Submitted:
12 September 2024
Posted:
14 September 2024
You are already at the latest version
Abstract
Context: The advent of new strains of resistant microbes and the concomitant growth in multidrug resistance have made antimicrobial resistance an urgent public health concern. New antimicrobials are desperately needed to boost the success rates of treating infectious diseases and save lives. There are many intriguing biomolecules with antibacterial action, which are mostly unexplored in microorganisms. Objectives: The objective of the present review article is to describe the importance of natural compounds against microorganisms using advanced techniques to protect people from diseases. Method: We have searched and incorporated many online literature to gather information from SCOPUS, SCI, PUBMED, ScienceDirect and Medline. We have discussed several microorganism sources for antimicrobials and antifungal drugs. In addition, describes microculturing techniques and applications of microbial-based microsystems. Results and Conclusion: Every kind of higher trophic life depends on microorganisms for sustenance. The unseen majority is essential to understanding how humans and other living forms can survive anthropogenic climate change. Antimicrobial substances and the latest methods, techniques, and strategies for developing effective treatments have been discussed in this article. Novel model systems and cutting-edge biomolecular and computational methodologies could help researchers enhance antimicrobial resistance by completely capitalizing on lead antimicrobials.
Keywords:
drug discovery
; microorganism
; natural products
; structure-activity
; antimicrobial resistance
1. Introduction
Multidrug-resistant organisms and antimicrobial-resistant diseases threaten the healthcare system and public health. Thousands die annually from resistant diseases (O'Neill et al., 2014). As antimicrobial resistance increases, their effectiveness becomes increasingly uncertain [2]. Consequently, it is crucial to investigate potential replacements for current antimicrobials. They account for over two-thirds of all newly authorized pharmaceutical applications [3].
In contrast to antibiotics produced by microorganisms, plant-based antimicrobials have attracted many researchers due to their promising effects with fewer side effects. Bioactive chemicals with antibacterial, antifungal, and cytotoxic bioactivity are produced by microorganisms [4,5,6,7]. Once again, their ability to produce functionally rich secondary metabolites contributes to their success in various habitats. Due to their unique biological features, microorganisms have recently attracted researchers' interest as a potential source of new antimicrobial drugs [8]. Modern molecular biology, genetic, genomic, and computational approaches make mining microbial structure systems for drug discovery easier [9,10,11].
Viruses, bacteria, archaea, fungi, and protists are all examples of microorganisms, which are biotic, ubiquitous, diverse entities. Novel antimicrobial agents are primarily discovered in bacteria and fungi. Clinically resistant germs like S. aureus, MRSA, M. luteus, B. subtilis, and E. faecalis are toxic to cyclic peptides, polyketides, and alkaloids found in bacteria, such as mathiapeptide A, destotamide B, Marfomycins A, B, and E; spirotetronates polyketides, abyssomycin C, Lobophorin F, and H; and sesquiterpene derivatives, alkaloids, and mafuraquinocins A and D,which are derived from bacterial sources (Table 1).
Inhibitors of Gram-negative and Gram-positive pathogenic bacteria include fungus-derived xanthones, emerixanthones, and engyodontiumsones [12]. Analogues of ambuic acid, such as penicyclone and depsidone, have this property. Yeast [16,17], microalgae[14,15], and cyanobacteria[13,14] are among the other groups whose compounds have demonstrated anti-infective potential in both animal and laboratory studies.
In order to combat the rise of antibiotic-resistant bacteria, modern scientific research has prepared the way for the development of synthetic antimicrobials by altering the chemical and structural makeup of naturally occurring goods. Novel antimicrobials have been fabricated and visualized using X-ray crystallography, structured-guided designs, and components based on primogenitor cell lines [18]. Oxepanoprolinamide is a derivative of lincosamide that has a higher therapeutic efficacy against resistant bacterial strains and an increased probability of overcoming multidrug resistance caused by ATP binding cassette (ABC) F-, Erm-, and Cfr genes [18]. Multiple resistant microbial strains have emerged recently, and new technologies have become available (Figure 1).
2. Bacterial Sources of Antimicrobials
Bacteriocin, organic acids, diacetyl, hydrogen peroxide, and other antimicrobials are produced by lactic acid bacteria (LAB). The growth of some bacteria can be inhibited by the bacteriocin produced by the boza-isolated Lactobacillus pentosus ST712BZ [19,20]. Bacteriocin was discovered by Andre Gratias in 1925 when he realized that colicin V hindered E. coli growth [21].
Klaenhammer categorized bacteriocins based on their molecular weight, susceptibility to enzymes, capacity to withstand high temperatures, presence of modified amino acids after translation, and their mechanism of action. [22]. Class I separates lantibiotics into Ia and Ib by chemical structure and charge. Bacteriocin classes IIa, IIb, and IIc exist [22]. Some heat-stable peptides weigh less than 10 kDa. Class IV consists of large peptides and carbohydrates or lipids, whereas Class III includes thermo-labile peptides with a high molecular weight, often exceeding 30 kDa, such as Helveticin J. [23]. Despite competing theories by researchers such as Cotter et al. [24] and Drider et al. [25], there is still no one-size-fits-all solution.
There are currently three main categories into which bacteriocins are classified based on their biochemical and genetic characteristics. Everything else is in Class III, Class II is non-lantibiotic, and Class I contains lantibiotics [25]. The antimicrobial effect of bacteriocins is determined by their main structure [26, [Table 2]. Some bacteriocins enter cells and interfere with protein synthesis and gene expression [26], while others interfere with the proton motive force of electrified membrane vesicles [27]. Antibiotics have a dual action against bacteria [26,28,29].
When non-lantibiotics bind to the mannose phosphotransferase permease (Man-PTS) subunits C and D, they establish an intra-membrane channel that allows ions to move, ultimately leading to cell death [29,30]. Because of their high net positive charges, circular bacteriocins electrostatically attract bacterial membranes that are negatively charged. Cell death, alterations to the membrane potential, ion efflux, and holes are all results of this interaction [31]. The cell wall hydrolyzes more quickly due to bacteriolysins, which gradually causes the cell to disintegrate [32,33]. Target cells' ability to take up glucose is inhibited by bacteriolysis, leading to their death from starvation [34,35,36].
Synergistic or antagonistic interactions can occur between antimicrobial drugs and the microorganisms they are effective against [37]. Nisin and other bancteriocins kill E. faecalis and other bacteria that cause periodontal disease, preventing gingivitis and dentobacterial plaque in dogs [26]. Some Pseudomonas species produce the anionic biosurfactant rhamnolipid [38,39]. Antimicrobial, antibiofilm, antitumor, and antioxidative properties have all been observed for these compounds [40,41,42].
They can modify viral membrane glycoproteins and interact with viral lipid membranes, especially in light of their use in combating bovine coronaviruses, HSV-1 and HSV-2 [43,44]. The rhamnolipids (M15RL) produced by the Antarctic bacterium Pseudomonas gessardii (P.gessardii) M15 have been shown to inhibit SARS-CoV-2 [45].
3. Bacterial Sources of Antifungal Compounds
The red pradimicins A, B, and C are produced by Actinomadura hibisca [56]. These pradimicins are effective against Aspergillus and Candida [57, [Table 3]. Pratimimins are benzonapthacenequinones that transport D-alanine and carbohydrates, as revealed by spectral analysis and chemical breakdown [56]. By forming a compound with the cell walls of vulnerable microbes, pradimicins damage fungal cells by binding to terminal D mannosides [57]. Many species of Actinoplanes create antifungal metabolites as well.
The Actinoplanes ianthinogenes-produced purpuromycin kills Trycophyton mentagrophytes [58]. Produced by Octamycini [58]. Streptomyces species produce branching bacilli that are aerobic and Gram-positive Chemical antifungals. The chemicals nystatin, phoslatomycins, phthoxazolin A, faeriefungin, butyrolactols, sultriecin, polyoxin, and dunaimycins are examples.
Aspergillus species, Saccharomyces spp., and Candida albicans can be effectively targeted by azobacilin, bacereutin, cispentacin, and mycocerein, which are generated from Bacillus cereus compounds, as stated by Kerr. [58] Bacillus licheniformis produces both the peptide A12-C and fungicin M-4. [63,64]. Enterobacter agglomerans becomes antibacterial against Aspergillus niger, Candida species, phytopathogenic fungi, and dermatophytes, when treated with pyrrolnitrin, according to research by Chernin et al.
A and B, two antifungal herbicolins, are again produced by Enterobacter agglomerans [65,66,67]. The dipeptide CB-25-1 from Serratia plymuthica [68], Dihydroaeruginoic acid [69], pyocyanin [70], and 1-hydroxyphenazine [71] are compounds created by P. aeruginosa, which is present in a healthy human stomach. Pseudomonas also produces the peptide family pseudomycin [72], the caryoynencins [73], and the cyclic hydroxamic acid, G1549 [74], among other antibacterial chemicals.
Antimicrobial chemicals can also be found in Burkholderia species, which are bacteria [75]. Cepalycin [76], xylocandins [77], and heptylmethylquinolinone [78] are some of the other compounds that B. cepacia can create. The antibacterial enacyloxcins are thought to have originated in the Burkholderia species [79]. There are eight different antibacterial chemicals in the enacyloxcin family [76,77]. Maltophilin gives the Rhizosphere strain of Stenotrophomonas maltophilia antifungal characteristics. Antifungal action against Neurospora crassa, but not yeast, has been observed for polyenic antibiotics from the genus Gluconobacter [80].
4. Fungal Sources of Antimicrobials
Since penicillin G was first isolated from a fungus species in 1928 [81], because of their prolific secondary metabolite production, they are promising candidates for use as starting materials in developing new antimicrobial drugs. Enfumafungin, a triterpenoid extracted from a Hormonema species, was first identified more than ten years ago and has since been demonstrated to be particularly efficient against Candida spp. and Aspergillus spp. [82]. Phase II clinical trials are underway for SCY-078, a semisynthetic derivative of enfumafungin [83,84].
However, in recent years, favolon, a metabolite of strobilurins generated by Favolaschia calocera (F. calocera), has been found and demonstrated to be less toxic while still displaying substantial antifungal action against human pathogens [85]. By disrupting quorum sensing, fungal metabolites prevent biofilm formation. Coprinuslactone, isolated from C. comatus, inhibits P. aeruginosa biofilm formation [86]. A Kenyan basidiomycete's microporenic acid A is lethal to biofilms of Staphylococcus aureus and Candida albicans [87]. Antibiotics that also work against biofilms hold great promise.
Influenza Rubylide S, produced by the marine fungus Aspergillus terreus OUCMDZ-1925, inhibited the activity of the H1N1 virus [88]. Cladosporium sphaerospermum 2005-01-E3 yielded a new hybrid polyketide called Cladosin C. Penicillium chrysogenum PJX-17 is the source of the potent H1N1-fighting sorbicillinoids sorbicatechols A and B [89].
Trypilepyrazinol has demonstrated antiviral activity against several viruses, including HIV and HCV [90]. Aspernigrin C and malformin C, which A. niger SCSIO Jcsw6F30 produced, had strong antiviral activity against HIV-1 [91]. The Streptomyces kaviengensis (F7E2f) isolate antimycin A blocks mitochondrial electron transport and pyrimidine synthesis, making it effective against WEEV [92] [Table 4].
