
Submitted:
16 October 2024
Posted:
17 October 2024
You are already at the latest version
Abstract
This study investigated commensal and pathogenic E. coli from pigs at farms and slaughterhouses in Sardinia, focusing on genetic relatedness and antimicrobial resistance (AMR). Samples were collected from six fattening pig farms (A-F) and five slaughterhouses (S1-S5). In the farms, environmental fecal sampling from the fattening pigs’ pens was carried out and information regarding farm management and biosecurity measures were collected. Pigs that had been in the sampled pens were selected for sampling at the slaughterhouse. Mesenteric lymph nodes, colon content and carcass surface samples were collected at the five slaughterhouse (S1-S5), in a total of 38 samples form 152 animals. At the slaughterhouses, also environmental samples were collected from food-contact surfaces and non-food-contact surfaces (36 samples overall). E. coli was detected in all farms, 97% of pigs, and 100% of slaughterhouses. Whole genome sequencing and antimicrobial susceptibility testing were per-formed on 96 isolates, revealing 13.5% pathogenic isolates, including ExPEC, ETEC, STEC-ETEC hy-brids, and UPEC. A total of 40 sequence types (STs) were identified, with ST10 being the most com-mon. High-risk clones (ST88, ST101, ST410, and ST648) were also detected. Over half of the isolates (52.1%) carried at least one AMR gene, with 42.7% harboring multiple AMR genes, particularly tet (37.5%) and blaTEM (32.3%). High phenotypic resistance was observed for tetracycline, ampicillin, and sulfamethoxazole-trimethoprim. The study highlights the widespread presence of pathogenic E. coli in pigs, even in isolated environments, and emphasizes the need for continuous surveillance due to the significant AMR found in both pathogenic and non-pathogenic isolates.
Keywords:
commensal
; fattening pigs
; virulence genes
; surveillance
; production chain
1. Introduction
Escherichia coli (E. coli) is a widespread bacterium and the isolates are classified as non-pathogenic or pathogenic based on virulence factors. Non-pathogenic E. coli are commensal members of the natural microbiota of humans and warm-blooded animals [1]. Pathogenic E. coli are causative agents of intestinal and extraintestinal diseases in humans and animals and are classified into different “pathotypes” based on the presence of specific virulence genes. Among the intestinal pathogens, E. coli are classified as enterotoxigenic (ETEC), enteropathogenic (EPEC), Shiga toxin-producing (STEC), enteroinvasive (EIEC), enteroaggregative (EAEC), and diffusely adherent (DAEC). Extra-intestinal pathogenic E. coli, usually called ExPEC, can be further classified as uropathogenic E. coli (UPEC) and sepsis or meningitis-associated E. coli (MNEC) [2]. Many pathotype-specific virulence markers are frequently carried on mobile genetic elements such as plasmids and phages. The ability of E. coli to acquire virulence and related genes via horizontal gene transfer leads to the development of pathogroups that are different from the pre-existing ones; such divergent pathogroups are often reported as “hybrids” [3].
Domestic pigs play an important role as reservoirs for different E. coli pathotypes. In particular, the stx2e gene-carrying STEC, which are the source of oedema disease commonly affecting post-weaning pigs, cause significant financial losses in the pig industry [4]. Moreover, enteric colibacillosis is a common disease in weaning pigs caused by ETEC [5]. Given the global significance of pathogenic E. coli as a foodborne pathogen [6] and the relevance of pork meat in foodborne outbreaks [7], the control of this microorganism in the pig food chain is crucial in food safety. In this framework, farms and slaughterhouses are the proper points of the food chain to carry out epidemiological studies on zoonotic agents.
Antimicrobial resistance (AMR) is a global problem that threatens human and animal health. AMR bacteria can be transmitted through the food chain, in which numerous points of entry are possible from farm to fork, and particularly through the consumption of raw or undercooked meat products [8]. The major aim of food control is to avoid the spread of pathogens, as stated in Regulation (EC) No 2073/2005. Nevertheless, carriers of antimicrobial resistance may pose health hazards regardless of an isolate's virulence characteristics. Especially, commensal bacteria may serve as a reservoir for genetic transmission in the gut: research on patients indicates that resistant bacteria obtained from oral ingestion of pork meat can persist in the gastrointestinal tract and be found in the faeces for as long as 14 days following ingestion [9]. In this framework, according to Commission Implementing Decision (EU) 2020/1729, monitoring of AMR is mandatory in Salmonella spp., Campylobacter coli and jejuni and indicator commensal E. coli in the major domestically produced animal populations and their derived meat. Specific monitoring of extended-spectrum β-lactamases (ESBL), AmpC β-Lactamases (AmpC) and carbapenemases producing Salmonella and indicator commensal E. coli is also required. ESBL and AmpC-producing E. coli have been isolated from various food-producing animals in several EU countries, including pigs. This suggests that food and animals may play an important role as reservoirs [10].
In this context, the objective of this work was to study the prevalence of pathogenic E. coli in fattening and to characterize commensal and pathogenic E. coli isolates in pigs in farms and at slaughter in Sardinia and to characterize commensal and pathogenic E. coli isolates.
2. Materials and Methods
2.1. Study Design
Six fattening pig farms (A, B, C, D, E, F) distributed throughout the region (Sardinia, Italy) were selected; farms were comparable to each other in terms of procedures adopted and with a minimum of 150 pigs and 50 fattening pigs. Three out of six (50%) of the farms reported no use of antibiotics on the animals. In the selected farms, the pigs were kept in paddocks of 8-10 pigs each and, by the end of the fattening period, the animals were transported to slaughterhouses located less than 100 km (less than 2 h) from each farm, and they were slaughtered within 15–18 h of fasting. On each farm, a group of finisher pigs of at least 16 weeks of age, kept within the same pen and intended to be slaughtered in the following 14 days, were selected for sampling at the farm and the slaughterhouse. In the farms, environmental sampling was carried out on the floor of the fattening pigs’ pens with sterile socks kits (Techinal Service Consultants Ltd; Heywood, NW, United Kingdom); a pooled faecal sample was collected from the pens, by walking on the floor, covering at least 50% of the pen, closely to the walls, into the corners, around the water supply and the trough. After sampling, the socks were placed into individual sterile plastic bags at refrigeration temperature. Using a checklist, the farmer was also questioned about the management and biosecurity practices used on the farm, including questions regarding the cleaning procedures and antibiotics used.
No more than 14 days after environmental samples at the farms, samples from the same pigs were collected at the slaughterhouse. Based on the slaughterhouses to which the farms of origin referred, 5 slaughterhouses were subjected to sampling (S1, S2, S3, S4 and S5). Two farms (Farm A and Farm B) referred to the same slaughterhouse (S1), which was sampled twice. Only pigs that had been in the sampled pens were selected for sampling at the slaughterhouse. As many of these were small farms and transported only a few animals at one time, all pigs coming from the same farm and slaughtered on the same day were considered a single batch. The sampling included all pigs from the sampled pens that were sent to the slaughterhouse within 14 days. Overall, 38 animals were sampled, divided as follows: 6 pigs from Farm A, 3 pigs from Farm B, 10 pigs from Farm C, 3 pigs from Farm D, 10 pigs from Farm E and 6 pigs from Farm F. A total of 114 animal samples were obtained, including mesenteric lymph nodes, colon content and carcass surface samples from each pig. In detail, immediately after evisceration, mesenteric lymph nodes and colon content samples were collected. Mesenteric lymph nodes (at least 25 g) were collected with a sterile, disposable scalpel. The colon was incised and at least 25 g of its contents were collected. Before chilling, carcass surface samples were taken by non-destructive method with a sterile sponge pre-moistened with 10 ml of sterile Buffered Peptone Water (3M Health Care, Milano, Italy); according to ISO 17604:2015 [11], sampling was carried out using the same sponge for four points in the carcass, namely ham, loins, abdomen and throat, with a sterile 10x10 cm2 delimiter (Copan, Brescia, Italy), from the least contaminated point (ham) to the most contaminated (throat). The sponges were handled with a sterile glove and placed inside sterile sponge bags.
Environmental samples were also collected at the slaughterhouses. Surfaces were sampled at the end of the slaughtering operations of the pig batch and before cleaning procedures, using a sterile sponge and a sterile delimiter (10x10 cm2). The following surfaces were sampled:
- Food contact surfaces (FC): cutting equipment (knives, saws) and hair removal equipment (brushes or whips)
- Non-food contact surfaces (NFC): walls near the stunning and killing area, walls and drain surface of the pre-chilling area
- Scalding water (SW): approximately 100 mL of scalding water, collected using a sterile sampler (Bibby Scientific Limited, Stone, UK).
As for environmental samples, 36 samples were collected overall, including 12 FC samples, 18 NFC samples and 6 SW samples.
All the samples were transported to the laboratory at +4±1 °C and processed within 24 h after collection.
2.2. Microbiological Analysis
E. coli detection was conducted on the samples using an in-house method, which included an enrichment phase in EC Broth selective medium (Biolife, Milan, Italy) and isolation in Levine EMB Blue Agar (Biolife, Milan, Italy), as previously described [12]. Species confirmation was conducted by mass spectrometry, using Matrix-Assisted Laser Desorption/Ionization–Time of Flight (MALDI–TOF).
2.3. Whole Genome Sequencing
Whole genome sequencing (WGS) was carried out on a selection of 100 E. coli isolates. In particular, the isolates selection ratio was as follows: 6 environmental samples from pig farms (one sample per farm), 81 samples from slaughtered pigs (one isolate for each type of positive sample, among lymph nodes, colon contents and carcass surface samples, for each pig) and 13 environmental samples from slaughterhouses (one isolate for each type of positive sample for each slaughterhouse).
Genomic DNA was extracted with an enzymatic pre-lysis step; automated purification was conducted according to the producer’s instruction using the MagNA Pure 96 DNA and Viral NA Small Volume Kit and DNA Blood ds SV 2.0 protocol (Roche Diagnostics, København, Denmark). Genomic libraries were constructed and sequencing was carried out on the NextSeq® 550 (Illumina, San Diego, USA) platform using the Nextera XT Kit (Illumina, San Diego, USA) and 300-cycle kits. Quality control of the obtained sequencing data was conducted using Bifrost software (Bifrost Inc, Westerly, Rhode Island, US) to ensure adequate sequencing depth, species verification and identify contamination issues.
