
Submitted:
18 September 2024
Posted:
19 September 2024
You are already at the latest version
Abstract
Salinity, one of the most important abiotic stress factors, affects plant growth and limits agricultural productivity. In the study, the effects of iron (FeNP; 0.025 and 0.05 mM), silver (AgNP; 0,2 ve 0,4 mg L-1) and silicon dioxide (SiNP;7,5 ve 15 mg L-1) nanoparticles on morphological and physiological parameters of in vitro blackberry plants grown under salinity stress (NaCl; 15 mM ve 35 mM) were investigated. According to our study results, it was determined that higher values obtained from SiNP application in terms of shoot development parameters, FeNP application found more succesfull for root development, AgNP application was effective in SPAD, leaf relative water content (LRWC) and relative growth rate (RGR), and FeNP application was increased superoxide dismutase (SOD) and catalase (CAT) enzyme activities. Salt stress was significantly affected the root development, SPAD value, LRWC and RGR, SOD and CAT enzyme activities. As a result, under salt stress conditions, SiNP, FeNP and AgNP applications can significantly reduce the negative effects of stress and promote vegetative development of the plant compared to control conditions.
Keywords:
micropropagation
; nanotechnology
; berries
; rubus
; abiotic
; stress
1. Introduction
Boysenberry, which is included in order Rosales, Rosaceae family, Rubus genus, is a shrub-shaped plant with high economic value traded all over the world with its pleasant taste, high nutritional value fruit [1,2,3]. The cultivation of Boysenberry, which was obtained by crossing Rubus baileyanus and Rubus loganobaccus in the USA between 1921-23, is increasing rapidly due to its suitability for fresh consumption and processing in the food industry, and higher fruit quality compared to other blackberry varieties [4]. Boysenberry fruits are preferred by the food industry due to their high aroma and appeal [5]. While boysenberry cultivation is a new agricultural production area for Turkey, it is produced at a very large rate in the world, especially in Mexico and the United States [6].
One of the most important problems of today's agricultural production, salinity, also poses a serious problem in berries production and many scientific studies in the literature seek solutions to this problem. [7,8]. Boysenberry is considered a salt-sensitive plant based on its ability to tolerate increased salt concentration in the cytosol [9]. Salinity limits the yield and quality of agricultural products and causes morphological, biochemical, physiological and molecular changes in the plant [10,11,12,13]. High salt concentrations in the growth medium of the plant cause restriction of water intake of the plant, loss of turgor, closure of stomata and decrease in photosynthesis [14]. In addition, the detrimental effects of salt stress include dehydration, ionic stress, osmotic stress and oxidative stress, inhibition of photosynthesis and uptake of protein synthesis, nitrogen deficiency (chlorosis and necrosis), slowing of plant growth, and production of highly reactive oxygen (ROS) [15,16]. Accumulation of ROS in the cell induces antioxidative enzymes (such as CAT, SOD, nonspecific peroxidase (POD) and ascorbate peroxidase (APX)), significantly reducing superoxide and hydrogen peroxide levels, resulting in nonspecific oxidation of proteins and nucleic acids [9,17,18,19]. Such conditions can cause plant growth arrest, increased stress, and tissue death [8,17]. Although the tolerance of plants to salt varies according to genetic differences, it can also be affected by environmental factors. For this reason, different methods are used to prevent the negative effects of salinity. However, these methods can be time-consuming and costly applications [20]. For this reason, studies on applications that can increase plant tolerance against salt stress are gaining importance today [21].
The rapid growth and development of nanotechnology has resulted in the rapidly increasing production and use of nanoparticles, and these technological products have begun to be used extensively in agricultural production [22]. The amount of chemicals used extensively, causing various problems in agricultural production, can be reduced with NPs [23], and agricultural product productivity can be increased [24,25,26]. NP applications in plants are generally aimed at increasing the adaptation and resistance of the plant to stress factors [27,28,29,30,31]. It is stated that exogenous application of NPs to plants helps to regulate the antioxidant system activity and cellular water balance in plants [27,32]. SiO2 nanoparticles (SiNP) are single particles of silica dioxide, an inorganic metal oxide with a diameter of less than 100 nm. The low-cost and easily produced SiO2 form of the Si element, which is considered one of the most valuable elements for plant life, has an effective feature to increase resistance against diseases and stress factors in plants [33,34,35]. It has also been reported that SiO2 is a plant growth inducer that strengthens the antioxidant system under salinity stress, increases silicification in the root endodermal layer, improves tolerance to abiotic stresses, and cell water balance [32]. The intake of Fe [36], which is a necessary and limiting micronutrient for plant growth, can be affected by many factors such as soil, climatic conditions, the way the fertilizer is applied, and the growth period of the plant [37]. Therefore, the application of this element as NP can increase the solubility and distribution of insoluble nutrients in the soil, reduce the inactivation of nutrients and increase their bioavailability [38]. As FeNPs are a very rich source of Fe, plants may be more useful due to the gradual release of Fe over a wide pH range [39,40]. It is stated that AgNPs [41,42,43], which are among the metallic nanoparticles, can be effective in promoting plant growth and development [27,44,45,46,47], increasing the chlorophyll content and photosynthesis rate [27,44,48]. In some studies, it has been reported that AgNPs are very effective in improving resistance to salinity in various plants [49,50]. Research is needed on the role of nanoparticles in improving plant tolerance to environmental stresses such as drought and salinity.
Although there are many reports on the effects of NPs in vegetables and field crops as well as on their microbiological activities [51,52,53,54,55], there are limited studies investigating their effects in berries, especially on boysenberry fruits. In this study, it was aimed to determine the effectiveness of some nanomaterials (SiNP, FeNP, AgNP) on the morphological and physiological properties of boysenberry plant grown under in vitro culture conditions and salinity stress (15 and 35 mM NaCl).
2. Materials and Methods
2.1. Materials
In the study, the boysenberry, obtained by crossing Rubus baileyanus and Rubus loganobaccus was used as plant material. Certified saplings obtained from a commercial company were taken into the greenhouse and explants from these plants were transferred to in vitro culture.
Sigma Aldrich brand NPs were used as Nano Materials. SiNPs were used as a dry white powder (Silica nanopowder), with a particle size of 10-20 nm in diameter, a surface area of 590-690 m2/g and a purity of 99.8%. FeNPs (Iron nanopowder) were used as black colored and high purity iron powders with a particle size of 35-45 nm in diameter, a bulk density of 0.5 g/cm3, and a true density of 7.9 g/cm3. AgNPs (Silver nanopowder) were used with an average particle size of <10 nm, an average surface area of 10.0-15.0 m2/g and a purity of 99.9%. Emission Scanning Electron Microscope (SEM) (ZEİSS-LEO 1430) images are shown in Figure 1.
2.2. Method
Two factors were applied in the experiment, different salt concentrations (15 and 35 mM NaCl) and different doses of three different nanomaterials (7.5 and 15 mg L-1SiNP, 0.025 and 0.05 mM FeNP, 0.2 and 0.4 mg L-1AgNP). Shoot tips and nodal cuttings taken from the boysenberry plant were kept in 70% ethanol for 1 minute for surface sterilization, then passed through distilled water 3 times and then sterilized for 15 minutes in 20% sodium hypochlorite solution with a few drops of Tween 20 added (Figure 2a and 2b) [56]. Afterwards, the explants were rinsed 3 times with distilled water in a sterile cabinet and transferred to the media. MS [57] basic medium composition with 3% sucrose and 0.7% agar added was used as the nutrient medium and the pH was adjusted to 5.8. Approximately 40 ml of medium was added to 190 cc glass jars and sterilized at 121°C at 1.5 psi pressure for 20 minutes. Disinfected plant materials were cut in a sterile cabinet with a length of 0.5-0.7 cm and a single seat bud was prepared for planting. The prepared plant materials were planted with four explants in each jar (Figure 2c). The plant materials taken into the environment were cultured for 4 weeks at 25 ± 1ºC, in a 16-hour photoperiod, under a fluorescent lamp (30-35 μmol m-² S-¹) in the climate chamber (Figure 2d).
2.2.1. NP Applications
Explants grown in vitro for 4-5 weeks after that they were transferred to 21 different full-strength MS mediums containing 0.5 ml BAP + 0.5 ml IBA added with different salt and NP concentrations (Table 1).
In order to prepare stock suspensions from NPs, the solutions prepared with deionized water at the determined concentrations and dissolved by sonication for 30 minutes using a probe type sonicator (Misonix, QSonica LLC, Newton, ABD) [58]. Deionized water in which the NPs were suspended was also used to prepare MS. Two different doses of sodium chloride (15-35 mM NaCl) were added to the solution prepared by adding 3% sucrose, 0.7% agar and 0.5 ml BAP + 0.5 ml IBA (pH 5.8).
Boysenberry plantlets obtained as a result of micropropagation under in vitro conditions, in culture rooms with a temperature of 23±1 C, 16 hours of daylight, 8 hours of darkness, and a light intensity of 3000 lux. The research was conducted in three replicates, and 10 jars containing 1 plant were used for each repetition.
2.2.2. Morphological Parameters
Plantlets were cultured in the prepared nutrient media for 4-5 weeks (Figure 3), after that, shoot fresh weight (SFW) (g), shoot dry weight (SDW) (g), shoot length (SL) (mm), stem diameter (SD) (mm), root fresh weight (RFW) (g), root dry weight (RDW) (g), root length (RL) (mm), number of leaves (LN) (per plant), leaf width (LW) (mm) and leaf length (LL) (mm) were determined. To determine the dry weights of shoots and roots, the samples were placed in paper packages and kept in an oven at 70 °C for 48 hours and their weights were determined.
2.2.3. Physiological Parameters
Leaf relative water contents (LRWC %) was performed according to Sanchez et al. [59]. It was calculated using the following equation:
RWC= [(FW-DW)/(TW-DW)]x100
FW: Fresh weight, DW: Dry weight, TW: Turgor weight
SPAD index was measured using chlorophyll content meter (SPAD– 502, Konica Minolta Sensing, Inc., Tokyo, Japan).
Relative Growth Rate (RGR): Plants were weighed in terms of total dry weight before exposure to salt stress (3 true leaf stage) and after the stress period was completed, and the difference between the 2 measurements was divided by the number of days to determine the growth rates of varieties exposed to different treatments during the stress period in g dry weight/day [60].
Superoxide dismutase (SOD) and catalase (CAT) enzyme extraction was done under 4 °C. Spectrophotometric analysis was conducted on a Shimadzu 2401 UV/visible light spectrophotometer. Leaf samples which were used for extract kept at -80 °C. Extract was prepared by homogenization of frozen 1 g leaf sample, containing 0.1 M phosphate buffer (pH 7.8). The homogenate was centrifuged at 10 000 rpm for 10 min at 4°C, and the supernatant was collected. The supernatant was used as assaying superoxide dismutase (SOD) and catalase (CAT).
Catalase activity (CAT) was measured spectrophotometrically by the disappearance of H2O2 by measuring the decrease in absorbance at 240 nm for 1 min due to H2O2 [61]. The reaction mixture contained 25 mM phosphate buffer (pH 7.0), 10 mM H2O2, and enzyme.
SOD activity was defined as meaasuring the inhibition in photoreduction of nitroblue tetrazolium (NBT) by SOD enzyme [62]. The reaction mixture contained 50 mM sodium phosphate buffer (pH 7.8), 0.66 mM EDTA, 10 mM L-methionine, 33 µM NBT, 3.3 µM riboflavin. For the SOD reaction, absorbance was recorded at 560 nm after incubating the mixture at room temperature for 15 minutes under white light.
2.2.4. Statistical Analysis
The study datas were evaluated in the “SPSS 23.0” statistical program. Differences between applications were evaluated in Duncan Multiple Comparison test at p ≤ 0.05 significance level.
3. Results
In this study, it was aimed to examine the effects of different doses of NaCl and NP applications on the physiology and morphological characteristics of the boysenberry plant under in vitro growing conditions. In order to determine the effect of the applications, the parameters which SFW, SDW, SL, SD, RFW, RDW, RL, NL, LW, LL, chlorophyll index (SPAD), LRW), RGR, SOD and CAT activity of the boysenberry plants were examined. Within the scope of the study, the significant differences between the applications were evaluated at the p ≤ 0.05 significance level in the Duncan Multiple Comparison test and the results are presented in the tables below.
3.1. Morphological
The data we obtained regarding shoot development as a result of the study are given in Table 2. When these data are examined, it is seen that the findings are statistically significant (P ≤ 0.05). Obtained data were evaluated, it could be said that NP applications had a significant effect on the shoot development of boysenberry plants grown in vitro. Highest SFW (0.841 g) value was obtained from application of 0.025 mM FeNP+15 mM NaCl and the lowest SFW (0.376 g) was obtained from the application of 15 mg L-1 SiNP+0 mM NaCl. Similarly, the highest SDW value (0.453 g and 0.446 g) was determined in the FeNP applications. On the other hand, it is possible to say that the most significant effect on SL was made by the application of 0.2 mg L-1 AgNP+15 mM NaCl (61.10 mm). When the effects of applications on the plant SD were examined in numerical terms, the highest plant SD was obtained from the application of 0 mM NaCl + 15 mg L-1 SiNP (2.46 mm), and the lowest plant stem diameter was obtained from the application of 15 mM NaCI + 7.5 mg L-1 SiNP.
