
Submitted:
02 April 2024
Posted:
02 April 2024
You are already at the latest version
Abstract
The expanding global population has increased the demand for sustainable protein sources, and microbial protein (MP) has emerged as a promising alternative. However, conventional carbon (glucose) and nitrogen (ammonia, urea) sources needed for MP production pose environmental and economic issues. This study aims to produce protein using lignocellulosic biomass (LCB) as a carbon source and the nitrogen fixation ability of Klebsiella oxytoca M5A1 as a nitrogen source. The study investigates the pretreatment of LCB (switchgrass), enzymatic hydrolysis, protein quantification, and high-performance liquid chromatography system (HPLC) analysis to track glucose utilization and organic acid production. K. oxytoca M5A1 harnessed free nitrogen from the atmosphere through nitrogen fixation and used abundant, cheap glucose from LCB to produce MP and organic acids as by-products. Protein production occurred in two phases: first within the initial 8 hours and secondly, within the last 16 hours. The highest protein concentration was at 40 hours, with approximately 68.3% protein content. HPLC analysis revealed a dynamic profile of glucose utilization and organic acids (Lactic acid, Propionic acid, Acetic acid, and Succinic acid). K. oxytoca M5A1 exhibited an early high rate of glucose consumption, and conversion to organic acids that were later used for second-phase protein production. The acids profile revealed intra-conversion from one acid to another via metabolic pathways (glycolysis and tricarboxylic acid cycle). Overall, leveraging LCB and the nitrogen-fixing ability of K. oxytoca M5A1 for MP production offers an eco-friendly and cost-effective alternative to traditional protein sources, contributing to a sustainable circular economy.
Keywords:
Microbial protein
; Lignocellulosic biomass
; switchgrass hydrolysate
; Klebsiella oxytoca M5A1
; Fermentation
; Organic acids.
1. Introduction
With the rapid growth of the world population, there's an increasing demand for sustainable and efficient protein sources. This demand stems from the global need for both feed and food, as well as the environmental considerations associated with traditional agriculture and livestock production [1]. While traditional sources such as plant-based and animal-based proteins have long been the cornerstone of human nutrition, they are increasingly facing challenges ranging from environmental impact to scalability and cost efficiency. Plant-based protein production often requires extensive land use, water consumption, and chemical inputs, leading to deforestation, soil degradation, and habitat loss [2]. Additionally, livestock farming contributes significantly to greenhouse gas emissions, water pollution, and biodiversity loss [2]. In recent years, significant biodiversity loss and deforestation are observed due to forest conversion. About 30–50 % of arable land, 20 % of global energy production and 70 % of fresh water are used for this purpose [3]. Therefore, in response to these challenges, microbial protein (MP) production has emerged as a promising alternative, offering significant advantages in terms of sustainability, low carbon footprint, and cost-effectiveness.
MP production involves the cultivation of microorganisms, such as bacteria, fungi, algae, and yeast, under controlled conditions to yield high-quality protein [4]. These microorganisms can be engineered or naturally occurring, and can efficiently convert simple raw materials into valuable products. The production of protein requires sugar, especially glucose, as a carbon and energy source [5]. Traditionally, conventional sugars have been the primary choice for this purpose. However, sugar production is often costly and relies on intensive agricultural practices that can lead to environmental degradation and increased carbon footprint [6]. High demand for these sugars in various industries can lead to competition for limited resources, potentially driving up prices and exacerbating issues of food security and affordability. In addition to carbon sources, nitrogen is also essential for MP synthesis, typically supplied in the form of ammonia or urea. However, the production of these nitrogen sources is non-renewable, energy-intensive, and can lead to environmental pollution if not properly managed [7].
Leveraging lignocellulosic biomass (LCB) as a carbon source and harnessing the nitrogen-fixing abilities of bacteria, such as K. oxytoca M5A1, offers a revolutionary approach to meet the rising demand for protein while addressing concerns related to sustainability, carbon footprint, and cost efficiency. LCB is a renewable and abundant feedstock that offers a sustainable substrate for MP production, eliminating the need for conventional glucose.
Over the years, several works have been done, specifically on the utilization of LCB or agro-waste materials for protein production, as well as other valuable chemicals. Rajoka et al. [8] used mixed substrates containing beet pulp hydrolysate supplemented with molasses and glucose for fermentation of Candida utilis and Brevibacterium lactofermentum during MP production. MP has been synthesized using soybean hull biomass feedstock obtained from soya bean oil extraction as substrate [9]. However, the protein yield are reportedly low to attract industrial interest. One possible reason is the lack of nitrogen sources during the fermentation of LCB in the aforementioned studies. To overcome this, we use K. oxytoca M5A1 that have the unique ability to fix atmospheric nitrogen using inherent Nitrogenase. Nitrogenase reduces atmospheric nitrogen (N2) by breaking the strong triple bond between nitrogen atoms, resulting in the formation of ammonia (NH3) [10]. We envisage that the use LCB as substrates reduces the cost of raw materials, while the nitrogen-fixing capability of bacteria eliminates the need nitrogen sources, thereby lowering the fermentation costs during MP synthesis. This approach would potentially not only makes MP production more economically viable but also aligns with global efforts to reduce the environmental footprint of food/feed production.
2. Methodology
2.1. Media
Luria-Bertani agar (LB agar) contains (in 500ml) 5 g tryptone, 7.5 g agar, 2.5 g yeast extract and 5 g sodium chloride (NaCl). All the reagents were initially mixed in 400ml distilled water, and the pH was adjusted to 7.0 by using sodium hydroxide (NaOH) solution (1 N). The mixture was then top up to 500ml and autoclaved for 20min. After autoclaving, the media cooled in a water bath and poured into petri dishes under aseptic conditions. The plate was stored in the fridge for further use. The minimal medium used for fermentation was prepared without sucrose and contains 0.1 g of CaCl2−2H2O, 25 g of Na2HPO4·7H20, 0.25 g of MgSO4·7H2O, 3 g of KH2PO4, 0.25 mg of Na2MoO4·2H2O, 1 g of NaCl, and 2.9 mg of FeCl2·4H2O in 1L distilled water. Super Optimal Broth (SOB) medium used as inoculum medium contains 0.5 g of NaCl, 2.4 g of MgSO4, 5 g of yeast extract, 0.186 g of KCl, and 20 g of Peptone in 1L distilled water.
