
Submitted:
11 October 2024
Posted:
14 October 2024
You are already at the latest version
Abstract
2-phenylchromen-4-one, commonly known as flavone, plays multifaceted roles in biological response, largely due to its abundance in natural sources. The methoxy group in naturally occurring flavones promotes cytotoxic activity in various cancer cell lines by targeting protein markers, in facilitating ligand-protein binding mechanisms and activating cascading downstream signaling pathways leading to cell death. However, the lipophilic nature of these analogs is a key concern as it impacts drug membrane transfer. While lipophilicity is crucial for drug efficacy, excessive effects in flavonoids can reduce water solubility and hinder drug transport to target sites. Recent in vitro studies suggest that the incorporation of polar hydroxyl groups which can form hydrogen bonds and stabilize free radicals may help overcome the challenges associated with methoxy groups while maintaining their essential lipophilic properties. Naturally coexisting with the methoxyflavones, this review explores the synergistic role of hydroxy and methoxy moieties through hydrogen bonding capacity in maximizing cytotoxicity against cancer cell lines. The physicochemical analysis revealed the potential intramolecular interaction and favorable electron delocalization region between both moieties to improve cytotoxicity levels. Together, the analysis provides a useful strategy for structural-activity relationship (SAR) of flavonoid analogs in distinct protein markers, suggesting optimal functional group positioning to achieve balanced lipophilicity, effective hydrogen bonding, and simultaneously minimized steric hindrance in targeting specific cancer cell types.
Keywords:
Medicinal plants
; methoxyflavones
; physicochemical properties
; structural-activity relationship
; anticancer mechanisms
1. Introduction
Flavones, a prominent subclass within the flavonoid family, exhibit a multi-target pharmacological profile owing to their intricate structural system. This structure features a highly soluble chromone core (1-benzopyran-4-one) with a hydrophobic phenyl ring attached to the ortho position of pyran ring [1]. The unique pharmacological properties of natural flavonoid, particularly flavones contribute to its diverse bioactivities in both traditional therapies and modern medicine [2,3,4,5]. The physicochemical properties of these flavones are largely attributed to two major group that coexist, predominantly the methoxy and hydroxy substituents in natural [6,7]. Both groups exhibit distinct characteristics and work synergistically in an electron donor-acceptor mechanism, influencing electron density distribution depending on their position in rings A, B, and C of the flavone structure [8,9,10]. As a result, the flavones pharmacophores significantly influence the bioavailability of flavones, one of the primary focus in discovering potential therapeutic drugs (Moldoveanu & David, 2022) [11]. Drug discovery originating from medicinal plants has fruitfully developed new potent therapeutics, such as morphine, an alkaloid derived from Papaver somniferum (opium poppy) [12]. Similarly, the discovery of taxol, a terpenoid compound from Taxus brevifolia (pacific yew tree) led to the commercialization of paclitaxel as an antitumor drug [13]. Despite the reported biological efficacy of flavones, a comprehensive systematic review authored by Xu et al. highlighted significant challenges in developing anti-tumor drugs based on flavones aglycones. These challenges are often due to physical and structural limitations that affect the bioavailability. Thus, efforts to delve into the chemistry of highly lipophilic flavones were enormous, particularly focusing on the hydrophobic region such as the unsaturated double bond between C2 and C3 on ring C and phenyl ring B.
The distinct physicochemical attributes of methoxy and hydroxy moieties add a prospect to the synergistic relationship between both functional groups in enhancing biological responses. Therefore, the in-depth analysis of the physicochemical properties, including lipophilic effects, intramolecular interactions and hydrogen bonding capacity will be useful in the structural-activity relationship (SAR) investigations of methoxyflavones’ analogs to determine its target of interest and eventually contribute to its distinct biological activities [15,16,17]. An optimal lipophilic capacity of a lead compound is part of the major limitation of flavones in anticancer drug discovery [18,19]. The partition coefficient, LogP is one of the parameters to predict hydrophobicity and lipophilicity and has been validated on various flavone scaffolds [20]. The lipid-water partitioning of flavones is crucial given its influence on the uptake and delivery of flavones to the target sites. In plant species, methoxy moieties within the flavones scaffold strengthen the hydrophobic interaction through methylation, possessing hydrogen bonding acceptor (HBA) capabilities and initiating a stabilized ligand-protein binding energy [17,21]. Despite its greater binding and interaction with the active sites, the methoxylated group is a metabolic liability to a compound that leads to biotransformation through the O-demethylation metabolic pathway [22,23]. Although not reported for every naturally isolated and synthesized methoxyflavones scaffold, methoxylation could alter bioavailability, solubility, and pharmacology in flavones [24,25,26]. Nevertheless, the presence of the hydroxy group within the methoxyflavones scaffolds could offset the drawbacks by altering the extreme lipophilicity of flavones through intramolecular interaction and greater electron delocalization region in stabilizing the free radicals and established strong interaction with the target binding sites as hydrogen bond donor (HBD) (Figure 1) [27,28]. However, the incompatible position of the highly polar hydroxyl group within the flavones’ pharmacophores may activate unfavourable thermodynamic properties and desolvation penalties which pose significant challenges in medicinal chemistry [29,30].
**This diagram illustrates the relationship between the methoxyflavones scaffolds and their effect on the IC50. The C7 position plays a key role in the cytotoxic activity on cancer cell lines. The strong effect of C7-OH scaffold was contributed by at least one methoxy moieties on ring A and -OH moieties on the C5 or C5’ position. C6’-OH could exert a similar effect if the pharmacophore of C7-OCH3 or -OH was preserved. The C7-OCH3 was widely studied and contributed the most in yielding strong IC50 (<20 µM), with few exception:
- With C5-OCH3, stronger IC50 could be achieved by either the presence of optimal polar region from -OH moieties or HBA strength of at least one -OCH3 on ring B. Having multiple methoxy moieties on ring B significantly deteriorates the IC50 of the scaffold on numerous cancer cell lines.
- C5-OH, in general, was the most favored to induce strong IC50, with a few exclusions. The established methoxylated backbone on ring A (C6,7 or 8) with the addition of both methoxy and hydroxy moieties in neighboring positions (C3’ & C4’) strengthens the IHB, HBD and HBA effect. Otherwise, the absence of hydroxy moieties on ring B could be offset by the single methoxylated substitution on the same ring. The C3-OCH3 could coalesce, dependent on cell types.
As illustrated in Figure 1, the subtle repositioning of hydroxy and methoxy substituents on the flavone ring greatly affects the biological activity and the application of the flavones’ analogs either in the hydrophobic or polar aqueous region [31]. The coexistence of a hydroxy functional group may expand the polar region through hydrogen bonding interactions, though this effect depends on its position (Veiko et al., 2021) [32]. The hydroxy group in flavonoids including flavones has been extensively researched for its ability to scavenge free radicals, diminish ROS levels, and contribute to its potency as antioxidant, anticancer, and anti-inflammatory agents [33,34,35,36,37]. Hydrogen bonding of the hydroxyl group is notably robust and crucial for acting as a hydrogen bond donor. Nevertheless, an enormous polar region might accelerate drug hydrolysis and impair lipid membrane permeation (Hawthorne et al., 2022) [20]. Despite challenges related to metabolic stability and lipophilicity, methoxy moieties in flavones analogs have shown selectively potent anti-inflammatory effects, enhanced by the C4’-methoxyl group and an increased anti-proliferative effect from C7-methoxy substitution [16]. In addition, the existence of both methoxy and hydroxy functional groups in adjacence and its impact on the stereospecificity of flavones merits further exploration [16]. These interactions could promote the formation of an intramolecular interaction between the two atoms, serving as a double-edged sword by improving the stability of the parent structure and potentially blocking polar interaction in favor of a lipophilic environment through the shielding effect of intramolecular hydrogen bonding (IHB) [39,40]. The IHB between a hydroxyl group adjacent to the carbonyl group in various aromatic and polyphenolic compounds has been reported in computational studies [41,42,43]. For instance, the C5-OH substituent within the flavone skeleton interacts with C4 carbonyl group through intramolecular hydrogen bonding, with multiple studies showing that the associated increase in lipophilic capacity enhances cytotoxic effects on cancer cell lines more than C5-OCH3. In addition, the methoxy group acting as a hydrogen bond acceptor (HBA), may also form intramolecular hydrogen bonds in optimally positioned flavones analogs, particularly when ortho to hydroxy moieties. The presence of C3’-OCH3 lowers the hydrogen bonding acidity of C4’-OH by 20%, suggesting a stronger potential for stable IHB formation between the OH and OCH3 groups, thereby increasing lipophilic character [44,45]. Nevertheless, stable IHB formation can restrict the hydrogen donor capacity of C4’-OH and reduce its affinity for target proteins, unless compensated for by the external interaction with protein markers. In flavones, the intramolecular hydrogen bonding with the adjacent atoms predominantly occurs in rings A and B. Given their distinct roles in polar-hydrophobic interactions, a balanced relationships between these moieties is crucial for maximizing the pharmacological potential of flavones, particularly in anticancer applications.
2. SAR and Mechanism of Anticancer Activity of Methoxyflavones Derivatives
2.1. Breast Cancer
Compound 5,3’-dihydroxy-3,6,7,8,4’-PeMF, isolated from Glycosmis ovoidea demonstrated potent cytotoxic effects on breast cancer cells, MCF-7 with IC50 values of 49.46, 17.56 & 3.71 μM [46]. Signs of apoptosis such as cell membrane blebbing, shrinkage, and fragmentation were observed. A loss of mitochondrial membrane potential at concentrations of 2 & 20 μM was noted, indicating that intrinsic apoptosis had occurred [47].
**This figure illustrates the effect of pharmacophore changes on ring B. The presence of both moieties on neighboring positions (C3’ & C4’) is a crucial factor for stronger IC50 of 5-OH methoxyflavones analogs. The C8-OCH3 plays a crucial role in stabilizing the lipophilic capacity of ring A flavones. Given the presence of C6 and C7-OCH3, the reduction of single C8-OCH3 will not significantly affect the IC50 of these scaffolds. for xanthomicrol, the absence of methoxy adjacent to the C4’-OH was a single factor in the expected weak cytotoxicity profile.
By swapping the hydroxy and methoxy groups’ positions from C4’ to C3’, and removing of C8-OCH3, chrysosplenetin (5,4'-dihydroxy-3,6,7,3'-TeMF) was isolated and effectively induced cancer cell death on MCF-7 in 72-hour treatment with stronger IC50 value of 0.3 μM [48]. Another methoxyflavones, xanthomicrol, 5,4’-dihydroxy-6,7,8-TMF was inactive with an IC50 exceeding 100 µM [49]. Based on the schematic diagram (Figure 1), the 3,6,7-TMF scaffold is the backbone that preserves the lipophilic character, acts as a potential hydrogen bond acceptor, and facilitates hydrophobic interaction with the target proteins. The delocalization region of electron density from C5-OH free radicals (Figure 3) enable a highly stabilized intramolecular hydrogen bond (IHB) with the adjacent carbonyl group. This IHB between C5 and C4 substituents elevates the lipophilic capacity of the flavones’ analogs enhancing drug-lipid transfer and cytotoxic activity [44,50]. Interestingly, the interchanging effect of neighboring moieties on ring B significantly influences the cytotoxicity in MCF-7 reducing the IC50 from 3.71 to 0.3 µM, which highlights the favorable positioning of the C4’ hydroxy group over C3’. Regardless, both positions are within the ideal region for maximizing the potential of the hydroxy group to intensify the anticancer effect of methoxyflavones analogs. Similarly, compared to C5-OH, C4’-OH free radicals generate a stronger resonance effect than the C3’ attributed to the extended pi-conjugation from C2-C3 unsaturated bond up to carbonyl ring C (Figure 3) [27]. The existence of stabilized hydrogen bonding between C5-OH and the neighboring carbonyl group, along with the resonance effect from C4’ free radicals assist in interaction with C3’-OCH3, preserving the conformation and shielding the polar surface on ring B against specific target [51,52,53,54]. As a result, the absence of a neighboring methoxy group next to C4’-OH (xanthomicrol) leads to poor bioactivity. Thus, greater reduction potential from the resonance effect paves the way for strong hydrogen bonding with protein markers. The removal of C8-OCH3 unexpectedly preserved the biological effect of the scaffold. Due to the limited data available to predict the roles of C8-OCH3, it is hypothesized that for specific cell lines, the methoxylation of at least three positions within the AC-ring system from C5-OH methoxyflavones would be accommodative in balancing its physicochemical properties. C8-OCH3 may not necessarily contribute to the cytotoxic effect against MCF-7, even with the presence of heavily substituted C3,6,7-OCH3 in the methoxyflavones scaffold.
Focusing on triple-negative breast cancer cell lines, a similar compound 5,3’-dihydroxy-3,6,7,8,4’-PeMF was compared with nobiletin, 5,6,7,8,3’,4’-HeMF for cytotoxicity activity against MDA-MB-231 cell lines. As anticipated, compound 5,3’-dihydroxy-PeMF significantly reduced cell viability with an IC50 of 21.27 μM, compared to nobiletin which showed ~25% reduction at 10 μM, with no IC50 reported after 72-hour treatment. A marginal increase in IC50 in MDA-MB-231 cells compared to MCF-7 may be attributed to the highly invasive and metastatic nature. Several factors contribute to nobiletin inactivity including, though not restricted to (i) extreme lipophilic effect of ring B, (ii) exacerbated steric hindrance, (iii) and absence of IHB that stabilize the pharmacophore, leading to compound degradation. Despite the similarities in functional groups between the two compounds except for C3 hydroxy group, the role of hydroxy groups in the antitumor activity of methoxyflavones was noteworthy. The direct apple-to-apple comparison may be premature to definitively claim that hydroxy flavones are the key markers in flavones analogs’ effectiveness as opposed to the methoxy groups, reports indicate that hydroxyflavone analogs’ cytotoxicity against MDA-MB-231 cell lines is considerably lower with IC50 values around 200 μM for treatments ranging from 24 to 72-hour, compared to nobiletin. This reduced effectiveness is attributed to the extensive polar surface of polyhydroxylated flavones [55,56,57]. Thus, strategically optimizing the placement of hydroxy moieties within the methoxyflavones pharmacophore could balance the extreme lipophilic and polar region features, enhancing their synergistic interaction.
OH methoxyflavones scaffolds, influenced by the C5’-OH. As summarized in Figure 1, excessive methoxylated effect on ring B yielding negative IC50 result on cancer cell lines.
The polymethoxyflavones were screened against human epidermal growth factor receptor 2 positive (Her-2+) HCC1954 breast cancer cell lines in vitro by [58]. Compound 4',5'-dihydroxy-5,7,3'-TMF exhibited strong cytotoxicity against these cancer cells with an IC50 value of 8.58 µM (Figure 4). The cytotoxicity of the treatment was marginally weaker compared to the previous study that had been discussed due to the more invasive and metastatic nature of the HCC1954 relative to MCF-7 [59]. Surprisingly, dehydroxylated moieties on ring B increased cytotoxicity activity. The formation of IHB between C4’ and C5’ hydroxy groups, which was found to be active and greatly stabilized the scaffold and preserved the lipophilic properties of the flavones [27]. However, modifying the skeleton by substituting C5'-hydroxy to C5’-methoxy group (4'-hydroxy-5,7,3',5'-TeMF) significantly weakened the cytotoxic effect, resulting in an IC50 exceeding 100 µM. The comparison between both compounds highlights several challenges including the bulkiness of the C5’-OCH3 group that hindered IHB formation with the carbonyl group and the presence of multiple methoxylated substitution on ring B (C3’ and C5’) which were unfavorable. Without C5-OH, the C4’-OH in compound 4'-hydroxy-5,7,3',5'-TeMF exhibited unexpectedly poor cytotoxicity, triggered by an increased potential for resonance effect arising from C4’-OH free radicals to the carbonyl group as both AC- and BC-ring electron delocalization were not in conjugated system [60,61]. The C4’-hydroxy regions of flavone pharmacophores are known for having the weakest hydrogen bonding energy in ring B and are the most active radicals, that readily distribute electron density with greater active region rise from the ring B up to carbonyl group of ring C from resonance and conjugation effects [62]. Thus, an excessive polar surface significantly weakens the bioactivity of the scaffold. In fact, fully methoxylated flavones, 5,7,3’,4’,5’-PeMF demonstrated a better IC50 at 53.84 µM, although the range was considered weak as the IC50 exceeded the threshold of 50 µM. With a limited polar surface, the massive lipophilic capacity from C3’,4’,5’-trimethoxy substitution may enhance membrane transfer but will restrict strong hydrogen bonding interaction with the protein target markers. As a result, the addition of two more methoxy groups on C-6 and C-8 (5,6,7,8,3′,4′,5′-heptamethoxyflavone) resulted in weak cytotoxic effects on the cell lines with an IC50 > 100 µM. We hypothesize that high methoxy substituents on ring A of the flavones structure, with the absence of a polar region worsens the active region of methoxyflavones, thus suppressing the antiproliferative effect on HCC1954 breast cancer cell lines. Nevertheless, the SAR analysis revealed that the C5’-OH contributes to enhancing the antiproliferative effect on HCC1954 cell lines. An in-depth investigation into the incorporation of C5’-OH with the methoxyflavones scaffold may be required as current data is limited to validate the strength of the C5’ position.
2.2. Prostate Cancer
The anticancer effects of methoxylated flavones have been extensively studied against various prostate cancer cell lines in vitro, including PC3, VCaP, LNCaP, and DU145. Recent research involving six methoxyflavones across five studies revealed that all flavones’ derivatives bearing C6-OCH3 incorporated into their skeleton structure (Figure 1). The variations in the antiproliferative effects against different cell lines can be thoroughly analyzed, given the similarities in parenthetical structures to determine the future potential target of these flavone derivatives and may postulate the crucial role of C6-OCH3 in combatting prostate cancer.
In studies targeting PC3 prostate cancer cell lines, both polymethoxyflavones, tangeritin (5,6,7,8,4'-PeMF) and 5-demethyltangeritin (5-hydroxy-6,7,8,4'-TeMF) exerted strong cytotoxicity in a concentration-controlled manner [63]. Within 48-hour treatment, both compounds significantly reduced PC3 cell viability with IC50 values of 17.2 and 11.8 µM, respectively (Table 1). The increase in logP magnitude correlates with enhanced hydrophobic capacity, facilitates strong intramolecular hydrogen bonding (IHB) between C5-OH and the neighboring C4 carbonyl group. While IHB was absent with C5-OCH3, this group likely interacts with the target site as HBA, enabling polar interactions with the carbonyl group and the binding target [64]. The SAR analysis suggests that both hydroxylation and methoxylation at the C5 position significantly contribute to the antiproliferative effects on PC3 cell lines. Conversely, another study on similar cancer cell lines showed minimal bioactivity of nobiletin (5,6,7,8,3′,4′-HeMF) in a concentration-dependent manner with IC50 values around 80 µM for the same treatment duration [65]. A comparison of cell viability between the two studies confirmed the inactive anti-proliferative effect of nobiletin. Treatment of 20 µM tangeritin reduced cell viability to below 50%, whereas a similar concentration of nobiletin maintained a 100% cell viability over the same treatment period. Interestingly, the only structural difference between tangeritin and nobiletin is the addition of C3’-OCH3 to nobiletin’s structure. The increased bulkiness of ring B in nobiletin, due to multiple methoxy substitutions at adjacent positions (C3’ & C4’), enhances steric hindrance and elevates both the partition coefficient (logP), and distribution coefficient (logD) (Figure 5) [66]. Therefore, the presence of an optimal polar surface provided by hydroxy moieties at ring B introduces an amphipathic characteristic, essential for a more potent cytotoxic effect on cancer cell lines.
