria, Azospirillum sp. B510, and the growth of rice (Oryza sativa L.) plants. Appl. Soil Ecol. 2020, 154, 103599. [Google Scholar] [CrossRef]
- He, C.; Wang, W.; Hou, J. Plant Growth and Soil Microbial Impacts of Enhancing Licorice With Inoculating Dark Septate Endophytes Under Drought Stress. Front. Microbiol. 2019, 10, 2277. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and Challenges of Microbial Application for Crop Improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef]
- Hirsch, P.R. Release of transgenic bacterial inoculants - rhizobia as a case study. Plant Soil 2005, 266, 1–10. [Google Scholar] [CrossRef]
- Bellanger, X.; Guilloteau, H.; Bonot, S.; Merlin, C. Demonstrating plasmid-based horizontal gene transfer in complex environmental matrices: A practical approach for a critical review. Sci. Total. Environ. 2014, 493, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Shankar, K.; Mehendale, H.M. Oxidative Stress. In Encyclopedia of Toxicology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 735–737. ISBN 978-0-12-386455-0. [Google Scholar]
Figure 1.
Land use variation over the millennia. (Modified from: https://www.weforum.org/agenda/2021/02/ice-age-forest-lost-demand-agriculture/).
Figure 1.
Land use variation over the millennia. (Modified from: https://www.weforum.org/agenda/2021/02/ice-age-forest-lost-demand-agriculture/).
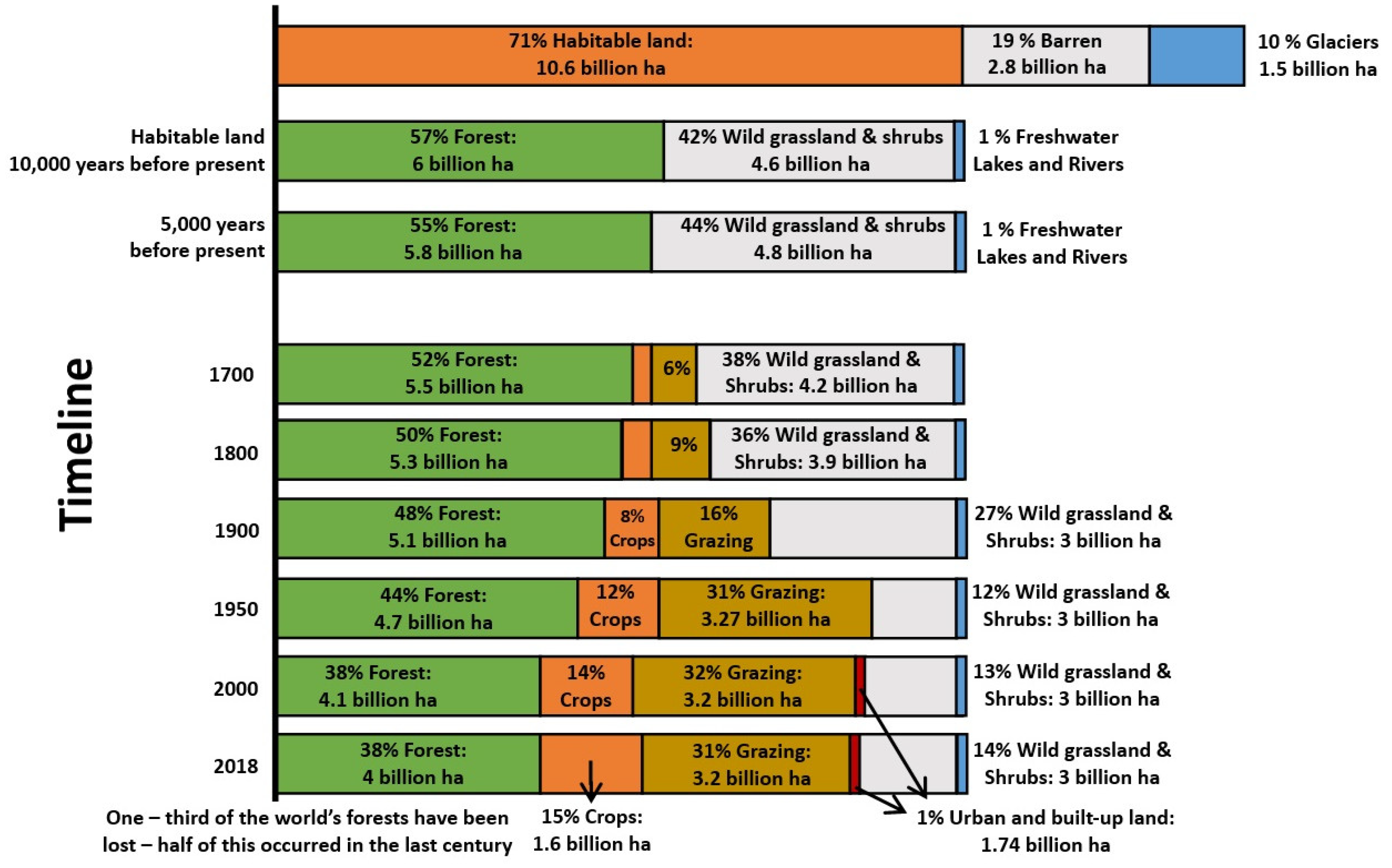
Figure 3.
Role of the rubisco molecule in plant primary production and cellular respiration. Modified from Gupta and Kim, 2015 [63].
Figure 3.
Role of the rubisco molecule in plant primary production and cellular respiration. Modified from Gupta and Kim, 2015 [63].
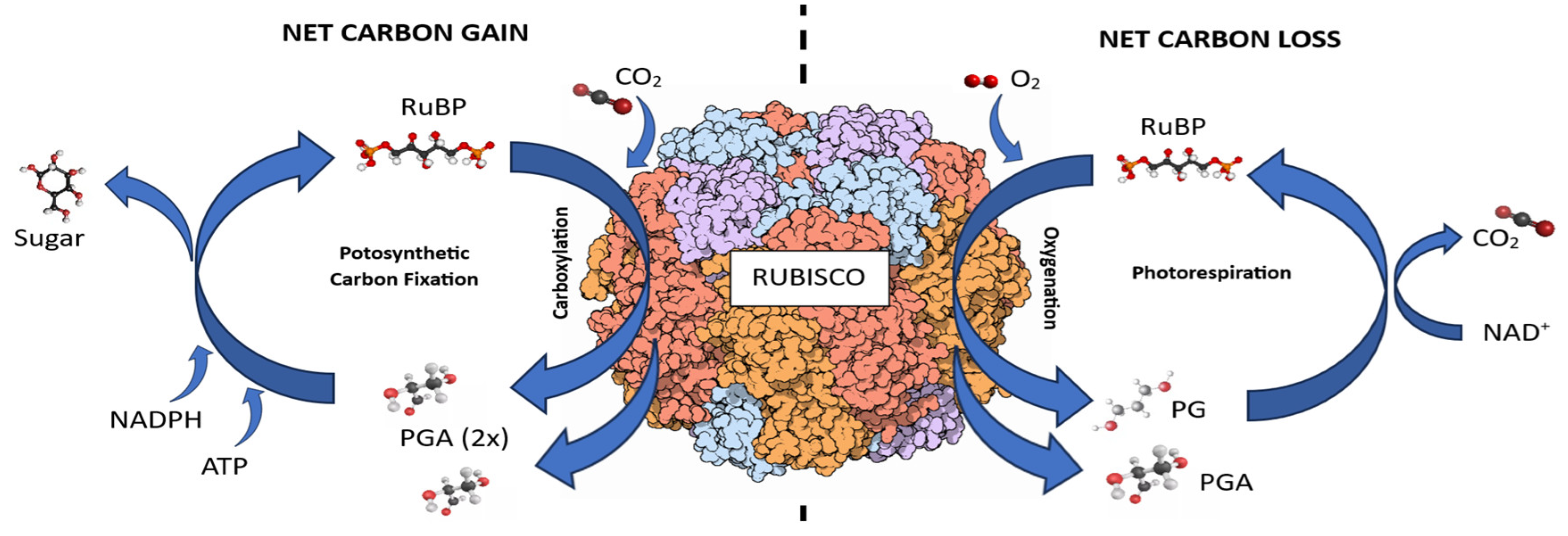
Figure 4.