2.2. Culturing of Klebsiella oxytoca M5A1
A fresh culture of K. oxytoca M5A1 (obtained from Microbiology department, North Dakota State University) was sub-cultured on LB agar plate. Using a sterile wire loop, a distinct colony of K. oxytoca M5A1 was carefully streaked on the LB agar plate under aseptic condition and the plates were incubated for 24 h at 30°C. After incubation the overnight culture was prepared by aseptically introducing a swab of K. oxytoca M5A1 into 10ml SOB broth and incubated for 24 h (30°C, 200rpm).
2.3. Biomass Pretreatment
The LCB (Switchgrass) was pretreated in alkaline using aqueous ammonia. According to Hammed et al. (2021), a 1-L screw-capped Pyrex bottle, 1:6 solid-liquid ratio of 10% (w/v) aqueous ammonia and biomass sample was mixed together and placed at 60°C for 24 h. After 24 h, the pretreated samples were washed (using vacuum filtration method) with distilled water until the pH reached approximately 7.5. The washed pretreated biomass was spread on the workbench and left to air dry for few hours [11].
2.4. Enzymatic Hydrolysis of Biomass
Alkaline pretreated switchgrass was enzymatically hydrolyzed with enzyme cellulase (Cellic HTec, Novozymes, Denmark) in 250ml Erlenmeyer flasks. The mixtures contained the pretreated switch grass in sodium citrate buffer, supplemented with 0.04% sodium azide to prevent an unwanted growth of microorganisms, and 3% (of biomass loading) enzyme cellulase. The mixture was hydrolyzed for 72hr at 50°C (130rpm). After hydrolysis, the Switchgrass hydrolysate was separated by filtration to remove residues from the mixture and autoclaved for 20min to prevent contamination.
2.5. Analytical Methods: Glucose Utilization and Organic Acids Production
The glucose content in the hydrolyzed biomass, glucose utilization and organic acids produced during fermentation were analyzed and quantified with HPLC system using LabSolutions Software for Shimadzu LC-2050C HPLC. The mobile phase was sulfuric acid (0.01 N) at a flow rate of 1.0 mL/min. The samples were filtered through a 0.45µm nylon filter then injected into the column. The microcentrifuge tube and 2mL autosampler vial with sample identifier(s) were prepared and labeled accordingly. The sample container was inverted for mixing and 2mL of homogenous solution was transferred to the labeled microcentrifuge tube. The sample aliquot was centrifuge for minimum of 3.5 minutes at 10,000 rpm. The supernatants were filtered through a 1cc syringe connected to a 0.45µm nylon filter into a 2mL autosampler vial. The samples were analyzed and results were reported in g/L.
2.6. Fermentation Conditions and Growth Profiling
The fermentation media was prepared using minimal salt solution and switchgrass hydrolysate. The solutions were separately prepared, autoclaved and placed in a water bath to cool. After cooling, the minimal salt solution and switchgrass hydrolysate (approximately 10g/L) were mixed in 1:1 v/v ratio in an Erlenmeyer flask, with pH of 6.5-7.5. The fermentation media consisting biomass hydrolysate was inoculated with 5% of overnight culture and incubated for 48 h (30°C, 200rpm). The samples were collected at 0, 8, 16, 24, 32, 40, and 48 h. The optical density of each collected samples were taken (OD 600nm) and microbial growth was determined, with the use of microplate Spectrophotometer (Infinite M Nano by Tecan).
2.7. Protein Quantification
At each time interval, the amount of protein was quantified using Bradford assay protein quantification protocol. The protein standard curve was established using the Bovine Serum Albumin (BSA) assay [12]. The sample was pipetted in a test-tube and the Bradford reagent was added in ration 1:5 v/v. The mixture was mixed and incubated at room temperature for 20min and the absorbance was taken at wavelength of 595nm using Tecan infinite M Nano microplate reader.
3. Result and Discussion
3.1. Growth Profile K. oxytoca
K. oxytoca strains as a nitrogen-fixing microorganisms require glucose in their fermentation media as a carbon source. However, the traditional production and price of refined glucose that is utilized during fermentation is expensive and may have a negative impact on the environment. In order to cost-effectively culture K. oxytoca, biomass hydrolysate is employed as an alternative source of carbon, substituting refined glucose. Biomass hydrolysates are more economical and derived from renewable sources that align with sustainability development (Mussatto & Teixeira, 2010). To demonstrate the suitability of K. oxytoca for industrial fermentation processes, its ability to grow in biomass hydrolysate is of paramount importance. Therefore, we conducted this experiment to observe how K. oxytoca will thrive in the presence of an alternative source of carbon such as switch grass hydrolysate.
Following the preliminary experiment and the assurance that nitrogen-fixing K. oxytoca M5A1 is able to utilize hydrolysate as a carbon source during fermentation, we decided to further our experiment on the activity of hydrolysate and the response of K. oxytoca M5A1. Fermentation process was carried out at 30°C for 48 h, under the pH of 6.5 and 200rpm. The medium contained 10 g/L equivalent of glucose from the hydrolysate, and we monitored the growth profile of the bacterial strain with 8 h intervals for 48 h (Figure 1).
The analysis of the growth rate provides a dynamic perspective on how the K. oxytoca M5A1 responds to the conditions in the fermentation broth. We discovered that from 0 to 8 h, there is a substantial increase in the average absorbance at OD600nm. This indicates a rapid growth rate during the initial phase. Beyond 8 h, the growth rate appears to slow down, but there is a consistent and steady increase in the average values. The highest average value was observed at 48 h, during the experiment. The initial phase of rapid growth is attributed to the easy access to bioavailable nutrients, while the slow growth phase is a result of nutrient depletion.
