Submitted:
08 October 2024
Posted:
09 October 2024
You are already at the latest version
Abstract
Fusobacterium nucleatum (F. nucleatum, Fn), an oral commensal that can become pathogenic, is associated with periodontitis (PD), adverse pregnancy outcomes, and colorectal cancer (CRC). MicroRNAs (miRNAs) are conserved, non-coding RNAs that play a regulatory role in gene expression and are detected in microbial infections. The aim of this study was to characterize the global microRNA expression kinetics in the mandibles of C57BL/6J mice infected with F. nucleatum for 8 and 16 weeks, and to identify miRNAs as biomarkers of the PD process using high-throughput NanoString nCounter® miRNA panels. Mice were divided into four groups: 8 weeks of Fn infection, 16 weeks of Fn infection, and their respective sham infection. Fn-infected mice at both 8- and 16 weeks showed 100% bacterial colonization on the gingival surface, along with a significant increase in alveolar bone resorption (p<0.0001) and intravascular dissemination to heart, indicating its invasive potential. Out of 577 miRNAs analyzed, seven miRNAs (miR-361-5p, miR-99b-5p) were upregulated, and two miRNAs (miR-362-3p and miR-720) were downregulated in the 8-week Fn infection group. In the 16-week Fn infection group, seven miRNAs (miR-361-5p, miR-99b-5p) were upregulated miRNAs and 13 miRNAs (miR-323-3p, miR-488) were downregulated. Notably, miRNAs such as miR-205, miR-210, miR-199a-3p which were differentially expressed (DE) in the Fn-infected mice at 8 weeks and miR-28 at 16 weeks, have been previously reported in human periodontitis. The 13 miRNAs induced by F. nucleatum (miR-361-5p) are linked to multiple malignancies, including esophageal, gastric, pancreatic, and colorectal cancers. Additionally, the DE miR-126-5p in the 8-week infection group has been identified as a potential biomarker for patients with PD and cardiovascular disease. The results indicate that F. nucleatum acts as a potent oncomicrobe, inducing several miRNAs associated with PD and linking it to systemic comorbidities. Furthermore, the study revealed similar expressions between PD-associated miRNAs and nine CRC tumor-expressed miRNAs.
Keywords:
F. nucleatum
; Periodontal disease
; miRNAs
; Cancer miRNAs
; Machine learning models
1. Introduction
The ubiquitous oral symbiont F. nucleatum (Fn) is a heterogeneous species with five known subspecies. It is the most predominant and abundant species in the oral cavity, present in both diseased and healthy individuals [1]. As a dominant microbe in the periodontium, it is a periodontal bacterium, a non-spore-forming obligate anaerobe, that does not possess fimbriae, pili, or flagellae, and is detected at higher levels in the oral cavity of periodontitis patients [2]. It is known to coaggregate/bridge/synergize with various microbial species and is a prominent intermediate oral pathobiont in the physical interaction and microbial complexes between Gram-positive and Gram-negative late colonizing bacteria, such as P. gingivalis, Treponema denticola, and Tannerella forsythia [3,4]. It is an intermediate colonizer that colonizes the tooth and epithelial surface [5,6], and is often implicated in various extra-oral diseases [4]. Previous in vivo subcutaneous co-infection [7] and oral co-infection studies [8] have demonstrated that the inclusion of F. nucleatum synergistically enhances bacterial virulence and disease severity. A recent study showed that F. nucleatum has invasive capacity and alters atherosclerosis risk factors (cholesterol, triglycerides, chylomicrons, VLDL, LDL, HDL), enhances inflammatory markers (CD30L, IL-4, IL-12), gene expression, and causes systemic infection in several organs with an atheroprotective immune response in ApoEnull mice [9].
One of the two most important virulence factors, Fusobacterium adhesion A (FadA) is known to play a critical role in bacterial dissemination and colonization in the placenta and is a potential diagnostic marker for F. nucleatum-associated diseases [4,10-15]. Fusobacterium apoptosis-inducing protein 2 (Fap2), an outer membrane protein involved in interspecies coaggregation, cell adhesion, cell nutrition, and antibiotic susceptibility is a large type V autotransporter. Furthermore, F. nucleatum induces a chronic inflammatory response by stimulating the production of pro-inflammatory cytokines (interleukin-1β, IL-6, IL-8, and MMPs) in gingival epithelial cells [16,17]. Several studies have also reported the association of F. nucleatum with pregnancy complications, including chorioamnionitis, spontaneous abortion, preterm birth, stillbirth, neonatal sepsis, and preeclampsia [18]. Emerging reports in the last two decades indicate a significant link between Fn (an oncomicrobe) and the progression of multiple digestive tract cancers, including esophageal [19], gastric [20], pancreatic [21], CRC [12,22], lung cancers [23], and oral squamous cell carcinoma (OSCC) [24]. Fn interacts with the molecular hallmarks of gastrointestinal cancers, inducing genomic mutations and promoting a permissive immune microenvironment by impairing anti-tumor checkpoints [25]. It is unknown how F. nucleatum, which is adapted as a symbiont in the oral microenvironment, can migrate to secondary extraoral niches by possessing gene regulatory mechanisms that allow it to thrive and proliferate in tumor environments.
microRNAs (miRNAs) are small (21-25 nucleotides) noncoding, regulatory RNAs that regulate gene expression by directly binding to the 3′ untranslated regions of their target miRNAs [26,27]. Approximately 60% of all genes in each mammalian genome are estimated to be regulated by a total of 2000 miRNAs [28,29]. Several studies on miRNAs have implicated them in many human systemic diseases. They regulate the immune response of hosts infected with exogenous pathogenic bacteria such as Helicobacter pylori [30], Treponema pallidum [31], M. tuberculosis [32], Salmonella [33], Listeria monocytogenes [34], Mycobacterium avium [35], and endogenous periodontal microbiome [36-39]. Hence understanding miRNA expression patterns could possibly lead to the development of novel diagnostic and therapeutic biomarkers for complicated inflammatory diseases driven by microbial infections, including periodontal disease (PD). Gingival tissue from chronic inflammatory PD has revealed a panel of microRNAs (hsa-miR-223-3p, hsa-miR-203b-5p, hsa-miR-146a-5p, hsa-miR-146b-5p, and hsa-miR-155-5p) [40] and also miR-146a in a rodent model [41]. Hence, understanding the expression pattern of miRNAs could potentially lead to the development of novel diagnostic biomarkers for PD. Recently, we reported sex-specific differential miRNA expression (miR-9, miR-148a, miR-669a, miR-199a-3p, miR-1274a, miR-377, miR-690) in mice infected with partial human mouth microbes (PAHMM) using a novel ecological time-sequential polybacterial periodontal infection (ETSPPI) mouse model [39]. In addition, we have also reported individual monobacterial periodontal infections with P. gingivalis, T. denticola, T. forsythia, S. gordonii in the mouse model, and identified several miRNAs linked with PD, various systemic diseases, and malignancies [36-38]. To date, the role of miRNAs in response to oral infection by the periodontal microbe F. nucleatum in a mouse model of PD has not been examined. Accordingly, this study aimed to enhance our understanding of whether intraoral infection of mice with F. nucleatum could lead to unique alterations in miRNA expression patterns and to assess the link between miRNAs and F. nucleatum-induced PD and multiple tumors. The present study was designed to analyze miRNA differential expression (DE) kinetics at two-time points (8 weeks and 16 weeks) in F. nucleatum-infected male and female C67BL/6J mice using high-throughput NanoString analysis with nCounter miRNA expression profiling. Furthermore, we employed the machine-learning (ML) algorithms XG Boost (XGB), Random Forest classifier (RFC), Logistic Regression (LR), Support Vector Classifier (SVC), and Multilayer Perceptron (MLP) to deepen our understanding of the complex interplay between Fn infection, PD, and oncogenic miRNAs. In recent studies, we analyzed miRNAs in response to oral monobacterial infection in the mouse model using several ML algorithms [38,42]. This approach provided deeper insights into the specific miRNAs associated with PD, its systemic comorbidities, and multiple tumors.
2. Results
2.1. Chronic Infection of F. nucleatum, Colonized in Mice Gingival Tissue
Intraoral infection of F. nucleatum induced time-dependent colonization on the gingiva and facilitated periodontal miRNA expression. Time-dependent gingival colonization for P. gingivalis, T. denticola, T. forsythia, and S. gordonii was observed in our earlier studies [36-38,42]. Fn bacterial colonization-initiated PD on chronic intraoral infection in a time-dependent manner. Testing of oral swab collected from the gingival plaque for 16S rRNA gene-specific PCR confirmed that gingival plaque samples in Group I mice had 50% Fn specific DNA at the 2 weeks of intraoral infection time point and tested positive for all the mice after 6 weeks of Fn infection. Group III mice were 60%, 90%, and 100% positive for Fn after 2-, 4-, and 6 weeks of infections, respectively (Table 1).
2.2. Alveolar Bone Resorption and Bacterial Genomic DNA in the Distal Organs
The pathogenic factors of periodontal bacteria and inflammatory cytokines mediated activation of osteoclasts together contribute to ABR [43]. Microscopic images of the mandibles in F. nucleatum-infected and sham-infected groups are shown in Figure 1B. Mice infected with F. nucleatum at both 8 weeks and 16 weeks’ time points showed significantly higher ABR in the mandible (lingual) p < 0.05 (Adjusted p-value = 0.001) for the 16 weeks group (Figure 1B-C). ABR measured in the maxilla did not show a significant difference between sham versus Fn infection in 8- and 16 weeks infection time points. Bacteria-specific DNA from 8 weeks F. nucleatum infection was identified in the heart (Table S1) in 3/10, in lung 2/10, and 1/10 in the liver. In the 16 weeks of infected mice, F. nucleatum gDNA was detected in 5/10 in the heart, in lungs 6/10, and 1/10 in the kidney (Supplementary Table S1). This finding suggests the physiological colonization/infection of the gingival epithelium and the intravascular dissemination of bacteria to the heart and distal organs, representing the invasive potential of F. nucleatum.
2.3. NanoString Analysis of miRNA in F. nucleatum-Infected Mandibles
The NanoString platform is an amplification-free technology using molecular barcodes and directly quantifies the RNA molecules without the events of reverse transcription and is strongly reliable in working with multiple sample types. The total RNA extracted from purified mandibles was further analyzed for global miRNA profiling in the 8 weeks and 16 weeks of F. nucleatum-infected mice (Table 2). nCounter miRNA expression profiling showed 7 upregulated miRNAs including most significantly miR-361-5p, miR-99b-5p and 2 downregulated miRNAs of miR-362-3p, miR-720 in 8 weeks of F. nucleatum infected mandibles compared to sham-infected mandibles (Table 2). Similarly, a total of 7 upregulated DE miRNAs (miR-361-5p, miR-99b-5p) and 13 downregulated miRNAs (miR-323-3p, miR-488) shown in 16 weeks of F. nucleatum infected mandibles compare to the sham-infected mandibles. The analysis between F. nucleatum infected female vs male mice showed 12 upregulated (miR-206, miR-210) and 12 downregulated miRNAs (miR-376a, miR-350) in the 8 weeks group and 5 upregulated (miR-152, miR-125b-5p) and 14 downregulated miRNAs (miR-375, miR-376a) in the 16 weeks infection group. The upregulated miR-210 in the 8 weeks Fn-infected female mice was found downregulated in the 16 weeks infected female mice. (Table 2). A p-value of < 0.05 and a fold change of 1.1 and above was considered for analysis and to be significant. DE upregulated miRNAs between 8- and 16 weeks analysis target function and target genes shown in Table 3 and Table 4 and downregulated miRNAs shown in supplementary table S2. The list of DE miRNAs (upregulated and downregulated) for 8- and 16 weeks analysis was shown in the supplementary Table S3. The upregulated and downregulated miRNAs in the female vs male comparison study for the 8 weeks of infection were shown in supplementary Table S4 and for 16 weeks was shown in Table S5.
2.4. Identification of Differentially Expressed (DE) miRNAs
Two Fn-infected (40 samples) and two sham-infected (20 samples) groups were analyzed using high throughput nCounter® miRNA Expression Panels. we performed volcano plot analysis to identify DE-miRNAs with statistical significance. It was plotted against log2 fold change on the x-axis and the negative log of p-value on the y-axis. The identified downregulated miRNAs (47) indicated in red and 14 upregulated miRNAs (green) showed a fold difference of +1.1 with a p-value of < 0.05 in 8 weeks F. nucleatum-infected mice compared to 16 weeks infected group (Figure 2A). All the black dots represent miRNAs that do not pass the filter parameters. In the 8 weeks infection study, 7 miRNAs showed higher expression (e.g., miR-361-5p, miR-99b-5p), and 7 miRNAs (e.g., miR-361-5p, miR-99b-5p) showed higher expression during 16 weeks of F. nucleatum-infected mandibles (Figure 2B, Table 3 and Table 4).
Upregulated miRNAs reported in the present 8 weeks study have been associated as diagnostic miRNAs in humans (miR-361-5p), identified in the gingiva of PD patients (miR-26a-5p), attenuating the sepsis-induced myocardial injury in the mice (miR-193a-3p), associated with heart failure diagnosis in human subjects (miR-324-5p), regulating the cell survival in periodontal ligament cells (miR-24-3p), associated in mice PD infections induced by T. denticola (miR-126-5p) and P. gingivalis (miR-99b-5p). The 16 weeks upregulated miRNAs were associated as PD diagnostic miRNA in humans (miR-361-5p), gingival tissue in chronic periodontitis patients (let-7a-5p), in the human periodontitis gingival tissue (let-7f-5p), reliable biomarker in patients with oral squamous cell carcinoma (miR-345-5p), clinical marker for atherosclerosis (miR-218-5p) and associated in mice PD infections induced by P. gingivalis (miR-99b-5p).

Table 3.
F. nucleatum-infection-induced upregulated miRNAs (8 weeks), reported functions, and target genes.
Table 3.