E. coli phylogroups were determined via Enterobase [13]. Core genome MLST for E. coli (according to the Applied Maths/Enterobase scheme) and single linkage clustering tree were calculated in Bionumerics 8.1 (Applied Maths, Sint Martems Latem, Belgium). In silico serotyping was analyzed from read mapping with the SerotypeFinder database [14] using the KMA mapping tool [15].
Resistance, virulence and plasmid-associated genes were analyzed in all samples using BioNumerics and CGE tools including AMRFinder [16], ResFinder [17], PlasmidFinder [18] and VirulenceFinder [19].
Aggregate virulence gene scores (VGS) were calculated for each isolate by summing the number of virulence genes detected from the genome sequences.
Raw sequence FASTQ data for this study are available at NCBI, under Bioproject PRJNA1171362.
2.4. Antimicrobial Susceptibility Testing
The disc-diffusion technique was utilized to evaluate the isolates' antibiotic resistance in accordance with the guidelines provided by the European Committee on Antimicrobial susceptibility Testing [20]. Commercial antimicrobial susceptibility discs (ThermoFisher Scientific, USA) and Mueller-Hinton agar (Microbiol, Cagliari, Italy) were utilized. All isolates were tested for amikacin (Ak, 30 μg), ampicillin (Amp, 10 μg) amoxicillin/clavulanic acid (Aug, 20 μg and 10 μg, respectively), azithromycin (Azm, 15 μg) cephazolin (Kz, 30 μg), cefoxitin (Fox, 30 μg), ceftriaxone (Cro, 30 μg), cefotaxime (CTX, 30 μg), ceftazidime (Caz, 5 μg), ciprofloxacin (Cip, 5 μg), imipenem (Ipm, 10 μg), kanamycin (K, 30 μg), levofloxacin (Lev, 5 μg), meropenem (Mem, 10 μg), nalidixic acid (Na, 30 μg), streptomycin (S10, 10 μg), tetracycline (Te, 30 μg), trimethoprim/sulfamethoxazole (Sxt, 1:19, 25 μg). According to the test results, isolates were categorized as susceptible or resistant according to the EUCAST recommendations; intermediate isolates were considered susceptible [20]. Isolates displaying resistance to at least three antimicrobial groups were considered multi-resistant (MR) [21].
2.5. Statistical Analysis
Differences in the prevalence of E. coli between samples (lymph nodes, colon content and carcass surface), farms and slaughterhouses were evaluated using One-way ANOVA with post-hoc Tukey HSD with Statgraphics-Centurion XIX software (Stat Point Technologies, Warrenton, VA, USA). The significance level was defined as p < 0.05.
3. Results
3.1. Farms
Farms were comparable to each other in terms of processing, breeding techniques and procedures adopted. Four of the pig farms also had piglet production in addition to fattening (A, B, C, E); one farm (E) purchased replacement piglets from outside Sardinia and the animals, before being introduced into the farm, had to spend 21 days in isolation and quarantine. The median value of fattening pigs present on the farms was 170 (ranging from 80 to 700). The fattening pigs were kept in paddocks with around ten pigs each. Three out of six farms had a slotted floor, and two had an external paddock for each pen. In 4/6 farms, a comprehensive cleaning on fully empty stables after a fattening cycle was carried out (all-in-all-out approach) in addition to the daily removal of excrement from the pens; in the two other cases (farms C and F), cleaning was not structured and the all-in-all-out approach was not applied. Animals were mainly fed commercial pelleted feed, with the occasional addition of whey (farm B and D). The most frequent pathologies in the farms were pneumonia, abortions and dermatitis. As regards the use of antibiotics, 3/6 (50%) farms reported not using any antibiotic substance, whilst the remaining farms reported using amoxicillin, trimethoprim-sulfamethoxazole and oxytetracycline. Most Farms reported the administration of antibiotics to individual animals in cases of disease, except for Farm B which reported the administration of trimethoprim-sulfamethoxazole along with the feed if a group of pigs had health issues. Management characteristics of the farms are summarized in Table 1.
3.2. Microbiological Analysis
Sequencing confirmed the E. coli species in 96/100 isolates. The isolates belonging to a different species were excluded from further analysis. As reported in Table 2, E. coli was detected in 100% of sampled farms (6/6), in 97.4% of sampled pigs (37/38) and in 100% (5/5) of sampled slaughterhouses.
Regarding the samples collected from pigs, E. coli isolates were detected overall in 81/114 (71.1%) samples, more specifically in 32/38 (84.2%) colon content samples, 26/38 (65.8%) lymph nodes samples and 24/38 (63.1%) carcass surface samples. Among the pigs, 17/38 (44.7%) animals tested simultaneously positive in all the samples tested (lymph nodes, colon content and carcass surface), 10/38 (26.4%) tested positive in two samples tested (of which 7/10 tested positive in lymph nodes and colon content, 2/10 in colon content and carcass surface and 1/10 in lymph nodes and carcass surface), and 10/38 (26.4%) were positive in only one out of three types of samples (of which 6/10 tested positive in colon content samples, 4/10 in carcass surface samples). A breakthrough of positive samples in pigs is shown in Figure 1.
Regarding the environmental samples collected from slaughterhouses, E. coli was detected in 13/36 (36.2%) samples, specifically in 6/12 of FC samples (namely, two from bristles removal equipment and four from cutting equipment), 4/18 NFC samples (one from walls in the stunning and bleeding area, two from the pre-chilling room's walls and one from the drain surface) and 1/6 SW samples. Higher occurrences were observed in slaughterhouses S3 and S4, in which 44% of samples (4/9) showed positivity for E. coli; in both slaughterhouses, E. coli was equally found in FC and NFC surfaces.
3.3. E. coli Characterization
Phylogenetic group A (58/96, occurrence of 60.4%) and B1 (28/96, 29.2%) were the most common among the isolates. Regarding the predicted pathotype, 7/96 (7.3%) of the isolates fit into the ExPEC pathotype, 3/96 (3.1%) of the isolates were ETEC, 2/96 (2.1%) were STEC/ETEC hybrids, and 1/96 (1%) of the isolates was UPEC. Table 3 shows the genotypic characterization of the isolates.
Overall, pathogenic E. coli isolates were detected in 10/38 (26.3%) of the studied pigs. As for sample types, colon content samples had the highest percentage of positivity (Table 6), showing positivity for STEC-ETEC in two pigs (2/38, 5.3%) and for ETEC in two more pigs. Lymph nodes samples accounted for ExPEC positivity in one pig. Moreover, one pig (1/38, 2.6%) was an ExPEC carrier only in the carcass surface sample. Simultaneous positivity was observed in two pigs: one (1/38, 2.6%) tested positive for ExPEC at colon content and carcass surface samples and another (1/38, 2.6%) at colon content and lymph node samples. No statistically significant difference was observed in the prevalence of pathogenic isolates in the samples (p>0.05).
At least one virulence gene was observed in all of sequenced E. coli isolates, with a mean value of 17.9 ± 10.9 and a median of 13.5 virulence genes per isolate; higher rates of virulence genes were observed in isolates with a predicted pathotype (p < 0.05). Among isolates that were not classified in any pathogenic group, a mean value of 15.3 ± 9.9 (median 11) virulence genes per isolate was observed, with 27/84 (32.1%) isolates carrying ≥ 20 and 10/84 (11.9%) carrying ≥ 30 (Table 3). The most common virulence genes in E. coli isolates were terC (96/96, 100%), mdtM (92/96, 95.8%) and acrF (88/96, 91.7%).
Most ExPEC isolates (4/7, 57.1%) had the heat-stable enterotoxin-1 gene astA, with two or more other virulence genes typical of ExPEC strains [22]. The three ETEC isolates identified possessed typical ETEC genes, namely the stb gene (2/3, 66.7%) and the astA gene (3/3, 100%), with two or more other virulence genes. The two STEC-ETEC hybrid isolates identified in our investigation possessed the stx2e gene (2/2, 100%) and other virulence genes. Complete genetic characterization and virulence genes of the isolates are reported in Table 3.
Overall, 47 serotypes were identified; the most common was O130:H26 (6/96, 6.2%). In 26 isolates, the in-silico serotyping was not able to identify the O antigen.
A total of 40 sequence types (ST) were detected in the isolates collected in the present survey. The most common ST was ST10 (15/96, 15.6%). ExPEC isolates detected in this investigation were ST88 (5/7), ST101 (1/7) and ST847 (1/7). The UPEC isolate detected in this investigation was ST648 (1/1). The three ETEC isolates identified in this investigation were ST196 (1/3), ST410 (1/3) and ST5771 (1/3). The two STEC-ETEC hybrid isolates identified in our investigation were ST46 (2/2).
3.4. Cluster Analysis
The cgMLST cluster analysis on E. coli isolates reported in Figure 2 shows that some isolates belonging to the same ST were genetically close with < 5 allelic differences (AD).
Isolates with the same STs collected from pigs coming from the same farm clustered together, as can be observed for ST10 isolates from Farm A or ST345 isolates from Farm C. In general, no mixing of isolates between farms was observed, with some exceptions: three isolates from farm F (ST10 and ST877) showed close genetic similarity with isolates collected from pigs coming from Farm A, Farm C and Farm E; furthermore, two isolates (ST14809) from pigs coming from Farm A and Farm E were also genetically closely related. The biggest cluster involved five ST877 isolates with less than 10 AD; isolates were collected from samples from three animals coming from Farm E and from an environmental sample from FC surface from slaughterhouse S5. Four ST88 clustered together, with less than 10 AD: these isolates were detected on samples from animals belonging to the same farm and were all ExPEC pathotypes.
Different STs were also detected in isolates collected from environmental samples at the slaughterhouses. As shown in Figure 2, two ST1114 isolates showed a high genetic similarity, with less than 10 AD; in particular, these isolates were identified in a SW sample and in a NFC sample of the drainage channel of the slaughterhouse S3, after the slaughter of pigs from farm D. Moreover, as mentioned, one ST877 isolate from an environmental sample from FC surface (chainsaw) in slaughterhouse S5 was genetically closely related (less than 10 AD) with other ST877 isolates recovered from pigs coming from Farm E. Another genetic similarity was observed between two ST746 isolates found from a FC surface (dehairing whip) and a pig sample during the slaughter of animals coming from farm E.