**This table illustrates pharmacophore changes on ring B flavones (C3’ and C4’) flavones significantly influenced the IC50 of the analogs. Green indicates favorable moieties and red signifies unfavorable position.
Besides PC3 cell lines, another androgen-independent DU145 cell line is known for its metastatic and invasive behavior due to its non-reliance on hormones for growth [70,71]. Compared to the vehicle group (0.5% DMSO), casticin (5,3’-dihydroxy-3,6,7,4’-TeMF) induced a weak cytotoxic effect maintaining over 80% cell viability at concentrations of 20-50 µM during 48-hour treatment [68]. Structural modifications on rings A and B specifically shifting C4’ > C8-OCH3 and C3’-OH to a more favorable hydroxy position at C4’-OH resulted in the creation of calycopterin (5,4’-dihydroxy-3,6,7,8-TeMF). However, the cytotoxic potential of calycopterin remained low with an IC50 value of 235 µM in 48-hour treatment [69]. Nevertheless, when comparing IC10 values of both compounds, casticin (IC10: 2.5 µM) was significantly more potent than the calycopterin (IC10: 20 µM). SAR analysis suggested similar cytotoxic behavior in both hormone-independent PC3 and DU145 cells upon treatment with these derivatives. The C6,7-OCH3 group was identified as the backbone for the potent antiproliferative effect through lipophilic capacity and pi-alkyl hydrophobic interactions. Meanwhile, the absence of at least one methoxy group (C3’ or C5’) adjacent to the C4’-OH adversely reversed the potency (Figure 5). Multiple anticancer studies on both cell lines depicted comparable cytotoxic capacities across different samples (Table 2).
** This diagram illustrates the structural modifications between various flavone derivatives and their associated cytotoxic effects. Blue highlights indicate regions of the molecule that enhance cytotoxicity, while red highlights denote regions that reduce it. The arrow directions show the transformation from one compound to another, detailing the specific chemical changes like the addition or removal of hydroxyl (OH) and methoxy (OMe) groups. For example, the transformation from nobiletin to 5-demethyltangeritin shows the replacement of methoxy groups with hydroxyl groups, correlated with an increase in cytotoxic activity.
** This table validate the similarities on cytotoxicity activity of some natural and synthetic compounds on both hormonal-dependent prostate cancer cell lines.
VCaP and LNCaP are androgen-dependent cell lines with weak metastatic potential. Interestingly, owing to the structural similarities of flavonoid families with reproductive hormones, these compounds demonstrate active interactions with androgen receptors [76,77]. The list of phytoestrogenic and phytoandrogenic compounds known to be polyphenolics which demonstrate strong radical scavengers [78]. Thus, we hypothesize that an increased polar region on ring B of methoxyflavones may induce stronger cytotoxic activity on hormone-dependent cell lines by targeting hormone receptors. Supporting this hypothesis, calycopterin exhibited potent cytotoxicity on LNCaP cells (IC50: 116.5 µM) compared to DU145 cells (235.0 µM) in 48-hour treatment [69], although, the IC50 still exceeded 100 µM. Similar to observations in PC3 cell lines, the influence of C4’-OH in enhancing free radicals scavenging capabilities was suppressed by the IHB formation between C5-OH and the carbonyl group, further limited by the absence of IHB in ring B resulting in a smaller region of electron delocalization (Figure 5) [27]. To support the hypothesis, cell viability remained above 80% (100 µM) after 48-hour treatment with nobiletin on VCaP cells, significantly weaker compared to treatment on hormone-independent PC3 cells [65]. Thus, the lack of a polar region within the aromatic ring B leads to an increased lipophilic capacity, that hinders the hydrogen donating capability. Likewise, the enormous hydroxylation in the flavone’s lipophilic backbone at ring A in C5,7,4’-OH (hispidulin / 5,7,4’-trihidroxy-6-methoxyflavone) failed to improve the cytotoxic effect, proving that the lipophilic effect of methoxy group on ring A is crucial in creating hydrophobic interactions, with hydroxy moieties on ring B to elevate its cell death effects [67,79]. We can conclude that both methoxylated and hydroxylated groups play a crucial and synergistic role in their respective region on rings A and B of the flavone structure, maximizing cytotoxic effects on both hormone-dependent and -independent cell lines.
2.3. Colon Cancer
The compound 5,7,3',4'-TeMF (60 µM) slightly reduced HCT116 cell viability to approximately 75% upon treatment for 72-hour [80]. Meanwhile, another tetramethoxyflavones, compound 7,8,3',4'-TeMF reduced viability marginally to 90% under the same conditions. The rearrangement of the methoxy group from C5 to C8 did not significantly enhance the cytotoxic effect of the TeMF derivatives on HCT116 cell lines (Figure 6). The strong hydrophobic effect from the methoxylated substituents, combined with a lack of polar interaction sites on ring B, may contribute to the impotent effect. Although the rearrangement of C5 to C8-OCH3 reduced the cytotoxic effect, it is hypothesized that the absence of C5-OCH3 or -OH plays a greater role in influencing the bioactivity of methoxyflavones analogs. Based on the behavior of C8-OCH3 on cancer cell lines, these moieties have little to no effect on anticancer efficacies. Nevertheless, despite higher cell viability, both tetramethoxyflavones (TeMF) reduced cell migration and invasion, consistently decreasing cell motility, and the expression of vimentin, and axin2 mRNA, key components of the Wnt-β-catenin signaling pathway in HCT116 cells [80].
**This diagram illustrates the molecular structures of various methoxyflavones and their respective impacts on cell viability. Key features include:
- 5,7,3',4'-TeMF at 60 µM resulting in 75% cell viability, highlighting its moderate cytotoxic effect.
- 7,8,3',4'-TeMF showing a slight reduction in cytotoxic efficacy with about 90% cell viability at the same concentration.
- Xanthomicrol (5,4'-dihydroxy-6,7,8-TMF) and sudachitin (5,7,4'-trihydroxy-6,8,3'-TMF) demonstrating enhanced cytotoxic effects with 42% cell viability at 15 µM and an IC50 of 56.23 µM at 48 hours, respectively.
- The arrows indicate the structural transitions between compounds, noting the shift from C5 to C8-OCH3 and the associated impact on cytotoxicity. Highlighted green regions emphasize the structural components correlated with increased cytotoxic effects, whereas red highlights denote areas associated with decreased effects. This visualization aids in understanding how specific structural modifications influence anticancer activity against HCT116 cells.
In vitro treatment of xanthomicrol, 5,4'-dihydroxy-6,7,8-TMF for 24 hours on HCT116 colon cancer cell lines potently reduced the cell viability to 42% at 15 μM and to 3% at higher concentrations (21 μM) [81]. Meanwhile, sudachitin, 5,7,4'-trihydroxy-6,8,3'-TMF moderately suppressed cell viability in HCT-116 and HT-29 colorectal cell lines in 48-hour treatment of yielding IC50 values of 56.23 µM and 37.07 µM, respectively [82]. The screening portrays hydroxy group at position C5 and C4’ in xanthomicrol were potent against HCT116, while the additional C7-OH in sudachitin disrupted the methoxyflavone scaffold, which moderately impacting the efficacy, owing to stabilized IHB between C3’-OCH3 and C4’-OH (Figure 6). As highlighted in our schematic diagram, the disproportionate substitution of the methoxy group, particularly on ring B (C3’ and C4’) of both tetramethoxyflavones leads to steric hindrance, that inhibits the formation of intermolecular bond and reducing the antiproliferative effects. Xanthomicrol and sudachitin demonstrate a perfect hypothesis whereby the weak cytotoxic effect by the extra bulkiness of the methoxylated substituent on ring A and absence of polar region on ring B were offset by hydroxylation of C4’, contributing to stronger bioactivity. The presence of methoxy and hydroxy moieties on position C5 ring A did not significantly influence the bioactivity of flavones derivatives, as both exert similar lipophilic characteristics through different mechanisms - methylation and electron delocalization. Nevertheless, the unexpected cytotoxicity of xanthomicrol on HCT116 gained our interest due to the deviation from the proposed SAR schematic in Figure 1. With the formation of IHB between C5-OH with carbonyl group C4, and the lack of methoxy group on ring B, we are expecting a moderate effect resulting from the exposure of C4’-OH polar region. For instance, xanthomicrol reported weak activity against HeLa and MCF-7 with IC50 exceeding 100 µM in 24- and 72-hour, respectively [49,83]. Meanwhile, calycopterin, with a similar ring B scaffold to xanthomicrol showed a weaker activity against prostate cancer cell [12]. Therefore, we suggest that the HCT116 viable cells may be selectively suppressed by the polar surface of C4’-OH in xanthomicrol indicating a role in cell death induction. The overwhelming cell death induced by xanthomicrol was due to high cell accumulation at the G2/M phase (15 μM) and G1/G0 phase (21 μM) through early and late apoptosis [81]. We believe that C4’-OH may be a critical substituent that triggering the apoptotic cell death pathway, evidenced by its potent IC50 values. These findings suggest that increased methoxylation on the flavone skeleton does not correlate to elevated cytotoxicity on cancer cells. Instead, cytotoxicity appears to depend on the specific positioning of substituents and the strategic placement of hydroxyl groups within the flavone structure.
2.4. Liver Cancer
Demethylated nobiletin's derivatives revealed significant antiproliferative potential against HepG2 liver cancer cell lines [84]. Compound 5,3'-didemethylnobiletin demonstrates the strongest IC50 (41.37 μM), followed by 5,3',4'-tridemethylnobiletin (46.18 μM), 5-demethylobiletin (47.31 μM) and 5,4'-didemethylnobiletin (54.46 μM) when treated for 24-hour.
** This diagram maps out the structural modifications of 5-demethylnobiletin and their corresponding IC50 values, indicating cytotoxic potency against HepG2 cells. Each derivative (1-4) shows a different substitution pattern on ring B methoxyflavones structure, highlighted in the blue circle. The IC50 values adjacent to each structure illustrate the effect of these modifications on cytotoxic efficacy. Notably, derivative 2 with a hydroxyl group at position R3' shows the lowest IC50 value of 41.37 µM, suggesting increased efficacy compared to other derivatives.
Demethylation of C3’ and C4' in compounds (1) to (4) significantly enhances cytotoxic effects on HepG2 cells as depicted in Figure 7. Hydroxylation at C5, together with C3’ or C4’ has been shown to improve cell death effects with C4’ displaying slightly superior potential, arguably. Nevertheless, based on our proposed figure, the cytotoxic potential between C3’- and C4’-OH derivatives narrows down to the target cell death markers, preferrable of the polar surface region on ring B and lipophilic capacity of the target compound. Although C4’-OH flavone derivatives have demonstrated strong cytotoxic activities across various cancer cell lines as compared to C3’-OH with the presence of C5-OH, it may not be the case for HepG2’s anti-cancer mechanisms. In principle, as illustrated in the breast cancer section and multiple studies, the potent effect of C4’-OH is contributed by the greater electron delocalization within rings B and C, which further stabilizes free radicals within the conjugated system and boosts electron donor capability. As a result, the potential of IHB interaction between the adjacent group intensified [44]. To add, C3’ and C4’ interaction with protein targets through respective electron donor and acceptor capability, could induce more potent cytotoxic effects. A highly stable conformation preserves the molecular structure, nonetheless, may hinder interactions with the target site, which is coherent with the multiple reports that additional hydroxylation on ring B does not significantly correlate with the potency effect on cancer cells. Interestingly, the interaction that exists between the hydroxy group at C3’ and C4’ on ring B (P4) achieved comparable IC50 values to its methoxylated counterpart (P1). To conclude, all four compounds show strong potency despite a variety of functional groups, especially the catechol moiety in ring B, which is commonly found in naturally isolated flavones [85]. Thus, we hypothesize the demethylation of C5, alongside 6,7,8-trimethoxylated substituent on ring A is crucial for anticancer activities on HepG2 cell lines.
** This diagram displays the structural variations among three flavonoid derivatives and correlates these changes with their cytotoxic effectiveness. Key features highlighted:
- Favored moieties (green) are indicated on structures where modifications have led to improved cytotoxic activity.
- Unfavored moieties (blue) are shown on structures where modifications have reduced cytotoxic effectiveness.
-
Each compound's impact on cell viability is quantified next to the respective molecular structure, with IC50 values provided:
- o
- 5-demethylnobiletin: 50% cell viability at 47.31 μM over 24 hours.
- o
- Sinensetin: 70-80% cell viability at 50 μM over 24 hours.
- o
- Sudachitin: 50% cell viability at 49.32 μM over 48 hours.
The arrows indicate structural transformations and their direct impact on cytotoxic efficacy, illustrating how specific modifications in the flavonoid skeleton influence cell death pathways in HepG2 cells. This visual representation aids in understanding the critical chemical groups responsible for the compounds' varying levels of potency against cancer cells.
To validate our hypothesis, results were compared with an earlier study by Kim et al. [86]. Disturbance on ring A, by removal of methoxy and hydroxy group on C8 and C5, respectively led to a significant drop in cytotoxic effect. In a similar treatment duration (24-hour), 50 μM of sinensetin (5,6,7,3’,4’-PeMF) reduced cell viability by only 30%, with no IC50 reported. Despite a longer treatment period (48h) was employed, it’s worth discussing the cytotoxicity effect of sudachitin due to its unique conformation [87]. While it retained the 5-hydroxy group, justifying a strong IC50 (49.32 μM), and hydroxylation was also observed on position C7 and C4’ on rings A and B, respectively. Note that as 24-hour treatment was not reported, we assumed the activity of sudachitin might be lower if the cytotoxic effect was in a time-controlled condition. The result, however, appeals to our interest in exploring the activity of C7-OH given its strong cytotoxicity activity on par with the C7-OCH3 methoxylated flavone analogs (Figure 8). Our analysis shows that current C7-OH methoxylated flavones demonstrate varied cytotoxicity strength against cancer cell lines [67,88]. Acacetin, 5,7-dihydroxy-4’-methoxyflavone was one of the earlier mono-methoxyflavones studied in anticancer that may untangle the potency of C7-OH. Acacetin reduced cell viability with an estimated IC50 of 25 µM after 24-hour treatment [89]. The variation may exist due to the preferential of specific moieties and the pathway that leads to the reduction of cell viability. Interestingly, the slight variation in IC50 values of the 5-demethylnobiletin may attributed to the different pathways leading to cell death. The strongest IC50 was observed with 5,3’-didemethylnobiletin which induced apoptotic cell death via cleavage on caspase-3 and cell cycle arrest at the G2/M phase [84]. In contrast, its methoxylated counterpart, sinensetin reduced cell viability by inducing autophagic cell death [86]. With muted expression of pro- and anti-apoptotic markers with early indicator of apoptotic cell death was not detected, Increased levels of the autophagic marker, beclin-1, and LC3B-II, along with downregulation of p62, confirmed the cell death mechanism of sinensetin. Simultaneous activation of the upstream autophagic marker, p-AMPK, and weakened p-mTOR by sinensetin treatment initiate autophagy cell death.
2.5. Acute & Chronic Myeloid Leukemia
FMS-like tyrosine kinase 3 (FLT3) is notable overexpressed in AML, making the inhibition of this kinase a critical focus in leukemia therapy [90,91]. FLT3 activation facilitated through its extracellular ligand-binding domain, triggers downstream protein cascades that stimulate cell proliferation and differentiation. However, it is a challenge in AML due to the aggressive nature of this cancer. Given the high FLT3 levels, the exploration of flavone derivatives as potential inhibitors is vital. FLT3-internal tandem duplication (FL3-ITD) positive cell lines such as MOLM-13 and MV4-11 serve as principle models to investigate drug resistance mechanism against tyrosine kinase inhibitors [92]. An in-vitro study by Yen et al. identified four flavone derivatives with potent cytotoxic activity in these cell lines over 72-hour treatment period:
- 5,7-dihydroxy-4'-MF (IC50: 9.1 & 6.8 µM)
- 5,4’-dihydroxy-6,7-DMF (IC50: 2.6 & 2.6 µM),
- 5,7-dihydroxy-6,4'-DMF (IC50: 5.9 & 7.9 µM) and
- 5,7,4’-trihydroxy-6-MF/Hispidulin (IC50: 7.0 & 6.8 µM),
The compound 5,4’-dihydroxy-6,7-dimethoxyflavone demonstrated the strongest cytotoxic effect, notably due to the C7-OCH3 (Table 3). Likewise, substituting the hydroxy group on C7 disrupts the lipophilic capacity with rich electron delocalization region, which is crucial for ring A. Upon C7 demethylation, pharmacophore changes on C6 and C4’s position are incapable of reversing the cytotoxicity potential.
2.5.1. Acute and Chronic Myeloid Leukemia: C5-OH as a Promising Methoxyflavones’ Anticancer Backbone
C5-OH has showcased strong evidence as a backbone for an anticancer marker within the flavone class. Inevitably, researchers are keenly focused on the demethylation of C5 in nobiletin structure to develop targeted anticancer strategies. According to Chen et al. [82], 5-demethyl nobiletin exhibited outstanding antiproliferative effects against multiple acute myeloid leukemia (AML) cell lines including HL-60, THP-1, U-937, and HEL as well as the chronic myeloid leukemia (CML) cell line, K562. When treated for 48 hours, this compound demonstrated IC50 values of 85.7, 32.3, 30.4, and 65.3 μM, respectively against all AML cell lines, compared to a vehicle control (0.1% DMSO).
2.5.2. Diverse Mechanisms in Different Cell Lines
A comprehensive review by Skopek et al. [93], pointed out that the cell death pathway of HL-60 is primarily driven by the apoptotic proteins [94]. In contrast, the survival of THP-1 and U-937 cells is largely controlled by the inflammatory markers [95], while proliferation in HEL cell lines is stimulated by the JAK-STAT3-ERK1/2 signaling pathway [96,97]. These pathways highlight the multi-target of flavones which contribute to strong IC50 values across all cancerous cells.
