Submitted:
06 September 2024
Posted:
06 September 2024
You are already at the latest version
Abstract
This study investigated the effects of dietary garlic powder and fermented garlic powder supplementation at 1% and 2% levels on growth performance, digestive tract efficacy, blood biochemistry, immunity, and antioxidant status of Liza ramada (n=225 fish; 86.00±0.42 g) over a 60-day period. Fish fed diets supplemented with both forms of garlic at both levels exhibited significantly improved final body weight, weight gain, specific growth rate, and feed conversion ratio compared to the control group. Digestive enzyme activities (amylase, lipase, and protease) were significantly enhanced in all supplemented groups. Blood biochemical analysis revealed reduced glucose levels and increased total protein in garlic-supplemented groups, with no adverse effects on liver or kidney function markers. Immune parameters, including lysozyme activity, bactericidal activity against Streptococcus agalactiae, alternative complement pathway (ACP), and, respiratory burst (NBT) were significantly enhanced in garlic-supplemented groups, with fermented garlic showing more pronounced effects. Antioxidant enzyme activities (SOD, CAT, and GPx) were also significantly increased in all supplemented groups, particularly in those fed fermented garlic. No significant differences in survival rates were observed among treatments. The results suggest that both garlic powder and fermented garlic powder supplementation, especially at the 2% level, can effectively improve growth, feed utilization, immune function, and antioxidant status in L. ramada. Fermented garlic generally demonstrated superior effects, indicating its potential as a beneficial feed additive in aquaculture. Based on these findings, it is recommended to incorporate fermented garlic powder at a 2% level in L. ramada diets to optimize growth performance and health status. Further research is warranted to investigate the long-term effects and cost-effectiveness of this supplementation strategy in commercial aquaculture settings.
Keywords:
Allium sativum
; Antioxidant status
; Digestive enzymes
; Feed additives
; Fermented garlic
; Growth performance
; Liza ramada
1. Introduction
Aquaculture, representing one of the fastest-growing sectors in global food production, is pivotal in addressing the increasing demand for fish protein [1]. Among the various species cultivated, grey mullet (Liza ramada) is particularly valued for its adaptability, resilience to varying environmental conditions, and high nutritional content, making it a prime candidate for sustainable aquaculture practices [2,3]. In recent years, L. ramada has become important for Egyptian fish farmers, aiming to enhance aquaculture practices in Egypt [4]. However, enhancing the growth performance and health of grey mullet remains a significant challenge due to the susceptibility of fish to diseases and suboptimal growth conditions in intensive farming systems [5].
In aquaculture, the primary challenges are ensuring a steady supply of feed and managing its cost [6]. Feed expenses significantly impact the sustainability of fish farming across various species, including mullets. As feed typically accounts for 30–70% of the total operational costs, it plays a critical role in determining the profitability of aquaculture ventures [7]. This makes the research into the ingredients, particularly additives that can enhance and consume the diets, crucial for gradually decreasing production costs [8,9]. The broad availability and affordability of eco-friendly, plant-based products have positioned them as the most promising alternatives to synthetic options [10]. The growing interest in the global aquaculture sector has been driven by the ongoing identification and development of plant ingredients as substitutes for antibiotics, aiming to mitigate their significant side effects [11,12].
The use of dietary supplements to enhance fish health and growth is a well-established practice in aquaculture [13]. o minimize antibiotic usage in intensive fish farming, garlic (Allium sativum) has gained attention as a powerful natural additive due to its wide range of bioactive compounds, including allicin, saponins, and flavonoids, which possess antimicrobial, antioxidative, and immunomodulatory properties [14]. These properties make garlic a promising candidate for improving fish health and the growth performance [15]. Fermentation, a traditional method to enhance the nutritional profile and bioavailability of food, further augments the benefits of garlic [16,17]. Fermented garlic, rich in enhanced bioactive components such as S-allyl mercaptocysteine and S-allyl cysteine, has shown superior antioxidative and immune-stimulating properties compared to its non-fermented counterpart [18]. The fermentation process increases the concentration of these beneficial compounds, potentially offering greater health benefits to animal species [19-21].
Garlic (A. sativum) has demonstrated notable benefits in aquaculture, improving growth, immune function, and disease resistance a wide range of fish species [15,22]. Allicin, a key compound in garlic, enhances growth by boosting digestion and nutrient absorption [23]. For instance, garlic supplementation of 1% enhanced growth in Oreochromis niloticus, with similar results in juvenile tilapia [24,25]. Garlic at 3% improved growth and feed conversion in Tilapia zillii [26], while 2% garlic polysaccharide extract significantly benefited Clarias gariepinus juveniles [27,28]. Garlic supplementation also enhanced growth in Mugil cephalus larvae [29] and Lates calcarifer [30].
In terms of immunostimulant effects, garlic boosts leukocyte counts and enhances phagocytic activity in various species. Studies have shown increased immune responses in O. niloticus, L. calcarifer, O. mykiss, and Cyprinus carpio with garlic supplementation [31-33]. Garlic also improves skin mucus lysozyme activity and blood protein levels, enhancing disease resistance [34,35].
Garlic's antimicrobial properties offer a safe alternative to antibiotics, effectively combating pathogens like Vibrio spp., E. coli, and Saprolegnia parasitica [15]. It has been shown to improve resistance to bacterial infections in species such as Labeo rohita, and O. mykiss [31,36]. Additionally, garlic has antifungal and antiparasitic effects, showing promise against pathogens such as Candida and Trichodina sp. [37,38]. However, garlic’s effectiveness can vary by species, dosage, and feed processing conditions, indicating the need for tailored applications to maximize benefits [39,40].
Despite the extensive research on the benefits of garlic in aquaculture, studies specifically examining the effects of fermented garlic on fish are notably scarce. To date, no comprehensive studies have explored the impact of fermented garlic on the growth, antioxidative capacity, immune responses, and intestinal histology of grey mullet (Liza ramada). This research thus represents the first report to investigate the comparative effects of non-fermented and fermented garlic in this species.
By elucidating the differential effects of these dietary interventions, this study aims to contribute to the development of sustainable and eco-friendly strategies for aquaculture, ultimately promoting the health and productivity of farmed fish. Understanding these effects could pave the way for innovative dietary approaches that enhance fish health and growth in an environmentally sustainable manner.
2. Materials and Methods
2.1. Ethical Considerations and Experimental Setup
This study was reviewed and approved by the Ethical Review Committee of the Faculty of Desert Agriculture at King Salman International University, Egypt (Reference No. KSIU/2024/DA-8). Experiments were carried out at the Baltim Research Station, National Institute of Oceanography and Fisheries, Egypt. L. ramada fish were obtained from a commercial farm in Kafr Elsheikh, Egypt, and acclimatized for two weeks in laboratory conditions. 225 healthy juvenile fish, each weighing approximately 86.00±0.42 grams, were evenly distributed into 15 well-aerated 100-liter tanks at a density of 15 fish per tank. They were fed experimental diets to satiety three times a day (7:00–7:30 AM, 1:00–1:30 PM, and 5:00–5:30 PM) over a 60-day period. Daily monitoring of water quality parameters showed dissolved oxygen (DO) at 6.71 ± 0.31 mg/L, temperature at 25.34 ± 0.22°C, and pH at 7.62 ± 0.54, using digital meters (Hanna, Romania, Do meter model HI 9,147, and pH meter model WT-80).
2.2. Source and Preparation of Garlic Forms
Standard garlic powder was prepared by initially peeling and slicing fresh garlic cloves. These were subsequently dehydrated using a vacuum oven at 60°C until a moisture content below 5% was achieved. The dried garlic was then ground into a fine powder using a laboratory-grade mill. Fermented garlic powder was produced through a controlled microbial process. Fresh garlic cloves were immersed in a 10% (w/v) sodium chloride solution inoculated with a starter culture of Lactobacillus plantarum and allowed to ferment for six weeks at a controlled temperature of 25°C. Following fermentation, the garlic cloves were meticulously rinsed with deionized water to eliminate residual salt and extraneous microorganisms. The cleaned cloves were then dehydrated to a moisture content below 5% using a vacuum oven at 60°C. Finally, the dehydrated garlic was ground into a fine powder using a laboratory-grade mill. HPLC analysis was employed to quantify the organosulfur compounds, key bioactive components in garlic. A reversed-phase C18 column was used with a gradient elution of methanol and water containing 0.1% formic acid. Detection was achieved using a UV detector at 254 nm.
2.3. Formulation of Experimental Diets
Five experimental diets were formulated, incorporating two forms of garlic (fermented and non-fermented) at levels of 1% and 2% per kg of diet, along with a control group without garlic. The diets were designed to be isonitrogenous (~33.42 ± 0.21 % crude protein) and isolipidic (~7.99 ± 0.27 % crude lipid) as outlined by Shehata, et al. [5]. Ingredients, including fish meal, soybean meal, fish oil, soybean oil, vitamins, and minerals [41], were sourced from a private feed factory in Kafr Elsheikh, Egypt (Table 1). Ingredients were weighed, ground, and mixed using an electric mixer, then water was added to form doughs, which were pelletized using a small-scale meat mincer (0.3 mm diameter, Regina Supernova pellet mill, Italy). The pellets were air-dried, packed into plastic bags, and stored at -20°C. Experimental diets were analyzed for moisture, dry matter, ash, crude protein, and crude fat following AOAC [42] procedures. Moisture content was determined by drying samples at 105°C, crude protein by the Kjeldahl method, crude fat by Soxhlet extraction, and ash content by incineration at 550°C.

Table 1.
Basal diet constituents and proximate composition.
| Ingredients | % | Proximate analysis | |
|---|---|---|---|
| Fish meal (65% CP) | 12 | Crude protein (%) | 33.42 ± 0.21 |
| Soybean meal (44% CP) | 38 | Crude lipids (%) | 7.99 ± 0.27 |
| Gluten | 6 | Fiber (%) | 5.11 ± 0.21 |
| Yellow corn | 20 | Ash (%) | 7.44 ± 0.22 |
| Wheat bran | 11 | Gross energy (MJ/Kg) 2 | 18.22 ± 0.33 |
| Rice bran | 7 | ||
| Fish oil | 4 | ||
| Mineral premix 1 | 0.5 | ||
| Vitamin premix 1 | 0.5 | ||
| Dicalcium phosphate | 1 | ||
| Total | 100 | ||
1 Vitamin and mineral mixtures described by Magouz et al. [41]. 2 The gross energy was multiplied using the values of 23.6 kJ/g for protein, 39.5 kJ/g for lipid, and 17.2 kJ/g for carbohydrates.
2.4. Sample Collection and Performance Metrics
After the 60-day feeding trial, fish were fasted for 24 hours and anesthetized with ethyl 3-aminobenzoate (MS-222; 100 μg/mL). Performance metrics and survival rates were assessed.
Where:
WG: weight gain; W60: weight at 60 days of feeding; W0: initial weight; 60: experimental period (day); SGR: specific growth rate; FI: feed intake; FCR: feed conversion ratio; SR: survival rate; N60: fish number at 60-day of feeding trial; N0: fish number at the beginning of the feeding period.
Twelve fish from each group (4 fish/tank) were dissected for gut samples for digestive enzyme and tissue evaluation. Four fish per tank were randomly selected for blood sampling using a 1 mL sterile syringe. Blood was stored at 4°C for 12 hours, centrifuged (4000 rpm, 4°C, 15 minutes) to obtain serum, and stored at –80°C for blood chemistry analysis.
2.5. Digestive Enzyme Activity
To prepare the gut samples, we were homogenized for 60 seconds in an ice-cold mannitol buffer (50 mM mannitol + 2 mM Tris-HCl, pH 7) at a 30:1 (v/w) ratio [43]. The homogenate was centrifuged at 9000 g for 10 minutes at 4°C, and the supernatant was further centrifuged at 34,000 g for 30 minutes at 4°C. The precipitate was resuspended in buffer (0.1 M KCl, 5 mM Tris-Hepes, 1 mM DTT; pH 7.5) for the evaluation of alkaline phosphatase (ALP), protease [44], α-amylase, and lipase [45].