3.5. AMR Characterization
Overall, 50/96 (52.1%) isolates had at least one AMR gene and 41/50 (82%) had two or more AMR genes. As for sample types, in 5/6 (83.4%) farms, 39/80 (48.7%) samples from pigs and 9/13 (69.2%) environmental samples from slaughterhouses at least one resistance gene was observed.
In the E. coli isolates, thirty antimicrobial resistance genes were identified overall, namely genes encoding resistance (from the most common to the least) to tetracyclines (tetA, tetB, tetM), ß-lactams (blaTEM-1A, blaTEM-1B, blaTEM-1C, blaTEM-1D), aminoglycosides (aadA1, aadA2, ant(3'')-Ia, aph(3')-Ia, aac(3)-IId, aph(6)-Id, aph(3'')-Ib), sulphonamide (sul1, sul2, sul3), trimethoprim (dfrA1, dfrA5, dfrA12), cloramphenicols (floR, cmlA1), streptothricin (sat2), quinolones (qnrS1, qnrB19), fosfomycin (fosA7.5), macrolides (mefB, mphA, lnuG) and colistin (mcr-10.1).
More specifically, the most detected AMR genes were the tet gene (36/96, 37.5% of isolates), which was mostly found in pig isolates (31/80, 38.7%), and blaTEM-1 genes (31/96, 32.3% of isolates), with blaTEM-1B being the most prevalent (25/31, 80.6%).
Regarding pathogenic isolates (13 overall), three ExPEC (3/13, 23.1%) and one ETEC (1/13, 7.7%) had AMR genes, more specifically one ExPEC isolate had the tet(B) and fosA7 genes, one ExPEC isolate had tet(B), one ExPEC isolate had the blaTEM-1B, tet(A) and floR genes and one ETEC isolate had the fosA7.5 gene.
If we divide the isolates based on the farm of origin, some discrepancies were observed. Although some farms declared not to use antibiotics, AMR genes were detected in all the sampled farms. For instance, farm B reported no use of antibiotic substances, however in E. coli isolates ß-lactams (blaTEM-1A and blaTEM-1B), aminoglycosides (aadA1, aph(6)-Id and aph(3'')-Ib), macrolides (inuG), tetracycline (tetA, tetB), trimethoprim (dfrA1), sulfonamides (sul3), chloramphenicol (floR) and streptothricin (sat2) resistance genes were detected. Moreover, also genes conferring resistance to substances not reported by farmers were observed, like in the case of Farm C. This Farm reported the use of amoxicillin-clavulanic acid and oxytetracycline, and in the E. coli isolates the ß-lactams genes blaTEM-1B and blaTEM-1D and the tetracycline resistance genes tet(A) and tet(B) were detected. However, also aminoglycosides (aadA1, aadA2, ant(3'')-Ia, aac(3)-IId, aph(6)-Id and aph(3'')-Ib), macrolides (mphA), trimethoprim (dfrA12), Fosfomycin (fosA), streptotricin (sat2) and sulfonamides (sul2) resistance genes were detected in Farm C samples.
Regarding the phenotypic AMR of isolates, 49/96 (51%) isolates showed resistance to at least one antimicrobial tested. Resistance against tetracycline was detected in 70% of the resistant isolates, followed by ampicillin (58%) and sulfamethoxazole-trimethoprim (36.4%). Twenty-three different phenotypic patterns were identified with the most common being Te (12/49, 24.5%), followed by AmpTe and AS16TeSxt (6/49, 12.2% each). Regarding pathogenic E. coli, 3/13 (23.1%) and only ExPEC isolates, showed phenotypic AMR against the antimicrobials tested: two isolated showed Te resistance pattern and one isolated AmpTe. Unexpectedly, an isolate from environmental sampling at a farm carrying the mcr-10.1 gene was detected. A complete AMR characterization of the isolates, divided by farm, is summarized in Figure 3.
4. Discussion
The E. coli population isolated from pig samples was diverse, with mostly non-pathogenic isolates from various phylogroups. Phylogroup A accounted for 60.4% of the isolates, the majority of which (97%) did not belong to known pathotypes. This indicates that E. coli isolates belonging to phylogroup A are often commensal and non-pathogenic [23]. E. coli isolates belonging to know pathotypes were not detected in environmental samples but were found in pigs from Farms C, E, and F. When comparing this outcome with farm management methods, the pens were never fully emptied during cleaning processes, and the all-in-all-out cleaning and disinfection strategy was not used in Farms C and F. The lack of a systematic cleaning process in these farms could have facilitated the spread of these pathogens from one batch to the next and cross-contamination across production cycles [24]. The Farm E was the only farm that purchased replacement piglets and did not use a closed cycle. Movement of animals poses a major risk for disease introduction [25], and despite monitoring and quarantine, the arrival of potentially ill animals may have introduced pathogenic strains. The importance of appropriate biosecurity measures in farms to ensure safety are confirmed.
All of the E. coli isolates carried at least one virulence gene. The average was 17.9 genes per isolate. Even isolates that did not belong to known pathotypes and were regarded as non-pathogenic may pose a risk of virulence due to this high number of virulence genes.
In our investigation, the most commonly detected E. coli pathotypes were ExPEC and UPEC. There is currently scarce information available on the epidemiology of these pathotypes in pigs and slaughterhouses in Italy; nevertheless, ExPEC and UPEC isolates have been found in wastewater from German pig slaughterhouses [26]. A concerning result was the detection of STs that are common in human infections worldwide. Particularly, ExPEC isolates belonging to ST88 and ST101 have been linked to a majority of human infections in both community- and health-related contexts [27,28].
Also, the ST648 associated with UPEC is among the most often reported STs in the human-animal-environment interface globally and are regarded as critical priority STs [29,30]. Pig meat has been linked to ExPEC isolates [31,32] and there has been discussion about food animals, especially pigs, acting as reservoirs for ExPEC and UPEC [33,34]. Even though ExPEC does not cause gastrointestinal illness, illness, it may spread through contaminated meat from the gastrointestinal tract to other sites, such as the blood or urinary tract [35]. This paradigm shifts of E. coli as a foodborne pathogen, highlights a different risk pathway. Case-control studies have shown a higher incidence of urinary tract infections caused by antimicrobial-resistant E. coli in individuals who frequently consume pork and chicken [31,32], further supporting the potential transmission of foodborne pathogens like UPEC through meat.
The ST410 is another extremely virulent pathogenic ST, which is often linked to genes that are characteristic for the ExPEC pathotype [36]. However, the isolate found in our study possessed common ETEC virulence genes, e.g. the heat-stable enterotoxin stb gene, which is mostly linked to pig ETEC infection [37,38].
We found two STEC-ETEC hybrid isolates (ST46) from pig colon content, which probably have scarce relevance for public health. The isolates had the stx2e gene, which encodes for a Shiga toxin 2 variant, typically causing an E. coli edema disease in pigs [39]. Isolates with this virulence profile are considered not particularly pathogenic for humans [40,41].
ST10 was the most prevalent sequence type (15.6%) among isolates regarded as non-pathogenic. ST10 is an emerging ST among E. coli isolates, known as a low-virulent, antibiotic-susceptible human intestine colonizer that is also connected to human infections and the development of ESBLs [33]. The fact that ST10 isolates were widely discovered in farms in our study confirms that this ST is widely distributed. Although ST10 isolates did not belong to any known pathotypes, the heat-stable enterotoxin-1 gene astA was detected in three (16.7%) isolates; moreover, ST10 isolates possessed up to 36 virulence genes, often typical for ExPEC (irp2, iss, ompT, terC) and ETEC (fdeC, mdtM, ymgB) pathotypes.
The single linkage tree (Figure 2) shows that some isolates from the same ST were genetically highly related, grouping by farm origin. For example, Farm A's ST10 isolates and Farm C's isolates grouped together, which shows that genetically highly related isolates circulate among pigs on the same farm. Very infrequent mixing between isolates was observed and this could be attributed to the farms' controlled environments and correct application of biosecurity measures.
Only three isolates from Farm F Farm F were genetically highly related to isolates from other farms:ST10 isolates from Farm A and C and a ST877 from Farm E. ST877 isolates had less than 10 allelic differences (AD) and were collected from three animals from Farm E and a FC sample (chainsaw) from slaughterhouse S5 after the slaughtering of farm F pigs. According to the operators' reports, the farms' practice of sending pigs to several slaughterhouses may account for the closely related isolates, and the detection may have been caused by an isolate residing in the slaughterhouse. However, it is difficult to determine the reason This might rely on the presence of shared operators among farms or a possible movement of animals or items between farms. In this regard, more investigation is required.
We found E. coli in 36.2% of the environmental samples, indicating possible areas of contamination in the slaughterhouse settings. E. coli was found on non-food contact surfaces (such as drainage areas and walls) and, frequently, on food contact surfaces (such as cutting and bristle removal equipment), underscoring the significance of stringent hygiene procedures across the slaughterhouse.
Some isolates from environmental and pig samples showed strong genetic similarities, suggesting potential cross-contamination or transmission within the slaughterhouse environment. Two ST746 isolates were found in a dehairing whip and a pig's lymph nodes during the slaughter of animals from farm E, indicating that equipment may act as a vehicle for E. coli spread. Two ST1114 isolates from slaughterhouse S3 were detected in the drainage channel and scalding water after slaughtering pigs from farm D. The contamination source could be pigs from farm D, which contaminated the environment. One ST877 isolate was found on a chainsaw surface in slaughterhouse S5 after pigs from farm F were slaughtered. This environmental isolate was closely related (AD < 1) to ST877 E. coli from pigs of Farm E. In this case too, the most likely cause in this instance is direct or indirect transmission from a strain present in the slaughterhouse environment to animals and equipment. These results underscore the role pigs play as possible sources of E. coli contamination in slaughterhouses, though further research is necessary to fully understand these dynamics.