The obtained result is similar to previous work on growth of K. oxytoca strains in switchgrass hydrolysates. Cha et al. [13] reported that a substantial increase in growth of K.oxytoca KCTC1686 strains (CHA004) was experienced between 0 –10 h during cultivation with sunflower stalk hydrolysate. Similar to this, we also experienced a substantial increase in growth rate of K. oxytoca M5A1 between the 0 - 8 h using switch grass hydrolysate. Park et al. [14] reported that engineered K. oxytoca MAK01 strains could grow using the microalgae hydrolysate as the main substrate, and all clones experienced exponential growth at 4 hours and the stationary phase at 10 hours. Contrary to this, our strain of interest K. oxytoca M5A1 experienced slower growth between 16 h and 24h. In another study conducted by Cheng et al. [15], it was observed that K. oxytoca M5A1 could grow in corncob hydrolysate. The strains of K. oxytoca M5A1 used in this study demonstrated proliferation in switchgrass hydrolysate. However, a thorough understanding of the observed variation in the growth phase may be achieved by studying the changes in glucose utilization and organic acid concentrations.
3.2. Glucose Utilization of K. oxytoca M5A during Fermentation
Glucose, particularly, is a common carbon source for microorganisms. Research has demonstrated that the production of microbial biomass is closely linked to the availability of reducing sugars [5]. Microorganisms assimilate glucose to build cellular components, such as amino acids, nucleotides, and other organic molecules required for growth and reproduction. Therefore, glucose utilization efficiency of K. oxytoca M5A1 is essential for the production of protein and organic acids. To ascertain whether the previously documented rapid growth is correlated with the glucose utilization rate of our strain, we monitored the glucose consumption rate of K. oxytoca M5A1 utilizing an HPLC system. The samples were taken at time intervals (every 8 h) and the glucose concentration was estimated (Figure 2).
The results obtained indicated a significant decrease in glucose concentration from 12.84 g/L to 3.80 g/L within the first 8 hours. This observation suggests active glucose consumption by K. oxytoca M5A1, which aligns with the previously observed growth pattern. This phase is typically characterized by rapid growth and metabolic activity of microorganisms, as they utilize available nutrients for energy and biomass production. Consequently, a faster rate of glucose utilization is generally associated with accelerated cell division. By 16 h, the glucose concentration had decreased to 0 g/L, indicating that K. oxytoca M5A1 had exhausted the available glucose resources. This stage coincides with the slowdown experienced during their growth.
In the context of sugar utilization, several studies have highlighted the capabilities of different microorganisms. Yu et al. [16] reported in their work that T. dermatis 32,903 exhibited a sugar utilization preference for glucose compared to xylose, as glucose utilization rate was higher in the first 24 h. Abdel-Rahman et al. [17] reported that Bacillus coagulans Azu-10 completely consumed glucose within 4–8 h. Another study using glucose from Golenkinia sp. hydrolysate reported that glucose was completely used by K. oxytoca MAK01 within 7.5 h [14]. A study also reported that lignocellulosic biomass glucose was completely utilized within 8-12 h of fermentation by K.oxytoca KCTC1686 strains [13].
Overall, K. oxytoca M5A1 effectively utilized glucose in switchgrass hydrolysate, with a rapid initial consumption followed by complete exhaustion. This utilization occur through metabolic pathways such as glycolysis and fermentation [18]. Bacteria can absorb multiple sugars into the cytoplasm and utilize them for ATP production (the energy currency of the cell) via glycolysis. Glycolysis is the most primitive metabolic system for energy acquisition and is responsible for the breakdown of glucose into organic acids such as pyruvic acid and lactic acid [19] . The fermentation pathway can lead to the production of various end products such as protein, depending on the conditions and strains used.
3.3. Microbial Protein Content of K. oxytoca M5A1
K. oxytoca M5A1 simultaneously fix nitrogen during fermentation. This involves the conversion of atmospheric nitrogen (N2) into ammonia (NH3) or other nitrogen compounds. This ammonia can then be utilized in the synthesis of amino acids, the building blocks of proteins. However, could nitrogen fixation capability of K. oxytoca M5A1 influence its protein synthesis efficiency, particularly when utilizing glucose from biomass hydrolysate? The production of microbial protein was carefully monitored at time interval using Bradford protein quantification assay at wavelength of 595nm. The Protein production of K. oxytoca M5A1 is shown in Figure 3.
During the initial phase of glucose utilization (0-8 h), there is a noticeable increase in protein concentration from 53.41 µg/ml to 284.73 µg/ml. This suggests active protein synthesis during the early growth phase when glucose is being consumed. The protein concentration continues to rise, but at a slower rate between 16 h and 24 h. This is the point where glucose utilization plateaus, but protein production is still occurring (slowly) as the microbes adjust to changing metabolic conditions. As glucose is completely consumed, the protein concentration continues to increase through 32 h, reaching a peak at 40 h (683.46 µg/ml). This marks the second phase of protein production, suggesting that K. oxytoca M5A1 may be utilizing alternative carbon sources present in the hydrolysate during protein production. Although, we only accounted for glucose in our study, but previous studies have reported that cellulosic biomasses display considerable diversity in their sugar composition upon hydrolysis, resulting in a range of sugar mixtures. This variability is expected to influence the utilization of sugars [20]. At 48 h, the protein concentration in the sample has diminished to 376.98 µg/ml, suggesting the occurrence of protein degradation or precipitation. This observed reduction may be attributed to alterations in the pH level, which are likely induced by the metabolic processes.
Previous research has demonstrated the utilization of various substrates for microbial protein production, highlighting the importance of substrate selection in achieving desirable protein yields. Rajoka et al. [8] reported in their study that the total sugars in the hydrolysate of beet pulp, along with glucose and molasses, were almost completely consumed by Candida utilis and B. lactofermentum, which resulted in production of more cell mass and 54.5% crude proteins. Thiviya et al. [5] also conducted a comparative study using different fruit peel wastes as substrate for palmyrah toddy yeast, and concluded that papaya peel waste is a suitable substrate for protein biomass production having 52.4% protein content. Two strains of Bacillus subtilis MR10 and TK8 were cultured in Soybean hull obtained from soya bean oil extraction, and the protein content after fermentation was 25.6% for MR10 and 26.6% for strain TK8 [21]. Kunasundari et al. [22] utilized a synthetic growth medium for cultivation of Cupriavidus necator to produce protein, and they reported between 40-46% protein content. Zheng et al. [23] reported that protein content of Candida arborea AS1.257 cells produced from the rice straw hydrolysate was improved by adding nitrogen source such as ammonia solution and urea to fermentation medium. Unlike these studies, such as that by Kunasundari et al. [22] and Zheng et al. [23], which utilize urea as a source of nitrogen for protein production, our study is unique in that we do not employ additional nitrogen sources in our fermentation medium. Instead, we have taken advantage of nitrogen-fixing bacteria, specifically K. oxytoca M5A1, to utilize free nitrogen from the atmosphere. In our study, we cultured nitrogen-fixing K. oxytoca M5A1 while utilizing switchgrass hydrolysate as a substrate for protein production, obtaining approximately 68.3% protein content. The variations observed in these studies confirm that the yield of microbial protein produced during fermentation is not only influenced by the type of substrate used but also microbial strain employed.
