Submitted:
01 October 2024
Posted:
02 October 2024
You are already at the latest version
Abstract
Newcastle disease is a global disease, spreading quickly during outbreaks in poultry and leading to significant economic loss due to high morbidity, mortality, trade restrictions and embargoes. This disease has been present in India for the past 96 years, since 1928. Antigenic differences among Newcastle disease virus (NDV) strains are resulting in massive outbreaks in vaccinated and un-vaccinated poultry flocks around the globe. Synanthropic bird reservoirs contribute significantly to the extensive genetic diversity of the virus. This diversity, along with the vulnerability of many avian species, has led to diagnostic and vaccination failures. The genetic attributes of the circulating strains in India, meanwhile, remain mostly undisclosed. Ongoing evolution of spectrum of NDV genotypes in India, this review paper underscores the need for vigilant monitoring and adaptation of vaccination strategies to address emerging variants.
Keywords:
newcastle disease virus
; chicken
; vaccines
1. Introduction
Poultry represents one of the most commonly available, cheapest and acceptable sources of protein that accounts for more than 40% among the livestock [1]. In recent decades, the poultry industry has experienced remarkable expansion due to increasing demand for eggs and meat. United States and China are the leading global producers in the poultry meat and egg segments respectively. Since 1990, there has been a remarkable increase of over 100 percent in global egg production [2]. Poultry production in India has witnessed a radical transition from handful rearing of native chickens in backyard to high-capacity farms with environmental controlled settings boasting annual turnover of more than 10 billion USD [3]. In India, the broiler and layer sectors are steadily growing, ranking third and fourth globally by producing 138.38 billion eggs [4] and 4.3 MMT of poultry meat [5], respectively. On the other hand, diversified poultry accounts more than 1.43% of total poultry population. However, this rapid expansion brings challenges like antimicrobial resistance, residues, and frequent disease outbreaks [6]. Repeated outbreaks of several infectious diseases specially of viral origin are hampering its growth rate with Newcastle Disease (ND) & Avian Influenza (AI) are the most concerned diseases. Globally, a total of 2606 lakh cases were recorded in poultry with 40.22% death rate (Table 1). The probability of NDV among laying chickens during first phase of egg production is around 10% with resultant 15% production loss [7]. The economic impact because of NDV outbreaks in US and Bangladesh poultry flocks was 162 and 288 million dollars respectively [8,9].
Newcastle disease (ND) is one of the most infectious avian diseases caused by avian paramyxovirus type1 (APMV-1) with an ability to infect over 250 species of poultry [10,11] and claiming loss of millions of dollars to the avian industry. World Organization for Animal Health [12] terrestrial animal health code defines the Newcastle Disease as an infection of birds caused APMV–1 that either having intracerebral pathogenicity index (ICPI) of 0.7 or greater in day-old chicks (Gallus gallus) or multiple basic amino acids at the C-terminus of the F2 protein and phenylalanine at residue 117, which is the N-terminus of the F1 protein. In poultry, NDV is reflected in four disease forms such as Velogenic (viscerotropic and neurotropic), Mesogenic, Lentogenic and Asymptomatic enteric form.
Newcastle disease Infection in chickens is characterized by high mortality, torticollis, greenish diarrhoea, gasping, watery albumens and affecting mainly trachea, proventriculus, intestinal segments and caecal tonsils (Figure 1). Along with Highly Pathogenic Avian Influenza, this disease is recognized worldwide as one of the two most destructive and dreaded diseases causing up to 100% deaths in unvaccinated poultry flocks [13]. Although, all species are susceptible, the disease progression and mortality vary among species and breed.
2. Newcastle Disease Virus (NDV) and Genotypes
2.1. NDV Genotypes: A Global Update
Newcastle Disease is the second most widespread disease globally, affecting many countries after rabies [14] with significant prevalence. High number of outbreaks have been recorded in Asian followed by African region with least impact in Europe and United States (Figure 2). Functional fusion (F) protein is the primary determinant of APMV-1 pathogenesis which is generated by its precursor F0 within the fusion protein cleavage site (FPCS) [15]. Based on FPCS, ND viruses have been categorized into two classes wherein the class-I represents 9 genotypes with genome size of 15,198 nucleotides and class II as 11 genotypes [16,17]. Class I genotypes exhibit a global distribution among wild birds and are often non-pathogenic to chickens but still could be traced in live bird markets. The class II encompasses a majority of highly pathogenic viruses, as well as certain non-pathogenic and vaccine strains [18] with worldwide circulation [19]. The lower genetic diversity of class I viruses than class II may be due to their circulation primarily among wild birds having no history of ND vaccination, resulting in reduced immune pressure on these viruses [20].
In 1926, ND was first documented in Java of Indonesia [21,22]. Subsequently, in 1927, the virus was isolated by Doyle in Newcastle upon Tyne, England, and designated as NDV. Following this discovery, numerous outbreaks of virulent NDV affecting various avian species were reported across continents, including Asia (China, Korea, India, Japan, Sri Lanka, Saudi Arabia, Kazakhstan, Philippines), Africa (Kenya), Australia, Europe (France, Italy, England, Scotland, Spain, and Russia) and North America (United States of America, Canada, and Costa Rica) [23,24]. Ever since the first emergence in 1926, there have been five global ND pandemics, each originated by distinct genotypes. The inaugural pandemic, spanning from 1920 to 1960, emerged concurrently in Southeast Asia and Europe, requiring approximately three decades to proliferate globally. This pandemic was driven by NDV variants possessing genotypes I, II, III, and IV [25]. Notably, chickens, waterfowl, and various bird species were primary targets of infection, with regional variations in disease manifestation [26]. During the 1960s and 1970s, a second pandemic potentially originated in the Middle East. This was driven by the intensified commercialization of the global poultry industry and increased international trade of parrots. This phase primarily featured NDV genotypes V and VI, posing a threat to ornamental and caged bird populations [27,28]. The third pandemic, attributed to genotype VIb isolates, emerged in the late 1970s initially among racing pigeons but swiftly spread globally, hampered by challenges in implementing rigorous husbandry practices within racing pigeon communities [19]. Presently, the fourth ongoing pandemic, believed to have commenced in the late 1980s, is linked to genotypes V, VI, VIII and more specifically VII.1. This pandemic has inflicted significant economic loss to the poultry industry across Southeast Asia, Africa, America, Middle East and Europe, [29,30,31].
Currently, fifth panzootic was originated in Asia in late 2000 with genotype VII.2 prominently. Given its worldwide prevalence in well-entrenched epidemic form, this disease poses a permanent threat to both small-scale and industrial poultry rearing. Across the regions, Asia followed by Africa, America, Europe and Oceania revealed to have highest number of NDV cases and associated deaths with several genotypes reported (Table 1). Since 2005, 131 countries have notified the NDV outbreaks in poultry. Iran is the country with highest burden of NDV outbreaks followed by Vietnam, China, India and Afghanistan. Genotype VII has been emerged as a dominant version in Iran since 2011 affecting various poultry species [32]. Recent outbreaks despite rigorous vaccination in Middle East, Asia, Europe, Africa & South America were due to Genotype VII.1.1 and Latin America was associated with genotypes V, VI, VII, XII, XVI [33]. Australia officially declared NDV free zone in 2002 and no outbreaks were detected till 2011 in commercial poultry populations [34,35].
Early genotypes (pre-1960s) have a genome length of 15,186-nt, while contemporary genotypes (post-1960s) have 15,192-nt. G124S and K192N alterations in the F gene of ancient genotypes III and IV viruses created genotype I viruses, which evolved into genotype II viruses by adding L69M and D82E [36]. Research relevant to Genotype VII revealed 273 publications since 2004, while Genotype XIII was reported since 2012 with 38 publications indicating the emerging scenario of Genotype XIII (Figure 3).
Genotype VII has played a significant role in the most recent fourth pandemic of Newcastle Disease Virus (NDV), becoming the dominant strain globally. This genotype exhibits a complex diversity, with sub-genotype VIIa primarily impacting countries in Asia and Europe, whereas genotype VIIb is more prevalent in South Africa. Furthermore, sub-genotypes VIIc, d, e, f, g, h, and i have been identified in isolates from China, Kazakhstan, and South Africa [19,37]. The newly characterized sub-genotype VIIi exhibits genetic similarity to strains identified in Indonesia, as well as those collected in Pakistan and Israel in 2013 [38]. This sub-genotype has been responsible for Newcastle Disease (ND) outbreaks in Pakistan since 2012 [37] and is deemed enzootic due to its widespread distribution, thus contributing to the ongoing fifth ND panzootic. In Israel, there has been a transition from the previously dominant sub-genotypes VIId and VIIb to VIIi since 2012 [39].

Table 1.
Global occurrence of NDV genotypes, cases and deaths reported from 2005-2024 in different regions [12].
Table 1.
Global occurrence of NDV genotypes, cases and deaths reported from 2005-2024 in different regions [12].
| Region | Total cases (in lakhs) |
Deaths (in lakhs) |
% Deaths | Genotype | References |
|---|---|---|---|---|---|
| Asia | 2135.32 | 828.61 | 38.80 | VIId | [40] |
| VII | [41] | ||||
| VIIi | [42] | ||||
| VIII | [43] | ||||
| XIII | [44] | ||||
| VII.1.1 | [45] | ||||
| XIII.2.1 | [46] | ||||
| VII.2 | [47] | ||||
| XIII.2 | [48] | ||||
| Africa | 411.33 | 203.33 | 49.43 | NT | - |
| America | 46.40 |
10.35 |
22.31 |
VII VIIa, VIId |
[49,50] |
| Europe | 13.01 | 5.83 | 44.81 | VIIa | [51] |
| VII.2 |
[52,53] | ||||
| Oceania | 0.00017 | 0.00017 | 100.00 | NT | No genotypes reported |
2.2. NDV Genotypes: An Indian Update
Newcastle disease in India was first witnessed in Ranikhet of Uttarakhand [Edwards, 1928, [54]] followed by Tamil Nadu region [55] and continued to pose a threat to both commercial and backyard poultry productions. Currently, India is under endemic situation for ND resulting in substantial economic losses. Recurring outbreaks affecting many avian species are still occurring in Southern, Northern, Central and Northeastern parts as detailed in Table 2 and Table 3. In India, virulent NDV strains were reported from diverse avian species viz. pigeons, Mynahs, Emu, Owl, peafowl and quails besides domesticated chickens. The relative risk ratio of Newcastle disease is higher for intensive production systems with respect to semi-intensive (6.7) and backyard or free range (2.1). Similarly, the risk ratio for broilers is of 4.5 and 4.9 with respect to commercial layers and backyard birds [56]. Khorajiya et al. [57] reported economic loss of 45,000 USD in vaccinated layer flocks of Gujarat, India due to GXIII NDV induced infection. Mixed poultry production systems, live and wet markets, porous borders, high density farms, open transportation of live birds supports the frequent NDV outbreaks [58].
Ever since from 2005, a total of 5071 outbreaks with 3.9 million attacks and 0.22 million deaths were noticed in India till 2023 [12] with a death rate of 5.96% in different classes of poultry (Figure 4; Table 2 and Table 3). Higher outbreak intensity was noticed in North-Eastern, Punjab and Tamil Nadu regions, while least outbreaks were recorded in Telangana, Madhya Pradesh, and Himachal Pradesh (Figure 5). Three genotypes (IV, VII and XIII) belonging to 5 different classes have been recorded in India till date [59,60,61]. Genotype XIIIb (KX372709-KX372711) was first reported in 2006 in commercial chickens at Nagpur [62]; while Genotype VII existence is dated back to 1989 in chickens at Tamil Nadu region [59]. In chickens, Genotype XIIIb (Acc. no. KX372709-KX372711) was first reported in 2006 in commercial chickens in Nagpur [Morla et al., 2016, 62], while the existence of Genotype VII dates back to 1989 in the Tamil Nadu region [59].
The study on NDV isolates from pigeons, using monoclonal antibodies for antigenic analysis, found evidence of atypical antigenic types of virus circulation in India, which may contribute to vaccine inefficacy [63]. The occurrence of NDV outbreaks in vaccinated poultry flocks with Genotype XIII such as Pandu-KR072665 from Ranchi, KX345397 from Kamrupa, and XIII from Bareilly) in India suggests ongoing virus evolution. Further, the isolation of Genotype XIIIb from vaccinated layer and broiler flocks from 2006-2014 from Central India with up to 93% mortality suggesting the panzootic potential in the country [62]. Velogenic NDV strains of Genotype-IV isolated from Indian chicken and pigeon in 1997 and 2000 respectively provides the strong evidence of its circulation in Indian sub-continent with an origin indigenous in nature [59].
In mixed viral infections, especially during bird-to-bird transmission NDV strain sequences may change. In India, Genotypes II, III, VI, VIII, XIIIa are reported from 1989-2013 in wild and domesticated poultry of which Genotypes II, III, VI are genetically distinct [64]. Jakhesara and colleagues [65] identified the circulation of Genotypes II, IV and XIII in vaccinated chicken flocks from different parts of India. ND viruses recovered from peafowl indicated the persistence of genotype II in non-domesticated avian host [66]. Higher mortality of 79.4% in layer and broiler flocks of Northern India revealed to caused by an isolate of Genotype XIIIa [60]. Phylogenetic and evolutionary distance analyses of NDV revealed a circulation of novel sub genotype of VIIIe in vaccinated chicken flocks of Tamil Nadu [67]. Exacerbation of clinical signs upon co-infection of NDV Genotype XIII and XIIIe with Low pathogenic avian influenza was reported in Southern region of India [68]. Very recently, whole genome sequencing of NDV isolated from commercial layer farms of Chhattisgarh region revealed the presence of genotype VII.2 caused outbreaks in 2023 [61]. In addition, emergence of novel genotypes XXII.1 and XXII.2 from North-Eastern states are being witnessed in the country [69]. A recent outbreak among commercial chickens in Kashmir was caused by Genotype VIIi and linked to the NDV strain Chicken/Israel/2011/1115 818 [70].
Different genotypes of NDV reported in various regions across the country was illustrated in Table 2. To understand the dynamics of vNDV genotypes across various geographical locations of India, the universally available nucleotide sequences submitted during various outbreaks were downloaded and analyzed. Recent analysis of nucleotide sequences reveals distinct geographic variations with GXIII prevalent in South and GVII in Northern parts of India (Figure 5a). This regional differentiation suggests that nucleotides are somewhat unique to specific districts or regions, indicating localized evolutionary pressures. Furthermore, recent strains from 2022 and 2023 showed a strong nucleotide similarity, suggesting a common ancestor closely related to Genotype VII (Figure 5b). These findings highlight the potential influence of geographic and environmental factors on strain evolution and distribution. Comparatively higher death rates in chickens and other poultry were observed to be associated with Genotype XIII than Genotype VII (Table 3).
The evolutionary history of Genotype VII and XIII strains was inferred by using the Maximum Likelihood method and Tamura-Nei model [71]. A bootstrap consensus tree, derived from 500 replicates [72], represents the evolutionary history of the analyzed taxa. Branches with less than 50% bootstrap support were collapsed. Initial trees for the heuristic search were automatically generated using the Neighbor-Join and BioNJ algorithms on a pairwise distance matrix estimated with the Tamura-Nei model, selecting the topology with the highest log likelihood value. This analysis included 43 nucleotide sequences, with a total of 479 positions in the final dataset. Evolutionary analyses were performed using MEGA software version 11 [73].
Table 2.
