
Submitted:
29 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
This review explores recent advances in the design of fermentation processes for producing alternative proteins, with a focus on the utilization of agro-industrial waste and renewable substrates. Key bioprocess strategies, such as optimized bioreactor designs and strict fermentation parameter control, are discussed to enhance substrate conversion efficiency into protein biomass. The role of enzyme technologies in improving digestibility and protein quality is also highlighted. The article underscores the importance of developing cost-effective fermentation solutions that minimize environmental impact while addressing technical challenges such as scalability and contamination control. Furthermore, it emphasizes the growing need for innovations in fermentation process design to ensure the sustainability of industrial protein production. The review concludes that improvements in process design are fundamental to overcoming technological and regulatory barriers, particularly in increasing the efficiency and competitiveness of non-meat proteins in the global market.
Keywords:
food alternatives
; precision fermentation
; non‐animal protein
; protein enzymatic hydrolysate
; single‐cell protein
1. Introduction
The growing global demand for non-animal alternative proteins, driven by environmental, ethical, and health concerns, has led the food industry to develop innovative and sustainable solutions. The production of meat and dairy products from animal sources requires high levels of natural resources, such as fresh water and land extensions, in addition to being directly associated with the emission of greenhouse gases (GHG). Many of these emissions come from enteric fermentation in ruminants, such as cows, which release methane (CH₄), a gas with a global warming potential 25 times greater than carbon dioxide (CO₂) over 100 years.[1].
It is estimated that each cow can release 500 liters of methane per day, which contributes significantly to the environmental impact of livestock farming [2]. In addition, manure management also emits methane and nitrous oxide, and deforestation for pasture and feed production reduces carbon sequestration capacity. The use of fossil fuels in transporting and processing these raw materials also contributes to further increasing CO₂ emissions. Studies show that the environmental impact of beef production includes the emission of 10.4 kg of CO₂ equivalent per kilogram of live weight, with feed production having the greatest impact on these emissions.[3]. Thus, these ambiental problems have increased the search for more environmentally friendly alternatives, such as protein from insects and microorganisms, which usually present a minor cost of production, consumption of natural resources, such as water, and space for production [4].
On the other hand, microbial protein production, also known as “single-cell protein,” uses microorganisms such as yeast, fungi, and bacteria to convert substrates such as sugars and agricultural waste into nutrient-rich protein biomass. This technology offers several advantages over animal-based protein production, including greater resource efficiency, rapid growth of microorganisms, and reduced environmental impact, especially regarding greenhouse gas emissions and land use. In addition, microorganisms can metabolize various substrates, from glucose derived from plant crops to more complex agro-industrial waste such as sugarcane bagasse or fruit peels, promoting sustainability and waste reduction [5].
Growing awareness of the environmental impact of meat production, combined with concerns about animal health and welfare, has contributed to a decrease in meat consumption by the population. This trend has stimulated the search for alternative protein sources, such as microbial or plant-based proteins, which can be a viable and sustainable solution to meet the demand for proteins without the exact environmental and health costs associated with conventional meat production [6,7].
Microorganisms and enzymes also present a high substrate conversion to biomass, reaching around 50% of the raw material converted into microbial protein [8]. When comparing animal protein sources, it can be seen that one kilo of beef contains approximately 250 grams of protein, while chicken meat has around 220 grams per kilo, pork around 210 grams, and lamb or sheep meat around 250 grams per kilo. In contrast, microbial protein production shows significant advantages, especially in substrate conversion efficiency, process speed, and reduced environmental impacts. The yield of the current biotechnological system is already promising; however, there remains significant potential for further optimization through advancements in fermentation processes and genetic engineering. By selecting and genetically modifying microorganisms, we can enhance nutrient efficiency and increase protein accumulation. Furthermore, implementing bioprocess optimization strategies—such as controlling pH, temperature, and bioreactor configurations—will maximize overall yield [9].
Despite technological advances, several challenges must be overcome to enable the large-scale adoption of these technologies involving the biotechnological production of non-meat proteins. The selection of microbial strains that produce proteins with sensory and functional properties comparable to proteins of animal origin is one of the main challenges. Advances in synthetic biology and genetic engineering have allowed customized strains to develop specific proteins with nutritional and organoleptic profiles suitable for food applications [10]. However, using genetically modified strains depends on regulatory issues and public acceptance. In markets such as the European Union, for example, regulations on the use of genetically modified organisms (GMOs) are more restrictive, which limits the introduction of these products into the market. At the same time, acceptance has been more positive in the United States, with a significant portion of North American consumers willing to try products based on fermented proteins [11].
In addition to microorganisms, using enzymes is an essential tool in optimizing fermentation processes and in post-production modification of proteins. Proteolytic enzymes, such as papain, bromelain, and pepsin, can be used strategically to develop alternative proteins to adjust the amino acid profile and improve protein digestibility [12]. These enzymes act by breaking peptide bonds in protein chains, converting them into smaller peptides or free amino acids, which can facilitate digestion in the human body, optimizing the nutritional value of proteins and making them more suitable to meet the specific needs of different populations, such as athletes or individuals with dietary restrictions.
Soy or pea-based products, for example, are widely used in the production of alternative proteins. Proteolytic enzymes can be applied to reduce the presence of anti-nutritional factors, such as trypsin inhibitors, which can interfere with nutrient absorption. Protein hydrolysis carried out by enzymes can increase the availability of essential amino acids, improving the nutritional quality of the product. This way, the amino acid profile becomes more similar to animal proteins, offering a more nutritionally complete alternative [13].
Additionally, proteolytic enzymes can modify the functionality of proteins regarding solubility, viscosity, and foaming ability, which is particularly useful in products such as plant-based protein drinks and nutritional supplements [14]. In meat analogs such as plant-based burgers and sausages, these enzymes can improve texture and mouthfeel, providing a more tender and juicy sensation, desirable characteristics that increase consumer acceptance. Crosslinking enzymes such as transglutaminase have been widely used in producing alternative proteins due to their ability to catalyze the formation of covalent bonds between protein chains, creating a more stable protein network [15]. This process significantly improves the texture of products such as vegetable burgers, meatballs, and sausage analogs, providing greater firmness and elasticity to the structure, fundamental characteristics to replicate the fibrous and succulent texture found in meats of animal origin [16].
Although many studies have shown promising laboratory-scale results, the transition to large-scale production in industrial-scale bioreactors often declines efficiency. The use of renewable and low-cost substrates, such as agricultural or industrial waste, can significantly reduce production costs and environmental impact. Recent innovations in fermentation include precision fermentation technologies, which combine genetic engineering with computational modeling to engineer microorganisms capable of producing proteins with specific characteristics. [17,18,19]. Computational modeling simulates the metabolic behavior of microorganisms, identifying the most efficient genetic modifications to optimize protein production. In this way, it is possible to adjust properties such as flavor, texture, and solubility, essential for developing meat and dairy substitutes that meet nutritional and sensory expectations [20]. This process also speeds up development, allowing modifications to be tested before being applied in the laboratory, which reduces time and costs. However, there are still challenges to be overcome, such as the high cost of implementing these technologies and regulatory acceptance, which may limit their commercial adoption.
Given these challenges, it is essential to review the current state of non-animal protein production by fermentation to identify how technological innovations can be applied practically and efficiently. This article aims to explore the main innovations and challenges in the optimization of fermentation processes, with an emphasis on scalability, sustainability, and critical evaluation of the engineering configurations involved. It seeks to improve efficiency, reduce costs, and ensure industrial viability.
2. Alternative to Meat Proteins
Interest in alternative proteins to meat has grown significantly, driven by environmental and health concerns. Livestock production is responsible for a high percentage of global greenhouse gas emissions. It requires large amounts of land and water as natural resources, reinforcing the urgent need for more sustainable alternatives. In this context, five main types of alternative proteins to meat stand out both for their technological feasibility and environmental benefits: plant-based proteins, insect proteins, cultured meat, single-cell proteins, and protein compounds obtained from enzymatic hydrolysis, the latter two standing out as promising biotechnological routes for obtaining these proteins. Figure 1 presents the main methods for obtaining non-meat proteins. These categories encompass different protein sources that have increasingly attracted the attention of the food industry and researchers due to their potential for reducing the environmental impact of conventional protein production [21]. These categories encompass different protein sources that have increasingly attracted the attention of the food industry and researchers due to their potential for reducing the environmental impact of conventional protein production [21].
Plant proteins have gained prominence, with products based on soy, peas, and other legumes widely used in the formulation of meat analogs. Plant proteins are commonly used in human and animal food formulations as a functional ingredient. Proteins from legume seeds can improve food formulations' nutritional quality, technological functionality, and nutraceutical properties. Most research has focused on legume seeds such as soybeans, peas, and beans, but other non-commercial legume seeds used only for animal feed have received little attention [22]. According to Meticulous Research (2022), the plant-based protein market is expected to reach US$23.4 billion by 2028, with the main drivers being the growing demand for protein-rich diets, the appeal to health and well-being, the increased search for alternatives to conventional animal products and the consequent need for the food and beverage industry to provide options to consumers. Vegetables and cereals with high protein content, such as soy, wheat, lentils, and chickpeas, can be used to make plant-based products.
Like unconventional plants, leafy vegetables are recognized as low-cost protein sources due to their ability to synthesize amino acids using sunlight, carbon dioxide, and atmospheric nitrogen. Although these vegetables have high protein content, their use in the food industry is still limited due to the lack of understanding of their functional and structural properties. These properties, such as solubility, water or oil absorption capacity, emulsification, and foaming or gel formation capacity, are essential to determine their specific applications in food formulation [23].
Protein extracted from soy, rich in essential amino acids, is one of the most versatile sources of vegetable proteins and is often compared to proteins of animal origin, which makes it an excellent option for the formulation of meat analogs. Soy products, such as tofu, soy milk, tempeh, and textured soy, are widely consumed as substitutes for meat proteins, particularly in vegetarian diets [6]. In addition to their nutritional contribution, soy proteins have important functional properties, such as solubility, water and oil absorption, and emulsification capacity, which are essential in the formulation of processed foods [24].
These plant-based protein foods can replicate animal meat's texture, flavor, and nutritional profile, providing a similar sensory experience but reducing environmental impact. Compared to beef, studies show that plant-based proteins generate up to 90% fewer greenhouse gas emissions and require fewer natural resources, making them a sustainable and attractive option for human nutrition [25]. Non-meat eaters, such as vegans, seek protein alternatives to maintain essential functions, such as growth, repair, and maintenance of body tissues. On the other hand, adopting a plant-based diet is motivated by factors such as ethics, sustainability, and health. Ethical reasons related to concern for animal welfare drove veganism and still represent the main reason for changing eating patterns and lifestyles [26]. In its current production model, livestock farming has been identified as one of the leading causes of climate change and environmental imbalances observed in recent years. In addition, prevention and health maintenance are another justification for adopting a dietary pattern based on plant-based proteins since studies demonstrate a strong association between plant-based diets and a reduced risk of chronic non-communicable diseases [27].
Insect proteins have also gained prominence due to their high protein content. Insects such as crickets and mealworms are particularly valued because they are rich in protein and can be farmed with less environmental impact, using less water and space than conventional livestock farming. It is estimated that more than US$1 billion has already been invested in the European insect industry, with promising results. In 2019, more than 6,000 tons of insect proteins were produced, and the global edible insect market is expected to grow at an annual rate of 26.5%, reaching a value of US$4.63 billion by 2027. Even higher projections indicate that the market could reach US$8 billion by 2030 [28].
Including insects in the diet can be nutritionally positive, as they provide high levels of essential amino acids, highly digestible proteins, unsaturated fatty acids, and micronutrients such as iron and calcium. In addition, they can improve the intestinal microbiota and have antioxidant properties. Despite the environmental and nutritional benefits, the acceptance of insects as a source of protein still faces resistance, mainly due to rejection, lack of custom, and concerns about food safety [29]. With the projected increase in global food demand, edible insects are emerging as a sustainable solution, as they are rich in proteins, antioxidants, prebiotics, and polyphenols, with potential health benefits. Consumed by more than two billion people, they can be eaten whole or processed. Still, food safety issues, such as allergenicity, toxins, and appropriate regulation, are essential to ensure quality and increase acceptance [30].
The use of insects as a source of protein has proven to be a sustainable alternative to global food production challenges. One example is the larva of Tenebrio molitor (yellow mealworm), which has stood out as a rich source of protein, with a protein content ranging from 41% to 66% (dry basis), in addition to a profile of essential amino acids, such as leucine, isoleucine, and methionine. This makes these insects an excellent option for enriching traditional food products, significantly increasing their nutritional value. Tenebrio molitor is also rich in fatty acids, with a lipid content ranging from 15% to 50%, and is composed mainly of monounsaturated and polyunsaturated fatty acids, such as oleic and linoleic acids. In addition to their high nutritional value, these insects have technological benefits. For example, incorporating mealworm powder into foods, such as bread and protein bars, increases the protein content and influences the final product's sensory and technological characteristics [31].
Cultured meat, in turn, represents an emerging innovation that has attracted considerable interest from both industry and research. Cultured meat is produced from animal cells grown in controlled environments to form muscle tissue. Although it still faces challenges related to scale and cost, this technology offers an alternative to conventional meat that uses less land and water and eliminates the need for animal slaughter. Several types of proteins can be used to produce these foods (Table 1), such as soy, fungal, and algal proteins, which can be used as a substrate for the production of proteins, such as basmati rice, which can be used to produce fungal proteins [32], or the generation of composites, such as hydrogels that simulate meat and can be consumed, like hamburgers. Although advances are still needed in flavor and texture, sensory analyses have demonstrated these products' high potential as meat substitutes [33].
Fu et al. [4] investigated the acceptance of cultured meat as a sustainable substitute for conventional meat. They found that the main facilitators of acceptance are perceived health benefits, animal welfare, and food quality. On the other hand, neophobia toward food technology emerged as a significant inhibitor, while the perception of the “unnaturalness” of cultured meat did not have a relevant impact on product acceptance. Thus, promoting health and environmental benefits and mitigating technological concerns may increase the acceptance of cultured meat as a viable and sustainable alternative to conventional meat. Choudhary et al. [38] investigated the perception and acceptance of cultured meat among meat scientists in India. The study revealed that only 25% of scientists support the development of cultured meat, with 38% willing to try it at least once, although only 6% are willing to buy it regularly or pay a higher price. Among the barriers identified, technology neophobia (21%) and high cost (19%) were highlighted, while the perception of “unnaturalness” was relevant for 46% of the respondents. Although they recognize environmental and animal welfare benefits, most scientists still believe there is insufficient scientific evidence on the safety and environmental impacts of cultured meat, and they consider a rapid transition to this type of production risky.
Single-cell proteins — produced by microorganisms such as yeast, fungi, and bacteria — have also proven to be a promising alternative in the new protein market. These proteins, known as Single Cell Protein (SCP), can be cultivated in bioreactors using a variety of substrates, including agro-industrial waste. Genetic engineering enables the development of microbial strains that produce proteins with adjusted nutritional profiles, adapting the amino acid composition to human needs. Biotechnology promotes the production of proteins efficiently and with less environmental impact, contributing to the circular economy by taking advantage of waste that would otherwise be discarded.