2.5.3. Comparative Efficacy and Binding Preferences
Notably, similar treatment on K562 CML cell lines yielded a slightly weaker IC50 value of 91.5 μM. Astonishingly, an earlier study by Yen et al. demonstrated a stronger IC50 value of 82.49 μM for nobiletin under comparable conditions, suggesting that selective binding associated with C5-OCH3 could be advantageous as observed in studies involving triple-negative breast cancer, pancreatic (MIA paca-2), and skin cancer cell lines. With limited studies on flavone families, methoxy moieties were reported to be highly favored compared to other cancer cell lines with greater lipophilic character and antiproliferative effect on K562 cell lines [99,100]. Both studies highlighted the contribution of the lipophilic region in facilitating the transport of active compounds to target proteins.
2.5.4. Casticin's Distinct Cytotoxic Profile
Nevertheless, 5,3’-dihydroxy-3,6,7,4’-TeMF, casticin, was investigated for its cytotoxic effects on WEHI-3 mouse leukemia cell lines where it reduced cell viability moderately to 53.4% at a concentration of 1.0 μM for 48-hour treatment, corresponding to an IC50 less than 1 μM, markedly more potent than its effects observed on DU145 prostate cancer cell lines [101]. The inhibitory activity reflects the selective characteristic of the flavones compound, which requires high specificity to induce a potent anti-tumor effect by targeting cell death protein markers.
2.6. Gastric Cancer
Three polymethoxyflavones; nobiletin, 5-demethyl nobiletin, and tangeritin have exhibited potent antiproliferative activity against several gastric cancer cell lines including AGS, BGC-823, and SGC-7901 (Table 3) [102]. In 48-hour treatment, tangeritin emerged as the most effective, reducing AGS cell viability to below 20% at an estimated concentration of 33.57 µM. However, its efficacy was moderate on the same cell lines maintaining about to ~30% viability at a concentration of 62.13 µM. In contrast, both nobiletin, 5-demethyl nobiletin showed no activity at concentrations up to 100 µM on both BGC-823 and SGC-7901 cell lines, and 5-demethyl nobiletin also was inactive on all tested cell lines.
2.6.1. Physicochemical and SAR Insights
Physicochemical and structure-activity relationship (SAR) analysis indicates that the C5-OH group is generally unfavorable against all tested gastric cancer cell lines although showing strong activity in other cancerous cell lines. This inconsistency underscores the need for further pharmacophore studies to fully understand the interactions. Interestingly, C5-OCH3 appears to maintain stable hydrogen bond interactions with protein markers, enhancing the cytotoxic potential of tangeritin significantly.
2.6.2. Molecular Interactions and Anticancer Effects
Based on our schematic diagram analysis, the heavy substitution of the methoxy group on ring B does diminish the cytotoxicity capacity of nobiletin compared to tangeritin. In addition, the presence of C5-OCH3 may facilitate the interaction of the polar carbonyl surface with the target site through an electron transfer/hydrogen acceptor mechanism [103], which is crucial for stronger binding with protein markers. This mechanism demonstrates how specific functional groups significantly affect the binding strength and overall anticancer efficacy of these compounds.
2.7. Skin Cancer
Among eight methoxyflavones isolated from Gardenia oudiepe (Table 3), only 2 PMF displayed a strong antiproliferative effect against A2058 melanoma cell lines [104]. Treatment with compounds 5,7-dihydroxy-3,6,4'-TMF (P1) and 7,5'-dihydroxy-3,6,3',4'-TeMF (P2) at 10 µM for 72-hour significantly reduced cancer cell growth by 66.52% and 42.86%, respectively. Meanwhile, both compounds 5,7,4’-trihydroxy-3,6-DMF (P3) and 5,7-dihydroxy-3,6,3’,4’,5’-PeMF (P4) showed the least cytotoxic potential due to amphipathicity disruption on ring B. The A2058 melanoma cell lines are highly invasive and metastasized, and having extra lipophilic and hydrophobic characteristics on ring B was expected to improve the cytotoxic effect of the methoxyflavones analogs.
The substitution of the methoxy group (P1) with hydroxy group (P4) at position C4’ reduced the hydrophobic strength on ring B and diminished its growth inhibition effect significantly from 66.52% to 18.55% under the same concentration and treatment duration. Nevertheless, the neighboring effect of a methoxy group (C4’) and a hydroxyl group (C5’) may triggered the formation of IHB, stabilizing the compound and improving its hydrophobic capacity with the addition of C3’-OCH3 (P2) despite the intense bulkiness within ring B. On another note, additional C5' methoxy (5,7-dihydroxy-3,6,3',4',5'-PeMF) replacing the hydroxy group reversed the cytotoxic effect and significantly reduced its effectiveness with >90% cancer cell viability. The massive hydrophobic effect introduced by the addition of a trimethoxy group on ring B without possible interaction from adjacent functional group is the main contributing factor in its reduced efficacy. However, both compounds P1 and P2 induced potent inhibitory effect with strong IC50 values of 3.92 and 8.18 μM, respectively through apoptotic cell death mechanism involving the cleavage of caspase-3.
2.8. Oral Cancer
Five methoxyflavone compounds isolated from Hottonia palustris were tested on SCC-25 squamous carcinoma cell line over 24- and 48-hour demonstrating distinct antiproliferative activity [105]. Of these, only one isolated PMF (5,6′-dihydroxy-2′,3′-DMF) showed potent cytotoxic effects with IC50 values of 78.2 and 40.6 µM, on respective duration treatment. Meanwhile, 5-hydroxy-2′-MF, 5-hydroxy-2′,6′-DMF, 5-hydroxy-2′,3′,6′-TMF and 5,2’-dihydroxy-6’-MF were ineffective with IC50 values exceeding 200 µM (Table 3).
2.8.1. Structural Insights and SAR Analysis
Limited SAR analysis suggests that a balance between both hydrophobic and polar interactions across rings A and B is crucial. Based on our screening, these compounds are notable among flavones derivatives for having methoxy or hydroxy groups bonded to the C6’ position on ring B. Based on the cytotoxic effect, it's interesting to highlight the structural similarities between 5,6′-dihydroxy-2′,3′-DMF (IC50: 40.6 µM) and 5-hydroxy-2′,3′,6′-TMF (IC50 > 200 µM). Position C6' was hypothesized to significantly influence the bioactivity of the isolated PMF on SCC-25 cell lines. Early screening demonstrates the favorable C6’-OH position, however, more SAR analysis is required to confirm the pharmacophore that contributes to a strong cytotoxicity effect.
2.8.2. Hydroxylation and its Impact
Unlike the active compound 5,6′-dihydroxy-2′,3′-DMF, hydroxylation on ring B was not observed in the other screened compounds, which is a crucial moiety for C5-OH methoxyflavones scaffold. Further investigation will assist in determining if the C6’-OH has a distinct feature particularly as the position C2’,4’, 6’-OH align within the golden triangle, optimizing free radicals electron delocalization. This configuration potentially contributes to IHB and enhances the polar surface region’s interaction with target proteins.
2.9. Bile duct and Pancreatic Cancer
In bile duct cancer, sudachitin (5,7,4'-trihydroxy-6,8,3'-TMF) significantly reduces cancer cell viability with lower IC50 values of HuCCT1 (53.21 µM) and RBE (24.1 µM) in 48-hour treatment (Table 3). As discussed previously, sudachitin is well known for its potent activity in various cancer types, including colon and pancreatic cancers. Nevertheless, the detailed cell death mechanisms have remained scarce and have not been investigated up to the recent studies. In contrast, nobiletin (5,6,7,8,3',4’–HeMF) unable to reduce cell viability in both TFK1 and RBE cell lines, even at concentrations up to 100 µM in both 24- and 48-hour treatments [106]. As previously discussed, inactivity of nobiletin against certain cancer cell lines, including breast and prostate cancer cell lines is attributed to its massive lipophilic effect and selective cell death pathways. The GSK3β pathway is identified as contributing to nobiletin’s antiproliferative effects based on western blot analysis. Resembling the bile duct cancer, sudachitin shows potent activity against MIA PaCa-2 and PANC-1 pancreatic cancer cell lines, in 48-hour treatment (IC50: 43.35 & 32.73 µM) [87]. Meanwhile, nobiletin displays weak antiproliferative effects on these cells with PANC-1 cell viability remaining above 80% even after treatment at the highest concentration (120 μM) and longest duration treatment (72-hour) [107]. Nonetheless, nobiletin shows significant cytotoxicity effects of MIA PaCa-2 cells after 72-hour treatment, reducing cell viability to less than 20% at 80 μM. The selective cytotoxicity of nobiletin may be attributed to the heterogeneity of the cells, with PANC-1 being reported to be more invasive and have a greater metastatic effect than MIA PaCa-2 cells.
2.10. Cervical and Ovarian cancer
2.10.1. Cervical Cancer and HPV Infection
The growth of cervical cancer cell lines has been associated with human papillomaviruses (HPVs) infections, notably its subgroup, HPV18+ (Hela) and HPV16+ (C33A), meanwhile, SiHa cells are HPV negative (HPV-) [108]. The HPV oncoproteins E6 and E7 are known to facilitate malignant tumor conversion by disrupting the cancer cell death markers, which is evident in HPV18+ Hela cells [109,110]. In both studies reported by Zhang et al. [111,112], compound 6-methoxyflavone exhibited potent cytotoxicity against cervical cancer cells. Over a 72-hour treatment period, this compound reduced Hela cell viability the most (IC50:55.31 µM), followed by C33A (109.57 µM and SiHa (208.53 µM) (Table 3). Although compound 6-methoxyflavone was the most potent to Hela, its IC50 value was considered moderate, compared to the other cancer cell lines. Conversely, another methoxyflavones, xanthomicrol, 5,4'-dihydroxy-6,7,8-TMF was less potent against Hela cells with an IC50 exceeding 100 µM in a 72-hour treatment [83]. Due to the distinct pharmacophore, SAR predictions on methoxyflavone selectivity against cervical cancer cell lines based on recent studies were found to be inadequate.
2.10.2. Ovarian Cancer and Treatment Efficacy
With limited studies utilizing methoxyflavones scaffold, the SAR profiling is limited on targeting ovarian cancer cell lines. Based on Table 3, treatment with wogonin on all cisplatin sensitive (SKOV3 and OV2008) and its respective resistance counterparts (SKOV3/DDP and C13*) ovarian cancer cell lines for 72 hours at doses up to 20 µM demonstrated a weak antiproliferative effect with cell viability remaining above 80% [113]. Nonetheless, at the highest concentration (160 µM), wogonin significantly reduced all cell viabilities to 20%. The induction of apoptotic cancer cell death was proposed by the presence of DNA laddering and confirmed through the downregulation of Akt phosphorylation.
3. Mechanism of Methoxy and Hydroxy Flavones Derivatives
3.1. Flavones modulate the apoptotic cell death pathway
Briefly, recent studies on the antitumor mechanism of flavones derivatives depict a multi-strategy approach. These compounds leverage the versatile positions available for hydroxy and methoxy substituents in flavones’ molecular structure, influencing various carcinogenic pathways. Apoptosis or programmed cell death has been a hallmark mechanism employed by flavones to prevent oncogenesis through both extrinsic and intrinsic mitochondrial pathways. The activation of mitochondrial outer membrane permeabilization (MOMP) is crucial in stimulating the expression of apoptotic proteins. Caspase-3, a critical player in apoptosis is often due to its central role in both mitochondrial pathways that leading to cell death [40,117,118,119]. For instance, 6-methoxyflavone induces apoptosis via the extrinsic or death receptor pathways, activating receptors such as Fas and tumor necrosis factor (TNF). Prior to caspase-3 cleavage, this is followed by the upregulation of the tumor suppressor p53 in the intrinsic mitochondrial pathway occurred due to mutation in the Tp53 gene, given its primary roles in cell cycle arrest, DNA repair, and apoptosis. Flavones derivatives are known to activate the Bcl-2 family’s protein, which comprises pro-apoptotic, anti-apoptotic, and BH3-only proteins, controlled by the expression level of p53 [81,111,112,115,120]. Elevated levels of pro-apoptotic proteins such as BID, BAX and BAK alongside the downregulation of anti-apoptotic proteins like MCL-1, Bcl-2, and Bcl-xL in the intrinsic mitochondrial pathway resulted in activation of caspase-3, -8 and -9, prior to cell death [46,63,101,102]. In some cases, the variation of pro- and anti-apoptotic protein expression levels induced the release of cytochrome C, subsequently inhibiting APAF-1 mRNA expression before caspase cleavage [111,112].
3.2. Flavones-Induced Cell Cycle Arrest
Flavones not only induce apoptosis but also impact cell cycle regulation through the tumor suppressor protein p53. The transcription factor p53 is a central tumor-blocking protein that activates dozens of downstream genes that modulate cell cycles. Numerous studies have illustrated the affiliation of p53 with its transcriptional target, p21Waf1/CIP1 in suppressing cyclins and cyclin-dependent kinase (CDK) expression by forming inhibitory complexes, that lead to cell cycle arrest [121,122]. Flavone derivatives such as 5,4'-dihydroxy-6,7,8-TMF and 5-demethyltangeritin by activation of p21Waf1/CIP1 via transcriptional target of P53 arrest cell cycle at the G0/G1 phase, consequently inhibiting cyclin D and CDK4 expression [81,106]. Nobiletin is known for its antitumor mechanism as it also induces cell cycle arrest. Concurrently, Lotfizadeh et al. and Yen et al. depict similar behavior of calycopterin and nobiletin, respectively by inducing cell cycle arrest at the G1 phase, with nobiletin specifically activating p21 and p27 expression, subsequently suppressed the cyclin D mRNA expression. Nevertheless, modifying nobiletin by replacing methoxy group with hydroxy at the C5 position (5-demethylnobiletin) notably influences the cell cycle arrest, shifting the cell cycle arrest towards the G1/S phase with weaker expression of cyclin E and A mediated by p21 [82]. Meanwhile, 6-methoxyflavone depicts cell cycle arrest at the S phase through p21Waf1/CIP1 activation resulting in suppression of cyclin A and CDK2 activities [111,112].
3.3. Mitogen-activated protein kinase (MAPK) pathway
The mitogen-activated protein kinase (MAPK) signalling pathway, centred around the RAS oncogene GTP-binding protein has been targeted for colorectal and pancreatic cancer therapies since the late 1970s [123,124]. The pathway involves the inhibition of RAS upstream novel target, SOS-1, a guanine nucleotide exchange factor. This inhibition prevents the conversion of GDP to GTP by RAS assisted by GRB2 (growth factor receptor bound protein 2), thus blocking RAF dimerization and subsequent downstream signalling [125]. Over 85% of cancers are marked by abnormal activity of MAPK, emphasizing the need for targeted intervention by suppressing the mitogen-activated proteins. The activation of SOS-1 triggers a series of interactions among RAF, MEK, and ERK1/2, resulted in the abnormal activation of the MAPK pathway [126]. Casticin has been shown to inhibit both SOS-1 and GRB2, reducing phosphorylation levels of ERK1/2, a downstream component of the MAPK pathway, indicating suppression of MAPK signalling [65,68]. Within the cytosol, casticin, has been shown inhibit both SOS-1 and GRB2, reducing phosphorylation levels of ERK1/2 level, a downstream component of the MAPK pathway, indicating suppression of MAPK signalling [65,68]. This inhibition also leads to suppression of c-myc, a transcription factor with positive correlation with this pathway [127]. Moreover, while regulating MAPK signalling, RAS and GRB2 are well-known to activate PI3K/Akt pathway, contributing to oncogenic behavior and cancer progression [128]. Stimulation of both oncogenes-induced RAS-activated PI3K together with the amplification of mutation on its major effector, AKT and mTOR drives the malignant resistance and tumor progression (Glaviano et al., 2023). Methoxyflavones derivatives like wogonin and casticin are noted for inhibiting angiogenesis by impacting these critical pathways, highlighting their potential in therapeutic strategies against cancer [68,113].
3.4. NF-κB Signaling
NF-κB, a key transcription factor plays a critical role in inflammation and cancer by promoting tumorigenesis through the activation of various genes including IκB, p65, and p50. This pathway, involved in both canonical nor non-canonical pathways, is crucial for DNA repair, drug resistance, inhibition of apoptosis, and promotion of cell proliferation [130,131]. Various studies consistently link NF-ĸB to cancer progression, notably its role in resisting TNF-α-induced apoptosis by enhancing ROS levels and antioxidant enzyme activities, thereby preventing cell death [132].
Methoxyflavones derivatives have been identified as anti-inflammatory agents that can moderate the activation of NF-κB triggered by LPS- and TNF-α as demonstrated by numerous studies [133,134]. Concurrent investigation on the activity of PARP-1 nuclear enzyme was important given its roles in DNA repair and reversing cancer cell death by triggering NF-ĸB expression [135]. Thus, the inhibition of PARP-1 have been shown to promote apoptosis, effectively leading to cancer cell death as discussed by Chen et al. [82]. For instance, phosphorylated NF-κB (p-p65) were markedly downregulated in LPS-induced cancer cells while TNF-α-induced NF-κB expression was suppressed with upregulation of IκBα [114]. Nonetheless, treatment with flavones has been associated with an overexpression of PARP-1 which suggests potential unique mechanism of action. This overexpression leads to an increase in Poly(ADP-ribosyl)ation (PAR) synthesis during the early apoptosis, subsequently activating caspase-3 and initiating the proteolysis of PARP-1, which contributes to the strong cytotoxic effects observed in Hela cell viability [136].
3.5. Matrix Metalloproteinase (MMP) cascades signaling
Cancer-associated fibroblast (CAF) is one of the largest groups in the matrix component and part of the tumor microenvironment and plays crucial role in cancer progression [116]. These cells, when activated from normal fibroblast within adipose tissue notably by cancer cells such as 4T1 breast cancer cell line, exhibited increased mRNA expression of fibroblast biomarker, alpha-smooth muscle actin (α-sma), vimentin, and others [116]. This interaction between CAFs and tumor cells contributes to the micro-interaction between the CAF and tumor cell, contributes to the development of highly metastatic and invasive cancer cells.
Research has shown that PMF, specifically oroxylin A (5,7-dihydroxy-6-methoxyflavone), successfully reduced the mRNA expression of α-sma, vimentin, and fibronectin without significant toxicity in killing normal cell L-929 and normal primary cell, NBF, indicating selective efficacy [116]. Furthermore, CAF activation leads to higher invasiveness in the 4T1 cells, which PMFs have been potently downregulating the mRNA expression of MMP-2, MMP-9, and MMP-14 in a dose-controlled manner. The inactivation of CAF by PMF is facilitated by targeting ACTN1 through interaction at ASP-781, as revealed by molecular docking analysis. This interaction leads to a significant reduction in the phosphorylation of downstream signaling pathway, including focal adhesion kinase (FAK), janus kinase 2 (JAK2), the mammalian target of rapamycin (mTOR), and signal transducer and activator of transcription 3 (STAT3), thereby inhibiting key process that promote cancer cell migration and invasion.
3.6. Autophagy in Cancer
Autophagy, a self-degradation process essential for removing damaged organelles and other intracellular components, involves the formation of autophagosomes that fuse with lysosomes for decomposition and recycling [137]. In cancer, autophagy plays a dual indefinite roles: it suppresses cancer growth in the earlier stage by eliminating defective components that could promote malignancy, and it may support tumor growth at later stages by aiding cancer cell survival under stress [138].