2.6. Blood Biochemical Parameters
Biochemical parameters were meticulously evaluated using a comprehensive array of Bio-diagnostic kits from Bio-diagnostic®, Egypt. These included Glucose (Ref. No. GL 13 20), Albumin (Ref. No. AB 10 10), Triglyceride (Ref. No. TG 20 11), Total Cholesterol (Ref. No. TC 20 10), Total Protein (Ref. No. TP 20 20), ALT (glutamic pyruvic transaminase, Ref. No. AT 10 34), AST (glutamic oxaloacetic transaminase, Ref. No. AT 10 45), Urea (Ref. No. UR 21 10), and Creatinine (Ref. No. CR 12 50). Each parameter was assessed with the utmost precision, strictly following the manufacturer's detailed instructions, ensuring the accuracy and reliability of the results [2].
2.7. Antioxidant Activity Assessment
Enzyme activities of superoxide dismutase (SOD, wavelengths (WL) = 550 nm), catalase (CAT, WL = 280 nm) [46], and glutathione peroxidase (GPx, WL=412 nm) [47] were measured using ELISA kits from Cusabio Biotech Company, Ltd. (Wuhan, China) according to the manufacturer's instructions. These analyses were performed using a microplate spectrophotometer, following the manufacturer's instructions meticulously.
2.7. Immune Function Tests
The serum lysozyme level (LYZ) was quantified using a turbidometric assay, as described by Lygren, et al. [48]. To gauge serum bactericidal activity, we used an Escherichia coli suspension and assessed inhibition at 570 nm, following the method outlined by El Basuini, et al. [49]. Additionally, the oxidative burst of neutrophils was evaluated through nitroblue tetrazolium (NBT) assay, a technique detailed by Anderson and Siwicki [50]. To determine alternative complement pathway activities (ACP), we also applied the methods described by Yano [51] using serum samples.
2.8. Histological Examination of Intestine and liver
Intestine and liver samples were rinsed with sterile cold PBS (pH ~7.4) and immersed in 10% buffered formalin for two days. Samples were dehydrated, embedded in paraffin, sectioned to 5μm, and stained with hematoxylin and eosin [52,53]. High-resolution photomicrographs of the stained sections were captured using a Leica EC3 digital camera mounted on a Leica DM500 microscope (Leica, Germany).
2.9. Data Collection and Statistical Analysis
The data were collected and assessed for normal distribution and homogeneity of variance. Statistical analysis was conducted using SPSS (version 20). A one-way ANOVA was employed to determine the effects of dietary garlic on the means, with Duncan's multiple range test applied to identify significant mean differences. Results are presented as means ± standard errors (SEs) and significance was defined as p < 0.05.
3. Results
3.1. Garlic organosulfur compounds
Table 2 provides a comparative analysis of the concentration of key organosulfur compounds in both fermented and non-fermented garlic powder. In non-fermented garlic powder, allicin is present at a concentration of 3.2 mg/g, whereas in fermented garlic powder, this concentration dramatically decreases to 0.1 mg/g. Conversely, the concentration of diallyl sulfide (DAS) increases from 0.9 mg/g in non-fermented garlic powder to 2.1 mg/g in fermented garlic powder, indicating that fermentation enhances the formation of DAS. Similarly, diallyl disulfide (DADS) shows a substantial increase, with concentrations rising from 1.6 mg/g in non-fermented garlic to 4.8 mg/g in its fermented counterpart, suggesting enhanced bioactivity through fermentation. The level of S-allyl cysteine remains relatively stable, with a slight increase from 4.9 mg/g in non-fermented garlic to 5.1 mg/g in fermented garlic powder, highlighting its preservation during fermentation.
Table 2.
Organosulfur compounds in fermented and non-fermented garlic powder.
| Compound (mg/g) | Garlic Powder | Fermented Garlic Powder |
|---|---|---|
| Allicin (mg/g) | 3.2 | 0.1 |
| Diallyl sulfide (DAS) (mg/g) | 0.9 | 2.1 |
| Diallyl disulfide (DADS) (mg/g) | 1.6 | 4.8 |
| S-allyl cysteine (mg/g) | 4.9 | 5.1 |
3.2. Growth, feed utilization, and survival rate
Table 3 presents the performance of Liza ramada (growth, feed efficacy, survival%) fed experimental diets supplemented with garlic powder and fermented garlic powder at 1% and 2% levels for 60 days. Fish fed diets containing garlic powder or fermented garlic powder at both 1% and 2% levels exhibited significantly higher final body weight (FBW), weight gain (WG), and specific growth rate (SGR) compared to the control group (P < 0.05). The final body weights ranged from 243.50 ± 2.08 g to 248.15 ± 4.33 g for the supplemented groups, compared to 225.43 ± 2.93 g for the control group. The specific growth rate (SGR) was significantly improved in all supplemented groups (1.73-1.77 %/day) compared to the control (1.60 ± 0.03 %/day). Interestingly, there were no significant differences in growth parameters between the different types (garlic powder vs. fermented garlic powder) or levels (1% vs. 2%) of supplementation. Feed conversion ratio (FCR) was significantly lower in all supplemented groups (1.48-1.52) compared to the control group (1.65 ± 0.05), indicating improved feed utilization efficiency with garlic supplementation (P < 0.05). The lowest FCR values were observed in the fermented garlic groups (1.48 ± 0.03 and 1.48 ± 0.04 for 1% and 2%, respectively), although these were not statistically different from the garlic powder groups. Survival rates (SR%) were high across all experimental groups, ranging from 97.78% to 100%, with no significant differences observed between treatments.
Table 3.
Growth variables, feed efficacy, and survival rate of Liza ramada fed test diets for 60 days.
Table 3.
Growth variables, feed efficacy, and survival rate of Liza ramada fed test diets for 60 days.
| Item | Experimental diets | ||||
|---|---|---|---|---|---|
| Control | Garlic 1% | Garlic 2% | Fermented garlic 1% | Fermented garlic 2% | |
| Initial Body weight (IBW, g) | 86.25±0.20 | 86.08±0.31 | 86.08±0.12 | 85.81±0.38 | 85.78±0.21 |
| Final Body weight (FBW, g) | 225.43±2.93 b | 243.50±2.08 a | 246.31±4.67 a | 247.35±3.90 a | 248.15±4.33 a |
| Weight gain (WG, g) | 139.18±3.05 b | 157.42±1.95 a | 160.23±4.56 a | 161.53±3.54 a | 162.37±4.40 a |
| Specific growth rate (SGR, %/day) | 1.60±0.03 b | 1.73±0.01 a | 1.75±0.03 a | 1.77±0.02 a | 1.77±0.03 a |
| Feed conversion ratio (FCR) | 1.65±0.05 a | 1.52±0.01 b | 1.50±0.02 b | 1.48±0.03 b | 1.48±0.04 b |
| Survival rate (SR, %) | 97.78±2.22 | 100.00±0.00 | 97.78±2.22 | 100.00±0.00 | 100.00±0.00 |
Results are expressed as mean values ± standard error of the mean (SEM). Different lowercase letters indicate statistically significant differences between groups (P < 0.05).
3.3. Digestive enzymes
Table 4 presents the digestive enzyme activity of L. ramada after 60 days on experimental diets supplemented with garlic powder and fermented garlic powder at 1% and 2% levels. All three digestive enzymes examined (amylase, lipase, and protease) showed significantly increased activity in fish fed diets supplemented with either garlic powder or fermented garlic powder compared to the control group (P < 0.05). Amylase activity was significantly higher in all supplemented groups (ranging from 15.85 ± 1.09 to 16.44 ± 1.21 U/mg) compared to the control group (11.71 ± 0.64 U/mg). The highest amylase activity was observed in the fermented garlic 2% group, although there were no significant differences among the supplemented groups. Lipase activity showed a similar trend, with all supplemented groups exhibiting significantly higher activity (23.19 ± 0.75 to 24.49 ± 1.36 U/mg) compared to the control group (18.95 ± 0.94 U/mg). The fermented garlic 2% group showed the highest lipase activity, but again, there were no significant differences among the supplemented groups. Protease activity followed the same pattern, with significantly higher activity in all supplemented groups (20.04 ± 0.75 to 20.26 ± 1.36 U/mg) compared to the control group (15.77 ± 0.62 U/mg). As with the other enzymes, there were no significant differences in protease activity among the different garlic supplementation groups.
Table 4.
Digestive enzyme of Liza ramada fed experimental diets for 60 days.
| Enzyme activity (U/ mg) |
Experimental diets | ||||
|---|---|---|---|---|---|
| Control | Garlic 1% | Garlic 2% | Fermented garlic 1% |
Fermented garlic 2% |
|
| Amylase | 11.71±0.64 b | 15.85±1.09 a | 15.89±1.10 a | 16.35±1.08 a | 16.44±1.21 a |
| Lipase | 18.95±0.94 b | 23.19±0.75 a | 24.34±1.55 a | 24.22±1.08 a | 24.49±1.36 a |
| Protease | 15.77±0.62 b | 20.23±0.82 a | 20.22±1.29 a | 20.04±0.75 a | 20.26±1.36 a |
Results are expressed as mean values ± standard error of the mean (SEM). Different lowercase letters indicate statistically significant differences between groups (P < 0.05).
3.4. Intestine and liver histology
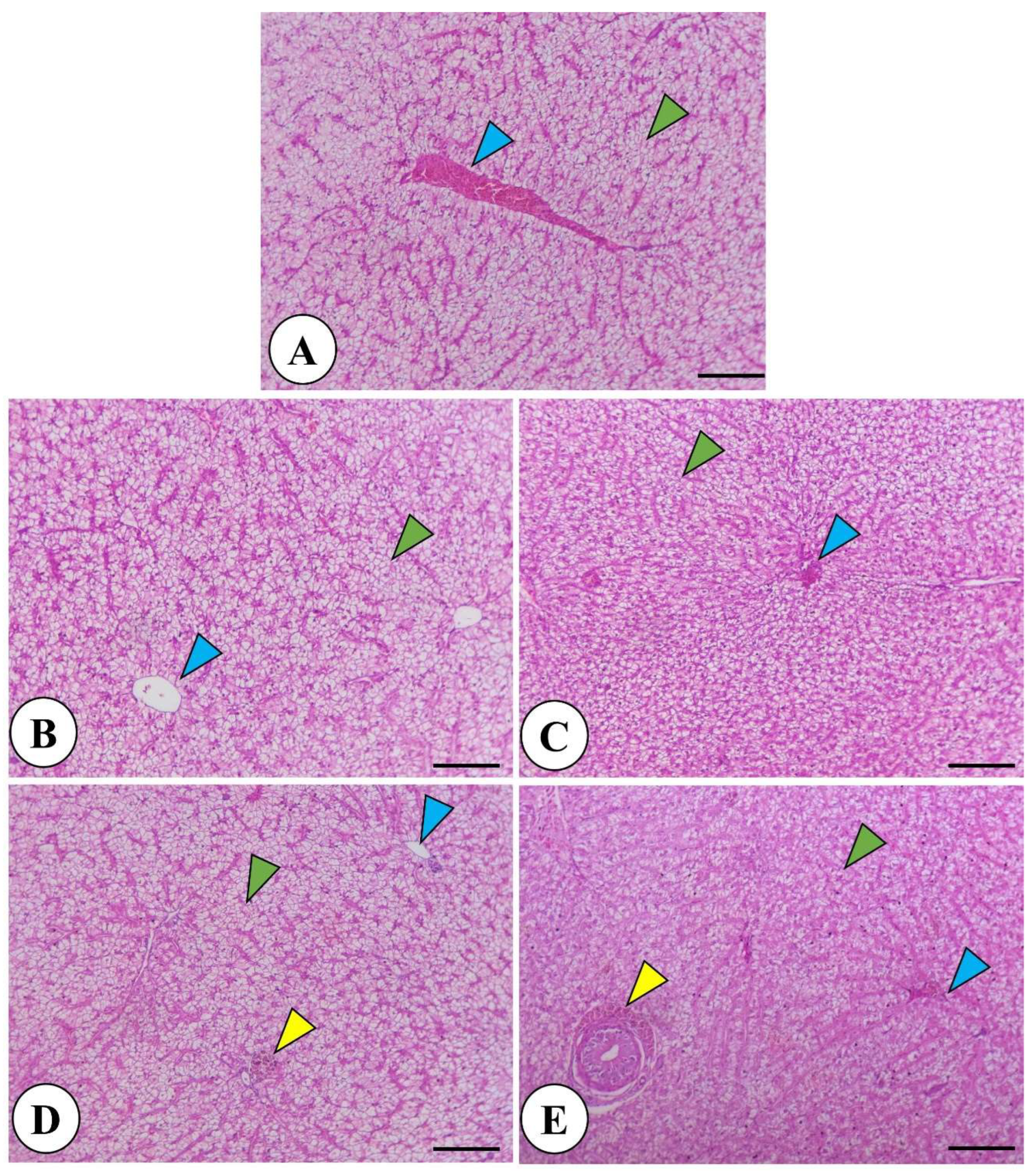
Histological examination of the L. ramada intestine in control and garlic-treated groups revealed a normal intestinal structure. Garlic supplementation led to increased intestinal villi length and branching. Intestine morphology showed normal mucosa and wall in all groups, with enhanced villi length, branching, and goblet cell number in garlic-supplemented fish. Immune cell infiltration was observed near villous crypts with the highest fermented garlic dose (Figure 1). The liver exhibited normal parenchyma with no inflammatory or degenerative changes in all groups. Fermented garlic powder significantly increased glycogen deposits and induced perivascular and periductal melanomacrophage accumulation (Figure 2).