SCP is produced from microorganisms such as algae, yeast, and bacteria and can be used as a rich source of protein, unsaturated fatty acids, vitamins, and minerals, a high-quality supplement for animal feed. The main raw materials for fermentation include agro-industrial by-products such as molasses and corn liquor, which are economical and nutrient-rich substrates. SCP has a high protein content, ranging from 30% to 80%, depending on the microorganism used, such as bacteria that can reach up to 80% protein. SCP can also be used for animal weight gain, such as sea bass growth and chickens' feed intake [39].
A study examined the production of SCP from various types of industrial food waste, including potato, banana, orange, and carrot peels. It was found that potato peels yielded the highest amount of SCP, particularly when incubated at 30°C for five days. The incorporation of nitrogen sources, such as ammonium sulfate, enhanced the yield by as much as 58%. Proximal analysis revealed that SCP has a protein content of 47.78%, significantly greater than wheat flour (12.47%). Furthermore, SCP contains high levels of essential amino acids, such as lysine (3.14%) and leucine (4.35%), which are considerably elevated compared to the amounts found in wheat flour. Sensory tests indicated that up to 4% of SCP added to enriched bread was well accepted by consumers, highlighting the potential of SCP as a sustainable source of protein to enhance foods and contribute to food security [40].
Zhu et al. [41] studied the production of SCP using photosynthetic bacteria fed with food waste fermentation liquids. The study demonstrated that SCP production could be significantly increased, with a yield of 2088.4 mg/L, using 20% of the fermentation liquid as the carbon source. The bacteria preferred volatile fatty acids, especially acetic acid, which accounted for 54.8% to 67.6% of the carbon consumed. Regulation of the carbon source also enhanced the metabolic pathways involved in SCP biosynthesis, such as the tricarboxylic acid (TCA) cycle and the conversion of acetate to acetyl-CoA. However, excess fermentation liquid (40%) inhibited bacterial growth and SCP production, indicating that a high organic load may harm cellular activity. These results suggest optimizing carbon source regulation is essential to maximize SCP production and provide a sustainable alternative for food waste valorization.
Bertasini et al. [42] explored the production of single-cell proteins (SCP) using sugar-rich effluent from candy production and nutrient-rich agricultural digestate as substrates for the growth of Saccharomyces cerevisiae. In continuous tests, a two-day hydraulic retention time (HRT) with a dilution rate of 0.50 d⁻¹ yielded the highest SCP productivity of 0.25 g/L per day and a protein content of 28% (w/w). The amino acid profile demonstrated that the obtained SCPs can be used as supplements in fish and monogastric animal feed but are unsuitable for pet food due to insufficient amino acid levels. This study suggests that agricultural anaerobic digestates can be transformed into biorefineries to produce microbial proteins.
The use of proteolytic enzymes is also a promising approach to developing alternative proteins, particularly for their role in improving digestibility and optimizing the amino acid profile of these proteins. These enzymes increase nutritional value by converting low-value by-products into functional ingredients, expanding food and nutrition industry applications. Enzymes such as papain, bromelain, Alcalase®, and others have been widely investigated due to their ability to modify protein structures and generate hydrolysates with valuable functional properties, such as antioxidant and antimicrobial activity.
Oliveira Filho et al. [43] studied using proteolytic enzymes to produce bioactive protein hydrolysates from cottonseed by-products. Three commercial enzymes were used: Alcalase®, Neutrase®, and Flavourzyme®, with additional heat treatment to increase the degree of hydrolysis. The study revealed that Alcalase® produced the best results, with superior antioxidant activity and angiotensin-converting enzyme (ACE) inhibition of up to 99.5%. In addition, the hydrolysates showed antimicrobial properties, inhibiting the growth of Colletotrichum gloeosporioides and Staphylococcus aureus by up to 73.89% and 32.38%, respectively. These results demonstrate that appropriate enzymes, combined with heat treatment, can convert low-value by-products into functional ingredients with antioxidant, antimicrobial, and antihypertensive potential.
Kopplin et al. [44] investigated the production of antioxidant hydrolysates from bovine sodium caseinate (NaCAS) and soy protein isolate (SPI) using three non-commercial proteases from Bacillus sp. CL14, CL18 and CL33A. The study showed that the proteases successfully hydrolyzed both substrates, increasing their antioxidant activities, such as radical scavenging capacity, Fe²⁺ chelating capacity, and reducing power. The highest antioxidant activities were observed after 2 to 3 hours of hydrolysis. For NaCAS hydrolysates, the CL14 protease showed the best performance, followed by CL18 and CL33A. CL14 also showed the highest antioxidant activity for SPI hydrolysates, with similar results between CL33A and CL18. These findings highlight the potential application of these proteases and hydrolysates in food and feed biotechnology.
Prikhodko et al. [45] obtained functional ingredients from casein and gluten protein fractions to develop hypoallergenic mixtures with functional properties and balanced amino acid composition. The research used enzymes such as Protex 6L, chymotrypsin, and pancreatin to hydrolyze gliadin, glutenin, and casein proteins. The resulting hydrolysates showed good emulsifying and foaming capacity, and the mixture of casein and glutenin hydrolysates (ratio 2:1) had a high amino acid score, making it promising for developing hypoallergenic ingredients in functional foods.
It is essential to highlight that the growing interest in sustainable food solutions has driven a significant expansion in research on alternative proteins, as evidenced in the data presented. Since 2015, there has been an increase in publications on plant-based proteins, reflecting the interest in plant-based diets and the concern about the environmental impacts of livestock farming (Figure 2A). Likewise, insect proteins have been increasingly investigated since 2010, consolidating themselves as a sustainable and efficient source (Figure 2B). In the field of cultivated meat, biotechnological advances and the search for alternatives to animal slaughter have intensified research since 2010 (Figure 2C). Regarding single-cell proteins, such as yeast and microalgae, the growing potential of this segment, especially for the use of agro-industrial waste, has been highlighted since 2015 (Figure 2D). Finally, enzymatic protein hydrolysates, focusing on the nutritional and functional improvement of proteins via enzymes, have gained relevance since the 2000s (Figure 2E).
Livestock production is responsible for a high percentage of global greenhouse gas emissions. It requires large amounts of land and water as natural resources, reinforcing the urgent need for more sustainable alternatives. In this context, five main types of alternative proteins to meat stand out both for their technological feasibility and environmental benefits: plant-based proteins, insect proteins, cultured meat, single-cell proteins, and protein compounds obtained from enzymatic hydrolysis; the latter two stand out as promising biotechnological routes for obtaining these proteins. These advances demonstrate the growing importance of alternative proteins in scientific research and the food industry, driven by the search for more sustainable solutions. This review addresses biotechnological advances in the production of proteins and protein ingredients, highlighting processes such as the production of single-cell proteins and the use of proteolytic enzymes. The work presents the leading technologies employed, highlighting the innovative methods shaping the future of food production, with the potential to meet the growing demand for efficient and sustainable protein sources.
In Table 2, several substrates produce different protein non-meat-based foods. It is possible to observe the usage of microorganisms and enzymes and the application of sensory analysis that may infer that they are suited to produce foods. However, improvements in flavor and texture may enhance their potential by simulating the characteristics of meats.
3. Biotechnological Configurations and Processes for Protein Production
3.1. Fermentation Processes for Single-Cell Protein Production
Protein production through fermentation processes requires significant technological advances, especially in bioreactor design and strict control of fermentation parameters, which may differ between the applied microorganisms (Figure 3). Microorganisms, such as fungi and bacteria, may be used to convert agro-industrial waste into protein biomass, but process optimization still requires challenges.
Van Peteghem et al. [46] produced microbial proteins from bioethanol using five microorganisms, including bacteria and yeasts, under different carbon and nitrogen availability conditions. Under carbon-limiting conditions, the biomass yield was maximum for Cyberlindnera saturnus and Wickerhamomyces anomalus (0.40-0.82 g CDW/g of ethanol consumed). At the same time, Corynebacterium glutamicum presented the lowest protein production cost under this condition. M. pulcherrima had high protein production (up to 0.15 g/g ethanol), standing out with C. glutamicum for its economic and nutritional potential. These results indicate that selecting carbon-limiting or balanced conditions (C/N of 20) is essential to optimize the cost and biomass yield and ensure a protein profile suitable for human consumption.
In an investigation of single-cell protein production by Saccharomyces cerevisiae in a submerged batch bioprocess, the culture medium composition was optimized to maximize protein production. Using experimental design methodology, the optimal concentrations were determined to be 0.3 g/L ammonium sulfate, 0.15 g/L iron sulfate, 1 g/L glycine, and 50 g/L glucose. These conditions resulted in a cell count of 8.84 log CFU/mL and a protein content of 44.6% in the dry biomass. Glycine and glucose were the most influential factors, contributing 39.32% and 36.15%, respectively, to cell growth. Additionally, ammonium and iron sulfate demonstrated significant interactions, with an interaction severity index of 50.71% [47]
Recent advancements in sustainable single-cell protein production include a study that developed a bioelectrosynthesis system to convert biogas into single-cell protein. This system achieved a protein concentration of 472.04 mg/L and an impressive conversion efficiency of CO₂ to CH₄ of 92.97%. By increasing the fermenter size from 100 mL to 1 L, the protein production tripled, resulting in a CH₄ to single-cell protein conversion efficiency of 70.67%. Furthermore, the system showed remarkable tolerance to H₂S, handling concentrations of up to 5000 ppm without significantly affecting protein production. The generated biomass was protein-rich, containing over 62.8% of total amino acids. While biogas presents a promising alternative to costly and unsustainable natural gas, its utilization requires purification to reduce undesirable components like CO₂ and H₂S, which can hinder process efficiency [48].
The specific requirements of the alternative protein industry require advances in bioreactor design, specially adapted to handle the high viscosity of certain cultures, such as filamentous fungi, ensuring precise control of fermentation conditions. In many cases, models such as air-lift bioreactors are ideal, as their structure allows the movement of the dense medium without the need for mechanical agitation parts, reducing energy consumption compared to conventional bioreactors. This configuration is especially efficient for the production of fungal biomass, where high viscosity can make it difficult to use conventional agitators [49].
Samsing et al. [50] explored the use of microbial single-cell proteins (SCP) as a replacement for fishmeal in aquaculture feeds, specifically in barramundi (Lates calcarifer) fry. The study found that replacing 25% and 50% of fishmeal with SCP from Methylococcus capsulatus led to reduced growth performance, with weight gain rates of 85.5% and 61.4%, respectively, compared to 106.2% in the control group. Additionally, survival rates dropped to 66.7% and 71.7% in the 25% and 50% SCP groups, respectively, compared to 96.7% in control. However, histopathological analysis revealed necrotizing enteritis, pancreatic atrophy, and liver pathology in fish-fed SCP diets, indicating potential health risks at higher inclusion of bacterial protein diets.
The design of appropriate bioreactors for each bioprocess is essential to maximize the efficient use of waste and promote more sustainable and economically viable biotechnological processes. Given the growing demand for alternative proteins, using lignocellulosic biomass residues presents a promising solution for producing mycoproteins. Reviews indicate that the application of optimized bioreactors, whether in solid-state, submerged, or surface culture fermentation, is central to these processes [51].
Voutilanen et al. [52] reported a pre-treatment analysis, such as steam explosion and enzymatic hydrolysis, for the production of SCP from wheat straw, obtaining a hydrolysate of 67.7 g/L of sugars, which would then be used for the production of SCP from Pekilo, Fusarium and Torula and recombinant protein. This method resulted in an economically viable process (5160-9007 €/ton) when compared with the production of other proteins, such as vegetable proteins (10,500 €/ton), indicating that the production of these compounds may be viable for food applications and present competitive market value; however, textural and flavor improvements may increase the final value of the product

Table 2.
Main Bioreactors and Conditions for Production of Single-Cell Proteins, Forms of Consumption.
Table 2.
Main Bioreactors and Conditions for Production of Single-Cell Proteins, Forms of Consumption.
| Microorganism | Bioreactor | Substrate | Volume (L) | Protein production | Temperature (°C) | pH | Stirring (rpm) | Oxygenation | Time | References |
| Methylococcus capsulatus MIR | Bioreactor | Methane | 1.5 | 4.72 g/L | 42 | 6.3 | 1000 | 18000 cm³ de ar/h | 10-14 days | [53] |
| Methylococcus capsulatus | Fixed-film anaerobic digester | Biological waste | 17.5 | 52% (w/w) | 45 | 7.0-8.0 | - | 0% | 12.25 days | [54] |
| Saccharomyces cerevisiae | Bioreactor | Food waste | 15 | 2.2 g/mL | - | 3.0-3.5 | - | - | 4 days | [55] |
| Fusarium venenatum KACC | Pressure cycle reactor | Malt extract broth | 150.000 | 300-350 kg biomass/h | 28-30 | 6.0 | - | - | Continuous | [56,57] |
| Fusarium venenatum | Stirred bioreactor | Glucose | 3.5 | 9.53 g/L | 28 | 6.0 | 100 up to 300 | 1 vvm | 72 h | [58] |
| Fusarium venenatum IR372C | Laboratory bioreactor | Modified date syrup in Vogel medium | 3.0 | 55% (w/w) | 28 | 5.6 | 400 | 1 vvm | 72 h | [59] |
| Fusarium venenatum CGMCC | Stirred bioreactor | Glucose | 3.7 | 10.2 g/L (61,9 %, m/m) | 29 | 6.0 | 100 up to 300 | 1 vvm | 48 h | [60] |
| Chlorella sorokiniana GT-1 | Pilot fermenter | Glucose | 1000 | 73.5 g/L/d | 30 | 6.0 | 180 | DO = 20% | 6 days | [61] |
| Spirulina platensis | Photobioreactor | Beet sugar extraction cake | 0.4 | 0.56 g/L (52.5%, m/m) | 26 | 8.0 | - | - | 7 days | [62] |
Gome et al. [63] explored the cultivation of bovine mesenchymal stem cells on plant-based scaffolds using a macrofluidic single-use bioreactor (MSUB) at 37 ºC in a 5% CO2. The study found that puffed rice, used as a scaffold, provides an optimal environment for cell adhesion and growth thanks to its porous structure and hydrophilic surface. Cells seeded on the rice scaffolds showed a threefold increase in proliferation after nine days in the MSUB compared to traditional flask cultures. Moreover, the MSUB's design minimized shear forces, which are detrimental to cell growth, and allowed efficient nutrient exchange, further enhancing cell proliferation. These results indicate that using food-grade plant-based scaffolds in microfluidic bioreactors could significantly reduce production costs and increase scalability in cultured meat bioprocessing.
Recent research also demonstrates the potential of edible mycelium to support larger-scale cultured meat production. Ogawa et al. [64] report that Aspergillus oryzae mycelium is effective as a carrier for anchorage-dependent cells, providing superior attachment and promoting cell proliferation and differentiation in dense cultures, increasing the viability of high-volume production. This natural support system can improve the growth and metabolic activity of C2C12 myoblasts, outperforming inedible Cytodex microcarriers in bovine satellite cell culture, highlighting an innovative and sustainable alternative for the production of cultured meat at scale.