As seen in the Figure 4 below, while there is no statistically significant difference between NP applications on SFW, in terms of other parameters, it is seen that NP applications and their different doses affect the shoot development of boysenberry. According to the obtained data, it can be said that SiNP application was significantly effective on the increase in shoot length, stem diameter and shoot dry weight. In addition, FeNP application was also effective in increasing the dry weight of the shoot. It is known that salt stress has a negative effect on shoot development. Although it can be said that the development of boysenberry plants grown in vitro under different salt concentrations was generally negatively affected as a result of our study, it is also shown in Figure 4 that this effect of salt stress applied at 15 and 35 mM concentrations was not statistically significant.
In our study, we also examined the effects of salt stress and NP applications on root development of in vitro plantlets. According to our study results, when the root development data in Table 3 were examined, the findings were found to be statistically significant (P ≤ 0.05), and it was determined that there were statistically significant differences between the applications. In the data obtained, the highest average values in terms of root development among all applications were determined in the 15 mM NaCl+0.025 mM FeNP application (RFW 0.613 g, RDW 0.571 g, RL 54.07 mm). The highest average value in terms of RFW was determined as 0.613 g (15 mM NaCl+0.025 mM FeNP), and it was found remarkable that there was a difference of approximately 6 times between this value and the lowest average value (35 mM NaCl+0 mg L-1 NP; 0.103 g) obtained from plants without NP application. A similar situation was observed in the RDW and RL parameters (Table 3).
When the graphs in Figure 5 are examined, the following situation is clearly seen; salt stress negatively affects the root development of boysenberry plants grown under in vitro conditions. SiNP, FeNP and AgNP added to the growth medium under salt stress can eliminate the negative effects of this stress and at the same time promote the root development of the plant compared to control conditions. In this study, the effects of different NPs and their different concentrations on root development were compared. As a result, it was determined that 0.05 mM FeNP application added to the medium was significantly effective compared to other NPs. It was also found that the application of 35 mM salt to the medium stressed the plants and that the presence of salt in these concentrations in the medium weakened the root development of the plants. The lowest RFW (0.368 g) and RDW (0.054 g) mean values were found in plants treated with 35 mM salt.
The development of leaves that meet the nutrients the plant needs through photosynthesis is an important criterion among plant growth and development parameters. In Table 4, the data of the NL, LL and LW of the boysenberry plants of the applications are presented. According to the findings we obtained as a result of our study, we determined the highest number of leaves per plant in the 0 mM salt +0.025 mM FeNP application (23.25 per plant). In this sense, the lowest value (4.50 per plant) was determined in the 35 mM salt+0 mg L-1 NP application, where no NP application was made and 35 mM salt was applied. As a different result from the findings we obtained from the criterion of the NL per plant, the highest average values in terms of LL (18.70 mm) and LW (14.46 mm) parameters were determined in the 15 mM salt + 0.2 mg L-1 AgNP application. However, as in the NL criterion, the lowest values in these parameters were determined in the plants without NP application (LL; 35 mM salt+0 mg L-1 NP 5.34 mm, LW; 35 mM salt+0 mg L-1 NP 5.21 mm).
Leaf growth and development data obtained as a result of the study, belonging to NP and salt stress applications, are shown in Figure 6. According to the results obtained, statistically significant differences were detected between NP applications at the P ≤ 0.05 significance level. As seen in Figure 6, the effects of different NPs on the number of leaves of plantlets in vitro were evaluated and it was determined that the best result was obtained from 0.025 mM FeNP application (18.43 per plant). Among the averages of LL and LW measurements performed to understand the size of the leaves, the highest values were obtained as a result of SiNP applications. Although it is seen in the graph in Figure 6 that salt applications at different concentrations had a negative effect on the leaf development of plantlets, it was determined that this difference was not significant.
In Table 5, the data of the SPAD value, LRWC and RGR of the boysenberry plants of the applications are presented. One of the questions sought to be answered within the scope of this research was to determine whether salt stress affects the chlorophyll content of plantlets in vitro, the water content of leaves and the growth rate of plants. It was also tried to determine whether the negative effects, if any, caused by salt stress can be eliminated with different NP applications. When the data in Table 5 is examined, the following situation is clearly seen; NP application of different types and concentrations to plants under salt stress had a significant effect on the SPAD, LRWC and RGR contents of plantlets. Among all applications, the highest SPAD value (89.63) was determined in 15 mM salt+0.4 mg L-1 AgNP application, the highest LRWC value was determined in 0 mM slat+7.5 mg L-1 SiNP (92.87) application, and the highest RGR value was determined in 0 mM salt+0.2 mg L-1 AgNP (0.056) application.
One of the aims of the study was to determine the superiority of different NPs over each other in eliminating the effects of salt stress, which is an important abiotic stress factor. If we compare different types of NP applications, it can be seen in Figure 7 that FeNP and AgNP applications are more successful than SiNP and control in terms of increasing the SPAD value. A similar situation occurs when we examine the RGR parameter. The 0.2 mg L-1 AgNP application was the application that obtained the highest average value (0.034) compared to all other applications. When the situation is examined in terms of LRWC, it is seen in Figure 4 below that 0.4 mg L-1 AgNP application is successful in increasing the average value. However, there is a difference here, which is that 7.5 mg L-1 SiNP (71.71) application and 0.4 mg L-1 AgNP (71.49) application give similar average results in this parameter and are statistically in the same group (Figure 7).
At the end of the study, CAT and SOD activity levels were determined in the leaves of plants obtained from all applications. According to our study results, when the data of the CAT and SOD enzyme activity of the boysenberry plants are in Table 6 were examined, statistically significant differences were determined between the applications at the significance level of P ≤ 0.05. For CAT and SOD, enhanced activities were seen in all plants grown under salinity stress conditions and treated with NP. Among all applications, the highest values for both CAT (17.82 U/g TA) and SOD (311.55 U/g TA) activity were reached in the 0.05 mM FeNP+35 mM NaCl application. A striking and expected situation in Table 4 was that enzyme activities increased with increasing salt application levels in almost all applications. The lower values were obtained from the 0 mM NaCl application without salt application. The lowest CAT activity values among all applications were determined in the 0 mg NP+0 mM NaCl (16.81 U/g TA) application.
According to our study results, when the data of the the plant CAT and SOD enzyme activity in Figure 8 were examined, statistically significant differences were determined between the applications at the significance level of P ≤ 0.05.
Within the scope of the study, the effects of different NP types and their use at different doses on CAT and SOD enzyme activities of boysenberry plants grown in vitro under salt stress conditions were evaluated. At the same time, whether salt stress had an effect on these enzyme activities was also the subject of the study. The enzyme activity data obtained from the study are given in Figure 8. When the graphs in Figure 8a are examined separately, it is seen that the most important applications that caused the increase in CAT enzyme activity among different NP applications are 0.025 mM and 0.050 mM FeNP and 0.2 mg AgNP applications.
In the SOD enzyme activity data, the effects of 0.050 mM FeNP and 0.2 mg AgNP applications were also found to be significant. However, here, the effectiveness of 0.025 mM FeNP application was lower, while 0.4 mg AgNP application was found to be significantly effective (Figure 8c). The effects of salt stress on enzyme activities were investigated and as a result, the highest CAT and SOD values were determined as a result of the analysis made from the leaves of plants applied with 35 mM NaCl (Figure 8b,d).
4. Discussion
Within the scope of the study, the effects of applications on boysenberry plant growth parameters which fresh shoot weight, fresh root weight, shoot length, root length, stem diameter, leaf number, leaf length, leaf width, dry stem weight, dry root weight, chlorophyll index, LRWC, RGR, SOD activity and CAT activity, were evaluated.
Shoot development is an important indicator in terms of production and growth of plant. Turhan and Eriş [63], in their study examining the effects of NaCl applications on two different strawberry cultivars grown in greenhouse conditions, reported that the fresh root weight was affected by NaCl applications, and 17 and 34 NaCl applications reduced the fresh shoot weight. Kalteh et al. [50], in their study investigating the effects of Si NPs on the basil plant, reported different NaCl concentrations reduced the shoot weight. In the findings obtained in our study, it was determined that there were significant increases in shoot development with FeNP and SiNP applications (Table 2). A study similar to our study findings was obtained in the study by Olur et al. [66], investigated the effects of endophyte bacteria on the growth and health of cucumber plants in plants growing under salt stress, and it was determined that some endophyte bacterial isolates provided a significant increase in plant shoot and root fresh dry weight. Yavuzlar et al. [65], investigated the effects of different glycine concentrations on shoot growth in explants at different NaCl levels in in vitro cultivation, and reported that salt concentrations had an effect on shoot length and reduced shoot length at NaCl concentrations. Mirmazlum et al. [66] investigated the effects of exogenous melatonin applications in different salinity environments and concentrations on buttercup plants, and reported that salinity stress had a negative effect on the growth parameters of buttercup plants and reduced shoot length. Şener and Kurt [56], in their study examining the effect of IBA and BAP applications on the micro-propagation of the shoot tip in vitro conditions in boysenberry plant, and reported the highest stem diameter of 1.44 mm in plant nutrients. This value is lower than our study findings due to the difference in plant nutrients. Haghighi and Pessarakli [49], reported that the plant stem diameter decreased with the addition of NaCl, that salinity had detrimental effects on the plant stem diameters. On the other hand, there was no significant difference between 25 and 50 mM NaCl applications, in their research using a hydroponic growing system, in which they examined the curative effects of different levels of Si and nano-silicon on plant growth in cherry tomatoes under NaCl stress. However, our study findings showed that different NP applications had a positive effect on plant stem diameter. In addition, similar to our study, a decrease in plant stem diameter was observed in NaCl doses compared to control plant groups. Dry weight can be a very useful and reliable indicator used to evaluate the results of any application carried out to increase crop yield or quality in the agricultural production process [67]. Smilarly to our study findings, Azimi et al. [68], in their research examining the effects of six different concentrations of SiO2 nanoparticles and three seed pre-cooling treatments on the germination and growth of wheatgrass, reported that SiO2 nanoparticle applications increased shoot dry weight in wheatgrass. Tahir et al. [69], in their study examining the effects of silicon application in salinity stress growing medium on the growth and ionic composition of different wheat genotypes, reported that the addition of silicon to the nutrient solution increased the amount of shoot dry matter in all genotypes. Koçak [70], in his study examining the determination of salt tolerance levels of 32 local green bean genotypes, reported that shoot dry weight decreased in all genotypes compared to the control. Şener et al. [35] reported that shoot development of strawberry plants grown in vitro under drought stress could be increased by SiNP applications. As listed above, the positive effects of NP applications on plant development are known from various studies in the literature. NPs, which have the capacity to penetrate into small systems due to their properties, can reach distant regions within the plant to trigger different biochemical and physiological pathways [71]. Plants are often exposed to stress factors such as salinity, drought, high and low temperatures, and heavy metals in the environments they are grown in. This can trigger cell toxicity, which leads to the breakdown of macromolecules and the disruption of membrane structure. Reactive oxygen species, known for their growth-inhibiting properties, proliferate under stress conditions, resulting in cytotoxicity and genotoxicity [72]. It has also been suggested that nanoparticles significantly affect the production of bioactive compounds in different plants and plant cell cultures and therefore can be used as standard elicitors in plant cell, tissue and organ cultures to produce valuable metabolites [73]. In this study, the effect of NPs on shoot development in micropropagation of berries, whose propagation by tissue culture is becoming more widespread day by day, was also investigated. When the obtained results were evaluated, it was determined that the use of different types and concentrations of NPs, in line with the information in the literature, stimulated the shoot development of boysenberry grown in vitro. As a result of comparing different NPs with each other, it was determined that SiNPs were more successful in increasing SD, SDW and SL in vitro. Silicon (Si), which is not included in the category of essential elements of higher plants and whose effects have not yet been fully understood, is considered one of the most beneficial elements for plant life [74]. In some studies, it is reported that Si application increases growth and yield by improving plant water status, changing the ultrastructure of leaf organelles, activating plant defense systems and reducing free radicals [35,75,76,77].
Plants need to have a strong root structure in order to continue their lives in a healthy way [78]. Roots, as in all plants, are the most important organs of the plant that take the water and plant nutrients required for the plant's growth and yield from the soil or media [78]. Hwang et al. [79] view that high-quality planting of strawberries should be done with well-grown and well-developed plant roots is actually valid for all plant production processes. Similar to our findings, it is reported in the literature that metal-based nanoparticles have inductive effects on plant growth and development, such as shoot/root development, seed germination, biomass production and physiological/biochemical activities [80]. Turhan and Eriş [63], in their study examining the effects of NaCl applications on two different strawberry cultivars grown in greenhouse conditions, reported that the fresh root weight was affected by NaCl applications, and 17 and 34 NaCl mM applications reduced the fresh root weight. Haghighi and Pessarakli [49] investigated the ameliorative effects of different levels of Si and nano-silicon on plant growth in cherry tomatoes under salt stress using hydroponic cultivation system. They reported that fresh root weight and root volume were significantly affected by stress and the lowest values were obtained from 25 mM NaCl applications. They also reported that silicon made a positive contribution to fresh shoot weight in applications at 25 mM NaCl level [49]. Siddiqui et al. [32], investigated the effects of Na SiO2 on the growth development of zucchini plants grown under salt stress conditions and reported that Na SiO2 alleviated the negative effects of salinity, improved the growth characteristics of pumpkin plants and increased the fresh root weight. Badawy et al. [81], studied the effects of nanoparticle (Si and Se) applications on two different rice varieties grown in saline conditions and reported that root length decreased significantly as a result of the high inhibitory effect of NaCl. Salih et al. [82] investigated the effects of biogenic AgNPs on parameters such as germination rate and growth of tomato and reported that Ag-containing nanoparticles had a significant effect on the development of stem and root systems. In the root dry weight, Azimi et al. [68], reported that SiO2 nanoparticle applications increased root dry weight in wheatgrass. Similarly, Tahir et al. [69], reported that the addition of silicon to the salinity stress growing medium increased the amount of shoot dry matter in all genotypes. Siddiqui et al. [32], reported that Na SiO2 improved the growth characteristics of zucchini plants by alleviating the negative effect of salinity n in their study examining the curative effect of Na SiO2 under salt stress conditions of zucchini plants. In another study, Doğan [83], in his research examining the effect of humic acid and silicon applications on eliminating the negative effects of heavy metals on strawberry cultivation, reported that humic acid and silicon applications had positive effects on the dry root weight of plants. According to the data obtained from our study, NP applications were able to protect the root development of boysenberry plants grown under in vitro conditions from the negative effects of salt stress. NP applications provided significant increases compared to the control in all parameters examined to monitor root development. As a result of our study, we determined that the use of FeNP had the best effect on root development of plants grown under salt stress under in vitro conditions.