NDV Genotypes isolated from different poultry species across India (2005-2023).
| Year | Region | Species | Genotype | References |
|---|---|---|---|---|
| 2006-2012 | Central India | Chicken | XIIIb | [62] |
| 2016-2020 | North-Eastern India | Chicken | XXII.1, XXII.2 & XIII | [69] |
| 2010-2012 | Southern India | Emu | XIII | [74] |
| 2011-2013 | Northern India | Pea fowl | VIIi & XIII | [75] |
| 2014 | Southern and Northern India | Chicken | XIII | [76] |
| 2015 | North-Eastern India | Chicken | XIII & XIIIc | [77] |
| 2015-2016 | Northern India | Chicken | XIIIa | [60] |
| 2015-2016 | Southern India | Chicken | XIII & XIIIe | [68,78] |
| 2019 | Eastern India | Commercial and backyard poultry | XIII | [79] |
| 2023 | Northern India | Chicken | VII.2 | [61] |
Table 3.
Details of cases reported, deaths and genotypes identified in different locations of India.
Table 3.
Details of cases reported, deaths and genotypes identified in different locations of India.
| Region | Cases | Deaths | % Deaths | Genotype | References |
|---|---|---|---|---|---|
| Andhra Pradesh | 2941 | 1581 | 53.76 | XIII | [65] |
| Andaman and Nicobar | 26843 | 2437 | 9.08 | ||
| Arunachal Pradesh | 1889 | 762 | 40.34 | ||
| Assam | 20496 | 5996 | 29.25 | XIII & XIIIc | [77] |
| Bihar | 35 | 15 | 42.86 | ||
| Chhattisgarh | 4869 | 1213 | 24.91 | ||
| Daman & Diu | 83 | 4 | 48.19 | ||
| Goa | 24 | 4 | 16.67 | ||
| Gujarat | 3773 | 442 | 11.71 | XIII | [57,80] |
| Haryana | 2624586 | 59284 | 2.26 | VIIi |
[64,75] |
| XIII | [65] | ||||
| Jammu & Kashmir | 30773 | 4642 | 15.08 | ||
| Jharkhand | 32 | 0 | 0.00 | ||
| Karnataka | 5720 | 4709 | 82.32 | ||
| Kerala | 55936 | 3701 | 6.62 | ||
| Lakshadweep | 26473 | 11735 | 44.33 | ||
| Maharashtra | 13496 | 12768 | 94.61 | XIIIb | [62] |
| Manipur | 73240 | 3516 | 4.80 | ||
| Meghalaya | 23847 | 7801 | 32.71 | ||
| Mizoram | 26694 | 9115 | 34.30 | ||
| Nagaland | 25934 | 5439 | 20.97 | ||
| Orissa | 205 | 25 | 12.20 | ||
| Pondicherry | 9552 | 2932 | 30.70 | ||
| Rajasthan | 13140 | 3130 | 23.82 | ||
| Tamil Nadu | 684845 | 17540 | 2.56 | XIII &XIIIe | [65,68,78] |
| Tripura | 144320 | 34160 | 23.67 | ||
| Uttar Pradesh | 12 | 9 | 75.00 | XIIIa | [60] |
| XIII, VIIi | [64] | ||||
| West Bengal | 95642 | 40390 | 42.23 | XIII | [65] |
3. Newcastle Disease: Vaccines and Vaccination
Vaccination is the most viable and practically feasible strategy for prevention of control of NDV outbreaks in endemic countries [81]. All the countries share common practices, including the use of commercial live or inactivated vaccines (prepared from genotype II) and similar vaccination regimes (number of doses, route of administration, combination of live and inactivated options). The seroprevalence rate of ND in India is as high as 83% [82]. In India, nearly all farmers carryout ND vaccination [83] with variable periodicity. Currently, Genotypes I and II are the most used in NDV vaccines worldwide including India. Several lentogenic and mesogenic strains have been in use for decades with varying ICPI efficacies (Figure 7) in different formats (Figure 8). Among lentogenic strains, cloned versions have lower ICPI and V4 and Ulster 2C HAS ‘0’ ICPI indicating their least pathogenic effects on chicks. Further cloned strains possess low virulence with better efficacy. For disease control, immunization using lentogenic and mesogenic strains in both live and inactivated/killed vaccine formats is currently practiced at different stages, depending on the birds’ production status.
Owing to the immediate immune spiking, live vaccines having lentogenic virus are commonly employed to prime the immune system of bird followed by vaccination with mesogenic strain and inactivated vaccines. However, the vaccination is tailored depending upon infection pressure and type of chicken production (Table 4). For example, administration of ND killed (concentrated) vaccines is common in commercial broilers upon hatch to ensure lasting immunity in addition to traditional live vaccines. In India, vaccines are available in various formats and combinations (Table 5), with some being imported and others manufactured locally.
R2B (Mukteshwar), a mesogenic ND vaccine strain, is widely utilized in the Indian subcontinent. It has demonstrated excellent efficacy in older birds (6–8 weeks) by providing long-lasting immunity; however, it has been shown to be pathogenic for young chicks. Phylogenetic analyses of vaccine strains in India, including R2B, based on F, HN, and whole genome sequences, categorize them within Genotype II [85]. This viral strain originated from the passage of one of three Indian field isolates at the Indian Veterinary Research Institute in 1945 and has been employed as a vaccine candidate for booster immunization since then [86]. R2B was integrated into vaccination programs within the Indian poultry industry during the 1980s and is currently produced by major Indian poultry vaccine manufacturers such as Indovax Private Limited (Haryana), Hester Biosciences (Gujarat), and Ventri Biologicals (Maharashtra), with billions of doses administered to date. The inclusion of this strain in vaccination protocols for commercial laying stocks has markedly increased egg production from 3 billion in 1961 to 139 billion by 2023 (Figure 9). This enhancement is attributed to a substantial reduction in Newcastle Disease (ND) incidence to below critical levels (Figure 10).
4. Does Heterologous Genotype-Based Vaccines Offer Better Protection than Homologous?
Regardless of vaccination regimes that depend on conventional heterologous vaccines for the circulating NDV, frequent outbreaks of ND or non-optimum control occur, with considerable virus shedding and severe economic loss (mortalities and/or drop in egg production). The examination of all the Indian isolates of Genotype II, VII, XIII and other documented viruses revealed a cleavage site (RRQKRF in velogenic and GRQGRL in lentogenic) indicating the high pathogenic potential in these circulating virulent strains (Figure 11). Amino acid sequences of vaccine Genotype II with Genotype VII and XIII showed great diversity at 4, 8, 9, 13, 16, 20, 22, 29, 30, 82, 112-117, 121, 124, 145, 192, 195, 203, 231, 232, 272, 288, 396, 421, 482, 552 and 553 (Table 6). Owing to 3.93-17.31% nucleotide (Table 7) and 10.14-14.96% amino acid (Table 8) variability among vaccine and circulating virulent field strains (Genotypes VII & XIII), the protection levels are questionable. Although vaccination with Genotype-II for ND control has been widely accepted strategy globally, its efficacy is variable concerning types and genotypes. Recent instance of 73.68% disease incidence in completely ND-vaccinated layer flocks of India [87] provides further evidence of the inefficacy of conventional genotype vaccines.
Genotype VII NDV homologous vaccine induced higher haemagglutination titers with reduced clinical disease and mortality in birds challenged with genotype VII NDV strain [88]. The integration of recombinant genotype VII-based NDV inactivated vaccines, such as NDV IBS002 and NDV AF2240-I, alongside the existing commercial immunization regimen, provides comprehensive protection with reduced virus shedding against corresponding virulent strains. This approach addresses the limitations of the LaSota strain, which has shown reduced efficacy in certain contexts [89]. Efficacy studies of commercially available live and killed Genotype VII based vaccines with a prime-boost vaccine regime against Genotype VII1.1 challenge in broilers demonstrated 93-100% survivability with the virus shedding continued till 10 days post-challenge (dpc) [90]. Vaccination with Genotype XIII.2 (killed BD-C161/2010) homologous to infection outbreak in chickens resulted in sustained liveability (100%) with no clinical illness, reduced viral shedding till 7 dpc [48]. Similarly, immunization of virulent NDV genotype VII infected broilers with recombinant live and autogenous inactivated GVII 1.1 based vaccine reduced mortality, clinical symptoms, mean severity index of internal organ lesions and reduced viral shedding of tracheal and cloacal swabs [91].
Homologous ND vaccines often reduce viral shedding in oro-pharyngeal swabs and decrease the number of birds shedding virus [92,93,94]. Further, this also achieves sterile immunity which is otherwise not possible with mismatched genotypes due to induction of suboptimal immunity [95]. Administration of a booster dose of inactivated genotype VII followed by Lasota live vaccine (genotype II) in layers under the Genotype VII experimental infection module witnessed a sustained egg production of up to 87.5% [96]. Genotype VII-based ND vaccine, when administered with montanide to birds infected with genotype VII NDV induced 100% protection with circulating antibodies, lasting 12 weeks post-vaccination and gradually declines at 16 weeks [97]. In a study by Wang et al. [98], chickens infected with the virulent DHN3 strain, when vaccinated with the inactivated form of genotype VII-based rDHN3-mF, exhibited a significantly stronger anti-DHN3 antibody response. This vaccination provided superior (100%) protection and faster viral clearance compared to Lasota strains. When, genotype VII-based NDV vaccine loaded with poly-lactic-co-glycolic (PLGA) nanoparticles afforded 100% protection to the chickens infected with genotype VII virulent NDV [99]. The adjuvant potential on induction of Th1 cell mediate and humoral immunity was also superior with enhanced IFN-γ expression.
Recombinant vaccines (Vectormune ND) with Genotype I based D-26 strain showed better protection rates of up to 100% to different genotypes i.e., IV, V VIIa, VIIb, VIId and VIII with significant reduction of virus titers (Ceva Animal Health study). Genotype II based vaccines in different strain formats yielded varied results to velogenic genotype VII challenge in chickens. Lasota, Clone, 12IR and B1 vaccines resulted in 90, 80 and 60% protection rate with no viral shedding noticed in any of the vaccinated groups [81]. Vaccination with LaSota strains accompanied by Toll-like Receptor adjuvants (Imiquimoid & OD-1826) increased antibody titers and reduced viral shedding in birds challenged with genotype VI velogenic NDV [100]. Although, heterologous/mismatch vaccines were advocated against NDV challenge their efficacy was reported with limited success restricting to genotype VII. Further, the success of genotype II based vaccines against genotype XIII strains is questionable. Matching genotype-based vaccines results in virus spill overs with limited protection (Table 9).
5. Strategies for Updating Vaccine Genotype to Mitigate Dominant Field Virus Strains
The occurrence of recombination in NDV strains has been a subject of controversy. Nevertheless, experimental evidence supporting the occurrence of recombination in NDV is currently lacking. However, it is necessary to adopt a comprehensive strategy to assess ND virulence. This involves characterizing novel strains using the protocols established by the World Organization for Animal Health (WOAH). Currently, it may be deemed daunting to assert that the utilization of reverse genetics has the potential to be harnessed in the development of a secure recombinant attenuated vaccination for poultry. Some of the plausible strategies for generating homologous vaccines from virulent strains in the field have been discussed under this section.
5.1. Classical Inactivation of Field Viruses
A killed or inactivated vaccine is one of the primary methods used to control and prevent the spread of this disease. The development of a ND killed vaccine involves virus isolation, inactivation, adjuvant addition, formulation and testing. A virulent strain of the NDV strain can be isolated from the infected birds and further propagation can be done by increasing the titre by inoculating in embryonated chicken eggs. The most common inactivated reagents that are available for inactivating the conventional killed vaccines are formaldehyde and β-propiolactone [101]. These reagents inactivate the virus ensuring that it should not replicate but retain its ability to induce an immune response. To boost the immune response, an adjuvant is added to the inactivated virus which will improve the vaccine’s effectiveness by prolonging the presence of antigen in the body. The commonly used adjuvants are aluminium hydroxide [102] or montanide [103]. Recently, to activate aluminium adjuvants to induce cell-mediated immunity the composite nano adjuvant N-2-HACC-Al NPs were synthesized by the N -2 Hydroxypropyl trimethyl ammonium chloride chitosan (N-2-HACC) and aluminum sulfate (Al2(SO4)3). It showed higher vaccine efficacy than those of the commercially combined vaccine [103]. Furthermore, encapsulation of inactivated NDV with PLGA particles induced strong immune responses compared to the commercial oil-based adjuvanted NDV-killed vaccine [98]. So, further formulation can be done by the addition of adjuvants and stabilizers. The use of killed vaccines in controlling ND in poultry is highly significant because it does not contain live virus making it safer as live vaccines might cause mild reactions and lead to complications. It can be formulated in such a way as to cover multiple strains, offering broader protection against various field strains as well as it will provide long-lasting immunity with adjuvants.
5.2. Reverse Genetics Approach
Reverse genetics technology has revolutionized the field of virology, particularly in the development of vaccines for avian diseases such as the Newcastle Disease Virus (NDV), which causes severe economic losses in the poultry industry. This technology allows scientists to manipulate the genome of RNA viruses, including NDV, facilitating the creation of improved, targeted vaccines. The application of reverse genetics in NDV vaccine development has led to significant breakthroughs in the production of more effective, safer, and targeted vaccines for poultry. Reverse genetics technology first emerged in the 1990s and was a major leap forward in molecular biology. For negative-sense RNA viruses like NDV, it was difficult to manipulate viral genomes because their RNA genome is not infectious on its own. However, in 1996, Palese., P [105] and his co-workers pioneered the reverse genetics technique for negative-strand RNA viruses, allowing recombinant viruses to be generated from cloned cDNA. In the case of NDV, reverse genetics was successfully applied in 1999 when recombinant NDV was recovered using this technology [106]. This opened new possibilities for studying and manipulating NDV’s genome for vaccine development.
Traditional live-attenuated vaccines for NDV have been effective, but they sometimes pose a risk of reverting to virulence [107]. Reverse genetics has addressed this issue by enabling the targeted attenuation of the virus, ensuring that the vaccine strains remain stable and do not revert to virulence. By introducing specific mutations in key genes such as the “F” the pathogenicity of NDV can be reduced while maintaining its ability to stimulate a strong immune response. Vectored vaccines use NDV as a vector to express foreign antigens from other pathogens [94]. This emerging NDV vector can be utilized for both bivalent and multivalent vaccine preparations, targeting not only NDV but also other important poultry viral diseases. NDV’s ability to stably accommodate foreign genes, combined with its capacity to induce mucosal, humoral, and cellular immunity, makes it an ideal candidate for such vaccines. A variety of recombinant poultry viral vaccines have been developed using NDV as a vector, offering protection against multiple pathogens. Notable examples include vaccines targeting Avian Influenza (AI) virus [108], Infectious Bursal Disease Virus (IBDV) [109], Infectious Laryngotracheitis Virus (ILTV) [110] Reovirus [111], Infectious Bronchitis Virus (IBV) [112], and Chicken Infectious Anemia (CIA) [113]. Chimeric recombinant based Genotype VII.1.1 vaccines have gained commercial success with ‘0’ ICPI and higher protection levels (up to 93%) in ND challenged birds. These vaccines involved Lasota and VG/GA strain as backbones into which the F and HN genes of KBNP-C4152R2L and CK/ME/19 strains were inserted [89]. These advancements highlight the versatility of NDV-vectored vaccines in offering broad-spectrum protection against multiple poultry diseases.
