Submitted:
04 September 2024
Posted:
05 September 2024
You are already at the latest version
Abstract
In mammals, fatty acid transport protein 1 (FATP1) is involved in cellular long-chain fatty acid (LCFA) uptake and activation, especially in processes of transportation, oxidation and triglyceride synthesis. However, the role of Fatp1 in invertebrates, especially decapod crustaceans, is still poorly understood. In this study, the cDNA of a fatp1 gene of a decapod crustacean, mud crab Scylla paramamosain, was cloned and functionally characterized. The fatp1 gene encoded a polypeptide of 643 amino acids that possessed all the typical features of the Fatp family, and shared high homology with the other Fatp orthologs of crustaceans. The relative expression levels of fatp1 were higher in metabolically active tissues such as hepatopancreas, stomach and gill than the other parts of crabs. Knockdown of the fatp1 gene in vivo reduced triglyceride and total lipid levels in the hepatopancreas, and increased the tissue expression levels of genes related to fatty acid transportation, allocation and hydrolysis including long-chain acyl-CoA synthetase 3/4 (acsl3/4) and carnitine palmitoyl transferase 1 (cpt1), and decreased expression levels of genes related to fatty acid synthesis such as acetyl-CoA carboxylase (acc) and fatty acid synthase (fas). Increased dietary n-3 LC-PUFA levels resulted in the up-regulation of fatp1 gene expression in the hepatopancreas, and the increase in LC-PUFA content, especially EPA and DHA, in both polar and nonpolar lipids in hepatopancreas and muscles of crabs. The data collectively suggested that the fatp1 gene, identified in S. paramamosain, might play important roles in regulating fatty acid absorption, utilization and allocation in crustaceans.
Keywords:
Scylla paramamosain
; Fatp1
; clone
; lipid accumulation
1. Introduction
Fatty acid transporter proteins (FATPs), also known as solute carrier family 27, are a family of proteins involved in the processes of cellular uptake and activation of long-chain fatty acids (LCFA) [1,2]. So far in mammalian genomes, six proteins have been identified as members of the FATP family, designated FATP1-6, all of which are 63-80 kDa integral membrane proteins with the N-terminus at the extracellular side, the C-terminus at the cytosolic side, and at least one transmembrane domain [2,3,4]. In addition, all FATP members have a highly conserved, 311-amino acid signature sequence known as the FATP sequence, as well as an AMP binding domain, located on the C-terminus [2], which is important for FATP-mediated fatty acid transport and commonly found in members of the long-chain acyl-CoA synthetase (ACSL) family [5,6]. Although the sequences of FATPs are similar, the expressed proteins exhibit highly distinct patterns of tissue distribution and each plays a discrete role in lipid homeostasis associated with fatty acid uptake [7,8].
FATP1, the first member of the FATP family to be identified through the screening of a cDNA library from 3T3-L1 adipocytes for LCFA uptake, showed a preference for a broad range of fatty acids and was highly expressed and abundant in tissues with rapid fatty acid metabolism, such as the heart, skeletal muscle, liver and adipose tissue [9,10,11]. Previous studies revealed that human FATP1 protein had 646 amino acids, containing the transmembrane domain, an AMP binding motif and multiple membrane-associated domains peripherally associated with the inner leaflet of the plasma membrane [4,9,12]. Gain- and loss-of-function studies have demonstrated that FATP1 is vital for cellular LCFA uptake and activation, and is involved in the processes of LCFA transportation and oxidation, triglyceride synthesis, lipid accumulation, and adipocyte differentiation [2,13]. Knockdown of FATP1 significantly reduced triglyceride accumulation and lipid droplet sizes in 3T3-L1 adipocytes [14], and LCFA uptake and lipid accumulation in adipocytes and skeletal muscle of Fatp1 knockout mice were severely reduced when compared to the non-knockdown control [15,16]. Similarly, using gain-of-function analysis, both LCFA uptake and lipid accumulation were found to be facilitated by overexpression of FATP1 in adipocytes, heart and skeletal muscle [17]. However, other studies reported that overexpression of Fatp1 did not increase lipid accumulation in rat L6E6 myotubes [18], and that lipid accumulation was inhibited by overexpression of Fatp1 in QM-7 (quail muscle) cells and mouse skeletal muscle [19,20]. While it was argued whether FATP1 can promote or inhibits fatty acid uptake and lipid accumulation in different cell types or tissues in mammals, further studies are clearly required to characterize and validate the function of Fatp1 in other non-mammalian animal species.
Therefore, although the mechanism is still poorly understood, FATP1 has been shown to play vital roles in LCFA absorption and activation in vertebrates [3,13]. Several Fatps or orthologs have also been identified and characterized in invertebrates. In wild-type Drosophila, Fatp is required for the expansion of lipid droplets (LD), which are indispensable for photoreceptor survival during aging in retinal pigment cells [21]. In silk moth Bombyx mori, Fatp was shown to facilitate LD accumulation and triacylglycerol synthesis within pheromone gland cells as a source of substrate for biosynthesis of the pheromone bombykol [22]. In Caenorhabditis elegans, mutants of two genes, acs-20 and acs-22, genes homologous to mammalian Fatp4, reduced the incorporation of exogenous very long chain fatty acids (VLCFA) into sphingomyelin, which is associated with severely disrupted cuticle surface barrier function [23]. Moreover, several studies have found that expression levels of fatp in decapoda crustaceans were affected by dietary fatty acid levels [24,25,26]. However, until now, the functions of Fatps involved in LCFA transport and activation in crustaceans have not yet been fully verified.
Mud crab (Scylla paramamosain), characterized by its large size, delicious meat, and high nutritional and economic value, is one of the most valuable crustaceans in the Indo-West Pacific region, and is cultured widely in the coastal areas of many Asian countries [27,28]. A high content of lipids and, especially, omega-3 long-chain polyunsaturated fatty acids (LC-PUFA), eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3), in the edible tissues such as hepatopancreas, ovary and muscle contribute to the nutritional value of mud crab [27,29]. Given the widely studied involvement of FATP1 in lipid metabolism of vertebrates, we identified and isolated the cDNA of a fatp1 gene from an invertebrate Scylla paramamosain and characterized its functional roles in lipid accumulation, with the aim of elucidating the involvement of Fatp1 in the absorption, utilization and allocation of dietary lipids in crustaceans. To our knowledge, this is the first report of the involvement of a Fatp1 in lipid accumulation in crustaceans.
