
Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
The reproductive biology of the invasive blue crab Callinectes sapidus was studied in the Thermaikos Gulf (Northwest Aegean Sea, Eastern Mediterranean). In the two-year survey 5,698 (2,897♂/ 2,801♀) crabs were caught with the use of fyke nets. Total sex ratio (♂/♀) reached equality (1.03:1). The female blue crab exhibited a protracted reproductive period. Mature and ovigerous females exhibit short migratory movements from estuarine and inshore waters, where the population mostly congregates (0-3 m), and move to slightly deeper waters (1-3 m) up to 9 m for spawning. A total of 340 ovigerous females were caught. Their number varied both spatially and temporally; they were observed for a 7-month period (April to October) with a clear peak in July - August and at a 3 m depth gradient corresponding to ≈60% of the total number of ovigerous females caught in both years. Size at first sexual maturity (CW50) was estimated at 113.1 mm CW. Average fecundity was ≈790,000 eggs. Experimental trawling showed that inshore waters (<1 m) in the estuaries serve as nursery areas for juveniles. Defining the spatiotemporal and bathymetrical distribution of ovigerous females in any invaded coastal habitat could be considered key information for the implementation of a management policy for the species.
Keywords:
invasive blue crab
; Callinectes sapidus
; Thermaikos Gulf
; Aegean Sea
; reproductive biology
; ovigerous females
; depth gradient
; size at first maturity
; fecundity
; population management policy
1. Introduction
The sexually dimorphic, euryhaline Atlantic blue crab, Callinectes sapidus Rathbun (Decapoda, Portunidae) exhibits a complex life history with distinctive life stage-dependent seasonal, ecological and geographical/spatial distribution [1]. The portunid crab has socio-economical and ecological importance and occupies a wide native range along tropical, subtropical, and temperate ecosystems in the Western Atlantic Ocean and the Gulf of Mexico from Nova Scotia and the Gulf of Maine to Northern Argentina including Bermuda, the West Indies, and the Caribbean Sea [2,3,4,5].
In the last century (1900) Callinectes sapidus was introduced accidentally or intentionally into the Atlantic coast of France [6]. In the mid 1930’s the blue crab C. sapidus appeared in the North Aegean Sea and subsequently in other Mediterranean sites most probably over multiple times through ballast waters [7,8,9,10,11]. In the 1950s’ it was well established in the eastern Basin along the coasts of the North Aegean Sea and of the Levantine Sea including northern Africa (Egypt) [12,13,14]. In the late 1950’s a significant increase in population abundance led to a significant commercial exploitation of the blue crab in the Thermaikos Gulf. However, due to an unknown reason or a combination of reasons the blue crab stock collapsed in the mid 1960’s followed by a serious decline. Afterwards the blue crab was only sporadically recorded in the North Aegean Sea and in the Thermaikos Gulf [15,16] and [references there]). In the late 1960’s a drastic decrease of C. sapidus production was recorded also along the Mediterranean coast of Egypt as a result of the construction of the High Dam and the complete prevention of the Nile floodwaters, which mainly affected the Northern Delta Lakes [17,18]. At the same period its occurrence was documented in the Black Sea [19]. In the 1980’s onwards the blue crab populations thrived along the Anatolian coast (Levantine Sea)(e.g., [20]) and intense fishing activities were deployed resulted gradually to a drastic decrease in landings (1.3 mt in 2023)[21]; Turkish Statistical Institute https://data.tuik.gov.tr/Bulten/Index?p=Fishery-Products-2023-53702/ assessed 21 August 2024).
However, in the early 2000s high abundance of the species was observed mainly along the coast of the north and south eastern Mediterranean (Aegean and Ionian Seas, Mediterranean coast of Egypt) (e.g., [15,16,18,22,23,24]) and in a relatively short time the invasive portunid has succeeded to spread almost throughout the Basin (e.g., [5,25]). Moreover, the blue crab nowadays is expanding along the coast of Morocco in the east Atlantic Ocean but also in the Black Sea (e.g., [5,26,27]). This remarkable expansion is probably caused by the rising average temperatures of the sea surface (SST) in the Mediterranean due to climate-driven change; temperature trends in the Mediterranean Sea during years 2003–2019 showed an SST increase of 0.040 ± 0.001 °C/yr. [28] that possibly influences invasive species by affecting their entry pathways and the establishment of new habitats followed initially by population out breaks in most cases. The blue crab C. sapidus is considered among the most prominent invasive species of all taxa within the Mediterranean Basin and the awareness of it is increased due to its tasty flavor.
Callinectes sapidus is a valuable resource and supports economically important fisheries along the Atlantic and Gulf coasts of USA, where however its populations have fluctuated dramatically during the past 30 years, contributing to significant fishery instability reaching 52,00 mt in 2022 [29,30]. In the Mediterranean the invasive portunid crab forms nowadays the subject of economically important fisheries at a local scale throughout the Basin (e.g., [15,16,22,31,32,33,34,35,36]). However, the blue crab is designated by the EE as an invasive species with a high risk of biodiversity loss and measures should be undertaken towards control of its population outbreaks either through eradication or minimization of its populations size (Regulation 1143/2014). Biological invasions pose a leading threat to biodiversity world-wide [37] and Clavero et al. [38] claimed that the blue crab has the potential to induce radical changes in the structure and the composition of coastal marine communities. Furthermore, the blue crab has significant negative socioeconomic impacts as it destroys fishing nets and predates on fishery ,https://www.rfi.fr/en/france/20220123-getting-snappy-concerns-over-blue-crab-invasion-in-the-mediterranean) and shellfish aquaculture production (https://www.reuters.com/sustainability/italys-clam-industry-under-threat-blue-crab-invasion-2023-08-03/ wreaking havoc on the fishing and shellfish industry and threatening local economies. Therefore, it seems that the management of the blue crab is crucial for biodiversity conservation and coastal industry in the Mediterranean Basin. However, managing invasive species is particularly challenging mainly because marine ecosystems are highly connected across broad spatial scales. Moreover, since for well-established species eradication is unlikely the aim of the management is generally to reduce their populations to levels that exert lower impacts considered as acceptable [39,40,41] Thus,the General Fisheries Commission for the Mediterranean (GFCM) forward the sustainable exploitation of the blue crab aimed at ensuring socioeconomic sustainability (Recommendation GFCM/42/2018/7). Similarly, FAOs proposal is to attain sustainable fisheries of the blue crabs that could act as a tool keeping them at nonexpanding level and minimizing its effect on Mediterranean ecosystems in the perspective to assure that this resource remain profitable for the north African countries. However, the management policy of the blue crab fishery either towards maintenance of sustainable fisheries or eradication/population size control, should require temporal and/or spatial management measures either for protection or a targeted fishing on the spawning stock of the blue crabs’ populations. Nevertheless, the implementation of any of the above management policies requires the knowledge of the life cycle, ecology and reproductive biology of the blue crab and especially of its critical life stage of spawning.
Reproduction in the blue crab is a complex process requiring precise coordination of physiological, behavioral, and ecological processes to ensure reproductive success [42,43]. In an estuarine environment typically involves specific seasonal and spatial migrations for copulation followed by the down-estuary migration of inseminated and ovigerous females towards higher-salinity coastal waters for egg laying and hatching e.g., [44,45,46,47]. The species is thought to be extremely fertile e.g., [48,49], a critical biological feature for its relative success as an invasive species in the Mediterranean Sea. However, diversified environmental conditions, such as temperature and salinity, prevailed in various geographical regions where the species has been established can modify aspects of blue crab mating (and other aspects of reproduction) because they influence the timing of molting, as well as the structure of local populations, including the spatial and temporal distribution of crabs e.g., [16,43,50,51].
Information on the reproduction of the blue crab in the Mediterranean Sea is very limited. Relevant studies on reproductive aspects and estimation of the fecundity of the species have been conducted in the Levantine Sea (Beymelek Lagoon, Iskenderun [= Alexandretta Gulf]), in the central Mediterranean (Sicily) and in the northwestern (Methoni Bay, Thermaikos Gulf) and northeastern Aegean Sea (Monolimni Lagoon, Evros Delta) [15,16,52,53,54]. The present work aims to study the reproductive biology of Callinectes sapidus in the Thermaikos Gulf, an estuarine – open sea system, in the northwestern Aegean Sea, Eastern Mediterranean. Furthermore, aims to contribute to the debate which is in progress in the EU and FAO but also within the Mediterranean countries for the development of a fisheries management strategy for the blue crab populations within the Basin. Biological data on the reproductive potential of a population are critical and stock assessments and management decisions often incorporate aspects of reproduction [48]. In this perspective the survey identifies a key knowledge on the reproductive behavior of the ovigerous female blue crab during spawning phase that it could be useful for the implementation of management measures and actions towards sustainable fisheries or size control (i.e., minimization) of the Atlantic blue crab populations within invaded areas of the Mediterranean Basin, respectively.
2. Materials & Methods
2.1. Study Area
The Thermaikos Gulf is a semi-enclosed, microtidal coastal basin of the mid-latitude east-central Mediterranean Sea. It is located in the NW Aegean Sea, and it covers about 3,300 km2 (Figure 1). It is characterized by a gentle slope and is divided into an inner and outer part; the inner gulf has a shallow bathymetry with its deeper zones around 50 m whereas the outer gradually reaches depths of 200 m towards its continental shelf break with the Aegean Sea and the deep basin of Sporades. The distribution of physical properties and the general circulation of Thermaikos are controlled by four major environmental factors: 1) The air sea interactions and especially the wind forcing, 2) the exchanges with the northern Aegean sea through the southern boundary, 3) the river plume dynamics and 4) the morphological setting of the enclosed basin [55]. The Thermaikos Gulf is a typical deltaic platform characterized by estuaries, numeral inlets, enclosures and small like lagoon complexities formed mainly by major Rivers Aliakmon, Loudias and Axios, discharging along the northwestern coast in the inner gulf (see Figure 1), and Pinios River further south discharging in the outer gulf. These major rivers are the main sources of freshwater and nutrient input and associated fine grained terrigenous material entering the gulf; the seabed is dominated by various soft substrate types, which are locally covered mainly by seagrass meadows of the Posidonia oceanica and Cymodocea nodosa and the seaweed Gracilaria sp. In the unvegetated sedimentary bottom muddy and muddy sands constitute the predominate type surface sediments especially over the inner and the western half of the shelf (central and outer parts) [56,57,58]. Industrial discharges, urban untreated sewage agricultural runoff, shipping, etc, resulted in a heavy eutrophication of the nearshore marine environment with persistent phytoplankton blooms. Especially the western coastal zone is under pressure due to intense agricultural activities and is characterized by a large development of long line mussel farming located near the deltas of the inner gulf [55].
2.2. Sampling Procedure
2.2.1. Monthly and Seasonally Samplings
During the two year survey from December 2020 to November 2022 fyke nets (10 pairs; mesh size 18-20 mm) were deployed monthly in twelve sampling stations at depths 1 m, 3 m, 6 m and 9 m along the northwestern coast of the inner Thermaikos Gulf. Additionally, fyke nets (10 pairs; mesh size 18-20 mm) were deployed seasonally (February, May, July and October) in nine sampling stations at depths 1 m, 3 m, 6 m, 9 m, 15 m and 25 m including a hang mussel culture unit with no mussels cultured in the course of the study at 3.5 m depth, a long line mussel culture unit with mussels cultured during the course of the study at 7-10 m depth and in Loudias River at 800 m distance from the sea at 2.5 m depth (Figure 1)(Table 1).
Sampling took place following local fishery practice with the use of an artisanal fishing boat (length 12 m, 400 hp). Fyke nets were deployed in the morning and collected after about 48 hours. None of the fyke nets contained bait. Bottom temperature (0C), salinity (psu) and dissolved oxygen (DO) (mg/l) were recorded with a HACH autometer in all sampling stations (monthly and seasonally) throughout the period of survey. All the collected Callinectes sapidus specimens were transferred to the laboratory and sexed according to the shape of their abdomen and counted. Females were also separated into immature and mature depending on the shape of their abdomen. For each crab, the carapace width (CW), including the lateral spines, was measured with a digital Vernier caliper to the nearest 0.01 mm. Crabs of CW <60 mm were considered as juveniles [59,60,61].
2.2.2. Experimental Sampling
Experimental samplings with the use of a parallelepiped dredge like gear (length 2.45 m, width 0.75 m) with a net (mesh size 14 mm) that was trawled for ≈5 minutes from the fishing boat were conducted in February 2021 and in March 2022 in Bara, an enclosure lagoon like formation sub-estuary of Aliakmon River. Samplings were carried `out in sampling stations Bar 1 (February 2021) and in Bar 3 (March 2022); Bar3 was a sampling station at a 500 m distance westerly to the sampling station Bar 1. In both stations trawling was conducted at a depth <1 m (Figure 1).
2.2.3. Macroscopical Examination of Ovaries
Every month from December 2020 to November 2022 visual inspection of ovaries was performed in a subsample of sexually mature females, including ovigerous females, according to the five ovarian stages development (I-V) proposed by Hard [62]: Stage I (ovary following the last molt); Stage II (maturation of the ovary); Stage III (mature ovary preceding the first ovulation); Stage IV (Ovary between the first and second ovulation); Stage V (ovary after the second ovulation). For the period May to October 2021 and from February, May to August and November 2022, ovaries of 53 and 21 females respectively, at maturation stages III and IV (ovigerous with fully mature ovaries), were dissected and weighted. In ovigerous females, the stage of egg development was based on the coloration of the egg mass and was classified into 5 stages: stage 1: bright orange, stage 2: pale yellow, stage 3: pale brown, stage 4: dark brown, stage 5 (advanced egg developmental stage): dark black (see [16]).
2.2.4. Ovarian Histological Analysis
Histological analysis of the different ovarian stages of female C. sapidus was conducted. Specimens were transferred to the laboratory and after dissection the ovaries were removed and fixed in fixative solution for 24 hours at 4 0C. The fixed tissues (ovaries) were first rinsed in running water for 24 hours and then embedded in paraffin. For embedding, the tissue pieces were immersed successively, for one hour, in alcohol solutions of increasing concentration (50%, 70%, 95%, 100% x2). They were then immersed for two hours in xylene solution and placed in an oven at 65°C (2 h) in a special container containing liquid paraffin to be impregnated with it throughout their thickness. Finally, they were placed in special metal moulds in liquid paraffin which solidified after a few minutes at room temperature. Using a paraffin microtome (MICROM HM315), the ovaries were cut along their longitudinal axis and all successive sections, 5 μm thick, were collected. Sections were then selected and stained with Harris hematoxylin solution (Scharlab S.L., Sentmenat, Barcelona, Spain) and eosin Y (yellowish) (VWR International, Leuven, Belgium). The ovarian tissues were then examined and observed under a light microscope connected to a digital camera (ProgRes Plus 2.1, JENOPTIC Optical Systems GmbH, Jena, Germany). Histological sections were photographed at an appropriate magnification scale using Progress Capture 2.1 software.
2.2.5. Fecundity
For the examination of fecundity ovigerous females sampled in June, July and August 2021 were used. The collected individuals were transferred to the laboratory for further analyses. The total wet weight of both the right and the left egg mass was measured by removing the pleopods to the nearest 0.001 g. Four replicate subsamples were weighted to the nearest 0.001 g and the number of eggs from each replicate was measured with the use of a Bogorov counting chamber. These data were then used to estimate the total number of eggs of each individual. Photographs of approximately 1,500 eggs were taken with the use of a stereomicroscope and then they were measured using the ImageJ software (± 0.1 μm accuracy). Due to the ovoid shape of the egg the lengths of the longest and shortest axes were measured to the nearest 1μm, and their mean was calculated as egg diameter [63](Sainte-Marie, 1993). The egg mas index (EMI) was calculated as Egg mass index (%) = 100* total egg wet weight / body wet weight, while egg volume was estimated as the volume of a prolate spheroid, using the long and short axes measurements [64](Kemberling & Darnell 2020).
2.2.6. Data Analysis
Pooled monthly data of the two year of survey (December 2020-November 2022) of bottom temperature (0C), salinity (psu) and oxygen (mg/l) at three sampling stations at a 1 m depth gradient (Meth1, Bar1, Loud 1), were used and mean, minimum and maximum values were estimated. The above stations were selected as they were representative of the three main sub-areas (Methoni Bay, Bara, Loudias Bay) where the blue crab is distributed during most of its life cycle both bathymetrically and spatially in the inner gulf.
Total and monthly sex ratio for the period of survey and sex ratio at 3 m depth gradient (♂/♀), including the hang mussel culture sampling station (3.5 m) were assessed and deviations from 1 were estimated applying a chi-square (χ2) test.
Females size-frequency distributions were calculated per 5 mm size (CW) class intervals for all intact specimens (mature, immature, juveniles) caught during the two years of survey. Significant differences on size distributions, mean ranks and medians between a) intact females caught in the first and in the second year, b) intact mature (including ovigerous) and immature females of both years, were tested with two-sample Kolmogorov-Smirnov (z) test, Mann Whitney (U) test, and independent samples median test (T), respectively. Spearman rank correlation (ρ) and Pearson correlation (r) were used to test the relationship between females size (CW) and: a) ovarian weight with ovaries at maturation stages III and IV, b) ovarian weight with ovaries at maturation stage IV, respectively. All females examined were caught during the period May – September of both years.
Pearson correlation (r) was used to test the relationship between a) ovigerous females caught from April to October and the mean temperature from pooled data recorded in all sampling stations in Methoni Bay, Bara, Loudias Bay, off Axios estuary (seasonal) and off Aliakmonas estuary, b) ovigerous females with depth and c) ovigerous females with eggs at developmental stage 5 with depth. Independent sample Kruskal Wallis test (H) was conducted to examine if there was any significant difference between the number of ovigerous females and a) spatial distribution, b) depth gradient. One-way Anova (F) was used to test if significant difference exists between the number of ovigerous females caught only in July and August and depth gradient, followed by LCD pairwise test. For the above cases the number of ovigerous from the sampling station of mussel hang culture (3.5 m depth) were pooled into those of Loudias Bay and at 3 m gradient. Independent sample Kruskal Wallis test (H) followed by pairwise comparisons and one-way Anova (F) were conducted to examine any significant difference in size (CW) of a) ovigerous females and b) ovigerous females with eggs at developmental stage 5, with depth gradient, respectively. All the above statistical analyses were performed using the SPSS software package. The mean size at sexual maturity of females (CW50) caught in both years was estimated with the package ‘Sizemat’ in R (ver. 4.2.0) with Rstudio.
To determine statistical differences of fecundity and egg size among months, a one-way ANOVA (F) was employed, combined with Tukey’s post hoc test. To determine the relationship of fecundity, total egg mass and egg volume with size (CW), linear regressions were constructed. All analyses were conducted using R studio (ver. 4.0.0)
3. Results
3.1. Physicochemical Parameters
3.1.1. Temperature
In the first year (December 2020 – November 2021) the bottom temperature ranged from 9 0C in January 2021 (MΕΤH 1) to 30,1 0C (LOUD 1) in June 2021. The following year (December 2021 – Νovember 2022 the temperature ranged from 3,8 0C in January 2022 (BAR 1) to 28,7 0C in July 2022 (AX 1) (Figure 2).
3.1.2. Salinity
In the first-year salinity ranged from 11,78 psu in February 2021 (BAR 1) to 37,6 psu in March 2021 (LOUD 1). In the second-year salinity ranged from 16 (BAR 1) in December 2021 to a maximum of 36,9 psu in March 2022 (AL 3).
3.1.3. Oxygen
Oxygen fluctuated significantly depending on month and sampling station. In the first and the second period both minimum and maximum oxygen values were recorded in LOUD 1 with 3.52 (July 2021) and 16.16 mg/l (May 2021) and 2.5 (June 2022) and 18.15 mg/l (March 2022), respectively.
