Submitted:
30 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
The rising incidence of prostate cancer necessitates innovative treatment approaches, particularly as diseases like the COVID-19 pandemic can disrupt traditional cancer care. This study aimed to evaluate the impact of hypofractionated versus conventionally fractionated radiotherapy on prostate cancer cell lines in vitro. Prostate cancer cell lines (PC-3 and DU-145) were exposed to varying doses of radiation alongside non-cancerous BPH-1 cells. We assessed radiation effects on cell proliferation, viability, colony formation, DNA repair, migration, invasion, and cytotoxicity. Results demonstrated that the prostate cell lines exhibited varying responses, with hypofractionation favourably impacting aggressive PC-3 cells while preserving non-cancerous cells. In contrast, conventional fractionation led to increased invasion and cytotoxicity in both prostate cancer cell lines. These findings advocate for personalised radiation therapy approaches that enhance treatment efficacy by considering the distinct behaviours of differing prostate cancer subtypes.
Keywords:
α/β ratio
; Cell lines
; Clonogenic survival assay
; Gamma-H2AX assay
; Hypofractionated radiotherapy
; Invasion assay
; Lactate dehydrogenase assay
; Migration assay
; Prostate cancer
; Radiobiology
1. Introduction
Prostate cancer is a prevalent cancer in men, accounting for roughly 15% of cancer cases worldwide, with even higher rates in Sub-Saharan Africa, where it constitutes 23% of male cancers [1]. Projections suggest that the number of new prostate cancer cases will surge from 1.4 million in 2020 to 2.9 million by 2040, underscoring the need for proactive government strategies alongside lifestyle and public health initiatives to manage this increase [2]. Radiation therapy (RT) has advanced significantly, becoming a cornerstone of cancer treatment as both a primary and adjunctive modality [3,4,5,6,7]. The COVID-19 pandemic has highlighted the importance of reassessing treatment strategies, bringing hypofractionated radiotherapy (HFRT) to the forefront as an effective approach that reduces hospital visits and infection risks while maintaining efficacy [8].
Hypofractionation (HF) delivers higher fractionated doses of radiation in fewer sessions, optimising therapeutic outcomes [9]. While some studies demonstrate its successful implementation, ongoing debates surround the optimal fractionation schemes and the underlying radiobiological mechanisms [10,11,12]. Comprehensive in vitro investigations are essential to further our understanding of HFRT and its application [13]. The complexity of radiobiological mechanisms is influenced by tissue sensitivity, fractionation, and dose delivery, with in vitro studies providing vital insights into biological responses to radiation [14]. These factors are critical in understanding HF within variable cancer contexts, where financial constraints, limited access to technology, and disparities in healthcare can impact treatment decisions [15]. Radiotherapy centres in Africa have adapted their approaches to hypofractionated dosing, prioritising patient care and safety [16,17], reflecting a broader global response to the pandemic that emphasises flexibility and preparedness [18].
Recent studies, including the CHHiP trial and the RTOG 0415 trial, have enhanced the understanding of hypofractionated radiation therapy for prostate cancer, indicating that its efficacy is comparable to conventional therapy, particularly in terms of disease-free survival for low-risk patients [19,20,21]. Evaluating the interplay of tissue sensitivity, fractionation, and dose delivery is essential for assessing treatment outcomes and toxicity profiles in prostate cancer radiotherapy [22,23]. Advanced modalities such as intensity-modulated radiation therapy (IMRT) and stereotactic body radiotherapy (SBRT) have been shown to enhance outcomes while minimising exposure to healthy tissue [24,25].
The linear-quadratic (LQ) model is fundamental for understanding the biological response to radiation, recognising the interplay between lethal and potentially lethal effects [22,26,27]. The α/β ratio derived from this model is critical for predicting treatment outcomes and understanding radiosensitivity and repair capacity [8,28,29]. A high α/β ratio indicates greater sensitivity to higher doses per fraction, informing clinical strategies for hypofractionation regimens through biological effective dose (BED) modelling [30,31,32,33].
The clonogenic survival assay (CSA) is a key method for evaluating cellular responses and determining α/β values, providing insights into the biological mechanisms underpinning treatment responses [34]. Notably, research into DNA repair processes, including non-homologous end joining (NHEJ) and homologous recombination repair (HRR), has crucial implications for understanding hypofractionated radiotherapy outcomes [35,36,37,38]. Additionally, radiation impacts cell growth and apoptosis, with the p53 gene playing a pivotal role in modulating these processes, particularly in the context of cancer behaviour [39,40].
Lactate dehydrogenase (LDH), a key enzyme in the glycolytic pathway, serves as an important biomarker for monitoring cancer treatment outcomes, correlating with tumour progression and response to therapy [41,42,43,44,45]. Elevated LDH levels often indicate aggressive disease behaviour and poor prognosis, offering insights into tumour metabolism and treatment response [46,47,48].
This study aimed to elucidate the radiobiological principles underlying HF and conventionally fractionated (CF) radiation therapy. Key concepts such as the α/β ratio, BED, and radiobiological assays were explored to investigate the differential effects of HF and CF radiation doses on tumour and normal cell responses. A hypofractionated regimen was designed to deliver a single larger dose in one treatment session, theoretically achieving similar BED as conventional fractionation. Cellular responses to radiation were examined through clonogenic survival assays, DNA damage assessment, migration, invasion, adaptive response, and LDH secretion. By integrating historical perspectives with contemporary research findings, this study seeks to enhance the understanding of the radiobiological effects of hypofractionation, ultimately informing the development of more effective treatment protocols and contributing to the growing body of knowledge in this area.
2. Materials and Methods
A systematic investigation was conducted to examine the effects of HF and CF radiation on prostate cell lines. Radiation doses were calculated based on clonogenic survival assays and adjusted to achieve a comparable BED between the two models. Radiation-induced responses were evaluated by monitoring cell growth and conducting radiobiological assays, aimed at elucidating the effects of HF and CF on cellular behaviour and informing optimal treatment strategies. Specifically, the use of in vitro models with isolated cells allowed for the control of various variables and isolation of the effects of different fractionation schedules. For HF, a single fraction was chosen to represent the higher dose delivered, as demonstrated by previous studies [49,50].
2.1. Prostate Cell Lines
This study utilised the following cell lines: the cancerous cell lines (PC-3 and DU-145) and a non-cancerous cell line (BPH-1). The PC-3 cell line was cultured in Ham's F-12 medium (Catalogue number 21765029) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin (100 U/mL penicillin and 100 μg/mL streptomycin) under standard culture conditions (37°C, 5% CO2). BPH-1 was maintained in Roswell Park Memorial Institute (RPMI) medium (Catalogue number 21875034) supplemented with 10% FBS, 1% penicillin-streptomycin, and 2 mM L-glutamine under similar conditions (37°C, 5% CO2). DU-145 was cultured in Eagle's minimum essential medium (EMEM) (Catalogue number 21575022) supplemented with 10% FBS and 1% penicillin-streptomycin, also under standard conditions (37°C, 5% CO2). All media and supplements were obtained from LTC Tech South Africa (Pty) Ltd. The cancerous cell lines (PC-3 and DU-145) are characterised by a mutant p53 and high metastatic potential, with PC-3 being androgen receptor-negative (AR-) and DU-145 being androgen receptor-positive (AR+). BPH-1, in contrast, is a non-cancerous prostate epithelial cell line exhibiting wild-type p53 [51,52,53].
2.2. Doubling time, Adaptive Response and Clonogenic Survival Assay (CSA)
The doubling time of the cell lines was calculated using a standard formula [54,55]. Cells were seeded at a predetermined density, and initial counts were recorded. Subsequent counts were taken at 24-hour intervals over 72 hours to monitor proliferation. Prior to experimentation, cells were cultured for several weeks to ensure active growth. A priming dose of 0.5 Gy was administered to assess the adaptive doubling time, with additional serial counts performed to evaluate the effect of low-dose radiation (0.5 Gy) on cell proliferation. X-ray irradiation was conducted using a Precision X-Ray 320 kV irradiator (Madison, Connecticut, USA), operated at 250 kV with a dose rate of 0.69 Gy/min.
The Clonogenic survival assay (CSA) was used to assess the surviving fraction of cells following exposure to varying doses of radiation [56]. The CSA served as the primary endpoint for evaluating radiation sensitivity and α/β ratios of the cell lines. Each cell line was seeded in 6-well plates at densities corresponding to each radiation dose and subsequently irradiated. A range of doses from 2 Gy to 10 Gy was administered in increments of 2 Gy. A total of 3 technical replicates were performed for each experimental condition, alongside 6 biological replicates per cell line and endpoint to ensure the reliability of the results. After incubating the cell cultures for six doubling times to facilitate colony formation, the surviving fractions at all doses were calculated. The survival data were fitted to the linear-quadratic (LQ) model using non-linear regression analysis with the Marquardt-Levenberg algorithm [57] to generate survival curves. The LQ model equation was employed to model the relationship between radiation dose and survival fraction:
where S(D) represents the surviving fraction at dose D, and α and β are parameters extracted from the curve fit. The fitted curve allowed for the extraction of the LQ model parameters (α) and (β), representing the linear and quadratic components of cell killing, respectively. Additionally, D50, the absorbed radiation dose required for 50% cell killing, was calculated using the LQ model equation. The relative sensitivity (RS), indicating the ratio of D50 values between non-cancerous and cancerous cell lines, was also determined. The extracted α and β values were interpreted in the context of radiosensitivity, with higher α/β ratio values suggesting greater sensitivity to radiation.