5. Advances in Micro-Culturing Technology
In 1951, Hungarian scientist Dr. Gyula Takatsy invented microwell plates, the first to minimize laboratory sample sizes. Figure 2 from Banks [94] illustrates how his idea completely changed laboratory titrations and serial dilutions. There are several drawbacks to using 96-well culture plates. It is not genuinely high-throughput, and evaporation makes it difficult to keep a constant volume in a culture over time. Furthermore, waste product accumulation within wells and restrictions in oxygen transmission restrict the expansion of confluent cells. Finally, Ingham et al. [95] point out that the low picture resolution of microwell plates makes it hard to visualize qualitative data.
The complicated interactions between microorganisms and high-throughput screening required 50 years of "micro-culturing" progression. Miniaturizing culture conditions allows for the isolation and compartmentalization of microorganisms. Fast-growing species often obscure slower-growing ones due to this type of competition. However, although they are relatively rare, slow-growing organisms account for a sizable fraction of microbial diversity [96]. Microenvironments are highly understudied, and the microtechnologies discussed here allow researchers to do so on spatiotemporal scales relevant to microorganisms [97].
6. Microarrays
The transition from mL-scale to mL-scale cell culture begins with microarray technology: automated and manual arrays print cell droplets in liquid or alginate gel onto microscope slides. The microarray has several possible uses, including but not limited to studying phenotypic, morphological, proliferative, and cell-cell communication outcomes of cell-culture miniaturization [98].
The "nano-biofilms" of Candida albicans were printed in a single study utilizing microarray printing techniques to create 1,200 tiny droplets of 30 nL each. When compared to normally grown Candida albicans, the "nano-biofilms" formed by Candida albicans were determined to be genetically and morphologically identical. Subsequently, 28 combinatorial synergistic antifungals were found to be effective against the microdroplets when tested for drug sensitivity.
In addition, microarrays are a useful tool for speeding up the discovery of novel metabolic pathways and developing novel drugs against well-established ADROs. Liquid droplets are easily damaged in studies, and secondly, they can only be interrogated in a static position on a Petri plate. Gel droplets protect cells from damage and provide a stable experimental environment, but the optimal conditions for investigating flow remain to be determined for microarray technology [99].
7. Antimicrobial Peptides
Various naturally occurring compounds known as antimicrobial peptides (AMPs) protect hosts from pathogens, including bacteria, fungi, parasites, and viruses [100]. Small in size, most AMPs are cationic because they contain several lysine or arginine residues. This positive charge allows AMPs to interact with the predominantly negative bacteria membranes. However, some AMPs have anionic properties [101,102].
The synergistic action with traditional antibiotics, low resistance risk, great efficacy at low concentrations, and broad spectrum activity of AMPs have all attracted attention [103,104]. They act on multiple plasma membrane and intracellular targets to destroy pathogenic organisms. Antimicrobial peptides (AMPs) are antibacterial, antifungal, antiviral, antiparasitic, and immunomodulatory [100]. While treating patients with VRE, Acinetobacter baumannii, or MRSA; S. Staphylococcus aureus, Listeria monocytogenes, and the bacterium E. Foodborne E. coli, Salmonella, and Vibrio parahaemolyticus infections can be prevented with the use of AMPs.
Defensins, nisin, and cecropins are AMPs that kill Gram-positive and Gram-negative bacteria. Both the Aristicluthys nobilia interferon-I inspired AMP P5 (YIRKIRRFFKKLKKILKK-NH2) and AMP P9 (SYERKINRHFKTLKKNLKKK-NH2) have been demonstrated to inhibit MRSA [105]. As a result of AMR, there has been a dramatic increase in demand for AMPs, which are used in both human healthcare and agricultural settings.
9. Microorganisms Affect Climate Change
Phytoplankton in the oceans reproduce far more quickly than tree species (days vs. decades)30, are more distributed, and are less affected by seasonal shifts. Phytoplankton are among the first organisms that respond rapidly to climate shifts. The cyclical structure of phytoplankton blooms makes it difficult to forecast how climate change will affect primary productivity. Grazing and viruses are examples of top-down variables, while the availability of key nutrients and vertical mixing are examples of bottom-up variables that affect phytoplankton blooms. [111,112,113,114,115,116]
Chemolithoautotrophic bacteria and archaea fix CO2, supplementing the role of marine phytoplankton in CO2 sequestration [119,120,121] in both deep-sea waters [117] and surface waters in polar winter [118]. Archaea and Marine bacteria are crucial for the ocean's surface respiration and cycling of many elements. We do not know much about how methanogens and methanotrophs add to the release of this greenhouse gas into the air. Marine viruses, for example, affect the efficiency with which carbon is stored and deposited in the deep ocean, and Eukaryotic grazers are integral components of the microbial food chain. Global biogeochemical cycles [122] are affected by climate change because of its effect on predator-prey relationships, particularly virus-host interactions.
Dissemination is typically simpler for bacteria than it is for larger creatures. Many microbial species vary biogeographically due to dispersal, lifestyle (including the influence of host association), and environmental variables that impact the composition and functioning of communities. [123,124,125,126]. Temperature and latitudinal gradients are particularly significant for marine ecosystems [127,128]. The adaptive potential [129] of microorganisms like bacteria, archaea, and microalgae is greatly enhanced by their vast population densities and fast asexual generation rates. Ocean acidification and other environmental variables related to climate change have received very little attention in evolutionary studies [129,130]. Little is known about the molecular mechanisms underlying physiological reactions or their biogeochemical consequences [131].
10. Terrestrial Biome
About half of the world's net primary production [120] comes from land plants. More carbon than is found in the atmosphere and plants is stored in the world's soils—a total of 2,000 billion tonnes. Land microorganisms number about 1029, like aquatic ones. Soil bacteria regulate the amount of organic carbon kept in soil and the amount released into the atmosphere. [133,134], in addition to controlling productivity with macronutrients like phosphate and nitrogen. Mycorrhizal fungi help plants get nitrogen and phosphate in various environments [134]. Plants provide their fungal symbionts with lots of carbon.
Up to 25% of all man-made carbon dioxide (CO2) is stored in forests [135,136], and forests account for 50% of all terrestrial primary productivity. Grasslands account for about 29% of Earth's surface area. These dry and semiarid regions respond differently to human-caused climate change than their forested counterparts [137,138], and they are crucial to the carbon budget. Lakes comprise around 4% of the Earth's non-glaciated surface area, and many methane gas emissions come from shallow lakes [139,140].
11. Climate Change Affects Microorganisms
Climate change affects microbial community structure and diversity through seasonality, temperature, plant composition, litter, and root exudates [141,142]. Natural geothermal warming in the lab and the long run (more than 50 years) boosted respiration and biomass of soil microbes, which released net carbon dioxide, while substrates were depleted, decreasing biomass and microbial activity [143]. That some microbial groups find it difficult to adjust to warmer environments is clear from this.
12. Infectious Diseases
Climate change impacts several environmental characteristics [144], which impacts illnesses in marine and terrestrial organisms [145]. (Figure 3). Modifying their defensive systems and nutrient cycling routes may make corals more vulnerable to bleaching and disease. [146] due to changes in their microbiome brought about by warming oceans. Damage to fish tissue from ocean acidification may impair their immune systems, allowing germs to colonize their bodies more easily [147]. Pathogens [148] potentially contribute to forest mortality due to drought and heat stress.
As a result of changes in host and parasite acclimation [149], climate change can raise disease risk. Temperature might increase susceptibility to infection for ectotherms (like amphibians), potentially by perturbing immune responses [149,150]. The Cuban tree frog is more vulnerable to Batrachochytrium dendrobatidis when temperatures fluctuate weekly and daily. Assessing host-pathogen reactions directly is of utmost importance rather than making assumptions based on research solely focused on the growth rates of individual microorganisms in order to determine the significance of climate change accurately. [149], as there is a discrepancy between the effect of rising temperatures on infection and the fungus' reduced growth capacity in pure culture. Recent research indicates that antibiotic resistance among human infections is projected to increase due to climate change [151].
Due to climate change, the transmission season may last longer, the rate at which diseases replicate in the vector may speed up, and the number and distribution of mosquitoes may grow. El Nio/Southern Oscillation (ENSO) and other large-scale climate phenomena significantly impact the spread of many infectious diseases, especially in vector-borne or aquatic environments. ENSO is associated with several significant human and animal illnesses, including malaria, dengue fever, Zika virus disease, cholera, the plague, African horse sickness, and many more. [152,153,154,155,156].
13. Tools and Techniques
Used for Identifying New Microorganism-Based Antimicrobial Drugs Finding new bioactive chemicals that are effective against disease-causing microorganisms is important because of the spread of antimicrobial resistance. The challenges encountered over the years include the inherent low concentrations of antimicrobials in natural sources, the absence of advanced and inventive drug discovery tools, and the substantial costs and limited investment in drug development and discovery. [157,158]. The following are examples of conventional methods for extracting useful chemicals from bacteria:
- Examples of diffusion processes include Agar discs, antimicrobial gradients, wells, plugs, cross streaks, and poisoned food. The agar disc diffusion method was created in 1940 to determine bacterial resistance [159]. Detecting some bacterial infections is difficult [160]. This comprises Streptococci, H. influenzae, N. gonorrhoea, N. meningitidis, and H. parainfluenzae. Agar-grown bacteria are exposed to a predefined dosage of the test chemical in this assay. The test compound's antimicrobial properties permeate into the agar, stopping the growth of sensitive bacteria. The zone's diameter is measured when a growth inhibitor is used [161,162]. Agar disc diffusion is a simple and cost-effective method for determining the minimum inhibitory concentration (MIC). Nevertheless, the outcomes are imprecise. [161].
- The antimicrobial gradient technique (Etest) may be utilized to calculate the minimum inhibitory concentration (MIC) of antibacterials, antifungals, and other antimicrobials, which combines dilution and diffusion techniques. This technique can also be utilized to analyze medication interactions [161,163,164,165].
- Several methods exist for screening and determining the susceptibility of microorganisms to antimicrobial medications. These include the time-kill test [168], the ATP bioluminescence assay [169,170,171,172], and the flow-cytofluorometric approach [173]. ATP bioluminescence has allowed for the evaluation of cellular ATP levels. The luciferin-luciferase bioluminescent test is popular due to its high sensitivity. A high quantum yield chemiluminescent reaction with MgATP2+ oxidizes luciferin catalytically via the luciferase. Light intensity correlates with ATP levels under ideal conditions. Stimulating ATP release from a disintegrating cell to react with luciferin-luciferase and produce light can determine cellular ATP. A luminometer is used to quantify the brightness of an object.
- A flow cytometer and flow cytofluorometric approach can identify antimicrobial resistance and predict the chemical's effect on microbe cell damage and viability [174].
14. Micromachined Devices
The grid layouts used in microfabrication allow for predictability and the use of readily available automated software for scoring growth, both of which contribute to the technology's high level of accuracy [95]. There is a wide range in mechanical strength, permeability, biocompatibility, and surface charge among the materials utilized to make these gadgets [175].
An unanticipated advantage of these microdevices is their portability, allowing in situ incubation in locations other than a laboratory. Hence, rather than creating and managing artificially altered growth circumstances, we may rely on the inherent microenvironment to facilitate the cultivation of previously unattainable species. [176]. Kaeberlein et al. [177] proposed in the early 2000s that it would be possible to isolate and culture microorganisms without understanding the precise components necessary to sustain the establishment of a given ecosystem by just employing their natural milieu. Since this was the case, they created diffusion chambers by placing an agar matrix containing an ambient sample between two membranes with pores of 0.03 m [178].
