Submitted:
15 October 2024
Posted:
16 October 2024
You are already at the latest version
Abstract
Acrostichum aureum is a halophytic pantropical invasive fern growing in mangroves and swamps. Its association with arbuscular mycorrhizal fungi (AMF) has been reported in Asia. AMF and their symbiosis (AM) commonly colonise the absorption organs of terrestrial plants worldwide. Fur-thermore, AMF/AM are well known for their capacity to bioaccumulate toxic elements and to alleviate biotic and abiotic stress (e.g. salinity stress) in their hosts. However, how AMF are in-volved in the halophytism of A. aureum, and the structures where NaCl accumulates remains unknown. This study shows that A. aureum forms AM in Neotropical thermal margins of natural ponds, NaCl accumulation occurs inside AMF vesicles, and CaSO4 precipitates in colonised roots. Therefore, AM would be contributing to the halophytic nature of this fern, allowing it to thrive in saline and thermal environments by capturing NaCl from fern tissues, compartmentalizing it inside its vesicles and precipitating CaSO4.
Keywords:
thermal ponds
; halophytic fern
; arbuscular mycorrhizas
; Glomus
; Glomeromycota
; coastal environments
; mangroves
; Neotropics
; plant invasions
1. Introduction
Salt surfeit within a plant has detrimental effects on vital processes, such as photosynthesis, protein synthesis, and lipid metabolism. Plant responses to salt stress are greatly complex, involving anatomical, morphological, physiological, biochemical, and molecular mechanisms of salt tolerance. Salinity tolerance differs greatly among plants. Some plants are negatively affected by the presence of excessive minerals; other salt-tolerant plants can survive it or even thrive, such as halophytes [1,2,3,4,5]. These plant species grow at a concentration ≥ 200 mM of sodium chloride (NaCl), tolerating or even demanding increased concentrations from the water they absorb [2,6]. In terms of tolerance to and demand for sodium salts, halophytes are classified as obligate or true, and facultative [7,8].
Halophytism in pteridophytes is notably rare [9]; furthermore, ferns do not grow at sites where salinities exceed fifty per thousand and are absent in arid coastlines with a high soil salinity. The best known halophytic fern is the Acrostichum genus, named “the mangrove fern”, which has three species exceptionally tolerant to salt stress [10,11]: A. danaeifolium, A. speciosum and A. aureum. Their rhizomes and adventitious roots often grow in flooded soil, although they are never completely submersed. A. danaeifolium is restricted to the Neotropics and is the least salt tolerant [9,12]; A. speciosum, from the Paleotropics, is considered the only obligate halophytic fern species [11]; and the pantropical species A. aureum is halophytic [10,11]. A. danaeifolium and A. aureum are distributed in Colombia [13], where A. aureum proliferates in compacted, saline and arid alkaline soils [14].
Arbuscular mycorrhizal fungi (AMF) are ubiquitous fungal symbionts belonging to the phylum Glomeromycota [15]. AMF form an obligate mutualistic symbiosis, arbuscular mycorrhiza (AM), with the roots and absorption organs of ca. 80% species of both higher and lower land plants, including ferns [16,17]. AMF receive carbon compounds from the host; in exchange, they provide plants with limiting nutrients, such as nitrogen and phosphorus, and contribute to the host’s mineral and water access and protection against biotic and abiotic stresses, including pathogens, droughts, heat and salinity [18]. The exchange of nutrients between symbionts takes place through the cellular interface between plant and fungus in the colonised host roots. Soil AMF spores colonise roots through hyphae. Hyphae grow inter- and intracellularly in cortical host parenchyma; they branch, forming arbuscules, i.e. fine dichotomously branched hyphae in the host cell, the interchanging fungal structures where the bidirectional plant-AMF exchange occurs [19]. Hyphae also develop storage structures, the vesicles. Moreover, AMF hyphae inhabit the soil, extending beyond the rhizosphere depletion zone, where they capture, hold and transport nutrients and water to the host [18]. Furthermore, plants have different degrees of dependence on (or mycorrhizal status of their AM symbiosis), with ferns being obligate, facultative or non-mycorrhizal [20,21].
AMF are an important group of rhizosphere soil microorganisms spread nearly in every ecosystem worldwide, including saline environments [22,23,24,25,26,27,28]. Saline habitats comprise approximately 7% of the global land surface [29,30,31], harbouring similar plant communities. In these plant communities, zonal distribution of species depends on the soil salt concentration, which, together with biotic factors such as associated fungi, seems to be the factors driving plant competitiveness. In saline environments, plant communities include several families of angiosperms considered glomalean fungi colonised (GFC), non- or facultative mycorrhizal plants [17,18]. Furthermore, saline soils are often waterlogged, compacted and with increasing salt concentrations; these characteristics reduce AMF spore germination and, therefore, hyphal growth and root colonisation success [32,33]. Although high salt and water content in soils are unfavourable to AMF growth, there are records of AMF colonisation of halophytes [22,23,24,28]. While halophytes are considered to have a low or null ability to associate with AMF [17], some halophytes are AM, such as the halophytic fern Acrostichum aureum, which was reported to be colonised by AMF in mangroves of China and India [34,35].
It is well known that plants have several anatomo-morphological, biochemical and functional tools to sort out salt stress [1,7]. Furthermore, AMF-plant associations were proposed as one of the plant’s mechanisms for alleviating and mitigating salinity stress [6,36,37,38,39] because AMF improves plant tolerance to abiotic environmental stresses, including salinity. Saline environments seem to negatively affect AMF; however, positive effects of colonization on AM plants under salt stress conditions were reported, such as host growth and performance by improved host plant nutrition, higher K+/Na+ ratio in plant tissues and a better osmotic adjustment by accumulation of compatible solutes such as proline, glycine, betaine or soluble sugars and proteins [6,40,41,42,43]. In host plants under salt stress, AMF also improve the photosynthetic rate as well as water use efficiency and uptake [30,44], stimulate the activity of antioxidant enzymes against the reactive oxygen species generated by salinity, and regulate the expression of the plant genes involved in the maintenance of a better water status in plant tissues, such as the biosynthesis of proline, aquaporins and late embryogenesis abundant proteins with chaperone activity [45]. Furthermore, gene expression patterns suggest a lower salt stress damage in plants associated with AMF than in non-mycorrhizal plants [6,36,37,38,39,46]. The bulk of information on AMF effects and benefits regarding salinity is related to cultivated hosts, mainly in experimental systems, with reports from native environments and ferns being scarce.
Moreover, AMF were found to be involved in the host tolerance to salt compounds in saline conditions in field soils, greenhouse experiments and in vitro culture [46,47,48]. Thus, AMF improve plant tolerance through different mechanisms, including the differential capture and bioaccumulation of elements in the inner root cells colonised by intraradical mycelium and vesicles [48] and in extraradical hyphae and spores [47]. Furthermore, AMF colonisation has effects on the genetic expression of ion transporters in the roots of host plants [46]. However, to our knowledge, whether NaCl could be bioaccumulated in the AMF structures colonising the host roots in saline soil remains unknown.
The concept of sustainable development has been evolved since it was coined in 1980 to our days when it is based on three foundational pillars which are economic, social and environmental sustainability [49,50]. In particular, the biophysical characteristics of the environment [50] are considered the best indicators of environmental sustainability [50,51], and they are directly related to green production strategies in terrestrial environments and blue strategies in aquatic ecosystems in general, and especially, in marine systems [51]. Thus, the environmental component of sustainable development, represents an important challenge to maintain and improve the integral quality condition of the ecosystems and their resources that support the life in our planet, not only for open water ecosystems but in particular for coastal and wetlands areas [52]. Coastal ecosystems such as mangroves, salt marshes and wetlands provide benefits to human that include mainly food, fishing, recreational and intangible resources as main components of the economic and social development together with regulating ecosystems services like protection from storms, flooding and erosion, with a strong effect in the less developed, and island states [52]. Furthermore, coastal systems in those less developed countries and island states are also under higher human and environmental pressure than in the rich and developed counties by addition of strong pressures such as climate change, invasive species, and fish resources overexploitation. Thus, management and conservation of costal ecosystems and their resources in growing countries are included in the SDG (Sustainable Development Goals) 2030 Agenda [52].
Growth-promoting microorganisms such as AMF also are considered as tools for environmental sustainability [53,54,55]. Especially, AMF are capable of the improvement of their host nutrition, health, osmotic response against soil salts, and plant detoxification by bioaccumulation of heavy metals, all these functions also contribute to their involvement in the bioremediation of soils [53] and to the environmental sustainability by means of the coastal system restoration [54] and coastal wetlands conservation [55]. The Córdoba River basin and its wetland coastal zone is considered one of the most important water sources in the Magdalena Department of Colombia due to its waters are used in agriculture, fishing, tourism and consumption in Ciénaga town and surrounding area. In the Córdoba River basin, agricultural activities generate waste such as fertilizers and pesticides that are dumped into the basin, in addition to human and animal excrements, with negative affect on the water quality [56], and other agricultural effect is the soil salinization. The halophytic fern Acrostichum aureum L. and its mycorrhizal interactions are potential tools for bioremediation of these brackish wetland areas and its study can provide basic information to implement its future use in the bio-capture of salts, heavy metals and other pollutants generated by agriculture and of anthropogenic origin. In this coastal wetland, our aims were to determine if the mangrove fern A. aureum was associated to AMF and to analyse NaCl salt bioaccumulation in the AMF structures into the roots growing in saline soils in northern South America.