3.2. Sex Ratio
In total, 5,698 crabs were collected in the two-year study (December 2020 – November 2022) in the inner Thermaikos Gulf. In the first and in the second year 2,626 and 3,072 crabs were caught, respectively. Out of the 5,698 crabs, 2,897 were males and 2,801 females with 1,326 and 1,571 and 1,300 and 1,501 specimens caught in the first and in the second year, respectively. Total sex ratio (♂/♀) reached equality (1.034) (χ2 = 1.62, p = 0.202). Monthly sex ratio exhibited a similar trend in both years except for December, February, April and August (Figure 3).
In the first year sex ratio did not deviate from equality for 7 months; January (χ2 = 0.14 , p = 0.71), May (χ2= 0.44, p = 0.5), June (χ2 = 2.37, p = 0.12), July (χ2 = 0.28, p = 0.59), September (χ2 = 3.66, p = 0.056), October (χ2 = 2.58, p = 0.11) and November (χ2 = 0.09, p = 0.75). Males predominated in December (χ2 = 18.06, p<0.001), February (χ2 = 4.88, p = 0.02), March (χ2 = 15.52, p<0.01) and April (χ2 = 9.52, p = 0.002) except for August (χ2 = 5.32, p = 0.02) when sex ratio was in favor of females. In the second-year females dominated in February (χ2 = 4, p = 0.04) and May (χ2= 16,41, p<0.001), whereas in August (χ2= 25.75, p<0.001) and in September (χ2 = 4.67, p = 0.03) males outnumbered females. Sex ratio reached equality in April (χ2 = 3.64, p = 0.06), June (χ2 = 1.58, p = 0.21), July (χ2 = 0.003, p = 0.95), October (χ2 = 0.004, p = 0.95) and November (χ2 = 0.04, p = 0.83).
Sex ratio at the 3 m depth gradient, including hang mussel culture sampling station (3.5 m), was in favor of females with a ratio of 1:2.02 (♂/♀) (χ2 = 150.22, p <0.001) (♂ = 449, ♀ = 899). Only 74 immature females were found which represented 8% of all females.
3.3. Females Monthly Distribution
Monthly distribution of all females caught during the survey is presented in Figure 4. For both years the highest number of females was caught during the summer (June – August) with a peak in July (N=944) followed by fall (September, October) and May whereas significant lower number was recorded from late fall to mid spring (November to April) (N <100 crabs) Similarly, by excluding the seasonal sampling stations the highest number of females was caught in July (Ν = 544) followed by June (Ν = 462) and August (Ν = 408) in both years, respectively.
3.4. Females Size (CW) Frequency Distribution
Out of the 2,801 females caught, sexually mature ones including ovigerous females made up ≈80% of the sample (N = 2,242) with 1,020 and 1,222 individuals collected in the first and in the second year, respectively. Immature females (>60 mm CW) caught were 540 (17% of the sample) with a similar number collected in the first (N = 268) and in the second year (N = 273), respectively. Only 19 juveniles (<60 mm )(range CW: 30 – 59,75 mm) were caught ( <1% of the sample) with 12 and 7 individuals collected in the first and the second year, respectively (Figure 5).
Out of the 2,801 females, 2,642 individuals were intact with a mean CW(±sd) = 125.69 (±21.04)mm (range: 30-176.64 mm). In the first and in the second year 1,242 (mean CW =125.52 ±21.90 mm; range: 30 – 172.97 mm) and 1,400 (mean CW = 125.78±20.29 mm; range: 41.67 – 176.64 mm) intact female crabs were measured. A Kolmogorov Smirnov test, Mann Whitney test and Independent-Samples Median Test showed that there was no significant difference in size frequency distributions, mean ranks and medians between females collected in the first and in the second year ; (Kolmogorov Smirnov test: z = 0.979, p=0.293); Mann-Whitney test: U = 857,642, p = 0,548, and Independent median test: T = 1.969, p<0.172, respectively.
Intact mature females (N = 2,111) ranged from 90.85 to 176.64 mm CW with a mean(±sd) = 133.71(±13.19) mm. The total number of intact mature females in the first and the second year was 978 and 1,133 with a mean CW(±sd) = 134.46 (±13.11) mm (range: 90.85-172.97 mm) and CW(±sd) = 133.11(±13.25) mm (range: 91.8 – 176.64 mm), respectively. Intact ovigerous females had a mean CW(±sd) = 130.33 (±14.26 mm; range 99.21-172.97 mm).
Intact immature females (N = 512) had a mean(±sd) = 95.33(±13.08 mm) and ranged from 60,66 to 133 mm CW. The total number of intact immature females in the first and the second year was 252 and 260, with a mean CW(±sd) = 94.73(±12.71) mm (range 60.82 - 133 mm) and a mean CW(±sd) = 95.92(±13.43mm) (range 60.66-131.91 mm), respectively. Size frequency distributions, mean ranks and medians were significantly different between intact mature and immature females; Kolmogorov Smirnov test: z = 17.362, p<0.000); Mann-Whitney test: U = 20,948.500, z = 9.3p<0.000; Independent median test: T = 625.797, p<0.000, respectively.
3.5. Ovarian Stages
3.5.1. Gross Appearance of Ovaries
Visual inspection of ovarian stages was performed in 1,282 sexually mature females. All five ovarian stages (I-V) proposed by Hard [62] were observed. Monthly frequency of ovarian stages in the first year (December 2020 to November 2021) was estimated from 666 sexually mature females and is shown in Figure 6a. Premature ovarian stages were observed throughout the year: stage I was observed in all months with high percentages in December and March and with a peak in April when all ovaries of females examined were at that stage; Stage II appeared all over the year except for April and with the highest percentage observed in January (56%). Females with mature ovaries (stage III) were observed almost all over the year, except for February, March and April. The highest percentages recorded from May to August (46-54%) but also in October (54%) whereas in September stage III was low (20%). Stage IV appeared in July (26%), August (20%) and September (4%). Stage V was observed from June to November with the highest percentage recorded in September (21%). In the second year the ovarian stages were examined in 616 sexually mature females (Figure 6b). Stage I was present in all months except for January and peaked in March (70%). Stage II was observed throughout the year with the highest percentage recorded in January when however, only two crabs were caught. Stage III was present in all months except for January. Lower percentages were observed from February to April and in September (20-25%) whereas the highest percentages were recorded from May to August (35-50%) but also from October to November (34-47%). Stage IV appeared from June to September with the highest percentage recorded in July (33%) and in August (27%). Stage V was present from May to November except for August whereas the highest proportions appeared in May (17%) and in July (14%).
3.5.2. Histological Analysis of Ovarian Stages
Histological analysis of the ovaries of female blue crab assigned five stages (stov1 – stov 5) in the ovarian maturation process: inactive (stov 1); growing (stov 2); development (stov 3); mature (stov 4) and resorption (stov 5) (Figure 7a-e). In the inactive stage the primary oocytes form the previtellogenic oocytes after meiotic division. An increase in cytoplasmic size is observed (Figure7a). In growing stage early gametogenesis occurs, the nucleus remains large with more than one nucleoid, a thin basement membrane appears, and a perinuclear vacuole in the cytoplasm is present (Figure 7b). As oogenesis progresses, at the development stage, there is an increase in the size of the oocytes and the formation of the Zona pellucida. Granulosa cells arranged radially around the Zona pellucida (Figure 7c). During the maturity stage, large oocytes without visible nucleus are present, yolk grains forming glassy plates with no gaps between them and with bright pink color (Figure 7d). In the resorption stage after the spawning period, residual oocytes in various stages of development are observed. Phagocytes participate in the process of absorbing residual gametes (Figure 7e).
3.6. Size (Carapace Width) - Ovarian Weight
In females with ovaries at stages III and IV (see [62]) Spearman rank order correlation demonstrated a significant weak positive correlation between carapace width and ovarian weight (ρ = 0,263, p = 0,025) (N = 73). However, when only females at stage IV (N = 20), i.e., between first and second ovulation, were examined the Pearson correlation test showed that CW and ovarian weight resulted in a highly significant strong positive correlation (r = 0.61, p = 0.003).
3.7. Ovigerous Females
3.7.1. Temperature – Ovigerous Females
A strong but non-significant correlation was found between mean pooled temperatures from April to October of both years and the number of ovigerous females collected in the respective months (Pearson correlation test, r = 0.631, p = 0.129).
3.7.2. Monthly Distribution of Ovigerous Females and Ovigerous Females with Eggs at Developmental Stage 5
During the two-year survey a total of 340 ovigerous females with eggs at all developmental stages (1-5) were caught (N = 156, 2021) (N = 184, 2022). Monthly distribution of ovigerous females in the first and in the second year is presented in Figure 8. In the first year ovigerous were collected from May to September whereas in the second year their presence extended from April to October. The highest number of ovigerous females in the first and in the second year was caught in July with 83 and 135 crabs which corresponded to 53 and 73 % of the total number of ovigerous in each year, respectively. It was followed by August with 61 and 20 crabs, in the first and the second year, respectively, but also by June 2021 with 14 ovigerous. In all other months in both years the number of ovigerous caught was ≤8 crabs. Chronologically, the first ovigerous females were caught at the end of April (N = 3) and the last ones at the end of October 2021 (N = 3). By excluding the seasonal stations, the number of ovigerous caught in monthly stations was 206 with 113 and 93 specimens found during the first and the second year, respectively. Specifically, in the first year the highest number was found in August (N = 61) followed by July (N = 42). Conversely, in the second year the highest number was caught in July (N = 45) followed by August (N = 20). For both years the highest number of ovigerous in monthly stations were collected in July with 87 individuals followed by 81 in August, represented 42 and 39 % of the total number of ovigerous caught in monthly stations, respectively.
A total of 97 ovigerous females with eggs at developmental stage 5 were caught, representing 30% of total ovigerous females. In the first and the second year 46 and 51 ovigerous were caught, respectively. In the first year the largest number was selected in July (N = 33) followed by August (11) whereas only one was caught in May and September, respectively. Similarly, in the second year the largest number was selected in July (N = 37) followed by August (N = 9); only one in June and two in September and October were caught, respectively. By excluding the seasonal stations, the number of ovigerous with eggs at stage 5 caught in monthly stations was 52 with 34 and 18 specimens found during the first and the second year, respectively. Specifically, in the first year the highest number was found in July (N = 23) followed by August (N = 11). In the second year all ovigerous females with eggs at developmental stage 5 were found in July and August with the same number caught (N = 9) in both months.
3.7.3. Spatial Distribution of Ovigerous Females and Ovigerous Females with Eggs at Developmental Stage 5
Spatial distribution of ovigerous females in the inner Thermaikos is presented in Figure 9a,b. In both years ovigerous were found at 18 out of 21 monthly and seasonal sampling stations with the highest number caught in both seasonal station AX1 and monthly station AL2 (N = 64) followed by the seasonal station AX2 (N = 49). In the first year 42 ovigerous were caught in AL2 followed by AX1 (N=35), METH 2 (N = 27) and BAR 2 (N = 14). In all other sampling stations ovigerous were ≤10 individuals. In the second year 45 ovigerous were caught in AX2, followed by AX1 (N = 29 ), AL2 (N = 22) , BAR 2 (N = 19), METH 2 (N = 16) and in the hang mussel culture (N = 11). In all other sampling stations ovigerous at stage 5 were ≤10 specimens. In both years 10 ovigerous, five in each year, were collected in the nursery area of Bar1. Only two ovigerous with eggs in early developmental stage (stages 1 and 2) were found in Loudias River one in each year. Not any crab or ovigerous female was found at the sampling stations AL3 (15 m depth) and AL4 (25 m depth) and at the long line mussel culture station where mussels were cultivated all year round (7-10 m depth). Independent sample Kruskal Wallis test (H) showed that the number of ovigerous females varied significantly among the sampling stations (χ2=28.104, df = 17, p = 0.044).
Ovigerous females with eggs at developmental stage 5 were found inshore in all sampling subareas in the inner Thermaikos i.e., Methoni Bay, Bara, off Aliakmon estuary, Loudias Bay (including the hang culture station) and off Axios estuary. From pooled data from all sampling stations in every sub-area, the highest number (N = 30) was selected both off Aliakmon and Axios estuaries, which represented each the 31% of all ovigerous with eggs at stage 5 caught. It was followed by Methoni Bay (N = 22) and Loudias Bay (N = 14), including the hang mussel culture sampling station, represented 23% and 14% of all ovigerous with eggs at stage 5 caught, respectively. In both years the highest number of ovigerous with eggs at developmental stage 5 was found in sampling station AL 2 (N = 28) which represented ≈30% of all ovigerous at that stage followed by AΧ 2 (N = 18) and METH 2 (N= 14). In the first and the second year the highest number of ovigerous with eggs at developmental stage 5 was caught in AL 2 (N = 11) (24% of all ovigerous with eggs at stage 5) and in AΧ 2 (N = 18) (35% of all ovigerous with eggs at stage 5), respectively. No one ovigerous with eggs at stage 5 was collected in Loudias River and in the sub-estuary area of Bara (sampling stations BAR 1, BAR 2) with the exclusion of one female with eggs in its pleopods, that it was previously hatched and found in BAR 2 probably migrating back to the sub-estuary from spawning site.
3.7.4. Bathymetrical Distribution of Ovigerous Females and Ovigerous Females with Eggs at Developmental Stage 5
Τhe highest number of ovigerous females in the first and the second year was caught at 3 m depth gradient with 90 and 120 crabs, corresponded to 58 and 65% of the total number of ovigerous females, respectively (Figure 10). Strong presence of ovigerous was observed at 1 m depth where 50 and 38 crabs were caught in the first and the second year, respectively. At 6 m depth were found 14 and 20 ovigerous whereas at 9 m depth 3 and 5 crabs were collected in the first and the second year, respectively. In Loudias River only two ovigerous with eggs at developmental stage 1 and 2 were caught, each in every year. A strong negative but non-significant correlation with depth and ovigerous females was found with depth gradient (Pearson correlation test; r = -0.65 p = 0.342).
Independent sample Kruskal Wallis (H) test (χ2=20.711, p = 0,000) showed that there was a significant difference between the number of ovigerous and depth gradient. However, pairwise comparisons showed that the number of ovigerous does not vary at 1, 3 and 6 m depths whereas it was significantly different with that in depth 9 m and in Loudias River. The number of ovigerous caught at 9 m and in Loudias River was not different. However, when only July and August were tested when 88% of all ovigerous were caught during the survey, one-way Anova (F) showed that there was a significant difference (F = 5.209, p = 0.008) between the number of the ovigerous and depth gradient. Μultiple pairwise comparisons (LCD) showed that the number of ovigerous at 3 m depth was significant different to all other depths (p = 0.001-0.027).
In both years the highest number of ovigerous with eggs at developmental stage 5 was caught at 3 m depth (N = 71), including the sampling station at the hang mussel culture (N=9), corresponded to 73% of the total number of ovigerous females found, followed by a significantly lower number at 1 m (N=12), 6 m (Ν = 8) and at 9 m (N = 6). No one ovigerous with eggs at stage 5 was collected at 15 and 25 m depth and at the long line mussel culture sampling station. A moderate negative non-significant correlation between ovigerous at stage 5 with depth gradient was found (Pearson correlation test; r = -0.41 p =0.593).
3.7.5. Size of Ovigerous Females and Ovigerous Females with Eggs at Developmental Stage 5
The mean CW of intact ovigerous females (N = 325) was 130.33(±14.26) mm (range CW: 98.21- 172.97 mm) whereas the mean CW of intact ovigerous females with eggs at developmental stage 5 (N = 94) was lower (CW = 127.27±15.19) mm (range CW: 98.21 -172.97 mm). The smaller and the larger ovigerous females in size (CW) had eggs at developmental stage 5.
3.7.6. Size of Ovigerous Females and Ovigerous Females with Eggs at Developmental Stage 5 with Depth Gradient
The mean CW(±sd) and the number (N) of ovigerous caught at the different depth gradients of 1 m, 3 m, 6 m and 9 m was 133.28(±13.98) mm (N = 80), 128.53 (±14.08) mm (N = 120), 132.75(± 13.76)mm (N = 35) and 138.76 ±17.12 mm (N= 7), respectively. Independent sample Kruskal Wallis (H) test showed that there was a significant difference in CW between ovigerous females according to depth (χ2 = 8.516, df = 3, p = 0.036) Multiple pairwise comparisons showed that mean CW of ovigerous found at 1 m depth was higher than that of ovigerous at depth 3 m (p = 0.024) but non-significant to that of ovigerous at 6 m (p = 0.966) and 9 m (p = 0.413) whereas CW between ovigerous females caught at 3, 6 and 9 m was non-significant (p = 0.09 – 0.488); however, the number of ovigerous found at 6 and 9 m depth was significant lower than at 1 and 3 m depth gradient .
Moreover, the mean CW(±sd) of intact ovigerous females with eggs at developmental stage 5 caught at the different depth gradients of 1 m, 3 m, 6 m and 9 m was 121.35(± 9.44)mm (range: 98.21-136.98 mm), 127.73(±15.93)mm (range: 100.17-172.97 mm), 131.84(±12.07) mm (range: 106.03 – 146.79 mm), 142.78(±22.72)mm (range:115.79-170.98 mm). One - way Anova (F) showed that there was not a significant difference in CW between ovigerous females at stage 5 according to depth (F = 2.664, df = 3, p = 0.053).
4. Size (CW50) at First Maturity
The mean size at maturity for females (CW50) collected in the two-years survey was calculated as 113.1 mm CW, using Frequentist regression; (CW50) = 113.1 mm (r2 = 0.97, confidence intervals = 112.8-113.4 mm) (Figure 11).
5. Experimental Sampling
Almost all crabs caught during the experimental trawling in sampling station BAR 1 (depth <1 m) and in BAR 3 (depth <1 m), in March 2022, in Bara sub-area were juveniles (CW <60 mm). In February 2021 were selected only juveniles (N = 51; 25 males and 26 females), with an equal sex ratio, ranged in size (CW) from 11- 34.23 mm with mean(±s.d) = 21.11(±5.35) mm. In March 2022, 52 individuals were caught with 28 males and 25 females and a sex ratio almost reaching equality. Out of 52 crabs, 49 were juveniles and 3 adults: two immature females with CW = 79.09 mm and CW = 88.17 mm, and one male (CW = 101.73 mm). Size of juveniles ranged from 12.51 to 54.7 mm with a mean CW(±s.d) = 28.05(±27.47) mm.
6. Fecundity
Absolute fecundity was measured from 30 ovigerous females caught in June (N=4), July (N = 16) and in August 2021 (N = 10). Mean size(±s.d) of ovigerous females was 133.2(±13.9) mm, bearing eggs of various developmental stages (1-5). The number of eggs ranged from 236,407 to 1,939,139 with a mean(±s.d) 788,031(±447,534). Total egg mass ranged from 5.1 to 27.38 g with an average(±s.d) 13.65(±6.65)g. EMI(%) ranged from a minimum of 4.18 to a maximum of 13.34% with an average(±s.d) 8.18 (±2.18)%. Egg volume fluctuated from 0.00729 to 0.01982 μm3 with an average(±s.d) 0.012 (±0.003) μm3 (N = 290 eggs) whereas egg diameter ranged from 195.5 to 265.25 μm with a mean 224.39 (±20.47) μm. One-way Anova showed that statistical differences were observed for fecundity (F = 4.23; p = 0.025) and egg mass index (EMI) (F = 5.65; p = 0.009) with month, while no significant variation was reported for total egg mass (F = 2.04; p = 0.15), diameter (F = 2.11; p = 0.177) and volume (F = 1.09; p = 0.5). Τukey’s HSD showed that both fecundity and EMI exhibited highest values in June compared to August with p= 0.02 and p < 0.005, respectively, while July showed no statistical differences with the other two months, (Figure 12) with p = 0.087 (July-June) and p = 0.87 (July-August) and p = 0.053 (July-June) and p =0.898 (July-August), respectively. Linear regressions showed that the highest correlation was observed between carapace width and total egg mass (r2 = 0.47, p<0.001), followed by the number of eggs (r2 = 0.41, p<0.001) while egg diameter showed no statistical relationship with carapace width (p>0.05) (Figure 13).