2. Materials and Methods
2.1. Experimental Animals
All procedures performed on crabs were in accordance with the National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978) and approved by the Institutional Animal Care and Use Committee of Shantou University (Guangdong, China).
Juvenile mud crabs (S. paramamosain) used in the feeding experiment and in the RNA interference experiment were collected from the estuary of the Rongjiang River in Shantou city, China. The crabs were maintained in an indoor recirculating aquaculture system for two weeks and acclimated to brackish water (15 g/L) during which time they were fed a commercial crab feed. In addition, adult crabs for provision of tissues were obtained from the Niutianyang coastal area of Shantou city (Guangdong province, China), and cultured artificially by feeding them with Chinese mystery snail Cipangopaludina chinensis and small saltwater clam Potamocorbula rubromuscula in offshore ponds, where the ambient water temperature was 20.0 to 32.0 °C, pH 7.4 to 8.1, salinity 5.5 to 10.2 ppt, dissolved oxygen between 4.0 and 7.8 ppm, and ammonia-nitrogen < 0.5 ppm.
2.2. Feeding Trial
Ninety healthy crabs (initial average weight, 56.66 ± 1.57 g) were placed into individual polypropylene tanks (32 cm × 20 cm × 14 cm; Zhongkehai, Qingdao, China) and allocated randomly into five dietary treatment groups with each treatment consisting of 18 crabs. Crabs were fed with 5 isoproteic (41.7 % crude protein) and isolipidic (7.8 % crude lipid) diets containing graded levels (0.50, 0.73, 1.04, 1.42 and 2.57 % of dry weight, respectively) of n-3 LC-PUFA, predominantly EPA and DHA, with a constant level (0.52 % of dry weight) of n-6 LC-PUFA, mainly arachidonic acid (ARA, 20:4n-6). The experimental diets were prepared and stored as described in detail previously (Zhang et al., 2019), and formulations and analyzed proximate compositions are shown in Supplemental Table S1. During the feeding trial, crabs were fed to apparent satiety twice a day at 7:00 and 18:00, the water temperature ranged between 25 °C and 28 °C, dissolved oxygen between 4.0 - 6.0 mg/L, ammonia nitrogen < 0.05 mg/L, and salinity was about 15 g/L. After the 9-week feeding trial, the crabs were starved for 24 h and chilled on ice to anesthetize, and then the hepatopancreas from 5 crabs per treatment were collected and frozen immediately in liquid nitrogen for RNA analysis. In addition, several tissues including hemolymph, eyestalk, heart, intestine, muscle, gill, stomach, hepatopancreas and testis were sampled from six healthy adult male mud crabs (body weight of 210.79 ± 10.58 g) with testis development stage III. Tissues were dissected rapidly and frozen immediately in liquid nitrogen as described previously [30]. All tissues sample were stored at -80 °C until RNA extraction.
2.3. Cloning and Sequencing of S. paramamosain fatp1 cDNA
Total RNA was isolated by RNA isolator reagent in accordance with the manufacturer’s protocol (Vazyme Biotech Co., China). The integrity, concentration and quality of total RNA was determined using a NanoDrop One Spectrophotometer (Thermo Scientific, USA) and 1.0 % agarose gel electrophoresis, respectively. First-strand cDNA was synthesized using HiScript III RT SuperMix for qPCR (+ gDNA wiper) reagent under the manufacturer’s instructions (Vazyme Biotech). The full-length cDNA of fatp1 was amplified based on the transcriptome database of S. paramamosain [31] with specific primers (Supplementary Table S2), using HiScript-TS 5’/3’ RACE Kit following the manufacturer’s protocol (Vazyme Biotech). The PCR products were purified and cloned into pCE2 TA/Blunt-Zero Vector (Vazyme Biotech), which was then transformed into competent cells E. coli DH5α as described previously [30]. After being identified by PCR, the positive clones were selected and subsequently sequenced commercially (Sangon Biotech (Shanghai) Co., Shanghai, China).
2.4. Bioinformatic Analysis
Based on the sequencing results, DNAMAN V9.0 was used to analyze the full-length cDNA sequence to obtain the complete open reading frame (ORF) and amino acid sequences. Biochemical information of encoding protein was analyzed by ProtParam online tools (http://web.expasy.org/protparam/). The protein sequences of Fatp1 orthologs from vertebrates and invertebrates were compared with that of S. paramamosain using multiple sequence alignment by ClustalW (https://www.genome.jp/tools-bin/clustalw). The phylogenic analysis was constructed with different Fatp1 and their orthologs from vertebrates and invertebrates using MEGA software V6.0.
2.5. RNA Interference
A prokaryotic expression vector was constructed to synthesize double-stranded RNA (dsRNA) for knocking down the expression of fatp1 gene in Scylla paramamosain as described in detail previously [30]. Briefly, a 438 bp fragment of Fatp1 full-length cDNA was chosen and amplified by PCR using specific primers (Supplementary Table S2) to produce the corresponding dsRNA according to its sequence characteristics, and amplified by PCR using specific primers. After purification, the PCR product was digested with EcoR I and Stu I and then ligated with the pLITMUS-28i vector (NEB, MA, USA), followed by transformation into E. coli HT115 (DE3) Competent Cells. After verification by restriction enzyme digestion and sequencing, the single positive clone was inoculated in LB medium containing 10 μg/mL ampicillin and tetracycline (Sigma-Aldrich, USA) at 37 °C with shaking at 250 rpm/min for 4 h (OD600 ≈ 0.4 - 0.6). Then, IPTG (isopropyl-β-d-thiogalactoside) was added to a final concentration of 0.1 mM/L and the mixture was cultured for another 4 h to induce the expression of single-stranded RNA. Subsequently, total RNA was extracted and annealed at 70 ℃ for 10 min to synthesize dsRNA. Redundant single-strand RNA and DNA were digested by RNase A and DNase Ⅰ, respectively (Sangon Biotech (Shanghai) Co.). After confirming the concentration, quality and reliability, all dsRNA were stored at -80 ℃ until further use.