S(D)=e−(αD+βD^2),
2.3. Biological Effective Dose (BED)
The biologically effective dose (BED) quantifies the biological effectiveness of radiation therapy by considering the dose per fraction and the radiobiological parameters, alpha (α) and beta (β), across different regimens [58]. BED provides a standardised measure for comparing the effects of radiation doses delivered over varying schedules, thereby facilitating the optimisation of radiation therapy protocols. In the HF model, the BED for a single fraction was calculated using Equation (2):
where D is the total dose, d is the dose per fractions, and α/β is the ratio of alpha to beta [58]. By substituting the calculated BED from conventional fractionation of 2 Gy per fraction over 4 fractions (totaling 8 Gy), the required dose for hypofractionation (D = d for a single fraction) can be determined. In this context, the dose (D) for hypofractionation can be calculated as the only unknown variable in the BED equation, given that all other variables have been established using the conventional fractionation scheme.
2.4. Split Dose Experiment and the Clonogenic Survival Assay
To investigate the effects of varying time gaps between radiation fractions on cell survival, cells were seeded in T25 flasks and irradiated with a total dose of 2 Gy, divided into 2 fractions with intervals of 1 to 6 hours. The cells first received 1 Gy, followed by incubation for the respective time. After this, a second 1 Gy fraction was delivered, and the plates were incubated for a duration equivalent to six doubling times. Following six doubling times to allow for colony formation, surviving fractions were determined. For staining, a 5% Crystal Violet solution was prepared by dissolving 500 mg of Crystal Violet (Merck Life Science, Catalogue number: 179337) in a mixture of 25 mL AR-grade methanol (Merck Life Science, Catalogue number: 179335) and 75 mL distilled water. This solution was utilised after the cells were fixed using ice-cold 100% methanol for 10 minutes. Cells were washed with cold PBS, then incubated with 3 mL of 0.5% Crystal Violet in 25% methanol for 10 minutes at room temperature. Excess dye was removed, and the plates were rinsed with water to halt the staining process, followed by air drying. Colonies were manually counted by visually identifying distinct morphologically viable colonies. The surviving fraction at each interval was compared to that of cells irradiated with the full 2 Gy dose without time gaps. The repair factor was calculated as the percentage change in survival due to each time interval, providing insight into the cells' capacity to repair DNA damage.
2.5. Gamma-H2AX Foci Assay
The γH2AX foci assay, as described by Nair et al. (2021), was employed to investigate DNA damage and repair kinetics following radiation exposure [59]. One million cells were irradiated with 2 Gy and incubated for 0-6 hours. Following the harvesting and trypsinisation of one million cells, they were irradiated with a dose of 2 Gy and incubated for time intervals ranging from 0 to 6 hours. For fixation, cells were treated with PBS containing 3% paraformaldehyde (PFA, freshly prepared) for 20 minutes, followed by overnight storage in PBS with 0.5% PFA. Immunostaining was performed using a primary anti-γ-H2AX antibody (catalogue number: 613402, Biocom Africa) and a secondary rabbit anti-mouse FITC antibody (catalogue number: 31561, LTC Tech South Africa), prior to quantification. The average number of γH2AX foci per cell was evaluated using the MetaCyte software module of the Metafer version 4 scanning system (MetaSystems, Germany). A minimum of three slides were scored for each exposure condition.
To illustrate the repair processes of DNA damage induced by radiation, the efficiency of radiation-induced DNA damage repair was calculated using equation 3:
Where REt represents the repair efficiency at timepoint (t), γ-H2AXmax represents the maximum amount of foci observed in response to 2 Gy, γ-H2AXt the amount of foci observed at the given timepoint in response to 2 Gy.
This methodology outlines the experimental steps and analysis involved in assessing DNA damage response through immunostaining of γ-H2AX foci following 2 Gy irradiation at different time points. The repair efficiency (RE) was calculated based on the methodology outlined by Panek and Miszczyk (2019), with slight modifications to accommodate our experimental conditions [60].
2.6. Migration Assay
The migration assay was conducted to investigate the migratory behaviour of the BPH-1 cell line, which serves as a non-cancerous model for benign prostatic hyperplasia (BPH). By using a non-cancerous cell line, this study aims to elucidate the effects of radiation on normal prostate tissue, thereby providing valuable insights into the response of non-cancerous cells in contrast to malignant counterparts. Understanding these dynamics is crucial for developing effective radiotherapy strategies that minimise damage to non-cancerous cells while effectively targeting tumour cells. Cells were seeded in T25 flasks at a density that ensured 100% confluence after 24 hours. Cell suspensions were prepared and added to each well of the Culture-Insert 2 Well. After confluence was reached, the insert (Culture-Insert 2 Well ibidi® GmbH, Catalogue number: 80209-150, ibidi®, GmbH was carefully removed, and non-attached cells and debris were washed away with cell-free medium.
Irradiation was administered at a dose of 2 Gy per fraction, totalling 8 Gy delivered in 4 fractions with a 3-hour interval between each conventional fractionation. For the hypofractionation approach, a single dose of 4.93 Gy was applied. Both fractionation schemes resulted in a similar biological effective dose (BED) of 13.93 Gy, allowing for a comparative analysis of the effects on cell migration under different radiotherapy regimens. Following irradiation, cells were monitored using the CytoSMART™ System, a compact live cell imaging system acquired from Lonza, (supplied locally by Whitehead Scientific), which enables time-lapse observation every 4 hours without disturbing the cultures. Images were captured at regular intervals with a 10x objective lens and analysed using ImageJ software (Version 1.54i) to quantify cell migration dynamics. The change in gap area over time was plotted, and linear regression analysis was conducted to calculate the slope of the linear phase, representing the migration speed of the cells.
2.7. Invasion Assay
The invasion assay is a crucial tool in cancer research for assessing the invasive potential of tumour cells, mimicking the intricate process of cancer metastasis [56]. This assay provides insights into the molecular and cellular determinants of tumour invasion. The transwell invasion assay was employed to investigate the invasive properties of prostate cancer cell lines (PC-3 and DU-145), using Geltrex® (catalogue number: A1413202), obtained from LTC Tech South Africa (Pty) Ltd, as a substrate. The assay was performed per the manufacturer’s instructions, involving preparation, staining, and quantification. Briefly, irradiated cells were treated with a total dose of 8.00 Gy in four fractions for both PC-3 and DU-145 cell lines, with each fraction delivered at 2.00 Gy and a 3-hour gap between fractions, while in the hypofractionation scheme, cells received single doses of 5.15 Gy and 4.85 Gy, respectively, before being added to the upper chamber, while complete media with 10% FBS was added to the lower well. The insert (LTC Tech South Africa Pty LTD, catalogue number 140640) was incubated for 24 hours at 37°C. After incubation, the insert was removed, washed with PBS, fixed with 4% paraformaldehyde, and stained with Crystal Violet. Invaded cells were quantified at 10x magnification using the CytoSMART Lux 10 imaging system (Lonza, Basel, supplied locally by Whitehead Scientific) and expressed as a percentage of the total number of seeded cells.
2.7. Lactate Dehydrogenase (LDH) Assay
The LDH assay, a well-established method for evaluating cellular health, was used to assess cytotoxicity by quantifying lactate dehydrogenase enzyme release into the culture supernatant. LDH, a ubiquitous cytoplasmic enzyme, indicates cellular damage when found extracellularly [56]. The LDH-Cytox Assay Kit (Biocom Africa, Catalogue: 426401) was employed according to the manufacturer's instructions, which included meticulous cell preparation and incubation with test substances. Both cancerous (DU-145, PC-3) and non-cancerous (BPH-1) prostate cell lines were cultured in complete growth medium under standard conditions (37°C, 5% CO2) and prepared at a concentration of 5 × 105 cells/ml. These cells were then subjected to either conventional fractionation (2.00 Gy × 4 fractions for a total of 8.00 Gy with a 3-hour inter-fractional interval) or hypofractionation (single doses of 4.93 Gy for BPH-1, 4.85 Gy for DU-145, and 5.15 Gy for PC-3). Following irradiation, the released LDH levels were measured using the LDH-Cytox Assay Kit (Biocom Africa, Catalogue: 426401) according to the manufacturer’s instructions, and absorbance was determined at 490 nm using a microplate reader (Berthold Technologies, Version 2.2.2.1).
2.8. Data Analysis
Data analysis was conducted meticulously to ensure the reliability and reproducibility of the results. This study aimed to compare the responses of various cell lines to two radiotherapy techniques: conventional fractionation and hypofractionation. Multiple assays were performed to assess the effects of these radiotherapy methods on prostate cell lines in vitro. Microsoft Excel (Microsoft Office Standard 2019) was utilised for initial data organisation and basic statistical analysis. Means and standard deviations for each experimental condition were calculated to describe the variability in cell responses, with standard deviation derived from the formula that quantifies the dispersion of data points around the mean.
Logarithmic plots were generated using GraphPad Prism (version 10.0.0 for Windows), with the radiation dose plotted on the x-axis and the survival fraction on the y-axis. To analyse the survival fraction data, nonlinear regression analysis was performed using the Marquardt-Levenberg algorithm in GraphPad Prism. This approach facilitated the fitting of survival curves to the LQ model, effectively modelling the relationship between radiation dose and cell survival. During this analysis, the estimated parameters (α and β) were derived, providing critical insights into the cellular responses to radiation exposure. The α and β values obtained from the survival curves were compared across the various cell lines. Statistical analyses were performed using Analysis of Variance (ANOVA) to assess differences among the different fractionation conditions. For all statistical tests, p-values were calculated to assess the significance of the observed differences. A p-value of less than 0.05 indicated that the differences in radiation response metrics among the treatment conditions were unlikely to have occurred by chance, thereby allowing for meaningful conclusions regarding the effects of conventional versus hypofractionation radiotherapy.