7. Discussion
Female blue crabs Callinectes sapidus have a complex reproductive strategy characterized by two life stage-dependent and directionally opposite migrations from estuarine to oceanic/marine habitats e.g., [43,47,50]. This strategy, part of the life cycle of the blue crab, has been studied thoroughly for more than 100 years in the temperate waters of the Mid and South Atlantic Bight along the east coast of the USA e.g., [42,43,46,47,48,65,66,67,68,69,70,71,72]. In a typical estuarine environment mating generally occurs in low- and mid-salinity areas from spring to fall and females store the sperm of the male to inseminate future egg masses; e.g., [42,43,73]. After mating, females migrate gradually, through two phases, to down-estuary to higher salinity regions (e.g., lower bays, bay mouths, and adjacent offshore areas) (phase I), and subsequently towards spawning grounds (phase II) [43,45,69,70,74,75]. They generally extrude eggs during the summer months and carry them as a mass under their abdomens for ≈2-week development period. Spawning usually takes place within 1 to 2 months after mating and up to 9 months for those inseminated by the end of previous summer and fall [42,45,70,74,76]. Females release larvae near estuary mouths and larvae are transported offshore [47,77,78] where larval development occurs. Wind events transport megalopae as recruits back to coastal regions and estuaries to mature into adults [70,79,80]. Most of the above critical stages of the complex life cycle and reproductive strategy of the female blue crab have been also described from coastal and estuarine habitats of the Mediterranean Sea where the blue crab has invaded and established thriving populations see [15,16,53,81,82]
Sex Ratio and Mating
Fitz and Wiegert [83] stated that sex ratio is presumed to be balanced in most blue crab populations. In Thermaikos total sex ratio of the population does not deviate from equality; unexpectedly, since blue crab stock has been intensively exploited for more than fifteen years in the inner gulf and large male blue crabs were scarcely found in the fished population (unpublished Kevrekidis et al.). During a study in Methoni Bay (Thermaikos Gulf) in the period 2011-2012 males outnumbered females with a ratio of 2:1 [15]. In Monolimni Lagoon (Evros River estuary) total sex ratio skewed towards males [16]. Males dominance is reported from Italian, Egyptian and Croatian lagoons [18,34,84,85] but also for blue crab populations along the mid-Atlantic Bight [83,86]. However, sex ratio was found to be highly biased towards females along the southwestern Coast of Turkey [21,52,54].
Female blue crabs have a single opportunity to mate, immediately after their final (pubertal) molt to maturity [42,87]. Mating involves intricate interactions between males and females [43,88,89]( and a sex ratio 1:1 is expected. However, in blue crabs, the structure of local populations, such as the number of males and females that are ready to mate (operational sex ratio), influences an individual’s ability to find and compete for receptive mates. Moreover, female blue crabs mature asynchronously and are thought to mate only once, so that the pool of receptive males is usually larger than that of receptive females. Additionally, males mature at a smaller size than females [43]. Rains et al. [90] found that although the operational sex ratio in six tributaries of Chesapeake Bay was variable, both of the forementioned factors can remain sex ratio skewed toward males even if the male : female sex ratio of a blue crab population is skewed toward females. However, in the Thermaikos a monthly sex ratio 1:1 in April 2021 and May 2022 and in June, July and September through November in both years was assessed, indicating that from mid spring to mid-summer and in fall mating is probably taking place and that estimated sex ratio corresponded with the operational sex ratio. In Monolimni Lagoon (Evros River estuary, northeastern Aegean) mating probably occurs in early summer (June) in hyposaline areas when a sex ratio of 1:1 was recorded [16].
Size at First Maturity
In the present study mean size (CW) of females, including immature and mature females, was lower (125.67 mm) than that estimated from Monolimni Lagoon (130 mm) with both studies contacted at the same period of time (2020-2022) [16]. However, mean size of mature females (including ovigerous females) and of ovigerous females were higher than those in the Levantine Sea (Inskederun Bay)[52] with 133.71 and 130.33 and 123.7 and 120.2 mm, respectively. Moreover, mean size at maturity (CW50) estimated in this survey (113 mm) was lower than that in Monolimni Lagoon (123.8 mm) [16] and in Trapani saltmarshes (120 mm) (Sicily, central Mediterranean) [53] where blue crab populations have been only recently exploited intensively. However, size at first maturity was similar than that reported from Iskenderun Bay (114.1 mm) [52] and lower than that in Beymelek lagoon (118.5 mm) in the Levantine Sea [21,52,54], indicating that an intense fishery pressure upon the blue crab population was taking place also in the Thermaikos Gulf. Lipcius and Stockhausen [71] reported that in the Chesapeake Bay, where the largest fishery for the blue crab both today and historically exists along the Atlantic coast, size at maturity ranged from 108 to 118 mm CW, as CW50 decreased from 118.4 mm between 1988 and 1991 to 107.9 mm CW between 1992 and 2000 [71]. The same authors proposed that this reduction was related to poor recruitment, in concert with high fishing mortality of the stock, which was at or near overexploitation, and of natural mortality. A significant variation in CW50 is reported from other native areas of the species; in New England tidal rivers, Taylor and Fehon [91] estimated size at maturity at 129 mm, in Maryland coastal Bays it was estimated at 116 mm CW [86], and in Florida estuaries in the Atlantic coast CW50 ranged from 123.7 to 129.5 mm CW whereas in North Gulf estuaries from 116.5 to 121.5 mm CW [92]. Significantly lower values (102-103.3 mm CW) were estimated on the southern coast of Brazil [93,94]. However, out of the effect of fishing intensity Fisher [95]stated that environmental factors also play an important role on estimating life history parameters with female blue crabs mature at smaller sizes as temperature and salinity increases.
Ovarian Development
During the two-year survey in the Thermaikos Gulf, females blue crabs exhibited a protracted reproductive period; macroscopical inspection of the ovarian development according to Hard [62] has shown that mature ovaries (Stage III) were present in all months with however low percentages recorded during the cold period ` (December to March) of the year. The highest percentages of mature ovaries (stage III) were observed from May to August but also in October and in November. In both years stage IV appeared from May to September and peaked in July, whereas stage V appeared from May to November with a peak in September. Stage I and II were present in all months throughout the two years with higher percentages during winter and early spring.
In the present study histological analysis assigned five stages of ovarian development (stov 1- stov 5) of female C. sapidus. The description of the five stages was similar to the one made by Carvalho-Saucedo et al. [96] who previously also assigned five stages on the ovarian development of the blue crab. These stages (stov 1- stov 5) however did not correspond to the five stages (I-V) proposed by Hard [62] based on gross appearance. In the present study ovaries assigned into stage II according to Hard, which were visually differentiated from stage I and stage III due to color and size, might histologically belong to gametogenesis (stov 2) or development (stov 3) stages. Secondly, ovaries at stage IV corresponded according to Hard [62]to ovaries between first and second ovulation was not identified as a separated stage and thus it was categorized histologically into the mature stage (stov 4).
In Evros estuary, NE Aegean Sea (temperature range November 2020- October 2021: 6-27 0C) females with mature ovaries (stage III) were observed for an 8-month period, from late winter (February) to October (mid-fall) whereas stage IV peaked in July and stage V appeared from June to October [16] . In Iskenderun Bay, Tureli et al. [52] reported that mature and ripe ovaries were found all year round with higher percentages from August to October. In the Atlantic coast ovarian development in female blue crabs may take from as little as 2 months to as much as 9 months after mating. Environmental conditions, especially temperature, play a role in determining the length of time it takes for the ovary to develop with warmer temperatures speed development. Moreover, time of mating determines the length of time it takes the ovary to develop; if mating occurs in late spring or early summer, the ovary should be well developed and ready for egg extrusion by late summer. If mating occurs late in the summer or early in the fall, the ovary develops during the winter and egg extrusion does not occur until late spring to mid-summer of the following year [69,97].
Thus, timing of brood production is influenced by environmental conditions and there is a temporal and spatial variation in the timing and duration of the spawning season with spawning initiated earlier in the spring at lower latitudes [43,73,74,98]. Monthly presence of ovigerous females and especially of ovigerous females bearing eggs in the late developmental stage 5, i.e prior to eggs’ release and hatching, showed that the spawning period of the blue crab in the Thermaikos Gulf extends from late April to late October. However, taking into consideration that a ) ovigerous females with eggs in early developmental stage were caught at the end of March 2012 during a study conducted in the period 2011-2012 in Methoni Bay (see [15] and b) the time needed for ovigerous females with eggs in early developmental stage to the time that eggs are released and hatched has been estimated approximately to 14 days ( e.g., [43], indicates that the spawning period of the blue crab in the Thermaikos Gulf extends for an approximately seven - month period, i.e., mid-April to late October. Ovigerous females were observed for a six month period (May to October) in Evros estuary [16], for an eight month period (February-September) in Beymelek Lagoon whereas in Iskenderun Bay for only 6 months (May to September)[52]. Along the east coast of the Atlantic the length of the blue crab spawning season increases as a function of water temperature lasting about three months in N. Jersey (June to August) [43], four months in Chesapeake Bay (May to September [42,77] five months in south Carolina [99] , nine months in Florida (February to October) [100] or almost year- round [92] and year-round in southeast Brazil [93,94]. Thus, a longer spawning season allows production of additional broods per year because blue crabs are multiparous (see below fecundity). However, Differences in reproductive timing between tropical, subtropical, and temperate populations of blue crab hint at potential effects of warming on the length of the spawning season, yet changes in the duration of the spawning season due to climate change within geographic locations are expected [48].
Ovigerous Females
In Thermaikos a strong but non-significant correlation was found between temperature and ovigerous females; the highest number of ovigerous females but also of ovigerous females with eggs at stage 5 recorded for both years in July followed by August. Therefore, spawning of the blue crab in Thermaikos Gulf peaked clearly in July and August at temperature ranged from 26 to 29 0C in inshore waters (see below),(see below) indicating a non-interannual temporal variation on spawning. However, a temporal variation on females spawning peak was found in the study conducted in the period 2011-2012 in Methoni Bay [15]. In this one-year study consisted by bimonthly samplings the highest number of ovigerous was caught in May (N = 22) followed by July (N = 9) with the remaining 2 specimens fished in March having eggs in early developmental stages. Moreover, the highest number of ovigerous crabs with eggs in late developmental stages was recorded also in May (N=13) with another two caught in July. However, in the present study no one ovigerous was found in May in any sampling station of Methoni Bay; inversely, in both spawning seasons (2021 and 2022) the greatest number of ovigerous in the Bay was caught in July followed by August. Similarly, in the southeastern Mediterranean the highest number of ovigerous females was observed in July and August (Iskenderun=) Bay) [52] , whereas in the Beymelek Lagoon spawning period lasted between July and September [54]. However, in Evros estuary ovigerous females at stage 5 peaked in September [16].
The spatial distribution of ovigerous females showed that they were found mostly in inshore waters in almost all sampled sub-areas of the inner gulf from the Axios estuary in the east to Methoni Bay to the west. Ovigerous females were found also in Loudias River at 800 m distance from the rivers’ opening to the sea, having eggs in early developmental stage, implying that ovigerous females bearing eggs at the early stages of development may also tolerate extremely low salinity values corresponding to freshwater conditions (see also [43]
No one ovigerous was found at the sampling station of the long line mussel culture station (10 m depth in Loudias bay and at 15 and 25 m offshore Aliakmon estuary (see below). The highest number of ovigerous females but also of ovigerous females with eggs at stage 5 were found in the seasonal sampling stations AX1 and AX2 and in AL2 and AL3 situated at the adjacent coastal waters of Axios and Aliakmon estuaries, respectively indicating probably a site selection by the ovigerous females. These stations are characterized by a bare substratum at depths from 0 to 3 m with sandy-mud substrate and with no seagrass/seaweed vegetation. Inversely, Loudias along with Bara and Methoni Bays from 0 to 2 m depth are characterized by a muddy and muddy-sand substrate and the seasonal presence of Gracilaria sp and permanent Cymodocea nodosa meadows, respectively, but also by a bare substrate at 3 m. However, it must be noted that stations AX1 and AX2 along with stations AL2 and AL3 are sites that are not fished intensively or not fished at all by the fishermen throughout the year, respectively, compared to Bara, Loudias and Methoni sub-areas. Thus, it is believed that no spatial-specific spawning site seems to be selected by the female’s blue crab in Thermaikos and that spawning occurs in inshore waters along the estuarine coastline of the northwestern part of the inner gulf of the inner gulf.
However, in Thermaikos depth plays a significant role in site selection by the spawning females; the number of both ovigerous and ovigerous with eggs at stage 5 in both years was higher at 3 m and 1 m than at 6 m and at 9 m depth gradients, whereas no one ovigerous was found at 10 m, 15 m and 25 m depth gradients. Moreover, in both years during the spawning peak (July-August) and the spawning periods the number of ovigerous and of ovigerous at stage 5 was higher at 3 m than at all other depth gradients, respectively. However, it must be noted that in the present study the highest frequency (75%) of females at stage 5 was found at 9 m gradient, with however a very small number caught, probably due to a need for these ovigerous to have further isolation and protection from predators during hatching and release of larvae. The above showed that in Thermaikos a) spawning sites are found in the inshore waters of the inner gulf along a depth gradient (see below) and c) a strong negative and moderate correlation exists between ovigerous females and ovigerous females at stage 5 with depth, respectively. Similarly, in Evros estuary a seaward migration of ovigerous females for spawning was observed with most of them found at the adjacent offshore waters of Monolimni lagoon at 1 m followed by 3 m and 6 m gradients indicating also a strong negative correlation between ovigerous females and depth. Moreover, the highest frequency of ovigerous at stage 5 was recorded at 6 m gradient (75%) indicating also a moderate correlation between ovigerous females at stage 5 and depth [16]. In native areas female blue crabs tend to migrate to lower bays, bay mouths, and adjacent offshore areas for spawning. In the Lower Chesapeake Bay distribution of adult females ranged from 6 to 14 m whereas the blue crab spawning stock shifts spatially both during the reproductive period and between years; [102,103]. In south Carolina (Charleston Harbour) 96% of spawning females were found at 7 to 11 m depth [99]. Moreover, a significant depth and distance differentiation of the spawning grounds from the coast is reported: In the Gulf of Mexico sand shoals 20–50 km offshore of the Louisiana coast have been identified as spawning grounds; these shoals are high-relief, subaqueous stands of mostly sandy sediment (water depth 3 to 10 m) [104,105]. In the relatively shallow bathymetry of Texas’ inshore areas, CPUE of egg-bearing females in the early era was highest in waters between 1 and 10 m whereas CPUE during the late era of the year was highest in depths approaching 15 m [106]. In Georgia summer peak spawning occurred primarily on the continental shelf as far as 13 km offshore [107].
Although the timing of various components of the model (phases I and II) for the seaward migration of females differ among estuaries in tropical, subtropical and temperate regions, the sequence of events seems to be similar. However, in Chesapeake Bay females’ seaward migration exhibits a significant spatiotemporal differentiation depending on mating site: Females that mate in the upper Bay must migrate long distances (>200 km) to reach high salinity spawning grounds. Thus, compared with females that mate in the lower Bay, females that mate in the upper Bay arrive at the spawning grounds later in the year [68] often overwintering en route [69]. However, Epifanio [47] stated that little is known about phase I migration in small estuaries, where the average depth is much shallower than that in Chesapeake Bay and the distance from mating grounds to brooding grounds is much shorter (85% moved < 15 km) [75,108,109]. In Thermaikos Gulf, Axios and Aliakmon estuarine riverine systems including enclosures and lagoon like formations are shallow and relatively small and the distance from mating sites to estuarine mouths and inshore coastal waters is up to 3 km at most cases. Thus, the duration of sea ward migration of inseminated and of ovigerous females [45,47] (phase I) is very short. Furthermore, in phase II ovigerous females from estuarine mouths and ovigerous females that inhabit inshore coastal waters up to 1 m and do not undertake phase I, migrate at slightly deeper waters for spawning, at 1 m and up to 9 m depth gradient but mainly at 3 m depth gradient which is the foremost selected spawning depth of the blue crab in the gulf. Thus, in Thermaikos due to its shallow and relatively small estuaries, the gentle slope and low bathymetry of the inner gulf, the reproductive strategy of female blue crab is marked by a short distance between spawning phases I and II.
Epifanio [47,72] stated that larval development of the blue crab includes seven zoeal stages followed by a single megalopa stage; development of zoeae occurs in the open coastal waters and requires 3-4 weeks under favorable conditions whereas recruitment to juvenile populations is dependent on advective transport of megalopa back to the estuary. Therefore, the extended spawning period observed in Thermaikos indicates that an extended recruitment is expected to take place, too; thus, several cohorts of megalopae of the blue crab are expected as recruits to engross the inlets canals and enclosures in the estuaries serving as nurseries in the estuarine areas of the gulf favored by southward winds and sea currents during summer and early fall. Once in the estuary, settlement and metamorphosis of the megalopa are stimulated by chemical cues associated with appropriate nursery habitat [110]. Newly metamorphosed juveniles are a few mm in carapace width and remain in juvenile habitat for several months until they reach a width of 20-30 mm [47] when they began disperse long distances into low salinity sub-estuaries to feed and grow to maturity in two years [111]. In western Mediterranean (Spain) the highest zoea density was detected at 1 km off Albufeira Lagoon and settlement peak occurs between September and November for megalops and juveniles (<20 mm CW) [82]. In Iskenderun Bay blue crab settlement occurred between mid-June and mid-October with the densest settlement recorded in August and September on various bottom substrates. In the Beymelek recruitment to the benthic population started at the end of January with juvenile found almost throughout the year. In Thermaikos during the two-year survey, very few juveniles (N=19) (CW range: 30-60 mm) were caught from February to April and in July in all sampled sub areas due mainly to selectivity of the mesh size of fyke nets. However, in the experimental trawlings conducted in February 2021 and March 2022 at depth <0,5 m in Bara subarea a significant number of juveniles with a mean CW = 22 mm and CW = 25 mm, respectively, was found in the muddy substrate. Additionally, in the trawling of March were caught only two adult crabs as it is believed that the others escaped due to their swimming capability. In both the experimental trawlings a 1:1 sex ratio was estimated for juveniles; sex ratio is presumed to be balanced especially in the juvenile stages of a blue crab population [83]; These samplings were conducted after a 6-7 month period from the spawning peak in the gulf (July-August) and the juveniles caught may represent the main cohort that enters the estuaries and coastal inlets in the inner gulf. Therefore, it seems that the newly crabs undergo a migratory coastward pattern and engross into estuarine coastline, inlets canals and lagoon like formations which serve as nursery areas. Thus, juveniles are distributed at very shallow waters (<0.5 m depth) and share the same micro-habitats with immature and mature adults in the different sub-areas of the inner gulf. Moreover, in the Thermaikos Gulf 90 and 88% of the total number of blue crabs were caught at ≤3m during the first and the second year of the survey, respectively (Kevrekidis et al., unpublished data). Thus, the blue crab population is aggregated in the adjacent inshore waters (e.g., Loudias Bay), embayments (e.g., Methoni Bay) and lagoon like formations (e.g., Bara) of the inner gulf where blue crabs become mature and mate, probably in sites associated with meadows of C. nodosa (METH 1) or Gracilaria sp. (BAR 2) and spent most of their life cycle. Consequently, inseminated and ovigerous crabs migrate from the above areas but also from Loudias River to release larvae at 3 m and up to 9m and at a distance up to 2 km offshore (Figure 14).
Fecundity
Female blue crabs are known for their highly reproductive rate and fecundity (see below) [43] characteristics of an r-selected strategy [112].Within the Mediterranean the high fecundity of the invasive portunid enhance the possibilities of a successful establishment in a selected habitat followed by initial population outbreaks in most cases [38] and by a further dynamic expansion into new coastal and estuarine habitats throughout the Basin.
Estimates of fecundity of the blue crab varied significantly; it ranged from 700,000 to a mean of 3.2 million of eggs in the Middle and south Atlantic Bight [42,48,49,65,92,113] and from 1.8 to 3.27 million eggs in northern Gulf of Mexico [64,92,114,115], whereas in South Atlantic [94] and [93] estimated a mean of 2 million and a median of 978,000 eggs per female, respectively. In the Mediterranean information is scarce and relevant studies have been conducted only in Sicily (central Mediterranean) [53] and in Iskenderun Gulf (Alexandretta) Levantine sea [52] with mean fecundity estimated at 1.5 million (985,000 to 2.1 million) and 1.91 million (667,950–4,669,853) respectively. In the present study mean fecundity (788,031) was close to the lower range of estimated fecundity in both native and invasive areas of the blue crab (788,031 eggs) (236,407 to 1,939139).