After the effective injection concentration and interval time for the dsRNA to induce the knockdown effect on the fatp1 gene in mud crab were verified in a preliminarily experiment, continuous injection of the dsRNA in vivo was used to conduct the RNA interference experiment in S. paramamosain. All crabs were acclimated in breeding boxes (32 cm × 20 cm × 14 cm) for two weeks. In total, 48 healthy crabs with an average weight of 5 g were randomly selected and equally divided into two groups, with 24 individuals in each group. The experimental group was treated with dsRNA of fatp1 diluted in phosphate buffered saline (PBS), while the negative control group was treated with an equal volume of PBS. Crabs were injected with dsRNA at 3 mg/kg body weight into the arthropodial membrane at the base of the walking legs every three days for four times. Crabs were fed the diet with 1.04 % n-3 LC-PUFA during the RNA interference in vivo injection experiment (see Supplementary Table S1). During the trial, the water temperature was maintained at 25 - 30 °C, pH at 7.5 to 8.1, ammonia nitrogen at < 0.05 mg/L, dissolved oxygen > 5.0 mg/L, and salinity of 15 g/L.
At the end of the RNA interference experiment, all surviving crabs were fasted for 24 h before dissection. Immediately prior dissection, they were chilled on ice for general anesthesia, and hemolymph, hepatopancreas and muscle were collected. Due to the low quantity of the tissues extracted, the hemolymph, hepatopancreas and muscle of 4 - 6 crabs were pooled into one replicate, and each treatment had 4 – 6 replicates (n = 4/6). The pooled samples were then stored at -80 °C for subsequent biochemical, fatty acid and gene expression analyses. Additionally, a part of the hepatopancreas from three crabs each group were collected and stored in 4 % paraformaldehyde (Biosharp, China) at room temperature for histological analysis.
2.6. Tissue Total Lipid, Triglyceride, and Cholesterol Contents, and Fatty Acid Composition Analysis
The contents of total triglyceride (TG) and total cholesterol (T-CHO) in the hepatopancreas of S. paramamosain were determined using commercial assay kits on the manufacturer’s protocols (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Total lipids of hepatopancreas and muscle were extracted with a mixture of chloroform/methanol (2:1, by vol.) and saponified using 0.5 M potassium hydroxide in methanol, and then subjected to methylation using boron trifluoride methanol (14 %, Acros Organics, NJ, USA). Subsequently, the fatty acid methyl esters were separated by gas chromatography (GC-2010 plus, Shimadzu, Kyoto, Japan) with the parameters and procedures as described in detail previously [32]. Individual fatty acids were identified using known commercial standards (Sigma-Aldrich, USA) and quantified in a GC solution workstation (Shimadzu, Kyoto, Japan) with the percentage of each fatty acid calculated by the peak area normalization method.
2.7. Histological Analysis
Hepatopancreas tissue samples stored in 4 % paraformaldehyde were frozen, fixed and sectioned (5 μm), and then stained with Oil Red O (ORO) as described in detail previously [33]. The sections were examined and photographed under a light microscope (Axio Imager 2, Zeiss, Germany) equipped with a camera (Axiocam 506, Zeiss) and software (ZEN 2, Zeiss) for image acquisition. Lipid droplets stained by ORO were quantified using Image-pro plus version 6.0 (Media Cybernetics, Maryland, MD, USA).
2.8. Quantitative PCR (qPCR) Analysis
The expression levels of genes including fatty acid transport protein 1/4 (fatp1/4), long-chain acyl-CoA synthetase 1/3 /4 (acsl1/3/4), acetyl CoA carboxylase (acc), fatty acid synthetase (fas), sterol regulatory element binding protein 1 (srebp1), hormone-sensitive lipase (hsl) and carnitine palmitoyltransferase 1 (cpt1) in hepatopancreas and muscle were detected by SYBR Green-based real-time quantitative PCR (qPCR). Total RNA was extracted as described above and 1μg of high-quality RNA was reverse-transcribed using HiScript III RT SuperMix for qPCR (+ gDNA wiper) kit (Vazyme Biotech). All qPCR assays were performed in a LightCycler®480 thermocycler (Roche, Switzerland) as described previously [30]. Each sample was run in triplicate, and reactions without templates were used as negative controls. Primers used in qPCR are shown in Supplementary Table S2. The 18 s rRNA gene was used as internal control (housekeeping) gene for normalizing experimental data, and relative transcript levels of each gene were calculated via the 2−ΔΔCt method.
2.9. Statistical Analysis
Results were expressed as means ± SEM (standard error of the mean) with n value as indicated. Normal distribution and homogeneity of variance of the data were first checked by the Kolmogorov-Smirnov and Barlett’ s tests before independent sample t-test for two groups, or one-way ANOVA and Tukey’s test for multiple group comparisons were performed. All statistical analyses were performed with SPSS software (version 26.0, IBM, USA). A P value < 0.05 was considered statistically significant. All graphics were prepared with Origin 2017 (OriginLab Corporation, Northampton, MA, USA).
3. Results
3.1. Sequence and Phylogenetic Analysis
The full-length sequence of the cDNA of the mud crab S. paramamosain fatp1 gene obtained by 3’-RACE and 5’-RACE contained a 1929 bp ORF, which encoded 643 amino acid residues. The full nucleotide sequence is available at Genbank database with accession number OR232206. The predicted molecular weight and theoretical isoelectric point of the putative Fatp1 polypeptide were 71,777 kDa and 8.06, respectively. The deduced putative Fatp1 polypeptide possessed all the typical features of the Fatp family including a conserved amino acid sequence IYTSGTTGXPK (Figure 1A), acyl-CoA synthase-related domain (Figure S2) [34], and three predicted transmembrane regions located at amino acid residues 15-43, 134-160 and 392-419 of the N-terminus. The results of homologous sequence alignment showed that the S. paramamosain Fatp1 shared low sequence identity with vertebrates (45 %) and relatively high identity with invertebrates (64 %), especially with Penaeus vannamei (65 %). The phylogenetic tree constructed by MEGA 6.0 indicated that the fatp1 gene of S. paramamosain clustered together with other Fatp1 orthologs from decapod crustaceans, especially Portunus trituberculatus, and separated from Fatp1 orthologs of vertebrates (Figure 1B).
3.2. Tissue Distribution Pattern of fatp1 in Mud Crabs
The fatp1 gene was expressed diversely in all mud crab tissues, with significantly highest expression level in hepatopancreas (P < 0.05), followed by stomach, gill, heart, eyestalk, gonad and intestine (Figure 2). Comparatively lower levels of expression of the fatp1 gene were found in ganglion, muscle and hemocyte, with hemocyte showing the lowest expression level of all tissues examined.