Inoculation of the ichip is accomplished by dilution-to-extinction of the ambient sample, resulting in the development of monocultures without the need for manual picking and isolation [179]. Many phylogenetically unique species, including those discovered in human mouths [180] and saltwater [181], have never been cultured using conventional methods. However, since the ichip was introduced, they have been cultivated and isolated.
Recently, novel antibiotics such as Novo10 and Neocitreamicin I and II have been identified thanks to the successful cultivation of rare environmental species utilizing the specified diffusion chips and chips [182,183]. Figure 4: Teixobactin was identified during the isolation and passage of Eleftheria terrae, a novel -proteobacteria [184]. It effectively killed Gram-positive bacteria, even those that had developed resistance to antibiotics.
Cultures of previously unknown species were established in a soil sample from the environment using the ichip [184]. Antibiotic activity against S. aureus was tested by culturing various species onto agar plates. The in vitro efficacy of bioactive compounds against a wide range of pathogens was determined after their extraction, purification, and characterization. Using this method, researchers found teixobactin, a new antibiotic molecule that prevented any mutants from becoming resistant in early testing. After initial discovery, innovative medications undergo extensive testing for safety and efficacy before being developed for consumer use.
15. Single Emulsion Droplet Microfluidics
Because each droplet functions as a miniature bioreactor, droplet microfluidics has recently gained importance for studying cells. The culture medium often has length scales similar to prokaryotic and eukaryotic cells. Therefore, droplets ranging in size from pico- to microliters facilitate the fast passage of gasses, nutrients, metabolic waste, and similar substances. [97].
Beneyton et al. [185] employed a microfluidic system to conduct a screening process for the enzymatic production of -amylase by the filamentous fungus Aspergillus niger. The droplets were generated at a frequency of 80-90 droplets per second and had a volume in the nanoliter range (10-18 nL). Agar droplets have also been widely used for cell confinement, in addition to the liquid, aqueous phase often associated with single emulsions. During encapsulation, the agar syringe is kept liquid by submerging it in a water bath that is heated to a regulated temperature [186]. Alkayyali et al. [187] created the Microbe Domestication Pod (MD Pod) to cultivate marine samples separated within agarose beads in a single emulsion. After the gel beads are injected inside the Pod, the device may be placed back in its natural home to continue its development.
Silicon molds can be etched, embossed, or lithographed using various methods available due to microelectromechanical system (MEMS) microfabrication processes. Soft photolithography, pioneered by Xia and Whitesides [188], is currently one of the most popular fabrication processes. In soft lithography, a master mold of the necessary microfluidic device is made of photo-cross-linkable polymers rather than silicon wafers. Commercially available chips may not be ideal for exploratory research, especially with environmental materials. Several university-based public foundries have been established to facilitate customization and ease of use in the fabrication process by enabling users to submit their designs for microfabricated chips without needing specialized training or financial outlay [189].
16. Double Emulsion Droplet Microfluidics and Polymer-Based Nanocultures
Microfluidic channels with hydrodynamic pressure flow and coflowing geometry can create double-emulsion droplets, which function similarly to single-emulsion droplets. The cell inoculum is in an aqueous phase at the system's center. The hydrophobic, hydrophilic intermediate phase produces double emulsion droplets, which are stabilized in solution by a surfactant in a continuous watery (hydrophilic) phase.
Because double-emulsion droplets partition chemical reactions into nanoliter bioreactors, droplet microfluidics enables high-throughput research. Products that have passed the chemical test can be separated using fluorescence-activated cell sorting. Zinchenko et al. [190] demonstrated the platform's efficacy by identifying enhanced cellular clones capable of producing catalytically active enzymes from 106 inactive enzymes in 10 m droplets. The researchers demonstrated that the assay can be heat-inactivated to prevent catalytic activation and that droplets may be frozen at 80°C and thawed for discontinuous workflows.
Modifying the polymeric membrane to fit the specific structural requirements of different uses is possible. PDMS's high mechanical toughness and elasticity make it an important material; its Young modulus is around 0.5 to 3 MPa [191]. While this aids in producing robust capsules for cell encapsulation, it complicates subsequent processing steps, such as emptying the capsules.
17. Applications of Microbial-based Microsystems and Perceived Challeanges
Optical transparency allows for the study of phylogenetic responses in isolated colonies that have been stimulated chemically. Observing increasingly complex consortia to examine intra- and inter-species interactions is vital in differentiating symbiotic and antagonistic behaviors across species and pathogenic switching in opportunistic illnesses.
Many difficult-to-manage species, however, have unidentified secondary metabolite needs [192]. Microorganisms can be cultivated in engineered microsystems, even in extreme environments that would have previously been impossible. Clostridium difficile was deemed an "Urgent Threat" [193], according to the 2019 report on Antimicrobial Resistance Threats from the United States Centres for Disease Control and Prevention (CDC). Monocultures provide a new perspective for studying the intricate interactions between gut microbiota and C. difficile in a low-oxygen environment by allowing optical access under a microscope.
The purpose of these ecosystems is not confined to the production of so-called "unculturable" species. Many applications that previously were impossible to accomplish with the level of control, accuracy, and resolution required to identify study targets have been made possible thanks to the development of miniaturizing culturing techniques [194]. Low-throughput, expensive approaches are often connected with drug and enzymatic metabolite screening in the biotechnology industry [195]. Microfluidics is superior because it streamlines the process by shrinking everything to a microscopic size. Rapid turnaround time (a few hours instead of days) reduces the overall expense of screening procedures. Diagnostic tools, biological assays, and analytical tools in the pharmaceutical business can greatly benefit from the replicability and automation provided by microfluidics [196].
Materials design is an example of a more specialized difficulty encountered during gadget production. According to Aleklett et al. [97], unlike naturally occurring habitats, PDMS, an essentially hydrophobic elastomer, does not offer cells a heterogeneous setting where they can be mechanically manipulated. However, the hydrophilic surface modifications induced by oxygen plasma treatment of PDMS are only transitory, as the surface quickly reverts to its lower free energy state due to the migration of oligomers inside the PDMS network [197].
18. Conclusions
In this day and age of antimicrobial resistance, the diversity of chemicals found in bacteria and other microorganisms and their powerful action is important. Expanding our search to include these organisms will lead us to potentially game-changing antimicrobials with mechanisms of action not yet seen in existing drugs. Antimicrobial resistance can thus be drastically decreased in this way. In order to fully understand the influence of microbial diversity and activity on various levels of interactions and system fluxes, it is necessary to perform experiments across several scales, ranging from individual organisms to communities and entire ecosystems. To accurately portray the Earth as a whole, models of the biosphere must account for both the biotic (consisting of bacteria, plants, and organic matter substrates) and abiotic (including mineral surfaces, ocean physics, and chemistry) environments in which microbes operate. There are now fascinating and unique ways to explore "microbial dark matter" and the accompanying microbial dynamics because of the downsizing of culturing technologies. Precise instruments created by microfabrication techniques are used to investigate the microenvironment of cells. Instead of using conventional nutrient-rich liquids, these devices utilize natural surroundings that exist everywhere.
Funding
Not applicable.
Data Availability Statement
All information produced or examined in this research is incorporated within this published article.
Acknowledgments
The authors wish to thank RAK Medical and Health Sciences University, Ras Al Khaimah, United Arab Emirates, for their research support for the research project.
Conflicts of Interest
The authors assert that they do not have any conflicts of interest.
References
- O'Neill, J. 2014. Antimicrobial Resistance: Tackling a crisis for the health and wealth of nations. Rev. Antimicrob. Resist.
- Laws, M.; Shaaban, A.; Rahman, K.M. 2019. Antibiotic resistance breakers: Current approaches and future directions. FEMS Microbiol. Rev., 43, 490–516.
- Newman, D.J.; Cragg, G.M. 2012. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod., 75, 311–335. [CrossRef]
- Elissawy, A.M.; Dehkordi, E.S.; Mehdinezhad, N.; Ashour, M.L.; Pour, P.M. 2021. Cytotoxic Alkaloids Derived from Marine Sponges: A Comprehensive Review. Biomolecules, 11, 258. [CrossRef]
- El-Demerdash, A.; Tammam, M.A.; Atanasov, A.G.; Hooper, J.N.A.; Al-Mourabit, A.; Kijjoa, A. 2018. Chemistry and Biological Activities of the Marine Sponges of the Genera Mycale (Arenochalina), Biemna and Clathria. Mar. Drugs, 16, 214. [CrossRef]
- Karpinski, T.M. 2019. Marine Macrolides with Antibacterial and/or Antifungal Activity. Mar. Drugs, 17, 241. [CrossRef]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. 2015. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs, 13, 3479–3513. [CrossRef]
- Swift, C.L.; Louie, K.B.; Bowen, B.P.; Olson, H.M.; Purvine, S.O.; Salamov, A.; Mondo, S.J.; Solomon, K.V.; Wright, A.T.; Northen, T.R.; et al. 2021. Anaerobic gut fungi are an untapped reservoir of natural products. Proc. Natl. Acad. Sci. USA, 118, e2019855118. [CrossRef]
- Maghembe, R.; Damian, D.; Makaranga, A.; Nyandoro, S.S.; Lyantagaye, S.L.; Kusari, S.; Hatti-Kaul, R. 2020. Omics for Bioprospecting and Drug Discovery from Bacteria and Microalgae. Antibiot, 9, 229. [CrossRef]
- Moir, D.T.; Shaw, K.J.; Hare, R.S.; Vovis, G.F. 1999. Genomics and Antimicrobial Drug Discovery. Antimicrob. Agents Chemother., 43, 439. [CrossRef]
- Schnappinger, D. 2015. Genetic Approaches to Facilitate Antibacterial Drug Development. Cold Spring Harb. Perspect. Med., 5, a021139. [CrossRef]
- Tortorella, E.; Tedesco, P.; Espositom, F.P.; January, G.G.; Fani, R.; Jaspars, M.; de Pascale, D. 2018. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs, 16, 355. [CrossRef]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. 2011. Cyanobacteria: An emerging source for drug discovery. J. Antibiot., 64, 401–412. [CrossRef]
- Rojas, V.; Rivas, L.; Cardenas, C.; Guzman, F. 2020. Cyanobacteria and Eukaryotic Microalgae as Emerging Sources of Antibacterial Peptides. Molecules, 25, 5804. [CrossRef]
- Alsenani, F.; Tupally, K.R.; Chua, E.T.; Eltanahy, E.; Alsufyani, H.; Parekh, H.S.; Schenk, P.M. 2020. Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds. Saudi. Pharm. J, 28, 1834–1841. [CrossRef]
- Santovito, E.; Greco, D.; Marquis, V.; Raspoet, R.; D’Ascanio, V.; Logrieco, A.F.; Avantaggiato, G. 2019. Antimicrobial Activity of Yeast Cell Wall Products Against Clostridium perfringens. Foodborne Pathog. Dis., 16, 638–647. [CrossRef]
- Hatoum, R.; Labrie, S.; Fliss, I. (2012) Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol., 3, 421. [CrossRef]
- Mitcheltree, M.J.; Pisipati, A.; Syroegin, E.A.; Silvestre, K.J.; Klepacki, D.; Mason, J.D.; Terwilliger, D.W.; Testolin, G.; Pote, A.R.; Wu, K.J.Y.; et al. 2021. A synthetic antibiotic class overcoming bacterial multidrug resistance. Nature, 599, 507–512. [CrossRef]
- Tenea, G.N.; Yépez, L. 2016. Bioactive Compounds of Lactic Acid Bacteria. Case Study: Evaluation of Antimicrobial Activity of Bacteriocin-producing Lactobacilli Isolated from Native Ecological Niches of Ecuador. Probiotics Prebiotics Hum. Nutr. Health, xxx, 149–167.