Amirvaresi and Ovissipour [65] investigated using plant- and microbial-derived protein hydrolysates as sustainable substitutes for fetal bovine serum (FBS) in seafood cell culture media. They found that low concentrations (0.001–0.1 mg/mL) of hydrolysates from algae, mushrooms, peas, and yeast promoted zebrafish embryonic stem cell growth and maintained cell viability in serum-reduced media, with cell growth comparable to that in media containing 10% serum. Algae protein hydrolysates provided the highest amino acid content and supported the highest cell viability while reducing greenhouse gas emissions by 90% compared to traditional FBS-based media. The greenhouse gas emissions of protein hydrolysates were higher than those of fetal bovine serum due to resource consumption during enzymatic hydrolysis. Still, at low concentrations (0.01 mg/mL), the hydrolysates showed a lower impact per liter of prepared medium. Algal proteins stood out with the highest emissions among all sources evaluated. This research demonstrates the potential of protein hydrolysates to reduce costs and environmental impact in cultivated meat production.
Wikandari et al. [66] analyzed mycoprotein production using Rhizopus oligosporus cultivated in wastewater from the tempeh manufacturing process as a substrate in a shaker flask. They observed that the highest biomass yield (7.76 g/L) was obtained when supplemented with yeast extract under optimal conditions of pH 4.5 and shaking at 125 rpm. The mycoprotein produced had a protein content of 19.44%, low-fat content (1.56%), and high fiber content (8.51%). In addition, it was rich in essential amino acids, such as lysine (3.05 g/100 g) and leucine (2.58 g/100 g), which are both critical for human nutrition. Thus, residual boiling water from tempeh production may be used as a medium for edible filamentous fungi, offering potential as a sustainable, high-quality protein source. However, further research is needed to improve digestibility and develop products.
3.2. Application of Enzymes to Obtain Protein Compounds
Proteases, such as Papain and bromelain, have been applied in the food industry for their potential to enhance the digestibility and production of bioactive compounds. However, microbial enzymes produced by Bacillus, Aspergillus, and other microorganisms have also been used due to their high production yield (Figure 4).
Enzymes present some advantages over microorganisms to produce protein hydrolysates, as they are more selective. The production of toxic compounds is unusual, being used to enhance digestibility and reduce allergenicity [67]. However, the application of enzymes is usually more related to the enhancement of the properties of protein food or the production of precursors for cultivated meat production [68].
Enzymes have also been applied to produce hydrolysates, especially bioactive peptides. For example, Alcalase, Flavourzyme, and other enzymes have been studied for their potential to hydrolysate peptides, promoting the production of more digestible compounds and enhancing their nutritional properties (Table 3).
Several enzymes may be used for the hydrolysis of vegetal materials, such as chia seed expeller [71], resulting in a high degree of hydrolysis (15% for Alcalase, and Neutrase ) and the production of several bioactivities compounds, including antioxidant, hypoglycemic, hypotensive, hypolipidemic, and anti-alcohol effects. Neutrase stood out with the highest angiotensin I-converting enzyme (ACE) inhibition, showing an IC50 of 0.219 ± 0.030 mg/mL, indicating its potential for antihypertensive applications. Additionally, Flavourzyme demonstrated excellent alcohol dehydrogenase (ADH) activation, with the lowest IC50 of 0.047 ± 0.002 mg/mL, suggesting its efficacy in preventing alcohol-induced liver damage. These results emphasize that selecting specific proteases can tailor the functional properties of protein hydrolysates, offering valuable applications for chia by-products as multifunctional ingredients in the food and health industries.
Soybean and peanut meals, also offer a promising route for producing inputs for cultured meat. The usage of enzymes, such as Viscozyme, may enhance protein extraction yields from the biomass, with the highest yields reaching 31.69% for soybean meal and 33.98% for peanut meal [70]. The enzymatic hydrolysis using Alcalase further optimized the process, achieving a maximum degree of hydrolysis of 31.76% for soybean meal and 30.59% for peanut meal under optimal conditions (50°C, pH 8.0, enzyme-to-substrate ratios of 3.5% and 5.0%, respectively). These results suggest that proteins and peptides obtained through these processes could serve as cost-effective, animal-free inputs for cultured meat production, potentially replacing the need for fetal bovine serum in cell culture media.
Some studies have applied combinations of enzymes, such as Alcalase and Flavourzyme [72]. The results showed that hydrolysates produced with Flavourzyme exhibited the highest antioxidant activity and water-holding capacity, likely due to its dual action as an endo- and exo-peptidase, generating smaller peptide fragments with greater bioactivity. In contrast, Alcalase, an endo-peptidase, produced hydrolysates with moderate antioxidant activity and a higher degree of hydrolysis (20.6%). Yet, its impact on water-holding capacity was lower than that of Flavourzyme. However, combining both enzymes did not significantly enhance hydrolysis or antioxidant activity beyond Alcalase alone, suggesting that Flavourzyme may not have been fully active in the combined treatment.
Some studies have also applied plant-based enzymes to explore the bioactive properties of protein hydrolysates, such as bromelain and papain, to produce hydrolysates of quinoa and amaranth proteins. The hydrolysates presented high antioxidant, antimicrobial, and anti-hemolytic activities. The highest 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) and 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activities were observed for quinoa hydrolysates generated with chymotrypsin after 2 hours (QC-2) and amaranth hydrolysates after 4 hours (AC-4), with values of 525.2 µmol TE/µg and 1442.5 µmol TE/µg, respectively. Antimicrobial activity against Staphylococcus aureus was strongest in the 6-hour bromelain-generated amaranth hydrolysate (AB-6), showing a 21.0 mm inhibition zone.
3. Biotechnological Configurations and Processes for Protein Production
The use of alternative substrates, such as agro-industrial waste, is a sustainable alternative for food production, using by-products or non-edible substances to produce protein compounds, presenting environmental and nutritional advantages compared to meats and products traditionally used to feed people and animals.[83,84,85]. The following topics address recent strategies for using different types of waste — including sludge, industrial effluents, and plant and food waste — highlighting the technological configurations and the impact of these approaches on the viability of the large-scale process (Figure 5).
4.1. Production of SCP from Sludge and Organic Waste
To make protein production more sustainable, studies have explored alternative sources, such as the conversion of biogas from sewage sludge into SCP, focusing on use in animal feed. In this context, methanotrophic bacteria have effectively converted biogas into high-protein biomass. The research found a yield of 0.66 g of biomass per gram of CH₄, with more than 41% protein (dry weight) and essential amino acids, such as histidine, valine, and leucine. Mixed cultures containing species such as Methylomonas and Methylophilus showed higher productivity, with Methylomonas being favored in anaerobic digestion supernatants. Despite the promising results, culture growth was affected by the presence of H₂S in the biogas and the lack of micronutrients, indicating that the removal of H₂S and the addition of nutrients are necessary interventions to optimize the process and obtain quality protein biomass for feed formulation [86].
Volatile fatty acids (VFAs) derived from the anaerobic digestion of organic waste, such as manure and food scraps, have great potential as a carbon source, especially in processes involving the cultivation of fungi. The use of Aspergillus oryzae in effluents rich in VFAs, for example, resulted in a biomass yield of 0.47 g of dry biomass per gram of VFAs consumed, with a protein content of up to 47% and a fat content of 5%. In addition, the process demonstrated a significant capacity for contaminant removal, with reductions of up to 58% in chemical oxygen demand (COD) and 48% in ammonium, suggesting that the biomass produced could be an effective and sustainable resource for animal feed supplementation, as indicated in previous studies [87].
Research has explored the reuse of urban waste through a specific technique. This method allows for a biomass yield ranging from 0.59 to 0.76 grams of cell dry weight per gram of methane (g-Cell Dry Weight/g-CH₄). Notably, the highest recorded yield reached 0.87 g-Cell Dry Weight/g-CH₄ when biomethane was used. The biomass produced is rich in essential amino acids, including aspartic acid, glutamic acid, and lysine, comparable to traditional protein sources. Additionally, the study showed that the digestate from anaerobic digestion can effectively serve as a nitrogen source for microbial growth. This is true for both centrifuged-filtered digestate and nitrogen extracted through an electrochemical process, achieving ammonium assimilation efficiency of up to 91.5%. The research emphasizes the potential of biogas as a substitute for natural gas in producing single-cell protein, providing a sustainable solution for utilizing urban waste [88].
Mutant strains can also increase protein yield, as reported by But et al. [53], who investigated single-cell protein production (SCP) from methane using the methanotrophic bacterium Methylococcus capsulatus MIR and glycogen-deficient mutants. The main results indicated that the mutant strain showed an 18-fold reduction in glycogen content (10.8 mg/g DCW) compared to the wild-type strain (187.5 mg/g DCW), which resulted in an increase in biomass protein content from 54% to 71% of dry weight (DCW). The study also observed that the mutation in the glucokinase gene drastically reduced glycogen content but resulted in an intracellular glucose accumulation twice as high as that observed in the wild-type strain. These findings suggest that inhibition of glycogen synthesis can improve SCP quality by increasing protein content without compromising the growth rate of the mutant strains, which is promising for application in large-scale bioprocesses.
4.2. Production of SCP from Industrial Effluents
SCP production from industrial effluents has attracted interest due to its potential to transform waste into high-value protein sources. In particular, using brewery wastewater as a substrate in bioreactors has proven effective, resulting in a biomass with 55% crude protein suitable for animal feed supplementation. In studies conducted in bioreactors, it was observed that microbial communities varied between production trials despite using the same influent. It was noted that Rhizospheric diazotrophs, such as Azospirillum and Azonexus, constituted up to 73% of the microbial sequences in the final product. Monitoring microbial diversity at each stage revealed lower diversity in the acidogenic basin. In contrast, the bioreactor, operating with an average of 2600 mg/L of suspended solids (MLSS) and low levels of dissolved oxygen (0.22 mg/L), favored a microbial composition that contributed significantly to SCP yield [83].
Also, Zhu et al. [89] investigated optimizing the extraction process of proteins and polypeptides from waste brewer's yeast (WBY) using ultrasound-assisted extraction and enzymatic hydrolysis. The maximum protein and polypeptide extraction rate was 73.94% and 61.24%, respectively. The protein composition included 17 amino acids, with emphasis on glutamate (6.46%), glycine (5.26%), and lysine (3.92%). Amino acid scoring revealed that threonine and sulfur amino acids (methionine + cysteine) were limiting. Furthermore, polypeptides with molecular weight less than 10 kDa demonstrated higher antioxidant activity against hydroxyl radicals (95.10%), DPPH (98.37%), and ABTS (69.41%) compared to higher molecular weight proteins. These findings suggest that proteins and polypeptides extracted from WBY can be used as functional ingredients in the food and health industry due to their high amino acid quality and potent antioxidant activity.
Kurcz et al. [90] produced SCP from yeast Candida utilis using glycerol as a carbon source and potato processing wastewater as a nitrogen source. The study revealed that adding 5% glycerol to the medium resulted in a biomass yield of 33.25 g/L, with a protein content of 36.7 g/100 g dry biomass and a protein production of 12.2 g/L after 72 hours of cultivation. The process also significantly reduced glycerol in the medium (83%) and nitrogen compounds (51%), in addition to reducing the chemical oxygen demand (COD) index by 91%, highlighting the potential use of industrial waste in the production of SCP.
Liu et al. [84] produced SCP from potato starch processing residues using a two-step fermentation process. In the first step, the residue was treated with the mutant Aspergillus niger H3, selected for its high cellulase activity after UV mutagenesis and ethyl sulfate, resulting in 80.54% degradation of the cellulose in the residue. In the second step, Bacillus licheniformis was used to ferment the released sugars, optimizing the fermentation conditions at 32.8°C, pH 6.67, and an inoculum concentration of 1.78%. After treatment, the crude protein content in the product increased from 6.24% to 38.21%, with a significant increase in true protein from 2.83% to 24.86%. The study demonstrated that the process has the potential for large-scale application, offering a solution for both pollution caused by industrial waste and animal feed production.
4.3. Production of SCP from Plant Residues
The use of yeasts in solid-state fermentation Candida tropicalis, Candida krusei, and Saccharomyces cerevisiae may also be an alternative for using plant residues and producing SCP. The combination of S. cerevisiae with C. tropicalis was the most efficient in a study carried out by Shahzad et al. [91], resulting in a SCP yield of 10.86 mg/g dry biomass without pretreatment, an increase of 63.6% compared to the control. After pretreatment of the plant residues with 4% sulfuric acid and 140°C, SCP production increased to 21.9 mg/g. Nutrient supplementation further increased the output to 31.7 mg/g, representing a total increase of 375.8% compared to the untreated substrate. These results highlight the potential of optimizing acid pretreatment and yeast co-culture for efficient SCP production from plant residues.
Through alkaline hydrolysis, SCP may be produced from olive fruit residues using the yeast Candida lipolytica [92]. The treatment of the residues with 0.8 M NaOH at 100°C for 60 min resulted in a maximum sugar extraction of 12.5%. The yeast achieved a maximum biomass growth of 17.55 g/L, with SCP production of 11.24 g/L, equivalent to 64% of the dry weight of the biomass, after 4 days of incubation at 30°C and an initial pH of 5.5. The study demonstrated that using 0.4% peptone as a nitrogen source was optimal to maximize SCP production. These results highlight the potential of olive residues as a substrate for sustainable SCP production while mitigating the environmental impact of these residues.
The simultaneous production of polyhydroxyalkanoates (PHA) from Carica papaya waste and SCP have also been reported [93]. The study showed that the yeast Saccharomyces cerevisiae NDRI 364 produced up to 4.86 g/L of SCP under optimized conditions of 25°C, pH 5, and 120 hours of fermentation. The best carbon source was sucrose (4.26 g/L), and the best nitrogen source was meat extract (4.56 g/L). In addition, the SCP produced had 31.10% protein, including all essential and non-essential amino acids, with high levels of tryptophan (6.49%) and glycine (5.72%). These results indicate the potential of papaya waste as an inexpensive and effective substrate for SCP production.
Hashem et al. [94] used wasted date molasses (WDM) as a substrate for cultivating unconventional yeasts. The yeasts Hanseniaspora guilliermondii, Hanseniaspora uvarum, Issatchenkia orientalis, and Cyberlindnera fabianii were identified as the most productive, reaching biomass yields of up to 75.82 g dry biomass/100 g WDM and protein content of 54.34%. The addition of organic or inorganic nitrogen sources significantly increased the conversion efficiency of WDM to SCP. The results showed that these new yeast strains have potential for industrial application in producing SCP, mainly for animal feed.
The production of high-value-added co-products parallel to SCP production has also been studied, such as the synthesis of SCP and xylitol by the yeast Candida intermedia FL023 from lignocellulosic hydrolysates. The strain showed high efficiency in the assimilation of hexoses, pentoses, and cellobiose, with a protein content of 48.4%. Using corncob hydrolysate and urea as carbon and nitrogen sources, the SCP productivity reached 0.86 g L⁻¹ h⁻¹, yielding 0.40 g of biomass per gram of sugar. When Miscanthus straw pretreated with NaOH, the productivity was 0.23 g L⁻¹ h⁻¹, yielding 0.17 g g⁻¹ of straw. Furthermore, the yeast showed high tolerance to furfural and acetic acid inhibitors during xylitol production, yielding 45.7 g L⁻¹. These results demonstrate the potential of C. intermedia for producing SCP and xylitol from lignocellulosic residues, providing a promising route for valorizing abundant biomasses [85].
