
Submitted:
18 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
In plants, melatonin (MT) and abscisic acid (ABA) are conventionally treated as molecules mitigating stress responses. To understand the mechanisms of ABA–MT interplay, we examined the effects of ABA and MT treatment in loss-of-function ABA and MT mutants exposed to high light (HL) stress. ABA constantly suppressed ASMT encoding N-acetylserotonin methyltransferase in the context of differential responses of other MT biosynthesis genes in both wild type (WT) and mutants. However, this response was absent in the mutant with the disrupted ABI4. Given that the ASMT promoter region contains several potential ABI4-binding elements, these data suggest that ASMT can be a potential target gene for ABI4. A role for ABI4 in the interactions between ABA and MT is supported by the finding that ABI4 is constitutively derepressed in the MT signaling mutants cand2 and gpa1, which exhibited elevated steady state levels of ABI4 transcripts and were not regulated by either stress or melatonin. In addition, the abi4 mutant showed increased modulations in the expression of the MT catabolic genes M2H and M3H in response to ABA treatment inferring that this transcription factor is a negative regulator of ABA-dependent changes in MT content. Furthermore, all tested mutants with impaired ABA synthesis or signaling displayed elevated steady state MT levels compared to WT, while MT treatment contributed to downregulation of key ABA synthesis and signaling genes. Collectively, our results suggest that ABA and melatonin act antagonistically modulating the expression of ABA and MT signaling and metabolism genes.
Keywords:
Arabidopsis thaliana
; abscisic acid
; gene expression
; high light stress
; melatonin
; mutants
1. Introduction
Melatonin is a multifunctional effector which plays diverse physiological roles in plants, from seed germination to post-harvest fruit storage and seed longevity [1]. However, the beneficial effects of this molecule as an anti-stress agent are of paramount importance due to its powerful antioxidant activity. Melatonin acts as a scavenger of reactive oxygen species (ROS) and reactive nitrogen species (RNS) such as O2−, OH−, NO and ONOO− and also enhances the expression of superoxide generating enzymes (RbOHs), superoxide dismutase and the enzymes involved in detoxification of excess H2O2, particularly CAT, POD, and APX [2]. At low concentrations, melatonin may act as a hormone-like signaling molecule involved in crosstalk with virtually all known plant hormones by modulating their signaling circuits and metabolism [3].
The Arabidopsis genome harbors several genes encoding four successive steps of melatonin biosynthesis. During the first two steps serotonin is derived from tryptophan by two enzymatic reactions carried out by TDC (tryptophan decarboxylase) and T5H (tryptamine 5-hydroxylase). The next step is committed SNAT (serotonin N-acyltransferase) responsible for the synthesis of N-acetylserotonin, which is further catalyzed into melatonin by O-methyltransferases COMT (caffeic acid O-methyltransferase) and ASMT (N-aceylserotonin O-methyltransferase) [4]. Two melatonin metabolic genes M2H (melatonin 2-hydroxylase) and M3H (melatonin 3-hydroxylase) convert melatonin into 2-hydroxymelatonin and cyclic 3-hydroxymelatonin respectively [5].
The signaling pathway of melatonin and its metabolites is proposed to act through the mitogen-activated protein kinase (MAPK) cascade, which is preceded by receptor-like kinases (RLKs), putative candidates for melatonin receptors in plants [6]. Alternative signaling circuit includes putative MT receptor gene CAND2 (also known as PMTR1), which is associated with GPA, encoding the α-subunit of the heterotrimeric G protein [7].
Abscisic acid is conventionally considered a stress hormone since its level is greatly increased under a variety of abiotic stresses [8]. ABA via binding to the PYR/PYL/RCAR (PYL) family of receptors, inhibits a class of A-type protein phosphatase 2Cs (PP2Cs). PP2C inactivation triggers phosphorylation of a collection of basic leucine zipper transcription factors (ABA-responsive element binding factors, ABFs) by a small set of class 3 sucrose nonfermenting-1-related protein kinase 2s (SNRK2s). Phosphorylated ABFs activate downstream target genes through the cis-acting ABA response elements (ABREs). In addition to the canonic PYL-PP2C-SnRK2 core pathway, ABA responses in plants require a network of other signaling pathways that involve calcium, ROS, NO, phospholipids, and the various kinases [9]. All these pathways can interact with each other through positive or negative feedback mechanisms producing a remarkable pleiotropy of effects in plant responses to abiotic stresses.
Melatonin-mediated action generally induces down-regulation of ABA biosynthesis genes and up-regulation of ABA catabolism genes resulting in a decrease of ABA level [2,3]. For example, in apple melatonin selectively down-regulated MdNCED3, an ABA synthesis gene, and triggered its catabolic genes, MdCYP707A1 and MdCYP707A2, reducing ABA contents in drought-stressed plants [10]. Under drought stress, exogeneous melatonin downregulated NCED1 expression in maize and induced the expression of ABA8ox1 (ABA-8- oxidase 1) and ABA8ox3 responsible for ABA breakdown [11]. Other examples include Chinese cabbage [12], Chinese hickory Carya cathayensis [13], mango [14], cucumber [15] and ryegrass [16]. A special mechanism by which melatonin antagonizes ABA action was recently described in Arabidopsis. Melatonin offsets ABA action to delay leaf senescence via RBOHD-dependent H2O2 production that triggers [Ca2+]cyt accumulation and subsequently inhibits K+ efflux and delays cell death/leaf senescence in response to ABA [17].
On the other hand, an increase in melatonin content can lead to up regulation of ABA-responsive genes testifying that crosstalk between MT and ABA is highly variable. Opposite responses were detected for Elymus nutans [18], Brassica napus [19] and Raphanus sativus [20]. Overexpression of AtASMT caused massive melatonin accumulation and synergized with ABA to inhibit seed germination in Arabidopsis [21]. In case of barley, exogenously applied melatonin resulted in higher ABA concentration in the drought-primed plants than in the nonprimed plants when exposed to cold stress [22]. Likewise, during arsenic stress, MT treatment of the susceptible rice cultivar Khitish up-regulated transcript levels of NCED3, and down-regulated transcript levels of ABA8ox1 elevating the endogenous ABA level. However, ABA concentration remained unaltered in tolerant rice cultivar Muktashri indicating that the response depends upon plant genotype [23].
While much of the work conducted on melatonin-ABA interplay in plants has been targeted on influence of melatonin on ABA content and signal transduction, few studies have focused on the intersecting effects of ABA and melatonin on the expression of signaling and metabolic genes of these regulators. In rice, melatonin was shown to regulate ROS homeostasis under nitrogen limitation via the module OsbZIP79-OsABI5, composed of two ABA related trans-factors. However, the module did not directly regulate melatonin biosynthesis [24]. On the other hand, treatments with abscisic and methyl jasmonic acids simultaneously induced three isoforms of N-acetylserotonin methyltransferase in rice, the final enzyme in melatonin biosynthetic pathway [25]
In this work, using exogenously applied ABA and melatonin as well as melatonin and ABA-related mutants, we showed that ABA is involved in downregulation of Arabidopsis ASMT gene under high light stress via the activity of the transcription factor ABI4 (ABSCISIC ACID INSENSITIVE 4). Additionally, we identified that MT-dependent regulation of ABA genes depends on CAND2 - GPA1 signaling circuit.
2. Results
2.1. ABA Treatment Differentially Regulates Physiological Parameters and Stress-Induced Genes in Col0 and MT Mutants
ABA treatment under mild light (60 μmol m-2 s-1, 24 h) was perceived by wild type plants as a mild stress, as evidenced by the reduced transcript accumulation of PsbS involved in quenching singlet excited chlorophylls and increased expression of stress marker gene AOX1a which participates in the fine-tuning of mitochondrial membrane potential and the alleviation of ROS production (Figure 1). Under HL stress (600 µmol m-2 s-1, 24 h), ABA additionally increased the accumulation of AOX1a transcripts, and also reduced stress-induced levels of ELIP2 transcripts (EARLY LIGHT-INDUCED PROTEIN 2), which is known to modulate chlorophyll synthesis to prevent photo-oxidative stress [26]. MT mutants, when treated with ABA, did not differ significantly from the wild type in their responses under either control or stress conditions.
In parallel, the content of TBARs, a marker of membrane damage caused by lipid peroxidation, was significantly lifted by ABA treatment of plants under mild light, whereas it was decreased under combined application of HL and ABA compared to stress-induced levels. Hence, ABA treatment of control plants had a positive effect only under stress conditions. Exogenous ABA did not significantly change chlorophyll content in WT and asmt mutant under mild light, but promoted increased chlorophyll levels under excessive light compared to stress-repressed levels. On the other hand, MT signaling mutants, cand2 and gpa1, exhibited reduced chlorophyll levels under mild light when treated with ABA which indicates their elevated sensitivity to the hormone. However, as with the wild type, the chlorophyll content of these mutants increased after ABA treatment compared to their HL control values confirming plasticity of plant responses to ABA.