During mating the blue crab female may store sperm sufficient for repeated future spawnings [42,43,115]. Brood production is a complicated process involving the temporal and spatial coordination of female physiology with particular environmental condition and the number of broods and size is influenced by a variety of factors including temperature salinity and size [43]. In temperate waters females appear to live about 3.5 yr and are capable of producing one to three broods per spawning season and up to 6–7 broods during their lifetime. [42,43,48,49,62,73] and the present study confirms that a female can produce at least two broods in single spawning season as there were found ovigerous females having ovaries at stage IV (female with full ovaries carrying egg mass) and stage V (female after the first ovulation carrying egg mass) (see [62]). However, in subtropical waters blue crabs can produce up to 6-7 broods per year and up to 16-18 broods over their lifespan [43,73,115,116].
The wide differences on mean fecundity estimated for C. sapidus could be partially explained by the number of broods that produced by the female blue crab and included in estimates of fecundity. Fecundity has been found to decrease with successive broods [76,116,117]. Graham et al. [115] stated that in the north Gulf of Mexico primiparous spring females were the most fecund and multiparous (second to more broods) summer/fall females the least fecund but they found no statistical difference in fecundity by brood class and season. However, [48] Schneider et al. (2024) stated that in Chesapeake Bay month had a substantial effect on female fecundity; spawners in June were mainly primiparous and had the highest quantities of stored sperm and reproductive potential and that across months primiparous females produced greater quantities of eggs than multiparous females with primiparous females in August exhibiting the highest mean fecundity. In the present study fecundity and egg mass index (EMI) exhibited highest values in June compared to August whereas in July showed no statistical differences with the other two months whereas no difference was found for total egg mass diameter and volume between months. Thus, the lower mean fecundity estimated in the present survey compared to all other studies could be partially due to that no one primiparous spring female (first brood) was examined and that the number of females analyzed from June was low (N=4) and that probably females examined in July and August were mostly multiparous. Moreover, in the present study abundance of ovigerous females varied by month indicating that annual egg production may fluctuate substantially as a result of interannual fluctuations in abundance and fishing pressure.
In the present study fecundity and total egg mass were related positively with size (CW); a positive relationship between fecundity and CW has been documented from mid-Atlantic and the north Mexico Gulf [44,48,49,115]. A weak positive linear relationship between fecundity and CW as well as with total egg weight was reported from the Levantine Sea [52]. Moreover, in Thermaikos no relationship between fecundity and CW with egg diameter was found whereas Graham [115] found an inverse relationship between fecundity and egg diameter and that CW and egg diameter were positively correlated. In the present study no difference was found for egg diameter among months whereas [115] found no difference between brood classes and egg diameter.
Management Implications
In the native areas such as in the mid and south Atlantic Bight the blue crab is highly exploited and among common strategies for management of its fishery are measures to protect the spawning stock of mature females, which have been employed in the Chesapeake Bay (Chesapeake Bay Program, 1997) and in the North Carolina (NCDMF, 2004)[48]. More specifically, spawning sanctuaries have been established and regulations have been implemented across Chesapeake Bay to prohibit the catch of female blue crabs during egg-bearing stages of the reproductive cycle [71] due to significant spawning stock-recruitment relationship [71,79,118,119].In Virginia, the winter dredge fishery, which mostly harvested females from the spawning grounds prior to the spawning season, was closed; the historic spawning sanctuary was expanded to protect 50% of the spawning stock and the harvest of females with dark egg clutches, which hatch within days, was prohibited [30,103] In Maryland, access to the female fishery was restricted and female daily catch limits were lowered (Chesapeake Bay Stock Assessment Committee, 2023)
On the contrary, in the Mediterranean where the invasive blue crab has developed nowadays thriving populations throughout the Basin its impacts on the structure and function of the diverse coastal habitats of the Basin remains up today largely unknown whereas significant negative socioeconomic impacts have been reported. Only recently the Commission (2022) has initiated the preparation of a risk assessment for Callinectes sapidus, which is required before considering whether to add it as an invasive alien species (IAS) of Union concern. Moreover, a research program has been developed by the General Fisheries Commission for the Mediterranean (GFCM) to understand what the best solutions are to manage and mitigate the impacts of this species in the Mediterranean. As rates of marine bioinvasions continue to increase and eradication of marine invasive species has been achieved in rare cases, characterized by early detection and rapid response in restricted areas (e.g [120] pressure will increase on marine managers to reduce the impacts of invaders, and on scientists to provide control options [39,121]. A major question is, therefore, how to distribute treatment efforts cost-effectively across space and time to prevent or slow the spread of invasive species [122]. Giakoumi et al. [41] proposed 11 management actions for the control of invasive species; the physically (mechanically) remove of the species, at early stages, however, of the species establishment is proposed among others as the first action that has to be taken.
However, for more than a decade the blue crab has begun a valuable resource for the Mediterranean artisanal fishery from the northern Aegean Sea in the east to Tunisia in the west and from Egypt in the south to the north Adriatic. Thus, commercial fisheries of local importance have been developed throughout the Basin and the blue crab is intensively exploited as no legislation rules are imposed for its fishery at least in the southern European countries. Therefore, for a considerable time a discussion on the policy that the EE and southern European countries should follow for the management of the fishery of this invasive portunid crab either towards maintenance of sustainable fisheries or eradication and population size control, is still in progress. Specifically, the management of the fishery of the blue crab should require temporal and/or spatial management measures and actions to be implemented either for the protection of the recruitment and spawning stock (e.g., season and area closures) or an intense fishery targeting on the spawning stock aiming to reduce reproductive output and recruitment, minimizing thus the population size, respectively [64]. However, the implementation of any of the above management policies require a good knowledge of the complex life cycle and reproductive strategy of the blue crab. Nevertheless, Mediterranean countries should gain the knowledge of how to control population outbreaks of the species especially when these produce a significant risk for socioeconomical activities, e.g., fisheries and aquaculture production on a local scale, as to mitigate or to minimize its impacts. Thus, the knowledge of the critical life history stage of spawning of the blue crab and especially the spatiotemporal and bathymetrical distribution of ovigerous females in every invaded coastal area (lagoon, estuaries, bays, etc), as to intervene through management measures and actions to control and probably drastically minimize their number is a prerequisite.
Under the above perspective the present study provides a key knowledge for the implementation of management measures and actions aimed to control population size and outbreaks of the blue crab, by targeting on the spawning stock. These measures will be based on the critical information acquired on spatiotemporal and bathymetrical distribution of ovigerous females in Thermaikos Gulf which can be used as a ‘case study’. The intensity and the time the actions needed to be conducted depend on the magnitude of the population outbreaks, the type of habitat and the effectiveness of the fishing gears. Especially, in Thermaikos Gulf the two-year survey of the reproductive biology of the blue crab confirmed the short migratory movements of mature and ovigerous females from estuarine areas and inshore waters of low depth to slightly deeper inshore waters for spawning. Thus, spatiotemporal distribution showed that 88% of all ovigerous were found in July and August along the shoreline from Axios estuary to Methoni Bay with 61%of ovigerous found at a depth gradient of 3 m and with a sex ratio 1:2 (♂/♀). Thus, management measures towards minimization of the population size of the blue crab could be a selective intense fishing of ovigerous females exclusively in July and August that it would be critical as to reduce the spawners before being able to reproduce. These measures could be accomplished with the use of suitable fishing gears (i.e., fyke nets or crab pots (Luisiana type) without a bait, deployed at the depth gradient of 3 m along the shoreline from Methoni Bay in the west to Axios Delta to the east in the inner Thermaikos Gulf. As recruitment – size of spawning stock relationship has been documented e.g., [71,79] ultimately the above measures will affect recruitment that will end up on a significant reduction of the spawning stock and the minimization of the size of the blue crab population in Thermaikos Gulf.
However, as the blue crab is commercially an important resource it is expected that its populations will remain in the future under intense fishery exploitation throughout the Mediterranean Basin. The remarkable viability of its populations against intense fishery exploitation through time is believed to be due to the peculiar biological characteristics of the species (e.g multiple spawning and high fecundity, omnivory feeding strategy, very few predators) enhanced by the climate change and the consequent increased of the sea surface temperature in the Mediterranean providing higher survival rates during overwintering. Furthermore, since until now no thorough research has been conducted that elucidates the blue crab impact on the various coastal ecosystems of the Basin it is more probable that the proposal of the sustainable exploitation of the blue crab will be advanced by the EE and the Mediterranean countries. This policy will however inevitably propose temporal and spatial measures for the protection of the blue crab population and its spawning stock. Nevertheless, this policy will not be probably applicable in every country, as each one of them encounters a different degree of risk against the blue crab, especially in regions where significant ecological reserves, fishery or aquaculture industry is developed. Thus, these countries will be forced to restrain the population outbreaks and/or control blue crabs populations size through implementation of management measures/actions targeting on spawning stock through intense fishery.
References
- Thongda, W.; Chung, J.S.; Tsutsui, N.; Zmora, N.; Katenta, A. Seasonal Variations in Reproductive Activity of the Blue Crab, Callinectes Sapidus: Vitellogenin Expression and Levels of Vitellogenin in the Hemolymph during Ovarian Development. Comp Biochem Physiol A Mol Integr Physiol 2015, 179, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.B. THE SWIMMING CRABS OF THE GENUS CALLINECTES (DECAPODA: PORTUNIDAE). Fishery Bulletin 1974, 72, 685–798. [Google Scholar]
- Millikin, M.R. Synopsis of Biological Data on the Blue Crab, Callinectes Sapidus Rathbun; National Oceanic and Atmospheric Administration, National Marine Fisheries Service, 1984. [Google Scholar]
- Johnson, D.S. The Savory Swimmer Swims North: A Northern Range Extension of the Blue Crab Callinectes Sapidus? Journal of Crustacean Biology 2015, 35, 105–110. [Google Scholar] [CrossRef]
- Mancinelli, G.; Bardelli, R.; Zenetos, A. A Global Occurrence Database of the Atlantic Blue Crab Callinectes Sapidus. Sci Data 2021, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, E.L. Sur Un Callinectes Sapidus M. Rathbun Trouvé à Rochefort. Bull. Mus. Hist. Nat. 1901, 7, 16–17. [Google Scholar]
- Giordani, S. Neptunus Pelagicus (L.) Nell’Alto Adriatico (Neptunus Pelagicus in the Northern Adriatic). Natura 1951, 42. [Google Scholar]
- Serbetis, C. Un Nouveau Crustacé Comestible En Mer Egée Callinectes Sapidus Rathb. (Décapode Brachyoure). Proc. Gen. Fish. Comm. Mediterran. 1959, 5, 505–507. [Google Scholar]
- Georgiadis, C.; Georgiadis, G. Zur Kenntnis Der Crustacea Decapoda Des Golfes von Thessaloniki. Crustaceana 1974, 26, 239–248. [Google Scholar] [CrossRef]
- Riedl, R. Fauna Und Flora Des Mittelmeeres. Internationale Revue der gesamten Hydrobiologie und Hydrographie 1985, 70, 442–442. [Google Scholar] [CrossRef]
- Nehring, S. Invasion History and Success of the American Blue Crab Callinectes Sapidus in European and Adjacent Waters. In the Wrong Place - Alien Marine Crustaceans: Distribution, Biology and Impacts; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-94-007-0591-3. [Google Scholar]
- Holthuis, L.B.; Gottlieb, E. The Occurrence of the American Blue Crab, Callinectes Sapidus Rathbun, in Israel Waters. Bull. Res. Counc. of Israel. 1955, 5, 154–156. [Google Scholar]
- Holthuis, L. Report on a Collection of Crustacea Decapoda and Stomatopoda from Turkey and the Balkans. Zoologische Verhandelingen 1961, 47, 1–67. [Google Scholar]
- Banoub, M.W. Survey of the Blue-Crab Callinectus Sapidus (Rath.), in Lake Edku in 1960. Alex. Institute. of Hydrobiology Egypt 1963, 1–17. [Google Scholar]
- Kevrekidis, K.; Antoniadou, C. Abundance and Population Structure of the Blue Crab Callinectes Sapidus (Decapoda, Portunidae) in Thermaikos Gulf (Methoni Bay), Northern Aegean Sea. Crustaceana 2018, 69, 641–657. [Google Scholar] [CrossRef]
- Kevrekidis, K.; Kevrekidis, T.; Mogias, A.; Boubonari, T.; Kantaridou, F.; Kaisari, N.; Malea, P.; Dounas, C.; Thessalou-Legaki, M. Fisheries Biology and Basic Life-Cycle Characteristics of the Invasive Blue Crab Callinectes Sapidus Rathbun in the Estuarine Area of the Evros River (Northeast Aegean Sea, Eastern Mediterranean). Journal of Marine Science and Engineering 2023, 11, 462. [Google Scholar] [CrossRef]
- Ramadan, Sh.E.; Dowidar, N.M. Brachyura (Decapoda Crustacea) from the Mediterranean Waters of Egypt. Thalassia Jugoslavica 1972, 8, 127–139. [Google Scholar]
- Abdel Razek, F.A.; Ismaiel, M.; Ameran, M.A.A. Occurrence of the Blue Crab Callinectes Sapidus, Rathbun, 1896, and Its Fisheries Biology in Bardawil Lagoon, Sinai Peninsula, Egypt. The Egyptian Journal of Aquatic Research 2016, 42, 223–229. [Google Scholar] [CrossRef]
- Bulgurkov, K.I. Callinectes Sapidus Rathbun in the Black Sea. Izvest. Niors. 1968, 9, 97–99. [Google Scholar]
- ENZENRO§, R.; ENZENRO§, L.; BİNGEL, F. Occurrence of Blue Crab, Callinectes Sapidus (RATHBUN, 1896) (Crustacea, Brachyura) on the Turkish Mediterranean and the Adjacent Aegean Coast and Its Size Distribution in the Bay of Iskenderun. Turkish Journal of Zoology 1997, 21, 113–122. [Google Scholar] [CrossRef]
- Türeli, C.; Miller, T.J.; Gündoğdu, S.; Yeşilyurt, İ.N. Growth and Mortality of Blue Crab (Callinectes Sapidus) in the North-Eastern Mediterranean Sea. Journal of FisheriesSciences.com 2016, 10, 55–62. [Google Scholar]
- F. Mehanna, S.; G. Desouky, M.; E. Farouk, A. Population Dynamics and Fisheries Characteristics of the Blue Crab Callinectes Sapidus (Rathbun, 1896) as an Invasive Species in Bardawil Lagoon, Egypt. Egyptian Journal of Aquatic Biology and Fisheries 2019, 23, 599–611. [Google Scholar] [CrossRef]
- Perdikaris, C.; Konstantinidis, E.; Gouva, E.; Ergolavou, A.; Klaoudatos, D.; Nathanailides, C.; Paschos, I. Occurrence of the Invasive Crab Species Callinectes Sapidus Rathbun, 1896, in NW Greece. Walailak Journal of Science and Technology (WJST) 2016, 13, 503–510. [Google Scholar]
- Kevrekidis, K. Callinectes Sapidus (Decapoda, Brachyura): An Allochthonous Species in Thermaikos Gulf. Fish. News 2010, 340, 44–49. [Google Scholar]
- Castriota, L.; Falautano, M.; Perzia, P. When Nature Requires a Resource to Be Used—The Case of Callinectes Sapidus: Distribution, Aggregation Patterns, and Spatial Structure in Northwest Europe, the Mediterranean Sea, and Adjacent Waters. Biology 2024, 13, 279. [Google Scholar] [CrossRef] [PubMed]
- Chaouti, A.; Belattmania, Z.; Nadri, A.; Serrão, E.; Encarnação, J.; Teodósio, A.; Reani, A.; Sabour, B. The Invasive Atlantic Blue Crab Callinectes Sapidus Rathbun, 1896 Expands Its Distributional Range Southward to Atlantic African Shores: First Records along the Atlantic Coast of Morocco. BIR 2022, 11, 227–237. [Google Scholar] [CrossRef]
- Aydin, M.; Karadurmuş, U.; VEREP, B.; Gözler, A. Expansion of the Distribution Range and Size of the Invasive Blue Crab on the Turkish Coast of the Black Sea. Journal of Anatolian Environmental and Animal Sciences 2024, 9, 127–131. [Google Scholar] [CrossRef]
- García-Gómez, G.; García-Herrero, Á.; Sánchez, N.; Pardos, F.; Izquierdo-Muñoz, A.; Fontaneto, D.; Martínez, A. Meiofauna Is an Important, yet Often Overlooked, Component of Biodiversity in the Ecosystem Formed By. Invertebrate Biology 2022, 141, e12377. [Google Scholar] [CrossRef]
- NOAAFisheries(2024). Available online: https://www.fisheries.noaa.gov/foss/f?p=215:200:11552239188158 (accessed on 29 August 2024).
- Schneider, A.K.; Fabrizio, M.C.; Lipcius, R.N. Reproductive Potential of the Blue Crab Spawning Stock in Chesapeake Bay across Eras and Exploitation Rates Using Nemertean Worms as Biomarkers. Marine Ecology Progress Series 2023, 716, 77–91. [Google Scholar] [CrossRef]
- Prado, P.; Baeta, M.; Mestre, E.; Solis, M.A.; Sanahuja, I.; Gairin, I.; Camps-Castellà, J.; Falco, S.; Ballesteros, M. Trophic Role and Predatory Interactions between the Blue Crab, Callinectes Sapidus, and Native Species in Open Waters of the Ebro Delta. 2024. [Google Scholar] [CrossRef]
- Rifi, M.; Basti, L.; Rizzo, L.; Tanduo, V.; Radulovici, A.; Jaziri, S.; Uysal, İ.; Souissi, N.; Mekki, Z.; Crocetta, F. Tackling Bioinvasions in Commercially Exploitable Species through Interdisciplinary Approaches: A Case Study on Blue Crabs in Africa’s Mediterranean Coast (Bizerte Lagoon, Tunisia). Estuarine, Coastal and Shelf Science 2023, 291, 108419. [Google Scholar] [CrossRef]
- Mancinelli, G.; Chainho, P.; Cilenti, L.; Falco, S.; Kapiris, K.; Katselis, G.; Ribeiro, F. The Atlantic Blue Crab Callinectes Sapidus in Southern European Coastal Waters: Distribution, Impact and Prospective Invasion Management Strategies. Marine Pollution Bulletin 2017, 119, 5–11. [Google Scholar] [CrossRef]
- Glamuzina, L.; Conides, A.; Mancinelli, G.; Glamuzina, B. A Comparison of Traditional and Locally Novel Fishing Gear for the Exploitation of the Invasive Atlantic Blue Crab in the Eastern Adriatic Sea. Journal of Marine Science and Engineering 2021, 9, 1019. [Google Scholar] [CrossRef]
- Öndes, F.; Gökçe, G. Distribution and Fishery of the Invasive Blue Crab (Callinectes Sapidus) in Turkey Based on Local Ecological Knowledge of Fishers. JAES 2021, 6, 325–332. [Google Scholar] [CrossRef]
- Chairi, H.; González-Ortegón, E. Additional Records of the Blue Crab Callinectes Sapidus Rathbun, 1896 in the Moroccan Sea, Africa. BIR 2022, 11, 776–784. [Google Scholar] [CrossRef]
- Levine, J.M. Biological Invasions. Current Biology 2008, 18, R57–R60. [Google Scholar] [CrossRef]
- Clavero, M.; Franch, N.; Bernardo-Madrid, R.; López, V.; Abelló, P.; Queral, J.M.; Mancinelli, G. Severe, Rapid and Widespread Impacts of an Atlantic Blue Crab Invasion. Marine Pollution Bulletin 2022, 176, 113479. [Google Scholar] [CrossRef]
- Thresher, R.E.; Kuris, A.M. Options for Managing Invasive Marine Species. Biological Invasions 2004, 6, 295–300. [Google Scholar] [CrossRef]
- Usseglio, P.; Selwyn, J.D.; Downey-Wall, A.M.; Hogan, J.D. Effectiveness of Removals of the Invasive Lionfish: How Many Dives Are Needed to Deplete a Reef? PeerJ 2017, 5, e3043. [Google Scholar] [CrossRef]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management Priorities for Marine Invasive Species. Science of The Total Environment 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Van Engel, W.A. The Blue Crab and Its Fishery in Chesapeake Bay. Part 1. Reproduction, Early Development, Growth, and Migration. Commercial Fisheries Review 1958, 20, 6–17. [Google Scholar]
- Jivoff, P.R.; Hines, A.H.; Quackenbush, S. Chapter 7: Reproduction and Embryonic Development. In Blue Crab: Callinectes Sapidus. 2007, 181–223.