More than half (52.1%) of the E. coli isolates carried at least one AMR gene and most (82%) of them carried two or more AMR genes. This high incidence suggests that AMR is significantly present in the populations that were examined. The identification of several resistance genes in a single isolate raises the possibility that these bacteria will survive and multiply in settings where several antibiotics are administered, contributing to the spread of multidrug-resistant bacteria [42]. Furthermore, our results show that the incidence of AMR genes varies between environmental and pig samples. At least one resistance gene was found in 48% of pig samples and 69% of environmental samples from slaughterhouses, suggesting that environmental contamination with resistant bacteria might be an important source of AMR spread.
Slaughterhouses represent a critical point where microorganisms from many animals and sources may converge. During the slaughtering and meat processing stages, microorganisms from one animal can spread to others via surfaces, tools, handlers, or through direct contact with meat. This is particularly concerning when resistant bacteria are present because they can easily spread throughout the facility, contaminating carcasses and products [43].
We could detect at least one AMR gene in the majority (83%) E. coli isolates from farm samples confirming that the farm environment can be a source of resistant bacteria that could lead to the (re)colonization of pigs [44]. Thus, as was previously mentioned, biosecurity measures, such as proper cleaning and sanitization between animal batches, allow for the control of the spread of harmful and resistant strains [24]. Our findings serve as a reminder that a "one-health" strategy that addresses both the animals and their environment is necessary to reduce AMR.
A critical aspect of the study is the observation of discrepancies in AMR gene detection across farms, particularly in cases where farms reported no antibiotic use. Farm B, for instance, had E. coli isolates with resistance genes for many antibiotic families, such as tetracyclines, aminoglycosides, and β-lactams, although declaring no usage of antibiotics. In Farm C, where the use of specific antibiotics like amoxicillin-clavulanic acid and oxytetracycline was reported, resistance genes were documented for these antibiotics and also for others not reported by the farm. This finding aligns with other studies showing that E. coli isolates, regardless of antibiotic exposure, may possess high baseline levels of AMR genes [45,46]. This suggests that resistance genes can persist even without direct antibiotic pressure, likely due to historical antibiotic use [47]. The persistence of AMR genes is concerning as it complicates the mitigation of resistance, showing that once established in bacterial populations, resistance can remain stable without selective pressure. Traditionally, the concept of antibiotic limitation assumes that in antibiotic-free environments, vulnerable bacteria outcompete resistant ones, as resistance mechanisms often come with fitness costs, such as maintaining resistance plasmids or expressing drug efflux pumps [48]. However, these fitness costs are highly variable and dependent on the environment and genetic background [49]. Moreover, compensatory mutations can arise, reducing the fitness cost of resistance and allowing AMR genes to persist in bacterial populations [50].
Moreover, the transmission and maintenance of AMR genes on farms may be influenced by factors other than the direct use of antibiotics. These include contamination from outside sources (such people, animals, or objects), improper use of the all-in-all-out and cleaning systems, and horizontal gene transfer between pathogenic or commensal bacteria [47]. This highlights the complexity of AMR dynamics in farm environments. A particularly noteworthy finding in this context was the detection of an isolate from environmental sampling at Farm D carrying the mcr-10.1 gene, which confers resistance to colistin. Colistin is classified by the World Health Organization (WHO) as a "highest priority" critically important antimicrobial due to its role as a last-resort treatment for severe infections caused by gram-negative bacteria in humans [51]. The Mobile colistin resistance (mcr) genes are particularly concerning because they are often carried on plasmids, which facilitates their spread through horizontal gene transfer [52]. The isolate identified in the current study was ST746, a genotype frequently associated with multiple resistances and previously detected in humans, livestock, and wastewater [53,54,55]. Interestingly, this isolate lacked additional AMR genes even though it had various plasmid types (ColpVC, IncFIB, IncFII, IncX, IncY, data not shown).
A similar case was reported in an Italian pig farm, where a single isolate carrying the mcr-9 gene was found, despite the absence of colistin use [56]. The Authors suggested that the isolate likely originated from external sources, such as contaminated well water, people, or animals. A similar scenario could explain the presence of mcr-10.1 in our study, but further investigation is required to confirm this hypothesis. These findings support the essentially useful application of molecular methods to find genes that might not be expressed in the carrier isolate but that could be transferred to other bacteria.
The most prevalent AMR genes were those that confer resistance to tetracyclines (e.g., tetA, tetB, tetM) and β-lactams (e.g., blaTEM-1A, blaTEM-1B). Tetracycline resistance was prevalent (37.5%), which is consistent with its extensive usage, especially in pig husbandry [57]. The wide detection of blaTEM genes, particularly blaTEM-1B, is also indicative of the ongoing selection pressure imposed by the widespread use of β-lactam antibiotics, which are commonly used to treat infections in both people and animals [58]. Overall, the AMR pattern found in the present survey is consistent with what is reported in E. coli isolates in Italy [59] and is also consistent with the European report on AMR in zoonotic and indicator bacteria, which identified resistance to ampicillin, tetracycline, sulfamethoxazole, and trimethoprim as the most prevalent resistances in indicator E. coli in member States and in all animal categories in 2021/2022 [60]. The absence of carbapenemase-, AmpC β-lactamases-, and extended-spectrum β-lactamases-producing E. coli isolates in our study is an encouraging outcome when taking into account the public health significance.
In the present investigation, AMR genes were observed in a lesser percentage of pathogenic E. coli isolates than in the entire E. coli population. This result differs from the hypothesis that pathogenic strains, which frequently encounter antibiotic pressure during infections, would exhibit higher levels of resistance. However, the possibility that pathogens like ExPEC and ETEC might produce infections that are challenging to cure is still raised by the presence of AMR genes in these strains.
Phenotypic resistance was observed in 51% of the E. coli isolates in our study, with tetracycline (70%) and ampicillin (58%) being the most common. The study identified 23 phenotypic resistance patterns, with tetracycline resistance alone or in combination with other antibiotics, like ampicillin, being the most frequent. These patterns align with genotypic data. Interestingly, only ExPEC isolates among pathogenic E. coli showed phenotypic resistance, particularly to tetracycline and ampicillin. This suggests that while pathogenic strains may carry fewer AMR genes overall, those with resistance genes express significant resistance, complicating treatment options [61].
The genotypic results and the phenotypic resistance were highly consistent in our study. Nevertheless, phenotypic resistance was not present in all isolates having resistance genes, and vice versa. This result may be dependent on the absence of resistance gene expression because of weak or absent promoters and low copy numbers, or, conversely, on the existence of various resistance mechanisms that are not associated with a particular gene or antimicrobial agent, like efflux pumps [62,63]. In order to properly understand and handle the spread of resistance, the results highlight the necessity of thorough AMR monitoring programs that include both genotypic and phenotypic data.
5. Conclusions
Our results demonstrate that fattening pigs can act as carriers and shedders of globally distributed pathogenic E. coli isolates including ExPEC ST88 and ST101, UPEC ST648, ETEC ST410, and E. coli ST10 also in a closed environment such as the island of Sardinia. This highlights the crucial role of biosecurity measures at the farm level in preventing the spread of pathogens along the supply chain. Additionally, numerous non-pathogenic E. coli isolates carrying virulence genes were observed, indicating that non-pathogenic/commensal E. coli can serve as a reservoir for different virulence gene and possibly transmit them to other bacteria.
The present study found high genetic relatedness between bacterial isolates from both pig and environmental samples, helping trace contamination sources in the slaughterhouse. In this regard, hygiene measures like GMP and GHP on tools and surfaces during processing are essential in preventing contamination that could lead to the spread of pathogens to carcasses and meat.
We could also detect a high AMR in E. coli isolates recovered from farms, fattening pigs and slaughterhouses, reflecting the national and European situation and suggesting that AMR is influenced not only by antibiotic use but also by historical practices, environmental contamination, and bacterial adaptation. The study revealed that non-pathogenic E. coli isolates, even without direct antimicrobial exposure, harbored high levels of resistance genes, posing a public health risk by acting as reservoirs for resistance genes.
These findings emphasize the need to include non-pathogenic isolates in AMR monitoring programs and highlight the importance of ongoing surveillance in animals and production chains to effectively fight the spread of AMR.
Author Contributions
C.S., F.P. and E.P.L.D.S. conceived, validated, and supervised the research; methodology was created by C.S., F.P., G.S. and P. G.; G.S., F.P., M.P.M. and P.G. carried out the formal analysis; G.S., F.P., P.G., M.F.-A., M.P.M. and M.M. contributed to writing, reviewing and editing the manuscript; funding acquisition was carried out by C.S. and E.P.L.D.S. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Sequencing datawere uploaded to NCBI under BioProject PRJNA1171362.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Gordon, D.M. The ecology of Escherichia coli. In Escherichia coli: Pathotypes and Principles of Pathogenesis, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 3–20. ISBN 9780123970480. [Google Scholar]
- Müller, D.; Greune, L.; Heusipp, G.; Karch, H.; Fruth, A.; Tschäpe, H.; Schmidt, M.A. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl. Environ. Microbiol. 2007, 73, 3380–3390. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M.; Fratamico, P.M.; Manning, S.D.; Funk, J.A. Shiga toxin-producing Escherichia coli in swine: the public health perspective. Anim. Health Res. Rev. 2014, 15, 63–75. [Google Scholar] [CrossRef]
- World Health Organization. Prioritization of pathogens to guide discovery, research and development of new antibiotics for drug-resistant bacterial infections, including tuberculosis; World Health Organization: Geneva, 2017. Available online: https://www.who.int/publications/i/item/WHO-EMP-IAU-2017.12.
- Warmate, D.; Onarinde, B.A. Food safety incidents in the red meat industry: A review of foodborne disease outbreaks linked to the consumption of red meat and its products, 1991 to 2021. Int. J. Food Microbiol. 2023, 398, 110240. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Lizarazo, C.M.; Sierra-Cadavid, A. Prevalence of Escherichia coli generic and pathogenic in pork meat: systematic review and meta-analysis. World J. Microbiol. Biotechnol. 2023, 39, 247. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, T.L.; Blom, M.; Monnet, D.L.; Frimodt-Møller, N.; Poulsen, R.L.; Espersen, F. Transient intestinal carriage after ingestion of antibiotic-resistant Enterococcus faecium from chicken and pork. N. Engl. J. Med. 2001, 345, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards. Scientific Opinion on the public health risks of bacterial isolates producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 2011, 9, 2322. [Google Scholar] [CrossRef]
- ISO 17604; Microbiology of the Food Chain. Carcass sampling for microbiological analysis. The International Organization for Standardization: Geneva, Switzerland, 2015.
