Submitted:
26 September 2024
Posted:
27 September 2024
You are already at the latest version
Abstract
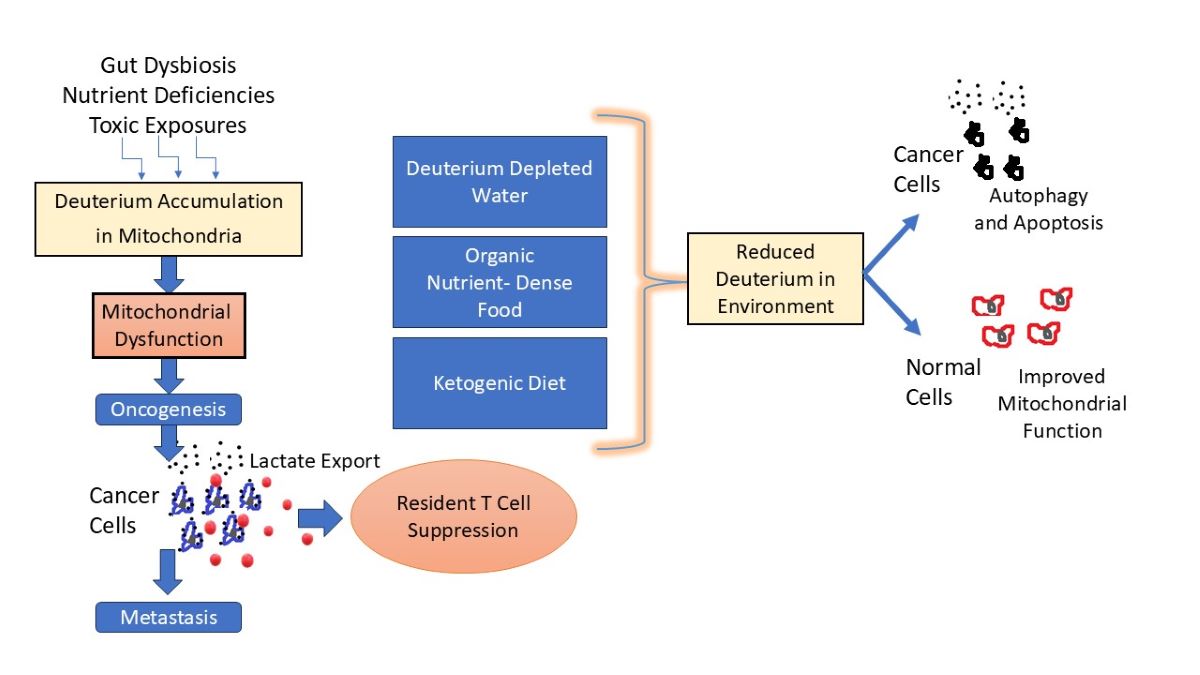
Deuterium is a non-radioactive isotope of hydrogen, containing a neutron as well as a proton, which makes it twice as heavy as hydrogen. Deuterium is a natural element found at 156 parts per million in seawater. Human metabolism employs clever strategies to minimize the amount of deuterium in mitochondrial water, because it causes a stutter in ATPase pumps, introducing excess reactive oxygen species and inefficiencies in ATP production. Gut microbes produce hydrogen gas that is 80% depleted in deuterium (deupleted), and this gas is recycled into organic matter that supplies deupleted nutrients to the host, such as acetate, butyrate, formate, methionine, and choline. Mitochondrial dysfunction is associated with many chronic diseases, most notably, cancer. Dehydrogenases are flavoproteins that typically have a high deuterium kinetic isotope effect, and they supply deupleted protons to NAD+ to produce NADH, which supplies protons to the ATPase pumps. Here, we examine the unique metabolic policies of cancer cells in a new light, and we propose that a tumor may arise as a strategy to help repair the mitochondria of tumor-resident immune cells overburdened with deuterium. While lactate accumulation in the tumor microenvironment promotes tumor growth, it also provides a valuable deuterium depleted nutrient to the tumor-resident immune cells. We provide strong evidence that deuterium depleted water (DDW) is a promising treatment for cancer.
Keywords:
Microbiome
; Deuterium
; Cancer
; Warburg effect
; Deuterium depleted water
; Lactate
; Formate
; Short chain fatty acids
; Apoptosis
; Autophagy
1. Introduction
The complex metabolic policies of biological organisms are very difficult to investigate, but one factor that has not received the attention it deserves is the role of deuterium. Deuterium is a natural heavy isotope of hydrogen, the most common atom in the universe. While hydrogen contains just one proton and one electron, deuterium also contains a neutron, and this makes it about twice as heavy as hydrogen, changing its biophysical and biochemical properties in important ways, compared to hydrogen. Wherever there is a hydrogen atom in a molecule, there is a small probability that it could be a deuterium atom. Deuterium is present in sea water at 156 parts per million, which sounds small, but, because hydrogen is so common, this translates into a level in the blood, atom for atom, that is six times more than calcium, and much more than the level of various essential elements, such as iron, iodine, copper, manganese, and cobalt [1].
The budding field of deutenomics can be defined as “the study of autonomic hydrogen isotope discrimination processes in nature.” Notably, in biological organisms, deuterium is fractionated differently in different biological compartments, with excess levels found in collagen in the extracellular compartments, especially the bone, and reduced levels in fatty tissues, lysosomes, and mitochondria [2]. Large amounts of deuterium depleted (deupleted) metabolic water are synthesized in the mitochondria during the production of the cell’s energy currency, ATP. The ATPase pumps use proton motive force to energize the reaction, and deuterium disrupts the process, causing the release of reaction oxygen species (ROS) and damaging the pumps.
Mice supplied with drinking water containing over 50% D2O suffer severe symptoms and die within a week to ten days [3]. Remarkably, deuterium is not toxic to microbes like Escherichia coli (E. coli). Even at 50% D2O, the maximum growth rate of E. coli is only reduced by 5% [4]. At more modest levels of D2O enrichment, E. coli growth rate is actually accelerated [4]. Protocols exist for developing strains of E. coli that are capable of growth on extremely high levels of deuterium. These strains are useful for producing highly deuterated proteins for nuclear magnetic resonance imaging experiments [5]. It is thus tempting to speculate that E. coli and other bacteria may be able to sequester deuterium in order to lower the deuterium load for the host.
Intriguingly, the growth of cancer cells in vitro is suppressed when their medium is deupleted (e.g., half the amount of deuterium found in seawater), whereas it is accelerated when the medium is deuterium enriched (e.g., twice the amount in seawater) [6]. This is opposite to the effect of deuterium on normal cells. This suggests that cancer cells are capable of sensing the amount of deuterium in the medium, and, when it is elevated, they are triggered through signaling mechanisms to proliferate. An experiment involving nasopharyngeal carcinoma (NPC) cells demonstrated that deuterium-depleted water (DDW) with deuterium concentrations at 100, 75, and 50 ppm suppressed the growth and invasiveness of tumor cells and caused cell cycle arrest. Notably, by contrast, DDW promoted the growth of normal control cells [7]. A provocative idea to explain this phenomenon is that a primary reason for the existence of cancer cells is to help restore deuterium homeostasis for the organism as a whole. It may be that, when living cells detect elevated levels of deuterium in their environment, they have an increased potential for oncogenesis. When the deuterium level in their surrounding medium is sufficiently reduced, cancer cells commit apoptosis, since the deuterium overload problem has been resolved.
In this paper, we examine the unusual metabolic policies of cancer cells in a new light. We propose that cancer cells are able to concentrate deuterium internally, while synthesizing deupleted nutrients that are released into the tumor microenvironment and into the circulation, and we argue that this helps alleviate the deuterium overload problem systemically. By greatly reducing the activity of the ATPase pumps in the mitochondria, cancer cells suppress the mitochondrial generation of ROS, thus maintaining survival despite deuterium overload. They derive their ATP mainly through massive processing of glucose through glycolysis, and they produce large amounts of lactate that is released into the circulation. The lactate that they release is a valuable deupleted nutrient that other cells, particularly the tumor-resident immune cells, can exploit.
In the following sections, we will first explain in greater depth how deuterium disrupts the function of the ATPase pumps. After describing how enzyme catalysis is affected by deuterium, we will show how gut microbial metabolism can be viewed as an elegant process to supply deupleted nutrients to support mitochondrial health of the host cells. Subsequently, we focus on two important nutrients, butyrate and choline, and show how they relate to deuterium homeostasis. A discussion on yeast overgrowth draws parallels between yeast and cancer cells, tied to deuterium toxicity. In extensive further analysis, we examine the unique aspects of cancer metabolic policies in light of the deuterium problem. We review what is known about the effects of DDW treatment on tumor cells, in both animals and humans, and provide evidence that it is a promising nontoxic therapy for cancer patients.
2. Mitochondrial ATPase Depends on Deupleted Protons
Mitochondrial ATP production depends upon the universal cofactor nicotinamide adenine dinucleotide (NAD). In multiple steps in the citric acid cycle in the mitochondrial matrix, NAD+ gets reduced to NADH by gaining two electrons and one proton from various substrates. NADH then provides protons to the intermembrane space to build up a proton gradient, while providing electrons to the electron transport chain in the inner membrane. Ultimately, the protons leave the intermembrane space to return to the matrix, exiting via ATPase pumps and providing the motive force to rotate the pump and fuel the production of ATP, while reducing oxygen to metabolic water [8]. The cell likely also needs to minimize deuterium concentrations in the DNA molecules in the mitochondria and the nucleus. The two strands of DNA are held together by hydrogen bonds between the two bases on opposite strands. Deuterium, being heavier, binds more strongly in covalent bonds, weakening its bond strength in ionic bonds [9,10]. Human stem cells grown in vitro on deuterium-enriched medium show a reduction in population doubling time, reflecting a slowing down of the cell cycle [11].
ATP synthase is the smallest known rotary motor in nature. F0F1 ATPase (F-ATPase) is a molecular motor positioned in the inner membrane of the mitochondrial intermembrane space that uses proton motive force as protons cross the membrane to generate the energy needed to phosphorylate ATP, while producing metabolic water from oxygen molecules. Protons moving through the ATPase pumps need to dissociate rapidly from Asp61 in F0 as an essential part of the catalytic reaction. Deuterons bind more tightly to organic molecules, and dissociate more slowly, resulting in a stutter in the pump [9,12].
The γ C-terminal helix of the ATPase molecular motor is the tip of the rotor that drives ATP synthesis. A hydrogen-deuterium exchange experiment showed that this helix accumulates significantly more deuterium from heavy water than other parts of the enzyme complex. Rotation of the γ rotor caused greatly enhanced deuteration in this C-terminal helix. The rotor tip is also prone to get stuck and unfold, being much less stable than other helices in the protein. Deuterium weakens hydrogen bonds, and their breakage likely causes unfolding, inducing a stutter in the motor rotation [13].
Repeated stutters caused by deuterons can be highly disruptive of the ATPase function, leading to inefficiencies in ATP production and increases in reactive oxygen species (ROS) [9,12]. Evidence suggests that the water inside mitochondria is highly viscous, because nearly all of it is interfacial water, i.e., water binding to hydrophilic surfaces. Water near hydrophilic surfaces has a much higher viscosity than bulk water. According to Sommer et al., “Nanoscopic water layers bound to hydrophilic surfaces (bound water) present viscosity values which are orders of magnitude larger than those of bulk water.” [14], p. 1. Reactive oxygen species increase the viscosity of water, and light, especially infrared light, reduces it [14].
Figure 1 shows a schematic representation of the electron transport chain (ETC), including the delivery of protons to the intermembrane space by Complex I-IV, the reduction of oxygen to metabolic water by Complex IV, and the synthesis of ATP in Complex V.
When ATP synthesis gets stalled by deuterons, Complex I and III, in particular, can release
reactive ROS, most notably the superoxide anion (O∗−). Mitochondrial dysfunction is a core feature of many chronic diseases, including neurodegenerative and neurodevelopmental diseases, metabolic diseases, autoimmune diseases, and, most notably, cancer [15,16,17]. It therefore behooves the cell to minimize the number of deuterons in the mitochondrial water, to the extent that this is possible.
Proton coupled electron transfer (PCET) reactions are also disturbed by deuterium. The electron transport chain is slowed down in the presence of deuterons, and this can cause upstream accumulation and subsequent leakage of electrons, increasing free radical release [12].
3. Deuterium Kinetic Isotope Effects
It is important to note that deuterons binding to other atoms have a very different exchange rate with protons in the water, depending on the atom they are bound to. The dominant atoms in organic molecules are oxygen, nitrogen, sulfur, and carbon. Carbon binds tightly to protons and deuterons, and so there is usually very little exchange between a proton or deuteron binding to carbon and protons/deuterons in the water, although there are important exceptions. Generally speaking, there are increasing degrees of exchange in order from carbon to sulfur to nitrogen to oxygen [1]. In many biological reactions catalyzed by special enzymes such as flavoproteins, protons are transferred from one molecule to another, exploiting proton tunneling to maintain the purity of the sequestered proton.
Because deuterium has different physical and chemical properties from hydrogen, it is often the case that an enzyme that extracts a hydrogen atom from a substrate may show significantly different kinetics if that hydrogen is replaced by deuterium. Researchers have defined a deuterium kinetic isotope effect (KIE) as a measure of the change in the rate of a reaction when deuterium replaces hydrogen, compared to the reaction when hydrogen is present. Generally, the KIE is greater than 1.0, meaning that deuterium weakens the enzyme’s kinetics. Some enzymes, such as soybean lipoxygenase, have a remarkable ability to reject deuterium, with KIEs over 100 [18].
Isomerases are an interesting class of enzymes that reversibly convert two isomers back and forth between each other. Many isomerases are able to scrub deuterium from the molecules involved via exchange of deuterium with protons from the water during the reaction, even when the protons were originally bound to carbon atoms. For example, it has been shown experimentally that the C(2) position of glucose- 6-phosphate from spinach leaf starch becomes deupleted as a consequence of the action of phosphoglucose isomerase, which has a high deuterium KIE [19]. Triosephosphate isomerase, a core enzyme of the glycolysis pathway, scrubs deuterium from the carbon atom that then supplies the deupleted proton to convert NAD+ to NADH in the next step in the pathway, catalyzed by glyceraldehyde-3-phosphate dehydrogenase [20].
At least half of all enzyme-catalyzed reactions involve hydrogen transfer. Furthermore, many hydrogen transfer reactions involve some amount of quantum mechanical hydrogen tunneling. Protons, being lighter, are much more capable of tunneling than are deuterons [21], and therefore these reactions tend to have a high deuterium KIE [22]. Flavoproteins are a large class of enzymes that exploit proton tunneling mediated by the bound flavin, such as flavin adenine dinucleotide (FAD). They tend to have a KIE for deuterium that ranges from 3.5 to 10 and can be as high as 25 [23]. Most dehydrogenases are flavoproteins, and therefore they are capable of depleting deuterium in the product of their reaction.
4. Microbes Supply Deupleted Nutrients to the Host
Intestinal microbial flora (commensals) can synthesize small organic molecules by using hydrogen gas as a reducing agent. These microbial metabolic products play a critical role in supplying deupleted nutrients to the host cells. Colonocytes thrive best on the four-carbon short chain fatty acid, butyrate, whose synthesis can be traced back to carbon dioxide and hydrogen gas. In fact, the gut microbes produce three short chain fatty acids (SCFAs) in abundance: acetate, propionate and butyrate, which often originate from dietary fiber. Their synthesis involves a critical step of extracting hydrogen gas from breakdown products of fiber, and then using the hydrogen gas to produce new organic molecules from carbon dioxide. Through this unusual strategy, these SCFAs can be expected to be depleted in deuterium, a feat largely achieved by the hydrogen gas. Eighteen percent of the world’s population experience bloating at least once a week [24]. This is an indicator of an impaired ability to recycle gases produced by gut microbes into organic nutrients.
Deuterium isotope fractionation occurs during evaporation and condensation, where deuterium tends towards condensation and protium is more inclined to evaporate [25]. This is logical, since deuterium is twice as heavy as protium. As a result, deuterium tends to concentrate in snow and ice, and the water that melts from glaciers is typically deupleted [26,27]. Furthermore, rain falling at the Equator tends to have higher deuterium content than rain falling at higher latitudes, because more of the water evaporates from the raindrop during its descent at higher atmospheric temperatures, leaving behind deuterium enriched liquid water. These observations become relevant when considering the synthesis of gases by gut microbes, because it appears that they take advantage of this principle to synthesize deupleted nutrients for the host.
Hydrogen gas is one of the main gases that accumulate in the gut as a consequence of microbial metabolism. Many gut microbes possess functioning hydrogenase enzymes that can synthesize hydrogen gas from simple molecules such as glucose and formate (HCOOH). During fermentation, the E. coli strains can use a membrane-bound hydrogenase coupled with formate dehydrogenase to produce H2 and CO2 from formate [28]. This reaction may play an essential role in the production of deupleted SCFAs, as acetogens can subsequently produce acetate from H2 and CO2, where the H2 could have been derived from formate metabolism. This sort of cycling through H2 synthesis is a clever strategy for deuterium fractionation. Recently, there has been a resurgent interest in the use of hydrogen gas therapeutically to treat a large list of diseases [29]. Firmicutes and Bacteroides are the most prevalent hydrogen-producing bacteria in the gut, together making up over 90% of the bacteria in the gut [30].
A paper published in 1961 involved investigations of a Pseudomonas species which showed that this microbe was able to synthesize hydrogen gas (H2) that was 80% depleted in deuterium, compared to the normal amounts present in water. The gas was produced, along with carbon dioxide (CO2), from formate. The enzymes that produced the hydrogen gas are similar to enzymes present in E. coli and other coliform bacteria that populate the human gut [31]. It is possible that a simple biophysical argument based on the idea that deuterium, being heavier, tends to remain in the liquid phase can explain this phenomenon, but it is also likely that the enzyme has a high deuterium KIE, further fractionating the deuterium.