2. Materials and Methods
Acrostichum aureum L. (Pteridaceae, Polypodales) [57] is distributed in the Caribbean lowlands and in Pacific coastal areas in Colombia, within an elevational range of 0-950 m a.s.l. Samples were taken from El Volcán thermal pond margins (11°01´12.10´´N; 74°12´26.88´´W), where soil is marshy or flooded; the ponds are located 5 km away from the Córdoba River mouth in the Caribbean Sea, Magdalena Department, Colombia (Figure 1), in March, 2023. This area is characterized by riparian vegetation of Fabaceae (Prosopis juliflora, Caesalpinia coriaria) and Arecaceae (Attalea butyracea and Chamaedorea pinnatifrons).
Samples of underground rhizomes with adventitious roots were collected from sporophytes of seven A. aureum individuals growing on the margins of thermal ponds located in El Volcán (Figure 1), and stored in plastic bags that were refrigerated at 4°C until processing. In the laboratory, the fine rhizomes of each sporophyte were washed, cleared and stained [58]. The finest adventitious roots of the rhizomes of each sporophyte were cut into segments of approximately 1 cm and mounted on semipermanent slides with water-glycerol 1:3 v/v solution. Root colonization by AMF was quantified using optical microscope at 400 magnifications by McGonigle et al. method [59]. Root slides were observed under the microscope and the AMF structures colonising roots were microphotographed using a Zeiss AXIO LAB A1.
Micrographs of scanning electron microscopy (SEM) were obtained with the Zeiss LEO 1450VP microscope at the Laboratory of Electron Microscopy and Microanalysis (LABMEM) of the National University of San Luis (UNSL), Argentina. Furthermore, the chemical composition of samples was analysed by means of the energy-dispersive X-ray spectrometry (EDX) with the EDAX Genesis 2000 spectrometer along with SEM (SEM-EDX). Samples were mounted on double-sided carbon adhesive tape on aluminium stubs and carbon-coated and observed at 15 KeV. In addition, vesicles were detached from roots by microdissection under a binocular stereomicroscope (Nikon 7200) at a magnification of 500 X for their posterior observation with SEM and microanalysis by EDX.
The pH (Standard Methods Nro. 4500-H, 2012), salinity and conductivity (Standard Methods Nro. 2510-B, 2012) from water and sediments were analysed in the Laboratorio de Calidad Ambiental Marina (LABCAM) of the Instituto de Investigaciones Marinas y Costeras (INVEMAR-Santa Marta, Colombia). Furthermore, rhizosphere soil composition was analysed using SEM-EDX and micrographs for soil element composition.
3. Results
The roots and rhizomes of Acrostichum aureum sporophytes growing on the margins of the thermal ponds (Figure 2) were colonised by AMF, which exhibited the characteristic structures in the host roots (Figure 3). Furthermore, extraradical structures such as hyphae, spores, and intraradical arbuscules and vesicles of the AMF associated with A. aureum showed dense cytoplasmic contents (Figure 4). Salt crystals were also observed inside the vesicles (Figure 5a).
EDX analyses showed that the crystals present in the vesicles are composed mainly of NaCl (Figure 5b), with very low proportions of K, Ca and S. In addition, the amorphous crystals accumulated inside the roots were composed of CaSO4 (Figure 6a-c). Other elements such as K and Na accounted for 1% of the total elements around the crystals, whereas Cl and Mg were also detected as traces lower than 1% (Figure 6d).
Thermal water (Table 1) of ponds where A. aureum ferns grow was characterized by acid pH; nitrates; orthophosphates; high level of silicates, Zn, and Mn; high content of Na, K and Ca; and electrical conductivity of 3.08 mS/cm. Furthermore, the pond sediments (Table 2) were muddy and presented ammonia, organic matter (OM), Cu, Zn, Fe, Mn, and similar low concentrations of Na and K. In the rhizosphere soil around roots of A. aureum, the elemental composition EDX analysis showed C, O, Na, Cl, K, Ca, Mg, Fe, Al, Si, and S (Table 3).
All mature A. aureum sporophytes were colonized by AMF in their roots; the percentage of total root length colonised (% RL) was high, followed by the hyphal colonization (%HC), arbuscules (%AC) and the lowest value was for the vesicles (%VC) (Table 4).
4. Discussion
In the Colombian wetlands of the Córdoba river, the roots and rhizomes of Acrostichum aureum sporophytes growing on the margins of the thermal ponds were found colonised by AMF, which formed their characteristic structures in the host roots. In Asian mangroves, roots of A. aureum were reported as highly to lowly colonised and forming arbuscules and vesicles in Goa, India [27], and China [34,35], where A. aureum was colonised, with percentages of colonisation varying with the flooding level [35]. In contrast, A. aureum was considered non-colonised in other Indian mangroves [61,62]. As far as we know, this is the first report of AMF colonisation in roots and rhizomes of A. aureum in thermal plant communities associated with Colombian mangroves and thermal ponds in a Neotropical area. In this Neotropical wetland, the AMF colonization was high as well as the percentage of arbuscules, the nutrient interchange host-fungus structures [18,19], suggesting that the symbiotic interaction between A. aureum -AMF in the thermal ponds were functioning actively as mutualistic.
In this work, we found AMF colonisation not only in the roots of A. aureum, in agreement with this halophytic fern in Asian mangroves, but also in the fern rhizomes; we also found AMF colonisation in thermal ponds in the Colombian Neotropical area which is not a mangrove but was a wetland in the past. Moreover, it is amazing that the water temperature (47 °C) of these thermal ponds where A. aureum roots and rhizomes were colonised is the highest value ever detected for this fern, with previous reports indicating 29-30 °C [60]. Thus, the finding of A. aureum in Colombian thermal ponds and its mycorrhizal association extends its known distribution, exceeding the one in mangrove communities, and provides the record of the highest growth temperatures recorded so far for this species.
In addition, in the Volcán thermal ponds, extraradical structures such as the hyphae and spores, as well as intraradical arbuscules and vesicles of the AMF associated with A. aureum showed dense cytoplasmic contents and salt crystals, which were found within the vesicles. In previous reports of AMF colonisation and structures [27,34,35,60] these dense contents inside AMF structures were not reported for A. aureum. It is known that AMF can improve plant tolerance to salt stress through different mechanisms, one of them being the bioaccumulation of compatible solutes and elements [62] in the inner root cells colonised by intraradical mycelium and vesicles [48], and extraradical hyphae and spores [47]. As far as we know, this is the first report about dense cytoplasmic content in AMF associated with the halophytic fern A. aureum growing in the Neotropics and thermal ponds.
Besides, the EDX crystal analysis of vesicles showed they were composed of NaCl inside the cortical root cells of A. aureum growing in the thermal ponds, in the vesicles, using PIXE in combination with STIM, Olsson et al. [48] found that P was the most common element, followed by K and Ca, in roots of leak growing under greenhouse conditions. Furthermore, the authors found Zn, Si, Al, S, Mn, Cu, and Cl inside the vesicles. On the other hand, Hammer et al. [47] studied Acacia cyanophylla-AMF association in the field using bioassays under high salinity conditions; the authors found significantly high levels of Ca, Cl, Mg, Fe, Si and K, except for Na, which was present at a much lower concentration than its Cl counterion in extraradical hyphae and spores. Although in field samples, Na and Cl were present inside extraradical hyphae and spores, Hammer et al. [47] concluded that extraradical AMF hyphae and spores are selective in capturing these elements and that they are able to avoid Na incorporation to the plant when the ratios of K/Na and Ca/Na are considered in field and in vitro culture experimental approaches [47]. However, we also found NaCl crystals inside root vesicles of A. aureum with the same Na and Cl levels as those of field-sampled roots and rhizomes; indeed, this is the first report of NaCl inside vesicles of AMF colonising the roots of the halophytic fern A. aureum. Furthermore, the presence of NaCl salt crystals in the vesicles suggests that the role of the AMF colonising the roots of this halophyte fern would be that of bioaccumulating salt inside the fungus instead of avoiding Na and Cl accumulation, thereby providing the plant with another mechanism to avoid salt incorporation.