- Siddi, G.; Piras, F.; Gymoese, P.; Torpdahl, M.; Meloni, M.P.; Cuccu, M.; Migoni, M.; Cabras, D.; Fredriksson-Ahomaa, M.; De Santis, E.P.L.; et al. Pathogenic profile and antimicrobial resistance of Escherichia coli, Escherichia marmotae, and Escherichia ruysiae detected from hunted wild boars in Sardinia (Italy). Int. J. Food Microbiol. 2024, 421, 110790. [Google Scholar] [CrossRef]
- Enterobase. Available online: https://enterobase.warwick.ac.uk (accessed on 14 October 2024).
- SerotypeFinder database. Available online: https://journals.asm.org/doi/pdf/10.1128/jcm.00008-15 (accessed on 14 October 2024).
- KMA mapping tool. Available online: https://doi.org/10.1186/s12859-018-2336-6 (accessed on 16 October 2024). [CrossRef]
- AMRFinder tool. Available online: https://github.com/ncbi/amr (accessed on 16 October 2024).
- ResFinder tool. Available online: https://cge.food.dtu.dk/services/ResFinder/ (accessed on 16 October 2024).
- PlasmidFinder tool. Available online: https://cge.cbs.dtu.dk/services/PlasmidFinder/ (accessed on 16 October 2024).
- VirulenceFinder tool. 2024. Available online: https://cge.food.dtu.dk/services/VirulenceFinder/ (accessed on 16 October 2024).
- EUCAST. Breakpoint tables for interpretation of MICs and zone diameters 2024; Version 14.0. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 16 October 2024).
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; Van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Kro, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachic, E.M.; Sato, M.I.Z.; Gomes, T.A.T.; Amaral, L.A.; Ottoboni, L.M.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Amass, S.F. Biosecurity: stopping the bugs from getting in. Pig J. 2005, 55, 104–114. [Google Scholar]
- Stark, K.D.; Wingstrand, A.; Dahl, J.; Mogelmose, V.; Lo Fo Wong, D.M. Differences and similarities among experts’ opinions on Salmonella enterica dynamics in swine pre-harvest. Prev. Vet. Med. 2002, 53, 7–20. [Google Scholar] [CrossRef]
- Savin, M.; Bierbaum, G.; Kreyenschmidt, J.; Schmithausen, R.M.; Sib, E.; Schmoger, S.; Käsbohrer, A.; Hammerl, J.A. Clinically relevant Escherichia coli isolates from process waters and wastewater of poultry and pig slaughterhouses in Germany. Microorganisms 2021, 9, 698. [Google Scholar] [CrossRef]
- Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Nishiyama, M.Y., Jr.; et al. Virulence potential of a multidrug-resistant Escherichia coli strain belonging to the emerging clonal group ST101-B1 isolated from bloodstream infection. Microorganisms 2020, 8, 827. [Google Scholar] [CrossRef]
- Vanstokstraeten, R.; Crombé, F.; Piérard, D.; Castillo Moral, A.; Wybo, I.; De Geyter, D.; Janssen, T.; Caljon, B.; Demuyser, T. Molecular characterization of extraintestinal and diarrheagenic Escherichia coli blood isolates. Virulence 2022, 13, 2032–2041. [Google Scholar] [CrossRef]
- Ewbank, A.C.; Fuentes-Castillo, D.; Sacristán, C.; Esposito, F.; Fuga, B.; Cardoso, B.; Godoy, S.N.; Zamana, R.R.; Gattamorta, M.A.; Catão-Dias, J.L.; et al. World Health Organization Critical Priority Escherichia coli clone ST648 in magnificent frigatebird (Fregata magnificens) of an uninhabited insular environment. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Kocsis, B.; Gulyás, D.; Szabó, D. Emergence and dissemination of extraintestinal pathogenic high-risk international clones of Escherichia coli. Life 2022, 12, 2077. [Google Scholar] [CrossRef]
- Manges, A.R.; Smith, S.P.; Lau, B.J.; Nuval, C.J.; Eisenberg, J.N.; Dietrich, P.S.; Riley, L.W. Retail meat consumption and the acquisition of antimicrobial resistant Escherichia coli causing urinary tract infections: A case-control study. Foodborne Pathog. Dis. 2007, 4, 419–431. [Google Scholar] [CrossRef]
- Jakobsen, L.; Spangholm, D.J.; Pedersen, K.; Jensen, L.B.; Emborg, H.; Agersø, Y.; Aarestrup, F.M.; Hammerum, A.M.; Frimodt-Møller, N. Broiler chickens, broiler chicken meat, pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients. Int. J. Food Microbiol. 2010, 142, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R.; Johnson, J.R. Food-borne origins of Escherichia coli causing extraintestinal infections. Clin. Infect. Dis. 2012, 55, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Goor, A.V.; Mellata, M. Evaluation of Escherichia coli isolates from healthy chickens to determine their potential risk to poultry and human health. PLOS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef]
- Nordstrom, L.; Liu, C.M.; Price, L.B. Foodborne urinary tract infections: A new paradigm for antimicrobial-resistant foodborne illness. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Ba, X.; Guo, Y.; Moran, R.A.; Doughty, E.L.; Liu, B.; Yao, L.; Li, J.; He, N.; Shen, S.; Li, Y.; et al. Global emergence of a hypervirulent carbapenem-resistant Escherichia coli ST410 clone. Nat. Commun. 2024, 15, 1–15. [Google Scholar] [CrossRef]
- Frydendahl, K. Prevalence of serogroups and virulence genes in Escherichia coli associated with postweaning diarrhoea and edema disease in pigs and a comparison of diagnostic approaches. Vet. Microbiol. 2002, 85, 169–182. [Google Scholar] [CrossRef]
- Shepard, S.M.; Danzeisen, J.L.; Isaacson, R.E.; Seemann, T.; Achtman, M.; Johnson, T.J. Genome sequences and phylogenetic analysis of K88- and F18-positive porcine enterotoxigenic Escherichia coli. J. Bacteriol. 2011, 194, 395–405. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef]
- Sonntag, A.; Bielaszewska, M.; Mellmann, A.; Dierksen, N.; Schierack, P.; Wieler, L.H.; Schmidt, M.A.; Karch, H. Shiga toxin 2e-producing Escherichia coli isolates from humans and pigs differ in their virulence profiles and interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2005, 71, 8855–8863. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel; Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Scientific Opinion on the pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. EFSA J. 2020, 18, 5967. [CrossRef]
- WHO. Antimicrobial resistance: global report on surveillance. 2014. Available online: https://www.who.int/publications/i/item/WHO-HSE-PED-AIP-2014.2 (accessed on 14 October 2024).
- Ghidini, S.; Luca, S.D.; Rodríguez-López, P.; Simon, A.C.; Liuzzo, G.; Poli, L.; Ianieri, A.; Zanardi, E. Microbial contamination, antimicrobial resistance and biofilm formation of bacteria isolated from a high-throughput pig abattoir. Ital. J. Food Saf. 2022, 11. [Google Scholar] [CrossRef]
- De Lucia, A.; Card, R.; Duggett, N.; Smith, R.; Davies, R.; Cawthraw, S.; Anjum, M.; Rambaldi, M.; Ostanello, F.; Martelli, F. Reduction in antimicrobial resistance prevalence in Escherichia coli from a pig farm following withdrawal of group antimicrobial treatment. Vet. Microb. 2021, 258, 109125. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.; Muwonge, A.; Hutchings, M.R.; Mainda, G.; Bronsvoort, B.M.; Gally, D.L.; Corbishley, A. Resistance to change: AMR gene dynamics on a commercial pig farm with high antimicrobial usage. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.P.; May, H.E.; AbuOun, M.; Stubberfield, E.; Gilson, D.; Chau, K.K.; Crook, D.W.; Shaw, L.P.; Read, D.S.; Stoesser, N.; et al. A longitudinal study reveals persistence of antimicrobial resistance on livestock farms is not due to antimicrobial usage alone. Front. Microbiol. 2023, 14, 1070340. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef]
- Hinz, A.; Amado, A.; Kassen, R.; Bank, C.; Wong, A. Unpredictability of the fitness effects of antimicrobial resistance mutations across environments in Escherichia coli. Mol. Biol. Evol. 2024, 41. [Google Scholar] [CrossRef]
- Martinez, J.L.; Baquero, F. Mutation frequencies and antibiotic resistance. Antimicrob. Agents Chemother. 2000, 44, 1771–1777. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 14 October 2019; Available online: https://www.who.int/publications/i/item/9789241515528 (accessed on 14 October 2024).