- Todorov, S.D.; Dicks, L.M.T. .2007. Bacteriocin production by Lactobacillus pentosus ST712BZ isolated from boza. Braz. J. Microbiol., 38, 166–172. [CrossRef]
- Oscáriz, J.C.; Pisabarro, A.G. 2001. Classification and mode of action of membrane-active bacteriocins produced by Gram-positive bacteria. Int. Microbiol., 4, 13–19. [CrossRef]
- Klaenhammer, T.R. 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev., 12, 39–85. [CrossRef]
- Güllüce, M.; Karadayı, M.; Barı¸s, Ö. 2013. Bacteriocins: Promising natural antimicrobials. Local Environ., 3, 6.
- Cotter, P.D.; Hill, C.; Ross, R.P. 2005. Food microbiology: Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol., 3, 777–788. [CrossRef]
- Drider, D.; Fimland, G.; Héchard, Y.; McMullen, L.M.; Prévost, H. 2006. The Continuing Story of Class IIa Bacteriocins. Microbiol. Mol. Biol. Rev., 70, 564–582. [CrossRef]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. 2021. Bacteriocins from lactic acid bacteria. A powerful alternative as antimicrobials, probiotics, and immunomodulators in veterinary medicine. Animals, 11, 979. [CrossRef]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. 2007. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biol. Technol., 50, 521–542. [CrossRef]
- Bauer, R.; Dicks, L.M.T. 2005 Mode of action of lipid II-targeting lantibiotics. Int. J. Food Microbiol., 101, 201–216. [CrossRef]
- Paiva, A.D.; Breukink, E.; Mantovani, H.C. (2011) Role of lipid II and membrane thickness in the mechanism of action of the lantibiotic bovicin HC5. Antimicrob. Agents Chemother., 55, 5284–5293. [CrossRef]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. 2010. Structure and Mode-of-Action of the Two-Peptide (Class-IIb) Bacteriocins. Probiotics Antimicrob. Proteins, 2, 52–60. [CrossRef]
- Perez, R.H.; Zendo, T.; Sonomoto, K. 2018 Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front Microbiol., 9, 2085. [CrossRef]
- Simmonds, R.S.; Pearson, L.; Kennedy, R.C. 2018. Mode of Action of a Lysostaphin-Like Bacteriolytic Agent Produced by Streptococcus zooepidemicus 4881. Appl. Environ. Microbiol., 62, 4536–4541. [CrossRef]
- Sun, Z.; Wang, X.; Zhang, X.; Wu, H.; Zou, Y.; Li, P.; Sun, C.; Xu, W. 2018. Class III bacteriocin Helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane. J. Ind. Microbiol. Biotechnol., 45, 213–227. [CrossRef]
- Swe, P.M.; Cook, G.M.; Tagg, J.R.; Jack, R.W. 2009. Mode of Action of Dysgalacticin: A Large Heat-Labile Bacteriocin. J. Antimicrob. Chemother., 63, 679–686. [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. 2020. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics, 9, 32. [CrossRef]
- Mclller, E.; Radler, F. Caseicin. 1993. a Bacteriocin from Lactobacillus casei. Folia Microbiol., 38, 441–446. [CrossRef]
- Bollenbach, T. 2015. Antimicrobial interactions: Mechanisms and implications for drug discovery and resistance evolution. Curr. Opin. Microbiol., 27, 1–9. [CrossRef]
- Herzog, M.; Tiso, T.; Blank, L.M.; Winter, R. 2020. Interaction of rhamnolipids with model biomembranes of varying complexity. Biochim. Biophys. Acta-Biomembr., 1862, 183431. [CrossRef]
- Jiang, J.; Zu, Y.; Li, X.; Meng, Q.; Long, X. 2020. Recent progress towards industrial rhamnolipids fermentation: Process optimization and foam control. Bioresour. Technol., 298, 122394. [CrossRef]
- Thakur, P.; Saini, N.K.; Thakur, V.K.; Gupta, V.K.; Saini, R.V.; Saini, A.K. 2021. Rhamnolipid the Glycolipid Biosurfactant: Emerging trends and promising strategies in the field of biotechnology and biomedicine. Microb. Cell Factories, 20, 1–15. [CrossRef]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. 2013. Chemical Structure and In Vitro Antitumor Activity of Rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol., 170, 676–689. [CrossRef]
- Borah, S.N.; Goswami, D.; Sarma, H.K.; Cameotra, S.S.; Deka, S. 2016. Rhamnolipid biosurfactant against Fusarium verticillioides to control stalk and ear rot disease of maize. Front. Microbiol., 7, 1505. [CrossRef]
- Jin, L.; Black, W.; Sawyer, T. 2021. Application of Environment-Friendly Rhamnolipids against Transmission of Enveloped Viruses Like SARS-CoV2. Viruses, 13, 322. [CrossRef]
- Remichkova, M.; Galabova, D.; Roeva, I.; Karpenko, E.; Shulga, A.; Galabov, A.S. 2008. Anti-herpesvirus activities of Pseudomonas sp. S-17 rhamnolipid and its complex with alginate. Zeitschrift Fur Naturforsch Sect. C J. Biosci., 63, 75–81. [CrossRef]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Esposito, F.P.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. 2021. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics, 13, 2121. [CrossRef]
- Broadbent, J.R.; Chou, Y.C.; Gillies, K.; Kondo, J.K. 1989. Nisin Inhibits Several Gram-Positive, Mastitis-Causing Pathogens. J. Dairy Sci., 72, 3342–3345. [CrossRef]
- Lebel, G.; Piché, F.; Frenette, M.; Gottschalk, M.; Grenier, D. 2013. Antimicrobial activity of nisin against the swine pathogen Streptococcus suis and its synergistic interaction with antibiotics. Peptides, 50, 19–23. [CrossRef]
- Carroll, J.; Draper, L.A.; O'Connor, P.M.; Coffey, A.; Hill, C.; Ross, R.P.; Cotter, P.D.; O'Mahony, J. 2010. Comparison of the activities of the lantibiotics nisin and lacticin 3147 against clinically significant mycobacteria. Int. J. Antimicrob. Agents, 36, 132–136. [CrossRef]
- Campion, A.; Casey, P.G.; Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. 2013. In vivo activity of Nisin A and Nisin v against Listeria monocytogenes in mice. BMC Microbiol., 13, 23. [CrossRef]
- Grilli, E.; Messina, M.; Catelli, E.; Morlacchini, M.; Piva, A. 2009. Pediocin a improves growth performance of broilers challenged with Clostridium perfringens. Poult. Sci., 88, 2152–2158. [CrossRef]
- Lauková, A.; Styková, E.; Kubašová, I.; Gancarˇcíková, S.; Plachá, I.; Mudro ˇnová, D.; Kandriˇcáková, A.; Miltko, R.; Belzecki, G.; Valocký, I. 2018. Enterocin M and its Beneficial Effects in Horses—A Pilot Experiment. Probiotics Antimicrob. Proteins, 10, 420–426. [CrossRef]
- Wang, Q.; Cui, Y.; Wang, W.; Xu, J.; Xu, L. 2012. Production of two bacteriocins in various growth conditions produced by gram-positive bacteria isolated from chicken cecum. Can. J. Microbiol., 58, 93–101. [CrossRef]
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. 2008. Isolation and Purification of Enterocin E-760 with Broad Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria. Antimicrob. Agents Chemother., 52, 1094–1100. [CrossRef]
- Ryan, M.P.; Flynn, J.; Hill, C.; Ross, R.P.; Meaney, W.J. 1999. The natural food grade inhibitor, lacticin 3147, reduced the incidence of mastitis after experimental challenge with Streptococcus dysgalactiae in nonlactating dairy cows. J. Dairy Sci., 82, 2625–2631. [CrossRef]
- Pieterse, R.; Todorov, S.D.; Dicks, L.M.T. 2010. Mode of action and in vitro susceptibility of mastitis pathogens to macedocin ST91KM and preparation of a teat seal containing the bacteriocin. Braz. J. Microbiol., 41, 133–145. [CrossRef]
- Tomita, K.; Nishio, M.; Saitoh, K.; Yamamoto, H.; Hoshino, Y.; Ohkuma, H.; Konishi, M.; Miyaki, T.; Oki, T. 1990. Pradimicins A, B and C: New antifungal antibiotics. I. Taxonomy, production, isolation and physico-chemical properties. J. Antibiot, 43, 755–762. [CrossRef]
- Walsh, T.J.; Giri, N. 1997. Pradimicins: A novel class of broad-spectrum antifungal compounds. Eur. J. Clin. Microbiol. Infect. Dis., 16, 93–97. [CrossRef]
- Kerr, J.R. 1999. Bacterial inhibition of fungal growth and pathogenicity. Microb. Ecol. Health Dis., 11, 129–142. [CrossRef]
- Boumehira, A.Z.; El-Enshasy, H.A.; Hacene, H.; Elsayed, E.A.; Aziz, R.; Park, E.Y. 2016. Recent progress on the development of antibiotics from the genus Micromonospora. Biotechnol. Bioprocess Eng., 21, 199–223. [CrossRef]
- Schwartz, R.E.; Giacobbe, R.A.; Monaghan, R.L. 1989. L-671,329, a new antifungal agent. I. Fermentation and isolation. J. Antibiotic, 42, 163–167. [CrossRef]
- Nishizawa, N.; Kondo, Y.; Koyama, M.; Omoto, S.; Iwata, M.; Tsuruoka, T.; Inouye, S. 1983. Studies on a new nucleotide antibiotic, Dapiramicin. J. Antibiot., 37, 1–5.