Moreover, Medina et al. [63] concluded that the distribution of A. aureum in three mangrove sites with different soil salinity “is not due to lack of salt resistance in the sporophyte”. In these mangrove areas in Puerto Rico, the Cl-/Na+ ratio in cell sap of A. aureum was twice the values of the other mangrove species. Furthermore, A. aureum has a ratio of 2.6 K+/Na+, whereas typical mangrove species have 0.5. According to Medina et al. [63], their results indicated the presence of ion selectivity probably of Na+ at root level. Furthermore, Na+ exclusion from roots and AMF structures was found to be a mechanism to avoid salinity in Acacia cyanophylla (a halophyte host of the desert of Turkey); this plant benefited from AMF, since it prevented Na+ capture while increasing K+ and Ca2+ capture [47]. In contrast to this mechanism, NaCl crystals present inside AMF vesicles of colonised A. aurem roots could be functioning as ion differential selectors in these saline environments, being a different mechanism to overcome environmental salinity in this halophytic fern. Therefore, this result may explain ion selectivity at the root level [63] inside the roots of this halophytic fern through AMF colonisation and NaCl bioaccumulation.
In Córdoba River thermal ponds, A. aureum presented CaSO4 crystal accumulation inside roots, which was mainly composed of CaSO4, in lower proportions by Na and K, and by trace elements such as Cl, and Mg. It is known that the Ca2+/Na+ ratio is an important parameter to measure salt stress in plants. Under salt stress conditions, Ca2+ absorption is interfered by the elevated rhizospheric Na+ concentration and Ca2+ is replaced at cell wall and plasma membrane level. Consequently, Ca2+ translocation is reduced, as well as Ca2+/Na+ ratio in plants under salt stresses. Furthermore, a low Ca2+/Na+ ratio causes a decrease in hydraulic conductivity and cell turgor, and interruptions in Ca2+ signalling. Although Mg2+ presence affects Ca2+ uptake, mycorrhizal plants accumulate more Mg2+ than non-mycorrhizal plants and show an advantageous Ca2+/Na+ ratio due to the Ca2+ uptake improvement under salt stress conditions [45]. Although the mechanism involved in these processes remains unknown, mycorrhizal colonisation was found to help the host plant overcome the deficiencies generated by Na+, Ca2+ and K+, maintaining beneficial K+/Na+, Ca2+/Na+, and Ca2+/Mg2+ ratios in host tissues [62].
Furthermore, Ca2+ works as the signal for root development and adaptation to optimize uptake of phosphate (Pi), an important plant growth limiting factor. However, Ca2+ has to be controlled due to its high toxicity to Pi metabolism [64]. Moreover, depending on the soil environment pH, Pi precipitates in the presence of Ca2+, Mg2+, and Zn2+ cations, becoming unavailable to plants [31,45], and generating salt-induced plant Pi deficiency [45]. Thus, inside A. aureum roots, Ca was retained as a CaSO4 salt, forming amorphous crystals that made Pi absorption possible for the plant. Moreover, Ca2+ formed salts with sulphates and contributed to the removal of SO4-- ions from root media, avoiding H2SO4 formation and the consequent acidification of the thermal water and rhizospheric soil where A. aureum inhabits. Therefore, CaSO4 amorphous crystals in A. aureum roots could have a dual alleviation function under saline stress: contributing to Pi absorption and balancing or buffering pH to the roots of this fern. Our results failed to explain whether the gypsum salt is formed by one of the partners or the A. aureum-AMF arbuscular mycorrhizal symbiosis or whether it is formed synergistically by the symbiosis. However, it is clear that CaSO4 presence inside A. aureum roots is a beneficial mechanism for this fern to thrive under saline stress.
Several studies suggest that sulphur-bioaccumulator plants (thiophores, gypsophiles and gypsovags) produce mainly foliar CaSO4 salts [65]. Instead, the presence of root CaSO4 was reported only for Acacia ancistrocarpa [66]. Gypsum salt bioaccumulation has been considered a biomineralogical tolerance mechanism against sulphate salinity and soluble calcium excess in plants capable of settling in abiotically stressed soils, such as deserts, coastal and riparian zones, where the presence of soil gypsum does not necessarily indicate that the plant is a bioaccumulator of this mineral [66,67]. Growth of gypsum-forming plants was also limited by Pi unavailability caused by the scarce soluble calcium-phosphate formation in the soils they inhabit. However, Robson et al. [65] stated that the capacity for gypsum biomineralization per se did not confer thiophores with tolerance to overcome Pi deficiency. In this study, surprisingly, gypsum crystals, which are usually stored in the leaves and shoots of most gypsophiles and thiophores, were detected inside A. aureum roots. In this mycorrhizal halophytic fern, these CaSO4 crystals may increase Pi availability and capture from saline soils, cooperating with the A. aureum-AMF association, which efficiently improves the host’s Pi nutrition [18]. Furthermore, as far as we know, this study is the first report of gypsum salts inside fern roots. Experimental studies are necessary to confirm whether A. aureum is a thiophore fern and to understand the process of amorphous crystal production and its role in osmoregulation mechanisms.
The results of this study show that the halophytic pioneer fern A. aureum, characteristic of the vegetation in pantropical mangroves and swamps, can also grow in thermal water from Neotropical Colombian springs. Furthermore, bioaccumulation of NaCl crystals was observed in AMF vesicles inside A. aureum roots, as well as amorphous CaSO4 crystals, suggesting that this species is a thiophore. Therefore, in this fern, mycorrhizal colonisation facilitates a wide range of mechanisms, either inherent to the species or provided by its AM symbionts; such mechanisms allow the plant to thrive in environments with high stress levels and extreme environmental conditions, such as saline and flooded soils and thermal waters, enabling availability of Pi in the rhizosphere, its capture and later translocation, consequently allowing A. aureum to be a “conqueror” fern. The capability of A. aureum-AMF association for the salts bioaccumulation in this coastal environments constitute an important attribute to the environmental sustainability [49,50,51,52] involvement of this fern in similar wetlands saline systems.
Besides, in mangroves, marshes, swamps and wetlands of Africa and Asia, A. aureum is widely used as an edible vegetal and this fern species also has various ethnobotanical uses in traditional medicine by its properties such as analgesic, antibacterial, antidiarrheal, anthelmintic, anti-inflammatory, antioxidant, and heling agent. This mangrove fern also produces numerous advantageous phytochemical compounds (e. g. kaempferol, di-(2-methylheptyl) phthalate, β-sitosterol, (2S,3S)-sulfated pterosin C, (+)-pinoresinol-4-O-sulfate, lupeol, α-amyrin and phytol); further, extracts of A. aureum have been used for the treatment of cancer, diabetes, ulcer, and viral diseases [68,69]. In the American continent, it has also been considered an edible plant widely consumed in Meso America since the times of the Maya people [70] and today [71]; in North America, it was used in Mexico as diuretic and expectorant [72] also, in Meso and South America [71,73]. In addition, A. aureum has been recorded in Colombia providing some different health consumables such as herbal medicine for constipation, fever, malaria, rheumatism, among others human diseases, and as an expectorant, fodder and bed for livestock, and ornamental use [71,72,73].
For more than 2,500 years, humans have used plants for medicinal purposes. Currently, they are used directly as a source of herbal medicine to cure diseases in most underdeveloped countries, particularly in tropical and subtropical areas of the planet; and indirectly, as providers of active compounds and chemical substances used in the pharmaceutical industry for the production of commercially available drugs [74]. Medicinal plants are threatened with extinction due to habitat loss and pollution. At the same time, the increase in their demand, the concomitant increase in their production, and the standardization required for their commercialization have contributed to a marked accrue in their intensive cultivation [75,76]. Likewise, the large-scale production of medicinal plants implied the detriment of their medicinal qualities such as the decrease in the contents of their active ingredients and their metabolic activity [77,78].
Symbiotic interactions between medicinal plants with endophyte root fungi such as AMF [78,79] have received special interest worldwide due to the benefits that these fungal symbionts provide to their hosts, improving their nutrition, increasing their productivity, resistance to pathogens and drought [18]. In particular, AMF and AM are essential for their cultivation, due to their primary role in increasing active compounds, medicinal secondary metabolites and the metabolic activity of the host, and consequently the increase in the pharmaceutical quality of mycorrhizal plants [80,81]. Furthermore, A. aureum has been shown a promising potential in phytoremediation of environmental contaminant like heavy metals by their removal from soil, waste water and effluents especially in marine wetlands together with its high degree of tolerance against arsenate toxicity [69], plus the bioaccumulation of soil and water salts NaCl found in this study though A. aureum-AMF colonisation. Thus, the association of A aureum –AMF in the tropical saline areas such as Córdoba River thermal ponds may suggest a synergistic potential of this fern to the economic and social pillars of the sustainability [49,50] in this coastal stressed environments where their sustainability is under huge threatened [52].