The hydrogen gas that is produced by gut microbes is a powerful reducing agent, and, in particular, it can reduce CO2 to methane gas (CH4), another prominent gas in the gut. In fact, there is a seemingly futile cycle that repeatedly takes place in the gut, reducing CO2 to CH4 and then metabolizing CH4 back to CO2, as illustrated in Figure 2. What can be noticed is that this process results in the conversion of 3 molecules of NAD+ to NADH, where the hydrogen atom attached to the NAD molecule will presumably be 80% deupleted, a remarkable feat. This proton eventually makes its way to the mitochondrial intermembrane space, through a multistep process that will be described later in this paper.
The highly reactive small molecule, formaldehdye, is critically involved in pathways that intersect this carbon cycle. Organisms called methylotrophs can assimilate (fixate) formaldehyde and use it for biomass production via one-carbon metabolism pathways [32]. Gut microbes metabolize formaldehyde in both an assimilatory and a dissimilatory pathway. We have already seen a dissimilatory pathway that produces deupleted H2 gas, but it can also yield NADH along with carbonate. In the assimilatory pathway, formaldehyde participates directly in one-carbon metabolism, beginning by reacting (spontaneously as well as enzymatically) with tetrahydrofolate (H4F) to produce methylene-tetrahydrofolate (CH2= H4F). This molecule, an important assimilatory intermediate of formaldehyde metabolism, plays a central role in the methylation pathways that deliver methyl groups to many different macromolecules such as proteins and DNA, and also supply methyl groups to produce critically important micronutrients such as methionine, melatonin, and choline [33]. A provocative thought is that these methylation pathways may be important in human metabolism in large part because the methyl groups can be expected to be highly deupleted, as they can be traced back to microbially produced H2 gas.
Figure 3 schematizes the methylation pathways utilized by human cells which ultimately populate the cytosines of DNA molecules with methyl groups. A most significant but little-known fact is that the methyl groups are incrementally removed from cytosines in place by ten- eleven translocation (TET) methylcytosine dioxygenases, whereby the hydrogen atoms are removed one by one, and succinate is synthesized from α-ketoglutarate each time. Succinate is a very important molecule in the mitochondria, as it supplies protons to coenzyme Q10 in the inner membrane of the mitochondrial intermembrane space, and this is probably also why it plays powerful signaling roles in biology [34]. These protons ultimately contribute to the pool of protons that supply proton motive force to the ATPase pumps, so they need to be significantly deupleted.
TET proteins play powerful roles in myeloid and lymphoid immune cell development and modulation of the inflammatory response. Dysregulated TET enzymes are linked to both myeloid and lymphoid leukemias [35]. Global DNA hypomethylation is a characteristic feature of many cancers [36,37], and this could be an indicator of insufficient methylation capacity, which could be traced back to deficiencies in the gut microbes that synthesize deupleted methane that ultimately feeds into the methylation pathways.
There are at least three broad classes of microbes that inhabit the gut that can use hydrogen gas as a reducing agent, methanogens, sulfate-reducing bacteria (SRB), and acetogens [38,39]. As their names suggest, methanogens produce methane from H2 and CO2, and acetogens produce acetate from H2 and CO2, according to the following equation:
4H2 + 2CO2 = CH3COOH + 2H2O; ∆G0 = −95kJ/mol. (1)
Studies on several different strains of acetogenic bacteria obtained from human feces found that all of them could grow when provided with only hydrogen gas and carbon dioxide, and acetate was the sole metabolite produced, via reductive acetogenesis [40,41]. The acetate that is produced by acetogenic bacteria is further processed by other gut microbes into butyrate, an essential nutrient for the epidermal cells lining the colon (colonocytes). Figure 4 schematizes the complex biological pathways by which gut microbes convert fiber to butyrate. Butyrate is sufficiently important that we are devoting the entire next section to it.
5. The Many Benefits of Butyrate to Human Health
About 300 to 600 mmol of SCFAs are produced on average every day in the human intestine, and most of it is absorbed by the colonocytes [42]. Members of the Firmicutes phylum, particularly Faecalibacterium prausnitzii, are the most common butyrate-producing bacteria in the gut [43,44]. These microbes combine two acetate molecules to synthesize butyrate, and, if the acetate is deupleted, then the butyrate will be, too. Butyrate is the preferred fuel for colonocytes lining the gut, and many studies have shown that inflammatory bowel disease (IBD) is linked to low butyrate levels [42,45].
There is considerable excitement in recent years about the potential for butyrate to treat cancer therapeutically [46,47]. Butyrate suppresses cell growth, migration, and invasion of cancer cells by inhibiting histone deacetylase (HDAC) activity [48,49]. HDACs remove acetyl groups from histones, which generally alters expression of proteins involved in cancer [50]. HDAC inhibitor drugs have recently shown promise as anti-cancer agents, as they induce growth arrest and apoptosis in several different human cancer cell lines [51].
Remarkably, butyrate causes apoptosis of cancer cells through multiple mechanisms. Butyrate induces death-associated protein kinase (DAPK) expression in human gastric cancer cells, leading to apoptosis [52]. This effect is likely mediated by its action as an HDAC inhibitor. Butyrate also activates the mitochondrial apoptotic pathway by increasing the ratio of pro-apoptic Bax to anti-apoptotic Bcl-2, which leads to cytochrome c release, caspase 3 activation and apoptosis [53].
Sirtuins are a large class of HDACs, and they can deacetylate many other proteins besides histones, causing major metabolic policy changes in cells. Two important protein complexes in the mitochondria that are regulated by the sirtuin SIRT3 are pyruvate decarboxylase complex (PDC) and Complex I of the electron transport chain. Butyrate has been demonstrated to strongly inhibit SIRT3, and this has a dramatic effect on cancer cells supplemented with butyrate. PDC links glycolysis to the TCA cycle by converting pyruvate to acetyl-CoA. Butyrate treatment decreases lactate production by activating PDC, and causes the accumulation of citrate in the mitochondria, due to suppression of Complex I together with activation of PDC. The result of essentially reversing the Warburg effect is to induce apoptosis in cancer cells [54].
It has even been said that the primary beneficial effect of gut microbes on host metabolism is directly due to the production of SCFAs. SCFAs are beneficial in maintaining gut barrier function, glucose homeostasis, immunomodulation, appetite regulation and obesity [49]. Butyrate can be passed into the blood stream by colonocytes, and it can cross the blood-brain barrier. It has been proposed that butyrate plays an important role in the communication channels of the gut-brain axis [55].
6. Aldehyde Detoxification Pathways and Cancer
As we have described, formaldehyde is a common organic product of gut microbial metabolism, and it is central to one-carbon metabolism and methylation pathways. The synthesis by gut microbes of hydrogen gas and carbon dioxide from formaldehyde and the reverse synthesis of formaldehyde from hydrogen gas and carbon dioxide is a cyclic pathway with a crucial outcome of producing deupleted methyl groups and supplying deupleted protons to the ATPase pumps.
Formaldehyde is classified as a carcinogen because it easily reacts with DNA causing DNA-interstrand crosslinks and DNA-protein crosslinks [56]. Because formaldehyde is highly reactive, it is essential to have on supply enzymes that can efficiently convert it to a less reactive product. We have already shown that formaldehyde reacts nonenzymatically with H4F, producing CH2= H4F, the first step in the methylation pathways. Formaldehyde also reacts spontaneously with glutathione, forming S-hydroxymethyglutathione [57].
The alcohol dehydrogenase family is a broad class of enzymes which are evolutionarily conserved from bacteria to humans. Many of them are responsible for the metabolism of short chain alcohols, notably ethanol and methanol. Genetic defects in these enzymes have been linked to alcoholism, schizophrenia, Parkinson’s disease, asthma, autism and cancer [57]. A unique member of this class is S-nitrosoglutathione reductase (GSNOR), also known as alcohol dehydrogenase 5 (ADH5). This enzyme is ubiquitously expressed in both plants and animals, and it plays an essential role in controlling reactive nitrogen species. However, it also metabolizes hydroxymethylglutathione, yielding formate and glutathione as the reaction products, while reducing NAD+ to NADH [58]. Hence, GSNOR is a glutathione-dependent formaldehyde dehydrogenase, that can help supply deupleted protons to NAD+ while detoxifying formaldehyde.
Ethanol can be converted to acetaldehyde and then acetate through the action of dehydrogenases. Each of these reactions yields a molecule of NADH. Like formaldehyde, acetaldehyde is highly reactive and can damage DNA. Many Asians are known to have defective versions of ALDH2, a mitochondrial enzyme highly expressed in the intestinal mucosa, and this defect is associated with facial flushing following alcohol consumption. This enzyme catalyzes both the oxidation of ethanol to acetaldehyde and the oxidation of acetaldehyde to acetate. When this enzyme is deficient, acetaldehyde accumulates to toxic levels. A deficiency in this enzyme is linked to esophageal cancer and head and neck cancers [59,60].
Fanconi anemia (FA) is a rare autosomal recessive disorder that causes severe impairment of stem cell maturation in the bone marrow (bone marrow failure), and a high predisposition to cancer, especially acute myeloid leukemia (AML). It is due to mutations in genes that collaborate in DNA repair mechanisms [61]. These elaborate repair mechanisms are essential for repairing chronic damage caused by formaldehyde and acetaldehyde. In a paper published in 2020, Yasuyoshi Oka et al. describe a severe genetic disorder called aplastic anemia, mental retardation, and dwarfism (AMeD) that is due to digenic mutations in genes coding for ALDH2 and AHD5. In patients who suffer from this condition, which is very rare, there is an inability of the DNA repair mechanisms to keep up with the DNA damage that is caused by accumulated formaldehyde, despite healthy DNA repair enzymes [62]. These authors wrote: “Our results suggest that the formaldehyde clearance is as important as the DNA repair system for normal development of both humans and mice.” Thus, while formaldehyde plays a central role in supplying the mitochondria with deupleted protons, it does so at a huge cost, whenever the dehydrogenases that supply NAD+ with deupleted protons are defective.
7. Choline, Cancer, and Cardiovascular Disease
Choline (2-hydroxyethyl)trimethylammonium) is an essential nutrient for humans, acting both as an important structural part of phospholipids and as a methyl donor in one-carbon chemistry. It is a precursor to multiple membrane lipids including phosphatidylcholine, sphingomyelin and plasmalogens. It is important to note that S-adenosylmethionine is used as the methyl donor to supply three methyl groups to membrane phosphatidylethanolamine to produce phosphatidylcholine. This implies that these three methyl groups, all attached to the nitrogen atom of choline, would be deupleted. The choline in these membrane lipids is used by cholinergic neurons to synthesize acetylcholine, an important neurotransmitter [63].
Colorectal cancer is the third most commonly diagnosed cancer type worldwide, and it is second only to lung cancer in mortality [64]. A combined deficiency in folate, choline and methionine causes DNA hypomethylation, hepatic steatosis, cirrhosis, and ultimately liver cancer in rodents, even when they are not exposed to carcinogenic agents [65]. A study investigating DNA methylation levels in colon cancer biopsies revealed that the tumor cells had significantly reduced DNA methylation compared to the adjacent healthy cells, which was associated with reduced expression of the folate receptor [66]. In a study based in China, higher intake of dietary choline was associated with a reduced risk of colorectal cancer [67]. The authors suggested that protection might be due to choline’s role as a methyl donor. Insufficient methyl donor capacity might disrupt DNA methylation and impair DNA repair mechanisms [65].
Choline is almost surely a valuable deupleted nutrient to support mitochondrial health. It is informative to trace the pathways of choline metabolism in the gut, which critically involve the gut microbes. In the gut, choline is converted to betaine (trimethylglycine) which then provides methyl groups to convert homocysteine back to methionine, via the enzyme betaine-homocysteine methyltransferase (BHMT). Choline can be metabolized by the gut microbes into trimethylamine (TMA), which is then passed to the liver where it is oxidized to trimethylamine oxide (TMAO). Betaine and carnitine can also be converted to TMAO. Elevated serum levels of TMAO are associated with cardiovascular disease [68].
There are several strains of methanogenic archaea that inhabit the human gut, and these strains can metabolize tri-, di-, and monomethylamine (all derivatives of choline) to methane, using hydrogen gas as a reducing agent [69,70]. A unique group of methanogenic archaea that populate the human gut, in the order Methanomassiliicoccales, are even restricted to utilizing only methyl compounds such as TMA as substrates to synthesize methane using H2 as a reducing agent [71,72]. This pathway, which involves oxidizing methyl groups to CO2 and H2, and then recombining these products to synthesize methane, may play an essential role in fractionating deuterium out of organic molecules.
The methane that is synthesized is likely to be extremely low in deuterium, since the methyl groups it was derived from most likely came from a previous cycle of deuterium scrubbing.
Probiotics containing these archaea species may be a way to reduce the levels of TMAO in the blood and hence to treat cardiovascular disease [73]. Indeed, gut colonization with Methanobrevibacter smithii in ApoE-/- mice led to a significant reduction in plasma TMAO levels, as well as attenuation of the atherosclerosis burden [74].
Acetylcholinesterase is the enzyme that converts acetylcholine to acetate and choline, freeing up choline for future metabolism. Organophosphate pesticides are well known acetyl-cholinesterase inhibitors. A study conducted in Sudan revealed that cancer patients had significantly reduced acetylcholinesterase activity compared to controls (53.4 vs. 93.8 (p-value 0.001)) [75]. It is possible that the reduced potential for converting choline into valuable deupleted nutrients via gut microbial activities explains this link.
8. Yeast Overgrowth and Serum Ethanol
An in vivo study on human non-small cell lung cancer showed that lactate is the preferred fuel used by tumor cells in the citric acid cycle. The cells release lactate derived from glucose through glycolysis into the circulation and then retrieve it later for further processing in the mitochondria [76]. It is likely that the primary purpose of these extra steps is to further reduce the likelihood that the proton that ultimately becomes the hydrogen atom in NADH, produced in the mitochondria, is a deuteron.
This same principle probably applies for yeast cells populating the gut, except that the glucose is converted to ethanol rather than lactate. A rare condition aptly named “auto- brewery syndrome” happens when a person experiences a massive overgrowth of yeast in the gut, which are producing large quantities of ethanol from glucose via fermentation and releasing it into the gut, from which it is ultimately absorbed into the bloodstream [77]. The person experiences symptoms of severe inebriation, despite not having consumed any alcohol. Two genera of yeast are most noted for causing this condition, namely Saccharomyces spp. and Candida spp. [78].
Ethanol and lactate have long been viewed as waste products of glycolysis, but this view is rapidly changing. Tracer studies conducted in 2020 on 13C-labeled lactate have shown that this nutrient is taken up by every tissue in the body, even by tumor cells [79]. A study on fermenting yeast cells (Saccharomyces cerevisiae) showed that these cells practice a near complete uncoupling of glycolysis from oxidative phosphorylation. These authors wrote: “Specifically, we show that fermenting budding yeast simultaneously release and uptake ethanol, much as many mammalian cells simultaneously produce and consume circulating lactate.” [80] Two dehydrogenase steps take ethanol to acetaldehyde and then acetyl-CoA, which directly feeds into the citric acid cycle. Both steps convert NAD+ to NADH. Under oxidative stress conditions, an even larger portion of the supply of protons for NADH production in the mitochondria is contributed by externally supplied ethanol, compared to pyruvate derived from glucose metabolism [80]. Figure 5 illustrates the common but surprising metabolic practice of yeast cells.
9. Yeast and Cancer cells have Much in Common
There is gaining support for the concept of treating cancer with DDW. A double-blind clinical trial showed that administration of DDW caused a 3-7-fold increase in mean survival time in lung cancer patients, and a 2-fold increased survival time in breast cancer [81]. A case-control study involved pancreatic cancer patients, where both groups received chemotherapy, but the treatment group also received orally deupleted water (45-65 ppm deuterium tailored down over a three-month period). The treated group survived on average for 19.6 months compared to only 6.36 months for the control group [9]. Glioblastoma is the most common brain cancer. A study involving 55 patients suffering from glioblastoma, who continued conventional chemotherapy while consuming DDW, found that their mean survival time (MST) (30 months) was considerably higher than the historical MST for this disease (under 15 months) [82]. Exposure of tumor cells to DDW decreases the number of cells in S phase and increases the number in G1 phase, essentially inducing G1-phase cell growth arrest [1].
Yeast cells and cancer cells have much in common. It seems that both have an end function that restores deuterium homeostasis to the organism, or perhaps more specifically to the immune cells that infiltrate their microenvironment. Candida albicans is an opportunistic pathogenic fungus, and it tends to infect those with impaired immune function, including cancer patients [83]. Ironically, Candida overgrowth is also a risk factor for cancer [84]. As was discussed previously, Candida overgrowth leads to excess accumulation of ethanol in the blood. Ethanol is metabolized into acetate in two steps involving ethanol dehydrogenase (producing acetaldehyde) and acetaldehyde dehydrogenase (Aldh), producing acetate. This acetate is expected to be deupleted.
The primary enzyme in the liver for metabolizing acetaldehyde is Aldh2, localized to the mitochondria. Genetic mutations causing deficiencies in Aldh2 increase the risk to hepatic cell carcinoma (HCC) following alcohol-related liver fibrosis. The description of this process by Seo et al. is precise and revealing: “Mechanistic studies revealed that after chronic alcohol exposure, Aldh2-deficient hepatocytes produce a large amount of harmful oxidized mitochondrial DNA via extracellular vesicles, which can be delivered into neighboring HCC cells and subsequently activate multiple oncogenic pathways, promoting HCC.” [85].
Chronic ethanol consumption increases cancer risk, not only in the liver but also in the digestive tract and breast tissues [86]. People with genetic defects in Aldh are especially vulnerable to cancer from chronic ethanol exposure, because acetaldehyde lingers long enough to cause DNA damage [86]. It may be for this reason that frequent episodes of Candida overgrowth are a risk factor for cancer [84]. Several Candida species, including Saccharomyces cerevisiae and Cyberlindnera jadinii, are transcriptionally active in gastrointestinal tumors and predictive of worse tumor outcomes [87]. It is also interesting from the standpoint of the potential for systemic deupletion that the development of cancer might alleviate the burden on the Candida fungi to repair the deuterium overload problem, a Faustian bargain. Chemotherapy can increase the risk of systemic Candidiasis in cancer patients, and this may simply be because the cancer cells can no longer adequately support deuterium detoxification for the host, so Candida species rise to the occasion [88].