2.2. Responses of MT-Related Genes to ABA Treatment of MT Mutants
To elucidate the regulatory interactions between ABA and melatonin, we next examined the responses of MT related genes to ABA treatment in WT and MT mutants (Figure 2). Under mild light, exogenous ABA drastically down-regulated ASMT (4 fold), and to a lesser extent SNAT1 both in WT and MT mutants and did not change transcript accumulation of COMT and T5H. TDK expression varied: it was not significantly affected in WT and gpa1, but was reduced in asmt and slightly increased in cand2. It should be noted, however, that the steady state levels of TDC transcripts in untreated MT mutants were 3 to 4 fold higher than in WT.
Under excessive irradiation, ABA maintained the expression of SNAT1 and COMT at a higher level as compared to untreated stressed plants, probably due to the stress mitigating effect of ABA. T5H was strongly inhibited in WT and all MT mutants except asmt, despite elevated steady state transcript level in this mutant. TDC transcript levels were decreased in WT and gpa1 and did not substantially differ from the control levels in asmt and cand2. However, ASMT expression was still downregulated by the hormone in WT and MT mutants under either control or stress conditions. These results indicate that ABA constantly suppresses ASMT in the context of differential responses of other MT biosynthesis genes.
The interactions between ABA and melatonin may also be mediated by the expression of MT catabolism genes, given that MT metabolites may have specific functions in plants. Melatonin is converted to 2-hydroxymelatonin (2-OM) and cyclic 3-hydroxymelatonin (3-OM) by members of the 2-oxoglutarate-dependent enzyme family M2H and M3H [27]. The expression of M2H, encoding melatonin 2-hydroxylase (M2H), was slightly reduced in WT and to a lesser extent in MT mutants upon exposure to HL or ABA and upon combined treatment with ABA and HL. In contrast, a second gene of plant melatonin metabolism, encoding melatonin 3-hydroxylase (M3H), was upregulated 6- to 10-fold under HL. Moreover, its transcripts increased 20- to 40-fold in WT and MT mutants after ABA treatment under mild and high irradiation. These data support the idea that ABA may antagonistically regulate MT levels modulating the expression of MT catabolism genes.
We also tested whether the putative MT signaling genes CAND2 and GPA1, responded to ABA treatment. Exogenous ABA did not affect CAND2 and GPA1 expression under control radiation and contributed to their slightly reduced levels in high light conditions.
2.3. ABA Mutations Affect Melatonin Content and the Expression of MT Related Genes
To further delineate possible links between melatonin and ABA, we assessed the effect of modulated endogenous ABA status on the responses of MT biosynthesis and signaling genes to stress and MT treatment The study included ABA biosynthesis mutants aba2 and aba3 with the lowered level of endogenous ABA [28], and mutants with impaired ABI3, ABI4 and ABI5 genes encoding three well-characterized transcription factors. These regulators of ABA signaling are members of the B3-, APETALA2- (AP2), and basic leucine zipper- (bZIP) domain families, respectively, which have been shown to regulate overlapping subsets of ABA-inducible genes [29].
Considering that the effect of MT treatment may depend on the level of endogenous MT, we first assessed melatonin content in ABA mutants under control conditions and under photooxidative stress (Figure 3). All the mutants displayed elevated steady state MT levels (2.5 fold) as compared to WT, especially in abi5, consistent with the increased steady state levels of ASMT transcripts in this mutant. Following HL exposure, melatonin content decreased in the mutants with altered ABA status.
The expression of MT synthesis genes SNAT and COMT was downregulated under HL in the mutants, though less than in WT or did not change (Figure 4). However, in contrast to WT, ASMT transcript levels increased, especially in aba3 and aba2 (5 and 6 fold respectively) and remained elevated after combined-treatment with HL and melatonin. Thus, defects of ABA synthesis or signaling contributed to the induction of ASMT. These results imply that ASMT expression negatively correlated with ABA content and signaling. T5H and TDC were also activated by stress and stress+MT, in ABA mutants, although in WT stress-induced elevation of both genes was mitigated by MT.
HL mildly reduced the expression of CAND2, and did not reliably affect GPA1 in ABA mutants, in contrast to WT in which both genes were dramatically downregulated (3-4 fold). Under combined MT+HL treatment no further changes of CAND2 and GPA1 expression occurred in tested ABA mutants while in WT plants the transcript levels of both genes increased as compared to those in untreated stressed plants. Hence, the expression patterns of CAND2 and GPA1 under stress conditions depend on ABA status with ABA deficiency promoting increased expression of MT signaling genes. The results also suggest possible involvement of additional factors in ABA dependent regulation of GPA1 expression since the expression pattern of GPA1 in ABA mutants did not strictly follow that of CAND2.
Given that melatonin signaling pathway may act through the mitogen-activated protein kinase (MAPK) cascade, [6] we also analyzed expression profiles of MAPK6. However, no statistically significant changes occurred following HL or HL+MT exposure in either WT or ABA mutants
2.4. MT Treatment Alters the Expression of ABA Related Genes in MT Mutants
Next, we studied whether melatonin regulated the expression of ABA related genes in MT mutants. In wild type plants, key ABA synthesis and signaling genes NCED3, ABA3, ABI1, ABI2, ABI4, ABI5 were upregulated and ABI3 was downregulated by excess irradiation, whereas melatonin attenuated these responses (Figure 5). Such alleviation may indicate a more stable status of the plant exposed to melatonin, when stress-induced response of ABA genes is less pronounced. At the same time, this reaction may be the result of competitive relations between these regulators for a common set of transcription targets under stress, leading to partial repression of ABA genes by melatonin. Of note, NCED4, which plays a major role in degradation of beta-carotene, a substrate for ABA synthesis [30] was strongly downregulated by HL stress in accord with stress protective role of ABA and to a lesser extent following MT+HL treatment alleviating the effects of stress.
The catabolism of ABA from ABA to phaseic acid is catalyzed by a cytochrome P450 monooxygenase (P450) encoded by CYP707As [31]. ABA can also be glucosylated by a UDP-glucosyltransferase encoded by UGT71C5 to ABA-glucose ester (ABA-GE), which is an inactive form of ABA [32]. ABA-GE, in its turn, can rapidly be transformed to active ABA via AtBG1 and AtBG2 encoded β-glucosidases when the environment conditions change. MT treatment under HL did not substantially affect CYP707A transcript levels compared to their increased values under HL. UGT71C5 expression was significantly downregulated while that of AtBG1 dramatically upregulated under HL which may contribute to increased ABA levels, and these alterations were attenuated by MT. Therefore, melatonin treatment primarily affects genes involved in the metabolism of reversible forms of ABA.
In the melatonin-deficient asmt mutant, the response of ABA genes to stress and melatonin was similar to that of the wild-type, although less pronounced for some genes. However, in cand2 and gpa1 mutants ABI4 was not regulated by either stress or melatonin Moreover, MT insensitive cand2 and gpa1 exhibited elevated steady state levels of ABI4 transcripts (3- and 4-fold, respectively, as compared to WT) suggesting that ABI4 is constitutively derepressed in the cand2 and gpa1 background and further response to stress treatment is not possible. Steady state levels of ABI4 transcripts in MT deficient asmt were also significantly increased (1.5-fold). These results imply that the transcription factor ABI4 may play a role in the interactions between ABA and melatonin.
2.5. ASMT as a Potential Target Gene for ABI4 under HL Stress
Finally, we tested whether MT metabolism and signaling genes were transcriptionally altered in ABA mutants (Figure 4). Steady state levels of TDC and T5H transcripts were significantly elevated and that of COMT, SNAT and GPA1 reduced in aba2 and aba3 mutants relative to WT. ASMT and TDC transcript abundance was increased in abi5.and abi3 while the expression of CAND2 was not significantly altered.
Under ABA treatment or ABA treatment coupled with HL, the expression of SNAT and COMT slightly decreased in WT and ABA mutants as compared to untreated samples. ABA+HL application also down regulated TDC and T5H in WT, aba2, aba3 and abi3 as compared to their stress induced levels, but had no impact in abi4 and abi5. These data suggest that ABA may act through ABI4 and ABI5 transcription factors in stress dependent regulation of TDC and T5H. It should be noted, however, that TDC and T5H have also been identified as participants in IAA biosynthesis. Therefore, ABA-dependent expression of these genes may be related to ABA/auxin interactions, given that the expression of downstream SNAT and COMT in the melatonin biosynthetic pathway followed that of WT in all ABA mutants tested upon ABA+HL.
ABA also negatively regulated ASMT transcript accumulation, under both mild and high light in WT plants and in aba2, aba3, abi3, and abi5. However, ASMT did not respond to ABA treatment in the abi4 mutant. These results suggest that ASMT can be a potential target gene for ABI4 thus supporting the notion that the transcription factor ABI4 is a key player in the interactions between ABA and melatonin.