Reverse genetics has also been instrumental in the development of DIVA vaccines, which allow for the differentiation between vaccinated animals and those naturally infected with NDV. This is done by modifying the NDV genome to delete or modify certain antigenic genes, providing a unique marker for distinguishing between vaccine-induced immunity and natural infection. This advancement is crucial for disease surveillance and control in poultry populations. The future of reverse genetics in NDV vaccine development is promising. Advances in gene-editing technologies, such as CRISPR, could further refine the manipulation of the NDV genome. Additionally, the development of more sophisticated vaccine delivery systems, such as in-ovo vaccination combined with reverse genetics-designed vaccines, could provide even greater efficiency and protection for poultry [114].
5.3. CRISPR-Based Gene Editing Vaccines
CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and the associated Cas9 protein represent a powerful third generation of programmable genome editing tools. When a virus invades a cell, CRISPR binds to viral RNA, and Cas9 induces double-strand breaks in targeted DNA sequences. A key advantage of the CRISPR/Cas9 system is that its expression plasmids transiently deliver the Cas9 enzyme and gRNA, allowing for gene disruption without integrating transgenes into the host genome [115]. This versatile system has been successfully applied in various avian species, including chickens and quail, to manipulate somatic cells [116], create eggs with modified biochemical compositions [117,118], produce myostatin knockout chickens [119], and study resistance to avian influenza [120]. CRISPR also plays a crucial role in understanding virus-host interactions, viral editing, and the functionality of virulent viral factors. With CRISPR, researchers can easily and precisely knock-in, knock-out, or knock-down genes [121].
In the field of vaccinology, CRISPR is becoming recognized as a precise, cost-effective, and efficient technology that is redefining the traditional recombinant vaccines. This innovative approach allows for the simultaneous insertion of larger gene fragments containing multiple antigens, facilitating the development of multi-valent vaccines that offer extended immunity [121]. Unlike conventional vaccines, CRISPR edited vaccines requires no plaque purification, attenuation and inactivation [122]. Herpes virus turkey (HVT) was developed as a vaccine vector with high immunogenic potential by targeted gene editing of gB, gI, gE [123] for generation of vaccines against Infectious bursal disease [124] Infectious laryngotracheitis (ILT) and avian influenza (HPAI) [125]. Development of homologous vaccines using Genotype VII and XIII for ND control using CRISPR tool mainly includes knock out of FPCS followed by knock-in of suitable genes from Lasota, F, B1 or other genotypes II for the creation of recombinant mutants. Direct editing of negative and positive senses single-stranded viruses can be possible with NHEJ-CRISPR/Cas 9 enzyme complex Cpf1 derived from Prevotella and Francisella [126]. Successful integration of NDV F gene into ILTV viral backbone vector by non- homologous end joining was reported by Atasoy et al. [127] using CRISPR/Cas9 and Cre-Lox system. This probably the first attempt to develop CRISPR mediated multi-valent vaccine. However, selection of protospacer adjacent motifs, off-target effects, delivery methods etc must be given due consideration for CRISPR edited vaccines
the text following an equation need not be a new paragraph. Please punctuate equations as regular text.
6. Conclusion
Newcastle disease threat to poultry and non-poultry existed for decades and continues in the globe. Its existence is endemic in India with complex genotype circulation in poultry flocks. Since the virus is a single serotype with great antigenic & genetic variations, mismatching of current live (G-I, II, V) and inactivated vaccines with dominant field strains (G-VII, VIII, XIII) is possible. Although, the conventional genotype-based vaccines provide clinical protection against different velogenic strains especially GVII and GXIII, their inability to block the viral shedding causes increased viral persistence in the environment. A genotype-matched NDV vaccination for India would be intriguing. A genotype XIII vaccination may improve immunity and protect birds from aggressive NDV epidemics. Administration of live vaccines to homologous field strains decreases viral shedding in the flock and spillage into environments besides eliciting higher hemagglutination titers. Mutations in live vaccines with F & L proteins from field variants guarantees safety. Reverse genetics of virulent strains; Modified live vaccines with F, HN proteins from prevalent field strain in lentogenic backbone and Recombinant vaccine technologies (rNDV_G-VIII) and Mutations in AA of F clevage site (A-VIII) are some of the strategies for the development of homologous NDV vaccines with matched genotype to circulating field viruses. Live recombinant NDV vaccines (HIMMVAC, RINNOVAC EL7, GENOVAC N5) are available globally for Genotype VII alone with its usage restricted to certain regions in the country. Commercial preparation and availability for Genotypes VII, VIII and XIII vaccines are lacking with their safety and immunogenicity features questionable. Updating and optimization of vaccines with homologous genotypes matching with dominant field strains (Genotype VII, VIII, XIII) or importing from other countries may offer sustainable solution for Indian poultry industry. CRISPR based gene edited vaccines offer promising solution for mitigating the infection pressure caused by diverse genotypes.
Author Contributions
G.K. Collected the data, written and edited the manuscript. S.S.K. Developed tabular and figure-based illustrations from the collected data. V.M. Retrieved the universally available sequences and analyzed them with interpretation. A.K.T. Conceived the idea and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: “This research received no external funding”.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Data will be available upon personal request from corresponding author
Acknowledgments
Authors are thankful to Indian Council of Agricultural Research for providing necessary facilities to carryout this study.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- FAO. https://ourworldindata.org/grapher/poultry-production tonnes, Accessed on 20.09.2024.
- Statista. Leading egg production countries. https://www.statista.com/statistics/263971/top-10-countries-worldwide-in-egg-production, Statista, 2024.
- Kolluri, G.; Tyagi, J.S.; Sasidhar, P.V.K. Research Note: Indian poultry industry vis-à-vis coronavirus disease 2019: a situation analysis report. Poult. Sci. 2020, 100, 100828–100828. [CrossRef]
- DAHD. Government of India, Ministry of Fisheries, Animal Husbandry and Dairying, Department of Animal Husbandry and Dairying, Krishi Bhavan, New Delhi. 2023.
- Statista. India: poultry meat production volume. https://www.statista.com/statistics/1358419/india-economic-contribution-of-poultry-meat, Statista, 2023.
- Kolluri, G. and Tiwari, A.K. Newcastle disease virus in chickens: Time to revisit the vaccine updation in India? In International symposium on animal viruses, vaccines and immunity (AVVI 2024), Orissa, India, 9-11, February.
- Charkhkar, S.; Bashizade, M.; Sotoudehnejad, M.; Ghodrati, M.; Bulbuli, F.; Akbarein, H. The evaluation and importance of Newcastle disease’s economic loss in commercial layer poultry. 2024, 2, 1–4. [CrossRef]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: a review of field recognition and current methods of laboratory detection. J. vet. Diagn. Invest. 2011, 23, 637-56.
- Khatun, M.; Islam, I.; Ershaduzzaman, M.; Islam, H.; Yasmin, S.; Hossen, A.; Hasan, M. Economic impact of newcastle disease on village chickens-a case of Bangladesh. Asian. Inst. Res. J. Econ. Bus. 2018, 1, 358-67.
- Alexander, D. J. Newcastle disease. Br. Poult. Sci. 2001, 42, 5–22.
- Aldous, E.W.; Mynn, J.K.; Banks, J.; Alexander, D.J. A molecular epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. Avian Pathol. 2003, 32, 237–255. [CrossRef]
- WOAH-WAHIS. World Organization for Animal Health Information System Interface (WAHIS). Available online: http://wahis.woah.org/#/home (accessed on 11 August 2022).
- World Organization for Animal Health. Infection with Newcastle disease, Chapter 10.9, Article 10.9.1. Terrestrial Code Online Access - WOAH - World Organisation for Animal Health. 2024, Accessed on September 10, 2023.
- World Bank. World livestock disease atlas: a quantitative analysis of global animal health data (2006-2009). Washington, DC: World Bank. http://documents.worldbank.org/curated/en/2011/11/15812714/worldlivestock-disease-atlas-quantitative-analysis-global-animal-health-data-2006-2009, 2011.
- Glickman, R.L.; Syddall, R.J.; Iorio, R.M.; Sheehan, J.P.; A Bratt, M. Quantitative basic residue requirements in the cleavage-activation site of the fusion glycoprotein as a determinant of virulence for Newcastle disease virus. J. Virol. 1988, 62, 354–356. [CrossRef]
- Kim, L.M.; King, D.J.; Curry, P.E.; Suarez, D.L.; Stallknecht, D.E.; Slemons, R.D.; Pedersen, J.C.; Senne, D.A.; Winker, K.; Afonso, C.L. Phylogenetic Diversity among Low-Virulence Newcastle Disease Viruses from Waterfowl and Shorebirds and Comparison of Genotype Distributions to Those of Poultry-Origin Isolates. J. Virol. 2007, 81, 12641–12653. [CrossRef]
- Kim, L.M.; King, D.J.; Suarez, D.L.; Wong, C.W.; Afonso, C.L. Characterization of class I Newcastle disease virus isolates from HongKong live bird markets and detection using real-time reverse transcription-PCR. J. Clin. Microbiol. 2007b, 45, 1310–1314.
- Czeglédi, A.; Ujvári, D.; Somogyi, E.; Wehmann, E.; Werner, O.; Lomniczi, B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res. 2006, 120, 36–48. [CrossRef]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2010, 10, 26–35. [CrossRef]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [CrossRef]
- Du, J.; Xia, J.; Li, S.; Shen, Y.; Chen, W.; Luo, Y.; Zhao, Q.; Wen, Y.; Wu, R.; Yan, Q.; et al. Evolutionary dynamics and transmission patterns of Newcastle disease virus in China through Bayesian phylogeographical analysis. PLOS ONE 2020, 15, e0239809. [CrossRef]
- Amoia, C.F.A.N.; Nnadi, P.A.; Ezema, C.; Couacy-Hymann, E. Epidemiology of Newcastle disease in Africa with emphasis on Côte d'Ivoire: A review. Veter- World 2021, 14, 1727–1740. [CrossRef]
- Turner, A.J.; Kovesdy, L. STUDIES ON THE EPIZOOTIOLOGY OF INFECTION WITH AVIRULENT NEWCASTLE DISEASE VIRUS IN BROILER CHICKEN FLOCKS IN VICTORIA. Aust. Veter- J. 1974, 50, 155–158. [CrossRef]
- Habib, M.; Shabbir, M.Z. Suppl-2, M3: Adaptation of Newcastle disease virus (NDV) in feral birds and their Potential Role in Interspecies Transmission. The Open Virol. J. 2018, 12, 52.
- Wang, J.; Liu, H.; Liu, W.; Zheng, D.; Zhao, Y.; Li, Y.; Wang, Y.; Ge, S.; Lv, Y.; Zuo, Y.; et al. Genomic Characterizations of Six Pigeon Paramyxovirus Type 1 Viruses Isolated from Live Bird Markets in China during 2011 to 2013. PLOS ONE 2015, 10, e0124261–e0124261. [CrossRef]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Diagnostic and Vaccination Approaches for Newcastle Disease Virus in Poultry: The Current and Emerging Perspectives. BioMed Res. Int. 2018, 2018, 1–18. [CrossRef]
- Walker, J.W.; Heron, B.R.; Mixson, M.A. Exotic Newcastle Disease Eradication Program in the United States. Avian Dis. 1973, 17, 486–503. [CrossRef]
- Pearson, G.L.; McCann, M.K. The role of indigenous wild, semidomestic, and exotic birds in the epizootiology of velogenic viscerotropic Newcastle disease in southern California, 1972-1973.. 1975, 167, 610–4.
- Herczeg, J.; Wehmann, E.; Bragg, R.R.; Dias, P.M.T.; Hadjiev, G.; Werner, O.; Lomniczi, B. Two novel genetic groups (VIIb and VIII) responsible for recent Newcastle disease outbreaks in Southern Africa, one (VIIb) of which reached Southern Europe. Arch. Virol. 1999, 144, 2087–2099. [CrossRef]
- Kwon, H.-J.; Cho, S.-H.; Ahn, Y.-J.; Seo, S.-H.; Choi, K.-S.; Kim, S.-J. Molecular epidemiology of Newcastle disease in Republic of Korea. Veter- Microbiol. 2003, 95, 39–48. [CrossRef]
- Tan, S.W.; Ideris, A.; Omar, A.R.; Yusoff, K.; Hair-Bejo, M. Sequence and phylogenetic analysis of Newcastle disease virus genotypes isolated in Malaysia between 2004 and 2005. Arch. Virol. 2009, 155, 63–70. [CrossRef]
- Goudarzi, H.; VAN Borm, S.; Bashashati, M.; Sabouri, F.; Abdoshah, M.; Nouri, A.; Banani, M.; Ebrahimi, M.M.; Molouki, A. Characterization and full genome sequencing of a velogenic Newcastle disease virus (NDV) strain Ck/IR/Beh/2011 belonging to subgenotype VII(L). Acta Virol. 2019, 63, 217–222. [CrossRef]
- Absalón, A.E.; Cortés-Espinosa, D.V.; Lucio, E.; Miller, P.J.; Afonso, C.L. Epidemiology, control, and prevention of Newcastle disease in endemic regions: Latin America. Trop. Anim. Heal. Prod. 2019, 51, 1033–1048. [CrossRef]
- Kirkland, P.D. Virulent Newcastle disease virus in Australia: in through the ‘back door’. Aust. vet. J. 2000, 78, 331-333.
- Murray, G. Australia officially free of Newcastle disease. Aust. Vet. J. 2003, 81, 460-462.
- Yu, L.; Wang, Z.; Jiang, Y.; Chang, L.; Kwang, J. Characterization of Newly Emerging Newcastle Disease Virus Isolates from the People's Republic of China and Taiwan. J. Clin. Microbiol. 2001, 39, 3512–3519. [CrossRef]
- Miller, P.J.; Haddas, R.; Simanov, L.; Lublin, A.; Rehmani, S.F.; Wajid, A.; Bibi, T.; Khan, T.A.; Yaqub, T.; Setiyaningsih, S.; et al. Identification of new sub-genotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Genet. Evol. 2015, 29, 216–229. [CrossRef]
- Munir, M.T.; Chowdhury, M.R; Ahmed, Z. Emergence of new sub-genotypes of Newcastle disease virus in Pakistan. J Avian Res, 2016, 2, 1-7.
