
Submitted:
23 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Pigeon paramyxovirus serotype 1 (PPMV-1), primarily originating from racing pigeons, has become a global panzootic, spreading to poultry and wild birds, thus posing a threat to the poultry industry. Egypt uses both inactivated pigeon paramyxovirus (PPMV-1) and conventional Newcastle disease virus (NDV) vaccines to protect pigeons. However, the impact of prevalent strains and the effectiveness of available vaccines in pigeons in Egypt are unclear. This study investigates the virulence of PPMV-1 isolated from vaccinated pigeons in Sharkia, Egypt (Pigeon/ Egypt/Sharkia-19/2015/KX580988). Ten-day-old specific-pathogen-free embryonated chicken eggs infected with this strain exhibited a mean death time (MDT) of 86.4±5.88 hours. The intracerebral pathogenicity index (ICPI) in day-old chicks was 0.8, while pigeons experienced an ICPI of 0.96 and an intravenous pathogenicity index (IVPI) of 2.11. These findings classify the strain as virulent and velogenic. Experimental infection with this PPMV-1 strain at 106 EID50/0.1 ml resulted in a 62.5% mortality rate in pigeons, displaying nervous and enteric distress. The virus caused extensive lesions in visceral organs, with strong immunohistochemistry signals in all examined organs, indicating the systemic spread of the virus concurrent to its neurotropic and viscerotropic tropism. Furthermore, vaccination using inactivated PPMV-1 and live NDV LaSota vaccine regimen protected 100% of pigeons against mortality, while a single NDV LaSota vaccine provided 62.5% protection. The PPMV alone or combined with NDV LaSota induced protective levels of hemagglutination inhibition antibody titers and reduced virus shedding from buccal and cloacal cavities. These findings suggest that using both inactivated PPMV-1 (G-VI) and live attenuated NDV (LaSota) vaccines is an effective prophylactic regimen for preventing and controlling PPMV-1 and NDV in pigeons, thereby reducing the risk of interspecies transmission.
Keywords:
igeon paramyxovirus serotype 1
; Avian Orthoavulavirus 1
; Pathogenicity
; Histopathology-Immunohistochemistry
; Vaccines efficacy
; Generalized Linear Gamma Model
1. Introduction
Pigeons (Columba livia) are hosts to a variety of diseases, including bacterial, viral, and parasitic. One infectious agent of particular importance in this species is pigeon paramyxovirus type 1 (PPMV-1) [1]. PPMV-1 is the primary cause of diseases in free-ranging and captive pigeons and doves [2,3]. It is closely related to Newcastle Disease virus (NDV), which causes severe outbreaks in chickens. Both PPMV-1 and NDV are enveloped viruses containing a negative-sense, single-stranded RNA genome that belongs to class II genotype VI (formerly known as genotype Vib or lineage 4b) of Paramyxovirus type 1 [4,5,6]. These viruses are classified into the same serotype within the Orthoavulavirus genus, Avulavirinae subfamily, and Paramyxoviridae family within the order Mononegavirales [7,8].
In the late 1970s, PPMV-1 was predicted to have originated in the Middle East and spread to Europe [9], where the first outbreak was reported in pigeons in the early 1980s [10,11]. Since then, PPMV-1 has become endemic in many countries globally [12,13,14]. The global spread of PPMV-1 is likely due to long-distance migration, competition flights, ornamental exhibitions, and the trade of live birds [15].
Since 1981, pigeons in Egypt exhibited clinical signs comparable to NDV infection, and the presence of NDV antigens was detected in serum samples taken from diseased pigeons in 1984 [9,16]. As the viruses of PPMV-1 genotype VI are known to infect the members of the family Columbidae they can also infect other wild birds and domestic poultry [17,18,19]. The virulence of NDV strains in chickens is quite variable and classified into three pathotypes: Lentogenic, mesogenic, and velogenic. These pathotypes are determined using assays such as mean death time (MDT) of chicken embryos, intracerebral pathogenicity index (ICPI), intravenous pathogenicity index (IVPI), and genomic sequences analysis [20,21,22]. However, most PPMV-1 strains are virulent in pigeons; their virulence in chickens is largely predicted based on the presence of multiple basic amino acids at the cleavage site motif of the viral fusion (F) protein [23,24,25]. This modulates the virulence phenotype from non-virulent to highly virulent phenotype in chickens [26,27]. Thus, the pigeon-originated viruses pose a constant threat to the poultry industry [23,28,29]. Depending on the genetic makeup of the infecting PPMV-1 strain, the pigeons experience a range of differential clinical signs, including nervous (tremors of the neck and wings, bilateral or unilateral locomotor disorders, torticollis, paralysis, and disturbed equilibrium), digestive (polydipsia, polyuria, anorexia, and diarrhea) [24,30,31] and/or respiratory (gasping, coughing, sneezing, and tracheal rales) signs [19].
PPMV-1 is enzootic in Egypt and causes significant negative economic impacts on commercial poultry production [19,32]. Proper and effective methods of prevention and control, in addition to diagnostic methods, are important for pigeon paramyxoviruses [13]. While vaccination of racing pigeons against NDV is compulsory in many countries [33], most pigeon-rearing strategies in Egypt vary and do not necessitate routine immunization against NDV [32]. Therefore, they are exposed to spillover from wild, unvaccinated pigeons. Thus, improving pigeon vaccination regimen against PPMV-1 could potentially reduce the disease burden in Egypt.
Several vaccination studies have evaluated the protective efficacy of available NDV poultry vaccines, such as live formulation of NDV LaSota or its variant Clone-30 strains which belong to Genotype II of Class II paramyxovirus [34]. The immunogenicity of pigeon-derived Genotype VI differs from the chicken-origin Genotype II vaccine strain of NDV [35,36]. However, little is known about the protective properties of an inactivated PPMV-1 vaccine in comparison to commercial NDV LaSota vaccine in pigeons [37].
Despite extensive research on the virus, further studies on the phenotypic characteristics of different strains of PPMV-1 prevalent in nature are needed. Experimental infections with contemporary strains dominating in the field could provide insights into their virulence phenotypes and assess the effectiveness of available commercial vaccines in mitigating their impacts.
This study evaluates the phenotypic properties (MDT, ICPI, IVPI) of the PPMV-1 strain isolated from pigeons (Pigeon/Egypt/Sharkia-19/2015/KX580988) in both chickens and pigeons. Additionally, it assesses the effectiveness of available commercial paramyxovirus vaccines in protecting pigeons from this PPMV-1 isolate.
These findings will enhance our understanding, diagnosis, and prevention of PPMV-1 infections in regions with intensive pigeon rearing. They will also help in reducing the risk of disease transmission and optimizing vaccination programs.
2. Materials and Methods
2.1. Ethics Considerations
The experiment design and protocol were approved by the Institutional Animal Care and Use Committee of Zagazig University, Egypt (approval ID ZU-IACUC/2/F/26/2020). This study was conducted in strict accordance with the approved guidelines for the Care and Use of Laboratory Animals. Birds were allowed to acclimatise to the facilities before the study began. Welfare checks were performed on the birds two to three times daily, and clinical scores were recorded to assess humane endpoints. Any bird showing signs of disease that met a humane endpoint was euthanised.
2.2. Birds
Domestic pigeons (Colomba livia) N=99 used in this study were hatched and reared in a backyard rearing system for 4 weeks and kept non-vaccinated against NDV and PPMV-1. One-day-old specific-pathogen free (SPF) white leghorn chicks N=20 (Koom Oshiem, Fayoum, Egypt) were used to characterise the PPMV-1 strain and evaluate the vaccine’s efficacy. All birds were housed in separate experimental units at the Faculty of Veterinary Medicine, Zagazig University.
Before the experiment, the health status of domestic pigeons was observed for 7–14 days. Five pigeons were randomly selected and euthanized, and then all tissues and organs were carefully examined for any macro-pathological lesions. Blood samples and tracheal and cloacal swabs were collected from the remaining pigeons.
It was found that the pigeons did not show any signs, and none of the euthanized birds had pathological lesions. The collected serum samples lacked avian paramyxovirus-1 (APMV-1) and avian influenza virus (AIV)-specific hemagglutination inhibition (HI) antibodies by the HI test. The HI test was performed using hyperimmune serum against NDV, PPMV-1 [38], and H5, H9 AIV (Department of Avian and Rabbit Medicine, Faculty of Veterinary Medicine, Zagazig University) according to the standard method of World Organisation of Animal Health (WOAH) [39]. Antibody titers of 1:4 or greater were used to indicate past infection, current exposure, maternal antibodies, or vaccination titers. Likewise, none of the swab samples had hemagglutinating agents using isolation in SPF embryonated chicken eggs (SPF-ECEs) and then identification by a rapid hemagglutination (HA) assay.
All animals, experimental procedures and protocols were approved by the Institutional Animal Care and Use Committee of Zagazig University, Egypt. The humane endpoints for chickens and pigeons inoculated with PPMV-1 were followed such that any infected bird showing signs of green watery diarrhoea, nervous symptoms (such as twisting of the head and neck or loss of balance), or difficulty in breathing (mouth breathing with an extended neck) was immediately euthanised and recorded as having reached severe disease signs.
2.3. Vaccines
Two vaccines used in this study. (i) Live freeze-dried form of LaSota lentogenic strain of Newcastle Disease virus (CEVAC® NEW L, Ceva Sante Animale Egypt, Al Sheikh Zayed, Giza) which contained 106 EID50 of vaccinal virus/dose and was delivered via eye drops of 50 µL per dose. (ii) Formalized inactivated pigeon paramyxovirus vaccine (containing 1010 EID50 /mL) obtained from the Veterinary Serum and Vaccine Research Institute (VSVRI, Abassia, Cairo, Egypt, Batch number 1907) and delivered via subcutaneous injection in the third middle of the neck at 0.5 mL dose per bird.
2.4. Pigeon Paramyxovirus-1 (PPMV-1) Isolate
The current study used the A PPMV-1 isolate named Pigeon/Egypt/Sharkia-19/2015/KX580988, (GenBank ID: KX580988) [32]. This isolate originated from a native pigeon flock in Al-Shabanat, Sharkia, Egypt, which had an outbreak in 2015. This flock was vaccinated with NDV LaSota vaccine and suffered from noteworthy nervous symptoms and diarrhea, with up to 80% mortality. Nephrosis, nephritis, enteritis, and a congested brain were recorded during necropsy. This isolate was identified as PPMV-1 by using hyperimmune serum against PPMV-1 prepared by Hamouda et al. [38] with a titer of 4 log2 in the HI test. Subsequently, RT-PCR was used to identify the viral RNA, and sequencing analysis of the isolate confirmed the presence of a poly-basic fusion protein cleavage site 112KRQKRF117, identifying the strain as velogenic PPMV-1 and belonged to subgenotype Vib.2, class II [32]. PPMV-1 isolate was propagated using inoculation in the allantoic sac of 10-day-old SPF embryonated chicken eggs (SPF-ECEs). After 5 days incubation at 37°C and relative humidity of 70%, ECEs with both live and dead embryos were investigated and allantoic fluids (Afs) harvested. Collected Afs were tested for hemagglutinating virus titer by a micro-plate hemagglutination (HA) test using 1% (v/v) washed chicken red blood cells (RBCs) as recommended by WOAH [39].
2.5. Biological Characterization
The pathogenic potential of the isolate was determined by mean death time (MDT), intracerebral pathogenicity index (ICPI), and intravenous pathogenicity index (IVPI) tests.
2.5.1. Mean Death Time (MDT) and Virus Titration
The MDT test was conducted using 10-day-old SPF-ECEs and calculated according to the method previously described [4]. The value of MDT was determined as the mean time in hours necessary for the death of all ECEs. Specifically, MDT values greater than 90 h indicate low virulence, between 60 and 90 h indicate moderate virulence, and less than 60 h indicate high virulence. The embryonic infectious dose of 50% (EID50) of the isolate was calculated using the Reed and Muench method [40] in SPF-ECEs. The virus stock was diluted in phosphate-buffered saline and standardized to 106 EID50/0.1 mL.
2.5.2. Intracerebral Pathogenicity Index (ICPI), and Intravenous Pathogenicity Index (IVPI)
A total of thirty clinically healthy, 5-week-old domestic pigeons (Colomba livia) and twenty, one-day-old SPF chicks were used to detect the ICPI [39,41] and IVPI [4]. To further elucidate the pathogenic potential of the virus via this pathway, ICPI was performed in SPF chicks as specified in all standard methods and was also applied in pigeons, which are natural hosts of the virus.
Three groups (G1P–G3P) of 10 pigeons per group and two groups (G4C–G5C) of 10 SPF chicks per group were prepared for inoculation. Groups G1P and G4C were inoculated intracerebrally (I/C) with 50 µL of the ten-fold dilution of the fresh virus AF stock containing HA titre of 210 (107.6 EID50) in 0.1 ml. Group G2P was inoculated intravenously (I/V) with 0.1 ml of the ten-fold dilution of the same fresh virus AF stock (Figure 1A). Groups G3P and G5C were used as mock-inoculated control birds and were housed separately from the other birds. The birds were observed and examined daily for 8 and 10 days in intracerebrally and intravenously inoculated groups, respectively. The observed clinical signs and mortalities were recorded and scored to determine the pathogenicity indexes.
The ICPI Involves scoring sick or dead (0 = normal; 1 = sick; and 2 = dead). ICPI below 0.7 was considered to be of low virulence, and those with an ICPI equal to or greater than 0.7 were considered to be virulent [39,42]. The IVPI involves scoring illness (0 = normal; 1 = sick; 2 = paralyzed or nervous signs; and 3 = dead) after IV inoculation. The velogenic strains have an IVPI score of 2–3, mesogenic 0–0.5, and lentogenic of zero [41].
2.6. Evaluating Vaccine Efficacy
2.6.1. Pigeon Experiment Design
Sixty-four 6-week-old clinically healthy domestic pigeons (Colomba livia) were divided into 4 groups (G1–G4) of 16 birds each. The experimental pigeons were kept in the experiment units under controlled conditions for the period of the study and received consistent feed and water ad libitum. The birds in groups 1, 2, and 3 were vaccinated with live NDV LaSota, inactivated PPMV, and dual vaccine- one dose each of NDV LaSota and PPMV, respectively (Figure 1B). The NDV LaSota vaccine was applied on the first day of the experiment and boosted after two weeks (8 weeks of age). On the second day of the experiment, the formalized-inactivated PPMV vaccine was administered once. Pigeons in group 4 were kept as the control (non-vaccinated) group.