Chronological aging can be defined as the diminished survival of nondividing cells due to senescence under growth in various conditions. In an experiment involving growing yeast cells on a medium with 50% D2O enrichment, it was found that D2O actually increased the chronological lifespan of the yeast cells by up to 85%. D2O substantially suppressed the production of reactive oxygen species, which was thought to be due to increased use of glycolysis and reduced use of oxidative phosphorylation for ATP synthesis. It was hypothesized that aging could be a consequence of the damaging effects of ROS produced primarily by the mitochondria [89]. Yeast cells can survive in both an aerobic and an anaerobic environment. They are quite capable of surviving long term on glycolysis alone, and apparently, they can alter their metabolism strategically towards aerobic glycolysis, in the presence of high deuterium exposure, shutting down ATP production by the mitochondria. Similarly, cancer cells switch to glycolysis as an energy source and release deupleted nutrients such as lactate and formate, likely triggered by elevated deuterium levels in their growth medium. Interestingly, reactivating oxidative phosphorylation can be a useful chemotherapy strategy, because it induces mitochondrial-stress-dependent apoptosis in cancer cells. The anticancer agent dichloroacetate (DCA) activates the TCA cycle in the mitochondria, which restores oxidative phosphorylation, increasing oxidative stress leading to apoptosis [90].
10. Explaining Cancer’s Unusual Metabolic Policies
In this section, we will examine several of the abnormal aspects of cancer cell metabolism from the perspective of deuterium homeostasis. Cancer cells adopt an unusual metabolic policy, exploiting aerobic glycolysis as the main source of ATP and invoking anabolic pathways in the mitochondria to provide resources to support proliferation and migration [91]. They upregulate vacuolar ATPase (V-ATPase) to pump protons out of the cell, reversing the membrane polarity and acidifying the external space [92]. V-ATPase plays a critical role in cancer pathology, and it is being actively considered as an attractive therapeutic target in cancer [93].
10.1. V-ATPase and Microenvironment Proton Deupletion
There are two main types of ATPases in biological organisms: F-ATPase and V-ATPase. We have seen that F-ATPase is the enzyme in the mitochondria that synthesizes ATP from ADP, while reducing oxygen to metabolic water. It relies on deupleted protons crossing the inner membrane of the mitochondria from a proton-dense to a proton-sparse medium as the motive force. V-ATPases have an opposite function. Protons are pumped across a membrane from a proton-sparse medium to a proton-dense medium, and this requires energy, which is supplied by converting ATP to ADP. When they are localized to the plasma membrane, they pump protons into the extracellular space, and when they are localized at the membrane of the endosome/lysosome organelle, they can acidify the endosome to convert it to a lysosome [94].
A seminal paper published in 1990 involved evaluating the deuterium KIE of yeast V-ATPase acting at the plasma membrane. The authors were surprised to find that the extrusion of H+ ions was reduced by as much as 90% when the cells were suspended in heavy water. In referring to V-ATPase, these authors wrote: “It thus appears that the binding site for protons (or hydronium ions) to be transported does not accept deuterons (or deuteronium ions) with equal ease or perhaps not at all.” [95]. In other words, yeast V-ATPase has an extremely high deuterium KIE.
It is significant that human breast cancer cells with high metastatic potential overexpress V-ATPase at the plasma membrane, and they use it presumably to transfer deupleted protons into the extracellular space, thus enriching deuterium levels in the cell’s cytoplasm [96]. This strange practice is consistent with a theory that a primary role of cancer cells is to maintain high internal levels of deuterium in order to promote reduced deuterium levels in the extracellular space.
Immune cells routinely invade the tumor microenvironment, but there are often signaling mechanisms that cause them to stand down, failing to kill the tumor cells, via a process called “tumor immune escape” [97]. These immune cells can presumably benefit from the deupleted protons released by the tumor cells to help them maintain low deuterium levels in their own organelles, with the ultimate goal of restoring their ability to kill and clear the cancer cells, once they regain mitochondrial health. An optimal T cell response is essential for efficiently killing cancer cells, and this is a hallmark of successful cancer immunotherapies [98]. In this regard, the study of BC Kovács et al.is revealing [99]. The researchers exposed mice to chemical carcinogens and then the animals were supplied with DDW with 30 ppm deuterium. The DDW treatment prevented tumor development as compared to control mice that lacked the DDW supplementation. Similar results from studies that same researcher conducted in humans show that DDW treatment inhibited cancer relapse. A possible explanation is that the immune cells in the tumor microenvironment benefitted sufficiently from the deupleted water to be able to restore mitochondrial health, allowing them to initiate anti-tumor surveillance and release cytotoxic metabolites.
10.2. Lactate
Cancer cells commonly overproduce lactate and then release it into the circulation, allowing the recipient cell to skip glycolysis and efficiently produce ATP directly from the tumor-provided lactate molecule, a process known as the Warburg effect. Meanwhile, the cancer cell can repurpose its mitochondria towards anabolic pathways, thus avoiding exposure to ROS generated by oxidative phosphorylation [100]. While lactate had long been viewed as a waste product of glycolysis, researchers have now come to appreciate the fact that it is actually a valuable nutrient, and that its presence in the microenvironment has powerful signaling capabilities, especially towards immune cells [101]. Lactate accumulation in the tissue microenvironment is a common feature of both inflammatory disease and cancer [102]. The Warburg effect is a hallmark of cancer cells: they produce ATP primarily through glycolysis even in the presence of adequate oxygen for oxidative phosphorylation [103]. This is similar to the strategy that yeast cells use in the presence of excess deuterium. Aerobic glycolysis produces lactate under a variety of stressful conditions, such as trauma, infection, myocardial infarction, and heart failure [102].
Lactate is a very useful fuel for the mitochondria, because it can easily be converted
to pyruvate via lactate dehydrogenase. Glycolysis, taking place in the cytoplasm, converts glucose to pyruvate. This pyruvate molecule can be transported into the mitochondria and used directly to generate large amounts of ATP via the citric acid cycle. However, most of the time this is not what happens. Instead, pyruvate is converted to lactate in the cytoplasm, and then the lactate is transported across the mitochondrial membrane and converted back to pyruvate [104].
While this appears to be a superfluous step, it is actually a very clever strategy for assuring that the hydrogen atoms in the mitochondrial NADH are deupleted. Cytoplasmic lactate dehydrogenase has a high deuterium KIE (∼3-4) [105], so it provides lactate with a deupleted proton, which is then delivered to NAD+ in the mitochondria, to finally produce a further deupleted NADH molecule within the mitochondria, again, most likely, since the enzyme is a flavoprotein, with a high deuterium KIE. The extra steps of passing a proton to lactate and then from lactate to NAD+ provide a simple mechanism for further shedding of deuterium. Figure 6 depicts the lactate shuttle.
Very frequently, the metabolism of glucose to produce ATP is carried out in two steps in two different cells, with the first cell producing lactate from glucose via glycolysis in the cytoplasm, and the second one metabolizing lactate to CO2 and water in the mitochondria. A paper published by Xiao et al. in 2022 brings awareness to the fact that lactate, rather than being a waste product, is a very common metabolic intermediate between glycolysis in one cell and the citric acid cycle in another distant cell. These authors wrote: “Indeed, it seems that most carbohydrate oxidation in mammals, rather than occurring by a tissue taking up glucose and fully oxidizing it to carbon dioxide, instead involves carbon flowing through circulating lactate as a metabolic intermediate.” [80]. For cancer cells, lactate seems to be a necessary molecule for energy transport. The release of lactate and acidification by tumor cells accompanies metastasis, angiogenesis and poor outcomes of cancer progression [106].
10.3. Considerations Around Lactate, V-ATPase, Acidification, and M2 Macrophages
Most tumors have a slightly elevated internal pH (7.3-7.7) compared to the low external pH (5.6 - 7.0), where the polarity is opposite to that of normal cells. Lactate concentrations in the tumor microenvironment can be as high as 40 millimolar, and it is generally erroneously believed that lactate is responsible for the low pH in the tumor external milieu. However, this is caused predominantly by the activities of V-ATPase pumps, which, as we’ve seen, are pumping deupleted protons out of the cell. The abnormal over-expression and unusual subcellular locations of V-ATPases in tumor cells are associated with a poor prognosis in many types of cancer. Plasma-membrane V-ATPase promotes tumor cell invasion, metastasis and drug resistance [107]. The release of lactate and acidification by tumor cells accompanies metastasis, angiogenesis and cancer progression [106]. Pérez-Tomás and Pérez-Guillén wrote: “Several studies conducted in the 1990s clearly showed that lactate is not responsible for extracellular acidosis; rather, ATP hydrolysis coupled with glycolysis is the main source of H+ production that leads to a decrease in the extracellular pH.” [106].
Researchers from Johns Hopkins University published a provocative paper in 2022 which proposed that maintaining lactate metabolism is essential for CD8+ T cells to promote their cytotoxic activity [108]. Lactate dehydrogenase (LDH) expression was found to be essential for this benefit to hold [108]. Notably, LDH results in the synthesis of NADH to support deupleted protons for the ATPase pumps. These authors maintain that it is the acidic environment more than the presence of lactate that induces immune suppression. They also claimed that lactate is a valuable nutrient to fuel the TCA cycle in regulatory T cells (Tregs) and bone-marrow derived macrophages. The lactate anion is well tolerated by CD8+ T cells in pH neutral conditions [109]. Experiments involving exposing CD8+ T cells in culture to a pH-neutral environment along with high lactate availability during the T-cell activation phase revealed that lactate promoted rapid expansion and an enhanced effector profile, increasing production of interferon γ and cytotoxic proteases such as granzymes [109].
A study published in 2022 showed that treatment of tumor-bearing mice with subcutaneous administration of sodium lactate solution at pH 7.4 leads to improved immune function of CD8+ T cells by inhibiting histone deacetylases and boosting stem-like properties. This resulted in significant suppression of tumor growth. These authors suggested that it is the low pH of the tumor microenvironment that promotes tumor progression rather than the lactate itself [110]
Although lactate is a beneficial fuel source for T cells, there is a large literature that shows that lactate acts as a signaling molecule to suppress immune function in tumors, supporting tumor escape. Tumor associated macrophages (TAMs) are the most abundant immune cells in the tumor microenvironment, and they play a powerful role in reprogramming T cells towards an immunosuppressive state, as well as promoting tumor initiation and progression [111]. Macrophages can be characterized as being in either an M1 (pro-inflammatory) or M2 (anti-inflammatory) state. Lactic acid skews macrophages towards an M2 state, particularly when the external environment has a low pH [112].
Excess lactic acid is an indicator of immunosuppression [101]. Elevated serum lactic acid levels are associated with poor clinical outcomes in COVID-19 [113]. Interestingly, in our recent study describing an unusual case of stress hemopoiesis of autoimmune origin due to mRNA COVID-19 vaccination, the patient had elevated levels of lactic acid in plasma [114]. Lactate in the tumor microenvironment facilitates the recruitment of immunosupppressive cells, including TAMs, regulatory T cells (Tregs), and myeloid-derived suppressor cells (MDSCs) [115].
Lactate can induce marked changes in gene expression, in part through histone lactylation, a process that was only first discovered in 2019 by Zhang et al. [116]. In response to a bacterial infection, M0 macrophages differentiate into pro-inflammatory M1 macrophages, with an associated increase in glycolysis. As time passes, lactate accumulates in the cell, and this leads to histone lactylation, which then induces a transformation into an M2 phenotype, in preparation for wound healing and resolution of the inflammatory state. Arg1 is a gene associated with M2 macrophages. In TAMs isolated from lung tumors and melanoma cultures, its overexpression was confirmed in association with lactylated histones, but not with acetylated histones. Exogenous lactate (e.g., released by cancer cells) also increased histone lactylation levels in immune cells and induced Arg1 expression, as well as other M2-like genes, such as Vascular Endothelial Growth Factor A (VEGFA) [116].
Several studies on various types of cancer have confirmed that M2 macrophages promote tumor survival. M2 macrophages facilitate tumor metastasis and augment drug resistance [117]. In an in vitro study on prostate cancer cells, it was found that acidic pH, independent from lactate, can promote the polarization of macrophages towards an M2 state, which promotes tumor survival, including enhanced tumor cell proliferation, loss of macrophage cytotoxicity, and release of angiogenic factors [118]. Tumor relapse after chemotherapy is a major clinical problem, because it often involves inoperable metastasis. M2 macrophages accumulate around blood vessels after chemotherapy, promoting tumor revascularization and relapse [119]. In glioblastoma cells, extracellular vesicles containing LDHA are released by macrophages recruited to the tumor microenvironment, to promote tumor cell glycolysis, proliferation, and survival [120].
Both LDH and monocarboxylate transporter 1 (MCT1), the transporter that imports lactate into the cell, show deuterium KIEs that would further support low deuterium levels in the mitochondria. Studies on MCT1 activity in heavy water showed that it has a significant deuterium KIE, with the uptake of lactate (and protons/deuterons) slowed significantly in D2O at pD 6.8 compared to H2O at pH 6.8 [121]. What this means in practice is that cells preferentially take up protons rather than deuterons when they take up lactate, further improving the internal H/D ratio. A study on the effects of deuterium on the activity of human LDH found a solvent isotope effect of 2.75 for D2O and a kinetic isotope of 2.74 for NADD instead of NADH, at pH 4.3 [122].
10.4. Formate Overflow and Cancer
Formate is essential for nucleotide synthesis in proliferating cells. However, cancer cells often synthesize even more formate than they actually need, a phenomenon that has been called “formate overflow” [123,124]. Cancer cells excrete the excess formate into the tumor microenvironment, and it makes its way into the blood stream. Stressed cancer cells, for example those subjected to chemotherapy, further increase formate release, and excess formate in the tumor microenvironment promotes invasion of glioblastoma cells, through activation of calcium signaling. Formate also promotes metastasis in breast cancer cells. Increased synthesis of matrix metalloproteinase (MMP) in response to formate signaling facilitates migration and extravasation [125].
Serine is probably the primary source of formate produced by human cells [126]. Serine can be catabolized to glycine and formate in the mitochondria [127]. Serine catabolism is induced upon energy stress: the pathway that converts serine to glycine and formate yields one molecule of ATP for each serine molecule catabolized. The de novo synthesis of serine is often upregulated in cancer cells, and hypoxia induces upregulation of the serine catabolic enzyme serine hydroxymethyltransferase [128]. Meiser et al. likened the phenomenon of serine one-carbon catabolism with formate overflow observed in cancer cells to the phenomenon of glucose catabolism with lactate overflow [123]. In both cases, a valuable source of low-deuterium protons is released into the circulation. Furthermore, deupleted hydrogen gas can be produced by the gut microbes from serum-derived formate, and used to produce deupleted SCFAs.
The mevalonate pathway is another possible source of formate synthesis in cancer cells. Processing of acetyl-CoA through the mevalonate pathway is often upregulated by cancer cells [129]. Increased demand for the mevalonate pathway is a hallmark of oncogenesis [130]. The mevalonate pathway results in the synthesis of cholesterol from three acetate molecules, and formate is released as a side product. Many other metabolites of the mevalonate pathway are also of use to cancer cells. Formate is also released in the demethylation of androstenedione and testosterone by aromatase to form estrone and estradiol, respectively [126]. Aromatase overexpression in breast cancer and endometrial cancer contributes significantly to neoplastic transformation induced by estrogen [131]. Overexpression of aromatase in the testes promotes testicular cancer [132].
Another source of formate is through metabolism of tryptophan through the kynurenine pathway. Various enzymes convert tryptophan to formyl-kynurenine, which then becomes kynurenine after the release of free formate. Many cancers upregulate the expression of tryptophan transporters and the activity of indoleamine dioxygenase (IDO), the enyzme that produces formyl-tryptophan [133]. IDO1 is overexpressed in the vast majority of cancers, and it plays an important role in tumor cell evasion of both the innate and the adaptive immune systems [134].
11. Deuterium-depleted water (DDW) therapy as a treatment for cancer
Accumulating experimental evidence from in vitro and in vivo studies, as well as human clinical studies, shows that treatment with DDW exerts strong anti-tumor activity [6,7]. The following subsections are dedicated to presenting, analyzing and interpreting the observations obtained from these clinical trials. Determining exactly how DDW suppresses cancer growth has been a challenging task for the research community.
11.1. Human Clinical Studies Show Promising Anti-Cancer Results with DDW Treatment
Human clinical studies provide data that describe a potent anti-cancer activity obtained by DDW treatment. Glioblastoma multiforme (GBM) is an aggressive brain tumor with a relatively short median survival time (MST), especially at later onsets of disease [135]. In the clinical study of G Somlyai et al., patients who had sustained surgical excision of a brain tumor consumed wholly DDW as a daily fluid, to reduce the deuterium content in their body [136]. The DDW intake was given as an adjuvant therapy to chemotherapy and radiation. The DDW treatment in this study extended their MST from 14.6 months to 30 months. DDW was especially effective for female patients; their MST was increased to 42 months. Moreover, patients who had started DDW therapy concurrently with radiation therapy lived for 47 months.
Pancreatic cancer is a more aggressive and invasive type of cancer, occurring not only in the elderly but also in younger individuals [137]. A study by LG Boros et al. showed that DDW treatment in combination with conventional chemotherapy in pancreatic cancer patients remarkably increased their MST from 6.36 months to 19.6 months. The authors paralleled the clinical study results with in vitro experiments. In these experiments, the pancreatic cancer cells were grown in DDW that had differential concentrations of low deuterium content. The in vitro results showed that the lower the deuterium concentration in DDW, the more the pancreatic cancer cells were inhibited in their proliferation capability. Moreover, the low proliferation rate prostate cancer cells (having low cell viability) had complementary, marked defects in their lipid membranes and glucose metabolism. In another clinical trial involving prostate cancer patients, it was shown that the consumption of DDW contributed to the overall decrease in tumor size and increased the MST of the patients. The DDW intake in the prostate cancer patients matched with the low concentrations of their prostate serum antigen (PSA). Moreover, DDW treatment prolonged significantly the MST of patients who suffered from distant metastasis [138].