`Furthermore, the ABI4-disrupted mutant showed a 2-3-fold decrease in M2H transcript accumulation following ABA treatment and a 2-fold increase in transcript levels compared to the dramatically ABA-induced M3H expression in WT and other ABA mutants. These results confirm that ABI4 is an important factor in the regulation of the interactions between melatonin and ABA, However, the exact biological meaning of ABA-dependent shifts in the expression of melatonin catabolism genes remains to be clarified, taking into account that, on the one hand, 2-OHM triggers the production of ROS [33], and on the other hand, that 3-OHM is a powerful antioxidant exhibiting even higher antioxidant activity, than melatonin [5].
2.6. In Silico Promotor Analysis of ASMT Gene
A role for the ABI4 transcription factor in the regulation of MT synthesis implies that promoter regions of MT related genes contain cis-regulatory elements that are potential binding sites for ABI4. ABI4 was shown to bind a number of DNA sequences including Coupling Element 1 (CE1) CACCG [34], two ABE motifs GC(C/G)GCTT(T) [35], B ELEMENT CGTGAT [36] and S-box and S-box similar sequence CACYKSCA [37]. A motif CCAC has also been suggested to bind ABI4 particularly when adjacent to, or overlapping with, a G box motif [38]. In addition, DRE and ABRE core sequences have the potential to be bound by ABI4 [35].
Detailed in silico analysis of ASMT promoter region within 1000 bp from ATG start codon (basal part), showed one potential ABI4-binding element CE1 (nt 467 to 472) (Figure 6). The binding affinity of CE1 (CACCG) is considered higher than that observed with other DNA-binding sites (Finkelstein et al., 2011). We also identified one ABRE-like element (nt 742 to749) within basal part. Five more elements, one CE1 (nt 1953 to 1958) and four CCAC motifs (nt 1005 to 1010; 1261 to 1266; 1954 to 1959; and 2113 to 2118) were found in distal region of the ASMT promoter as its maximum length can reach 2,483 bp according to the AGRIS database (https://agris knowledgebase.org/AtcisDB/getpromseq.html?id=At4g35160). We therefore hypothesize that ASMT could be a direct transcriptional target of ABI4.
It should be noted that sequence recognition by ABI4 may be more flexible than known canonical sequences. According to the results of [29] many of the direct ABI4 transcriptional targets do not contain the previously characterized ABI4 binding motifs. Nevertheless, some of them were able to interact with ABI4 in electrophoretic mobility shift assays. Moreover, ABI4-regulated promoters could be synergistically induced by specific combinations of ABI4 and some bZIP factors, even when no canonical ABI4 binding sites were present. Given a relatively small number of direct transcriptional targets for ABI4 within proximal part, we can also hypothesize that ASMT promoter can be regulated indirectly via cis-regulatory elements recognized by ABI4-inducible transcription factors.
Functional characterization of the ASMT promoter was carried out by testing the ability to drive expression of the reporter β-glucuronidase (GUS) gene. To this end three 5′-deletion fragments (−1000 bp, −1847 bp, and −2483 bp) upstream of the ASMT translation start codon were fused to the open reading frame of the GUS reporter gene. ABA treatment (50 µM ABA for 6 h) of ASMT pro::GUS fusions in the wild type background resulted in a considerable decrease in accumulation of ASMT pro::GUS transcripts as compared to untreated leaf discs (Figure S1). The signal also decreased in plants containing1000 bp ASMT promoter fragment in abi4 background. At the same time, plants containing the 1847 bp and 2483 bp fragments in abi4 background did not differ from untreated plants when exposed to ABA suggesting that ABI4 was required for ABA induced decrease in ASMT promoter activity. Given that distal region of the ASMT promoter includes one CE1 and four CCAC motifs it was concluded that ABI4 can directly play role in the ability of the ASMT promoter to respond to ABA treatment.
3. Discussion
Overall, we found that melatonin and ABA are able to mutually influence the expression of the genes involved in their metabolism and signaling. In particular, the ASMT which encodes enzyme of the final step of melatonin biosynthesis was suppressed by melatonin treatment under both mild and HL in the wild type. However, the expression remained virtually unchanged in the mutants with the disrupted genes for ABA synthesis and signaling under mild light and was even upregulated under combined MT+HL treatment. Therefore, deficiencies in ABA content or perception contribute to increased expression of ASMT gene, which has been proposed to be stress-regulated [39].
In parallel, the ASMT was downregulated by ABA treatment in the wild type plants and to a lesser extent in aba2, aba3, abi3 and abi5 mutants with the compromised ABA status. But the ASMT did not respond to ABA in abi4 with inactivated gene for transcription factor ABI4. We therefore hypothesize that application of ABA induces repression in ASMT promoter activity in WT via interaction with the ABI4 transcriptional factor, and this repression is absent in the abi4 mutant. These results are consistent with the notion that ASMT is a potential target gene for ABI4 which is probably involved in melatonin biosynthesis as a negative regulator.
It should be noted that ABI4 has been suggested to act as both a transcription activator and a repressor, which is determined by its interaction with other proteins, protein modification or binding to discrete DNA binding sites leading to either activation or repression of target genes [35]. ABI4 binds to a number of GC-enriched DNA sequences providing regulation of target genes involved in a large functional spectrum of activities. At least 2 Coupling Element1 (CE1) sequences (CACCG) are present in ASMT promotor region and one ABRE-like element (TACGTGTA), in addition to 7 CCAC motifs which also correlated with ABI4 signaling [38].
Furthermore, in this study, we found that ASMT pro::GUS fusions did not respond to ABA treatment in abi4 mutant background, although ABA decreased ASMT promoter activity in WT background. These results imply that a functional ABI4 is necessary for ABA dependent regulation of ASMT expression. Consistent with the fact that ABI4 is involved in ABA-melatonin crosstalk, MT-insensitive cand2 and gpa1 exhibited increased steady state levels of ABI4 transcripts, suggesting that ABI4 is constitutively derepressed in genotypes defective in MT signaling.
Although MT may act directly as an anti-stress agent, the complexity of MT-dependent gene expression suggests that MT mediates many important functions through a melatonin receptor(s) and downstream signal transduction pathways in plants. To date, studies of the melatonin- CAND2-GPA1-signaling pathway have documented its involvement in osmotic, salt, drought and HL stress tolerance, in addition to biotic stresses. Here we have showed that an intricate network of ABA related genes under HL, at least in part, depends on CAND2. This is particularly relevant for ABA signaling, since, in contrast to WT, the expression levels of ABA signaling genes in cand2 and gpa1 mutants did not respond significantly to MT treatment.
The identification of molecular links between MT and ABA synthesis and signaling provides a basis for understanding how disruptions of these two regulators can feed back to influence each other. According to results of this study, inactivating components of MT signaling circuit results in compensatory effects in ABA signaling. On the other hand, deficiencies in ABA synthesis and signaling entail increased expression of MT synthesis genes and elevated steady state MT levels. The existence of such compensatory mechanisms gives rise to a certain parallelism in the action of these two regulatory molecules and can have a broad impact beyond the immediate activity of the inactivated components.
In this context, ABA-dependent modulations in the expression of melatonin catabolism are of special interest. ABA treatment promoted downregulation of M2M, which may result in decreased levels of 2-OM with its ROS-inducing mode of action and consequently decreased ROS production. In parallel, M3H upregulation, particularly in the abi4 mutant, may trigger 3-OM overproduction and enhance antioxidant activity. Therefore, the ABA-induced decrease in MT content, which can be regarded as a manifestation of the antagonistic relationship between ABA and MT, may actually lead to an increase in antioxidant capacity and stress resistance
The functional redundancy in the effects of MT and ABA allows the plant to trigger alternative pathways in response to environmental challenges or developmental demands. At the same time, they provoke antagonism between ABA and MT under both control and stress conditions. As we have already outlined, examples of antagonistic relations between MT and ABA were shown for a number of plant species [11,12,13,14,15,16,17]. On the other hand, synergism in MT and ABA effects was also demonstrated in several studies [18,19,20,21,22]. Moreover, dual role of melatonin as a signaling molecule in ABA-melatonin interplay was revealed during seed formation and seed germination of Arabidopsis. Knock-out mutants for melatonin receptor PMTR1 contained higher ABA concentrations in developing seeds, but accumulated lower ABA levels in dry and imbibed seeds than the wild-type Col-0 [40]. Hence, multiplicity of possible functions defined by ABA-melatonin crosstalk imply multivariant responses depending on physiological state and genetic background.
The interactions between melatonin and ABA are likely to be integrated with other hormone signals. Mutual impact of ABA, gibberellic acid (GA) and MT has been shown for cucumber. Exogenous melatonin increased the expression of ABA catabolism genes (monooxygenase, CYP707A1 and CYP707A2) and GA biosynthesis genes (GA20ox and GA3ox), and also suppressed the ABA biosynthesis gene 9-cis-epoxycarotenoid dioxygenase (NCED2). This led to a decrease in ABA levels and an increase in GA3 and GA4 content, and accelerated seed germination [15]. A similar picture was observed during the germination of cotton seeds (Gossypium hirsutum L.) under salt stress conditions [41].