At 9 weeks of age, four pigeon groups were subdivided into 8 subgroups (G1a, G2a, G3a, and G4a) and (G1b, G2b, G3b, and G4b). The G1b, G2b, G3b and G4b subgroups were challenged intra-oculonasally with dose of 0.1 mL containing 106 EID50 of the challenge virus “Pigeon/Egypt/Sharkia-19//2015/KX580988”. The G1a, G2a, G3a and G4a subgroups remained unchallenged. Serum samples were collected weekly from each subgroup (3 birds per subgroup) to determine the avian paramyxovirus-specific antibody titers using the hemagglutination inhibition (HI) test. Post-challenge, pigeons were monitored twice daily for 14 days to record clinical signs and mortality. The clinical signs of pigeons in the different groups were scored daily according to their clinical condition (0 = healthy; 1 = diseased; 2 = nervous signs; 3 = dead). The clinical index was calculated analogously to determining the intravenous pathogenicity index (IVPI). As determined by IVPI, the course of disease from inoculation was nearly the same as that from natural infection via the oculonasal route. In addition, the observation period was extended to 14 days to cover the time period from productive infection till complete recovery. Pigeons that died suddenly during the experiment were immediately necropsied. Birds that were culled upon reaching humane endpoints, as well as those that recovered from infection and did not experience humane endpoints, were euthanised at the end of the observation period for post-mortem examination and tissue collection to detect disease lesions. Three pooled tracheal and cloacal swabs were collected from 6 randomly selected pigeons (2 swabs per pool) at 3-, 5-, and 7-day post-challenge (dpc) to determine the virus shedding.
2.6.2. Immune Response
The HI test was performed on the collected serum samples according to the procedures listed in the WOAH [39] using 4 hemagglutination units (HAU) of the PPMV-1 strain (Pigeon/Egypt/Sharkia-19/2015/KX580988) as a whole virus, the LaSota vaccine strain of NDV (Pestikal® LASOTA SPF), and 1% chicken red blood cells (RBCs). The serum samples were thermally inactivated for 30 minutes at 56 °C. Twofold serial dilutions were carried out, and the HI titer was expressed as log2 of the reciprocal of the highest serum dilution, which suppressed the HA activity.
2.6.3. Virus Shedding Determination
As previously mentioned, tracheal and cloacal swabs were collected on the third, fifth and 7th dpc from different subgroups. These swabs were suspended in 1.5 mL minimum essential media (MEM) containing antibiotics (Penstrept, Lonza) then clarified by centrifugation at 3,000 rpm for 10 minutes at 4°C. Virus titration was implemented using Real-Time Quantitative Reverse Transcription polymerase chain reaction (RT-qPCR). Viral RNA was extracted from the supernatant using the QIAamp MinElute Virus Spin kit (QiagenGmbH, Hilden, Germany), according to the manufacturers’ protocol. The extracted RNA was subjected to one-step real time RT-PCR using AgPath-IDTM one –step RT-PCR Kit (Applied Biosystems, Thermo Fisher Scientific Inc., USA) for detection and titration of PPMV-1. The following primers and probe specific to F protein gene as designed by Sabra et al. [43] were used: forward primer 5′- TGATTCCATCCGCAGGATACAAG -3′, reverse primer 5′- GCTGCTGTTATCTGTGCCGA-3′, and probe: F-4876 5–[6-FAM] AAGCGYTTCTGTCTCYTTCCT CCT [BHQ_1]–3′. The amplification was performed in an Applied Biosystems™ StepOne™ Real-Time PCR System (Thermo Fisher Scientific Inc.), with cycle conditions as mentioned by Fuller et al. [44]. Overall, the amount of viral RNA in the swab samples was based on the threshold cycle (Ct) values obtained by RT- qPCR.
2.7. Microscopic Examination to Track the Pathological Pathway of PPMV-1 (Pigeon/Egypt/Sharkia-19/2015/KX580988) Strain in Infected Pigeons
2.7.1. Histopathology
In the non-vaccinated challenged group, in which the pigeons were oculonasaly inoculated with PPMV-1 at 9 weeks of age, tissue samples were collected from different organs (brain, trachea, lung, heart, liver, pancreas, spleen, proventriculus, intestine and kidneys) from the freshly dead and euthanized symptomatic pigeons. The collected tissues were fixed in 10% neutral-buffered formalin before being embedded in paraffin and sectioned in duplicate to 3 µm thickness. For histopathology, slides were stained by hematoxylin and eosin (H&E) stain and examined by a light microscope [45].
2.7.2. Immunohistochemistry
To produce hyperimmune serum against PPMV-1, rabbits were given a series of injections in accordance with the schedule outlined by Samiullah et al. [46]. MagneProtein G Beads for Antibody Purification (Promega Corporation, Madison, Wisconsin, USA) were used to purify antibodies in accordance with the manufacturer's instructions.
Tissue sections were placed on slides coated with poly-L-lysine, which were then deparaffinized and rehydrated. Heat-induced antigen retrieval, blocking of nonspecific protein binding and endogenous peroxide application were performed. Tissue sections were incubated overnight with a monoclonal primary antibody (rabbit anti-PPMV-1 Ig), followed by incubation with horseradish peroxidase-conjugated goat polyclonal secondary antibody to rabbit Ig (SM802 EnVisionTM FLEX/HRP). Color was developed with a 3,3'-Diaminobenzidine (DAB) substrate (DM827 EnVisionTM FLEX DAB þ chromogen) [47]. Positive results appeared as a brown precipitate localized at the site of binding and were observed under an optical microscope. Negative tissue slides were established by adding negative serum instead of the primary antibody on non-vaccinated challenged pigeon (G4b) tissue specimens and used as a reaction guide. Additionally, tissue specimens of non-vaccinated non-challenged pigeons (G4a) were incubated with the primary anti-PPMV-1 antibodies.
2.8. Statistical Analysis
Several linear and generalized linear models were performed, and the Akaike Information Criterion (AIC) was used to select the model with the best quality to fit the data. A generalized linear model (GLM) with a gamma distribution and log link function was employed to analyze hemagglutination inhibition (HI) titers as it had the lowest AIC. The HI titers are continuous, positively skewed data, and the gamma distribution is well-suited for modeling such data. The log link function ensures that predicted values remain positive on the original HI titer scale.
The generalized linear model gamma regression with log link function is used to assess the pattern of the HI titer change using two different antigens within five groups over the subsequent five weeks of the experiment. The probability function of the gamma distribution is as follows:
for y (HI titre) > 0, α > 0 (the shape parameter) and β > 0 (the scale or the spread parameter), where E[y] = αβ and var[y] = αβ2. Note that Γ () is the gamma function. The y variable is now changed to z as: y → z = exp{y} to receive the density of the log-gamma distributed random variable Z = exp{Y} [48].
The analysis was conducted in R (version 2023.06.1) using the “glm” function. Maximum likelihood estimation was used to fit the model. Goodness-of-fit was evaluated using chi-square tests and residual deviance. Estimated coefficients and their p-values were reported to assess the significance of predictor variables on the log-transformed mean change in HI titers. P value < 0.05 is considered a significant level. The model coefficients were exponentiated to provide insights into the actual effects of the predictors (antigen type, treatment groups, and weeks of experiment) on HI titer change.
3. Results
3.1. The Selected PPMV-1 Strain Caused Severe Disease Symptoms and Was Lethal to Chicken Embryos
Virulence of the selected PPMV-1 strain to chicken embryos was determined by inoculating ten-day-old SPF ECEs with the Pigeon/Egypt/Sharkia-19/2015/KX580988 strain. Inoculated embryos showed severe congestion of visible veins, along with subcutaneous hemorrhages, and stunted growth, in the first passage in 80% of inoculated embryos at 96 hours and increased to 100% within 48-72 hours post-inoculation (pi) in the third passage. The embryo death reached to 100% of inoculated embryos within 72 hours pi. The virus present in the collected allantoic fluid (AF) at 72 hours pi contained an HA titer of 10 log2 using the micro-plate hemagglutination (HA) test.
3.2. Selected PPMV-1 Strain Caused Severe Clinical Disease and Was Lethal to Chickens and Pigeons
The virulence of the selected PPMV-1 strain (Pigeon/Egypt/Sharkia-19/2015/KX580988) in pigeons and chickens was determined by adopting standard MDT [40], ICPI [39,41], and IVPI [4] protocols. The MDT was 86.4±5.88 hours in SPF-ECEs, suggesting that the virulence of this isolate was moderate. The observed ICPI in one-day-old SPF infected chicks was 0.8, while it was 0.96 in 5-week-old infected pigeons. This supports the classification of this PPMV-1 strain as a virulent pathotype; thus, outbreaks with closely related strains come under the notifiable outbreaks to WOAH. The observed IVPI score in 5-week-old pigeons was found to be 2.11, which deemed the strain velogenic. The presence of a multi-basic cleavage site motif in the virus F protein together with in ovo and in vivo infection scores determined via MDT, ICPI, and IVPI assays (Figure 2) confirms that the Pigeon/Egypt/Sharkia-19/2015/KX580988 strain is a virulent pathotype for both pigeons and chickens.
3.3. Pigeons Vaccinated with PPMV or Dual Vaccine (LaSota and PPMV) Showed Protection from Clinical Diseases and Mortalities when Challenged with PPMV-1 strain (Pigeon/Egypt/Sharkia-19/2015/KX580988)
To determine the protective efficacy of available vaccines against PPMV-1 strains infecting pigeons in Egypt, sixty-four six-week-old pigeons were divided into four groups (n=16/group) and vaccinated with live NDV LaSota (G1), inactivated PPMV-1 (G2), or dual vaccine—one dose per each NDV LaSota and PPMV-1 (G3). The fourth group (G4) was kept as an unvaccinated control. Three weeks post-vaccination, the birds from each group were divided into two subgroups (a and b, each n=8/subgroup). One of each subgroup (G1b, G2b, G3b, G4b) was challenged with the PPMV-1 strain (Pigeon//Egypt/Sharkia-19/2015/KX580988), while the second subgroups (G1a, G2a, G3a, G4a) were kept as a non-challenged control (Figure 1). Clinical signs and mortality rates were recorded (Figure 3a).
Survival curves post-challenge in vaccinated and unvaccinated pigeons using tested vaccines are shown in Figure 3a. The clinical signs of PPMV-1 infection in the non-vaccinated challenged group (G4b) started on the fifth-day post-infection (dpi). All infected birds suffered from ruffled feathers, lethargy, and anorexia; one bird showed respiratory signs and greenish diarrhea was seen in 5 birds at 7 dpi. Six out of 8 birds showed signs of head tremors, torticollis, opisthotonus position, and wing or complete paralysis, which appeared at 8 dpi (Table 1 and Figure 4A–D). Almost all birds experienced moderate to severe disease signs and the cumulative clinical score was 1.03 (Figure 3b)
100% of the infected pigeons showed the above-mentioned clinical signs with clinical score equal to 1.03 (Figure 3b). Among the eight infected pigeons, five birds died from 7 to 12 dpi (mortality rate (62.5%), and the remaining three (37%) pigeons survived with clinical distress.
Pigeons infected with PPMV-1 after vaccination with either PPMV alone (G2b) or a dual vaccine (NDV LaSota and PPMV) (G3b) showed no apparent clinical disease signs and remained nearly normal, with only one bird in the group of eight showed mild diarrhea (G2b) and head tremor (G3b) at 13 dpc, but all recovered; thus the vaccine protection efficacy from mortality was 100%. The pigeons in the NDV LaSota-only vaccinated, virus-challenged group (G1b) showed relatively reduced protection from clinical disease signs compared to those that received PPMV alone or the dual (NDV LaSota and PPMV) vaccine. Clinical manifestations in these pigeons began at 7 days post-challenge (dpc), and the overall clinical score remained at 62.5%. This was relatively less pronounced in terms of severity and frequency compared to the non-vaccinated, challenged group (Table 1 and Figure 4E,F). Overall, the clinical score of the NDV LaSota vaccinated challenged group was 0.67 (Figure 3b), with a survival rate of 62.5%. The vaccinated and non-vaccinated groups of pigeons (G1a-G4a) that were kept as controls and not challenged with the virus (to monitor natural exposure from the environment) showed no clinical disease signs or mortality, confirming that the experimental birds were not exposed to PPMV-1 or any other natural infections commonly causing clinical disease in pigeons.
3.4. Pigeons Vaccinated with PPMV or Dual Vaccines (NDV LaSota and PPMV) Showed Low Severity of Postmortem Lesions when Challenged with PPMV-1 strain (Pigeon/Egypt/Sharkia-19/2015/KX580988)
The severity of disease induced by virus challenge in vaccinated versus non-vaccinated pigeons was assessed by observing post-mortem lesions. Following the virus challenge, all pigeons (n=8) in the non-vaccinated control group (G4b) either died or were culled because of reaching the humane endpoint. The severity of gross lesions in these birds was apparent including congestion of all visceral organs (septicemia), a congested and hemorrhagic brain, and pancreas with punctate hemorrhages or necrosis. The spleen was enlarged, congested (n = 4), and atrophied and the kidneys displayed nephrosis or nephritis-like condition. Hemorrhages were noticed between the esophagus and proventriculus; additionally, enteritis was recorded with greenish content in the gizzard and intestine. In 2 out of 8 pigeons, the heart showed minute petechial hemorrhages. The thymus and bursa of Fabricius were atrophied in 5 and 4 of the infected pigeons, respectively (Figures 4G–L and Table 2). In the group vaccinated with NDV LaSota (G1b), five pigeons either died or were euthanized after reaching humane endpoints. These birds exhibited post-mortem lesions with moderate severity (Figures 4M-Q and Table 2). In contrast, in the groups vaccinated with PPMV (G2b) or dual vaccinated with both LaSota and PPMV (G3b), only one pigeon from each group became reached the humane end point and was euthanized. Post mortem showed relatively mild, less pronounced gross lesions in their visceral organs (Table 2).