5. Conclusions
In this coastal and anthropic stressed environment of the Caribbean basin, AMF root colonisation in A. aureum could enhance phytoremediation by promoting the species growth as a hyperaccumulating, halophytic and salt tolerant plant capable of bioameliorating saline soils. Additional experimental studies are necessary to understand the mechanisms involved in salt bioaccumulation within AMF structures, NaCl and CaSO4 osmoregulation in A. aureum, as well as fern-AMF symbiosis and nutrition processes. This dominant fern species also would be useful as a green tool and piece to build the environmental sustainability of the tropical wetlands that it inhabits. Furthermore, the relevant properties of A. aureum as an edible plant, its numerous ethnobotanical uses and medicinal properties, and its capability for the production of pharmacological substances applicable to human health, would project this mycorrhizal-halophytic-bioaccumulating fern as an excellent renewable natural resource and could position it as a natural resource directly related to the other two pillars of sustainable development, that is economic and social sustainability. A. aureum has the potential to be a fundamental species in the sustainable development of coastal environments whose sustainability is seriously threatened, especially in developing countries such as those found in the Caribbean area of the Neotropics.
Author Contributions
M.A.N. and M.L. conceived and designed the project, carried out the experimental work and wrote most of the result discussion. M.L. was responsible for the AMF root colonisation detection and quantification, while M.L., M.A.N., E.C. and S.M. participated in the sample processing and chemical plant analysis. M.A.N. and M.L. initiated and promoted the writing of the manuscript, all the authors interpreted the results and wrote the manuscript, M. C. P. revised also critically the manuscript, and E.C. carried out the EDX analysis.
Funding and Acknowledgments
This project was carried out thanks to the financial support of the Research Vice-Rectory from the University of Magdalena to the Researchers M.A.N. and S.N., Secretary of Science and Technology of the University of San Luis for financial support of the PROICO 02-2023. Moreover, the participation of Dr. Mónica A. Lugo, a researcher at Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), was fundamental to this project.
Data Availability Statement
Data available on request.
Conflicts of Interest
The authors declare no competing interests.
References
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: a review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Coleman, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Coleman, T.D. Plant salt tolerance: adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Bhowmik, P.C.; Hossain, A.; Rahman, M.M.; Vara Prasad, M.N.; Ozturk, M.; Fujita, M. Potential use of halophytes to remediate saline soils. BioMed Res. Int. 2014, Volume 2014, Article ID 589341, 1–12. [CrossRef]
- Shabala, S. Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruíz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Aslam, R.; Bostan, N.; Nabgha-e-Amen; Maleeha, M.; Safdar, W. A critical review on halophytes: Salt tolerant plants. J. Medic. Plants Res. 2011, 5, 7108–7118. [Google Scholar] [CrossRef]
- Lloyd, R.M.; Buckley, D.P. Effects of salinity on gametophyte of Acrostichum aureum and A. danaefolium. Fern Gaz. 1986, 13, 97–102. [Google Scholar]
- Medina, E.; Francisco, A.M.; Wingfield, R.; Casañas, O.L. Halofitismo en plantas de la costa Caribe de Venezuela: halófitas y halotolerantes. Acta Bot. Venez. 2008, 31, 49–80. [Google Scholar]
- Mehltreter, K. Phenology and habitat specificity of tropical ferns. In Biology and Evolution of Ferns and Lycophytes; Ranker, T.A., Haufler, C.H., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 201–221. [Google Scholar] [CrossRef]
- Mehltreter, K.; Palacios-Ríos, M. Phenological studies of Acrostichum danaeifolium (Pteridaceae, Pteridophyta) at a mangrove site on the Gulf of Mexico. J. Trop. Ecol. 2003, 19, 155–162. [Google Scholar] [CrossRef]
- Murillo-A., J.; Orozco-O., N.; Rivera-Díaz, O. Los helechos y los licófitos del Caribe colombiano. In Colombia Diversidad Biótica XII. La Región Caribe de Colombia; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia, 2012; pp. 333–348. [Google Scholar]
- Blanco, J.F.; Estrada, E.A.; Ortiz, L.F.; Urrego, L.E. Ecosystem-wide impacts of deforestation in mangroves. In The Urabá Gulf (Colombian Caribbean) Case Study. Internat. Scholar. Res. Network ISRN Ecol. Article ID 958709, 2012, Volume 2012, 1–14. [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal symbiosis, 2nd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Harrison, M.J. Molecular and cellular aspects of the arbuscular mycorrhizal symbiosis. Annu. Rev. Plant Physiol. Plant. Mol. Biol. 1999, 50, 361–389. [Google Scholar] [CrossRef]
- Trappe, J.M. Phylogenetic and ecologic aspects of mycotrophy in the angiosperms from an evolutionary standpoint. In Ecophysiology of VA Mycorrhizal Plants; Safir, G.R., Ed.; CRC Press: Boca Raton, USA, 1987; pp. 5–25. [Google Scholar]
- Pressel, S.; Bidartondo, M.I.; Field, K.J.; Rimington, W.R.; Duckett, J.G. Pteridophyte fungal associations: current knowledge and future perspectives. J. Syst. Evol. 2016, 54, 666–678. [Google Scholar] [CrossRef]
- Kahn, A.G. The occurrence of mycorrhizas in halophytes, hydrophytes and xerophytes, and of Endogone spores in adjacent soils. J. Gen. Microbial. 1974, 81, 7–14. [Google Scholar] [CrossRef]
- Kahn, A.G. Effects of various soil environmental stresses on the occurrence, distribution and effectiveness of va mycorrhizae. Biotropica 1995, 8, 39–44. [Google Scholar] [CrossRef]
- Rozema, J.; Arp, W.; van Diggelen, J.; van Esbroek, M.; Broekman, R.; Punte, H. Occurrence and ecological significance of vesicular arbuscular mycorrhiza in the salt marsh environment. Acta Bot. Neerl. 1986, 35, 457–467. [Google Scholar] [CrossRef]
- Sengupta, A.; Chaudhuri, S. Atypical root endophytic fungi of mangrove plant community of Sundarban and their possible significance as mycorrhiza. J. Mycopathol. Res. 1994, 32, 29–39. [Google Scholar]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum, may be partly related to elevated Kþ/Naþ ratios in root and shoot tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef] [PubMed]
- D´Souza, J.; Rodrigues, B.F. Biodiversity of Arbuscular Mycorrhizal (AM) fungi in mangroves of Goa in West India. J. Forestry Res. 2013, 24, 515–523. [Google Scholar] [CrossRef]
- Becerra, A.G.; Cofré, M.N.; García, I. Mycorrhizal simbiosis in salt-tolerance species and halophytes growing in salt affected soils of South America. In Mycorrhizal Fungi in South America; Pagano, M.C., Lugo, M.A., Eds.; Springer: Cham, Switzerland, 2019; pp. 295–314. [Google Scholar]
- Ruiz-Lozano, J.M.; Azcón, R.; Gómez, M. Alleviation of salt stress by arbuscular-mycorrhizal Glomus species in Lactuca sativa plants. Physiol. Plant. 1996, 98, 767–772. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Symbiotic efficiency and infectivity of an autochthonous arbuscular mycorrhizal Glomus sp. from saline soils and G. deserticola under salinity. Mycorrhiza 2000, 10, 137–143. [Google Scholar] [CrossRef]
- Parihar, M.; Rakshit, A. Arbuscular mycorrhiza: a versatile component for alleviation of salt stress. Nat. Environ. Pollut. Technol. 2016, 15, 417–428. [Google Scholar]
- Juniper, S.; Abbott, L. Vesicular-arbuscular mycorrhizas and soil salinity. Mycorrhiza 1993, 4, 45–57. [Google Scholar] [CrossRef]
- Juniper, S.; Abbott, L. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza 2006, 16, 371–379. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, Q.; Yang, Z.; Hu, Z.; Tam, N.F.-Y.; Xin, G. Arbuscular mycorrhizal fungi in two mangroves in South China. Plant Soil 2010, 331, 181–191. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.; Qiu, Q.; Xin, G.; Yang, Z.; Shi, S. Flooding greatly affects the diversity of arbuscular mycorrhizal fungi communities in the roots of wetland plants. PLoS ONE 2011, 6, e24512. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: a review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef]
- Abdel Lafet, A.A.H.; Miransari, M. The role of arbuscular mycorrhizal fungi in alleviation of salt stress. In Use of Microbes for the Alleviation of Soil Stresses; Miransari, M., Ed.; Springer: New York, USA, 2014; pp. 23–38. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boughttas, S.; Hu, S.; Oh, S.H.; Sa, T. A meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Dilfuza, E.; Abd-Allah, E.F.; Hashem, A.; Kumar, A.; Ahmad, P. Salinity stress and arbuscular mycorrhizal symbiosis in plants. In Use of Microbes for the Alleviation of Soil Stresses; Miransari, M., Ed.; Springer: New York, USA, 2014; pp. 139–159. [Google Scholar] [CrossRef]