Other studies also report the use of Rhodopseudomonas palustris P1, stimulated by fermented pineapple extract (FPE), to treat effluents from natural rubber sheet processing, producing single-cell protein (SCP) as a byproduct [95]. The main results indicated that the removal of chemical oxygen demand (COD) was 98%, suspended solids (SS) 79%, and total sulfide (TtS) 72% after 4 days of treatment under microaerobic light conditions. The biomass generated consisted of 65% protein, 8% carbohydrate, and 3% fat, making it a potential source of SCP for animal feed. These results demonstrate the efficiency of the process, in addition to eliminating the H₂S odor in the treatment, suggesting the feasibility of using this approach to treat wastewater at low cost.
4.3. Production of SCP from Food Waste
Food waste has also been reported for the synthesis of SCP. The solid-state fermentation (SSF) of food industry waste mixtures using Saccharomyces cerevisiae, Kluyveromyces marxianus, and kefir to produce single-cell protein (SCP), volatile aroma compounds, and lipids. The main results indicated that the highest protein content (38.5% on a dry basis) was obtained with S. cerevisiae. In comparison, K. marxianus produced the highest sum of protein and lipids (59.2% on a dry basis), with a lipid content of 25.5%. The study also highlighted the production of volatile aroma compounds, such as e-pinene, whose concentration reached 4208 mg/kg in the kefir fermented product. These findings suggest that the SSF process can produce animal feed enriched in protein, lipids, and value-added compounds, such as e-pinene, in a biorefinery approach[96].
Tropea et al. [97] evaluated the use of a multicomplex substrate composed of food waste of animal and vegetable origin, such as fish, pineapple, banana, apple, and citrus peels, fermented by the yeast Saccharomyces cerevisiae. The fermentation process was optimized with enzyme supplementation, resulting in a protein content of up to 40.19%. In addition, the final substrate presented 14.46% lipids and a significant reduction in soluble (from 20.5% to 6.10%) and insoluble (from 19.15% to 2.14%) sugars. These results indicate the potential for simultaneous valorization of different food wastes, without prior separation, to produce SCP suitable for animal feed.
Additionally, the production of SCP from industrial food waste, such as potato peel, banana peel, carrot pulp, and orange peel, using Saccharomyces cerevisiae in submerged fermentation, has presented high yields [98]. The maximum SCP yield was obtained from potato peel, with 47.78% crude protein, while orange peel showed the lowest yield. The addition of ammonium sulfate as a nitrogen source significantly increased SCP production, with a 58% increase in yield over potato peel. In addition, the SCP produced was rich in essential amino acids, such as lysine, valine, and leucine, with concentrations up to 9.8 times higher than in wheat flour. Sensory analysis of the SCP-enriched bread indicated that adding up to 4% SCP did not negatively affect the organoleptic properties, making it a viable solution to increase the protein content of foods.
5. Technological, Regulatory, and Consumer Acceptance Challenges Regarding Non-Meat Proteins
The development of non-animal fermented proteins is driving significant changes in the food industry. However, incorporating these products faces substantial regulatory and sociocultural challenges, including the need to demonstrate safety, efficacy, and consumer acceptance. Gaps in current regulations and public caution aggravate this scenario.
Technological innovations in the production of non-animal proteins range from developing products that mimic animal tissues to optimizing processes to increase protein yield. Hosseini and Alireza (2022) [99] described using microreactors that replicate ground meat structures, using components such as fat emulsions and non-human plasma to simulate the texture and flavor of conventional meat. Bubner et al. (2024) [100] and Reed et al. (2021) [101] investigated methods of muscle fiber replication and cell culture processes. Bubner et al. (2024) [100] highlight a method of producing muscle spheroids, which allows cell expansion in controlled environments with growth factors to optimize yield. Reed et al. (2021) [101] propose an alternative approach using the microorganism Cupriavidus necator to generate protein-rich biomass by CO₂ fermentation, converting it into a protein mass with a meat-like texture after an extrusion process.
Simpson et al. (2020) [102] used gaseous substrates rich in CO, CO₂, and H₂ with anaerobic microorganisms to convert industrial waste into protein biomass, reducing dependence on agricultural sources. Techniques such as using filamentous fungi, such as Fusarium venenatum, to produce biomaterials with high protein density in membrane bioreactors have also shown promise. This system, described by Macur et al. (2024) [103], allows the control of the fungus's growth, maximizing the protein content and minimizing toxins.
Pearlman et al. (2022) [104] reported the genetic modification of Cupriavidus necator, reducing the production of polyhydroxybutyrate (PHB) and increasing the protein content, an advantageous trait for animal feed and aquaculture. Research by Kragh et al. (2021) [105] explores the use of the enzyme tripeptidyl peptidase, derived from Aspergillus fumigatus, for the production of protein hydrolysates with lower immunogenicity and bitterness, suitable for food formulations.
Fermentative processes have both advantages and disadvantages when producing SCP by fermentation. However, pH and temperature may be controlled during the process, as they may lead to a reduction in production or contamination; thus, bioreactors are used. Also, substrate consumption and the production of inhibitors by-products may decrease yields; thus, for bacteria and yeast, fed-batch or continuous stirred-tank reactors are usually applied in those productions. Additionally, some substrates require complex pre-treatments, increasing costs and making large-scale implementation difficult. Strict regulations and high energy consumption in some operations also represent obstacles to be overcome (Table 4).
However, although technology in non-meat protein production has advanced consistently in recent years, accepting these products by a large part of the population is still a barrier to overcome. Product characteristics, psychological factors, and external variables, such as health, taste, and sustainability, influence this acceptance. Products such as plant-based proteins and cultured meat attract different consumer profiles. In contrast, psychosocial resistance, such as food neophobia, tends to be more significant for options such as cultured meat and insect-derived proteins (Onwezen et al. 2021) [106].
Cultured meat, for example, is seen as an environmentally beneficial alternative, eliminating the need for slaughter. However, the perception of artificiality, in addition to the high cost of production and ethical and religious challenges, limits its popularity (Peker et al. 2024) [107]. Plant-based protein products also face resistance, especially due to taste and texture issues. Still, their environmental appeal and health benefits extend their potential among restrictive diets and dietary choices, such as veganism. Processing methods such as fermentation and hydrolysis have been explored to improve the sensory profile of these products.
Recent studies indicate that acceptance of alternative proteins depends on familiarity and perception of environmental and nutritional benefits. Hanan et al. (2024) [108] identified seven main factors that influence the acceptance of cultured meat, including neophobia, perceived artificiality, and situational context. Plant-based and algae products are more accepted due to familiarity and perceived health and environmental benefits. At the same time, cultured meat and insect proteins encounter greater resistance due to a lack of information and the disgust factor (Can, Majoo, and Öztürkcan 2024) [109].
A factor to consider in this field is the regulatory actions of each country. Since this is an innovative product, there are still no consistent and specific regulations for many of these alternative proteins, generating uncertainty for industry and consumers. In the United States, for example, the regulation of alternative proteins is divided between the FDA (Food and Drug Administration) and the USDA (Department of Agriculture), which can generate ambiguity, especially in products derived from microorganisms. A case that illustrates this situation well is that of Quorn, a product based on mycoprotein derived from the fungus Fusarium venenatum. For its regulatory approval, it was necessary to present detailed evidence of safety due to concerns about fungal toxicity [110] [111]. Furthermore, labeling these products is a critical point: it is essential to clearly describe the production process and ingredients, as recommended by the UK Food Standards Agency. Although transparency is a benefit, terms such as “microorganisms” can still generate consumer resistance. Thus, the usage of GRAS microorganisms is important for the safety of SCP, as some microbial proteins may also present some unwanted effects, as was observed in fish that were fed by bacterial protein and presented mild necrotizing enteritis in their gastrointestinal tracts [50].
Consumer acceptance is also profoundly influenced by cultural factors. In regions where fermented products are standard, such as Asia, the acceptance of alternative proteins, especially SCP, tends to be higher. Consumers in Europe and the United States prefer plant-based proteins associated with "natural" sources. Communication strategies emphasizing these alternatives' environmental and nutritional benefits have sought to mitigate distrust.
In addition to regulatory and acceptance barriers, environmental and ethical issues have driven the demand for alternative proteins. Life cycle studies indicate that fermented proteins produce significantly lower carbon emissions than conventional animal proteins, especially in mycoproteins requiring less water and arable land. However, industrial fermentation processes still present sustainability challenges that must be explored in future research to validate environmental viability.
In the face of a rigorous and constantly evolving regulatory landscape, fermented proteins of non-animal origin still face obstacles to achieving market consolidation. Cultural barriers, consumer distrust, and regulatory complexity represent significant challenges. The success of these food alternatives depends on transparent regulatory policies, educational communication strategies, and a joint effort between industry, regulators, and consumers to establish these proteins as safe, sustainable, and affordable options in the global diet.
6. Conclusions
The growing demand for alternative non-animal proteins requires technological innovations that promote sustainability and ensure economic viability and market competitiveness. This review highlighted significant advancements in fermentation processes to produce single-cell proteins and protein hydrolysates, emphasizing the optimization of agro-industrial waste as substrates. While challenges related to scalability, contamination control, and regulatory acceptance remain substantial obstacles, the technologies discussed, such as precision fermentation and the use of proteolytic enzymes, show great potential for improving the efficiency and quality of alternative proteins. For these innovations to be widely adopted, continued development of cost-effective processes and increased public acceptance will be essential. Ultimately, fermented alternative proteins can play a central role in creating a more sustainable food system capable of meeting the nutritional needs of the growing global population. Thus, future research should focus on improving public acceptance of alternative protein products and enhancing the economic viability of emerging technologies, such as the large-scale use of agro-industrial waste and the effects of long-term diets composed of SCP or protein hydrolysates.
Author Contributions
Emanuel do Nascimento Muniz: Investigation, data curation, writing—original draft preparation. Rebeca Teixeira de Queiroz Montenegro: Investigation, data curation, writing—original draft preparation. Daniel Nascimento da Silva: Investigation, data curation, writing—original draft preparation. Alan Portal D’Almeida: Conceptualization, formal analysis, investigation, writing—original draft preparation, visualization, writing—review and editing. Luciana Rocha Barros Gonçalves: Conceptualization, investigation, supervision, writing—review and editing, writing—review and editing. Tiago Lima de Albuquerque: Conceptualization, methodology, resources, investigation, supervision, project administration, writing—original draft preparation, writing—review and editing.
Data Availability Statement
The data will be available at request.
Acknowledgments