Overall, the data suggests a dynamic relationship between glucose utilization and protein production. K. oxytoca M5A1 demonstrates the capability to fix nitrogen from the atmosphere, and utilize sugars derived from hydrolyzed biomass, such as switch grass, for protein synthesis. This ability to leverage cheap and readily available biomass sugars and use free nitrogen for protein production significantly reduces the cost of industrial-scale production. Furthermore, the availability of biomass as a substrate enhances its sustainability and renewability compared to traditional refined sugars. This innovative approach not only contributes to a reduction in the carbon footprint but also promotes an environmentally friendly and economically viable protein production process.
3.4. Organic Acids Profile During Nitrogen Fermentation to Protein
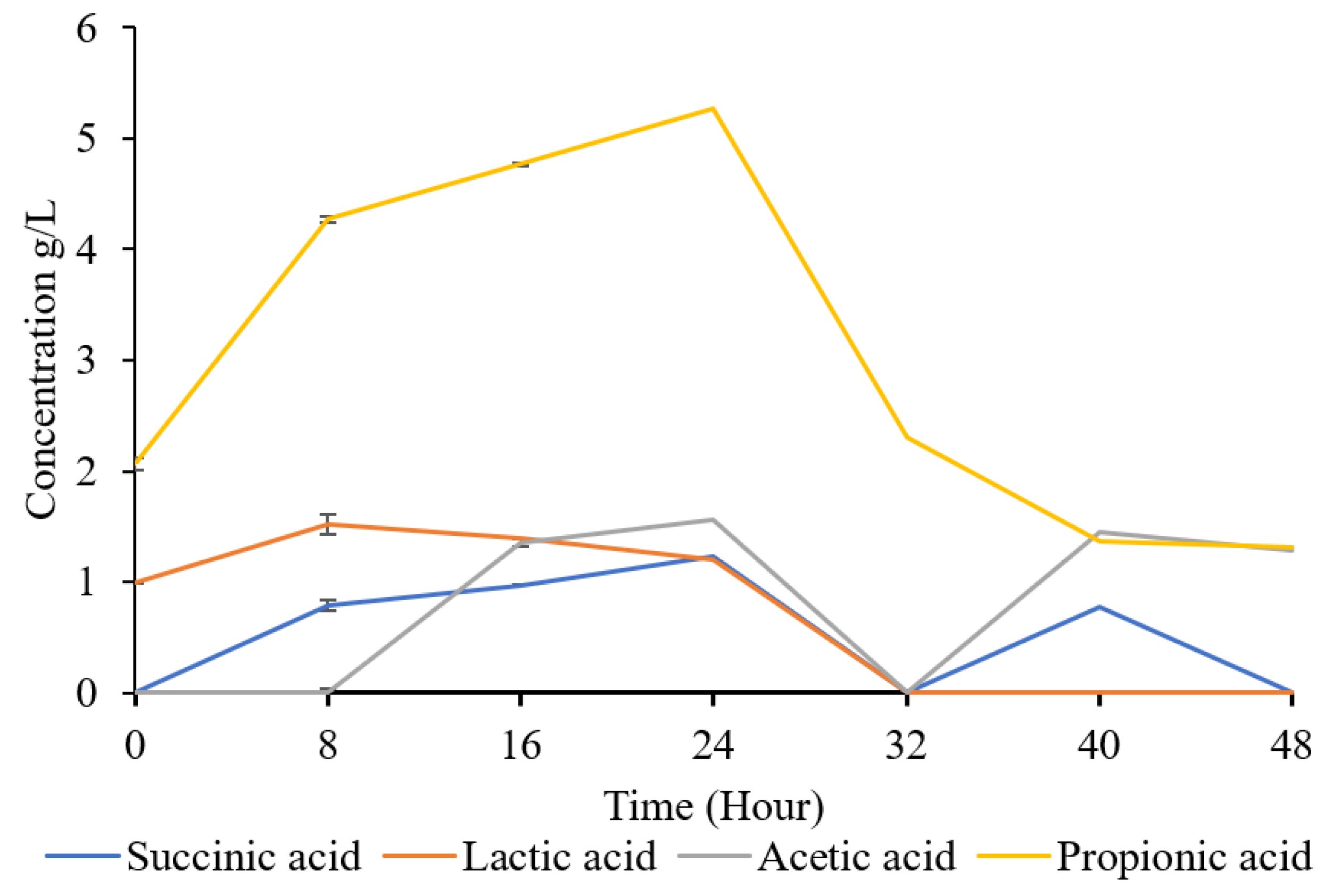
During fermentation, microorganisms utilize substrates through enzymatic reactions, leading to the production of organic acids as metabolic by-products. Various organic acids can be produced, including but not limited to acetic acid, lactic acid, succinic acid, propionic acid, citric acid, and formic acid [24]. The specific types and quantities of organic acids produced depend on factors such as the microbial strains used, fermentation conditions (e.g., pH, temperature, nutrient availability), and the substrate [25] [26]. An understanding of organic acid profile during fermentation will reveal the active metabolic pathway and activities. Since the metabolic dynamics of organic acid during nitrogen fixation of K. oxytoca M5A1 is unclear, we studied the production of organic acid over the course of fermentation (Figure 4).
We observed the presence of lactic acid (LA), succinic acid (SA), acetic acid (AA), and propionic acid (PA) during the fermentation process. Unlike SA and AA without noticeable quantity at 0 h, LA and PA started at moderate concentrations of 0.99 g/L and 2.06 g/L, respectively, because of their initial presence in the hydrolysate (preliminary result not shown), suggesting a relatively stable environment.