3.5. The PPMV-1 Strain (Pigeon/Egypt/Sharkia-19//2015/KX580988) Induced Extensive Microscopic Lesions in the Tissue of Visceral Organs of Non-Vaccinated Infected Pigeons
Microscopic lesions in the brain, pancreas, spleen, liver, kidneys, and heart were examined after H&E staining. Histopathological pictures of the brain revealed congested capillaries, sub-meningeal hemorrhages, and focal cortical hemorrhages. Multifocal degeneration and swelling of neurons were also observed, along with focal gliosis (Figure 5a–c). The pancreas of infected pigeons showed necrotic pancreatitis characterized by severe congestion, multifocal hemorrhage, and multifocal aggregations of lymphocytic infiltrations, as well as degeneration and necrosis of pancreatic acini (Figure 5d–f). Multifocal hemorrhages, congestion of sinusoids and splenic blood vessels, and lymphoid depletion of the white pulp of the spleen were observed (Figure 5g–i). Microscopic examination of livers showed severe congestion, focal degeneration, and discrete necrosis of some hepatic cells, along with mild leukocytes around bile ducts (Figure 5j). The kidney displayed tubulo-interstitial nephritis characterized by inter-tubular hemorrhages, degeneration of renal tubular epithelium, and focal necrosis with leukocytic infiltrations (Figure 5k). Inter-muscular congestion and hemorrhages were found in the heart (Figure 5l). By contrast, no histopathological changes were observed in respective tissues of the non-vaccinated non-challenged pigeons (Supplementary Figure S1).
3.6. The PPMV-1 Strain (Pigeon/Egypt/Sharkia-19/2015/KX580988) Expressed Strong Immunohistochemistry Signals in the Tissue Organs of Non-Vaccinated Infected Pigeons
The PPMV-1 antigen was found to be expressed in various tissue organs of pigeons, where a strong positive peroxidase reaction was detected. This reaction was shown in the brain neurons (Figure 6a), tracheal epithelium, with the inflammatory cells infiltrated in the propria submucosa (Figure 6b), and air capillaries of the lung (Figure 6c). The exocrine pancreatic acini (Figure 6d), splenic parenchyma (Figure 6e), and cardiomyocytes of some affected pigeons (Figure 6f) had viral antigen expression. Furthermore, the acinar epithelium of the proventriculus with the desquamation epithelial cells (Figure 6g), the intestinal mucosa (Figure 6h), and the renal tubular epithelium (Figure 6i) with the inflammatory cells occupying the interstitial tissue showed positive immunolabeling. This suggests that PPMV-1 is systemically distributed in internal organs with clear neuro-viscero tropism. Negative immune staining was observed in the non-vaccinated non-challenged pigeon (G4a) using the prepared specific anti-PPMV-1 antibody. Similarly, negative immunostaining was detected in non-vaccinated challenged pigeons (G4b) following staining with negative serum (Supplementary Figure S2).
3.7. Vaccination with PPMV-1 or Dual with NDV LaSota and PPMV-1 Produced Predicted Protective Levels of HI Antibody Titers in Pigeons
Serum was collected from all vaccinated pigeons and subjected to HI assays. The data show that, 3 weeks post-vaccination, pigeons vaccinated with PPMV and challenged (G2b) had an HI titre of 6±0.47 and 5.67±0.45 using LaSota and PPMV-1 antigens, respectively. Besides, pigeons vaccinated with LaSota and PPMV and challenged (G3b) had an HI titre of 5±0.39 using both antigens of LaSota and PPMV-1 (Table 3). Initially, plotting the density distribution of the HI titer revealed a positive skewness in the data (Figure 7a). Therefore, the log of the HI titer using the gamma distribution was recommended to represent the model with the data, which was selected as the best fit after comparing other routinely used models such as linear, general linear, and generalized linear models. A comparison to the linear or generalized linear models revealed that the generalized linear model with gamma distribution and log link function was the best, having the lowest AIC (Figure 7b). The predicted values of the generalized linear gamma model for the mean antibody titers using the HI test clarified that there were no differences between the use of LaSota (as a heterologous antigen) and PPMV-1 (as a homologous antigen), as shown in Figures 8b and c.
The model coefficients (Figure 8a) and estimated marginal means (Table 3) indicated that the antibody titers were significantly developed at 3 weeks post-vaccination (the time of challenge) in groups vaccinated with inactivated PPMV (G2b) (5.67±0.45:6±0.47) and NDV LaSota with inactivated PPMV (G3b) (5±0.39) compared to the NDV LaSota group (G1b) and unvaccinated groups (G4a and G4b). The antibody (Ab) levels of the same groups (G2b and G3b) continuously increased post-challenge and reached the highest titers at 5 weeks post-vaccination (Figures 8b and 8c), particularly in G2b, either using PPMV or NDV LaSota antigens, with a coefficient of 1.56. This indicated that this group at week 5 post-vaccination was related to a highly significant (56 times) increase in HI titer with a P value equal to 2.2e-16, compared to other groups and weeks.
In the NDV LaSota vaccinated group (G1b), the development of HI antibodies (Abs) was slow and not detected until the fifth week (Figures 8b and 8c). Although its Abs level was high (coefficient value = 1.19 in the fifth week), it was lower than from the time of the PPMV-1 challenge, as shown in Figure 8a, and the increase in HI titer at the 5th week wasn’t significant (P = 0.06).
3.8. The Inactivated PPMV Vaccine and LaSota with PPMV Vaccines Reduced the Virus Shedding in the Challenged Pigeons with Pigeon/ Egypt/Sharkia-19/2015/KX580988
The vaccinated group with the inactivated PPMV vaccine (G2b) inhibited virus shedding. The group that received live LaSota with inactivated PPMV vaccines (G3b) prevented the tracheal shedding and markedly minimized the cloacal shedding, with a Ct value of 37 in one out of 3 vial swabs at 7 dpc. In the live NDV LaSota vaccination group (G1b), virus shedding was detected in both tracheal and cloacal swabs as early as day 5 post-challenge; however, it reduced the number of birds that shed the virus, contrasting that in the unvaccinated challenged group (G4b) (Table 4).
4. Discussion
PPMV-1 is now a serious global hazard to bird populations, with pigeons being particularly vulnerable [49,50]. The strain "Pigeon/Egypt/Sharkia-19//2015/KX580988," previously isolated from a vaccinated native pigeon flock in Egypt, was classified as a velogenic strain that belonged to subgenotype VIb.2, class II. The study was designated to determine the virus pathotype characteristics in pigeons and chickens and to evaluate the protective efficacy of available commercial paramyxovirus vaccines against the velogenic strains of PPMV-1. [35]. Based on the observed MDT value (86.4±5.88 in 10-day-old SPF-ECEs), ICPI value (0.8 ad 0.96 in 1-day-old SPF-chicks and 5-week-old pigeons respectively), and IVPI scores (insert relevant IVPI score), the Pigeon/Egypt/Sharkia-19//2015/KX580988 strain exhibits moderate virulence pathotype characteristics. Earlier reported studies displayed similar pathogenicity for PPMV-1 using MDT and ICPI in chicks [51,52,53,54,55,56], although the poly-basic fusion protein in the cleavage site identified this strain as velogenic PPMV-1 [32]. The majority of PPMV-1 strains are mesogenic or lentogenic for chickens, according to ICPI and MDT tests, even though they have been demonstrated to have deduced amino acid motifs, suggesting high pathogenicity [25,57]. These imply that other factors affect the pathogenic phenotype of these viruses and that the cleavage site is not the only factor regulating PPMV-1's pathogenicity in various species [26,35]. According to WOAH standards, the ICPI of the strain used in present study belongs to the virulent or velogenic category [39]. Notably, the ICPI (0.96) was relatively high in pigeons, with more pigeons showing nervous signs (n = 4/10) and mortalities (n = 6/10) than chicks. Considering that pigeons are the natural host of the PPMV-1, it may not be suitable for its assessment to be carried out in chickens only [37]. However, when the PPMV-1 strain was tested for pathogenicity and clinical symptoms after successive passages in chickens, it became more virulent [1,13,26]. This pilot study and others [37] concluded that in order to evaluate the real virulence of PPMV-1, pigeons should be employed for pathogenicity testing.
It is interesting to note that the IVPI value suggested the PPMV-1 was velogenic with a score of 2.11 in pigeons, accompanied by mortality (n = 10/10) within 7 days post-inoculation. The different degrees of virus pathogenicity in pigeons relate to different inoculation routes. In the intravenous route, the virus enters the bloodstream immediately and subsequently spreads to different organs at a faster rate [58]. This systemic route of virus inoculation is the cause of the nervous involvement [59], and high mortality that reaches 100%. According to the results of the bioassays used in this prospective study, the PPMV-1 strain used in the present study showed pathogenicity in both chicks and pigeons.
Considering the increase in PPMV-1 infections in recent years, disease prevention is required. Effective vaccinations or other preventative and control measures need to be developed and applied in pigeon lofts to reduce losses and the risk of disease transmission to other birds [60]. The commercial vaccines usually used in most countries to control variant paramyxovirus-1 (PPMV-1) are classical paramyxovirus-1 and chicken NDV-live or inactivated vaccines [34]. In contrast to most other countries, Egypt provides commercially accessible inactivated pigeon paramyxovirus (PPMV) in addition to the conventional NDV vaccines. The available commercial paramyxovirus vaccines under investigation in this study were the live attenuated NDV vaccine (LaSota) and the formalized inactivated pigeon paramyxovirus (PPMV).
To complete the understanding of PPMV-1 pathogenicity and identify the markers of its natural pathologic pathway, the pigeons infected via the pigeons oculonasal route were thoroughly investigated. Clinical findings and morbidity and mortality rates were recorded daily. Histopathology examination was used to document the microscopic lesions in the various tissue organs. Additionally, immunohistochemistry (IHC) was used to determine the gene expression of the viral strain and confirm that the lesions were the result of PPMV-1 infection.
Consistent with previous studies [53,55,56,61], the non-vaccinated challenged pigeons exhibited typical clinical signs and gross lesions. It is important to note that the nervous signs and greenish diarrhea were the common signs as documented by Śmietanka et al. [1], Zhan et al. [55], Shalaby et al. [62], and were accompanied by lesions recorded in the brain and digestive tract of 100% of pigeons. Primarily through the pattern of noticed signs and lesions, it is expected that this PPMV-1 strain has neurotropic and viscerotropic potentiality. Moreover, Chang et al. [53] have concluded that virus titer was higher in the brain, intestine, and lung than in other tissues.
The signs in the current study started at 5 dpi, as previously recorded by Xie et al. [36] and Chang et al. [53]. However, other studies showed signs as early as 2-4 dpi [55,56,61]. This variation in the incubation period depends on the virus’s properties, the dose in the inoculum, and host features such as age, breed, and immune response [1]. Additionally, the clinical score recorded by the infected pigeons reached 1.03, and the morbidity and mortality rates were 100% and 62.5%, respectively, factoring in birds that died or reached the humane endpoint within 2 days of the onset of clinical signs.
The microscopic lesions observed in multiple organs of PPMV-1 infected pigeons were similar to those noted in previous reports [37,61,62,63,64]. This in-depth microscopic examination of the infected organs revealed the extent of virus dissemination and the direct damage it causes, negatively impacting the birds' health and contributing to morbidity and mortality. Among the common gross lesions and microscopic changes that caught our attention were in the brain, informing sub-meningeal and focal cortical hemorrhages, multifocal degeneration, and swelling of neurons with focal gliosis. The involvement of the central nervous system (CNS) in PPMV-1 disease was prevalent, particularly given the neurologic clinical signs often observed. This indicates the tropism of the PPMV-1 strain for the nervous tissues. Necrotizing pancreatitis was most often observed in pigeons and is considered an acute change which might cause animal death due to systemic inflammatory response syndrome and multi-organic failure as a result of releasing activated proteolytic enzymes and pro-inflammatory cytokines into the bloodstream [65]. Tubulointerstitial nephritis was observed which could contribute to the loss of homeostasis. Only a few studies had reliable confirmation of lymphocytosis [63]. On the other hand, notable alterations in lymphoid organs have not always been described in PPMV-1 outbreaks [66,67]. In the present study, lymphoid depletion of the spleen's white pulp was recorded in the presence of viral antigen expression in the IHC, which suggests that PPMV-1 may cause lymphoid depletion. Although IHC analysis has been adopted a few times in prior studies [62,63], it confirmed incrimination of PPMV-1 responsibility for changes in the collected tissue organs by detecting a strong PPMV-1 antigen expression. This suggests that the use of IHC to detect PPMV-1 infections in tissues could be a potential tool to trace the viral pathway in tissues and confirm its lesions.
PPMV-1 was able to cause systemic infection, which had a wide range of tissue distribution in pigeons, along with the prominent neurological and intestinal symptoms, and was broadly consistent with pronounced lesions in these organs. This suggests that PPMV-1 is a systemic virus with neuro- and viscero-tropism.
All the vaccinated groups had a lower severity and frequency of the signs of disease compared to the non-vaccinated pigeons. Importantly, the pigeons were more protected against the PPMV-1 challenge by both programs of the inactivated PPMV vaccine and LaSota with PPMV, which inhibited the appearance of clinical symptoms. The clinical scores for both vaccination programs were 0.01 and 0.02, respectively, and their corresponding morbidity rates were 12.5%. On the other hand, the LaSota vaccination had the lower protection rate, which the clinical score was 0.67 and a 62.5% morbidity rate. Both vaccine programs containing inactivated PPMV protected 100% of pigeons against mortality, while the LaSota vaccine was 62.5%. This indicated that the inactivated PPMV vaccine provided good protection. Similar observations were reported by Zhang et al. [37], Amer et al. [60], Stone [68], Hassan, [69], who conducted their investigation on the inactivated PPMV vaccine either locally commercial [60,69] or prepared from an isolated strain [37,68] and ND(V?) vaccines against PPMV-1 challenge in pigeons. They recommended that the homologous vaccine is more efficacious than the ND(V?) vaccines such as Ulster, LaSota, and Hitchner B1. And the protection rate reached 100% against morbidity and mortality.
It is well known that antibodies against viruses protect the host from damage caused by the virus [70,71]. Comparing the immune efficacies in the vaccinated groups using the HI test, the vaccination containing inactivated PPMV whether alone or with LaSota developed significant high antibody levels in pigeons from 3 weeks post-vaccination (initial PPMV-1 challenge) than in the LaSota vaccine alone. Antibody level also increased over time post-challenge. This suggests that the PPMV-1-matched vaccine could induce elevated humoral response. This finding mirrors the results of Zhang et al. [37].
Using either LaSota or PPMV-1 as diagnostic antigens against HI antibodies of PPMV-1 showed similar inhibition. This could be attributed to the use of polyclonal antibodies rather than monoclonal ones since only monoclonal antibodies prepared against the PPMV-1 variant virus can inhibit all PPMV-1 isolates in HI test but no other NDV strains [72]. Monoclonal specific epitopes on the F polypeptide induce greater neutralization than those against HN in vitro and in vivo tests [73].