Figure 1.
Histomicrograph of Liza ramada intestine in the control (A) and other treated groups by garlic powder at levels of 1 and 2% (B and C) and fermented garlic powder at levels of 1 and 2% (D and E). The morphological examination revealed normal intact structure of intestinal mucosa and wall in all groups with increased length and branching of intestinal villi (green arrowhead) as well as increased number of goblet cells (blue arrowhead) in line with supplemented garlic or fermented garlic in the fish diet. Immune cell infiltration (yellow arrowhead) was prominent near the villous crypts with fermented garlic at 2% (E). Stain H&E. Bar = 100 µm.
Figure 1.
Histomicrograph of Liza ramada intestine in the control (A) and other treated groups by garlic powder at levels of 1 and 2% (B and C) and fermented garlic powder at levels of 1 and 2% (D and E). The morphological examination revealed normal intact structure of intestinal mucosa and wall in all groups with increased length and branching of intestinal villi (green arrowhead) as well as increased number of goblet cells (blue arrowhead) in line with supplemented garlic or fermented garlic in the fish diet. Immune cell infiltration (yellow arrowhead) was prominent near the villous crypts with fermented garlic at 2% (E). Stain H&E. Bar = 100 µm.
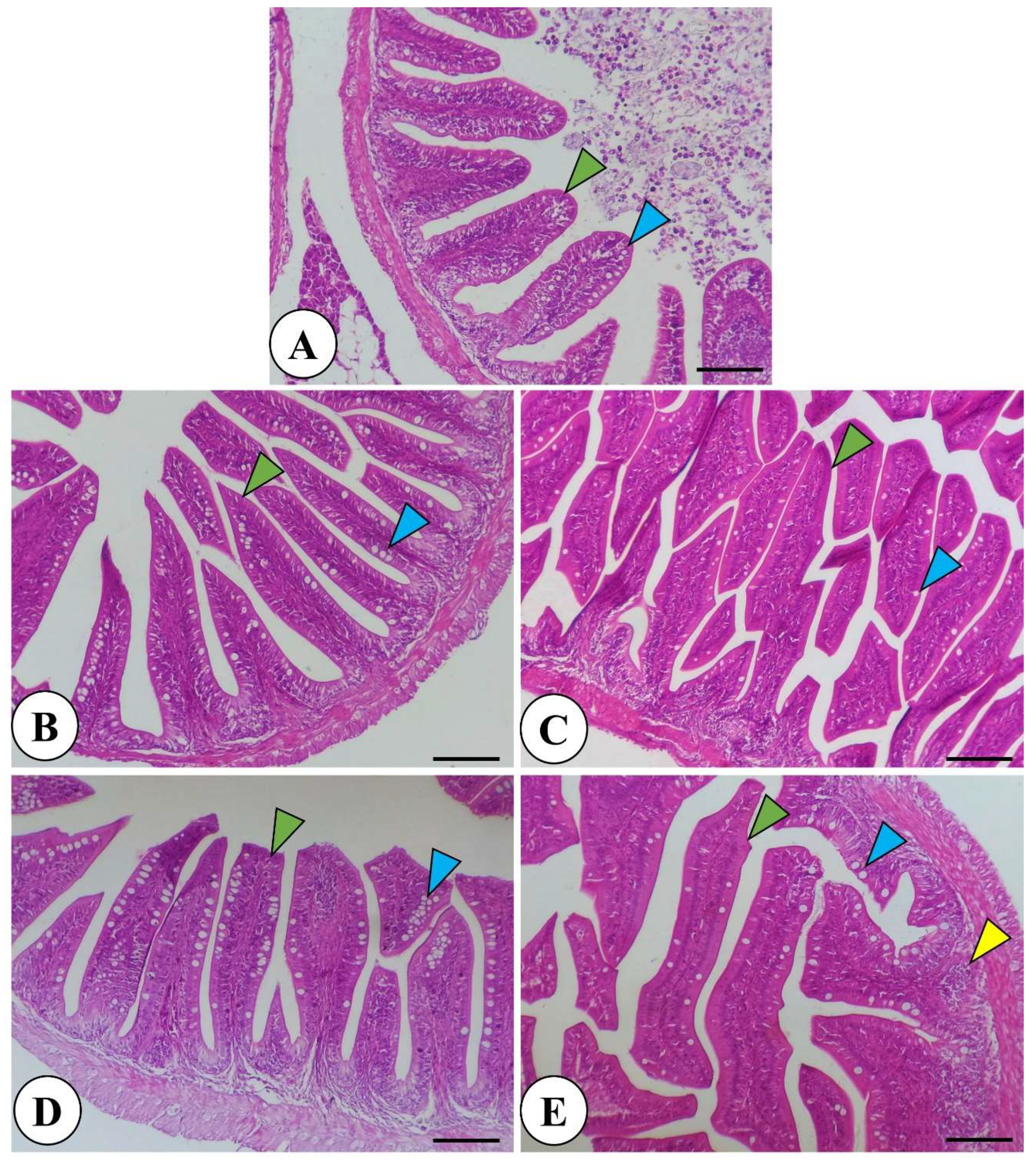
Figure 2.
Histomicrograph of Liza ramada liver in the control (A) and other treated groups by garlic powder at levels of 1 and 2% (B and C) and fermented garlic powder at levels of 1 and 2% (D and E). The morphological appearance revealed normal structure including hepatocytes (green arrowhead), central veins (blue arrowhead) and bile duct (yellow arrowhead). There is an increased glycogen deposit with fermented garlic powder at levels of 1 and 2%, in addition to periductal and perivascular accumulation of melanomacrophages (yellow arrowhead). Stain H&E. Bar = 100 µm.
Figure 2.
Histomicrograph of Liza ramada liver in the control (A) and other treated groups by garlic powder at levels of 1 and 2% (B and C) and fermented garlic powder at levels of 1 and 2% (D and E). The morphological appearance revealed normal structure including hepatocytes (green arrowhead), central veins (blue arrowhead) and bile duct (yellow arrowhead). There is an increased glycogen deposit with fermented garlic powder at levels of 1 and 2%, in addition to periductal and perivascular accumulation of melanomacrophages (yellow arrowhead). Stain H&E. Bar = 100 µm.
3.5. Blood biochemical parameters
Table 5 presents the blood biochemical parameters of L. ramada following a 60-day experimental period with diets supplemented with garlic powder and fermented garlic powder at 1% and 2% levels. Glucose levels were significantly lower (P < 0.05) in all garlic-supplemented groups (ranging from 88.00 ± 3.46 to 93.67 ± 4.26 mg/dl) compared to the control group (114.00 ± 5.20 mg/dl). The lowest glucose concentration was observed in the fermented garlic 2% group, although there were no significant differences among the supplemented groups. Total protein levels were significantly higher (P < 0.05) in all garlic-supplemented groups (6.68 ± 0.70 to 6.82 ± 0.25 g/dl) compared to the control group (5.10 ± 0.37 g/dl). The fermented garlic 2% group showed the highest total protein concentration, but there were no significant differences among the supplemented groups. Globulin levels showed a trend towards higher values in the garlic-supplemented groups (4.90 ± 0.38 to 5.09 ± 0.51 g/dl) compared to the control (3.48 ± 0.25 g/dl), although these differences were not statistically significant. No statistically significant differences (P > 0.05) were observed in blood levels of albumin, total cholesterol, triglycerides, ALT, AST, urea, or creatinine between the control and garlic-supplemented groups. However, there was a trend towards lower total cholesterol levels in the garlic-supplemented groups (9.00 ± 1.15 to 10.33 ± 1.45 mg/dl) compared to the control (13.33 ± 1.20 mg/dl), though this difference was not statistically significant.
Table 5.
Blood biochemical indices of Liza ramada fed test diets for 60 days.
| Items | Experimental diets | ||||
|---|---|---|---|---|---|
| Control | Garlic 1% | Garlic 2% | Fermented garlic 1% | Fermented garlic 2% | |
| Glucose (mg/ dl) | 114.00±5.20 a | 93.67±4.26 b | 92.00±4.00 b | 91.67±2.60 b | 88.00±3.46 b |
| Total Protein (g/ dl) | 5.10±0.37 b | 6.68±0.70 a | 6.69±0.40 a | 6.76±0.55 a | 6.82±0.25 a |
| Globulin (g/ dl) | 3.48±0.25 | 4.97±0.69 | 4.90±0.38 | 5.03±0.57 | 5.09±0.51 |
| Albumin (g/ dl) | 1.62±0.31 | 1.71±0.06 | 1.79±0.06 | 1.73±0.03 | 1.74±0.34 |
| Total Cholesterol (mg/ dl) | 13.33±1.20 | 10.33±1.45 | 10.00±1.15 | 10.00±1.53 | 9.00±1.15 |
| Triglyceride (mg/ dl) | 65.67±3.76 | 67.67±3.48 | 68.00±8.19 | 69.33±7.13 | 70.00±7.23 |
| ALT (IU/ l) | 54.67±4.26 | 56.67±3.28 | 57.67±4.81 | 61.33±5.46 | 60.67±5.36 |
| AST (IU/ l) | 56.00±4.36 | 57.00±4.73 | 56.00±5.86 | 57.67±7.22 | 54.67±5.81 |
| Urea (mg/ dl) | 10.00±0.58 | 10.33±0.88 | 11.00±1.53 | 9.33±1.86 | 9.67±1.45 |
| Creatinine (mg/ dl) | 0.27±0.07 | 0.27±0.03 | 0.30±0.10 | 0.23±0.09 | 0.30±0.12 |
Results are expressed as mean values ± standard error of the mean (SEM). Different lowercase letters indicate statistically significant differences between groups (P < 0.05).
3.6. Immunity
Figure 3 presents the immune parameters of L. ramada after a 60-day feeding experiment with diets supplemented with garlic powder and fermented garlic powder at 1% and 2% levels. Lysozyme activity showed a significant increase in all garlic-supplemented groups compared to the control (P < 0.05). The activity increased in a dose-dependent manner, with the highest values observed in the fermented garlic 2% group, followed by fermented garlic 1%, garlic 2%, and garlic 1%. The control group had the lowest lysozyme activity.
Figure 3.
Lysozyme activity, bactericidal activity, respiratory burst (Nitro-blue Tetrazolium, NBT) and serum alternative complement pathway (ACP) in the serum of Liza ramada fed the experimental diets for 60 days.
Figure 3.
Lysozyme activity, bactericidal activity, respiratory burst (Nitro-blue Tetrazolium, NBT) and serum alternative complement pathway (ACP) in the serum of Liza ramada fed the experimental diets for 60 days.