3. Results
3.1. Optimisation of Comparable Treatment Regimens and Adaptive Doubling Time
Results showed significant variability in growth rates among the cell lines. Table 1 presents the baseline and adaptive doubling times for each cell line. The adaptive doubling time, after exposure to 0.5 Gy radiation, showed an increase compared to the baseline doubling time for each prostate cell line. However, statistical analysis did not reveal significant differences in the adaptive doubling times compared to the baseline values for any of the prostate cell lines.
Table 2 summarises the radiobiological parameters and dose-response characteristics of the prostate cell lines. The α values for BPH-1, PC-3, and DU-145 were 0.08, 0.12, and 0.10 Gy−1, respectively, while the β values for BPH-1, PC-3, and DU-145 were 0.03, 0.03, and 0.04 Gy−2, respectively. The D50 values also varied: BPH-1 showed 3.66 Gy, PC-3 had 3.21 Gy, and DU-145 presented 3.10 Gy. Statistical analysis indicated no significant differences in D50 between the non-cancerous BPH-1 and the cancerous cell lines (PC-3 and DU-145). The relative sensitivity (RS) values were 1.14 and 1.18 for PC-3 and DU-145, respectively, although these differences were not statistically significant (p = 0.447 and 0.794, respectively).
Figure 1 illustrates the repair efficiency and maximum foci observed in prostate cell lines BPH-1, PC-3, and DU-145 after irradiation with 2 Gy. The graph shows distinct repair dynamics across the cell lines over time. The maximum foci were observed at different times post-irradiation: 0.5 hours for BPH-1 and PC-3 cells, and 1 hour for DU-145 cells. Subsequently, the maximum disappearance of foci was observed uniformly across all cell lines at 3 hours post-irradiation, with corresponding repair efficiencies of 99.15% for BPH-1, 93.62% for PC-3, and 89.57% for DU-145. The graph also shows the maximum recovery factors, which indicate the ability of cells to recover after receiving two 1 Gy doses with a time gap compared to a single 2 Gy dose with no time gap.
3.2. Surviving Fraction and Adaptive Response
The results depicted in Figure 2 illustrate that the surviving fractions for all prostate cell lines were higher under the HF regimen compared to CF. BPH-1 cells exhibited percentage survivals of 15.28% for CF and 21.34% for HF (p < 0.01). Similarly, PC-3 cells demonstrated percentage survivals of 17.23% for CF and 22.92% for HF (p < 0.05). While DU-145 cells showed survival fractions of 19.08% for CF and 21.34% for HF (p = 0.060), the difference was statistically insignificant.
The effects of the priming dose of 0.5 Gy resulted in an overall increase in survival rates. For the BPH-1 cell line, both CF and HF resulted in significant increases in percentage survival. Specifically, survival increased from 15.28 to 17.36% after CF, reflecting a difference of 2.08% (p = 0.0073), while it increased from 21.34 to 24.51% following HF, representing a difference of 3.17% (p = 0.0006). Similarly, in the DU-145 cell line, survival rose from 19.08 to 22.88% after CF, resulting in a difference of 3.80% (p = 0.0002), whereas it increased from 25.48 to 31.34% after HF, corresponding to a difference of 5.86% (p < 0.0001). In the PC-3 cell line, significant increases in percentage survival were also exhibited after both CF and HF with a priming dose. CF resulted in an increase from 17.23 to 20.27%, indicating a difference of 3.04% (p = 0.0036), and HF increased survival from 22.92 to 27.34%, with a difference of 4.42% (p < 0.0001).
As in Table 3, the hypofractionation and intrinsic survival (IS) rates differed for all cell lines. The percentage IS was calculated using the LQ model, which incorporated the α/β components to estimate the percentage cell survival as a function of the radiation doses used for HF (or single-dose.
The non-cancerous cell line, BPH-1, exhibited statistically significant differences between the intrinsic and single-dose survival rates (p = 0.0012).
3.3. Prostate Cell Line Migration and Invasion
Figure 3a illustrates the gap closure over time for the control group and groups treated with CF and HF. The slope values for gap closure were as follows: -4.9121 × 104 μm2h−1 for the control, -2.7125 × 104 μm2h−1 for CF treatment, and -3.0554 × 104 μm2h−1 for HF treatment. The steeper slope in the control group indicates the fastest migration, followed by the HF-treated group and then the CF-treated group. Comparison of slopes between the CF and HF treatment groups revealed a significant difference in migration capability (p < 0.01), suggesting a difference between hypofractionation and conventional fractionation. Additionally, both irradiation groups exhibited a significantly slower migration rate compared to the control group (p < 0.001).
The invasion percentages of prostate cell lines into the extracellular matrix under different fractionation regimes including the unirradiated groups are presented in Figure 3b.
For PC-3 cells, the invasion percentage was 87% (±10%) for the control group, 94% (±3%) for the CF-treated group, and 91% (±7%) for the HF-treated group. For DU-145 cells, the invasion percentage was 84% (±5%) for the control group, 95% (±3%) for the CF-treated group, and 87% (±6%) for the HF-treated group. Comparing the invasion percentages, the CF-treated group exhibited the highest invasion capacity for both PC-3 and DU-145 cells, while the control group had the lowest invasion capacity although this increase was statistically insignificant. The p-values for the significant test between the CF and HF fractionation regimes for PC-3 cells was 0.766, and for DU-145 cells was 0.111, indicating no significant difference in invasion capability between the two regimes for both cell lines.
3.4. Lactate Dehydrogenase (LDH)
Figure 4 presents the results of the LDH assay for the prostate cell lines under HF and CF regimes.
For BPH-1 cells, the percentage of LDH released was 24% (±0.009) under HF and 25% (±0.005) under CF, with a p-value less than 0.01. For DU-145 cells, the percentage of LDH released was 19% (±0.014) under HF and 21% (±0.016) under CF, with a p-value of 0.084. Similarly, for PC-3 cells, the percentage of LDH released was 18% (±0.026) under HF and 19% (±0.007) under CF, with a p-value of 0.314. The results indicate a statistically significant difference in LDH release between HF and CF for BPH-1 cells (p < 0.01), while no significant differences were observed for DU-145 (p = 0.084) and PC-3 (p = 0.314) cells.
4. Discussion
The distinct radiobiological characteristics of prostate cell lines are elucidated through the analysis of their α and β values, which inform their radiosensitivity and potential responses to radiation therapy (see Table 2). Among the tested cell lines, PC-3 cells exhibited the highest α value of 0.12 Gy⁻1, indicating a greater sensitivity to radiation-induced cell death compared to BPH-1 (α = 0.08 Gy⁻1) and DU-145 (α = 0.10 Gy⁻1). Notably, while PC-3 cells shared the same β value of 0.03 Gy⁻2 as BPH-1, Du-145 cells demonstrated a higher β value of 0.04 Gy⁻2, indicating more efficient DNA repair mechanisms. Consequently, the lowest α/β ratio of the PC-3 cells, highlights their heightened radiosensitivity which is attributed to its increased susceptibility to radiation. The low α/β ratios observed in both PC-3 and DU-145 further indicate responsiveness to fractionation, where higher doses per fraction may overwhelm repair mechanisms, facilitating increased cell death or growth inhibition. Jayakumar et al. (2014) reported reduced radiosensitivity in DU-145 cells compared to PC-3, indicating that individual cellular characteristics significantly influence treatment outcomes [61]. Though, Lövey et al. (2013) noted that, although PC-3 cells demonstrate comparable radiosensitivity at lower doses (0.5 and 2 Gy), their surviving fraction declines more steeply relative to DU-145 as the radiation dose increases, thereby highlighting the complex relationship between radiation dose and cellular response [62]. Despite these differences, the relative sensitivity (RS) values of PC-3 and DU-145 compared to BPH-1 showed no statistically significant differences, suggesting that both cancerous lines have similarly elevated sensitivity to radiation. In terms of fractionation schemes, the α/β ratios provide valuable insights into potential responses, with PC-3 displaying the highest overall ratio and thus indicating greater sensitivity than both DU-145 and BPH-1.
The concept of a "therapeutic window" pertains to radiation doses that effectively target and kill cancer cells while minimising damage to normal tissues. The observed RS values, coupled with the narrower therapeutic window in cancerous cell lines, underscore the potential for enhanced treatment efficacy [63]. Predominant β values of 0.03 Gy⁻2 across all cell lines imply comparable DNA repair efficiencies, revealing that variations in radiation sensitivity are primarily governed by differences in α values. These initial findings align with the work of van Leeuwen et al. (2018), where α values across prostate cell lines varied from 0.1 to 0.3 Gy⁻1, and β values ranged from 0.01 to 0.06 Gy⁻2 [22]. The combined evidence of lower α/β ratios, reduced D50 values, and consistent β values in cancerous prostate cell lines supports the hypothesis that hypofractionated radiotherapy could leverage the differential responses between cancerous and non-cancerous prostate cells. This necessitates further inquiry into optimal fractionation regimens in treating prostate cancer, taking into account the dynamics of radiation-induced cell death and DNA repair mechanisms.