Likewise, the two vaccination programs containing inactivated PPMV resulted in no pigeons shedding virus. In contrast, the LaSota vaccine did not prevent virus shedding from the cloaca or orally, which was detectable from the fifth day post-challenge, although it decreased the number of birds that shed the virus in comparison with non-vaccinated pigeons.
A notable problem of this disease is the virus shedding from the infected pigeons [51,53]. The main risk from the virus is its ability to transmit from infected birds to other bird populations (wild or domesticated) and evolutionary changes which potentially increases its virulence phenotype. So, protection against virus shedding is considered one of the important priorities for evaluating the success of vaccination programs. In this study, LaSota vaccine provided insufficient protection against morbidity, mortality, virus shedding with low antibody level pre- and post-challenge. Previous studies detected that the ND-derived strain vaccines were inadequate for the protection of pigeons against PPMV-1, such as LaSota vaccine [37,74] and Hitchner B1 vaccine [60]. Additionally, confirmed by field practices, it is shown that despite the use of LaSota vaccine in pigeons, the disease outbreaks continue [19,35], and this is the case in the pigeons from which the strain under study was obtained. This is attributed to the presence of biological, serological, and genetic differences between the prevalence both the NDV and the PPMV-1 strain [34,37,51,75] even if it was minor [35]. The known transmitted NDV strains have undertaken major shifts in their genotypes over the past few decades, although they still belong to a single serotype [70].
5. Conclusions
To maintain the continued growth of the poultry sector, PPMV-1 infection in pigeons must be controlled [35]. Applying live NDV chicken-adapted vaccines alone like LaSota won't provide full protection against PPMV-1. This study provided a potential application of inactivated PPMV, which has the potential to provide full protection against PPMV-1. A more suitable and sufficient prophylactic program to prevent and control both PPMV-1 and NDV in pigeons is the combination of both the live attenuated NDV vaccine such as LaSota and the inactivated PPMV-1 (G-VI). This diminishes the risk of both viruses from interspecies transmission. The preference to use both live LaSota in combination with inactivated PPMV-1 may induce both humoral immune response to PPMV-1 and the live NDV potentially driving cell-mediated immunity providing faster and higher levels of protection [76]. This ensures early protection against the challenges of field viruses. Along with this, in countries like Egypt that have commercial PPMV vaccines, it is necessary to develop them by targeting the currently circulating strains and then applying it to all pigeon lofts, while countries that have not yet implemented vaccines should look into developing and producing antigenically matched inactivated vaccines.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.A.M.E. and M.I.; methodology, A.A.M.E., R.M.E., E.E.H., H.F.G., A.A.D., and A.H.E..; formal analysis, H.F.G., A.A.M.E., and R.M.E.; investigation, A.A.M.E., R.M.E., and E.E.H.; writing—original draft preparation, A.A.M.E., R.M.E., E.E.H., and H.F.G.; writing—review and editing, A.A.M.E., M.I., and R.M.E.; visualization, A.A.M.E., R.M.E., E.E.H., H.F.G., A.A.D., A.H.E., and M.I.; project administration, A.A.M.E., and M.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grant #27667 from the Newton Mosharafa and British Council Newton Fund Institutional Links grant #IL3261727271, MI is supported through UK Research and Innovation (UKRI), Biotechnology and Biological Sciences Research Council (BBSRC) grants (BBS/E/PI/230001A, BBS/E/PI/230001C, BBS/E/PI/230002B).
Informed Consent Statement
Not applicable.
Data Availability Statement
Participant data will be made available upon reasonable requests directed to the corresponding author. The investigator and collaborators will review and approve proposals based on scientific merit. After approval of a proposal, data can be shared through a secure online platform after signing a data access agreement.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
References
- Śmietanka, K.; Olszewska, M.; Domańska-Blicharz, K.; Bocian, Ł.; Minta, Z. Experimental infection of different species of birds with pigeon paramyxovirus type 1 Virus—Evaluation of clinical outcomes, viral shedding, and distribution in tissues. Avian Dis. 2014, 58, 523-530. [CrossRef]
- Alexander, D.J.; Parsons, G. Avian paramyxovirus type 1 infections of racing pigeons: 2 pathogenicity experiments in pigeons and chickens. Vet. Rec. 1984, 114, 466–469. [CrossRef]
- Wang, H.Y.; Wu, M.C.; Chen, H.W.; Lai, Y.C.; Huang, W.H.; Chang, H.W.; Jeng, C.R.; Cheng, C.H.; Wang, P.J.; Lai, Y.H.; Chang, Y.C. Isolation, full sequence analysis, and in situ hybridization of pigeon paramyxovirus-1 genotype VI. 2.1. 1.2. 2 from oriental turtle doves (Streptopelia orientalis). Poult. Sci. 2023, 102, 102974. [CrossRef]
- Alexander, D.J.; Senne, D.A. Newcastle disease. In Diseases of Poultry, 12th ed.; Saif, Y.M., Barnes, H.J., Glisson, J.R., Fadly, A.M., McDougald, L.R., Swayne, D.E., Eds.; Blackwell Publishing: Ames, IA, USA, 2008; pp. 75–100.
- Kim, L.M.; King, D.J.; Guzman, H.; Tesh, R.B.; Da Rosa, A.P.T.; Bueno Jr, R.; Dennett, J.A.; Afonso, C.L. Biological and phylogenetic characterization of pigeon paramyxovirus serotype 1 circulating in wild North American pigeons and doves. J. Clin. Microbiol. 2008, 46, 3303-3310. [CrossRef]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect. Genet. Evol. 2016, 39, 22-34. [CrossRef]
- Amarasinghe, G.K.; Bào, Y.; Basler, C.F.; Bavari, S.; Beer, M.; Bejerman, N.; Blasdell, K.R.; Bochnowski, A.; Briese, T.; Bukreyev, A.; Calisher, C.H.; Chandran, K.; Collins, P.L.; Dietzgen, R.G.; Dolnik, O.; Dürrwald, R.; Dye, J.M.; Easton, A.J.; Ebihara, H.; Fang, Q.; Formenty, P.; Fouchier, R.A.M.; Ghedin, E.; Harding, R.M.; Hewson, R.; Higgins, C.M.; Hong, J.; Horie, M.; James, A.P.; Jiāng, D.; Kobinger, G.P.; Kondo, H.; Kurath, G.; Lamb, R.A.; Lee, B.; Leroy, E.M.; Li, M.; Maisner, A.; Mühlberger, E.; Netesov, S.V.; Nowotny, N.; Patterson, J.L.; Payne, S.L.; Paweska, J.T.; Pearson, M.N.; Randall, R.E.; Revill, P.A.; Rima, B.K.; Rota, P.; Rubbenstroth, D.; Schwemmle, M.; Smither, S.J.; Song, Q.; Stone, D.M.; Takada, A.; Terregino, C.; Tesh, R.B.; Tomonaga, K.; Tordo, N.; Towner, J.S.; Vasilakis, N.; Volchkov, V.E.; Wahl-Jensen, V.; Walker, P.J.; Wang, B.; Wang, D.; Wang, F.; Wang, L.; Werren, J.H.; Whitfield, A.E.; Yan, Z.; Ye G.; Kuhn, J.H. Taxonomy of the order Mononegavirales: update 2017. Arch. Virol. 2017, 162, 2493-2504. [CrossRef]
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasinghe, G.K.; Anthony, S.J.; Avsic- Zupanc, T.; Ayllon, M.A.; Bahl, J.; Balkema-Buschmann, A.; Ballinger M.J. 2020 taxonomic update for phylum Negarna-viricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020, 165, 3023–3072. [CrossRef]
- Kaleta, E.F.; Alexander, D.J.; Russell, P.H. The first isolation of the avian pmv-1 virus responsible for the current panzootic in pigeons?. Avian Pathol. 1985, 14, 553-557. [CrossRef]
- Biancifiori, F.; Fioroni, A. An occurrence of Newcastle disease in pigeons: virological and serological studies on the isolates. Comp. Immunol. Microbiol. Infect. Dis. 1983, 6, 247–252. [CrossRef]
- Alexander, D.J.; Russell, P.H.; Collins, M.S. Paramyxovirus type 1 infections of racing pigeons: 1 characterisation of isolated viruses. Vet. Rec. 1984, 114, 444–446. [CrossRef]
- Werner, O.; Römer- Oberdörfer, A.; Köllner, B.; Manvell, R.J.; Alexander, D.J. Characterization of avian paramyxovirus type 1 strains isolated in Germany during 1992 to 1996. Avian Pathol. 1999, 28, 79–88. [CrossRef]
- Alexander, D.J. Newcastle disease in the European Union 2000 to 2009. Avian Pathol. 2011, 40, 547–558. [CrossRef]
- Napp, S.; Alba, A.; Rocha, A.I.; Sánchez, A.; Rivas, R.; Majó, N.; Perarnau, M.; Massot, C.; Miguel, E.S.; Soler, M.; Busquets, N. Six-year surveillance of Newcastle disease virus in wild birds in north-eastern Spain (Catalonia). Avian Pathol. 2017, 46, 59–67. [CrossRef]
- Aldous, E.W.; Fuller, C.M.; Ridgeon, J.H.; Irvine, R.M.; Alexander, D.J.; Brown, I.H. The Evolution of Pigeon Paramyxovirus Type 1 (PPMV-1) in Great Britain: A Molecular Epidemiological Study. Transbound. Emerg. Dis. 2014, 61, 134-139. [CrossRef]
- Vindevogel, H.; Duchatel, J.P. Panzootic Newcastle disease virus in pigeons. In Newcastle Disease, Alexander, D.J., Ed.; Kluwer Academic Publishers: Boston, 1988; pp. 184 -196.
- Terregino, C.; Cattoli, G.; Grossele, B.; Bertoli, E.; Tisato, E.; Capua, I. Characterization of Newcastle disease virus isolates obtained from Eurasian collared doves (Streptopelia decaocto) in Italy. Avian Pathol. 2003, 32, 63-68. [CrossRef]
- Marlier, D.; Vindevogel, H. Viral infections in pigeons. Vet. J. 2006, 172, 40–51. [CrossRef]
- Guo, H.; Liu, X.; Han, Z.; Shao, Y.; Chen, J.; Zhao, S.; Kong, X.; Liu, S. Phylogenetic analysis and comparison of eight strains of pigeon paramyxovirus type 1 (PPMV-1) isolated in China between 2010 and 2012. Arch. Virol. 2013, 158, 1121–1131. [CrossRef]
- Pienaar, A.C.E.; Cillier, J.A. The isolation of a paramyxovirus from pigeons in South Africa. Onderstepoort J. Vet. Res. 1987, 54, 653-654.
- Ide, P.I. Virological studies of paramyxovirus type 1 infection of pigeons. Can. Vet. J. 1987, 28, 601-603.
- Bogoyavlenskiy, A.; Berezin, V.; Prilipov, A.; Usachev, E.; Korotetskiy, I.; Zaitceva, I., Kydyrmanov, A.; Sayatov, M. Characterization of pigeon paramyxoviruses (Newcastle disease virus) isolated in Kazakhstan in 2005. Virologica. Sinica. 2012, 27, 93-99. [CrossRef]
- Collins, M.S.; Strong, I.; Alexander, D.J. Evaluation of the molecular-basis of pathogenicity of the variant Newcastle-disease viruses termed “Pigeon Pmv-1 Viruses”. Arch. Virol. 1994, 134, 403–11. [CrossRef]
- Meulemans, G.; Berg, T.V.D.; Decaesstecker, M.; Boschmans, M. Evolution of pigeon Newcastle disease virus strains. Avian Pathol. 2002, 31, 515-519. [CrossRef]
- Dortmans, J.C.; Fuller, C.M.; Aldous, E.W.; Rottier, P.J.; Peeters, B.P. Two genetically closely related pigeon paramyxovirus type 1 (PPMV-1) variants with identical velogenic fusion protein cleavage sites but with strongly contrasting virulence. Vet. Microbiol. 2010, 143, 139–144. [CrossRef]
- Dortmans, J.C.F.M.; Rottier, P.J.M.; Koch, G.; Peeters, B.P.H. Passaging of a Newcastle disease virus pigeon variant in chickens results in selection of viruses with mutations in the polymerase complex enhancing virus replication and virulence. J. Gen. Virol. 2011, 92, 336–345. [CrossRef]
- Wang, J.J.; Liu, H.L.; Liu, W.; Zheng, D.X.; Zhao, Y.L.; Li, Y.; Wang,Y.; Ge, S.; Lv, Y.; Zuo, Y.; Yu, S.; Wang Z. Genomic characterizations of six pigeon paramyxovirus type 1 viruses isolated from live bird markets in China during 2011 to 2013. PLoS ONE. 2015, 10, e0124261. [CrossRef]
- Collins, M.S.; Strong, I.; Alexander, D.J. Pathogenicity and phylogenetic evaluation of the variant Newcastle disease viruses termed “pigeon PMV-1 viruses” based on the nucleotide sequence of the fusion protein gene. Arch. Virol. 1996, 141, 635–47. [CrossRef]
- Irvine, R.M.; Aldous, E.W.; Marvell, R.J.; Cox, W.J.; Ceeraz, V.; Fuller, C.M.; Alexander, D.J.; Brown, I.H.; Wood, A.M.; Milne, J.C.; Wilson, M.; Hepple, R.G.; Hurst, A.; Sharpe, C.E. Outbreak of Newcastle disease due to pigeon paramyxovirus type I in grey partridges (Perdix perdix) in Scotland in October 2006. Vet. Rec. 2009, 165, 531–535. [CrossRef]
- Aldous, E.W.; Fuller, C.M.; Mynn, J.K.; Alexander; D.J. A molecular epidemiological investigation of isolates of the variant avian paramyxovirus type 1 virus (PPMV-1) responsible for the 1978 to present panzootic in pigeons. Avian Pathol. 2004, 33, 258–269. [CrossRef]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: a review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Invest. 2011, 23, 637–656. [CrossRef]
- Mansour, S.M.; Mohamed, F.F.; Eid, A.A.; Mor, S.K.; Goyal, S.M. Co-circulation of paramyxo-and influenza viruses in pigeons in Egypt. Avian Pathol. 2017, 46, 367-375. [CrossRef]
- Toro, H.; Hoerr, F.J.; Farmer, K.; Dykstra, C.C.; Roberts, S.R.; Perdue, M. Pigeon paramyxovirus: association with common avian pathogens in chickens and serologic survey in wild birds. Avian Dis. 2005, 49, 92-98. [CrossRef]
- Liu, M.; Qu, Y.; Wang, F.; Liu, S.; Sun, H. Genotypic and pathotypic characterization of Newcastle disease virus isolated from racing pigeons in China. Poult. Sci. 2015, 94, 1476–1482. [CrossRef]
- He, Y.; Lu, B.; Dimitrov, K.M.; Liang, J.; Chen, Z.; Zhao, W.; Qin, Y.; Duan, Q.; Zhou, Y.; Liu, L.; Li, B. Complete genome sequencing, molecular epidemiological, and pathogenicity analysis of pigeon paramyxoviruses type 1 isolated in Guangxi, China during 2012–2018. Viruses 2020, 12, 366. [CrossRef]
- Xie, P.; Chen, L.; Zhang, Y.; Lin, Q.; Ding, C.; Liao, M.; Xu, C.; Xiang, B.; Ren, T. Evolutionary dynamics and age-dependent pathogenesis of sub-genotype VI. 2.1. 1.2. 2 PPMV-1 in pigeons. Viruses 2020, 12, 433. [CrossRef]
- Zhang, Y.; Wang, W.; Li, Y.; Liu, J.; Wang, W.; Bai, J.; Yang, Z.; Liu, H.; Xiao, S. A pigeon paramyxovirus type 1 isolated from racing pigeon as an inactivated vaccine candidate provides effective protection. Poult. Sci. 2022, 101, 102097. [CrossRef]
- Hamouda, E.E.; Mohamed, N.A.; Eid, A.A.; Ismail, A.N.; El-Sisi, M.A.; Hassanin, O.A.; Abou-Hashem, N.A. Utilization of RT-PCR and restriction enzyme analysis in detection and differentiation of pigeon paramyxovirus-1 and Newcastle disease virus in pigeons. Zagazig Vet. J. 2017, 45, 37-47. [CrossRef]
- WOAH Terrestrial Manual. Newcastle Disease (Infection with Newcastle Disease Virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; WOAH: Paris, France, 2021; Chapter 3.3.14. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.14_NEWCASTLE_DIS.pdf (accessed on 15 March 2022).