In ryegrass (Lolium perenne L.), exogenous melatonin reduced the ABA content by suppressing the expression of biosynthesis (LpZEP and LpNCED1), downregulated ABA signaling genes (LpABI3 and LpABI5), but activated cytokinin (CK) synthesis genes (LpIPT2 and LpOG1), thereby increasing the content of endogenous CK and activating CK signaling circuits [42]. In our prior research we showed that melatonin may act synergistically with CK in Arabidopsis via melatonin-mediated activation or repression of the CK synthesis and signaling genes [16]. Conversely, CK was involved in expression regulation of the genes for melatonin metabolism and signal transduction, with the melatonin biosynthesis gene ASMT being a key component in the crosstalk between the CK and MT metabolic pathways. In this study we revealed that ASMT is a potential target for ABI4 in ABA-melatonin intersection. Therefore, fluctuation in melatonin activity induced by various hormones may, at least in part, be mediated by ASMT. These data are consistent with RNA sequencing and qRT-PCR data obtained by [43], which showed that in the walnut, Juglans regia, members of the ASMT gene family participate in regulatory networks of phytohormones such as ABA, salicylic acid and CK, regulating flower development and stress tolerance. In addition, in silico analysis of the promoter regions revealed 11 hormone-response motifs including the ABA response element ABRE, auxin response element AuxRR-core, jasmonic acid (JA) response elements CGTCA and TGACG gibberellin response element P-box and salicylic acid response element TCA.
Hormone-like behavior of MT while interacting with classical hormones, as well as participation of PMTR1/CAND2 receptor in such reactions, makes it possible to classify melatonin as a new phytohormone especially when it acts in physiologically effective concentrations using high-affinity binding sites. At the same time, MT is believed to emerge as the first-line antioxidant capable of operating in cardinally higher concentrations [44,45]. The evolution in modes of action and the acquisition of secondary hormone-like properties have required the incorporation of melatonin into highly interconnected web involved in hormone metabolism and signaling [46]. This contributed to the generation of joint pathways that ensured the inclusion of melatonin in the plant hormone system. The interplay between ABA and melatonin, resulting from a considerable overlap in effects, is an example of such integration, with MT-dependent regulation of ABA genes via CAND2/GPA1 signaling circuit and ABI4/ASMT acting as elements of this bond.
4. Material and Methods
4.1. Plant Materials and Stress Treatments
Arabidopsis thaliana WT (ecotype Columbia) and mutant lines aba2-1 (NASC 156), aba3-1(NASC 157), abi3 (NASC 503216, abi4 (NASC580095 and abi5 (NASC513163), asmt (NASC680911), cand2 (NASC678658), and gpa1 (NASC6534) were used in this study. Seeds were surface-sterilized and stratified for 48 h at 4°C in the dark. Plants were grown on half strength Murashige and Skoog (MS) medium with 1% sucrose and 0.5% agar in a growth chamber at 23°C with a 16-h photoperiod and photosynthetic photon flux density of 60 μmol m-2 s-1. Two-week-old plants were pretreated with 50 μM of melatonin for 72 h or 50 μM of ABA for 24 h. Prolonged pre-treatment was chosen because of the amphiphilic nature of melatonin, which implies relatively low penetrating ability. For HL treatment, plants were exposed for 24 h under the HPI-T2 2000 W/646 lamp (Philips, Netherlands) with a luminous energy flux of 600 μmol m-2 s-1. A combination of air and water cooling was used to minimize the thermal impact of the lamp. Control plants were left under growing conditions. At the end of the exposure plants were immediately frozen in liquid nitrogen and subjected to further analysis
4.2. RNA Extraction and qRT-PCR
Total RNA was extracted from frozen samples utilizing the TRIzol (Thermo Fisher Scientific) method. cDNAs synthesis was performed from 2 μg of total RNA using a mixture of oligo (dT) primers and random hexamers Relative gene expression levels were quantified via qRT-PCR using LightCycler 96 (Roche, Switzerland) with the hot start SYBR Green I technology. Standard thermal profile for all PCRs included following steps:95°C for 5 min, 40 cycles of 95°C for 15 s, 58°C for 15 s and 72°C for 25 s. Nuclear-encoded polyubiquitin UBQ10 gene, was used as an internal control. Primers used for qRT-PCR are presented in Table S1.
4.3. Melatonin Measurement
Determination of endogenous melatonin concentration was performed as described by Lee and Back [47] utilizing ELISA Kit CEA908GE (Cloud-Clone Corp., USA) according to the manufacturer's protocol. The optical density was measured at 450 nm with a Multiskan MS Microplate Reader LabSystems 352 (Thermo/LabSystems, USA).
4.4. Quantifications of Chlorophyll Content
Pigments were extracted from plant rosette leaves using 80% (v/v) acetone. The concentrations of chlorophyll (A+B) and carotenoids were determined by examining the absorbance of 440, 649 and 665 nm in the centrifugated supernatant as described by Lichtenthaler [48].
4.5. Determination of TBARs
Secondary products of membrane lipid peroxidation (TBARs) were determined by the reaction with thiobarbituric acid, as described by Heath and Packer [49]. The measurement was performed on a spectrophotometer (Pharmacia Biotech ultrospec 2000, UK) at wavelengths of 532 and 600 nm. The concentration of TBARs (μmol/g of fresh weight) was calculated using the formula: C = D/EL, where C is the concentration of MDA, μmol, D is the optical density, E is the molar extinction coefficient equal to 1.56 ×105 cm-1 M-1. The amount of TBARs was calculated in μmol/g of wet weight.
4.6. In Silico Promoter Analysis
Genomic sequence of ASMT gene upstream of ATG codon was obtained from arabidopsis.org database. ASMT promoter and cis-acting elements were predicted using PlantProm DB [50] database and the Arabidopsis Gene Regulatory Information Server (AGRIS) [51] databases, respectively. Nucleotide sequences analysis and localization of regulatory elements were performed with Vector NTI Advance 9.0 (Invitrogen) and visualized in Figure 6.
4.7. Construction of the ASMT pro::GUS Reporter Gene Fusions and Generation of ASMT pro::GUS Transgenic Plants
Generation of ASMT pro::GUS transgenic plants were performed as we previously described [52]. In brief, three fragments (1000 bp,1847 bp, and 2483 bp) upstream of the translation start codon of ASMT were amplified from the genomic DNA via polymerase chain reaction using different forward (P1000: 5’-aatggatccAGCTAGTTGTGGATCTGTAA-3’; P1847: 5’- aatggatccATCAAGGTCAAGTGATCAGAT-3’; P2483: 5’- atggatccCTATCTTTGACTTCTATTTGCT-3’) and general reverse (5’-aatctgcagGGTGAATTAGAGTTGAGGAAT-3’) primers carrying BamHI and PstI restriction sites, respectively. The resulting BamHI/PstI fragments were cloned upstream of the promoterless uidA reporter gene (GUS) of the binary vector pCambia-1381Z (Clontech, USA), producing pCambia ASMT pro::GUS constructs. The constructs obtained were introduced into Agrobacterium tumefaciens strain GV3101 (C58) (GoldBio, St Louis, MO, USA) using the freeze‒thaw method. Five-weeks-old wild-type Arabidopsis plants in the Col-0 and abi4 background were transfected with A. tumefaciens cells harboring P1000, P1847, and P2483 plasmids by the floral dip method. T1 transgenic seeds from each transformant plant were tested for germination on half MS medium containing 0.8% (w/v) phyto agar (Duchefa Biochemie) and 30 mg/L hygromycin (Hyg). The Hyg-resistant seedlings were then grown in soil for further analysis of ASMT pro::GUS expression.
4.8. Statistical Data Processing
All the experiments were repeated at least three times with similar results. Statistical analyses of gene expression data were performed with ANOVA with post hoc Holm multiple-comparison calculation using the online calculator (astatsa.com/ OneWay_Anova_with_TukeyHSD/). All data are presented as the mean values ± standard errors (SEs).
5. Conclusions
A plant’s ability to cope with stresses is highly correlated with two regulatory molecules melatonin and ABA which exhibit parallelism in their action. The overlapping effects of ABA and melatonin on the expression of their signaling and metabolic genes may provide a basis for understanding mechanisms of their interaction in plant responses to adverse environmental conditions. Here we have showed that compromised MT signaling results in compensatory effects in ABA signaling, whereas deficiencies in ABA synthesis and signaling cause increased expression of MT synthesis genes and elevated steady state MT levels implying that ABA and melatonin act antagonistically. On the other hand, ABA-dependent regulation of melatonin catabolism genes M3H and M2H promoting a decrease of MT levels may, in turn, modulate the content of MT metabolites and, as a result, augment plant antioxidant activity. This indirect effect highlights multifaceted output of ABA-MT interplay.