Interestingly, the intrinsic survival estimated through the LQ model did not consistently align with the observed survival following a single hypofractionated dose (see Table 3). For instance, the intrinsic survival of BPH-1 was calculated at 32.5 ± 3.5%, while the observed survival posts a 4.93 Gy dose was lower at 21.3 ± 4.0%, implying that the LQ model underestimates the radiosensitivity of these cells. Conversely, PC-3 cells exhibited an observed survival of 25.5 ± 2.0% after a 5.15 Gy application, which surpassed predicted values, indicating an overestimation of their radiosensitivity by the LQ model. In contrast, DU-145 cells displayed an observed survival closely resembling their intrinsic survival at 22.9 ± 2.0%, thus indicating a more accurate prediction by the LQ model. Statistically significant differences were found between predicted and observed survivals for BPH-1 cells (p = 0.0012), while no significant divergence was observed for PC-3 or DU-145 (p = 0.5612 and p = 0.3148, respectively). These results underscore the limitations of the LQ model in predicting radiosensitivity, suggesting that more nuanced models may be necessary to accurately account for cellular responses in specific contexts. Investigations by Cui et al. (2020) indicate that converting conventionally fractionated doses to a single high dose may necessitate the use of alternative formulas or a higher α/β ratio within standard LQ models [64].
Following the administration of a 0.5 Gy priming dose, significant adaptive responses were noted across the prostate cell lines, indicative of inherent radioresistance (p < 0.05). The CF and HF with a 0.5 Gy priming dose resulted in increased survival percentages for all tested cell lines, specifically BPH-1 (15.28 to 17.36%, p < 0.01), PC-3 (19.08 to 22.88, p < 0.05), and DU-145 (17.23 to 20.27%, p < 0.05). These results suggest robust DNA repair mechanisms among prostate cell lines, as illustrated in Figure 1a, and imply that efficient repair of DNA damage correlates with increased survival rates. Prostate cancer cells such as PC-3 and DU-145 typically present alterations in cell cycle regulation and apoptosis pathways, including mutations in tumour suppressor genes (notably p53) and overexpression of anti-apoptotic proteins. This allows them to evade programmed cell death [65]. Diverging from the expected outcomes, the adaptive responses observed in both BPH-1 and PC-3 cells imply that factors beyond p53 status contribute to their observed radioresistance [66]. Integral to cancer treatment strategies, these findings indicate that non-cancerous BPH-1 cells exhibited enhanced survival rates and adaptive response under HF, highlighting pronounced repair mechanisms (p < 0.001). Meanwhile, the aggressive PC-3 cells exhibited a statistically significant preference for CF over HF (p < 0.05), suggesting that HF strategies may benefit normal prostate tissue, whereas CF should be prioritised for aggressive cancer cells such as PC-3. This advocates for an integrated therapeutic approach designed to protect healthy tissues while optimising treatment outcomes based on tumour aggressiveness. Notably, the addition of a 0.5 Gy priming dose to CF enhanced the survival rate in non-cancerous BPH-1 cells, indicating a potentially beneficial effect of priming on these cells (p < 0.01). In contrast, the inclusion of a 0.5 Gy priming dose to CF and HF failed to reduce survival rates in PC-3 and DU-145 cells, suggesting that priming may not have a significant effect on the survival of these cancerous cells (p < 0.05). These findings suggested that a personalised approach to radiation therapy may be necessary, considering the specific characteristics of each tumour type and its sensitivity to different fractionation schemes and priming doses. This study underscores the importance of accounting for individual differences in DNA repair mechanisms, cell cycle regulation, and responses to cellular stress in developing radiation therapy protocols for prostate cancer. Combining HF and CF may improve therapeutic outcomes by leveraging the differential adaptive responses of non-cancerous and aggressive cancerous prostate cell lines. Therefore, a personalised approach to radiation therapy is essential, considering the unique characteristics of each tumour type and their sensitivity to varying fractionation schemes and priming doses.
The migration assay conducted on BPH-1 prostate cells provided valuable insights into the impact of radiation therapy on cell motility. The non-irradiated group exhibited rapid migration, whereas both the CF-treated and HF-treated groups displayed reduced speeds. This observation implies that radiation exposure inhibits the migration ability of non-cancerous cells and aligns with findings from other studies [67,68]. The comparison of migration rates between the CF and HF treatment groups indicated significant differences in slope values (p = 0.0002), suggesting that fractionation regimens differentially affect prostate cell motility. Importantly, both irradiation groups exhibited significantly slower migration rates compared to the control group (p < 0.001), underscoring the impact of radiation therapy on cell migration regardless of the fractionation regimen employed. The slower migration observed in the HF group may be attributed to enhanced cellular processes, such as activated repair mechanisms and metabolic activity, facilitating recovery despite reduced motility. Furthermore, the influence of radiation on cell adhesion is crucial in understanding the overall effects of radiation therapy on cellular behaviour within prostate tissue, particularly the observed reductions in motility alongside potential increases in tissue integrity and cellular communication [69,70].
The invasion assay conducted on prostate cancer cell lines (PC-3 and DU-145) revealed their invasive potential following exposure to different radiation fractionation regimes. While both cell lines are characterised by p53 mutations and originate from metastatic prostate adenocarcinoma, they inherently display a high degree of aggressiveness and a propensity for invasion. The irradiated groups showed increased invasion compared to control groups, highlighting the potential impact of radiation on enhancing invasive capabilities. These findings corroborate those reported by Chang et al. (2014), who observed increased invasive behaviour in prostate cell lines following irradiation [71]. Although both PC-3 and DU-145 exhibited slightly higher invasion capacities under CF compared to HF, these differences were not statistically significant. This suggests that the invasiveness of these cell lines may not be significantly influenced by the radiation fractionation schedule. The irradiation of immortalised cells may exacerbate p53 mutations while impairing the regulatory pathways that control cell cycle arrest and DNA repair [65]. This suggests that radiation may enhance the invasive potential of cells by disrupting key cellular mechanisms, although the lack of significant difference between CF and HF indicates that both fractionation schemes may similarly impact their invasive behaviour due to shared underlying molecular pathways.
In summary, the lactate dehydrogenase (LDH) assays demonstrate varying responses to radiation fractionation schemes among non-cancerous and cancerous prostate cell lines. For BPH-1 cells, the LDH release showed significant differences between CF and HF treatments (24% vs. 25%, p < 0.01), indicating that HF induces lower levels of cytotoxicity. This excess survival observed under HF suggests enhanced DNA repair capability among non-cancerous cells, making HF a potentially less damaging option and reducing the risk of adverse effects. Conversely, for the cancerous cell lines, DU-145 and PC-3, LDH release did not show significant differences between CF and HF. DU-145 cells exhibited 19% LDH release under HF and 21% under CF (p = 0.084), while PC-3 cells showed 18% under HF and 19% under CF (p = 0.314). The lack of significant differences in LDH release might be attributable to their p53 mutant status, which could lead to consistent levels of membrane damage irrespective of the fractionation scheme. However, the notable increase in surviving fraction under HF for PC-3 suggests more effective repair of radiation-induced damage, thereby reducing cytotoxic effects.
Moreover, the potential involvement of senescence in LDH release among cancerous cell lines warrants investigation, as radiation has been shown to increase the expression of γH2AX, a marker for senescence [72]. The similar LDH release observed despite increased survival for HF may indicate a rise in senescence within CF-treated cancer cells, which may not be immediately reflected in LDH assays that measure acute cytotoxicity. These findings highlight the complexity of cellular responses to different fractionation schemes and underscore the need for tailored treatment strategies that consider the specific characteristics of each tumour type and their sensitivity to varying fractionation schemes. Further research is required to fully understand the long-term effects and potential therapeutic implications of different fractionation strategies, particularly concerning their roles in influencing senescence and cancer progression. Optimising radiation therapy protocols for metastatic prostate cancer remains a critical endeavour, with the potential to significantly improve therapeutic outcomes based on the unique responses of the respective cell lines.
5. Conclusions
Prostate cancer is the second most common cancer diagnosis globally among men, accounting for over 1.4 million new cases each year. In the United States alone, it is estimated that 1 in 9 men will be diagnosed with prostate cancer during their lifetime. In sub-Saharan Africa, prostate cancer accounts for 23% of all male cancers, a significant burden in regions where resources are already strained. The field of cancer treatment faces numerous challenges, particularly when it comes to managing prostate cancer. One significant issue is the risk of excessive pre-treatment radiation doses inducing a radiation adaptive response, which can lead to increased radioresistance in some prostate cell types. This underscores the need for individualised treatment approaches to effectively counteract unintended radiation exposure. In response, advancements in radiation therapy, such as the adoption of shortened fractionation regimens, aim to improve oncologic outcomes and patient comfort. This highlights the importance of tailoring radiation therapy approaches to the unique characteristics of prostate cancer. The findings suggest that HF may be a more effective treatment option for preserving non-cancerous prostate cells, while CF may be more suitable for controlling aggressive prostate cancer subtypes. The observed differences in survival rates, migration, and invasion patterns between non-cancerous and cancerous prostate cells underscore the need for personalised radiation therapy strategies.
HF can result in lower LDH release and higher survival fractions in non-cancerous prostate cells, suggesting reduced cytotoxicity and better preservation of cell integrity. In contrast, cancerous cell lines exhibited varying responses to fractionation schemes, with PC-3 cells showing higher survival fractions under HF and DU-145 cells exhibiting no significant difference in survival. These findings have important implications for the development of optimal treatment strategies for prostate cancer patients.
In light of these results, it is recommended that healthcare providers consider personalised radiation therapy approaches for patients with prostate cancer, taking into account the unique characteristics of each patient's cancer. To improve treatment outcomes for prostate cancer patients, it is essential to prioritise timely and effective access to radiation therapy, particularly in resource-constrained settings. Further research is needed to investigate the long-term effects of radiation therapy on patients with prostate cancer, including the potential for delayed effects and impact on quality of life. Additionally, cost-effective strategies must be developed to deliver radiation therapy to underserved communities, ensuring equitable access to this important treatment modality.