The figures were created using the free version of BioRender.com. Enzyme images were produced using UCSF ChimeraX.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cavaliere, V.N.; Mindiola, D.J. Methane: A New Frontier in Organometallic Chemistry. Chem. Sci. 2012, 3, 3356. [Google Scholar] [CrossRef]
- Bačėninaitė, D.; Džermeikaitė, K.; Antanaitis, R. Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission. Animals 2022, 12, 2687. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Valappil, G.; Dutta, B.; Imbeault-Tétreault, H.; Ominski, K.H.; Cordeiro, M.R.C.; Kröbel, R.; Pogue, S.J.; McAllister, T.A. An Assessment of the Environmental Sustainability of Beef Production in Canada. Can. J. Anim. Sci. 2024, 104, 221–240. [Google Scholar] [CrossRef]
- Fu, W.; Zhang, H.; Whaley, J.E.; Kim, Y.K. Do Consumers Perceive Cultivated Meat as a Sustainable Substitute to Conventional Meat? Assessing the Facilitators and Inhibitors of Cultivated Meat Acceptance. Sustain. 2023, 15. [Google Scholar] [CrossRef]
- Cuadrado-Osorio, P.D.; Ramírez-Mejía, J.M.; Mejía-Avellaneda, L.F.; Mesa, L.; Bautista, E.J. Agro-Industrial Residues for Microbial Bioproducts: A Key Booster for Bioeconomy. Bioresour. Technol. Reports 2022, 20, 101232. [Google Scholar] [CrossRef]
- D’Almeida, A.P.; Infante Neta, A.A.; de Andrade-Lima, M.; de Albuquerque, T.L. Plant-Based Probiotic Foods: Current State and Future Trends. Food Sci. Biotechnol. 2024, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Mattila, A.S. The Impact of Environmental Messages on Consumer Responses to Plant-Based Meat: Does Language Style Matter? Int. J. Hosp. Manag. 2022, 107, 103298. [Google Scholar] [CrossRef]
- Meng, J.; Liu, S.; Gao, L.; Hong, K.; Liu, S.; Wu, X. Economical Production of Pichia Pastoris Single Cell Protein from Methanol at Industrial Pilot Scale. Microb. Cell Fact. 2023, 22, 198. [Google Scholar] [CrossRef]
- Zhao, M.; Ma, J.; Zhang, L.; Qi, H. Engineering Strategies for Enhanced Heterologous Protein Production by Saccharomyces Cerevisiae. Microb. Cell Fact. 2024, 23, 32. [Google Scholar] [CrossRef]
- Wang, G.; Wu, X.; Yin, Y. Synthetic Biology-Driven Customization of Functional Feed Resources. Trends Biotechnol. 2022, 40, 777–780. [Google Scholar] [CrossRef]
- Kühl, S.; Schäfer, A.; Kircher, C.; Mehlhose, C. Beyond the Cow: Consumer Perceptions and Information Impact on Acceptance of Precision Fermentation-Produced Cheese in Germany. Futur. Foods 2024, 10, 100411. [Google Scholar] [CrossRef]
- Saengsuk, N.; Laohakunjit, N.; Sanporkha, P.; Kaisangsri, N.; Selamassakul, O.; Ratanakhanokchai, K.; Uthairatanakij, A.; Waeonukul, R. Comparative Physicochemical Characteristics and in Vitro Protein Digestibility of Alginate/Calcium Salt Restructured Pork Steak Hydrolyzed with Bromelain and Addition of Various Hydrocolloids (Low Acyl Gellan, Low Methoxy Pectin and κ-Carrageenan). Food Chem. 2022, 393, 133315. [Google Scholar] [CrossRef]
- Vogelsang-O’Dwyer, M.; Sahin, A.W.; Arendt, E.K.; Zannini, E. Enzymatic Hydrolysis of Pulse Proteins as a Tool to Improve Techno-Functional Properties. Foods 2022, 11, 1307. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, G.; Goksen, G.; Mousavi Khaneghah, A. Plant-Based Protein Modification Strategies towards Challenges. J. Agric. Food Res. 2024, 15, 101017. [Google Scholar] [CrossRef]
- Lin, Y.; Maloney, K.; Drake, M.; Zheng, H. Synergistic Functionality of Transglutaminase and Protease on Modulating Texture of Pea Protein Based Yogurt Alternative: From Rheological and Tribological Characterizations to Sensory Perception. Food Hydrocoll. 2024, 150, 109652. [Google Scholar] [CrossRef]
- Nowacka, M.; Trusinska, M.; Chraniuk, P.; Drudi, F.; Lukasiewicz, J.; Nguyen, N.P.; Przybyszewska, A.; Pobiega, K.; Tappi, S.; Tylewicz, U.; et al. Developments in Plant Proteins Production for Meat and Fish Analogues. Molecules 2023, 28, 2966. [Google Scholar] [CrossRef] [PubMed]
- Markova, E.A.; Shaw, R.E.; Reynolds, C.R. Prediction of Strain Engineerings That Amplify Recombinant Protein Secretion through the Machine Learning Approach MaLPHAS. Eng. Biol. 2022, 6, 82–90. [Google Scholar] [CrossRef]
- Vinestock, T.; Short, M.; Ward, K.; Guo, M. Computer-Aided Chemical Engineering Research Advances in Precision Fermentation. Curr. Opin. Food Sci. 2024, 58, 101196. [Google Scholar] [CrossRef]
- Boukid, F.; Ganeshan, S.; Wang, Y.; Tülbek, M.Ç.; Nickerson, M.T. Bioengineered Enzymes and Precision Fermentation in the Food Industry. Int. J. Mol. Sci. 2023, 24, 10156. [Google Scholar] [CrossRef]
- Colarusso, A. V.; Goodchild-Michelman, I.; Rayle, M.; Zomorrodi, A.R. Computational Modeling of Metabolism in Microbial Communities on a Genome-Scale. Curr. Opin. Syst. Biol. 2021, 26, 46–57. [Google Scholar] [CrossRef]
- Hadi, J.; Brightwell, G. Safety of Alternative Proteins: Technological, Environmental and Regulatory Aspects of Cultured Meat, Plant-Based Meat, Insect Protein and Single-Cell Protein. Foods 2021, 10, 1226. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Altamirano, B.; Méndez-Lagunas, L.L.; Rodríguez-Ramírez, J.; Sandoval-Torres, S.; Aquino-González, L.V.; Barriada-Bernal, L.G. Techno-Functional Properties and Antioxidant Capacity of the Concentrate and Protein Fractions of Leucaena Spp. Seeds. Arch. Latinoam. Nutr. 2022, 72, 196–204. [Google Scholar] [CrossRef]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, S.O.; Taiwo, K.A.; Oyedele, D.J.; Adebooye, O.C.; Aluko, R.E. Comparative Study of the Structural and Functional Properties of Protein Isolates Prepared from Edible Vegetable Leaves. Int. J. Food Prop. 2020, 23, 955–970. [Google Scholar] [CrossRef]
- Cao, Z.-H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.-Y. Bioactivity of Soy-Based Fermented Foods: A Review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef]
- Lombardi, L.; Consalvo, S.; Esposito, C.; Tammaro, D. Tailoring Texture and Functionality of Vegetable Protein Meat Analogues through 3D Printed Porous Structures. Food Hydrocoll. 2025, 159, 110611. [Google Scholar] [CrossRef]
- MacDonald, J.; Brauer, P.; Yi, S. Meat Reduction among Post-Secondary Students: Exploration of Motives, Barriers, Diets and Preferences for Meals with Partial and Full Meat Substitution. Appetite 2023, 188, 106977. [Google Scholar] [CrossRef]
- Woodside, J. V.; Nugent, A.P.; Moore, R.E.; McKinley, M.C. Fruit and Vegetable Consumption as a Preventative Strategy for Non-Communicable Diseases. Proc. Nutr. Soc. 2023, 82, 186–199. [Google Scholar] [CrossRef]
- van Huis, A.; Rumpold, B.A.; van der Fels-Klerx, H.J.; Tomberlin, J.K. Advancing Edible Insects as Food and Feed in a Circular Economy. J. Insects as Food Feed 2021, 7, 935–948. [Google Scholar] [CrossRef]
- Ros-Baró, M.; Sánchez-Socarrás, V.; Santos-Pagès, M.; Bach-Faig, A.; Aguilar-Martínez, A. Consumers’ Acceptability and Perception of Edible Insects as an Emerging Protein Source. Int. J. Environ. Res. Public Health 2022, 19, 15756. [Google Scholar] [CrossRef]
- Rivas-Navia, D.M.; Dueñas-Rivadeneira, A.A.; Dueñas-Rivadeneira, J.P.; Aransiola, S.A.; Maddela, N.R.; Prasad, R. Bioactive Compounds of Insects for Food Use: Potentialities and Risks. J. Agric. Food Res. 2023, 14, 100807. [Google Scholar] [CrossRef]
- Andrzejczyk, B.; Łobacz, A.; Ziajka, J.; Lis, A.; Małkowska-Kowalczyk, M. Comprehensive Analysis of Yoghurt Made with the Addition of Yellow Mealworm Powder (Tenebrio Molitor). Foods 2024, 13, 2416. [Google Scholar] [CrossRef] [PubMed]
- Zwinkels, J.; Wolkers-Rooijackers, J.; Smid, E.J. Solid-State Fungal Fermentation Transforms Low-Quality Plant-Based Foods into Products with Improved Protein Quality. Lwt 2023, 184, 114979. [Google Scholar] [CrossRef]
- Tanaka, R. ichiro; Sakaguchi, K.; Yoshida, A.; Takahashi, H.; Shimizu, T. Efficient Expansion Culture of Bovine Myogenic Cells with Differentiation Capacity Using Muscle Extract-Supplemented Medium. Food Biosci. 2024, 61, 104610. [Google Scholar] [CrossRef]
- Yuan, X.; Zhu, X.; Sun, R.; Jiang, W.; Zhang, D.; Liu, H.; Sun, B. Sensory Attributes and Characterization of Aroma Profiles of Fermented Sausages Based on Fibrous-like Meat Substitute from Soybean Protein and Coprinus Comatus. Food Chem. 2022, 373, 131537. [Google Scholar] [CrossRef]
- Flaibam, B.; da Silva, M.F.; de Mélo, A.H.F.; Carvalho, P.H.; Galland, F.; Pacheco, M.T.B.; Goldbeck, R. Non-Animal Protein Hydrolysates from Agro-Industrial Wastes: A Prospect of Alternative Inputs for Cultured Meat. Food Chem. 2024, 443. [Google Scholar] [CrossRef]
- Fedeli, R.; Mazza, I.; Perini, C.; Salerni, E.; Loppi, S. New Frontiers in the Cultivation of Edible Fungi: The Application of Biostimulants Enhances the Nutritional Characteristics of Pleurotus Eryngii (DC.) Quél. Agric. 2024, 14. [Google Scholar] [CrossRef]
- Vega-Gómez, L.M.; Molina-Gilarranz, I.; Fontes-Candia, C.; Cebrián-Lloret, V.; Recio, I.; Martínez-Sanz, M. Production of Hybrid Protein-Polysaccharide Extracts from Ulva Spp. Seaweed with Potential as Food Ingredients. Food Hydrocoll. 2024, 153. [Google Scholar] [CrossRef]
- Choudhary, F.; Khandi, S.A.; Aadil, R.M.; Bekhit, A.E.D.A.; Abdi, G.; Bhat, Z.F. What Do Meat Scientists Think about Cultured Meat? Appl. Food Res. 2023, 3, 100360. [Google Scholar] [CrossRef]
- Molnár, J.; Vasas, D.; Kalocsai, R.; Szakál, T.; Ahmed, M.H. Production of Single Cell Protein by the Fermentation Biotechnology for Animal Feeding. Elelmiszervizsgalati Kozlemenyek 2022, 68, 3896–3903. [Google Scholar] [CrossRef]
- Khan, M.K.I.; Asif, M.; Razzaq, Z.U.; Nazir, A.; Maan, A.A. Sustainable Food Industrial Waste Management through Single Cell Protein Production and Characterization of Protein Enriched Bread. Food Biosci. 2022, 46, 101406. [Google Scholar] [CrossRef]
- Zhu, Z.; Wu, Y.; Hu, W.; Zheng, X.; Chen, Y. Valorization of Food Waste Fermentation Liquid into Single Cell Protein by Photosynthetic Bacteria via Stimulating Carbon Metabolic Pathway and Environmental Behaviour. Bioresour. Technol. 2022, 361, 127704. [Google Scholar] [CrossRef] [PubMed]
- Bertasini, D.; Binati, R.L.; Bolzonella, D.; Battista, F. Single Cell Proteins Production from Food Processing Effluents and Digestate. Chemosphere 2022, 296, 134076. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Filho, J.G.; Rodrigues, J.M.; Valadares, A.C.F.; de Almeida, A.B.; Valencia-Mejia, E.; Fernandes, K.F.; Lemes, A.C.; Alves, C.C.F.; Sousa, H.A. de F.; da Silva, E.R.; et al. Bioactive Properties of Protein Hydrolysate of Cottonseed Byproduct: Antioxidant, Antimicrobial, and Angiotensin-Converting Enzyme (ACE) Inhibitory Activities. Waste and Biomass Valorization 2021, 12, 1395–1404. [Google Scholar] [CrossRef]
- Kopplin, B.W.; Bernardo, B. da S.; Moraes, G.P.; Goettems, T.L.; Clerici, N.J.; Lermen, A.M.; Daroit, D.J. Production of Antioxidant Hydrolysates from Bovine Caseinate and Soy Protein Using Three Non-Commercial Bacterial Proteases. Biocatal. Biotransformation 2024, 1–9. [Google Scholar] [CrossRef]
- Prikhodko, D. V.; Krasnoshtanova, A.A. Using Casein and Gluten Protein Fractions to Obtain Functional Ingredients. Foods Raw Mater. 2023, 11, 223–231. [Google Scholar] [CrossRef]
- Van Peteghem, L.; Sakarika, M.; Matassa, S.; Rabaey, K. The Role of Microorganisms and Carbon-to-Nitrogen Ratios for Microbial Protein Production from Bioethanol. Appl. Environ. Microbiol. 2022, 88. [Google Scholar] [CrossRef]
- Hezarjaribi, M.; Ardestani, F.; Ghorbani, H.R. Single Cell Protein Production by Saccharomyces Cerevisiae Using an Optimized Culture Medium Composition in a Batch Submerged Bioprocess. Appl. Biochem. Biotechnol. 2016, 179, 1336–1345. [Google Scholar] [CrossRef]
- Xu, M.; Zhao, D.; Zhu, X.; Su, Y.; Angelidaki, I.; Zhang, Y. Biogas Upgrading and Valorization to Single-Cell Protein in a Bioinorganic Electrosynthesis System. Chem. Eng. J. 2021, 426. [Google Scholar] [CrossRef]
- Knychala, M.M.; Boing, L.A.; Ienczak, J.L.; Trichez, D.; Stambuk, B.U. Precision Fermentation as an Alternative to Animal Protein, a Review. Fermentation 2024, 10. [Google Scholar] [CrossRef]
- Samsing, F.; Sullivan, R.; Truong, H.; Rombenso, A.; Sangster, C.R.; Bannister, J.; Longshaw, M.; Becker, J.A. Replacement of Fishmeal with a Microbial Single-Cell Protein Induced Enteropathy and Poor Growth Outcomes in Barramundi (Lates Calcarifer) Fry. J. Fish Dis. 2024, 47, 1–16. [Google Scholar] [CrossRef]
- Ng, Z.Y.; Kee, P.E.; Abdullah, R.; Lan, J.C.; Ling, T.C.; Jiang, J.; Lim, J.W.; Khoo, K.S. Conversion of Lignocellulosic Biomass Waste into Mycoprotein : Current Status and Future Directions for Sustainable Protein Production. Biomass Convers. Biorefinery 2024. [Google Scholar] [CrossRef]
- Voutilainen, E.; Pihlajaniemi, V.; Parviainen, T. Economic Comparison of Food Protein Production with Single-Cell Organisms from Lignocellulose Side-Streams. Bioresour. Technol. Reports 2021, 14, 100683. [Google Scholar] [CrossRef]
- But, S.Y.; Suleimanov, R.Z.; Oshkin, I.Y.; Rozova, O.N.; Mustakhimov, I.I.; Pimenov, N. V.; Dedysh, S.N.; Khmelenina, V.N. New Solutions in Single-Cell Protein Production from Methane: Construction of Glycogen-Deficient Mutants of Methylococcus Capsulatus MIR. Fermentation 2024, 10. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Kronyak, R.E.; House, C.H. Coupling of Anaerobic Waste Treatment to Produce Protein- and Lipid-Rich Bacterial Biomass. Life Sci. Sp. Res. 2017, 15, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, T.; Pellizzeri, V.; Calabrese, G.; Di Bella, G.; Cicero, N.; Dugo, G. Production of Single Cell Protein (SCP) from Food and Agricultural Waste by Using Saccharomyces Cerevisiae. Nat. Prod. Res. 2018, 32, 648–653. [Google Scholar] [CrossRef]