One of the critical functions of autophagy in cancer is its ability to prevent the accumulation of ROS that leads to oxidative stress and DNA damage. This prevention is achieved by blocking the lipidation of LC3 protein, converting it from LC3-I to LC3-II via ATG4, an enzyme essential for autophagosome formation [139,140]. An extensive review by Zhao et al. discussed the regulatory roles of mTOR-ATG4- LC3-II pathway in regulating ROS levels and autophagy. A study by Wang et al. explored the potential antiproliferative effects of hispidulin, a compound known to influence this pathway, on DU145 and VCaP prostate cancer cell lines. Hispidulin potency in reduced cell migration and invasion after 48-hour treatment, suggesting its potential to elevate autophagy in these cancer cells. Evidence of increased rate of autophagy flux, characterized by the conversion of LC3 I to LC3 II, and elevated PPARγ expression, in tandem with weak phosphorylation of mTOR and S6K1, supports the hypothesis that hispidulin can modulate autophagy to suppress tumor progression.
3.7. ER-Stress-Induced Apoptosis By Flavone
The progression of malignant tumors is often influenced by the stress induced within the cells intrinsically and extrinsically resulting to disruption within the tumor microenvironment [142]. One significant source of cellular stress is the endoplasmic reticulum (ER), where irregularities and disruptions in protein folding can activate stress-induced signalling pathways. These pathways not only promote cancer cell growth but also enhance metastatic potential [143,144]. A key component in the ER stress response is the PERK signalling pathway. PERK plays vital role in mediating apoptosis through the mitochondrial pathway via BAX/Bcl-2 regulation or through triggering autophagy by activating downstream markers. Recent studies have shown that flavone derivatives can induce autophagy through elevated mRNA expression of downstream proteins such as PERK, EIF2α, IRE1α, ATF4, and CHOP [111,112]. This enhanced autophagic response to ER stress presents a significant mechanism by which flavones may exert their anticancer effects, providing a dual approach to controlling tumor progression through both apoptosis and autophagy.
3.8. Targeting Topoisomerase Enzymes In Highly Expressed Cancer Cells
Topoisomerases I and II are critical enzymes in DNA replication and transcription, and they are often highly expressed in various cancers including breast cancer [145]. Methoxyflavones derivatives have been identified as potent inhibitors of these enzymes exploiting their ability to interfere with unwinding process of DNA, leading to cell death [146]. For instance, chrysosplenetin, a specific methoxyflavones derivative has shown a strong inhibitory effect on topoisomerase I [48]. The strong potency of the compound may be attributed to the targeting inhibition of the topoisomerase enzyme. The strong inhibitory effect of chrysosplenetin toward topoisomerase I leads to a higher form I percentage in concentration concentration-dependent manner. The Form I of pBR322 plasmid DNA increased from 31.0% to 69.0%, whereas the percentage of Forms II and III declined. Meanwhile, topoisomerase II was inhibited by the compound in a similar concentration with the form I accumulation increased to 96.9% at the highest concentration. These findings highlight the potent anticancer properties of methoxyflavones derivatives through their targeted action against topoisomerase enzymes providing a promising avenue for therapeutic intervention in cancers with high topoisomerase expression.
3.9. Wnt-β-Catenin Pathway
The Wnt-β-catenin signaling pathway is critically involved in uncontrolled cancer cell proliferation, especially noted in colorectal cancer cells. Abnormalities in this pathway contribute to the cancer's invasive and metastatic characteristics [147]. Dysregulation of the key signaling cofactor β-catenin is known to influence the development of a cancer stem cell phenotype, characterized by high levels of cyclin D and c-myc, which are crucial for tumorigenesis [148]. The activation of this pathway leads to the elevated expression of downstream Wnt target genes which includes axin2 and vimentin which drive the cancer progression [149]. Studies using the RT-PCR have shown that tetramethoxyflavone specifically 5,7,3',4'-TeMF & 7,8,3',4'-TeMF can downregulate the mRNA expression of c-Myc, axin2, and vimentin in HCT116 colorectal cells at a concentration of 30 µM [80].
4. Conclusions
The multi-target of methoxyflavones has contributed to various biological activities in recent in-vitro studies. Being naturally abundant, the flavonoid subclass could exist in a variety range of scaffolds with distinct features. Our review indicates major parts of the methoxyflavones scaffold have not been fully explored with limited findings. The pharmacophore of the anticancer effect of methoxyflavones was centered on ring A, strengthened by C3’ and C4’ ring B, with few compounds bearing C2’, C5’ and C6’ moieties. The superiority of the anticancer effect of methoxyflavones against other natural flavones such as hydroxyflavones and the flavones glycosides was established. The methoxy group governed the lipophilic capacity and facilitated drug-membrane transfer, thus preserving the bioavailability of the scaffolds. The lipophilic character of methoxyflavones was the main contribution to a remarkable activity against various cancer cell lines, based on our analysis. Additionally, critical investigation summarized the methoxyflavones themselves could not sustain a stronger cytotoxic effect of IC50 less than 20 µM without the presence of the hydroxy group. The preservation of lipophilic properties on ring A flavones, with a balance capacity between a polar surface and lipophilic effect on ring B is the pathway to achieve stronger IC50. Generally, methoxy moieties occupied at least two out of three positions from the C6, 7, and 8 positions, and the addition of hydroxy group on position C5 was fundamental to stronger anticancer activity. Next, having hydroxy moieties alongside with methoxy group on either C3’ or C4’ arrangement could amplify the cytotoxic capacity. Both methoxy or hydroxy groups in the respective position on ring B could achieve similarly strong IC50, yet it came down to the conformational changes on rings A & B and on the cell types. Some C5-OCH3 derivatives could induce strong IC50, with few exceptions such as a maximum of single hydroxy or methoxy group on ring B. To maximize interaction with the target protein marker, balancing between the methoxy and hydroxy moieties in terms of the number and position of both groups in ring A and B flavones is crucial. Additionally, the collaborative efficacy of both groups will maximize the resonance effect and electron delocalization of free radical flavones and initiate the formation of IHB. Concurrently, both interaction mechanisms stabilized and preserved the lipophilicity while simultaneously offering an optimal polar surface for stronger interaction with the cancer cell death marker. The review solely focused on the pharmacophore specificity of methoxyflavones analogs in-vitro, with other factors such as drug metabolism, bioavailability, and toxicity, which may influence the resultant efficacy. The review may narrow down future research for targeting and isolating active natural methoxyflavones and assisting in the synthetic designing of flavones scaffolds with favorable positions of both methoxy and hydroxy moieties.
Author Contributions
“Conceptualization, M.A. and M.A.M.; methodology, M.A.; validation, M.A.; formal analysis, M.A. and M.A.M.; investigation, M.A.; resources, M.A.M., R.R., N.N.N.R., S.G.H.T and S.H.A; data curation, M.A.; writing-original draft preparation, M.A.; writing-review and editing, M.A. and M.A.M.; visualization, M.A.; supervision, M.A.M. and R.R.; project administration, M.A. and M.A.M.. All authors have read and agreed to the published version of the manuscript.”.
Funding
This article review was funded by the Ministry of Higher Education, Fundamental Research Grant Scheme, FRGS/1/2021/SKK0/MSU/02/1 and the MSU Publication & Conference Grant, (MPCG-004-022023-SGS) from the Management & Science University, Shah Alam, Malaysia.
Acknowledgments
We are grateful to the contribution by all co-authors and research members towards the completion of this article review.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
SAR; structural-activity relationship; HBA; hydrogen bond acceptor; HBD; hydrogen bond donor; OCH3; methoxy group; OH; hydroxy group; IHB; intramolecular hydrogen bond; MF; methoxyflavone; DMF; dimethoxyflavone; TMF; trimethoxyflavone; TeMF; tetramethoxyflavone; PeMF; pentamethoxyflavone; HeMF; hexamethoxyflavone; Her-2+, epidermal growth factor receptor 2; AML; acute myeloid leukaemia; CML; chronic myeloid leukaemia; logP; lipid partition coefficient; logD; distribution coefficient; TNFR; tumor necrosis factor receptor; MCL-1; Myeloid cell leukemia-1; BID; BH3 interacting-domain death agonist; BAX; Bcl-2-associated X protein; APAF-1; Apoptotic protease activating factor-1; CDK; cyclin-dependent kinase; MAPK; mitogen-activated protein kinase; GDP; guanosine diphosphate; GTP; guanosine triphosphatase; RAF; rapidly accelerated fibrosarcoma; MEK; mitogen-activated protein kinase kinase; ERK1/2; extracellular signal-regulated kinase 1/2; GRB2; Growth Factor Receptor-bound protein 2; PI3K; phosphatidylinositol 3-kinase; AKT; protein kinase B; mTOR; mammalian target of rapamycin; NF-κB; nuclear factor kappa B; kappaB kinase; LPS; lipopolysaccharide; TNF-α; tumor necrosis factor alpha; PARP-1; Poly (ADP-ribose) polymerase 1; CAF; cancer-associated fibroblast; αSMA ; alpha smooth muscle actin; MMP; matrix metalloproteinase; FAK; focal-adhesion kinase; JAK2; janus kinase 2; STAT3; signal transducer and activator of transcription 3; ACTN1; alpha-actinin-1; LC3; microtubule-associated protein light chain 3; PPARγ; peroxisome proliferator-activated receptor-gamma; ER; endoplastic reticulum; PERK; protein kinase R-like endoplasmic reticulum kinase; EIF2α. eukaryotic translation initiation factor 2 alpha; IRE1α; inositol-requiring enzyme 1 alpha; ATF4; activating transcription factor 4; CHOP; C/EBP homologous protein
References
- Lewandowski, W.; Lewandowska, H.; Golonko, A.; Świderski, G.; Świsłocka, R.; Kalinowska, M. Correlations between molecular structure and biological activity in "logical series" of dietary chromone derivatives. PLOS ONE 2020, 15, e0229477. [Google Scholar] [CrossRef] [PubMed]
- Mutalib, M. A., Shamsuddin, A. S., Ramli, N. N. N., Tang, S. G. H., & Adam, S. H. (2023, May 1). Antiproliferative Activity and Polyphenol Analysis in Tomato (Solanum Lycopersicon). https://malaysianjournalofmicroscopy.org/ojs/index.php/mjm/article/view/737.
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. BioMed Res. Int. 2022, 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brodowska, K.M. Natural flavonoids: Classification, potential role, and application of flavonoid analogues Eur. J. Biol. Res. 2017. [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Goel, N.; Garg, A.; Nagendra, C.; Reddy, A.M.; Biswas, R.; Natarajan, R.; Jaisankar, P. In-vitro and in-silico cholinesterase inhibitory activity of bioactive molecules isolated from the leaves of Andrographis nallamalayana J.L. Ellis and roots of Andrographis beddomei C.B. Clarke. J. Mol. Struct. 2024, 1301, 137406. [Google Scholar] [CrossRef]
- Le, H.L.; Nguyen, V.H.; Nguyen, T.D.; Nguyen, T.V.A.; Le, D.H. Potential antiaggregatory and anticoagulant activity of Kaempferia parviflora extract and its methoxyflavones. Ind. Crop. Prod. 2022, 192, 116030. [Google Scholar] [CrossRef]
- Wang, D.; Li, Z.; Jiang, Z.; Li, Y.; Chen, Q.; Zhou, Z. Polymethoxylated flavone variations and in vitro biological activities of locally cultivated Citrus varieties in China. Food Chem. 2024, 463, 141047. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Y.; Chen, S.; Lin, J.; Bian, J.; Huang, D. Anti-Inflammation Activity of Flavones and Their Structure–Activity Relationship. J. Agric. Food Chem. 2021, 69, 7285–7302. [Google Scholar] [CrossRef]
- Zhao, L.; Yuan, X.; Wang, J.; Feng, Y.; Ji, F.; Li, Z.; Bian, J. A review on flavones targeting serine/threonine protein kinases for potential anticancer drugs. Bioorganic Med. Chem. 2019, 27, 677–685. [Google Scholar] [CrossRef]
- Moldoveanu, S. C., & David, V. (2022). Characterization of analytes and matrices. In Elsevier eBooks (pp. 179–205). [CrossRef]
- Nakhaee, S.; Ghasemi, S.; Karimzadeh, K.; Zamani, N.; Alinejad-Mofrad, S.; Mehrpour, O. The effects of opium on the cardiovascular system: a review of side effects, uses, and potential mechanisms. Subst. Abus. Treat. Prev. Policy 2020, 15, 1–13. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Patra, J.K.; Singh, Y.D.; Panda, M.K.; Das, G.; Adetunji, C.O.; Michael, O.S.; Sytar, O.; Polito, L.; et al. Paclitaxel: Application in Modern Oncology and Nanomedicine-Based Cancer Therapy. Oxidative Med. Cell. Longev. 2021, 2021, 1–24. [Google Scholar] [CrossRef]
- Xu, K.; Ren, X.; Wang, J.; Zhang, Q.; Fu, X.; Zhang, P.-C. Clinical development and informatics analysis of natural and semi-synthetic flavonoid drugs: A critical review. J. Adv. Res. 2023, 63, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, J.; Ma, L.; Li, J.; Shahzad, N.; Kim, C.K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci. Rep. 2020, 10, 2611. [Google Scholar] [CrossRef]
- Aidiel, M.M.; Maisarah, A.; Khalid, K.; Ramli, N.N.; Tang, S.; Adam, S. Polymethoxyflavones transcends expectation, a prominent flavonoid subclass from Kaempferia parviflora: A critical review. Arab. J. Chem. 2023, 17. [Google Scholar] [CrossRef]
- Chiodi, D.; Ishihara, Y. The role of the methoxy group in approved drugs. Eur. J. Med. Chem. 2024, 273, 116364. [Google Scholar] [CrossRef]
- Dong, X.; Li, X.; Ruan, X.; Kong, L.; Wang, N.; Gao, W.; Wang, R.; Sun, Y.; Jin, M. A deep insight into the structure-solubility relationship and molecular interaction mechanism of diverse flavonoids in molecular solvents, ionic liquids, and molecular solvent/ionic liquid mixtures. J. Mol. Liq. 2023, 385, 122359. [Google Scholar] [CrossRef]
- Qiu, T.; Wu, D.; Yang, L.; Ye, H.; Wang, Q.; Cao, Z.; Tang, K. Exploring the Mechanism of Flavonoids Through Systematic Bioinformatics Analysis. Front. Pharmacol. 2018, 9, 918. [Google Scholar] [CrossRef]
- Hawthorne, D.; Pannala, A.; Sandeman, S.; Lloyd, A. Sustained and targeted delivery of hydrophilic drug compounds: A review of existing and novel technologies from bench to bedside. J. Drug Deliv. Sci. Technol. 2022, 78, 103936. [Google Scholar] [CrossRef]
- Dash, S.; Rathi, E.; Kumar, A.; Chawla, K.; Kini, S.G. Identification of DprE1 inhibitors for tuberculosis through integrated in-silico approaches. Sci. Rep. 2024, 14, 1–28. [Google Scholar] [CrossRef]
- Lombardo, M.; G-Dayanandan, N.; Keshipeddy, S.; Zhou, W.; Si, D.; Reeve, S.; Alverson, J.; Barney, P.; Walker, L.; Hoody, J.; et al. Structure-Guided In Vitro to In Vivo Pharmacokinetic Optimization of Propargyl-Linked Antifolates. Drug Metab. Dispos. 2019, 47, 995–1003. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, W. Drug metabolism in drug discovery and development. Acta Pharm. Sin. B 2018, 8, 721–732. [Google Scholar] [CrossRef]