Author Contributions
PdP, SN, PBN and JA conceptualised and designed the experiments. PdP performed the irradiation experiments, dosimetry, and laboratory work. PdP and JA analysed the data and PdP performed the statistical analysis. PdP, SN, PBN and JA wrote the paper. All authors contributed to and approved the final version of the article.
Funding
This project was funded by SAINTS (Southern African Institute for Nuclear Technology & Sciences).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
We would like to thank Professor Pauline Busisiwe Nkosi (PBN) from Durban University of Technology for covering the publication costs associated with this research.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Musekiwa, A.; Moyo, M.; Mohammed, M.; Matsena-Zingoni, Z.; Twabi, H.S.; Batidzirai, J.M.; Singini, G.C.; Kgarosi, K.; Mchunu, N.; Nevhungoni, P.; Silinda, P. Mapping evidence on the burden of breast, cervical, and prostate cancers in Sub-Saharan Africa: a scoping review. Frontiers in Public Health 2022, 10, 908302. [Google Scholar] [CrossRef] [PubMed]
- James, N.D.; Tannock, I.; N'Dow, J.; Feng, F.; Gillessen, S.; Ali, S.A.; Trujillo, B.; Al-Lazikani, B.; Attard, G.; Bray, F.; Compérat, E. The Lancet Commission on prostate cancer: planning for the surge in cases. The Lancet 2024, 403, 1683–1722. [Google Scholar] [CrossRef] [PubMed]
- Burnet, N.G.; Benson, R.J.; Williams, M.V.; Peacock, J.H. Improving cancer outcomes through radiotherapy: Lack of UK radiotherapy resources prejudices cancer outcomes. British Medical Journal 2000, 320, 198–199. [Google Scholar] [CrossRef] [PubMed]
- Gasinska, A. The contribution of women to radiobiology: Marie Curie and beyond. Reports of Practical Oncology and Radiotherapy 2016, 21, 250–258. [Google Scholar] [CrossRef]
- Casas, F.; Abdel-Wahab, S.; Filipovic, N.; Jeremic, B. Radiation therapy. [CrossRef]
- Ndlovu, N. Radiotherapy treatment in cancer control and its important role in Africa. Ecancermedicalscience 2019, 13, 942. [Google Scholar] [CrossRef]
- Valentini, V.; Boldrini, L.; Mariani, S.; Massaccesi, M. Role of radiation oncology in modern multidisciplinary cancer treatment. Molecular Oncology 2020, 14, 1431–1441. [Google Scholar] [CrossRef]
- Laine, A.M.; Pompos, A.; Timmerman, R.; Jiang, S.; Story, M.D.; Pistenmaa, D.; Choy, H. The role of hypofractionated radiation therapy with photons, protons, and heavy ions for treating extracranial lesions. Frontiers in Oncology 2016, 5, 302. [Google Scholar] [CrossRef]
- Insley, B.; Hsu, I.C.; Cunha, J.A.M. Paradigm shift in radiation treatment planning over multiple treatment modalities. Journal of Medical Physics 2021, 46, 135–139. [Google Scholar] [CrossRef]
- Feofanova, N.; Geraldo, J.M.; Andrade, L.M.D. Radiation oncology in vitro: trends to improve radiotherapy through molecular targets. BioMed Research International 2014, 2014, 461687. [Google Scholar] [CrossRef]
- Shibamoto, Y.; Miyakawa, A.; Otsuka, S.; Iwata, H. Radiobiology of hypofractionated stereotactic radiotherapy: what are the optimal fractionation schedules? Journal of Radiation Research 2016, 57, i76–i82. [Google Scholar] [CrossRef]
- Shuryak, I.; Hall, E.J.; Brenner, D.J. Optimized Hypofractionation Can Markedly Improve Tumor Control and Decrease Late Effects for Head and Neck Cancer. Radiation Oncology Biology 2019, 104, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Aitken, K.; Mukherjee, S. When Less is More: The Rising Tide of Hypofractionation. Clinical Oncology (Royal College of Radiologists (Great Britain)) 2022, 34, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, F.; Hernández, A. Optimization of radiotherapy fractionation schedules based on radiobiological functions. The British Journal of Radiology 2017, 90, 20170400. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.D.; Gao, D.; Redfern, A.; Thompson, E.W. Controversies around epithelial-mesenchymal plasticity in cancer metastasis. Nature Reviews Cancer 2019, 19, 716–732. [Google Scholar] [CrossRef]
- Kochbati L, Vanderpuye V, Moujahed R, Rejeb M Ben, Naimi Z, Olasinde T. Cancer care and COVID-19: tailoring recommendations for the African radiation oncology context. Ecancermedicalscience 2020, 14, 1144.
- Swanson, W.; Kamwa, F.; Samba, R.; Ige, T.; Lasebikan, N.; Mallum, A.; Ngoma, T.; Sajo, E.; Elzawawy, A.; Incrocci, L.; Ngwa, W. Hypofractionated radiotherapy in African cancer centers. Frontiers in Oncology 2021, 10, 618641. [Google Scholar] [CrossRef]
- Bernabucci, L.; Cornacchione, P.; Boldrini, L.; Pasini, D.; Dinapoli, L.; Smiljanic, L.; Valentini, V.; Dinapoli, N. Radiotherapy during the COVID-19: a review about management and treatment strategies. Reports of Practical Oncology and Radiotherapy 2022, 27, 291–302. [Google Scholar] [CrossRef]
- Fowler, J.; Chappell, R.; Ritter, M. Is alpha/beta for prostate tumors really low? International Journal of Radiation Oncology Biology Physics 2001, 50, 1021–1031. [Google Scholar] [CrossRef]
- Dearnaley, D.; Syndikus, I.; Mossop, H.; Khoo, V.; Birtle, A.; Bloomfield, D.; Graham, J.; Kirkbride, P.; Logue, J.; Malik, Z.; Money-Kyrle, J. Conventional versus hypofractionated high-dose intensity-modulated radiotherapy for prostate cancer: 5-year outcomes of the randomised, non-inferiority, phase 3 CHHiP trial. The Lancet Oncology 2016, 17, 1047–1060. [Google Scholar] [CrossRef]
- Lee, W.R. , Dignam, J. J., Amin, M., Bruner, D., Low, D., Swanson, G.P., Shah, A., D'Souza, D., Michalski, J.M., Dayes, I. and Seaward, S.A. NRG Oncology RTOG 0415: A randomized phase III non-inferiority study comparing two fractionation schedules in patients with low-risk prostate cancer. Journal of Clinical Oncology 2016, 4, 2325–2332. [Google Scholar]
- Van Leeuwen, C.M.; Oei, A.L.; Crezee, J.; Bel, A.; Franken, N.A.P.; Stalpers, L.J.A.; Kok, H.P. The alpha and beta of tumours: a review of parameters of the linear-quadratic model, derived from clinical radiotherapy studies. Radiation Oncology 2018, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.S.; Lasley, F.D.; Das, I.J.; Mendonca, M.S.; Dynlacht, J.R. Normal Tissue Radiation Response BT - Basic Radiotherapy Physics and Biology. In: Chang DS, Lasley FD, Das IJ, Mendonca MS, Dynlacht JR, editors. Cham: Springer International Publishing; 2021, 261–272. [CrossRef]
- Janopaul-Naylor, J.R.; Shen, Y.; Qian, D.C.; Buchwald, Z.S. The Abscopal Effect: A Review of Pre-Clinical and Clinical Advances. International Journal of Molecular Sciences 2021, 22, 11061. [Google Scholar] [CrossRef] [PubMed]
- Hassan, I.M.; Attalla, E.M.; El-Gohary, M.I. Impact of 3D conformal and Intensity Modulated Radiotherapy on secondary cancer risk for patients with early prostate cancer. Journal of Radiation Research and Applied Sciences 2022, 15, 46–54. [Google Scholar] [CrossRef]
- Hobbs, R.F.; Howell, R.W.; Song, H.; Baechler, S.; Sgouros, G. Redefining relative biological effectiveness in the context of the EQDX formalism: implications for alpha-particle emitter therapy. Radiation Research 2014, 181, 90–98. [Google Scholar] [CrossRef]
- McMahon, S.J.; Prise, K.M. Mechanistic modelling of radiation responses. Cancers 2019, 11, 205. [Google Scholar] [CrossRef]
- Dörr, W.; Schmidt, M. Normal Tissue Radiobiology. In: Brahme ABTCBP, editor. Oxford: Elsevier; 2014: 75–95. Available online: https://www.sciencedirect. 9780. [Google Scholar]
- Hawkins, R.B. Effect of heterogeneous radiosensitivity on the survival, alpha beta ratio and biological effective dose calculation of irradiated mammalian cell populations. Clinical and Translational Radiation Oncology 2017, 4, 32–38. [Google Scholar] [CrossRef]
- Gustafsson, J.; Nilsson, P.; Gleisner, K.S. On the biologically effective dose (BED)-using convolution for calculating the effects of repair: I. Analytical considerations. Physics in Medicine and Biology 2013, 58, 1507–1527. [Google Scholar] [CrossRef]