Leaves are an important organ that ensures the survival of plants, so leaf number, leaf width, leaf length, chlorophyll index and LRWC parameters are important indicators of healthy plant growth [84]. In the leaf number, Şener and Kurt [56], reported the highest leaf number of 48.20 mm in the boysenberry plant. This value was higher than our study findings due to the difference in plant nutrients. Kalteh et al. [50], in their study examining the effect of silicon nanoparticle applications on salinity stress in basil, reported that silicon nanoparticle applications increased the amount of photosynthesis of the plant by decreasing the salinity stress, increasing the leaf number and leaf area. Bora [85], in his research examining the effects of NaCl stress applications on Jalapeno pepper cultivar in hydroponic growing system, reported that the lowest number of leafs was obtained in NaCl applications. Similar results were obtained from our study findings. Bernal et al. [86] studied the effect of paclobutrazol and AgNPs on the in vitro regeneration of potato (Solanum tuberosum L.) and reported that leaf number, root number, stomata density and chlorophyll content increased when PAC was combined with AgNPs compared to when PAC was used alone. Avestan et al. [87] reported that NaSiO2 applications in strawberry reduced the negative effects of salinity and improved plant vegetative growth. In the leaf area (length and width), Şener and Kurt [56], reported the highest leaf length of 14.09 mm and the highest leaf width of 11.72 mm in the boysenberry plant. These values were lower than our study findings due to the difference in plant nutrients. Akhoundnejad et al. [88], in their study examining the effects of different doses of AgNP applications on the growth and development of onion plants, and reported that the leaf width was higher than the control, while the leaf length was lower. Iqbal et al. [51], reported that different doses of AgNP applications in wheat significantly increased leaf area. In their study, Abou-Shlell et al. [89], examined the effect of foliar application of different growth promoting nanoparticles in different concentrations of salinity environment on the growth, biochemical and anatomical properties of Moringa oliefera L. plant, and reported that FeNP applications increased leaf area. As a result of our study, the effect of salt stress on the NL, LL and LW was not found to be significant. It was expected that the effect of stress on this green part of the plant, which contains a lot of water, would be significant. Numerical differences were detected between the averages. The effects of stress may not have been measured significantly due to reasons such as the fact that the study was conducted in in vitro conditions, was more controlled than in vivo conditions and had a shorter duration. One of the hypotheses of the research was that NPs alleviate the effects of stress and have a positive effect on plant growth and development. The findings supported this hypothesis. The use of FeNP and SiNP at different concentrations had a significant effect on the leaf development of boysenberry plants grown under in vitro conditions. A striking point in the findings is that AgNPs fell behind FeNPs and SiNPs in this regard. In many studies, the inhibitory properties of AgNPs are mentioned [90,91,92]. When the data obtained from our study are evaluated, it would not be correct to mention an inhibitory property for AgNPs. Only in some parameters, lower average values were recorded from AgNP applications compared to other NPs or the control application. However, due to reasons such as the fact that the study was conducted under in vitro cultivation conditions and was conducted on a plant from a botanically different family and species, the results obtained are more diverse and cannot be directly compared with the studies of other authors. This shows that the same nanoparticles have a multifaceted effect.
In the chlorophyll index (SPAD value), Al-aghabary et al. [93], in their study investigating the effects of different nanoparticle applications and salt stress on the chlorophyll content, chlorophyll fluorescence, and malondialdehyde concentration in tomato, reported that salt stress significantly reduced the chlorophyll content, but added SiNP increased the chlorophyll content under salt stress 10 days after the application. Similarly, to our study findings, Kuşvuran et al. [94], in their study examining the changes in the amount of Na+, K+, Cl- ions and the amount of chlorophyll in Cucumis genotypes treated with 100 mM NaCl, reported a decrease in the amount of chlorophyll in plant leafs under NaCl stress. Ghafariyan et al. [39] examined the effects of FeNP applications in soybean on chlorophyll variations and the possibilities of reducing iron deficiency in soybeans, and reported that even low concentrations of FeNP treatments under hydrophobic conditions significantly increased the chlorophyll content in soybean leafs. In another study, Bora [85], in his research examining the effects of different salt concentrations applied during different vegetation periods on the physiological, morphological and chemical properties of pepper plants, reported that NaCl stress applications in pepper plant decreased chlorophyll values. These research results are similar to the NaCl applications in our study findings, and our study findings show that, the negative effect of salt stress reduces AgNPs and FeNPs. Another study with similar results to our study was conducted by Gökçe [95]. Gökçe, in his study examining the effects of AgNPs on tomato seeds, determined that the increase in AgNP concentrations was statistically significant on pigment values [95]. LRWC is an important indicator of leaf water status, which reflects the balance between water uptake and transpiration rate in plants [96]. In the LRWC, Fan et al., [97] in their study evaluating the effects of Si and K applications on plant growth and ion-selective absorption under saline-alkali stress at different concentrations in ryegrass, reported that Si and K applications increased LRWC. Zahedi et al. [98], in their study examining the effects of drought on plant growth and fruit yield of SiO2 and Se nanoparticle applications in strawberry, reported SiO2 and Se nanoparticle applications increased the LRWC. Bora [85], in his study examining the effects of NaCl stress applications on the Jalapeno pepper cultivar in the hydroponic growing system, reported that LRWC at 0 mM dose was 96.87%, while LRWC at 100 mM dose decreased to 67.29% in NaCl stress applications. Similarly, according to our study findings, it was determined that LRWC decreased under the effect of NaCl stress, but the effect of stress was alleviated by NP applications. NP application can lead to changes in the transcription of genes involved in the biosynthesis of plant hormones or signal transduction [45,99]. Transcriptomic studies have revealed that the expression of genes related to abiotic and biotic stress responses is also increased by nanoparticle application [100,101]. In some cases, various NPs can increase the activity and expression of genes related to phytohormones that promote root growth, such as ABA and cytokinin. Thus, water and nutrient uptake by plants can be accelerated and increased [102,103,104]. RGR is an important indicator used to evaluate plant growth performance and productivity as an important determinant of plant competitiveness [105]. Abou-Shlell et al. [89], reported that NP applications (Zn, Fe, Cu and Si) decreased the negative effects of salty environment and improved vegetative growth in Moringa oliefera L.. Siddiqui et al. [106], in their literature review study to emphasize the key role of nanoparticle applications in plants, reported that the effect of NPs varies from plant to plant, depending on the application concentration and shape. Accordingly, they reported that SiNPs increased germination and antioxidant activity in applications in different plants, ZnNPs increased plant growth and development, AgPNs increased plant growth. In various studies, the inhibitory effects of AgNPs, which are metal-based NPs, have been mentioned [90,91,92]. The results obtained from our study are the opposite, and indicate that AgNPs promote growth and development. It is thought that this is due to the fact that AgNPs, when compared to AgNO3, release ions (Ag+) that can block the ethylene receptor by changing the copper ion cofactor of the ethylene binding site [86]. According to Elatafi and Fang [107], AgNPs have a larger surface area to volume ratio compared to other forms of silver such as AgNO3, which can make them more effective. The ions (Ag+) of AgNPs, which have lower toxicity effects, have the ability to inhibit the formation of 1-aminocyclopropane-1-carboxylic acid and thus the effect of ethylene.
Antioxidants (such as SOD and CAT) are the first defence block against damage caused by free radicals caused by biotic and abiotic environmental factors that affect plant growth, development and productivity. NP applications often result in increased levels of reactive oxygen species (ROS) and subsequent activation of the antioxidant system [108]. In the CAT enzyme activity, Zahedi et al. [98], reported SiO2 and Se nanoparticle applications increased the CAT enzyme activity. Al-aghabary et al. [93], reported that salt stress inhibited CAT activity, but added Si improved CAT activity under salt stress. Günalp [109], in his study examining the effects of Jasmonic Acid (JA) application on salt tolerance in eggplant embryo culture, and reported that CAT enzyme activity, one of the antioxidative enzymes, was at very low levels compared to the control group, while on the other hand, it increased catalase enzyme activities in very high amounts. Similarly, in our study findings, it was concluded that CAT activities in boysenberry plants were found at a low rate compared to the control group. According to our study findings, the high result of CAT activity in 35 mM NaCl + 0.05 mM FeNP application shows that stress factors activate the defense mechanisms by activating the antioxidant defense system. Another study with similar findings was conducted by Marček et al [19], which they examined the effects of different concentrations of NaCl (15 and 35 mM) applications on blackberry plants in vitro, 35 mM NaCl application caused a significant increase in CAT activities, which proved to be an effective antioxidant defense mechanism in blackberry plants. In the SOD enzyme activity, Zahedi et al. [98], reported SiO2 and Se nanoparticle applications increased the SOD enzyme activity. Al-aghabary et al. [93], reported that salt stress significantly reduced SOD activity, but added Si improved SOD activity under salt stress. Günalp [109] examined the effects of Jasmonic Acid (JA) application on salt tolerance in eggplant embryo culture. According to Günalp's study findings, the control group determined that SOD activity was found in applications containing JA and applications containing only NaCl, and SOD activity was reported to be significantly increased in all JA or salt treatments compared to the control group [109]. Our study findings showed the same results, showing that SOD activity increased with increasing salt concentrations, as in the reference studies. Unlike these similar results, Gökçe [95], reported a decrease in SOD activities in the stem tissues of tomato plants in his study, in which he examined the physiological, morphological and biochemical effects of AgNPs in tomato seeds. It is known that nanoparticles play an important role in affecting antioxidant defense systems and triggering some other important metabolic activities in plants under salt stress [110]. In our study, the roles of NPs against oxidative stress were investigated under in vitro cultivation conditions and salinity conditions and significantly increased activities of SOD and CAT enzymes in the leaves of boysenberry plants were determined. Antioxidant enzymes such as SOD and CAT are induced in plants in response to abiotic stress. Therefore, their levels are expected to increase under stress conditions. These enzymes are also a vital part of the antioxidant defense system to prevent hydroxyl radicals responsible for lipid peroxidation in cell membranes and biomarkers of oxidative stress [111].
5. Conclusions
Agricultural production activities, which are in a dynamic process, produce solutions to the main problems encountered for the continuation of human existence. It is an important research area to develop new solution approaches to the problems that arise in agricultural production activities by using the latest developments in science and technology, such as nano technology. One of these problems is salinity caused by factors such as excessive irrigation and fertilization. This study was carried out to investigate the effects of different concentrations of NaCl and different doses of NP on the morphological and physiological properties of the boysenberry plant under in vitro growing conditions.
According to our findings obtained from the parameters in which the effects of the applications were monitored, 7.5 and 15 mg L-1 SiNP application was significantly effective on the increase in SL, SD and SDW. When the effects of NPs applied at different concentrations on root development were compared, it was determined that 0.05 mM FeNP application added to the medium was significantly effective on RFW and RDW compared to other NPs. While the highest results in terms of NL of in vitro plants were obtained from 0.025 mM FeNP application, the highest average LL values were determined in 0.2 mg L-1 AgNP and 15 mg L-1 SiNP application, and the highest average LW value was determined in 7.5 mg L-1 SiNP application. While 0.4 mg L-1 AgNP application was found to be more effective than other applications in terms of SPAD value, the highest RGR average value was determined in 0.2 mg L-1 AgNP application. The highest LRWC values were determined in 0.4 mg L-1 AgNP and 7.5 mg L-1 SiNP applications. Application of 0.2 mg L-1 AgNP, 0.025 and 0.05 mM FeNP was found to be significantly effective on CAT enzyme activity. The highest SOD enzyme activity values were recorded from 0.4 mg L-1 AgNP and 0.05 mM FeNP applications. As a conclusion, it has been concluded that the applications of SiNP, FeNP and AgNP at different doses in vitro growing conditions and under salinity stress have a positive effect on the vegetative growth of the boysenberry plant, as it reduces the negative effects of salinity.
Author Contributions
“Conceptualization, S.A. and Z.K.; methodology, S.A. and Z.K.; data curation, S.A., and Z.K.; writing—original draft preparation, S.A. and Z.K.; writing—review and editing, S.A.; supervision, S.A.; project administration, S.A..; funding acquisition, S.A. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was funded by Akdeniz University, Scientific Research Projects Coordination Unit, Antalya, Turkey, grant number FYL-2020-5362 (only for research funding).