F. nucleatum-infection-induced upregulated miRNAs (8 weeks), reported functions, and target genes.
| miRs /(FC) |
p-value | Reported Functions | Target genes |
|---|---|---|---|
| miR-361 -5p (1.3) | 0.0311 | Downregulated in the 13 types of cancer, including CRC [44]. Upregulated in-stable coronary heart disease, acute coronary heart syndrome [45]. | 16 (E.g., Ctbp2, Tfam, Nol7) |
| miR-26a-5p -5p (1.23) | 0.0008 | Downregulated in the gingiva of periodontitis patients [46]. Associated with cardiovascular diseases [47]. | 426 (e.g., Kpna2, Nus1, Rgs17) |
| miR193a -3p (1.21) | 0.0150 | Downregulated in plasma and salivary exosomes of Chronic periodontitis patients [48]. Downregulated in the P. gingivalis-LPS-treated human periodontal ligament cells [49]. Upregulated miR-193 improved cardiac function, and attenuated myocardial injury, inflammation, and cardiomyocyte apoptosis in septic mice [50]. Dysregulated in Pancreatic cancer [51]; colorectal cancer [52]; endometrial cancer [53]. A novel biomarker for Alzheimer's disease diagnosis [54]. Downregulated in the microbiota-mediated ulcerative colitis [55]. | 8(e.g., Gla, Ifngr2, Metap2) |
| miR-126 -5p (1.2) | 0.0308 | Upregulated miR in T. denticola-induced periodontitis [37]. Preventing alveolar bone resorption in diabetic periodontitis [56]. Upregulated miR in the gingiva of PD patients [57]. Associated in colorectal cancer [58]; promoting chemoresistance of ovarian cancer cells [59]. Upregulated in coronary artery ectasia patients [60]. Upregulated in thoracic aorta aneurism patients [61]. Reported as a molecular target for myocardial infarction treatment [62]. Reported as a poultry meat quality and food safety marker miRNA [63]. | 55 (e.g., Gfpt1, H2-D1, Il10rb, Kras) |
|
miR-324 -5p (1.18) |
0.0164 | Downregulated in the gingival tissue of periodontitis patients [64]. Reported as a therapeutic-miR in cervical cancer, colorectal cancer, gastric cancer, brain tumors, and hepatocellular carcinoma [65]. Human cases with heart failure have reported AUC for miR-324-5p and are considered as a diagnostic miR [66]. Downregulated in osteoporosis patients [67]. | 12 (E.g., Zfp295, Zscan12, Kcnk6) |
|
miR-24-3p (1.17) |
0.0160 | Downregulated in the unstimulated saliva of chronic periodontitis patients [48]. Reported as a defensive miR against periodontal inflammation [68]. A diagnostic marker for multiple cancers [69]. | 375 (e.g., Oxt, Chrna1, Birc5) |
| miR-99b -5p (1.14) | 0.0120 | Downregulated in the gingiva of periodontitis patients [64]. Upregulated in the P. gingivalis-induced periodontitis [36]. Downregulated in the saliva of chronic periodontitis patients [48]. Upregulated in M. tuberculosis-infected murine dendritic cells [70]. Novel chemosensitizing miRNAs in high-risk neuroblastoma [71,72]. Associated in the cancer conditions to ovaries [73]; prostate [74]; colorectal [75]; gastric [76]; liver [77]; and lungs [78]. | 4 (E.g., Comp, Grik3, Slc35d2) |
The upregulated miRNAs for 8 weeks of F. nucleatum-infection were associated with aggressive periodontitis in the saliva and gingival tissue of human subjects (e.g., miR-26a-5p; miR-193a-3p). Bold red, blue, and green color indicates miRNAs associated with multiple cancers, periodontitis, and its systemic comorbidities, respectively.
Table 4.
F. nucleatum-infection induced upregulated miRNAs (16 weeks), reported functions, and target genes.
Table 4.
F. nucleatum-infection induced upregulated miRNAs (16 weeks), reported functions, and target genes.
| miRs /FC |
p-value | Reported Functions | Target genes |
|---|---|---|---|
| let-7a-5p (1.28) |
0.0011 | Downregulated in T. denticola-induced periodontitis [37]. Downregulated in the saliva of patients with aggressive periodontitis [79]. Upregulated in gingival tissue of chronic periodontitis patients [80]. IL-13 a cytokine essential for allergic lung diseases is regulated by mmu-let-7a-5p [81]. Downregulated in bronchial biopsy of severe Asthma patients [82]. | 28 (e.g., Lin28a, IL6, Hoxa9) |
| miR-127 -3p (1.28) | 0.0217 | Upregulated in T. forsythia-induced rodent periodontitis models [38]. Upregulated in the inflamed primary human gingival fibroblasts [83]. Upregulated in the human advanced carotid atheroma [84]. May play an important role in acute myocardial injury [85]. Tumor suppressor miRNA in triple-negative breast cancer cells [86]. | 10 (E.g., Rtl1, Gpi1, Ghdc) |
| miR-361 -5p (1.19) | 0.0357 | Reported in 8 weeks analysis study | |
|
miR-345 -5p (1.16) |
0.0034 | Reliable biomarker in patients with oral squamous cell carcinoma [87]. Acting as an anti-inflammatory miRNA in mice with allergic rhinitis [88]. Reported as a protective miRNA during gestational diabetes mellitus subjects [89]. | 10 (e.g., Ccdc127, Eaf1, Atic) |
| let-7f-5p (1.16) |
0.0298 | Upregulated in human periodontitis gingival tissue [57]. Potential biomarker for abdominal aortic aneurysm [90]. Involvement in the pathogenesis of SLE-lupus nephritis [91]. | 16 (e.g., Atp2b2, Ifnar1, Nf2). |
| miR-99b -5p (1.15) | 0.0299 | Reported in 8 weeks analysis study | |
| miR-218 -5p (1.15) | 0.0333 | Upregulated in the T. forsythia-induced rodent periodontitis models [38]. Downregulated in the gingival fibroblasts and is essential for myofibroblast differentiation [92]. Upregulated in the inflamed gingiva [93]. Reduced expression was observed in the atherosclerosis cohort and considered a clinical marker for atherosclerosis [94]. Downregulated in smokers without airflow limitation and in patients with chronic obstructive pulmonary disease [95]. | 20 (e.g., Epg5, Prdm1, Bsn, Eno2) |
Details of the biological function and target genes were given for the top ten significantly expressed miRNAs in 16 weeks infected mice mandibles. The upregulated miRNAs for 16 weeks of F. nucleatum-infection were associated with aggressive periodontitis in the saliva and gingival tissue of human subjects (mmu-let-7a-5p; mmu-let-7f-5p), acute myocardial injury (miR-127-3p), and clinical marker for atherosclerosis (miR-218-5p). Bold red, blue, and green color indicates miRNAs associated with multiple cancers, periodontitis, and its systemic comorbidities, respectively.
2.5. DE miRNAs and Functional Pathway Analysis
Functional enrichment analysis for the up-and down-regulated miRNAs was performed using DIANA-miRPath software to predict the biological function. The resulting KEGG pathway analysis revealed that the upregulated miRs in the 8 weeks analysis were associated with the Pathways in cancer, Axon guidance pathway, ErbR signaling pathway, Renal cell carcinoma, mTOR signaling pathway, Porphyrin metabolism, Adherens junction, Glioma, cAMP signaling pathway, etc. (Figure 2C). The upregulated miRNAs in the 16 weeks analysis (miR-99b-5p, miR-361-5p, miR-345-5p, miR-218-5p and miR-127-3p) associated with the proteoglycans in cancer, pathways in cancer, axon guidance pathway, cell adhesion molecules pathway, adherens junction pathway, GABAergic synapse pathway, circadian entrainment, bacterial invasion of epithelial cells and proteoglycans in cancer (Figure 2D). The miRs of miR-361-5p (Cblb, Rhoa, Rac1), miR-218-5p (Pik3r1, Arpc1b, Elmo1, Shc4), and miR-345-5p (Pxn) were associated with bacterial invasion of epithelial cells pathways having the target genes in the brackets (Figure 3 and Table S6). Pathways in cancer in both 8- and 16 weeks upregulated miRNAs have a single miRNA (miR-361-5p) that has the potential to interact with all of the genes in the pathway. The DE miRNAs upregulated in the 8- and 16 weeks of Fn infection have a regulatory role on 50, and 37 genes (Figure 4) respectively in the pathways of cancer. The miR-361-5p which is associated with pathways in cancer has a regulatory target on 11 genes including the Crebbp gene which has the unusual largest interactions in the pathways of cancer. The reviewed information revealed that genetic aberrations of CREBBP/EP300 were observed in various types of solid tumors and hematologic malignancies and considered promising therapeutic targets [96]. Detailed reported functions of the upregulated miRNAs during 8- and 16 weeks of Fn infection are shown in Table 3 and Table 4, respectively. The list of upregulated miRNAs associated with bacterial invasion of epithelial cells (Supplementary Table S6), and pathways in cancers are shown in Tables S7 and S8. The number of target genes for each upregulated miRNA in the 8- and 16 weeks infection group was analyzed using miRTarBase. We used mmu-miR-361-5p as the example for an upregulated DE miRNA during 8 weeks of infection in identifying the target genes using miRTarBase (Supplementary Table S9). Each miRNA has different target genes along with a specific miRTarBase ID. F. nucleatum-infection induced DE-upregulated mmu-miR-361-5p has 15 different target genes with 15 different miRTarBase IDs as stated in Table S9. The list of target genes for 16 weeks upregulated miRNAs and the miRTarBase IDs are shown in supplementary Table S10.
2.6. Machine Learning Analysis of NanoString miRNA Copies
Our ML analysis results are divided into two groups: tree-based methods consisting of XGB and RFC, and non-tree-based methods consisting of LR, SVC, and MLP. In the 8 weeks dataset, mmu-miR-1899 had the highest impact on the tree-based models (Figure 4A1, XGB; Figure 4B1, RFC), LR and SCV were most impacted by mmu-miR-22, and miR-720 in MLP had the highest impact (Figure 4C1, LR; Figure 4D1, SVC; Figure 4E1, MLP). For the 16 weeks dataset, mmu-miR-339-5p had the highest impact on the tree-based models (XGB, RFC; Figure 4A2 and Figure 4B2), while mmu-miR-7a had the highest impact on LR and SVC, and miR-1 in MLP was most impacted by miR-1 (LR, SVC, Figure 4C2, Figure 4D2 and MLP; Figure 4E2). Finally, in the combined 8- and 16 weeks dataset, the most impactful miRNAs were mmu-miR-26a-5p in XGB, miR-704 in RFC was again the most impactful for the tree-based models (XGB, RFC; Figure 4A3 and Figure 4B3), mmu-miR-22 in LR and SVC, and mmu-miR-720 in MLP (LR, SVC, Figure 4C3, Figure 4D3 and MLP, Figure 4E3). The results of the SHAP value analysis are summarized in Supplementary figure S1-5, Table 5 summarizes the most impactful miRNAs for each ML model as well as descriptions of the miRNA functions, and Table 6 summarizes the top five important miRNAs.
Table 5.
Summary of the miRNA and its importance for each machine learning model.
| miRNA (Feature Rank) |
ML Model | MIMAT# | Target Functions |
|---|---|---|---|
| 8 Weeks Analysis | |||
| mirR-22 (1) | LR, SVC | MIMAT0000531 | Upregulated in periodontal disease and obesity [97]. |
| miR-205 (2) | LR, SVC, MLP | MIMAT0000238 | Downregulated in chronic periodontitis patients [98,99]. |
| miR-720 (3) | LR, SVC | MIMAT0003484 | Novel miRNA regulating the differentiation of Dental pulp cells [100]. |
| miR-1899 (1) | XGB, RFC | MIMAT0007869 | |
| 16 Weeks Analysis | |||
| let-7a-5p (1) | LR, SVC | MIMAT0000521 | Downregulated in aggressive periodontitis patients [79]. Promotes the osteogenesis of bone marrow mesenchymal stem cells [101]. |
| miR-1 (2) | LR, SVC | MIMAT0000123 | Noninvasive biomarker for breast cancer [102]. Upregulated in patients with myocardial infarction [103]. |
| miR-22 (3) | LR, SVC | MIMAT0000531 | Shown in the 8 weeks analysis |
| miR-205 (4) | LR, SVC | MIMAT0000238 | Shown in the 8 weeks analysis |
| miR-125b-5p (5) | LR, SVC | MIMAT0000135 | Associated with osteogenic differentiation [104]. Overlapping miRNA between periodontitis and Nonalcoholic fatty liver disease [105]. |
| miR-339-5p (1) | XGB, RFC | MIMAT0000584 | Most predictive periodontal miRNA in the T. forsythia-induced mice periodontitis [38]. |
| 8- and 16 Weeks Analysis | |||
| miR-22 (1) | LR, SVC | MIMAT0000531 | Shown in the 8 weeks analysis |
| miR-133a (2) | LR, MLP | MIMAT0000145 | Upregulated in the T. denticola-induced mice periodontitis [37]. |
| miR-1 (4) | SVC, MLP | MIMAT0000123 | Shown in the 16 weeks analysis |
The rank (or importance) of the miRNA in predicting if the mouse was infected is in parenthesis. The higher the rank, the more important the miRNA was for making the prediction. For the 8- and 16 weeks cohort XGB and RFC did not agree on the importance of a miRNA. Models: logistic regression (LR), C-support vector classifier (SVC), multilayer perceptron (MLP), random forest classifier (RFC), and extreme gradient boosting (XGB).
In addition to determining which miRNAs were most impactful on the ML models (Table 5), we also found high levels of agreement in the miRNAs ranked 2 to 5 (Table 6). Interestingly, the tree-based models did not agree on the importance of miRNAs for ranks between 2-5 in the 8 weeks, 16 weeks, and the combined 8- and 16 weeks data sets. In the 8 weeks dataset, the non-tree-based models ranked miR-205 as the second most important feature, and LR and SVC ranked miR-720 third. In the 16 weeks dataset, LR and SVC ranked miR-1 as second, miR-22 as third, miR-205 as fourth, and miR-125b-5p as the fifth most important feature. Lastly, in the combined 8- and 16 weeks dataset, LR and MLP ranked miR-133a as the second most important feature, and SVC and MLP ranked miR-1 as fourth.
Table 6.
Summary of the importance and miRNA for each machine learning model.
| miRNA Feature Rank | ||||||
|---|---|---|---|---|---|---|
| Cohort | miRNA | 1 | 2 | 3 | 4 | 5 |
| 8 weeks | miR-22 | LR, SVC | MLP | |||
| miR-205 | LR, SVC, MLP | |||||
| miR-720 | MLP | LR, SVC | ||||
| miR-1 | LR | MLP | ||||
| let-7c | MLP | LR | ||||
| miR-125b-5p | SVC | |||||
| miR-148a | SVC | |||||
| miR-1899 | XGB, RFC | |||||
| miR-670 | XGB | |||||
| miR-706 | XGB | |||||
| miR-878-3p | XGB | |||||
| miR-302b | XGB | |||||
| miR-453 | RFC | |||||
| miR-1970 | RFC | |||||
| miR-694 | RFC | |||||
| miR-m108-2-5p.2 | RFC | |||||
| 16 weeks | let-7a-5p | LR, SVC | MLP | |||
| miR-1 | MLP | LR, SVC | ||||
| miR-22 | MLP | LR, SVC | ||||
| miR-205 | LR, SVC | |||||
| miR-125b-5p | LR, SVC | |||||
| miR-720 | MLP | |||||
| let-7b | MLP | |||||
| miR-339-5p | XGB, RFC | |||||
| miR-878-3p | XGB | |||||
| miR-323-3p | XGB | |||||
| miR-128 | XGB | |||||
| miR-18a | XGB | |||||
| 8 and 16 weeks |
miR-22 | LR, SVC | MLP | |||
| miR-133a | LR, MLP | SVC | ||||
| miR-1 | LR | SVC, MLP | ||||
| miR-205 | SVC | LR | ||||
| miR-16 | LR | |||||
| miR-720 | MLP | |||||
| let-7a-5p | SVC | |||||
| let-7c | MLP | |||||
| miR-26a-5p | XGB | RFC | ||||
| miR-128 | XGB | |||||
| miR-1971 | XGB | |||||
| miR-18a | XGB | |||||
| miR-743b-5p | XGB | |||||
| miR-704 | RFC | |||||
| miR-324-5p | RFC | |||||
| miR-762 | RFC | |||||
| miR-130b | RFC | |||||
The higher the rank, the more important the model found the miRNA to be for predicting if the mouse was infected. Models: logistic regression (LR), C-support vector classifier (SVC), multilayer perceptron (MLP), random forest classifier (RFC), and extreme gradient boosting (XGB). LR-Logistic regression, SVC-C-support vector classifier, MLP-multilayer perceptron, RFC-random forest classifier, and XGB-extreme gradient boosting.