An overview of the phosphorus cycle in nature (Modified from Shrivastava et al., 2018 [146]).
Figure 4.
An overview of the phosphorus cycle in nature (Modified from Shrivastava et al., 2018 [146]).
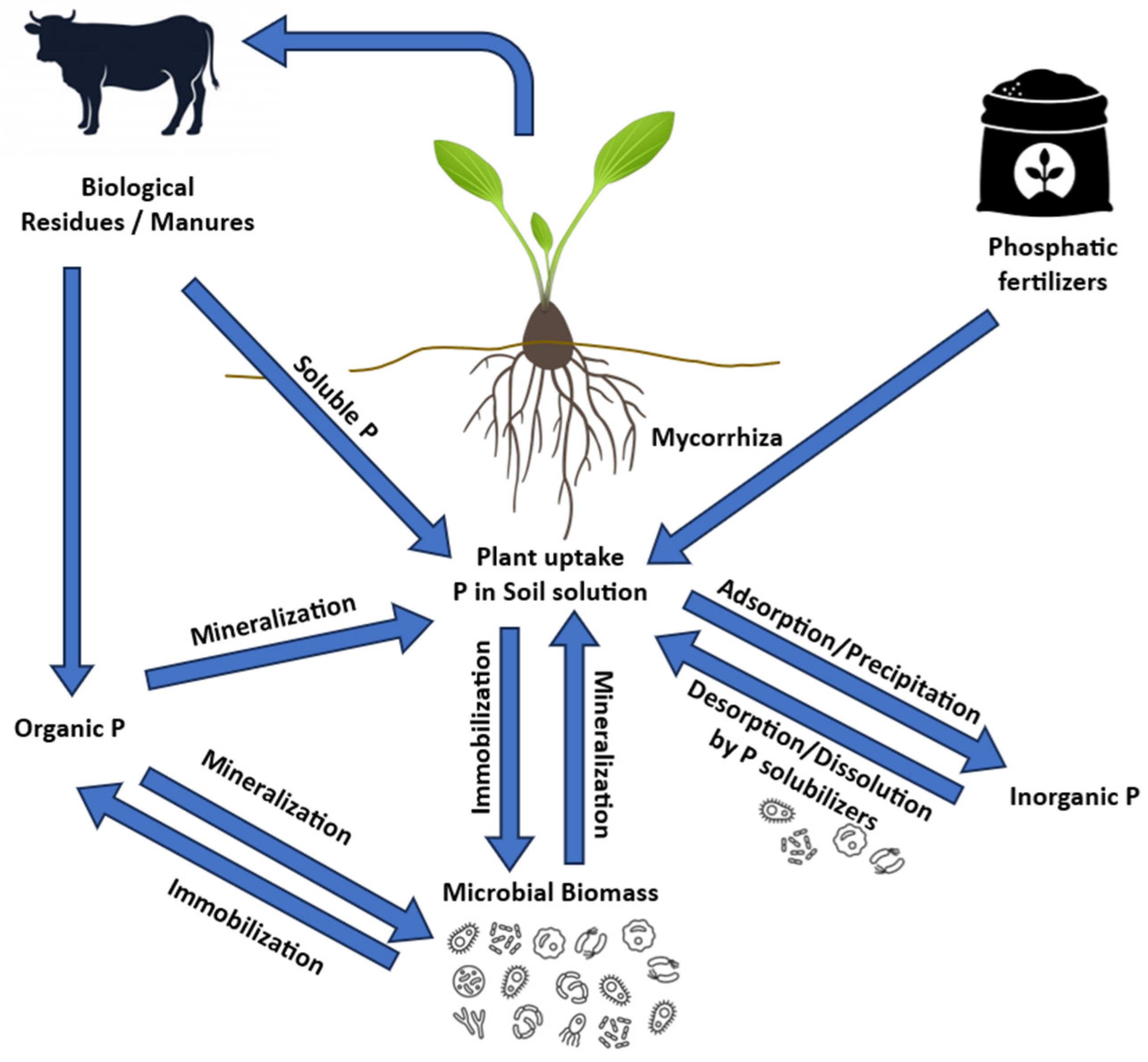
Figure 5.