- Ghalyanchilangeroudi, A.; Hosseini, H.; Jabbarifakhr, M.; Mehrabadi, M.H.F.; Najafi, H.; Ghafouri, S.A.; Mousavi, F.S.; Ziafati, Z.; Modiri, A. Emergence of a virulent genotype VIIi of Newcastle disease virus in Iran. Avian Pathol. 2018, 47, 509–519. [CrossRef]
- Zhang, Y.-Y.; Shao, M.-Y.; Yu, X.-H.; Zhao, J.; Zhang, G.-Z. Molecular characterization of chicken-derived genotype VIId Newcastle disease virus isolates in China during 2005–2012 reveals a new length in hemagglutinin–neuraminidase. Infect. Genet. Evol. 2014, 21, 359–366. [CrossRef]
- Ewies, S.S.; Ali, A.; Tamam, S.M.; Madbouly, H.M. Molecular characterization of Newcastle disease virus (genotype VII) from broiler chickens in Egypt. Beni-Suef Univ. J. Basic Appl. Sci. 2017, 6, 232–237. [CrossRef]
- Wajid, A.; Dimitrov, K.M.; Wasim, M.; Rehmani, S.F.; Basharat, A.; Bibi, T.; Arif, S.; Yaqub, T.; Tayyab, M.; Ababneh, M.; et al. Repeated isolation of virulent Newcastle disease viruses in poultry and captive non-poultry avian species in Pakistan from 2011 to 2016. Prev. Veter- Med. 2017, 142, 1–6. [CrossRef]
- Sabouri, F.; Marandi, M.V.; Bashashati, M. Characterization of a novel VIIl sub-genotype of Newcastle disease virus circulating in Iran. Avian Pathol. 2018, 47, 90–99. [CrossRef]
- Kabiraj, C.K.; Mumu, T.T.; Chowdhury, E.H.; Islam, M.R.; Nooruzzaman, M. Sequential Pathology of a Genotype XIII Newcastle Disease Virus from Bangladesh in Chickens on Experimental Infection. Pathogens 2020, 9, 539. [CrossRef]
- Ahmed, H.M.; Amer, S.A.; Abdel-Alim, G.A.; Elbayoumi, K.M.; Kutkat, M.A.; and Amer, M.M. Molecular characterization of recently classified Newcastle disease virus genotype VII. 1.1 isolated from Egypt. Int. J. Vet. Sci. 2022, 11, 295-301.
- Hejazi, Z.; Tabatabaeizadeh, S.; Toroghi, R.; Farzin, H.; Saffarian, P. First detection and characterisation of sub-genotype XIII.2.1 Newcastle disease virus isolated from backyard chickens in Iran. Veter- Med. Sci. 2022, 8, 2521–2531. [CrossRef]
- Kariithi, H.M.; Volkening, J.D.; Chiwanga, G.H.; Goraichuk, I.V.; Olivier, T.L.; Msoffe, P.L.M.; Suarez, D.L. Virulent Newcastle disease virus genotypes V.3, VII.2, and XIII.1.1 and their coinfections with infectious bronchitis viruses and other avian pathogens in backyard chickens in Tanzania. Front. Veter- Sci. 2023, 10, 1272402. [CrossRef]
- Hossain, I.; Subarna, J.F.; Kabiraj, C.K.; Begum, J.A.; Parvin, R.; Martins, M.; Diel, D.G.; Chowdhury, E.H.; Islam, M.R.; Nooruzzaman, M. A Booster with a Genotype-Matched Inactivated Newcastle Disease Virus (NDV) Vaccine Candidate Provides Better Protection against a Virulent Genotype XIII.2 Virus. Vaccines 2023, 11, 1005. [CrossRef]
- Perozo, F.; Marcano, R.; Afonso, C.L. Biological and Phylogenetic Characterization of a Genotype VII Newcastle Disease Virus from Venezuela: Efficacy of Field Vaccination. J. Clin. Microbiol. 2012, 50, 1204–1208. [CrossRef]
- Diel, D.G.; Susta, L.; Garcia, S.C.; Killian, M.L.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Complete Genome and Clinicopathological Characterization of a Virulent Newcastle Disease Virus Isolate from South America. J. Clin. Microbiol. 2012, 50, 378–387. [CrossRef]
- Fuller, C.; Löndt, B.; Dimitrov, K.M.; Lewis, N.; van Boheemen, S.; Fouchier, R.; Coven, F.; Goujgoulova, G.; Haddas, R.; Brown, I. An Epizootiological Report of the Re-emergence and Spread of a Lineage of Virulent Newcastle Disease Virus into Eastern Europe. Transbound. Emerg. Dis. 2015, 64, 1001–1007. [CrossRef]
- Steensels, M.; Van Borm, S.; Mertens, I.; Houdart, P.; Rauw, F.; Roupie, V.; Snoeck, C.J.; Bourg, M.; Losch, S.; Beerens, N.; et al. Molecular and virological characterization of the first poultry outbreaks of Genotype VII.2 velogenic avian orthoavulavirus type 1 (NDV) in North-West Europe, BeNeLux, 2018. Transbound. Emerg. Dis. 2021, 68, 2147–2160. [CrossRef]
- Turan, N.; Ozsemir, C.; Yilmaz, A.; Cizmecigil, U.Y.; Aydin, O.; Bamac, O.E.; Gurel, A.; Kutukcu, A.; Ozsemir, K.; Tali, H.E.; et al. Identification of Newcastle disease virus subgenotype VII.2 in wild birds in Turkey. BMC Veter- Res. 2020, 16, 1–8. [CrossRef]
- Edwards J. A New Fowl Disease. March 31st, Mukteshwar, Vet. Res. 1928, 14–15.
- Kylasamaier, K. A. Study on Madras Fowl Pest. 1931.
- Ravishankar, C.; Ravindran, R.; John, A.A.; Divakar, N.; Chandy, G.; Joshi, V.; Chaudhary, D.; Bansal, N.; Singh, R.; Sahoo, N.; et al. Author response for "Detection of Newcastle disease virus and assessment of associated relative risk in backyard and commercial poultry in Kerala, India". 2021. [CrossRef]
- Khorajiya, J.; Joshi, B.; Mathakiya, R.; Prajapati, K.; Sipai, S. Economic Impact of Genotype- Xiii Newcastle Disease Virus Infection on Commercial Vaccinated Layer Farms in India. Int. J. Livest. Res. 2018, 8, 280. [CrossRef]
- Das, M.; Kumar, S. Evidence of independent evolution of genotype XIII Newcastle disease viruses in India. Arch. Virol. 2016, 162, 997–1007. [CrossRef]
- Tirumurugaan, K.G.; Kapgate, S.; Vinupriya, M.K.; Vijayarani, K.; Kumanan, K.; Elankumaran, S. Genotypic and Pathotypic Characterization of Newcastle Disease Viruses from India. PLOS ONE 2011, 6, e28414. [CrossRef]
- Mariappan, A.K.; Munusamy, P.; Kumar, D.; Latheef, S.K.; Singh, S.D.; Singh, R.; Dhama, K. Pathological and molecular investigation of velogenic viscerotropic Newcastle disease outbreak in a vaccinated chicken flocks. VirusDisease 2018, 29, 180–191. [CrossRef]
- Reddy, N.; Patil, K.; Shah, N.; Rathod, P.; Chavda, N.; Ruparel, F.; Chhikara, M.K. Deciphering whole genome sequence of a Newcastle disease virus genotype VII. 2 isolate from a commercial poultry farm in India. Gene Rep. 2024, 34, 101884.
- Morla, S.; Shah, M.; Kaore, M.; Kurkure, N.V.; Kumar, S. Molecular characterization of genotype XIIIb Newcastle disease virus from central India during 2006–2012: Evidence of its panzootic potential. Microb. Pathog. 2016, 99, 83–86. [CrossRef]
- Roy, P.; Venugopalan, A.; Koteeswaran, A. Antigenetically unusual Newcastle disease virus from racing pigeons in India.. Trop. Anim. Heal. Prod. 2000, 32, 183–188. [CrossRef]
- Desingu, P.A.; Singh, S.D.; Dhama, K.; Karthik, K.; Kumar, O.R.V.; Malik, Y.S. Phylogenetic analysis of Newcastle disease virus isolates occurring in India during 1989–2013. VirusDisease 2016, 27, 203–206. [CrossRef]
- Jakhesara, S.J.; Prasad, V.V.S.P.; Pal, J.K.; Jhala, M.K.; Prajapati, K.S.; Joshi, C.G. Pathotypic and Sequence Characterization of Newcastle Disease Viruses from Vaccinated Chickens Reveals Circulation of Genotype II, IV and XIII and in India. Transbound. Emerg. Dis. 2014, 63, 523–539. [CrossRef]
- Khulape, S.A.; Gaikwad, S.S.; Chellappa, M.M.; Mishra, B.P.; Dey, S. Complete Genome Sequence of a Newcastle Disease Virus Isolated from Wild Peacock ( Pavo cristatus ) in India. Genome Announc. 2014, 2, e00495-14. [CrossRef]
- Gowthaman, V.; Ganesan, V.; Murthy, T.R.G.K.; Nair, S.; Yegavinti, N.; Saraswathy, P.V.; Kumar, G.S.; Udhayavel, S.; Senthilvel, K.; Subbiah, M. Molecular phylogenetics of Newcastle disease viruses isolated from vaccinated flocks during outbreaks in Southern India reveals circulation of a novel sub-genotype. Transbound. Emerg. Dis. 2018, 66, 363–372. [CrossRef]
- Gowthaman, V.; Singh, S.D.; Dhama, K.; Ramakrishnan, M.A.; Malik, Y.P.S.; Murthy, T.R.G.K.; Chitra, R.; Munir, M. Co-infection of Newcastle disease virus genotype XIII with low pathogenic avian influenza exacerbates clinical outcome of Newcastle disease in vaccinated layer poultry flocks. VirusDisease 2019, 30, 441–452. [CrossRef]
- Rajkhowa, T.K.; Zodinpuii, D.; Bhutia, L.D.; Islam, S.J.; Gogoi, A.; Hauhnar, L.; Kiran, J.; Choudhary, O.P. Emergence of a novel genotype of class II New Castle Disease virus in North Eastern States of India. Gene 2023, 864, 147315. [CrossRef]
- Maqbool, R.; Gul, I.; Wani, S.; Kashoo, Z.; Gul, N.; Islam, S.U.; Ahmad, W.; Wali, A.; Qureshi, S. Molecular Characterisation and Dynamics of the Fusion Protein of an Emerging Genotype VIIi of Newcastle Disease Virus. Agric. Res. 2024, 1–12. [CrossRef]
- Tamura K.; Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512-526.
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evol. 1985, 39, 783-791.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [CrossRef]
- Gowthaman, V.; Singh, S.D.; Dhama, K.; Desingu, P.A.; Kumar, A.; Malik, Y.S.; Munir, M. Isolation and characterization of genotype XIII Newcastle disease virus from Emu in India. VirusDisease 2016, 27, 315–318. [CrossRef]
- Desingu, P.A.; Singh, S.D.; Dhama, K.; Vinodhkumar, O.R.; Barathidasan, R.; Malik, Y.S.; Singh, R.; and Singh, R.K. Molecular characterization, isolation, pathology and pathotyping of peafowl (Pavo cristatus) origin Newcastle disease virus isolates recovered from disease outbreaks in three states of India. Avian Pathol. 2016b, 45, 674-682.
- Jakhesara, S.J.; Prasad, V, V.; Pal, J.K.; Jhala, M.K.; Prajapati, K, S.; Joshi, C, G.; Pathotypic and Sequence Characterization of Newcastle Disease Viruses from Vaccinated Chickens Reveals Circulation of Genotype II, IV and XIII and in India. Transbound. Emerg. Dis. 2016, 63, 523-39.
- Nath, B.; Kumar, S. Emerging variant of genotype XIII Newcastle disease virus from Northeast India. Acta Trop. 2017, 172, 64–69. [CrossRef]
- Gowthaman, V.; Ganesan, V.; Murthy, T.R.G.K.; Nair, S.; Yegavinti, N.; Saraswathy, P.V.; Kumar, G.S.; Udhayavel, S.; Senthilvel, K.; Subbiah, M. Molecular phylogenetics of Newcastle disease viruses isolated from vaccinated flocks during outbreaks in Southern India reveals circulation of a novel sub-genotype. Transbound. Emerg. Dis. 2018, 66, 363–372. [CrossRef]
- Deka, P.; Nath, M.K.; Das, S.; Das, B.C.; Phukan, A.; Lahkar, D.; Bora, B.; Shokeen, K.; Kumar, A.; Deka, P. A study of risk factors associated with Newcastle disease and molecular characterization of genotype XIII Newcastle disease virus in backyard and commercial poultry in Assam, India. Res. Veter- Sci. 2022, 150, 122–130. [CrossRef]
- Jakhesara, S.J.; Patel, A.K.; Malsaria, P.; Pal, J.K.; Joshi, C.G. Seroconversion Studies of Indian Newcastle Disease Virus Isolates of Genotype XIII in 3 week Old Chickens. Biosci. Biotechnol. Res. Asia 2019, 16, 27–31. [CrossRef]
- Hassanzadeh, M.; Abedi, M.; Bashashati, M.; Yousefi, A.R.; Abdoshah, M.; Mirzaie, S. Evaluation of the Newcastle disease virus genotype VII–mismatched vaccines in SPF chickens: A challenge efficacy study. Veter- Anim. Sci. 2024, 24, 100348. [CrossRef]
- Geetha, M.; Malmarugan, S.; Dinakaran, A.M.; Sharma, V.K.; Mishra, R.K.; Jagadeeswaran, D. Seroprevalence of New Castle disease, infectious Bursal disease and egg drop syndrome 76 in ducks. Ind. J. Vet. Anim. Res. 2008, 4, 200–202.
- Vegad, J. Drift variants of low pathogenic avian influenza virus: observations from India. World's Poult. Sci. J. 2014, 70, 767–774. [CrossRef]
- Kolluri, G. and Tyagi, J.S. Biosecurity and vaccination approaches in commercial (layers, broilers and broiler breeders) and backyard Poultry stocks. In: Tyagi, J.S. Gautham K. , Gopi, M., and Rokade, J.J., Eds.; 2019. Capacity Building of Field Functionaries on Diversified Poultry Production and Processing Technology under Off-Campus Collaborative Training Programme for Field Functionaries with MANAGE, Hyderabad ed.; Tyagi, J.S., G Kolluri., Gopi, M. and Rokade, J.J., Eds.; ICAR-CARI, Bareilly, India. 2019: Volume 1, pp. 114-133.
- Dey, S.; Chellappa, M.M.; Gaikwad, S.; Kataria, J.M.; Vakharia, V.N. Genotype Characterization of Commonly Used Newcastle Disease Virus Vaccine Strains of India. PLOS ONE 2014, 9, e98869. [CrossRef]
- Iyer, G.S.; Hashmi, Z.A. Studies on Newcastle (Ranikhet) disease virus strain differences in amenability to attenuations. Indian J. Vet. Sci. 1945, 15, 155–157.
- Sharma, R.; Sandeep, S.; Sanjay, K. Estimated Incremental Benefits of Complete Vaccination Against Newcastle Disease in Layers”. Acta. Sci. Vet. Sci. 2023, 5, 55-57.
- Hu, Z.; Hu, S.; Meng, C.; Wang, X.; Zhu, J.; Liu, X. Generation of a Genotype VII Newcastle Disease Virus Vaccine Candidate with High Yield in Embryonated Chicken Eggs. Avian Dis. 2011, 55, 391–397. [CrossRef]
- Roohani, K.; Tan, S.W.; Yeap, S.K.; Ideris, A.; Bejo, M.H.; Omar, A.R. Characterisation of genotype VII Newcastle disease virus (NDV) isolated from NDV vaccinated chickens, and the efficacy of LaSota and recombinant genotype VII vaccines against challenge with velogenic NDV. J. Veter- Sci. 2015, 16, 447–457. [CrossRef]
- Dewidar, A.A.A.; Kilany, W.H.; El-Sawah, A.A.; Shany, S.A.S.; Dahshan, A.-H.M.; Hisham, I.; Elkady, M.F.; Ali, A. Genotype VII.1.1-Based Newcastle Disease Virus Vaccines Afford Better Protection against Field Isolates in Commercial Broiler Chickens. Animals 2022, 12, 1696. [CrossRef]
- Sultan, H.A.; Elfeil, W.K.; Nour, A.A.; Tantawy, L.; Kamel, E.G.; Eed, E.M.; El Askary, A.; Talaat, S. Efficacy of the Newcastle Disease Virus Genotype VII.1.1-Matched Vaccines in Commercial Broilers. Vaccines 2021, 10, 29. [CrossRef]
- Cho, S.-H.; Kwon, H.-J.; Kim, T.-E.; Kim, J.-H.; Yoo, H.-S.; Park, M.-H.; Park, Y.-H.; Kim, S.-J. Characterization of a Recombinant Newcastle Disease Virus Vaccine Strain. Clin. Vaccine Immunol. 2008, 15, 1572–1579. [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of Viral Shedding Following Vaccination With Inactivated and Live Newcastle Disease Vaccines Formulated With Wild-Type and Recombinant Viruses. Avian Dis. 2009, 53, 39–49. [CrossRef]
- Xiao, S.; Nayak, B.; Samuel, A.; Paldurai, A.; Kanabagattebasavarajappa, M.; Prajitno, T.Y.; Bharoto, E.E.; Collins, P.L.; Samal, S.K. Generation by Reverse Genetics of an Effective, Stable, Live-Attenuated Newcastle Disease Virus Vaccine Based on a Currently Circulating, Highly Virulent Indonesian Strain. PLOS ONE 2012, 7, e52751. [CrossRef]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136.