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Wu, L.; Chen, J.; Wang, L.; Wu, Z. Whole genome sequence of an MCR-1-carrying, extended-spectrum β-lactamase (ESBL)-producing Escherichia coli ST746 isolate recovered from a community-acquired urinary tract infection. J. Glob. Antimicrob. Resist. 2018, 13, 171–173. [Google Scholar] [CrossRef]
- Shin, H.; Kim, Y.; Han, D.; Hur, H. Emergence of high-level carbapenem and extensively drug-resistant Escherichia coli ST746 producing NDM-5 in influent of wastewater treatment plant, Seoul, South Korea. Front. Microbiol. 2021, 12, 645411. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, Z.; Wang, Y.; Chen, Y.; Sun, J.; Yang, Y.; Si, H. Antimicrobial resistance and transconjugants characteristics of sul3 positive Escherichia coli isolated from animals in Nanning, Guangxi Province. Animals 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Guarneri, F.; Bertasio, C.; Romeo, C.; Formenti, N.; Scali, F.; Parisio, G.; Canziani, S.; Boifava, C.; Guadagno, F.; Boniotti, M.B.; et al. First detection of mcr-9 in a multidrug-resistant Escherichia coli of animal origin in Italy is not related to colistin usage on a pig farm. Antibiotics 2023, 12, 689. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Koutsoumanis, K.; Allende, A.; Alvarez-Ordoñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Scientific Opinion on the maximum levels of cross-contamination for 24 antimicrobial active substances in non-target feed. Part 12: Tetracyclines: tetracycline, chlortetracycline, oxytetracycline, and doxycycline. EFSA Journal 2021, 19, 6864. [CrossRef]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Koutsoumanis, K.; Allende, A.; Alvarez-Ordoñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Scientific Opinion on the role played by the environment in the emergence and spread of antimicrobial resistance (AMR) through the food chain. EFSA Journal 2021, 19, 6651. [CrossRef]
- Bassi, P.; Bosco, C.; Bonilauri, P.; Luppi, A.; Fontana, M.C.; Fiorentini, L.; Rugna, G. Antimicrobial Resistance and Virulence Factors Assessment in Escherichia coli Isolated from Swine in Italy from 2017 to 2021. Pathogens 2022, 12, 112. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) & ECDC (European Centre for Disease Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2021–2022. EFSA Journal 2024, 22, e8583. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nature Medicine 2004, 10(12 Suppl), S122–S129. [Google Scholar] [CrossRef]
- Toprak, E.; Veres, A.; Michel, J.; Chait, R.; Hartl, D.L.; Kishony, R. Evolutionary paths to antibiotic resistance under dynamically sustained drug selection. Nature Genetics 2011, 44, 101–105. [Google Scholar] [CrossRef]
- Moran, R.A.; Anantham, S.; Holt, K.E.; Hall, R.M. Prediction of antibiotic resistance from antibiotic resistance genes detected in antibiotic-resistant commensal Escherichia coli using PCR or WGS. Journal of Antimicrobial Chemotherapy 2017, 72, 700–704. [Google Scholar] [CrossRef]
Figure 1.
Presence of E. coli in different samples of pigs at slaughter. LN: mesenteric lymph nodes; CC: colon content; CS: carcass surface.
Figure 1.
Presence of E. coli in different samples of pigs at slaughter. LN: mesenteric lymph nodes; CC: colon content; CS: carcass surface.
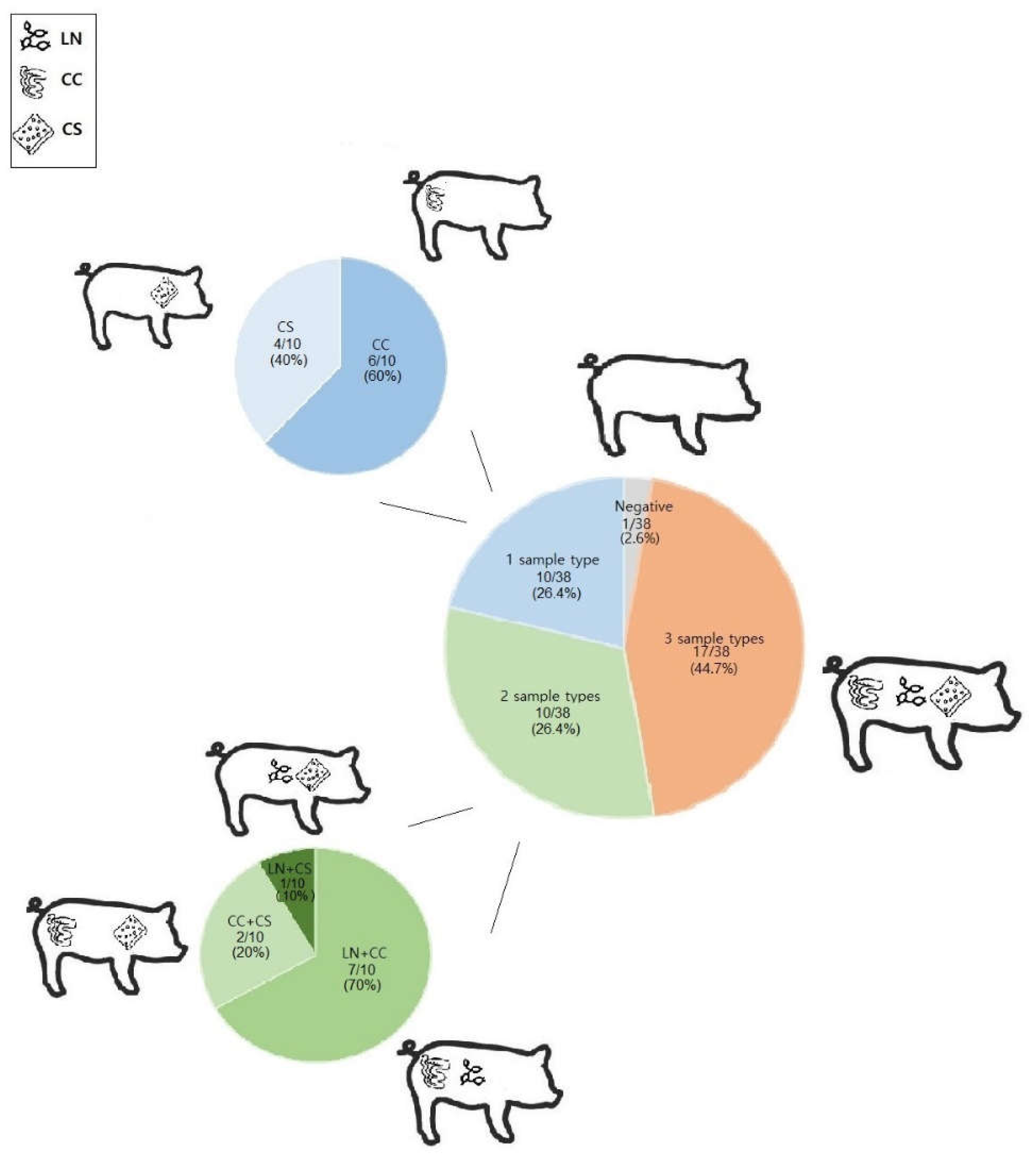
Figure 2.
Core genome Multi-locus sequence type (cgMLST) single linkage clustering tree showing genetic relatedness, sequence types (ST), phylogenetic groups and pathotypes of E. coli isolates. Branch length is in the number of allelic differences (AD) and the tree is shown with a branch length cut off at 200 AD.
Figure 2.
Core genome Multi-locus sequence type (cgMLST) single linkage clustering tree showing genetic relatedness, sequence types (ST), phylogenetic groups and pathotypes of E. coli isolates. Branch length is in the number of allelic differences (AD) and the tree is shown with a branch length cut off at 200 AD.
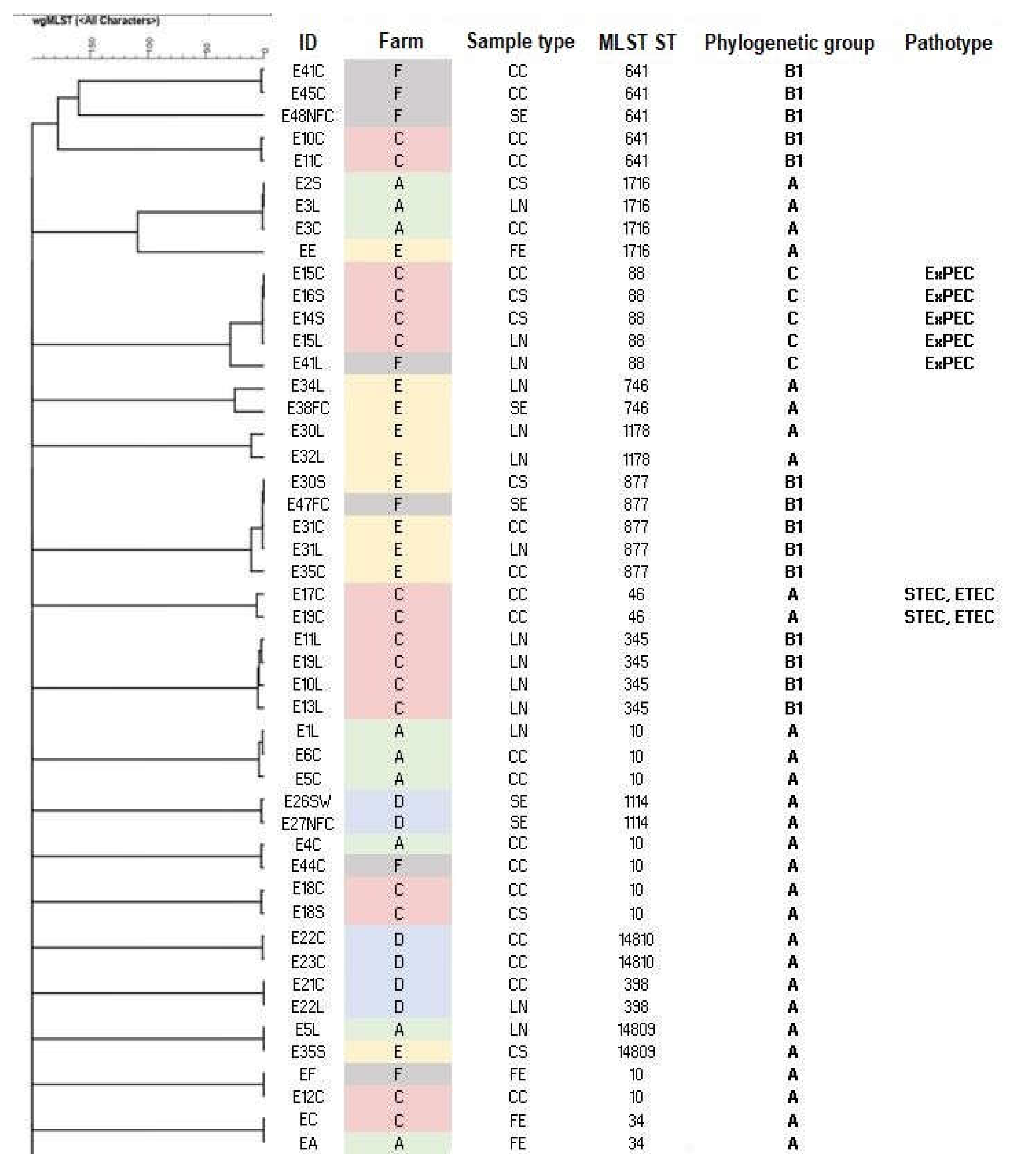
Figure 3.