Environmental Parameters Influencing Microbial Fertilization or Bioremediation.
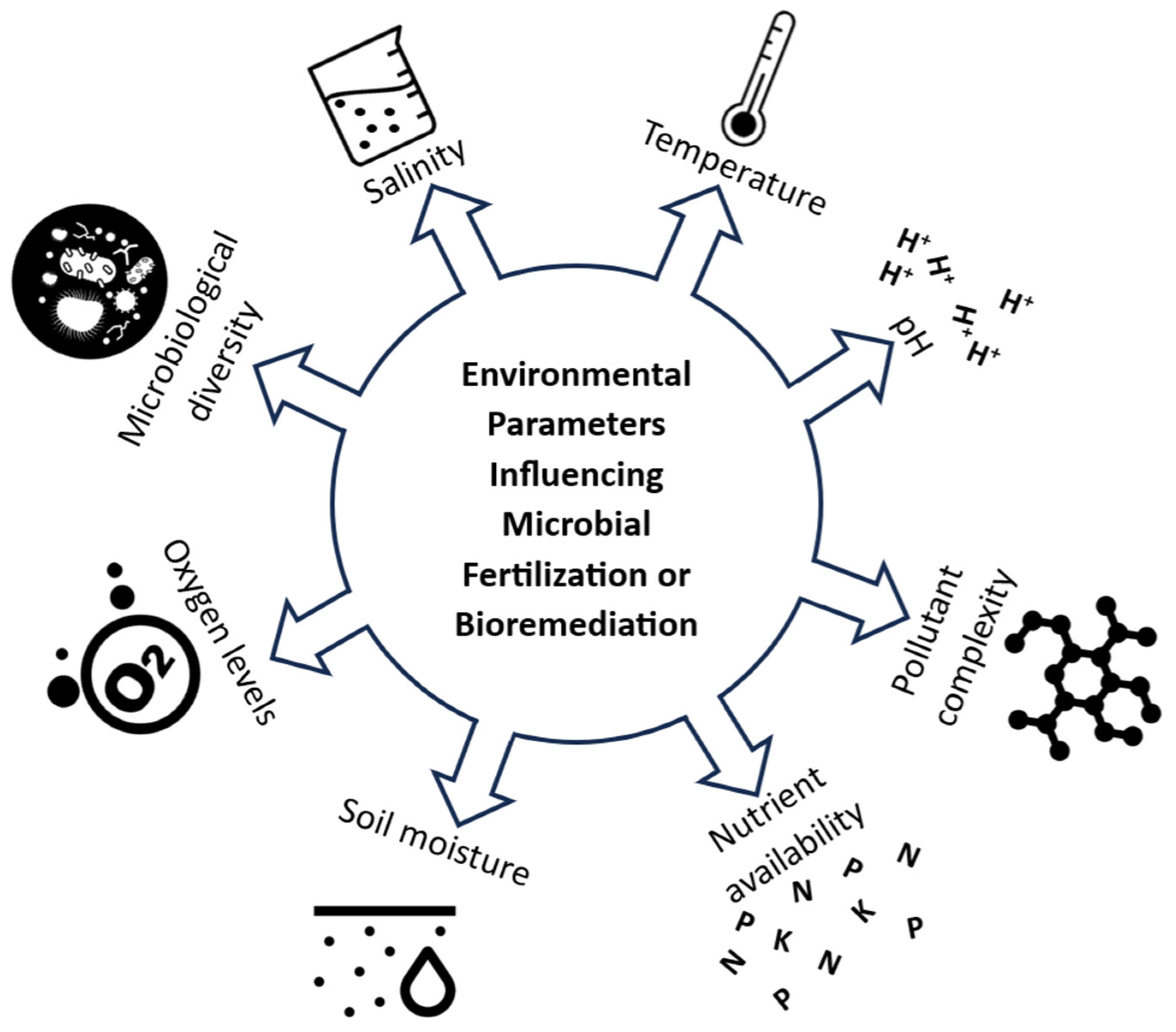

Table 1.