Changes in the expression of M2H and M3H were especially pronounced in abi4 inferring that this transcription factor is a negative regulator of ABA dependent changes in MT content. Furthermore, ABA induced downregulation of MT synthesis gene ASMT. However, the repression was absent in the abi4 mutant suggesting that ASMT can be a potential target gene for ABI4. An intricate network of ABA and MT related genes under HL, at least in part, is dependent on MT receptor CAND2. ABI4 was not regulated by either stress or melatonin in cand2 and gpa1 which had elevated steady state ABI4 transcript levels, specifying that ABI4 is constitutively derepressed. Overall, our results suggest that the functional redundancy of the effects of ABA and MT may contribute to their antagonistic relationship, superimposed on their exclusive transcriptional programs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, N. K. and V.K.; Funding acquisition, V. K. and N. K.; Investigation, I. B., E.S.P. and N. K.; Project administration, V. K.; Writing—original. draft, N. K.; Writing—review and editing, N. K., V. K., I. B. and E.S.P. All authors have read and agreed to the published version of the manuscript.
Funding
The work was carried out at the expense of the Russian Science Foundation (grant no. 23-14-00011). The data shown in Figure 2 were obtained with the support of the Ministry of Science and Higher Education of Russian Federation (theme No. 122042700044-6).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, T.; Wang, J; Sun, Y.; Zhang, L; Zheng, S. Versatile Roles of Melatonin in Growth and Stress Tolerance in Plants J. Plant Growth Regul. 2022, 41, 507–523.
- Arnao, M.B.; Hernandez-Ruiz, J. Melatonin as a regulatory hub of plant hormone levels and action in stress situations. Plant Biol. 2021, 23, 7-19. [CrossRef]
- Samanta, S.; Roychoudhury, A. Crosstalk of melatonin with major phytohormones and growth regulators in mediating abiotic stress tolerance in plants. South Afric. J. Bot. 2023, 163, 201-216. [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. The Antioxidant Cyclic 3-Hydroxymelatonin Promotes the Growth and Flowering of Arabidopsis thaliana. Antioxidants 2022, 11, 1157. [CrossRef]
- Back, K. Melatonin metabolism, signaling and possible roles in plants. Plant J. 2021, 105, 376–391. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA Perception and Signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tan, D.-X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J; Wang, Y.; Yang, Q. Melatonin Improves Drought Resistance in Maize Seedlings by Enhancing the Antioxidant System and Regulating Abscisic Acid Metabolism to Maintain Stomatal Opening Under PEG-Induced Drought. J. Plant Biol. 2021, 64, 299–312. [CrossRef]
- Tan, X.-L.; Fan, Z.-Q.; Kuang, J.-F.; Lu, W.-J.; Reiter, R.J.; Lakshmanan, P.; Su, X.-G.; Zhou, J.; Chen, J.Y.; Shan, W. Melatonin delays leaf senescence of Chinese flowering cabbage by suppressing ABFs-mediated abscisic acid biosynthesis and chlorophyll degradation. J. Pineal Res. 2019, 67, e12570. [CrossRef]
- Sharma, A.; Wang, J., Xu, D.; Tao, S.; Chong, S.; Yan, D.; Li, Z.; Yuan, H.; Zheng, B. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci Total Environ. 2020, 15, 713:136675. [CrossRef]
- Liu, S.; Huang, H.; Huber, D.J.; Pan, Y.; Shi, X.; Zhang, Z. Delay of ripening and softening in "Guifei" mango fruit by postharvest application of melatonin. Postharvest Biol. & Technol. 2020, 163, 111136.
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; Guo, Y.D. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA- interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [CrossRef]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat induced leaf senescence involves changes in abscisic acid and cytokinin biosynthesis and signaling pathways in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36-45. [CrossRef]
- Guo, Y; Zhu, J; Liu, J; Xue, Y; Chang, J; Zhang, Y; Ahammed, G.J.; Wei, C.; Ma, J.; Li, P.; Zhang, X,; Li, H. Melatonin delays ABA-induced leaf senescence via H2 O2 -dependent calcium signalling. Plant Cell Environ. 2023, 46, 171-184. [CrossRef]
- Fu, J; Wu, Y; Miao, Y; Xu. Y.; Zhao, E.; Wang, J.; Sun, H.; Liu, Q.; Xue, Y.; Xu, Y.; Hu, T. Improved cold tolerance in Elymus nutans by exogenous application of melatonin may involve ABA-dependent and ABA-independent pathways. Sci. Rep. 2017, 7. 39865. [CrossRef]
- Tan, X.; Long, W.; Zeng, L.; Ding, X.; Cheng, Y.; Zhang, X.; Zou, X. Melatonin-induced transcriptome variation of rapeseed seedlings under salt stress. Int. J. Mol. Sci. 2019, 20, 5355. [Google Scholar] [CrossRef]
- Jia, C.; Yu, X.; Zhang, M.; Liu, Z.; Zou, P.; Ma, J.; Xu, Y. (2020) Application of melatonin-enhanced tolerance to high-temperature stress in cherry radish (Raphanus sativus L. var. radculus pers). J. Plant Growth Regul. 2020, 39, 631-640. [CrossRef]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.J.; Li, X.; Mou, Z.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin inhibits seed germination by crosstalk with abscisic acid, gibberellin, and auxin in Arabidopsis. J. Pineal Res. 2021, 70, e12736. [Google Scholar] [CrossRef]
- Li, X.; Tan, D.X.; Jiang, D.; Liu, F. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef]
- Samanta, S.; Banerjee, A.; Roychoudhury, A. Exogenous melatonin regulates endogenous phytohormone homeostasis and thiol-mediated detoxification in two indica rice cultivars under arsenic stress. Plant Cell Rep. 2021, 40, 1585–1602. [Google Scholar] [CrossRef]
- Jiang, M.; Song, Y.; Yang, R.; Zheng, C.; Zheng, Y.; Zhang, H.; Li, S.; Tan, Y.; Huang, J.; Shu, Q.; Li, R. Melatonin activates the OsbZIP79–OsABI5 module that orchestrates nitrogen and ROS homeostasis to alleviate nitrogen-limitation stress in rice. Plant Comm. 2023, 4, 100674. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Byeon, Y.; Back, K. Functional analyses of three ASMT gene family members in rice plants. J Pineal Res. 2013, 55, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Tzvetkova-Chevolleau, T.; Franck, F.; Alawady, A.E.; Dall’Osto, L.; Carrière, F.; Bassi, R.; Grimm, B.; Nussaume, L.; Havaux, M. The light stress-induced protein ELIP2 is a regulator of chlorophyll synthesis in Arabidopsis thaliana. Plant J. 2007, 50, 795–809. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. 2-Hydroxymelatonin, Rather Than Melatonin, Is Responsible for RBOH-Dependent Reactive Oxygen Species Production Leading to Premature Senescence in Plants. Antioxidants 2021, 10, 1728. [CrossRef]
- Léon-Kloosterziel, K.M; Gil, M.A.; Ruijs, G.J.; Jacobsen, S.E.; Olszewski, N.E.; Schwartz, S.H.; Zeevaart, J.A.; Koornneef, M. Isolation and characterization of abscisic acid-deficient Arabidopsis mutants at two new loci. Plant J. 1996, 10, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.M.; Lynch, T.J.; Mobin, R.; Finkelstein, R.R. Direct targets of the transcription factors ABA-Insensitive(ABI)4 and ABI5 reveal synergistic action by ABI4 and several bZIP ABA response factors. Plant Mol. Biol. 2011, 75, 347–363. [Google Scholar] [CrossRef]
- Gonzalez-Jorge, S.; Ha,S.-H.; Magallanes-Lundback, M.; Gilliland, L.U.; Zhou, A.; Lipka, A.E.; Nguyen, Y.-N.; Angelovici, R.; Lin, H.; Cepela, J.; Little, H.; Buell, C.R.; Gore, M.A.; Dellapennaa D.D. Carotenoid cleavage dioxygenase4 Is a Negative Regulator of β-Carotene Content in Arabidopsis Seeds. Plant Cell 2013, 25, 4812–4826. [CrossRef]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K., Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8'-hydroxylases: key enzymes in ABA catabolism. EMBO J. 2004, 23, 1647-1656. [CrossRef]
- Liu, Z; Yan, J.P.; Li, D.K.; Luo, Q.; Yan, Q.; Liu, Z.B.; Ye, L.M.; Wang, J.M.; Li, X.F.; Yang, Y. UDP-glucosyltransferase71c5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis. Plant Physiol. 2015, 167, 1659-1670. [CrossRef]
- Lee H.J.; Back, K.. 2-Hydroxymelatonin Promotes Seed Germination by Increasing Reactive Oxygen Species Production and Gibberellin Synthesis in Arabidopsis thaliana. Antioxidants 2022, 11, 737. [CrossRef]
- Niu, X.P.; Helentjaris, T.; Bate, N.J. Maize ABI4 binds coupling element1 in abscisic acid and sugar response genes. Plant Cell 2002, 14, 2565–2575. [Google Scholar] [CrossRef]
- Shkolnik-Inbar, D.; Adler, G.; Bar-Zvi, D. ABI4 downregulates expression of the sodium transporter HKT1;1 in Arabidopsis roots and affects salt tolerance. Plant J. 2013, 73: 993-1005. [CrossRef]
- Giraud, E.; Van Aken, O.; Ho, L.H.; Whelan, J. The transcription factor ABI4 is a regulator of mitochondrial retrograde expression of ALTERNATIVE OXIDASE1a. Plant Physiol., 2009, 150, 1286-1296. [CrossRef]
- Acevedo-Hernandez, G.J.; Leon, P.; Herrera-Estrella, L.R. Sugar and ABA responsiveness of a minimal RBCS light-responsive unit is mediated by direct binding of ABIPlant J. 2005, 43, 506–519. [CrossRef]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F; Sachetto-Martins, G.; Surpin, M.; Lim, J., Mittler, R.; Chory, J. Signals from chloroplasts converge to regulate nuclear gene expression. Science 2007, 316, 715–719.