- Hao, Y.; Wei, Z.; Wang, Z.; Li, G.; Yao, Y.; Dun, B. Biotransformation of Flavonoids Improves Antimicrobial and Anti-Breast Cancer Activities In Vitro. Foods 2021, 10, 2367. [Google Scholar] [CrossRef]
- Štellerová, D.; Michalík, M.; Lukeš, V. Methoxylated flavones with potential therapeutic and photo-protective attributes: Theoretical investigation of substitution effect. Phytochemistry 2022, 203, 113387. [Google Scholar] [CrossRef]
- Li, S.; Pan, M.-H.; Lo, C.-Y.; Tan, D.; Wang, Y.; Shahidi, F.; Ho, C.-T. Chemistry and health effects of polymethoxyflavones and hydroxylated polymethoxyflavones. J. Funct. Foods 2008, 1, 2–12. [Google Scholar] [CrossRef]
- Spiegel, M.; Andruniów, T.; Sroka, Z. Flavones’ and Flavonols’ Antiradical Structure–Activity Relationship—A Quantum Chemical Study. Antioxidants 2020, 9, 461. [Google Scholar] [CrossRef]
- Messaadia, L.; Bekkar, Y.; Benamira, M.; Lahmar, H. Predicting the antioxidant activity of some flavonoids of Arbutus plant: A theoretical approach. Chem. Phys. Impact 2020, 1, 100007. [Google Scholar] [CrossRef]
- Huggins, D.J.; Sherman, W.; Tidor, B. Rational Approaches to Improving Selectivity in Drug Design. J. Med. Chem. 2012, 55, 1424–1444. [Google Scholar] [CrossRef]
- Cramer, J.; Sager, C.P.; Ernst, B. Hydroxyl Groups in Synthetic and Natural-Product-Derived Therapeutics: A Perspective on a Common Functional Group. J. Med. Chem. 2019, 62, 8915–8930. [Google Scholar] [CrossRef]
- Łodyga-Chruścińska, E.; Kowalska-Baron, A.; Błazińska, P.; Pilo, M.; Zucca, A.; Korolevich, V.M.; Cheshchevik, V.T. Position Impact of Hydroxy Groups on Spectral, Acid–Base Profiles and DNA Interactions of Several Monohydroxy Flavanones. Molecules 2019, 24, 3049. [Google Scholar] [CrossRef]
- Veiko, A.G.; Lapshina, E.A.; Zavodnik, I.B. Comparative analysis of molecular properties and reactions with oxidants for quercetin, catechin, and naringenin. Mol. Cell. Biochem. 2021, 476, 4287–4299. [Google Scholar] [CrossRef]
- De Souza, L.A.; Da Silva, H.C.; De Almeida, W.B. Structural Determination of Antioxidant and Anticancer Flavonoid Rutin in Solution through DFT Calculations of 1H NMR Chemical Shifts. ChemistryOpen 2018, 7, 902–913. [Google Scholar] [CrossRef]
- Parthiban, A.; Sachithanandam, V.; Lalitha, P.; Elumalai, D.; Asha, R.N.; Jeyakumar, T.C.; Muthukumaran, J.; Jain, M.; Jayabal, K.; Mageswaran, T.; et al. Isolation and biological evaluation 7-hydroxy flavone fromAvicennia officinalisL: insights from extensivein vitro, DFT, molecular docking and molecular dynamics simulation studies. J. Biomol. Struct. Dyn. 2022, 41, 2848–2860. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.; Kaur, P.; Shukla, S.; Abbas, A.; Fu, P.; Gupta, S. Plant flavone apigenin inhibits HDAC and remodels chromatin to induce growth arrest and apoptosis in human prostate cancer cells: In vitro and in vivo study. Mol. Carcinog. 2011, 51, 952–962. [Google Scholar] [CrossRef]
- Mohod, S.M.; Kandhare, A.D.; Bodhankar, S.L. Gastroprotective potential of Pentahydroxy flavone isolated from Madhuca indica J. F. Gmel. leaves against acetic acid-induced ulcer in rats: The role of oxido-inflammatory and prostaglandins markers. J. Ethnopharmacol. 2016, 182, 150–159. [Google Scholar] [CrossRef]
- Lima, J. C. S., De Oliveira, R. G., Silva, V. C., De Sousa, P. T., Jr, Violante, I. M. P., Macho, A., & De Oliveira Martins, D. T. (2018). Anti-inflammatory activity of 4′,6,7-trihydroxy-5-methoxyflavone from Fridericia chica (Bonpl.) L.G.Lohmann. Natural Product Research, 34(5), 726–730. [CrossRef]
- Phung, H.M.; Lee, S.; Hong, S.; Lee, S.; Jung, K.; Kang, K.S. Protective Effect of Polymethoxyflavones Isolated from Kaempferia parviflora against TNF-α-Induced Human Dermal Fibroblast Damage. Antioxidants 2021, 10, 1609. [Google Scholar] [CrossRef]
- Over, B.; McCarren, P.; Artursson, P.; Foley, M.; Giordanetto, F.; Grönberg, G.; Hilgendorf, C.; Lee, M.D.; Matsson, P.; Muncipinto, G.; et al. Impact of Stereospecific Intramolecular Hydrogen Bonding on Cell Permeability and Physicochemical Properties. J. Med. Chem. 2014, 57, 2746–2754. [Google Scholar] [CrossRef]
- Alex, A.; Millan, D.S.; Perez, M.; Wakenhut, F.; Whitlock, G.A. Intramolecular hydrogen bonding to improve membrane permeability and absorption in beyond rule of five chemical space. MedChemComm 2011, 2, 669–674. [Google Scholar] [CrossRef]
- Rusinska-Roszak, D. Energy of Intramolecular Hydrogen Bonding in ortho-Hydroxybenzaldehydes, Phenones and Quinones. Transfer of Aromaticity from ipso-Benzene Ring to the Enol System(s). Molecules 2017, 22, 481. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Charisiadis, P.; Primikyri, A.; Pappas, C.G.; Exarchou, V.; Tzakos, A.G.; Gerothanassis, I.P. Hydrogen bonding probes of phenol –OH groups. Org. Biomol. Chem. 2012, 11, 1013–1025. [Google Scholar] [CrossRef]
- Abraham, R.J.; Mobli, M. An NMR, IR and theoretical investigation of 1H Chemical Shifts and hydrogen bonding in phenols. Magn. Reson. Chem. 2007, 45, 865–877. [Google Scholar] [CrossRef]
- Whaley, W.L.; Okoso-Amaa, E.M.; Womack, C.L.; Vladimirova, A.; Rogers, L.B.; Risher, M.J.; Abraham, M.H. Summation Solute Hydrogen Bonding Acidity Values for Hydroxyl Substituted Flavones Determined by NMR Spectroscopy. Nat. Prod. Commun. 2013, 8, 85–98. [Google Scholar] [CrossRef]
- Huque, F. T. T., & Platts, J. A. (2003). The effect of intramolecular interactions on hydrogen bond acidityElectronic supplementary information (ESI) available: summary of retrained regression using DFT methods. Organic & Biomolecular Chemistry, 1(8), 1419–1424. [CrossRef]
- Carcache, P.J.B.; Eugenio, G.D.A.; Ninh, T.N.; Moore, C.E.; Rivera-Chávez, J.; Ren, Y.; Soejarto, D.D.; Kinghorn, A.D. Cytotoxic constituents of Glycosmis ovoidea collected in Vietnam. Fitoterapia 2022, 162, 105265–105265. [Google Scholar] [CrossRef] [PubMed]
- Al-Aamri, H.M.; Irving, H.R.; Bradley, C.; Meehan-Andrews, T. Intrinsic and extrinsic apoptosis responses in leukaemia cells following daunorubicin treatment. BMC Cancer 2021, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Şöhretoğlu, D.; Barut, B.; Sari, S.; Özel, A.; Arroo, R. In vitro and in silico assessment of DNA interaction, topoisomerase I and II inhibition properties of chrysosplenetin. Int. J. Biol. Macromol. 2020, 163, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, W.S.; Tarqui, S.; Huang, X.; Almanza, G.; Oredsson, S. Breast cancer cell line toxicity of a flavonoid isolated from Baccharis densiflora. BMC Complement. Med. Ther. 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Miyanabe, K.; Akiba, H.; Kuroda, D.; Nakakido, M.; Kusano-Arai, O.; Iwanari, H.; Hamakubo, T.; Caaveiro, J.M.M.; Tsumoto, K. Intramolecular H-bonds govern the recognition of a flexible peptide by an antibody. J. Biochem. 2018, 164, 65–76. [Google Scholar] [CrossRef]
- de Vicente, J.; Lemoine, R.; Bartlett, M.; Hermann, J.C.; Hekmat-Nejad, M.; Henningsen, R.; Jin, S.; Kuglstatter, A.; Li, H.; Lovey, A.J.; et al. Scaffold hopping towards potent and selective JAK3 inhibitors: Discovery of novel C-5 substituted pyrrolopyrazines. Bioorganic Med. Chem. Lett. 2014, 24, 4969–4975. [Google Scholar] [CrossRef]
- Sakamoto, T.; Koga, Y.; Hikota, M.; Matsuki, K.; Murakami, M.; Kikkawa, K.; Fujishige, K.; Kotera, J.; Omori, K.; Morimoto, H.; et al. Design and synthesis of novel 5-(3,4,5-trimethoxybenzoyl)-4-aminopyrimidine derivatives as potent and selective phosphodiesterase 5 inhibitors: Scaffold hopping using a pseudo-ring by intramolecular hydrogen bond formation. Bioorganic Med. Chem. Lett. 2014, 24, 5175–5180. [Google Scholar] [CrossRef]
- Ettorre, A.; D’andrea, P.; Mauro, S.; Porcelloni, M.; Rossi, C.; Altamura, M.; Catalioto, R.M.; Giuliani, S.; Maggi, C.A.; Fattori, D. hNK2 receptor antagonists. The use of intramolecular hydrogen bonding to increase solubility and membrane permeability. Bioorganic Med. Chem. Lett. 2011, 21, 1807–1809. [Google Scholar] [CrossRef]
- McDonagh, A.F.; Lightner, D.A. Influence of Conformation and Intramolecular Hydrogen Bonding on the Acyl Glucuronidation and Biliary Excretion of Acetylenic Bis-Dipyrrinones Related to Bilirubin. J. Med. Chem. 2007, 50, 480–488. [Google Scholar] [CrossRef]
- Dilek, B.; Meltem, . Quercetin suppresses cell proliferation using the apoptosis pathways in MCF-7 and MDA-MB-231 human breast carcinoma cells in monolayer and spheroid model cultures. South Afr. J. Bot. 2023, 162, 259–270. [CrossRef]
- Jin, H.; Lee, W.S.; Eun, S.Y.; Park, H.-S.; Kim, G.; Choi, Y.H.; Ryu, C.H.; Jung, J.M.; Hong, S.C.; Shin, S.C.; et al. Morin, a flavonoid from Moraceae, suppresses growth and invasion of the highly metastatic breast cancer cell line MDA-MB-231 partly through suppression of the Akt pathway. Int. J. Oncol. 2014, 45, 1629–1637. [Google Scholar] [CrossRef]
- Chien, S.-Y.; Wu, Y.-C.; Chung, J.-G.; Yang, J.-S.; Lu, H.-F.; Tsou, M.-F.; Wood, W.; Kuo, S.-J.; Chen, D.-R. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum. Exp. Toxicol. 2009, 28, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Vongdeth, K.; Han, P.; Li, W.; Wang, Q.-A. Synthesis and Antiproliferative Activity of Natural and Non-Natural Polymethoxychalcones and Polymethoxyflavones. Chem. Nat. Compd. 2019, 55, 11–17. [Google Scholar] [CrossRef]
- Pereira, D.d.A.; Sandim, V.; Fernandes, T.F.B.; Almeida, V.H.; Rocha, M.R.; Amaral, R.J.F.C.D.; Rossi, M.I.D.; Kalume, D.E.; Zingali, R.B. Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2. Int. J. Mol. Sci. 2022, 23, 10194. [Google Scholar] [CrossRef]
- Amić, D., Davidović-Amić, D., Bešlo, D. and Trinajstić, N. (2003). Structure-Radical Scavenging Activity Relationships of Flavonoids. Croatica Chemica Acta, 76(1), 55-61. https://hrcak.srce.hr/103057.
- Heijnen, C.; Haenen, G.; van Acker, F.; van der Vijgh, W.; Bast, A. Flavonoids as peroxynitrite scavengers: the role of the hydroxyl groups. Toxicol. Vitr. 2001, 15, 3–6. [Google Scholar] [CrossRef]
- Milenković, D.; Đorović, J.; Petrović, V.; Avdović, E.; Marković, Z. Hydrogen atom transfer versus proton coupled electron transfer mechanism of gallic acid with different peroxy radicals. React. Kinet. Catal. Lett. 2017, 123, 215–230. [Google Scholar] [CrossRef]
- Tsai, H.-Y.; Yang, J.-F.; Chen, Y.-B.; Guo, J.-L.; Li, S.; Wei, G.-J.; Ho, C.-T.; Hsu, J.-L.; Chang, C.-I.; Liang, Y.-S.; et al. Acetylation Enhances the Anticancer Activity and Oral Bioavailability of 5-Demethyltangeretin. Int. J. Mol. Sci. 2022, 23, 13284. [Google Scholar] [CrossRef]
- Whaley, W.L.; Rummel, J.D.; Kastrapeli, N. Interactions of Genistein and Related Isoflavones with Lipid Micelles. Langmuir 2006, 22, 7175–7184. [Google Scholar] [CrossRef]
- Ma, Y.; Ren, X.; Patel, N.; Xu, X.; Wu, P.; Liu, W.; Zhang, K.; Goodin, S.; Li, D.; Zheng, X. Nobiletin, a citrus polymethoxyflavone, enhances the effects of bicalutamide on prostate cancer cellsviadown regulation of NF-κB, STAT3, and ERK activation. RSC Adv. 2020, 10, 10254–10262. [Google Scholar] [CrossRef]
- Echeverría, J.; Opazo, J.; Mendoza, L.; Urzúa, A.; Wilkens, M. Structure-Activity and Lipophilicity Relationships of Selected Antibacterial Natural Flavones and Flavanones of Chilean Flora. Molecules 2017, 22, 608. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, S.; Jia, Y.; Yu, X.; Mou, R.; Li, X. Hispidulin inhibits proliferation, migration, and invasion by promoting autophagy via regulation of PPARγ activation in prostate cancer cells and xenograft models. Biosci. Biotechnol. Biochem. 2020, 85, 786–797. [Google Scholar] [CrossRef]
- Lin, C.; Chen, K.; Tsai, C.; Tsai, F.; Huang, C.; Tang, C.; Yang, J.; Hsu, Y.; Peng, S.; Chung, J. Casticin inhibits human prostate cancer DU 145 cell migration and invasion via Ras/Akt/NF-κB signaling pathways. J. Food Biochem. 2019, 43, e12902. [Google Scholar] [CrossRef] [PubMed]
- Lotfizadeh, R.; Sepehri, H.; Attari, F.; Delphi, L. Flavonoid Calycopterin Induces Apoptosis in Human Prostate Cancer Cells In-vitro. IJPR 2020, 19, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Kalra, R.; Bhagyaraj, E.; Tiwari, D.; Nanduri, R.; Chacko, A.P.; Jain, M.; Mahajan, S.; Khatri, N.; Gupta, P. AIRE promotes androgen-independent prostate cancer by directly regulating IL-6 and modulating tumor microenvironment. Oncogenesis 2018, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Feldman, B.J.; Feldman, D. The development of androgen-independent prostate cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Shin, E.A.; Kim, B.; Kim, B.-I.; Chitsazian-Yazdi, M.; Iranshahi, M.; Kim, S.-H. Auraptene Induces Apoptosis via Myeloid Cell Leukemia 1-Mediated Activation of Caspases in PC3 and DU145 Prostate Cancer Cells. Phytotherapy Res. 2017, 31, 891–898. [Google Scholar] [CrossRef]
- Yang, C.; Ma, X.; Wang, Z.; Zeng, X.; Hu, Z.; Ye, Z.; Shen, G. Curcumin induces apoptosis and protective autophagy in castration-resistant prostate cancer cells through iron chelation. Drug Des. Dev. Ther. 2017, ume11, 431–439. [Google Scholar] [CrossRef]
- Cevik, O.; Turut, F.A.; Acidereli, H.; Yildirim, S. Cyclosporine-A induces apoptosis in human prostate cancer cells PC3 and DU145 via downregulation of COX-2 and upregulation of TGFβ. Turk. J. Biochem. 2018, 44, 47–54. [Google Scholar] [CrossRef]
- Yang, D.-L.; Zhang, Y.-J.; He, L.-J.; Hu, C.-S.; Gao, L.-X.; Huang, J.-H.; Tang, Y.; Luo, J.; Tang, D.-Y.; Chen, Z.-Z. Demethylzeylasteral (T-96) initiates extrinsic apoptosis against prostate cancer cells by inducing ROS-mediated ER stress and suppressing autophagic flux. Biol. Res. 2021, 54, 1–14. [Google Scholar] [CrossRef]
- D’arrigo, G.; Gianquinto, E.; Rossetti, G.; Cruciani, G.; Lorenzetti, S.; Spyrakis, F. Binding of Androgen- and Estrogen-Like Flavonoids to Their Cognate (Non)Nuclear Receptors: A Comparison by Computational Prediction. Molecules 2021, 26, 1613. [Google Scholar] [CrossRef]
- Tousi, M.S.; Sepehri, H.; Khoee, S.; Farimani, M.M.; Delphi, L.; Mansourizadeh, F. Evaluation of apoptotic effects of mPEG-b-PLGA coated iron oxide nanoparticles as a eupatorin carrier on DU-145 and LNCaP human prostate cancer cell lines. J. Pharm. Anal. 2020, 11, 108–121. [Google Scholar] [CrossRef]
- Smeriglio, A.; Trombetta, D.; Marcoccia, D.; Narciso, L.; Mantovani, A.; Lorenzetti, S. Intracellular distribution and biological effects of phytochemicals in a sex steroid- sensitive model of human prostate adenocarcinoma. Anti-Cancer Agents Med. Chem. 2014, 14, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.B.; Mir, H.; Kapur, N.; Gales, D.N.; Carriere, P.P.; Singh, S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J. Surg. Oncol. 2018, 16, 1–12. [Google Scholar] [CrossRef]
- Hino, S.-I.; Inenaga, K.; Miyazaki, T.; Tanaka-Mizota, C. Suppression of HCT116 Human Colon Cancer Cell Motility by Polymethoxyflavones is Associated with Inhibition of Wnt/β-Catenin Signaling. Nutr. Cancer 2022, 74, 1–8. [Google Scholar] [CrossRef]
- Wu, X.; Li, Z.; Sun, Y.; Li, F.; Gao, Z.; Zheng, J.; Xiao, H. Identification of Xanthomicrol as a Major Metabolite of 5-Demethyltangeretin in Mouse Gastrointestinal Tract and Its Inhibitory Effects on Colon Cancer Cells. Front. Nutr. 2020, 7, 103. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Wang, C.-Y.; Tsao, E.-C.; Chen, Y.-T.; Wu, M.-J.; Ho, C.-T.; Yen, J.-H. 5-Demethylnobiletin Inhibits Cell Proliferation, Downregulates ID1 Expression, Modulates the NF-κB/TNF-α Pathway and Exerts Antileukemic Effects in AML Cells. Int. J. Mol. Sci. 2022, 23, 7392. [Google Scholar] [CrossRef]