3. Discussion
Prior chronic oral infection established F. nucleatum colonization in the oral cavity, induced significant humoral IgG and IgM antibody response, and resulted in significant ABR and detection of genomic DNA in systemic organs (heart, aorta, liver, kidney, lung) indicating bacteremia [9]. In addition, vascular inflammation was detected by enhanced systemic cytokines (CD30L, IL-4, IL-12), oxidized LDL and serum amyloid A, as well as an altered serum lipid profile (cholesterol, triglycerides, chylomicrons, VLDL, LDL, HDL), in infected mice. Altered aortic gene expression in infected ApoEnull hyperlipidemic mice further indicated its virulence [9]. The frequent involvement of F. nucleatum in extra-oral systemic infections and comorbidities [2] including adverse pregnancy outcomes, and various malignancies such as CRC [12,22] supports a role for this species in multiple disease pathogenesis. This is because bacteria can survive, spread hematogenously, and replicate at sites distant from the oral cavity. F. nucleatum uses its FadA adhesion to bind to and invade both endothelial cells and epithelial cells [16], giving it a high potential for actively inducing PD and its systemic comorbidities. Although the etiologic role of F. nucleatum as an important bacterium in the progression of PD is known, the underlying molecular genetic mechanisms are not completely understood. Recently, several studies have suggested that aberrant expression of miRNAs is involved in both infectious and non-infectious diseases, influencing the initiation and progression of pathology and the development of miRNAs as diagnostic biomarker in CRC. To the best of our knowledge, this study will be the first investigation to examine F. nucleatum oral infection, global miRNA induction, and genes involved in periodontitis.
In this report, we focused on how F. nucleatum selectively modulates host gingival epithelial cell responses with robust and specific miRNA expression during the progression of PD. We demonstrate that chronic oral infection with F. nucleatum in male and female mice results in physiological colonization/infection of the gingival surfaces after 4 infection cycles, invades gingival epithelium, and induces PD outcome measures such as ABR, which are significantly higher in mice at both 8- and 16 weeks infection time points. Additionally, F. nucleatum is able to spread via intravascular dissemination to distal organs, including the heart suggesting its invasive potential and ability to modulate host immune response. This invasive potential to the heart and other tissues was robust when F. nucleatum was administered with late colonizers such as P. gingivalis, T. denticola, and T. forsythia and the early colonizer S. gordonii [39], suggesting their physiological, nutritional, and metabolic synergistic interactions in the gingiva. This co-dependence enhances microbial multiplication and invasion of gingival epithelium, leading to systemic intravascular dissemination. The partial human mouth microbes (PHAMM) ecological time sequential polybacterial periodontal infection (ETSPPI) model involves an array of five bacteria-mediated intraoral infection, which colonizes bacteria on the gingiva, induces periodontitis and leads to induced sex-specific miRNA expression. Several of these miRNAs are linked to numerous systemic diseases comorbidities [39]. Monobacterial intraoral infections with P. gingivalis, T. denticola, T. forsythia, and S. gordonii also induced periodontitis with robust alterations in miRNAs in mice models [36-38,42].
This study analyzed 577 mouse miRNAs in mandibles from Fn-infected and sham-infected mice using the high-throughput NanoString nCounter miRNA profiling. Most of the DE miRNAs were unique, except for two miRNAs (361, 99b), and specific to the time point, indicating that miRNA induction is transient and time-dependent. A total of 29 miRNAs were DE in mandible tissue during 8- and 16 weeks F. nucleatum infection compared with sham infection. Of these, 14 miRNAs were upregulated and 15 miRNAs were downregulated. Among all the 14 upregulated miRNAs at 8- and 16 weeks, 4 miRNAs were reported in human periodontitis studies, indicating that these upregulated miRNAs are associated with the induction of PD. Specifically, miR-26a-5p was downregulated in the gingiva of periodontitis patients [46], mmu-let-7a-5p was found in the saliva of patients with aggressive periodontitis [79] and in the gingival tissue of chronic periodontitis patients [80], and mmu-let-7f-5p was found in human periodontitis gingival tissue [57], indicating that preclinical in vivo miRNA data corroborate with clinical PD miRNA data. Three of the upregulated miRNAs have been associated with preclinical mouse studies; miR-99b-5p in P. gingivalis-induced PD [36], miR-126-5p in T. denticola-induced PD [37], and miR-127-3p in T. forsythia-induced PD [38], and were also expressed during F. nucleatum infection. Furthermore, the two miRNAs, miR-361-5p and miR-99b-5p, were commonly expressed and upregulated at both time points.
In addition, the miRNAs miR-361-5p, miR-193a-3p, miR-324-5p, miR-99, miR-127-3p, miR-345-5p, miR-218-5p, let-7f-5p are associated with cardiovascular diseases [45,50,66,85,94,106-108] miR-324-5p and miR-345-5p are linked to adipocyte differentiation [109,110], while miR-127-3p, miR-345-5p, and miR-99 are associated with tumor malignancies [86,87,111-113]. Interestingly, a single miRNA can be associated with different disease conditions. For example, miR-324-5p is linked to osteoporosis [67], and multiple myeloma [114], let-7a-5p is associated with various inflammatory conditions [81,115], chemotherapy-exposed mice ovaries [116], an antifibrotic role under hypoxic stress [117], and asthma [82]. miR-24-3p is involved in phagocytosis [68], periodontal ligament cells [118], and gingival fibroblasts [92], let-7f-5p is linked to abdominal aortic aneurysm [90] and lupus nephritis [91]. Different miRNAs are also associated with microbial infection-exposed immune cells. For example, miR-99 is linked to M. tuberculosis-infected murine dendritic cells [70], miR-127-3p is involved in macrophage anti-microbial responses [119], and miR-24-3p is associated with S. aureus caused-osteomyelitis [120].
Recent reports clearly demonstrate microbiological, molecular, and genetic links between Fn and multiple digestive tract cancers including esophageal, gastric, pancreatic, and CRC. Interestingly, our study showed 37 significantly increased gene expressions in multiple cancers and several miRNAs associated with both multiple cancers including CRC and PD in humans [121], some of which also link PD with inflammatory systemic comorbidities. Several studies identified oncogenic marker miRNAs (oncomiRs) in CRC tumors (miR-126-5p, miR-99b-5p, miR-26a-5p, miR-24-3p, miR-361-5p, miR-193a-3p, miR-218-5p, let-7a-5p, and let-7f-5p), which were also expressed in mouse mandibles during F. nucleatum infection. This indicates a link between invasive F. nucleatum and multiple tumors, including CRC, further strengthening the association between periodontitis and CRC. In contrast, P. gingivalis oral infection in mice showed DE miR-185, miR-22, miR-152, miR-423, miR-151, miR-28, and miR-145, which were also found to be upregulated in various malignancies such as prostate, breast, glioma, gastric, and hepatocellular carcinoma [36]. This data indicates that periodontal bacteria-expressed miRNA in gingival tissues and other systemic diseases have some common mRNA targets that mediate disease progression.
KEGG pathway analysis revealed that the most target genes of upregulated miRNAs during the 8-week period were linked with several pathways, including those involved in cancer, axon guidance, GABAergic synapse, proteoglycans in cancer, drug metabolism-cytochrome P450, ErbR signaling pathway, acute myeloid leukemia, phosphatidylinositol signaling system, renal cell carcinoma, adherens junction, glioma, and cAMP signaling pathway. Additionally, 10 genes were found to be associated with F. nucleatum bacterial invasion of epithelial cells and regulate the gene expression that mediates invasion of gingival epithelial cells, regulating gene expression that mediates the invasion of gingival epithelial cells. Other critical pathways linked with bacterial infection and host-cell associations were also identified.
Periodontal disease-causing infectious oral microbes are commonly detected in atherosclerotic plaques and are associated with many systemic diseases. F. nucleatum is linked with more systemic diseases than other known periodontal microbes and has been isolated from more than 10 cites if systemic infection [2]. Periodontal infection is associated with atherosclerosis, where atherosclerotic arteries of patients tested positive for F. nucleatum genomic DNA (84%) [122]. There is evidence that F. nucleatum sub-species have fine-tuning capabilities to drive disease progression and niche colonization in the disease habitat. Zepeda-Rivera et al. revealed that the F. nucleatum strains causing colorectal cancer (Fna C2 strains) were present in the oral cavity of patients with colorectal cancer [121]. Polymicrobial oral infection in the mice, including F. nucleatum as a member, caused the hematogenous dissemination of Fn organisms into the heart and aorta [123]. In rodent models, the predicted hypothesis of transient bacteremia caused by F. nucleatum facilitated its transmission from the oral cavity to the uterus, resulting in premature delivery, stillbirths, and no sustained live births [124]. The abundance/enrichment of F. nucleatum in colorectal tumor specimens confirmed its correlation with colorectal cancer [125]. The tumor miRNAs widely observed among the polybacterial infection and different monobacterial infections were analyzed and presented in Table S9.
The machine learning model analysis for F. nucleatum showed some similarities with the ML algorithm in the S. gordonii (Sg) monoinfection. In our present study, tree-based models identified miR-339-5p and miR-323-3p as significant features, which were also observed in S. gordonii-induced PD mice. The miRNA features miR-22, miR-205, miR-720, miR-1, mmu-let-7c, and mmu-miR-7a were identified in non-tree-based models in both the S. gordonii study and our present study. The miR-125b-5p feature was uniquely observed in the MLP model of S. gordonii infection and in the LR, and CVC models of our present study. Similarly, miR-133a was an observed feature in the CVC model of the S. gordonii PD studies and was reported in the non-tree-based models of our present study [42]. Additionally, tree-based models in our present study and in T. forsythia-induced PD mice identified common features including miR-18a, miR-339-5p, miR-130b, and miR-704 [38]. We acknowledge that further research is necessary to elucidate how Fn influence the development of PD versus cancer. Further, we recognize that key factors may still be unidentified which direct the progression towards PD rather than cancer, despite the robust induction of the many oncogenic miRNAs. This area requires more investigation to fully understand the underlying mechanisms and potential therapeutic targets.
4. Materials and Methods
4.1. Animal Models, Ethical Statement, and Grouping
Male (20) and female (20) wild-type C57BL/J mice aged 8 weeks were purchased from the Jackson Laboratory. All the animal procedures were performed according to the guidelines of the University of Florida Institutional Animal Care and Use Committee protocol # 202200000223. Mice were divided into four groups (n = 10; five male and five female), Group I was infected with F. nucleatum for 8 weeks; Group II was sham-infected for 8 weeks; Group III was infected with F. nucleatum for 16 weeks and Group IV was sham-infected mice for 16 weeks (Table 1).
4.2. Bacterial Strain, Culture, and Mice-Oral Administration
F. nucleatum ATCC 49256 subspecies vincentii was isolated from periodontal pocket [126] grown on blood agar plates in a Coy anaerobic chamber at 37°C for 2-3 days, harvested, and prepared for intra-oral infections in mice as described previously [9,39,127-130]. Mice in Group I was intraorally infected with 108 Fn cells/mouse, suspended in reduced transport fluid (RTF) and equal volumes of carboxymethyl cellulose (6% CMC) for four times a week every other week for a total of 8 weeks. In Group III, the infection scheme was for a total of 16 weeks. Group II and IV mice were treated 1:1 ratio of RTF and 6% CMC as sham infection for 8 and 16 weeks, respectively. After the designated infection period, mice were euthanized (CO2 inhalation), and blood, and organ samples (mandibles, maxilla, brain, heart, liver, lungs, spleen, and kidney) were collected. RNAlater solution was used to preserve the left maxilla and mandibles for miRNA analysis, whereas the right maxilla and mandibles for horizontal alveolar bone resorption (ABR) morphometry measurements [39].
4.3. DNA Isolation and Molecular Detection of Bacteria Genome
Gingival plaque samples from the mice that received oral Fn infection were collected in Tris EDTA (TE) buffer [39,131]. Fn genomic DNA (gDNA) present in the oral plaque samples and the distal organs of the heart, lungs, brain, liver, kidney, and spleen were detected using 16S rRNA gene-specific primers 5′-TAAAGCGCGTCTAGGTGGTT-3′, and reverse primer 5′-ACAGCTTTGCGACTCTCTGT-3′ [39]. Colony PCR was adopted to detect Fn gDNA in the gingival plaque samples. Qiagen Dneasy Blood and Tissue Kit (Qiagen, Germantown, MD, USA) was used to extract the gDNA from the distal organs and stored at –20°C [39]. F. nucleatum culture DNA was used as positive control and sterile milli Q water was considered negative control in the PCR reaction. The amplified Fn-specific DNA was run through agarose electrophoresis and visualized in the UVP GelStudio touch Imaging System (Analytik Jena US LLC, CA, USA).
4.4. Measurement of Alveolar Bone Resorption (ABR)
After euthanasia, the mandibles and maxilla were dissected and placed in a beaker, autoclaved to remove the soft flesh over the jawbone. Two-dimensional alveolar bone imaging was performed using a stereo dissecting microscope (Stereo Discovery V8, Carl Zeiss Microimaging, Inc., Thornwood, NY, USA). The pattern of the horizontal ABR area was measured by histomorphometry, as described previously [39,131]. Two examiners were blinded to measure ABR, and the data acquired were used for quantitative analysis.
Total RNA isolation and quality assessment
Total RNA was isolated from left mandibles using the mirVanaTM miRNA Isolation Kit (Ambion, Austin, TX, USA). The final RNA yield, quality assessments, and purity were determined using the standard method [36,37,39]. RNA with an OD 260/230 ratio of >2 and OD 260/280 ratio of >2 was quantified using an Epoch Microplate Spectrophotometer (BioTek, USA, Winooski, VT, USA) and taken for NanoString analysis.