- Lee, D.J.; Kang, A.N.; Lee, J.; Kwak, M.; Mun, D.; Lee, D.; Oh, S.; Kim, Y. Molecular Characterization of Fusarium Venenatum- Based Microbial Protein in Animal Models of Obesity Using Multi-Omics Analysis. 2024, 7, 1–14. [CrossRef]
- Wiebe, M.G. Myco-Protein from Fusarium Venenatum : A Well-Established Product for Human Consumption. 2002, 2684, 421–427. [CrossRef]
- Tong, S.; Hong, R.; Chen, W.; Chai, M.; Zhang, Y.; Sun, Y.; Wang, Q.; Li, D. Synchronous Bioproduction of Betanin and Mycoprotein in the Engineered Edible Fungus Fusarium Venenatum. 2024. [CrossRef]
- Hashempour-Baltork, F.; Hosseini, S.M.; Assarehzadegan, M.A.; Khosravi-Darani, K.; Hosseini, H. Safety Assays and Nutritional Values of Mycoprotein Produced by Fusarium Venenatum IR372C from Date Waste as Substrate. J. Sci. Food Agric. 2020, 100, 4433–4441. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Chen, W.; Hong, R.; Chai, M.; Sun, Y.; Wang, Q.; Li, D. Efficient Mycoprotein Production with Low CO 2 Emissions through Metabolic Engineering and Fermentation Optimization of Fusarium Venenatum. 2024. [CrossRef]
- Jin, H.; Chuai, W.; Li, K.; Hou, G.; Wu, M.; Chen, J.; Wang, H.; Jia, J.; Han, D.; Hu, Q. Ultrahigh-Cell-Density Heterotrophic Cultivation of the Unicellular Green Alga Chlorella Sorokiniana for Biomass Production. Biotechnol. Bioeng. 2021, 118, 4138–4151. [Google Scholar] [CrossRef]
- Saad, S.; Hussien, M.H.; Abou-ElWafa, G.S.; Aldesuquy, H.S.; Eltanahy, E. Filter Cake Extract from the Beet Sugar Industry as an Economic Growth Medium for the Production of Spirulina Platensis as a Microbial Cell Factory for Protein. Microb. Cell Fact. 2023, 22, 1–21. [Google Scholar] [CrossRef]
- Gome, G.; Chak, B.; Tawil, S.; Shpatz, D.; Giron, J.; Brajzblat, I.; Weizman, C.; Grishko, A.; Schlesinger, S.; Shoseyov, O. Cultivation of Bovine Mesenchymal Stem Cells on Plant-Based Scaffolds in a Macrofluidic Single-Use Bioreactor for Cultured Meat. Foods 2024, 13. [Google Scholar] [CrossRef]
- Ogawa, M.; Kermani, A.S.; Huynh, M.J.; Baar, K.; Leach, J.K.; Block, D.E. Edible Mycelium as Proliferation and Differentiation Support for Anchorage-Dependent Animal Cells in Cultivated Meat Production. npj Sci. Food 2024, 8, 1–10. [Google Scholar] [CrossRef]
- Amirvaresi, A.; Ovissipour, R. Evaluation of Plant- and Microbial-Derived Protein Hydrolysates as Substitutes for Fetal Bovine Serum in Cultivated Seafood Cell Culture Media. bioRxiv 2024, 10. [Google Scholar] [CrossRef]
- Wikandari, R.; Tanugraha, D.R.; Yastanto, A.J.; Manikharda; Gmoser, R. ; Teixeira, J.A. Development of Meat Substitutes from Filamentous Fungi Cultivated on Residual Water of Tempeh Factories. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Pedrosa, M.M.; Rodríguez, J.; Muzquiz, M.; Maleki, S.J.; Cuadrado, C.; Burbano, C.; Crespo, J.F. Influence of Enzymatic Hydrolysis on the Allergenicity of Roasted Peanut Protein Extract. Int. Arch. Allergy Immunol. 2011, 157, 41–50. [Google Scholar] [CrossRef]
- Flaibam, B.; Goldbeck, R. Effects of Enzymes on Protein Extraction and Post-Extraction Hydrolysis of Non-Animal Agro-Industrial Wastes to Obtain Inputs for Cultured Meat. Food Bioprod. Process. 2024, 143, 117–127. [Google Scholar] [CrossRef]
- Batish, I.; Brits, D.; Valencia, P.; Miyai, C.; Rafeeq, S.; Xu, Y.; Galanopoulos, M.; Sismour, E.; Ovissipour, R. Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia Illucens) Protein. Insects 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Flaibam, B.; Goldbeck, R. Food and Bioproducts Processing Effects of Enzymes on Protein Extraction and Post-Extraction Hydrolysis of Non-Animal Agro-Industrial Wastes to Obtain Inputs for Cultured Meat. 2024, 143, 117–127. [CrossRef]
- Chen, J.; Zhu, L.; Wu, Q.; Chen, Y.; Wu, G.; Zhang, H. Structure Characterization and Bioactivities of Protein Hydrolysates of Chia Seed Expeller Processed with Different Proteases in Silico and in Vitro. Food Biosci. 2023, 55, 102781. [Google Scholar] [CrossRef]
- Cumby, N.; Zhong, Y.; Naczk, M.; Shahidi, F. Antioxidant Activity and Water-Holding Capacity of Canola Protein Hydrolysates. Food Chem. 2008, 109, 144–148. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Sussmann, D.; Schweiggert-Weisz, U.; Eisner, P. Enzymatic Treatment of Soy Protein Isolates: Effects on the Potential Allergenicity, Technofunctionality, and Sensory Properties. Food Sci. Nutr. 2016, 4, 11–23. [Google Scholar] [CrossRef]
- Kong, X.; Zhou, H.; Qian, H. Enzymatic Preparation and Functional Properties of Wheat Gluten Hydrolysates. Food Chem. 2007, 101, 615–620. [Google Scholar] [CrossRef]
- Clemente, A.; Vioque, J.; Sánchez-Vioque, R.; Pedroche, J.; Bautista, J.; Millán, F. Protein Quality of Chickpea (Cicer Arietinum L.) Protein Hydrolysates. Food Chem. 1999, 67, 269–274. [Google Scholar] [CrossRef]
- Vaštag, Ž.; Popović, L.; Popović, S.; Krimer, V.; Peričin, D. Production of Enzymatic Hydrolysates with Antioxidant and Angiotensin-I Converting Enzyme Inhibitory Activity from Pumpkin Oil Cake Protein Isolate. Food Chem. 2011, 124, 1316–1321. [Google Scholar] [CrossRef]
- Wan Mohtar, W.A.A.-Q.I.; Hamid, A.A.; Abd-Aziz, S.; Muhamad, S.K.S.; Saari, N. Preparation of Bioactive Peptides with High Angiotensin Converting Enzyme Inhibitory Activity from Winged Bean [Psophocarpus Tetragonolobus (L.) DC.] Seed. J. Food Sci. Technol. 2014, 51, 3658–3668. [Google Scholar] [CrossRef] [PubMed]
- Wongsa, P.; Yuenyongrattanakorn, K.; Pongvachirint, W.; Auntalarok, A. Improving Anti-Hypertensive Properties of Plant-Based Alternatives to Yogurt Fortified with Rice Protein Hydrolysate. Heliyon 2022, 8, e11087. [Google Scholar] [CrossRef]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of Enzymatic Hydrolysis on Conformational and Functional Properties of Chickpea Protein Isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Yust, M. del M.; Pedroche, J.; Millán-Linares, M. del C.; Alcaide-Hidalgo, J.M.; Millán, F. Improvement of Functional Properties of Chickpea Proteins by Hydrolysis with Immobilised Alcalase. Food Chem. 2010, 122, 1212–1217. [Google Scholar] [CrossRef]
- Mudgil, P.; Omar, L.S.; Kamal, H.; Kilari, B.P.; Maqsood, S. Multi-Functional Bioactive Properties of Intact and Enzymatically Hydrolysed Quinoa and Amaranth Proteins. LWT 2019, 110, 207–213. [Google Scholar] [CrossRef]
- Kamal, H.; Mudgil, P.; Bhaskar, B.; Fisayo, A.F.; Gan, C.-Y.; Maqsood, S. Amaranth Proteins as Potential Source of Bioactive Peptides with Enhanced Inhibition of Enzymatic Markers Linked with Hypertension and Diabetes. J. Cereal Sci. 2021, 101, 103308. [Google Scholar] [CrossRef]
- Lee, J.Z.; Logan, A.; Terry, S.; Spear, J.R. Microbial Response to Single-Cell Protein Production and Brewery Wastewater Treatment. Microb. Biotechnol. 2015, 8, 65–76. [Google Scholar] [CrossRef]
- Liu, B.; Li, Y.; Song, J.; Zhang, L.; Dong, J.; Yang, Q. Production of Single-Cell Protein with Two-Step Fermentation for Treatment of Potato Starch Processing Waste. Cellulose 2014, 21, 3637–3645. [Google Scholar] [CrossRef]
- Wu, J.; Hu, J.; Zhao, S.; He, M.; Hu, G.; Ge, X.; Peng, N. Single-Cell Protein and Xylitol Production by a Novel Yeast Strain Candida Intermedia FL023 from Lignocellulosic Hydrolysates and Xylose. Appl. Biochem. Biotechnol. 2018, 185, 163–178. [Google Scholar] [CrossRef]
- Zha, X.; Tsapekos, P.; Zhu, X.; Khoshnevisan, B.; Lu, X.; Angelidaki, I. Bioconversion of Wastewater to Single Cell Protein by Methanotrophic Bacteria. Bioresour. Technol. 2021, 320, 124351. [Google Scholar] [CrossRef] [PubMed]
- Uwineza, C.; Sar, T.; Mahboubi, A.; Taherzadeh, M.J. Evaluation of the Cultivation of Aspergillus Oryzae on Organic Waste-Derived Vfa Effluents and Its Potential Application as Alternative Sustainable Nutrient Source for Animal Feed. Sustain. 2021, 13. [Google Scholar] [CrossRef]
- Khoshnevisan, B.; Tsapekos, P.; Zhang, Y.; Valverde-Pérez, B.; Angelidaki, I. Urban Biowaste Valorization by Coupling Anaerobic Digestion and Single Cell Protein Production. Bioresour. Technol. 2019, 290, 121743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wang, J.; Feng, Y.; Yin, H.; Lai, H.; Xiao, R.; He, S.; Yang, Z.; He, Y. Process Optimization, Amino Acid Composition, and Antioxidant Activities of Protein and Polypeptide Extracted from Waste Beer Yeast. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Kurcz, A.; Błażejak, S.; Kot, A.M.; Bzducha-Wróbel, A.; Kieliszek, M. Application of Industrial Wastes for the Production of Microbial Single-Cell Protein by Fodder Yeast Candida Utilis. Waste and Biomass Valorization 2018, 9, 57–64. [Google Scholar] [CrossRef]
- Shahzad, H.M.A.; Asim, Z.; Mahmoud, K.A.; Abdelhadi, O.M.A.; Almomani, F.; Rasool, K. Optimizing Cultural Conditions and Pretreatment for High-Value Single-Cell Protein from Vegetable Waste. Process Saf. Environ. Prot. 2024, 189, 685–692. [Google Scholar] [CrossRef]
- Rages, A.A.; Haider, M.M.; Aydin, M. Alkaline Hydrolysis of Olive Fruits Wastes for the Production of Single Cell Protein by Candida Lipolytica. Biocatal. Agric. Biotechnol. 2021, 33, 101999. [Google Scholar] [CrossRef]
- Umesh, M.; Priyanka, K.; Thazeem, B.; Preethi, K. Production of Single Cell Protein and Polyhydroxyalkanoate from Carica Papaya Waste. Arab. J. Sci. Eng. 2017, 42, 2361–2369. [Google Scholar] [CrossRef]
- Hashem, M.; Al-Qahtani, M.S.; Alamri, S.A.; Moustafa, Y.S.; Lyberatos, G.; Ntaikou, I. Valorizing Food Wastes: Assessment of Novel Yeast Strains for Enhanced Production of Single-Cell Protein from Wasted Date Molasses. Biomass Convers. Biorefinery 2022, 12, 4491–4502. [Google Scholar] [CrossRef]
- Kornochalert, N.; Kantachote, D.; Chaiprapat, S.; Techkarnjanaruk, S. Use of Rhodopseudomonas Palustris P1 Stimulated Growth by Fermented Pineapple Extract to Treat Latex Rubber Sheet Wastewater to Obtain Single Cell Protein. Ann. Microbiol. 2014, 64, 1021–1032. [Google Scholar] [CrossRef]
- Aggelopoulos, T.; Katsieris, K.; Bekatorou, A.; Pandey, A.; Banat, I.M.; Koutinas, A.A. Solid State Fermentation of Food Waste Mixtures for Single Cell Protein, Aroma Volatiles and Fat Production. Food Chem. 2014, 145, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Tropea, A.; Ferracane, A.; Albergamo, A.; Potortì, A.G.; Turco, V. Lo; Di Bella, G. Single Cell Protein Production through Multi Food-Waste Substrate Fermentation. Fermentation 2022, 8, 1–11. [Google Scholar] [CrossRef]
- Khan, M.K.I.; Asif, M.; Razzaq, Z.U.; Nazir, A.; Maan, A.A. Sustainable Food Industrial Waste Management through Single Cell Protein Production and Characterization of Protein Enriched Bread. Food Biosci. 2022, 46, 101406. [Google Scholar] [CrossRef]
- Hosseini, S.; Alireza, K. Methods and Systems of Preparing Cultivated Meat From Blood or Cellular Biomass 2022, 75.
- Bubner, P.; Daley, L.; Levesque-Tremblay, G. Using Organoids And/or Spheroids to Cultivate Meat 2024, 51.
- Reed, J.; Robertson, D.; Rao, K. STRUCTURED HIGH - PROTEIN MEAT ANALOGUE COMPOSITIONS 2021.
- Simpson, S.; Allen, W.E.; Conrado, R.J.; Molloy, S. GAS FERMENTATION FOR THE PRODUCTION OF PROTEIN OR FEED 2020.
- Macur, R.E.; Avniel, Y.C.; Black, R.U.; Hamilton, M.D.; Harney, M.J.; Eckstrom, E.B.; Kozubal, M.A. FOOD MATERIALS COMPRISING FILAMENTOUS FUNGAL PARTICLES AND MEMBRANE BIOREACTOR DESIGN 2024.
- Pearlman, P.S.; Rojas, H.F.C.; Smith, G.; Kirby, G.S. Nutritive Compositions and Methods Related Thereto 2022.
- Kragh, K.M.; Haaning, S.; Meihjohanns, E.; Degn, P.E.; Bak, S.Y.; Eisele, T. METHOD FOR PRODUCING A PROTEIN Publication Classification HYDROLYSATE EMPLOYING AN ASPERGILLUS FUMIGATUS TRIPEPTIDYL PEPTIDASE 2021.
- Onwezen, M.C.; Bouwman, E.P.; Reinders, M.J.; Dagevos, H. A Systematic Review on Consumer Acceptance of Alternative Proteins: Pulses, Algae, Insects, Plant-Based Meat Alternatives, and Cultured Meat. Appetite 2021, 159, 105058. [Google Scholar] [CrossRef]
- Peker, A.; Orkan, Ş.; Aral, Y.; İplikçioğlu Aral, G. A Comprehensive Outlook on Cultured Meat and Conventional Meat Production. Ankara Üniversitesi Vet. Fakültesi Derg. 2024. [CrossRef]
- Hanan, F.A.; Karim, S.A.; Aziz, Y.A.; Ishak, F.A.C.; Sumarjan, N. Consumer’s Cultured Meat Perception and Acceptance Determinants: A Systematic Review and Future Research Agenda. Int. J. Consum. Stud. 2024, 48. [Google Scholar] [CrossRef]
- Can, B.; Majoo, F.; Öztürkcan, A. Consumer Acceptance, Attitude and Knowledge Studies on Alternative Protein Sources: Insight Review. Gıda 2024, 49, 682–702. [Google Scholar] [CrossRef]
- Rothschild-Mancinelli, K.; Germann, S.M.; Andersen, M.R. Bottlenecks and Future Outlooks for High-Throughput Technologies for Filamentous Fungi; 2020; ISBN 9783030295400.
- Finnigan, T.J.A.; Wall, B.T.; Wilde, P.J.; Stephens, F.B.; Taylor, S.L.; Freedman, M.R. Mycoprotein: The Future of Nutritious Nonmeat Protein, a Symposium Review. Curr. Dev. Nutr. 2019, 3, 1–5. [Google Scholar] [CrossRef]
Figure 1.
TAlternative Non-Meat Protein Sources for Sustainable Food Production.