- Dunlap, C.A.; Bowman, M.J.; Rooney, A.P. 2019. Iturinic lipopeptide diversity in the bacillus subtilis species group-important antifungals for plant disease biocontrol applications. Front. Microbiol., 10, 1794. [CrossRef]
- Lebbadi, M.; Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Fungicin M4: 1994. A narrow spectrum peptide antibiotic from Bacillus licheniformis M-4. J. Appl. Bacteriol., 77, 49–53. [CrossRef]
- Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Lebbadi, M.; Valdivia, E. 1993. Isolation and physico-chemical characterization of an antifungal and antibacterial peptide produced by Bacillus licheniformis A12. Appl. Microbiol. Biotechnol., 39, 438–442. [CrossRef]
- Chernin, L.; Brandis, A.; Ismailov, Z.; Chet, I. 1996. Pyrrolnitrin production by an Enterobacter agglomerans strain with a broad spectrum of antagonistic activity towards fungal and bacterial phytopathogens. Curr. Microbiol., 32, 208–212. [CrossRef]
- Greiner, M.; Winkelmann, G. 1991. Fermentation and isolation of herbicolin A, a peptide antibiotic produced by Erwinia herbicola strain A 111. Appl. Microbiol. Biotechnol., 34, 565–569. [CrossRef]
- Winkelmann, G.; Lupp, R.; Jung, G. 1980. Herbicolins—New peptide antibiotics from erwinia herbicola. J. Antibiot., 33, 353–358. [CrossRef]
- Shoji, J.; Hinoo, H.; Sakazaki, R.; Kato, T.; Hattori, T.; Matsumoto, K.; Tawara, K.; Kikuchi, J.; Terui, Y. 1989. Isolation of CB-25-I, an antifungal antibiotic, from serratia plymuthica. J. Antibiot., 42, 869–874. [CrossRef]
- Serino, L.; Reimmann, C.; Visca, P.; Beyeler, M.; Chiesa, V.D.; Haas, D. 1997. Biosynthesis of pyochelin and dihydroaeruginoic acid requires the iron-regulated pchDCBA operon in Pseudomonas aeruginosa. J. Bacteriol., 179, 248–257. [CrossRef]
- Kerr, J.R.; Taylor, G.W.; Rutman, A.; Høiby, N.; Cole, P.J.; Wilson, R. 1999. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol., 52, 385–387. [CrossRef]
- Vincent, M.N.; Harrison, L.A.; Brackin, J.M.; Kovacevich, P.A.; Mukerji, P.; Weller, D.M.; Pierson, E.A. 1991. Genetic analysis of the antifungal activity of a soilborne Pseudomonas aureofaciens strain. Appl. Environ. Microbiol., 57, 2928–2934. [CrossRef]
- Harrison, L.; Teplow, D.B.; Rinaldi, M.; Strobel, G. Pseudomycins. 1991. a family of novel peptides from Pseudomonas syringae possessing broad-spectrum antifungal activity. J. Gen. Microbiol., 137, 2857–2865. [CrossRef]
- Yamaguchi, M.; Park, H.J.; Ishizuka, S.; Omata, K.; Hirama, M. 1995. Chemistry and Antimicrobial Activity of Caryoynencins Analogs. J. Med. Chem., 38, 5015–5022. [CrossRef]
- Barker, W.R.; Callaghan, C.; Hill, L.; Noble, D.; Acred, P.; Harper, P.B.; Sowa, M.A.; Fletton, R.A. G1549. 1979. A new cyclic hydroxamic acid antibiotic, isolated from culture broth of pseudomonas alcaligenes. J. Antibiot., 32, 1096–1103. [CrossRef]
- Lim, Y.; Suh, J.W.; Kim, S.; Hyun, B.; Kim, C.; Lee, C. 1994. hoon Cepacidine A, A novel antifungal antibiotic produced by pseudomonas cepacia. II. Physico-chemical properties and structure elucidation. J. Antibiot., 47, 1406–1416. [CrossRef]
- Abe, M.; Nakazawa, T. 1994. Characterization of Hemolytic and Antifungal Substance, Cepalycin, from Pseudomonas cepacia.Microbiol. Immunol, 38, 1–9. [CrossRef]
- Bisacchi, G.S.; Parker, W.L.; Hockstein, D.R.; Koster, W.H.; Rathnum, M.L.; Unger, S.E. 1987. Xylocandin: A new complex of antifungal peptides. II. Structural studies and chemical modifications. J. Antibiot., 40, 1520–1529. [CrossRef]
- Saalim, M.; Villegas-Moreno, J.; Clark, B.R. 2020. Bacterial Alkyl-4-quinolones: Discovery, Structural Diversity and Biological Properties. Molecules, 25, 5689. [CrossRef]
- Knappe, T.A.; Linne, U.; Zirah, S.; Rebuffat, S.; Xie, X.; Marahiel, M.A. 2008. Isolation and structural characterization of capistruin, a lasso peptide predicted from the genome sequence of Burkholderia thailandensis E264. J. Am. Chem. Soc., 130, 11446–11454. [CrossRef]
- Watanabe, T.; Izaki, K.; Takahashi, H. 1982. New polyenic antibiotics active against gram-positive and -negative bacteria: I. Isolation and purification of antibiotics produced by gl uconobacter sp. W-315. J. Antibiot., 35, 1141–1147. [CrossRef]
- Agrawal, S.; Deshmukh, S.K.; Reddy, M.S.; Prasad, R.; Goel, M. 2020. Endolichenic fungi: A hidden source of bioactive metabolites. S. Afr. J. Bot., 134, 163–186. [CrossRef]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; Del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. 2000. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol., 23, 333–343. [CrossRef]
- Kuhnert, E.; Li, Y.; Lan, N.; Yue, Q.; Chen, L.; Cox, R.J.; An, Z.; Yokoyama, K.; Bills, G.F. 2018. Enfumafungin synthase represents a novel lineage of fungal triterpene cyclases. Environ. Microbiol., 20, 3325–3342. [CrossRef]
- Sauter, H.; Steglich, W.; Anke, T. 1999. Strobilurins: Evolution of a new class of active substances. Angew. Chem.-Int. Ed., 38, 1328–1349.
- Chepkirui, C.; Richter, C.; Matasyoh, J.C.; Stadler, M. 2016. Monochlorinated calocerins A–D and 9-oxostrobilurin derivatives from the basidiomycete Favolaschia calocera. Phytochemistry, 132, 95–101. [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. 2019. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers., 97, 1–136. [CrossRef]
- Chepkirui, C.; Cheng, T.; Matasyoh, J.; Decock, C.; Stadler, M. 2018. An unprecedented spiro [furan-2,1’-indene]-3-one derivative and other nematicidal and antimicrobial metabolites from Sanghuangporus sp. (Hymenochaetaceae, Basidiomycota) collected in Kenya. Phytochem. Lett., 25, 141–146. [CrossRef]
- Zhu, T.; Chen, Z.; Liu, P.; Wang, Y.; Xin, Z.; Zhu, W. 2014. New rubrolides from the marine-derived fungus Aspergillus terreus OUCMDZ-1925. J. Antibiot., 67, 315–318. [CrossRef]
- Peng, J.; Zhang, XS.; Du, L.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. 2014. Sorbicatechols A and B, antiviral sorbicillinoids from the marinederived fungus Penicillium chrysogenum PJX-17. J. Nat. Prod., 77, 424–428. [CrossRef]
- Li, J.; Wang, Y.; Hao, X.; Li, S.; Jia, J.; Guan, Y.; Peng, Z.; Bi, H.; Xiao, C.; Cen, S.; et al. 2019. Broad-Spectrum Antiviral Natural Products from the Marine-Derived Penicillium sp. IMB17-046. Molecules, 24, 2821. [CrossRef]
- Zhou, X.; Fang, W.; Tan, S.; Lin, X.; Xun, T.; Yang, B.; Liu, S.; Liu, Y. 2016. Aspernigrins with anti-HIV-1 activities from the marine-derived fungus Aspergillus niger SCSIO Jcsw6F30. Bioorg. Med. Chem. Lett., 26, 361–365. [CrossRef]
- Raveh, A.; Delekta, P.C.; Dobry, C.J.; Peng, W.; Schultz, P.J.; Blakely, P.K.; Tai, A.W.; Matainaho, T.; Irani, D.N.; Sherman, D.H.; et al. 2013. Discovery of potent broad spectrum antivirals derived from marine actinobacteria. PLoS ONE, 8, e82318. [CrossRef]
- Wu, G.; Sun, X.; Yu, G.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. 2014. Cladosins A–E, hybrid polyketides from a deep-sea-derived fungus, Cladosporium sphaerospermum. J. Nat. Prod., 77, 270–275. [CrossRef]
- Banks, P. 2009. The Microplate Market Past, Present and Future [Online]. Drug Discovery World. Available online: https://www.ddw-online.com/themicroplate-market-past-present-and-future-1127-200904/ (accessed on 1 April 2021).
- Ingham, C.J.; Sprenkels, A.; Bomer, J.; Molenaar, D.; Van Den Berg, A.; Van Hylckama Vlieg, J.E.; et al. 2007. The micro-Petri dish, a million-well growth chip for the culture and high-throughput screening of microorganisms. Proc. Natl. Acad. Sci. U.S.A. 104, 18217–18222. [CrossRef]
- Jiang, C.-Y.; Dong, L.; Zhao, J.-K.; Hu, X.; Shen, C.; Qiao, Y.; et al. 2016. Highthroughput single-cell cultivation on microfluidic streak plates. Appl. Environ. Microbiol. 82:2210. [CrossRef]
- Aleklett, K.; Kiers, E.T.; Ohlsson, P.; Shimizu, T.S.; Caldas, V.E.A.; Hammer, E.C. 2018. Build your own soil: exploring microfluidics to create microbial habitat structures. ISME J. 12, 312–319. [CrossRef]
- Ge, Z.; Girguis, P.R.; Buie, C.R. 2016. Nanoporous microscale microbial incubators. Lab Chip 16, 480–488. [CrossRef]
- Srinivasan, A.; Leung, K.P.; Lopez-Ribot, J.L.; Ramasubramanian, A.K. 2013. High-throughput nano-biofilm microarray for antifungal drug discovery. mBio 4:e00331-00313. [CrossRef]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. 2019. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev., 39, 831–859. [CrossRef]
- Narayana, J.L.; Chen, J.Y. 2015. Antimicrobial peptides: Possible anti-infective agents. Peptides, 72, 88–94. [CrossRef]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. DRAMP 2.0. 2019. an updated data repository of antimicrobial peptides. Sci. Data, 6, 1–10. [CrossRef]
- da Cunha, N.B.; Cobacho, N.B.; Viana, J.F.C.; Lima, L.A.; Sampaio, K.B.O.; Dohms, S.S.M.; Ferreira, A.C.R.; de la Fuente-Núñez, C.; Costa, F.F.; Franco, O.L.; et al. 2017. The next generation of antimicrobial peptides (AMPs) as molecular therapeutic tools for the treatment of diseases with social and economic impacts. Drug Discov. Today, 22, 234–248. [CrossRef]
- Huerta-Cantillo, J.; Navarro-García, F. 2016. Properties and design of antimicrobial peptides as potential tools againstpathogens and malignant cells. Investig. En Discapac., 5, 96–115.
- Li, C.; Zhu, C.; Ren, B.; Yin, X.; Shim, S.H.; Gao, Y.; Zhu, J.; Zhao, P.; Liu, C.; Yu, R.; et al. 2019. Two optimized antimicrobial peptides with therapeutic potential for clinical antibiotic-resistant Staphylococcus aureus. Eur. J. Med. Chem., 183, 111686. [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. 2020. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol., 11, 582–779. [CrossRef]
- Carriel-Gomes, M.C.; Kratz, J.M.; Barracco, M.A.; Bachére, E.; Barardi, C.R.M.; Simões, C.M.O. 2007. In vitro antiviral activity of antimicrobial peptides against herpes simplex virus 1, adenovirus, and rotavirus. Mem. Inst. Oswaldo Cruz, 102, 469–472. [CrossRef]
- Kobori, T.; Iwamoto, S.; Takeyasu, K.; Ohtani, T. 2007. Biopolymers Volume 85/Number 4295. Biopolymers, 85, 392–406.