- Hines, A.H. Allometric Constraints and Variables of Reproductive Effort in Brachyuran Crabs. Mar. Biol. 1982, 69, 309–320. [Google Scholar] [CrossRef]
- Tankersley, R.A.; Wieber, M.G.; Sigala, M.A.; Kachurak, K.A. Migratory Behavior of Ovigerous Blue Crabs Callinectes Sapidus: Evidence for Selective Tidal-Stream Transport. The Biological Bulletin 1998, 195, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.H. Chapter 14: Ecology of Juvenile and Adult Blue Crabs. In Biology of the Blue Crab; Kenney, V.S., Cronin, E., Eds.; Maryland Sea Grant Program: College Park, MD, USA, 2007; pp. 575–665. [Google Scholar]
- Epifanio, C.E. Early Life History of the Blue Crab Callinectes Sapidus: A Review. shre 2019, 38, 1–22. [Google Scholar] [CrossRef]
- Schneider, A.K.; Shields, J.D.; Fabrizio, M.C.; Lipcius, R.N. Spawning History, Fecundity, and Potential Sperm Limitation of Female Blue Crabs in Chesapeake Bay. Fisheries Research 2024, 278, 107094. [Google Scholar] [CrossRef]
- Prager, M.; McConaugha, J.; Jones, C.; Geer, P. Fecundity of Blue Crab, Callinectes Sapidus, in Chesapeake Bay: Biological, Statistical and Management Considerations. Bulletin of Marine Science 1990, 46, 170–179. [Google Scholar]
- Hines, A.H.; Lipcius, R.N.; Haddon, A.M. Population Dynamics and Habitat Partitioning by Size, Sex, and Molt Stage of Blue Crabs Callinectes Sapidus in a Subestuary of Central Chesapeake Bay. Marine Ecology-Progress Series 1987, 36, 55–64. [Google Scholar] [CrossRef]
- Steele, P.; Bert, T.M. Population Ecology of the Blue Crab, Callinectes Sapidus Rathbun. In A Subtropical Estuary: Population Structure, Aspects of Reproduction, and Habitat Partitioning; Florida Marine Research Institute Publication: St. Petersburg, FL, USA, 1994. [Google Scholar]
- Türeli, C.; Yeşilyurt, İ.N.; Nevşat, İ.E. Female Reproductive Pattern of Callinectes Sapidus Rathbun, 1896 (Brachyura: Portunidae) in Iskenderun Bay, Eastern Mediterranean. Zoology in the Middle East 2018, 64, 55–63. [Google Scholar] [CrossRef]
- Marchessaux, G.; Gjoni, V.; Sarà, G. Environmental Drivers of Size-Based Population Structure, Sexual Maturity and Fecundity: A Study of the Invasive Blue Crab Callinectes Sapidus (Rathbun, 1896) in the Mediterranean Sea. PLOS ONE 2023, 18, e0289611. [Google Scholar] [CrossRef]
- Sumer, C.; Teksam, İ.; Karatas, H.; Beyhan, T.; Aydin, C.M. Growth and Reproduction Biology of the Blue Crab, Callinectes sapidus Rathbun, 1896, in the Beymelek Lagoon (Southwestern Coast of Turkey). Turkish Journal of Fisheries and Aquatic Sciences 2013, 13. [Google Scholar] [CrossRef]
- Androulidakis, Y.; Makris, C.; Kombiadou, K.; Krestenitis, Y.; Stefanidou, N.; Antoniadou, C.; Krasakopoulou, E.; Kalatzi, M.-I.; Baltikas, V.; Moustaka-Gouni, M.; et al. Oceanographic Research in the Thermaikos Gulf: A Review over Five Decades. Journal of Marine Science and Engineering 2024, 12, 795. [Google Scholar] [CrossRef]
- Lykousis, V.; Chronis, G. Mechanisms of Sediment Transport and Deposition: Sediment Sequences and Accumulation during the Holocene on the Thermaikos Plateau, the Continental Slope, and Basin (Sporadhes Basin), Northwestern Aegean Sea, Greece. Marine Geology 1989, 87, 15–26. [Google Scholar] [CrossRef]
- Lykousis, V.; Collins, M.B.; Ferentinos, G. Modern Sedimentation in the N.W. Aegean Sea. Marine Geology 1981, 43, 111–130. [Google Scholar] [CrossRef]
- Poulos, S.E.; Chronis, G.T.; Collins, M.B.; Lykousis, V. Thermaikos Gulf Coastal System, NW Aegean Sea: An Overview of Water/Sediment Fluxes in Relation to Air–Land–Ocean Interactions and Human Activities. Journal of Marine Systems 2000, 25, 47–76. [Google Scholar] [CrossRef]
- Olmi, E.J., III; Bishop, J.M. Variations in Total Width-Weight Relationships of Blue Crabs, Callinectes Sapidus, in Relation to Sex, Maturity, Molt Stage, and Carapace Form. Journal of Crustacean Biology 1983, 3, 575–581. [Google Scholar] [CrossRef]
- Cadman, L.R.; Weinstein, M.P. Size-Weight Relationships of Postecdysial Juvenile Blue Crabs ( Callinectes Sapidus Rathbun) from the Lower Chesapeake Bay. Journal of Crustacean Biology 1985, 5, 306–310. [Google Scholar] [CrossRef]
- Rugolo, L.J.; Knotts, K.S.; Lange, A.M.; Crecco, V.A. Stock Assessment of Chesapeake Bay Blue Crab (Callinectes Sapidus Rathbun). J. Shellfish Res. 1998, 17, 1321–1345. [Google Scholar]
- Hard, W.L. Ovarian Growth and Ovulation in the Mature Blue Crab, Callinectes Sapidus Rathbun; Chesapeake Biological Laboratory: Solomons, ML, USA, 1942. [Google Scholar]
- Sainte-Marie, B. Reproductive Cycle and Fecundity of Primiparous and Multiparous Female Snow Crab, Chionoecetes Opilio, in the Northwest Gulf of Saint Lawrence. Can. J. Fish. Aquat. Sci. 1993, 50, 2147–2156. [Google Scholar] [CrossRef]
- Kemberling, A.A.; Darnell, M.Z. Distribution, Relative Abundance, and Reproductive Output of Spawning Female Blue Crab (Callinectes Sapidus) in Offshore Waters of the Gulf of Mexico. FB 2020, 118, 346–358. [Google Scholar] [CrossRef]
- Churchill, E.P., Jr. Life History of the Blue Crab. Bulletin of the United States Bureau of Fisheries 1919, 36, 95–128. [Google Scholar]
- Lipcius, R. Blue-Crab Population-Dynamics In Chesapeake Bay - Variation In Abundance (York River, 1972-1988) And Stock-Recruit Functions. Bulletin of Marine Science 1990, 46, 180–194. [Google Scholar]
- Forward, R.B.; Cohen, J.H. Factors Affecting the Circatidal Rhythm in Vertical Swimming of Ovigerous Blue Crabs, Callinectes Sapidus, Involved in the Spawning Migration. Journal of Experimental Marine Biology and Ecology 2004, 299, 255–266. [Google Scholar] [CrossRef]
- Aguilar, R.; Hines, A.H.; Wolcott, T.G.; Wolcott, D.L.; Kramer, M.A.; Lipcius, R.N. The Timing and Route of Movement and Migration of Post-Copulatory Female Blue Crabs, Callinectes Sapidus Rathbun, from the Upper Chesapeake Bay. Journal of Experimental Marine Biology and Ecology 2005, 319, 117–128. [Google Scholar] [CrossRef]
- Turner, H.V.; Wolcott, D.L.; Wolcott, T.G.; Hines, A.H. Post-Mating Behavior, Intramolt Growth, and Onset of Migration to Chesapeake Bay Spawning Grounds by Adult Female Blue Crabs, Callinectes Sapidus Rathbun. Journal of Experimental Marine Biology and Ecology 2003, 295, 107–130. [Google Scholar] [CrossRef]
- Carr, S.D.; Tankersley, R.A.; Hench, J.L.; Forward, R.B.; Luettich, R.A. Movement Patterns and Trajectories of Ovigerous Blue Crabs Callinectes Sapidus during the Spawning Migration. Estuarine, Coastal and Shelf Science 2004, 60, 567–579. [Google Scholar] [CrossRef]
- Lipcius, R.N.; Stockhausen, W.T. Concurrent Decline of the Spawning Stock, Recruitment, Larval Abundance, and Size of the Blue Crab Callinectes Sapidus in Chesapeake Bay. Marine Ecology-Progress Series 2002, 226, 45–61. [Google Scholar] [CrossRef]
- Epifanio, C.E. Larval Biology. In Biology of the Blue Crab,edited by Kenney, V. S. and E. Cronin.; College Park, MD: Maryland Sea Grant, 2007; pp. 513–533. [Google Scholar]
- Hines, A.H. Ecology of Juvenile and Adult Blue Crabs: Summary of Discussion of Research Themes and Directions. Bulletin of Marine Science. 2003, 72, 423–433. [Google Scholar]
- Millikin, M.R.; Williams, A.B. Synopsis of Biological Data on the Blue Crab, Callinectes Sapidus Rathbun; National Oceanic and Atmospheric Administration, National Marine Fisheries Service, 1984. [Google Scholar]
- Darnell, M.Z.; Kemberling, A.A. Large-Scale Movements of Postcopulatory Female Blue Crabs Callinectes Sapidus in Tidal and Nontidal Estuaries of North Carolina. Transactions of the American Fisheries Society 2018, 147, 716–728. [Google Scholar] [CrossRef]
- Darnell, M.; Rittschof, D.; Darnell, K.; McDowell, R. Lifetime Reproductive Potential of Female Blue Crabs Callinectes Sapidus in North Carolina, USA. Mar. Ecol. Prog. Ser. 2009, 394, 153–163. [Google Scholar] [CrossRef]
- Provenzano, A.J.; McConaugha, J.R.; Philips, K.B.; Johnson, D.F.; Clark, J. Vertical Distribution of First Stage Larvae of the Blue Crab, Callinectes Sapidus, at the Mouth of Chesapeake Bay. Estuarine, Coastal and Shelf Science 1983, 16, 489–499. [Google Scholar] [CrossRef]
- Epifanio, C.E.; Valenti, C.C.; Pembroke, A.E. Dispersal and Recruitment of Blue Crab Larvae in Delaware Bay, U.S.A. Estuarine, Coastal and Shelf Science 1984, 18, 1–12. [Google Scholar] [CrossRef]
- Lipcius, R.; Engel, W.A.V. Blue-Crab Population-Dynamics In Chesapeake Bay - Variation In Abundance (York River, 1972-1988) And Stock-Recruit Functions. Bulletin of Marine Science 1990, 46, 180–194. [Google Scholar]
- Epifanio, C.E.; Garvine, R.W. Larval Transport on the Atlantic Continental Shelf of North America: A Review. Estuarine, Coastal and Shelf Science 2001, 52, 51–77. [Google Scholar] [CrossRef]
- Yesilyurt, I.N.; Tureli, C. Settlement of Blue Crab (Callinectes Sapidus) Megalopae in the North Shore of Yumurtalik Cove (Turkey). Fresenius Environmental Bulletin 2016, 25, 1007–1018. [Google Scholar]
- Gil-Fernández, A.; Rodilla, M.; Prado, P.; Falco, S. Early Life Stages of the Invasive Atlantic Blue Crab Callinectes Sapidus in the Western Mediterranean Sea. Estuarine, Coastal and Shelf Science 2024, 296, 108593. [Google Scholar] [CrossRef]
- Fitz, H.C.; Wiegert, R.G. Local Population Dynamics of Estuarine Blue Crabs: Abundance, Recruitment and Loss. Marine Ecology Progress Series 1992, 87, 23–40. [Google Scholar] [CrossRef]
- Mancinelli, G.; Carrozzo, L.; Costantini, M.L.; Rossi, L.; Marini, G.; Pinna, M. Occurrence of the Atlantic Blue Crab Callinectes Sapidus Rathbun, 1896 in Two Mediterranean Coastal Habitats: Temporary Visitor or Permanent Resident? Estuarine, Coastal and Shelf Science 2013, 135, 46–56. [Google Scholar] [CrossRef]
- Carrozzo, L.; Potenza, L.; Carlino, P.; Costantini, M.L.; Rossi, L.; Mancinelli, G. Seasonal Abundance and Trophic Position of the Atlantic Blue Crab Callinectes Sapidus Rathbun 1896 in a Mediterranean Coastal Habitat. Rend. Fis. Acc. Lincei 2014, 25, 201–208. [Google Scholar] [CrossRef]
- Lycett, K.A.; Shields, J.D.; Chung, J.S.; Pitula, J.S. Population Structure of the Blue Crab Callinectes Sapidus in the Maryland Coastal Bays. shre 2020, 39, 699–713. [Google Scholar] [CrossRef]
- Jivoff, P. A Review of Male Mating Success in the Blue Crab, Callinectes Sapidus, in Reference to the Potential for Fisheries-Induced Sperm Limitation. Bulletin of Marine Science. 2003, 72, 273–286. [Google Scholar]
- Gleeson, R.A. Pheromone Communication in the Reproductive Behavior of the Blue Crab, Callinectes Sapidus †. Marine Behaviour and Physiology 1980, 7, 119–134. [Google Scholar] [CrossRef]
- Jivoff, P.; Hines, A.H. Female Behaviour, Sexual Competition and Mate Guarding in the Blue Crab, Callinectes Sapidus. Anim Behav 1998, 55, 589–603. [Google Scholar] [CrossRef]
- Rains, S.A.M.; Wilberg, M.J.; Miller, T.J. Sex Ratios and Average Sperm per Female Blue Crab Callinectes Sapidus in Six Tributaries of Chesapeake Bay. Marine and Coastal Fisheries 2016, 8, 492–501. [Google Scholar] [CrossRef]
- Taylor, D.L.; Fehon, M.M. Blue Crab (Callinectes Sapidus) Population Structure in Southern New England Tidal Rivers: Patterns of Shallow-Water, Unvegetated Habitat Use and Quality. Estuaries and Coasts 2021, 44, 1320–1343. [Google Scholar] [CrossRef] [PubMed]
- Hart, H.R.; Crowley, C.E.; Walters, E.A. Blue Crab Spawning and Recruitment in Two Gulf Coast and Two Atlantic Estuaries in Florida. Marine and Coastal Fisheries 2021, 13, 113–130. [Google Scholar] [CrossRef]
- Pereira, M.J.; Branco, J.O.; Christoffersen, M.L.; Junior, F.F.; Fracasso, H.A.A.; Pinheiro, T.C. Population Biology of Callinectes Danae and Callinectes Sapidus (Crustacea: Brachyura: Portunidae) in the South-Western Atlantic. Journal of the Marine Biological Association of the United Kingdom 2009, 89, 1341–1351. [Google Scholar] [CrossRef]
- Severino-Rodrigues, E.; Musiello-Fernandes, J.; Mour, Á.A.S.; Branco, G.M.P.; Canéo, V.O.C. Fecundity, Reproductive Seasonality and Maturation Size of Callinectes Sapidus Females (Decapoda: Portunidae) in the Southeast Coast of Brazil. Revista de Biología Tropical 2013, 61, 595–602. [Google Scholar] [CrossRef]
- Fisher, M.R. Effect of Temperature and Salinity on Size at Maturity of Female Blue Crabs. Transactions of the American Fisheries Society 1999, 128, 499–506. [Google Scholar] [CrossRef]
- Carvalho-Saucedo, L.; Ramírez-Santiago, C.; Pérez, C. Histological Description of Gonadal Development of Females and Males of Callinectes Sapidus Rathbun, 1896 (Decapoda: Portunidae). jzoo 2015, 32, 188–194. [Google Scholar] [CrossRef]
- Harris, R. 1982.