- Duke, E.R.; Johnson, C.R.; Koch, K.E. Accumulation of phosphorus, dry matter and betaine during NaCl stress of split-root citrus seedlings colonized with vesicular arbuscular mycorrhizal fungi on zero, one or two halves. New Phytol. 1986, 104, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Zhang, F.S.; Li, X.L.; Tian, C.Y.; Tang, C.; Rengel, Z. Improved tolerance of maize plants to salt stress by arbuscular mycorrhiza is related to higher accumulation of soluble sugars in roots. Mycorrhiza 2002, 12, 185–190. [Google Scholar] [CrossRef]
- Ibrahim, A.H.; Abdel-Fattah, G.M.; Eman, F.M.; Abb El-Aziz, M.H.; Shohr, A.E. Arbuscular mycorrhizal fungi and spermine alleviate the adverse effects of salinity stress on electrolyte leankage and productivity of wheat plants. Phyton 2011, 51, 261–276. [Google Scholar]
- Abdel-Fattah, G.M.; Abdul-Wasea, A.A. Arbuscular mycorrhizal fungal application to improve growth and tolerance of wheat (Triticum aestivum L.) plants grown in saline soil. Acta Physiol. Plant 2012, 34, 267–277. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Zou, Y.-N.; He, X.-H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Evelin, H.; Sarda Devi, T.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: current understanding and new challenges. Front. Plant Sci. 2019, 10, 1–21. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Maathuis, F.J.M.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef]
- Hammer, E.C.; Nasr, H.; Pallon, J.; Olsson, P.A.; Wallander, H. Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 2011, 21, 117–129. [Google Scholar] [CrossRef]
- Olsson, P.A.; Hammer, E.C.; Pallon, J.; Van Aarle, I.M.; Wallander, H. Elemental composition in vesicles of an arbuscular mycorrhizal fungus, as revealed by PIXE analysis. Fungal Biol. 2011, 30, 1–6. [Google Scholar] [CrossRef]
- Moldan, B.; Janouŝka, S.; Hák, T. How to understand and measure environmental sustainability: Indicators and targets. Ecol. Indicators 2012, 17, 4–13. [Google Scholar] [CrossRef]
- Hak, T.; Moldan, B.; Dahl, A. L. Editorial. Ecol. Indicators 2012, 17, 1–3. [Google Scholar] [CrossRef]
- Kaswan, V.; Choudhary, M.; Kumar, P.; Kaswan, S.; Bajya, P. Green production strategies. In Encyclopedia of Food Security and Sustainability; Ferranti, P., Berry, E., Jock, A., Eds.; Elsevier: Amsterdam, Netherlands, 2019; pp. 492–500. [Google Scholar] [CrossRef]
- Neuman, B.; Ott, K.; Kenchington, R. Strong sustainability in coastal areas: a conceptual interpretation of SDG 14. Sust. Sci. 2017, 12, 1019–1035. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.; Prasad Tollamadugu, N.V.K.V. Role of plant growth-promoting microorganisms as a tool for environmental sustainability. In Recent Developments in Applied Microbiology And Biochemistry; Elsevier: Amsterdam, Netherlands, 2019; Volume 179, pp. 209–222. [Google Scholar] [CrossRef]
- Farrer, E.C.; Van Bael, S.A.; Clay, K.; Smith, M.K.H. Sulfur accumulation in gypsum-forming thiophores has its roots firmly in calcium. Estuaries Coasts 2022, 45, 1805–1822. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Wang, Y.-H.; Song, Y.-B.; Dong, M. Formation and functions of arbuscular mycorrhizae in coastal wetland ecosystems: A review. Ecosys. Health Sustain. 2022, 8, 2144465. [Google Scholar] [CrossRef]
- Fontalvo-Julio, F.A.; Tamaris-Turizo, C.E. Calidad del agua de la parte baja del río Córdoba (Magdalena, Colombia), usando el ICA-NSF. Intropica 2018, 13, 101–111. [Google Scholar] [CrossRef]
- Schuettpelz, E.; Schneider, H.; Smith, A.R.; Hovenkamp, P....; Zhou, X.-M. A community-derived classification for extant lycophytes and ferns. PPG I. J. Syst. Evol. 2016, 54, 563–603. [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedure of clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infections. Trans. Br. Mycol. Soc. 1970, 55, 159–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.L. A new method which gives an objective measure of colonization of roots by vesicular arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Sridhar, K.R.; Roy, S.; Sudheep, N.M. Assemblage and diversity of arbuscular mycorrhizal fungi in mangrove plant species of the southwest coast of India. In Mangroves Ecology, Biology and Taxonomy; Metras, J.N., Ed.; Nova Science Publishers Inc.: New York, USA, 2011; pp. 257–274. [Google Scholar] [CrossRef]
- Gupta, N.; Bihari, K.M.; Sengupta, I. Diversity of arbuscular mycorrhizal fungi in different salinity of mangrove ecosystem of Odisha, India. Adv. Plant Agric. Res. 2016, 3, 19–23. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Cuevas, E.; Popp, M.; Lugo, A.E. Soil salinity, sun exposure, and growth of Acrostichum aureum, the mangrove fern. Bot. Gaz. 1990, 151, 41–49. [Google Scholar] [CrossRef]
- Matthus, E.; Wilkins, K.A.; Swarbreck, A.M.; Doddrell, N.H.; Doccula, F.G.; Costa, A.; Daviesa, J.M. Phosphate starvation alters abiotic-stress-induced cytosolic free calcium increases in roots. Plant Physiol. 2019, 179, 1754–1767. [Google Scholar] [CrossRef] [PubMed]
- Robson, T.; Jason Stevens, J.; Dixond, K.; Nathan Reida, N. Sulfur accumulation in gypsum-forming thiophores has its roots firmly in calcium. Environ. Exp. Bot. 2017, 137, 208–219. [Google Scholar] [CrossRef]
- He, H.; Veneklass, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef]
- Moore, M.J.; Mota, J.F.; Douglas, N.A.; Olvera, H.F.; Ochoterena, H. The ecology, assembly and evolution of gypsophile floras. In Plant Ecology and Evolution in Harsh Environments; Rajakaruna, N., Boyd, R.S., Harris, T.B., Eds.; Nova Science Publishers Inc.: New York, USA, 2014; pp. 97–128. [Google Scholar]
- Awang, N.A.; Ali, A.M.; Mat, N. Alternative medicine from edible bitter plants of Besut, Malaysia. J. Agrobiotechnol. 2018, 9, 80–91. [Google Scholar]
- Akinwumi, K.A.; Abam, E.O.; Oloyede, S.T.; Adeduro, M.N.; Adeogun, Y.A.; Uwagboe, J.E. Acrostichium aureum Linn: traditional use, phytochemistry and biological activity. Clin. Phytosci. 2022, 8, 2–18. [Google Scholar] [CrossRef]
- Fedick, S.L. The Maya Forest: Destroyed or cultivated by the ancient Maya? PNAS 2010, 107, 953–954. www.pnas.org/content/107/3/953. [CrossRef]
- Hellmuth, N. Edible mangrove fern, Acrostichum aureum. Municipio de Livingston, Izabal, Guatemala. FLAAR (USA) and FLAAR. Mesoamérica (Guatemala). Wetlands 2022, series 3: rivers, lagoons, swamps, or ocean, Wetlands #17.
- Lara Rosales, Y.; Ocampo Castrejon, M.E. Ensayo etnobotánico de las Pteridofitas mexicanas. Tesis de Grado, Universidad Autónoma Metropolitana, Iztapalapa, México, 1991.
- Jimenez Álvarez, S.E. Estado actual de conocimiento del uso de algunos de los helechos presentes en Colombia. Tesis de Grado, Pontificia Universidad Javeriana, Facultad de Ciencias, Carrera de Biología, Bogotá D. C., 7 julio de 2011.
- World Health Organization-WHO. Available online: https://www.who.int/news-room/feature-stories/detail/traditional-medicine-has-a-long-history-of-contributing-to-conventional-medicine-and-continues-to-hold-promise; WHO report, 2019 (accessed on 7 October 2024).
- Mofokeng, M.M.; Du Plooy, C.P.; Araya, H.T.; Amoo, S.O.; Mokgehle, S.N.; Pofu, K.M.; Mashela, P.W. Medicinal plant cultivation for sustainable use and commercialisation of high-value crops. S. Afr. J. Sci. 2022, 118, #12190. [Google Scholar] [CrossRef]
- Porwal, O.; Singh, S.K.; Patel, D.K.; Gupta, S.; Tripathi, R.; Katekhaye, S. Cultivation, collection and processing of medicinal plants. In Bioactive Phytochemicals: Drug Discovery to Product Development; Ahmad, J., Ed.; Bentham Science Publishers: Saif Zone, Sharjah, U.A.E, 2020; pp. 14–30. [Google Scholar] [CrossRef]
- Wang, C.C.; Cai, H.; Zhao, H.; Yan, Y.; Shi, J.; Chen, S.; Tan, M.; Chen, J.; Zou, L.; Chen, C.; Liu, Z.; Xu, C.; Liu, X. Distribution patterns for metabolites in medicinal parts of wild and cultivated licorice. J. Pharm. Biomed. Anal. 2018, 161, 464–473. [Google Scholar] [CrossRef]
- Zeng, Y.; Guo, L.P.; Chen, B.D.; Hao, Z.P.; Wang, J.Y.; Huang, L.Q.; Yang, G.; Cui, X.M.; Yang, L.; Wu, Z.X.; Chen, M.L.; Zhang, Y. Arbuscular mycorrhizal symbiosis and active ingredients of medicinal plants: current research status and prospectives. Mycorrhiza 2013, 23, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Zubek, S.; Blaszkowski, J. Medicinal plants as hosts of arbuscular mycorrhizal fungi and dark septate endophytes. Phytochem. Rev. 2009, 8, 571–580. [Google Scholar] [CrossRef]
- Yuan, M.-L.; Zhang, M.-H.; Shi, Z.-Y.; Yang, S.; Zhang, M.-G.; Wang, Z.; Wu, S.-W.; Gao, J.-K. Arbuscular mycorrhizal fungi enhance active ingredients of medicinal plants: a quantitative analysis. Front. Plant Sci. 2023, 14, 1276918. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cartabia, A.; Lalaymia, I.; Declerck, S. Arbuscular mycorrhizal fungi and production of secondary metabolites in medicinal plants. Mycorrhiza 2022, 32, 221–256. [Google Scholar] [CrossRef]
Figure 1.