Acknowledgments
This research was funded by Akdeniz University, Scientific Research Projects Coordination Unit, Antalya, Turkey, grant number FYL-2020-5366 (only for research funding). The measurements and analyses in the study were carried out in the laboratory under the responsibility of Assoc. Prof. Dr. Kamile Ulukapı and Assoc. Prof. Dr. Ayşe Gül Nasırcılar. We thank them for their support and assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hummer, K.E.; Janick, J. Rubus iconography: Antiquity to the renaissance. Acta Hort. 2007, 759, 89–105. [Google Scholar] [CrossRef]
- Verma, R.; Gangrade, T.; Punasiya, R.; Ghulaxe, C. Rubus fruticosus (blackberry) use as an herbal medicine. G.R.Y. Institute of Pharmacy, "Vidhya Vihar" Borawan, Khargone, Madhya Pradesh, India. 2014. [CrossRef]
- Zia-Ul-Haq, M.; Riaz, M.; de Feo, V.; Jaafar, H.; Moga, M. Rubus fruticosus L.: Constituents, biological activities and health related uses. Molecules 2014, 19, 10998–11029. [Google Scholar] [CrossRef] [PubMed]
- Gruner, L.A.; Kornilov, B.B. Priority trends and prospects of blackberry breeding in conditions of Central Russia. Vavilovskii Zhurnal Genet Selektsii. 2020, 24, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Hall, H.K.; Langford, G. The'Boysenberry': development of the cultivar and industries in California, Oregon and New Zealand. In IX International Rubus and Ribes Symposium. 2005, 777, 103–108. [Google Scholar] [CrossRef]
- Balcı, G.; Keles, H. Bazı böğürtlen çeşitlerinin yozgat ekolojisinde adaptasyon yeteneklerinin belirlenmesi. Adnan Menderes Üniversitesi Ziraat Fakültesi Dergisi. 2019, 16, 45–52. [Google Scholar] [CrossRef]
- Cardona, W.A.; Gutiérrez, J.S.; Monsalve, O.I.; Bonilla, C.R. Salinity effect on the vegetative growth of Andean blackberry plants (Rubus glaucus Benth.) inoculated and non-inoculated with mycorrhizal fungi. Revista Colombiana de Ciencias Hortícolas. 2017, 11, 253–266. [Google Scholar] [CrossRef]
- Gonzalez-Jimenez, S.L.; Castillo-Gonzalez, A.M.; del Rosario Garcia-Mateos, M.; Valdez-Aguilar, L.A.; Ybarra-Moncada, C.; Avitia-Garcia, E. Response of blackberry (Rubus spp.) Cv. tupy to salinity. Revısta Fıtotecnıa Mexıcana. 2020, 43, 299–306. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Gill, B.S.; Appels, R.; Botha-Oberholster, A.-M.; Buell, C.R.; Bennetzen, J.L.; Chalhoub, B.; Chumley, F.; Dvorak, J.; Iwanaga, M.; Keller, B.; Li, W.; McCombie, W.R.; Ogihara, Y.; Quetier, F.; Sasaki, T. A Workshop report on wheat genome sequencing: international genome research on wheat consortium. Genetics. 2004, 168, 1087–1096. [Google Scholar] [CrossRef]
- Zhu, J.-K. Plant Salt Stress. In eLS, (Ed.). 2007. [CrossRef]
- Ayala-Astorga, G.I.; Alcaraz-Melendez, L. Salinity effects on protein content, lipid peroxidation, pigments, and proline in Paulownia imperialis (Siebold & Zuccarini) and Paulownia fortunei (Seemann & Hemsley) grown in vitro. Electronic Journal of Biotechnology. 2010, 13, 1–15. [Google Scholar] [CrossRef]
- Agastian, P.; Kingsley, S.J.; Vivekanandan, M. Effect of salinity on photosynthesis and biochemical characteristics in Mulberry genotypes. Photosynthetica. 2000, 38, 287–290. [Google Scholar] [CrossRef]
- Hannachi, S.; Steppe, K.; Eloudi, M.; Mechi, L.; Bahrini, I.; Van Labeke, M.-C. Salt Stress induced changes in photosynthesis and metabolic profiles of one tolerant (‘Bonica’) and one sensitive (‘Black Beauty’) eggplant cultivars (Solanum melongena L.). Plants. 2022, 11, 590. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Improving plant abiotic-stress resistance by exogenous application of osmoprotectants glycinebetaine and proline. Environmental and Experimental Botany, 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant, Cell and Environment. 2007, 25, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Miranda, R.S.; Mesquita, R.O.; Freitas, N.S.; Prisco, J.T.; Filho, E.G. Nitrate: ammonium nutrition alleviates detrimental effects of salinity by enhancing photosystem II efficiency in sorghum plants. Revista Brasileira de Engenharia Agrícola e Ambiental, 2014; ISSN 1807-1929. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant and Cell Environment. 2009, 33, 453–467. [Google Scholar] [CrossRef]
- Marček, T.; Velić, D.; Sabo, M.; Dugalić, K.; Velić, N.; Pranjić, A.; Klarić, I.; et al. Salinity effects on blackberry plants (Rubus fructicosus L.) grown in vitro. In 8th International Scientific/Professional Conference, Agriculture in Nature and Environment Protection, Vukovar, Croatia. 2015, 298-302. Croatian Soil Tillage Research Organization (CROSTRO). Available online: https://www.cabdirect.org/cabdirect/abstract/20173162760 (accessed on 09 March 2023).
- Deliboran, A.; Savran, Ş. Toprak tuzluluğu ve tuzluluğa bitkilerin dayanım mekanizmaları. Türk Bilimsel Derlemeler Dergisi. 2015, 1, 57–61. Available online: https://dergipark.org.tr/tr/pub/derleme/issue/35094/389309 (accessed on 09 March 2023).
- Winicov, I. Characterization of rice (Oryza sativa L.) plants regenerated from salt tolerant cell lines. Plant Science, 1996, 113, 105–111. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Mthiyane, D.M.N.; Onwudiwe, D.C.; Babalola, O.O. Harnessing the known and unknown impact of nanotechnology on enhancing food security and reducing postharvest losses: constraints and future prospects. Agronomy. 2022, 12, 1657. [Google Scholar] [CrossRef]
- Johnston, C.T. Probing the nanoscale architecture of clay minerals. Clay Minerals. 2010, 45, 245–279. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gill, S.S.; Duarte, A.C.; Pereira, E.; Ahmad, I. Silver nanoparticles in soil–plant systems. Journal of Nanoparticle Research. 2013, 15, 1896. [Google Scholar] [CrossRef]
- Delfani, M.; Firouzabadi, M.B.; Farrokhi, N.; Makarian, H. Some physiological responses of black-eyed pea to iron and magnesium nanofertilizers. Commun. Soil Sci. Plant Anal. 2014, 45, 530–540. [Google Scholar] [CrossRef]
- Tudi, M.; Daniel Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D.T. Agriculture development, pesticide application and its impact on the environment. Int J Environ Res Public Health. 2021, 18, 1112. [Google Scholar] [CrossRef]
- Şener, S.; Sayğı, H. The role of silver nanoparticles in response of in vitro boysenberry plants to drought stress. Horticulturae 2023, 9, 1177. [Google Scholar] [CrossRef]
- Ocsoy, I.; Paret, M.L.; Ocsoy, M.A.; Kunwar, S.; Chen, T.; You, M.; Tan, W. Nanotechnology in plant disease management: DNA-directed silver nanoparticles on graphene oxide as an antibacterial against Xanthomonas perforans. Acs Nano. 2013, 7, 8972–8980. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, Z.M. Expression Profiling of certain MADS-Box Genes in Arabidopsis Thaliana plant treated by silver nanoparticles. Czech Journal of Genetics and Plant Breeding. 2017, 53, 30–36. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Nair, R.; Giraldo, J.P.; Prasad, P.V.V. Cerium oxide nanoparticles decrease drought induced oxidative damage in sorghum leading to higher photosynthesis and grain yield. ACS omega. 2018, 3, 14406–14416. [Google Scholar] [CrossRef]
- Wu, H.; Shabala, L.; Shabala, S.; Giraldo, J. P. Hydroxyl radical scavenging by cerium oxide nanoparticles improves Arabidopsis salinity tolerance by enhancing leaf mesophyll potassium retention. Environmental Science: Nano. 2018, 5, 1567–1583. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Environmental Toxicology Chemistry, 2014, 33, 2429–2437. [Google Scholar] [CrossRef]
- Brecht, M.O.; Datnoff, L.E.; Kucharek, T.A.; Nagata, R.T. Influence of silicon and chlorothalonil on the suppression of gray leaf spot and increase plant growth in St. Augustinegrass. Plant Disease. 2004, 88, 338–344. [Google Scholar] [CrossRef]
- Karimi, J.; Mohsenzadeh, S. 'Effects of silicon oxide nanoparticles on growth and physiology of wheat seedlings'. Russian Journal of plant physiology. 2016, 63, 119–123. [Google Scholar] [CrossRef]
- Şener, S.; Sayğı, H.; Duran, C.N. Responses of in vitro strawberry plants to drought stress under the influence of nano-silicon dioxide. Sustainability 2023, 15, 15569. [Google Scholar] [CrossRef]
- Jelali, N.; Dell'Orto, M.; Rabhi, M.; Zocchi, G.; Abdelly, C.; Gharsalli, M. Physiological and biochemical responses for two cultivars of Pisum sativum ("Merveille de Kelvedon" and "Lincoln") to iron deficiency conditions. Scientia horticulturae. 2010, 124, 116–121. [Google Scholar] [CrossRef]
- Liu, S.Y.; Shieh, J.P.; Tzeng, J.I.; Chia-Hui, H.; Cheng, Y.L.; Huang, K.L.; Wang, J.J. Noveldepots of ketorolac esters have long-acting antinociceptive and antiinflammatory effects. Anesth. Analg. 2005, 101, 785–92. [Google Scholar] [CrossRef] [PubMed]
- Askary, M.; Talebi, S.; Amini, F.; Bangan, A. D. Effect of NaCl and iron oxide nanoparticles on Mentha Piperita essential oil composition. Environmental and Experimental Biology. 2016, 14, 27–32. [Google Scholar] [CrossRef]
- Ghafariyan, M.H.; Malakouti, M.J.; Dadpour, M.R.; Stroeve, P.; Mahmoudi, M. Effects of magnetite nanoparticles on soybean chlorophyll. Environ. Sci. Technol. 2013, 47, 10645–10652. [Google Scholar] [CrossRef] [PubMed]
- Armin, A.; Fotouhi, R.; Szyszkowski, W. On the FE modeling of soil-blade interaction in tillage operations. Finite Elements in Analysis and Design, 2014, 92, 1–11. [Google Scholar] [CrossRef]
- Beykaya, M.; Çağlar, A. Bitkisel özütler kullanılarak gümüş nanopartikül (AgNP) sentezlenmesi ve antimikrobiyal etkinlikleri üzerine bir araştırma. Afyon Kocatepe Üniversitesi Fen Ve Mühendislik Bilimleri Dergisi 2016, 16, 631–641. [Google Scholar] [CrossRef]
- Prathna, T.C.; Sharma, S.K.; Kennedy, M. Nanoparticles in household level water treatment: an overview. Separation and Purification Technology, 2018, 199, 260–270. [Google Scholar] [CrossRef]
- Mehmood, A. Brief overview of the application of silver nanoparticles to improve growth of crop plants. IET Nanobiotechnol. 2018, 12, 701–705. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J Bot. 2012, 12, 134–136. [Google Scholar] [CrossRef]
- Kaveh, R.; Li, Y.S.; Ranjbar, S.; Tehrani, R.; Brueck, C.L.; Van Aken, B. Changes in Arabidopsis thaliana gene expression in response to silver nanoparticles and silver ions. Environ Sci Technol. 2013, 47, 10637–44. [Google Scholar] [CrossRef]
- Vannini, N.; Girotra, M.; Naveiras, O.; Nikitin, G.; Campos, V.; Giger, S.; Roch, A.; Auwerx, J.; Lutolf, M.P. Specification of haematopoietic stem cell fate via modulation of mitochondrial activity. Nat Commun 2016, 7, 13125. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. Impacts of silver nanoparticles on plants: A focus on the phytotoxicity and underlying mechanism. Int J Mol Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Hatami, M.; Ghorbanpour, M. Effect of nanosilver on physiological performance of pelargonium plants exposed to dark storage. J. Hort. Res. 2013, 21, 15–20. [Google Scholar] [CrossRef]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherrytomatoes (Solanum lycopersicum L.) at early growth stage. Scientia Horticulturae 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Aliabadi, M.M.; Nosratabadi, A.F. Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J Chem Health Risks. 2014, 4, 49–55. [Google Scholar]
- Iqbal, M.; Raja, N.; Mashwani, Z.R.; Hussain, M.; Ejaz, M.; Yasmeen, F. Effect of silver nanoparticles on growth of wheat under heat stress. Iran J Sci Technol Trans Sci, 2017, 43, 387–395. [Google Scholar] [CrossRef]
- Nair, P.G.M.; Chung, I.M. Impact of copper oxide nanoparticles exposure on Arabidopsis thaliana growth, root system development, root lignificaion, and molecular level changes. Environ Sci Pollut Res 2019, 21, 12709–12722. [Google Scholar] [CrossRef]
- Thuesombat, P.; Hannongbua, S.; Akasit, S.; Chadchawan, S. Effect of silver nanoparticles on rice (Oryza sativa L. cv. KDML 105) seed germination and seedling growth. Ecotoxicol Environ Saf, 2014, 104, 302–309. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Latta, D.E.; McLean, J.E.; Britt, D.W.; Boyanov, M.I.; Anderson, A.J. Fate of CuO and ZnO nano- and microparticles in the plant. Environ. Sci. Technol. 2013, 47, 4734–4742. [Google Scholar] [CrossRef]
- Pei, Z.; Ming, D.; Liu, D.; Wan, G.; Geng, X.; Gong, H.; Zhou, W. Silicon improves the tolerance to water-deficit stress induced by polyethylene glycol in wheat (Triticum aestivum L.) seedlings. J. Plant Growth Regul. 2010, 29, 106–115. [Google Scholar] [CrossRef]
- Şener, S.; Kurt, Z. The effects of Benzyl Amino Purine (BAP) and Indole Butyric Acid (IBA) on in vitro shoot proliferation and rooting of Boysenberry. COMU J. of Agr. F. 2022, 10, 161–168. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A. Revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiological Plantarum, 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yin, L.; Colman, B.P.; McGill, B.M.; Wright, J.P.; Bernhardt, E.S. Effects of silver nanoparticle exposure on germination and early growth of eleven wetland plants. PLoS ONE 2012, 7, e47674. [Google Scholar] [CrossRef]
- Sanchez, F.J.; de Andrés, E.F.; Tenorio, J.L.; Ayerbe, L. Growth of epicotyls, turgor maintenance and osmotic adjustment in pea plants (Pisum sativum L.) subjected to water stress. Field Crop. Res. 2004, 86, 81–90. [Google Scholar] [CrossRef]
- Sugiyama, S. Relative contribution of meristem activities and specific leaf area to shoot relative growth rate in C3 grass species. Functional Ecology, 2005, 19, 925–931. [Google Scholar] [CrossRef]
- Cakmak, I. Activity of ascorbate-dependent H2O2-scavenging enzymes and leaf chlorosis are enhanced in magnesium-and potassium-deficient leaves, but not in phosphorus-deficient leaves. J. Exp. Bot. 1994, 45, 1259–1266. [Google Scholar] [CrossRef]
- Bayer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Turhan, E.; And Eriş, A. Growth and stomatal behaviour of two strawberry cultivars under long-term salinity stress. Turkish Journal of Agriculture and Forestry, 2007, 31, 54–61. [Google Scholar]
- Olur, Ü.; Uçar, C.; Akköprü, A. Tuz stresi altında gelişen bitkilerden izole edilen endofit bakterilerin bazı bitki gelişimini teşvik etme mekanizmalarının ve hıyar fide gelişimine etkilerinin belirlenmesi. Mustafa Kemal Üniversitesi Tarım Bilimleri Dergisi 2021, 26, 635–648. [Google Scholar] [CrossRef]
- Yavuzlar, E.E.; Karadal, S.; Adak, N. in vitro koşullarda farklı glisin konsantrasyonlarının çileklerde tuzluluk stresi üzerine etkileri. Anadolu Tarım Bilimleri Dergisi 2021, 36, 162–166. [Google Scholar] [CrossRef]
- Mirmazloum, I.; Eisa, E.A.; Honfi, P.; Tilly-Mándy, A. Exogenous melatonin application ınduced morpho-physiological and biochemical regulations conferring salt tolerance in Ranunculus asiaticus L. Horticulturae 2023, 9, 228. [Google Scholar] [CrossRef]
- Hort Americas. Fresh Weight vs. Dry Weight. 2022. Available online: https://hortamericas.com/blog/fresh-weight-vs-dry-weight/ (accessed on 9 March 2023).