- Nieddu, M.; Pollastro, F.; Caria, P.; Salamone, S.; Rosa, A. Xanthomicrol Activity in Cancer HeLa Cells: Comparison with Other Natural Methoxylated Flavones. Molecules 2023, 28, 558. [Google Scholar] [CrossRef]
- Xin, Y.; Zheng, T.; Zhang, M.; Zhang, R.; Zhu, S.; Li, D.; Zhao, D.; Ma, Y.; Ho, C.-T.; Huang, Q. Demethylnobiletin and its major metabolites: Efficient preparation and mechanism of their anti-proliferation activity in HepG2 cells. Food Sci. Hum. Wellness 2022, 11, 1191–1200. [Google Scholar] [CrossRef]
- Naróg, D.; Sobkowiak, A. Electrochemistry of Flavonoids. Molecules 2023, 28, 7618. [Google Scholar] [CrossRef]
- Kim, S.M.; Ha, S.E.; Lee, H.J.; Rampogu, S.; Vetrivel, P.; Kim, H.H.; Venkatarame Gowda Saralamma, V.; Lee, K.W.; Kim, G.S. Sinensetin Induces Autophagic Cell Death through p53-Related AMPK/mTOR Signaling in Hepatocellular Carcinoma HepG2 Cells. Nutrients 2020, 12, 2462. [Google Scholar] [CrossRef]
- Chen, S.; Nishi, M.; Morine, Y.; Yoshikawa, K.; Tokunaga, T.; Kashihara, H.; Takasu, C.; Wada, Y.; Yoshimoto, T.; Nakamoto, A.; et al. Polymethoxylated flavone sudachitin is a safe anticancer adjuvant that targets glycolysis in cancer-associated fibroblasts. Oncol. Lett. 2022, 24, 1–7. [Google Scholar] [CrossRef]
- Yen, S.-C.; Chen, L.-C.; Huang, H.-L.; Ngo, S.-T.; Wu, Y.-W.; Lin, T.E.; Sung, T.-Y.; Lien, S.-T.; Tseng, H.-J.; Pan, S.-L.; et al. Investigation of Selected Flavonoid Derivatives as Potent FLT3 Inhibitors for the Potential Treatment of Acute Myeloid Leukemia. J. Nat. Prod. 2021, 84, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alfwuaires, M.; Elsawy, H.; Sedky, A. Acacetin Inhibits Cell Proliferation and Induces Apoptosis in Human Hepatocellular Carcinoma Cell Lines. Molecules 2022, 27, 5361. [Google Scholar] [CrossRef] [PubMed]
- Novatcheva, E.D.; Anouty, Y.; Saunders, I.; Mangan, J.K.; Goodman, A.M. FMS-Like Tyrosine Kinase 3 Inhibitors for the Treatment of Acute Myeloid Leukemia. Clin. Lymphoma Myeloma Leuk. 2021, 22, e161–e184. [Google Scholar] [CrossRef] [PubMed]
- Ruglioni, M.; Crucitta, S.; Luculli, G.I.; Tancredi, G.; Del Giudice, M.L.; Mechelli, S.; Galimberti, S.; Danesi, R.; Del Re, M. Understanding mechanisms of resistance to FLT3 inhibitors in adult FLT3-mutated acute myeloid leukemia to guide treatment strategy. Crit. Rev. Oncol. 2024, 201, 104424. [Google Scholar] [CrossRef]
- Yan, B.; Chen, Q.; Xu, J.; Li, W.; Xu, B.; Qiu, Y. Low-frequency TP53 hotspot mutation contributes to chemoresistance through clonal expansion in acute myeloid leukemia. Leukemia 2020, 34, 1816–1827. [Google Scholar] [CrossRef]
- Skopek, R.; Palusińska, M.; Kaczor-Keller, K.; Pingwara, R.; Papierniak-Wyglądała, A.; Schenk, T.; Lewicki, S.; Zelent, A.; Szymański, Ł. Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research. Int. J. Mol. Sci. 2023, 24, 5377. [Google Scholar] [CrossRef]
- Sak, K.; Everaus, H. Established Human Cell Lines as Models to Study Anti-leukemic Effects of Flavonoids. Curr. Genom. 2016, 18, 3–26. [Google Scholar] [CrossRef]
- Pandur, E.; Micalizzi, G.; Mondello, L.; Horváth, A.; Sipos, K.; Horváth, G. Antioxidant and Anti-Inflammatory Effects of Thyme (Thymus vulgaris L.) Essential Oils Prepared at Different Plant Phenophases on Pseudomonas aeruginosa LPS-Activated THP-1 Macrophages. Antioxidants 2022, 11, 1330. [Google Scholar] [CrossRef]
- Venugopal, S.; Bar-Natan, M.; Mascarenhas, J.O. JAKs to STATs: A tantalizing therapeutic target in acute myeloid leukemia. Blood Rev. 2019, 40, 100634. [Google Scholar] [CrossRef]
- del Rosario, H.; Saavedra, E.; Brouard, I.; González-Santana, D.; García, C.; Spínola-Lasso, E.; Tabraue, C.; Quintana, J.; Estévez, F. Structure-activity relationships reveal a 2-furoyloxychalcone as a potent cytotoxic and apoptosis inducer for human U-937 and HL-60 leukaemia cells. Bioorganic Chem. 2022, 127, 105926. [Google Scholar] [CrossRef]
- Yen, J.-H.; Lin, C.-Y.; Chuang, C.-H.; Chin, H.-K.; Wu, M.-J.; Chen, P.-Y. Nobiletin Promotes Megakaryocytic Differentiation through the MAPK/ERK-Dependent EGR1 Expression and Exerts Anti-Leukemic Effects in Human Chronic Myeloid Leukemia (CML) K562 Cells. Cells 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Al-Nuaimi, A.; Al-Hiari, Y.; Kasabri, V.; Haddadin, R.; Mamdooh, N.; Alalawi, S.; Khaleel, S. A Novel Class of Functionalized Synthetic Fluoroquinolones with Dual Antiproliferative - Antimicrobial Capacities. Asian Pac. J. Cancer Prev. 2021, 22, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Cytotoxicity evaluation and the mode of cell death of K562 cells induced by organotin (IV) (2-methoxyethyl) methyldithiocarbamate compounds. J. Appl. Pharm. Sci. 2019, 9, 10–15. [CrossRef]
- Cheng, Z.; Chueh, F.; Peng, S.; Lin, C.; Kuo, C.; Huang, W.; Chen, P.; Way, T.; Chung, J. Combinational treatment of 5-fluorouracil and casticin induces apoptosis in mouse leukemia WEHI-3 cells in vitro. Environ. Toxicol. 2020, 35, 911–921. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Zhang, H.; Chen, J.; Cao, J.; Chen, Q.; Li, X.; Sun, C. Polymethoxyflavones from citrus inhibited gastric cancer cell proliferation through inducing apoptosis by upregulating RARβ, both in vitro and in vivo. Food Chem. Toxicol. 2020, 146, 111811. [Google Scholar] [CrossRef]
- Ragi, C.; Muraleedharan, K. Hydrogen bonding interactions between Hibiscetin and ethanol/water: DFT studies on structure and topologies. Chem. Phys. Impact 2023, 8. [Google Scholar] [CrossRef]
- de Oliveira-Júnior, R.G.; Marcoult-Fréville, N.; Prunier, G.; Beaugeard, L.; Filho, E.B.d.A.; Mourão, E.D.S.; Michel, S.; Quintans-Júnior, L.J.; Almeida, J.R.G.d.S.; Grougnet, R.; et al. Polymethoxyflavones from Gardenia oudiepe (Rubiaceae) induce cytoskeleton disruption-mediated apoptosis and sensitize BRAF-mutated melanoma cells to chemotherapy. Chem. Interactions 2020, 325, 109109. [Google Scholar] [CrossRef]
- Strawa, J.W.; Jakimiuk, K.; Szoka, Ł.; Brzezinski, K.; Drozdzal, P.; Pałka, J.A.; Tomczyk, M. New Polymethoxyflavones from Hottonia palustris Evoke DNA Biosynthesis-Inhibitory Activity in An Oral Squamous Carcinoma (SCC-25) Cell Line. Molecules 2022, 27, 4415. [Google Scholar] [CrossRef]
- You, L.; Lin, J.; Yu, Z.; Qian, Y.; Bi, Y.; Wang, F.; Zhang, L.; Zheng, C.; Zhang, J.; Li, W.; et al. Nobiletin suppresses cholangiocarcinoma proliferation via inhibiting GSK3β. Int. J. Biol. Sci. 2022, 18, 5698–5712. [Google Scholar] [CrossRef]
- Ren, X.; Ma, Y.; Wang, X.; Xu, X.; Wu, P.; Liu, W.; Zhang, K.; Goodin, S.; Li, D.; Zheng, X. Nobiletin Inhibits Cell Growth, Migration and Invasion, and Enhances the Anti-Cancer Effect of Gemcitabine on Pancreatic Cancer Cells. Nat. Prod. Commun. 2021, 16. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Y.; Jiang, W.; Ji, H.; Wang, Z.-W.; Zhu, X. Discovery of key genes as novel biomarkers specifically associated with HPV-negative cervical cancer. Mol. Ther. - Methods Clin. Dev. 2021, 21, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2020, 10, 3116. [Google Scholar] [CrossRef] [PubMed]
- Narisawa-Saito, M.; Kiyono, T. Basic mechanisms of high-risk human papillomavirus-induced carcinogenesis: Roles of E6 and E7 proteins. Cancer Sci. 2007, 98, 1505–1511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Quan, Y.; Bai, Y.; Yang, L.; Yang, Y. The effect and apoptosis mechanism of 6-methoxyflavone in HeLa cells. Biomarkers 2022, 27, 470–482. [Google Scholar] [CrossRef]
- Zhang, C.; Quan, Y.; Yang, L.; Bai, Y.; Yang, Y. 6-Methoxyflavone induces S-phase arrest through the CCNA2/CDK2/p21CIP1 signaling pathway in HeLa cells. Bioengineered 2022, 13, 7277–7292. [Google Scholar] [CrossRef]
- Xing, F.; Sun, C.; Luo, N.; He, Y.; Chen, M.; Ding, S.; Liu, C.; Feng, L.; Cheng, Z. Wogonin Increases Cisplatin Sensitivity in Ovarian Cancer Cells Through Inhibition of the Phosphatidylinositol 3-Kinase (PI3K)/Akt Pathway. Med Sci. Monit. 2019, 25, 6007–6014. [Google Scholar] [CrossRef]
- Kim, E., Kim, Y., Ji, Z., Kang, J., Wirianto, M., Paudel, K. R., Smith, J. A., Ono, K., Kim, J., Eckel-Mahan, K., Zhou, X., Lee, H. K., Yoo, J. Y., Yoo, S., & Chen, Z. (2022). ROR activation by Nobiletin enhances antitumor efficacy via suppression of IκB/NF-κB signaling in triple-negative breast cancer. Cell Death and Disease, 13(4). [CrossRef]
- Zehra, B.; Ahmed, A.; Sarwar, R.; Khan, A.; Farooq, U.; Ali, S.A.; Al-Harrasi, A. Apoptotic and antimetastatic activities of betulin isolated from Quercus incana against non-small cell lung cancer cells. Cancer Manag. Res. 2019, ume 11, 1667–1683. [Google Scholar] [CrossRef]
- Cao, Y.; Cao, W.; Qiu, Y.; Zhou, Y.; Guo, Q.; Gao, Y.; Lu, N. Oroxylin A suppresses ACTN1 expression to inactivate cancer-associated fibroblasts and restrain breast cancer metastasis. Pharmacol. Res. 2020, 159, 104981. [Google Scholar] [CrossRef]
- Feng, S.; Zhou, Y.; Huang, H.; Lin, Y.; Zeng, Y.; Han, S.; Huang, K.; Liu, Q.; Zhu, W.; Yuan, Z.; et al. Nobiletin Induces Ferroptosis in Human Skin Melanoma Cells Through the GSK3β-Mediated Keap1/Nrf2/HO-1 Signalling Pathway. Front. Genet. 2022, 13, 865073. [Google Scholar] [CrossRef]
- Goldar, S.; Khaniani, M.S.; Derakhshan, S.M.; Baradaran, B. Molecular Mechanisms of Apoptosis and Roles in Cancer Development and Treatment. Asian Pac. J. Cancer Prev. 2015, 16, 2129–2144. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Wei, Z.; Yang, W.; Huang, J.; Yang, Y.; Wang, J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front. Oncol. 2022, 12, 985363. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The Roles of Cyclin-Dependent Kinases in Cell-Cycle Progression and Therapeutic Strategies in Human Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef] [PubMed]
- Duo, L.; Chen, Y.; Liu, Q.; Ma, Z.; Farjudian, A.; Ho, W.Y.; Low, S.S.; Ren, J.; Hirst, J.D.; Xie, H.; et al. Discovery of novel SOS1 inhibitors using machine learning. RSC Med. Chem. 2024, 15, 1392–1403. [Google Scholar] [CrossRef]
- Rapp, U.R.; Todaro, G.J. Generation of New Mouse Sarcoma Viruses in Cell Culture. Science 1978, 201, 821–824. [Google Scholar] [CrossRef]
- Dillon, M.; Lopez, A.; Lin, E.; Sales, D.; Perets, R.; Jain, P. Progress on Ras/MAPK Signaling Research and Targeting in Blood and Solid Cancers. Cancers 2021, 13, 5059. [Google Scholar] [CrossRef]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Chang, F.; Steelman, L.S.; Lee, J.T.; Shelton, J.G.; Navolanic, P.M.; Blalock, W.L.; A Franklin, R.; A McCubrey, J. Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: potential targeting for therapeutic intervention. Leukemia 2003, 17, 1263–1293. [Google Scholar] [CrossRef]
- Ballou, L.M.; Chattopadhyay, M.; Li, Y.; Scarlata, S.; Lin, R.Z. Gαq binds to p110α/p85α phosphoinositide 3-kinase and displaces Ras. Biochem. J. 2006, 394, 557–562. [Google Scholar] [CrossRef]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y.; Yap, K.C.H.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B.; et al. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, L.; Li, W.; Zou, D.; Yu, J.; Wang, L.; Wong, C.C. Transcription factors in colorectal cancer: molecular mechanism and therapeutic implications. Oncogene 2020, 40, 1555–1569. [Google Scholar] [CrossRef] [PubMed]
- Micheau, O.; Tschopp, J. Induction of TNF Receptor I-Mediated Apoptosis via Two Sequential Signaling Complexes. Cell 2003, 114, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sun, L.; Xu, F. NF-κB in Cell Deaths, Therapeutic Resistance and Nanotherapy of Tumors: Recent Advances. Pharmaceuticals 2023, 16, 783. [Google Scholar] [CrossRef] [PubMed]
- Habib, E.S.; El-Bsoumy, E.; Ibrahim, A.K.; Helal, M.A.; El-Magd, M.A.; Ahmed, S.A. Anti-inflammatory effect of methoxyflavonoids fromChiliadenus montanus(Jasonia Montana)growing in Egypt. Nat. Prod. Res. 2021, 35, 5909–5913. [Google Scholar] [CrossRef]
- Chagas, M.D.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-De-Albuquerque, C.F. Flavonols and Flavones as Potential anti-Inflammatory, Antioxidant, and Antibacterial Compounds. Oxidative Med. Cell. Longev. 2022, 2022, 1–21. [Google Scholar] [CrossRef]
- Gobeil, S.; Boucher, C.C.; Nadeau, D.; Poirier, G.G. Characterization of the necrotic cleavage of poly(ADP-ribose) polymerase (PARP-1): implication of lysosomal proteases. Cell Death Differ. 2001, 8, 588–594. [Google Scholar] [CrossRef]
- Simbulan-Rosenthal, C.M.; Rosenthal, D.S.; Iyer, S.; Boulares, A.H.; Smulson, M.E. Transient Poly(ADP-ribosyl)ation of Nuclear Proteins and Role of Poly(ADP-ribose) Polymerase in the Early Stages of Apoptosis. J. Biol. Chem. 1998, 273, 13703–13712. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Paskeh, M.D.A.; Mirzaei, S.; Gholami, M.H.; Zarrabi, A.; Hashemi, F.; Hushmandi, K.; Hashemi, M.; Nabavi, N.; Crea, F.; et al. Targeting autophagy in prostate cancer: preclinical and clinical evidence for therapeutic response. J. Exp. Clin. Cancer Res. 2022, 41, 1–37. [Google Scholar] [CrossRef]
- Lim, S.M.; Hanif, E.A.M.; Chin, S.-F. Is targeting autophagy mechanism in cancer a good approach? The possible double-edge sword effect. Cell Biosci. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- El-Aziz, Y.S.A.; Leck, L.Y.W.; Jansson, P.J.; Sahni, S. Emerging Role of Autophagy in the Development and Progression of Oral Squamous Cell Carcinoma. Cancers 2021, 13, 6152. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Chen, Y.; Gibson, S.B. Regulation of Autophagy by Reactive Oxygen Species (ROS): Implications for Cancer Progression and Treatment. Antioxidants Redox Signal. 2009, 11, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-Z.; Jiang, S.; Zhang, L.; Yu, Z.-B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cubillos-Ruiz, J.R. Endoplasmic reticulum stress signals in the tumour and its microenvironment. Nat. Rev. Cancer 2020, 21, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Liu, Z.; Liang, M.-X.; Fei, Y.-J.; Zhang, W.; Wu, Y.; Tang, J.-H. Endoplasmic reticulum stress targeted therapy for breast cancer. Cell Commun. Signal. 2022, 20, 1–12. [Google Scholar] [CrossRef]
- Madden, E.; Logue, S.E.; Healy, S.J.; Manie, S.; Samali, A. The role of the unfolded protein response in cancer progression: From oncogenesis to chemoresistance. Biol. Cell 2019, 111, 1–17. [Google Scholar] [CrossRef]
- Liu, J.; Qu, L.; Meng, L.; Shou, C. Topoisomerase inhibitors promote cancer cell motility via ROS-mediated activation of JAK2-STAT1-CXCL1 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 1–12. [Google Scholar] [CrossRef]
- Yakkala, P.A.; Penumallu, N.R.; Shafi, S.; Kamal, A. Prospects of Topoisomerase Inhibitors as Promising Anti-Cancer Agents. Pharmaceuticals 2023, 16, 1456. [Google Scholar] [CrossRef]
- Ji, Y.; Lv, J.; Sun, D.; Huang, Y. Therapeutic strategies targeting Wnt/β-catenin signaling for colorectal cancer (Review). Int. J. Mol. Med. 2021, 49, 1–17. [Google Scholar] [CrossRef]
- Koury, J.; Zhong, L.; Hao, J. Targeting Signaling Pathways in Cancer Stem Cells for Cancer Treatment. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, W.; Guo, L.; Kril, L.M.; Begley, K.L.; Sviripa, V.M.; Chen, X.; Liu, X.; Lee, E.Y.; He, D.; et al. Potent Synergistic Effect on C-Myc-Driven Colorectal Cancers Using a Novel Indole-Substituted Quinoline with a Plk1 Inhibitor. Mol. Cancer Ther. 2021, 20, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram on anticancer activity of methoxyflavones scaffold.
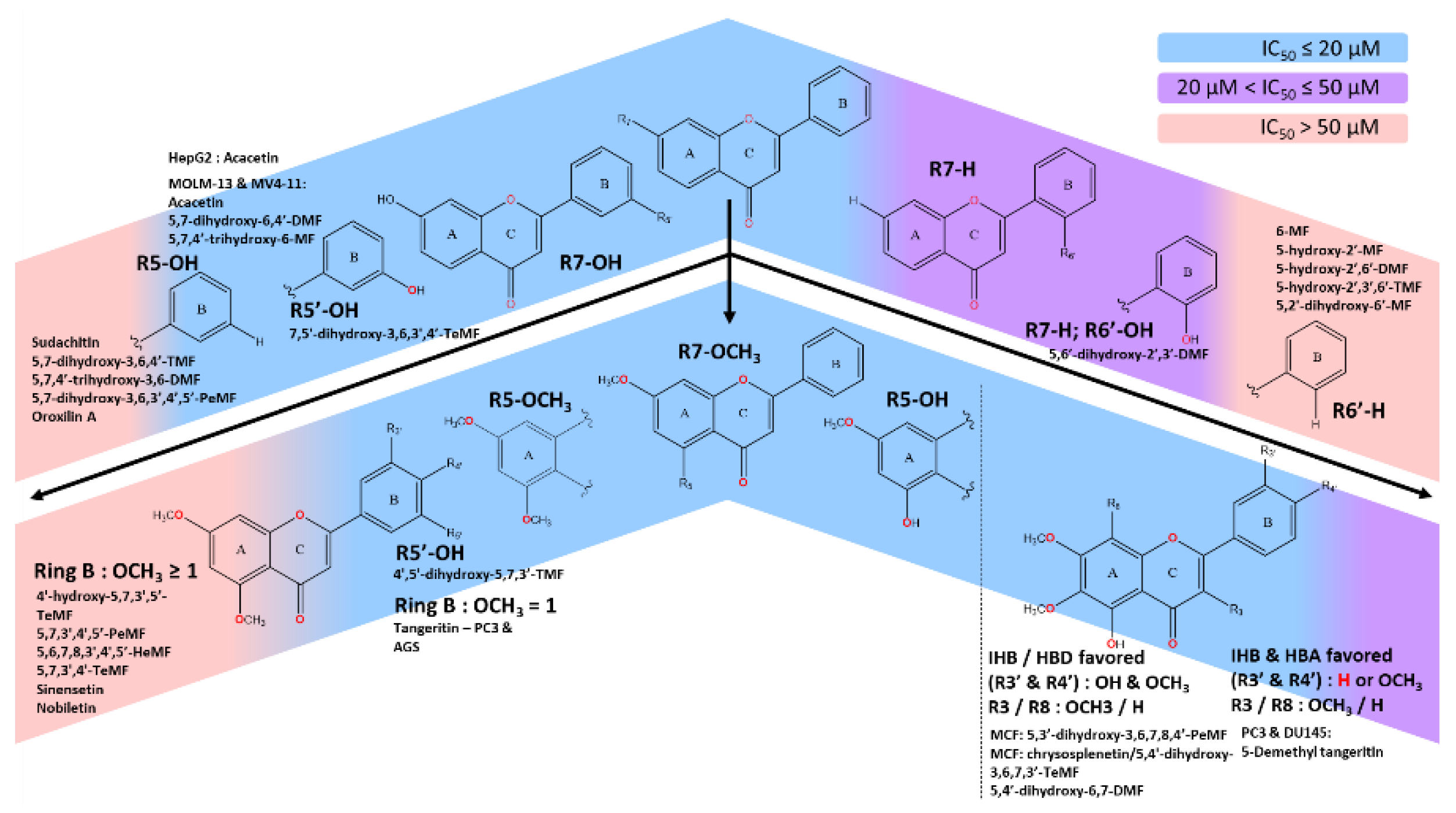
Figure 2.