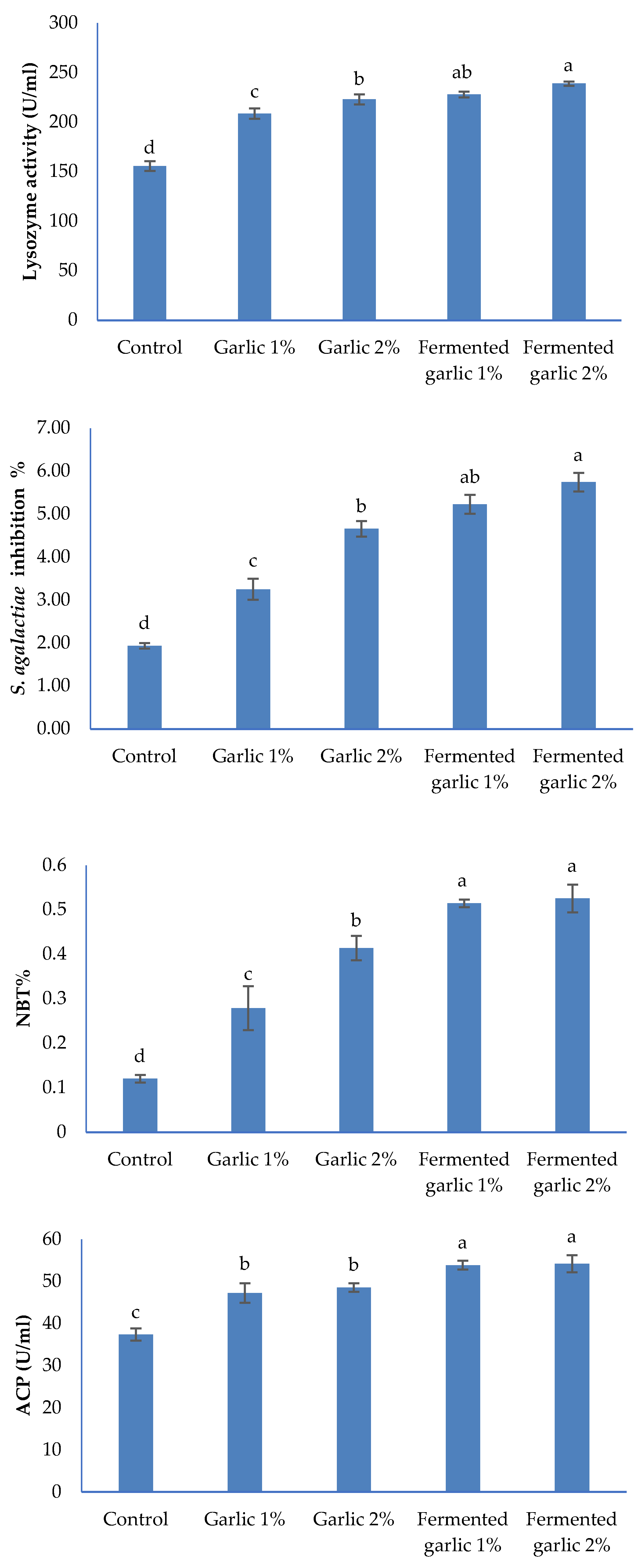
Bactericidal activity against S. agalactiae also demonstrated a significant increase in all supplemented groups compared to the control (P < 0.05). The fermented garlic 2% group showed the highest inhibition percentage, followed closely by fermented garlic 1%. Both garlic powder groups (1% and 2%) showed similar levels of inhibition, which were significantly higher than the control but lower than the fermented garlic groups.
The respiratory burst activity, measured by NBT reduction, exhibited a similar trend. All garlic-supplemented groups had significantly higher NBT% compared to the control (P < 0.05). The fermented garlic groups (1% and 2%) showed the highest NBT%, with no significant difference between them. The garlic powder groups also showed significantly increased NBT% compared to the control, with the 2% group having a higher value than the 1% group.
Serum alternative complement pathway (ACP) activity was significantly enhanced in all garlic-supplemented groups compared to the control (P < 0.05). Both fermented garlic groups (1% and 2%) showed the highest ACP activity, with no significant difference between them. The garlic powder groups (1% and 2%) also exhibited significantly increased ACP activity compared to the control, but at a lower level than the fermented garlic groups.
3.7. Antioxidants
Figure 4 demonstrates the antioxidant status in the serum of L. ramada after a 60-day feeding period with diets supplemented with garlic powder and fermented garlic powder at 1% and 2% levels. Superoxide dismutase (SOD) activity showed a significant increase in all garlic-supplemented groups compared to the control (P < 0.05). The highest SOD activity was observed in the fermented garlic groups (1% and 2%), with no significant difference between them. The garlic powder groups also exhibited significantly increased SOD activity compared to the control, with the 2% group showing higher activity than the 1% group.
Figure 4.
Superoxide dismutase (SOD),catalase (CAT), and glutathione peroxidase(GPx) of Liza ramada fed the experimental diets for 60 days.
Figure 4.
Superoxide dismutase (SOD),catalase (CAT), and glutathione peroxidase(GPx) of Liza ramada fed the experimental diets for 60 days.
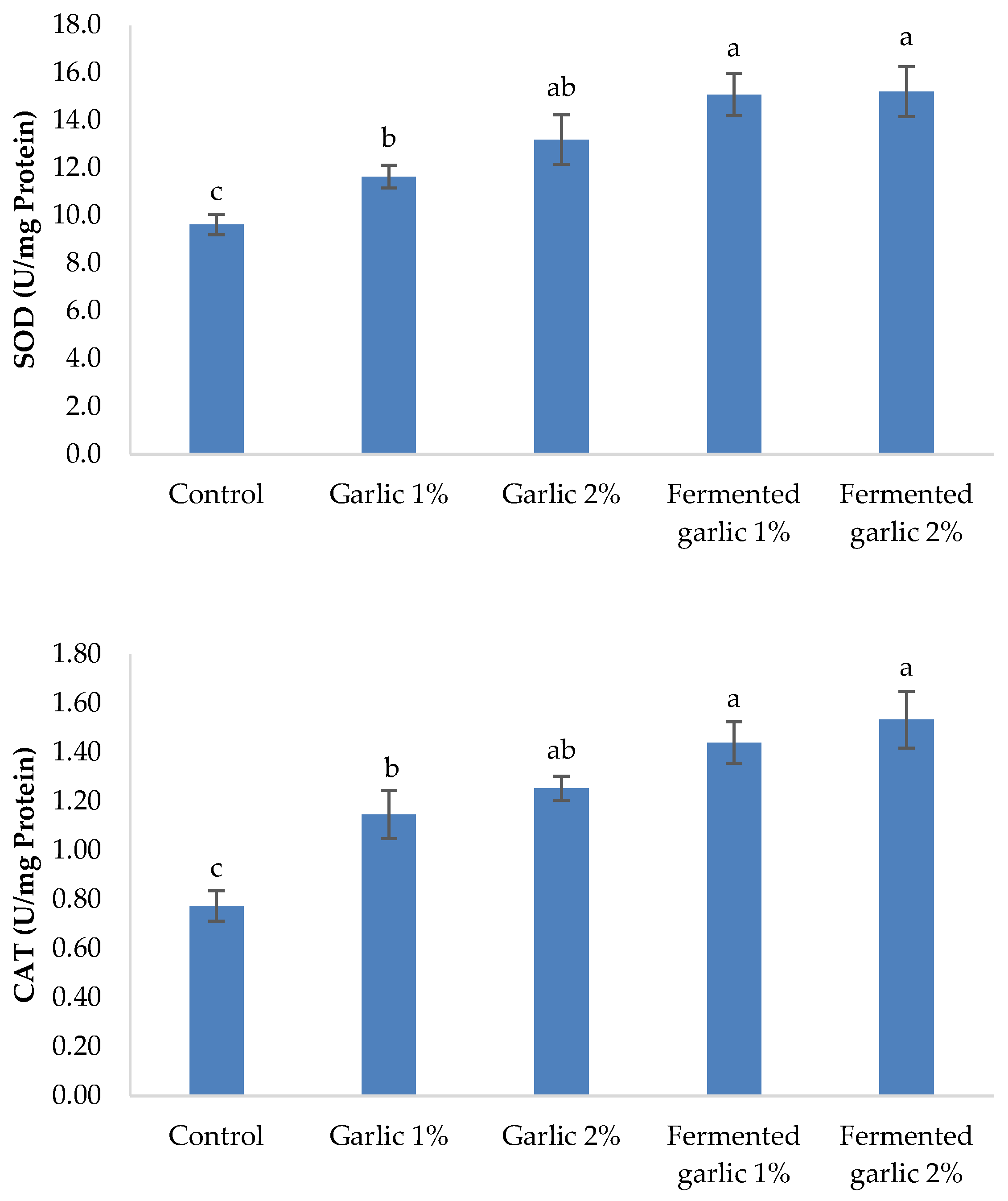
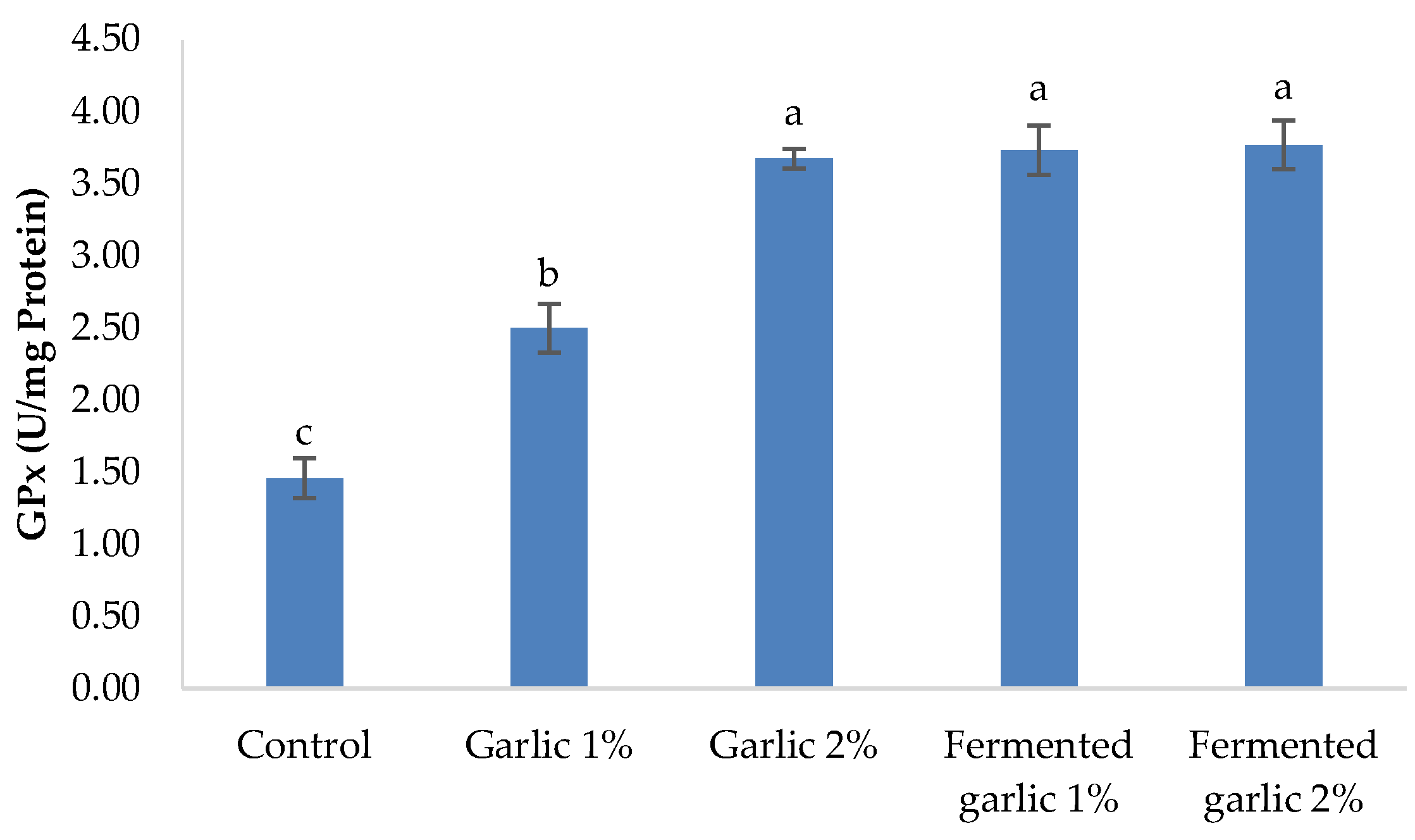
Catalase (CAT) activity demonstrated a similar trend, with all garlic-supplemented groups showing significantly higher activity compared to the control (P < 0.05). The fermented garlic groups (1% and 2%) exhibited the highest CAT activity, with no significant difference between them. The garlic powder groups also showed significantly increased CAT activity compared to the control, with the 2% group having higher activity than the 1% group, although this difference was not statistically significant.