Phenotypic resistance profile and AMR genes of E. coli isolates divided by farm.


Table 1.
Management characteristics of the farms.
| Farm | Herd size (n) | Fattening pigs (n) | Fattening period (days) | Floor of the fattening pen | Cleaning of the fattening pen | Feed | Water | Pest control | Antibiotic compounds used |
|---|---|---|---|---|---|---|---|---|---|
| A | 300 | 220 | Approx. 150 | S | Daily, AFAE | CP | Well | Rodents | Aug, Sxt |
| B | 180 | 130 | Approx. 180 | NS, external paddock | Daily | CP + whey | Mains | Rodents | - |
| C | 150 | 80 | Approx. 210 | S, external paddock | Twice a day | CP | Mains + well | Rodents | Aug, Ox |
| D | 150 | 80 | Approx. 270 | NS | AFAE | CP + whey | Well | Rodents | Aug |
| E | 5000 | 500 | Approx. 120 | NS | Daily, AFAE | CP | Well | Rodents | - |
| F | 1400 | 700 | Approx. 120 | S | Daily, AFAE | CP | Mains | Rodents | - |
S: slatted; NS: not slatted; AFAE: all in-all out; CP: commercial pellet; Aug: amoxicillin-clavulanic acid; Sxt: trimethoprim-sulfamethoxazole; Ox: oxytetracycline.
Table 2.
Presence of E. coli in mesenteric lymph nodes, colon content and carcass surface samples of fattening pigs; brackets indicate the number and type of pathogenic isolates.
Table 2.
Presence of E. coli in mesenteric lymph nodes, colon content and carcass surface samples of fattening pigs; brackets indicate the number and type of pathogenic isolates.
| Farm | Slaughterhouse | Number of tested pigs | Number of positive lymph nodes samples (pathogenic isolates) | Number of positive colon content samples (pathogenic isolates) | Number of positive carcass surface samples (pathogenic isolates) |
|---|---|---|---|---|---|
| A | S1 | 6 | 6 (0) | 6 (0) | 5 (0) |
| B | S1 | 3 | 2 (0) | 1 (0) | 0 |
| C | S2 | 10 | 9 (1 ExPEC) | 10 (2 ExPEC, 2 STEC-ETEC, 1 ETEC) | 6 (3 ExPEC) |
| D | S3 | 3 | 2 (0) | 3 (0) | 3 (0) |
| E | S4 | 10 | 4 (1 UPEC) | 5 (0) | 9 (0) |
| F | S5 | 6 | 2 (1 ExPEC) | 6 (2 ETEC) | 0 |
Table 3.
Characterization of E. coli isolates.
| ID | Sample type | Farm | Slaughterhouse | MLST ST | Serotype | Phylogenetic group | Predicted pathotype |
Virulence genes |
|---|---|---|---|---|---|---|---|---|
| EA | E | A | S1 | 34 | O101:H37 | A | - | acrF, astA, cea, emrE, fdeC, mdtM, terC, traT, ymgB |
| E1C | CC | A | S1 | 34 | O9:H10 | A | - | acrF, emrE, fdeC, mdtM, terC, ymgB |
| E1L | LN | A | S1 | 10 | O?:H9 | A | - | acrF, cea, fdeC, hra, iss, mdtM, terC |
| E1S | CS | A | S1 | 58 | O8:H30 | B1 | - | capU, cba, cia, cma, cvaC, etsC, fdeC, hlyF, iroBCDEN, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, sitA, terC, traT |
| E2C | CC | A | S1 | 542 | O?:H45 | A | - | acrF, fdeC, mdtM, pcoABCDERS, silABCEFPRs, terC |
| E2L | LN | A | S1 | 542 | O?:H45 | A | - | acrF, mdtM, pcoBCDER, silAF, terC, traT, ymgB |
| E2S | CS | A | S1 | 1716 | O130:H26 | A | - | acrF, emrE, fdeC, iss, mdtM, terC, traT, tsh, ymgB |
| E3C | CC | A | S1 | 1716 | O130:H26 | A | - | acrF, emrE, fdeC, iss, mdtM, terC, traT, tsh, ymgB |
| E3L | LN | A | S1 | 1716 | O130:H26 | A | - | acrF, emrE, fdeC, iss, mdtM, terC, traT, tsh, ymgB |
| E4C | CC | A | S1 | 10 | O111:H27 | A | - | acrF, astA, iss, mdtM, terC, traT, ymgB |
| E4L | LN | A | S1 | 48 | O99:H9 | A | - | espX1, fdeC, mdtM, terC, ymgB |
| E4S | CS | A | S1 | 48 | O26:H12 | A | - | acrF, astA, emrE, fdeC, fyuA, hra, irp2, mdtM, ompT, terC, traT, ybtP, ybtQ |
| E5C | CC | A | S1 | 10 | O101:H9 | A | - | acrF, fdeC, hra, iss, mdtM, terC, ymgB |
| E5L | LN | A | S1 | 14809 | O4:H45 | A | - | acrF, mdtM, terC, traT, tsh |
| E5S | CS | A | S1 | 5995 | O?:H27 | A | - | acrF, fdeC, mdtM, silABCEFPRS, terC, ymgB |
| E6C | CC | A | S1 | 10 | O89/O162/O101:H9 | A | - | acrF, cea, fdeC, hra, iss, mdtM, terC, ymgB |
| E6L | LN | A | S1 | 1716 | O126:H11 | A | - | acrF, fdeC, hra, iss, mdtM, pcoE, terC, ymgB |
| E6S | CS | A | S1 | 10 | O69:H32 | A | - | acrF, cba, cea, celb, cia, cma, fdeC, iss, katP, mdtM, ompT, pcoAER, silBCFRS, terC, ymgB |
| EB | A | B | S1 | 540 | O?:H30 | A | - | acrF, emrE, fyuA, hra, irp2, mdtM, terC, ybtP, ybtQ, ymgB |
| E7S | CS | B | S1 | 4442 | O54:H16 | B1 | - | acrF, cba, celb, cma, cvaC, ehxA, fdeC, iha, ireA, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, terBCDWZ, traT, tsh, ymgB |
| E8L | LN | B | S1 | 9598 | O168:H12 | A | - | acrF, astA, emrE, mdtM, pcoABCDERS, silABCEFPRS, terC, ymgB |
| E9C | CC | B | S1 | 744 | O101:H9 | A | - | acrF, astA, fdeC, hra, mdtM, merCPRT, silABCEFPRS, terC, traT, ymgB |
| EC | A | C | S2 | 34 | O?:H37 | A | - | acrF, astA, cea, emrE, fdeC, mdtM, terC, traT, ymgB |
| E10C | CC | C | S2 | 641 | O121:H10 | B1 | - | fedF, lpfA, sepA, terC, traT |
| E10L | LN | C | S2 | 345 | O8:H45 | B1 | - | acrF, fdeC, hlyA-alpha, iss, lpfA, mdtM, ompT, sepA, sitA, terC, traT, tsh, ymgB |
| E10S | CA | C | S2 | 58 | O25:H21 | B1 | - | acrF, afaA, afaB, astA, emrE, f17AG, fdeC, hra, iss, lpfA, mdtM, ompT, papC, terC, traT, tsh, ymgB |
| E11C | CC | C | S2 | 641 | O121:H10 | B1 | - | acrF, astA, f17AG, fdeC, fedF, lpfA, mdtM, sepA, terC, traT, ymgB |
| E11L | LN | C | S2 | 345 | O8:H45 | B1 | - | acrF, fdeC, hlyA-alpha, iss, lpfA, mdtM, ompT, sepA, sitA, terC, traT, tsh, ymgB |
| E12C | CC | C | S2 | 10 | O101:H9 | A | - | acrF, cea, emrE, espX1, fdeC, fyuA, irp2, iss, iucC, iutA, lpfA, mdtM, ompT, sitA, terC, traT, tsh, ymgB |
| E13C | CC | C | S2 | 847 | O?:H2 | B1 | ExPEC | acrF, cia, cma, cvaC, etsC, fdeC, hlyF, hra, iroN, iss, iucABCCD, iutA, lpfA, mchF, mdtM, ompT, papA_F13, papC, sitA, terC, traT, ybtP, ybtQ, ymgB |
| E13L | LN | C | S2 | 345 | O8:H45 | B1 | - | acrF, cvaC, emrE, fdeC, iroBCDEN, iss, iucABCD, lpfA, mchF, mdtM, ompT, papACEG-IIIH, sitA, terC, traT, tsh, ymgB |
| E13S | CS | C | S2 | 101 | O11:H10 | B1 | ExPEC | acrF, cba, cia, cma, cnf1, cvaC, emrE, etsC, fdeC, fyuA, hlyF, hra, iroN, irp2, iss, iucC, iutA, lpfA, mchF, mdtM, ompT, papA_F1651A, papC, sitA, terC, tsh, tsh, ymgB |
| E14C | CC | C | S2 | 10 | O101:H9 | A | - | acrF, aslA, csgA, cvaC, emrE, fdeC, fimH, gad, hlyA-alpha, hlyE, iroBCDEN, iucABCD, lpfA, mchF, mdtM, papCEFG-IIIH, terC, traJ, tsh, ybtPQ, yeh, ymgB |
| E14L | LN | C | S2 | 1277 | O?:H28 | A | - | aslA, csgA, espY, fimH, hlyE, terC, yeh |
| E14S | CS | C | S2 | 88 | O8:H9 | C | ExPEC | acrF, astA, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iroB, iroCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, papC, sitA, terC, traT, tsh, ybtP, ybtQ, ymgB |
| E15C | CC | C | S2 | 88 | O8:H9 | C | ExPEC | acrF, astA, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, papC, sitA, terC, traT, tsh, ybtPQ, ymgB |
| E15L | LN | C | S2 | 88 | O8:H9 | C | ExPEC | acrF, astA, cvaC, emrE, etsC, fdeC, fyuA, gad, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, papC, sitA, terC, traT, tsh, ybtPQ, ymgB |