In summary, DDW treatment is a promising anti-cancer approach as an adjuvant to either chemotherapy and/or radiation therapy, since cancer cells are sensitized to die upon deuterium depletion. DDW therapy is a non-toxic anti-cancer strategy that has proved its efficacy in experimental, animal and human clinical trials [139].
11.2. The Role of Cell Cycle Arrest
Deuteration (the exchange of hydrogen with deuterium) of active biomolecule structures involved in mitochondrial metabolism has long been shown to affect the normal mechanisms of hydrolysis of ATP [140]. In the study of A. Olgun, the author concluded: Deuteronation, as exemplified by ATP synthase and the electron transport chain (ETC), may interfere with the conformations and functions of many macromolecules and contribute to some pathologies like heavy water toxicity and aging [12].
In the case of cancer, one of the hallmarks to achieve anti-tumor activity is to increase the rate of autophagy and/or apoptosis of malignant cells at certain stages of cancer [141]. Stimulation of autophagy can reduce the toxic effects of deuterium through the turnover of defective mitochondria and heavily deuterated macromolecules. Autophagic mechanisms must be working efficiently in order to discard defective highly deuterated organelles [12]. Thus, the effects of deuteronation can be delayed or prevented by decreasing the intake of deuterium and/or increasing the turnover of organelles and macromolecules by stimulating autophagy.
It has recently become possible to produce water that is severely depleted in deuterium, and this has led to experimental trials to see whether deuterium depleted water (DDW) is useful therapeutically [142]. Experimental trials revealed that the application of DDW treatment promotes senescence of breast cancer cells [143]. Moreover, experiments on colorectal cancer cells treated with DDW showed similar senescent phenomena as in breast cancer cells, but also revealed a pro-apoptotic effect, suggesting a DDW-related promotion of autophagic and cancer cell death mechanisms [144]. In both cases the promotion of senescence and cancer cell death was accompanied by a certain micro-RNA (miRNA) profile that was induced by the treatment with DDW. Most intriguingly, further experiments on breast cancer cells showed that treatment with DDW had a direct growth inhibitory effect and promoted the anti-cancer activity of fluorouracil (5FU) [145]. The growth inhibition of the above-mentioned cancer cells was due to a G1/S cell cycle arrest. More recent experiments showed that DDW induces a similar G1/S phase cell cycle arrest that causes the death of colorectal cancer cells [146]. In another study, DDW was found to inhibit the proliferation, migration and invasion of nasopharyngeal carcinoma cells grown in culture. Strikingly, by contrast, growth of normal pre-osteoblast cells (mesenchymal progenitors of osteoblasts) was promoted in the presence of DDW [147].
11.3. Understanding the Mechanism of Tumor Suppression by DDW
Probably the most thorough study ever published involving attempts to understand exactly how DDW arrests tumor growth, causes oxidative stress, and induces apoptosis in tumor cells is a paper published by Zhang et al. in 2019 [148]. This paper used a proteomics approach to reveal a disbalance between ROS production and ROS neutralization brought on by DDW exposure, leading to suppression of activity of a number of proteins involved in maintaining tumor growth. Tumor cells (specifically, A549 cells) grown in culture exposed to DDW (80 ppm deuterium) experienced a 32% inhibition of cell proliferation.
The regulatory protein p62, also called sequestosome 1, was highly oxidized in response to the treatment [148]. p62 is a powerful signaling protein that is involved in regulating cancer metabolism [149]. p62 is highly expressed in most cancer cells, and decreasing its levels lowers proliferation rates [149]. p62 is an autophagy receptor protein that activates autophagy pathways and plays a critical role in apoptosis [150]. p62 upregulation and/or reduced degradation have been implicated in tumor formation, cancer promotion, and resistance to therapy [150]. Oxidation of p62 promotes its oligomerization which in turn leads to increased autophagy, including autophagy (and clearance) of p62 itself [151].
Extensive proteomics analysis revealed several specific enzymes whose expression was significantly downregulated in response to DDW treatment. The downregulated enzymes highlighted in the Zhang et al. paper are listed in Table 1. It is remarkable that most of these enzymes carry out reactions that involve either NADH or NADPH. Downregulation of ALDH4A1 would impair the ability of the tumor cell to convert proline to glutamate, decreasing its supply of glutamate, which is an important alternative fuel in the TCA cycle in cancer cells [152]. Glutamate deprivation in prostate cancer cells decreased proliferation, migration, and invasion, leading to apoptotic cell death [153]. Both FDXR and H6PD replenish NADPH, which is needed to maintain glutathione in its reduced state. In parallel, GPX downregulation suppresses the ability to detoxify H2O2 in the mitochondria via glutathione oxidation. DHFR regenerates H4F from H2F, so its suppression leads to a reduced supply of H4F. H4F is an essential coenzyme for de novo synthesis of both pyrimidine and purine nucleotides, needed for DNA synthesis in proliferating cells. DHFR inhibitors such as methotrexate are common antifolate drugs used in cancer therapy [154].
The most highly oxidized enzyme determined by Zhang et al. was hydroxymethylglutaryl CoA synthase 1 (HMGCS1), the first step in the mevalonate pathway. This pathway leads to the synthesis of cholesterol, which is essential for assembling the plasma membranes of proliferating cells. Suppression of the mevalonate pathway lowers the mitochondrial membrane potential and increases the release of pro-apoptotic factors [155]. Several other products of the mevalonate pathway are also essential for cellular health. Depletion of isoprenoids such as dolichol, ubiquinone, and farnesylated and geranygeranylated proteins leads to ER stress and apoptosis in liver cells [156].
Overall, it appears that exposure to DDW results in a complex signaling cascade in tumor cells that induces metabolic changes that interfere with proliferation and promote autophagy and apoptosis. It may not be the case that DDW is actually toxic to the tumor cells, but rather that they have been programmed to respond to low deuterium in such a way so as to shut down tumor growth and induce apoptotic cellular death.
12. Discussion
Deuterium is a natural heavy isotope of hydrogen that has distinct biophysical and biochemical properties, and it can damage mitochondrial ATPase pumps, leading to inefficient ATP synthesis and excessive production of reactive oxygen species [12]. Biological organisms have evolved to have clever mechanisms to reduce deuterium levels in the mitochondria, and the gut microbiome plays an essential role in supplying deupleted nutrients for the host, including the short chain fatty acids acetate, propionate and butyrate, and the methyl groups that participate in methylation pathways. The microbes collectively achieve this feat by synthesizing significantly deupleted H2 gas and using it as a reducing agent to synthesize deupleted small organic molecules [31,38].
Mitochondrial dysfunction is linked to many diseases, most notably cancer [17]. Here, we have presented extensive evidence that many of the unusual aspects of cancer metabolic policies can be explained as a strategy to produce and release deupleted nutrients and deupleted protons into the external milieu, while concentrating deuterium internally. By severely restricting the activity of the ATPase pumps, tumor cells protect their defective mitochondria from the primary damaging effects of deuterium. M2-like TAMs infiltrating the tumor microenvironment promote tumor growth, angiogenesis and metastasis, and signal to resident T cells to stand down [117]. We hypothesize that nutrients supplied by the tumor can support mitochondrial and lysosomal healing of the immune cells, eventually enabling them to become activated and destroy the tumor, unless the deuterium overload problem is too severe.
12.1. DDW, Autophagy and Apoptosis
When the medium is enriched in deuterium, tumor cells thrive, whereas they commit apoptosis in deupleted water [6]. The opposite is true for normal cells [7]. DDW has been found in many studies to be a promising therapy to treat cancer, with minimal side effects [139,145,148]. It is still not clear exactly how DDW works, but it appears that cancer cells can sense deuterium levels and respond through signaling pathways to launch an apoptotic program. DDW treatment may restore oxidative phosphorylation and result in an accumulation of ROS in cancer cells, which results in accelerated aging and shrinkage of the tumor, due perhaps to deficient antioxidant defenses and impaired ability to repair damaged DNA [148]. More research is needed to help clarify the effects of DDW on tumor cells.
The molecular mechanisms generating an autophagic response in cancer cells are tightly linked with the final outcome of apoptosis [157]. Targeting autophagy to induce apoptosis in tumor cells, however, is a complicated and unpredictable therapeutic strategy. The response depends on which kind of tumor is being targeted, and at which stage of oncogenesis autophagy is inhibited. Generally, at initial stages of oncogenesis, the cancer cells need autophagic recycled molecules to maintain their excessive proliferative metabolism. When autophagy is inhibited at this stage, a provoked cancer cell death by apoptosis can be achieved [158]. In contrast, in certain cancers, such as lung cancer, restoring the autophagy response during early oncogenesis can have an antitumor effect. However, in this type of cancer, it is the autophagy at the later stages of oncogenesis that promotes cancer progression via a dysregulated T regulatory cell response and an impaired ability of mutant p53 activity to safeguard the genome [159]. Therefore, using DDW to promote autophagy and apoptosis in cancer cells needs further clinical research evaluations to define at which stage of oncogenesis and which type of cancer this can be applied to to gain the maximum anti-cancer effects.
12.2. The Crosstalk between Senescence, Apoptosis and Cell Cycle Arrest Induced by DDW Treatment in Cancer
Under normal conditions, the cell will be promoted to induce cell cycle arrest via the transcriptional activation of the p21 gene by the tumor suppressor protein p53. p53 forms a complex with p21 that initiates a molecular cascade to inhibit cell cycle progression to mitosis (for a detailed review see [160]). Autophagy, on the other hand, is inhibited as the cell progresses to mitotic division (for a comprehensive review see [161]). Initiation of autophagy and cell cycle arrest share common molecular pathways, where, in both cases, the activity of p53 is catalytic. In most cases of cancer, however, either p53 is mutated or the cell suffers from biological inactivation of its related pathways that otherwise safeguard the genome [162]. Indeed, by contrast, in cancer cells, the mutant isoforms of p53 do not induce cell cycle arrest, but rather promote the continuation of cellular proliferation and oncogenesis that leads to cancer [163].
What is most intriguing about the gene activity of Tp53 and other oncogenes, namely c-myc, and Ha-ras, during deuterium depletion, is that they become inhibited, as shown in experiments with animals exposed to carcinogens [164]. In the seminal study of X Zhang et al., it was found that the p53 signaling pathway was the most downregulated signaling pathway of DDW-treated lung cancer cells [165]. A possible explanation for how DDW might induce cell cycle arrest in cancer cells, as shown in in vitro, animal and human studies, is the reduction of intracellular content of deuterium which then blocks the expression of cancer-related genes [166]. DDW, by delaying the expression rate of Tp53, also reduces the concentration of mutant p53 proteins that induce cellular proliferation and oncogenesis. This will prevent or can even stop cancer proliferation and development. Moreover, the shift (lowering) of the D to H ratio in the cytosol in malignant cells by DDW treatment reduces the metabolic activity that cancer cells are in most need of, and this will initiate apoptosis, autophagy and senescence, molecular and cellular phenomena that are coupled with cell cycle arrest [1,143,144].
Senescence is a permanent phenomenon of cell cycle arrest in which the cells age and are unresponsive to mechanisms that induce the repair of their DNA damage events [167]. The incorporation of deuterium into nucleic acid chains can break the hydrogen bonds between the nitrogenous bases of the DNA, and this creates opportunities, upon deuterium exchange with the lighter hydrogen, for DNA breaks [168]. It is therefore plausible to hypothesize that, during deupletion of the DNA (which will occur during DDW treatment), cancer cells, especially the cancer cells that lack the necessary DNA repair machinery, can’t cope with DNA damage events [1]. This aging would be a death mechanism for the cancer cells.
Furthermore, the incubation of lung cancer cells in DDW shows a substantial accumulation of reactive oxygen species (ROS). Notably, in the cancer cells, the normal mitochondrial function for depleting deuterium is disrupted, and therefore deuterium accumulates. It is plausible that, during deuterium depletion in DDW treated cancer cells, and when the antioxidant defenses (e.g., glutathione metabolism) of these cancer cells is insufficient [165], ROS accumulate, and the cancer cells finally suffer from accelerated aging and apoptosis [139].
12.3. DDW and V-ATPase
It is possible that one way in which DDW affects cancer cells is by decreasing the activity of V-ATPase pumps. These pumps reverse the polarity of the membrane by pumping protons out, and they have a very high deuterium KIE [95]. Researchers have discovered that there exist organic chromophores that can distinguish D2O from H2O based on their slight difference in acidity. This feature has been exploited in the design of D2O optical sensors to quantitatively detect levels of D2O in water [169]. It is therefore conceivable that biological organisms possess deuterium sensors that could have signaling capabilities to suppress the membrane-bound ATPase pumps under low deuterium conditions. V- ATPase pumps play a major role in evading apoptosis, increasing multidrug resistance, and promoting cell proliferation, migration and invasion [92]. V-ATPase inhibitors have been shown to cause slowed growth and increased cell death of cancer cells, simply by restoring the membrane polarity to the normal state [170].
There are many aspects of deuterium homeostasis that require further exploration. For example, heavy deuteration stabilizes tubulin [171]. Tubulin forms an intracellular code for cancer cells to progress in extensive mitosis and uncontrolled proliferation. Nowadays, the inhibition of microtubule formation of cancer cells constitutes a hallmark of anti-cancer therapy [172]. It is likely that DDW treatment disrupts deuterated tubulin formations, and, for this reason, deupletion also prevents cancer cells from proliferating, thus discontinuing their cell cycle.
12.4. Food as Medicine
Through our investigations into deuterium homeostasis, we have come to believe that the best way to both prevent and treat cancer is to maintain a wholesome diet of organic whole foods that are rich in micronutrients. Ultra-processed foods, which are deficient in micronutrients and highly contaminated with toxic additives, have become a growing percentage of the diet in modern times, and their overconsumption is associated with many adverse health outcomes [173]. It is essential to consume adequate amounts of the B vitamins, niacin, riboflavin and folate to assure abundant supplies of NAD, FAD, and methyltetrahydrofolate. Butter is an excellent source of butyrate, and it is low in deuterium. Animal-based fats and coconut oil are other low-deuterium nutrients. A high fiber diet can supply prebiotics to support butyrate synthesis by the gut microbiota. Choline (abundant in animal-based foods) is a good source of methyl groups, as well as betaine and the amino acids glycine, alanine and methionine. Eating certified organic foods will minimize exposure to toxic agrichemicals that could be disrupting the gut microbiome. Fermented foods are a great source of acetate, and they also support renewal of the gut microbiome.
13. Conclusions
Deuterium, a heavy isotope of hydrogen, is a pervasive natural element that presents special challenges to biological organisms. The mitochondrial ATPase pumps are very sensitive to deuterium, which accumulates in the motor and causes stutters, leading to an increase in reactive oxygen species and inefficiencies in ATP production. The gut microbiome plays a significant role in supplying deupleted nutrients to the host, via hydrogen recycling. We have presented evidence that oncogenesis may be driven by deuterium overload in mitochondria, and that the unusual metabolic policies of cancer cells can be explained as a strategy to hoard deuterium and supply deupleted nutrients to the resident immune cells as well as to distant organs. The importance of methylation pathways may be primarily due to the low deuterium content in the methyl groups. Deuterium depleted water has been found to have significant therapeutic value in treating cancer. An organic diet that is rich in low-deuterium nutrients, prebiotics, probiotics, and B vitamins is a good strategy for reducing cancer risk. DDW treatment for cancer needs a thorough laboratory clinical evaluation to discover its influence in autophagic related death by apoptosis at different stages of oncogenesis in different cancer types.
Funding
This research was funded in part by Quanta Computer, Inc., Taiwan, under grant number 6941564.
Conflicts of Interest
The authors deny any conflicts of interest.