El Volcán thermal pond sampling site of the Córdoba River mouth in the Caribbean Sea, Magdalena Department, Colombia.
Figure 1.
El Volcán thermal pond sampling site of the Córdoba River mouth in the Caribbean Sea, Magdalena Department, Colombia.
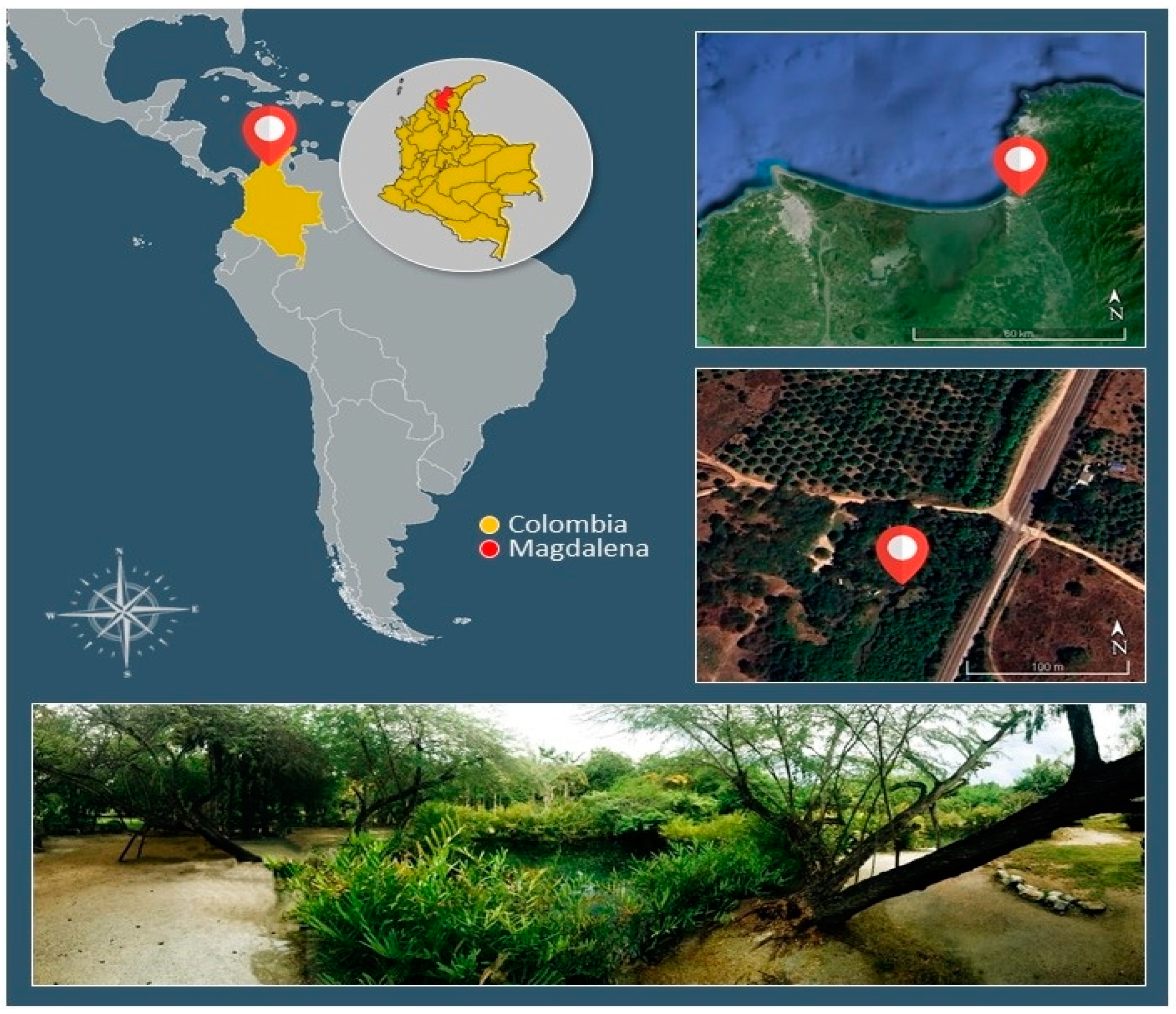
Figure 2.
Acrostichum aureum in the riparian zone in thermal ponds of “El Volcán”, Córdoba River, Colombia (a) Fern sporophytes growing on the margin (b) Frond of A. aureum in the sampling site, with fertile and non-fertile pinnae.
Figure 2.
Acrostichum aureum in the riparian zone in thermal ponds of “El Volcán”, Córdoba River, Colombia (a) Fern sporophytes growing on the margin (b) Frond of A. aureum in the sampling site, with fertile and non-fertile pinnae.

Figure 3.
Colonisation by AMF in fine roots of Acrostichum aureum (a) General view of a fern root colonised by AMF (b) Vesicles of AMF (c) Hyphal coils and arbuscules of AMF (d) Arbuscules of AMF.
Figure 3.
Colonisation by AMF in fine roots of Acrostichum aureum (a) General view of a fern root colonised by AMF (b) Vesicles of AMF (c) Hyphal coils and arbuscules of AMF (d) Arbuscules of AMF.
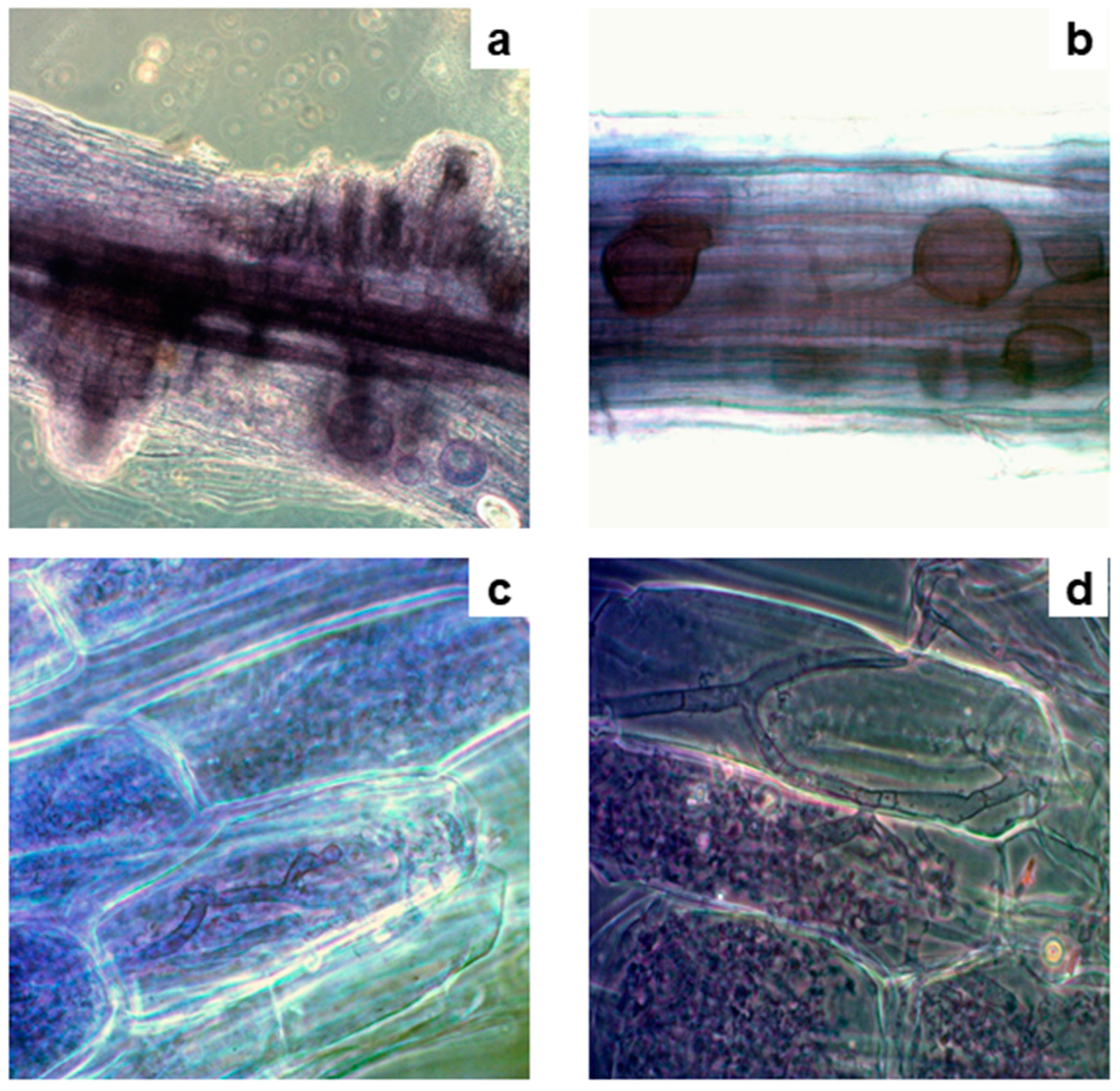
Figure 4.