- Hopewell, J.W.; Millar, W.T.; Lindquist, C.; Nordström, H.; Lidberg, P.; Gårding, J. Application of the concept of biologically effective dose (BED) to patients with vestibular schwannomas treated by radiosurgery. Journal of Radiosurgery and SBRT 2013, 2, 257–271. [Google Scholar]
- Millar, W.T.; Hopewell, J.W.; Paddick, I.; Lindquist, C.; Nordström, H.; Lidberg, G.; Gårding, J. The role of the concept of biologically effective dose (BED) in treatment planning in radiosurgery. Physica Medica 2015, 31, 627–633. [Google Scholar] [CrossRef]
- Parisi, A.; Beltran, C.J.; Furutani, K.M. Clonogenic Survival RBE Calculations in Carbon Ion Therapy: The Importance of the Absolute Values of α and β in the Photon Dose-Response Curve and a Strategy to Mitigate Their Anticorrelation. Quantum Beam Science 2023, 7, 3. [Google Scholar] [CrossRef]
- Franken, N.A.P.; Rodermond, H.M.; Stap, J.; Haveman, J.; Van Bree, C. Clonogenic assay of cells in vitro. Nature Protocols 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Zahnreich, S.; Ebersberger, A.; Kaina, B.; Schmidberger, H. Biodosimetry Based on γ-H2AX Quantification and Cytogenetics after Partial- and Total-Body Irradiation during Fractionated Radiotherapy. Radiation Research 2015, 183, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ren, S.; Yu, S.; Pan, H.; Li, T.; Ge, S.; Zhang, J.; Xia, N. Methods favoring homology-directed repair choice in response to CRISPR/Cas9 induced-double strand breaks. International Journal of Molecular Sciences 2020, 21, 6461. [Google Scholar] [CrossRef]
- Nuijens, A.C.; Oei, A.L.; van Oorschot, B.; Visser, J.; van Os, R.M.; Moerland, D.; Franken, N.A.; Rasch, C.R.; Stalpers, L.J. Gamma-H2AX foci decay ratio as a stronger predictive factor of late radiation toxicity than dose-volume parameters in a prospective cohort of prostate cancer patients. International Journal of Radiation Oncology Biology Physics 2022, 112, 212–221. [Google Scholar] [CrossRef]
- Someya, M.; Hasegawa, T.; Nakamura, A.J.; Tsuchiya, T.; Kitagawa, M.; Gocho, T.; Mafune, S.; Ikeuchi, Y.; Tauchi, H.; Sakata, K.I. Prediction of late adverse events in pelvic cancer patients receiving definitive radiotherapy using radiation-induced gamma-H2AX foci assay. Journal of Radiation Research 2023, 64, 948–953. [Google Scholar] [CrossRef]
- Roger, L.; Gadea, G.; Roux, P. Control of cell migration: a tumor suppressor function for p53? Biology of the Cell 2006, 98, 141–152. [Google Scholar] [CrossRef]
- Millar, F.R.; Janes, S.M.; Giangreco, A. Epithelial cell migration as a potential therapeutic target in early lung cancer. European Respiratory Review 2017, 26, 1–6. [Google Scholar] [CrossRef]
- Porporato, P.E.; Dhup, S.; Dadhich, R.K.; Copetti, T.; Sonveaux, P. Anticancer targets in the glycolytic metabolism of tumors: a comprehensive review. Frontiers in Pharmacology 2011, 2, 49. [Google Scholar] [CrossRef]
- Erez, A.; Shental, O.; Tchebiner, J.Z.; Laufer-Perl, M.; Wasserman, A.; Sella, T.; Guzner-Gur, H. Diagnostic and prognostic value of very high serum lactate dehydrogenase in admitted medical patients. Israel Medical Association Journal 2014, 16, 439–443. [Google Scholar]
- Bok, R.; Lee, J.; Sriram, R.; Keshari, K.; Sukumar, S.; Daneshmandi, S.; Korenchan, D.E.; Flavell, R.R.; Vigneron, D.B.; Kurhanewicz, J.; Seth, P. The role of lactate metabolism in prostate cancer progression and metastases revealed by dual-agent hyperpolarized 13C MRSI. Cancers 2019, 11, 257. [Google Scholar] [CrossRef]
- Mishra, D.; Banerjee, D. Lactate Dehydrogenases as Metabolic Links between Tumor and Stroma in the Tumor Microenvironment. Cancers (Basel) 2019, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Pelizzari, G.; Basile, D.; Zago, S.; Lisanti, C.; Bartoletti, M.; Bortot, L.; Vitale, M.G.; Fanotto, V.; Barban, S.; Cinausero, M.; Bonotto, M. Lactate dehydrogenase (LDH) response to first-line treatment predicts survival in metastatic breast cancer: first clues for a cost-effective and dynamic biomarker. Cancers 2019, 11, 1243. [Google Scholar] [CrossRef] [PubMed]
- Bouten, R.M.; Dalgard, C.L.; Soltis, A.R.; Slaven, J.E.; Day, R.M. Transcriptomic profiling and pathway analysis of cultured human lung microvascular endothelial cells following ionizing radiation exposure. Scientific Reports 2021, 11, 24214. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Distel, L.V.R.; Neuhuber, W. Caffeic Acid, Quercetin and 5-Fluorocytidine-Functionalized Au-Fe3O4 Nanoheterodimers for X-ray-Triggered Drug Delivery in Breast Tumor Spheroids. Nanomaterials 2021, 11, 1167. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Y.; Qin, Z.; Guo, H.; Chen, W.; Ding, T. Stereotactic Body Radiotherapy and Conventional Radiotherapy Induce Cytoskeleton Extension and Enlargement of Cell Morphology in Non-Small Cell Lung Cancer. Dose-Response 2021, 19, 15593258211064499. [Google Scholar] [CrossRef]
- Otsuka, S.; Shibamoto, Y.; Iwata, H.; Murata, R.; Sugie, C.; Ito, M.; Ogino, H. Compatibility of the linear-quadratic formalism and biologically effective dose concept to high-dose-per-fraction irradiation in a murine tumor. International Journal of Radiation Oncology Biology Physics 2011, 81, 1538–1543. [Google Scholar] [CrossRef]
- Zhang, H.; Wan, C.; Huang, J.; Yang, C.; Qin, Y.; Lu, Y.; Ma, J.; Wu, B.; Xu, S.; Wu, G.; Yang, K. In vitro radiobiological advantages of hypofractionation compared with conventional fractionation: early-passage NSCLC cells are less aggressive after hypofractionation. Radiation Research 2018, 190, 584–595. [Google Scholar] [CrossRef]
- Scaccianoce, E.; Festuccia, C.; Dondi, D.; Guerini, V.; Bologna, M.; Motta, M.; Poletti, A. Characterization of prostate cancer DU145 cells expressing the recombinant androgen receptor. Oncology Research 2003, 14, 101–112. [Google Scholar] [CrossRef]
- Tai, S. , Sun, Y. , Squires, J.M., Zhang, H., Oh, W.K., Liang, C.Z. and Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. The Prostate 2011, 71, 1668–1679. [Google Scholar]
- Moya, L.; Walpole, C.; Rae, F.; Srinivasan, S.; Seim, I.; Lai, J.; Nicol, D.; Williams, E.D.; Clements, J.A.; Batra, J. Characterisation of cell lines derived from prostate cancer patients with localized disease. Prostate Cancer and Prostatic Diseases 2023, 26, 614–624. [Google Scholar] [CrossRef]
- Vidal, M.A.; Kilroy, G.E.; Johnson, J.R.; Lopez, M.J.; Moore, R.M.; Gimble, J.M. Cell growth characteristics and differentiation frequency of adherent equine bone marrow-derived mesenchymal stromal cells: adipogenic and osteogenic capacity. Veterinary Surgery 2006, 35, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Cha, S.H.; Kang, H.W.; Song, J.Y.; Lee, K.W.; Ko, K.B.; Lee, H.T. Effects of serial passage on the characteristics and chondrogenic differentiation of canine umbilical cord matrix derived mesenchymal stem cells. Asian-Australasian Journal of Animal Sciences 2013, 26, 588. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. In vitro assays and techniques utilized in anticancer drug discovery. Journal of Applied Toxicology 2019, 39, 38–71. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.J. The linear quadratic model: Usage, interpretation and challenges. Physics in Medicine and Biology 2018, 64, 0–24. [Google Scholar] [CrossRef]
- Dale, R.; Carabe-Fernandez, A. The radiobiology of conventional radiotherapy and its application to radionuclide therapy. Cancer Biology and Radiopharmaceuticals 2005, 20, 47–51. [Google Scholar] [CrossRef]
- Nair, S.; Cairncross, S.; Miles, X.; Engelbrecht, M.; du Plessis, B.; Bolcaen, J.; Fisher, R.; Ndimba, R.; Cunningham, C.; Martínez-López, W.; Schunck, C. An automated microscopic scoring method for the γ-H2AX foci assay in human peripheral blood lymphocytes. Journal of Visualized Experiments (JoVE) 2021, 178, e62623. [Google Scholar] [CrossRef]
- Panek A, Miszczyk J. DNA repair processes in human lymphocytes irradiated with a 60-MeV proton radiotherapeutic beam. RAD Conference Proceedings 2019, 3, 10–14.