During the course of fermentation, the organic acid profile follow different patterns. PA production was highest and continue to increase until 24 h, LA increase slightly till 24 h, SA increase steadily till 24 h and then decline while AA synthesis started after 8 h and continue to increase until 24 h. Although LA, SA and AA were undetectable at 32 h, SA and AA were detected after.
Earlier findings have affirmed that these organic acids can be a primary product or by-product of many microorganisms. Phosriran et al. [27] reported that an engineered strain of K. oxytoca M5A1 (KMS006) produced SA at a concentration of 4.82 g/L, while the wild-type K. oxytoca M5A1 strain produce 1.47g/L SA. Although our study observed a concentration of 1.2 g/L of SA, the yield of 1.47 g/L by the wild-type strain in Phosriran et al.'s study aligns closely with our findings, suggesting consistency between our results and those of the wild-type strain. However, the engineered strain's significant higher yield may be attributed to its specific modifications for enhanced SA production, highlighting the impact of genetic engineering on metabolic pathways. Abdel-Rahman et al. [17] in their study reported that Bacillus coagulans Azu-10 exhibits potential in the production of LA from substrates of xylose and glucose derived from cellulosic biomass. In contrast to our study, Abdel-Rahman et al. [20] obtained as high as 50-102 g/L LA and 0.280-2.61 g/L AA as a by-product. However, they utilized a higher concentration of sugar ranging from 50 to 150 g/L, compared to 10 g/L used in our study.
The dynamic production of organic acids during fermentation with K. oxytoca M5A1 is through various metabolic pathways, including glycolysis, the TCA cycle, and amino acid metabolism. K. oxytoca, like other lactic acid bacteria, can produce lactic acid via homolactic fermentation, where glucose is converted into pyruvate via glycolysis, and then into lactic acid by lactate dehydrogenase [28]. Similar to lactic acid, succinic acid is also synthesized from pyruvate through the tricarboxylic acid (TCA) cycle [27]. During succinate synthesis, a significant amount of NADH is consumed. This process may also result in the generation of acetic acid through the conversion of acetyl-CoA to NADH, thereby co-occurring with succinic acid formation [29]. Propionic acid is produced through the deamination of amino acids, which involves the removal of an amino group from the amino acid molecule such as threonine, methionine, isoleucine and valine [30].
Similar to the rate at which K. oxytoca M5A1 consumes sugar, it also utilizes alternative carbon sources. The growth and protein synthesis was not inhibited by the presence of the organic acids, rather they utilized them as carbon source. This can be as a result of K.oxytoca species capability of growing in acidic conditions, as low as pH of 5 [31]. The time point at 32 hours appears crucial, as all the acids exhibit decrease in concentration concise with the beginning of second phase of protein production. K. oxytoca M5A1 completely consumed lactic acid, acetic acid, succinic acid, and a large percentage of propionic acid. In the second phase of protein synthesis the concentration doubled. When the supply of sugars is exhausted, bacteria can metabolize these organic acids to generate ATP. This process is facilitated by enzymes that catalyze the reduction of organic acids, converting them back into simpler molecules that can be further metabolized for energy production [32]. Overall, the observed trend in this study indicates that organic acids are been produced as a by-product and utilized as energy sources during fermentation after the complete depletion of glucose. Presumably, K. oxytoca M5A1 synthesis of organic acid occur for different reasons: (1) to produce acids that create an unfavorable environment for the growth of other microbes, (2) to make glucose unavailable for other microbes and (3) to produce acids for energy storage and survival when glucose is completely depleted.
4. Conclusion
Nitrogen-fixing bacteria K. oxytoca M5A1 demonstrated efficiency in utilizing lignocellulosic switchgrass hydrolysate as a carbon source for microbial protein production. Meaning that the strain is able to use a cheaper source of carbon while harnessing free nitrogen from the atmosphere. This offers a sustainable alternative to the use of conventional glucose and nitrogen in microbial protein synthesis. The dynamic patterns observed in organic acid profile highlight the complex interplay of microbial metabolism during fermentation. This study does not only advance our understanding of microbial behavior in response to different carbon sources but also holds promising implications for industrial applications. The utilization of LCB hydrolysate presents itself as a sustainable and economically viable strategy, aligning with global initiatives to reduce environmental impact and promote the circular economy. Future research endeavors should further investigate the underlying mechanisms governing the observed dynamics, optimization of microbial fermentation processes, molecular engineering, and techno- economic analysis of the procedures for enhanced productivity and efficiency.
References
- Pexas, G.; Doherty, B.; Kyriazakis, I. The Future of Protein Sources in Livestock Feeds: Implications for Sustainability and Food Safety. Front. Sustain. Food Syst. 2023, 7. [Google Scholar] [CrossRef]
- Sobhi, M.; Zakaria, E.; Zhu, F.; Liu, W.; Aboagye, D.; Hu, X.; Cui, Y.; Huo, S. Advanced Microbial Protein Technologies Are Promising for Supporting Global Food-Feed Supply Chains with Positive Environmental Impacts. Sci. Total Environ. 2023, 894, 165044. [Google Scholar] [CrossRef] [PubMed]
- Owsianiak, M.; Pusateri, V.; Zamalloa, C.; De Gussem, E.; Verstraete, W.; Ryberg, M.; Valverde-Pérez, B. Performance of Second-Generation Microbial Protein Used as Aquaculture Feed in Relation to Planetary Boundaries. Resour. Conserv. Recycl. 2022, 180, 106158. [Google Scholar] [CrossRef]