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. 2020. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res., 30, 343–355. [CrossRef]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. 2000. Evaluation of the inactivation of infectious herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis., 19, 187–194. [CrossRef]
- Hutchins, D.A.; Boyd, P.W. 2016. Marine phytoplankton and the changing ocean iron cycle. Nat. Clim. Change 6, 1072–1079. [CrossRef]
- Behrenfeld, M. J. 2014. Climate-mediated dance of the plankton. Nat. Clim. Change 4, 880–887. [CrossRef]
- De Baar, H. J. W.; et al. 1995. Importance of iron for plankton blooms and carbon dioxide drawdown in the Southern Ocean. Nature 373, 412–415. [CrossRef]
- Boyd, P. W.; et al. 2007. Mesoscale iron enrichment experiments 1993-2005: synthesis and future directions. Science 315, 612–617. [CrossRef]
- Behrenfeld, M. J.; et al. 2016. Revaluating ocean warming impacts on global phytoplankton. Nat. Clim. Change 6, 323–330. [CrossRef]
- Behrenfeld, M. J.; et al. 2017. Annual boom-bust cycles of polar phytoplankton biomass revealed by space-based lidar. Nat. Geosci. 10, 118–122. [CrossRef]
- Pachiadaki, M. G.; et al. 2017. Major role of nitrite-oxidizing bacteria in dark ocean carbon fixation. Science 358, 1046–105. [CrossRef]
- Grzymski, J. J.; et al. 2012. A metagenomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters. ISME J. 6, 1901–1915. [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. 1998. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240. [CrossRef]
- Behrenfeld, M. J.; et al. 2001. Biospheric primary production during an ENSO transition. Science 291, 2594–2597. [CrossRef]
- Boetius, A.; et al. 2013. Massive export of algal biomass from the melting Arctic sea ice. Science 339, 1430. [CrossRef]
- Danovaro, R.; et al. 2011. Marine viruses and global climate change. FEMS Microbiol. Rev. 35, 993–1034. [CrossRef]
- Brum, J. R.; et al. 2015. Patterns and ecological drivers of ocean viral communities. Science 348, 1261498. [CrossRef]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. 2012. Beyond biogeographic patterns: processes shaping the microbial landscape. Nat. Rev. Microbiol. 10, 497–506. [CrossRef]
- Zinger, L.; Boetius, A.; Ramette, A. 2014. Bacterial taxa-area and distance-decay relationships in marine environments. Mol. Ecol. 23, 954–964. [CrossRef]
- Archer, S. D. J.; et al. 2019. Airborne microbial transport limitation to isolated Antarctic soil habitats. Nat. Microbiol. 4, 925–932. [CrossRef]
- Wilkins, D.; van Sebille, E.; Rintoul, S.R.; Lauro, F.M.; Cavicchioli, R. 2013. Advection shapes Southern Ocean microbial assemblages independent of distance and environment effects. Nat. Commun. 4, 2457. [CrossRef]
- Cavicchioli, R. 2015. Microbial ecology of Antarctic aquatic systems. Nat. Rev. Microbiol. 13, 691–706. [CrossRef]
- Riebesell, U.; Gattuso, J.-P. 2015. Lessons learned from ocean acidification research. Nat. Clim. Change 5, 12–14. [CrossRef]
- Hutchins, D.A.; Fu, F.X. 2017. Microorganisms and ocean global change. Nat. Microbiol. 2, 17508. [CrossRef]
- Bunse, C.; et al. 2016. Response of marine bacterioplankton pH homeostasis gene expression to elevated CO2. Nat. Clim. Change 5, 483–491. [CrossRef]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. 2010. Microorganisms and climate change: terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 8, 779–790. [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. 2014. Belowground biodiversity and ecosystem functioning. Nature 515, 505–511. [CrossRef]
- Fellbaum, C.R.; Mensah, J.A.; Pfeffer, P.E.; Kiers, E.T.; Bücking, H. 2012. The role of carbon in fungal nutrient uptake and transport Implications for resource exchange in the arbuscular mycorrhizal symbiosis. Plant Signal. Behav. 7, 1509–1512. [CrossRef]
- Bonan, G. B. 2012. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320, 1444–1449. [CrossRef]
- Pan, Y.; et al. 2011. A large and persistent carbon sink in the world's forests. Science 333, 988–993. [CrossRef]
- Hovenden, M. J.; et al. 2019. Globally consistent influences of seasonal precipitation limit grassland biomass response to elevated CO2. Nat. Plants 5, 167–173. [CrossRef]
- Evans, R. D.; et al. 2014. Greater ecosystem carbon in the Mojave Desert after ten years exposure to elevated CO2. Nat. change.4, 394–397. [CrossRef]
- Davidson, T. A.; et al. 2018. Synergy between nutrients and warming enhances methane ebullition from experimental lakes. Nat. Clim. Change 8, 156–160. [CrossRef]
- van Bergen, T. J. H. M.; et al. Seasonal and diel variation in greenhouse gas emissions from an urban pond and its major drivers. Limnol. Oceanogr. [CrossRef]
- Jing, X.; et al. 2015. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nat. Commun. 6, 8159. [CrossRef]
- Delgado-Baquerizo, M.; et al. 2016. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 7, 10541. [CrossRef]
- Walker, T. W. N.; et al. 2018. Microbial temperature sensitivity and biomass change explain soil carbon loss with warming. Nat. Clim. Change 8, 885–889. [CrossRef]
- Altizer, S.; Ostfeld, R.S.; Johnson, P.T.; Kutz, S.; Harvell, C.D. 2013. Climate change and infectious diseases: from evidence to a predictive framework. Science 341, 514–519. [CrossRef]
- Harvell, C. D.; et al. 2002. Climate warming and disease risks for terrestrial and marine biota. Science 296, 2158–2162. [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. 2016. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 70, 317–340. [CrossRef]
- Frommel, A. Y.; et al. 2012A. Severe tissue damage in Atlantic cod larvae under increasing ocean acidification. Nat. Clim. Change 2, 42–46. [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. 2013. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Change 3, 30–36. [CrossRef]
- Raffel, T. R.; et al. 2013. Disease and thermal acclimation in a more variable and unpredictable climate. Nat. Clim. Change 3, 146–151. [CrossRef]
- Pounds, J. A.; et al. 2006. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439, 161–167. [CrossRef]
- MacFadden, D.R.; McGough, S.F.; Fisman, D.; Santillana, M.; Brownstein, J.S. 2018. Antibiotic resistance increases with local temperature. Nat. Clim. Change 8, 510–514. [CrossRef]
- Bouma, M.J.; Dye, C. 1997. Cycles of malaria associated with El Niño in Venezuela. JAMA 278, 1772–1774. [CrossRef]
- Baylis, M.; Mellor, P.S.; Meiswinkel, R. 1999. Horse sickness and ENSO in South Africa. Nature 397, 574. [CrossRef]
- Rohani, P. 2009. The link between dengue incidence and El Nino Southern Oscillation. PLOS Med. 6, e1000185. [CrossRef]
- Kreppel, K. S.; et al. 2014. A non-stationary relationship between global climate phenomena and human plague incidence in Madagascar. PLOS Neglect. Trop. Dis. 8, e3155. [CrossRef]
- Caminade, C.; et al. 2017. Global risk model for vector-borne transmission of Zika virus reveals the role of El Niño 2015. Proc. Natl Acad. Sci. USA 114, 119–124. [CrossRef]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. 2016. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ., 47, 20–33. [CrossRef]
- Trussell, P.C.; Baird, E.A. 1947. A rapid method for the assay of penicillin. Can. J. Res., 25, 5–8. [CrossRef]
- Rex, J.; Ghannoum, M.A.; Alexander, D.B.; Andes, D.; Brown, D.A.; Diekema, D.J.; Espinel-Ingroff, A.; Fowler, C.L.; Johnson, E.J.; Knapp, C.C.; et al. 2010. Method for Antifungal Disk Diffusion Susceptibility Testing of Nondermatophyte Filamentous Fungi; Approved Guideline; CLSI Document M51-A; Clinical and Laboratory Standards Institute: Wayne, PA, USA,; Volume 30, pp. 1–29.
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. 2016. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal., 6, 71–79. [CrossRef]
- Weinstein, M.P.; Patel, J.B.; Bobenchik, A.M.; Campeau, S.; Cullen, S.K.; Gallas, M.F.; Gold, H.; Humphries, R.M.; Kirn, T.J.; Lewis, J.S.; et al. 2019. Performance Standards for Antimicrobial Disk Susceptibility Tests: Approved Standard, 29th ed.; Clinical and Laboratory Standards Institute Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA,; Volume 32.
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. 1996 Comparison of three different in vitro methods of detecting synergy: Time-kill, checkerboard, and E test. Antimicrob. Agents Chemother., 40, 1914–1918. [CrossRef]
- Jiménez-Esquilín, A.E.; Roane, T.M. 2005. Antifungal activities of actinomycete strains associated with high-altitude sagebrush rhizosphere. J. Ind. Microbiol. Biotechnol., 32, 378–381. [CrossRef]
- Kumar, S.N.; Nambisan, B.; Sundaresan, A.; Mohandas, C.; Anto, R.J. 2014. Isolation and identification of antimicrobial secondary metabolites from Bacillus cereus associated with a rhabditid entomopathogenic nematode. Ann. Microbiol., 64, 209–218. [CrossRef]
- Menon, T.; Umamaheswari, K.; Kumarasamy, N.; Solomon, S.; Thyagarajan, S.P. 2001. Efficacy of fluconazole and itraconazole in the treatment of oral candidiasis in HIV patients. Acta Trop., 80, 151–154. [CrossRef]
- Imhof, A.; Balajee, S.A.; Marr, K.A. 2003. New Methods To Assess Susceptibilities of Aspergillus Isolates to Caspofungin. J. Clin. Microbiol., 41, 5683–5688. [CrossRef]
- Pfaller, M.A.; Sheehan, D.J.; Rex, J.H. 2004. Determination of Fungicidal Activities against Yeasts and Molds: Lessons Learned from Bactericidal Testing and the Need for Standardization. Clin. Microbiol. Rev., 17, 268–280. [CrossRef]
- Crouch, S.P.M.; Kozlowski, R.; Slater, K.J.; Fletcher, J. 1993. Use of ATP as a measure of cell proliferation and cell toxicity. J. Immunol. Methods, 160, 81–88. [CrossRef]
- Paloque, L.; Vidal, N.; Casanova, M.; Dumètre, A.; Verhaeghe, P.; Parzy, D.; Azas, N. 2013. A new, rapid and sensitive bioluminescence assay for drug screening on Leishmania. J. Microbiol. Methods, 95, 320–323. [CrossRef]
- Finger, S.; Wiegand, C.; Buschmann, H.J.; Hipler, U.C. 2013. Antibacterial properties of cyclodextrin-antiseptics-complexes determined by microplate laser nephelometry and ATP bioluminescence assay. Int. J. Pharm., 452, 188–193. [CrossRef]
- Galiger, C.; Brock, M.; Jouvion, G.; Savers, A.; Parlato, M.; Ibrahim-Granet, O. 2013. Assessment of efficacy of antifungals against Aspergillus fumigatus: Value of real-time bioluminescence imaging. Antimicrob. Agents Chemother., 57, 3046–3059. [CrossRef]
- Peyron, F.; Favel, A.; Guiraud-Dauriac, H.; El Mzibri, M.; Chastin, C.; Duménil, G.; Regli, P. 1997. Evaluation of a flow cytofluorometric method for rapid determination of amphotericin B susceptibility of yeast isolates. Antimicrob. Agents Chemother., 41, 1537–1540. [CrossRef]
- Paparella, A.; Taccogna, L.; Aguzzi, I.; Chaves-López, C.; Serio, A.; Marsilio, F.; Suzzi, G. 2008. Flow cytometric assessment of the antimicrobial activity of essential oils against Listeria monocytogenes. Food Control, 19, 1174–1182. [CrossRef]
- Landaburu, L.U.; Berenstein, A.J.; Videla, S.; Maru, P.; Shanmugam, D.; Chernomoretz, A.; Agüero, F. TDR Targets 6. 2020. Driving drug discovery for human pathogens through intensive chemogenomic data integration. Nucleic Acids Res., 48, D992. [CrossRef]
- Wondraczek, L.; Pohnert, G.; Schacher, F.H.; Köhler, A.; Gottschaldt, M.; Schubert, U.S.; et al. 2019. Artificial microbial arenas: materials for observing and manipulating microbial consortia. Adv. Mater. 31:1900284. [CrossRef]
- Stewart, E. J. 2012. Growing unculturable bacteria. J. Bacteriol. 194, 4151–4160. [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. 2002. Isolating "uncultivable" microorganisms in pure culture in a simulated natural environment. Science 296, 1127–1129. [CrossRef]
- Bollmann, A.; Lewis, K.; Epstein, S.S. 2007. Incubation of environmental samples in a diffusion chamber increases the diversity of recovered isolates. Appl. Environ. Microbiol. 73, 6386–6390. [CrossRef]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. 2017. In situ cultivation of previously uncultivable microorganisms using the ichip. Nat. Protoc. 12, 2232–2242. [CrossRef]
- Sizova, M. V.; Hohmann, T.; Hazen, A.; Paster, B.J.; Halem, S.R.; Murphy, C.M.; et al. 2012. New approaches for isolation of previously uncultivated oral bacteria. Appl. Environ. Microbiol. 78, 194–203. [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; et al. 2010. Use of Ichip for high-throughput in situ cultivation of "uncultivable" microbial species. Appl. Environ. Microbiol. 76, 2445–2450. [CrossRef]
- Sherpa, R.T.; Reese, C.J.; Montazeri Aliabadi, H. 2015. Application of iChip to grow "Uncultivable" microorganisms and its impact on antibiotic discovery. J. Pharm. Pharm. Sci. 18, 303–315. [CrossRef]
- Lodhi, A.F.; Zhang, Y.; Adil, M.; Deng, Y. 2018. Antibiotic discovery: combining isolation chip (iChip) technology and co-culture technique. Appl. Microbiol. Biotechnol. 102, 7333–7341. [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; et al. 2015. A new antibiotic kills pathogens without detectable resistance. Nature 517, 455–459. [CrossRef]
- Beneyton, T.; Wijaya, I.P.M.; Postros, P.; Najah, M.; Leblond, P.; Couvent, A.; et al. 2016. High-throughput screening of filamentous fungi using nanoliterrange droplet-based microfluidics. Sci. Rep. 6:27223. [CrossRef]
- Lin, Y.S.; Yang, C.H.; Lu, K.; Huang, K.S.; Zheng, Y.Z. 2011. Synthesis of agar microparticles using temperature-controlled microfluidic devices for Cordyceps militaris cultivation. Electrophoresis 32, 3157–3163. [CrossRef]
- Alkayyali, T.; Pope, E.; Wheatley, S.K.; Cartmell, C.; Haltli, B.; Kerr, R.G.; et al. 2021. Development of a microbe domestication pod (MD Pod) for in situ cultivation of micro-encapsulated marine bacteria. Biotechnol. Bioeng. 118, 1166–1176. [CrossRef]
- Xia, Y.; Whitesides, G.M. 1998. Soft Lithography. Angew. Chem. Int. Ed. 37, 550–575.