AMF structures outside and inside fine roots of Acrostichum aureum with dense cytoplasmic contents (arrowhead) (a) Hypha of AMF (b) Spores of AMF outside roots (c) Vesicles of AMF (d) Arbuscules of AMF inside roots.
Figure 4.
AMF structures outside and inside fine roots of Acrostichum aureum with dense cytoplasmic contents (arrowhead) (a) Hypha of AMF (b) Spores of AMF outside roots (c) Vesicles of AMF (d) Arbuscules of AMF inside roots.
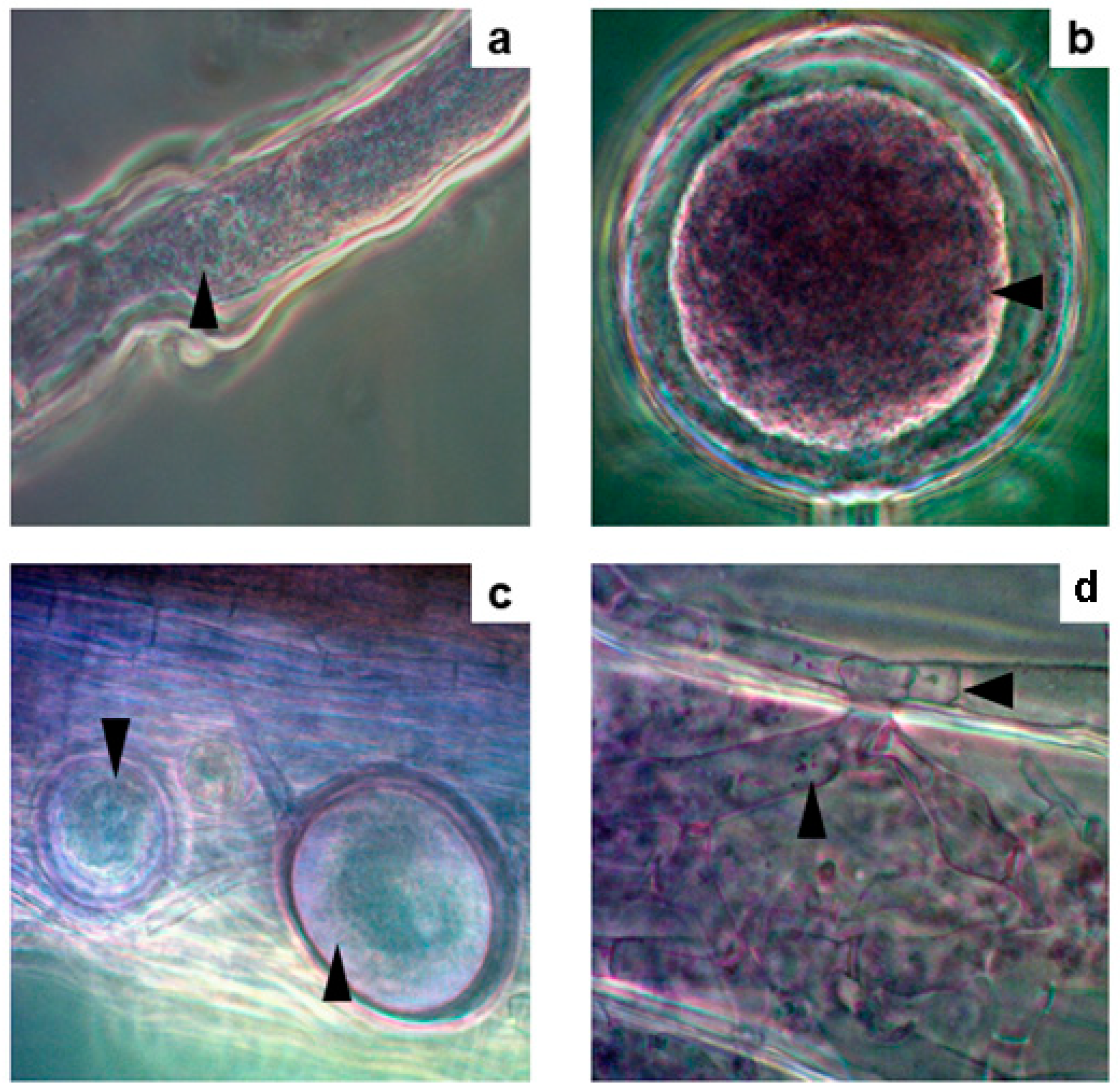
Figure 5.
Salt crystals within the vesicles of AMF (a) Vesicle of AMF with NaCl crystals (indicated by the arrowhead) observed under an optical microscope (b) The EDX analyses of crystals in the vesicles showed the elemental composition of NaCl (indicated by the arrowhead).
Figure 5.
Salt crystals within the vesicles of AMF (a) Vesicle of AMF with NaCl crystals (indicated by the arrowhead) observed under an optical microscope (b) The EDX analyses of crystals in the vesicles showed the elemental composition of NaCl (indicated by the arrowhead).
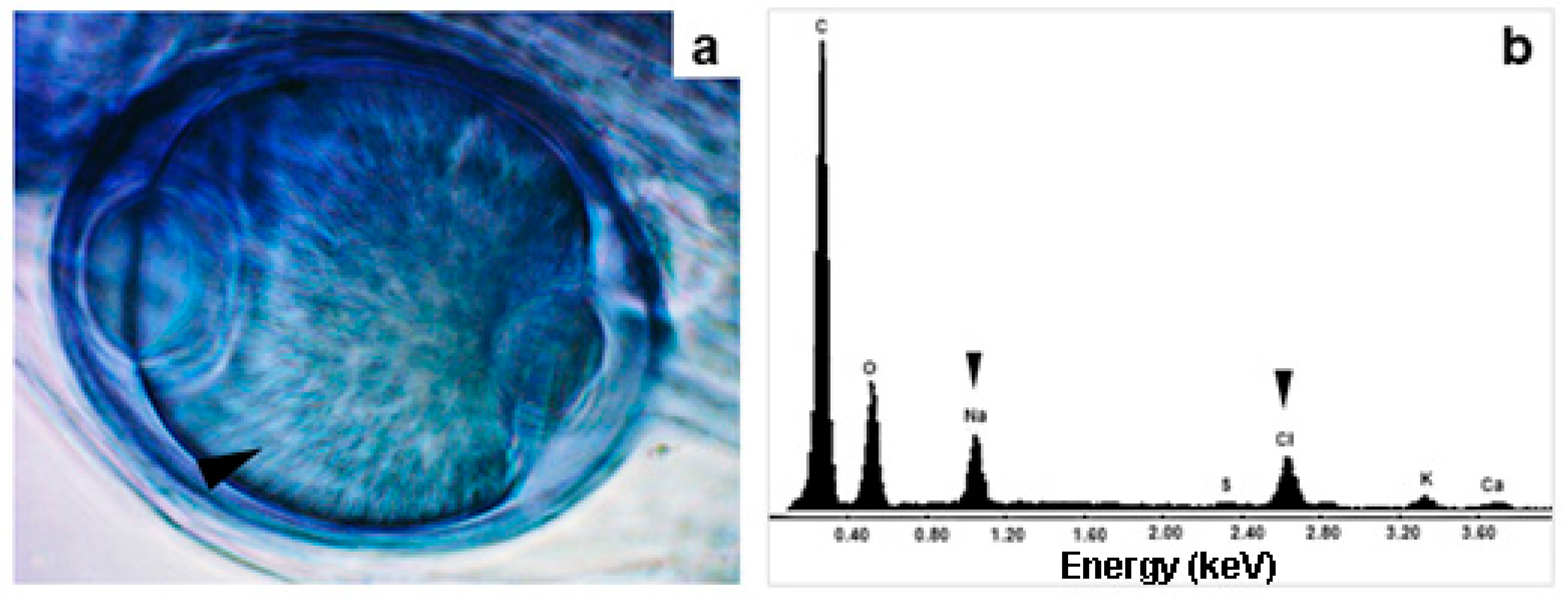
Figure 6.