3.3. Expression of fatp1 in Hepatopancreas of Crabs Fed Different Dietary n-3 LC-PUFA Levels
The expression level of the fatp1 gene in hepatopancreas increased significantly as dietary levels of n-3 LC-PUFA increased, and crabs fed the diets with 0.73, 1.04, 1.42 and 2.57 % n-3 LC-PUFA levels showed significantly higher expression levels of the fatp1 gene than those of crabs fed the diet with the lowest (0.50 %) level of n-3 LC-PUFA (P < 0.05) (Figure 3A).
3.4. Total Lipid and Fatty Acid Composition in Hepatopancreas and Muscle of Crabs Fed Different Dietary n-3 LC-PUFA Levels
Total lipid contents in hepatopancreas of crabs significantly decreased with increasing dietary level of n-3 LC-PUFA (Figure 3B), while the opposite pattern was observed for total lipid contents of muscle (P < 0.05) (Figure 3C). Among lipids, polar (e.g. membrane) lipids in hepatopancreas and muscle increased significantly as the dietary level of n-3 LC-PUFA increased, while the nonpolar (storage) lipids decreased significantly (P < 0.05) (Figure 3D, E). Moreover, the LC-PUFA contents, especially EPA and DHA (see Supplementary Table S3 and S4), in both polar and nonpolar lipids in hepatopancreas (Figure 3F) and muscle (Figure 3G) increased significantly with increased dietary level of n-3 LC-PUFA (P < 0.05).
3.5. Total Lipid, Triglyceride and Cholesterol Contents, and Fatty Acid Compositions in Hepatopancreas and Muscle after Knockdown of the fatp1 Gene in Mud Crab
RNA interference was used to study the functional role(s) of Fatp1 in lipid accumulation in mud crab. Analysis by qPCR showed that the expression level of the fatp1 gene in hepatopancreas decreased significantly compared with the control group after 2 weeks of continuous dsRNA injection (Figure 4A). Compared with the control group, the contents of total lipid and the concentration of TG were reduced significantly in hepatopancreas of crabs after knockdown of the fatp1 gene (P < 0.05) (Figure 4B, C), while no difference was found in the content of total cholesterol (Figure 4D). As evident in hepatopancreas stained with ORO, significantly fewer lipid droplets were observed in crabs treated with fatp1 gene dsRNA than in crabs of the control group (P < 0.01) (Figure 4E, F).
There were no significant differences in the fatty acid compositions of hepatopancreas between crabs treated with fatp1 gene dsRNA and the negative control, other than higher levels of 20:1n-9 and total monounsaturated fatty acids (MUFA), and a lower level of 12:0 (Table 1). Total saturated fatty acids (SFA) and LC-PUFA contents in hepatopancreas were slightly, but not significantly decreased when compared with the control group. Similarly, significantly increased MUFA contents and a non-significant trend for lower total LC-PUFA contents were also observed in muscle of crabs treated with fatp1 dsRNA (Table 1). However, compared with the control group, no difference was found in total SFA contents in muscle of crabs after treatment with fatp1 dsRNA, although the contents of 12:0 and 14:0 decreased significantly (P < 0.05).
3.6. Expression of Lipid Metabolism-Related Genes in Hepatopancreas of Crabs after Knockdown of the fatp1 Gene
Compared to the negative control, the expression levels of fatp4, acsl1, srebp1, acc and fas were decreased significantly (P < 0.05), while the expression levels of acsl3, acsl4 and cpt1 genes were increased significantly in crabs after knockdown of the fatp1 gene using RNA interference (P < 0.05) (Figure 5). Although not reaching statistical significance, relatively higher transcript level of hsl was also found in hepatopancreas of crabs with fatp1 knocked down compared to the negative control group.
4. Discussion
The FATP proteins are a family of membrane-bound proteins that contribute to the selective cellular absorption and activation of LCFA in mammals [1,2]. Among the members of the FATP family, FATP1 is involved in the process of LCFA transportation, oxidation, triglyceride synthesis, lipid accumulation, and adipocyte differentiation in tissues with rapid fatty acid metabolism, such as the heart, skeletal muscle, liver and adipose tissue[2,9,11,13]. The role of FATP1 in lipid metabolism has been studied extensively in vertebrates, especially mammals, whereas considerably less is known about Fatp1 in invertebrates such as crustaceans. In the present study, the cDNA of a fatp1 gene was cloned from a crustacean species, mud crab S. paramamosain, and shown to encode a putative Fatp1 of 643 amino acids, similar to human FATP1 protein that contains 646 amino acids [4,9]. The deduced putative Fatp1 polypeptide of mud crab contained the common representative characteristics of the FATP family, including a conserved amino acid sequence IYTSGTTGXPK, an acyl-CoA synthase related domain, and three predicted transmembrane segments [2,4,34]. Bioinformatic analysis showed that the Fatp1 of S. paramamosain clustered together and shared relatively high sequence identity with Fatp orthologs of other crustaceans. Overall, the above results were entirely consistent with the identity of the Fatp1 of S. paramamosain as a member of the Fatp family.
Previous studies in invertebrates demonstrated that a fatp homologue cloned from Eilema japonica was expressed predominantly in the pheromone gland, and its functional expression in E. coli enhanced the uptake of LCFA (C18 and C20), but not pheromone precursor hydrocarbons [35]. In the present study, the S. paramamosain fatp1 gene was highly expressed in hepatopancreas, stomach, gill and heart, which was different to the tissue distribution pattern in vertebrates, in which the Fatp1 gene was expressed abundantly in tissues with high activity of fatty acid consumption and oxidation such as heart and skeletal muscle, as well as liver and adipose tissue [9,11]. However, the significantly highest expression of fatp1 in hepatopancreas of crustaceans, the main organ involved in lipid metabolism and energy storage in crustaceans [36,37], showed its potential function in lipid absorption and storage, and further confirmed the Fatp1 of mud crab as an Fatp1 ortholog.