- Jayakumar, S.; Kunwar, A.; Sandur, S.K.; Pandey, B.N.; Chaubey, R.C. Differential response of DU145 and PC3 prostate cancer cells to ionizing radiation: Role of reactive oxygen species, GSH and Nrf2 in radiosensitivity. Biochimica et Biophysica Acta (BBA) - General Subjects 2014, 1840, 485–494. [Google Scholar] [CrossRef]
- Lövey, J.; Nie, D.; Tóvári, J.; Kenessey, I.; Tímár, J.; Kandouz, M.A.; Honn, K.V. Radiosensitivity of human prostate cancer cells can be modulated by inhibition of 12-lipoxygenase. Cancer Letters 2013, 335, 495–501. [Google Scholar] [CrossRef]
- Reda, M.; Bagley, A.F.; Zaidan, H.Y.; Yantasee, W. Augmenting the therapeutic window of radiotherapy: A perspective on molecularly targeted therapies and nanomaterials. Radiotherapy and Oncology 2020, 150, 225–235. [Google Scholar] [CrossRef]
- Cui, M.; Gao, X.; Lyu, F.; Sun, D. Mechanism of the DNA Damage Repair Gene Work on Elevated α/β Value after Hypofractionated Radiotherapy for Prostate Cancer. International Journal of Radiation Oncology Biology Physics 2019, 108, e526. [Google Scholar] [CrossRef]
- Okazaki, R. Role of p53 in Regulating Radiation Responses. Life 2022, 12, 1099. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhong, R.; Sun, L.; Jia, J.; Ma, S.; Liu, X. Ionizing radiation-induced adaptive response in fibroblasts under both monolayer and 3-dimensional conditions. PLoS One 2015, 10, e0121289. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.C.; Nguyen, E.V.; Niranjan, B.; Wu, Y.; Lim Kam Sian, T.C.; Horvath, L.G.; Taylor, R.A.; Daly, R.J. Cell-type-specific signaling networks impacted by prostate epithelial-stromal intercellular communication. Cancers 2023, 15, 699. [Google Scholar] [CrossRef]
- Torabinejad, S.; Soleymanifard, S.; Sayyah, S.; Behnam Rasouli, F. High-dose Irradiation Stimulated Breast Tumor Microenvironment to Enhance Tumor Cell Growth and Decrease Tumor Cell Motility. Journal of Biomedical Physics and Engineering 2023, 13, 209–216. [Google Scholar]
- Tang, X.; Cappa, T.; Kuhlenschmidt, T.; Kuhlenschmidt, M.; Saif, T. Specific and Non-Specific Adhesion in Cancer Cells with Various Metastatic Potentials BT - Mechanobiology of Cell-Cell and Cell-Matrix Interactions. In: Wagoner Johnson A, Harley BAC, editors. Boston, MA: Springer US; 2011: 105–22. [CrossRef]
- Sideri, S. , Petragnano, F. , Maggio, R., Petrungaro, S., Catizone, A., Gesualdi, L., De Martino, V., Battafarano, G., Del Fattore, A., Liguoro, D. and De Cesaris. Radioresistance Mechanisms in Prostate Cancer Cell Lines Surviving Ultra-Hypo-Fractionated EBRT: Implications and Possible Clinical Applications. Cancers 2022, 14, 5504. [Google Scholar]
- Chang, L.; Graham, P.H.; Hao, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. Emerging roles of radioresistance in prostate cancer metastasis and radiation therapy. Cancer and Metastasis Reviews 2014, 33, 469–496. [Google Scholar] [CrossRef]
- Fang, Y.; DeMarco, V.G.; Nicholl, M.B. Resveratrol enhances radiation sensitivity in prostate cancer by inhibiting cell proliferation and promoting cell senescence and apoptosis. Cancer Science 2012, 103, 1090–1098. [Google Scholar] [CrossRef]
Figure 1.
Analysis of prostate cancer cell lines to evaluate (a) repair efficiency following 2 Gy radiation exposure, assessed by the number of foci formations at various time intervals; and (b) repair factor determined from 2 Gy split-dose experiments utilising the clonogenic survival assay.
Figure 1.
Analysis of prostate cancer cell lines to evaluate (a) repair efficiency following 2 Gy radiation exposure, assessed by the number of foci formations at various time intervals; and (b) repair factor determined from 2 Gy split-dose experiments utilising the clonogenic survival assay.
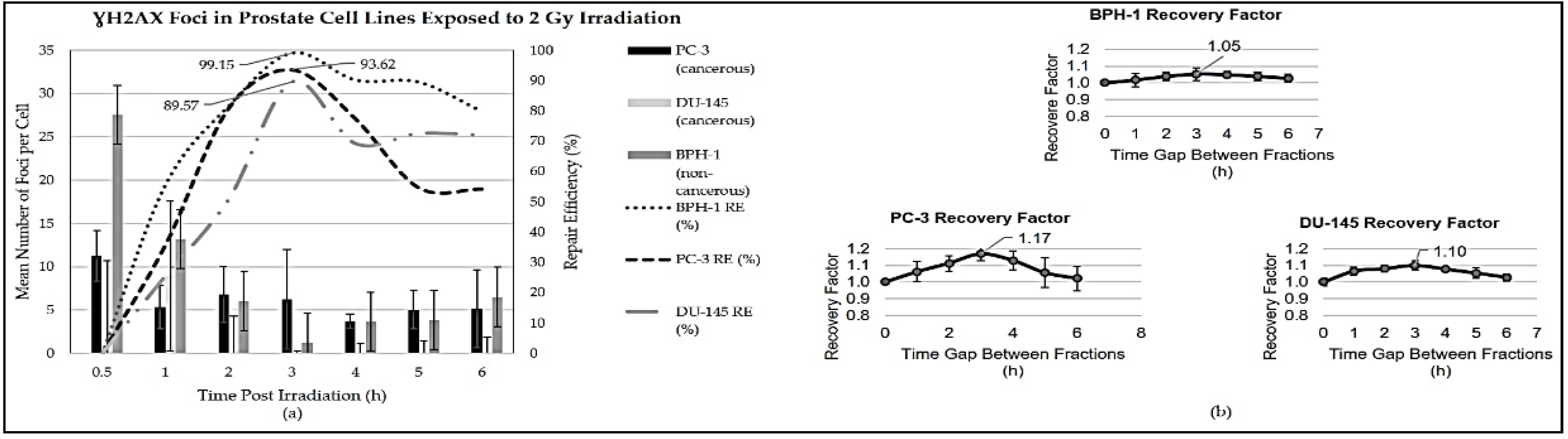
Figure 2.
Clonogenic survival comparisons between the two fractionation schemes for the prostate cell lines with and without a prime dose prior to fractionation.
Figure 2.
Clonogenic survival comparisons between the two fractionation schemes for the prostate cell lines with and without a prime dose prior to fractionation.
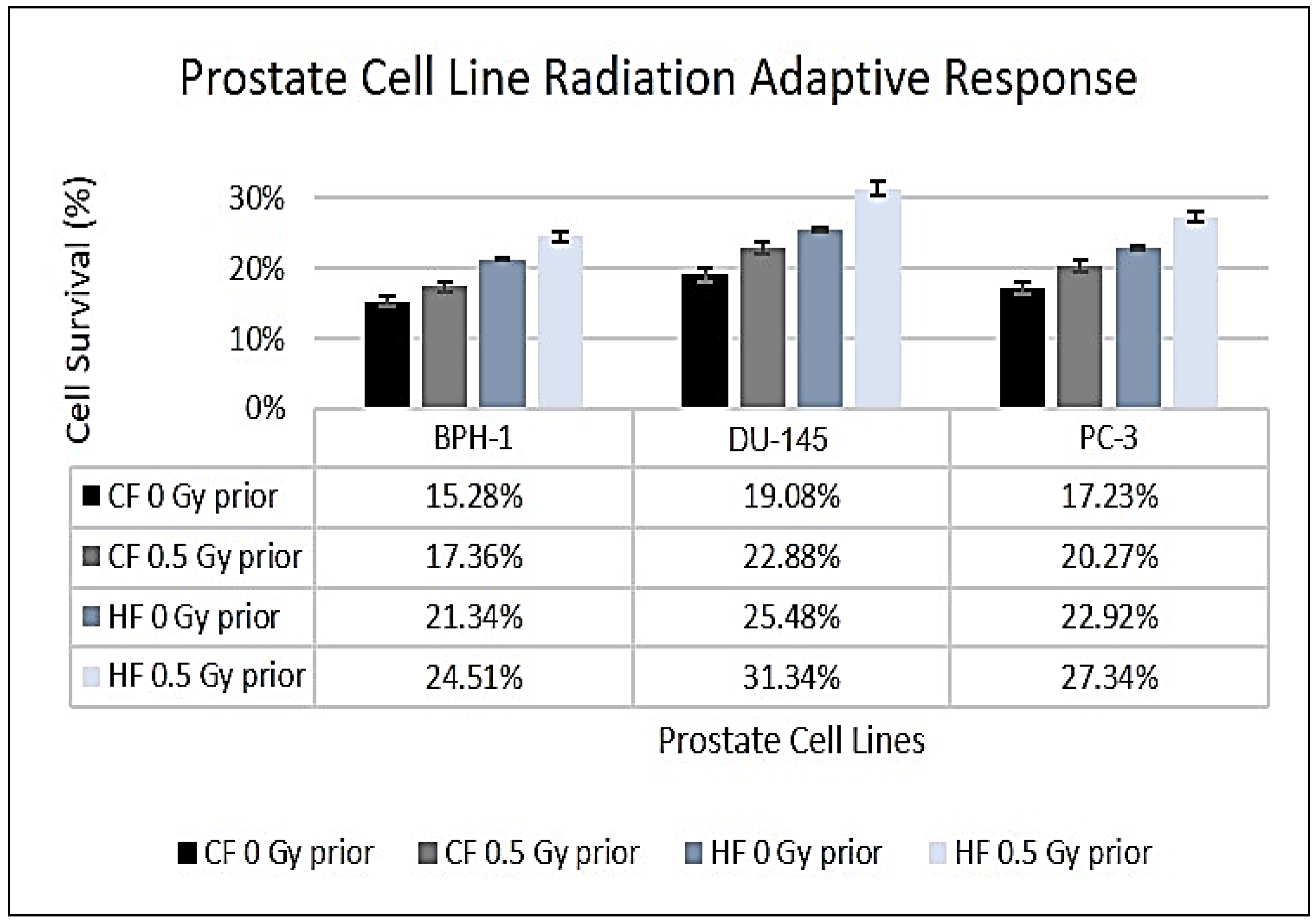
Figure 3.