4.5. miRNA-Transcriptome Expression Profiling
Left mandibles of the mice were selected for miRNA-transcriptome analysis using NanoString nCounter® Mouse miRNA Assay kit v1.5, a high-throughput nCounter® miRNA Expression Panels (NanoString Technologies, Seattle, WA, USA) which can evaluate 577 mouse specific miRNAs (36-39, 42). Total RNA extracted from each mouse mandible was hybridized with 100 ng of miRNA for 18 h at 65°C. The hybridized probes were purified and counted using the nCounter Prep station and digital analyzer after 24 h. The nCounter results were evaluated using nSolverTM 4.0 software as described (36-39, 42). Data Availability Statement: The data that support the findings of this study are openly available at https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSM7915885 (Accessed on 4th December 2023).
4.6. Bioinformatics Analysis
The normalized miRNA data was analyzed using the HyperScale architecture developed by ROSALIND®, Inc. (San Diego, CA) (https://rosalind.bio/ (Accessed on 16th September 2023) [132]. Fold changes in the miRNA were calculated following the standard formula [38] and limma R library [133]. The DE miRNAs were validated for miRNA-target gene interactions using the MiRTarBase database [134]. Venn diagram for the DE miRNAs was drawn using Venny 2.1 [39].
4.7. Kyoto Encyclopedia of Genes and Genomes (KEGG)
4.8. Multiple Machine Learning (ML) Model Analysis
The ML models used in our study were created and executed using version 3.11.4 of the Python programming language. We used XGBoost version 1.7.6 and the Scikit-learn version 1.3.0 implementations of LR, SVC, MLP, and RFC. A full list of the hyperparameters used for each ML model is available in Appendix S1 (Supplementary). SHAP version 0.42.1 was used to obtain feature importance results [42]. The code for executing the ML models on the NanoString copy data is available on GitHub at (https://github.com/uflcod/miRNA-periodontal-disease (accessed on February, 1st 2024) in the ‘notebooks’ directory. The notebooks for analyzing the F. nucleatum NanoString data begin with the prefix ‘Fn_’ (e.g., Fn_randomforest_miRNA.ipynb, Fn_xgboost_miRNA.ipynb).
4.9. Statistical Analysis
All the data in the graphs were presented as mean ± SEM. Data for the alveolar bone resorption were analyzed using ordinary two-way ANOVA, with Tukey’s multiple comparison test with a single pooled variance in Prism 9.4.1 (GraphPad Software, San Diego, CA, USA) [39,42,137]. p-value <0.05 was considered statistically significant. The differentially expressed miRNAs with FC of ±1 were considered significant. The Identification of significant DE-miRNA was done based on two-tailed t-tests. The Welch–Satterthwaite equation was used to calculate the distribution of the t-statistics. The volcano plot was drawn using GraphPad Software [39].
5. Conclusion
The studies carried out in the rodent models are the first in vivo F. nucleatum intraoral infection-induced periodontitis with emphasis on global miRNA profiling. Elevated miR-361 (1.3 FC) expression during 8 weeks of infection and 16 weeks of infection may play an important role in deciphering the information related to multiple malignancies. miR-127 and miR-26a could play a role in the initial immune response against F. nucleatum infection. miR-361 and miR-99b unique in both 8- and 16 weeks of Fn infection also expressed in the S. gordonii monoinfection. It is interesting to note that miR-126-5p has been shown as a potential biomarker in patients with periodontitis and coronary artery disease expressed uniquely in T. denticola monoinfection and the present Fn study. KEGG pathway analysis revealed the important roles of miR-361-5p, miR-218-5p, and miR-345-5p in regulating the genes in the bacterial invasion of epithelial cells pathway are also diagnostic markers for Fn induced periodontitis. The miRNA data presented provide clear evidence for the multi-pathogenic role of F. nucleatum in PD, several systemic comorbidities and nine oncogenic marker miRNAs (oncomiRs) in the CRC tumors.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
C.A. performed mouse experiments and analyzed the data. K.M.V. and S.J. did the molecular analysis of distal organ dissemination. S.J. performed bioinformatic and statistical analyses, initial drafting, and submission of data at NCBI. W.D.D. performed multiple ML models; L.K. was responsible for the conception, experimental design, analysis, initial drafting, editing, supervision, project administration, interpretation, and funding acquisition. L.K. and S.J. were involved in the final revision and editing of the manuscript. L.K. and E.K.L.C. are the Principal Investigators of the NIH (NIDCR) study. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the NIH National Institute of Dental and Craniofacial Research (NIDCR) (R01 DE028536) to L. Kesavalu and E.K.L. Chan. The funders had no role in study design, data collection, analysis, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
All animal procedures were approved by the University of Florida Institutional Animal Care and Use Committee (IACUC) under protocol number 202200000223.Informed Consent Statement
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are openly available at https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSM7915885 (Accessed on 4th December 2023).
Acknowledgments
The authors acknowledge Dr. Andreas Gonzalez, M.D for editing the manuscript and Dr. Yiping Han, Ph.D for previewing introduction section.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Krieger, M.; AbdelRahman, Y.M.; Choi, D.; Palmer, E.A.; Yoo, A.; McGuire, S.; Kreth, J.; Merritt, J. Stratification of Fusobacterium nucleatum by local health status in the oral cavity defines its subspecies disease association. Cell Host Microbe 2024, 32, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Bolstad, A.I.; Jensen, H.B.; Bakken, V. Taxonomy, biology, and periodontal aspects of Fusobacterium nucleatum. Clin Microbiol Rev 1996, 9, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J Clin Periodontol 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium nucleatum: a commensal-turned pathogen. Curr Opin Microbiol 2015, 23, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; London, J. Adhere today, here tomorrow: oral bacterial adherence. J Bacteriol 1993, 175, 3247–3252. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E. Oral microbial communities: biofilms, interactions, and genetic systems. Annu Rev Microbiol 2000, 54, 413–437. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Feuille, F.; Kesavalu, L.; Holt, S.C. Host modulation of tissue destruction caused by periodontopathogens: effects on a mixed microbial infection composed of Porphyromonas gingivalis and Fusobacterium nucleatum. Microb Pathog 1997, 23, 23–32. [Google Scholar] [CrossRef]
- Settem, R.P.; El-Hassan, A.T.; Honma, K.; Stafford, G.P.; Sharma, A. Fusobacterium nucleatum and Tannerella forsythia induce synergistic alveolar bone loss in a mouse periodontitis model. Infect Immun 2012, 80, 2436–2443. [Google Scholar] [CrossRef]
- Velsko, I.M.; Chukkapalli, S.S.; Rivera-Kweh, M.F.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Fusobacterium nucleatum Alters Atherosclerosis Risk Factors and Enhances Inflammatory Markers with an Atheroprotective Immune Response in ApoE(null) Mice. PLoS One 2015, 10, e0129795. [Google Scholar] [CrossRef]
- Han, Y.W.; Ikegami, A.; Rajanna, C.; Kawsar, H.I.; Zhou, Y.; Li, M.; Sojar, H.T.; Genco, R.J.; Kuramitsu, H.K.; Deng, C.X. Identification and characterization of a novel adhesin unique to oral fusobacteria. J Bacteriol 2005, 187, 5330–5340. [Google Scholar] [CrossRef]
- Xu, M.; Yamada, M.; Li, M.; Liu, H.; Chen, S.G.; Han, Y.W. FadA from Fusobacterium nucleatum utilizes both secreted and nonsecreted forms for functional oligomerization for attachment and invasion of host cells. J Biol Chem 2007, 282, 25000–25009. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/beta-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/beta-catenin modulator Annexin A1. EMBO Rep 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Gao, Q.; Mehrazarin, S.; Tangwanichgapong, K.; Wang, Y.; Huang, Y.; Pan, Y.; Robinson, S.; Liu, Z.; Zangiabadi, A.; et al. Fusobacterium nucleatum secretes amyloid-like FadA to enhance pathogenicity. EMBO Rep 2021, 22, e52891. [Google Scholar] [CrossRef]
- Baik, J.E.; Li, L.; Shah, M.A.; Freedberg, D.E.; Jin, Z.; Wang, T.C.; Han, Y.W. Circulating IgA Antibodies Against Fusobacterium nucleatum Amyloid Adhesin FadA are a Potential Biomarker for Colorectal Neoplasia. Cancer Res Commun 2022, 2, 1497–1503. [Google Scholar] [CrossRef]
- Han, Y.W.; Shi, W.; Huang, G.T.; Kinder Haake, S.; Park, N.H.; Kuramitsu, H.; Genco, R.J. Interactions between periodontal bacteria and human oral epithelial cells: Fusobacterium nucleatum adheres to and invades epithelial cells. Infection and immunity 2000, 68, 3140–3146. [Google Scholar] [CrossRef]
- Bui, F.Q.; Johnson, L.; Roberts, J.; Hung, S.C.; Lee, J.; Atanasova, K.R.; Huang, P.R.; Yilmaz, O.; Ojcius, D.M. Fusobacterium nucleatum infection of gingival epithelial cells leads to NLRP3 inflammasome-dependent secretion of IL-1beta and the danger signals ASC and HMGB1. Cell Microbiol 2016, 18, 970–981. [Google Scholar] [CrossRef]
- Ghosh, A.; Jaaback, K.; Boulton, A.; Wong-Brown, M.; Raymond, S.; Dutta, P.; Bowden, N.A.; Ghosh, A. Fusobacterium nucleatum: An Overview of Evidence, Demi-Decadal Trends, and Its Role in Adverse Pregnancy Outcomes and Various Gynecological Diseases, including Cancers. Cells 2024, 13. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, J.; Zhang, H.; Li, Q.; Qiu, H.; Hong, K.; Wang, W.; Xiao, Y.; Yu, B. Fusobacterium nucleatum promotes proliferation in oesophageal squamous cell carcinoma via AHR/CYP1A1 signalling. FEBS J 2023, 290, 837–854. [Google Scholar] [CrossRef]
- Lehr, K.; Nikitina, D.; Vilchez-Vargas, R.; Steponaitiene, R.; Thon, C.; Skieceviciene, J.; Schanze, D.; Zenker, M.; Malfertheiner, P.; Kupcinskas, J.; et al. Microbial composition of tumorous and adjacent gastric tissue is associated with prognosis of gastric cancer. Sci Rep 2023, 13, 4640. [Google Scholar] [CrossRef]
- Hayashi, M.; Ikenaga, N.; Nakata, K.; Luo, H.; Zhong, P.; Date, S.; Oyama, K.; Higashijima, N.; Kubo, A.; Iwamoto, C.; et al. Intratumor Fusobacterium nucleatum promotes the progression of pancreatic cancer via the CXCL1-CXCR2 axis. Cancer Sci 2023, 114, 3666–3678. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Cheng, Z.; Yin, Z.; Xu, J.; Wu, F.; Jin, Y.; Yang, G. Airway Fusobacterium is Associated with Poor Response to Immunotherapy in Lung Cancer. Onco Targets Ther 2022, 15, 201–213. [Google Scholar] [CrossRef] [PubMed]
- McIlvanna, E.; Linden, G.J.; Craig, S.G.; Lundy, F.T.; James, J.A. Fusobacterium nucleatum and oral cancer: a critical review. BMC Cancer 2021, 21, 1212. [Google Scholar] [CrossRef]
- Baima, G.; Ribaldone, D.G.; Romano, F.; Aimetti, M.; Romandini, M. The Gum-Gut Axis: Periodontitis and the Risk of Gastrointestinal Cancers. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Xie, Y.F.; Shu, R.; Jiang, S.Y.; Liu, D.L.; Zhang, X.L. Comparison of microRNA profiles of human periodontal diseased and healthy gingival tissues. Int J Oral Sci 2011, 3, 125–134. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Faulkner, J.L.; Sullivan, J.C. Circulating cell-free micro-RNA as biomarkers: from myocardial infarction to hypertension. Clin Sci (Lond) 2022, 136, 1341–1346. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Li, X.; Zhu, M.; Zhao, G.; Zhou, A.; Min, L.; Liu, S.; Zhang, N.; Zhu, S.; Guo, Q.; Zhang, S.; et al. MiR-1298-5p level downregulation induced by Helicobacter pylori infection inhibits autophagy and promotes gastric cancer development by targeting MAP2K6. Cell Signal 2022, 93, 110286. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, J.; Ke, W.; Zhang, X.; Chen, W.; Yang, J.; Liao, Y.; Liang, F.; Mei, S.; Li, M.; et al. MicroRNA expression profiling of peripheral blood mononuclear cells associated with syphilis. BMC Infect Dis 2020, 20, 165. [Google Scholar] [CrossRef] [PubMed]
- Davuluri, K.S.; Chauhan, D.S. microRNAs associated with the pathogenesis and their role in regulating various signaling pathways during. Front Cell Infect Microbiol 2022, 12, 1009901. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Cruz, A.R.; Rodrigues Lopes, I.; Maudet, C.; Sunkavalli, U.; Silva, R.J.; Sharan, M.; Lisowski, C.; Zaldívar-López, S.; Garrido, J.J.; et al. Functional screenings reveal different requirements for host microRNAs in Salmonella and Shigella infection. Nat Microbiol 2020, 5, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, B.R.; Zhang, M.; Sonntag, W.E.; Drevets, D.A. Neuroinvasive Listeria monocytogenes infection triggers accumulation of brain CD8. J Neuroinflammation 2020, 17, 259. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Tamizu, E.; Uno, S.; Uwamino, Y.; Fujiwara, H.; Nishio, K.; Nakano, Y.; Shiono, H.; Namkoong, H.; Hoshino, Y.; et al. hsa-miR-346 is a potential serum biomarker of Mycobacterium avium complex pulmonary disease activity. J Infect Chemother 2017, 23, 703–708. [Google Scholar] [CrossRef]