- Lipcius, R.N.; Eggleston, D.B.; Heck, K.L.; Seitz, R.D.; Montfrans, J.V. Post-Settlement Abundance, Survival, and Growth of Postlarvae and Young Juvenile Blue Crabs in Nursery Habitats. In Biology of the Blue Crab; Kenney, V.S., Cronin, E., Eds.; Maryland Sea Grant Program: College Park, MD, USA, 2007; pp. 535–565. [Google Scholar]
- Archambault, J.A.; Wenner, E.L.; Whitaker, J.D. Life History and Abundance of Blue Crab, Callinectes Sapidus Rathbun, at Charleston Harbor, South Carolina. Bulletin of Marine Science 1990, 46, 145–158. [Google Scholar]
- Tagatz, M.E. Biology of the Blue Crab, Callinectes Sapidus Rathbun, in the St. Johns River, Florida. United States Fish and Wildlife Service Fishery Bulletin 1968, 67, 17–33. [Google Scholar]
- Hines, A.H.; Johnson, E.G.; Darnell, M.Z.; Rittschof, D.; Miller, T.J.; Bauer, L.J.; Rodgers, P.; Aguilar, R. Predicting Effects of Climate Change on Blue Crabs in Chesapeake Bay. In Proceedings of the Biology and Management of Exploited Crab Populations under Climate Change; Alaska Sea Grant, University of Alaska Fairbanks, April 27 2011; pp. 109–127. [Google Scholar]
- Epifanio, C.E.; Masse, A.K.; Garvine, R.W. Transport of Blue Crab Larvae by Surface Currents off Delaware Bay, USA. Marine Ecology Progress Series 1989, 54, 35–41. [Google Scholar] [CrossRef]
- Lipcius, R.; Stockhausen, W.T.; Seitz, R.; Geer, P.J. Spatial Dynamics and Value of a Marine Protected Area and Corridor for the Blue Crab Spawning Stock in Chesapeake Bay. Bulletin of Marine Science 2003, 72, 453–469. [Google Scholar]
- Gelpi, C.; Condrey, R. e; Fleeger, J. w; Dubois, S.; Fleeger, J. w; Dubois, S. Discovery, Evaluation, and Implications of Blue Crab, Callinectes Sapidus, Spawning, Hatching, and Foraging Grounds in Federal (Us) Waters Offshore of Louisiana. Bulletin Of Marine Science 2009, 85, 203–222. [Google Scholar]
- Gelpi, C.; Fry, B.; Condrey, R.; Fleeger, J.; Dubois, S. Using δ13C and δ15N to Determine the Migratory History of Offshore Louisiana Blue Crab Spawning Stocks. Mar. Ecol. Prog. Ser. 2013, 494, 205–218. [Google Scholar] [CrossRef]
- Anderson, J.; Olsen, Z.; Glen Sutton, T.W.; Gelpi, C.; Topping, D. Environmental Drivers of the Spatial and Temporal Distribution of Spawning Blue Crabs Callinectes Sapidus in the Western Gulf of Mexico. North American Journal of Fisheries Management 2017, 37, 920–934. [Google Scholar] [CrossRef]
- Ogburn, M.B.; Habegger, L.C. Reproductive Status of Callinectes Sapidus as an Indicator of Spawning Habitat in the South Atlantic Bight, USA. Estuaries and Coasts 2015, 38, 2059–2069. [Google Scholar] [CrossRef]
- Darnell, M.Z.; Wolcott, T.G.; Rittschof, D. Environmental and Endogenous Control of Selective Tidal-Stream Transport Behavior during Blue Crab Callinectes Sapidus Spawning Migrations. Mar Biol 2012, 159, 621–631. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Millstein, E.; Plaia, G. Timing and Route of Migration of Mature Female Blue Crabs in a Tidal Estuary. Biology Letters 2015, 11, 20140936. [Google Scholar] [CrossRef]
- Epifanio, C.E.; Cohen, J.H. Behavioral Adaptations in Larvae of Brachyuran Crabs: A Review. Journal of Experimental Marine Biology and Ecology 2016, 482, 85–105. [Google Scholar] [CrossRef]
- Hines, A.H.; Wolcott, T.G.; González-Gurriarán, E.; González-Escalante, J.L.; Freire, J. Movement Patterns and Migrations in Crabs: Telemetry of Juvenile and Adult Behaviour in Callinectes Sapidus and Maja Squinado. Journal of the Marine Biological Association of the United Kingdom 1995, 75, 27–42. [Google Scholar] [CrossRef]
- Van Engel, W. Factors Affecting The Distribution And Abundance Of The Blue Crab In Chesapeake Bay; Pennsylvania Academy of Science, 1987. [Google Scholar]
- Truitt, R.V. The Blue in Our Water Resources and Their Eonscrvstlon. Chesapeake Biol. Lab. Corttríb. 1939, 27, 10–38. [Google Scholar]
- Hsueh, P.-W.; McClintock, J.B.; Hopkins, T.S. Population Dynamics and Life History Characteristics of the Blue Crabs Callinectes Similis and C. Sapidus in Bay Environments of the Northern Gulf of Mexico. Marine Ecology 1993, 14, 239–257. [Google Scholar] [CrossRef]
- Graham, D.J.; Perry, H.; Biesiot, P.; Fulford, R. Fecundity and Egg Diameter of Primiparous and Multiparous Blue Crab Callinectes Sapidus (Brachyura: Portunidae) in Mississippi Waters. Journal of Crustacean Biology 2012, 32, 49–56. [Google Scholar] [CrossRef]
- Dickinson, G.H.; Rittschof, D.; Latanich, C. Spawning Biology of the Blue Crab, Callinectes Sapidus, in North Carolina. Bulletin of Marine Science 2006, 79, 273–285. [Google Scholar]
- Darnell, M.Z.; Rittschof, D.; Forward, R.B. Endogenous Swimming Rhythms Underlying the Spawning Migration of the Blue Crab, Callinectes Sapidus: Ontogeny and Variation with Ambient Tidal Regime. Mar Biol 2010, 157, 2415–2425. [Google Scholar] [CrossRef]
- Tang, Q. Modification of the Ricker Stock Recruitment Model to Account for Environmentally Induced Variation in Recruitment with Particular Reference to the Blue Crab Fishery in Chesapeake Bay. Fisheries Research 1985, 3, 13–21. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Lipcius, R.N. Population Dynamics and Fisheries. In Biology of the Blue Crab,edited by Kenney, V. S. and E. Cronin.; College Park, MD: Maryland Sea Grant, 2007; pp. 711–755. [Google Scholar]
- Willan, R.C.; Russell, B.C.; Murfet, N.B.; Moore, K.L.; McEnnulty, F.R.; Horner, S.K.; Hewitt, C.L.; Dally, G.M.; Campbell, M.L.; Bourke, S.T. Outbreak of Mytilopsis Sallei (Récluz, 1849) (Bivalvia: Dreissenidae) in Australia. Molluscan Research 2000, 20, 25–30. [Google Scholar] [CrossRef]
- Bax, N.; Carlton, J.T.; Mathews-Amos, A.; Haedrich, R.L.; Howarth, F.G.; Purcell, J.E.; Rieser, A.; Gray, A. The Control of Biological Invasions in the World’s Oceans. Conservation Biology 2001, 15, 1234–1246. [Google Scholar] [CrossRef]
- Lampert, A. Optimizing Strategies for Slowing the Spread of Invasive Species. PLOS Computational Biology 2024, 20, e1011996. [Google Scholar] [CrossRef]
Figure 1.
Map of the of study area in the Thermaikos Gulf, NW Aegean Sea, NE Mediterranean Sea (A: inner part; B: outer part) indicating monthly (yellow), seasonal (green) and experimental (blue) sampling stations of Callinectes sapidus in the inner gulf.
Figure 1.
Map of the of study area in the Thermaikos Gulf, NW Aegean Sea, NE Mediterranean Sea (A: inner part; B: outer part) indicating monthly (yellow), seasonal (green) and experimental (blue) sampling stations of Callinectes sapidus in the inner gulf.
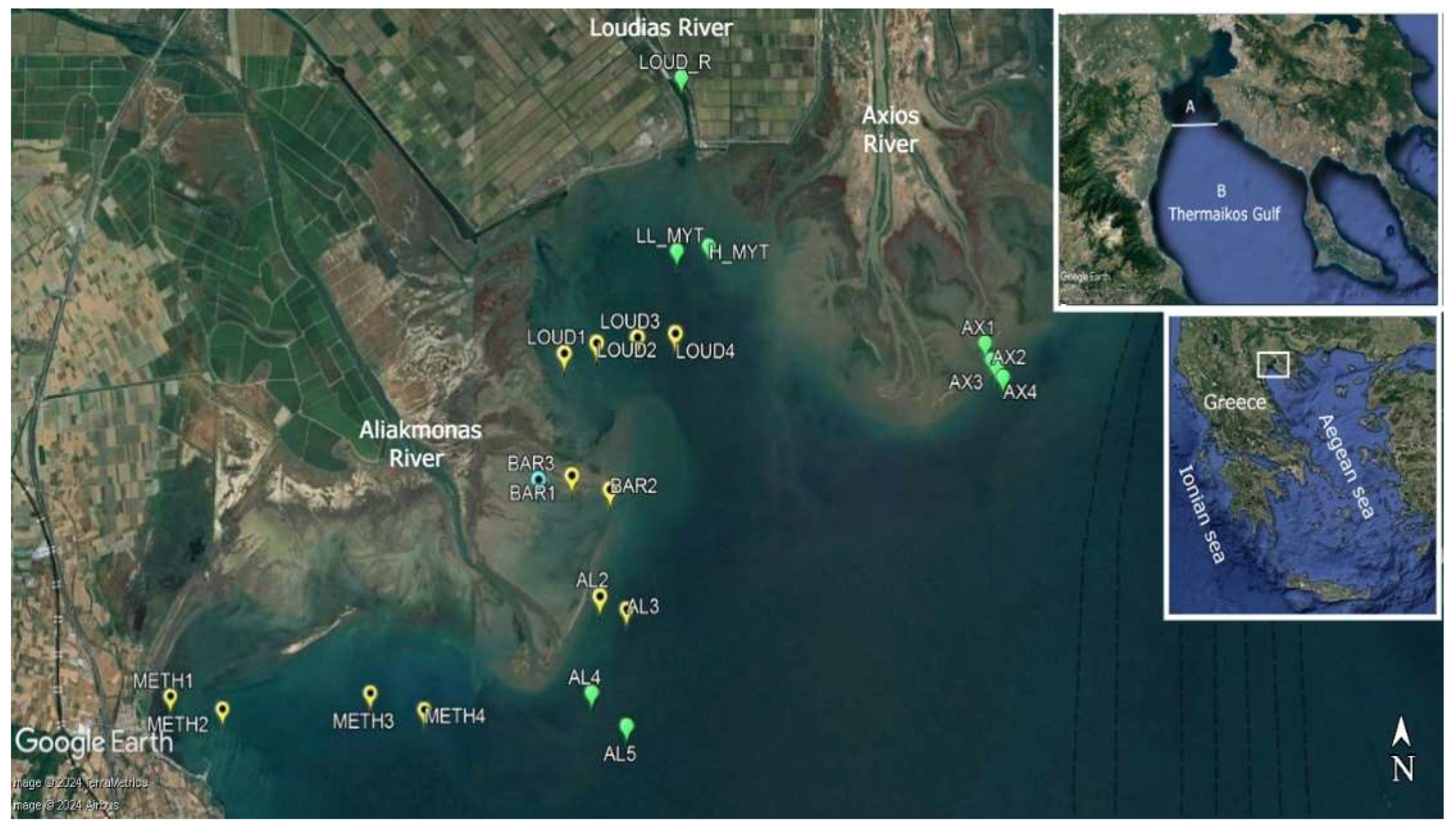
Figure 2.
Bottom temperature (0C) in three sampling stations (LOUD 1, METH 1, BAR 1) in the inner Thermaikos Gulf during the survey period (December 2020-November 2022).
Figure 2.
Bottom temperature (0C) in three sampling stations (LOUD 1, METH 1, BAR 1) in the inner Thermaikos Gulf during the survey period (December 2020-November 2022).
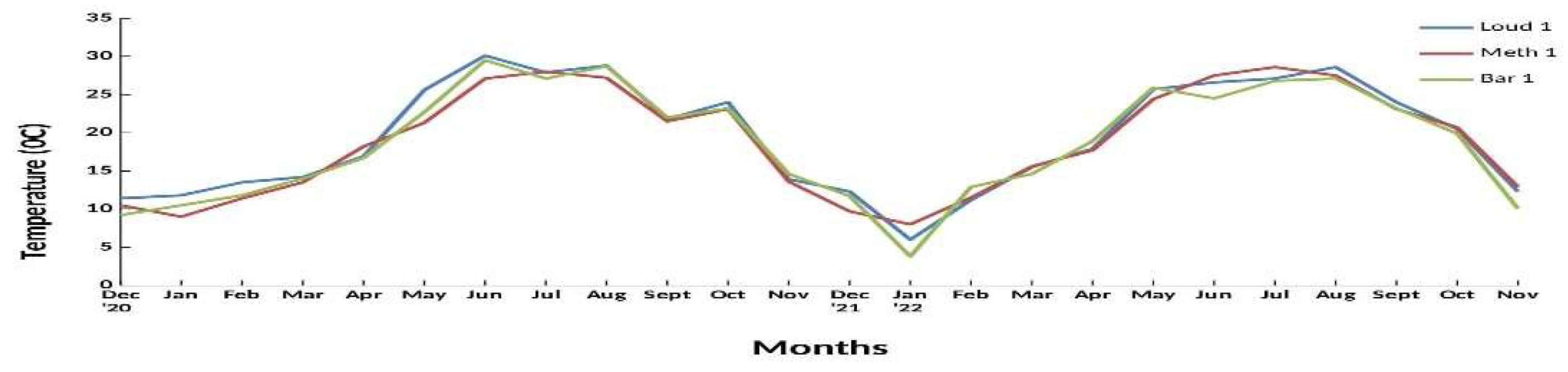
Figure 3.
Monthly sex ratio of the blue crab C. sapidus during the first (December 2020-November 2021) and the second year (December 2020-November 2022) in the inner Thermaikos Gulf.
Figure 3.
Monthly sex ratio of the blue crab C. sapidus during the first (December 2020-November 2021) and the second year (December 2020-November 2022) in the inner Thermaikos Gulf.
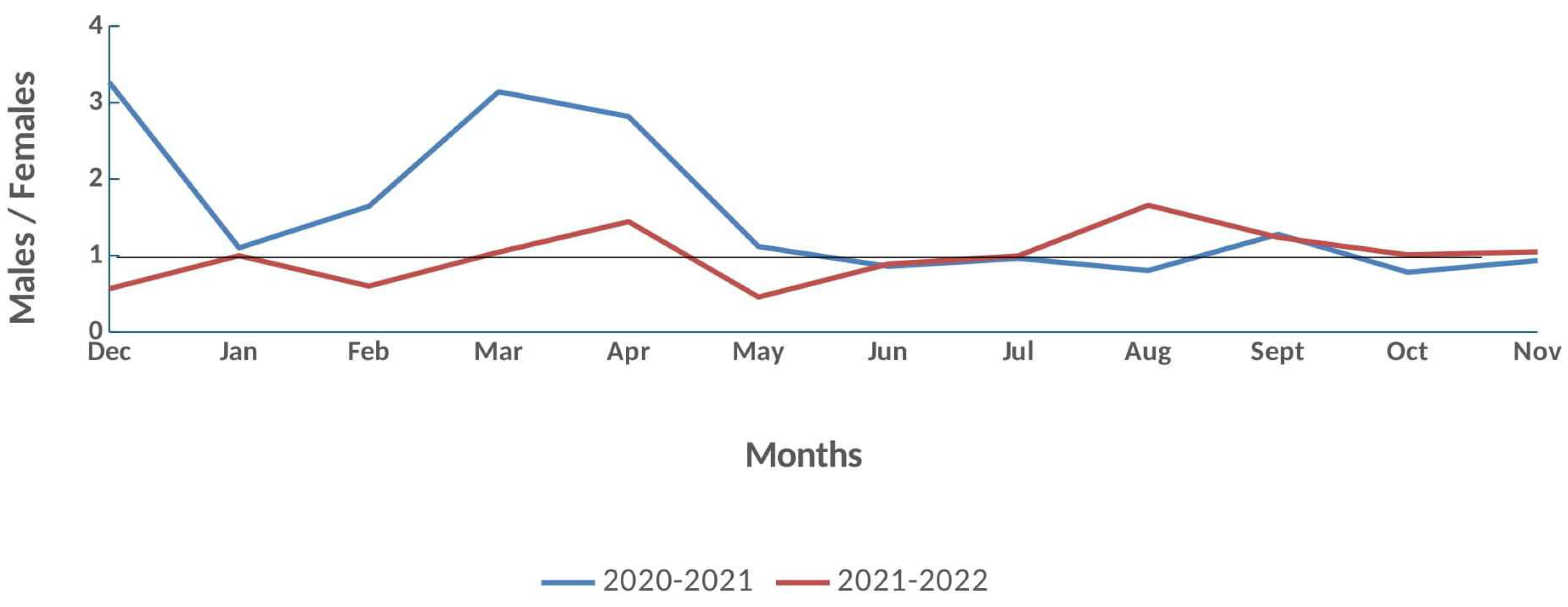
Figure 4.
Monthly distribution of female C. sapidus during the first (December 2020-November 2021) and the second year (December 2020-November 2022) in the inner Thermaikos Gulf.
Figure 4.
Monthly distribution of female C. sapidus during the first (December 2020-November 2021) and the second year (December 2020-November 2022) in the inner Thermaikos Gulf.
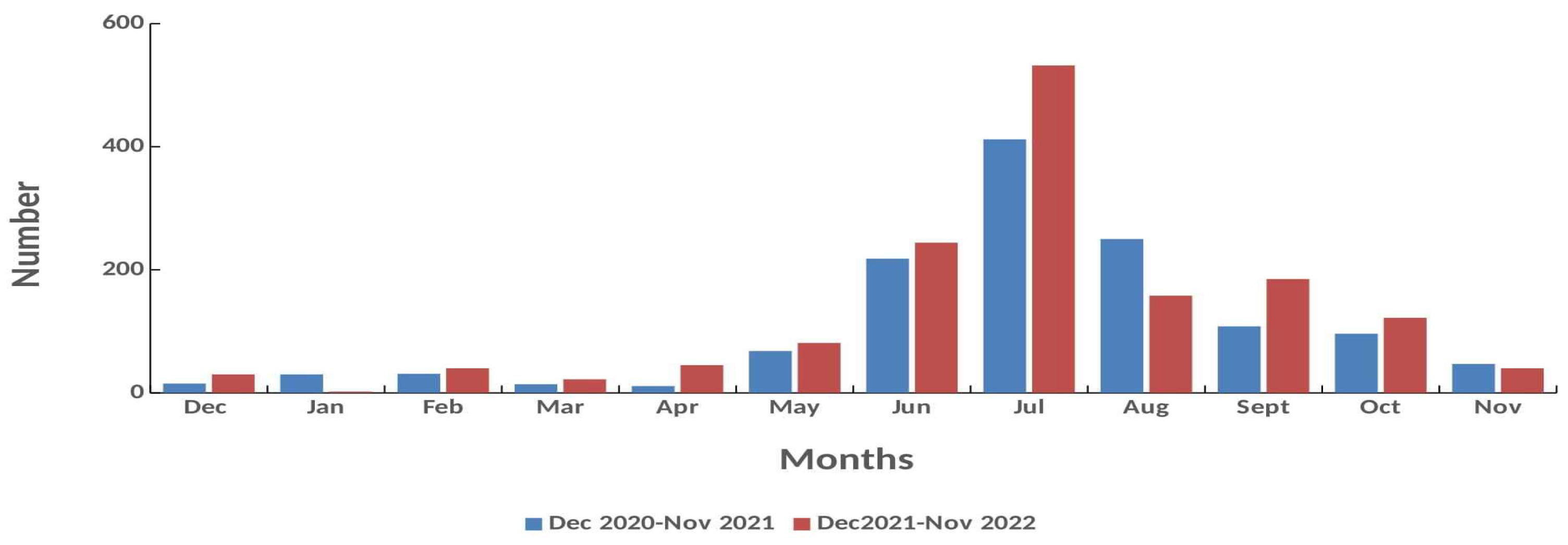
Figure 5.
Size - frequency distributions of mature and immature (including juveniles) females C. sapidus during the survey (December 2020- November 2022) in the inner Thermaikos Gulf.
Figure 5.
Size - frequency distributions of mature and immature (including juveniles) females C. sapidus during the survey (December 2020- November 2022) in the inner Thermaikos Gulf.
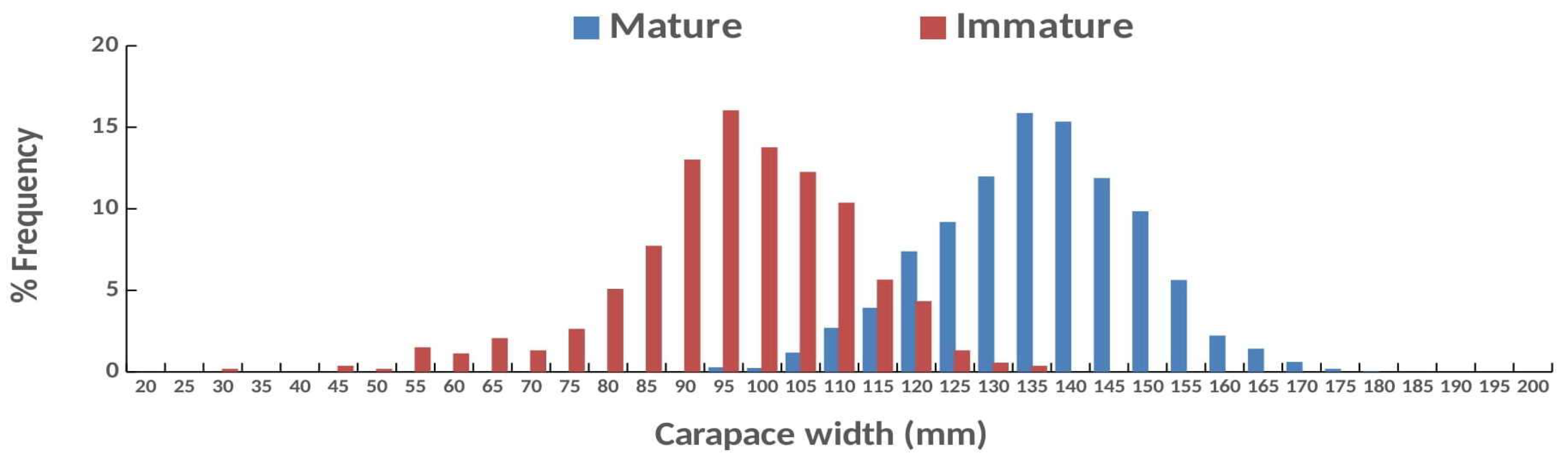
Figure 6.
a,b Frequency (%) of ovarian stages in female C. sapidus in the inner Thermaikos Gulf (December 2020 – November 2021 (N=666) (above) and December 2021- November 2022 (N=616) (below).
Figure 6.
a,b Frequency (%) of ovarian stages in female C. sapidus in the inner Thermaikos Gulf (December 2020 – November 2021 (N=666) (above) and December 2021- November 2022 (N=616) (below).