In mammals, however, it is debatable as to whether FATP1 promotes or inhibits the esterification and oxidation of fatty acids in different cell types and/or tissues [13]. In the present study, the reduction of both TG and total lipid contents of hepatopancreas in crabs after knockdown of fatp1 in vivo, while there was no change in the content of CHO, suggesting that the expression of fatp1 in hepatopancreas was important for lipid uptake and accretion in that tissue in mud crabs. This was consistent with previous studies that reported overexpression of Fatp1 promoted preadipocyte differentiation and fat deposition in bovine adipocytes, while Fatp1 interference had the opposite effect on adipocyte differentiation [14,38].
In mammals, FATP4 is the transport protein most closely related to FATP1, and is highly expressed in skeletal muscle, heart, and adipose tissue [39]. Previous study found that the loss of Fatp1 in 3T3-L1 cell lines did not affect the expression of FATP1 in FATP4 knockdown cell but lead to reduced triacylglycerol deposition [14]. It seems like the loss of key Fatp1 functions wouldn’t affect the expression of Fatp4. However, in the present study, knockdown of fatp1 affect the expression of fatp4. This implies that the expression of genes involved in lipid metabolism like fatp4 would more likely be an impact of the loss of key Fatp1 functions. Moreover, the expression levels of acsl genes, which encode proteins that are members of the Acsl (acyl-CoA synthetase) family [40,41] were also impacted by fatp1 knockdown with acsl3 expression being decreased, similar to fatp4, in hepatopancreas of crabs after knockdown of fatp1. Activation of fatty acids by acyl-CoA synthetase enzymes is the first step for exogenous or endogenous fatty acids to enter metabolic pathways, including de novo lipid synthesis, fatty acid oxidation and catabolism, and biological membrane remodeling, among others [42]. This suggested that the Fatp1 of mud crab may be involved in the regulation of hepatopancreatic lipid accumulation by affecting the transport and partitioning of fatty acids. In the present study, the expression levels of genes related to lipogenesis, including acc, fas and srebp1 all decreased in hepatopancreas of crabs subjected to knockdown of fatp1. Acc are the rate-limiting enzymes in de novo fatty acid synthesis (carried out by Fas), catalyzing the ATP-dependent carboxylation of acetyl-CoA to form malonyl-CoA [43], while Srebp1 is a key transcription factor regulating several genes of lipid biosynthesis. Thus, knockdown of fatp1 gene expression was associated with a general down-regulation of lipogenesis in hepatopancreas suggesting that Fatp1 and its function were important for stimulating lipogenesis. In contrast, the expression levels of genes related to lipid catabolism and fatty acid oxidation, hsl and cpt1, were increased in hepatopancreas of crabs subjected to fatp1 knockdown. Hsl is the hormonally regulated neutral lipase responsible for the hydrolysis of triglycerides [44], while Cpt1 is the rate-limiting enzyme in the p-oxidation of fatty acids. Therefore, fatp1 knockdown was associated with increased expression of these genes related to lipid catabolism suggesting that Fatp1 and its function were important for inhibiting lipid catabolism. However, increased hsl in hepatopancreas may also be one possible reason for the increased content of total lipids by promoting lipid deposition in this tissue in crabs subjected to fatp1 knockdown. It appears unlikely that these effects on the expression of other genes involved in lipid metabolism could be a direct effect of the gene knockdown, and would more likely be an impact of the loss of key Fatp1 functions. However, the precise mechanism of the effects of fatp1 knockdown on the expression of other genes of lipid metabolism is currently unclear and requires further study. Interestingly, in this respect, the expression levels of acsl3/4, which have substrate preferences for PUFA substrates like EPA and ARA [40,41], were also increased in hepatopancreas of crabs subjected to fatp1 knockdown, opposite to the effect on acsl1, and also not consistent with the fatty acid composition of hepatopancreas that tended to show reduced n-3 LC-PUFA in crabs subjected to fatp1 knockdown. Therefore, the interaction between the roles of Fatp1 and Acsl and the differential impact of fatp1 knockdown on the expression of different members of the acsl family also requires further investigation.
The FATP family of membrane-associated proteins have been suggested to be an important mediator of LCFA-trafficking into cells [45]. The use of stable isotopically labeled fatty acids demonstrated FATP2a was involved in the uptake and activation of exogenous fatty acids, with a preference toward n-3 PUFA (18:3 and 22:6) as well as very long-chain saturated fatty acids (24:0) [46]. Using siRNA interference to knock down expression of Fatp4 in Neuro2a cells decreased the activation of both long-chain (16:0) and very long-chain saturated fatty acids (24:0), while overexpression of FATP4 in COS-1 cells resulted in enhanced uptake of 16:0 [47]. It was also shown that FATP1 played a vital role in insulin-stimulated LCFA absorption and activation because insulin-stimulated fatty acid uptake was completely abolished in FATP1-null adipocytes and greatly reduced in skeletal muscle of FATP1-knockout animals while basal LCFA uptake by both tissues was unaffected [16]. In terms of fatty acid composition, there were clear trends for decreased total LC-PUFA contents in both muscle and hepatopancreas of crabs in the present study after treatment with fatp1 dsRNA, suggesting mud crab Fatp1 may have a possible substrate preference for LC-PUFA. It was also found that increasing dietary levels of n-3 LC-PUFA levels significantly up-regulated the expression level of fatp1 in hepatopancreas of mud crab, along with the increased contents of LC-PUFA in polar and nonpolar lipids in hepatopancreas and muscle. Similarly, in mammals, overexpression of FATP1 in rats and mice increased the rate of LCFA transportation, and enhanced both systemic deposition of fatty acids and intramuscular lipid accumulation [19,48,49]. Overall, these results suggest a key role for Fatp1in LC-PUFA uptake and esterification in hepatopancreas and muscle tissues in mud crab.