- Ye, T.; Yin, X.; Yu, L.; Zheng, S.J.; Cai, W.J.; Wu, Y.; Feng, Y.Q. Metabolic analysis of the melatonin biosynthesis pathway using chemical labeling coupled with liquid chromatography-mass spectrometry. J. Pineal Res 2019, 66, e12531. [Google Scholar] [CrossRef]
- Yin, X.; Bai, Y. L.; Gong, C.; Song, W.; Wu, Y.; Ye, T.; Feng, Y.Q. The phytomelatonin receptor PMTR1 regulates seed development and germination by modulating abscisic acid homeostasis in Arabidopsis thaliana. J. Pineal. Res. 2022, 72, e12797. [Google Scholar] [CrossRef]
- Chen, L.; Lu, B.; Liu, L.; Duan, W.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Li, C.; Bai, Z. Melatonin promotes seed germination under salt stress by regulating ABA and GA(3) in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem 2021, 162, 506–516. [Google Scholar] [CrossRef]
- Bychkov, I.A.; Andreeva, A.A.; Kudryakova, N.V.; Kusnetsov, V.V. Cytokinin Modulates Responses to Phytomelatonin in Arabidopsis thaliana under High Light Stress. Int. J. Mol. Sci. 2023, 24, 738. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Burgess, O.; Rossi, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in plants-diversity of levels and multiplicity of functions. Front. Plant Sci. 2016, 7, 198. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J. An evolutionary view of melatonin synthesis and metabolism related to its biological functions in plants. J. Exp. Bot. 2020, 71, 4677–4689. [Google Scholar] [CrossRef]
- Kusnetsov, V.V.; Bychkov, I.A.; Kudryakova, N.V. Phytomelatonin As an Element of the Plant Hormonal System. Russ. J. Plant Physiol. 2024, 71, 134. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin induction and its role in high light stress tolerance in Arabidopsis thaliana. J. Pineal. Res. 2018, 65, e12504. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods.Enzymol 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Heath, L.R.; Packer, L. Photoperoxidation in isolated chloroplasts. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: a database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–1147. [Google Scholar] [CrossRef]
- Palaniswamy, S.K.; James, S.; Sun, H.; Lamb, R.S.; Davuluri, R.V.; Grotewold, E. (2006)AGRIS and AtRegNet: A platform to link cis-regulatory elements and transcription factors into regulatory networks. Plant Physiology 2006, 140, 818-829 PMID 16524982. [CrossRef]
- Bychkov, I.A.; Pojidaeva, E.S.; Doroshenko, A.S.; Khripach, V.A.; Kudryakova, N.V.; Kusnetsov. V.V. Phytohormones as Regulators of Mitochondrial Gene Expression in Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 16924. [CrossRef]
Figure 1.
Effect of high light stress and ABA treatment on the TBARs content, сhlorophyll content and expression of stress marker genes. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). ABA was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences among variants within the same genotype at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test).
Figure 1.
Effect of high light stress and ABA treatment on the TBARs content, сhlorophyll content and expression of stress marker genes. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). ABA was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences among variants within the same genotype at p < 0.05 (ANOVA with post hoc Tukey's multiple-comparison test).
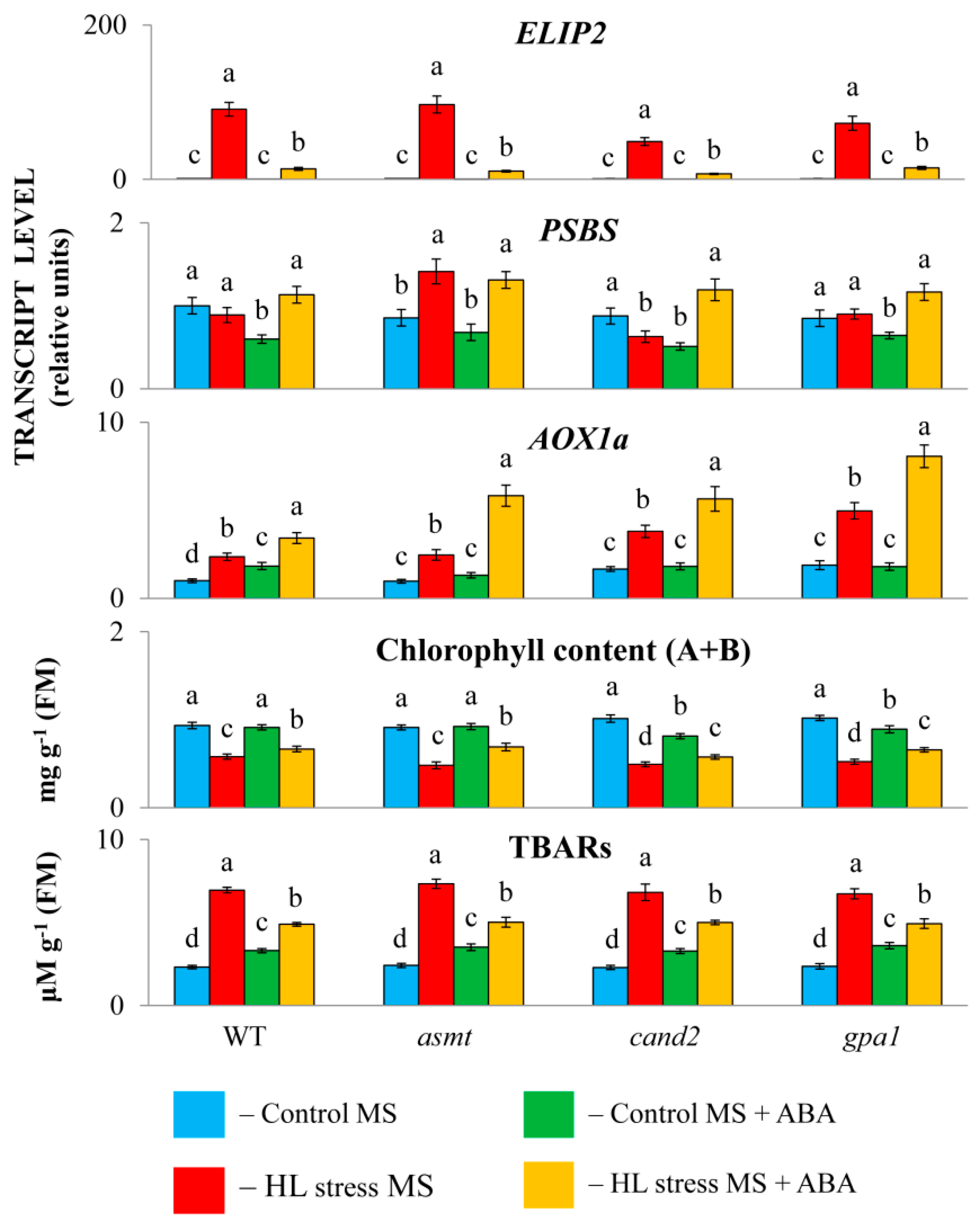
Figure 2.
Effect of high light stress and ABA on the expression of melatonin metabolism and signaling genes in MT-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). ABA was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
Figure 2.