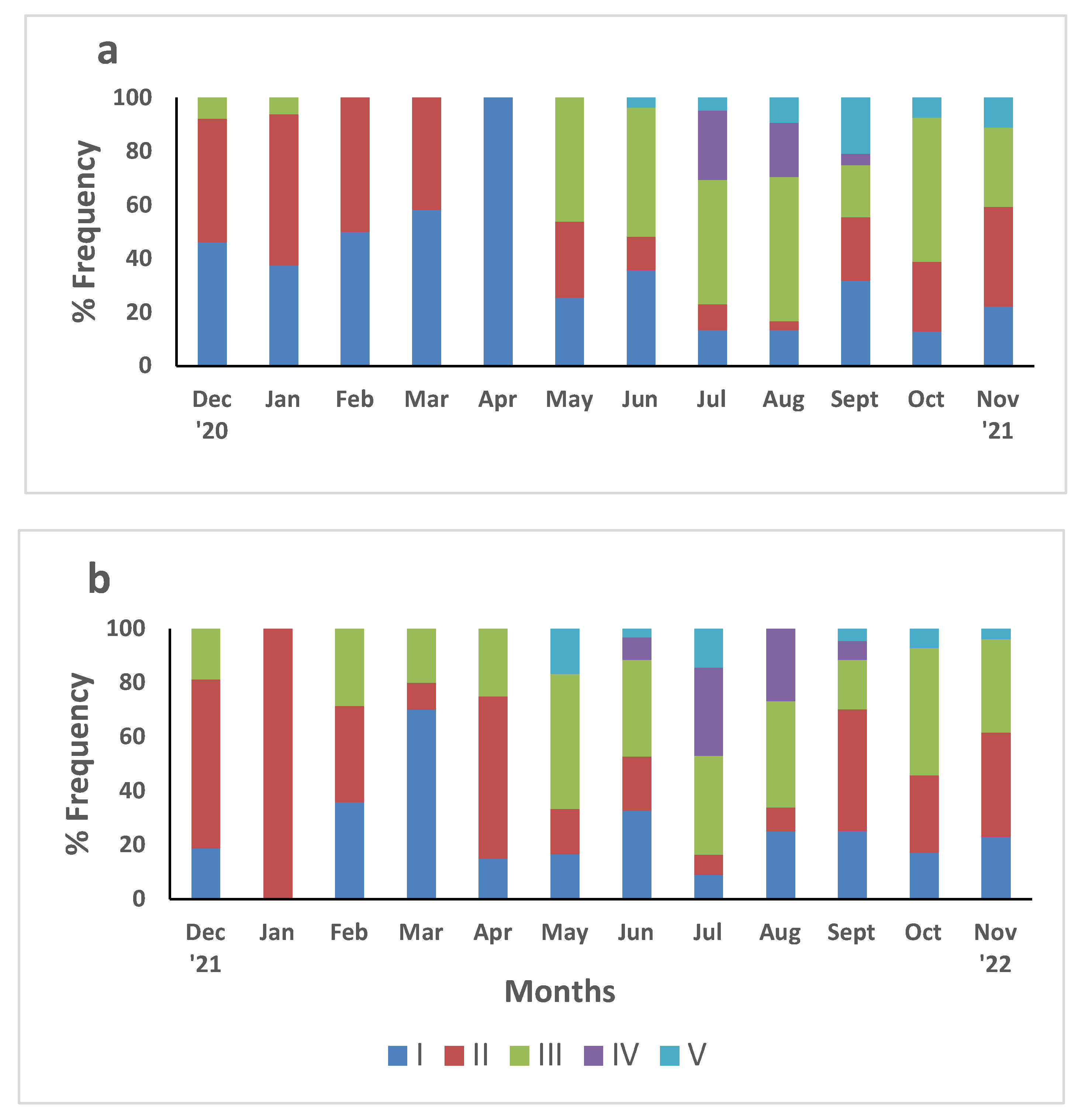
Figure 7.
Ovarian developmental stages of female Callinectes sapidus as determined by microscopic examination of histological preparations of gonads on a monthly basis (A) Undifferentiated germ cells; Stage 1 ov , (B) Nucleus remains large with more than one nucleoid and appearance of a thin basement membrane, nuclear gap (arrow) in early oocytes; Stage 2ov , (C) Development of Immature oocytes with accumulation of granules and a thin basement membrane (arrow); stage 3ov, (D) Mature oocytes (Arrow) in the mature gonad; Stage 4 ov , (E) Process of resorption of immature oocytes with presence of phagocytes (arrow); Stage 5ov. (n: nucleus, gv1: granulosa cells, gv2: nutrients) Hematoxylin-eosin staining.
Figure 7.
Ovarian developmental stages of female Callinectes sapidus as determined by microscopic examination of histological preparations of gonads on a monthly basis (A) Undifferentiated germ cells; Stage 1 ov , (B) Nucleus remains large with more than one nucleoid and appearance of a thin basement membrane, nuclear gap (arrow) in early oocytes; Stage 2ov , (C) Development of Immature oocytes with accumulation of granules and a thin basement membrane (arrow); stage 3ov, (D) Mature oocytes (Arrow) in the mature gonad; Stage 4 ov , (E) Process of resorption of immature oocytes with presence of phagocytes (arrow); Stage 5ov. (n: nucleus, gv1: granulosa cells, gv2: nutrients) Hematoxylin-eosin staining.
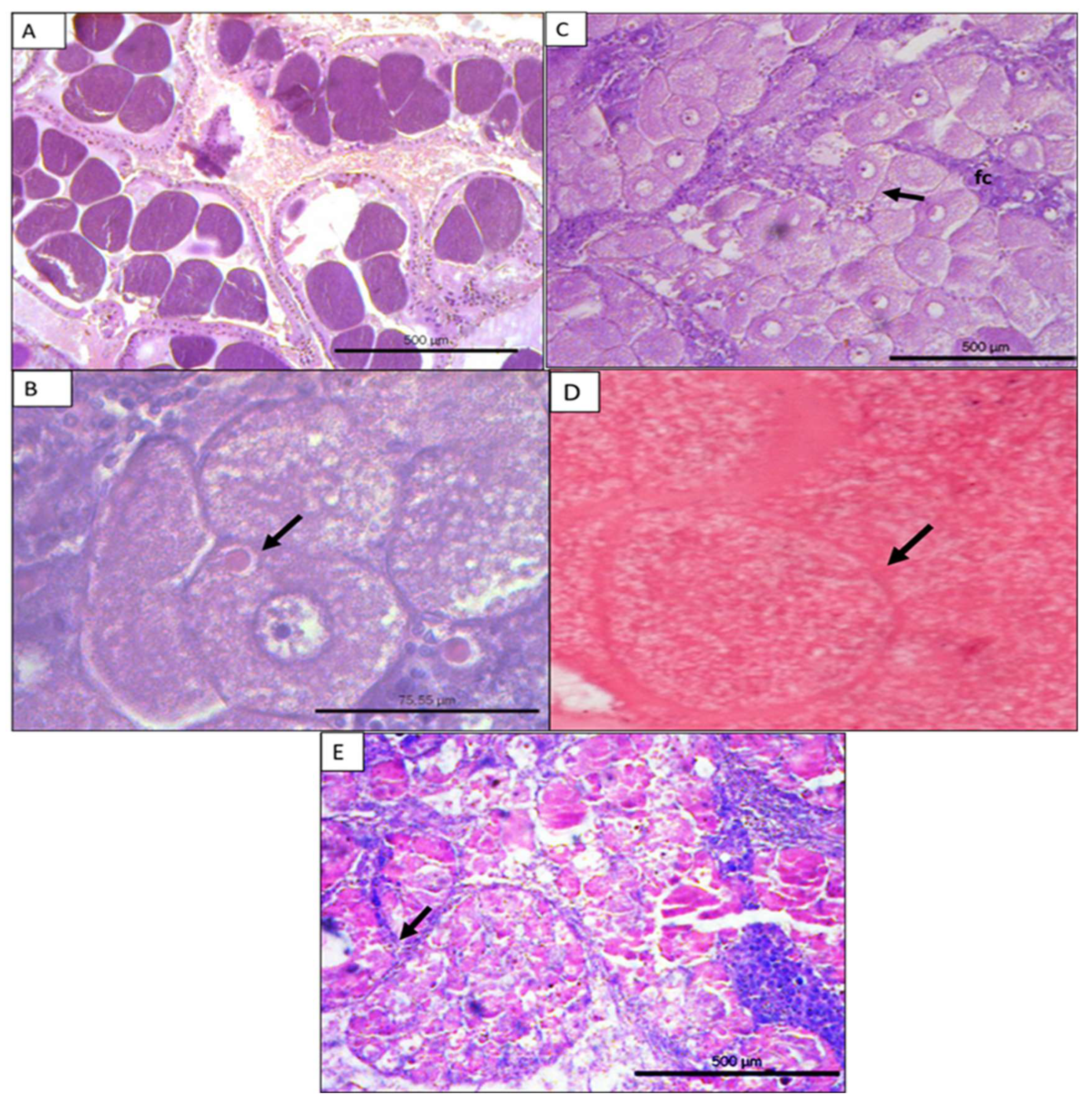
Figure 8.
Monthly distribution of ovigerous females C. sapidus in the inner Thermaikos Gulf during the first (December 2020 – November 2021 and the second year (December 2021- November 2022 (below).
Figure 8.
Monthly distribution of ovigerous females C. sapidus in the inner Thermaikos Gulf during the first (December 2020 – November 2021 and the second year (December 2021- November 2022 (below).
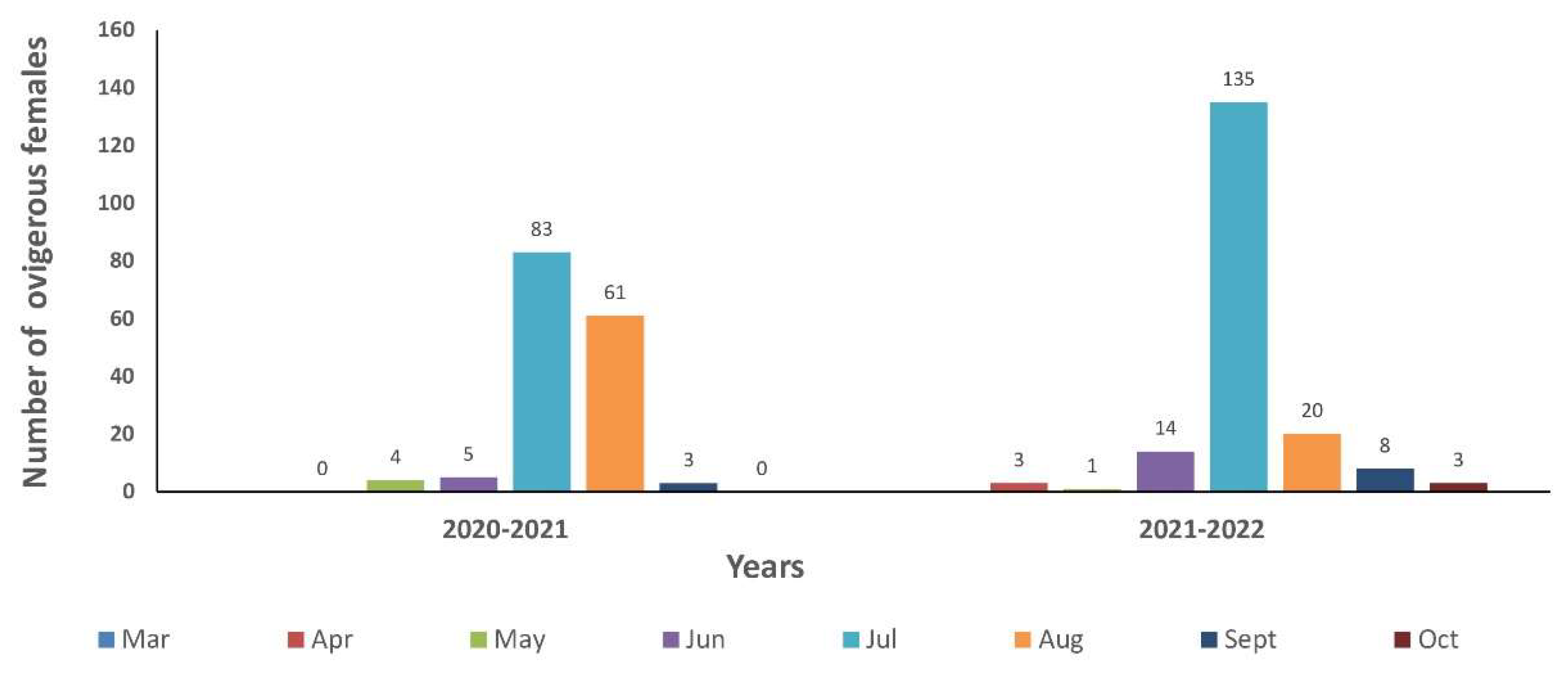
Figure 9.
a,b. Spatial distribution of ovigerous females in the inner Therrmaikos Gulf (December 2020 – November 2021)(above) and the second year (December 2021- November 2022 (below).
Figure 9.
a,b. Spatial distribution of ovigerous females in the inner Therrmaikos Gulf (December 2020 – November 2021)(above) and the second year (December 2021- November 2022 (below).
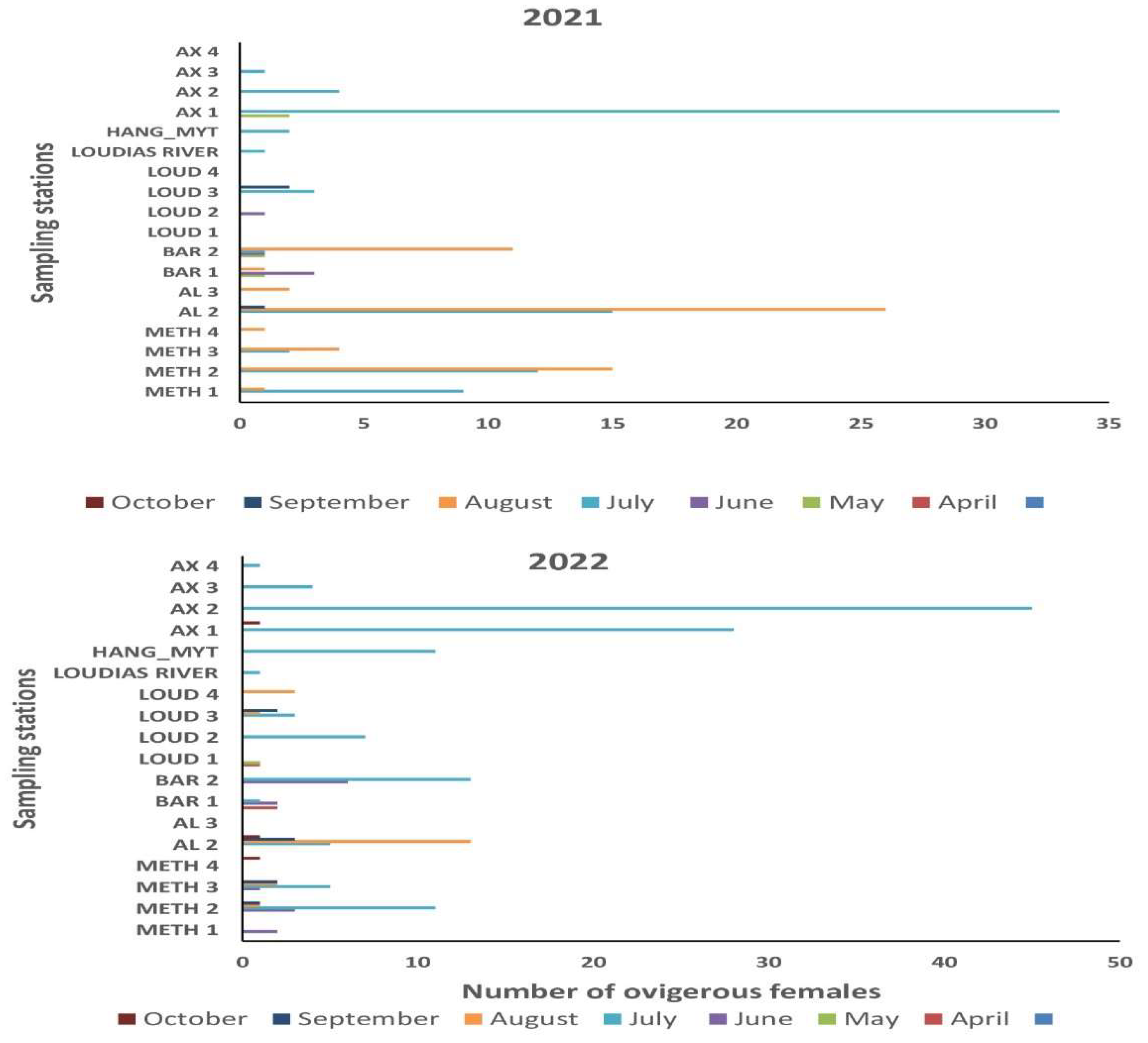
Figure 10.
Bathymetrical distribution of ovigerous females and ovigerous with eggs at developmental stage 5 during the period of survey in the inner Thermaikos Gulf.
Figure 10.
Bathymetrical distribution of ovigerous females and ovigerous with eggs at developmental stage 5 during the period of survey in the inner Thermaikos Gulf.
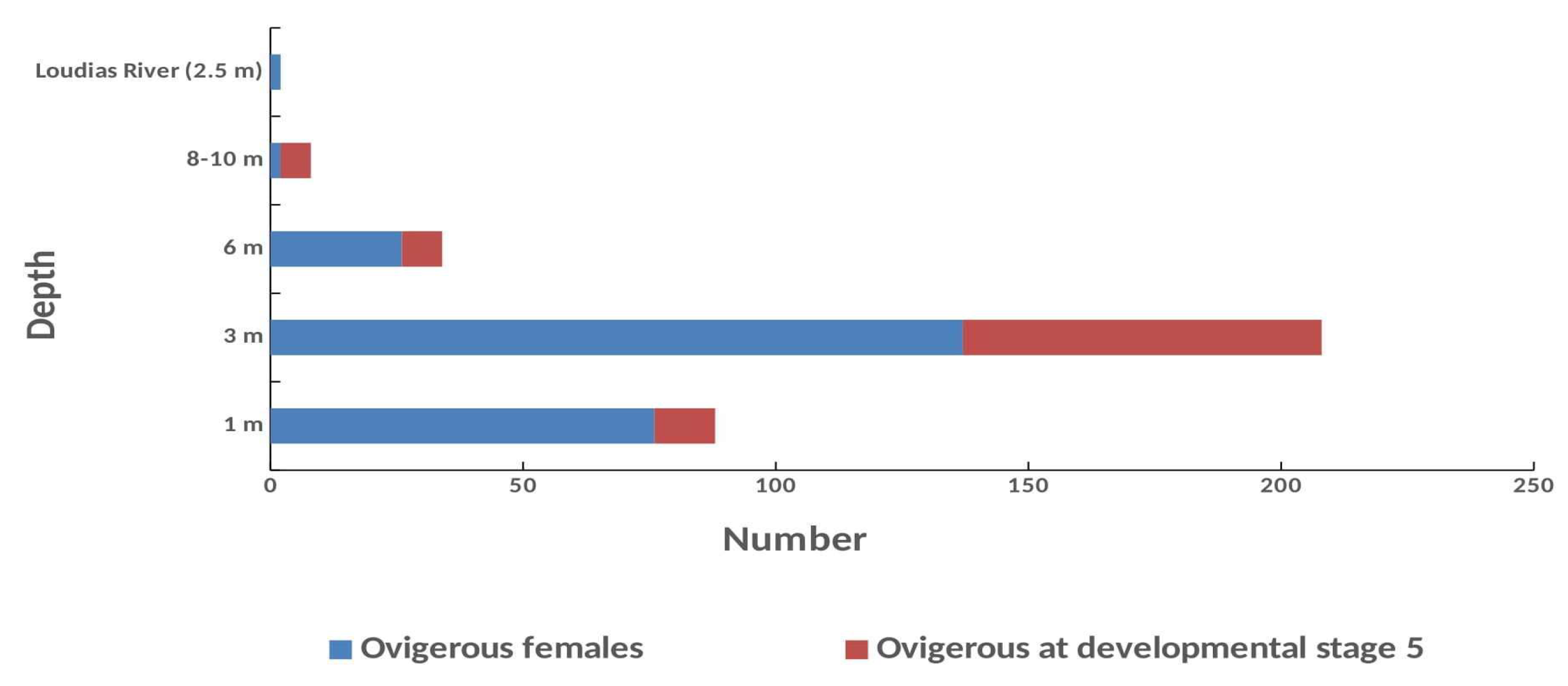
Figure 11.