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497.
- Alexander, D.J. Newcastle disease other avian para-myxoviruses. In A Laboratory Manual for the Isolation, Identification Characterization of Avian Pathogens, 4th ed.; Swayne, D.E., Glisson, J.R., Jackwood, M.W., Pearson, J.E., Reed, W.M., Eds.; American Association of Avian Pathologists: Kenneth Square, IA, 1998; pp. 156–163.
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle disease, other avian paramyxoviruses, and avian metapneumovirus infections. In Diseases of poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., de Witit, S., Grimes, T., Johnson, D., Kromm, M., Prajitno, T.Y., Rubinoff, I., Zavala, G. Eds., Wiley-Blackwell: Hoboken, NJ, 2019; pp. 109–166.
- Sabra, M.; Dimitrov, K.M.; Goraichuk, I.V.; Wajid, A.; Sharma, P.; Williams-Coplin, D.; Basharat, A.; Rehmani, S.F.; Muzyka, D.V.; Miller, P.J.; Afonso, C.L. Phylogenetic assessment reveals continuous evolution and circulation of pigeon-derived virulent avian avulaviruses 1 in Eastern Europe, Asia, and Africa. BMC Vet. Res. 2017, 13, 1-13. [CrossRef]
- Fuller, C.M.; Collins, M.S.; Alexander, D.J. Development of a real-time reverse-transcription PCR for the detection and simultaneous pathotyping of Newcastle disease virus isolates using a novel probe. Arch. Virol. 2009, 154, 929-937. [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D.. Bancroft’s theory and practice of histological techniques, 8th ed. Elsevier Health Sciences: England, 2018.
- Samiullah, M.; Rizvi, F.; Anjum, A.D.; Shah, M.F.A. Rising hyperimmune serum against avian paramyxovirus (APMV-1) and pigeon paramyxovirus (PPMV-1) in rabbits and their cross-reactivity. Pak. J. Biol. Sci. 2006, 9, 2184-2186. [CrossRef]
- Burns, R. Immunochemical protocols, 3rd ed.; Humana Press: Totowa, NJ, 2005.
- Dunn, P.K.; Smyth, G.K. Positive Continuous Data: Gamma and Inverse Gaussian GLMs, In Generalized Linear Models with Examples in R, Dunn, P.K., Smyth, G.K., Eds.; Springer New York: New York, NY, 2018; pp. 425-456.
- Xiang, B.; You, R.; Kang, Y.; Xie, P.; Zhu, W.; Sun, M.; Gao, P.; Li, Y.; Ren, T. Host immune responses of pigeons infected with Newcastle disease viruses isolated from pigeons. Microb. Pathog. 2019, 127, 131-137. [CrossRef]
- Zhan, T.; Hu, S.; Chen, Y.; Lu, X.; Liao, T.; Guo, L.; Gao, X.; Xu, X.; Hu, Z.; Liu, X. Comparative pathogenicity of two closely related Newcastle disease virus isolates from chicken and pigeon respectively. Virus Res. 2020, 286, 198091. [CrossRef]
- Qiu, X.; Meng, C.; Zhan, Y.; Yu, S.; Li, S.; Ren, T.; Yuan, W.; Xu, S.; Sun, Y.; Tan, L.; Song, C. Phylogenetic, antigenic and biological characterization of pigeon paramyxovirus type 1 circulating in China. Virol. J. 2017, 14, 1-13. [CrossRef]
- Esmaeelzadeh Dizaji, R.; Ghalyanchi Langeroudi, A.; Vasfi Marandi, M.; Hosseini, H.; Karimi, V.; Molouki, A. Characterization of Pigeon Paramyxovirus Type 1 Viruses (PPMV-1) Isolated in Iran. Iran. J. Virol. 2020, 14, 12-20.
- Chang, H.; Feng, S.; Wang, Y.; Li, F.; Su, Q.; Wang, B.; Du, J.; He, H. Isolation and pathogenic characterization of pigeon paramyxovirus type 1 via different inoculation routes in pigeons. Front. Vet. Sci. 2021, 7, 569901. [CrossRef]
- Zhan, T.; He, D.; Lu, X.; Liao, T.; Wang, W.; Chen, Q.; Liu, X.; Gu, M.; Wang, X.; Hu, S.; Liu, X. Biological characterization and evolutionary dynamics of pigeon paramyxovirus type 1 in China. Front. Vet. Sci. 2021, 8, 721102. [CrossRef]
- Zhan, T.; Lu, X.; He, D.; Gao, X.; Chen, Y.; Hu, Z.; Wang, X.; Hu, S.; Liu, X. Phylogenetic analysis and pathogenicity assessment of pigeon paramyxovirus type 1 circulating in China during 2007–2019. Transbound. Emerg. Dis. 2022, 69, 2076-2088. [CrossRef]
- Zhang, Y.; Ren, S.; Yuan, X.; Yang, J.; Ai, W.; Meng, K. Biological characterization of isolated pigeon Paramyxovirus-1 and its pathogenicity analysis in SPF chickens. Vet. Arh. 2023, 93, 65-70. [CrossRef]
- Dortmans, J.C.; Koch, G.; Rottier, P.J.; Peeters, B.P. Virulence of pigeon paramyxovirus type 1 does not always correlate with the cleavability of its fusion protein. J. Gen. Virol. 2009, 90, 2746–2750.
- Flint, S.J.; Enquist, L.W.; Racaniello, V.R.; Skalka, A.M. Principles of virology, molecular biology, pathogenesis and control of animal viruses. ASM press: Washington, DC, USA, 2004; pp. 126 –176.
- Rahman, S.; Nizamani, Z.A.; Soomro, N.M.; Kalhoro, N.H.; Rasool, F. Velogenic viscerotropic Newcastle disease virus produces variable pathogenicity in two chicken breeds. J. Anim. Health Prod. 2014, 2, 46-50. [CrossRef]
- Amer, M.I.S.; El-Bagoury, G.F.; Khodeir, M.H. Evaluation of the immune response of pigeons to Newcastle disease and pigeon paramyxo virus vaccines. Benha Vet. Med. J. 2013, 24, 148-156.
- Wang, F.; Gao, M.; Han, Z.; Hou, Y.; Zhang, L.; Ma, Z.; Ma, D. Innate immune responses of domestic pigeons to the infection of pigeon paramyxovirus type 1 virus. Poult. Sci. 2021, 100, 603–614. [CrossRef]
- Shalaby, S.M.; Awadin, W.F.; Hamed, M.F.; El-Tholoth, M.; Ibrahim, I.; El-Shaieb, A.F. Pathological and ultrastructural characteristics of Newcastle and pox diseases in naturally infected pigeons in Egypt. Adv. Anim. Vet. Sci. 2021, 9, 1995-2004. [CrossRef]
- Isidoro-Ayza, M.; Afonso, C.L.; Stanton, J.B.; Knowles, S.; Ip, H.S.; White, C.L.; Fenton, H.; Ruder, M.G.; Dolinski, A.C.; Lankton, J. Natural infections with pigeon paramyxovirus serotype 1: pathologic changes in Eurasian collared-doves (Strep-topelia decaocto) and rock pigeons (Columba livia) in the United States. Vet. Pathol. 2017, 54, 695–703. [CrossRef]
- Al-Alhially, A.A.; Al-Hamdany, E.K.; Ismail, H.K. Investigation of histopathological alteration of paramyxovirus-1 in naturally infected racing pigeons. Iraqi J. Vet. Sci. 2024, 38, 309-316. [CrossRef]
- Isenmann, R.; Henne-Bruns, D.; Adler, G. Shock and acute pancreatitis. Best. Pract. Res. Clin. Gastroenterol. 2003, 17, 345–355. [CrossRef]
- Barton, J.T.; Bickford, A.A.; Cooper, G.L.; Charlton, B.R.; Cardona, C.J. Avian paramyxovirus type 1 infections in racing pigeons in California, I: clinical signs, pathology, and serology. Avian Dis. 1992, 36, 463–468. [CrossRef]
- Johnston, K.M.; Key, D.W. Paramyxovirus-1 in feral pigeons (Columba livia) in Ontario. Can. Vet. J. 1992, 33, 796–800.
- Stone, H.D. Efficacy of oil-emulsion vaccines prepared with pigeon paramyxovirus-1, Ulster, and La Sota Newcastle disease viruses. Avian Dis. 1989, 157-162. [CrossRef]
- Hassan, E.A. Studies on Protection of Pigeon Squabs against Infection with Pigeon Paramyxovirus-L Using Different Strains of Newcastle Disease Vaccine in the Period Before The Age of Vaccination with PMV-1 Vaccine. Egyptian J. Agric. Res. 2005, 83, 1317-1324. [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447-453. [CrossRef]
- Wang, X.; Ren, S.; Wang, X.; Wang, C.; Fan, M.; Jia, Y.; Gao, X.; Liu, H.; Xiao, S.; Yang. Z. Genomic characterization of a wild-bird-origin pigeon paramyxovirus type 1 (PPMV-1) first isolated in the northwest region of China. Arch. Virol. 2017, 162, 749–761. [CrossRef]
- Alexander, D.J.; Manvell1, R.J.; Kemp, P.A.; Parsons, G.; Collins, M.S.; Brockman, S.; Russell, P.H.; Lister, S.A. Use of monoclonal antibodies in the characterisation of avian paramyxovirus type 1 (Newcastle disease virus) isolates submitted to an international reference laboratory. Avian Pathol. 1987, 16, 553–565. [CrossRef]
- Meulemans, G.; Letellier, C.; Gonze, M.; Carlier, M.C.; Burny, A. Newcastle disease virus F glycoprotein expressed from a recombinant vaccinia virus vector protects chickens against live-virus challenge. Avian Pathol. 1988, 17, 821-7. [CrossRef] [PubMed]
- Ye, H.; Qiu, W.; Li, M.; Liang, Z.; Wang, X.; Luo, K. Development and immune efficacy of Newcastle disease virus vaccine for pigeon. China Poult. 2012, 34, 20–24.
- Zhang Rui, Z.R.; Pu Juan, P.J.; Su JingLiang, S.J.; Zhao JiXun, Z.J.; Wang XiaoTing, W.X.; Zhang ShouPing, Z.S.; Li XiaoJiao, L.X.; Zhang GuoZhong, Z.G. Phylogenetic characterization of Newcastle disease virus isolated in the mainland of China during 2001-2009. Vet. Microbiol. 2010, 141, 246–257.
- Alexander, D.J. Newcastle disease and other avian paramyxovirus infections, In Diseases of poultry, 9th ed.; Calnek, B.W., Barnes, H.J., Beard, C.W., Reid, W.M., Yoder Jr, H.W., Eds.; Iowa State University Press: Ames, 1991; pp. 496-519.
Figure 1.
Summary of experimental design: A) Design of biological characterization. B) Design of vaccines assessment against challenged with field strain of PPMV-1 "Pigeon/Egypt/ Sharkia-19/2015/KX580988". G1a, G2a, G3a, and G4a were kept as control groups to monitor any natural infection exposure from the environment to these pigeons. G1b, G2b, G3b, and G4b were challenged with PPMV-1 strain.
Figure 1.
Summary of experimental design: A) Design of biological characterization. B) Design of vaccines assessment against challenged with field strain of PPMV-1 "Pigeon/Egypt/ Sharkia-19/2015/KX580988". G1a, G2a, G3a, and G4a were kept as control groups to monitor any natural infection exposure from the environment to these pigeons. G1b, G2b, G3b, and G4b were challenged with PPMV-1 strain.