References
- Yaglova, N.V.; Timokhina, E.P.; Obernikhin, S.S.; Yaglov, V.V. Emerging role of deuterium/protium disbalance in cell cycle and apoptosis. Int J Mol Sci. 2023, 24, 3107. [Google Scholar] [CrossRef] [PubMed]
- Boros, L.G.; Seneff, S.; Tric, M.; Palcsuc, L.; Roman, A.; Zubarev, R.A. Active involvement of compartmental, inter- and intramolecular deuterium disequilibrium in adaptive biology. PNAS 2024, 121, e2412390121. [Google Scholar] [CrossRef]
- Bachner, P.; McKay, D.G.; Rittenberg, D. The pathologic anatomy of deuterium intoxication. PNAS 1964, 51, 464–471. [Google Scholar] [CrossRef]
- Xie, X.; Zubarev, R.A. Effects of low-level deuterium enrichment on bacterial growth. PLoS One. 2014, 17, 9–e102071. [Google Scholar] [CrossRef] [PubMed]
- Paliy, O.; Bloor, D.; Brockwell, D.; Gilbert, P.; Barber, J. Improved methods of cultivation and production of deuteriated proteins from E. coli strains grown on fully deuteriated minimal medium. J Appl Microbiol. 2003, 94, 580–6. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Zubarev, R.A. Slight deuterium enrichment in water acts as an antioxidant: is deuterium a cell growth regulator? Mol Cell Proteomics. 2020, 19, 1790–1804. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhu, B.; Liu, C.; Fang, W.; Yang, H. Deuterium-depleted water selectively inhibits nasopharyngeal carcinoma cell proliferation in vitro. Nan Fang Yi Ke Da Xue Xue Bao. [article in Chinese]. 2021, 32, 1394-9. [Google Scholar]
- Stein, L.R.; Imai, S. The dynamic regulation of NAD metabolism in mitochondria. Trends Endocrinol Metab. 2012, 23, 420–8. [Google Scholar] [CrossRef]
- Boros, L.G.; Somlyai, I.; Kovács, B.Z.; Puskás, L.G.; Nagy, L.I.; Dux, L.; Farkas, G.; Somlyai, G. Deuterium depletion inhibits cell proliferation, RNA and nuclear membrane turnover to enhance survival in pancreatic cancer. Cancer Control. 2021, 28, 1073274821999655. [Google Scholar] [CrossRef]
- Jiang, C.; Liu, Y.; Wang, L.; Lu, F. Interaction between heavy water and single-strand DNA: A SERS study. Molecules. 2022, 27d, :6023. [CrossRef]
- Zlatska, A.; Gordiienko, I.; Vasyliev, R.; Zubov, D.; Gubar, O.; Rodnichenko, A.; Syroeshkin, A.; Zlatskiy, I. In vitro study of deuterium effect on biological properties of human cultured adipose-derived stem cells. Scientific World Journal. 2018, 2018, 5454367. [Google Scholar] [CrossRef]
- Olgun, A. Biological effects of deuteronation: ATP synthase as an example. Theor Biol Med Model 2007, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Murcia Rios, A.; Vahidi, S.; Dunn, S.D.; Konermann, L. Evidence for a partially stalled γ rotor in F1-ATPase from hydrogen-deuterium exchange experiments and molecular dynamics simulations. J Am Chem Soc. 2018, 140, 14860–14869. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A.P.; Haddad, M.Kh.; Fecht, H.-J. Light effect on water viscosity: Implication for ATP biosynthesis. Sci Rep 2015, 5, 12029. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Zhang, K.; Arakawa, H. Recent advances and new perspectives in mitochondrial dysfunction. Sci Rep 2023, 13, 7977. [Google Scholar] [CrossRef]
- Nicolson, G.L. Mitochondrial dysfunction and chronic disease: treatment with natural supplements. Integr Med (Encinitas). 2014, 13, 35–43. [Google Scholar]
- Luo, Y.; Ma, J.; Lu, W. The significance of mitochondrial dysfunction in cancer. Int J Mol Sci. 2020, 21, 5598. [Google Scholar] [CrossRef]
- Navratil, A.R.; Shchepinov, M.S.; Dennis, E.A. Lipidomics reveals dramatic physiological kinetic isotope effects during the enzymatic oxygenation of polyunsaturated fatty acids ex vivo. J Am Chem Soc. 2018, 140, 235–243. [Google Scholar] [CrossRef]
- Schleucher, J.; Vanderveer, P.; Markley, J.L.; Sharkey, T.D. Intramolecular deuterium distributions reveal disequilibrium of chloroplast phosphoglucose isomerase. Plant, Cell & Environment 1999, 22, 525–533. [Google Scholar] [CrossRef]
- Leadlay, P.F.; Albery, W.J.; Knowles, J.R. Energetics of triosephosphate isomerase: deuterium isotope effects in the enzyme-catalyzed reaction. Biochemistry. 1976, 15, 5617–20. [Google Scholar] [CrossRef]
- Henkel, S.; Ertelt, M.; Sander, W. Deuterium and hydrogen tunneling in the hydrogenation of 4-oxocyclohexa-2,5-dienylidene. Chemistry. 2014, 20, 7585–8. [Google Scholar] [CrossRef]
- Sutcliffe, M.J.; Scrutton, N.S. A new conceptual framework for enzyme catalysis. Hydrogen tunnelling coupled to enzyme dynamics in flavoprotein and quinoprotein enzymes. Eur J Biochem. 2002, 269, 3096–102. [Google Scholar] [CrossRef] [PubMed]
- Hay, S.; Pudney, C.R.; Scrutton, N.S. Structural and mechanistic aspects of flavoproteins: probes of hydrogen tunnelling. FEBS J. 2009, 276, 3930–41. [Google Scholar] [CrossRef] [PubMed]
- Ballou, S.; Singh, P.; Nee, J.; Rangan, V.; Iturrino, J.; Geeganage, G.; Lwe, B.; Bangdiwala, S.I.; Palsson, O.S.; Sperber, AD.; et al. Prevalence and associated factors of bloating: Results from the Rome Foundation global epidemiology study. Gastroenterology 2023, 165, 647-655.e4. [Google Scholar] [CrossRef] [PubMed]
- Vargas, D.; Chimborazo, O.; László, E.; Temovski, M.; Palcsu, L. Rainwater isotopic composition in the Ecuadorian Andes and Amazon reflects cross-equatorial flow seasonality. Water 2022, 14, 2121. [Google Scholar] [CrossRef]
- Petit, J.R.; White, W.C.; Young, N.W.; Jouzel, J.; Korotkevich, Y.S. Deuterium excess in recent Antarctic snow. J Geophys Res. 1991, 96(D3), 5113–5122. [Google Scholar] [CrossRef]
- Ciais, P.; White, J.W.C.; Jouzel, J.; Petit, J.R. The origin of present-day Antarctic precipitation from surface snow deuterium excess data. J Geophys Res. 1985, 100, 1891718927. [Google Scholar]
- Sokol, K.P.; Robinson, W.E.; Oliveira, A.R.; Zacarias, S.; Lee, C.Y.; Madden, C.; Bassegoda, A.; Hirst, J.; Pereira, I.A.C.; Reisner, E. Reversible and selective interconversion of hydrogen and carbon dioxide into formate by a semiartificial formate hydrogenlyase mimic. J Am Chem Soc. 2019, 141, 17498–17502. [Google Scholar] [CrossRef]
- Ohta, S. Recent progress toward hydrogen medicine: potential of molecular hydrogen for preventive and therapeutic applications. Curr Pharm Des. 2011, 17, 2241–52. [Google Scholar] [CrossRef]
- Ichikawa, Y.; Yamamoto, H.; Hirano, S.I.; Sato, B.; Takefuji, Y.; Satoh, F. The overlooked benefits of hydrogen-producing bacteria. Med Gas Res. 2023, 13, 108–111. [Google Scholar] [CrossRef]
- Krichevsky, M.I.; Friedman, I.; Newell, M.F.; Sisler, F.D. Deuterium fractionation during molecular hydrogen formation in marine pseudomonad. J Biol Chem. 1961, 236, 2520–5. [Google Scholar] [CrossRef]
- Klein, V.J.; Irla, M.; Gil López, M.; Brautaset, T.; Fernandes Brito, L. Unravelling formaldehyde metabolism in bacteria: Road towards synthetic methylotrophy. Microorganisms. 2022, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Marx, C.J.; Van Dien, S.J.; Lidstrom, M.E. Flux analysis uncovers key role of functional redundancy in formaldehyde meftabolism. PLoS Biol. 2005, 3, e16. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; Chouchani, E.T. Why succinate? Physiological regulation by a mitochondrial coenzyme Q sentinel. Nat Chem Biol. 2022, 18, 461–469. [Google Scholar] [CrossRef]
- Gerecke, C.; Egea Rodrigues, C.; Homann, T.; Kleuser, B. The role of ten-eleven translocation proteins in inflammation. Front Immunol. 2022, 13, 861351. [Google Scholar] [CrossRef]
- Torao, E.G.; Petrus, S.; Fernandez, A.F.; Fraga, M.l.F. Global DNA hypomethylation in cancer: review of validated methods and clinical significance. Clin Chem Lab Med. 2012, 50, 1733-42. [Google Scholar] [CrossRef]
- Li, J.; Huang, Q.; Zeng, F.; Li, W.; He, Z.; Chen, W.; Zhu, W.; Zhang, B. The prognostic value of global DNA hypomethylation in cancer: a meta-analysis. PLoS One. 2014, 9, e106290. [Google Scholar] [CrossRef]
- Smith, N.W.; Shorten, P.R.; Altermann, E.H.; Roy, N.C.; McNabb, W.C. Hydrogen cross-feeders of the human gastrointestinal tract. Gut Microbes. 2019, 10, 270–288. [Google Scholar] [CrossRef]
- Lajoie, S.F.; Bank, S.; Miller, T.L.; Wolin, M.J. Acetate production from hydrogen and [13C]carbon dioxide by the microflora of human feces. Appl Environ Microbiol. 1988, 54, 2723–7. [Google Scholar] [CrossRef] [PubMed]
- Bernalier, A.; Lelait, M.; Rochet, V.; Grivet, J.-P.; Gibson, G.R. Durand M. Acetogenesis from H, and CO, by methane- and non-methane-producing human colonic bacterial communities. FEMS Microbiology Ecology. 1996, 19, 193–202. [Google Scholar] [CrossRef]
- Bernalier, A.; Rochet, V.; Leclerc, M.; Dor, J.; Pochart, P. Diversity of H2/ CO2-utilizing acetogenic bacteria from feces of non-methane-producing humans. Curr Microbiol. 1996, 33, 94–9. [Google Scholar] [CrossRef]
- Recharla, N.; Geesala, R.; Shi, X.Z. Gut microbial metabolite butyrate and its therapeutic role in inflammatory bowel disease: A literature review. Nutrients. 2023, 15, 2275. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, S.; Layec, S.; Auger, S.; Juste, C.; Henry, C.; Charif, S.; Jaszczyszyn, Y.; Sokol, H.; Beney, L.; Langella, P.; et al. E. Faecalibacterium duncaniae A2-165 regulates the expression of butyrate synthesis, ferrous iron uptake, and stress-response genes based on acetate consumption. Sci Rep. 2024, 14, 987. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Holtrop, G.; Lobley, G.E.; Calder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of acetate to butyrate formation by human faecal bacteria. Br J Nutr. 2004, 91, 915–23. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, MA. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Sun, J.; Chen, S.; Zang, D.; Sun, H.; Sun, Y.; Chen, J. Butyrate as a promising therapeutic target in cancer: From pathogenesis to clinic (Review). Int J Oncol. 2024, 64, 44. [Google Scholar] [CrossRef] [PubMed]
- Son, M.Y.; Cho, H.S. Anticancer Effects of Gut Microbiota-Derived Short-Chain Fatty Acids in Cancers. J Microbiol Biotechnol. 2023, 33, 849–856. [Google Scholar] [CrossRef]
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 2003, 133, 2485S2493S. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of gut microbiota-generated short-chain fatty acids in metabolic and cardiovascular health. Curr Nutr Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Glozak, M.A.; Seto, E. Histone deacetylases and cancer. Oncogene. 2007, 26, 5420–32. [Google Scholar] [CrossRef]
- Di Gennaro, E.; Bruzzese, F.; Caraglia, M.; Abruzzese, A.; Budillon, A. Acetylation of proteins as novel target for antitumor therapy: review article. Amino Acids. 2004, 26, 435–41. [Google Scholar] [CrossRef]
- Shin, H.; Lee, Y.S.; Lee, Y.C. Sodium butyrate-induced DAPK-mediated apoptosis in human gastric cancer cells. Oncol Rep. 2012, 27, 1111–5. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, K.N.; Vitetta, L. Effects of intestinal microbial-elaborated butyrate on oncogenic signaling pathways. Nutrients. 2019, 11, 1026. [Google Scholar] [CrossRef]
- Xu, S.; Liu, C.X.; Xu, W.; Huang, L.; Zhao, J.Y.; Zhao, S.M. Butyrate induces apoptosis by activating PDC and inhibiting complex I through SIRT3 inactivation. Signal Transduct Target Ther. 2017, 2, 16035. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol (Lausanne). 2020, 11, 25. [Google Scholar] [CrossRef]
- Oka, Y.; Nakazawa, Y.; Shimada, M.; Ogi, T. Endogenous aldehyde-induced DNA-protein crosslinks are resolved by transcription-coupled repair. Nat Cell Biol. 2024, 26, 784–796. [Google Scholar] [CrossRef]
- Barnett, S.D.; Buxton, I.L.O. The role of S-nitrosoglutathione reductase (GSNOR) in human disease and therapy. Crit Rev Biochem Mol Biol. 2017, 52, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Umansky, C.; Morellato, A.E.; Rieckher, M.; Scheidegger, M.A.; Martinefski, M.R.; Fernndez, G.A.; Pak, O.; Kolesnikova, K.; Reingruber, H.; Bollini, M.; et al. Endogenous formaldehyde scavenges cellular glutathione resulting in redox disruption and cytotoxicity. Nat Commun. 2022, 13, 745. [Google Scholar] [CrossRef]
- Chang, J.S.; Hsiao, JR.; Chen, CH. ALDH2 polymorphism and alcohol-related cancers in Asians: a public health perspective. J Biomed Sci. 2017, 24, 19. [Google Scholar] [CrossRef]
- Wu, M.; Chang, S.C.; Kampman, E.; Yang, J.; Wang, X.S.; Gu, X.P.; Han, R.Q.; Liu, A.M.; Wallar, G.; Zhou, J.Y.; et al. Single nucleotide polymorphisms of ADH1B, ADH1C and ALDH2 genes and esophageal cancer: a population-based case-control study in China. Int J Cancer. 2013, 132, 1868–77. [Google Scholar] [CrossRef]
- Alter, B.P. Fanconi anemia and the development of leukemia. Best Pract Res Clin Haematol. 2014, 27, 214-21. [Google Scholar] [CrossRef]
- Oka, Y.; Hamada, M.; Nakazawa, Y.; Muramatsu, H.; Okuno, Y.; Higasa, K.; Shimada, M.; Takeshima, H.; Hanada, K.; Hirano, T.; et al. Digenic mutations in ALDH2 and ADH5 impair formaldehyde clearance and cause a multisystem disorder, AMeD syndrome. Sci Adv. 2020, 6, eabd7197. [Google Scholar] [CrossRef] [PubMed]
- Blusztajn, J.K.; Liscovitch, M.; Richardson, U.I. Synthesis of acetylcholine from choline derived from phosphatidylcholine in a human neuronal cell line. Proc Natl Acad Sci U S A. 1987, 84, 5474–7. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Davis, C.D.; Uthus, E.O. DNA methylation, cancer susceptibility, and nutrient interactions. Exp Biol Med (Maywood). 2004, 229, 988–95. [Google Scholar] [CrossRef] [PubMed]
- Szigeti, K.A.; Kalmár, A.; Galamb, O.; Valcz, G.; Barták, B.K.; Nagy, Z.B.; Zsigrai, S.; Felletr, I.; V. Patai, A.; Micsik, T.; et al. Global DNA hypomethylation of colorectal tumours detected in tissue and liquid biopsies may be related to decreased methyl-donor content. BMC Cancer 2022, 22, 605. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.S.; Fang, Y.J.; Pan, Z.Z.; Zhong, X.; Zheng, M.C.; Chen, Y.M.; Zhang, C.X. Choline and betaine intake and colorectal cancer risk in Chinese population: a case-control study. PLoS One. 2015, 10, e0118661. [Google Scholar] [CrossRef]