- Azimi, R.; Borzelabad, M.J.; Feizi, H.; Azimi, A. Interaction of SiO2 nanoparticles with seed prechilling on germination and early seedling growth of tall wheatgrass (Agropyron elongatum L.). Polish J Chem Technol, 2014, 16, 25–29. [Google Scholar] [CrossRef]
- Tahir, M.; Rahmatullah, A.; Aziz, T.; Ashraf, M. Wheat genotypes differed signifi cantly in their response to silicon nutrition under salinity stress. Journal of Plant Nutrition, 2010, 33, 1658–1671. [Google Scholar] [CrossRef]
- Koçak, A. Bazı Yerel Taze Fasulye Genotiplerinin Tuza (NaCl) Tolerans Düzeylerinin Belirlenmesi. Selçuk Üniversitesi, Fen Bilimleri Enstitüsü, Bahçe Bitkileri Anabilim Dalı, 2012, 54. Available online: https://tez.yok.gov.tr/UlusalTezMerkezi/tezSorguSonucYeni.jsp (accessed on 9 March 2023).
- Omar, R.A.; Afreen, S.; Talreja, N.; Chauhan, D.; Ashfaq, M. Impact of nanomaterials in plant systems. In Plant Nanobionics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 117–140. [Google Scholar]
- Can, B.; Gürel, A. Nanopartiküllerin bitki sistemlerinde ve bitki doku kültürlerinde uygulamalarına yönelik genel bir bakış. International Journal of Life Sciences and Biotechnology, 2023, 6, 335–370. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Karimi, J.; Mohsenzadeh, S. Effects of silicon oxide nanoparticles on growth and physiology of wheat seedlings. Russ. J. Plant Physiol. 2016, 63, 119–123. [Google Scholar] [CrossRef]
- Şener, S.; Duran, C.N.; Kurt, Z. The effect of silica gel application on plant growth, yield and fruit quality in greenhouse strawberry production. ANADOLU Ege Tarımsal Araştırma Enstitüsü Dergisi 2021, 31, 265–273. [Google Scholar] [CrossRef]
- Parveen, N.; Ashraf, M. Role of silicon in mitigating the adverse effects ofsalt stress on growth and photosynthetic attributes of two maize (Zea mays L.) cultivars grown hydroponically. Pak. J. Bot. 2010, 42, 1675–1684. [Google Scholar]
- El-Kady, M.E.; El-Boray, M.S.; Shalan, A.M.; Mohamed, L.M. Effect of silicon dioxide nanoparticles on growth improvement of banana shoots in vitro within rooting stage. Journal of Plant Production 2017, 8, 913–916. [Google Scholar] [CrossRef]
- Tajima, R. Importance of individual root traits to understand crop root system in agronomic and environmental contexts. Breed Sci. 2021, 71, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.-S.; Jeong, H.-W.; Jo, H.-G.; Kang, J.-H.; Hwang, S.-J. Rooting and growth characteristics of 'Maehyang' strawberry cutting transplants affected by different growing media including decomposed granite. Rhizosphere 2022, 22, 100520. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Kumar, D.S. Effect of nanoparticles on plants with regard to physiological attributes. In Plant Nanotechnology; Springer Science and Business Media LLC: Cham, Switzerland, 2016; pp. 119–153. [Google Scholar]
- Badawy, S.A.; Zayed, B.A.; Bassiouni, S.M.A.; Mahdi, A.H.A.; Majrashi, A.; Ali, E.F.; Seleiman, M.F. Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions. Plants 2021, 10, 1657. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.M.; Qahtan, A.A.; Al-Qurainy, F.; Al-Munqedhi, B.M. Impact of biogenic ag-containing nanoparticles on germination rate, growth, physiological, biochemical parameters, and antioxidants system of tomato (Solanum tuberosum L.) in vitro. Processes 2022, 10, 825. [Google Scholar] [CrossRef]
- Doğan, M. Çilekte Kadmiyum Toksitesi Altındaki Bitkiler Üzerine Hümik Asit ve Silikonun Etkilerinin İncelenmesi. Yüksek Lisans Tezi, Harran Üniversitesi, Fen Bilimleri Enstitüsü, Bahçe Bitkileri Anabilim Dalı, 2018, 46. Available online: https://tez.yok.gov.tr/UlusalTezMerkezi/tezSorguSonucYeni.jsp (accessed on 9 March 2023).
- Wang, C.; He, J.; Zhao, T.-H.; Cao, Y.; Wang, G.; Sun, B.; Yan, X.; Guo, W.; Li, M.-H. The smaller the leaf ıs, the faster the leaf water loses in a temperate forest. Front. Plant Sci. 2019, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Bora, M. Değişik Vejetasyon Dönemlerine Kadar Uygulanan Farklı Tuz Konsantrasyonlarının Biberde Meydana Getirdiği Fizyolojik, Morfolojik ve Kimyasal Değişikliklerin Belirlenmesi. Yüksek Lisans Tezi, Namık Kemal Üniversitesi, Fen Bilimleri Enstitüsü, Tekirdağ. 2014. Available online: https://tez.yok.gov.tr/UlusalTezMerkezi/tezSorguSonucYeni.jsp (accessed on 9 March 2023).
- Bernal, O.B.; Spinoso-Castillo, J.L.; Mancilla-Álvarez, E.; Trujillo, R.A.M.M.; Bello-Bello, J.J. In vitro conservation and regeneration of potato (Solanum tuberosum L.): role of paclobutrazol and silver nanoparticles. Horticulturae 2023, 9, 676. [Google Scholar] [CrossRef]
- Avestan, S.; Ghasemnezhad, M.; Esfahani, M.; Byrt, C.S. Application of nano-silicon dioxide improves salt stress tolerance in strawberry plants. Agronomy 2019, 9, 246. [Google Scholar] [CrossRef]
- Akhoundnejad, Y.; Baran, Ş.; Karakaş, Ö.; Mısırdalı, H. Farklı Dozlardaki Gümüş Nanopartiküllerinin Taze Soğan (Allium Cepa) Üzerine Etkisi. Şırnak Üniversitesi. Şırnak Üniversitesi Fen Bilimleri Dergisi. 2019. Available online: https://dergipark.org.tr/tr/pub/sufbd/issue/46848/551480 (accessed on 9 March 2023).
- Abou-Shlell, M.K.; El-Emary, F.A.; KHalifa, A.A. Effect of nanoparticle on growth, biochemical and anatomical characteristics of moringa plant (Moringa oleifera L.) under salinity stress condition. Archives of Agricultural Sciences Journal 2020, 3, 186–213. [Google Scholar] [CrossRef]
- Almutairi, Z.; Alharbi, A. Effect of silver nanoparticles on seed germination of crop plants. J. Adv. Agric. 2015, 4, 280–285. [Google Scholar] [CrossRef]
- Parveen, N.; Ashraf, M. Role of silicon in mitigating the adverse effects ofsalt stress on growth and photosynthetic attributes of two maize (Zea mays L.) cultivars grown hydroponically. Pak. J. Bot. 2010, 42, 1675–1684. [Google Scholar]
- Bayramzadeh, V.; Ghadiri, M.; Davoodi, M.H. Effects of silver nanoparticle exposure on germination and early growth of Pinus sylvestris and Alnus subcordata. Sains Malays. 2019, 48, 937–944. [Google Scholar] [CrossRef]
- Al-aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. Journal of Plant Nutrition 2005, 27, 2101–2115. [Google Scholar] [CrossRef]
- Kuşvuran, Ş.; Yaşar, F.; Abak, K.; Ellialtıoğlu, Ş. tuz stresi altında yetiştirilen tuza tolerant ve duyarlı Cucumis sp.’nin bazı genotiplerinde lipid peroksidasyonu, klorofil ve iyon miktarlarında meydana gelen değişimler. YY Üniversitesi Tarım Bilimleri Dergisi (J. Agric. Sci.) 2008, 18, 13–20. Available online: https://dergipark.org.tr/tr/pub/yyutbd/issue/21987/236073 (accessed on 09 March 2023).
- Gökçe, Z.Z. Gümüş Nanopartiküllerinin Domates (Lycopersicon Esculentum Mill.) Tohumları Üzerindeki Fizyolojik ve Biyokimyasal Etkilerinin Araştırılması. Yüksek Lisans Tezi, İstanbul Üniversitesi, Fen Bilimleri Enstitüsü, Biyoloji Anabilim Dalı. 2019. Available online: https://tez.yok.gov.tr/UlusalTezMerkezi/tezSorguSonucYeni.jsp (accessed on 9 March 2023).