- Weibel, D.B.; Diluzio, W.R.; Whitesides, G.M. 2007. Microfabrication meets microbiology. Nat. Rev. Microbiol. 5, 209–218. [CrossRef]
- Zinchenko, A.; Devenish, S.R.A.; Kintses, B.; Colin, P.-Y.; Fischlechner, M.; Hollfelder, F. 2014. One in a million: flow cytometric sorting of single celllysate assays in monodisperse picolitre double emulsion droplets for directed evolution. Anal. Chem. 86, 2526–2533. [CrossRef]
- Wang, Z.; Volinsky, A.A.; Gallant, N.D. 2014. Crosslinking effect on polydimethylsiloxane elastic modulus measured by custom-built compression instrument. J. Appl. Polym. Sci. 134:2014. [CrossRef]
- Nichols, D.; Lewis, K.; Orjala, J.; Mo, S.; Ortenberg, R.; Connor, P.; et al. 2008. Short peptide induces an "uncultivable" microorganism to grow in vitro. Appl. Environ. Microbiol. 74, 4889. [CrossRef]
- CDC (2020). Antibiotic Resistance Threats in the United States 2019. Atlanta, GA: CDC.
- Hu, B.; Xu, P.; Ma, L.; Chen, D.; Wang, J.; Dai, X.; et al. 2021. One cell at a time: droplet-based microbial cultivation, screening and sequencing. Mar. Life Sci. Technol. 3, 169–188. [CrossRef]
- Boedicker, J.Q.; Li, L.; Kline, T.R.; Ismagilov, R.F. 2008. Detecting bacteria and determining their susceptibility to antibiotics by stochastic confinement in nanoliter droplets using plug-based microfluidics. Lab Chip 8, 1265–1272. [CrossRef]
- Li, Y.; Yang, X.; Zhao, W. 2017. Emerging microtechnologies and automated systems for rapid bacterial identification and antibiotic susceptibility testing. Slas Technol. 22, 585–608. D. [CrossRef]
- Lee, J.N.; Park, C.; Whitesides, G.M. 2003. Solvent compatibility of poly(dimethylsiloxane)-based microfluidic devices. Anal. Chem. 75, 6544–6554. [CrossRef]
Figure 1.
Illustrates how his idea completely changed laboratory titrations and serial dilutions.
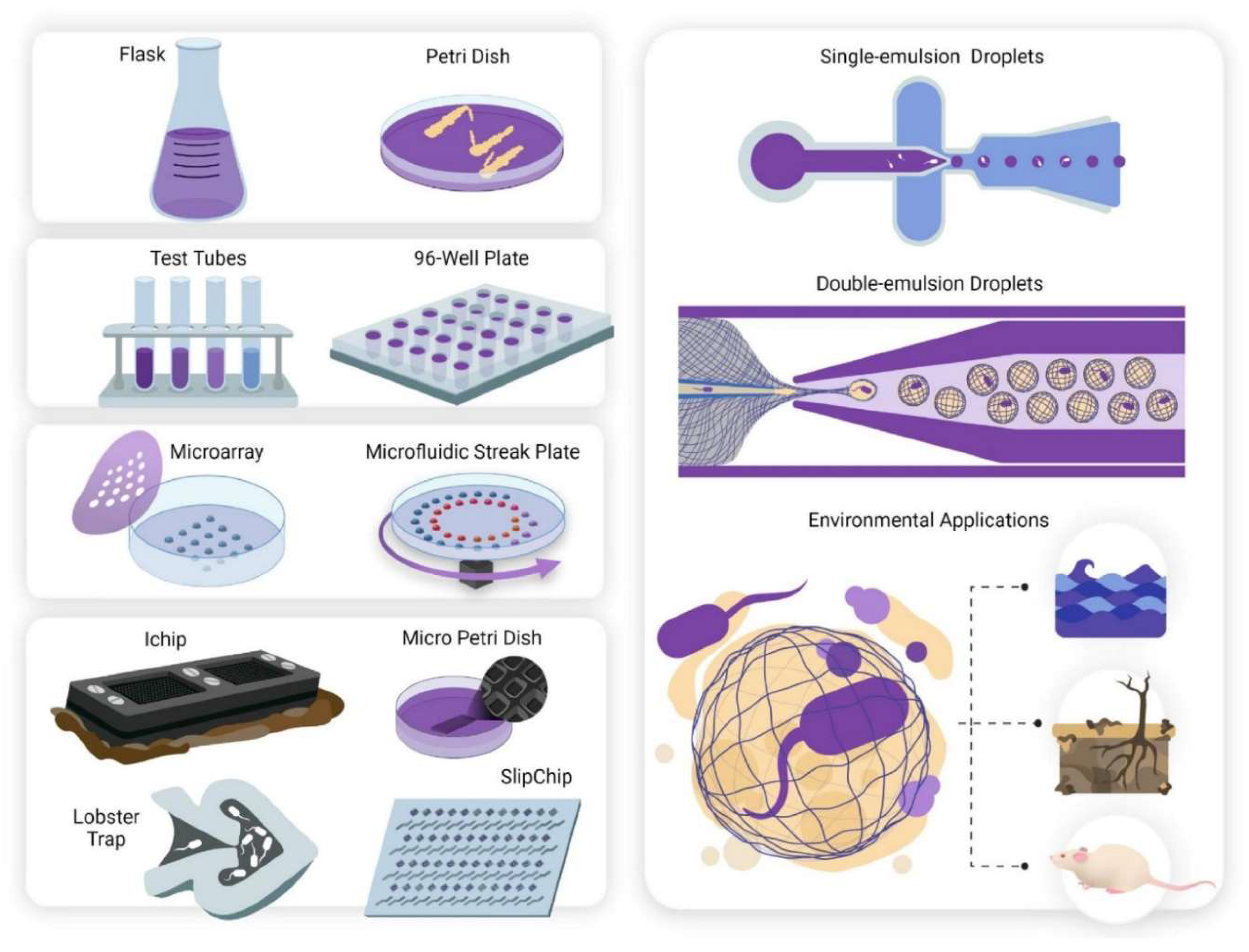
Figure 2.
Mechanism of action of antimicrobial peptides.
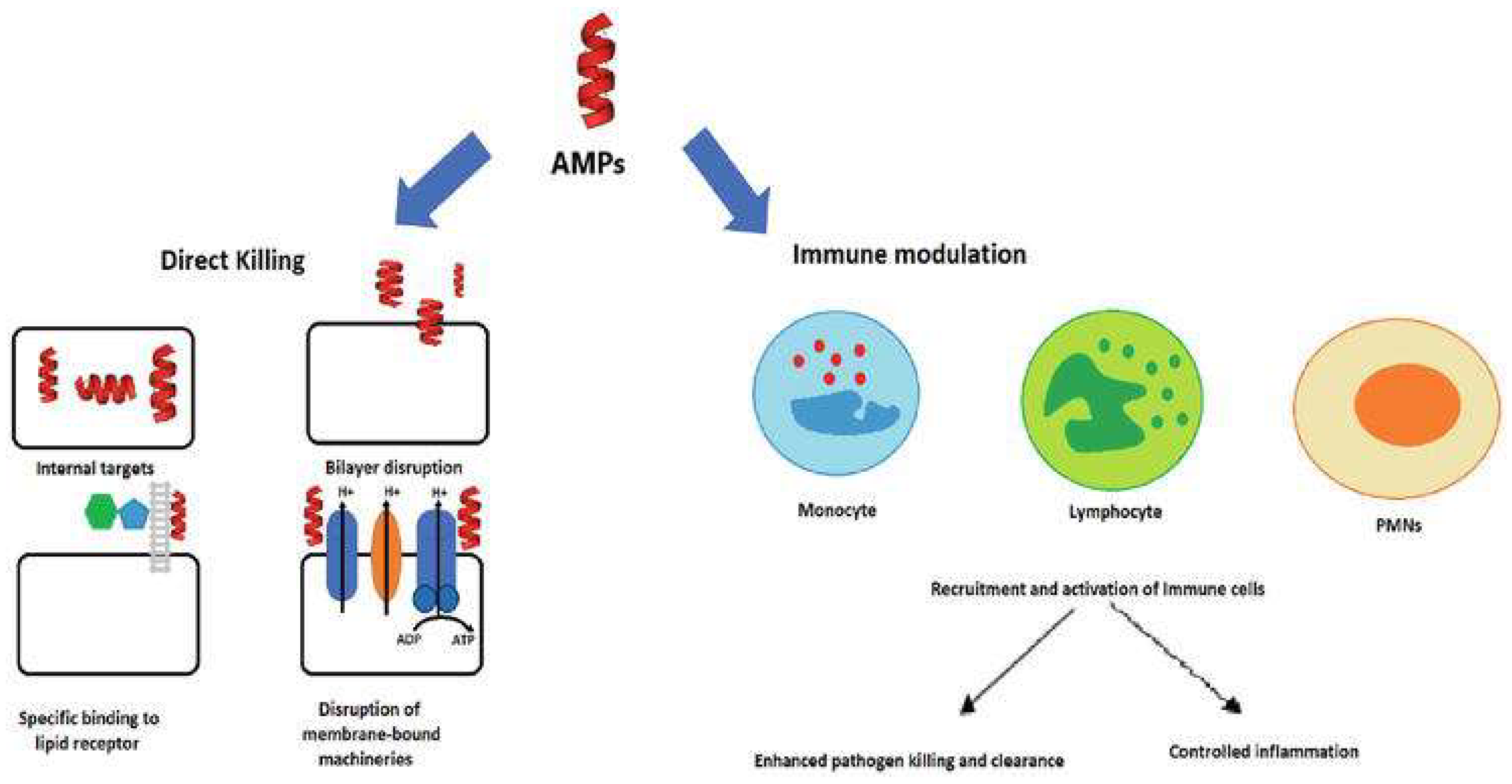
Figure 3.