The influence of the position of both functional groups in ring B on the IC50 value in MCF-7 in 72-hour treatment duration.
Figure 2.
The influence of the position of both functional groups in ring B on the IC50 value in MCF-7 in 72-hour treatment duration.
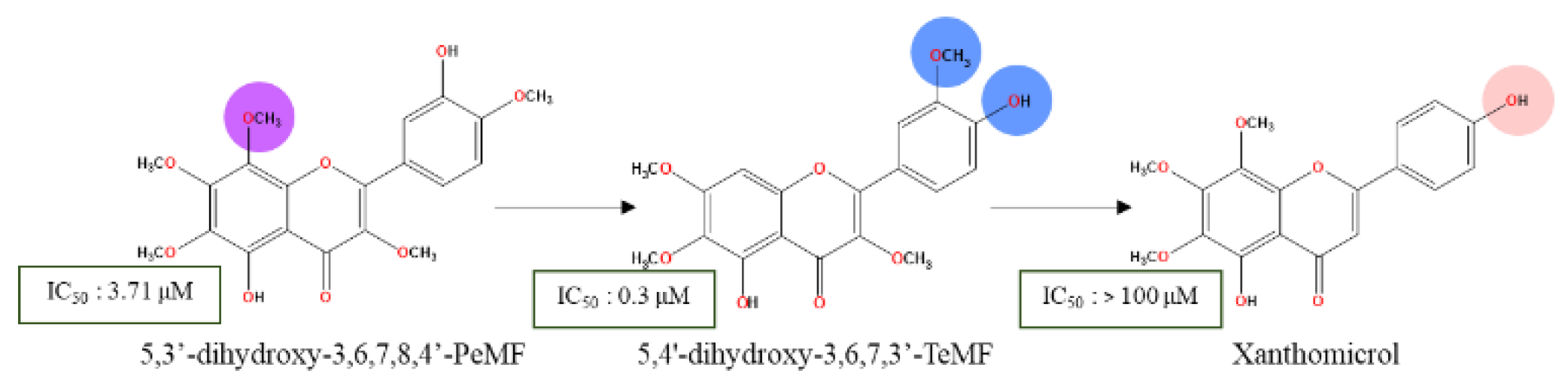
Figure 3.
Lewis’s resonance structures illustrating electron delocalization in (A) C3’, (B) C4’, (C) C5 free radicals. Ring AC and BC are not conjugated. According to a 2013 study, a strong IHB between C5-OH and the C4 oxygen atom of the carbonyl group facilitates the formation of IHB’s involving (D) C4’-OH and (E) C3’-OH with an adjacent methoxy groups.
Figure 3.
Lewis’s resonance structures illustrating electron delocalization in (A) C3’, (B) C4’, (C) C5 free radicals. Ring AC and BC are not conjugated. According to a 2013 study, a strong IHB between C5-OH and the C4 oxygen atom of the carbonyl group facilitates the formation of IHB’s involving (D) C4’-OH and (E) C3’-OH with an adjacent methoxy groups.
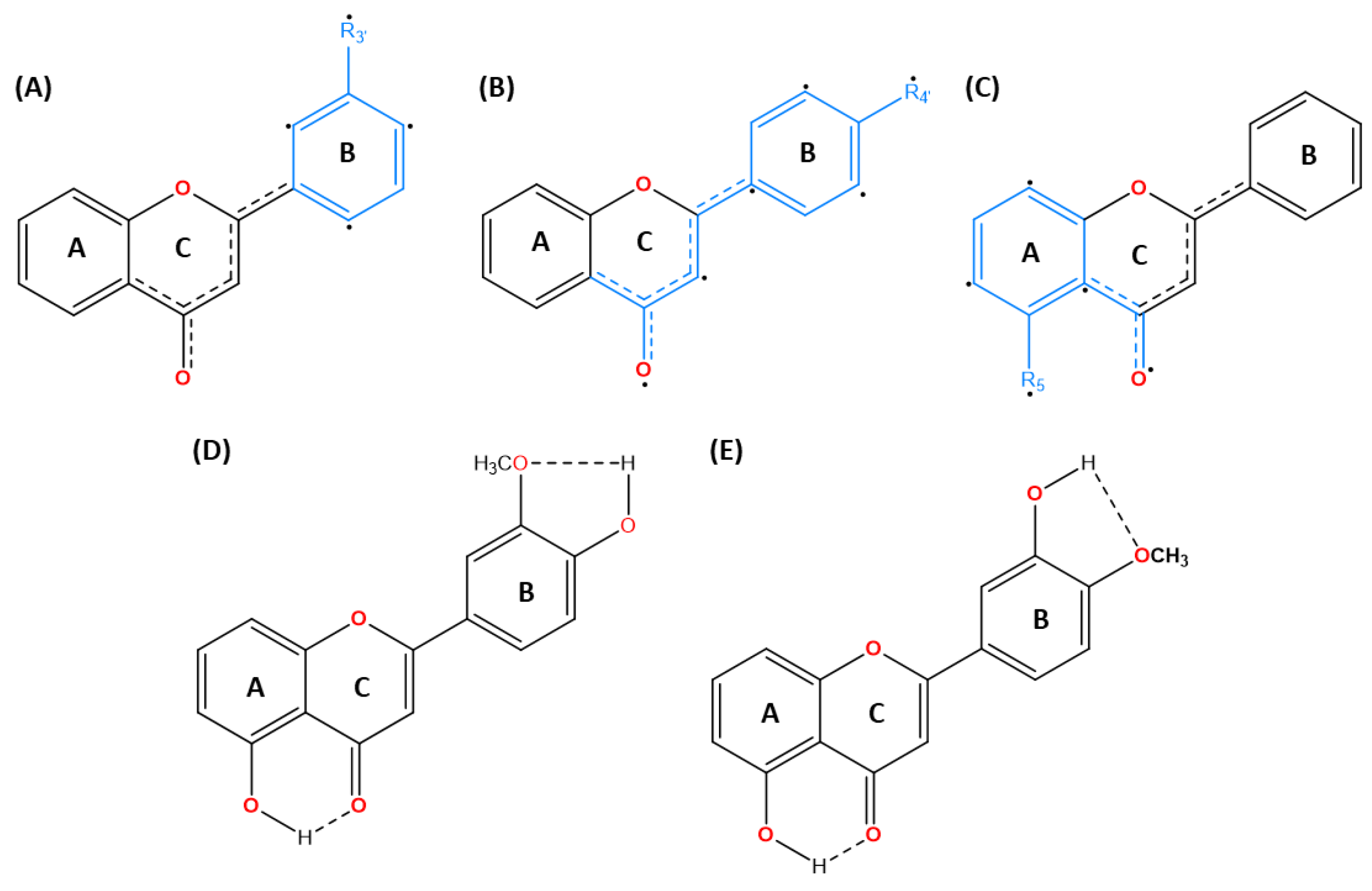
Figure 4.
Cytotoxic effects governed by dihydroxylated moieties on ring B highlighting the impact of C5’ scaffold composition. The presence of C5-OCH3 shows enhanced cytotoxicity similar to the C5-
Figure 4.
Cytotoxic effects governed by dihydroxylated moieties on ring B highlighting the impact of C5’ scaffold composition. The presence of C5-OCH3 shows enhanced cytotoxicity similar to the C5-
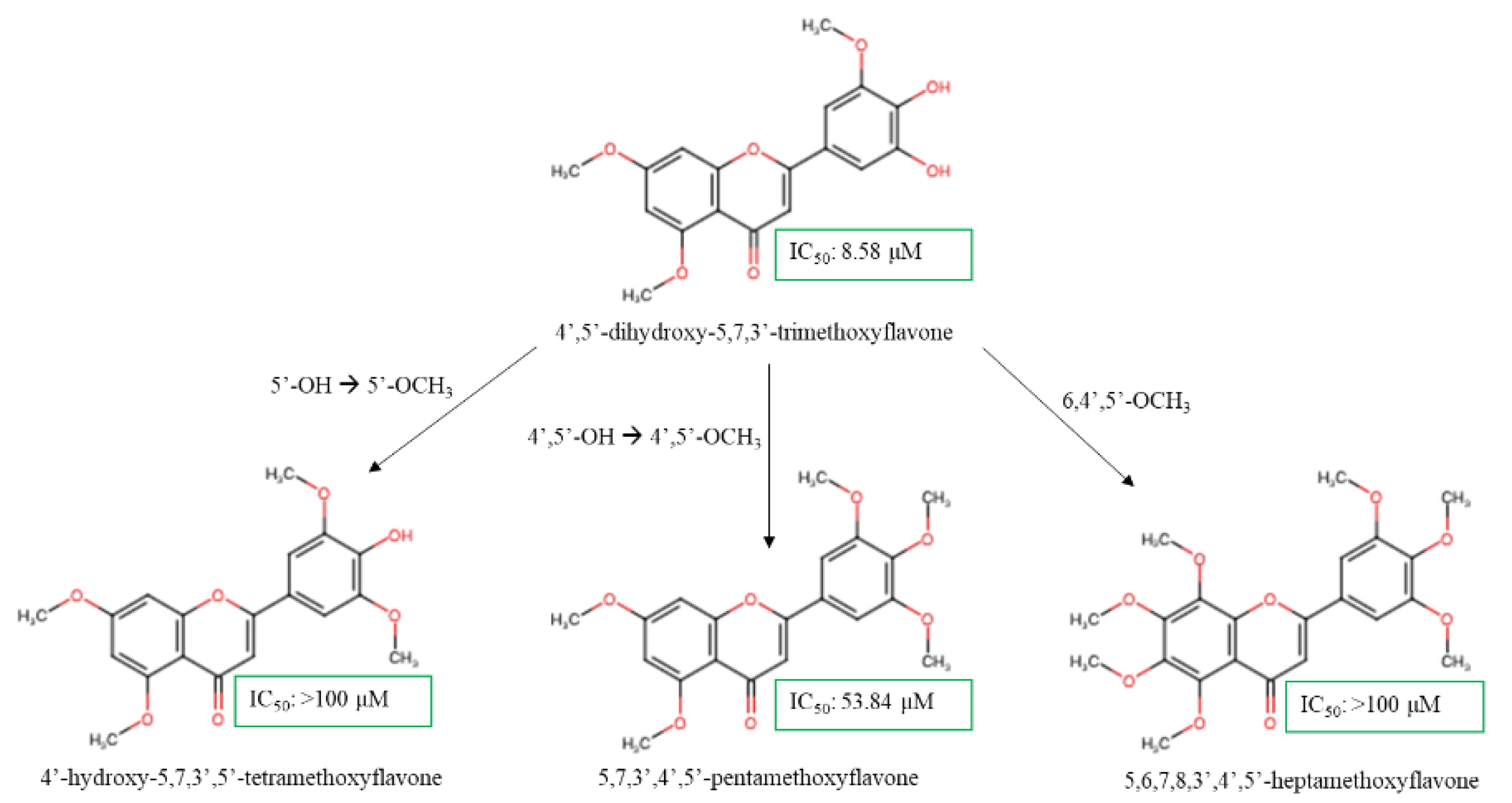
Figure 5.
SAR analysis of methoxyflavones analogs and their cytotoxic effects on prostate cancer cell lines.
Figure 5.
SAR analysis of methoxyflavones analogs and their cytotoxic effects on prostate cancer cell lines.
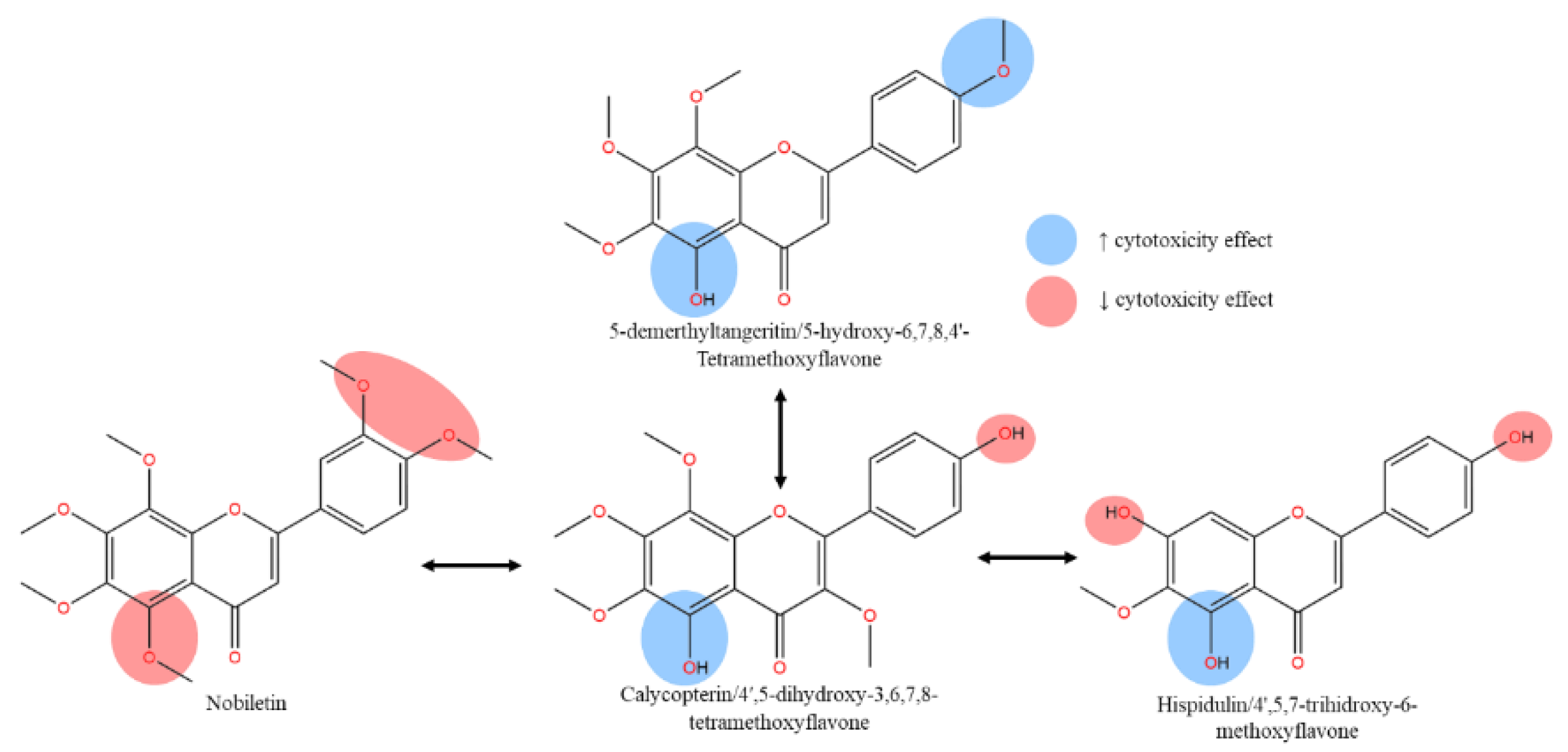
Figure 6.
SAR analysis on cytotoxicity activity of methoxyflavones analogs on HCT116 colon cancer cell lines.
Figure 6.
SAR analysis on cytotoxicity activity of methoxyflavones analogs on HCT116 colon cancer cell lines.
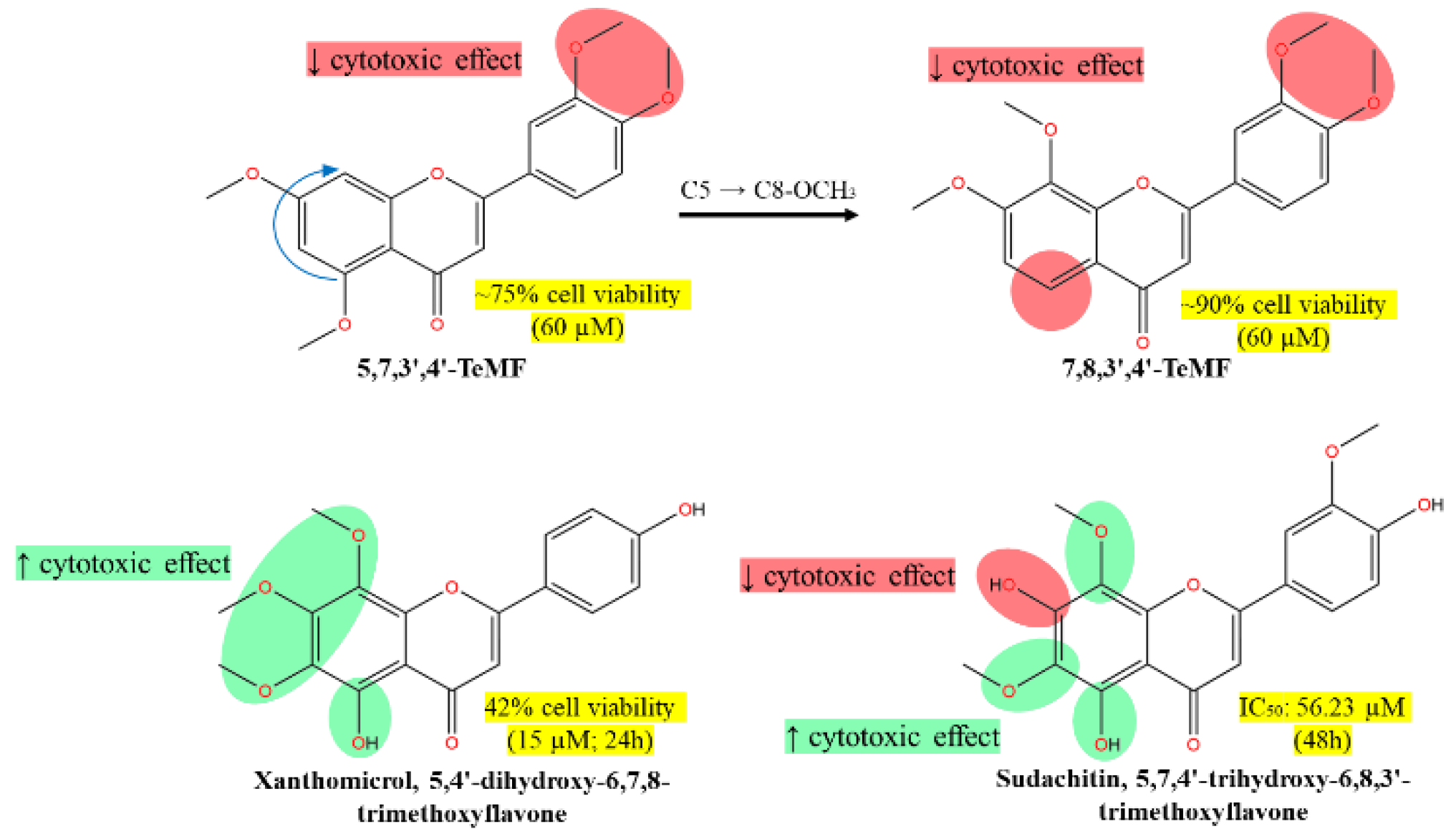
Figure 7.