- Aravindraja, C.; Vekariya, K.M.; Botello-Escalante, R.; Rahaman, S.O.; Chan, E.K.L.; Kesavalu, L. Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Aravindraja, C.; Jeepipalli, S.; Vekariya, K.M.; Botello-Escalante, R.; Chan, E.K.L.; Kesavalu, L. Oral Spirochete Treponema denticola Intraoral Infection Reveals Unique miR-133a, miR-486, miR-126-3p, miR-126-5p miRNA Expression Kinetics during Periodontitis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Aravindraja, C.; Jeepipalli, S.; Duncan, W.; Vekariya, K.M.; Bahadekar, S.; Chan, E.K.L.; Kesavalu, L. Unique miRomics Expression Profiles in Tannerella forsythia-Infected Mandibles during Periodontitis Using Machine Learning. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Aravindraja, C.; Kashef, M.R.; Vekariya, K.M.; Ghanta, R.K.; Karanth, S.; Chan, E.K.L.; Kesavalu, L. Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Guzeldemir-Akcakanat, E.; Sunnetci-Akkoyunlu, D.; Balta-Uysal, V.M.; Ozer, T.; Isik, E.B.; Cine, N. Differentially expressed miRNAs associated with generalized aggressive periodontitis. Clin Oral Investig 2023, 28, 7. [Google Scholar] [CrossRef]
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE-/- mice during experimental periodontal disease. Infection and immunity 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Aravindraja, C.; Jeepipalli, S.; Duncan, W.D.; Vekariya, K.M.; Rahaman, S.O.; Chan, E.K.L.; Kesavalu, L. Streptococcus gordonii Supragingival Bacterium Oral Infection-Induced Periodontitis and Robust miRNA Expression Kinetics. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Usui, M.; Onizuka, S.; Sato, T.; Kokabu, S.; Ariyoshi, W.; Nakashima, K. Mechanism of alveolar bone destruction in periodontitis - Periodontal bacteria and inflammation. Jpn Dent Sci Rev 2021, 57, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Dong, P.; Xiong, Y.; Yue, J.; Ihira, K.; Konno, Y.; Kobayashi, N.; Todo, Y.; Watari, H. MicroRNA-361: A Multifaceted Player Regulating Tumor Aggressiveness and Tumor Microenvironment Formation. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chang, G.; Cao, L.; Ding, G. Dysregulation of serum miR-361-5p serves as a biomarker to predict disease onset and short-term prognosis in acute coronary syndrome patients. BMC Cardiovasc Disord 2021, 21, 74. [Google Scholar] [CrossRef]
- Uttamani, J.R.; Naqvi, A.R.; Estepa, A.M.V.; Kulkarni, V.; Brambila, M.F.; Martinez, G.; Chapa, G.; Wu, C.D.; Li, W.; Rivas-Tumanyan, S.; et al. Downregulation of miRNA-26 in chronic periodontitis interferes with innate immune responses and cell migration by targeting phospholipase C beta 1. J Clin Periodontol 2023, 50, 102–113. [Google Scholar] [CrossRef]
- Mildeberger, L.; Bueto, J.; Wilmes, V.; Scheiper-Welling, S.; Niess, C.; Gradhand, E.; Verhoff, M.A.; Kauferstein, S. Suitable biomarkers for post-mortem differentiation of cardiac death causes: Quantitative analysis of miR-1, miR-133a and miR-26a in heart tissue and whole blood. Forensic Sci Int Genet 2023, 65, 102867. [Google Scholar] [CrossRef]
- Nik Mohamed Kamal, N.N.S.; Awang, R.A.R.; Mohamad, S.; Shahidan, W.N.S. Plasma- and Saliva Exosome Profile Reveals a Distinct MicroRNA Signature in Chronic Periodontitis. Front Physiol 2020, 11, 587381. [Google Scholar] [CrossRef]
- Du, A.; Zhao, S.; Wan, L.; Liu, T.; Peng, Z.; Zhou, Z.; Liao, Z.; Fang, H. MicroRNA expression profile of human periodontal ligament cells under the influence of Porphyromonas gingivalis LPS. J Cell Mol Med 2016, 20, 1329–1338. [Google Scholar] [CrossRef]
- Pan, J.; Alexan, B.; Dennis, D.; Bettina, C.; Christoph, L.I.M.; Tang, Y. microRNA-193-3p attenuates myocardial injury of mice with sepsis via STAT3/HMGB1 axis. J Transl Med 2021, 19, 386. [Google Scholar] [CrossRef]
- Girolimetti, G.; Pelisenco, I.A.; Eusebi, L.H.; Ricci, C.; Cavina, B.; Kurelac, I.; Verri, T.; Calcagnile, M.; Alifano, P.; Salvi, A.; et al. Dysregulation of a Subset of Circulating and Vesicle-Associated miRNA in Pancreatic Cancer. Noncoding RNA 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Wanram, S.; Klaewkla, N.; Pinyosri, P. Downregulation of Serum miR-133b and miR-206 Associate with Clinical Outcomes of Progression as Monitoring Biomarkers for Metastasis Colorectal Cancer Patients. Microrna 2024, 13, 56–62. [Google Scholar] [CrossRef]
- Wang, X.; Dai, L.; Liu, J.; Ge, J. Expression of miR-128-3p, miR-193a-3p and miR-193a-5p in endometrial cancer tissues and their relationship with clinicopathological parameters. Cell Mol Biol (Noisy-le-grand) 2022, 68, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Liu, Z.; Sun, G. Diagnostic value of miR-193a-3p in Alzheimer's disease and miR-193a-3p attenuates amyloid-beta induced neurotoxicity by targeting PTEN. Exp Gerontol 2020, 130, 110814. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Chen, X.; Chen, Q.; Shi, L.; Liang, H.; Zhou, Z.; Liu, Q.; Pang, W.; Hou, D.; Wang, C.; et al. MicroRNA-193a-3p Reduces Intestinal Inflammation in Response to Microbiota via Down-regulation of Colonic PepT1. J Biol Chem 2015, 290, 16099–16115. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Lai, W.; Song, L.; Deng, J.; Li, C.; Jiang, S. MicroRNA-126 regulates macrophage polarization to prevent the resorption of alveolar bone in diabetic periodontitis. Arch Oral Biol 2023, 150, 105686. [Google Scholar] [CrossRef]
- Buragaite-Staponkiene, B.; Rovas, A.; Puriene, A.; Snipaitiene, K.; Punceviciene, E.; Rimkevicius, A.; Butrimiene, I.; Jarmalaite, S. Gingival Tissue MiRNA Expression Profiling and an Analysis of Periodontitis-Specific Circulating MiRNAs. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Lin, H.; Hu, S.; Li, Y.; Li, S.; Teng, D.; Yang, Y.; Liu, B.; Du, X. H3K27ac-activated LncRNA NUTM2A-AS1 Facilitates the Progression of Colorectal Cancer Cells via MicroRNA-126-5p/FAM3C Axis. Curr Cancer Drug Targets 2024. [Google Scholar] [CrossRef]
- Bi, X.; Lv, X.; Liu, D.; Guo, H.; Yao, G.; Wang, L.; Liang, X.; Yang, Y. METTL3-mediated maturation of miR-126-5p promotes ovarian cancer progression via PTEN-mediated PI3K/Akt/mTOR pathway. Cancer Gene Ther 2021, 28, 335–349. [Google Scholar] [CrossRef]
- Yalim, Z.; Onrat, S.T.; Dural, I.E.; Onrat, E. Could Aneurysm and Atherosclerosis-Associated MicroRNAs (miR 24-1-5p, miR 34a-5p, miR 126-5p, miR 143-5p, miR 145-5p) Also Be Associated with Coronary Artery Ectasia? Genet Test Mol Biomarkers 2023, 27, 290–298. [Google Scholar] [CrossRef]
- Ekedi, A.; Rozhkov, A.N.; Shchekochikhin, D.Y.; Novikova, N.A.; Kopylov, P.Y.; Bestavashvili, A.A.; Ivanova, T.V.; Zhelankin, A.V.; Generozov, E.V.; Konanov, D.N.; et al. Evaluation of microRNA Expression Features in Patients with Various Types of Arterial Damage: Thoracic Aortic Aneurysm and Coronary Atherosclerosis. J Pers Med 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zou, Y.; Zhang, H. MicroRNA-126-5p Facilitates Hypoxia-Induced Vascular Endothelial Cell Injury via HIPK2. Ann Clin Lab Sci 2022, 52, 918–926. [Google Scholar] [PubMed]
- Baraldo, N.; Buzzoni, L.; Pasti, L.; Cavazzini, A.; Marchetti, N.; Mancia, A. miRNAs as Biomolecular Markers for Food Safety, Quality, and Traceability in Poultry Meat-A Preliminary Study. Molecules 2024, 29. [Google Scholar] [CrossRef] [PubMed]
- Baru, O.; Raduly, L.; Bica, C.; Chiroi, P.; Budisan, L.; Mehterov, N.; Ciocan, C.; Pop, L.A.; Buduru, S.; Braicu, C.; et al. Identification of a miRNA Panel with a Potential Determinant Role in Patients Suffering from Periodontitis. Curr Issues Mol Biol 2023, 45, 2248–2265. [Google Scholar] [CrossRef] [PubMed]
- Kadkhoda, S.; Hussen, B.M.; Eslami, S.; Ghafouri-Fard, S. A review on the role of miRNA-324 in various diseases. Front Genet 2022, 13, 950162. [Google Scholar] [CrossRef]
- Ellis, K.L.; Cameron, V.A.; Troughton, R.W.; Frampton, C.M.; Ellmers, L.J.; Richards, A.M. Circulating microRNAs as candidate markers to distinguish heart failure in breathless patients. Eur J Heart Fail 2013, 15, 1138–1147. [Google Scholar] [CrossRef]
- Gu, H.; Wu, L.; Chen, H.; Huang, Z.; Xu, J.; Zhou, K.; Zhang, Y.; Chen, J.; Xia, J.; Yin, X. Identification of differentially expressed microRNAs in the bone marrow of osteoporosis patients. Am J Transl Res 2019, 11, 2940–2954. [Google Scholar]
- Naqvi, A.R.; Fordham, J.B.; Nares, S. miR-24, miR-30b, and miR-142-3p regulate phagocytosis in myeloid inflammatory cells. J Immunol 2015, 194, 1916–1927. [Google Scholar] [CrossRef]
- Wang, H.; Chen, C.; Ding, K.; Zhang, W.; Hou, J. MiR-24-3p as a prognostic indicator for multiple cancers: from a meta-analysis view. Biosci Rep 2020, 40. [Google Scholar] [CrossRef]
- Singh, Y.; Kaul, V.; Mehra, A.; Chatterjee, S.; Tousif, S.; Dwivedi, V.P.; Suar, M.; Van Kaer, L.; Bishai, W.R.; Das, G. Mycobacterium tuberculosis controls microRNA-99b (miR-99b) expression in infected murine dendritic cells to modulate host immunity. The Journal of biological chemistry 2013, 288, 5056–5061. [Google Scholar] [CrossRef]
- Holliday, H.; Yang, J.; Dodson, E.; Nikolic, I.; Kamili, A.; Wheatley, M.; Deng, N.; Alexandrou, S.; Davis, T.P.; Kavallaris, M.; et al. miR-99b-5p, miR-380-3p, and miR-485-3p are novel chemosensitizing miRNAs in high-risk neuroblastoma. Mol Ther 2024, 32, 2031–2033. [Google Scholar] [CrossRef] [PubMed]
- Holliday, H.; Yang, J.; Dodson, E.; Nikolic, I.; Kamili, A.; Wheatley, M.; Deng, N.; Alexandrou, S.; Davis, T.P.; Kavallaris, M.; et al. miR-99b-5p, miR-380-3p, and miR-485-3p are novel chemosensitizing miRNAs in high-risk neuroblastoma. Mol Ther 2022, 30, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Wang, B.D. Combination of miR-99b-5p and Enzalutamide or Abiraterone Synergizes the Suppression of EMT-Mediated Metastasis in Prostate Cancer. Cancers (Basel) 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Gujrati, H.; Wang, B.D. Tumor suppressive miR-99b-5p as an epigenomic regulator mediating mTOR/AR/SMARCD1 signaling axis in aggressive prostate cancer. Front Oncol 2023, 13, 1184186. [Google Scholar] [CrossRef]
- Ning, S.; Chen, Y.; Li, S.; Liu, M.; Liu, H.; Ye, M.; Wang, C.; Pan, J.; Wei, W.; Li, J.; et al. Exosomal miR-99b-5p Secreted from Mesenchymal Stem Cells Can Retard the Progression of Colorectal Cancer by Targeting FGFR3. Stem Cell Rev Rep 2023, 19, 2901–2917. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Z.; Yang, Y.; Luo, M.; Zhang, M.; Wang, X.; Liu, L.; Hou, N.; Guo, Q.; Song, T.; et al. Author Correction: MiR-99b-5p and miR-203a-3p Function as Tumor Suppressors by Targeting IGF-1R in Gastric Cancer. Sci Rep 2023, 13, 906. [Google Scholar] [CrossRef]
- Nguyen, T.B.; Do, D.N.; Nguyen, T.T.P.; Nguyen, T.L.; Nguyen-Thanh, T.; Nguyen, H.T. Immune-related biomarkers shared by inflammatory bowel disease and liver cancer. PLoS One 2022, 17, e0267358. [Google Scholar] [CrossRef]
- Xia, J.; Luo, M.; Dai, L.; Wang, L.; Wang, L.; Zhu, J. Serum exosomal microRNAs as predictive markers for EGFR mutations in non-small-cell lung cancer. J Clin Lab Anal 2021, 35, e23743. [Google Scholar] [CrossRef]
- Lee, N.H.; Lee, E.; Kim, Y.S.; Kim, W.K.; Lee, Y.K.; Kim, S.H. Differential expression of microRNAs in the saliva of patients with aggressive periodontitis: a pilot study of potential biomarkers for aggressive periodontitis. J Periodontal Implant Sci 2020, 50, 281–290. [Google Scholar] [CrossRef]
- Venugopal, P.; Koshy, T.; Lavu, V.; Ranga Rao, S.; Ramasamy, S.; Hariharan, S.; Venkatesan, V. Differential expression of microRNAs let-7a, miR-125b, miR-100, and miR-21 and interaction with NF-kB pathway genes in periodontitis pathogenesis. J Cell Physiol 2018, 233, 5877–5884. [Google Scholar] [CrossRef]
- Polikepahad, S.; Knight, J.M.; Naghavi, A.O.; Oplt, T.; Creighton, C.J.; Shaw, C.; Benham, A.L.; Kim, J.; Soibam, B.; Harris, R.A.; et al. Proinflammatory role for let-7 microRNAS in experimental asthma. The Journal of biological chemistry 2010, 285, 30139–30149. [Google Scholar] [CrossRef] [PubMed]
- Rijavec, M.; Korosec, P.; Zavbi, M.; Kern, I.; Malovrh, M.M. Let-7a is differentially expressed in bronchial biopsies of patients with severe asthma. Sci Rep 2014, 4, 6103. [Google Scholar] [CrossRef] [PubMed]
- Ogata, Y.; Matsui, S.; Kato, A.; Zhou, L.; Nakayama, Y.; Takai, H. MicroRNA expression in inflamed and noninflamed gingival tissues from Japanese patients. J Oral Sci 2014, 56, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, Y.; Wang, C.; Hu, W.; Zou, S.; Ren, H.; Zuo, Y.; Qu, L. MiR-127-3p enhances macrophagic proliferation via disturbing fatty acid profiles and oxidative phosphorylation in atherosclerosis. J Mol Cell Cardiol 2024, 193, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Wang, X.; Wang, S.; Xia, Y.; Chen, H.; Yin, L.; Hu, K. Screening for Regulatory Network of miRNA-Inflammation, Oxidative Stress and Prognosis-Related mRNA in Acute Myocardial Infarction: An in silico and Validation Study. Int J Gen Med 2022, 15, 1715–1731. [Google Scholar] [CrossRef]