BPH-1 cell line () migration with error bars showing the standard deviation (SD) of each measurement and (b) cancerous prostate cell lines invasion capacity with error bars showing the standard deviation (SD) of each measurement.
Figure 3.
BPH-1 cell line () migration with error bars showing the standard deviation (SD) of each measurement and (b) cancerous prostate cell lines invasion capacity with error bars showing the standard deviation (SD) of each measurement.
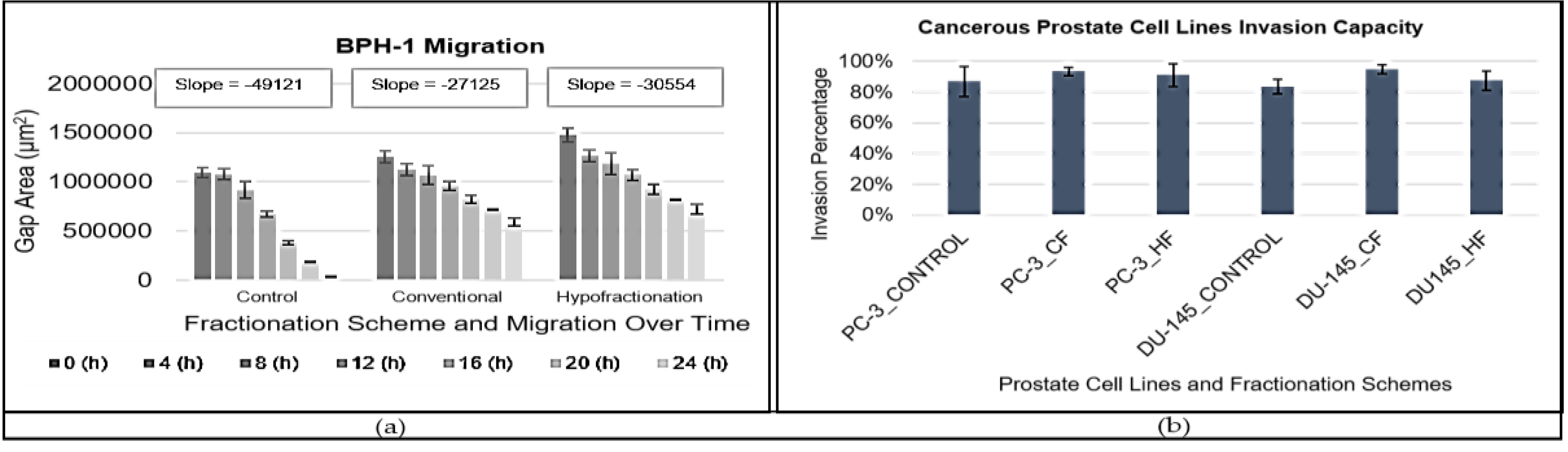
Figure 4.
Percentage of lactate dehydrogenase (LDH) release from three different prostate cell lines following treatment with hypofractionated and conventional fractionated radiation. Results are presented as mean ± standard error of the mean (SEM).
Figure 4.
Percentage of lactate dehydrogenase (LDH) release from three different prostate cell lines following treatment with hypofractionated and conventional fractionated radiation. Results are presented as mean ± standard error of the mean (SEM).
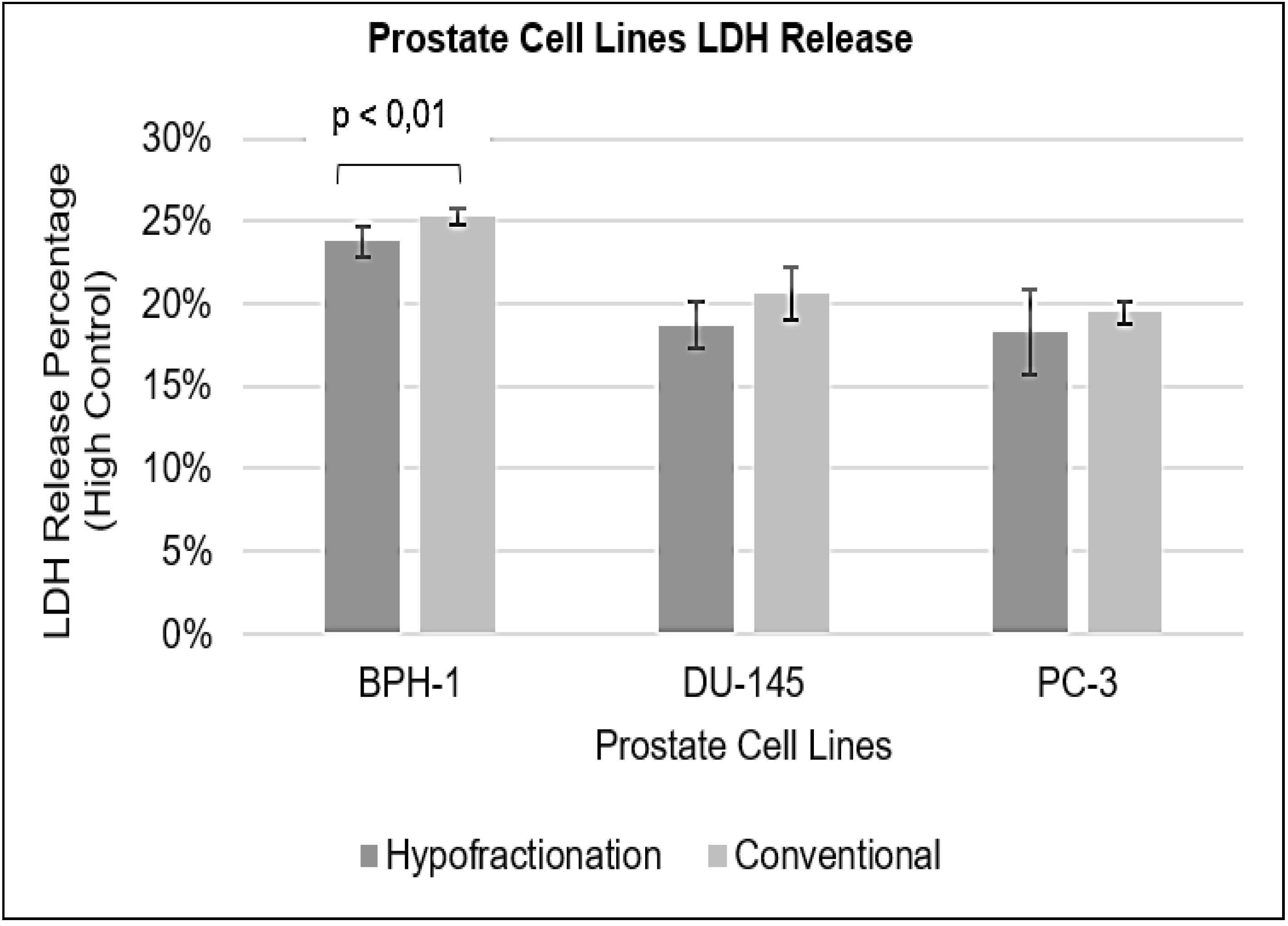

Table 1.
Prostate cell line doubling time comparison.
| Cell Line (Breast) |
Doubling Time (h) | Standard Deviation | Adaptive Doubling Time (h) | Standard Deviation (h) | p-value |
|---|---|---|---|---|---|
| BPH-1 | 35.21 | 5.80 | 37.61 | 2.30 | p = 0.235 |
| PC-3 | 36.20 | 2.80 | 40.63 | 1.30 | p = 0.572 |
| DU-145 | 18.40 | 2.90 | 18.83 | 0.30 | p = 0.857 |
Table 2.
Radiobiological parameters and dose-response characteristics of prostate cell lines.
| Cell Line (Prostate) |
α (Gy−1) | β (Gy−2) | α/β Ratio | D50 (Gy) | P-value | RS | Conventional Fractionation | Hypofractionation | Biological Effective Dose (BED) |
|---|---|---|---|---|---|---|---|---|---|
| BPH-1 (non-cancerous) |
0.08 ± 0.01 | 0.03 ± 0.00* | 2.7 ± 0.80 | 3.66 ± 0.10 | - | - | 2.00 Gy x 4 = 8.00 Gy with 3 hours gap | 4.93 Gy x 1 = 4.93 Gy | 13.93 Gy |
| PC-3 (cancerous) |
0.12 ± 0.00* | 0.03 ± 0.00* | 3.7 ± 0.90 | 3.21 ± 0.00* | 0.447 | 1.14 ± 0.00* | 2.00 Gy x 4 = 8.00 Gy with 3 hours gap | 5.15 Gy x 1 = 5.15 Gy | 14.67 Gy |
| DU-145 (cancerous) | 0.10 ± 0.00* | 0.04 ± 0.00* | 2.4 ± 0.80 | 3.10 ± 0.00* | 0.794 | 1.18 ± 0.00* | 2.00 Gy x 4 = 8.00 Gy with 3 hours gap | 4.85 Gy x 1 = 4.85 Gy | 12.32 Gy |
* Indicates values < 0.001.
Table 3.
Comparison of intrinsic and single-dose Survival rates for the prostate cell lines.
| Cell Lines | Single-Dose (Gy) | Intrinsic Survival (%) | Hypofractionation Survival (%) | p-value |
|---|---|---|---|---|
| BPH-1 | 4.93 | 32.5 ± 3.5 | 21.3 ± 4.0 | 0.0012 |
| PC-3 | 5.15 | 24.3 ± 3.1 | 25.5 ± 2.0 | 0.5612 |
| DU-145 | 4.85 | 24.0 ± 4.0 | 22.9 ± 2.0 | 0.3148 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