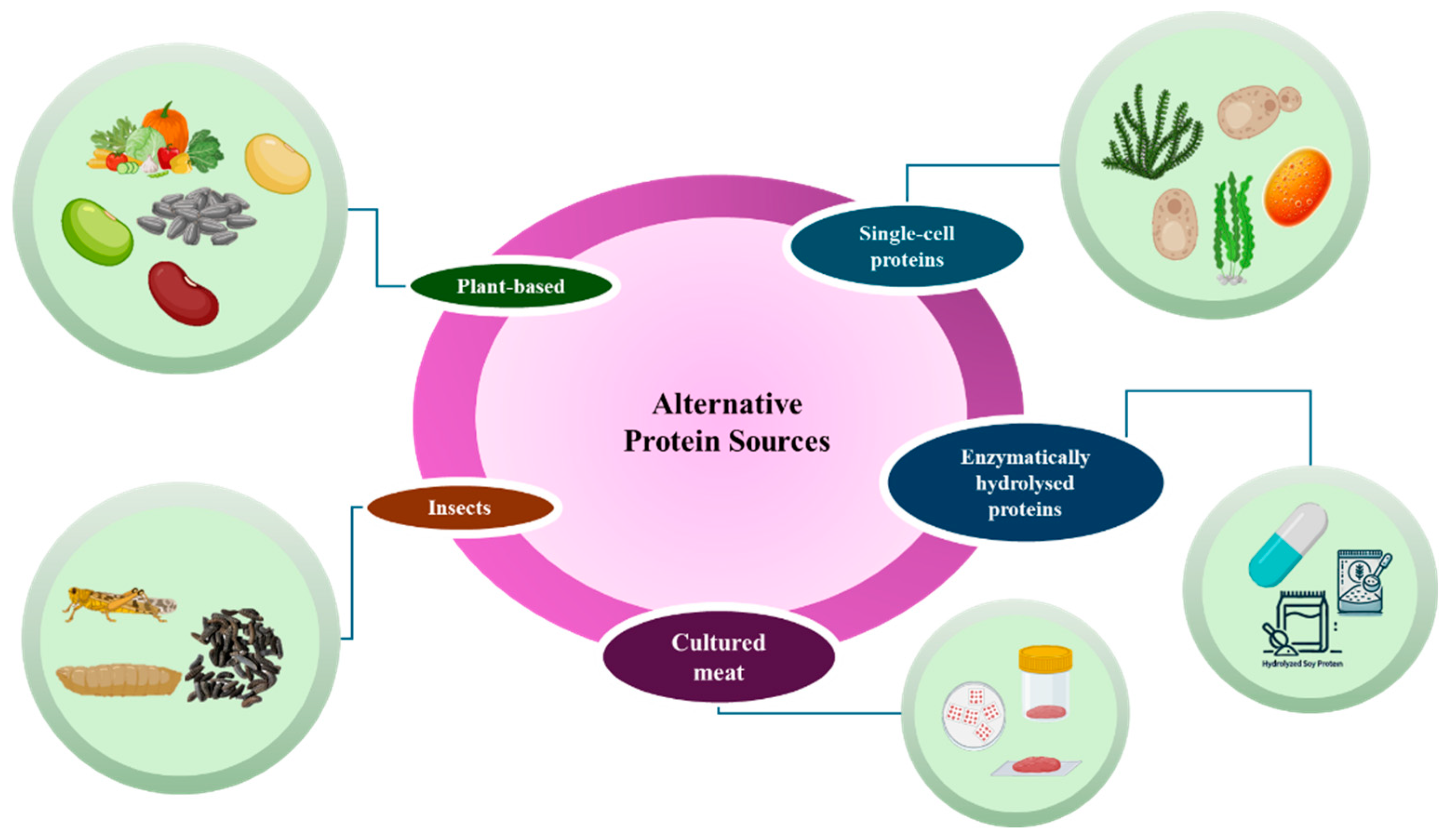
Figure 2.
Evolution of scientific publications over the last 20 years on different non-meat alternative protein sources. (A) Plant-based proteins. (B) Insect proteins are used in food. (C) Cultured meat. (D) Single-cell proteins. (E) Enzymatically hydrolyzed proteins used in food.
Figure 2.
Evolution of scientific publications over the last 20 years on different non-meat alternative protein sources. (A) Plant-based proteins. (B) Insect proteins are used in food. (C) Cultured meat. (D) Single-cell proteins. (E) Enzymatically hydrolyzed proteins used in food.
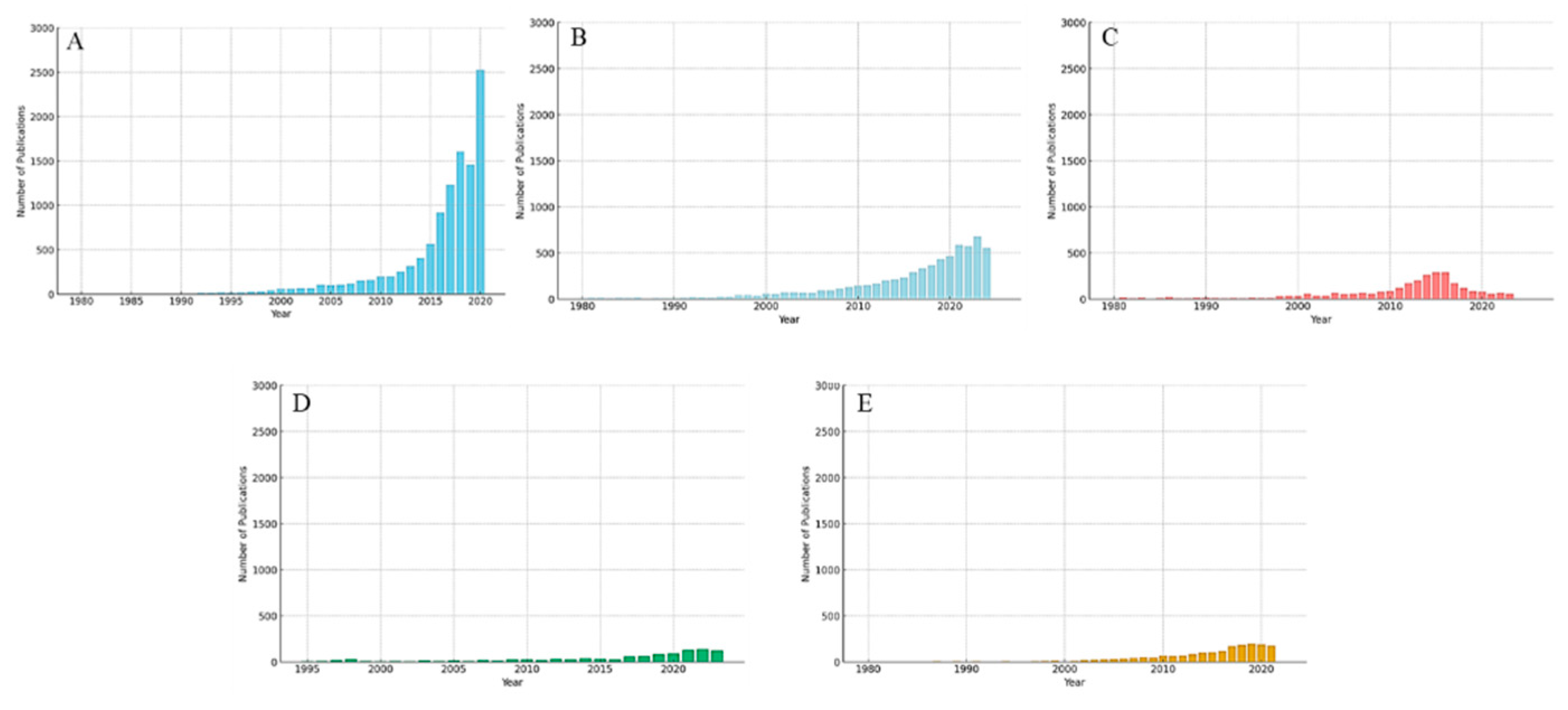
Figure 3.
Single-cell protein production by bacteria, yeast, filamentous fungi and algae by different industrial scale bioreactors.
Figure 3.
Single-cell protein production by bacteria, yeast, filamentous fungi and algae by different industrial scale bioreactors.
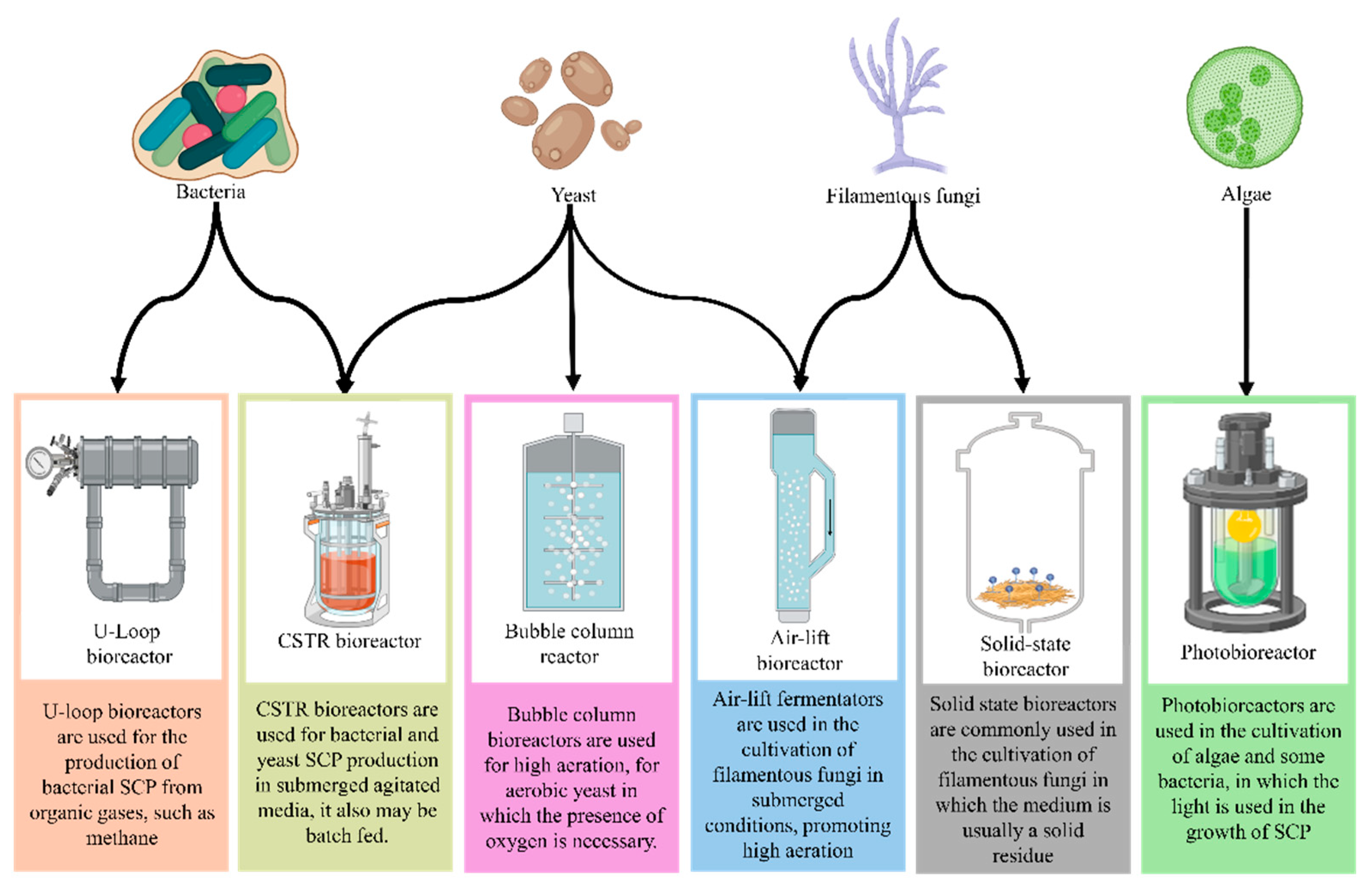
Figure 4.
Exemples of microbial and vegetal enzymes which are used in the food industry for the production of flavours, protein hydrolisates and to improve digestibility of proteins.
Figure 4.
Exemples of microbial and vegetal enzymes which are used in the food industry for the production of flavours, protein hydrolisates and to improve digestibility of proteins.
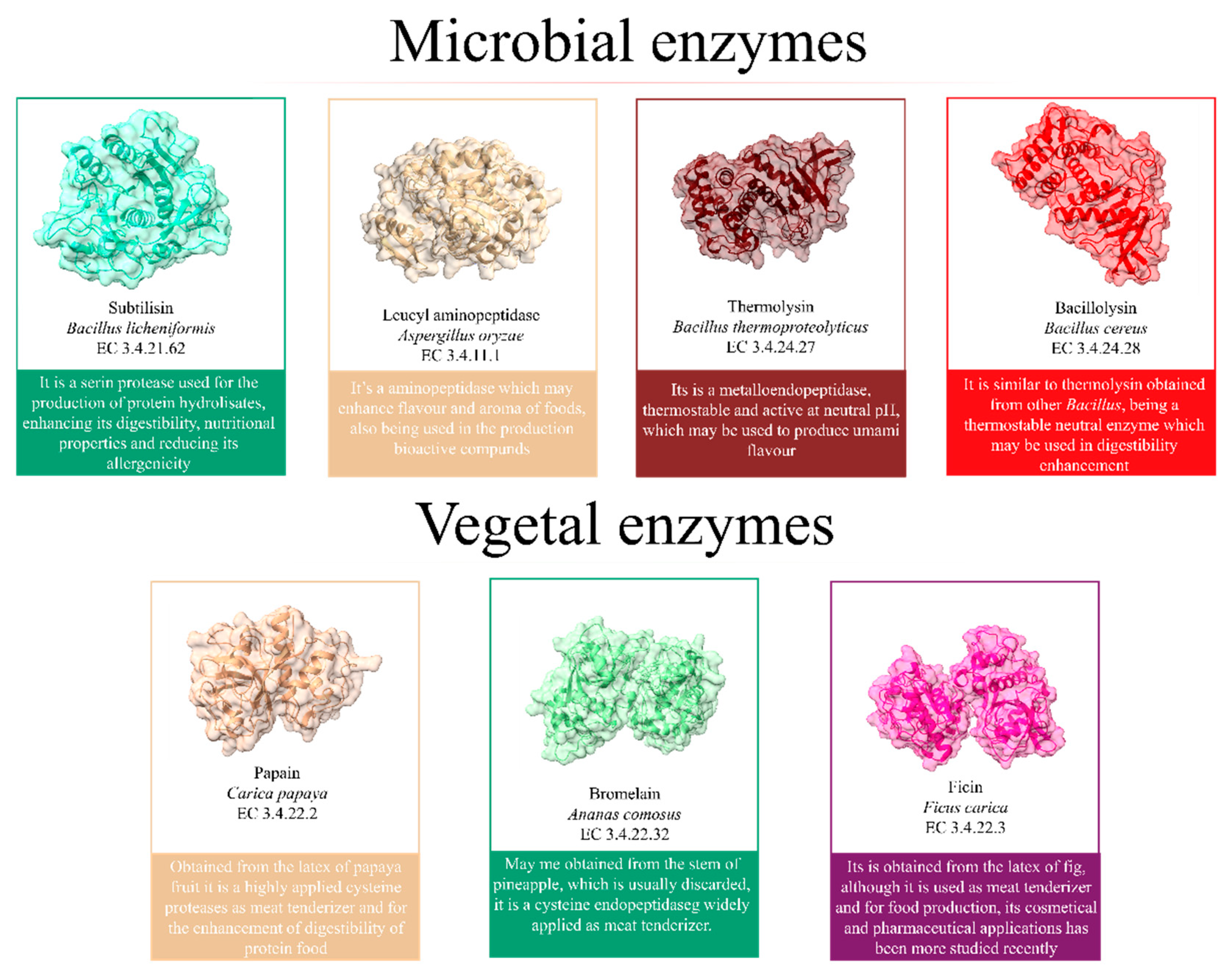
Figure 5.
Advantages of the usage of agroindustrial wastes as the carbon source for single-cell protein synthesis for food application.