Glutathione peroxidase (GPx) activity was significantly enhanced in all garlic-supplemented groups compared to the control (P < 0.05). The fermented garlic groups (1% and 2%) and the garlic powder 2% group showed the highest GPx activity, with no significant differences among them. The garlic powder 1% group also exhibited significantly increased GPx activity compared to the control, but at a lower level than the other supplemented groups.
4. Discussion
With the therapeutic efficacy of antibiotics diminishing for treating various bacterial infections in humans, several European countries have banned the use of dietary antibiotics [54]. This shift has spurred significant interest in identifying alternatives to antibiotics as growth promoters in the aquaculture industry. Among these alternatives, garlic (A. sativum) supplementation has emerged as a promising option for managing fish health due to its accessibility, affordability, and broad-spectrum effectiveness against pathogens. Research has demonstrated that garlic not only promotes fish growth but also enhances the immune system and controls pathogens, particularly bacteria and fungi [55].
The biological activity of organosulfur compounds, such as S-allyl-1-cysteine, diallyl disulfide, diallyl trisulfide, ajoene, and allicin, is fundamental to garlic's renowned healing properties [56]. Our study aligns with previous research, highlighting the significant impact of fermentation on the concentration and efficacy of these compounds. Notably, we observed a dramatic decrease in allicin concentration from 3.2 mg/g in non-fermented garlic to 0.1 mg/g in fermented garlic. Allicin, the primary bioactive component in raw garlic, is converted from alliin by the enzyme alliinase when garlic cloves are crushed [39]. Despite its numerous antimicrobial effects-including antibacterial, antifungal, antiviral, and antiparasitic activities [57] -allicin's instability results in its rapid decomposition during fermentation.
In contrast, our results show an increase in diallyl sulfide (DAS) from 0.9 mg/g in non-fermented garlic to 2.1 mg/g in fermented garlic, and a substantial rise in diallyl disulfide (DADS) from 1.6 mg/g to 4.8 mg/g. These findings indicate that fermentation enhances the formation of these more stable sulfur compounds. Diallyl disulfide and diallyl trisulfide, derivatives of allicin found in garlic essential oils, are known for their potent antimicrobial and antioxidant activities [58]. The increased concentrations of DAS and DADS suggest that fermentation could enhance garlic's antimicrobial efficacy, as sulfide compounds with more sulfur atoms exhibit stronger antimicrobial activity [59].
Additionally, the slight increase in S-allyl cysteine from 4.9 mg/g in non-fermented garlic to 5.1 mg/g in fermented garlic underscores the preservation of this important compound during fermentation. S-allyl cysteine is recognized for its antioxidant and neuroprotective properties, and its stability is beneficial for maintaining the health-promoting qualities of fermented garlic. Previous studies have shown that allicin induces an immediate and total inhibition of RNA synthesis, with RNA polymerase being a likely target due to allicin's reaction with thiol groups in enzymes [60]. However, the reduction of allicin and the corresponding increase in its derivatives like DADS and DAS in fermented garlic suggest that these compounds may play a more crucial role in the prolonged antimicrobial efficacy of fermented garlic products. These derivatives, identified as major components in garlic oil, contribute significantly to garlic’s overall bioactivity [61].
Generally, the fermentation process significantly alters the organosulfur compound profile of garlic, reducing allicin levels while increasing concentrations of more stable and bioactive compounds such as DAS and DADS. This transformation enhances the antimicrobial and antioxidant potential of garlic, potentially offering superior health benefits compared to non-fermented garlic. The preservation of S-allyl cysteine further highlights the benefits of fermentation in maintaining the therapeutic properties of garlic. These findings emphasize the importance of fermentation in enhancing the bioactivity and health-promoting qualities of garlic for dietary applications.
In the present study, garlic powder and fermented garlic powder significantly improve growth and feed efficiency in L. ramada. Both types of garlic supplementation at 1% and 2% levels resulted in higher FBW, WG, and SGR compared to the control group. Supplemented groups showed a lower FCR, indicating better feed utilization, with fermented garlic showing the lowest FCR values, though not significantly different from garlic powder groups. Survival rates were consistently high across all experimental groups.
These findings are consistent with previous studies showing the growth-promoting effects of garlic. For example, garlic supplementation at 1% level has been shown to improve growth parameters such as final weight and SGR in O. niloticus [62] and other species. Garlic powder has also been effective at 2% in Clarias gariepinus [28], and 3% in Mugil cephalus [29]. Similar positive effects have been reported for L. calarifer with varying garlic concentrations [30] and O. mykiss with garlic supplements [39].
However, garlic supplementation does not uniformly enhance growth across all fish species. Studies have shown no significant growth response in Perca fluviatilis juveniles [63] and Tambaqui (Colossoma macropomum) with varying garlic doses [64]. Red tilapia and hybrid tilapia (O. niloticus x O. aureus) also showed no significant growth improvement with garlic [65,66]. Additionally, Poecilia reticulata [67] and some studies on C. gariepinus found no significant weight gain with garlic supplementation [68].
Variations in growth responses may be due to factors such as fish species, diet composition, garlic dosage, and developmental stage [39]. The effectiveness of garlic might also be influenced by feed processing and the instability of allicin [69]. Overall, garlic powder and fermented garlic powder are effective at improving fish growth and feed utilization. While the specific type of garlic and dosage (1% and 2%) do not significantly affect growth parameters, fermentation seems to enhance feed efficiency.
It is important to note that the strong odor of garlic may significantly influence feed intake. Fermentation has been shown to improve the flavor of garlic products [70], potentially reducing the pungency and enhancing feed intake. Consequently, the fermented garlic powder used in this study might have contributed to increased feed consumption. However, to our knowledge, there are no existing studies evaluating the impact of fermented garlic powder on growth performance across various diets and species. Therefore, further research is needed to assess the effects of fermented garlic powder on growth performance in different fish species, particularly with different dietary formulations.
Garlic supplementation, as noted by Nya and Austin [71], boosts digestive enzyme activity in rainbow trout, which is consistent with our findings. Garlic enhances digestion by increasing bile acid secretion and stimulating enzymes such as lipase, trypsin, alkaline phosphatase, protease, and amylase [72]. This improved enzyme activity aids in the digestion of proteins, fats, and carbohydrates, promoting growth [73]. In the current study, fish-fed garlic powder or fermented garlic powder had higher amylase, lipase, and protease activities compared to the control. The 2% fermented garlic group had the highest enzyme levels, though differences among supplementation groups were not significant. Overall, both garlic forms effectively increased digestive enzyme activity, likely contributing to enhanced growth.
Histological examination of the intestine and liver is essential for evaluating the structural integrity and functional health of these organs in fish [74]. This study demonstrated that garlic supplementation led to significant improvements in intestinal morphology. Fish fed garlic-supplemented diets showed increased villi length, branching, and goblet cell number, while maintaining normal mucosa and intestinal wall structure. Notably, immune cell infiltration near villous crypts was observed, particularly at the highest dose of fermented garlic, suggesting an immunomodulatory effect. These enhancements in intestinal structure provide valuable insights into the fish's gastrointestinal health and nutrient absorption capacity [75].
Previous research supports these findings. Adibmoradi, et al. [76] reported histological and morphological changes in the intestines of fowl with 2% garlic supplementation, indicating garlic's potential to affect intestinal structure across species. Conversely, Agbebi, et al. [77] found no histological changes in Clarias gariepinus fed garlic-containing diets, highlighting species-specific responses. Mohammad [78] observed that garlic powder improved growth performance and histological properties in Cyprinus carpio, further supporting our results. These improvements indicate increased absorptive surface area and nutrient uptake capacity.
The liver, critical for detoxification and metabolic functions [79], also responded significantly to garlic supplementation. All groups exhibited normal liver parenchyma with no inflammatory or degenerative changes. However, fish fed fermented garlic powder showed a significant increase in glycogen deposits and perivascular and periductal melanomacrophage accumulation. These findings align with the outcomes observed in the present study, where garlic supplementation positively affected liver histology in Nile tilapia [80]. These effects might be attributed to specific interactions between garlic in different forms and the fish's metabolism, potentially influencing lipid metabolism and vascular integrity. These findings are consistent with the literature review by Valenzuela-Gutiérrez, et al. [15], which explored similar metabolic disruptions caused by dietary changes..
Taken together, the intestinal and liver changes in garlic-treated fish may reflect natural defense responses or adaptation to garlic. Garlic’s value lies in its disease-controlling effects with minimal side effects. While improvements in intestinal villi and glycogen deposition suggest benefits, further research is needed to understand the mechanisms and potential long-term impacts.
Evaluating biochemical parameters in fish is essential for understanding their physiological status, health, and response to dietary interventions [81]. In this study, glucose levels were significantly lower in all garlic-supplemented groups compared to the control group, with the lowest concentration observed in the 2% fermented garlic group. However, no significant differences were noted among the supplemented groups. These findings are consistent with those of Lee, et al. [82], who reported a hypoglycemic effect in juvenile Acipenser ruthenus fed a diet with 0.5% garlic extract, showing blood plasma glucose depletion after 1 hour (50.8 mg/dL) and 24 hours (57.6 mg/dL) post-feeding. Similarly, Thomson and Ali [83] and Kumar and Reddy [84] found that feeding mice 45 mg of garlic per kg body weight for 28 days significantly decreased serum glucose levels. Lower plasma glucose levels in fish have also been documented in studies assessing the physiological effects of A. sativum [63]. Garlic’s ability to decrease blood glucose is likely due to its effect on increasing serum insulin levels [85]. According to Ademiluyi, et al. [86] and Vazquez-Prieto, et al. [87], the S-allyl cysteine sulfoxide in garlic is responsible for its hypoglycemic activity.
Total protein levels were higher in all garlic-supplemented groups, with the 2% fermented garlic group showing the highest concentration, though differences among these groups were not statistically significant. There was also a trend towards higher globulin levels in the garlic-supplemented groups, although these differences were not significant. This aligns with findings that higher serum protein profiles, including total protein, albumin, and globulin levels, are linked to a stronger innate immune response and improved survival rates in fish [39,88]. Previous research has indicated that total plasma protein levels in fish typically range from 2 to 8 g/dL [89]. Sahu, et al. [36] found increased serum total protein, albumin, and glucose levels in L. rohita after 60 days of feeding with A. sativum compared to a control diet. Similarly, Nwabueze [90] observed elevated plasma protein levels in C. gariepinus when fed diets containing various concentrations of garlic.
No statistically significant differences were observed in blood levels of albumin, total cholesterol, triglycerides, ALT, AST, urea, or creatinine between the control and garlic-supplemented groups in this study. However, there was a trend towards lower total cholesterol levels in the garlic-supplemented groups. These results align with the findings of Abdelwahab, et al. [91], who reported significant decreases in serum total lipids and cholesterol in juvenile L. calcarifer treated with garlic. ÖZ and DİKel [92] also found a significant decrease in serum total lipids O. mykiss after garlic administration. Conversely, other studies, such as one on C. carpio, showed slight increases in serum triglyceride and cholesterol levels with traditional Chinese medicine extracts, though differences were not significant [93]. The variability in findings across studies may be due to differences in the composition and quantity of sulfur components in various garlic preparations, highlighting the need for standardization [18]. Other contributing factors could include subject recruitment, study duration, dietary control, lifestyle, and lipid analysis methods [94]. This study found that garlic extract is rich in proteins but low in lipids, which supports Lee, et al. [82], who reported that garlic inhibits cholesterol and fatty acid synthesis in the liver, though the precise mechanisms are not fully understood. Consequently, additional research is required to clarify the relationship between garlic dosage, application duration, biochemical effects, and fish species.