Effect of high light stress and ABA on the expression of melatonin metabolism and signaling genes in MT-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). ABA was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
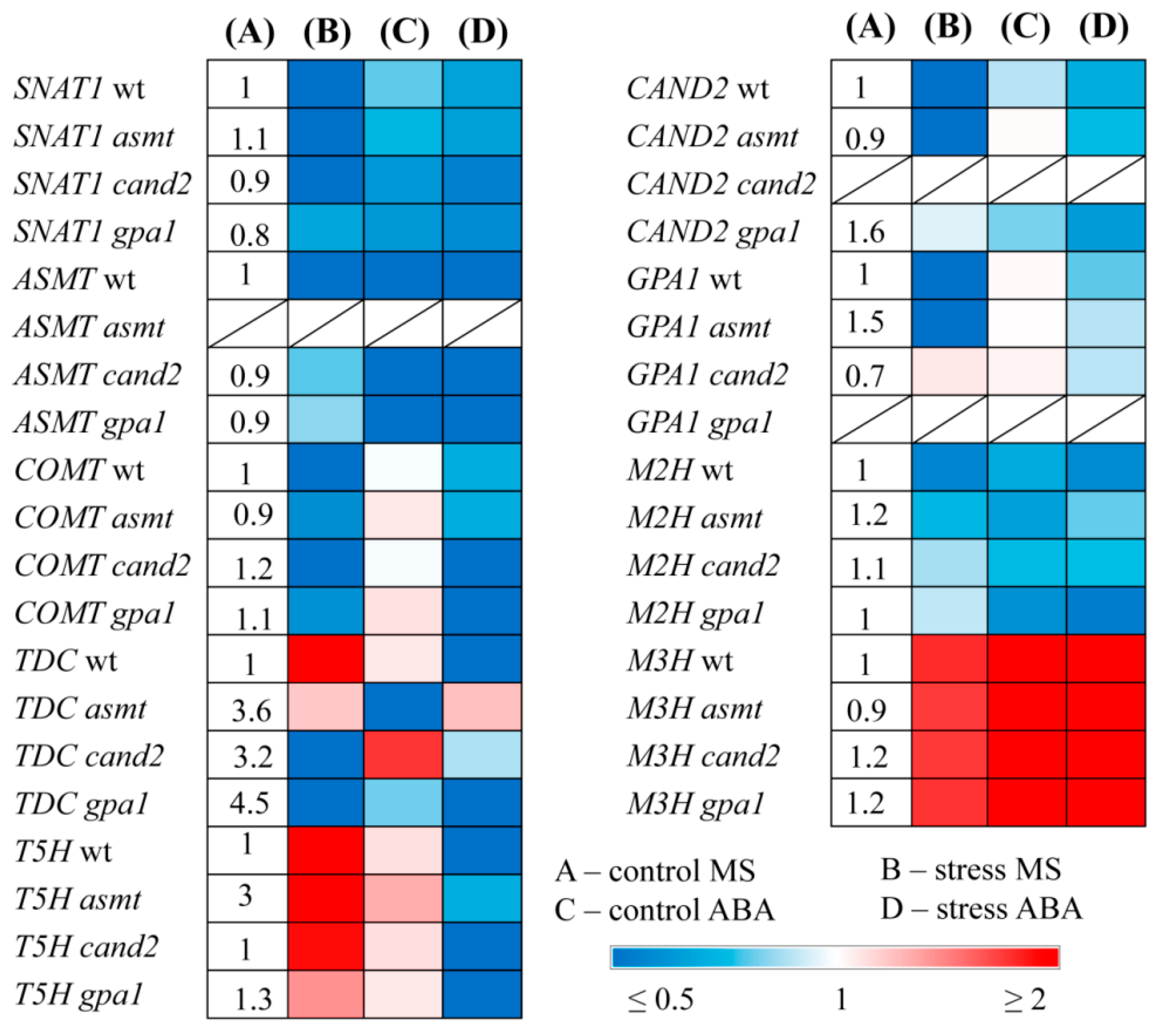
Figure 3.
Effect of high light stress on melatonin content. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences among variants within the same genotype at p < 0.05, asterisks indicate statistically significant differences between the mutants and the wild type under corresponding type of treatment at p < 0.05 (t-test).
Figure 3.
Effect of high light stress on melatonin content. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The data presented in the figure are the mean values (n ≥ 3). Error bars represent SEs. Different letters denote statistically significant differences among variants within the same genotype at p < 0.05, asterisks indicate statistically significant differences between the mutants and the wild type under corresponding type of treatment at p < 0.05 (t-test).
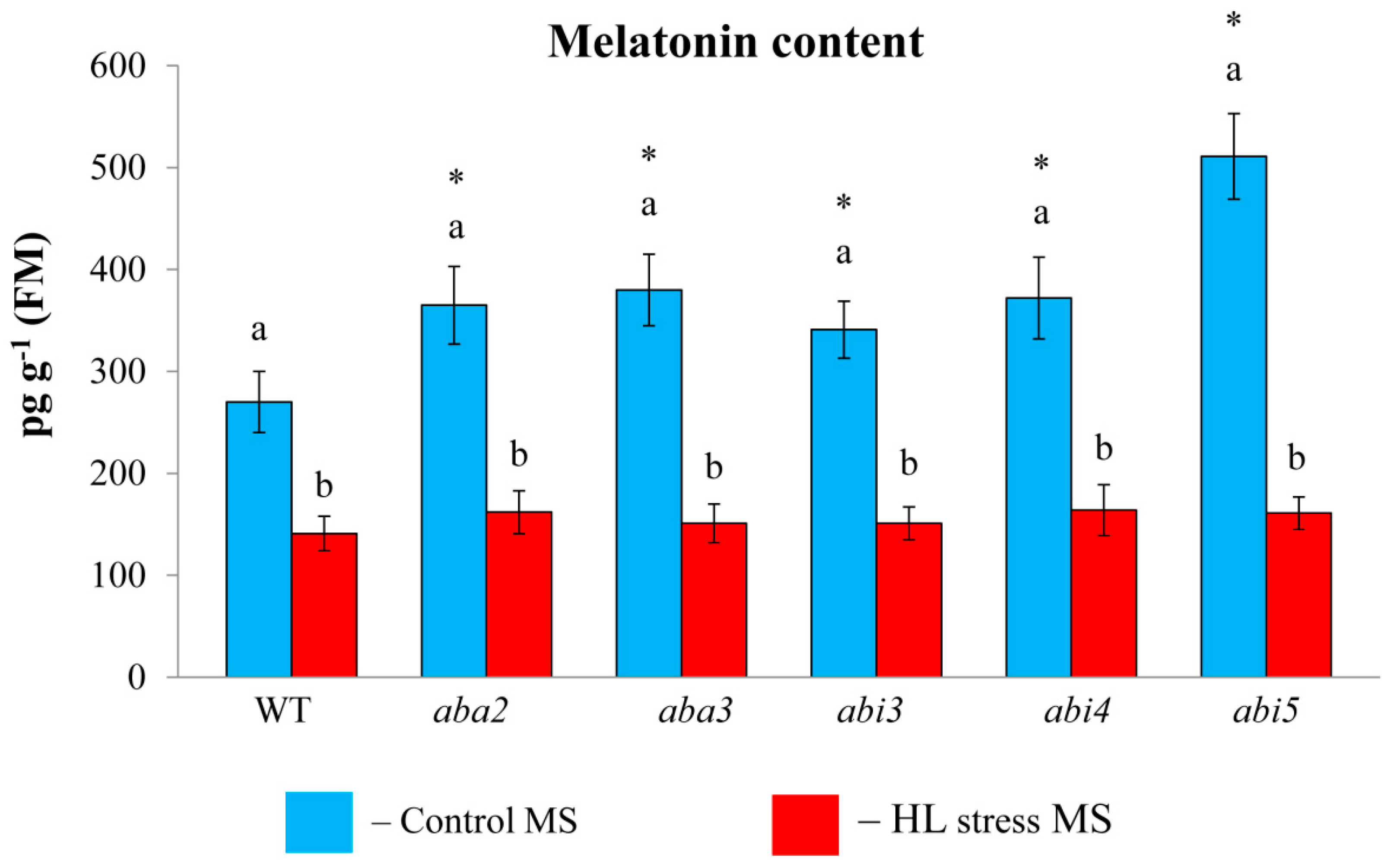
Figure 4.
Effect of high light stress, melatonin and ABA on the expression of melatonin metabolism and signaling genes in ABA-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Melatonin and ABA were used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
Figure 4.