SAR analysis on cytotoxicity activities of 5-demethylnobiletin derivatives on HepG2 liver cancer cell lines.
Figure 7.
SAR analysis on cytotoxicity activities of 5-demethylnobiletin derivatives on HepG2 liver cancer cell lines.
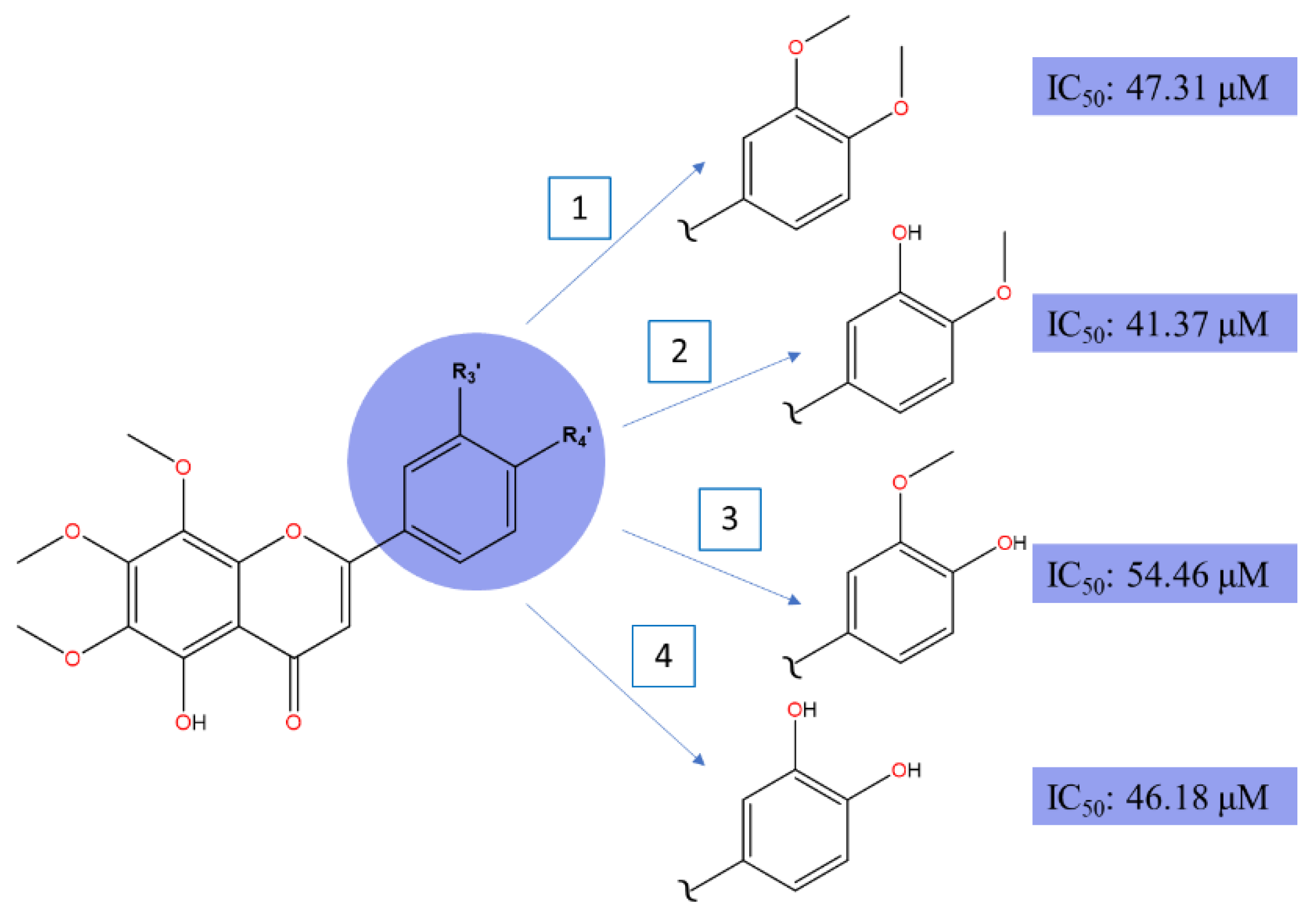
Figure 8.
SAR analysis on cytotoxicity activities of 5-demethylnobiletin, sinensetin, and sudachitin analogs on HepG2 colon cancer cell lines.
Figure 8.
SAR analysis on cytotoxicity activities of 5-demethylnobiletin, sinensetin, and sudachitin analogs on HepG2 colon cancer cell lines.
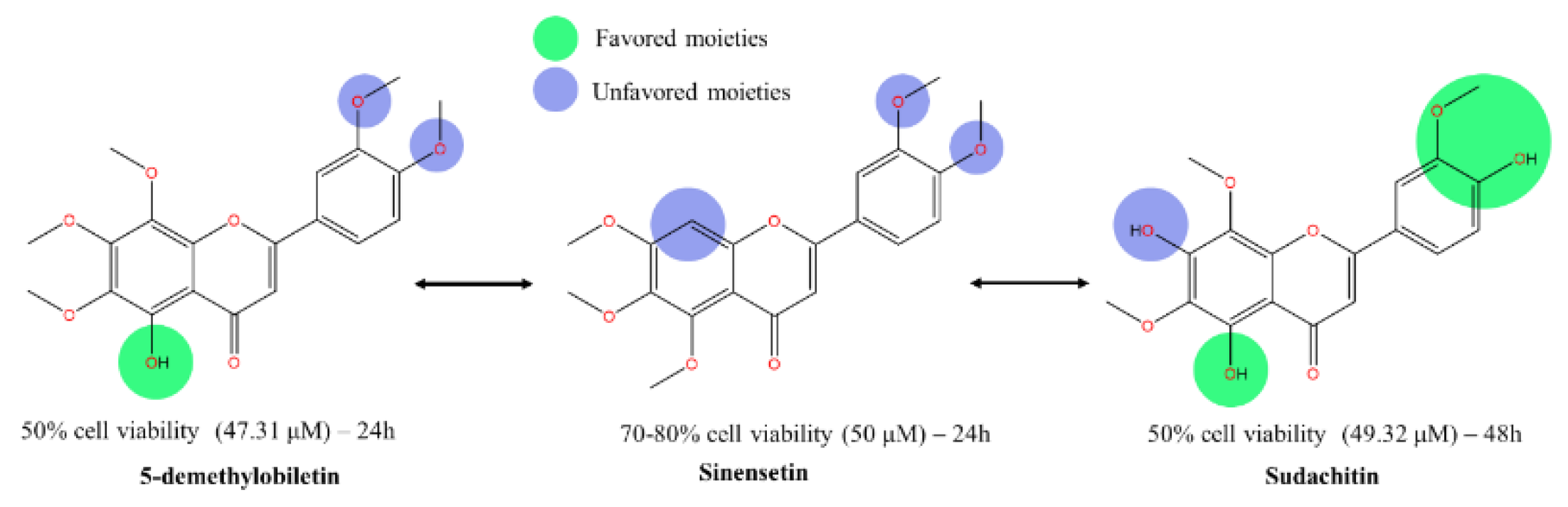

Table 1.
Comparative Antiproliferative Effects of Methoxyflavones against Prostate Cancer Cell Lines.
Table 1.
Comparative Antiproliferative Effects of Methoxyflavones against Prostate Cancer Cell Lines.
| Prostate cancer cell lines | C3 | C5 | C7 | C8 | C3’ | C4’ | IC50 (µM) / Cell viability (%) | Reference |
|---|---|---|---|---|---|---|---|---|
| PC3 | H | OMe | OMe | OMe | H | OMe | 17.2 | [63] |
| H | OH | OMe | OMe | H | OMe | 11.8 | ||
| H | OMe | OMe | OMe | OMe | OMe | 80.0 | [65] | |
| VCaP | H | OMe | OMe | OMe | OMe | OMe | > 80% (120 µM) | |
| H | OH | OH | H | H | OH | >50% (50 µM) | [67] | |
| DU145 | OMe | OH | OMe | H | OH | OMe | < 80% (50 µM) | [68] |
| OMe | OH | OMe | OMe | H | OH | 235.0 | [69] | |
| LNCaP | OMe | OH | OMe | OMe | H | OH | 116.5 |
Table 2.
Comparison of IC50 value of various compounds on hormone-independent prostate cancer cell lines (PC3 and DU145).
Table 2.
Comparison of IC50 value of various compounds on hormone-independent prostate cancer cell lines (PC3 and DU145).
| Sample | IC50 (µM) / cell viability (%) | Reference | |
| PC3 | DU145 | ||
| Auraptene | <50% (120 µM) | <50% (120 µM) | [72] |
| Curcumin | <60% (50 µM) | <60% (50 µM) | [73] |
| Cyclosporine-A | 8.80 μM | 12.14 μM | [74] |
| Demethylzeylasteral (T-96) | 13.10 µM | 11.47 µM | [75] |
Table 3.
Cytotoxicity assay of methoxyflavones analogs.
| Natural sources | Cancer | Cells | Treatment (hr) | IC50 / % cell viability (%, µM) | References | |
|---|---|---|---|---|---|---|
| 5,3’-dihydroxy-3,6,7,8,4’-PeMF | Glycosmis ovoidea Pierre | Breast | MCF-7 | 24 | 49.46 µM | [46] |
| 48 | 17.56 µM | |||||
| 72 | 3.71 µM | |||||
| MDA-MB-231 | 24 | 49.46 µM | ||||
| 48 | 49.46 µM | |||||
| 72 | 21.27 µM | |||||
| Nobiletin | Citrus sinensis / Citrus reticulata | Gastric | AGS | 48 | >20% (62.13 µM) | [102] |
| BGC-823 | >90% (100 µM) | |||||
| SGC-7901 | >90% (100 µM) | |||||
| CML | K562 | 48 | 82.49 µM | [98] | ||
| Prostate | VCaP | 48 | > 80% (100 µM) | [65] | ||
| PC3 | 48 | 80 - 100 µM | [63] | |||
| Pancreas | MIA PaCa-2 | 72 | 80% (120 µM) | [107] | ||
| PANC-1 | 72 | 20% (80 µM) | ||||
| Breast | MDA-MB-231 | 72 | 70% (10.04 µM) | [114] | ||
| Bile duct | TFK1 | 48 | Inactive | [82] | ||
| RBE | 48 | Inactive | ||||
| Compound | ||||||
| 5-demethylnobiletin | Citrus reticulata | Liver | HepG2 | 24 | 47.31 µM | [84] |
| AML | HL-60 | 48 | 85.7 µM | [98] | ||
| THP-1 | 32.3 µM | |||||
| U-937 | 30.4 µM | |||||
| HEL | 65.3 µM | |||||
| CML | K562 | 91.5 µM | ||||
| Gastric | AGS | >90% (100 µM) | [102] | |||
| BGC-823 | >90% (100 µM) | |||||
| SGC-7901 | >90% (100 µM) | |||||
| 5,7,4'-trihydroxy-6,8,3'-TMF / sudachitin | Citrus sudachi | Colon | HCT116 | 48 | 56.23 µM | [87] |
| HT-29 | 48 | 37.07 µM | ||||
| Liver | HepG2 | 48 | 49.32 µM | |||
| Bile duct | HuCCT1 | 48 | 53.21 µM | |||
| RBE | 48 | 24.1 µM | ||||
| Pancreas | MIA PaCa-2 | 48 | 43.35 µM | |||
| PANC-1 | 48 | 32.73 µM | ||||
| 5,6′-dihydroxy-2′,3′-DMF | Hottonia palustris | Oral | SCC-25 | 24 | 78.2 µM | [105] |
| 5,6′-dihydroxy-2′,3′-DMF | 48 | 40.6 µM | ||||
| 5-hydroxy-2′-MF | Inactive | |||||
| 5-hydroxy-2′,6′-DMF | Inactive | |||||
| 5-hydroxy-2′,3′,6′-TMF | Inactive | |||||
| 5,2'-dihydroxy-6'-MF | Inactive | |||||
| 5,7-dihydroxy-4'-MF / acacetin | Robinia pseudoacacia | MOLM-13 | 72 | 9.1 µM | [88] | |
| 5,4’-dihydroxy-6,7-DMF | Quercus incana | 2.6 µM | ||||
| 5,7-dihydroxy-6,4'-DMF / Pectolinarigenin | Artemisia capillaris | 5.9 µM | ||||
| 5,7,4’-trihydroxy-6-MF/ hispidulin | Artemisia capillaris | 7.0 µM | ||||
| 5,7,4’-trihydroxy-6-MF/hispidulin | Artemisia capillaris | MV4-11 | 6.8 µM | |||
| 5,4’-dihydroxy-6,7-DMF | Quercus incana | 2.6 µM | ||||
| 5,7-dihydroxy-6,4'-DMF / Pectolinarigenin | Artemisia capillaris | 7.9 µM | ||||
| 5,7-dihydroxy-4'-MF | Robinia pseudoacacia | 6.8 µM | ||||
| 5,7-dihydroxy-3,6,4'-TMF | Gardenia oudiepe (Rubiaceae) | Skin | A2058 | 72 | 66.52% (10 µM) | [104] |
| 7,5'-dihydroxy-3,6,3',4'-TeMF | 72 | 42.86% (10 µM) | ||||
| 5,7,4’-trihydroxy-3,6-DMF | 72 | Inactive | ||||
| 5,7-dihydroxy-3,6,3’,4’,5’-PeMF | 72 | Inactive | ||||
| 5,7-dihydroxy-8-MF / wogonin | Scutellaria baicalensis | Ovarian | SKOV3 | 72 | 80% (20 µM) | [113] |
| OV2008 | ||||||
| SKOV3/DDP | ||||||
| C13* | ||||||
| 4',5'-dihydroxy-5,7,3'-TMF | Synthetic compound | Breast | HCC1954 | NA | 8.58 µM | [58] |
| 4'-hydroxy-5,7,3',5'-TeMF | >100 µM | |||||
| 5,7,3',4',5'-PeMF | 53.84 µM | |||||
| 5,6,7,8,3',4',5'-heptamethoxyflavone | >100 µM | |||||
| 5,6,7,8,4'-PeMF / tangeritin | Citrus reticulata | Prostate | PC3 | 48 | 17.2 µM | [63] |
| Gastric | AGS | 48 | <20% (33.57 µM) | [102] | ||
| BGC-823 | 48 | >90% (100 µM) | ||||
| SGC-7901 | ||||||
| 5,4'-dihydroxy-6,7,8-TMF/ xanthomicrol | Achillea erba-rotta subsp. moschata | Colon | HCT116 | 24 | 42% (15 µM) | [81] |
| Breast | MCF7 | 72 | > 100 µM | [49] | ||
| Cervical | Hela | 72 | > 100 µM | [83] | ||
| 6-MF | Pimelea decora | Cervical | Hela | 72 | 55.31 µM | [111,112] |
| C33A | 72 | 109.57 µM | ||||
| SiHa | 72 | 208.53 µM | ||||
| 5,3'-didemethylnobiletin | Citrus reticulata | Liver | HepG2 | 24 | 41.37 µM | [84] |
| 5,4'-didemethylnobiletin | 24 | 54.46 µM | ||||
| 5,3',4'-tridemethylnobiletin | 24 | 46.18 µM | ||||
| 5,7,3',4'-TeMF | Kaempferia parviflora | Colon | HCT116 | 72 | 75% (60 µM) | [80] |
| 7,8,3',4'-TeMF | Citrus reticulata | 72 | 90% (60 µM) | |||
| 5,4’-dihydroxy-3,6,7,8-TeMF/ calycopterin | D. kotschyi Boiss | Prostate | DU145 | 48 | 235 µM | [69] |
| LNCaP | 48 | 116.5 µM | ||||
| 5,4’-dihydroxy-6,7-DMF / cirsimaritin | Quercus incana | Lung | NCI-H460 | 24 | 26.23 µM | [115] |
| 5,3′-dihydroxy-6,7,4′-TMF / eupatorin | 24 | 37.50 µM | ||||
| 5,7-dihydroxy-6-MF / acacetin | Robinia pseudoacacia | Liver | HepG2 | 24 | 25 µM | [89] |
| 5,6,7,3',4'-PeMF / Sinensetin | O. aristatus leaves | HepG2 | 24 | 80% (50 µM) | [86] | |
| 5,7-dihydroxy-6-MF/ Oroxylin A | Scutellaria baicalensis | Breast | 4T1 | 24 | ~90% (40 µM) | [116] |
| 5,7,4’-trihydroxy-6-MF | Artemisia capillaris | Prostate | VCaP | 48 | NA | [67] |
| 5,4’-dihidroxy-3,6,7,3'-TeMF / chrysosplenetin | Artemisia annua L. | Breast | MCF | 72 | 0.3 µM | [48] |
| 5-hydroxy-6,7,8,4'-TeMF / 5-Demethyltangeritin | Citrus reticulata | Prostate | PC3 | 48 | 11.8 µM | [63] |
| 5,3’-dihydroxy-3,6,7,4’-TeMF / casticin | Dracocephalum kotschyi Boiss | Prostate | DU145 | 48 | >80% | [68] |
**This table illustrates the cytotoxicity of all methoxyflavones analogs on various cancer cell lines. The cytotoxicity effect of methoxyflavones was centered on Nobiletin and its demethylation derivatives. Bolded IC50 indicates strong cytotoxicity activity, aligned with the proposed schematic diagram Figure 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