- Zhen, J.; Zhou, Z.; He, M.; Han, H.X.; Lv, E.H.; Wen, P.B.; Liu, X.; Wang, Y.T.; Cai, X.C.; Tian, J.Q.; et al. The gut microbial metabolite trimethylamine N-oxide and cardiovascular diseases. Front Endocrinol (Lausanne). 2023, 14, 1085041. [Google Scholar] [CrossRef] [PubMed]
- Dridi, B.; Fardeau, M.L.; Ollivier, B.; Raoult, D.; Drancourt, M. Methanomassiliicoccus luminyensis gen. nov., sp. nov., a methanogenic archaeon isolated from human faeces. Int J Syst Evol Microbiol. 2012, 62(Pt 8), 1902-1907. [Google Scholar] [CrossRef]
- Chhibber-Goel, J.; Gaur, A.; Singhal, V.; Parakh, N.; Bhargava, B.; Sharma, A. The complex metabolism of trimethylamine in humans: endogenous and exogenous sources. Expert Rev Mol Med. 2016, 18, e19. [Google Scholar] [CrossRef]
- Brugre, J.F.; Borrel, G.; Gaci, N.; Tottey, W.; O’Toole, P.W.; Malpuech-Brugre, C. Archaebiotics: proposed therapeutic use of archaea to prevent trimethylaminuria and cardiovascular disease. Gut Microbes. 2014, 5, 5–10. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Spector, T.D.; Youngblut, N.D.; Ley, R.E. Genomic insights into adaptations of trimethylamine-utilizing methanogens to diverse habitats, including the human gut. mSystems. 2021, 6, e00939–20. [Google Scholar] [CrossRef]
- Janeiro, M.H.; Ramírez, M.J.; Milagro, F.I.; Martínez, J.A.; Solas, M. Implication of trimethylamine N-oxide (TMAO) in disease: Potential biomarker or new therapeutic target. Nutrients. 2018, 10, 1398. [Google Scholar] [CrossRef]
- Ramezani, A.; Nolin, T.D.; Barrows, I.R.; Serrano, M.G.; Buck, G.A.; Regunathan-Shenk, R.; West, R.E. 3rd.; Latham, P.S.; Amdur, R.; Raj, D.S. Gut colonization with methanogenic archaea lowers plasma trimethylamine N-oxide concentrations in apolipoprotein E-/- mice. Sci Rep. 2018, 8, 14752. [Google Scholar] [CrossRef]
- Ahmed, G.Y.; Osman, A.A.; Mukhtar, A. Acetylcholinesterase enzyme among cancer patients a potential diagnostic and prognostic indicator a multicenter case-control study. Sci Rep. 2024, 14, 5127. [Google Scholar] [CrossRef]
- Faubert, B.; Li, K.Y.; Cai, L.; Hensley, C.T.; Kim, J.; Zacharias, L.G.; Yang, C.; Do, Q.N.; Doucette, S.; Burguete, D.; et al. Lactate metabolism in human lung tumors. Cell 2017, 171, 358-371.e9. [Google Scholar] [CrossRef]
- Zewude, R.T.; Croitoru, K.; Das, R.; Goldman, B.; Bogoch, II. Auto-brewery syndrome in a 50-year-old woman. CMAJ 2024, 196, E724–E727. [Google Scholar] [CrossRef]
- Tameez Ud Din, A.; Alam, F.; Tameez-Ud-Din, A.; Chaudhary, F.M.D. Auto-brewery syndrome: a clinical dilemma. Cureus 2020, 12, e10983. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; Enerbäc, S. Lactate: the ugly duckling of energy metabolism. Nat Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef]
- Xiao, T.; Khan, A.; Shen, Y.; Chen, L.; Rabinowitz, J.D. Glucose feeds the tricarboxylic acid cycle via excreted ethanol in fermenting yeast. Nat Chem Biol 2022, 18, 1380–1387. [Google Scholar] [CrossRef]
- Somlyai, G.; Gyöngyi, Z.; Somlyai, I.; Boros, L.G. Pre-clinical and clinical data confirm the anticancer effect of deuterium depletion. Eur. J. Integr. Med. 2016, 8, 28. [Google Scholar] [CrossRef]
- Somlyai, G.; Kovács, B.Z.; Papp, A.; Somlyai, I. A preliminary study indicating improvement in the median survival time of glioblastoma multiforme patients by the application of deuterium depletion in combination with conventional therapy. Biomedicines. 2023, 13, 11–1989. [Google Scholar] [CrossRef]
- Yu, D.; Liu, Z. The research progress in the interaction between Candida albicans and cancers. Front Microbiol. 2022, 13, 988734. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, W.; Wu, W.; Wu, S.; Young, A.; Yan, Z. Is Candida albicans a contributor to cancer? A critical review based on the current evidence. Microbiol Res. 2023, 272, 127370. [Google Scholar] [CrossRef]
- Seo, W.; Gao, Y.; He, Y.; Sun, J.; Xu, H.; Feng, D.; Park, S.H.; Cho, Y.E.; Guillot, A.; Ren, T.; et al. ALDH2 deficiency promotes alcohol-associated liver cancer by activating oncogenic pathways via oxidized DNA-enriched extracellular vesicles. J Hepatol. 2019, 71, 1000–1011. [Google Scholar] [CrossRef]
- Seitz, H.K.; Stickel, F. Acetaldehyde as an underestimated risk factor for cancer development: role of genetics in ethanol metabolism. Genes Nutr. 2010, 5, 121–8. [Google Scholar] [CrossRef]
- Dohlman, A.B.; Klug, J.; Mesko, M.; Gao, I.H.; Lipkin, S.M.; Shen, X.; Iliev, I.D. A pan-cancer mycobiome analysis reveals fungal involvement in gastrointestinal and lung tumors. Cell 2022, 185, 3807-3822.e12. [Google Scholar] [CrossRef]
- Teoh, F.; Pavelka, N. How Chemotherapy increases the risk of systemic Candidiasis in cancer patients: current paradigm and future directions. Pathogens. 2016, 5, 6. [Google Scholar] [CrossRef]
- Li, X.; Snyder, M.P. Yeast longevity promoted by reversing aging-associated decline in heavy isotope content. NPJ Aging Mech Dis. 2016, 2, 16004. [Google Scholar] [CrossRef]
- Bhat, T.A.; Kumar, S.; Chaudhary, A.K.; Yadav, N.; Chandra, D. Restoration of mitochondria function as a target for cancer therapy. Drug Discov Today. 2015, 20, 635–43. [Google Scholar] [CrossRef]
- Park, J.H.; Pyun, W.Y.; Park, H.W. Cancer metabolism: Phenotype, signaling and therapeutic targets. Cells. 2020, 9, 2308. [Google Scholar] [CrossRef]
- Stransky, L.; Cotter, K.; Forgac, M. The function of V-ATPases in cancer. Physiol Rev. 2016, 96, 1071–91. [Google Scholar] [CrossRef]
- Whitton, B.; Okamoto, H.; Packham, G.; Crabb, S.J. Vacuolar ATPase as a potential therapeutic target and mediator of treatment resistance in cancer. Cancer Med. 2018, 7, 3800–3811. [Google Scholar] [CrossRef]
- Perzov, N.; Padler-Karavani, V.; Nelson, H.; Nelson, N. Features of V-ATPases that distinguish them from F-ATPases. FEBS Lett. 2001, 504, 223–8. [Google Scholar] [CrossRef]
- Kotyk, A.; Dvorkov, M.; Koryta, J. Deuterons cannot replace protons in active transport processes in yeast. FEBS Lett. 1990, 264, 203–5. [Google Scholar] [CrossRef]
- Sennoune, S.R.; Bakunts, K.; Martínez, G.M.; Chua-Tuan, J.L.; Kebir, Y.; Attaya, M.N.; Martínez-Zaguiln, R. Vacuolar H+-ATPase in human breast cancer cells with distinct metastatic potential: distribution and functional activity. Am J Physiol Cell Physiol. 2004, 286, C1443–52. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol Cancer 2019, 18, 10. [Google Scholar] [CrossRef]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: from T cell basic science to clinical practice. Nat Rev Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef]
- Kovács, B.Z.; Puskás, L.G.; Nagy, L.I.; Papp, A.; Gyöngyi, Z.; Fórizs, I.; Czuppon, G.; Somlyai, I.; Somlyai, G. Blocking the Increase of Intracellular Deuterium Concentration Prevents the Expression of Cancer-Related Genes, Tumor Development, and Tumor Recurrence in Cancer Patients. Cancer Control. 2022, 29, 10732748211068963. [Google Scholar] [CrossRef]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem Soc Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef]
- Caslin, H.L.; Abebayehu, D.; Pinette, J.A.; Ryan, J.J. Lactate is a metabolic mediator that shapes immune cell fate and function. Front Physiol. 2021, 12, 688485. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Zhang, B.; Lin, X.; Fu, X.; An, Y.; Zou, Y.; Wang, J.X.; Wang, Z.; Yu, T. Lactate metabolism in human health and disease. Signal Transduct Target Ther. 2022, 7, 305. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef]
- Brooks, G.A.; Dubouchaud, H.; Brown, M.; Sicurello, J.P.; Butz, C.E. Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc Natl Acad Sci U S A. 1999, 96, 1129–34. [Google Scholar] [CrossRef]
- Grimshaw, C.E.; Cleland, W.W. Deuterium isotope effects on lactate dehydrogenase using L-2-hydroxysuccinamate and effect of an inhibitor in the variable substrate on observed isotope effects. Biochemistry. 1980, 19, 3153–7. [Google Scholar] [CrossRef]
- Pérez-Tomás, R.; Pérez-Guillén, I. Lactate in the tumor microenvironment: An essential molecule in cancer progression and treatment. Cancers (Basel). 2020, 12, 3244. [Google Scholar] [CrossRef]
- Chen, F.; Kang, R.; Liu, J.; Tang, D. The V-ATPases in cancer and cell death. Cancer Gene Ther. 2022, 29, 1529–1541. [Google Scholar] [CrossRef]
- Apostolova, P.; Pearce, E.L. Lactic acid and lactate: revisiting the physiological roles in the tumor microenvironment. Trends Immunol. 2022, 43, 969–977. [Google Scholar] [CrossRef]
- Barbieri, L.; Velia, P.; Gameiro, P.A.; Cunha, P.P.; Foskolou, I.P.; Rullman, E.; Bargiela, D.; Johnson, R.S.; Rundqvist, H. Lactate exposure shapes the metabolic and transcriptomic profile of CD8+ T cells. Front Immunol. 2023, 14, 1101433. [Google Scholar] [CrossRef]
- Feng, Q.; Liu, Z.; Yu, X.; Huang, T.; Chen, J.; Wang, J.; Wilhelm, J.; Li, S.; Song, J.; Li, W.; et al. Lactate increases stemness of CD8 + T cells to augment anti-tumor immunity. Nat Commun. 2022, 13, 4981. [Google Scholar] [CrossRef]
- Li, L.; Tian, Y. The role of metabolic reprogramming of tumor-associated macrophages in shaping the immunosuppressive tumor microenvironment. Biomed Pharmacother. 2023, 161, 114504. [Google Scholar] [CrossRef]
- Noe, J.T.; Rendon, B.E.; Geller, A.E.; Conroy, L.R.; Morrissey, S.M.; Young, L.E.A.; Bruntz, R.C.; Kim, E.J.; Wise-Mitchell, A.; Barbosa de Souza Rizzo, M.; et al. Lactate supports a metabolic-epigenetic link in macrophage polarization. Sci Adv. 2021, 7, eabi8602. [Google Scholar] [CrossRef] [PubMed]
- Nechipurenko, Y.D.; Semyonov, D.A.; Lavrinenko, I.A.; Lagutkin, D.A.; Generalov, E.A.; Zaitceva, A.Y.; Matveeva, O.V.; Yegorov, Y.E. The Role of Acidosis in the Pathogenesis of Severe Forms of COVID-19. Biology (Basel). 2021, 10, 852. [Google Scholar] [CrossRef] [PubMed]
- Kyriakopoulos, A.M.; Seneff, S. Pathology deterioration in a pure β-zero thalassemia heterozygote after mRNA COVID-19 vaccination: A case report and literature review. IJVTPR 2024, 3, 1316–1344. [Google Scholar] [CrossRef]
- Hayes, C.; Donohoe, C.L.; Davern, M.; Donlon, N.E. The oncogenic and clinical implications of lactate induced immunosuppression in the tumour microenvironment. Cancer Lett. 2021, 500, 75–86. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature. 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Chen, Z.; Luo, J.; Guo, W.; Sun, L.; Lin, L. Targeting M2-like tumor-associated macrophages is a potential therapeutic approach to overcome antitumor drug resistance. NPJ Precis Oncol. 2024, 8, 31. [Google Scholar] [CrossRef]
- El-Kenawi, A.; Gatenbee, C.; Robertson-Tessi, M.; Bravo, R.; Dhillon, J.; Balagurunathan, Y.; Berglund, A.; Vishvakarma, N.; Ibrahim-Hashim, A.; Choi, J.; et al. Acidity promotes tumour progression by altering macrophage phenotype in prostate cancer. Br J Cancer. 2019, 121, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.; Qian, B.Z.; Rowan, C.; Muthana, M.; Keklikoglou, I.; Olson, O.C.; Tazzyman, S.; Danson, S.; Addison, C.; Clemons, M.; et al. Macrophages Stimulate Tumor Relapse after Chemotherapy. Cancer Res. 2015, 75, 3479–91. [Google Scholar] [CrossRef]
- Khan, F.; Lin, Y.; Ali, H.; Pang, L.; Dunterman, M.; Hsu, W.-H.; Frenis, K.; Rowe, R.G.; Wainwright, D.A.; McCortney, K.; et al. Lactate dehydrogenase A regulates tumor-macrophage symbiosis to promote glioblastoma progression. Nat Commun. 2024, 15, 1987. [Google Scholar] [CrossRef]
- Geistlinger, K.; Schmidt, J.D.R.; Beitz, E. Human monocarboxylate transporters accept and relay protons via the bound substrate for selectivity and activity at physiological pH. PNAS Nexus. 2023, 2, pgad007. [Google Scholar] [CrossRef]
- Wang, Z.; Chang, E.P.; Schramm, V.L. Triple Isotope Effects Support Concerted Hydride and Proton Transfer and Promoting Vibrations in Human Heart Lactate Dehydrogenase. J Am Chem Soc. 2016, 16, 138–15004. [Google Scholar] [CrossRef] [PubMed]
- Meiser, J.; Tumanov, S.; Maddocks, O.; Labuschagne, C.F.; Athineos, D.; Van Den Broek, N.; Mackay, G.M.; Gottlieb, E.; Blyth, K.; Vousden, K.; et al. Serine one-carbon catabolism with formate overflow. Sci Adv. 2016, 2, e1601273. [Google Scholar] [CrossRef] [PubMed]
- Meiser, J. , Schuster, A., Pietzke, M. et al. Increased formate overflow is a hallmark of oxidative cancer. Nat Commun. 2018, 9, 1368. [Google Scholar] [CrossRef]
- Delbrouck, C.; Pozdeev, V.I.; Oudin, A.; Grzyb, K.; Neises, L.; Kiweler, N.; Skupin, A.; Letellier, E.; Niclou, S.P.; Meiser, J. FSMP-09. Formate promotes cancer cell invasion and metastasis via calcium signaling. Neurooncol Adv. 2021, 3 (Suppl 1), i18. [Google Scholar] [CrossRef]
- Pietzke. M.; Meiser, J.; Vazquez, A. Formate metabolism in health and disease. Mol Metab. 2020, 33, 23–37. [Google Scholar] [CrossRef]
- Delbrouck, C.; Kiweler, N.; Chen, O.; Pozdeev, V.I.; Haase, L.; Neises, L.; Oudin, A.; Fouquier d’Hroul, A.; Shen, R.; Schlicker, L.; et al. Formate promotes invasion and metastasis in reliance on lipid metabolism. Cell Rep. 2023, 42, 113034. [Google Scholar] [CrossRef]
- Ye, J.; Fan, J.; Venneti, S.; Wan, Y.W.; Pawel, B.R.; Zhang, J.; Finley, L.W.; Lu, C.; Lindsten, T.; Cross, J.R.; et al. Serine catabolism regulates mitochondrial redox control during hypoxia. Cancer Discov. 2014, 4, 1406–17. [Google Scholar] [CrossRef]
- Guerra, B.; Recio, C.; Aranda-Tavo, H.; Guerra-Rodríguez, M.; Garca-Castellano, J.M.; Fernndez-Pérez, L. The mevalonate pathway, a metabolic target in cancer therapy. Front Oncol. 2021, 11, 626971. [Google Scholar] [CrossRef]
- Juarez, D.; Fruman, D.A. Targeting the Mevalonate Pathway in Cancer. Trends Cancer. 2021, 7, 525–540. [Google Scholar] [CrossRef]
- Jongen, V.H.; Hollema, H.; Van Der Zee, A.G.; Heineman, M.J. Aromatase in the context of breast and endometrial cancer. A review. Minerva Endocrinol. 2006, 31, 47–60. [Google Scholar]