Antiviral peptide P9, produced from mouse β-defensin, inhibits SARS-CoV-2 infection by binding to viral glycoprotein and masking host cell ACE2 receptors.
Figure 3.
Antiviral peptide P9, produced from mouse β-defensin, inhibits SARS-CoV-2 infection by binding to viral glycoprotein and masking host cell ACE2 receptors.
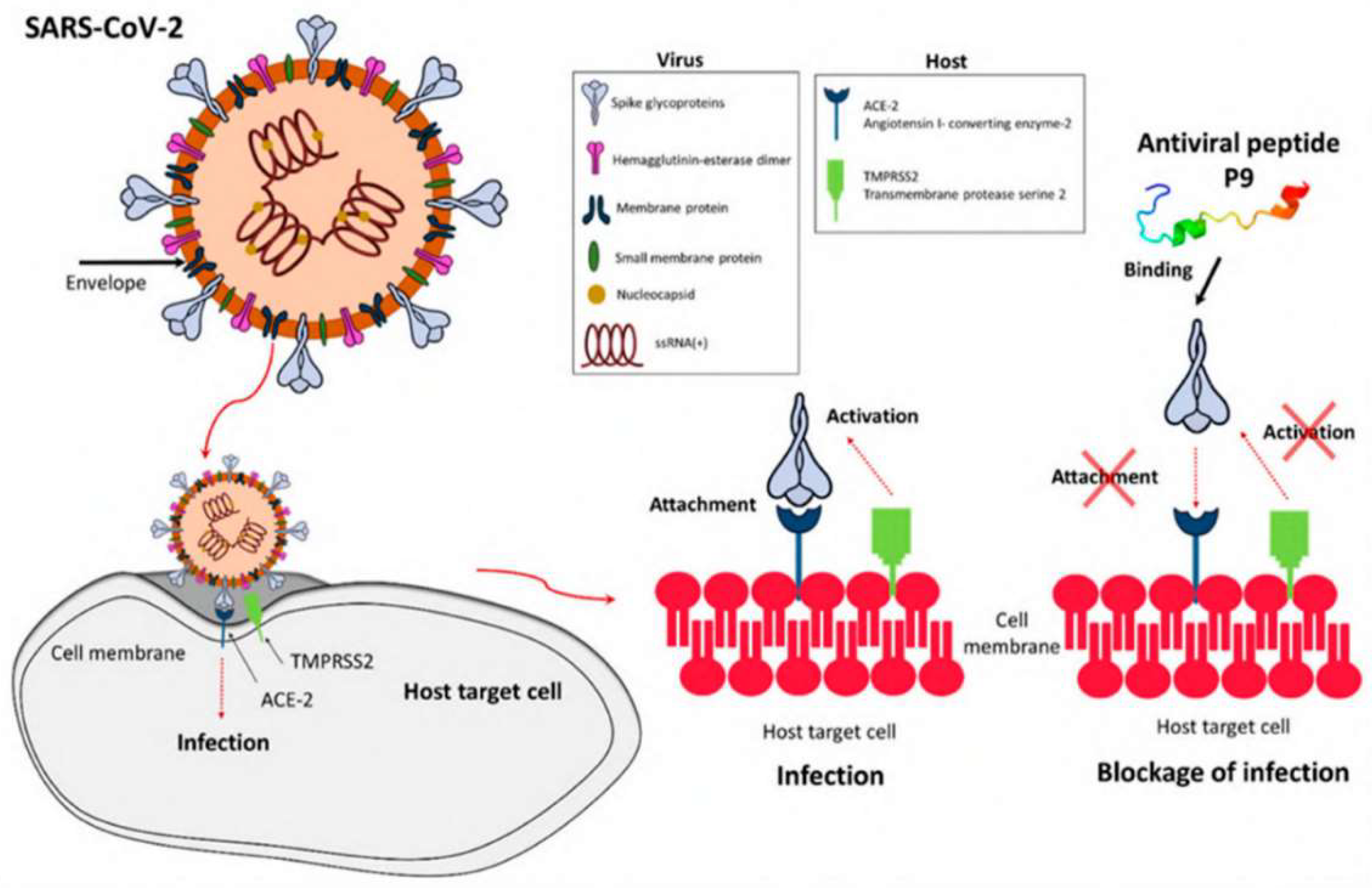
Figure 4.
Teixobactin was identified during the isolation and passage of Eleftheria terrae, a novel -proteobacteria.
Figure 4.
Teixobactin was identified during the isolation and passage of Eleftheria terrae, a novel -proteobacteria.

Table 1.
Chemical substances derived from microbes and their antimicrobial activity.
| Microorganism | Molecular Class | Chemical Compound | Antimicrobial Activity | Reference |
|---|---|---|---|---|
| Streptomyces scopuliridis | Cyclic peptide | Desotamide B | S. aureus, S. aureus | (Tortorella et al.,2018) |
| Marinactinospora thermotolerans | Cyclic peptide | Marthiapeptide A | S. aureus, M. luteus, B. subtillis, B. thuringiensis | |
| Verrucosispora spp. | Spirotetronate polyketides | Abyssomicin C | Staphylococcus aureus that is resistant to both methicillin and vancomycin | |
| Streptomyces drozdowiczii | Cyclic peptide | Marfomycins A, B, E | M. luteus | |
| Streptomyces spp. | Spirotetronate polyketides | Lobophorin H | B. subtilis | |
| Streptomyces spp. | Spirotetronate polyketides | Lobophorin F | S. aureus, E. feacalis | |
| Streptomyces niveus | Sesquiterpene derivative | Marfuraquinocin A, D | Methicillin-resistant Staphylococcus aureus, or simply S. aureus | |
| Streptomyces spp. | Alkaloid | Caboxamycin | S. epidermis, S. lentus, B. subtillis |
Table 2.
Specific bacteria that are vulnerable to bacteriocins and the species that make them.
| Bacteriocin | Susceptible Microorganisms | Bacteriocin Producer | Reference(s) |
|---|---|---|---|
| Nisin A | S. equinus, E. faecalis,. S. suis. S. epidermidis, S. aureus, S. dysgalactiae, S. uberis, S. agalactiae | Lactococcus lactic subsp. lactis | (Broadbent et al.,1989,Lebel et al.,2013,Carroll et al.,2010) |
| Nisin ANisin V | Listeria monocytogenes | L. lactis | (Campion et al.,2013) |
| Pediocin A | Clostridium perfringens | Pediococcus pentosaceus FBB61 | (Grili et al.,2009) |
| Enterocin M | Clostridium spp., Campylobacter spp. | Enterococcus faecium AL41 | (Laukova et al.,2018) |
| Enterocin CLE34 | Salmonella pullorum | Enterococcus faecium CLE34 | (Hernandez-Gonzalez et al.,2021),Wang et al.,2012) |
| Enterocin E-760 | Yersinia enterocolitica, E. coli, S. aureus, and Salmonella enterica serovars Enteritidis, Choleraesuis, Campylobacter jejuni; Typhimurium | Enterococcus faecium, Enterococcus durans, Enterococcus hirae | (Line et al.,2008) |
| Lacticin 3147 | S. agalactiae, S. dysgalactiae, S. aureus, S. uberis, paratuberculosis, Mycobacterium avium subsp. | Lactococcus lactis | (Carroll et al.,2010,Ryan et al.,1999) |
| Macedocin ST91KM | S. dysgalactiae, S. agalactiae, S. aureus, S. uberis | Streptococcus gallolyticus | (Pieterse et al.,2010) |
Table 3.
Bacterial origins of chemicals that inhibit fungal growth.
| Microorganism | Susceptible Organism(s) | Compound(s) | Reference |
|---|---|---|---|
| A. hibisca | Aspergillus spp., Candida spp. | Pradimicins A, B, C | (Tomita et al.,1990) |
| Actinoplanes spp. | T. mentagrophytes | Purpuromycin | (kerr et al.,1999) |
| B. cereus | Aspergillus spp., Saccharomyces spp, and C. albicans | Azoxybacilin, Bacereutin, Cispentacin, and Mycocerein | (kerr et al.,1999) |
| M. neiheumicin | S. cerevisiae | Neihumicin | (Boumehira et al.,2016) |
| Micromonospora species | A. cladosporium, C. albicans, and Cryptococcus spp. | Spartanamycin B | (Boumehira et al.,2016) |
| Micromonospora species SCC 1792 | Dermatophytes and Candida spp. | Sch 37137 | (Schwartz et al.,1989) |
| Micromonospora species SF-1917 | R. solania | Dapiramicins A and B | (Nishizawa et al.,1983) |
| B. subtilis | Phytopathogens | Iturin A | (kerr et al.,1999,Dunlap et al.,2019) , |
| B. lichenformis | Mucor spp., Microsporum canis, Sporothrix spp. | Fungicin M-4 | (Lebbadi et al.,1994,Galvez et al.,1993) , |
Table 4.
Biochemical substances derived from fungi that exhibit antimicrobial action.
| Microorganism | Antimicrobial Activity | Compounds | Reference(s) |
|---|---|---|---|
| Hormonema spp. | Aspergillus spp., Candida spp. | Enfumafungin | (Pelaez at al.,2000) |
| F. calocera | Mucor plumbeus, Candida tenuis | Favolon | (Chepkirui et al.,2016) |
| C. comatus | P. aeruginosa | Coprinuslactone | (Hyde et al.,2019) |
| Sanghuangporus spp. | C. albicans, S. aureus | Microporenic acid A | (Chepkirui et al.,2018) |
| Aspergillus terreus | Influenza A virus | Rubrolide S | (Zhu et al.,2014) |
| Cladosporium sphaerospermum | Influenza A H1N1 | Cladosin C | (Wu et al.,2014) |
| Penicillium sp. | HCV, HIV | β-hydroxyergosta-8,14,24 (28)-trien-7-one, Trypilepyrazinol | (Li et al.,2019) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