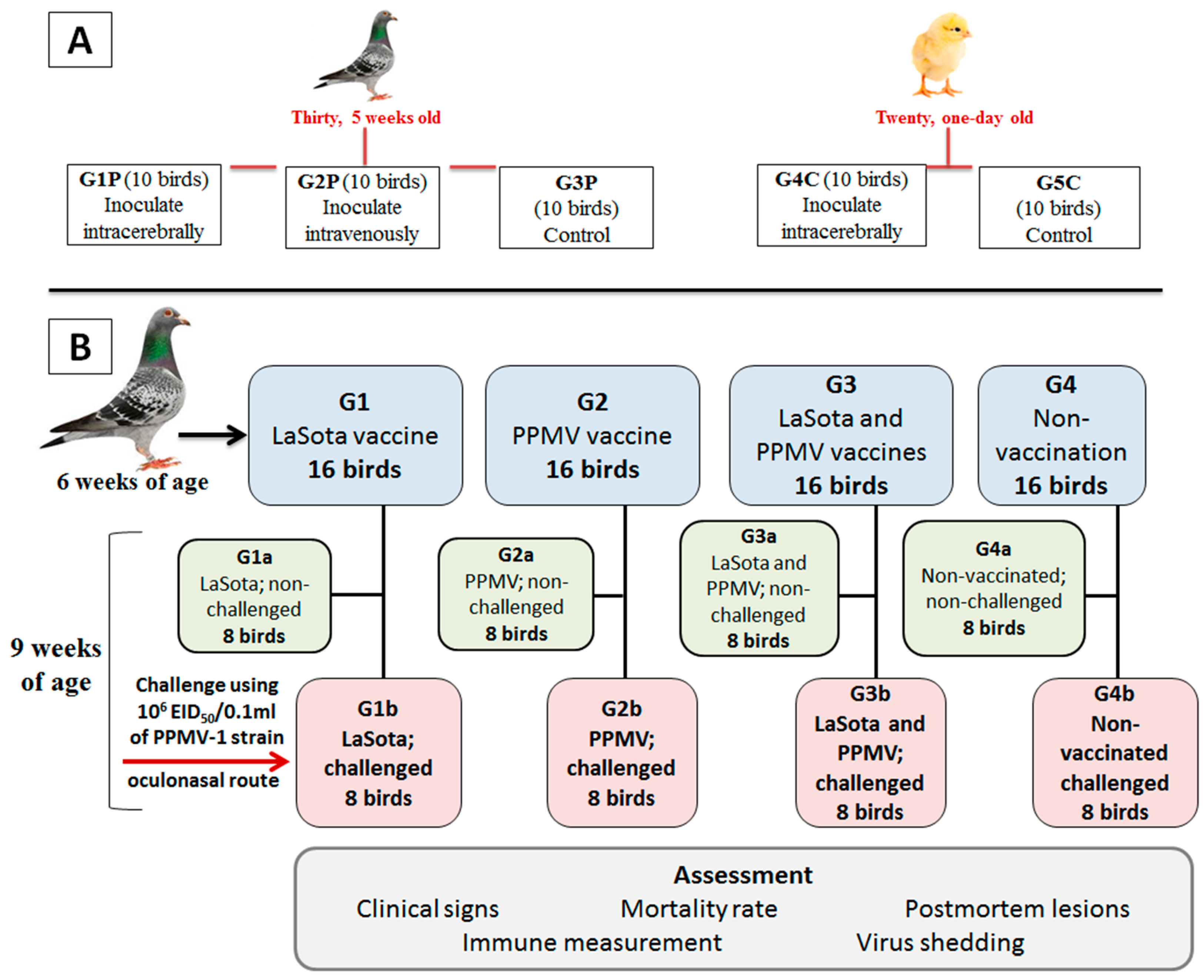
Figure 2.
Virulence (ICPI, and IVPI) assessment of the PPMV-1 strain (Pigeon/ Egypt/Sharkia-19/2015/KX580988) in pigeons and chickens. The birds were inoculated via intracerebral (I/C) and intravenous (I/V) routes to 1-day-old chicks and 5-week-old pigeons. The clinical scores were recorded until 10 days post infection (dpi). The clinical scores are represented as healthy (green color), displaying variable clinical signs, such as ruffled feather, depression, anorexia, and greenish diarrhea (light blue color, except nervous signs with (dark blue color)), and dead (red color).
Figure 2.
Virulence (ICPI, and IVPI) assessment of the PPMV-1 strain (Pigeon/ Egypt/Sharkia-19/2015/KX580988) in pigeons and chickens. The birds were inoculated via intracerebral (I/C) and intravenous (I/V) routes to 1-day-old chicks and 5-week-old pigeons. The clinical scores were recorded until 10 days post infection (dpi). The clinical scores are represented as healthy (green color), displaying variable clinical signs, such as ruffled feather, depression, anorexia, and greenish diarrhea (light blue color, except nervous signs with (dark blue color)), and dead (red color).
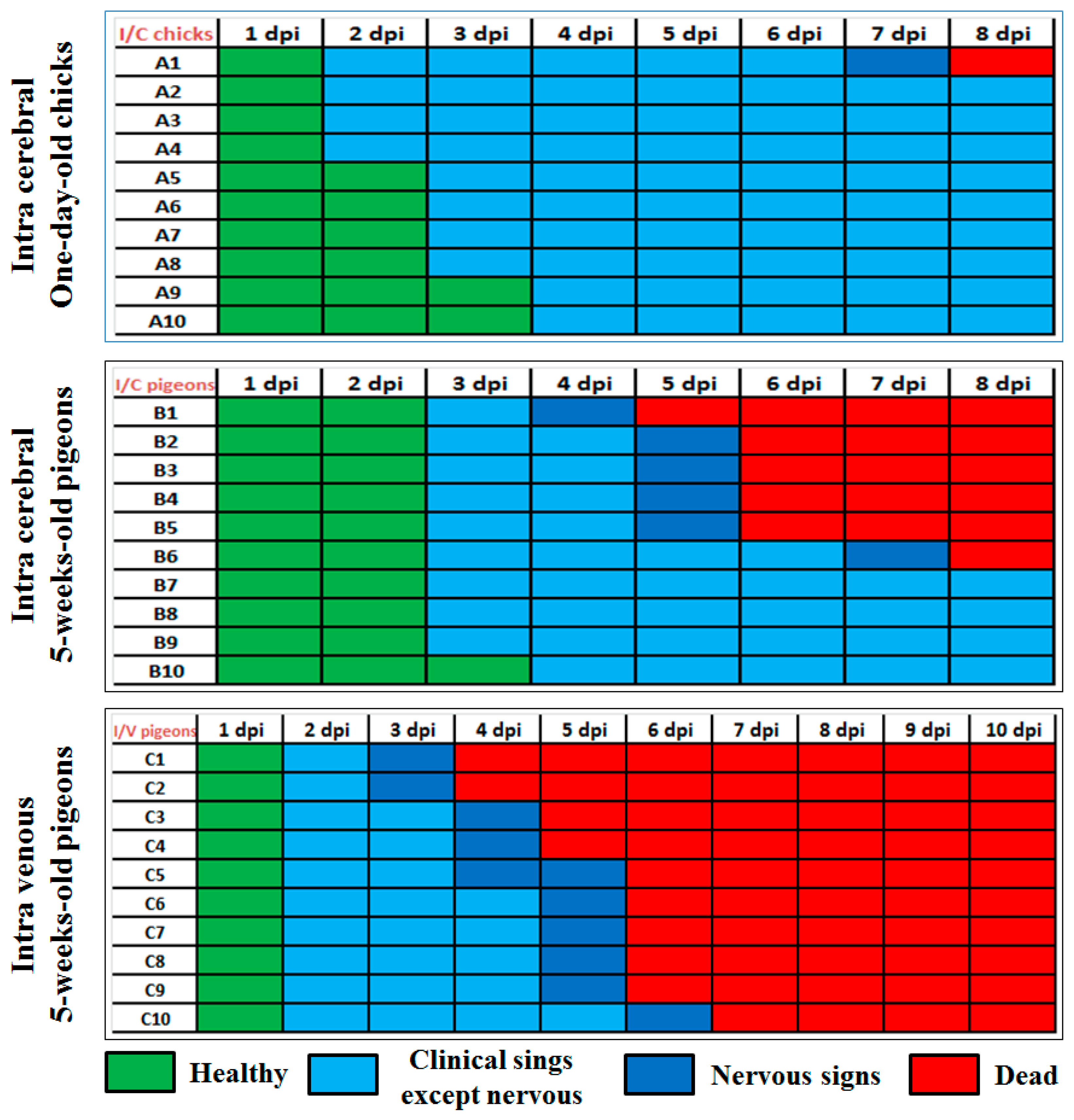
Figure 3.
Survival rates and clinical indices. a) Survival curves of vaccinated pigeons with tested vaccines post-challenge. b) Clinical indices of vaccinated pigeons with tested vaccines post-challenge. G1b: vaccinated with NDV LaSota and challenged, G2b: vaccinated with PPMV and challenged, G3b: dual vaccinated with LaSota and PPMV and challenged, G4a: non-vaccinated non-challenged, G4b: non-vaccinated challenged.
Figure 3.
Survival rates and clinical indices. a) Survival curves of vaccinated pigeons with tested vaccines post-challenge. b) Clinical indices of vaccinated pigeons with tested vaccines post-challenge. G1b: vaccinated with NDV LaSota and challenged, G2b: vaccinated with PPMV and challenged, G3b: dual vaccinated with LaSota and PPMV and challenged, G4a: non-vaccinated non-challenged, G4b: non-vaccinated challenged.
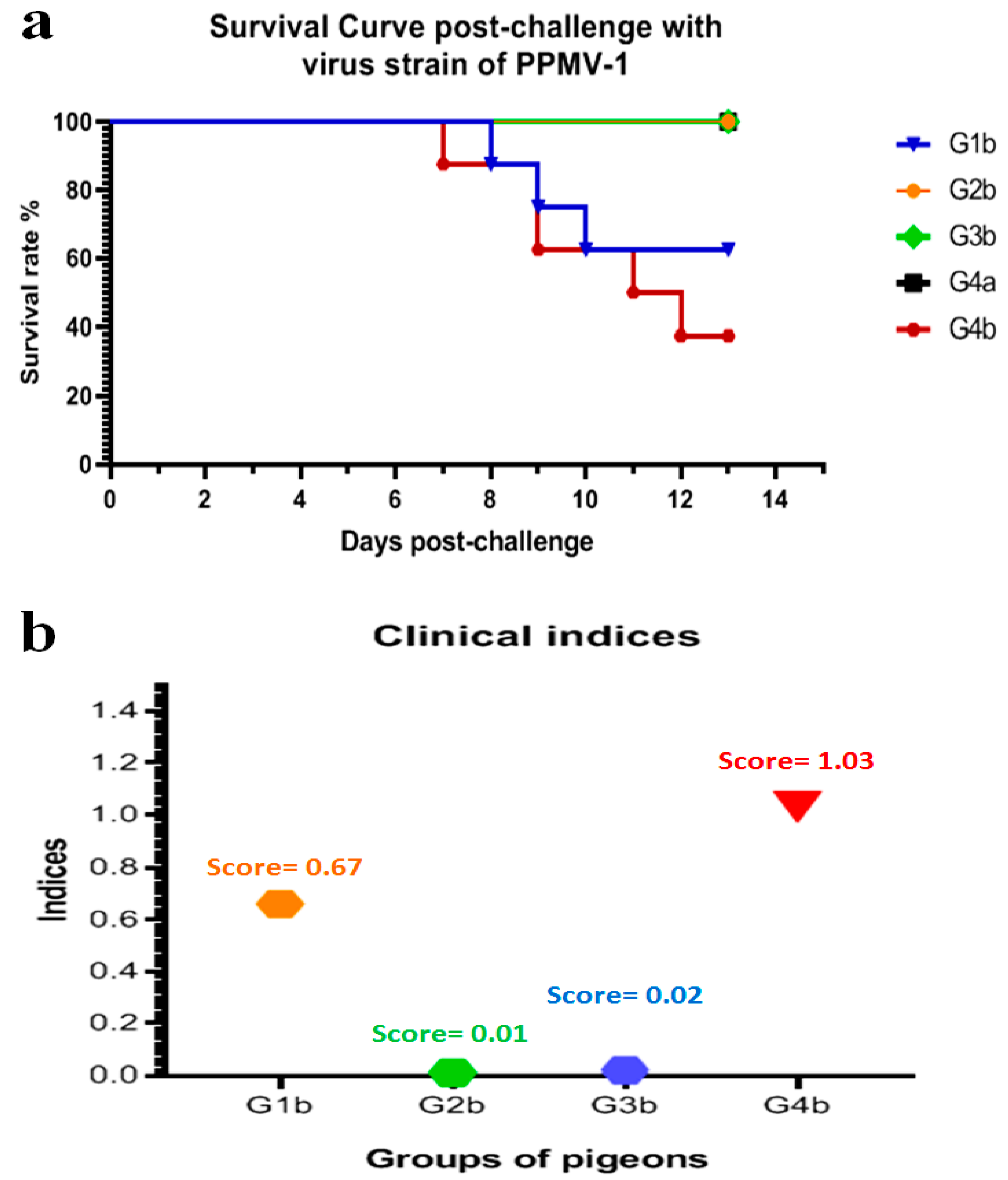
Figure 4.
Clinical and post-mortem findings of pigeons infected/challenged with PPMV-1 strain. A) Torticollis in non-vaccinated challenged pigeon (G4b). B) Wing paralysis in non-vaccinated challenged pigeon (G4b). C) Complete paralysis in non-vaccinated challenged pigeon (G4b). D) Severe greenish diarrhea in non-vaccinated challenged pigeon (G4b). E) Opisthotonus position in pigeon of NDV LaSota group (G1b). F) Greenish diarrhea in pigeon of NDV LaSota group (G1b). G) Severe congested and hemorrhagic brain in non-vaccinated challenged pigeon (G4b). H) Congested heart with minute petechial hemorrhages in non-vaccinated challenged pigeon (G4b). I) Enlarged and severe congested spleen with hemorrhages in non-vaccinated challenged pigeon (G4b). J) Pancreatitis with punctate hemorrhages in non-vaccinated challenged pigeon (G4b). K) Severe nephrosis in non-vaccinated challenged pigeon (G4b). L) Hemorrhages at junction between esophagus and proventriculus with greenish content in the gizzard of non-vaccinated challenged pigeon (G4b). M) Moderate congestion in brain of pigeon in NDV LaSota group (G1b). N) Mild congestion (apparently normal) of heart in pigeon of NDV LaSota group (G1b). O) Enlarged spleen with few hemorrhages in pigeon of NDV LaSota group (G1b). P) Pancreatitis with necrosis in pigeon of NDV LaSota group (G1b). Q) Moderate nephrosis in pigeon of NDV LaSota group (G1b).
Figure 4.