Allicin, the primary bioactive component of garlic, is well-documented for its antibacterial, antiparasitic, and antifungal properties [95]. It has the potential to reduce intestinal inflammation and exert an inhibitory immunomodulatory effect on intestinal epithelial cells [96]. Arreola, et al. [97] further suggested that Allium species enhance immune activities, including phagocytosis, cytokine release, natural killer cell activity, and lymphocyte synthesis.
Lysozyme, an enzyme crucial for its antimicrobial properties, is produced by leukocytes. It functions by breaking down microbial cell walls and activating the immune complement system [98]. In this study, lysozyme activity significantly increased in all garlic-supplemented groups compared to the control, with a dose-dependent effect. The highest activity was observed in the 2% fermented garlic group, followed by the 1% fermented garlic, 2% garlic powder, and 1% garlic powder groups, with the control group having the lowest activity. Previous studies have shown that garlic increases serum lysozyme activity in various fish species [36,99,100] and enhances skin mucus lysozyme activity, thereby boosting immunity and pathogen resistance [101]. Similar findings were reported in Salmo caspius [99], Sparus aurata [102] and Poecilia reticulata [67].
In fish, phagocytosis is a key defense mechanism that involves lysozyme, NBT, and ACH50 [45]. In the current study, all garlic-supplemented groups exhibited significantly increased bactericidal activity, respiratory burst activity, and ACP activity compared to the control. The 2% fermented garlic group showed the highest effects, followed by the 1% fermented garlic group. The garlic powder groups also showed significant increases but to a lesser extent than the fermented garlic groups.
Garlic enhances immunity by increasing phagocytic activity and respiratory burst in species such as Asian sea bass and rainbow trout [30,31], and stimulates superoxide anion production in Labeo rohita [36]. In some fish, the immunomodulatory effects of garlic are dose-dependent, as observed in hybrid tilapia [66]. However, these effects are not universal, as seen in juvenile Perca fluviatilis where no significant impact on respiratory burst or phagocytic activity was reported [63].
SOD, CAT, and GPx are integral components of the primary antioxidant defense system, protecting cells from free radicals generated during normal metabolism [103]. SOD is the initial enzyme responsible for converting superoxide radicals into hydrogen peroxide and oxygen [104]. Subsequently, catalase breaks down hydrogen peroxide into water and oxygen in peroxisomes, while GPx performs a similar function in mitochondria, where catalase is absent [103]. In this study, garlic supplementation led to a significant increase in SOD activity across all groups, with the highest levels observed in the 1% and 2% fermented garlic groups. CAT activity also showed substantial increases, particularly in the fermented garlic groups. Similarly, GPx activity was significantly elevated in all garlic-supplemented groups, with the highest levels found in the 2% fermented garlic group and 2% garlic powder group.
Garlic’s antioxidant benefits are attributed to its phenolic and saponin compounds, which inhibit free radical formation, enhance endogenous radical scavenging, and boost antioxidant enzyme activities, including SOD, CAT, and GPx [61]. These compounds also support cellular defense mechanisms against oxidative stress [105] and protect low-density lipoproteins from oxidative damage [106].
Previous research supports these findings. Metwally [24] demonstrated that garlic supplementation improved antioxidant enzyme activities in O. niloticus, including GPx, SOD, and CAT. Similarly, Mohebbi, et al. [107] found that garlic reduced lipid peroxidation and increased antioxidant enzyme activities in O. mykiss. Valenzuela-Gutiérrez, et al. [15] observed significant increases in GPx, CAT, and GR activities in trout-fed garlic and onion oils, further confirming garlic's role in enhancing antioxidant defenses. Overall, garlic supplementation effectively boosts key antioxidant enzymes in fish, supporting its use as a dietary additive to improve oxidative stress management and overall health in aquaculture.
5. Conclusions
This study reveals the remarkable potential of garlic supplementation in enhancing the growth, health, and metabolic efficiency of L. ramada. By shifting from non-fermented to fermented garlic, we observed a transformation in organosulfur compounds, with a dramatic increase in diallyl sulfide and diallyl disulfide, which are crucial for boosting the fish’s vitality and immune response. The incorporation of garlic, particularly in its fermented form, not only improved growth parameters and feed efficiency but also enriched digestive enzyme activity and antioxidant defenses. These enhancements were accompanied by better intestinal and liver health, underscoring the significant role of garlic in aquaculture nutrition. Based on these compelling results, it is recommended to include fermented garlic powder at a 2% level in L. ramada diets to achieve optimal growth and health outcomes. To fully capitalize on these benefits, further research is essential to explore the long-term impacts and economic feasibility of this supplementation in commercial aquaculture systems.
Author Contributions
Mohammed F. El Basuini: Conceptualization, Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Supervision, Project administration, Funding acquisition, Writing - Original Draft, Writing - Review & Editing. Mahasen M. E. A. Shaban: Methodology, Formal analysis, Investigation, Visualization. Abdelaziz M. El-Hais: Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Supervision. Ali A. Soliman: Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization. Nermeen M. Abu-Elala: Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Supervision. Islam I. Teiba: Conceptualization, Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Supervision, Project administration, Funding acquisition, Writing - Review & Editing. Mayada Alhoshy: Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Writing - Review & Editing. Ghada R. Sallam: Methodology, Software, Validation, Investigation, Resources, Data Curation, Visualization. Ronick Spenly Shadrack: Investigation, Resources, Data Curation, Visualization, Supervision, Funding acquisition, Writing - Original Draft, Writing - Review & Editing. Kumbukani Mzengereza: Resources, Data Curation, Visualization, Supervision, Funding acquisition, Writing - Original Draft. Akram Ismael Shehata: Conceptualization, Methodology, Software, Validation, Formal analysis, Investigation, Resources, Data Curation, Visualization, Supervision, Project administration, Funding acquisition, Writing - Original Draft, Writing - Review & Editing..
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of the Faculty of Desert Agriculture, King Salman International University (KSIU/2024/DA-8).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO, I., UNICEF, WFP, WHO. The State of Food Security and Nutrition in the World 2023. 2023.
- El Basuini, M.F.; Abdel Fattah, A.M.; El-Hais, A.M.; Soliman, A.A.; Amer, A.A.; Gewaily, M.; Zaki, M.A.A.; Zaineldin, A.I.; Dossou, S.; Teiba, I.I.; et al. Dietary co-enzyme Q10 boosted the growth performance, antioxidative capacity, immune responses, and intestinal and hepatic histomorphology of grey mullet (Liza ramada). Aquaculture Reports 2024, 36, 102147. [CrossRef]
- Shehata, A.I.; Shahin, S.A.; Elmaghraby, A.M.; Alhoshy, M.; Soliman, A.A.; Amer, A.A.; Habib, Y.J.; Gewaily, M.S.; El Basuini, M.F. Synergistic benefits of dietary silymarin and selenium on growth, immune functions, antioxidants, and gut/liver health of Thinlip mullet (Liza ramada) juveniles. Annals of Animal Science 2024, 0. [CrossRef]
- Shaalan, M.; El-Mahdy, M.; Saleh, M.; El-Matbouli, M. Aquaculture in Egypt: Insights on the Current Trends and Future Perspectives for Sustainable Development. Reviews in Fisheries Science & Aquaculture 2018, 26, 99-110. [CrossRef]
- Shehata, A.I.; Shahin, S.A.; Elmaghraby, A.M.; Alhoshy, M.; Toutou, M.M.; Soliman, A.A.; Amer, A.A.; Habib, Y.J.; Gewaily, M.S.; Teiba, I.I.; et al. Stevioside mitigates lead toxicity in thinlip mullet juveniles: Impacts on growth, metabolism, and immune function. Aquatic Toxicology 2024, 271, 106910. [CrossRef]
- Boyd, C.E.; D'Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso Jr, J.R.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. Journal of the World Aquaculture Society 2020, 51, 578-633. [CrossRef]
- Araujo, G.S.; Silva, J.W.A.d.; Cotas, J.; Pereira, L. Fish Farming Techniques: Current Situation and Trends. Journal of Marine Science and Engineering 2022, 10, 1598. [CrossRef]
- Pandey, A.K.; Kumar, P.; Saxena, M.J. Feed Additives in Animal Health. In Nutraceuticals in Veterinary Medicine, Gupta, R.C., Srivastava, A., Lall, R., Eds.; Springer International Publishing: Cham, 2019; pp. 345-362.
- Shehata, A.I.; Rasheed, M.; Rafiq, H.; Khalid, N.; Rafique, A.; Alhoshy, M.; Habib, Y.J.; El Basuini, M.F. Multi-functional application of octacosanol as a feed additive in animal and aquaculture: A review. Journal of Animal Physiology and Animal Nutrition 2024, n/a. [CrossRef]
- Hossain, M.S.; Small, B.C.; Kumar, V.; Hardy, R. Utilization of functional feed additives to produce cost-effective, ecofriendly aquafeeds high in plant-based ingredients. Reviews in Aquaculture 2024, 16, 121-153. [CrossRef]
- Eroldoğan, O.T.; Glencross, B.; Novoveska, L.; Gaudêncio, S.P.; Rinkevich, B.; Varese, G.C.; de Fátima Carvalho, M.; Tasdemir, D.; Safarik, I.; Nielsen, S.L.; et al. From the sea to aquafeed: A perspective overview. Reviews in Aquaculture 2023, 15, 1028-1057. [CrossRef]
- Alagawany, M. Antibiotic alternatives in poultry and fish feed; Bentham Science Publishers: 2022.
- Gatlin, D.M.; Yamamoto, F.Y. Chapter 11 - Nutritional supplements and fish health. In Fish Nutrition (Fourth Edition), Hardy, R.W., Kaushik, S.J., Eds.; Academic Press: 2022; pp. 745-773.
- Gupta, A.J.; Mainkar, P.; Mahajan, V. Exploring the Nutritional-Nutraceutical Composition and Phytochemical Potential of Garlic Agents in Preclinical and Clinical Studies With a Focus on Drug Likeness. Journal of Herbal Medicine 2024, 46, 100911. [CrossRef]
- Valenzuela-Gutiérrez, R.; Lago-Lestón, A.; Vargas-Albores, F.; Cicala, F.; Martínez-Porchas, M. Exploring the garlic (Allium sativum) properties for fish aquaculture. Fish Physiology and Biochemistry 2021, 47, 1179-1198. [CrossRef]
- Afzaal, M.; Saeed, F.; Rasheed, R.; Hussain, M.; Aamir, M.; Hussain, S.; Mohamed, A.A.; Alamri, M.S.; Anjum, F.M. Nutritional, biological, and therapeutic properties of black garlic: a critical review. International Journal of Food Properties 2021, 24, 1387-1402. [CrossRef]
- Shah, A.M.; Tarfeen, N.; Mohamed, H.; Song, Y. Fermented Foods: Their Health-Promoting Components and Potential Effects on Gut Microbiota. Fermentation 2023, 9, 118. [CrossRef]
- Ozma, M.A.; Abbasi, A.; Ahangarzadeh Rezaee, M.; Hosseini, H.; Hosseinzadeh, N.; Sabahi, S.; Noori, S.M.A.; Sepordeh, S.; Khodadadi, E.; Lahouty, M.; et al. A Critical Review on the Nutritional and Medicinal Profiles of Garlic’s (Allium sativum L.) Bioactive Compounds. Food Reviews International 2023, 39, 6324-6361. [CrossRef]
- Adi, A.C.; Susanti, Y.; Rachmawati, H.; Ishaura, E.R.; Farapti, F.; Salisa, W.; Rasyidi, M.F.; Sutthiwong, N. Effects of fermented and non-fermented garlic as an anti-diabetic on blood glucose levels of wistar rat. Journal of Public Health in Africa 2023.