- Umeh-Garcia, M.; Simion, C.; Ho, P.Y.; Batra, N.; Berg, A.L.; Carraway, K.L.; Yu, A.; Sweeney, C. A Novel Bioengineered miR-127 Prodrug Suppresses the Growth and Metastatic Potential of Triple-Negative Breast Cancer Cells. Cancer Res 2020, 80, 418–429. [Google Scholar] [CrossRef]
- Scholtz, B.; Horvath, J.; Tar, I.; Kiss, C.; Marton, I.J. Salivary miR-31-5p, miR-345-3p, and miR-424-3p Are Reliable Biomarkers in Patients with Oral Squamous Cell Carcinoma. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, Y.; Han, M.; Jiang, L.; Liang, D.; Li, S.; Xu, Z.; Wang, L.; Li, N. MicroRNA-345-5p acts as an anti-inflammatory regulator in experimental allergic rhinitis via the TLR4/NF-kappaB pathway. Int Immunopharmacol 2020, 86, 106522. [Google Scholar] [CrossRef]
- Li, Y.; Zhuang, J. miR-345-3p serves a protective role during gestational diabetes mellitus by targeting BAK1. Exp Ther Med 2021, 21, 2. [Google Scholar] [CrossRef]
- Spear, R.; Boytard, L.; Blervaque, R.; Chwastyniak, M.; Hot, D.; Vanhoutte, J.; Lamblin, N.; Amouyel, P.; Pinet, F. Let-7f: A New Potential Circulating Biomarker Identified by miRNA Profiling of Cells Isolated from Human Abdominal Aortic Aneurysm. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Geng, L.; Tang, X.; Wang, S.; Sun, Y.; Wang, D.; Tsao, B.P.; Feng, X.; Sun, L. Reduced Let-7f in Bone Marrow-Derived Mesenchymal Stem Cells Triggers Treg/Th17 Imbalance in Patients With Systemic Lupus Erythematosus. Front Immunol 2020, 11, 233. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Carter, D.E.; Leask, A. miR-218 regulates focal adhesion kinase-dependent TGFbeta signaling in fibroblasts. Mol Biol Cell 2014, 25, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Na, H.S.; Jeong, S.Y.; Jeong, S.H.; Park, H.R.; Chung, J. Comparison of inflammatory microRNA expression in healthy and periodontitis tissues. Biocell 2011, 35, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, Z.; Chen, Z.; Wei, Y.; Liang, H.; Zhang, X.; Gao, Z.; Zhu, H. MicroRNA-218-5p regulates inflammation response via targeting TLR4 in atherosclerosis. BMC Cardiovasc Disord 2023, 23, 122. [Google Scholar] [CrossRef] [PubMed]
- Conickx, G.; Mestdagh, P.; Avila Cobos, F.; Verhamme, F.M.; Maes, T.; Vanaudenaerde, B.M.; Seys, L.J.; Lahousse, L.; Kim, R.Y.; Hsu, A.C.; et al. MicroRNA Profiling Reveals a Role for MicroRNA-218-5p in the Pathogenesis of Chronic Obstructive Pulmonary Disease. Am J Respir Crit Care Med 2017, 195, 43–56. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Li, Y.; Peng, H.; Liu, J.; Zhang, J.; Xiao, X. The Role of CREBBP/EP300 and Its Therapeutic Implications in Hematological Malignancies. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Perri, R.; Nares, S.; Zhang, S.; Barros, S.P.; Offenbacher, S. MicroRNA modulation in obesity and periodontitis. J Dent Res 2012, 91, 33–38. [Google Scholar] [CrossRef]
- Jiang, F.; Zhou, Y.; Zhang, R.; Wen, Y. miR-205 and HMGB1 expressions in chronic periodontitis patients and their associations with the inflammatory factors. Am J Transl Res 2021, 13, 9224–9232. [Google Scholar]
- Stoecklin-Wasmer, C.; Guarnieri, P.; Celenti, R.; Demmer, R.T.; Kebschull, M.; Papapanou, P.N. MicroRNAs and their target genes in gingival tissues. J Dent Res 2012, 91, 934–940. [Google Scholar] [CrossRef]
- Hara, E.S.; Ono, M.; Eguchi, T.; Kubota, S.; Pham, H.T.; Sonoyama, W.; Tajima, S.; Takigawa, M.; Calderwood, S.K.; Kuboki, T. miRNA-720 controls stem cell phenotype, proliferation and differentiation of human dental pulp cells. PLoS One 2013, 8, e83545. [Google Scholar] [CrossRef]
- Yang, S.; Gao, J.; Chen, M.; Sun, Y.; Qiao, X.; Mao, H.; Guo, L.; Yu, Y.; Yang, D. Let-7a promotes periodontal bone regeneration of bone marrow mesenchymal stem cell aggregates via the Fas/FasL-autophagy pathway. J Cell Mol Med 2023, 27, 4056–4068. [Google Scholar] [CrossRef]
- Peng, J.; Lin, Y.; Sheng, X.; Yuan, C.; Wang, Y.; Yin, W.; Zhou, L.; Lu, J. Serum miRNA-1 may serve as a promising noninvasive biomarker for predicting treatment response in breast cancer patients receiving neoadjuvant chemotherapy. BMC Cancer 2024, 24, 789. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, F.; Seyed Mohammadzad, M.H. Up-Regulation of Cell-Free MicroRNA-1 and MicroRNA-221-3p Levels in Patients with Myocardial Infarction Undergoing Coronary Angiography. Adv Pharm Bull 2021, 11, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wu, X.; Zhang, W.; Chu, F.; Zhang, Q.; Gao, M.; Xu, Y.; Wu, Y. Effect of psoralen on the regulation of osteogenic differentiation induced by periodontal stem cell-derived exosomes. Hum Cell 2023, 36, 1389–1402. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, Z.; Yao, L.; Xue, B.; Xi, H.; Wang, X.; Sun, S. Exploration of Shared Gene Signatures and Molecular Mechanisms Between Periodontitis and Nonalcoholic Fatty Liver Disease. Front Genet 2022, 13, 939751. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Zhang, Y.H.; Ding, Y.Q.; Bi, X.Y.; Yuan, J.; Zhou, H.; Wang, P.X.; Zhang, L.L.; Ye, J.T. MicroRNA-99b-3p promotes angiotensin II-induced cardiac fibrosis in mice by targeting GSK-3beta. Acta Pharmacol Sin 2021, 42, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Cheon, E.J.; Lee, M.H.; Kim, H.A. MicroRNA-127-5p regulates matrix metalloproteinase 13 expression and interleukin-1beta-induced catabolic effects in human chondrocytes. Arthritis Rheum 2013, 65, 3141–3152. [Google Scholar] [CrossRef]
- Ma, L.; Cao, Y.; Zhang, L.; Li, K.; Yan, L.; Pan, Y.; Zhu, J. Celastrol mitigates high glucose-induced inflammation and apoptosis in rat H9c2 cardiomyocytes via miR-345-5p/growth arrest-specific 6. J Gene Med 2020, 22, e3201. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, X.; Wang, J.; Zhang, X.; Xu, Y.; Liu, Y.; Li, X.; Yang, G. miR-324-5p promotes adipocyte differentiation and lipid droplet accumulation by targeting Krueppel-like factor 3 (KLF3). J Cell Physiol 2020, 235, 7484–7495. [Google Scholar] [CrossRef]
- Liu, X.; He, Y.; Feng, Z.; Sheng, J.; Dong, A.; Zhang, M.; Cao, L. miR-345-5p regulates adipogenesis via targeting VEGF-B. Aging (Albany NY) 2020, 12, 17114–17121. [Google Scholar] [CrossRef]
- Mou, T.; Xie, F.; Zhong, P.; Hua, H.; Lai, L.; Yang, Q.; Wang, J. MiR-345-5p functions as a tumor suppressor in pancreatic cancer by directly targeting CCL8. Biomed Pharmacother 2019, 111, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Y.; Gui, S.Y.; Zhang, P.; Wang, M. Upregulation of miR-345-5p suppresses cell growth of lung adenocarcinoma by regulating ras homolog family member A (RhoA) and Rho/Rho associated protein kinase (Rho/ROCK) pathway. Chin Med J (Engl) 2021, 134, 2619–2628. [Google Scholar] [CrossRef]
- Jakob, M.; Mattes, L.M.; Kuffer, S.; Unger, K.; Hess, J.; Bertlich, M.; Haubner, F.; Ihler, F.; Canis, M.; Weiss, B.G.; et al. MicroRNA expression patterns in oral squamous cell carcinoma: hsa-mir-99b-3p and hsa-mir-100-5p as novel prognostic markers for oral cancer. Head Neck 2019, 41, 3499–3515. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, Y.; Martinez-Moreno, M.; Alonso, L.; Sanchez-Vencells, A.; Arranz, A.; Daga-Millan, R.; Sevilla-Movilla, S.; Valeri, A.; Martinez-Lopez, J.; Teixido, J. Regulation of miRNA expression by alpha4beta1 integrin-dependent multiple myeloma cell adhesion. EJHaem 2023, 4, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Sui, C.; Zhang, L.; Hu, Y. MicroRNA-let-7a inhibition inhibits LPS-induced inflammatory injury of chondrocytes by targeting IL6R. Mol Med Rep 2019, 20, 2633–2640. [Google Scholar] [CrossRef] [PubMed]
- Alexandri, C.; Stamatopoulos, B.; Rothe, F.; Bareche, Y.; Devos, M.; Demeestere, I. MicroRNA profiling and identification of let-7a as a target to prevent chemotherapy-induced primordial follicles apoptosis in mouse ovaries. Sci Rep 2019, 9, 9636. [Google Scholar] [CrossRef]
- Lo, C.H.; Li, L.C.; Yang, S.F.; Tsai, C.F.; Chuang, Y.T.; Chu, H.J.; Ueng, K.C. MicroRNA Let-7a, -7e and -133a Attenuate Hypoxia-Induced Atrial Fibrosis via Targeting Collagen Expression and the JNK Pathway in HL1 Cardiomyocytes. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Liu, C.; Chen, Z.; Wang, J.; Hu, H. Overexpression of X chromosome-linked inhibitor of apoptosis by inhibiting microRNA-24 protects periodontal ligament cells against hydrogen peroxide-induced cell apoptosis. Cell Mol Biol (Noisy-le-grand) 2016, 62, 6–13. [Google Scholar]
- Liu, X.; Mao, Y.; Kang, Y.; He, L.; Zhu, B.; Zhang, W.; Lu, Y.; Wu, Q.; Xu, D.; Shi, L. MicroRNA-127 Promotes Anti-microbial Host Defense through Restricting A20-Mediated De-ubiquitination of STAT3. iScience 2020, 23, 100763. [Google Scholar] [CrossRef]
- Jin, T.; Lu, Y.; He, Q.X.; Wang, H.; Li, B.F.; Zhu, L.Y.; Xu, Q.Y. The Role of MicroRNA, miR-24, and Its Target CHI3L1 in Osteomyelitis Caused by Staphylococcus aureus. J Cell Biochem 2015, 116, 2804–2813. [Google Scholar] [CrossRef]
- Zepeda-Rivera, M.; Minot, S.S.; Bouzek, H.; Wu, H.; Blanco-Miguez, A.; Manghi, P.; Jones, D.S.; LaCourse, K.D.; Wu, Y.; McMahon, E.F.; et al. A distinct Fusobacterium nucleatum clade dominates the colorectal cancer niche. Nature 2024, 628, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Ford, P.J.; Gemmell, E.; Hamlet, S.M.; Hasan, A.; Walker, P.J.; West, M.J.; Cullinan, M.P.; Seymour, G.J. Cross-reactivity of GroEL antibodies with human heat shock protein 60 and quantification of pathogens in atherosclerosis. Oral Microbiol Immunol 2005, 20, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Zheng, D.; Lucas, A.R.; Kesavalu, L. Polymicrobial Oral Infection with Four Periodontal Bacteria Orchestrates a Distinct Inflammatory Response and Atherosclerosis in ApoE null Mice. PLoS One 2015, 10, e0143291. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.W.; Redline, R.W.; Li, M.; Yin, L.; Hill, G.B.; McCormick, T.S. Fusobacterium nucleatum induces premature and term stillbirths in pregnant mice: implication of oral bacteria in preterm birth. Infect Immun 2004, 72, 2272–2279. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Dzink, J.L.; Sheenan, M.T.; Socransky, S.S. Proposal of three subspecies of Fusobacterium nucleatum Knorr 1922: Fusobacterium nucleatum subsp. nucleatum subsp. nov., comb. nov.; Fusobacterium nucleatum subsp. polymorphum subsp. nov., nom. rev., comb. nov.; and Fusobacterium nucleatum subsp. vincentii subsp. nov., nom. rev., comb. nov. Int J Syst Bacteriol 1990, 40, 74–78. [Google Scholar] [CrossRef]
- Kesavalu, L.; Sathishkumar, S.; Bakthavatchalu, V.; Matthews, C.; Dawson, D.; Steffen, M.; Ebersole, J.L. Rat model of polymicrobial infection, immunity, and alveolar bone resorption in periodontal disease. Infection and immunity 2007, 75, 1704–1712. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Ambadapadi, S.; Varkoly, K.; Jiron, J.; Aguirre, J.I.; Bhattacharyya, I.; Morel, L.M.; Lucas, A.R.; Kesavalu, L. Impaired innate immune signaling due to combined Toll-like receptor 2 and 4 deficiency affects both periodontitis and atherosclerosis in response to polybacterial infection. Pathog Dis 2018, 76. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Easwaran, M.; Rivera-Kweh, M.F.; Velsko, I.M.; Ambadapadi, S.; Dai, J.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Sequential colonization of periodontal pathogens in induction of periodontal disease and atherosclerosis in LDLRnull mice. Pathog Dis 2017, 75. [Google Scholar] [CrossRef]
- Velsko, I.M.; Chukkapalli, S.S.; Rivera-Kweh, M.F.; Zheng, D.; Aukhil, I.; Lucas, A.R.; Larjava, H.; Kesavalu, L. Periodontal pathogens invade gingiva and aortic adventitia and elicit inflammasome activation in alphavbeta6 integrin-deficient mice. Infect Immun 2015, 83, 4582–4593. [Google Scholar] [CrossRef]
- Aravindraja, C.; Sakthivel, R.; Liu, X.; Goodwin, M.; Veena, P.; Godovikova, V.; Fenno, J.C.; Levites, Y.; Golde, T.E.; Kesavalu, L. Intracerebral but Not Peripheral Infection of Live Porphyromonas gingivalis Exacerbates Alzheimer's Disease Like Amyloid Pathology in APP-TgCRND8 Mice. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Rutigliano, D.; Puglielli, A.; Fazio, V.M. Butein-instigated miR-186-5p-dependent modulation of TWIST1 affects resistance to cisplatin and bioenergetics of Malignant Pleural Mesothelioma cells. Cancer Drug Resist 2022, 5, 814–828. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Lin, Y.C.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: an informative resource for experimentally validated miRNA-target interactions. Nucleic Acids Res 2022, 50, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: deciphering microRNA function with experimental support. Nucleic Acids Res 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Velsko, I.M.; Rivera-Kweh, M.F.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Global TLR2 and 4 deficiency in mice impacts bone resorption, inflammatory markers and atherosclerosis to polymicrobial infection. Mol Oral Microbiol 2017, 32, 211–225. [Google Scholar] [CrossRef]
Figure 1.