Clinical and post-mortem findings of pigeons infected/challenged with PPMV-1 strain. A) Torticollis in non-vaccinated challenged pigeon (G4b). B) Wing paralysis in non-vaccinated challenged pigeon (G4b). C) Complete paralysis in non-vaccinated challenged pigeon (G4b). D) Severe greenish diarrhea in non-vaccinated challenged pigeon (G4b). E) Opisthotonus position in pigeon of NDV LaSota group (G1b). F) Greenish diarrhea in pigeon of NDV LaSota group (G1b). G) Severe congested and hemorrhagic brain in non-vaccinated challenged pigeon (G4b). H) Congested heart with minute petechial hemorrhages in non-vaccinated challenged pigeon (G4b). I) Enlarged and severe congested spleen with hemorrhages in non-vaccinated challenged pigeon (G4b). J) Pancreatitis with punctate hemorrhages in non-vaccinated challenged pigeon (G4b). K) Severe nephrosis in non-vaccinated challenged pigeon (G4b). L) Hemorrhages at junction between esophagus and proventriculus with greenish content in the gizzard of non-vaccinated challenged pigeon (G4b). M) Moderate congestion in brain of pigeon in NDV LaSota group (G1b). N) Mild congestion (apparently normal) of heart in pigeon of NDV LaSota group (G1b). O) Enlarged spleen with few hemorrhages in pigeon of NDV LaSota group (G1b). P) Pancreatitis with necrosis in pigeon of NDV LaSota group (G1b). Q) Moderate nephrosis in pigeon of NDV LaSota group (G1b).
Figure 5.
Histopathological lesions in different tissue organs of nin-vaccinated infected pigeons with PPMV-1 strain, H&E X400, Bar 50um. Brain: a) Congestion of brain capillaries (long arrows). b) Sub meningeal hemorrhages (H). c) Multifocal degeneration and swelling of neurons gliosis (arrow heads), degenerated neurons (short arrows). Pancreas: d) Severe congestion (stars). e) Multifocal hemorrhages (H) with degeneration and necrosis of pancreatic acini (N). f) Multifocal aggregations of lymphocytic infiltrations (arrow head). Spleen: g) Severe congestion of sinusoids and splenic blood vessels (stars). h) Multifocal hemorrhages (H), severe depletion of subcapsular lymphoid follicles (long arrows). i) Severe depletion of subcapsular lymphoid follicles (long arrows). j) Liver; severe congestion, focal degeneration (long arrow) and discrete necrosis of some hepatic cells, along with mild leukocytes around bile duct (arrow head). k) Kidney; intertubular hemorrhages (H), degeneration of renal tubular epithelium (long arrow) and focal necrosis (N) with leukocyte infiltration (arrow head). l) Heart; Inter-muscular congestion (long arrows), and hemorrhages (H).
Figure 5.
Histopathological lesions in different tissue organs of nin-vaccinated infected pigeons with PPMV-1 strain, H&E X400, Bar 50um. Brain: a) Congestion of brain capillaries (long arrows). b) Sub meningeal hemorrhages (H). c) Multifocal degeneration and swelling of neurons gliosis (arrow heads), degenerated neurons (short arrows). Pancreas: d) Severe congestion (stars). e) Multifocal hemorrhages (H) with degeneration and necrosis of pancreatic acini (N). f) Multifocal aggregations of lymphocytic infiltrations (arrow head). Spleen: g) Severe congestion of sinusoids and splenic blood vessels (stars). h) Multifocal hemorrhages (H), severe depletion of subcapsular lymphoid follicles (long arrows). i) Severe depletion of subcapsular lymphoid follicles (long arrows). j) Liver; severe congestion, focal degeneration (long arrow) and discrete necrosis of some hepatic cells, along with mild leukocytes around bile duct (arrow head). k) Kidney; intertubular hemorrhages (H), degeneration of renal tubular epithelium (long arrow) and focal necrosis (N) with leukocyte infiltration (arrow head). l) Heart; Inter-muscular congestion (long arrows), and hemorrhages (H).
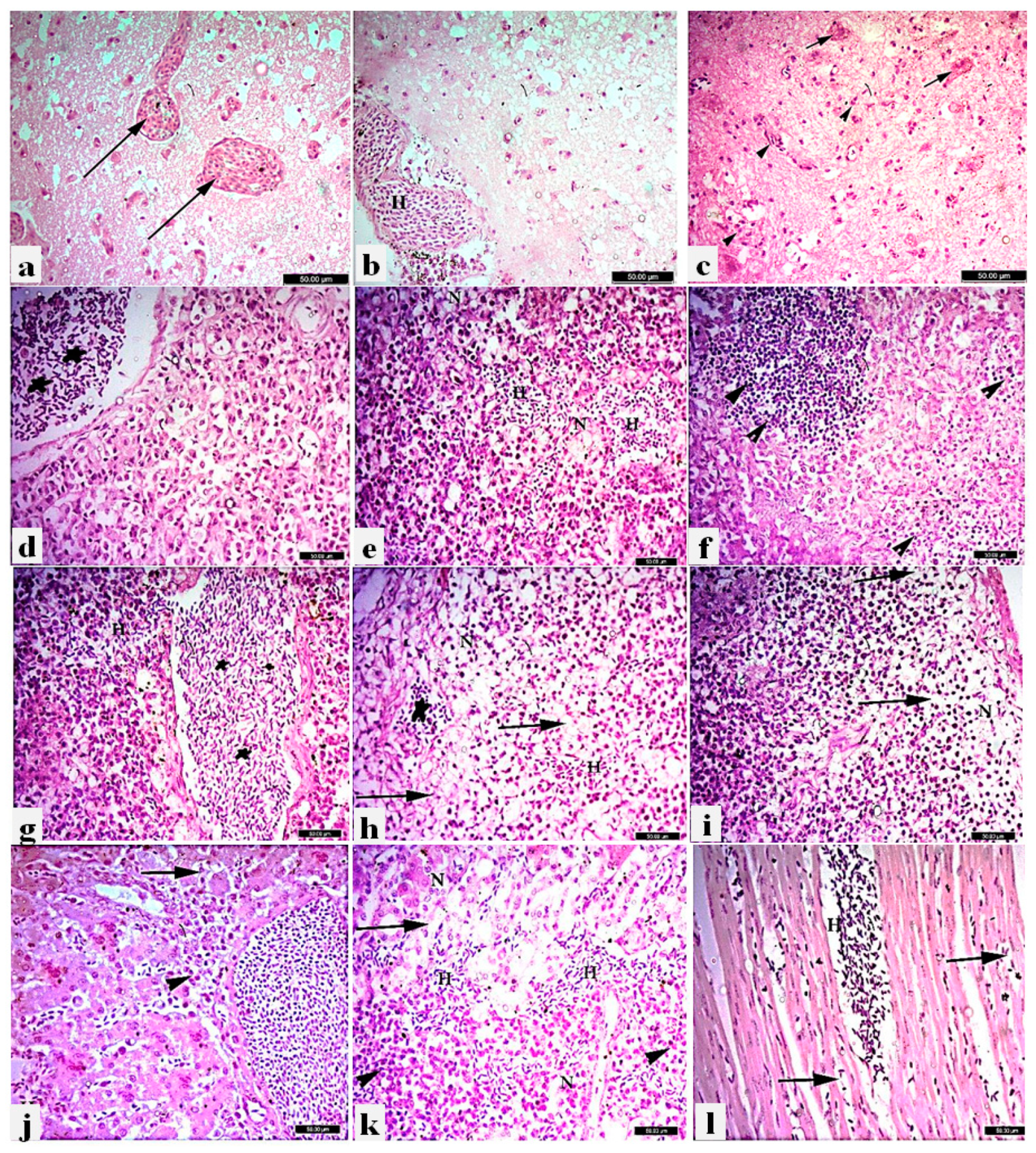
Figure 6.
Photomicrograph of immunohistochemistry (IHC) in different tissue organs of non-vaccinated and infected pigeons with PPMV-1 strain, Bar 25um. a) Brain showing positive expression of viral antigen in the degenerated neurons. b) Trachea showing positive expression of viral antigen in the tracheal epithelium and in the inflammatory cells in the propria submucosa. c) Lung showing positive expression of viral antigen in air capillaries. d) Pancreas showing positive expression of viral antigen in the exocrine epithelial cells. e) Spleen showing positive expression of viral antigen in the splenic parenchyma. f) Heart showing positive expression of viral antigen in the cardiac muscle. g) Proventriculus showing positive expression of viral antigen in the glandular epithelium and desquamated epithelial cells. h) Intestine showing positive expression of intestinal mucosa. i) Kidney showing positive expression of viral antigen in the renal tubular epithelium.
Figure 6.
Photomicrograph of immunohistochemistry (IHC) in different tissue organs of non-vaccinated and infected pigeons with PPMV-1 strain, Bar 25um. a) Brain showing positive expression of viral antigen in the degenerated neurons. b) Trachea showing positive expression of viral antigen in the tracheal epithelium and in the inflammatory cells in the propria submucosa. c) Lung showing positive expression of viral antigen in air capillaries. d) Pancreas showing positive expression of viral antigen in the exocrine epithelial cells. e) Spleen showing positive expression of viral antigen in the splenic parenchyma. f) Heart showing positive expression of viral antigen in the cardiac muscle. g) Proventriculus showing positive expression of viral antigen in the glandular epithelium and desquamated epithelial cells. h) Intestine showing positive expression of intestinal mucosa. i) Kidney showing positive expression of viral antigen in the renal tubular epithelium.
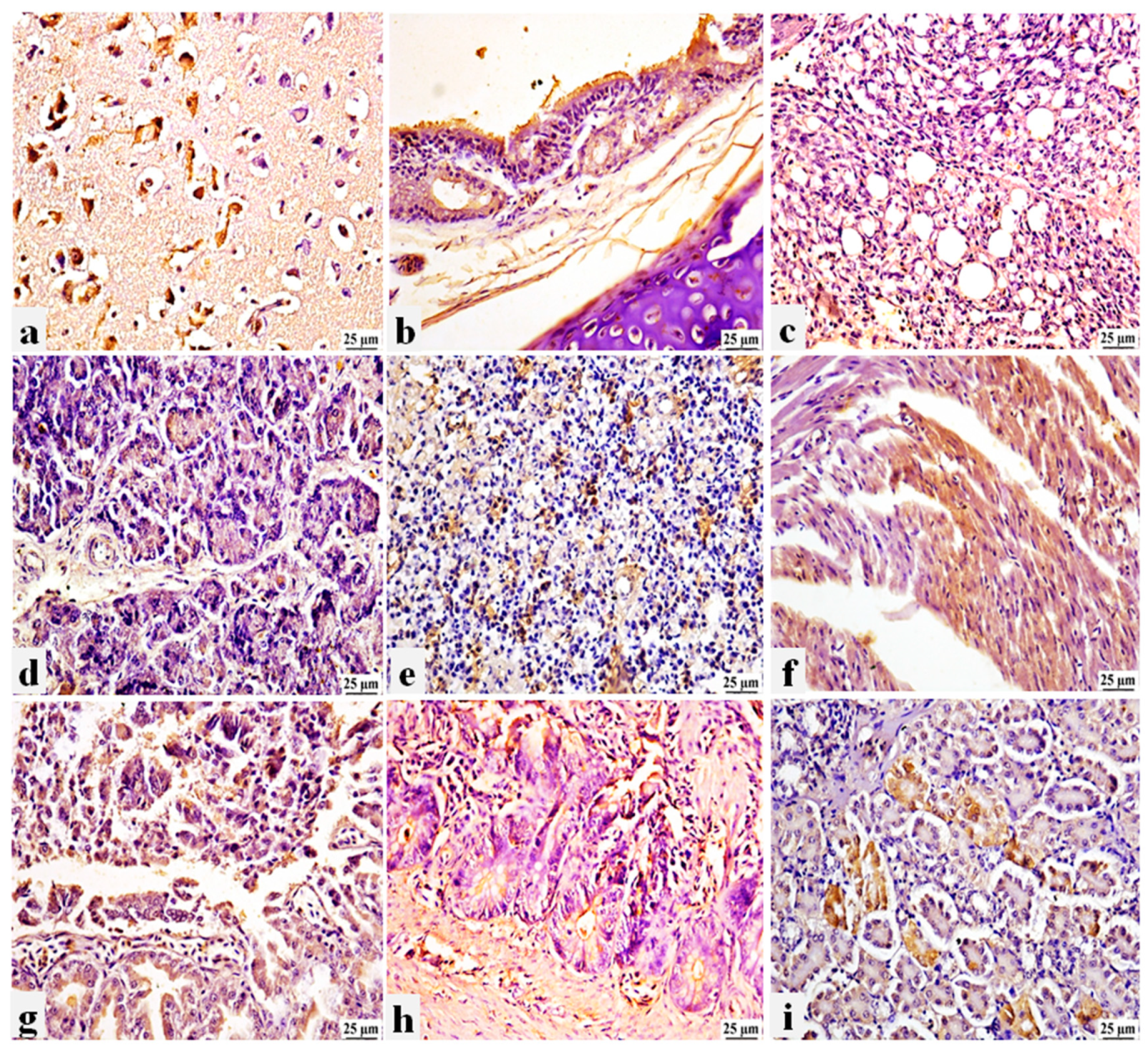
Figure 7.
a) Density plot showing the positive skewness in the HI titer distribution. b) Model selection criteria for linear and generalized linear designs based on Akaike Information Criterion (AIC).
Figure 7.
a) Density plot showing the positive skewness in the HI titer distribution. b) Model selection criteria for linear and generalized linear designs based on Akaike Information Criterion (AIC).