- Lee, K.; Lee, K.; Kim, G.; Kim, J.; Yeon, J.; Cho, S.; Chang, B.; Kim, S. Effects of dietary fermented garlic on the growth performance, relative organ weights, intestinal morphology, cecal microflora and serum characteristics of broiler chickens. Revista Brasileira de Ciência Avícola 2016, 18, 511-518. [CrossRef]
- Lee, W.-D.; Kothari, D.; Moon, S.-G.; Kim, J.; Kim, K.-I.; Ga, G.-W.; Kim, Y.-G.; Kim, S.-K. Evaluation of Non-Fermented and Fermented Chinese Chive Juice as an Alternative to Antibiotic Growth Promoters of Broilers. Animals 2022, 12, 2742. [CrossRef]
- Chen, J.; Wang, F.; Yin, Y.; Ma, X. The nutritional applications of garlic (Allium sativum) as natural feed additives in animals. PeerJ 2021, 9, e11934. [CrossRef]
- Huang, X.; Zhao, Z.; Yang, Y.; Song, Q.; Ding, J.; Han, B.; Zhao, C. Dietary allicin improves behavior, physiology, growth, and disease resistance in the sea cucumber Apostichopus japonicus. Aquaculture 2024, 593, 741321. [CrossRef]
- Metwally, M. Effects of garlic (Allium sativum) on some antioxidant activities in tilapia nilotica (Oreochromis niloticus). World Journal of fish and marine sciences 2009, 1, 56-64.
- Setijaningsih, L.; Setiadi, E.; Taufik, I.; Mulyasari. The effect of garlic Allium sativum addition in feed to the growth performance and immune response of tilapia Oreochromis niloticus. IOP Conference Series: Earth and Environmental Science 2021, 744, 012072. [CrossRef]
- Ajiboye, O.O.; Qari, R. Short-term evaluation of graded levels of dietary garlic powder (allium sativum L.) as growth promoter on growth, survival and feed utilization of redbelly Tilapia, Tilapia zillii reared in glass aquaria tanks. International Journal of Marine Science 2016, 6. [CrossRef]
- Ayoola, S.; Uzoamaka, O. Effect of Allium sativum on growth, feed utilization and haematological parameters of Clarias gariepinus juvenile. African Journal of Livestock Extension 2013, 12, 1-7.
- Gabriel, N.N.; Wilhelm, M.R.; Habte-Tsion, H.-M.; Chimwamurombe, P.; Omoregie, E. Dietary garlic (Allium sativum) crude polysaccharides supplementation on growth, haematological parameters, whole body composition and survival at low water pH challenge in African catfish (Clarias gariepinus) juveniles. Scientific African 2019, 5, e00128. [CrossRef]
- Akbary, P.; Negahdari Jafarbeigi, Y.; Sondakzehi, A. Effects of garlic (Allium sativum L) extract on growth, feed utilization and carcass composition in Mugil cephalus (Linnaeus, 1758) larvae. Iranian Journal of Fisheries Sciences 2016, 15, 552-557.
- Talpur, A.D.; Ikhwanuddin, M. Dietary effects of garlic (Allium sativum) on haemato-immunological parameters, survival, growth, and disease resistance against Vibrio harveyi infection in Asian sea bass, Lates calcarifer (Bloch). Aquaculture 2012, 364-365, 6-12. [CrossRef]
- Nya, E.J.; Austin, B. Use of garlic, Allium sativum, to control Aeromonas hydrophila infection in rainbow trout, Oncorhynchus mykiss (Walbaum). Journal of Fish Diseases 2009, 32, 963-970. [CrossRef]
- Güroy, D.; Emre, N.; Yalım, F.B.; Karadal, O.; Kaya, D.; Arifoğlu, N. Interaction of dietary garlic (Allium sativum), onion (Allium cepa), and probiotic on the growth performance and health status of juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture International 2024, 32, 4515-4528. [CrossRef]
- Karimi Pashaki, A.; Ghasemi, M.; ZorriehZahra, M.J.; Shrif Rohani, M.; Hosseini, S.M. Effects of dietary garlic (Allium sativum) extract on survival rate, blood and immune parameters changes and disease resistance of Common carp (Cyprinus carpio carpio Linnaeus, 1758) against Spring Viremia of Carp (SVC). IFRO 2020, 19, 1024-1039.
- Ahmadniaye Motlagh, H.; Safari, O.; Selahvarzi, Y.; Baghalian, A.; Kia, E. Non-specific immunity promotion in response to garlic extract supplemented diets in female Guppy (Poecilia reticulata). Fish & Shellfish Immunology 2020, 97, 96-99. [CrossRef]
- Ghehdarijani, M.S.; Hajimoradloo, A.; Ghorbani, R.; Roohi, Z. The effects of garlic-supplemented diets on skin mucosal immune responses, stress resistance and growth performance of the Caspian roach (Rutilus rutilus) fry. Fish & Shellfish Immunology 2016, 49, 79-83. [CrossRef]
- Sahu, S.; Das, B.K.; Mishra, B.K.; Pradhan, J.; Sarangi, N. Effect of Allium sativum on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Journal of Applied Ichthyology 2007, 23, 80-86. [CrossRef]
- Sharma, N.; Behl, T.; Singh, S.; Bansal, A.; Singh, S.K.; Zahoor, I. Expatiating the therapeutic profile of garlic (Allium sativum): a bench to bedside approach. Biointerface Res. Appl. Chem 2021, 11, 14225-14239. [CrossRef]
- Chitmanat, C.; Tongdonmuan, K.; Nunsong, W. The use of crude extracts from traditional medicinal plants to eliminate Trichodina sp. in tilapia (Oreochromis niloticus) fingerlings. Songklanakarin J. Sci. Technol 2005, 27, 359-364.
- Adineh, H.; Harsij, M.; Jafaryan, H.; Asadi, M. The effects of microencapsulated garlic (Allium sativum) extract on growth performance, body composition, immune response and antioxidant status of rainbow trout (Oncorhynchus mykiss) juveniles. Journal of Applied Animal Research 2020, 48, 372-378. [CrossRef]
- Muahiddah, N.; Diamahesa, W.A. The use of garlic (Allium sativum) as an immunostimulant in aquaculture. Journal of Fish Health 2023, 3, 11-18. [CrossRef]
- Magouz, F.; Abu-Ghanima, H.; Zaineldin, A.I.; Gewaily, M.S.; Soliman, A.; Amer, A.A.; Moustafa, E.M.; Younis, E.M.; Abdel-Warith, A.-W.A.; Davies, S.J.; et al. Dietary Bacillus subtilis relieved the growth retardation, hepatic failure, and antioxidative depression induced by ochratoxin A in Thinlip Mullet (Liza ramada). Aquaculture Reports 2022, 22, 100984. [CrossRef]
- AOAC. Official methods of analysis of the Association of Official Analytical Chemists; Association of official analytical chemists: 2008.
- Gisbert, E.; Giménez, G.; Fernández, I.; Kotzamanis, Y.; Estévez, A. Development of digestive enzymes in common dentex Dentex dentex during early ontogeny. Aquaculture 2009, 287, 381-387. [CrossRef]
- Au - Cupp-Enyard, C. Sigma's Non-specific Protease Activity Assay - Casein as a Substrate. JoVE 2008, e899. [CrossRef]
- El Basuini, M.F.; Zalat, R.Y.I.; El-Hais, A.M.; Soliman, A.A.; Amer, A.A.; Gewaily, M.; Gabr, S.A.; Zaineldin, A.I.; Dossou, S.; Teiba, I.I.; et al. Bee venom enhances performance and immune function in thinlip mullet: A promising approach for sustainable aquaculture. Fish & Shellfish Immunology 2024, 151, 109713. [CrossRef]
- Paoletti, F.; Mocali, A. [18] Determination of superoxide dismutase activity by purely chemical system based on NAD(P)H oOxidation. In Methods in Enzymology; Academic Press: 1990; Volume 186, pp. 209-220.
- Carlberg, I.; Mannervik, B. [59] Glutathione reductase. Methods in Enzymology 1985, 113, 484-490. [CrossRef]
- Lygren, B.; Hjeltnes, B.; Waagbø, R. Immune response and disease resistance in Atlantic salmon (Salmo salar L.) fed three levels of dietary vitamin E and the effect of vaccination on the liver status of antioxidant vitamins. Aquaculture International 2001, 9, 401-411. [CrossRef]
- El Basuini, M.F.; Zaki, M.A.A.; El-Hais, A.M.; Elhanafy, M.G.; El-Bilawy, E.H.; Zaineldin, A.I.; Abdel-Aziz, M.F.A.; Abouelsaad, I.A.; El-Ratel, I.T.; Mzengereza, K.; et al. Microbial, immune and antioxidant responses of Nile tilapia with dietary nano-curcumin supplements under chronic low temperatures. Aquaculture and Fisheries 2024, 9, 57-65. [CrossRef]
- Anderson, D.; Siwicki, A. Basic hematology and serology for fish health programs. 1995.
- Yano, T. Assay of hemolytic complement activity. Techniques in Fish Immunology 1992, 131-141.
- Bancroft, J.D.; Gamble, M. Theory and practice of histological techniques; Elsevier health sciences: 2008.
- Gewaily, M.S.; Abumandour, M.M.A. Gross morphological, histological and scanning electron specifications of the oropharyngeal cavity of the hooded crow (Corvus cornix pallescens). Anatomia, Histologia, Embryologia 2021, 50, 72-83. [CrossRef]
- More, S.J. European perspectives on efforts to reduce antimicrobial usage in food animal production. Irish Veterinary Journal 2020, 73, 2. [CrossRef]
- Lee, J.-Y.; Gao, Y. Review of the Application of Garlic, Allium sativum, in Aquaculture. Journal of the World Aquaculture Society 2012, 43, 447-458. [CrossRef]
- Bisen, P.; Emerald, M. Nutritional and therapeutic potential of garlic and onion (Allium sp.). Current Nutrition & Food Science 2016, 12, 190-199.
- Nakamoto, M.; Kunimura, K.; Suzuki, J.I.; Kodera, Y. Antimicrobial properties of hydrophobic compounds in garlic: Allicin, vinyldithiin, ajoene and diallyl polysulfides. Experimental and therapeutic medicine 2020, 19, 1550-1553.
- Mnayer, D.; Fabiano-Tixier, A.-S.; Petitcolas, E.; Hamieh, T.; Nehme, N.; Ferrant, C.; Fernandez, X.; Chemat, F. Chemical Composition, Antibacterial and Antioxidant Activities of Six Essentials Oils from the Alliaceae Family. Molecules 2014, 19, 20034-20053. [CrossRef]
- Gong, X.; Su, X.; Liu, H. Diallyl Trisulfide, the Antifungal Component of Garlic Essential Oil and the Bioactivity of Its Nanoemulsions Formed by Spontaneous Emulsification. Molecules 2021, 26, 7186. [CrossRef]
- Gao, Y.; Wang, B.; Qin, G.; Liang, S.; Yin, J.; Jiang, H.; Liu, M.; Li, X. Therapeutic potentials of allicin in cardiovascular disease: advances and future directions. Chinese Medicine 2024, 19, 93. [CrossRef]
- Shang, A.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Tang, G.-Y.; Corke, H.; Mavumengwana, V.; Li, H.-B. Bioactive Compounds and Biological Functions of Garlic (Allium sativum L.). Foods 2019, 8, 246.
- Soltan, M.; El-L, S. Effect of probiotics and some spices as feed additives on the performance and behaviour of the Nile tilapia, Oreochromis niloticus. Egyptian Journal of Aquatic Biology and Fisheries 2008, 12, 63-80. [CrossRef]
- Zare, M.; Tran, H.Q.; Prokešová, M.; Stejskal, V. Effects of Garlic Allium sativum Powder on Nutrient Digestibility, Haematology, and Immune and Stress Responses in Eurasian Perch Perca fluviatilis Juveniles. Animals 2021, 11, 2735. [CrossRef]
- Inoue, L.A.K.A.; Oliveira Maciel, P.; Gusmão Affonso, E.; de Lima Boijink, C.; Tavares-Dias, M. Growth, parasitic infection and hematology in Colossoma macropomum Cuvier, 1818 fed diets containing Allium sativum. Journal of Applied Ichthyology 2016, 32, 901-905. [CrossRef]
- Samson, J. Effect of garlic (Allium sativum) supplemented diets on growth, feed utilization and survival of red tilapia (Oreochromis sp.). International Journal of Agricultural Technology 2019, 15, 637–644.