| E16C | CC | C | S2 | 5771 | O7:H24 | D | ETEC | anr, aslA, astA, chuA, cia, csgA, eilA, eltlAB, espY, estb-STb1, fdeC, fimH, hha, hlyE, kpsEMII, lpfA, neuC, sitA, terC, traT, yeh |
| E16L | LN | C | S2 | 10 | O84:H21 | A | - | acrF, cea, emrE, gad, mdtM, ompT, terC, ymgB |
| E16S | CS | C | S2 | 88 | O8:H9 | C | ExPEC | acrF, astA, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, papC, sitA, terC, traT, tsh, ybtPQ, ymgB |
| E17C | CC | C | S2 | 46 | O9:H4 | A | STEC, ETEC | acrF, cba, cma, iss, mdtM, ompT, sepA, sta1, stb, stx2Ae, stx2e, terC,traT, ymgB |
| E17L | LN | C | S2 | 1147 | O128:H2 | B1 | - | fdeC, gad, lpfA, mdtM, terC, ymgB |
| E18C | CC | C | S2 | 10 | O?:H40? | A | - | acrF, cea, hra, mdtM, terC |
| E18L | LN | C | S2 | 1632 | O182:H38 | A | - | acrF, fdeC, hra, mdtM, terC, ymgB |
| E18S | CS | C | S2 | 10 | O?:H40? | A | - | acrF, cea, hra, mdtM, terC, ymgB |
| E19C | CC | C | S2 | 46 | O9:H4 | A | STEC, ETEC | acrF, cba, cma, iss, mdtM, ompT, sepA,s ta1,stb, stx2Ae, stx2e, terC,traT, ymgB |
| E19L | LN | C | S2 | 345 | O8:H45 | B1 | - | acrF, emrE, fdeC, iss, lpfA, mdtM, ompT, sitA, terC, traT, tsh, ymgB |
| E19S | CS | C | S2 | 58 | O165:H25 | B1 | - | acrF, cea, ehxA, fdeC, focCG, iroBCDEN, iss, lpfA, mchBCF, mcmA, mdtM, ompT, sfaDEF, terC, traT, ymgB |
| E20FC | FC | C | S2 | 711 | O120:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, terC, ymgB |
| ED | E | D | S3 | 746 | O21:H10 | A | - | acrF, arsADR, astA, fdeC, fyuA, irp2, iss, kpsEM_K11, mdtM, sitA, terC, ybtPQ, ymgB |
| E21C | CC | D | S3 | 398 | O8:H20 | A | - | acrF, astA, cma, cvaC, emrE, hra, iucABCD, iutA, mchF, mdtM, ompT, sitA, terC, traT, tsh, ymgB |
| E21L | LN | D | S3 | 542 | O179:O8:H45 | A | - | acrF, fdeC, hra, terC, ymgB |
| E21S | CS | D | S3 | 23 | O9:H32 | C | - | acrF, cvaC, etsC, fdeC, fyuA, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, sitA, terC, traT, tsh, ybtPQ, ymgB |
| E22C | CC | D | S3 | 14810 | O?:H45 | A | - | cia, cma, cvaC, etsC, fdeC, hlyF, iroBCDEN, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, sitA, terC, traT, ymgB |
| E22L | LN | D | S3 | 398 | O8:H20 | A | - | acrF, astA, cma, cvaC, emrE, hra, iucABCD, iutA, mchF, mdtM, ompT, sitA, terC, traT, tsh, ymgB |
| E22S | CS | D | S3 | 10 | O160:H4 | A | - | acrF, emrE, fdeC, fyuA, hra, irp2, mdtM, terC, ybtPQ, ymgB |
| E23C | CC | D | S3 | 14810 | O?:H45 | A | - | cia, cma, cvaC, etsC, fdeC, hlyF, iroBCDEN, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, sitA, terC, traT, ymgB |
| E24NFC | SE | D | S3 | 10 | O13:H11 | A | - | acrF, fdeC, fyuA, irp2, mdtM, ompT, terC, traT, tsh, ybtPQ, ymgB |
| E25FC | SE | D | S3 | 48 | O8:H18 | A | - | acrF, astA, fdeC, hra, mdtM, terC |
| E26SW | SE | D | S3 | 1114 | O117:H5 | A | - | acrF, fdeC, mdtM ,terC |
| E27NFC | SE | D | S3 | 1114 | O117:H5 | A | - | acrF, fdeC, gad, mdtM, terC |
| EE | FE | E | S4 | 1716 | O130:H26 | A | - | acrF, emrE, fdeC, iss, mdtM, terC, ymgB |
| E28S | CS | E | S4 | 1001 | O40:O8:H2 | B1 | - | acrF, cea, fdeC, lpfA, mdtM, terC, ymgB |
| E29S | CS | E | S4 | 542 | O184:H30 | A | - | acrF, astA, fdeC, mdtM, pcoABCDERS, silABCEDPRS, terC, ymgB |
| E30C | CC | E | S4 | 4156 | O113:H32 | A | - | acrF, fdeC, mdtM, merCPRT, terC, ymgB |
| E30L | LN | E | S4 | 1178 | O130:H26 | A | - | acrF, hra, mdtM, merCPRT, pcoABCDERS, silABEFPRS, terC, traT, tsh |
| E30S | CS | E | S4 | 877 | O?:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, ompT, pcoABCDERS, silABCEFPRS, terC, ymgB |
| E31C | CC | E | S4 | 877 | O?:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, ompT, pcoABCDERS, silABCEFPR, terC, ymgB |
| E31L | LN | E | S4 | 877 | O?:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, ompT, pcoABCDERS, silABCEFPRS, terC, ymgB |
| E31S | CS | E | S4 | 898 | O?:H48 | A | - | acrF, emrE, espX1, fdeC, lpfA, mdtM, terC, traT, ymgB |
| E32C | CC | E | S4 | 10 | O71:H27 | A | - | acrF, fdeC, fyuA, irp2, mdtM, ompT, pcoABCDERS, silABCEFPRS, silS, terC, ybtPQ, ymgB |
| E32L | LN | E | S4 | 1178 | O130:H26 | A | - | acrF, hra, mdtM, merCPRT, pcoABCDERS, silABEFPRS, terC, traT, tsh |
| E32S | CS | E | S4 | 542 | O8:H45 | A | - | acrF, fdeC, gad, hra, mdtM, terC, ymgB |
| E33L | LN | E | S4 | 648 | O2:H42 | F | UPEC | acrF, air, chuA, cia, eilA, emrDE, fdeC, iss, kpsEMII, lpfA, mdtM, ompT, terC, traT, yfcV, ymgB |
| E33S | CS | E | S4 | 165 | O180:H51 | A | - | aaiC, acrF, astA, cba, cea, emrE, katP, mcbA, mdtM, terC, traT, ymgB |
| E34L | LN | E | S4 | 746 | O?:H19 | A | - | acrF, astA, emrE, fdeC, mdtM, terC, traT, ymgB |
| E34S | CS | E | S4 | 2077 | O?:H8 | B1 | - | acrF, cea, emrE, fdeC, gad, iss, lpfA, mdtM, pcoABCDERS, pcoS, silABCEFPRS, terC, ymgB |
| E35C | CC | E | S4 | 877 | O?:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, ompT, pcoABCDERS, silABCEFPRS, terC, traT, ymgB |
| E35S | CS | E | S4 | 14809 | O4:H45 | A | - | acrF, mdtM, terC, traT, tsh |
| E37NFC | SE | E | S4 | 295 | O?:H16 | B1 | - | acrF, fdeC, lpfA, mdtM, terC, ymgB |
| E38FC | SE | E | S4 | 746 | O?:H19 | A | - | acrF, astA, emrE, fdeC, mdtM, terC, traT |
| E39FC | SE | E | S4 | 227 | O162:H10 | A | - | acrF, capU, emrE, etpD, fdeC, katP, mdtM, terC, traT, ymgB |
| E40NFC | SE | E | S4 | 88 | O?:H12 | C | - | acrF, cia, cib, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, merPRT, ompT, sitA, terC, traT, ybtPQ, ymgB |
| EF | FE | F | S5 | 10 | O101:H9 | A | - | acrF, cea, fdeC, fyuA, irp2, iss, iucABCD, iutA, mdtM, ompT, sitA, terC, traT, ybtPQ, ymgB |
| E41C | CC | F | S5 | 641 | O121:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, terC, ymgB |
| E41L | LN | F | S5 | 88 | O8:H9 | C | ExPEC | acrF, cea, cib, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchF, mdtM, ompT, papC, sitA, terC, traT, tsh, ybtPQ, ymgB |
| E42C | CC | F | S5 | 1115 | O102:H40 | A | - | acrF, cea, clpK, fdeC, hdeD-GI, hsp20, kefB-GI, mdtM, merCPRT, psi-GI, shsP, terC, trxLHR, yfdX1X2, ymgB |
| E42L | LN | F | S5 | 877 | O7:H10 | B1 | - | acrF, astA, fdeC, lpfA, mdtM, ompT, terC, ymgB |
| E43C | CC | F | S5 | 196 | O?:H7 | B1 | ETEC | acrF, astA, fdeC, lpfA, mdtM, stb, terC, ymgB |
| E44C | CC | F | S5 | 10 | O111:H27 | A | - | acrF, astA, iss, mdtM, terC, traT, ymgB |
| E45C | CC | F | S5 | 641 | O121:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, terC, ymgB |
| E46C | CC | F | S5 | 410 | O15:H12 | C | ETEC | acrF, astA, emrE, fdeC, iss, lpfA, mdtM, stb, terC, ymgB |
| E47FC | SE | F | S5 | 877 | O?:H10 | B1 | - | acrF, fdeC, lpfA, mdtM, ompT, pcoABCDERS, silABCEFPRS, terC, ymgB |
| E48NFC | SE | F | S5 | 641 | O121:H10 | B1 | - | acrF, fdeC, fedF, lpfA, mdtM, terC, traT, ymgB |
| E50FC | SE | F | S5 | 156 | O?:H28 | B1 | - | acrF, astA, cea, cvaC, emrE, etsC, fdeC, fyuA, hlyF, iha, iroBCDEN, irp2, iss, iucABCD, iutA, lpfA, mchBCF, mdtM, ompT, sitA, terC, traT, tsh, ybtPQ, ymgB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