- Fowler, K.A.; Gill, K.; Kirma, N.; Dillehay, D.L.; Tekmal, R.R. Overexpression of aromatase leads to development of testicular leydig cell tumors : an in vivo model for hormone-mediated testicular cancer. Am J Pathol. 2000, 156, 347–53. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.C.; Falcone, M.; Huerta Uribe, A.; Zhang, T.; Athineos, D.; Pietzke, M.; Vazquez, A.; Blyth, K.; Maddocks, O.D.K. Immune-regulated IDO1-dependent tryptophan metabolism is source of one-carbon units for pancreatic cancer and stellate cells. Mol Cell. 2021, 81, 2290-2302.e7. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, X.; Wang, L.; Ma, X.; Gong, Z.; Zhang, S.; Li, Y. Targeting the IDO1 pathway in cancer: from bench to bedside. J Hematol Oncol. 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Klockow, J.L.; Zhang, M.; Lafortune, F.; Chang, E.; Jin, L.; Wu, Y.; Daldrup-Link, H.E. Glioblastoma multiforme (GBM): An overview of current therapies and mechanisms of resistance. Pharmacol Res. 2021, 171, 105780. [Google Scholar] [CrossRef] [PubMed]
- Somlyai, G.; Kovács, B.Z.; Papp, A.; Somlyai, I. A preliminary study indicating improvement in the median survival time of glioblastoma multiforme patients by the application of deuterium depletion in combination with conventional therapy. Biomedicines 2023, 11, 1989. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, W. Pancreatic cancer: A review of risk factors, diagnosis, and treatment. Technol Cancer Res Treat. 2020, 19, 1533033820962117. [Google Scholar] [CrossRef]
- Kovács, A.; Guller, I.; Krempels, K.; Somlyai, I.; Jnosi, I.; Gyöngyi, Z.; Szab, I.; Ember, I.; Somlyai, G. Deuterium depletion may delay the progression of prostate cancer. J Cancer Therapy 2011, 2, 548–556. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, H. Deuterium-depleted water in cancer therapy: A systematic review of clinical and experimental trials. Nutrients 2024, 16, 1397. [Google Scholar] [CrossRef]
- Dorgan, L.J.; Schuster, S.M. The effect of nitration and D2O on the kinetics of beef heart mitochondrial adenosine triphosphatase. J Biol Chem. 1981, 256, 3910–6. [Google Scholar] [CrossRef]
- Mulcahy Levy, J.M.; Thorburn, A. Autophagy in cancer: moving from understanding mechanism to improving therapy responses in patients. Cell Death Differ 2020, 27, 843857. [Google Scholar] [CrossRef]
- Smirnov, A.Y.; Sulaberidze, G.A. Production of water with reduced content of deuterium for water supply system with desalination installation. J. Phys. Conf. Ser. 2018, 1099, 012035. [Google Scholar] [CrossRef]
- Lajos, R.; Braicu, C.; Jurj, A.; Chira, S.; Cojocneanu-Petric, R.; Pileczki, V.; Berindan-Neagoe, I. A miRNAs profile evolution of triple negative breast cancer cells in the presence of a possible adjuvant therapy and senescence inducer. J BUON. 2018, 23, 692–705. [Google Scholar] [PubMed]
- Chira, S.; Raduly, L.; Braicu, C.; Jurj, A.; Cojocneanu-Petric, R.; Pop, L.; Pileczki, V.; Ionescu, C.; Berindan-Neagoe, I. Premature senescence activation in DLD-1 colorectal cancer cells through adjuvant therapy to induce a miRNA profile modulating cellular death. Exp Ther Med. 2018, 16, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Yavari, K.; Kooshesh, L. Deuterium depleted water inhibits the proliferation of human MCF7 breast cancer cell lines by inducing cell cycle arrest. Nutr Cancer 2019, 71, 1019–1029. [Google Scholar] [CrossRef]
- Haseli, R.; Honarvar, M.; Yavari, K.; Ghavami, M. Synergistic anticancer effects of crocin combined with deuterium-depleted water on HT-29 cells. Anticancer Drugs. 2023, 34, 1162–1170. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, B.; He, Z.; Fu, H.; Dai, Z.; Huang, G.; Li, B.; Qin, D.; Zhang, X.; Tian, L.; Fang, W.; Yang, H. Deuterium-depleted water (DDW) inhibits the proliferation and migration of nasopharyngeal carcinoma cells in vitro. Biomed Pharmacother. 2013, 67, 489–96. [Google Scholar] [CrossRef]
- Zhang, X.; Gaetani, M.; Chernobrovkin, A.; Zubarev, R.A. Anticancer effect of deuterium depleted water -- redox disbalance leads to oxidative stress. Mol Cell Proteomics 2019, 18, 2373–2387. [Google Scholar] [CrossRef]
- Zhang, X.; Dai, M.; Li, S.; Li, M.; Cheng, B.; Ma, T.; Zhou, Z. The emerging potential role of p62 in cancer treatment by regulating metabolism. Trends Endocrinol Metab. 2023, 34, 474–488. [Google Scholar] [CrossRef]
- Islam, M.A.; Sooro, M.A.; Zhang, P. Autophagic regulation of p62 is critical for cancer therapy. Int J Mol Sci. 2018, 8, 19–1405. [Google Scholar] [CrossRef]
- Carroll, B.; Otten, E.G.; Manni, D.; Stefanatos, R.; Menzies, F.M.; Smith, G.R.; Jurk, D.; Kenneth, N.; Wilkinson, S.; Passos, J.F.; et al. Oxidation of SQSTM1/p62 mediates the link between redox state and protein homeostasis. Nat Commun. 2018, 9, 256. [Google Scholar] [CrossRef]
- Yi, H.; Talmon, G.; Wang, J. Glutamate in cancers: from metabolism to signaling. J Biomed Res. 2019, 34, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Koochekpour, S.; Majumdar, S.; Azabdaftari, G.; Attwood, K.; Scioneaux, R.; Subramani, D.; Manhardt, C.; Lorusso, G.D.; Willard, S.S.; Thompson, H.; et al. Serum glutamate levels correlate with Gleason score and glutamate blockade decreases proliferation, migration, and invasion and induces apoptosis in prostate cancer cells. Clin Cancer Res. 2012, 18, 5888–901. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.C. Design and synthesis of inhibitors of folate-dependent enzymes as antitumor agents. Adv Exp Med Biol. 1993, 338, 387–408. [Google Scholar] [CrossRef]
- Tricarico, P.M.; Crovella, S.; Celsi, F. Mevalonate pathway blockade, mitochondrial dysfunction and autophagy: A possible link. Int J Mol Sci. 2015, 16, 16067–84. [Google Scholar] [CrossRef]
- De Giorgi, M.; Jarrett, K.E.; Burton, J.C.; Doerfler, A.M.; Hurley, A.; Li, A.; Hsu, R.H.; Furgurson, M.; Patel, K.R.; Han, J.; et al. Depletion of essential isoprenoids and ER stress induction following acute liver-specific deletion of HMG-CoA reductase. J Lipid Res. 2020, 61, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy Levy, J.M.; Thorburn, A. Autophagy in cancer: moving from understanding mechanism to improving therapy responses in patients. Cell Death Differ. 2020, 27, 843–857. [Google Scholar] [CrossRef]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int J Mol Sci. 2018, 19, 3466. [Google Scholar] [CrossRef]
- Rao, S.; Tortola, L.; Perlot, T.; Wirnsberger, G.; Novatchkova, M.; Nitsch, R.; Sykacek, P.; Frank, L.; Schramek, D.; Komnenovic, V.; et al. A dual role for autophagy in a murine model of lung cancer. Nat Commun. 2014, 5, 3056. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ 2022, 29, 946960. [Google Scholar] [CrossRef]
- Mathiassen, S.G.; De Zio, D.; Cecconi, F. Autophagy and the cell cycle: A complex landscape. Front Oncol. 2017, 7, 51. [Google Scholar] [CrossRef]
- Hernández Borrero, L.J.; El-Deiry, W.S. Tumor suppressor p53: Biology, signaling pathways, and therapeutic targeting. Biochim Biophys Acta Rev Cancer 2021, 1876, 188556. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Nakagawara, A. Role of p53 in cell death and human cancers. Cancers (Basel) 2011, 3, 994–1013. [Google Scholar] [CrossRef] [PubMed]
- Gyöngyi, Z.; Somlyai, G. Deuterium depletion can decrease the expression of C-myc Ha-ras and p53 gene in carcinogen-treated mice. In Vivo 2000, 14, 437–9. [Google Scholar]
- Zhang, X.; Gaetani, M.; Chernobrovkin, A.; Zubarev, R.A. Anticancer effect of deuterium depleted water - redox disbalance leads to oxidative stress. Mol Cell Proteomics 2019, 18, 2373–2387. [Google Scholar] [CrossRef]
- Kovács, B.Z.; Pusks, L.G.; Nagy, L.I.; Papp, A.; Gyöngyi, Z.; Frizs, I.; Czuppon, G.; Somlyai, I.; Somlyai, G. Blocking the increase of intracellular deuterium concentration prevents the expression of cancer-related genes, tumor development, and tumor recurrence in cancer patients. Cancer Control 2022, 29, 10732748211068963. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Jat, P. Mechanisms of cellular senescence: Cell cycle arrest and senescence associated secretory phenotype. Front Cell Dev Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Basov, A.; Drobotenko, M.; Svidlov, A.; Gerasimenko, E.; Malyshko, V.; Elkina, A.; Baryshev, M.; Dzhimak, S. Inequality in the Frequency of the Open States Occurrence Depends on Single 2H/1H Replacement in DNA. Molecules 2020, 25, 3753. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, C.; Zhu, W.; Zheng, X.; Huang, Y.; Lu, Z. A facile strategy for the construction of purely organic optical sensors capable of distinguishing D2O from H2O. Angew Chem Int Ed Engl. 2019, 58, 6280–6284. [Google Scholar] [CrossRef]
- De Milito, A.; Canese, R.; Marino, M.L.; Borghi, M.; Iero, M.; Villa, A.; Venturi, G.; Lozuponem, F.; Iessi, E.; Logozzi, M.; et al. pH-dependent antitumor activity of proton pump inhibitors against human melanoma is mediated by inhibition of tumor acidity. Int J Cancer. 2010, 127, 207–19. [Google Scholar] [CrossRef]
- Das, A.; Sinha, S.; Acharya, B.R.; Paul, P.; Bhattacharyya, B.; Chakrabarti, G. Deuterium oxide stabilizes conformation of tubulin: A biophysical and biochemical study. BMB Rep. 2008, 41, 62–7. [Google Scholar] [CrossRef]
- Lopes, D.; Maiato, H. The tubulin code in mitosis and cancer. Cells 2020, 9, 2356. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth, L.; Machado, P.; Zinöcker, M.; Baker, P.; Lawrence, M. Ultra-Processed Foods and Health Outcomes: A Narrative Review. Nutrients. 2020, 12, 1955. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic of the electron transport chain (ETC) in the inner membrane of the mitochondrial intermembrane space, showing the release of damaging ROS at Complex I and III that comes about due to stalling of the ATPase pumps by deuterium. The various reactions that take place at each stage, aimed at filling the intermembrane space with protons, are also specified. The protons ultimately provide the motive force that drives ATP production.
Figure 1.
Schematic of the electron transport chain (ETC) in the inner membrane of the mitochondrial intermembrane space, showing the release of damaging ROS at Complex I and III that comes about due to stalling of the ATPase pumps by deuterium. The various reactions that take place at each stage, aimed at filling the intermembrane space with protons, are also specified. The protons ultimately provide the motive force that drives ATP production.
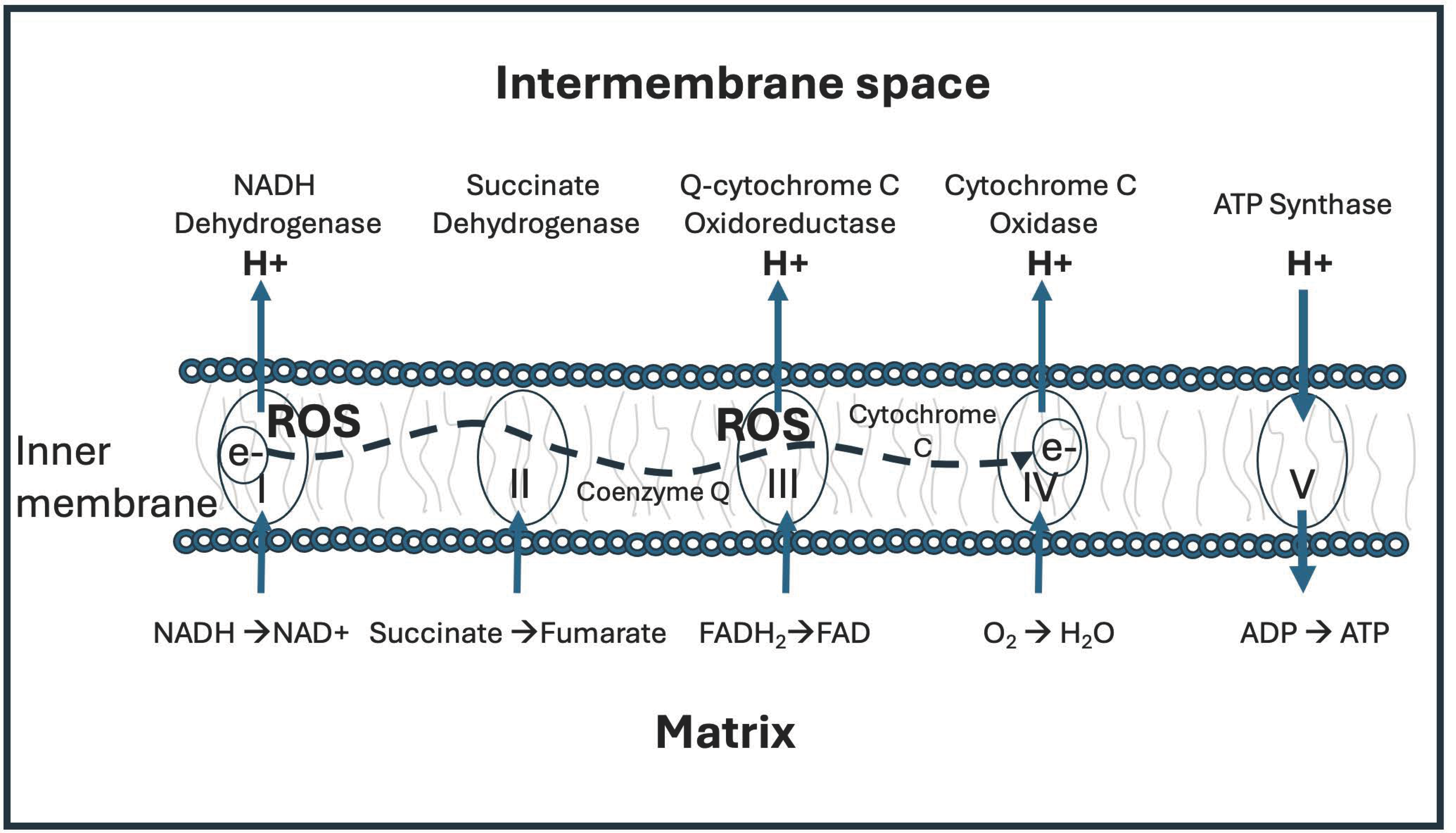
Figure 2.
The seemingly futile carbon cycle in the gut may serve an essential function, which is to capture and preserve deupleted protons by reducing three molecules of NAD+ to NADH. It begins with the reduction of carbon dioxide to methane gas using severely deupleted hydrogen gas as a reducing agent. A series of steps convert methane to methanol to formaldehyde to formate to carbonate and finally back to carbon dioxide, completing the cycle. The NADH, via another circuitous pathway, discussed in the text, eventually delivers a deupleted proton to the mitochondrial intermembrane space of the host cells. Some of the formaldehyde, which is highly reactive, is siphoned off to eventually deliver deupleted methyl groups in the various methylation pathways.
Figure 2.
The seemingly futile carbon cycle in the gut may serve an essential function, which is to capture and preserve deupleted protons by reducing three molecules of NAD+ to NADH. It begins with the reduction of carbon dioxide to methane gas using severely deupleted hydrogen gas as a reducing agent. A series of steps convert methane to methanol to formaldehyde to formate to carbonate and finally back to carbon dioxide, completing the cycle. The NADH, via another circuitous pathway, discussed in the text, eventually delivers a deupleted proton to the mitochondrial intermembrane space of the host cells. Some of the formaldehyde, which is highly reactive, is siphoned off to eventually deliver deupleted methyl groups in the various methylation pathways.
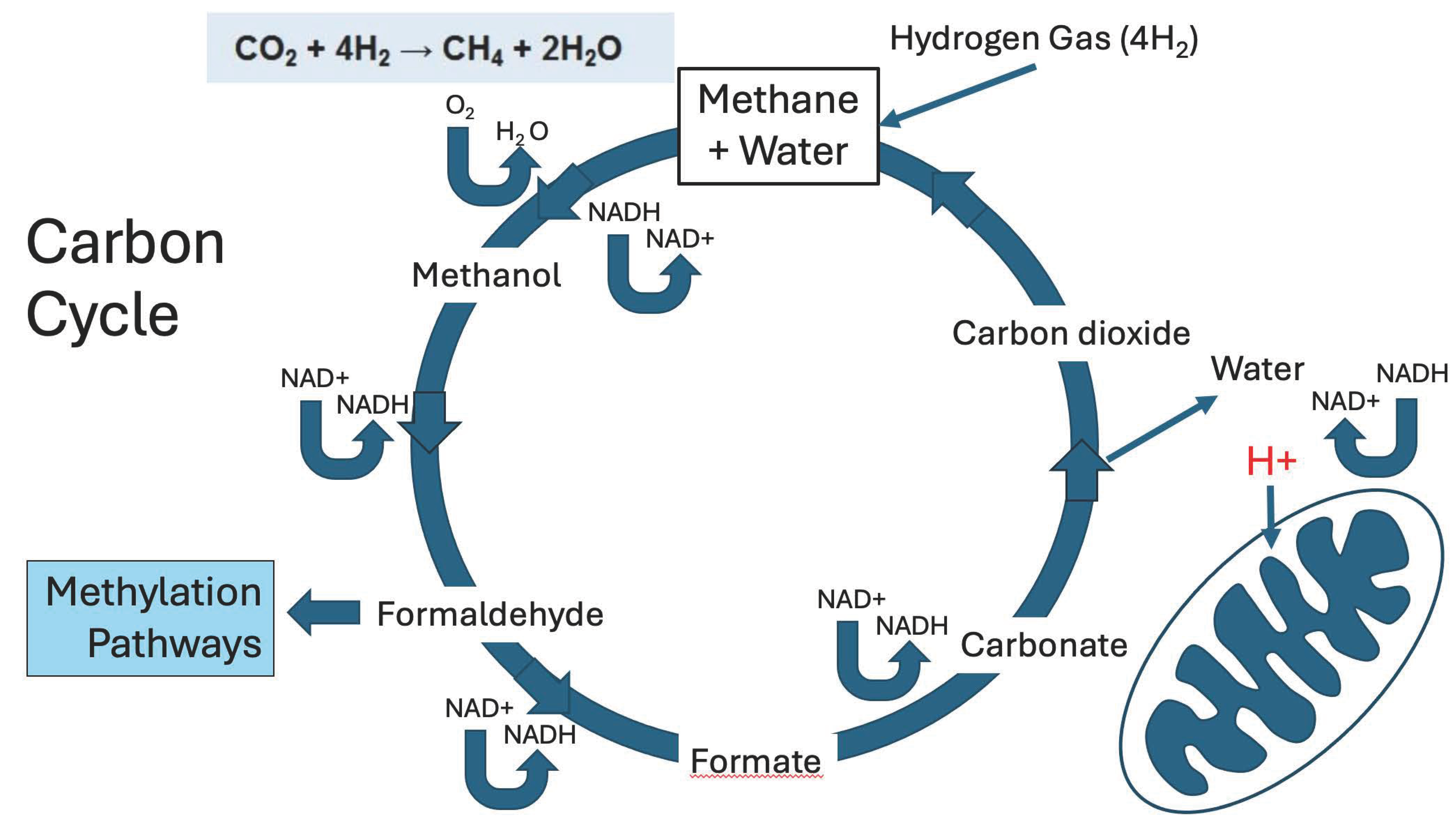
Figure 3.
A schematic presentation of various pathways whereby the deupleted protons in microbially produced methanol are ultimately delivered to α-ketoglutarate to produce succinate, an important organic molecule in the mitochondria that delivers protons to the inner membrane of the intermembrane space via the mitochondrial enzyme, succinate dehydrogenase. It may be the case that methylation pathways play such an important role in metabolism because the methyl groups that they pass around are significantly deupleted. All of the molecules that are highlighted in red carry deupleted protons derived originally from hydrogen gas. Serine, methionine and NADH also become carriers of low-deuterium protons that are widely used in metabolism. CH2= H4F: methylene-tetrahydrofolate; CH3-H4F: methyltetrahydrofolate; SAMe: S-adenosyl-methionine; CH3-cytosine: methyl-cytosine.
Figure 3.
A schematic presentation of various pathways whereby the deupleted protons in microbially produced methanol are ultimately delivered to α-ketoglutarate to produce succinate, an important organic molecule in the mitochondria that delivers protons to the inner membrane of the intermembrane space via the mitochondrial enzyme, succinate dehydrogenase. It may be the case that methylation pathways play such an important role in metabolism because the methyl groups that they pass around are significantly deupleted. All of the molecules that are highlighted in red carry deupleted protons derived originally from hydrogen gas. Serine, methionine and NADH also become carriers of low-deuterium protons that are widely used in metabolism. CH2= H4F: methylene-tetrahydrofolate; CH3-H4F: methyltetrahydrofolate; SAMe: S-adenosyl-methionine; CH3-cytosine: methyl-cytosine.