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of 'Katahdin'-derived potato cultivars. Breeding science 2016, 66, 328–331. [Google Scholar] [CrossRef]
- Fan, Y.; Shen, W.; Vanessa, P.; Cheng, F. Synergistic effect of Si and K in improving the growth, ion distribution and partitioning of Lolium perenne L. under saline-alkali stress. Journal of Integrative Agriculture. 2021, 20, 1660–1673. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Scientific reports 2020, 10, 17672. [Google Scholar] [CrossRef]
- Syu, Y.Y.; Hung, J.H.; Chen, J.C.; Chuang, H.W. Impact of size and shape of silver nanoparticles on Arabidopsis plant growth and gene expression. Plant Physiol. Biochem. 2014, 83, 57–64. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Gurunathan, S.; Chung, I.-M. Physiological, metabolic, and transcriptional effects of biologically-synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma 2015, 252, 1031–1046. [Google Scholar] [CrossRef]
- Landa, P.; Prerostova, S.; Petrova, S.; Knirsch, V.; Vankova, R.; Vanek, T. The transcriptomic response of Arabidopsis thaliana to zinc oxide: A comparison of the impact of nanoparticle, bulk, and ionic zinc. Environ. Sci. Technol. 2015, 49, 14537–14545. [Google Scholar] [CrossRef]
- Ragab, S.M.; Turoop, L.; Runo, S.; Nyanjom, S. The effect of foliar application of zinc oxide nanoparticles and Moringa oleifera leaf extract on growth, biochemical parameters and in promoting salt stress tolerance in faba bean. Afr. J. Biotechnol. 2022, 21, 252–266. [Google Scholar]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.F.; Abd El-Mageed, T.A.; Abd El-Mageed, S.A.; Rady, M.M.; Ali, E.F. Foliar application of zinc oxide nanoparticles promotes drought stress tolerance in eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-S.; Li, K.; Wang, Q.-M.; Song, X.-Y.; Su, H.-N.; Xie, B.-B.; Zhang, X.-Y.; Huang, F.; Bai-Cheng, Z.; Zhou, B.-C.; et al. Nitrogen starvation impacts the photosynthetic performance of porphyridium cruentum as revealed by chlorophyll a fluorescence. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pommerening, A.; Muszta, A. Methods of modelling relative growth rate. For. Ecosyst. 2015, 2, 5. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Firoz, M.; Al-Khaishany, M.Y. Role of Nanoparticles in Plants. In Nanotechnology and Plant Sciences; Siddiqui, M., Al-Whaibi, M., Mohammad, F., Eds.; Springer: Cham, 2015. [Google Scholar] [CrossRef]
- Elatafi, E.; Fang, J. Effect of Silver Nitrate (AgNO3) and Nano-Silver (Ag-NPs) on physiological characteristics of grapes and quality during storage period. Horticulturae 2022, 8, 419. [Google Scholar] [CrossRef]
- Zhao, L.; Peng, B.; Hernandez-Viezcas, J.A.; Rico, C.; Sun, Y.; Peralta-Videa, J.R.; Tang, X.; Niu, G.; Jin, L.; Varela-Ramirez, A.; et al. Stress response and tolerance of Zea mays to CeO2 nanoparticles: Cross talk among H2O2, Heat Shock Protein and Lipid Peroxidation. ACS Nano 2012, 6, 9615–9622. [Google Scholar] [CrossRef]
- Günalp, B. Patlıcan (Solanum melongena L.) Embriyo Kültüründe, Jasmonik Asit ve Tuz Stresi Etkileşiminin İncelenmesi. Ankara Üniversitesi Fen Bilimleri Enstitüsü (Basılmamış), Yüksek Lisans Tezi. Ankara, 2011, 77. Available online: https://tez.yok.gov.tr/UlusalTezMerkezi/tezSorguSonucYeni.jsp (accessed on 9 March 2023).
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef]
Figure 1.
Field Emission Scanning Electron Microscope SEM image of (a); SiNP, (b); FeNP, (c); AgNP.

Figure 2.
Micropropagation stages of boysenberry plants: (a) pre-sterilization; (b) surface sterilization; (c) appearance of explants 4 weeks after culture; (d) in vitro plants.
Figure 2.
Micropropagation stages of boysenberry plants: (a) pre-sterilization; (b) surface sterilization; (c) appearance of explants 4 weeks after culture; (d) in vitro plants.

Figure 3.
in vitro plants prepared for measurements and analyses.

Figure 4.
Shoot development of in vitro plants according to different types and concentrations of NP applications (a) SFW; (b) SDW; (c); SL, and NaCL stress: (d) SD; (e) SFW; (f) SDW; (g); SL; (h) SD.
Figure 4.
Shoot development of in vitro plants according to different types and concentrations of NP applications (a) SFW; (b) SDW; (c); SL, and NaCL stress: (d) SD; (e) SFW; (f) SDW; (g); SL; (h) SD.
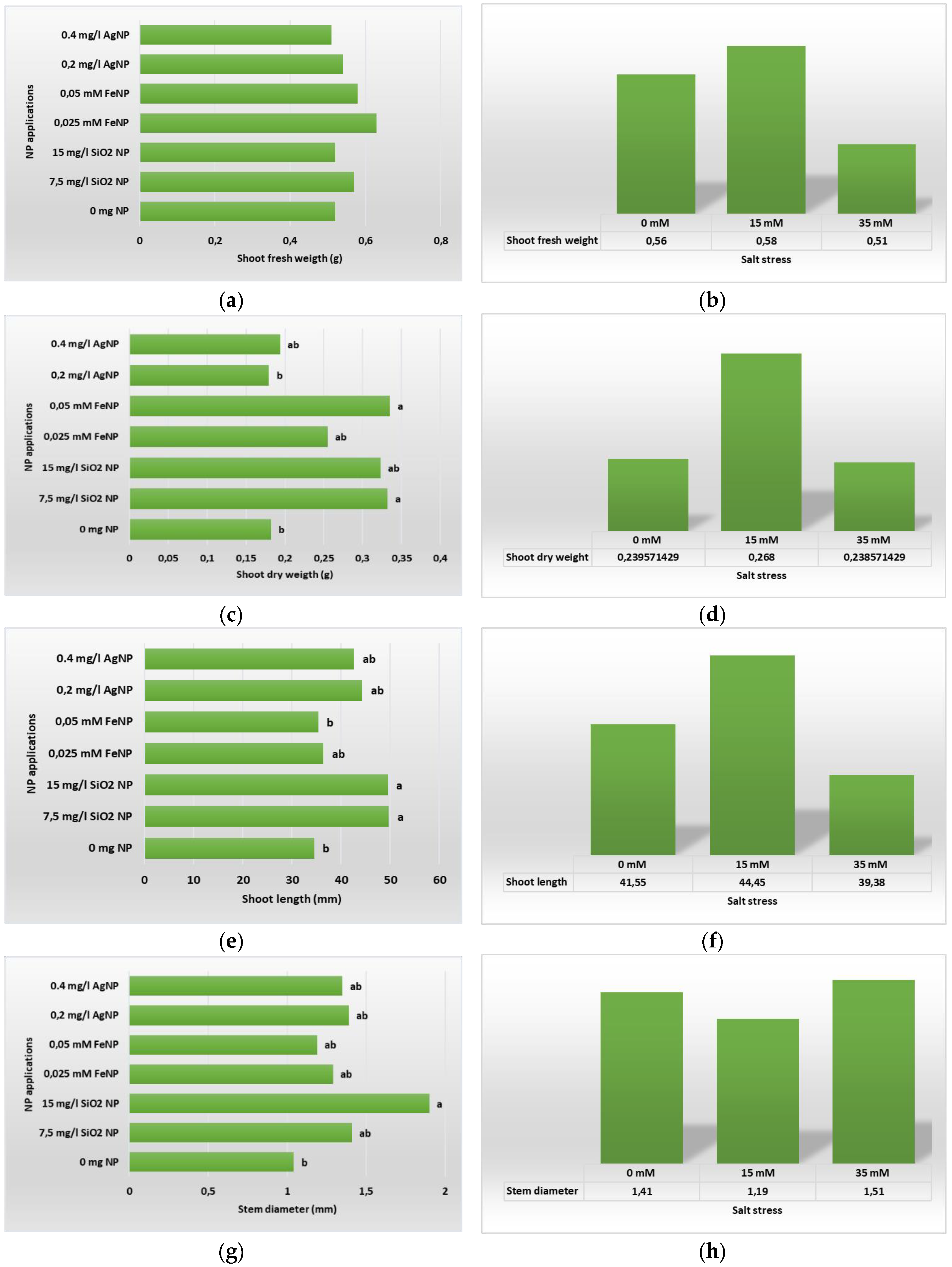
Figure 5.
Root development of in vitro plants according to different types and concentrations of NP applications (a) RFW; (b) RDW; (c); RL, and NaCL stress: (d) RFW; (e) RDW; (f) RL.
Figure 5.
Root development of in vitro plants according to different types and concentrations of NP applications (a) RFW; (b) RDW; (c); RL, and NaCL stress: (d) RFW; (e) RDW; (f) RL.
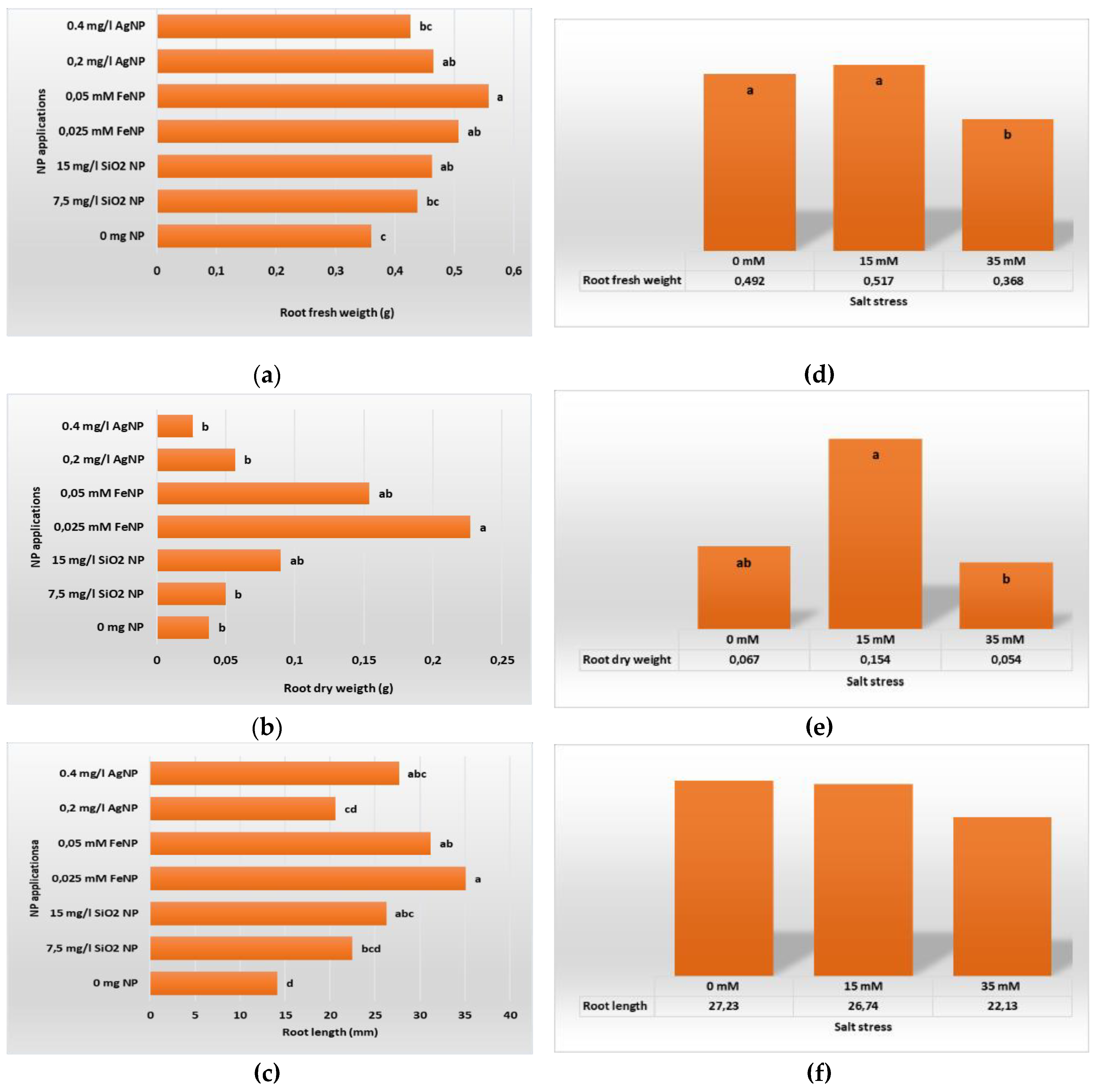
Figure 6.
Leaf development of in vitro plants according to different types and concentrations of NP applications (a) NL; (b) LL; (c); LW, and NaCL stress: (d) NL; (e) LL; (f) LW.
Figure 6.
Leaf development of in vitro plants according to different types and concentrations of NP applications (a) NL; (b) LL; (c); LW, and NaCL stress: (d) NL; (e) LL; (f) LW.