Some microorganisms that have been used for bioremediation of heavy metal-contaminated soils.
Table 1.
Some microorganisms that have been used for bioremediation of heavy metal-contaminated soils.
| Microbial inoculum | Heavy metal(s) | Reference |
|---|---|---|
| Bacteria: | ||
| Arthrobacter spp. | Cd | Roane et al., 2001 [197] |
| Pseudomonas veronii | Cd, Zn, Cu | Vullo et al., 2008 [198] |
| Burkholderia spp. | Zn, Pb, Mn, Cd, Cu, As | Yang et al., 2016 [199] |
| Kocuria flava | Cu | Achal et al., 2011 [200] |
| Bacillus cereus | Pb, Cd, Cr | Ayangbenro and Babalola, 2020 [201] |
| Sporosarcina ginsengisoli | As | Achal et al., 2012 [202] |
| Serratia sp. | Ni, Cd | Li et al., 2016 [203] |
| Enterobacter cloacae KJ-46, E. cloacae KJ-47, Sporosarcina soli B-22, and Viridibacillus arenosi B-21 | Cd, Pb, Cu | Kang et al., 2016 [204] |
| Enterobacter cloacae B2-DHA | Cr(VI) | Rahman et al., 2015 [205] |
| Brevibacillus parabrevis OZF 5 | Cr(VI), Zn | Wani et al., 2023 [206] |
| Cupriavidus metallidurans LBJ and Pseudomonas stutzeri LBR | Pb | Ridene et al., 2023 [207] |
| Acinetobacter sp. LSN-10 | Mn(II) | [208] |
| Achromobacter denitrificans, Klebsiella oxytoca, and Rhizobium radiobacter | Cd, Hg, As, Pb, Ni | [188] |
| Cyanobacteria | ||
| Gloeomargarita lithophora and Cyanothece sp | Sr | [209] |
| Nostoc minutum and Anabaena spiroides | Pb, Cd, Ni | [210] |
| Fungi | ||
| Simplicillium chinense | Cd, Pb | [211] |
| Penicillium sp. | Hg | [212] |
| Penicillium chrysogenum FMS2 | Cd | [213] |
| Aspergillus versicolor | Cr, Ni, Cu | [214] |
| Aspergillus fumigatus | Pb(II) | [215] |
| Aspergillus spp. | Cd, Cu | [216] |
| Gloeophyllum sepiarium | Cr | [200] |
| Perenniporia subtephropora, Daldinia starbaeckii, Phanerochaete concrescens, Cerrena aurantiopora, Fusarium equiseti, Polyporales sp., Aspergillus niger, Aspergillus fumigatus, and Trametes versicolor | As, Mn, Cu, Cr, and Fe | [217] |
| Yeasts | ||
| Candida tropicalis | Cr(VI) | [218] |
| Saccharomyces cerevisiae | Pb, Cd | [219] |
| Saccharomyces cerevisiae | Cd, Hg | [220] |
| Mixed consortia | ||
| Chlorella thermophila SM01, Leptolyngbya sp. XZMQ and Bacillus XZM | As | [221] |
| Leptolyngbya sp. XZ1 and Bacillus sp. S1 | Cd | [222] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