- Matassa, S.; Boon, N.; Pikaar, I.; Verstraete, W. Microbial Protein: Future Sustainable Food Supply Route with Low Environmental Footprint. Microb. Biotechnol. 2016, 9, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Thiviya, P.; Gamage, A.; Kapilan, R.; Merah, O.; Madhujith, T. Production of Single-Cell Protein from Fruit Peel Wastes Using Palmyrah Toddy Yeast. Fermentation 2022, 8, 355. [Google Scholar] [CrossRef]
- Aguilar-Rivera, N. Bioindicators for the Sustainability of Sugar Agro-Industry. Sugar Tech 2022, 24, 651–661. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of Nitrogen in Agriculture and Environment: Agronomic, Eco-Physiological and Molecular Approaches to Improve Nitrogen Use Efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef]
- Rajoka, M.I.; Ahmed, S.; Hashmi, A.S.; Athar, M. Production of Microbial Biomass Protein from Mixed Substrates by Sequential Culture Fermentation of Candida Utilis and Brevibacterium Lactofermentum. Ann. Microbiol. 2012, 62, 1173–1179. [Google Scholar] [CrossRef]
- Wongputtisin, P.; Khanongnuch, C.; Kongbuntad, W.; Niamsup, P.; Lumyong, S.; Sarkar, P.K. Use of Bacillus Subtilis Isolates from Tua-Nao towards Nutritional Improvement of Soya Bean Hull for Monogastric Feed Application. Lett. Appl. Microbiol. 2014, 59, 328–333. [Google Scholar] [CrossRef]
- Burén, S.; Rubio, L.M. State of the Art in Eukaryotic Nitrogenase Engineering. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Hammed, A.; Polunin, Y.; Voronov, A.; Pryor, S.W. Glucan Conversion and Membrane Recovery of Biomimetic Cellulosomes During Lignocellulosic Biomass Hydrolysis. Appl. Biochem. Biotechnol. 2021, 193, 2830–2842. [Google Scholar] [CrossRef] [PubMed]
- Spencer, V.A.; Davie, J.R. Isolation of Proteins Cross-Linked to DNA by Cisplatin. In Protein Protocols Handbook, The; Humana Press: New Jersey, 2002; ISBN 978-1-59259-169-5. [Google Scholar]
- Cha, J.W.; Jang, S.H.; Kim, Y.J.; Chang, Y.K.; Jeong, K.J. Engineering of Klebsiella Oxytoca for Production of 2,3-Butanediol Using Mixed Sugars Derived from Lignocellulosic Hydrolysates. GCB Bioenergy 2020, 12, 275–286. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, M.A.; Kim, Y.J.; Kim, Y.-C.; Chang, Y.K.; Jeong, K.J. Engineering of Klebsiella Oxytoca for Production of 2,3-Butanediol via Simultaneous Utilization of Sugars from a Golenkinia Sp. Hydrolysate. Bioresour. Technol. 2017, 245, 1386–1392. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Liu, Q.; Zhang, J.-A.; Li, J.-P.; Xu, J.-M.; Wang, G.-H. Improved 2,3-Butanediol Production from Corncob Acid Hydrolysate by Fed-Batch Fermentation Using Klebsiella Oxytoca. Process Biochem. 2010, 45, 613–616. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, S.; Zhang, Y.; Lu, M.; Sha, Y.; Zhai, R.; Xu, Z.; Jin, M. A Novel Fermentation Strategy for Efficient Xylose Utilization and Microbial Lipid Production in Lignocellulosic Hydrolysate. Bioresour. Technol. 2022, 361, 127624. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Hassan, S.E.-D.; Alrefaey, H.M.A.; Elsakhawy, T. Efficient Co-Utilization of Biomass-Derived Mixed Sugars for Lactic Acid Production by Bacillus Coagulans Azu-10. Fermentation 2021, 7, 28. [Google Scholar] [CrossRef]
- Förster, A.H.; Gescher, J. Metabolic Engineering of Escherichia Coli for Production of Mixed-Acid Fermentation End Products. Front. Bioeng. Biotechnol. 2014, 2. [Google Scholar]
- Kawada-Matsuo, M.; Oogai, Y.; Komatsuzawa, H. Sugar Allocation to Metabolic Pathways Is Tightly Regulated and Affects the Virulence of Streptococcus Mutans. Genes 2016, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Xiao, Y.; Tashiro, Y.; Wang, Y.; Zendo, T.; Sakai, K.; Sonomoto, K. Fed-Batch Fermentation for Enhanced Lactic Acid Production from Glucose/Xylose Mixture without Carbon Catabolite Repression. J. Biosci. Bioeng. 2015, 119, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Wongputtisin, P.; Khanongnuch, C.; Kongbuntad, W.; Niamsup, P.; Lumyong, S.; Sarkar, P.K. Use of Bacillus Subtilis Isolates from Tua-Nao towards Nutritional Improvement of Soya Bean Hull for Monogastric Feed Application. Lett. Appl. Microbiol. 2014, 59, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Kunasundari, B.; Murugaiyah, V.; Kaur, G.; Maurer, F.H.J.; Sudesh, K. Revisiting the Single Cell Protein Application of Cupriavidus Necator H16 and Recovering Bioplastic Granules Simultaneously. PLoS ONE 2013, 8, e78528. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-G.; Chen, X.-L.; Wang, Z. Microbial Biomass Production from Rice Straw Hydrolysate in Airlift Bioreactors. J. Biotechnol. 2005, 118, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Naraian, R.; Kumari, S. Microbial Production of Organic Acids. In Microbial Functional Foods and Nutraceuticals; John Wiley & Sons, Ltd, 2017; pp. 93–121 ISBN 978-1-119-04896-1.
- Coban, H.B. Organic Acids as Antimicrobial Food Agents: Applications and Microbial Productions. Bioprocess Biosyst. Eng. 2020, 43, 569–591. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Lv, Y.; Qian, X.; Jiang, Y.; Wu, M.; Zhang, W.; Zhou, J.; Dong, W.; Xin, F.; Jiang, M. Current Advances in Organic Acid Production from Organic Wastes by Using Microbial Co-Cultivation Systems. Biofuels Bioprod. Biorefining 2020, 14, 481–492. [Google Scholar] [CrossRef]
- Phosriran, C.; Wong, N.; Jantama, K. An Efficient Production of Bio-Succinate in a Novel Metabolically Engineered Klebsiella Oxytoca by Rational Metabolic Engineering and Evolutionary Adaptation. Bioresour. Technol. 2024, 393, 130045. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Lin, C.-J.; Lee, W.-C.; Teng, H.-Y.; Chuang, M.-H. Production of Succinic Acid through the Fermentation of Actinobacillus Succinogenes on the Hydrolysate of Napier Grass. Biotechnol. Biofuels Bioprod. 2022, 15, 9. [Google Scholar] [CrossRef]
- Ammar, E.M.; Philippidis, G.P. Fermentative Production of Propionic Acid: Prospects and Limitations of Microorganisms and Substrates. Appl. Microbiol. Biotechnol. 2021, 105, 6199–6213. [Google Scholar] [CrossRef]
- Joshi, B.; Joshi, J.; Bhattarai, T.; Sreerama, L. Chapter 15 - Currently Used Microbes and Advantages of Using Genetically Modified Microbes for Ethanol Production. In Bioethanol Production from Food Crops; Ray, R.C., Ramachandran, S., Eds.; Academic Press, 2019; pp. 293–316 ISBN 978-0-12-813766-6.