Salts accumulated inside roots of Acrostichum aureum analysed by SEM-EDX (a) SEM image of amorphous crystals obtained with the backscattered electron detector (black arrowhead) (b) X-ray mapping indicating the distribution of S in an A. aureum root (red arrowhead) (c) X-ray mapping indicating the distribution of Ca in the root (yellow arrowhead) (d) EDX spectrum of amorphous crystals showing its CaSO4 composition (red, and yellow arrowheads).
Figure 6.
Salts accumulated inside roots of Acrostichum aureum analysed by SEM-EDX (a) SEM image of amorphous crystals obtained with the backscattered electron detector (black arrowhead) (b) X-ray mapping indicating the distribution of S in an A. aureum root (red arrowhead) (c) X-ray mapping indicating the distribution of Ca in the root (yellow arrowhead) (d) EDX spectrum of amorphous crystals showing its CaSO4 composition (red, and yellow arrowheads).
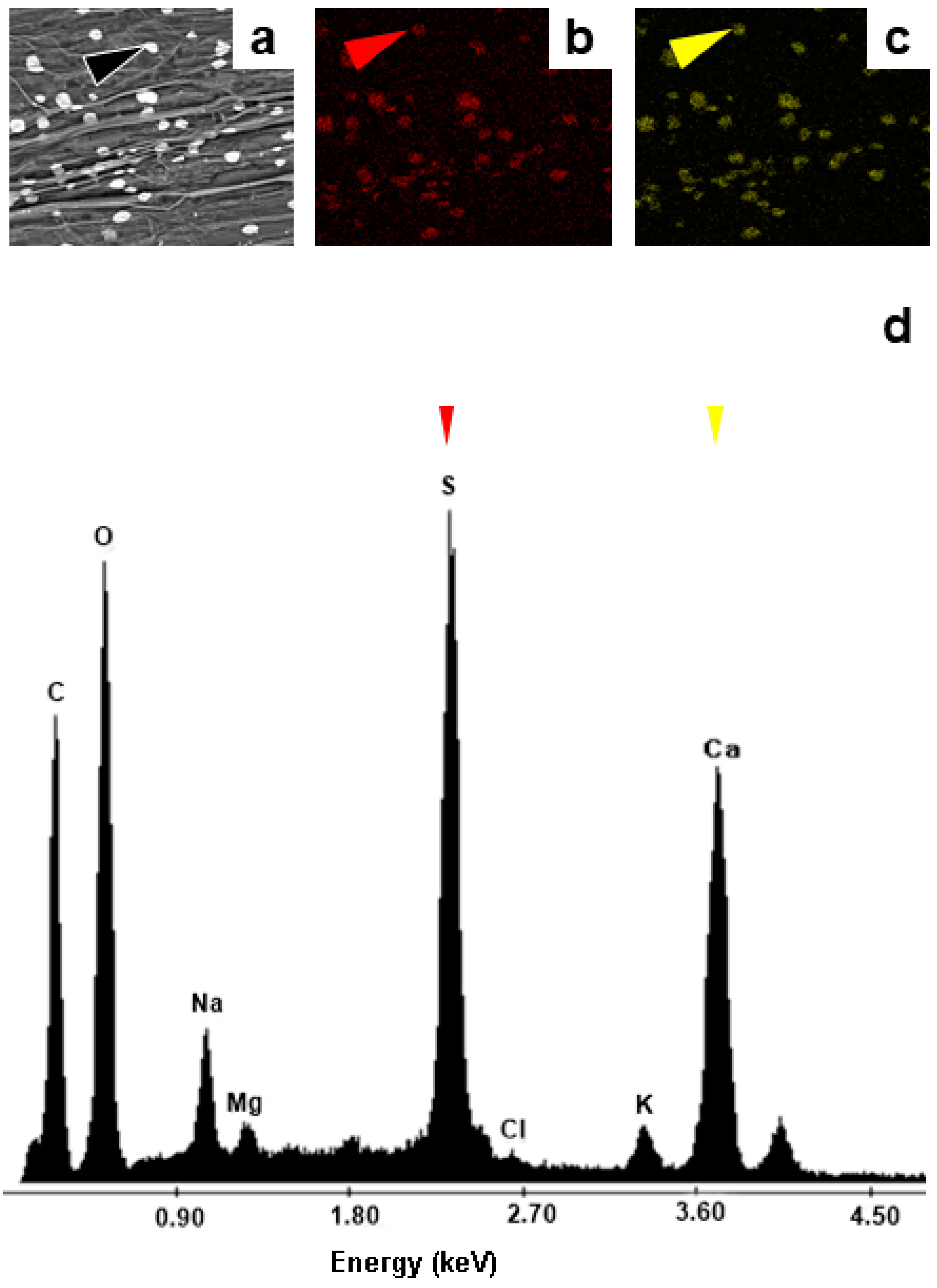

Table 1.
Chemical analysis of the thermal water where roots and rhizomes of Acrostichum aureum grow in the study area.
Table 1.
Chemical analysis of the thermal water where roots and rhizomes of Acrostichum aureum grow in the study area.
| Physico-chemical variables | Values |
|---|---|
| pH Electrical Conductivity (Ms/cm) |
5.45 |
| 3.08 | |
| N-NO2 (µg/L) | <LD* |
| N-NO3 (µg/L) | 2.5 |
| N-NH4 (µg/L) | <LD* |
| P-PO4 (µg/L) | 2.4 |
| Si-SiO4(µg/L) | 13916.1 |
| Pb (µg/L) | <LD* |
| Cd (µg/L | <LD* |
| Cr (µg/L) | <LD* |
| Cu (µg/L) | <LD* |
| Zn (µg/L) | 13.4 |
| Ni (µg/L) | <LD* |
| Fe (µg/L) | <LD* |
| Mn (mg/L) | 0.5 |
| Na (mg/L) | 240.3 |
| K (mg/L) | 4.6 |
| Mg (mg/L) | <LD* |
| Ca (mg/L) | 157.5 |
*LD: lower detection value than the methodological detection power.
Table 2.
Physico-chemical analysis of thermal sediments where roots and rhizomes of Acrostichum aureum were growing.
Table 2.
Physico-chemical analysis of thermal sediments where roots and rhizomes of Acrostichum aureum were growing.
| Physico-chemical features texture | Values |
|---|---|
| 2 mm 1 mm |
3.3% |
| 11.5% | |
| 0.5 mm | 16.9% |
| 250 µm | 16.3% |
| 125 µm | 12.3% |
| 63 µm | 7.1% |
| ≤ 63 µm | 32.7% |
| Chemical composition | Values |
| OM ox (mg/g) | 179.5 |
| OM vol (mg/g) | 266.3 |
| Humidity (% H2O/wet weight) | 60.0 |
| N-NO3 (µg/g) | *< LD |
| N-NH4 (µg/g) | 23.4 |
| P-PO4 (µg/g) | *< LD |
| Cu (µg/g) | 7.8 |
| Zn (µg/g) | 56.7 |
| Fe (µg/g) | 20.4 |
| Mn (mg/g) | 248.2 |
| Na (mg/g) | 1.8 |
| K (mg/g) | 2.1 |
* LD: lower detection value than the methodological detection power.
Table 3.
EDX chemical analysis of rhizospheric soil of Acrostichum aureum in thermal soils. The values are the mean of four replicates of EDX measurement. .
Table 3.
EDX chemical analysis of rhizospheric soil of Acrostichum aureum in thermal soils. The values are the mean of four replicates of EDX measurement. .
| Elements-K | Values* (% element/soil sample weight) |
|---|---|
| C K O K |
32.75± 2.16 |
| 32.18± 0.35 | |
| Na K | 1.75± 0.19 |
| Mg K | 0.62± 0.07 |
| Al K | 5.61± 0.33 |
| Si K | 18.43± 1.45 |
| S K | 0.41± 0.26 |
| Cl K | 0.60± 0.05 |
| K K | 1.05± 0.16 |
| Ca K | 3.91± 0.33 |
| Fe K | 2.71± 0.47 |
*Values lower than methodological detection power with percentages ˂ 1%, and main elements >10%, K= atomic energetic level of each element used as a measuring reference value; data are mean value ± standard deviation.
Table 4.
Arbuscular mycorrhizal fungi (AMF) root colonization in mature sporophytes of Acrostichum aureum in thermal soils.
Table 4.
Arbuscular mycorrhizal fungi (AMF) root colonization in mature sporophytes of Acrostichum aureum in thermal soils.
| AMF structures | Percentage of colonisation |
|---|---|
| %RL %HC |
57.93± 3.05 |
| 25.00± 10.64 | |
| %AC | 23.75± 24.62 |
| %VC | 9.29± 11.15 |
%RL= percentage of total root length colonization, %HC= percentage of hyphal colonisation, %AC= percentage of arbuscule colonisation, %VC= percentage of vesicle colonisation; *data are mean value ± standard deviation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