- Sultan, H.A.; Talaat, S.; Elfeil, W.K.; Selim, K.; Kutkat, M.A.; Amer, S.A.; Choi, K.-S. Protective efficacy of the Newcastle disease virus genotype VII–matched vaccine in commercial layers. Poult. Sci. 2020, 99, 1275–1286. [CrossRef]
- Fawzy, M.; Ali, R.R.; Elfeil, W.K.; Saleh, A.A.; El-Tarabilli, M.M.A. Efficacy of inactivated velogenic Newcastle disease virus genotype VII vaccine in broiler chickens. 2020, 11, 113–120. [CrossRef]
- Wang, N.; Huang, M.; Fung, T.S.; Luo, Q.; Ye, J.X.; Du, Q.R.; Wen, L.H.; Liu, D.X.; Chen, R.A. Rapid Development of an Effective Newcastle Disease Virus Vaccine Candidate by Attenuation of a Genotype VII Velogenic Isolate Using a Simple Infectious Cloning System. Front. Veter- Sci. 2020, 7. [CrossRef]
- Kumar, B.A.; Panickan, S.; Bindu, S.; Kumar, V.; Ramakrishnan, S.; Saxena, S.; Shrivastava, S.; Dandapat, S. Immunogenicity and protective efficacy of an inactivated Newcastle disease virus vaccine encapsulated in poly-(lactic-co-glycolic acid) nanoparticles. Poult. Sci. 2023, 102, 102679. [CrossRef]
- Lee, C.-W.; Bakre, A.; Olivier, T.L.; Alvarez-Narvaez, S.; Harrell, T.L.; Conrad, S.J. Toll-like Receptor Ligands Enhance Vaccine Efficacy against a Virulent Newcastle Disease Virus Challenge in Chickens. Pathogens 2023, 12, 1230. [CrossRef]
- Hongzhuan, Z.; Ying, T.; Xia, S.; Jinsong, G.; Zhenhua, Z.; Beiyu, J.; Yanyan, C.; Lulu, L.; Jue, Z.; Bing, Y.; et al. Preparation of the inactivated Newcastle disease vaccine by plasma activated water and evaluation of its protection efficacy. Appl. Microbiol. Biotechnol. 2019, 104, 107–117. [CrossRef]
- Baylor, N.W.; Egan, W.; Richman, P. Aluminum salts in vaccines—US perspective. Vaccine. 2002, 20, S18-S23.
- Moni, S.S.; Abdelwahab, S.I.; Jabeen, A.; Elmobark, M.E.; Aqaili, D.; Gohal, G.; Oraibi, B.; Farasani, A.M.; Jerah, A.A.; Alnajai, M.M.A.; et al. Advancements in Vaccine Adjuvants: The Journey from Alum to Nano Formulations. Vaccines 2023, 11, 1704. [CrossRef]
- Liu, J.; Guo, S.; Jin, Z.; Zhao, K. Adjuvanted quaternized chitosan composite aluminum nanoparticles-based vaccine formulation promotes immune responses in chickens. Vaccine 2023, 41, 2982–2989. [CrossRef]
- Palese, P.; Zheng, H.; Engelhardt, O.G.; Pleschka, S.; García-Sastre, A. Negative-strand RNA viruses: genetic engineering and applications.. Proc. Natl. Acad. Sci. 1996, 93, 11354–11358. [CrossRef]
- Peeters, B.P.H.; de Leeuw, O.S.; Koch, G.; Gielkens, A.L.J. Rescue of Newcastle Disease Virus from Cloned cDNA: Evidence that Cleavability of the Fusion Protein Is a Major Determinant for Virulence. J. Virol. 1999, 73, 5001–5009. [CrossRef]
- Ravikumar, R.; Chan, J.; Prabakaran, M. Vaccines against Major Poultry Viral Diseases: Strategies to Improve the Breadth and Protective Efficacy. Viruses 2022, 14, 1195. [CrossRef]
- Veits, J.; Wiesner, D.; Fuchs, W.; Hoffmann, B.; Granzow, H.; Starick, E.; Mundt, E.; Schirrmeier, H.; Mebatsion, T.; Mettenleiter, T.C.; et al. Newcastle disease virus expressing H5 hemagglutinin gene protects chickens against Newcastle disease and avian influenza. Proc. Natl. Acad. Sci. 2006, 103, 8197–8202. [CrossRef]
- Dey, S.; Chellappa, M.M.; Pathak, D.C.; Gaikwad, S.; Yadav, K.; Ramakrishnan, S.; Vakharia, V.N. Newcastle Disease Virus Vectored Bivalent Vaccine against Virulent Infectious Bursal Disease and Newcastle Disease of Chickens. Vaccines 2017, 5, 31. [CrossRef]
- Basavarajappa, M.K.; Kumar, S.; Khattar, S.K.; Gebreluul, G.T.; Paldurai, A.; Samal, S.K. A recombinant Newcastle disease virus (NDV) expressing infectious laryngotracheitis virus (ILTV) surface glycoprotein D protects against highly virulent ILTV and NDV challenges in chickens. Vaccine 2014, 32, 3555–3563. [CrossRef]
- Saikia, D.P.; Yadav, K.; Pathak, D.C.; Ramamurthy, N.; D’silva, A.L.; Marriappan, A.K.; Ramakrishnan, S.; Vakharia, V.N.; Chellappa, M.M.; Dey, S. Recombinant Newcastle Disease Virus (NDV) Expressing Sigma C Protein of Avian Reovirus (ARV) Protects against Both ARV and NDV in Chickens. Pathogens 2019, 8, 145. [CrossRef]
- Shirvani, E.; Paldurai, A.; Manoharan, V.K.; Varghese, B.P.; Samal, S.K. A Recombinant Newcastle Disease Virus (NDV) Expressing S Protein of Infectious Bronchitis Virus (IBV) Protects Chickens against IBV and NDV. Sci. Rep. 2018, 8, 11951, Correction in Sci. Rep. 2020, 10, 762. [CrossRef]
- Chellappa, M.M.; Dey, S.; Pathak, D.C.; Singh, A.; Ramamurthy, N.; Ramakrishnan, S.; Mariappan, A.K.; Dhama, K.; Vakharia, V.N. Newcastle Disease Virus Vectored Chicken Infectious Anaemia Vaccine Induces Robust Immune Response in Chickens. Viruses 2021, 13, 1985. [CrossRef]
- Palya, V.; Kiss, I.; Tatar-Kis, T.; Mato, T.; Felfoldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: recombinant HVT NDV provides high clinical protection and reduction challenge virus shedding, with the absence of vaccine reaction. Avian Dis. 2012, 56, 282–287.
- Lee, K.; A Mackley, V.; Rao, A.; Chong, A.T.; A Dewitt, M.; E Corn, J.; Murthy, N.; Inc, G.; Berkeley; States, U. Synthetically modified guide RNA and donor DNA are a versatile platform for CRISPR-Cas9 engineering. eLife 2017, 6. [CrossRef]
- Véron, N.; Qu, Z.; Kipen, P.A.; Hirst, C.E.; Marcelle, C. CRISPR mediated somatic cell genome engineering in the chicken. Dev. Biol. 2015, 407, 68–74. [CrossRef]
- Oishi, I.; Yoshii, K.; Miyahara, D.; Kagami, H.; Tagami, T. Targeted mutagenesis in chicken using CRISPR/Cas9 system. Sci. Rep. 2016, 6, 23980–23980. [CrossRef]
- Tan, J.W.; Joshi, P. Egg allergy: An update. J. Paediatr. Child Heal. 2013, 50, 11–15. [CrossRef]
- Kim, G.D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. 2020, 34, 5688-5696.
- Idoko-Akoh, A.; Goldhill, D.H.; Sheppard, C.M.; Bialy, D.; Quantrill, J.L.; Sukhova, K.; Brown, J.C.; Richardson, S.; Campbell, C.; Taylor, L.; et al. Creating resistance to avian influenza infection through genome editing of the ANP32 gene family. Nat. Commun. 2023, 14, 1–15. [CrossRef]
- Naeem, M.; Alkhodairy, H.F.; Ashraf, I.; Khalil, A.B. CRISPR/Cas System Toward the Development of Next-Generation Recombinant Vaccines: Current Scenario and Future Prospects. Arab. J. Sci. Eng. 2022, 48, 1–11. [CrossRef]
- Vilela, J.; Rohaim, M.A.; Munir, M. Application of CRISPR/Cas9 in Understanding Avian Viruses and Developing Poultry Vaccines. Front. Cell. Infect. Microbiol. 2020, 10. [CrossRef]
- Yao, Y.; Bassett, A; Nair, V. Targeted editing of avian herpesvirus vaccine vector using CRISPR/Cas9 nucleases. Vaccine Technol. 2016, 1,1-7.
- Tang, N.; Zhang, Y.; Pedrera, M.; Chang, P.; Baigent, S.; Moffat, K.; Shen, Z.; Nair, V.; Yao, Y. A simple and rapid approach to develop recombinant avian herpesvirus vectored vaccines using CRISPR/Cas9 system. Vaccine 2017, 36, 716–722. [CrossRef]
- Tang, N.; Zhang, Y.; Sadigh, Y.; Moffat, K.; Shen, Z.; Nair, V.; Yao, Y. Generation of A Triple Insert Live Avian Herpesvirus Vectored Vaccine Using CRISPR/Cas9-Based Gene Editing. Vaccines 2020, 8, 97. [CrossRef]
- Dronina, J.; Samukaite-Bubniene, U.; Ramanavicius, A. Towards application of CRISPR-Cas12a in the design of modern viral DNA detection tools (Review). J. Nanobiotechnology 2022, 20, 1–15. [CrossRef]
- Atasoy, M.O.; Rohaim, M.A.; Munir, M. Simultaneous Deletion of Virulence Factors and Insertion of Antigens into the Infectious Laryngotracheitis Virus Using NHEJ-CRISPR/Cas9 and Cre–Lox System for Construction of a Stable Vaccine Vector. Vaccines 2019, 7, 207. [CrossRef]
Figure 1.
Velogenic Newcastle Disease: Typical clinical signs and post-mortem lesions in vaccinated flocks of Rhode Island Red and White Leghorn grower chickens.
Figure 1.
Velogenic Newcastle Disease: Typical clinical signs and post-mortem lesions in vaccinated flocks of Rhode Island Red and White Leghorn grower chickens.
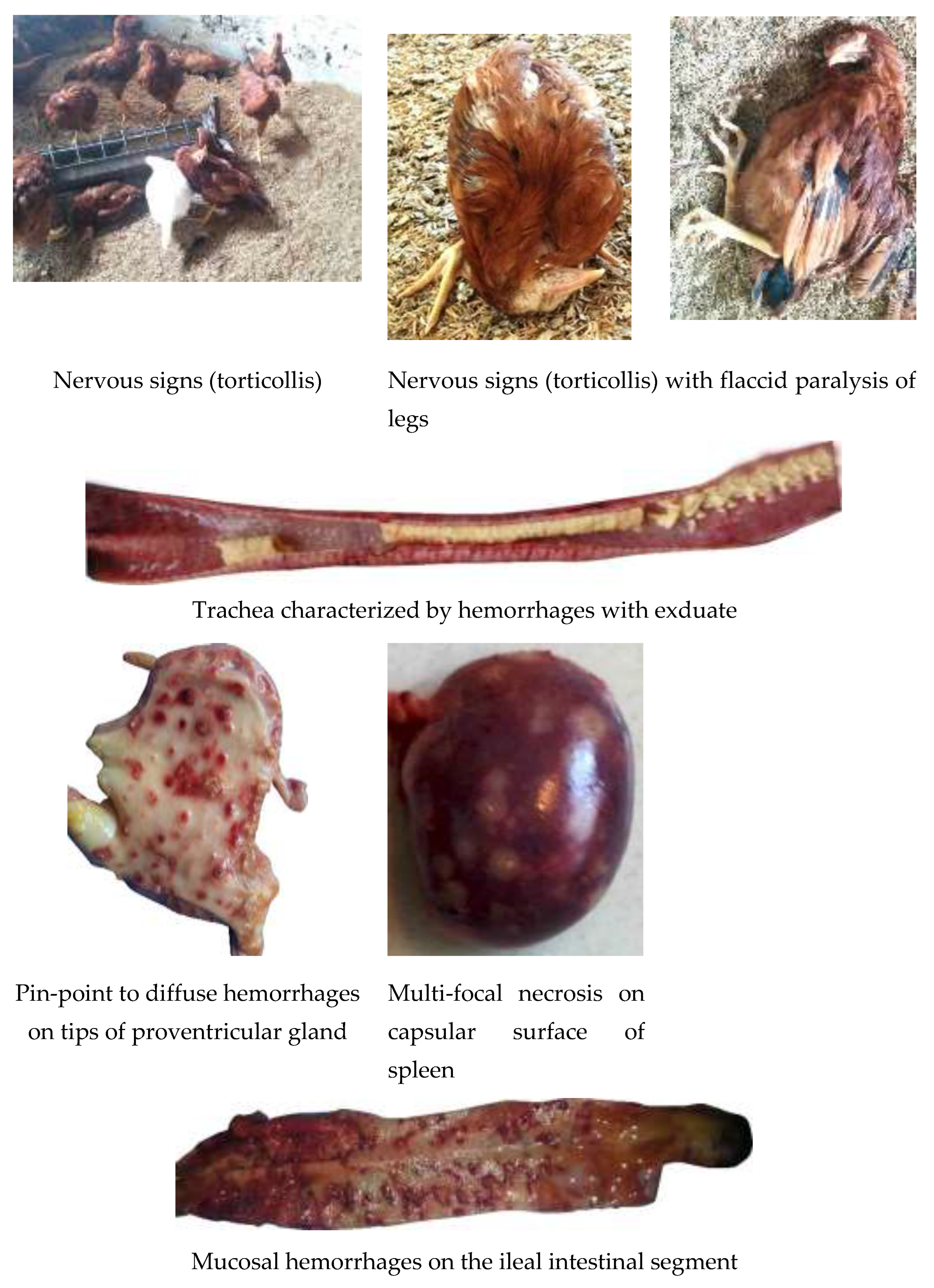
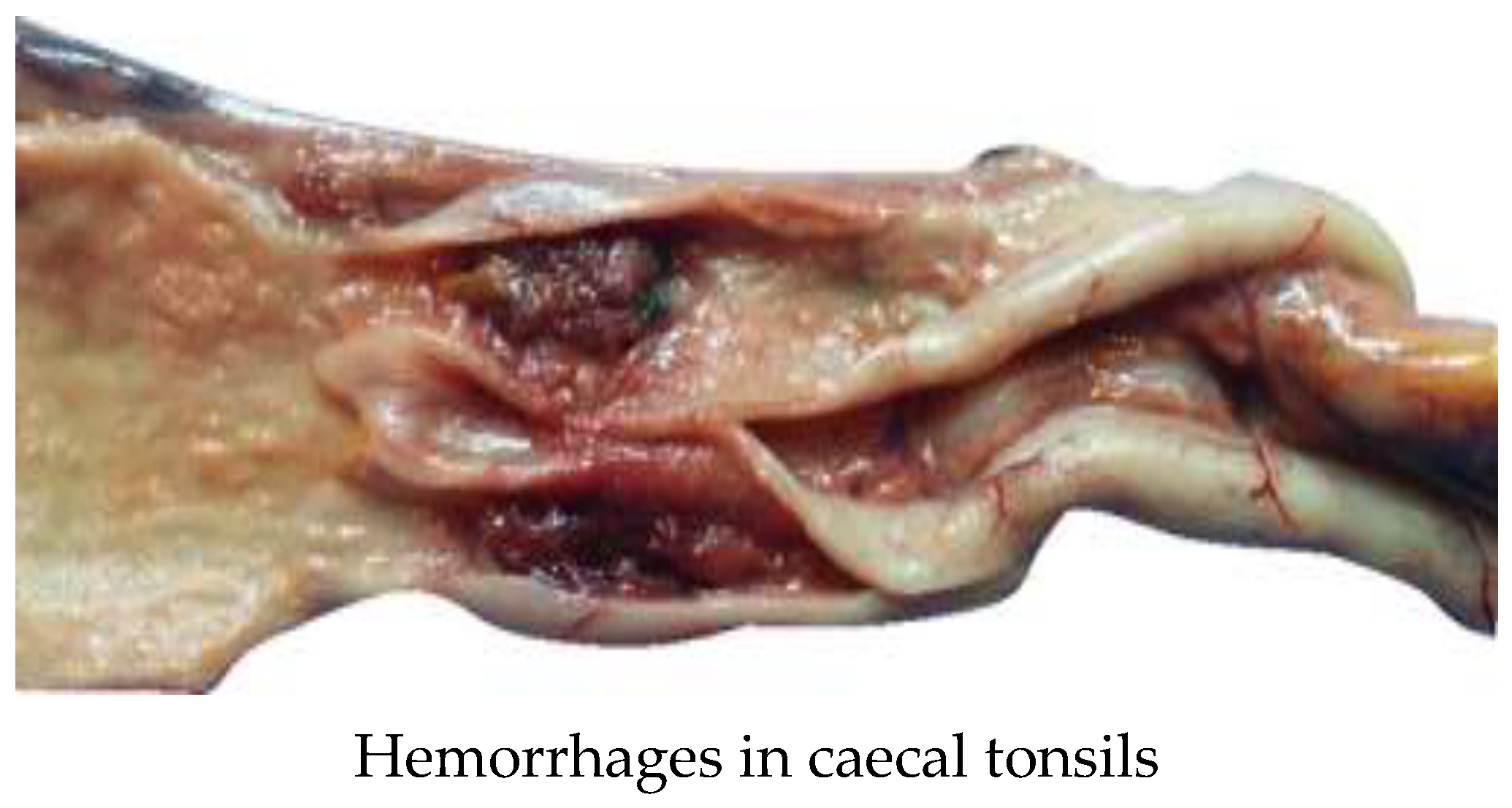
Figure 2.