- Ndong, D.; Fall, J. The effect of garlic (allium sativum) on growth and immune responses of hybrid tilapia (Oreochromis niloticus x Oreochromis aureus). Journal of Clinical Immnunology and Immunopathology Research 2011, 3, 1-9.
- Motlagh, A., Hamidreza; Safari, O.; Selahvarzi, Y.; Baghalian, A.; Kia, E. Non-specific immunity promotion in response to garlic extract supplemented diets in female Guppy (Poecilia reticulata). Fish & Shellfish Immunology 2020, 97, 96-99. [CrossRef]
- Onomu, A.J. Growth and haematological response of Clarias gariepinus to garlic (Allium sativum) supplemented diet. Sustainable Agriculture Research 2018, 8, 67-73. [CrossRef]
- Mattos, D.C.; Manhães, J.; Cardoso, L.; Aride, P.; Lavander, H.; Oliveira, A.; Radael, M.; Azevedo, R.; Junior, V. Influence of garlic extract on larval performance and survival of juvenile angelfish Pterophyllum scalare during transport. Brazilian Journal of Biology 2021, 83, e244480. [CrossRef]
- Breidt, F.; McFeeters, R.F.; Perez-Diaz, I.; Lee, C.-H. Fermented Vegetables. In Food Microbiology; 2012; pp. 841-855.
- Nya, E.J.; Austin, B. Dietary modulation of digestive enzymes by the administration of feed additives to rainbow trout, Oncorhynchus mykiss Walbaum. Aquaculture Nutrition 2011, 17, e459-e466. [CrossRef]
- Platel, K.; Rao, A.; Saraswathi, G.; Srinivasan, K. Digestive stimulant action of three Indian spice mixes in experimental rats. Food / Nahrung 2002, 46, 394-398. [CrossRef]
- Esmaeili, N.; Abedian Kenari, A.; Rombenso, A.N. Effects of fish meal replacement with meat and bone meal using garlic (Allium sativum) powder on growth, feeding, digestive enzymes and apparent digestibility of nutrients and fatty acids in juvenile rainbow trout (Oncorhynchus mykiss Walbaum, 1792). Aquaculture Nutrition 2017, 23, 1225-1234. [CrossRef]
- Shehata, A.I.; Soliman, A.A.; Ahmed, H.A.; Gewaily, M.S.; Amer, A.A.; Shukry, M.; Abdel-Latif, H.M.R. Evaluation of different probiotics on growth, body composition, antioxidant capacity, and histoarchitecture of Mugil capito. Scientific Reports 2024, 14, 7379. [CrossRef]
- Abdel-Latif, H.M.R.; Soliman, A.A.; Gewaily, M.S.; Amer, A.A.; Shukry, M.; Khalil, R.H.; Shehata, A.I. Dietary effects of Saccharomyces cerevisiae and Allium sativum on growth, antioxidant status, hepatic and intestinal histoarchitecture, expression of growth- and immune-related genes, and resistance of Oreochromis niloticus to Aeromonas sobria. Fish & Shellfish Immunology 2024, 148, 109493. [CrossRef]
- Adibmoradi, M.; Navidshad, B.; Seifdavati, J.; Royan, M. Effect of Dietary Garlic Meal on Histological Structure of Small Intestine in Broiler Chickens. The Journal of Poultry Science 2006, 43, 378-383. [CrossRef]
- Agbebi, O.; Ogunmuyiwa, T.; Herbert, S. Effect of dietary garlic source on feed utilization, growth and Histopathology of the African catfish (Clarias gariepinus). Journal of Agricultural Science 2013, 5, 26. [CrossRef]
- Mohammad, M.A. Effect of adding garlic Allium sativum powder in diet on hematological, biochemical and histopathological criteria of common carp Cyprinus carpio L. Iraqi Journal of Agricultural Sciences 2023, 54, 1040-1049.
- Camargo, M.M.; Martinez, C.B. Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream. Neotropical ichthyology 2007, 5, 327-336. [CrossRef]
- Öz, M.; Inanan, B.E.; Üstüner, E.; Karagoz, B.; Dikel, S. Effects of dietary garlic (Allium sativum) oil on growth performance, haemato-biochemical and histopathology of cypermethrin-intoxicated Nile tilapia (Oreochromis niloticus). Veterinary Medicine and Science 2024, 10, e1449. [CrossRef]
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: a review of dietary interventions. Reviews in Aquaculture 2021, 13, 2190-2247. [CrossRef]
- Lee, D.H.; Lim, S.R.; Ra, C.S.; Kim, J.D. Effects of Dietary Garlic Extracts on Whole Body Amino Acid and Fatty Acid Composition, Muscle Free Amino Acid Profiles and Blood Plasma Changes in Juvenile Sterlet Sturgeon, Acipenser ruthenus. Asian-Australas J Anim Sci 2012, 25, 1419-1429. [CrossRef]
- Thomson, M.; Ali, M. Garlic [Allium sativum]: a review of its potential use as an anti-cancer agent. Current cancer drug targets 2003, 3, 67-81. [CrossRef]
- Kumar, G.R.; Reddy, K.P. Reduced nociceptive responses in mice with alloxan induced hyperglycemia after garlic (Allium sativum Linn.) treatment. Indian Journal of Experimental Biology 1999, 37, 662-666, http://nopr.niscair.res.in/handle/123456789/19100.
- Padiya, R.; Khatua, T.N.; Bagul, P.K.; Kuncha, M.; Banerjee, S.K. Garlic improves insulin sensitivity and associated metabolic syndromes in fructose fed rats. Nutrition & Metabolism 2011, 8, 53. [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Owoloye, T.R.; Agbebi, O.J. Modulatory effects of dietary inclusion of garlic (Allium sativum) on gentamycin–induced hepatotoxicity and oxidative stress in rats. Asian Pacific Journal of Tropical Biomedicine 2013, 3, 470-475. [CrossRef]
- Vazquez-Prieto, M.A.; Rodriguez Lanzi, C.; Lembo, C.; Galmarini, C.R.; Miatello, R.M. Garlic and Onion Attenuates Vascular Inflammation and Oxidative Stress in Fructose-Fed Rats. Journal of Nutrition and Metabolism 2011, 2011, 475216. [CrossRef]
- Gholipour Kanani, H.; Nobahar, Z.; Kakoolaki, S.; Jafarian, H. Effect of ginger- and garlic-supplemented diet on growth performance, some hematological parameters and immune responses in juvenile Huso huso. Fish Physiology and Biochemistry 2014, 40, 481-490. [CrossRef]
- Ravi, S.; Jitender, K. Fish Biology and Ecology Theory: Circulatory system of fish. University College of Sciences, Osmania University, Hyderabad 2005, 347.
- Nwabueze, A.A. The effect of garlic (Allium sativum) on growth and haematological parameters of Clarias gariepinus (Burchell, 1822). Sustainable Agriculture Research 2012, 1. [CrossRef]
- Abdelwahab, A.M.; El-Bahr, S.M.; Al-Khamees, S. Influence of Dietary Garlic (Allium sativum) and/or Ascorbic Acid on Performance, Feed Utilization, Body Composition and Hemato-Biochemical Parameters of Juvenile Asian Sea Bass (Lates calcarifer). Animals 2020, 10, 2396. [CrossRef]
- ÖZ, M.; DİKel, S. Effect of garlic (Allium sativum) - supplemented diet on growth performance, body composition and fatty acid profile of rainbow trout (Oncorhynchus mykiss). Cellular and Molecular Biology 2022, 68, 217-225. [CrossRef]
- Jia, R.; Hou, Y.; Feng, W.; Nomingerel, M.; Li, B.; Zhu, J. Multi-Omics Analysis to Understand the Effects of Dietary Proanthocyanidins on Antioxidant Capacity, Muscle Nutrients, Lipid Metabolism, and Intestinal Microbiota in Cyprinus carpio. Antioxidants 2023, 12. [CrossRef]
- Warshafsky, S.; Kamer, R.S.; Sivak, S.L. Effect of garlic on total serum cholesterol: a meta-analysis. Annals of internal medicine 1993, 119, 599-605. [CrossRef]
- Okoro, B.C.; Dokunmu, T.M.; Okafor, E.; Sokoya, I.A.; Israel, E.N.; Olusegun, D.O.; Bella-Omunagbe, M.; Ebubechi, U.M.; Ugbogu, E.A.; Iweala, E.E.J. The ethnobotanical, bioactive compounds, pharmacological activities and toxicological evaluation of garlic (Allium sativum): A review. Pharmacological Research - Modern Chinese Medicine 2023, 8, 100273. [CrossRef]
- Salehi, B.; Zucca, P.; Orhan, I.E.; Azzini, E.; Adetunji, C.O.; Mohammed, S.A.; Banerjee, S.K.; Sharopov, F.; Rigano, D.; Sharifi-Rad, J.; et al. Allicin and health: A comprehensive review. Trends in Food Science & Technology 2019, 86, 502-516. [CrossRef]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and Anti-Inflammatory Effects of Garlic Compounds. Journal of Immunology Research 2015, 2015, 401630. [CrossRef]
- Nawaz, N.; Wen, S.; Wang, F.; Nawaz, S.; Raza, J.; Iftikhar, M.; Usman, M. Lysozyme and Its Application as Antibacterial Agent in Food Industry. Molecules 2022, 27, 6305. [CrossRef]
- Zaefarian, A.; Yeganeh, S.; Adhami, B. Dietary effects of garlic powder (Allium sativum) on growth, blood indices, carcass composition, and lysozyme activity in brown trout (Salmo caspius) and resistance against Yersinia ruckeri infection. Aquaculture International 2017, 25, 1987-1996. [CrossRef]
- Jahanjoo, V.; Yahyavi, M.; Akrami, R.; Bahri, A.H. Influence of adding garlic (Allium sativum), Ginger (Zingiber officinale), thyme (Thymus vulgaris) and their combination on the growth performance, haematoimmunological parameters and disease resistance to Photobacterium damselae in sobaity sea bream (Sparidentex hasta) Fry. Turkish Journal of Fisheries and Aquatic Sciences 2018, 18, 633-645. [CrossRef]
- Dash, S.; Das, S.; Samal, J.; Thatoi, H. Epidermal mucus, a major determinant in fish health: a review. Iranian journal of veterinary research 2018, 19, 72.
- Firmino, J.P.; Fernández-Alacid, L.; Vallejos-Vidal, E.; Salomón, R.; Sanahuja, I.; Tort, L.; Ibarz, A.; Reyes-López, F.E.; Gisbert, E. Carvacrol, Thymol, and Garlic Essential Oil Promote Skin Innate Immunity in Gilthead Seabream (Sparus aurata) Through the Multifactorial Modulation of the Secretory Pathway and Enhancement of Mucus Protective Capacity. Frontiers in Immunology 2021, 12. [CrossRef]
- Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria journal of medicine 2018, 54, 287-293. [CrossRef]
- Winterbourn, C.C. Biological chemistry of superoxide radicals. ChemTexts 2020, 6, 7. [CrossRef]
- Liu, J.; Guo, W.; Yang, M.; Liu, L.; Huang, S.; Tao, L.; Zhang, F.; Liu, Y. Investigation of the dynamic changes in the chemical constituents of Chinese “Laba” garlic during traditional processing. RSC advances 2018, 8, 41872-41883. [CrossRef]
- Adegbola, P.I.; Fadahunsi, O.S.; Ajilore, B.S.; Akintola, A.O.; Olorunnisola, O.S. Combined ginger and garlic extract improves serum lipid profile, oxidative stress markers and reduced IL-6 in diet induced obese rats. Obesity Medicine 2021, 23, 100336. [CrossRef]
- Mohebbi, A.; Nematollahi, A.; Dorcheh, E.E.; Asad, F.G. Influence of dietary garlic (Allium sativum) on the antioxidative status of rainbow trout (Oncorhynchus mykiss). Aquaculture Research 2012, 43, 1184-1193. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