The mean size at maturity of females C. sapidus in the Thermaikos Gulf during the period of study (2020-2022) using the Frequentist regression.
Figure 11.
The mean size at maturity of females C. sapidus in the Thermaikos Gulf during the period of study (2020-2022) using the Frequentist regression.
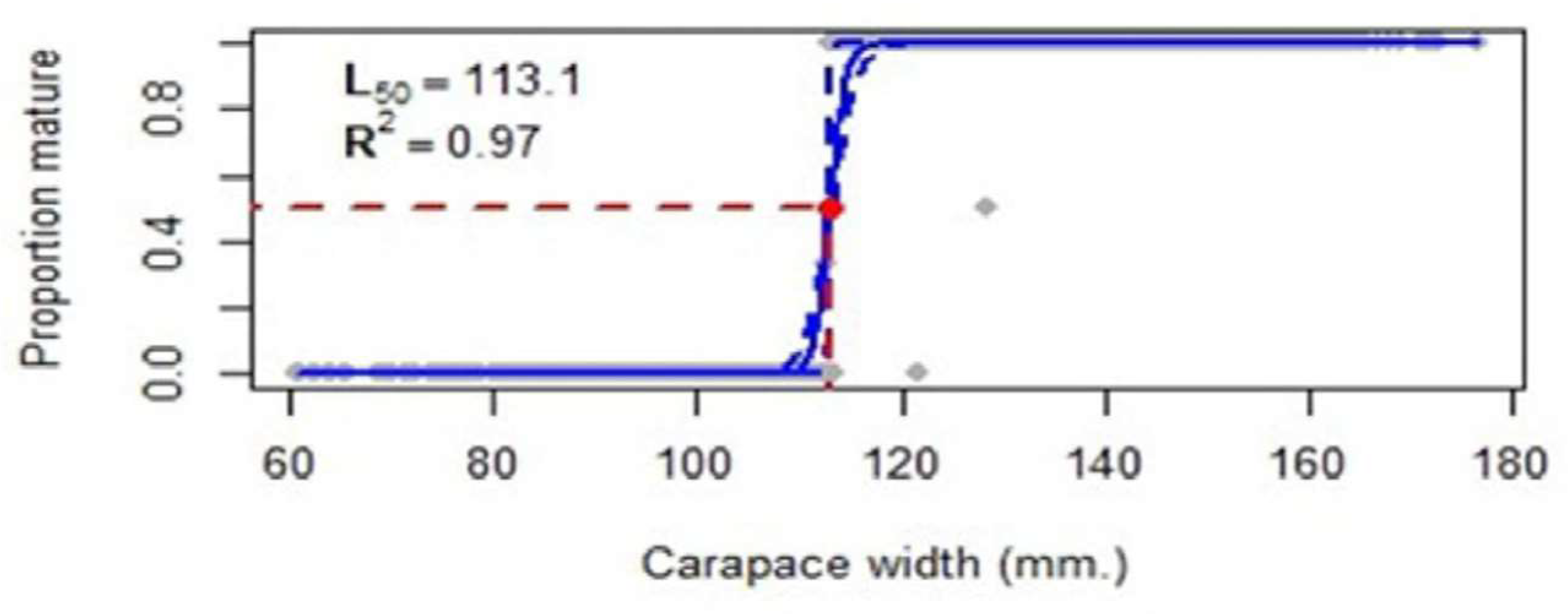
Figure 12.
Bar plots for monthly number of eggs and egg mass index (EMI) of females C. sapidus in the Thermaikos Gulf during the period of study (2020-2022) Different letters indicate statistical difference. Error bars show (± 2) standard deviations.
Figure 12.
Bar plots for monthly number of eggs and egg mass index (EMI) of females C. sapidus in the Thermaikos Gulf during the period of study (2020-2022) Different letters indicate statistical difference. Error bars show (± 2) standard deviations.
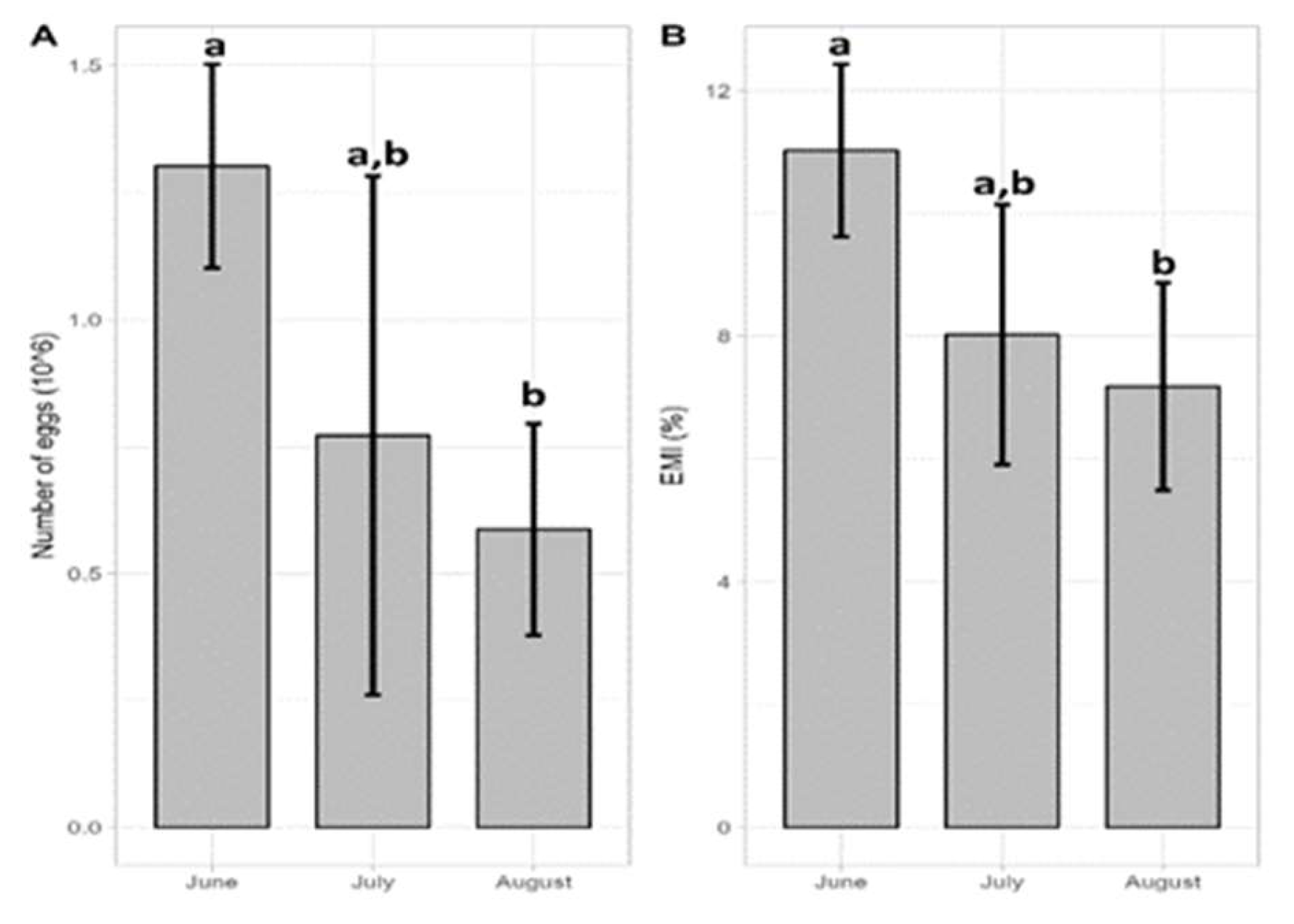
Figure 13.
Linear regression plots with 95% confidence intervals (gray areas) of (A) number of eggs – carapace width A), and (B) egg mass – carapace width (CW) of females C. sapidus in the Thermaikos Gulf.
Figure 13.
Linear regression plots with 95% confidence intervals (gray areas) of (A) number of eggs – carapace width A), and (B) egg mass – carapace width (CW) of females C. sapidus in the Thermaikos Gulf.
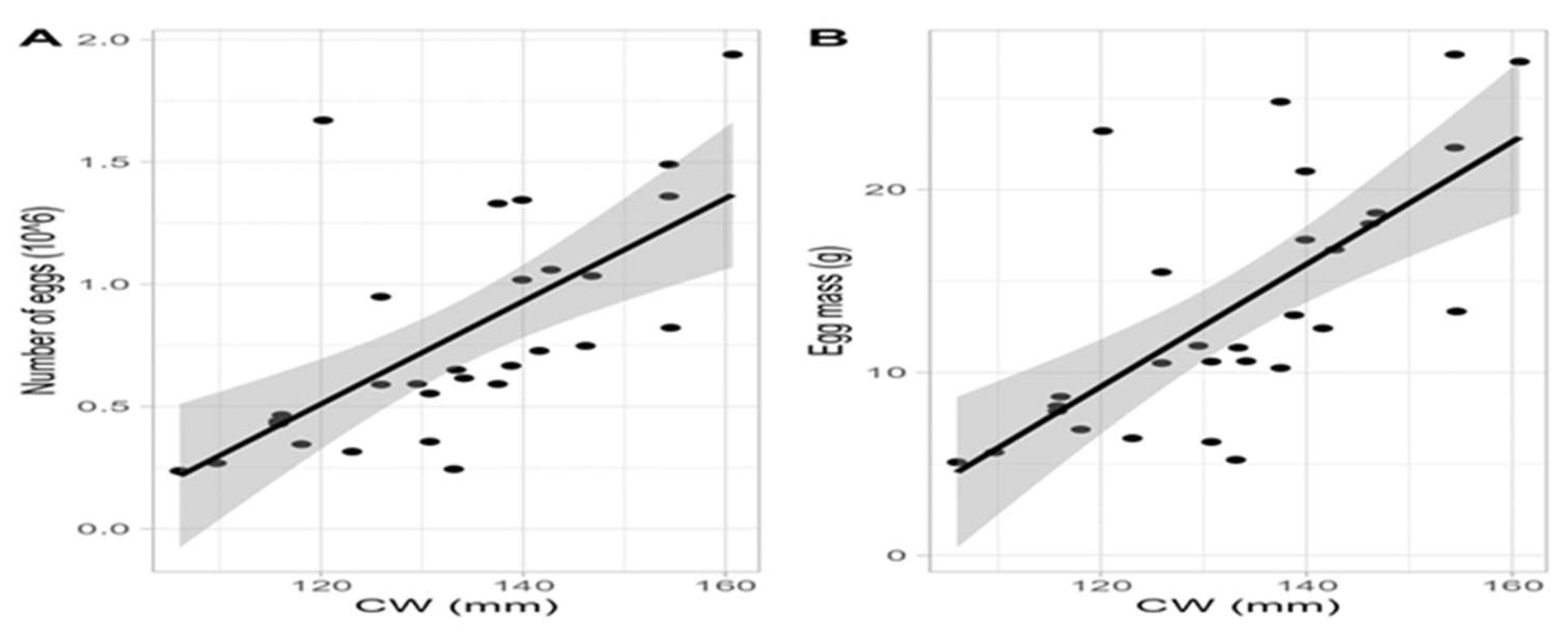
Figure 14.
Schematic diagram of the proposed life cycle of the blue crab in a semi-enclosed lagoon like formation sub-estuary of Aliakmonas Delta (Bara subarea) in the inner Thermaikos Gulf. Sampling stations Bar 1 (depth <1m), Bar 3 (depth <1 m), Bar 2 (depth 2.5 m, characterized by meadows of Gracilaria sp.) and Al 2 (depth 3m, in the adjacent marine waters) are shown.
Figure 14.
Schematic diagram of the proposed life cycle of the blue crab in a semi-enclosed lagoon like formation sub-estuary of Aliakmonas Delta (Bara subarea) in the inner Thermaikos Gulf. Sampling stations Bar 1 (depth <1m), Bar 3 (depth <1 m), Bar 2 (depth 2.5 m, characterized by meadows of Gracilaria sp.) and Al 2 (depth 3m, in the adjacent marine waters) are shown.
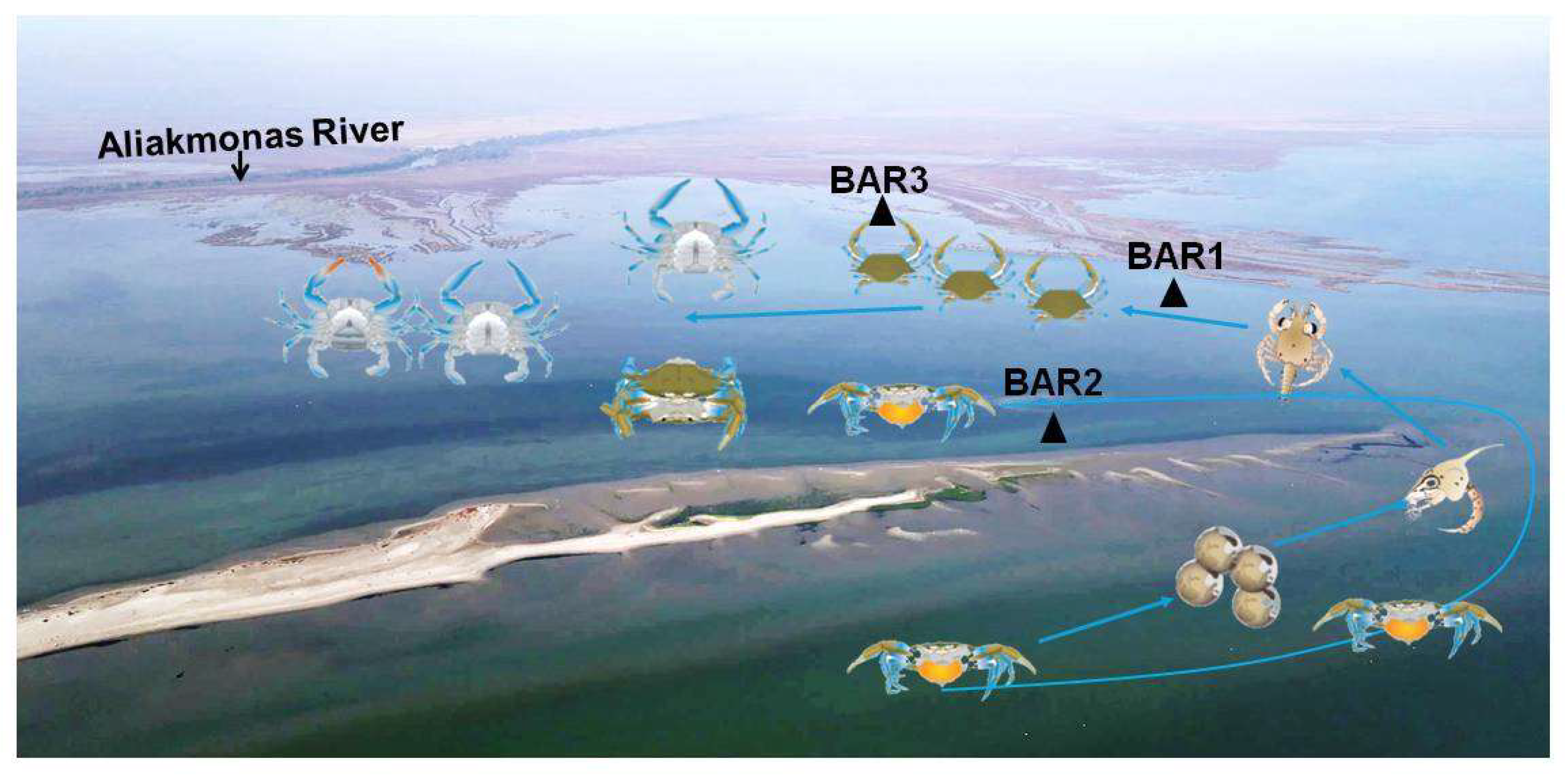

Table 1.
Monthly (A), seasonal (B) and experimental (C) sampling stations (code, depth and coordinates) of Callinectes sapidus in the inner Thermaikos Gulf (NW Aegean Sea).
Table 1.
Monthly (A), seasonal (B) and experimental (C) sampling stations (code, depth and coordinates) of Callinectes sapidus in the inner Thermaikos Gulf (NW Aegean Sea).
| A. Monthly sampling stations. | |||||
|---|---|---|---|---|---|
| Sampling stations (and code) | Depth (m) | Latitude | Longitude | ||
| Methoni Bay (ΜETH 1) | ≤1 | 40°27’37.50”N | 22°35’23.88”E | ||
| Methoni Bay (ΜETH 2) | 3 | 40°27’31.98”N | 22°35’57.00”E | ||
| Methoni Bay (ΜETH 3) | 6 | 40°27’38.70”N | 22°37’30.48”E | ||
| Methoni Bay (ΜETH 4) | 9 | 40°27’31.62”N | 22°38’4.74”E | ||
| Bara (BAR 1) | ≤1 | 40°29’10.98”N | 22°39’37.56”E | ||
| Bara (BAR 2) | 2.5 | 40°29’4.92”N | 22°40’1.98”E | ||
| Off Aliakmonas Delta (AL 2) | 3 | 40°28’18.90”N | E | ||
| Off Aliakmonas Delta (AL 3) | 9 | 40°28’14.46”N | 22°40’12.60”E | ||
| Loudias Bay (LOUD 1) | ≤1 | 40°30’4.08”N | 22°39’32.40”E | ||
| Loudias Bay (LOUD 2) | 3 | 40°30’8.58”N | 22°39’53.16”E | ||
| Loudias Bay (LOUD 3) | 6 | 40°30’10.98”N | 22°40’19.32”E | ||
| Loudias bay (LOUD 4) | 9 | 40°30’12.66”N | 22°40’43.62”E | ||
| B. Seasonal sampling stations. | |||||
| Sampling stations (and code) | Depth (m) | Latitude | Longitude | ||
| Off Aliakmonas Delta (AL 4) | 15 | 40°27’38.82”N | 22°39’50.64”E | ||
| Off Aliakmonas Delta (AL 5) | 25 | 40°27’24.60”N | 22°40’12.90”E | ||
| Off Axios Delta (AX 1) | ≤1 | 40°30’9.30”N | 22°44’2.34”E | ||
| Off Axios Delta (AX 2) | 3 | 40°30’1.86”N | 22°44’6.42”E | ||
| Off Axios Delta (AX 3) | 6 | 40°29’58.74”N | 22°44’9.54”E | ||
| Off Axios Delta (AX 4) | 9 | 40°29’54.42”N | 22°44’13.56”E | ||
| Long line mussel (Mytilus galloprovinciallis) culture unit (LL_MYT) |
7-10 | 40°30’50.04”N | 22°40’44.64”E | ||
| Hang mussel (Mytilus galloprovinciallis culture unit (H_MYT) |
3.5 | 40°30’52.08”N | 22°41’5.10”E | ||
| Loudias River (LOUD_R) | 2.5-3 (800 m from the sea) |
40°32’6.72”N 40°32’7.56”N |
22°40’47.58”E 22°40’46.14”E |
||
| C. Experimental sampling stations. | |||||
| Sampling stations (and code) | Depth (m) | Latitude | Longitude | ||
| Bara (BAR 1) | <1 | 40°29’10.98”N | 22°39’37.56”E | ||
| Bara (BAR 3) | <1 | 400 29’ 9.19’’ Ν | 220 39’16. 31’’Ε | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