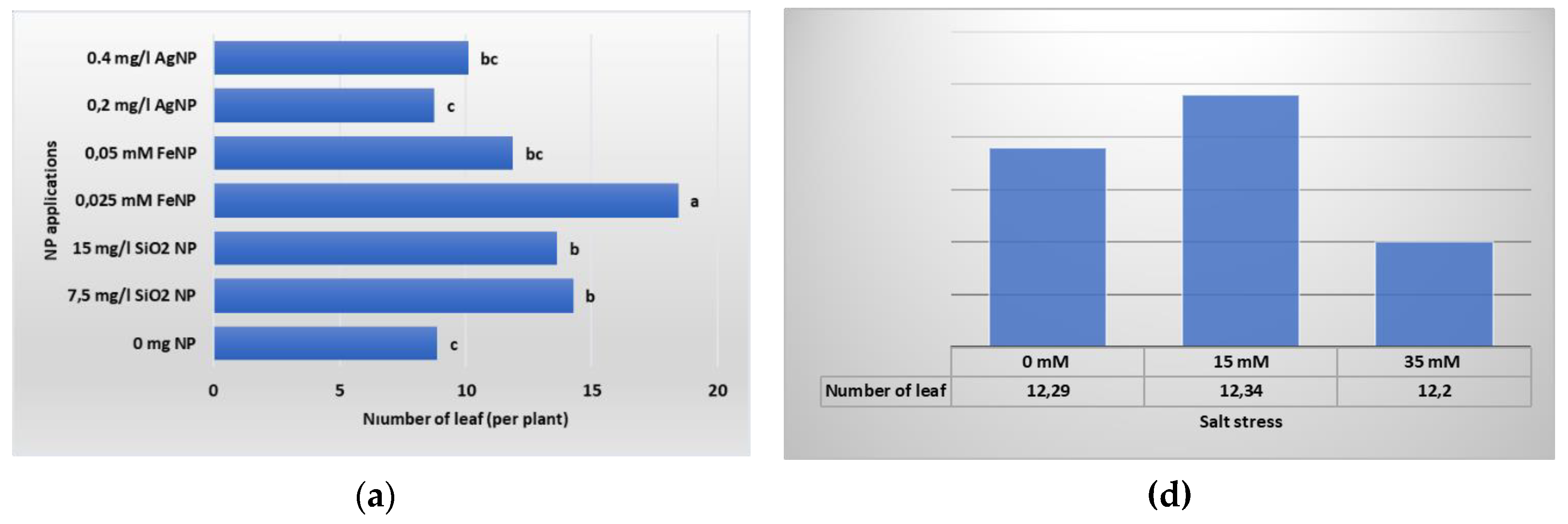
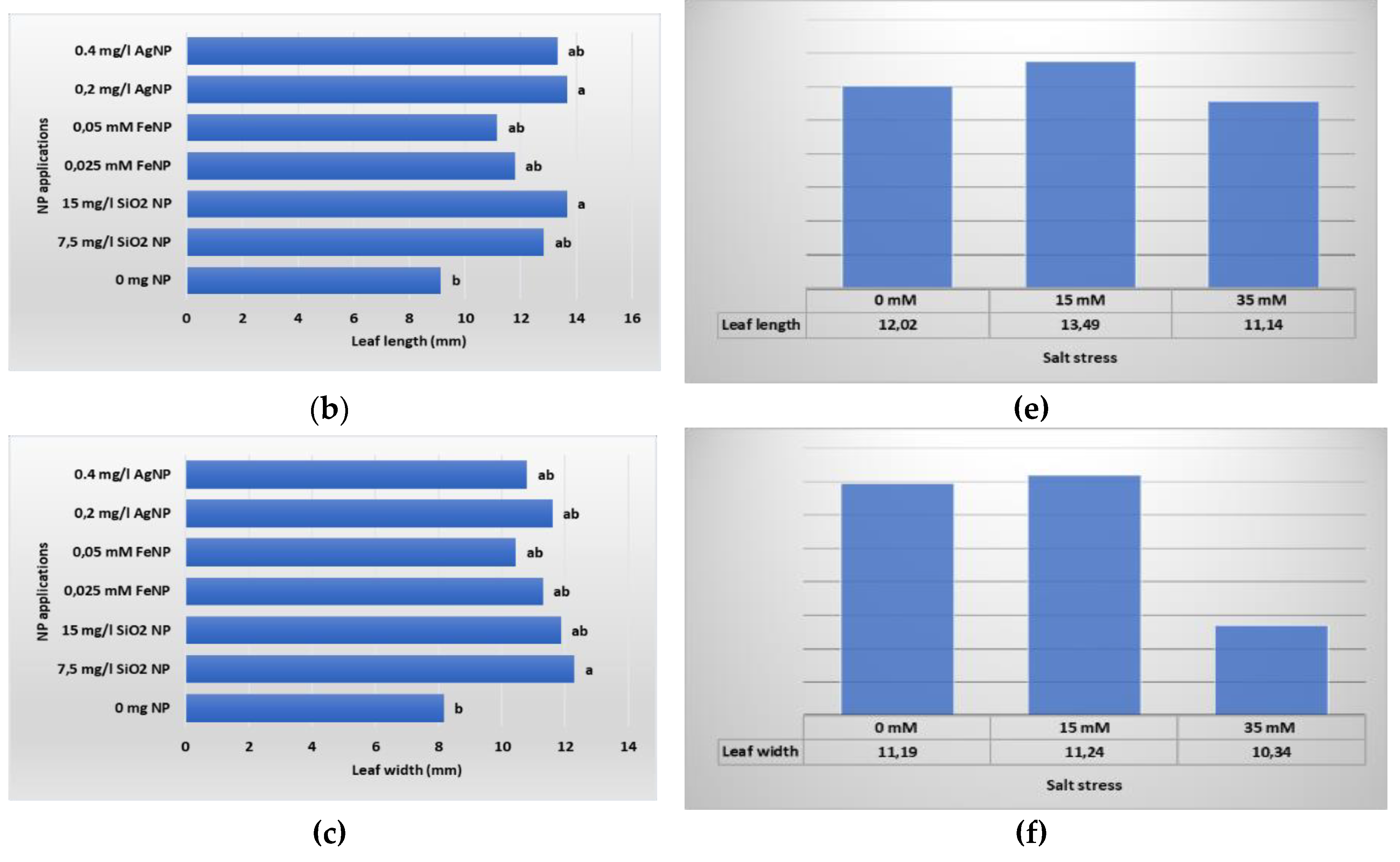
Figure 7.
Physiological growth parameters of in vitro plants according to different types and concentrations of NP applications (a) SPAD value; (b) LRWC; (c); RGR, and NaCL stress: (d) SPAD value; (e) LRWC; (f) RGR.
Figure 7.
Physiological growth parameters of in vitro plants according to different types and concentrations of NP applications (a) SPAD value; (b) LRWC; (c); RGR, and NaCL stress: (d) SPAD value; (e) LRWC; (f) RGR.
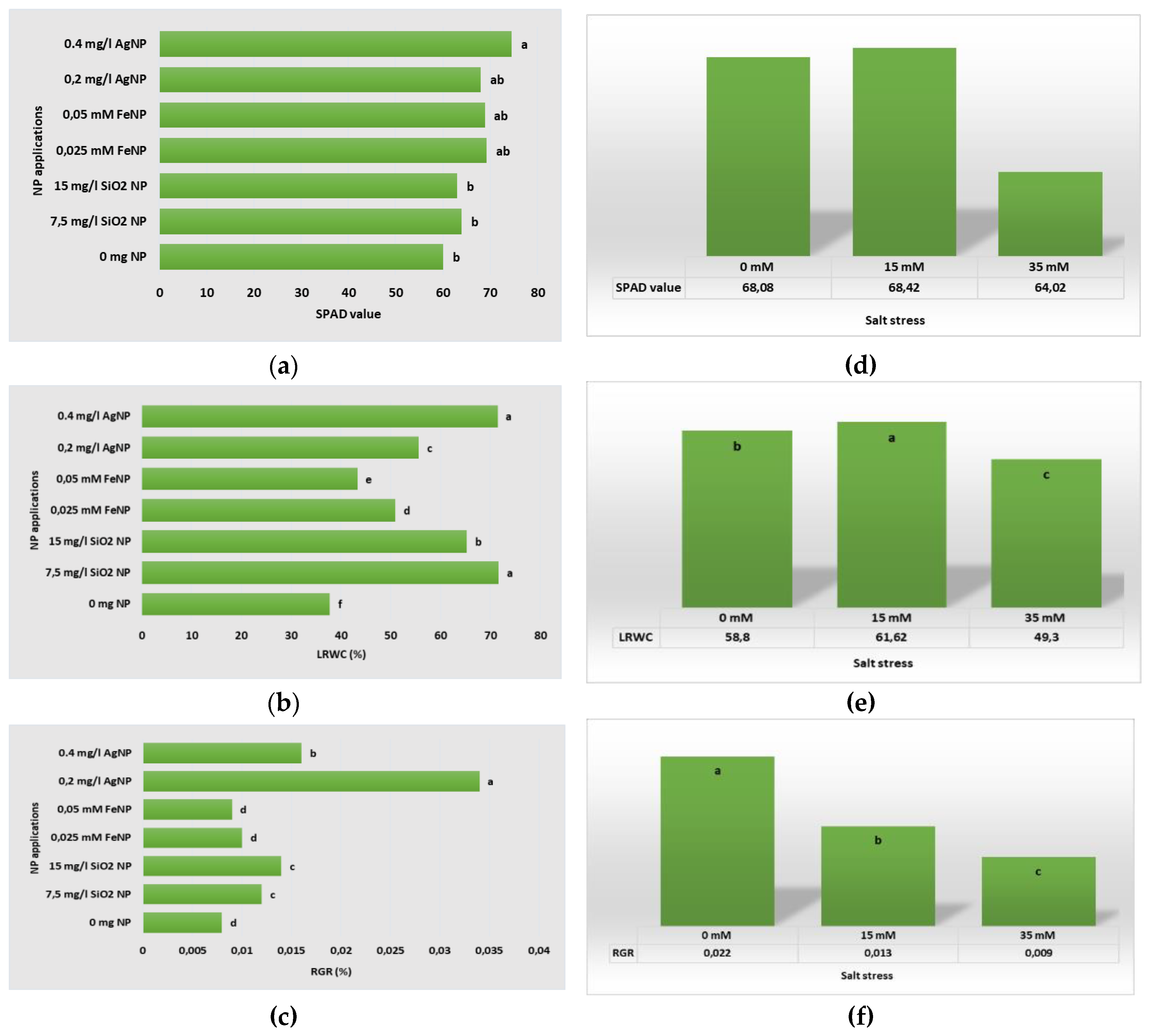
Figure 8.
CAT and SOD enzyme activity of in vitro plants according to different types and concentrations of NP applications (a) CAT; (b) SOD, and NaCL stress: (c) CAT; (d) SOD.
Figure 8.
CAT and SOD enzyme activity of in vitro plants according to different types and concentrations of NP applications (a) CAT; (b) SOD, and NaCL stress: (c) CAT; (d) SOD.
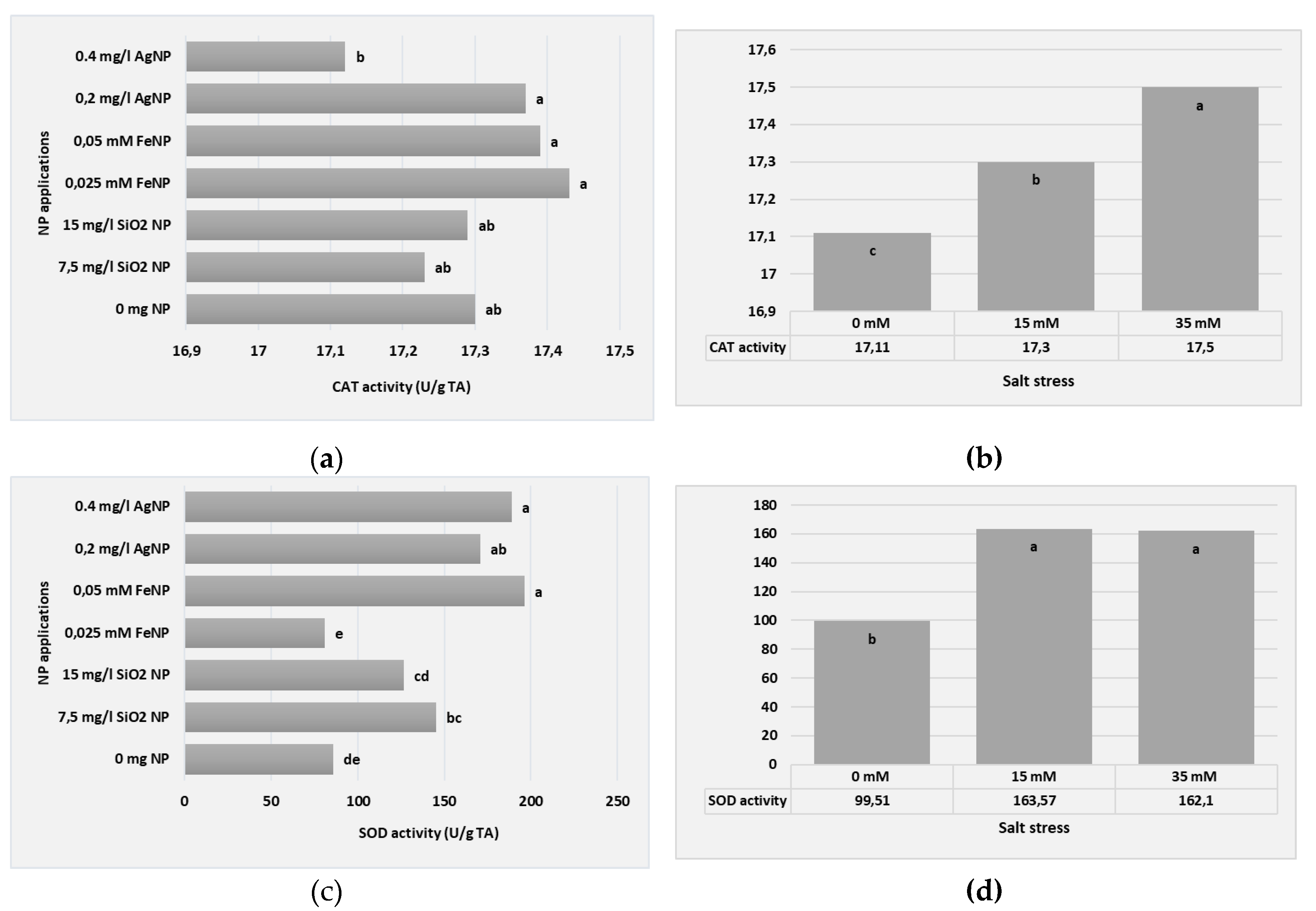

Table 1.