- Jurtshuk, P. Bacterial Metabolism. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston (TX), 1996 ISBN 978-0-9631172-1-2.
Figure 1.
Growth profile of nitrogen-fixing K. oxytoca M5A1 in biomass hydrolysate.
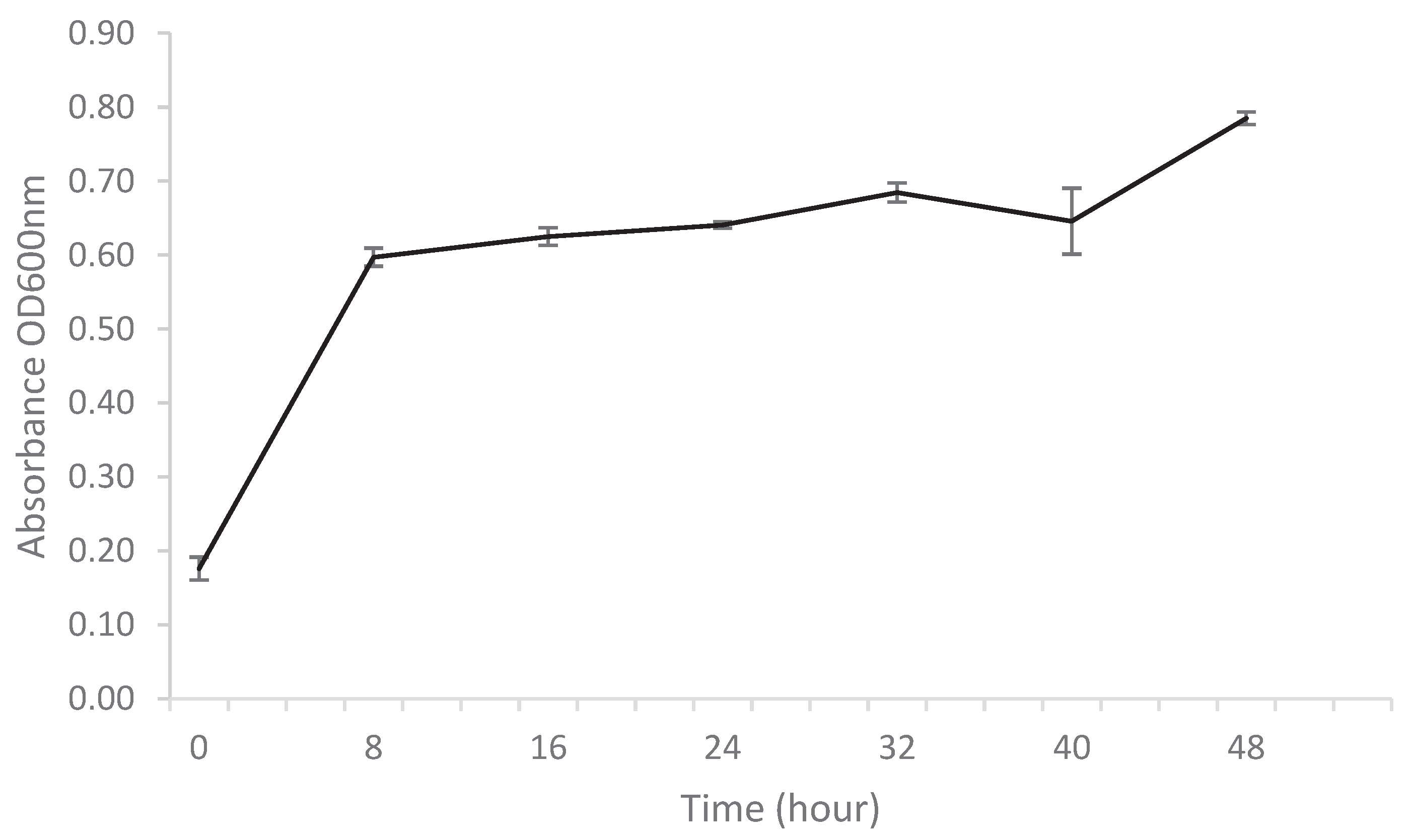
Figure 2.
Utilization rate of glucose during fermentation.