5. Conclusions
In conclusion, a novel fatp1 gene was successfully cloned and characterized from a decapod crustacean mud crab S. paramamosain. The mud crab Fatp1 contained all the typical features of the Fatp family, and shared high homology with other Fatp orthologs identified from crustaceans. The fatp1 mRNA was widely expressed in tissues, particularly in the more metabolically active tissues such as hepatopancreas, stomach and gill. Knockdown of fatp1 in vivo decreased lipid accumulation in the hepatopancreas associated with reduced expression levels of genes related to fatty acid uptake, transportation, oxidation and synthesis. Crabs fed increased dietary levels of n-3 LC-PUFA showed an up-regulated expression level of the fatp1 gene in hepatopancreas, as well as increasing contents of LC-PUFA, especially EPA and DHA, in both polar and nonpolar lipids in hepatopancreas and muscle. Overall, the present study indicated that the Fatp1 identified from S. paramamosain might play functional roles in absorption, utilization and allocation of fatty acid, especially PUFA in crustaceans, and has provided new insights into mechanisms of lipid accumulation in invertebrates.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, C.Y.C.; validation and investigation, W.Z., C.S.C. and X.H.; resources, X.W., S.W.; data curation, C.Y.C.; writing-original draft preparation, W.Z. and C.Y.C.; writing-review and editing, W.Z., S.T., D.R.T. K.W. and C.Y.C. visualization, W.Z., C.S.C. and X.H.; project administration, C.Y.C.; funding acquisition, C.Y.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31972806), the Special Fund Project of Science and Technology Application in Guangdong Province (STKJ2021106 & STKJ202209037), Guangdong Provincial Key Areas R&D Program (2021B0202050001), Special Science Found of Nansha-South China Agricultural University Fishery Research Institute, Guangzhou (NSYYKY202301), and the Department of Education of Guangdong Province (018KCXTD012).
Institutional Review Board Statement
All procedures performed on crabs were in accordance with the National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978) and approved by the Institutional Animal Care and Use Committee of Shantou University (Guangdong, China).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The availability of the data is restricted to investigators based at academic institutions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gimeno, R.E. Fatty acid transport proteins. Curr. Opin. Lipidol. 2007, 18, 271–276. [Google Scholar] [CrossRef]
- Anderson, C.M.; Stahl, A. SLC27 fatty acid transport proteins. Mol. Aspects Med. 2013, 34, 516–528. [Google Scholar] [CrossRef]
- Doege, H.; Stahl, A. Protein-mediated fatty acid uptake: Novel insights from in vivo models. Physiology 2006, 21, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.; Listenberger, L.L.; Ory, D.S.; Schaffer, J.E. Membrane topology of the murine fatty acid transport protein 1. J. Biol. Chem. 2001, 276, 37042–37050. [Google Scholar] [CrossRef]
- Hirsch, D.; Stahl, A.; Lodish, H.F. A family of fatty acid transporters conserved from mycobacterium to man. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 8625–8629. [Google Scholar] [CrossRef] [PubMed]
- Faergeman, N.J.; DiRusso, C.C.; Elberger, A.; Knudsen, J.; Black, P.N. Disruption of the Saccharomyces cerevisiae homologue to the murine fatty acid transport protein impairs uptake and growth on long-chain fatty acids. J. Biol. Chem. 1997, 272, 8531–8538. [Google Scholar] [CrossRef]
- DiRusso, C.C.; Li, H.; Darwis, D.; Watkins, P.A.; Berger, J.; Black, P.N. Comparative biochemical studies of the murine fatty acid transport proteins (FATP) expressed in yeast. J. Biol. Chem. 2005, 280, 16829–16837. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A. A current review of fatty acid transport proteins (SLC27). Pflug. Arch. Eur. J. Phy. 2004, 447, 722–727. [Google Scholar] [CrossRef]
- Schaffer, J.E.; Lodish, H.F. Expression cloning and characterization of a novel adipocyte long chain fatty acid transport protein. Cell 1994, 79, 427–436. [Google Scholar] [CrossRef]
- Stahl, A.; Evans, J.G.; Pattel, S.; Hirsch, D.; Lodish, H.F. Insulin causes fatty acid transport protein translocation and enhanced fatty acid uptake in adipocytes. Dev. Cell 2002, 2, 477–488. [Google Scholar] [CrossRef]
- Martin, G.; Nemoto, M.; Gelman, L.; Geffroy, S.; Najib, J.; Fruchart, J.C.; Roevens, P.; de Martinville, B.; Deeb, S.; Auwerx, J. The human fatty acid transport protein-1 (SLC27A1; FATP-1) cDNA and gene: organization, chromosomal localization, and expression. Genomics 2000, 66, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Pohl, J.; Ring, A.; Hermann, T.; Stremmel, W. Role of FATP in parenchymal cell fatty acid uptake. BBA-Mol. Cell Bio. L. 2004, 1686, 1–6. [Google Scholar] [CrossRef]
- Huang, J.P.; Zhu, R.R.; Shi, D.S. The role of FATP1 in lipid accumulation: a review. Mol. Cell Biochem. 2021, 476, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Lobo, S.; Wiczer, B.M.; Smith, A.J.; Hall, A.M.; Bernlohr, D.A. Fatty acid metabolism in adipocytes: functional analysis of fatty acid transport proteins 1 and 4. J. Lipid Res. 2007, 48, 609–620. [Google Scholar] [CrossRef]
- Wu, Q.W.; Kazantzis, M.; Doege, H.; Ortegon, A.M.; Tsang, B.; Falcon, A.; Stahl, A. Fatty acid transport protein 1 is required for nonshivering thermogenesis in brown adipose tissue. Diabetes 2006, 55, 3229–3237. [Google Scholar] [CrossRef]