Effect of high light stress, melatonin and ABA on the expression of melatonin metabolism and signaling genes in ABA-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Melatonin and ABA were used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress). The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
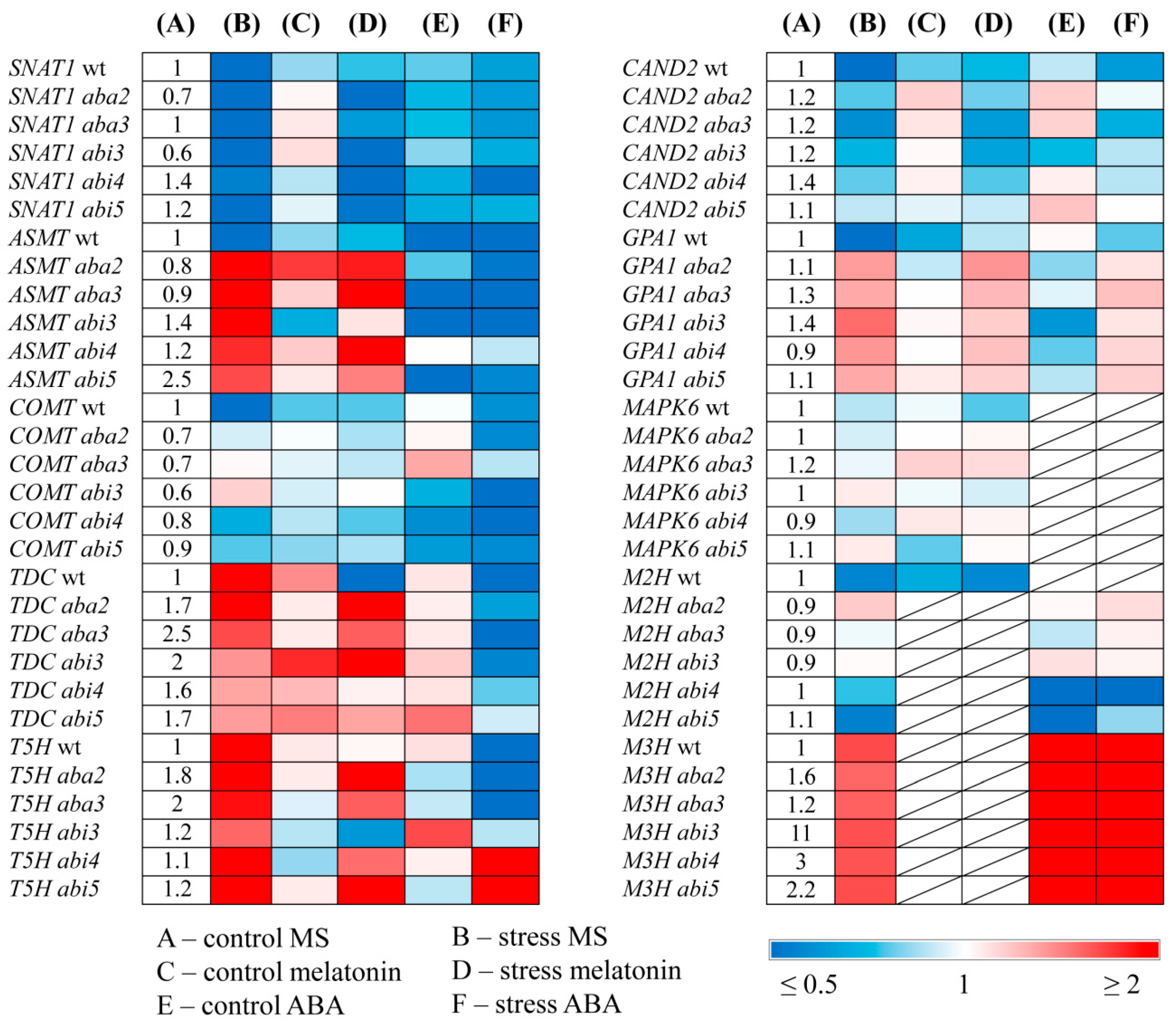
Figure 5.
Effect of high light stress and melatonin on the expression of ABA metabolism and signaling genes in MT-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Melatonin was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress).The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
Figure 5.
Effect of high light stress and melatonin on the expression of ABA metabolism and signaling genes in MT-mutants. WT and mutant plants were grown on Murashige and Skoog (MS) medium in Petri dishes for two weeks under a 16 h light / 8 h dark photoperiod at 23°C with 60 μm m-2 s-1 (control). Melatonin was used for treatment at a concentration of 50 μM. Experimental plants were exposed to high light (HL) for 24 hours at 600 μm m-2 s-1 (stress).The numbers in the “A” column indicate the baseline ratio of the expression of each gene in the wild type and mutant without treatments.
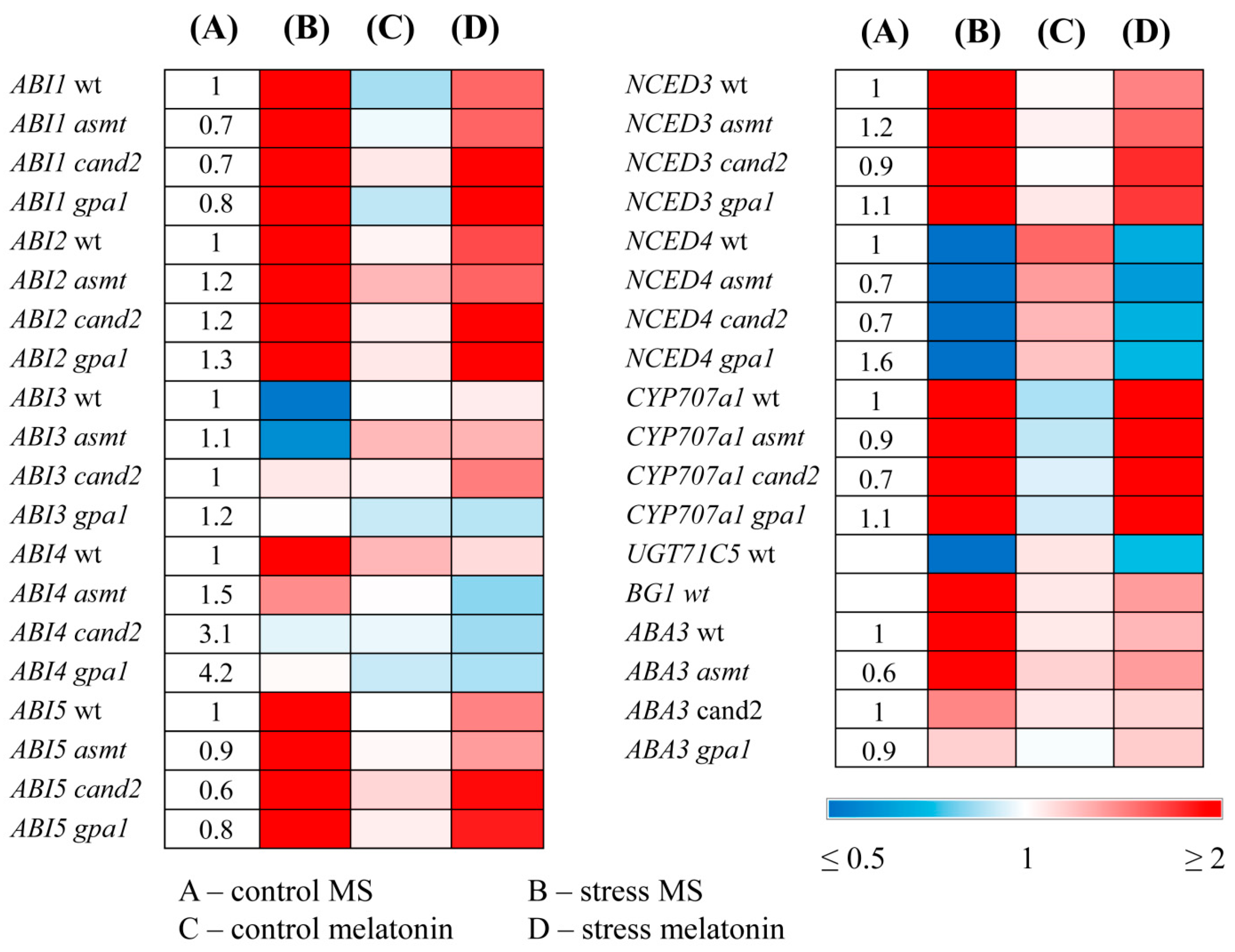
Figure 6.
The promoter region of the ASMT gene, 2483 bp in length, including the 5’-UTR region. The nucleotide sequence for the 5’-UTR is highlighted in blue. The basal promoter sequence of the gene, −1000 bp from the ATG initiation codon, identified using the PlantProm DB program, is shown in gray. CCAC motifs are shown in green. ABI4-binding elements CE1 CACCG are shown in yellow. ABRE-like motif TACGTGT is shown in red. According to the program https://agris-knowledgebase.org/, the length of the promoter of this gene can reach a longer region of 2483 bp. The TATA box sequence is highlighted in bold and ATG is start codon in red.
Figure 6.
The promoter region of the ASMT gene, 2483 bp in length, including the 5’-UTR region. The nucleotide sequence for the 5’-UTR is highlighted in blue. The basal promoter sequence of the gene, −1000 bp from the ATG initiation codon, identified using the PlantProm DB program, is shown in gray. CCAC motifs are shown in green. ABI4-binding elements CE1 CACCG are shown in yellow. ABRE-like motif TACGTGT is shown in red. According to the program https://agris-knowledgebase.org/, the length of the promoter of this gene can reach a longer region of 2483 bp. The TATA box sequence is highlighted in bold and ATG is start codon in red.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