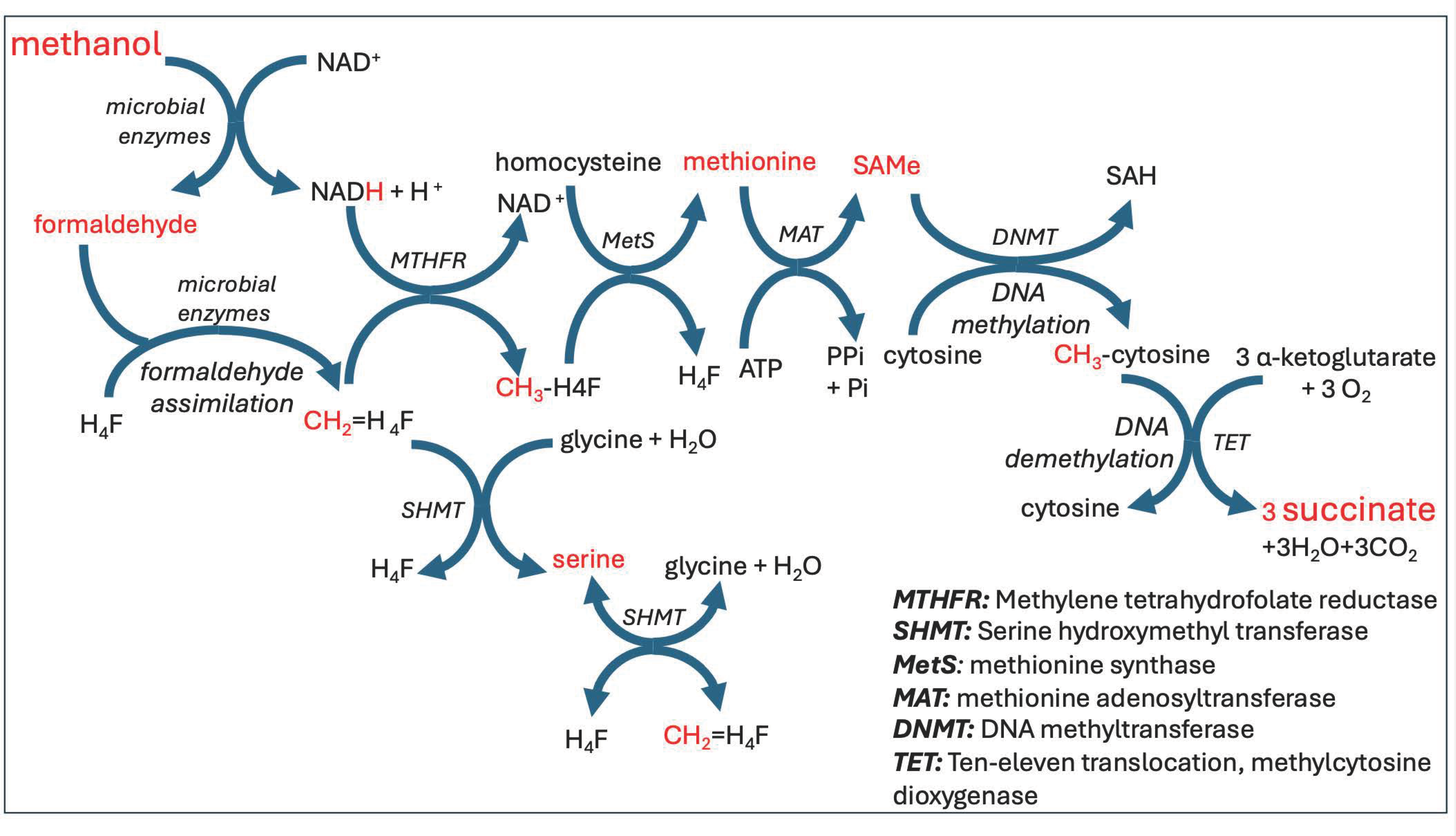
Figure 4.
Schematic of the processes by which gut microbes produce deupleted hydrogen gas and then use it to reduce carbon dioxide to acetate (acetogenic bacteria). Ultimately, further microbial enzymatic action converts acetate to butyrate. Butyrate is the preferred fuel of the colonocytes lining the colon, possibly because it is almost surely low in deuterium. HMOs: Human Milk Oligosaccharides. SCFAs: Short Chain Fatty Acids.
Figure 4.
Schematic of the processes by which gut microbes produce deupleted hydrogen gas and then use it to reduce carbon dioxide to acetate (acetogenic bacteria). Ultimately, further microbial enzymatic action converts acetate to butyrate. Butyrate is the preferred fuel of the colonocytes lining the colon, possibly because it is almost surely low in deuterium. HMOs: Human Milk Oligosaccharides. SCFAs: Short Chain Fatty Acids.
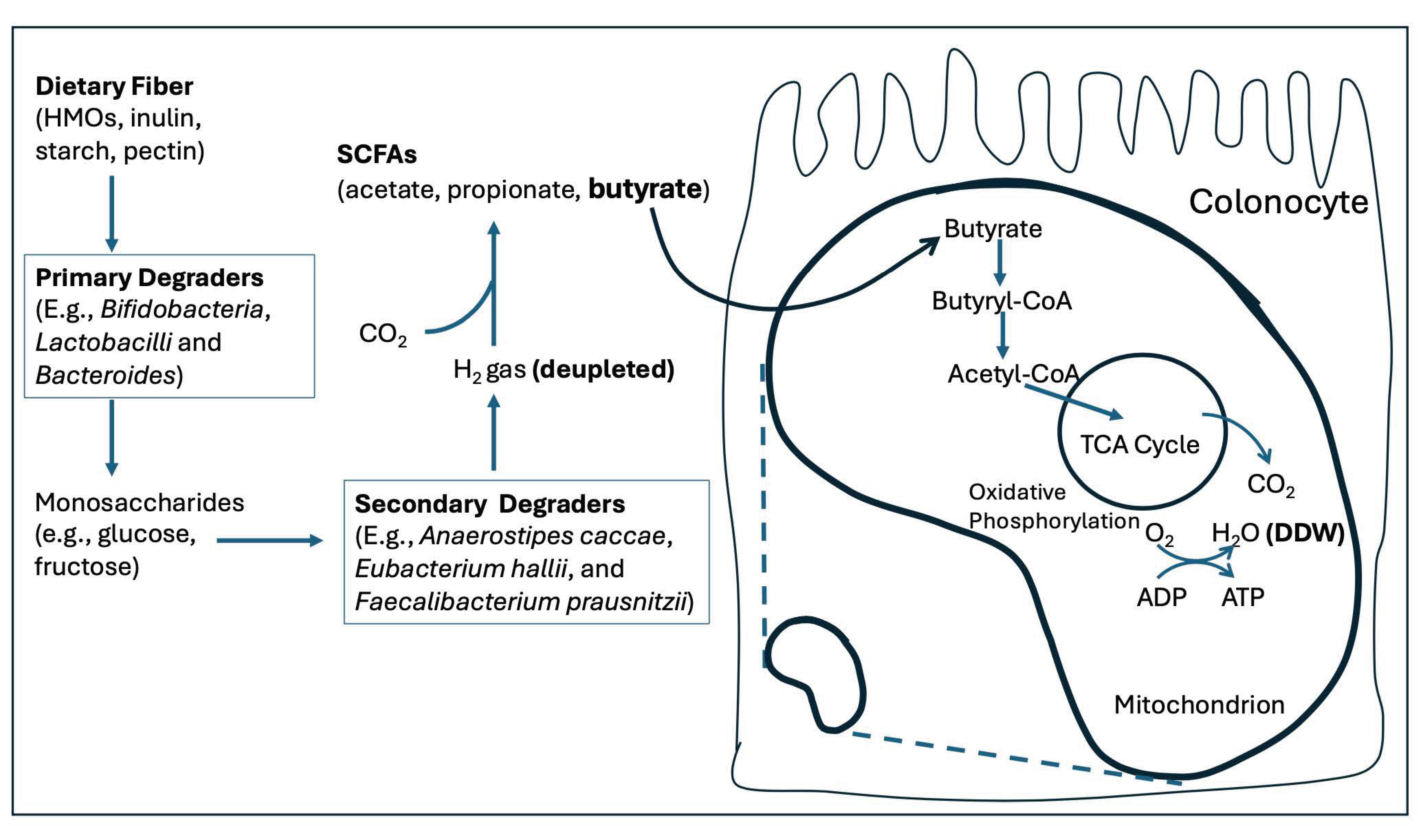
Figure 5.
During fermentation, yeast cells convert glucose to ethanol in the cytoplasm and release the ethanol into the extracellular space. During oxidative phosphorylation, they take up external ethanol from a collective pool and use it to fuel the citric acid cycle.
Figure 5.
During fermentation, yeast cells convert glucose to ethanol in the cytoplasm and release the ethanol into the extracellular space. During oxidative phosphorylation, they take up external ethanol from a collective pool and use it to fuel the citric acid cycle.
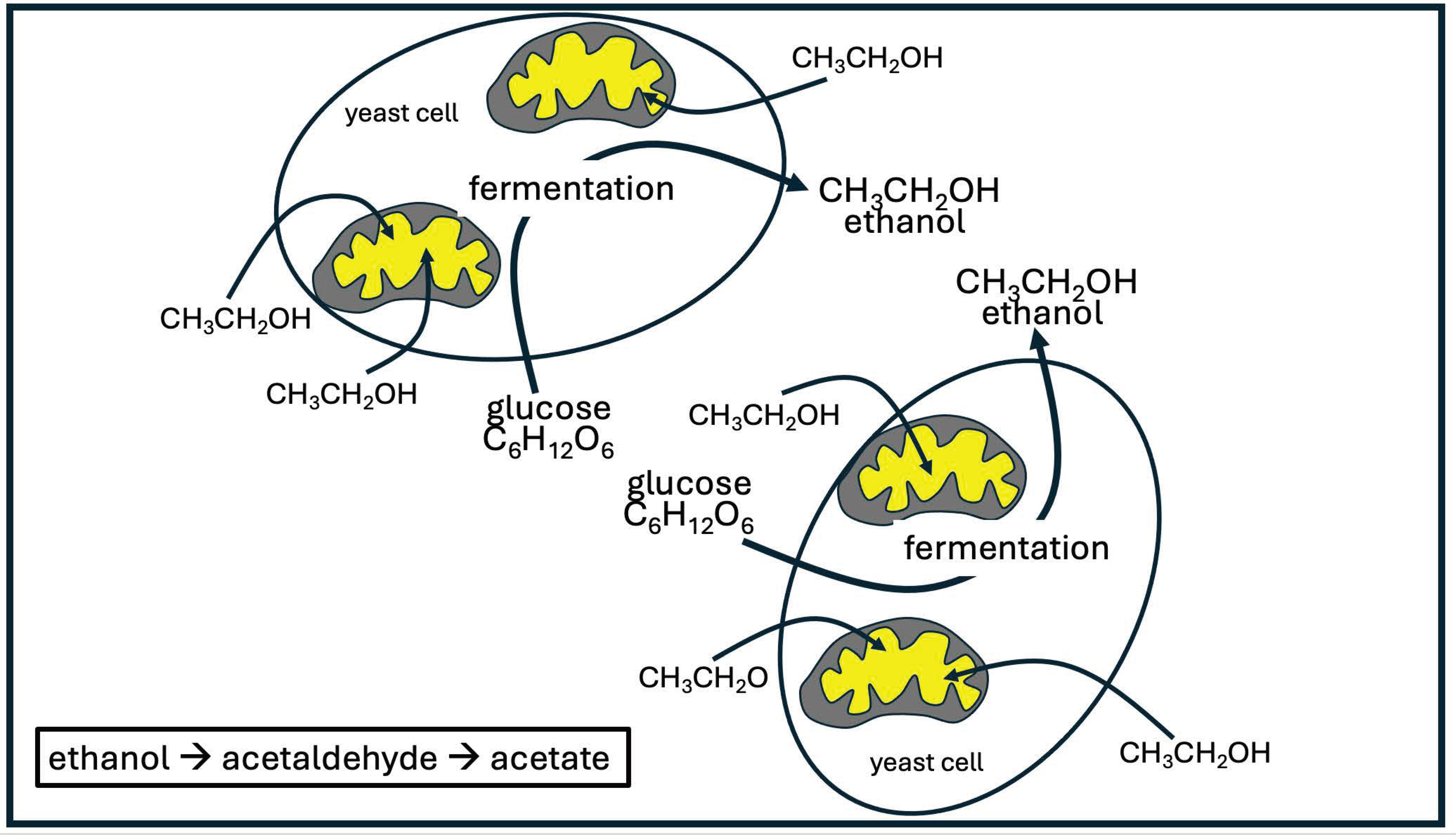
Figure 6.
Illustration of the lactate shuttle. Most of the pyruvate produced in the cytoplasm gets metabolized to lactate in the cytoplasm, and then the lactate is taken up into the mitochondrial matrix, utilizing mitochondrial MCT. Mitochondrial LDH is primarily responsible for converting lactate back to pyruvate, which can then enter the citric acid cycle for metabolism to CO2 and H2O. This assures that the proton attached to NADH in the matrix is derived from matrix metabolic water, i.e., is low in deuterium. mMCT = mitochondrial monocarboxylate transporter. ETC = electron transport chain. mLDH = mitochondrial lactate dehydrogenase. cLDH = cytoplasmic lactate dehydrogenase. Figure adapted from Figure 5 in Brooks et al., 1999 [104].
Figure 6.
Illustration of the lactate shuttle. Most of the pyruvate produced in the cytoplasm gets metabolized to lactate in the cytoplasm, and then the lactate is taken up into the mitochondrial matrix, utilizing mitochondrial MCT. Mitochondrial LDH is primarily responsible for converting lactate back to pyruvate, which can then enter the citric acid cycle for metabolism to CO2 and H2O. This assures that the proton attached to NADH in the matrix is derived from matrix metabolic water, i.e., is low in deuterium. mMCT = mitochondrial monocarboxylate transporter. ETC = electron transport chain. mLDH = mitochondrial lactate dehydrogenase. cLDH = cytoplasmic lactate dehydrogenase. Figure adapted from Figure 5 in Brooks et al., 1999 [104].
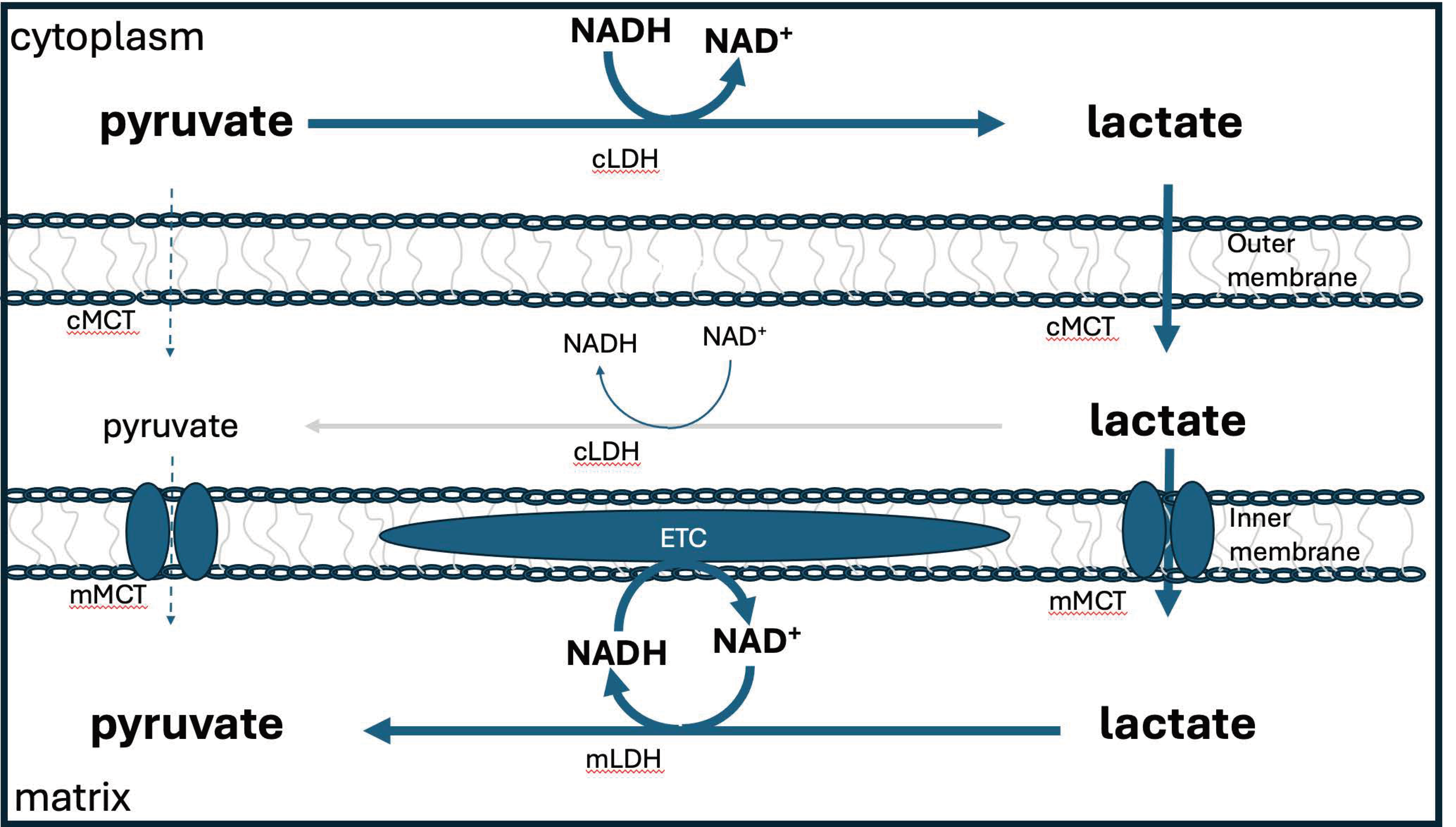

Table 1.
Proteins shown to be downregulated in tumor cells in response to exposure to DDW. ALDH4A1: aldehyde deyhydrogenase 4 A1; FDXR: ferredoxin reductase; H6PD: hexose-6-phosphate dehydrogenase; DHFR: dihydrofolate reductase; GPX4 glutathione peroxidase 4.
Table 1.
Proteins shown to be downregulated in tumor cells in response to exposure to DDW. ALDH4A1: aldehyde deyhydrogenase 4 A1; FDXR: ferredoxin reductase; H6PD: hexose-6-phosphate dehydrogenase; DHFR: dihydrofolate reductase; GPX4 glutathione peroxidase 4.
| Enzyme | Significance |
|---|---|
| ALDH4A1 | Mitochondrial NAD-dependent dehydrogenase; Essential role in pathway converting proline to glutamate |
| FDXR | Mitochondrial flavoprotein that converts NADP+ to NADPH |
| H6PD | A dehydrogenase that converts NADP+ to NADPH |
| DHFR | Regenerates FH$_4$ by reducing NADPH to NADP+ |
| GPX4 | Oxidizes glutathione, important antioxidant enzyme in mitochondria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