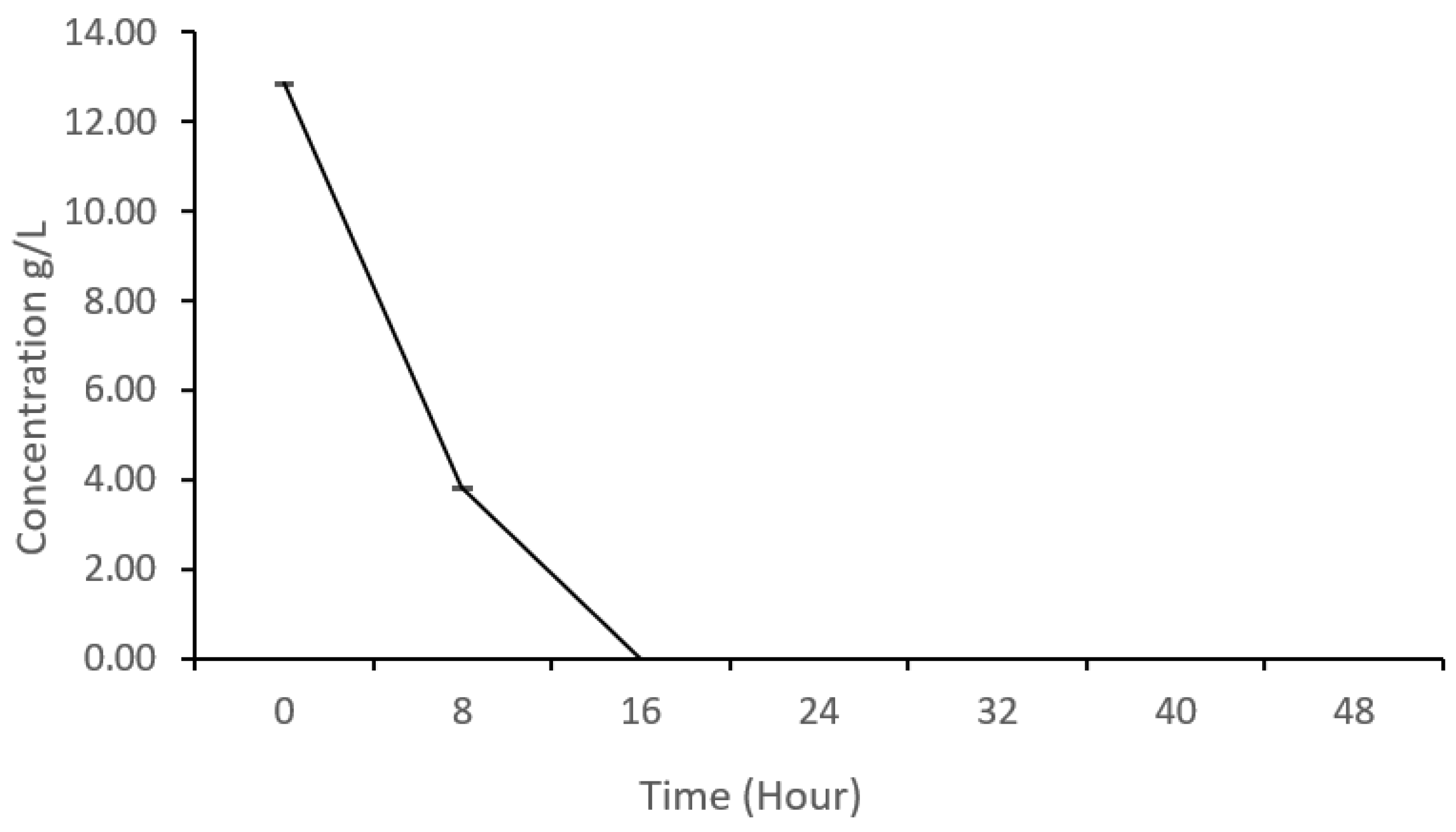
Figure 3.
Microbial protein concentration of K. oxytoca M5A1.
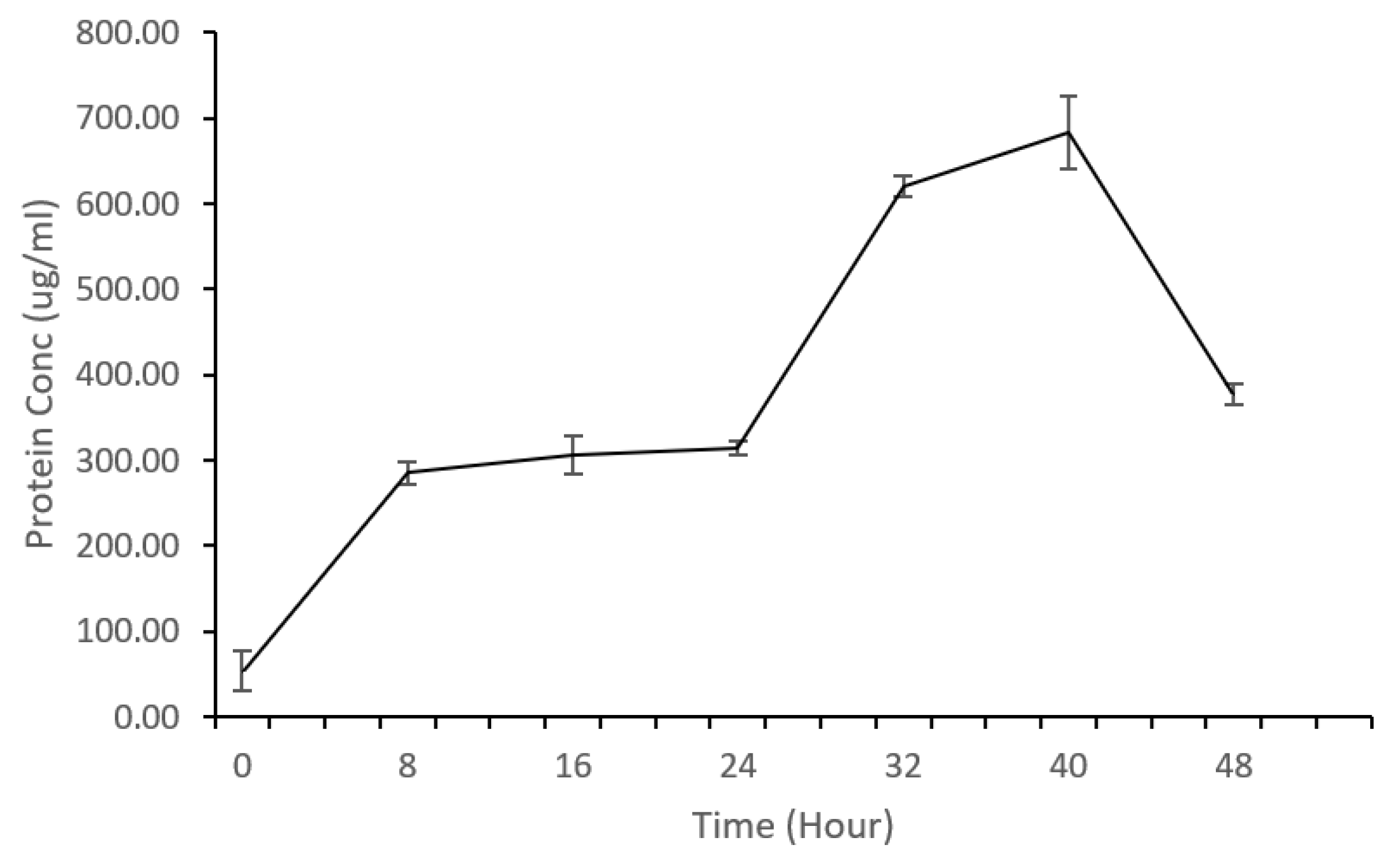
Figure 4.
Organic acids profile during fermentation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