Figure 5.
Advantages of the usage of agroindustrial wastes as the carbon source for single-cell protein synthesis for food application.
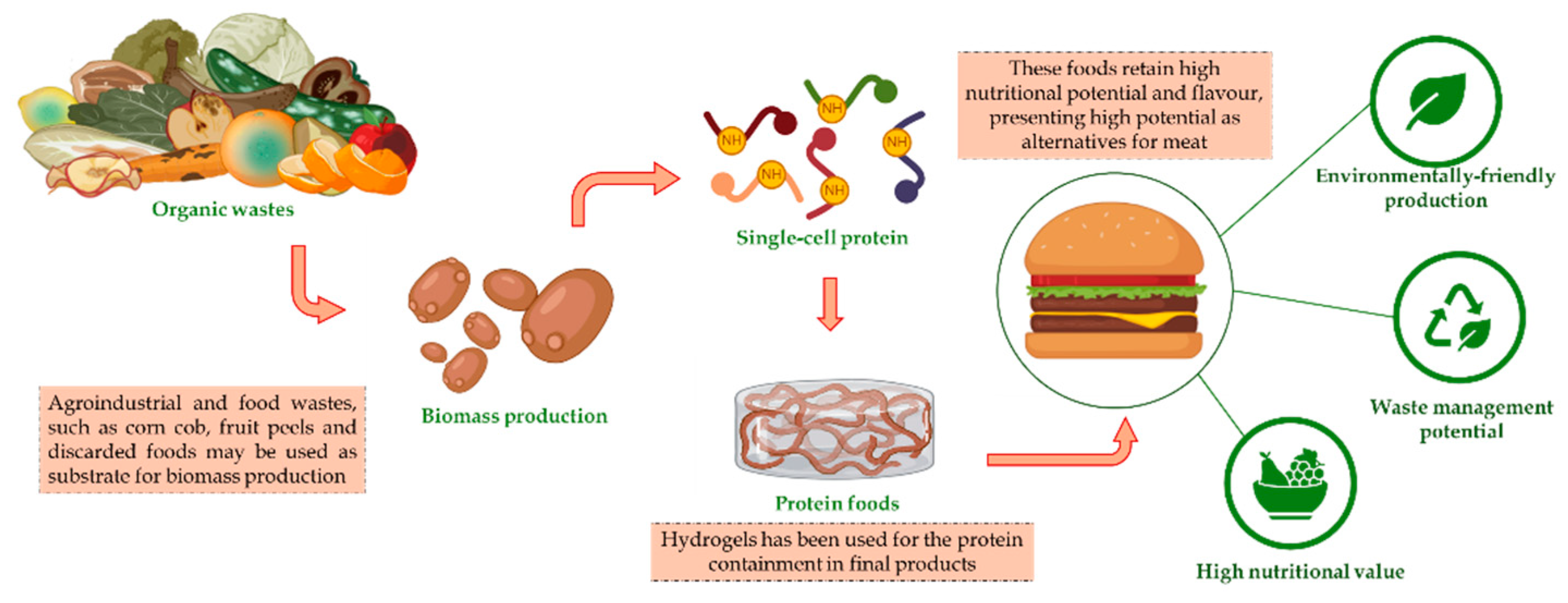
Table 1.
Production of non-meat proteins by biological route for use in cultured meat.
| Substrate used | Protein type | Microorganism/Enzyme | Growing conditions | Sensory analysis | Ref. |
| Soy protein blend with Coprinus comatus powder, distilled water, cornstarch, sunflower oil, konjac flour, carrageenan, egg white powder, etc. | Soy and fungal protein | Coprinus comatus (edible fungi) | 30 °C, 70% relative humidity, for 18h in a fermentation room | Sensory evaluation with 29 participants. The product obtained good scores in texture, flavor, and odor, surpassing other variations processed with different starters | [34] |
| Basmati rice, pearled barley | Fungal (Aspergillus oryzae, Rhizopus microsporus) | Aspergillus oryzae, Rhizopus microsporus | Solid state fermentation, 28-30 °C, 7-10 days. | - | [32] |
| DMEM medium supplemented with Fetal Bovine Serum and penicillin-streptomycin | Fungal (single cell protein) + alginate fiber | Mycelium (fungi) + alginate fiber | Humidified medium with 5% CO2 at 37 °C | Made to simulate meat in a hybrid burger with texture and viability analysis | [33] |
| Soy flour, peanut flour, and residual brewer's yeast | Protein from agro-industrial waste | Alcalase 2.4L and Neutrase 0.8L | Enzymatic hydrolysis at 50 °C and 200 rpm | - | [35] |
| Agricultural waste: wheat straw, beet pulp | Pleurotus eryngii (edible fungi) | Pleurotus eryngii | Temperature of 26 °C for 60 days with 70% hydration | - | [36] |
| Biomass of Ulva spp. collected from the Atlantic Ocean | Algae protein | Proteins from Ulva spp. (green seaweed) | Application of ultrasonic pretreatment (5 min) followed by pH adjustment for protein solubilization and precipitation (pH 12 and 3, respectively | - | [37] |
Table 3.
Production of non-meat-based protein hydrolysates by enzymatic hydrolysis.
| Enzyme | Source | Substrate | Pretreatment | Buffer | Conditions | Separation | Yield (%, w/w) | Reference |
| Alcalase | Bacillus licheniformis | Cottonseed meal | thermal pretreatment | 0.1 M TRIS-HCl | 55º C, pH 8.0 for 100 min | Centrifugation (5000 g for 10 min) | DH: 40.3-47.2% | [43] |
| Black soldier fly larvae | sodium hydroxide | - | 60 ºC and pH 6.85 | Centrifugation (2500 g for 5 min) | 51.4 | [69] | ||
| Soybean protein | - | 0.1 M TRIS-HCl | 50 ºC, pH 8.0, 200 rpm for 120 min | Centrifugation (5000 g for 20 min) | 22.44 | [70] | ||
| Peanut protein | - | 0.1 M TRIS-HCl | 50 ºC, pH 8.0, 200 rpm for 120 min | Centrifugation (5000 g for 20 min) | 16.44 | [70] | ||
| Chia seed expeller | Protein extraction by organic and enzymatic extraction | 45 ºC, pH 9.0 | Centrifugation (5000 g for 10 min) | DH: 15% | [71] | |||
| Canola protein | Grinding and hexane pretreatment | 50 ºC, pH 8.0 | Filtration |
20.6 | [72] | |||
| Soy protein | n-hexane extraction | 50 ºC, pH 8.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 13,0% | [73] | |||
| Wheat gluten protein | 100 mM phosphate buffer (pH 8.5) | 60 ºC, pH 8,5, for 360 minutes | Centrifugation (10000 g for 20 minutes) | DH: 16% | [74] | |||
| Chickpea protein | 100 mM phosphate buffer | 50 ºC, pH 8.0 | Centrifugation (4000 g for 30 minutes) | DH: 27% | [75] | |||
| Protein isolate from pumpkin oil cake | Extraction with hexane | Tris/HCl 0,1 mol/L pH 8.00 | 50 ºC, pH 8.0, For 60 minutes |
- | DH: 53.3% | [76] | ||
| winged bean seed | Extraction with petroleum ether | 60 ºC, pH 8.0, for 300 minutes |
Centrifugation (10000 g for 20 minutes) | DH: 16.1% | [77] | |||
| rice protein | sodium citrate (pH 7.0) | 60 ºC, pH 7.0, for 180 minutes |
Centrifugation (15000 g for 15 minutes) | DH: 33.96%, |
[78] | |||
| chickpea protein | Extraction with hexane | 50 ºC, pH 8.0, for 210 minutes |
Centrifugation (5000 g for 20 minutes) | DH: 14.7% |
[79] | |||
| chickpea seed protein | Treatment with sodium sulphite | 50 ºC, pH 8.0, for 10,50,150 e 300 minutes |
Filtration | DH: 10% |
[80] | |||
| Alcalase + Flavourzyme | Bacillus licheniformis+ Aspergillus oryzae | Canola protein | Grinding and hexane pretreatment | 50 ºC, pH 7.0/8,0 | Filtration |
18.9% | [72] | |
| Chickpea protein | 100 mM phosphate buffer | 50 ºC, pH 8.0 e 50 ºC, pH 7.0 | Centrifugation (4000 g for 30 minutes) | DH: 52% | [75] | |||
| Protein isolate from pumpkin oil cake | Extraction with hexane | Tris/HCl 0,1 mol/L pH 8.00 | 50 ºC, pH 8.0, For 60 minutes |
- | DH: 53.2% | [76] | ||
| Flavourzyme | Aspergillus oryzae | Cottonseed meal | thermal pretreatment | 0.1 M phosphate buffer | 60 ºC, pH 7.0 for 100 min | Centrifugation (5000 g for 10 min) | DH: 27.2-27.5% | [43] |
| Chia seed expeller | Protein extraction by organic and enzymatic extraction | 50 ºC, pH 7.0 | Centrifugation (5000 g for 10 min) | DH:11% | [71] | |||
| Canola protein | Grinding and hexane pretreatment | 50 ºC, pH 7.0 | Filtration |
6.33 | [72] | |||
| Soy protein | Extraction by hexane | 50 ºC, pH 6.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | 8.5% | [73] | |||
| Chickpea protein | - | 100 mM phosphate buffer | 50 ºC, pH 7.0 | Centrifugation (4000 g for 30 minutes) | DH: 27% | [75] | ||
| Protein isolate from pumpkin oil cake | Extraction by hexane | Tris/HCl 0,1 mol/L pH 8,00 | 50 ºC, pH 8.0, For 60 minutes |
- | DH: 37.2% | [76] | ||
| Winged bean seed | Extraction by petroleum ether | 55 ºC, pH 8.0, for 300 minutes |
Centrifugation (10000 g for 20 minutes) | DH: 23.1% | [77] | |||
| rice protein | sodium citrate (pH 7.0) | 50ºC, pH 7.0, for 180 minutes |
Centrifugation (15000 g for 15 minutes) |
DH: 33.94%, |
[78] | |||
| Neutrase | Bacillus amyloliquefaciens | Cottonseed meal | thermal pretreatment | 0.1 M phosphate buffer | 60 ºC, pH 7.0 for 100 min | Centrifugation (5000 g for 10 min) | DH: 29.7-36.4% | [43] |
| Chia seed expeller | Protein extraction by organic and enzymatic extraction | 45 ºC, pH 7.0 | Centrifugation (5000 g for 10 min) | DH: 15% | [71] | |||
| Soy protein | Extraction by hexane | 50 ºC, pH 6,5, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 6.3% | [73] | |||
| Protamex | Bacillus licheniformis e Bacillus amyloliquefaciens | Soy protein | Extraction by hexane | 60 ºC, pH 8.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 5.4% | [73] | |
| Chia seed expeller | Protein extraction by organic and enzymatic extraction | 55 ºC, pH 8.0 | Centrifugation (5000 g for 10 min) | DH: 8% | [71] | |||
| Corolase ® 7089 | Bacillus subtilis | Soy protein | Extraction by hexane | 55 ºC, pH 7.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 6.8% | [73] | |
| Corolase ® 2TS | Bacillus stearothermophilus | Soy protein | Extraction by hexane | 70 ºC, pH 7.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 7.8% | [73] | |
| N-01 | Bacillus subtilis | Soy protein | Extraction by hexane | 55 ºC, pH 7.2, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 4.8% | [73] | |
| Protease | Streptomyces griseus | Amaranth protein |
Extraction by hexane | 50 ºC, pH 8.0, for 360 minutes |
Centrifugation (10000 g for 10 minutes) | DH: 20% | [81] | |
| Quinoa protein | Extraction by hexane | 50 ºC, pH 8.0, for 360 minutes |
Centrifugation (10000 g for 10 minutes) | DH: 78% | [81] | |||
| Papain | Carica papaya | Black soldier fly larvae | sodium hydroxide | 60 ºC and pH 6.85 | Centrifugation (2500 g for 5 min) | 37.8% | [69] | |
| Chia seed expeller | Protein extraction by organic and enzymatic extraction | 55 ºC, pH 7.0 | Centrifugation (5000 g for 10 min) | DH: 6.5% | [71] | |||
| Soy protein | Extraction by hexane | 80 ºC, pH 7.0, for 120 minutes | Centrifugation (5600 g for 130 minutes) | DH: 4.6% | [73] | |||
| Winged bean seed | Extraction by Petroleum ether | 70 ºC, pH 6,5, for 300 minutes |
Centrifugation (10000 g for 20 minutes) | DH: 94.8% | [77] | |||
| Bromelain | Pineapple stem | Winged bean seed | Extraction by Petroleum ether | 45 ºC, pH 6,5, for 300 minutes |
Centrifugation (10000 g for 20 minutes) | DH: 74.1% | [77] | |
| Amaranth protein |
Extraction by hexane | 50 ºC, pH 8.0, for 360 minutes |
Centrifugation (10000 g for 10 minutes) | DH: 46% | [81] | |||
| Quinoa protein | Extraction by hexane | 50 ºC, pH 8.0, for 360 minutes |
Centrifugation (10000 g for 10 minutes) | DH: 76% | [81] | |||
| Amaranth protein |
Extraction by hexane | 50 ºC, pH 7.0, For 120 minutes |
Centrifugation (10000 g for 10 minutes) | DH: 35.4% | [82] |
Table 4.
Advantages and Disadvantages of Fermentation Processes for Single-Cell Protein (SCP) Production.
Table 4.
Advantages and Disadvantages of Fermentation Processes for Single-Cell Protein (SCP) Production.
| Aspect | Advantages | Disadvantages |
|---|---|---|
| Scalability | Fermentation processes can be easily scaled in industrial bioreactors, increasing productivity. | The transition from lab to industrial scale may face challenges in optimization and cost. |
| Growth Rate | Microorganisms grow rapidly under controlled conditions, allowing for short production cycles. | Some microorganisms may require specific and challenging conditions to maintain at large scales. |
| Use of Diverse Substrates | Can utilize agro-industrial residues as substrates, promoting sustainability. | Specific residues require expensive and complex pretreatments to be utilized by microorganisms. |
| Conversion Efficiency | High efficiency in converting substrates into protein-rich biomass. | The presence of inhibitory compounds in the substrate may hinder conversion. |
| Process Control | Fermentation in a controlled environment (pH, temperature, oxygen) ensures product quality and standardization. | Requires constant monitoring and sophisticated equipment, raising operational costs. |
| Production Versatility | Various microorganisms (fungi, yeast, algae, bacteria) can be used to produce different types of SCP. | Regulations or infrastructure needs may limit the selection of the appropriate microorganism. |
| Sustainability | Lower land and water usage compared to traditional agriculture. | Energy consumption in industrial fermentation can be significant, depending on scale and technology. |
| Contamination | Contamination can be prevented through effective sterilization processes. | If sterility control fails, there is a high risk of cross-contamination with other microorganisms. |
| Final Product Flexibility | SCP can be tailored for different nutritional or textural profiles to meet market needs. | The process to modify product characteristics may be expensive and complex. |
| Regulations | Fermentation production models already have established regulatory guidelines for food safety. | New fermented products may face stricter or slower regulatory approval processes. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