- Wu, Q.W.; Ortegon, A.M.; Tsang, B.; Doege, H.; Feingold, K.R.; Stahl, A. FATP1 is an insulin-sensitive fatty acid transporter involved in diet-induced obesity. Mol. Cell. Biol. 2006, 26, 3455–3467. [Google Scholar] [CrossRef]
- Chen, X.L.; Luo, Y.L.; Wang, R.S.; Zhou, B.; Huang, Z.Q.; Jia, G.; Zhao, H.; Liu, G.M. Effects of fatty acid transport protein 1 on proliferation and differentiation of porcine intramuscular preadipocytes. Anim. Sci. J. 2017, 88, 731–738. [Google Scholar] [CrossRef]
- Sebastian, D.; Guitart, M.; Garcia-Martinez, C.; Mauvezin, C.; Orellana-Gavalda, J.M.; Serra, D.; Gomez-Foix, A.M.; Hegardt, F.G.; Asins, G. Novel role of FATP1 in mitochondrial fatty acid oxidation in skeletal muscle cells. J. Lipid Res. 2009, 50, 1789–1799. [Google Scholar] [CrossRef]
- Guitart, M.; Osorio-Conles, O.; Pentinat, T.; Cebria, J.; Garcia-Villoria, J.; Sala, D.; Sebastian, D.; Zorzano, A.; Ribes, A.; Jimenez-Chillaron, J.C.; Garcia-Martinez, C.; Gomez-Foix, A.M. Fatty acid transport protein 1 (FATP1) localizes in mitochondria in mouse skeletal muscle and regulates lipid and ketone body disposal. Plos One 2014, 9, e98109. [Google Scholar]
- Qiu, F.F.; Xie, L.; Ma, J.E.; Luo, W.; Zhang, L.; Chao, Z.; Chen, S.H.; Nie, Q.H.; Lin, Z.M.; Zhang, X.Q. Lower expression of SLC27A1 enhances intramuscular fat deposition in chicken via down-regulated fatty acid oxidation mediated by CPT1A. Front. Physiol. 2017, 8, 449. [Google Scholar] [CrossRef]
- Van den Brink, D.M.; Cubizolle, A.; Chatelain, G.; Davoust, N.; Girard, V.; Johansen, S.; Napoletano, F.; Dourlen, P.; Guillou, L.; Angebault-Prouteau, C.; Bernoud-Hubac, N.; Guichardant, M.; Brabet, P.; Mollereau, B. Physiological and pathological roles of FATP-mediated lipid droplets in Drosophila and mice retina. Plos Genetics 2018, 14, e1007627. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, A.; Hashimoto, K.; Imai, K.; Matsumoto, S. Functional characterization of the Bombyx mori fatty acid transport protein (BmFATP) within the silkmoth pheromone gland. J. Biol. Chem. 2009, 284, 5128–5136. [Google Scholar] [CrossRef] [PubMed]
- Kage-Nakadai, E.; Kobuna, H.; Kimura, M.; Gengyo-Ando, K.; Inoue, T.; Arai, H.; Mitani, S. Two very long chain fatty acid acyl-CoA synthetase genes, acs-20 and acs-22, have roles in the cuticle surface barrier in Caenorhabditis elegans. Plos One 2010, 5, e8857. [Google Scholar] [CrossRef]
- Wang, X.; Jin, M.; Cheng, X.; Hu, X.; Zhao, M.; Yuan, Y.; Sun, P.; Jiao, L.; Tocher, D.R.; Zhou, Q. Hepatopancreas transcriptomic and lipidomic analyses reveal the molecular responses of mud crab (Scylla paramamosain) to dietary ratio of docosahexaenoic acid to eicosapentaenoic acid. Aquaculture 2022, 551, 737903. [Google Scholar] [CrossRef]
- Dai, Y.J.; Jiang, G.Z.; Liu, W.B.; Abasubong, K.P.; Zhang, D.D.; Li, X.F.; Chi, C.; Liu, W.B. Evaluation of dietary linoleic acid on growth as well as hepatopancreatic index, lipid accumulation oxidative stress and inflammation in Chinese mitten crabs (Eriocheir sinensis). Aquaculture Reports 2022, 22, 100983. [Google Scholar] [CrossRef]
- Liu, M.; Wang, L.; Cheng, Y.; Gong, J.; Zeng, C.; Wu, X. Effect of estradiol on hepatopancreatic lipid metabolism in the swimming crab, Portunus trituberculatus. Gen. Comp. Endocrinol. 2019, 280, 115–122. [Google Scholar] [CrossRef]
- Wu, Q.; Shi, X.; Fang, S.; Xie, Z.; Guan, M.; Li, S.; Zheng, H.; Zhang, Y.; Ikhwanuddin, M.; Ma, H. Different biochemical composition and nutritional value attribute to salinity and rearing period in male and female mud crab Scylla paramamosain. Aquaculture 2019, 513, 734417. [Google Scholar] [CrossRef]
- Chen, C.Y.; Nie, X.F.; Cheng, Y.; Shen, J.J.; He, X.D.; Wang, S.Q.; You, C.H.; Li, Y.Y. Comparative analysis of growth performance, feed utilization, and antioxidant capacity of juvenile mud crab, Scylla Paramamosian, reared at two different conditions. Crustaceana 2021, 94, 1021–1033. [Google Scholar] [CrossRef]
- Wu, Q.Y.; Waiho, K.; Huang, Z.; Li, S.K.; Zheng, H.P.; Zhang, Y.L.; Ikhwanuddin, M.; Lin, F.; Ma, H.Y. Growth performance and biochemical composition dynamics of ovary, hepatopancreas and muscle tissues at different ovarian maturation stages of female mud crab, Scylla paramamosain. Aquaculture 2020, 515, 734560. [Google Scholar] [CrossRef]
- Zhong, W.J.; Chen, C.S.; Tan, S.Y.; He, X.D.; Wang, S.Q.; Tocher, D.R.; Lin, F.; Sun, Z.J.; Wen, X.B.; Li, Y.Y.; Waiho, K.; Wu, X.G.; Chen, C.Y. Identification of a novel crustacean vascular endothelial growth factor b-like in the mud crab Scylla paramamosain, and examination of its role in lipid accumulation. Aquaculture 2023, 575, 739793. [Google Scholar] [CrossRef]
- Lin, J.L.; Shi, X.; Fang, S.B.; Zhang, Y.; You, C.H.; Ma, H.Y.; Lin, F. Comparative transcriptome analysis combining SMRT and NGS sequencing provides novel insights into sex differentiation and development in mud crab (Scylla paramamosain). Aquaculture 2019, 513, 734447. [Google Scholar] [CrossRef]
- Li, Y.; Monroig, O.; Zhang, L.; Wang, S.; Zheng, X.; Dick, J.R.; You, C.; Tocher, D.R. Vertebrate fatty acyl desaturase with Δ4 activity. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 16840–16845. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jiang, M.; Lian, G.L.; Liu, Q.; Shi, M.; Li, T.Y.; Song, L.T.; Ye, J.; He, Y.; Yao, L.M.; Zhang, C.X.; Lin, Z.Z.; Zhang, C.S.; Zhao, T.J.; Jia, W.P.; Li, P.; Lin, S.Y.; Lin, S.C. AIDA selectively mediates downregulation of fat synthesis enzymes by ERAD to retard intestinal fat absorption and prevent obesity. Cell Metab. 2018, 27, 843. [Google Scholar] [CrossRef]