Intraoral infection of F. nucleatum significantly induced ABR. (A). Schematic diagram of the experimental design depicting the monobacterial F. nucleatum infection (4 days per week on every alternate week), plaque sampling for PCR, and euthanasia. (B). Representative images showing horizontal ABR (mandible lingual view) of F. nucleatum-infected and sham-infected mice with the area of ABR outlined from the alveolar bone crest (ABC) to the cementoenamel junction (CEJ). (C). Morphometric analysis of the mandible and maxillary ABR in mice. A significant increase in ABR was observed in F. nucleatum-infected mice compared to sham-infected mice at both 8 weeks and 16 weeks infected mice (****, p < 0.0001; **, p < 0.01; *, p < 0.05) ordinary two-way ANOVA). Data points and error bars are mean ± SEM (n =10).
Figure 1.
Intraoral infection of F. nucleatum significantly induced ABR. (A). Schematic diagram of the experimental design depicting the monobacterial F. nucleatum infection (4 days per week on every alternate week), plaque sampling for PCR, and euthanasia. (B). Representative images showing horizontal ABR (mandible lingual view) of F. nucleatum-infected and sham-infected mice with the area of ABR outlined from the alveolar bone crest (ABC) to the cementoenamel junction (CEJ). (C). Morphometric analysis of the mandible and maxillary ABR in mice. A significant increase in ABR was observed in F. nucleatum-infected mice compared to sham-infected mice at both 8 weeks and 16 weeks infected mice (****, p < 0.0001; **, p < 0.01; *, p < 0.05) ordinary two-way ANOVA). Data points and error bars are mean ± SEM (n =10).
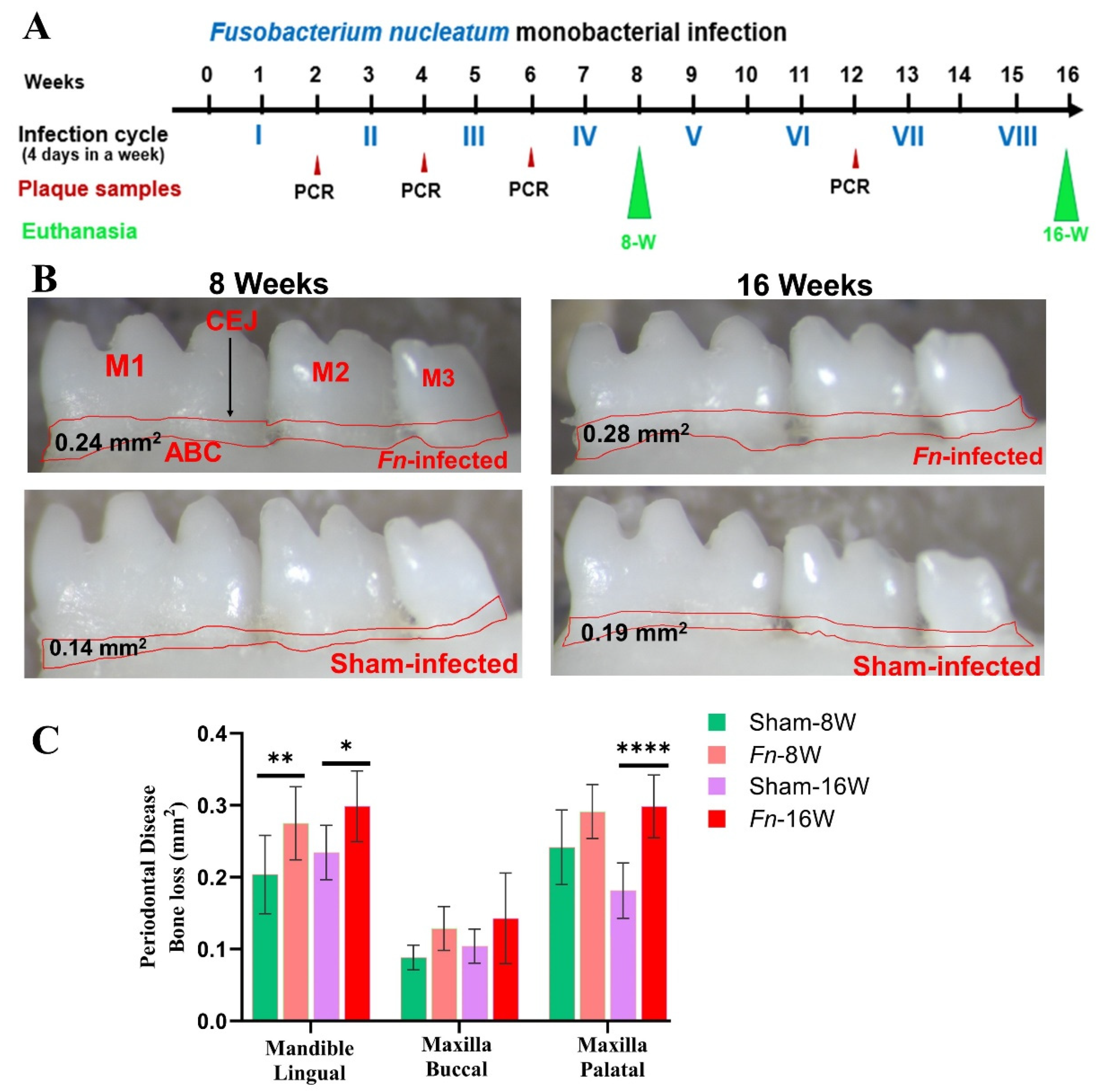
Figure 2.
DE miRNAs in F. nucleatum-infected mandibles (8- and 16 weeks). A. The volcano plot depicts the upregulated (green) and downregulated (red) miRNAs that showed a fold difference of ±1.1 with a p-value of < 0.05. The log2 fold change is on the x-axis, and the negative log of the p-value is on the y-axis. The black dots represent the miRNAs that do not pass the filter parameters. 7 significant upregulated miRNAs and 2 downregulated miRNAs were identified in 8 weeks of F. nucleatum-infected mice compared to 8 weeks sham-infected mice (n = 10). 7 significant upregulated miRs and 13 downregulated miRs were identified in 16 weeks of F. nucleatum-infected mice compared to 16 weeks of sham-infected mice (n = 10). B. Venn diagram analysis illustrates the distribution of DE miRNAs in 8 weeks and 16 weeks infections with F. nucleatum. C&D. Predicted functional pathway analysis of DE miRNAs from F. nucleatum-infected mandibles. Bubble Plot of KEGG analysis on predicted target genes of DE miRNAs in F. nucleatum-infected mice at 16 weeks of infection compared to sham-infected mice. The KEGG pathways are displayed on the y-axis, and the x-axis represents the false discovery rate (FDR) which means the probability of false positives in all tests. The size and color of the dots represent the number of predicted genes and corresponding p-value, respectively.
Figure 2.
DE miRNAs in F. nucleatum-infected mandibles (8- and 16 weeks). A. The volcano plot depicts the upregulated (green) and downregulated (red) miRNAs that showed a fold difference of ±1.1 with a p-value of < 0.05. The log2 fold change is on the x-axis, and the negative log of the p-value is on the y-axis. The black dots represent the miRNAs that do not pass the filter parameters. 7 significant upregulated miRNAs and 2 downregulated miRNAs were identified in 8 weeks of F. nucleatum-infected mice compared to 8 weeks sham-infected mice (n = 10). 7 significant upregulated miRs and 13 downregulated miRs were identified in 16 weeks of F. nucleatum-infected mice compared to 16 weeks of sham-infected mice (n = 10). B. Venn diagram analysis illustrates the distribution of DE miRNAs in 8 weeks and 16 weeks infections with F. nucleatum. C&D. Predicted functional pathway analysis of DE miRNAs from F. nucleatum-infected mandibles. Bubble Plot of KEGG analysis on predicted target genes of DE miRNAs in F. nucleatum-infected mice at 16 weeks of infection compared to sham-infected mice. The KEGG pathways are displayed on the y-axis, and the x-axis represents the false discovery rate (FDR) which means the probability of false positives in all tests. The size and color of the dots represent the number of predicted genes and corresponding p-value, respectively.
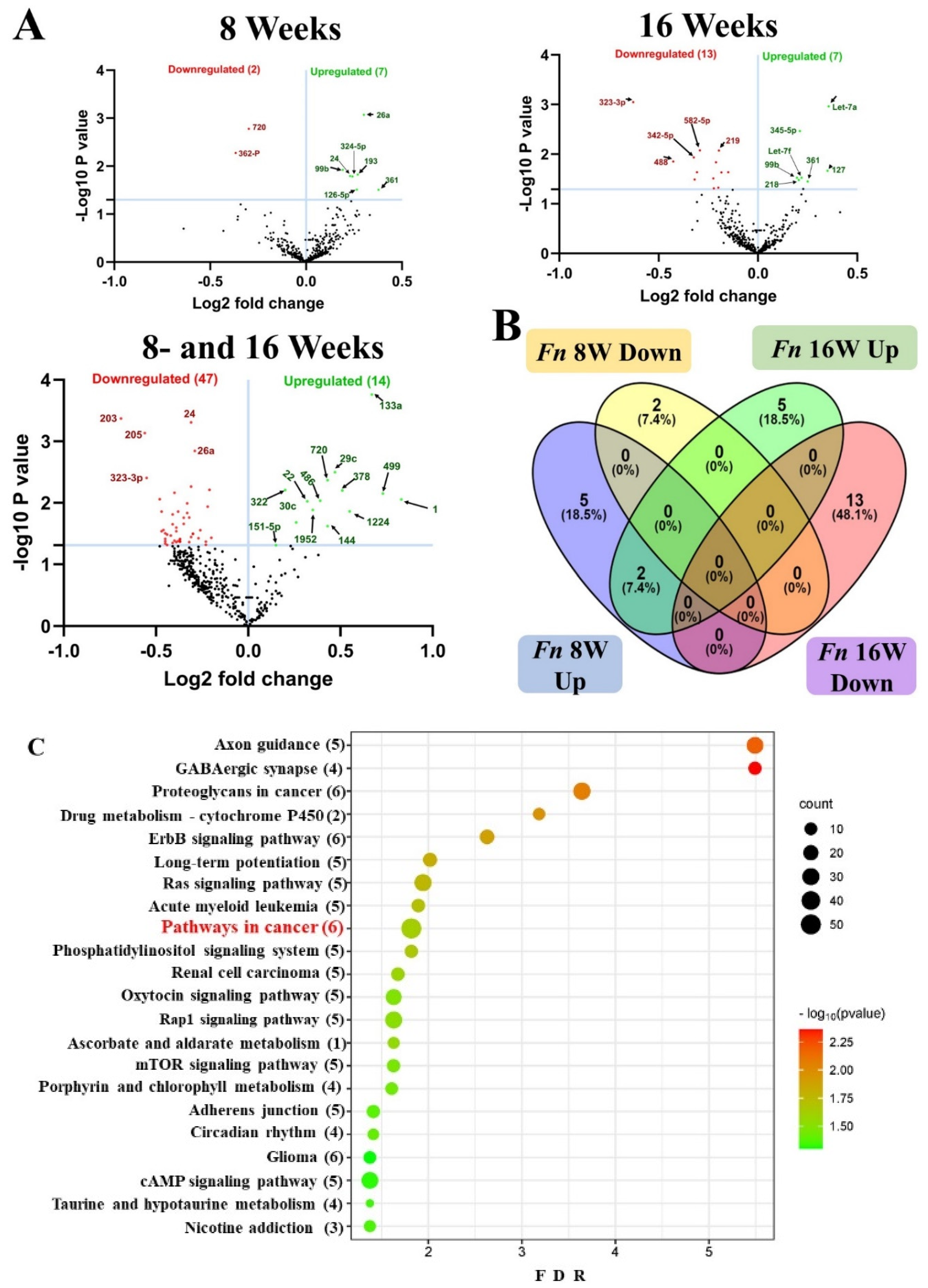
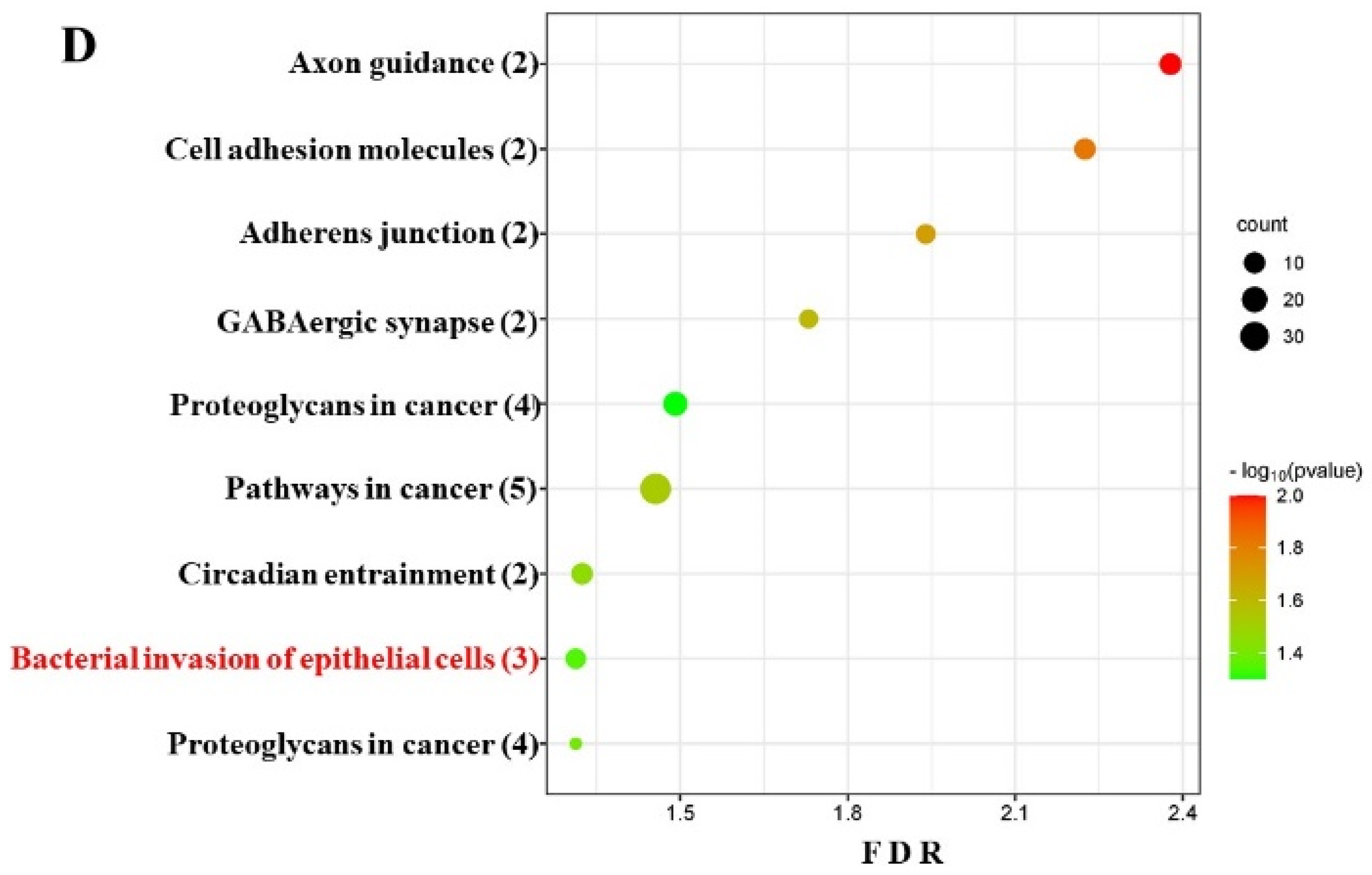
Figure 3.