The NDV outbreaks (in numbers) across the countries during 2005-2024 [12].
Figure 2.
The NDV outbreaks (in numbers) across the countries during 2005-2024 [12].
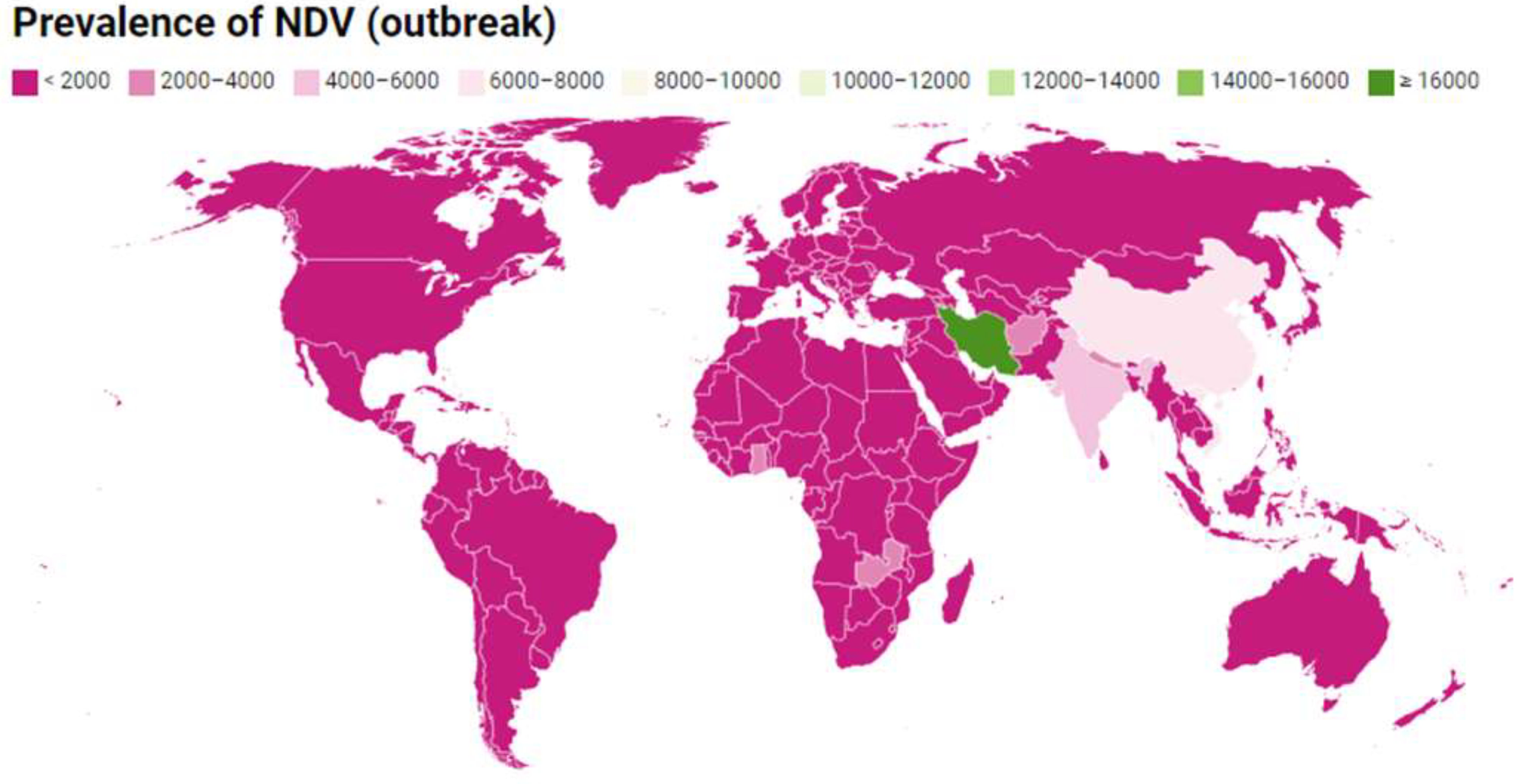
Figure 3.
Number of publications (year wise) reporting the Genotypes VII and XIII in poultry across the globe (Source: PubMed, 2024 (https://pubmed.ncbi.nlm.nih.gov/) Search strings: Search query: Newcastle disease genotypes VII in poultry and Search query: Newcastle disease genotypes XIII in poultry.
Figure 3.
Number of publications (year wise) reporting the Genotypes VII and XIII in poultry across the globe (Source: PubMed, 2024 (https://pubmed.ncbi.nlm.nih.gov/) Search strings: Search query: Newcastle disease genotypes VII in poultry and Search query: Newcastle disease genotypes XIII in poultry.
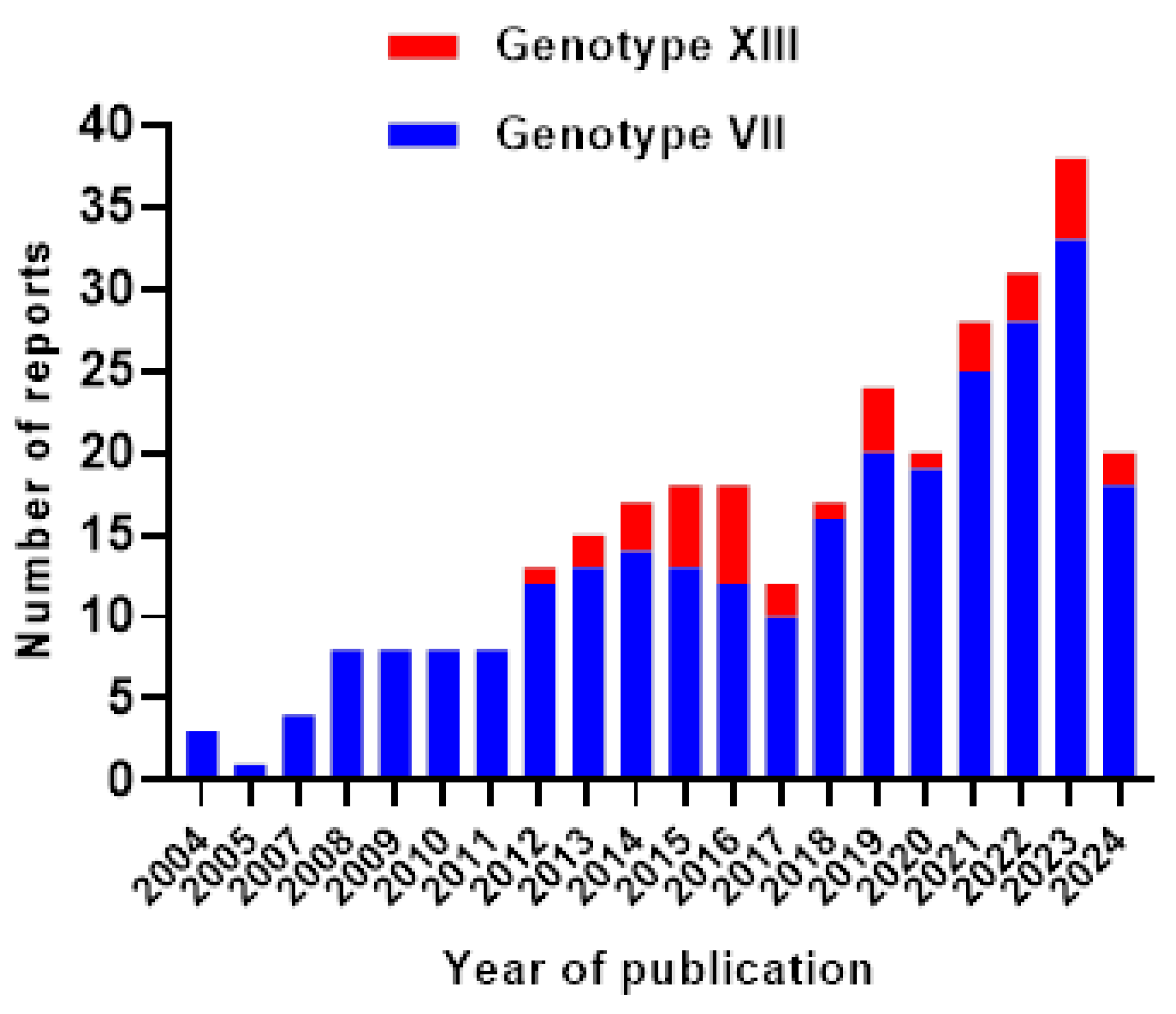
Figure 4.
Year wise velogenic NDV cases reported and deaths occurred in India during different years.
Figure 4.
Year wise velogenic NDV cases reported and deaths occurred in India during different years.
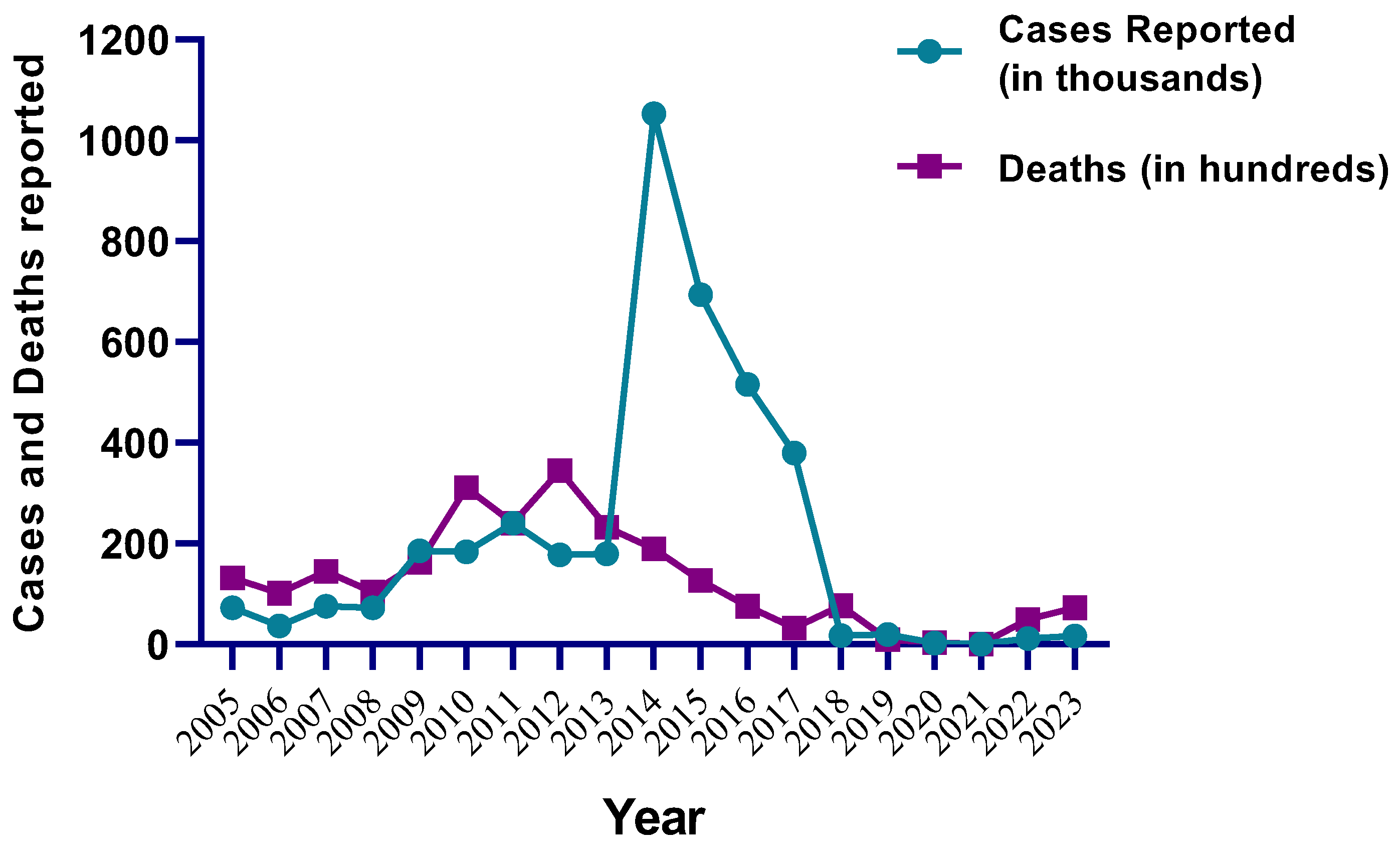
Figure 5.