Medias used in the research.
| Media | Applications |
|---|---|
| MS1 | 0 mM NaCl |
| MS2 | 15 mM NaCI |
| MS3 | 35 mM NaCI |
| MS4 | 7.5 mg L-1 SiNP |
| MS5 | 15 mM NaCI + 7.5 mg L-1 SiNP |
| MS6 | 35 mM NaCI +7.5 mg L-1 SiNP |
| MS7 | 15 mg L-1 SiNP |
| MS8 | 15 mM NaCI + 15 mg L-1 SiO2 NP |
| MS9 | 35 mM NaCI + 15 mg L-1 SiO2 NP |
| MS10 | 0.025 mM FeNP |
| MS11 | 15 mM NaCI + 0.025 mM FeNP |
| MS12 | 35 mM NaCI + 0.025 mM FeNP |
| MS13 | 0.05 mM FeNP |
| MS14 | 15 mM NaCI + 0.05 mM FeNP |
| MS15 | 35 mM NaCI + 0.05 mM FeNP |
| MS16 | 0.2 mg L-1 Ag |
| MS17 | 15 mM NaCI + 0.2 mg L-1 Ag |
| MS18 | 35 mM NaCI + 0.2 mg L-1 Ag |
| MS19 | 0.4 mg L-1 Ag |
| MS20 | 15 mM NaCI + 0.4 mg L-1 Ag |
| MS21 | 35 mM NaCI + 0.4 mg L-1 Ag |
Table 2.
Shoot growth parameters of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
Table 2.
Shoot growth parameters of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
| NP Applications | NaCl Concentrations | SFW (g) | SDW (g) | SL (mm) | SD (mm) |
|---|---|---|---|---|---|
| 0 mg L-1 NP | 0 mM | 0.423 cd | 0.137 c | 33.62 b | 0.95 b |
| 15 mM | 0.617 abcd | 0.186 ab | 40.42 ab | 1.06 ab | |
| 35 mM | 0.526 bcd | 0.258 ab | 29.56 b | 1.12 ab | |
| 7.5 mg L-1 SiNP | 0 mM | 0.614 abcd | 0.137 b | 46.14 ab | 1.21 ab |
| 15 mM | 0.487 bcd | 0.180 ab | 51.30 ab | 0.83 b | |
| 35 mM | 0.610 abcd | 0.230 ab | 51.71 ab | 2.20 ab | |
| 15 mg L-1 SiNP | 0 mM | 0.651 abc | 0.356 ab | 52.19 ab | 2.46 a |
| 15 mM | 0.537 bcd | 0.363 ab | 47.39 ab | 1.65 ab | |
| 35 mM | 0.376 d | 0.250 ab | 48.88 ab | 1.60 ab | |
| 0.025 mM FeNP | 0 mM | 0.516 bcd | 0.132 b | 35.19 b | 1.39 ab |
| 15 mM | 0.841 a | 0.446 a | 35.01 b | 1.29 ab | |
| 35 mM | 0.522 bcd | 0.355 ab | 39.09 ab | 1.19 ab | |
| 0.05 mM FeNP | 0 mM | 0.688 ab | 0.453 a | 49.46 ab | 1.18 ab |
| 15 mM | 0.565 bcd | 0.293 ab | 29.78 b | 1.09 ab | |
| 35 mM | 0.477 bcd | 0.328 ab | 27.08 b | 1.29 ab | |
| 0.2 mg L-1AgNP | 0 mM | 0.537 bcd | 0.168 b | 29.41 b | 1.60 ab |
| 15 mM | 0.519 bcd | 0.232 ab | 61.10 a | 1.14 ab | |
| 35 mM | 0.572 bcd | 0.136 b | 42.47 ab | 1.42 ab | |
| 0.4 mg L-1AgNP | 0 mM | 0.478 bcd | 0.287 ab | 44.83 ab | 1.07 ab |
| 15 mM | 0.528 bcd | 0.176 ab | 46.14 ab | 1.24 ab | |
| 35 mM | 0.520 bcd | 0.120 b | 36.88 ab | 1.73 ab |
* Each value is the mean of 3 replicated samples of 10 plants each. For each factor, values in a column followed by the same letter are not significantly different, according to the Duncan test, significant at p < 0.05.
Table 3.
Root growth parameters of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
Table 3.
Root growth parameters of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
| NP Applications | NaCl Concentrations | RFW (g) | RDW (g) | RL (mm) |
|---|---|---|---|---|
| 0 mg L-1 NP | 0 mM | 0.495 abcde | 0.066 b | 15.64 ef |
| 15 mM | 0.483 abcde | 0.031 b | 15.76 ef | |
| 35 mM | 0.103 f | 0.018 b | 11.09 ef | |
| 7.5 mg L-1 SiNP | 0 mM | 0.513 abcd | 0.117 b | 47.16 ab |
| 15 mM | 0.429 abcde | 0.023 b | 11.87 ef | |
| 35 mM | 0.371 cde | 0.010 b | 8.50 f | |
| 15 mg L-1 SiNP | 0 mM | 0.475 abcde | 0.042 b | 29.28 bcde |
| 15 mM | 0.508 abcd | 0.120 b | 19.47 def | |
| 35 mM | 0.402 bcde | 0.107 b | 30.10 bcde | |
| 0.025 mM FeNP | 0 mM | 0.355 de | 0.099 b | 36.89 abcd |
| 15 mM | 0.613 a | 0.571 a | 54.07 a | |
| 35 mM | 0.552 abc | 0.012 b | 14.34 ef | |
| 0.05 mM FeNP | 0 mM | 0.596 bc | 0.076 b | 15.56 ef |
| 15 mM | 0.587 ab | 0.246 b | 36.11 abcd | |
| 35 mM | 0.489 abcde | 0.141 b | 41.95 abc | |
| 0.2 mg L-1AgNP | 0 mM | 0.529 abcd | 0.043 b | 21.80 def |
| 15 mM | 0.519 abcd | 0.057 b | 19.37 def | |
| 35 mM | 0.347 de | 0.070 b | 20.72 def | |
| 0.4 mg L-1AgNP | 0 mM | 0.483 abcde | 0.027 b | 24.28 cdef |
| 15 mM | 0.452 abcde | 0.029 b | 30.53 bcde | |
| 35 mM | 0.313 e | 0.021 b | 28.20 bcde |
* Each value is the mean of 3 replicated samples of 10 plants each. For each factor, values in a column followed by the same letter are not significantly different, according to the Duncan test, significant at p < 0.05.
Table 4.
Leaf development of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
Table 4.
Leaf development of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
| NP Applications | NaCl Concentrations | NL (per plant) | LL (mm) | LW (mm) |
|---|---|---|---|---|
| 0 mg L-1 NP | 0 mM | 8.75 cdef | 14.85 abc | 12.85 ab |
| 15 mM | 13.25 bcde | 7.19 bcd | 6.43 bc | |
| 35 mM | 4.50 f | 5.34 e | 5.21 c | |
| 7.5 mg L-1 SiNP | 0 mM | 12.50 bcdef | 11.44 abcd | 11.54 abc |
| 15 mM | 17.50 ab | 13.26 abcd | 11.75 abc | |
| 35 mM | 12.75 bcdef | 10.66 abcd | 10.60 abc | |
| 15 mg L-1 SiNP | 0 mM | 10.75 bcdef | 14.56 abc | 13.26 ab |
| 15 mM | 12.75 bcdef | 13.66 abc | 10.75 abc | |
| 35 mM | 17.25 abc | 12.75 abcd | 11.62 abc | |
| 0.025 mM FeNP | 0 mM | 23.25 a | 14.24 abc | 13.34 ab |
| 15 mM | 17.50 ab | 13.82 abc | 11.93 abc | |
| 35 mM | 14.50 bcd | 10.43 bcd | 11.59 abc | |
| 0.05 mM FeNP | 0 mM | 10.75 bcdef | 6.61 cd | 7.58 abc |
| 15 mM | 11.00 bcdef | 12.46 abcd | 11.84 abc | |
| 35 mM | 13.75 bcde | 14.40 abc | 11.89 abc | |
| 0.2 mg L-1AgNP | 0 mM | 8.25 def | 12.32 abcd | 10.65 abc |
| 15 mM | 9.00 bcdef | 18.70 a | 14.46 a | |
| 35 mM | 9.00 bcdef | 9.97 bcd | 9.70 abc | |
| 0.4 mg L-1AgNP | 0 mM | 11.50 bcdef | 10.14 bcd | 9.08 abc |
| 15 mM | 5.25 ef | 15.37 ab | 11.55 abc | |
| 35 mM | 13.50 bcde | 14.45 abc | 11.77 abc |
* Each value is the mean of 3 replicated samples of 10 plants each. For each factor, values in a column followed by the same letter are not significantly different, according to the Duncan test, significant at p < 0.05.
Table 5.
Physiological growth parameters of boysenberry plants grown in vitro conditions containing different levels of NaCl and NPs.
Table 5.
Physiological growth parameters of boysenberry plants grown in vitro conditions containing different levels of NaCl and NPs.
| NP Applications | NaCl Concentrations | SPAD value | LRWC (%) | RGR (%) |
|---|---|---|---|---|
| 0 mg L-1 NP | 0 mM | 65.35 cd | 46.05 g | 0.011 ef |
| 15 mM | 87.48 ab | 32.06 ı | 0.008 fgh | |
| 35 mM | 55.04 d | 35.23 hı | 0.006 h | |
| 7.5 mg L-1 SiNP | 0 mM | 69.16 cd | 58.33 ef | 0.020 d |
| 15 mM | 63.98 cd | 92.87 a | 0.008 fgh | |
| 35 mM | 58.62 d | 63.92 d | 0.008 fgh | |
| 15 mg L-1 SiNP | 0 mM | 58.52 d | 54.35 ef | 0.024 c |
| 15 mM | 58.30 d | 52.71 f | 0.011 ef | |
| 35 mM | 72.36 bcd | 45.67 g | 0.007 gh | |
| 0.025 mM FeNP | 0 mM | 78.70 abc | 62.83 d | 0.013 e |
| 15 mM | 64.95 cd | 78.89 c | 0.010 efg | |
| 35 mM | 63.12 cd | 53.33 e | 0.006 h | |
| 0.05 mM FeNP | 0 mM | 65.81 cd | 52.74 f | 0.007 gh |
| 15 mM | 57.58 d | 32.25 ı | 0.011 ef | |
| 35 mM | 56.73 d | 44.95 g | 0.008 fgh | |
| 0.2 mg L-1AgNP | 0 mM | 69.85 cd | 52.59 f | 0.056 a |
| 15 mM | 57.03 d | 77.42 c | 0.034 b | |
| 35 mM | 77.25 abc | 36.75 h | 0.013 e | |
| 0.4 mg L-1AgNP | 0 mM | 69.20 cd | 84.71 b | 0.025 c |
| 15 mM | 89.63 a | 65.17 d | 0.010 efg | |
| 35 mM | 65.00 cd | 64.60 d | 0.013 e |
* Each value is the mean of 3 replicated samples of 10 plants each. For each factor, values in a column followed by the same letter are not significantly different, according to the Duncan test, significant at p < 0.05.
Table 6.
CAT and SOD enzyme activity of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
Table 6.
CAT and SOD enzyme activity of boysenberry plants grown in vitro conditions containing different type and concentrations of NP and NaCl applications.
| NP Applications | NaCl Concentrations | CAT activity (U/g TA) | SOD activity (U/g TA) |
|---|---|---|---|
| 0 mg L-1 NP | 0 mM | 16.81 h | 59.20 h |
| 15 mM | 17.42 bcd | 96.34 fgh | |
| 35 mM | 17.67 ab | 101.54 fgh | |
| 7.5 mg L-1 SiNP | 0 mM | 17.27 cdef | 89.19 fgh |
| 15 mM | 16.98 fgh | 146.07 defg | |
| 35 mM | 17.43 bcd | 200.22 bcd | |
| 15 mg L-1 SiNP | 0 mM | 16.86 gh | 53.93 h |
| 15 mM | 17.57 abc | 263.49 ab | |
| 35 mM | 17.45 abcd | 61.85 h | |
| 0.025 mM FeNP | 0 mM | 17.44 bcd | 94.61 fgh |
| 15 mM | 17.37 bcde | 101.71 fgh | |
| 35 mM | 17.45 abcd | 68.23 gh | |
| 0.05 mM FeNP | 0 mM | 16.99 fgh | 112.98 efgh |
| 15 mM | 17.42 bcd | 162.75 cdef | |
| 35 mM | 17.82 a | 311.55 a | |
| 0.2 mg L-1AgNP | 0 mM | 17.38 bcde | 88.04 fgh |
| 15 mM | 17.23 cdefg | 239.31 abc | |
| 35 mM | 17.50 abcd | 184.79 bcde | |
| 0.4 mg L-1AgNP | 0 mM | 17.03 efgh | 198.63 bcd |
| 15 mM | 17.13 defgh | 155.70 def | |
| 35 mM | 17.19 cdefg | 212.81 bcd |
* Each value is the mean of 3 replicated samples of 10 plants each. For each factor, values in a column followed by the same letter are not significantly different, according to the Duncan test, significant at p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