A. A total of 37 genes (identified by KEGG) were involved in the pathways in cancer and (B) 10 genes were involved in bacterial invasion of epithelial cells signaling pathway during 16 weeks F. nucleatum infection. Red boxes indicate genes significantly increased expression during pathways in cancer and bacterial invasion of epithelial cells. Green boxes indicate no change in gene expression. Many pathogenic bacteria can invade phagocytic and non-phagocytic cells and colonize them intracellularly, then become disseminated to other cells. Invasive bacteria induce their uptake by non-phagocytic host cells (e.g. epithelial cells) using two mechanisms referred to as the zipper model and trigger model. Listeria, Staphylococcus, Streptococcus, and Yersinia are examples of bacteria that can enter using the zipper model. These bacteria express proteins on their surfaces that interact with cellular receptors, initiating signaling cascades that result in close apposition of the cellular membrane around the entering bacteria. Shigella and Salmonella are examples of bacteria entering cells using the trigger model. An arrow indicates a molecular interaction and a line without an arrowhead indicates a molecular interaction resulting in inhibition.
Figure 3.
A. A total of 37 genes (identified by KEGG) were involved in the pathways in cancer and (B) 10 genes were involved in bacterial invasion of epithelial cells signaling pathway during 16 weeks F. nucleatum infection. Red boxes indicate genes significantly increased expression during pathways in cancer and bacterial invasion of epithelial cells. Green boxes indicate no change in gene expression. Many pathogenic bacteria can invade phagocytic and non-phagocytic cells and colonize them intracellularly, then become disseminated to other cells. Invasive bacteria induce their uptake by non-phagocytic host cells (e.g. epithelial cells) using two mechanisms referred to as the zipper model and trigger model. Listeria, Staphylococcus, Streptococcus, and Yersinia are examples of bacteria that can enter using the zipper model. These bacteria express proteins on their surfaces that interact with cellular receptors, initiating signaling cascades that result in close apposition of the cellular membrane around the entering bacteria. Shigella and Salmonella are examples of bacteria entering cells using the trigger model. An arrow indicates a molecular interaction and a line without an arrowhead indicates a molecular interaction resulting in inhibition.
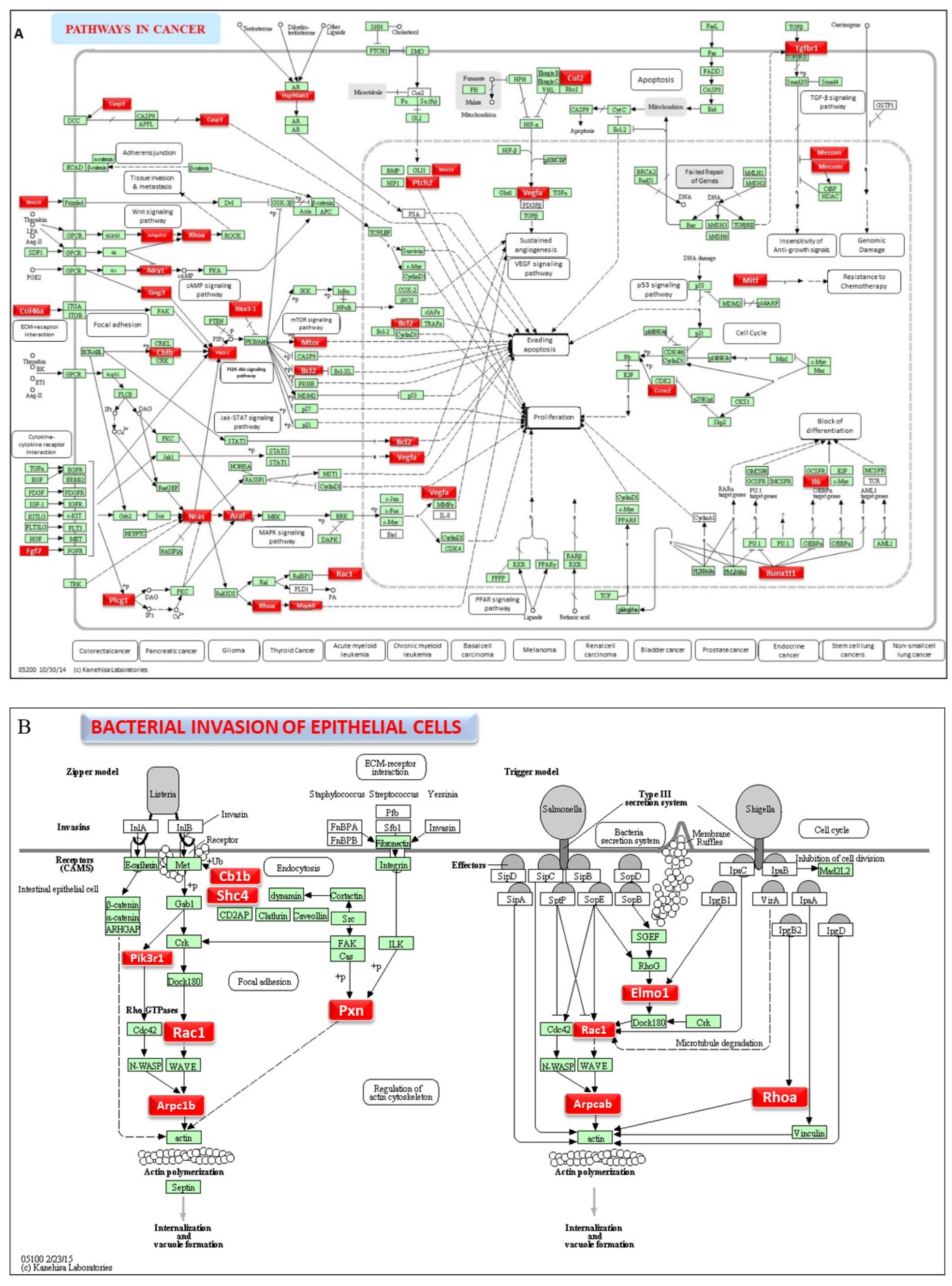
Figure 4.
A summary of the most important features in the five machine learning models using SHAP values. In figures (A–E), feature importance is ranked from the top (the most important) to the bottom (the least important). The x-axis shows the impact that a feature has on the model. The bar charts show the overall impact of a feature whereas the swarm plot shows both the positive and negative impacts. In the swarm plots, each dot represents an instance of a miRNA variable and the color bar shows the value (high to low) of the variable. The three study groups are (1), mice tested at 8 weeks. (2), mice tested at 16 weeks are shown, and (3), 8- and 16 weeks together. .
Figure 4.
A summary of the most important features in the five machine learning models using SHAP values. In figures (A–E), feature importance is ranked from the top (the most important) to the bottom (the least important). The x-axis shows the impact that a feature has on the model. The bar charts show the overall impact of a feature whereas the swarm plot shows both the positive and negative impacts. In the swarm plots, each dot represents an instance of a miRNA variable and the color bar shows the value (high to low) of the variable. The three study groups are (1), mice tested at 8 weeks. (2), mice tested at 16 weeks are shown, and (3), 8- and 16 weeks together. .
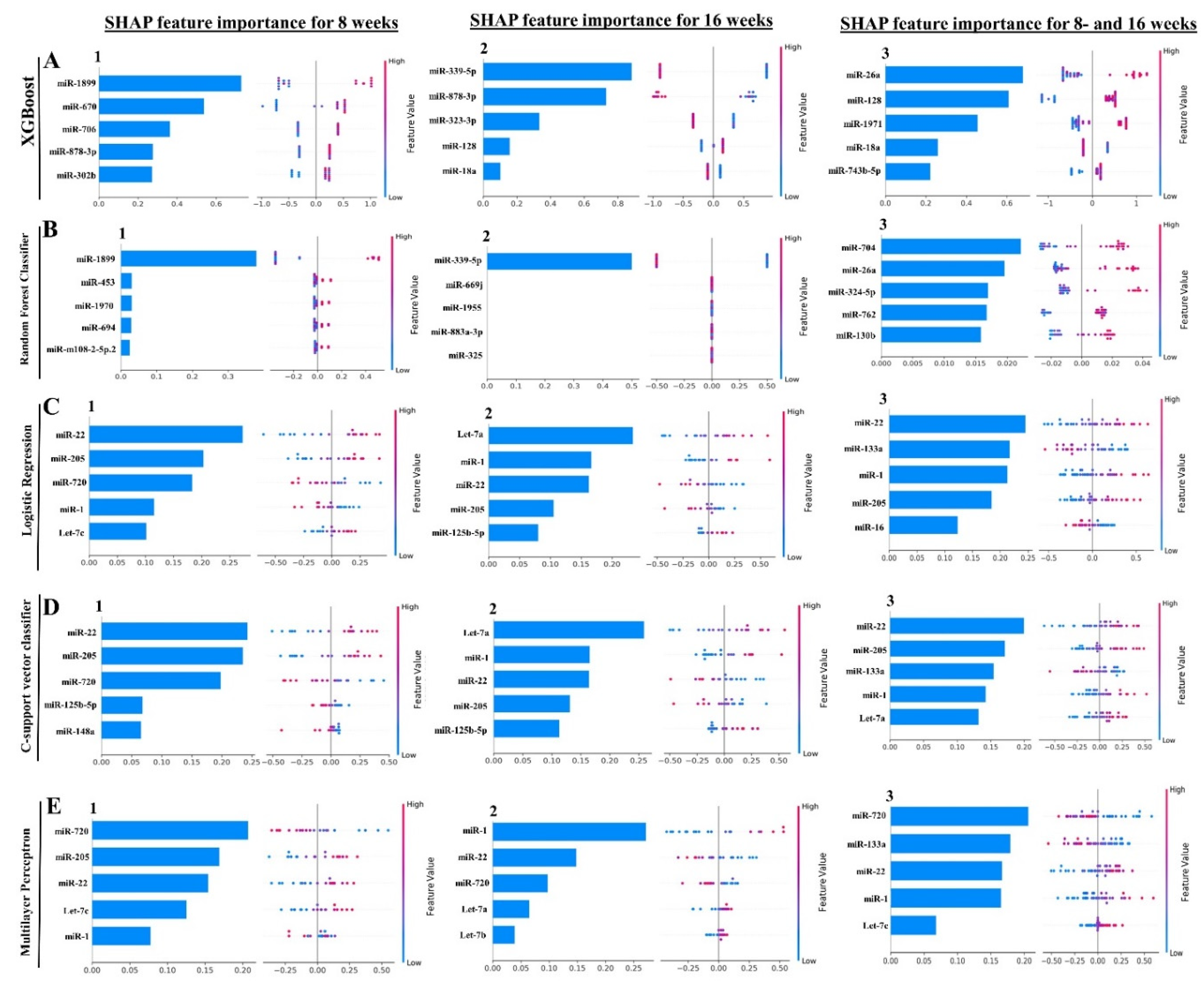
Table 1.
Gingival plaque samples tested positive for F. nucleatum gDNA using PCR.
| Group/Bacteria/Weeks | Positive gingival plaque samples (n=10) | |||
|---|---|---|---|---|
| 8 weeks’ Time point | 16 weeks’ time point | |||
| 2 weeks | 4 weeks | 6 weeks | 12 weeks | |
| Group I/ F. nucleatum ATCC 49256 [8 weeks] | 5/10a | 3/10 | 10/10 | --- |
| Group II/ Sham-infection [8 weeks] | 0/10 | NC | --- | --- |
| Group III/ F. nucleatum ATCC 49256 [16 weeks] | 6/10 | 9/10 | 10/10 | --- |
| Group IV/ Sham-infection [16 weeks] | 0/10 | NC | NC | 0/10 |
a Total numbers of gingival plaque samples that were collected after infections (2, 4, 6, and 12 weeks) and positive infections were determined by PCR analysis. NC – not collected to allow bacterial biofilm to adhere to the gingival surface, invade epithelial cells, and multiply. a The first value corresponds to the number of mice that tested positive for the respective genomic DNA and the second value corresponds to the total number of mice in the group.
Table 2.
Differentially expressed miRs during 8- and 16 weeks F. nucleatum infection in mice.
| Weeks/infection/sex | Upregulated miRNAs (p < 0.05) |
Downregulated miRNAs (p < 0.05) |
|---|---|---|
| 8 weeks – F. nucleatum-infected Vs 8 weeks – Sham infection (n = 10) |
7 (miR-361-5p, miR-99b-5p) | 2 (miR-362-3p, miR-720, miR-362-3p) |
| 8 weeks – F. nucleatum-infected female Vs male (n = 5) |
12 (miR-206, miR-210) | 12 (miR-376a, miR-350) |
| 16 weeks- F. nucleatum-infected Vs 16 weeks – sham infection (n = 10) |
7 (miR-361-5p, miR-99b-5p) | 13 (miR-323-3p, miR-488, miR-350) |
| 16 weeks – F. nucleatum-infected Female vs. male (n = 5) |
5 (miR-152, miR-125b-5p) | 14 (miR-375, miR-210, miR-376a, miR-362-3p) |
| 8 weeks – F. nucleatum-infected Vs 16 weeks F. nucleatum-infected (n = 10) |
14 (miR-133a, miR-22) | 47 (miR-323-3p, miR-1902) |
The number of DE miRNAs was shown for F. nucleatum-infected mice after 8- and 16 weeks infections. The commonly expressed miRNAs between 8- and 16 weeks bacterial infected groups are shown in brackets. Two of the miRNAs expressed in bacterial-infected groups were unique and specific to the 8- and 16 weeks infections.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