Infection pressure of NDV in different regions of India as indicated by number of outbreaks. Light intensity colour indicates lower number.
Figure 5.
Infection pressure of NDV in different regions of India as indicated by number of outbreaks. Light intensity colour indicates lower number.
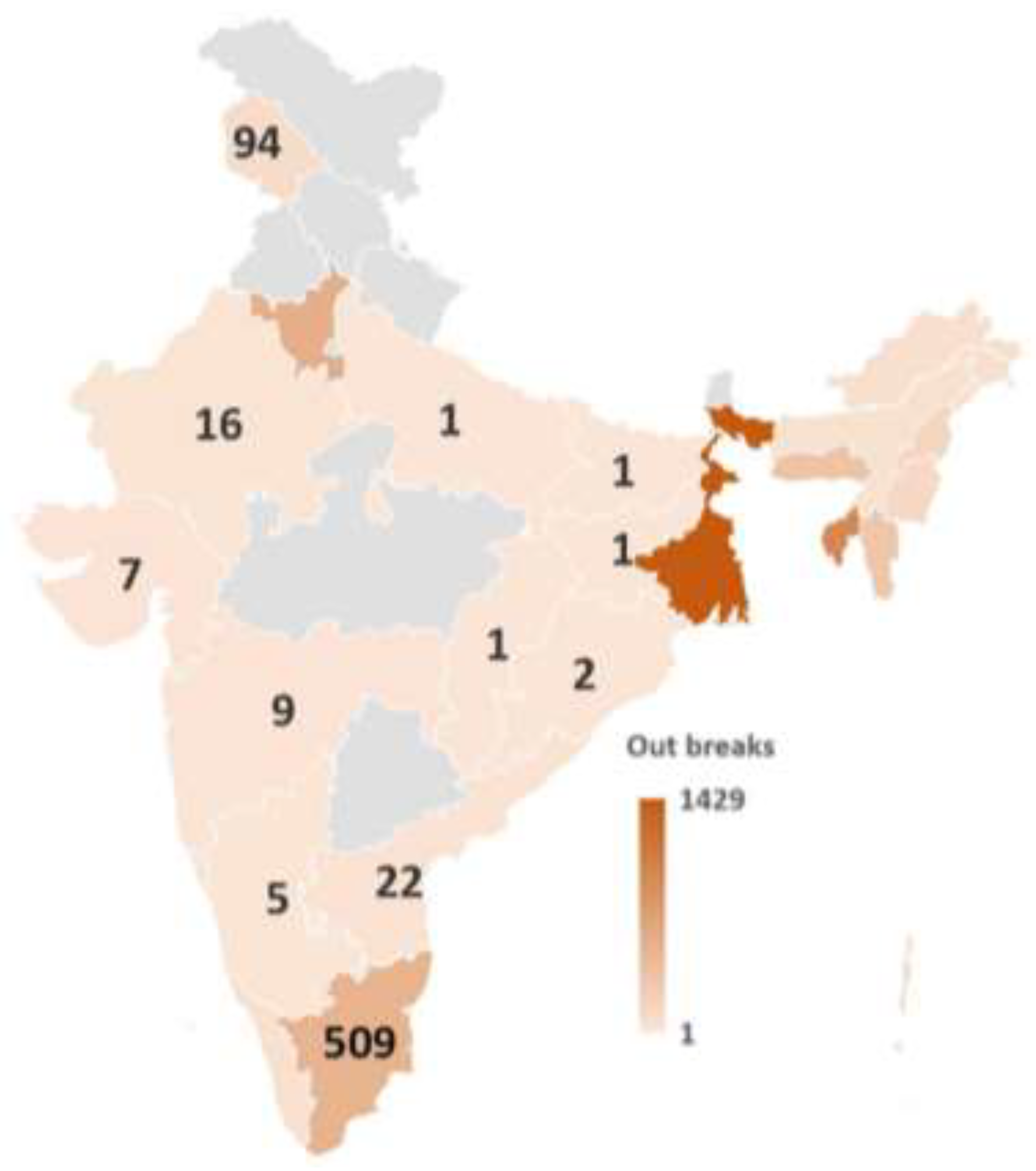
Figure 6.
(a) Phylogenetic tree (traditional/branch style) illustrating the evolutionary and distance relationships between various Newcastle Disease Virus (NDV) strains and genotypes circulating in India. The tree is constructed based on nucleotide sequences from previous reports and shows the distinct clustering of strains according to geographic regions. (b) The phylogenetic analysis (Circle tree/branch style) of Genotype VII and XIII NDV strains-based nucleotide sequences of F gene in the highly variable region including F gene cleavage site.
Figure 6.
(a) Phylogenetic tree (traditional/branch style) illustrating the evolutionary and distance relationships between various Newcastle Disease Virus (NDV) strains and genotypes circulating in India. The tree is constructed based on nucleotide sequences from previous reports and shows the distinct clustering of strains according to geographic regions. (b) The phylogenetic analysis (Circle tree/branch style) of Genotype VII and XIII NDV strains-based nucleotide sequences of F gene in the highly variable region including F gene cleavage site.
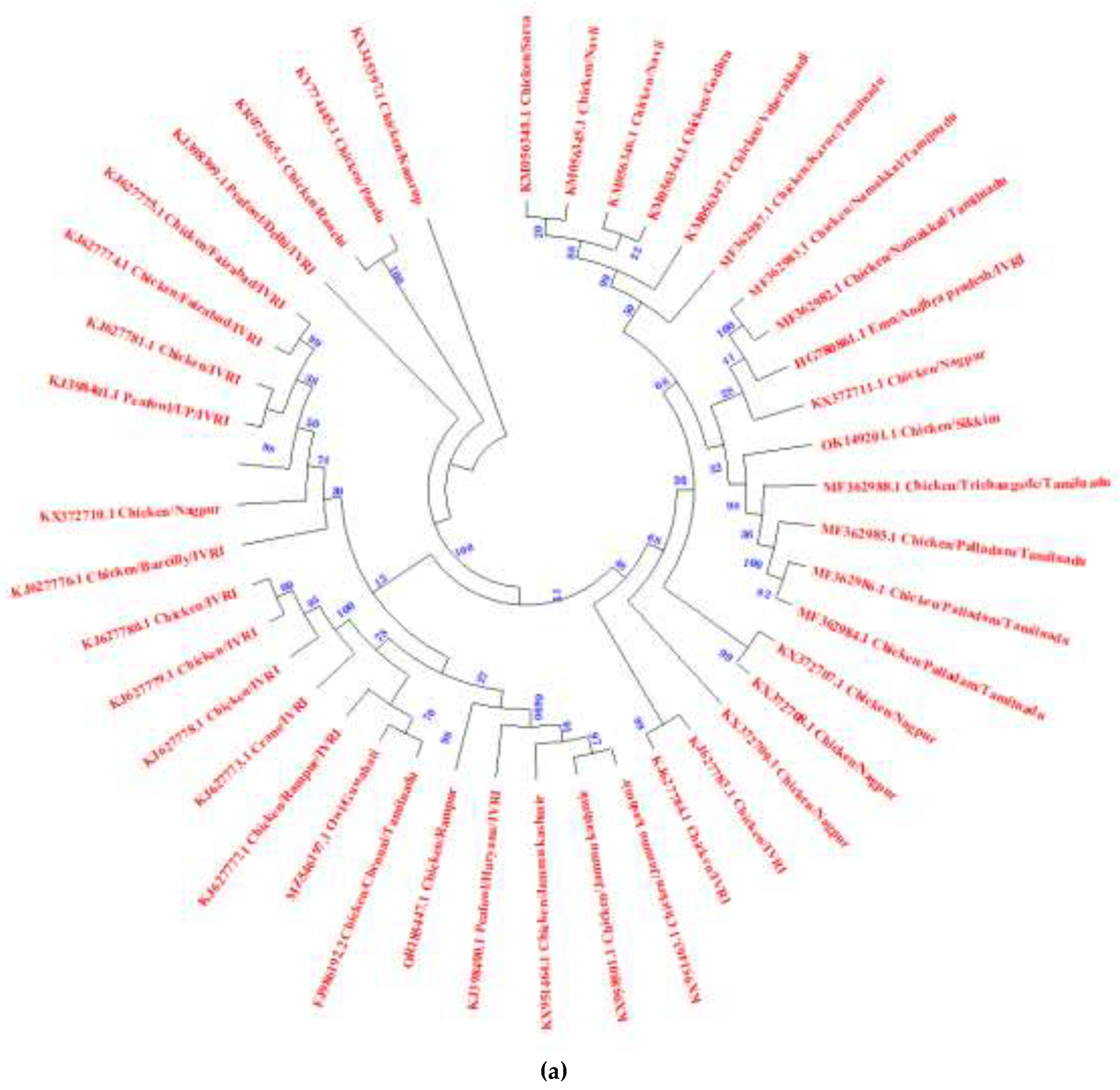
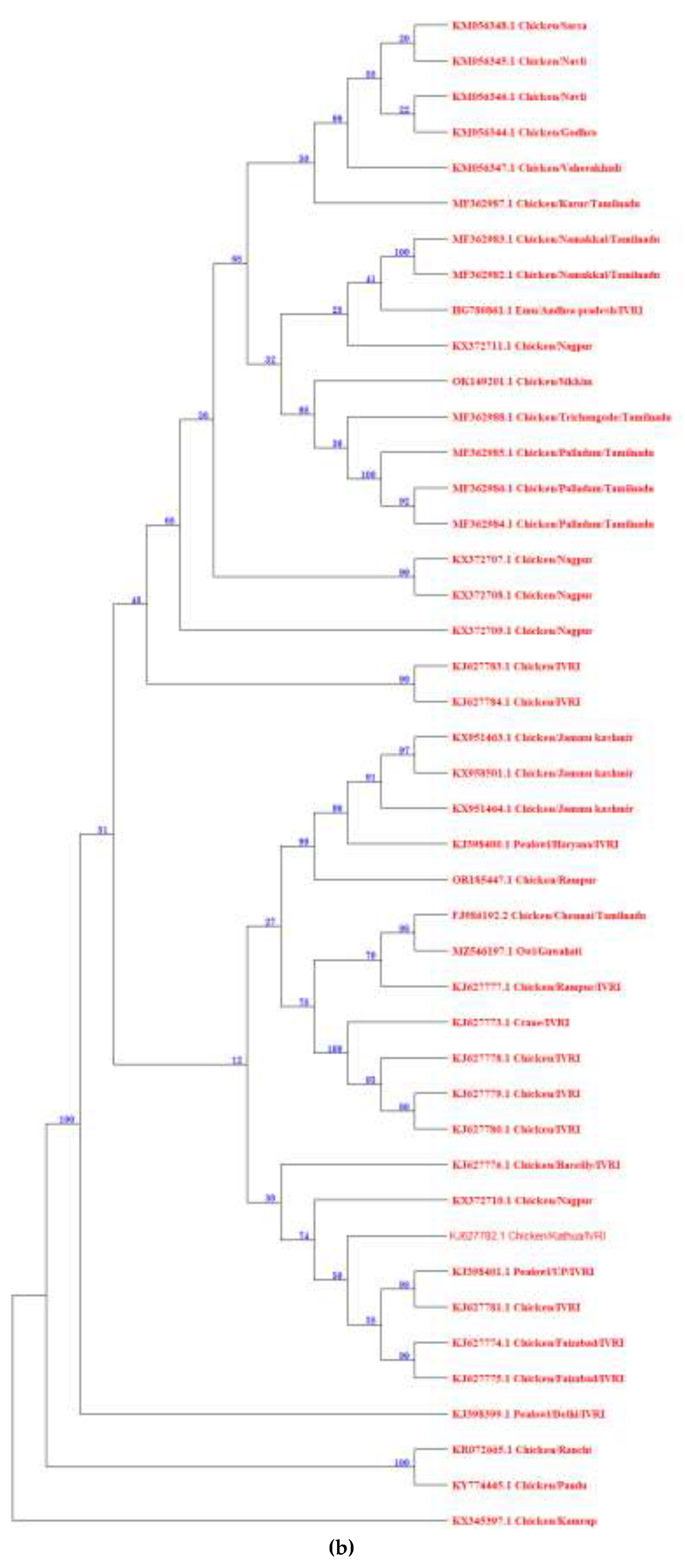
Figure 7.
Different vaccine strains of NDV being used in poultry and their residual virulence from 0 in asymptomatic enteric to 1.4 in mesogenic strains.
Figure 7.
Different vaccine strains of NDV being used in poultry and their residual virulence from 0 in asymptomatic enteric to 1.4 in mesogenic strains.
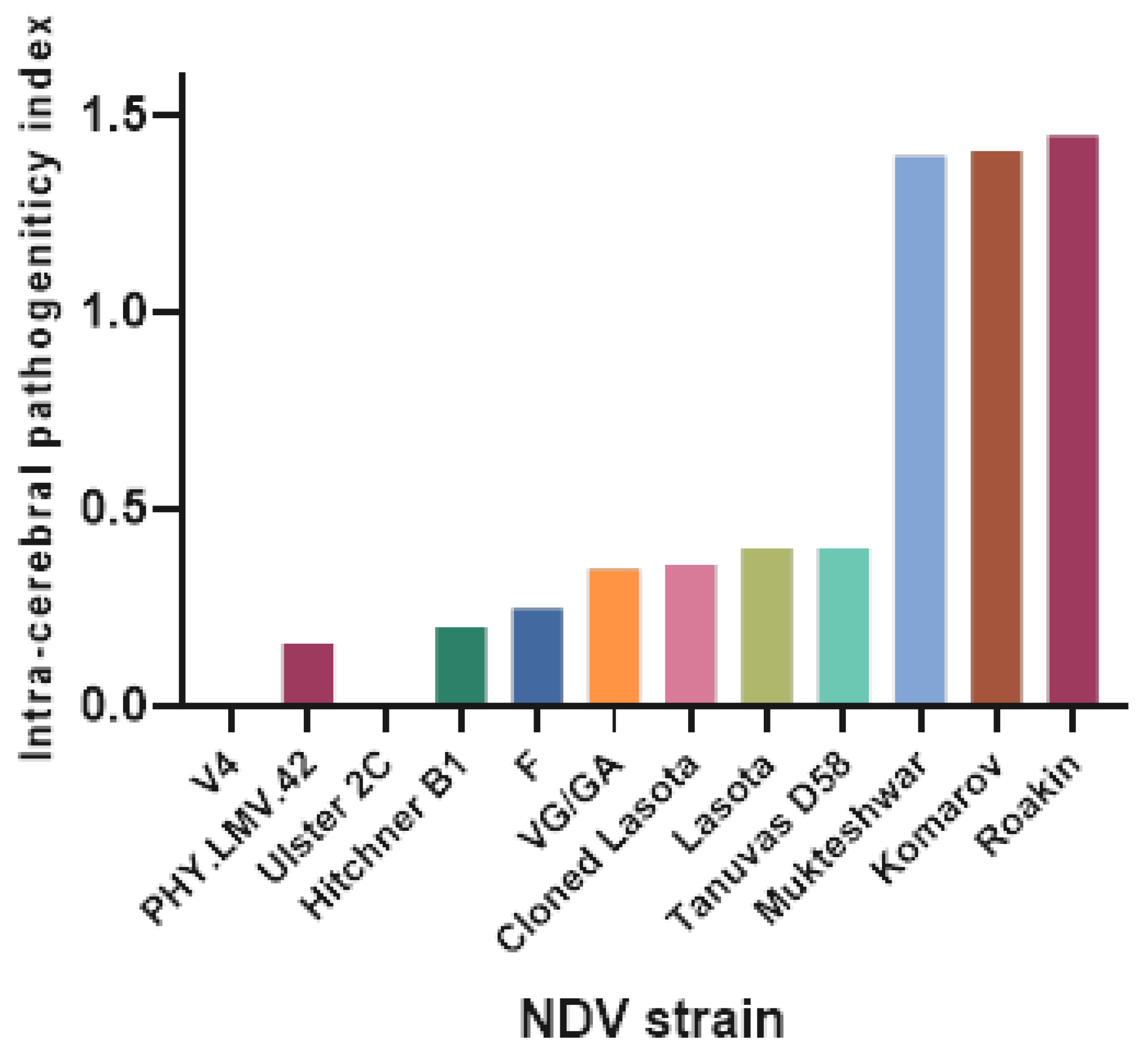
Figure 8.