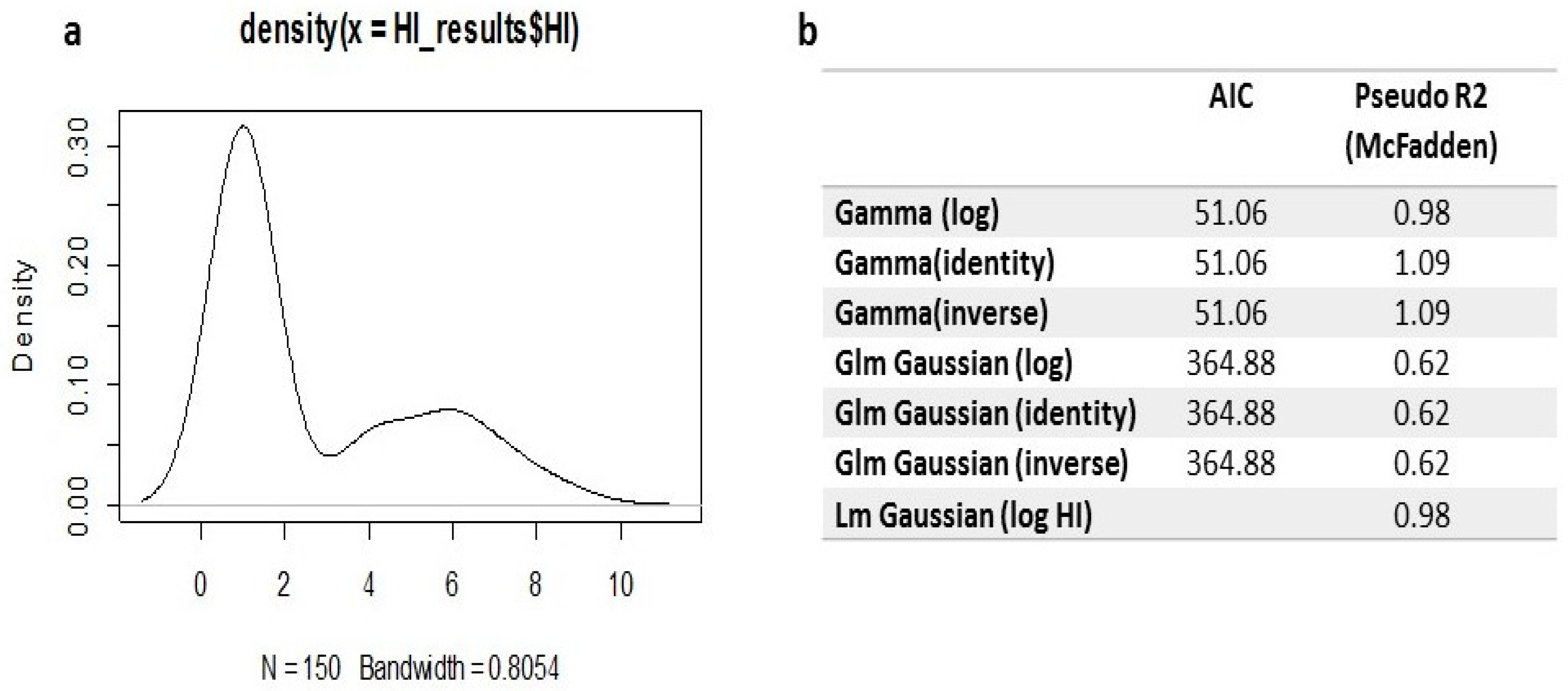
Figure 8.
a) The GLM model estimates with 95% confidence interval; *** representing the significance of the model coefficient estimates. b) Predicted values of the GLM for HI titers to detect the difference between using LaSota as heterologous antigen (antigen 1) and PPMV-1 as homologous antigen (antigen 2) within the 5 groups. c) Predicted values of the GLM for HI titers to detect the difference between the 5 groups within NDV LaSota (antigen 1) and PPMV-1 (antigen 2) at consecutive 5 weeks.
Figure 8.
a) The GLM model estimates with 95% confidence interval; *** representing the significance of the model coefficient estimates. b) Predicted values of the GLM for HI titers to detect the difference between using LaSota as heterologous antigen (antigen 1) and PPMV-1 as homologous antigen (antigen 2) within the 5 groups. c) Predicted values of the GLM for HI titers to detect the difference between the 5 groups within NDV LaSota (antigen 1) and PPMV-1 (antigen 2) at consecutive 5 weeks.
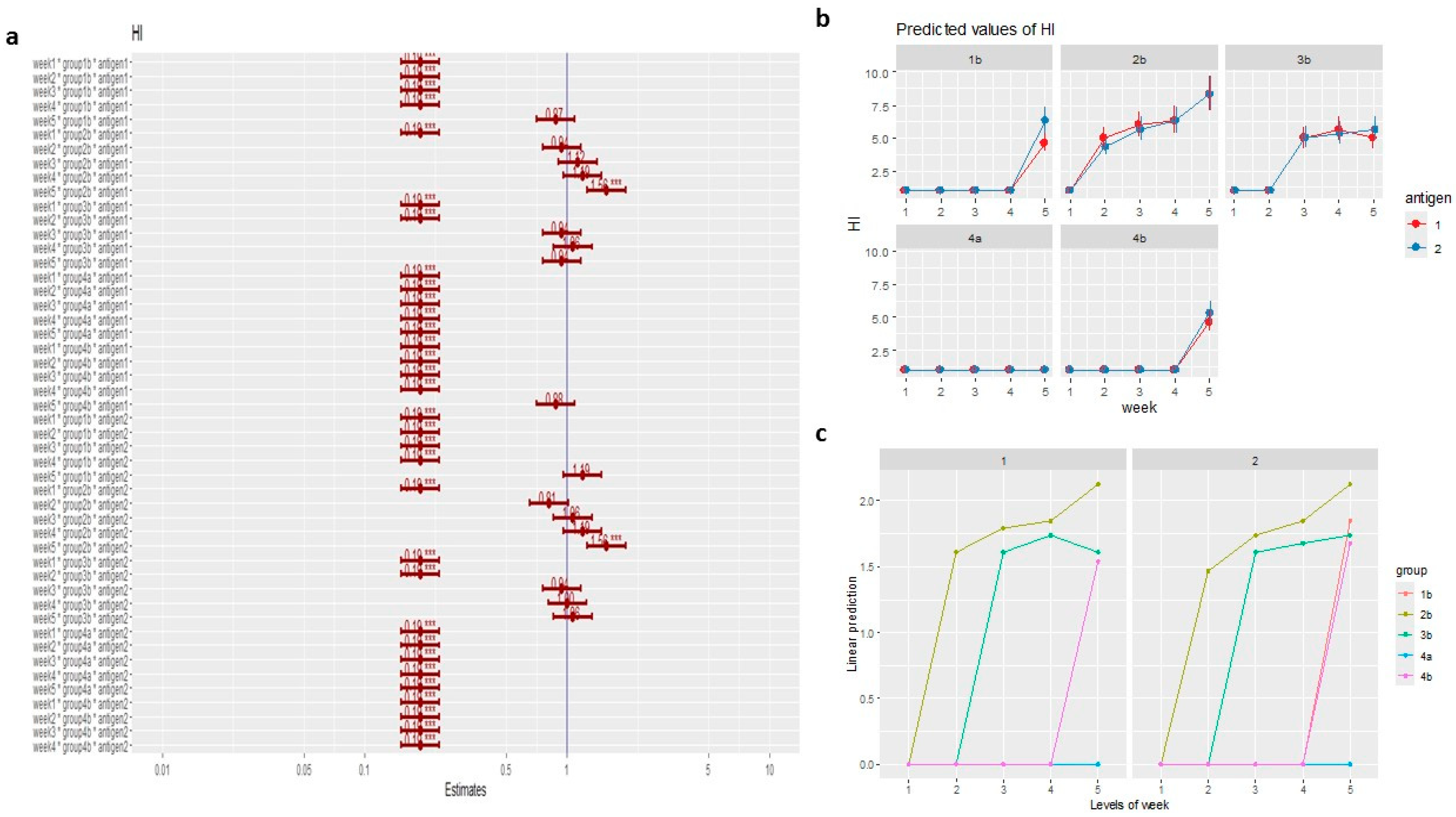

Table 1.
General and specific clinical signs, morbidities and mortalities of differentially PPMV-1 virus−vaccinated groups 14 days post-challenge, with clarification of the healthy condition in the vaccinated-non-challenged groups.
Table 1.
General and specific clinical signs, morbidities and mortalities of differentially PPMV-1 virus−vaccinated groups 14 days post-challenge, with clarification of the healthy condition in the vaccinated-non-challenged groups.
| Groups | Subgroups | General signs * | Respiratory Signs | Greenish diarrhea | Nervous signs | Morbidity number (%) | Clinical indices ** | Mortality number (%) |
|---|---|---|---|---|---|---|---|---|
| G1 | A | 0/8 | 0/8 | 0/8 | 0/8 | 0/8 (0) | 0 | 0/8 (0) |
| B | 5/8 | 0/8 | 2/8 | 2/8 | 5/8 (62.5) | 0.66 | 3/8 (37.5) | |
| G2 | A | 0/8 | 0/8 | 0/8 | 0/8 | 0/8 (0) | 0 | 0/8 (0) |
| B | 1/8 | 0/8 | 1/8 | 0/8 | 1/8 (12.5) | 0.01 | 0/8 (0) | |
| G3 | A | 0/8 | 0/8 | 0/8 | 0/8 | 0/8 (0) | 0 | 0/8 (0) |
| B | 1/8 | 0/8 | 0/8 | 1/8 | 1/8 (12.5) | 0.02 | 0/8 (0) | |
| G4 | A | 0/8 | 0/8 | 0/8 | 0/8 | 0/8 (0) | 0 | 0/8 (0) |
| B | 8/8 | 1/8 | 5/8 | 6/8 | 8/8 (100) | 1.03 | 5/8 (62.5) |
(*) General signs mean ruffled feathers, lethargy, and anorexia, (**) Clinical indices were calculated as previously mentioned in the materials Subgroups (G1a, G2a, G3a) were vaccinated with live NDV LaSota, inactivated PPMV, and simultaneously NDV LaSota and PPMV vaccines, respectively without challenge. Whereas, subgroups (G1b, G2b, G3b) were vaccinated with live NDV LaSota, inactivated PPMV, and simultaneously NDV LaSota and PPMV vaccines, respectively, and challenged with PPMV-1 strain. However, G4a and G4b were kept non-vaccinated non-challenged and challenged, respectively.
Table 2.
Pathological lesions among the dead and sacrificed morbid pigeons of differentially PPMV-1 virus−vaccinated groups post-challenge with unvaccinated challenged group.
Table 2.
Pathological lesions among the dead and sacrificed morbid pigeons of differentially PPMV-1 virus−vaccinated groups post-challenge with unvaccinated challenged group.
| Sub-groups | Total No. | No. of dead and euthanized morbid pigeons | Congested/ hemorrhagic brain | Enlarged/ atrophied spleen | Pancreas with hemorrhages/ necrosis | Hemorrhages on heart | Hemorrhages between esophagus and proventriculus | Enteritis with greenish content | Kidneys | Atrophy thymus | Atrophy of Bursa of fabricius |
|---|---|---|---|---|---|---|---|---|---|---|---|
| G1b | 8 | 5 | 4 Congested |
3 | 3 (1 hemorrhagic, 2 necrosis) |
0 | 1 | 3 | 3 (1 nephritis, 2 nephrosis) |
3 | 3 |
| G2b | 8 | 1 | 0 | 0 | 1 Necrosis |
0 | 0 | 1 | 0 | 0 | 0 |
| G3b | 8 | 1 | 1 Congested |
0 | 1 Necrosis |
0 | 0 | 0 | 1 nephrosis |
0 | 0 |
| G4b | 8 | 8 | 8 congested and hemorrhagic |
5 | 8 (6 hemorrhagic, 2 necrosis) |
2 | 2 | 8 | 7 (2 nephritis, 5 nephrosis) |
5 | 4 |
Subgroups (G1b, G2b, G3b) were vaccinated with live NDV LaSota, inactivated PPMV, and dual LaSota and PPMV vaccines, respectively, and challenged with PPMV-1 strain. However, G4b were kept unvaccinated- challenged.
Table 3.
Estimated marginal means ± SEM for the HI titer values over 5 weeks for 5 groups with 2 antigens.
Table 3.
Estimated marginal means ± SEM for the HI titer values over 5 weeks for 5 groups with 2 antigens.
| Antigen | Groups | Weeks post-vaccination | ||||
|---|---|---|---|---|---|---|
| Week1 | Week2 | Week3 | Week4 | Week5 | ||
| LaSota | 1b | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 4.67±0.37 b |
| 2b | 1±0.08 c | 5±0.39 b | 6±0.47 ab | 6.33±0.49ab | 8.33±0.66 a | |
| 3b | 1±0.08 c | 1±0.08 c | 5±0.39b | 5.67±0.45 ab | 5±0.39 b | |
| 4a | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | |
| 4b | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 4.67±0.37 b | |
| PPMV-1 | 1b | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 6.33±0.49 ab |
| 2b | 1±0.08 c | 4.33±0.34 b | 5.67±0.45 ab | 6.33±0.49ab | 8.33±0.66 a | |
| 3b | 1±0.08 c | 1±0.08 c | 5±0.39b | 5.33±0.42 ab | 5.67±0.45 ab | |
| 4a | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | |
| 4b | 1±0.08 c | 1±0.08 c | 1±0.08 c | 1±0.08 c | 5.33±0.42 b | |
acb Means with different superscript showed a statistical significance p < 0.05.
Table 4.
Detection of viral shedding from collected swabs after the challenge of vaccinated and unvaccinated pigeons with the PPMV-1 isolate at 3, 5, and 7 days post challenge (dpc) and estimated by quantitative real-time-PCR (RT-qPCR).
Table 4.
Detection of viral shedding from collected swabs after the challenge of vaccinated and unvaccinated pigeons with the PPMV-1 isolate at 3, 5, and 7 days post challenge (dpc) and estimated by quantitative real-time-PCR (RT-qPCR).
| Groups | Tracheal swab | Cloacal swab | ||||||||||
| 3dpc | 5dpc | 7dpc | 3dpc | 5dpc | 7dpc | |||||||
| No of swabs * | Ct value ** | No of swabs | Ct value | No of swabs | CT value | No of swabs | Ct value | No of swabs | Ct value | No of swabs | Ct value | |
| G1b; LaSota-challenged | 0/3 | - | 1/3 | 37.6 | 1/3 | 30.9 | 0/3 | - | 2/3 | 32.6-34.9 | 1/3 | 28.4 |
| G2b; PPMV-challenged | 0/3 | - | 0/3 | - | 0/3 | - | 0/3 | - | 0/3 | - | 0/3 | - |
| G3b; LaSota &PPMV-challenged | 0/3 | - | 0/3 | - | 1/3 | 39.3*** | 0/3 | - | 0/3 | - | 1/3 | 37 |
| G4b; unvaccinated challenged | 0/3 | - | 1/3 | 37.3 | 3/3 | 35-36 | 0/3 | - | 3/3 | 32.8-33.9 | 3/3 | 33.9-35.4 |
dpc: days post-challenge, (*) Number of vials swabs (n=3), each vial containing 2 swabs from trachea or cloaca of 2 different pigeons/subgroup. (**) Average of the threshold cycle (Ct) values obtained by RT- qPCR according to number of positive swabs and was indication to the amount of viral RNA. (***) Ct value ˃ 39 considered the viral RNA undetectable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