The makeup of ND vaccines being used in the poultry industry.
Figure 9.
Year wise deployment of ND vaccine (R2B/Mukteshwar strain of Genotype-II) doses and egg production trend in India (Calculated based on FAOSTAT, 2024 database).
Figure 9.
Year wise deployment of ND vaccine (R2B/Mukteshwar strain of Genotype-II) doses and egg production trend in India (Calculated based on FAOSTAT, 2024 database).
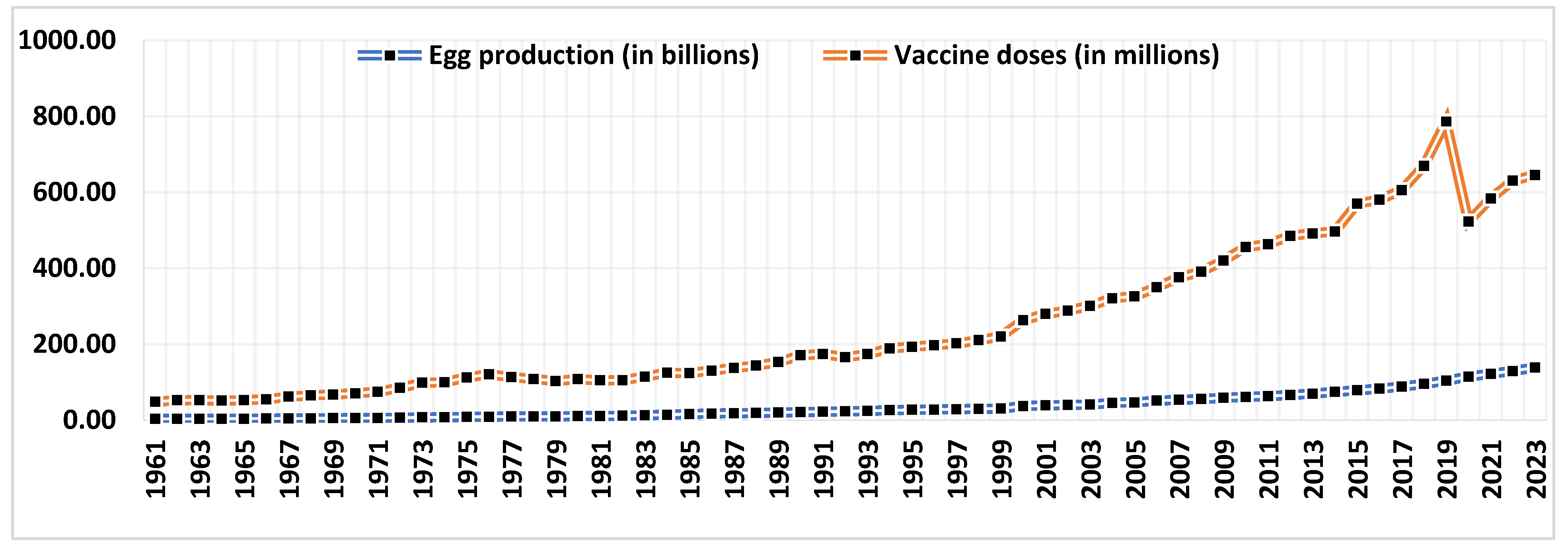
Figure 10.
Trends in ND vaccination with Mukteshwar strain and NDV cases reported (Calculated based on FAOSTAT, 2024 database).
Figure 10.
Trends in ND vaccination with Mukteshwar strain and NDV cases reported (Calculated based on FAOSTAT, 2024 database).
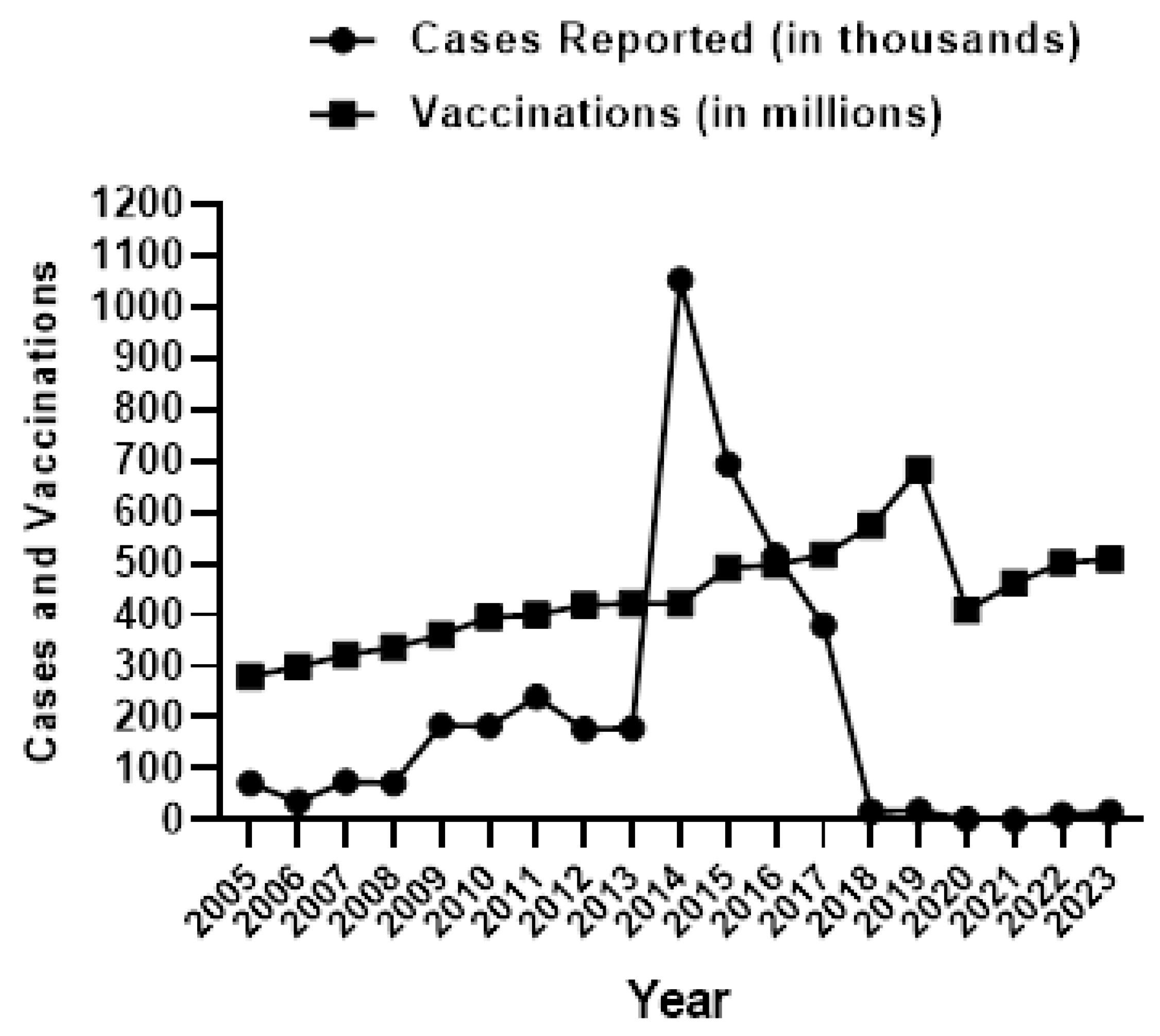
Figure 11.
Protein sequences of different genotypes of Newcastle disease isolated in India indicating differentiation of Fusion protein clevage site (FPCS) (1-9).
Figure 11.
Protein sequences of different genotypes of Newcastle disease isolated in India indicating differentiation of Fusion protein clevage site (FPCS) (1-9).

Table 4.
Consensus vaccination regime in birds of varied production potentials in India [84].
Table 4.
Consensus vaccination regime in birds of varied production potentials in India [84].
| Type of bird | Vaccination regime |
|---|---|
| Commercial layers (BV300, IB) | 5-6 days ND Lasota, + ND Killed, 11 wk R2B/RDVK, 16 wk ND Killed |
| Commercial broilers (Cobb 430Y, Ross) | DOH: ND Killed, 5-6 days ND Lasota, 10 days ND Killed, 28 days ND Live Booster |
| Dual-purpose and coloured birds | 0 day: ND D58, 21 days: D58, 56 days: R2B, 14 wk: D58 clone, 16 wk: ND Killed |
Table 5.
Commercially available vaccine strains (Genotype II) in different formats in India.
| Vaccine strains | Combination (if any) | |
|---|---|---|
| Live | Killed | |
| F/B1/Lasota/D58/ D26+FC126/Nobilis ND clone 30/VH clone/R2B/CH-80 cloned/CL-79 | Lasota/VH | ND+IB ND+IB+IBD |
Table 6.
Identities of amino acid sequences at different regions of NDV sequence between vaccine genotypes (G-II) and velogenic genotypes (G-VII, G-XIII).
Table 6.
Identities of amino acid sequences at different regions of NDV sequence between vaccine genotypes (G-II) and velogenic genotypes (G-VII, G-XIII).
| Strains | F1 region | Fusion protein clevage site | F2 region | |||||||||||||||||||||||||||||||
| 4 | 8 | 9 | 13 | 16 | 20 | 22 | 29 | 30 | 82 | 112 | 113 | 114 | 115 | 116 | 117 | 121 | 124 | 145 | 192 | 195 | 203 | 231 | 232 | 272 | 288 | 396 | 421 | 482 | 552 | 553 | ||||
| AEL75045.1 (LaSota) | R | K | N | M | T | A | V | A | N | D | G | R | Q | G | R | L | I | G | K | K | Q | A | N | K | N | T | M | K | E | K | M | |||
| AHN09749.1 (F) | R | K | N | M | T | A | V | A | N | D | G | R | Q | G | R | L | I | G | K | K | Q | A | N | K | N | T | M | K | E | K | M | |||
| AFX98109.1 (R2B) | R | K | N | M | T | A | V | A | N | D | R | R | Q | K | R | F | I | G | K | K | Q | A | N | K | N | T | M | K | E | K | M | |||
| UQW17861.1 (Genotype 7) | K | R | I | L | I | M | I | T | S | E | K | R | Q | K | R | F | V | S | N | N | R | T | T | Q | Y | N | I | R | A | R | A | |||
| AHG26218.1 (Genotype VII) | K | R | I | L | I | M | I | T | S | E | R | R | Q | K | R | F | V | S | N | N | R | T | T | Q | Y | N | I | R | A | R | A | |||
| ATU83336.1 (Genotype XIII) | K | R | I | L | I | M | I | T | S | E | R | R | Q | K | R | F | V | S | N | N | R | T | T | Q | Y | N | I | R | A | R | A | |||
| AIQ78308.1 (Genotype XIII) | K | R | I | L | I | M | I | T | S | E | R | R | Q | K | R | F | V | S | N | N | R | T | T | Q | Y | N | I | R | A | R | A | |||
Table 7.
Percentage identity between genotypes of vaccine and velogenic NDV strains in India based on nucleotide sequences.
Table 7.
Percentage identity between genotypes of vaccine and velogenic NDV strains in India based on nucleotide sequences.
| Strain | JF950510.1_Lasota | KC987036.1_F | JX316216.1_R2B |
|---|---|---|---|
| JF950510.1_Lasota | - | 96.07 | 93.616 |
| KC987036.1_F | 96.07 | - | 93.04 |
| JX316216.1_R2b | 93.62 | 93.04 | - |
| KF740478.1_ Genotype VII | 84.58 | 84.05 | 84.73 |
| MZ546197.1_Genotype VII | 83.05 | 84.96 | 82.69 |
| KY774445.1_Genotype XIII | 83.22 | 82.88 | 83.81 |
| KM056347.1_Genotype XIII | 83.20 | 85.43 | 83.56 |
Table 8.
Percentage identity between genotypes of vaccine and velogenic NDV strains in India based on amino acid sequences.
Table 8.
Percentage identity between genotypes of vaccine and velogenic NDV strains in India based on amino acid sequences.
| F gene | AEL75045.1_Lasota | AHN09749.1 _F | AFX98109.1_ R2B |
|---|---|---|---|
| AEL75045.1 _Lasota | - | 98.37 | 94.03 |
| AHN09749.1_F | 98.37 | - | 94.94 |
| AFX98109.1_R2B | 94.03 | 94.94 | - |
| AHG26218.1_Genotype VII | 89.67 | 89.67 | 89.86 |
| UQW17861.1_Genotype VII | 88.77 | 88.77 | 88.77 |
| ATU83336.1 _Genotype XIII | 88.77 | 88.59 | 88.95 |
| AIQ78308.1_Genotype XIII | 88.77 | 88.77 | 88.95 |
| HN gene | AEL75044.1_ Lasota | AHN09747.1_ F | AFX98110.1_ R2B |
| AEL75044.1_Lasota | - | 96.19 | 92.55 |
| AHN09747.1_F | 96.19 | - | 90.99 |
| AFX98110.1_R2B | 92.55 | 90.99 | - |
| AHG26219.1_Genotype VII | 86.97 | 86.27 | 88.01 |
| UQW17859.1_Genotype VII | 87.52 | 86.29 | 88.05 |
| ATU83337.1_Genotype XIII | 88.25 | 87.19 | 87.72 |
| AIQ78309.1_Genotype XIII | 85.92 | 85.04 | 86.60 |
Table 9.
Overview of traditional and newer genotype-based vaccines for control of NDV in poultry.
| Features | Vaccine strains | |
|---|---|---|
| Traditional genotypes | Newer genotypes | |
| GI, GII across the globe | GV (America), GVI (Pigeons), GVII (Asia and Africa) | |
| Protection from morbidity & mortality |
++ |
+++ |
| Reduction in virus shedding | + | +++ |
| Field virus silent dissemination & spill over | +++ | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