- Stuhlsatz-Krouper, S.M.; Bennett, N.E.; Schaffer, J.E. Molecular aspects of fatty acid transport: mutations in the IYTSGTTGXPK motif impair fatty acid transport protein function. Prostaglandins Leukotrienes Essent. Fatty Acids 1999, 60, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.G.; Fujii, T.; Ito, K.; Nakano, R.; Ishikawa, Y. Cloning and functional characterization of a fatty acid transport protein (FATP) from the pheromone gland of a lichen moth, Eilema japonica, which secretes an alkenyl sex pheromone. Insect Biochem. Mol. Biol. 2011, 41, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, X.G.; Liu, Z.J.; Zheng, H.J.; Cheng, Y.X. Insights into hepatopancreatic functions for nutrition metabolism and ovarian development in the crab Portunus trituberculatus: gene discovery in the comparative transcriptome of different hepatopancreas stages. Plos One 2014, 9, e84921. [Google Scholar]
- Vogt, G. Functional cytology of the hepatopancreas of decapod crustaceans. J. Morphol. 2019, 280, 1405–1444. [Google Scholar] [CrossRef]
- Liu, X.; Li, S.; Wang, L.; Zhang, W.; Wang, Y.; Gui, L.; Zan, L.; Zhao, C. The effect of FATP1 on adipocyte differentiation in Qinchuan beef cattle. Animals 2021, 11, 2789. [Google Scholar] [CrossRef]
- Stahl, A.; Gimeno, R.E.; Tartaglia, L.A.; Lodish, H.F. Fatty acid transport proteins: a current view of a growing family. Trends Endocrin. Met. 2001, 12, 266–273. [Google Scholar] [CrossRef]
- Soupene, E.; Kuypers, F.A. Mammalian long-chain Acyl-CoA synthetases. Exp. Biol. Med. 2008, 233, 507–521. [Google Scholar] [CrossRef]
- Ellis, J.M.; Frahm, J.L.; Li, L.O.; Coleman, R.A. Acyl-coenzyme A synthetases in metabolic control. Curr. Opin. Lipidol. 2010, 21, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Marques, M.; Cunha, I.; Reis-Henriques, M.A.; Santos, M.M.; Castro, L.F.C. Diversity and history of the long-chain acyl-CoA synthetase (Acsl) gene family in vertebrates. BMC Evol. Bio. 2013, 13, 271. [Google Scholar] [CrossRef] [PubMed]
- Polyak, S.W.; Abell, A.D.; Wilce, M.C.J.; Zhang, L.; Booker, G.W. Structure, function and selective inhibition of bacterial acetyl-coa carboxylase. Appl. Microbiol. Biot. 2012, 93, 983–992. [Google Scholar] [CrossRef]
- Recazens, E.; Mouisel, E.; Langin, D. Hormone-sensitive lipase: sixty years later. Prog. Lipid Res. 2021, 82, 101084. [Google Scholar] [CrossRef]
- Pohl, J.; Ring, A.; Ehehalt, R.; Herrmann, T.; Stremmel, W. New concepts of cellular fatty acid uptake: role of fatty acid transport proteins and of caveolae. Proc. Nutr. Soc. 2004, 63, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Melton, E.M.; Cerny, R.L.; Watkins, P.A.; Dirusso, C.C.; Black, P.N. Human fatty acid transport protein 2a/very long chain acyl-CoA synthetase 1 (FATP2a/Acsvl1) has a preference in mediating the channeling of exogenous n-3 fatty acids into phosphatidylinositol. J. Biol. Chem. 2011, 286, 30670–30679. [Google Scholar] [CrossRef]
- Jia, Z.; Pei, Z.; Maiguel, D.; Toomer, C.J.; Watkins, P.A. The fatty acid transport protein (FATP) family: Very long chain acyl-CoA synthetases or solute carriers? J. Mol. Neurosci. 2007, 33, 25–31. [Google Scholar] [CrossRef]
- Nickerson, J.G.; Alkhateeb, H.; Benton, C.R.; Lally, J.; Nickerson, J.; Han, X.-X.; Wilson, M.H.; Jain, S.S.; Snook, L.A.; Glatz, J.F.C.; Chabowski, A.; Luiken, J.J.F.P.; Bonen, A. Greater transport efficiencies of the membrane fatty acid transporters FAT/CD36 and FATP4 compared with FABPpm and FATP1 and differential effects on fatty acid esterification and oxidation in rat skeletal muscle. J. Biol. Chem. 2009, 284, 16522–16530. [Google Scholar] [CrossRef]
- Holloway, G.P.; Chou, C.J.; Lally, J.; Stellingwerff, T.; Maher, A.C.; Gavrilova, O.; Haluzik, M.; Alkhateeb, H.; Reitman, M.L.; Bonen, A. Increasing skeletal muscle fatty acid transport protein 1 (FATP1) targets fatty acids to oxidation and does not predispose mice to diet-induced insulin resistance. Diabetologia 2011, 54, 1457–1467. [Google Scholar] [CrossRef]
Figure 1.
The mud crab Fatp1 is a member of the FATP family. (A) Conserved amino acid sequence alignment of IYTSGTTGXPK from mud crab Fatp1 and its orthologs of other species. (B) Phylogenetic tree of amino acid sequences of mud crab Fatp1 with its orthologs of other vertebrates and invertebrates. The Fatp1 orthologs of Homo sapiens (NP_940982.1), Mus musculus (NP_036107.1), Penaeus vannamei (XP_015907328.1) and Parasteatoda tepidariorum (KY271629.1/ARO77488.1) were compared with S. paramamosain Fatp1. Mud crab Fatp1 is marked by the black triangle. The neighbor-joining (NJ) polygenetic tree was tested by bootstrapping using 1000 replications.
Figure 1.
The mud crab Fatp1 is a member of the FATP family. (A) Conserved amino acid sequence alignment of IYTSGTTGXPK from mud crab Fatp1 and its orthologs of other species. (B) Phylogenetic tree of amino acid sequences of mud crab Fatp1 with its orthologs of other vertebrates and invertebrates. The Fatp1 orthologs of Homo sapiens (NP_940982.1), Mus musculus (NP_036107.1), Penaeus vannamei (XP_015907328.1) and Parasteatoda tepidariorum (KY271629.1/ARO77488.1) were compared with S. paramamosain Fatp1. Mud crab Fatp1 is marked by the black triangle. The neighbor-joining (NJ) polygenetic tree was tested by bootstrapping using 1000 replications.
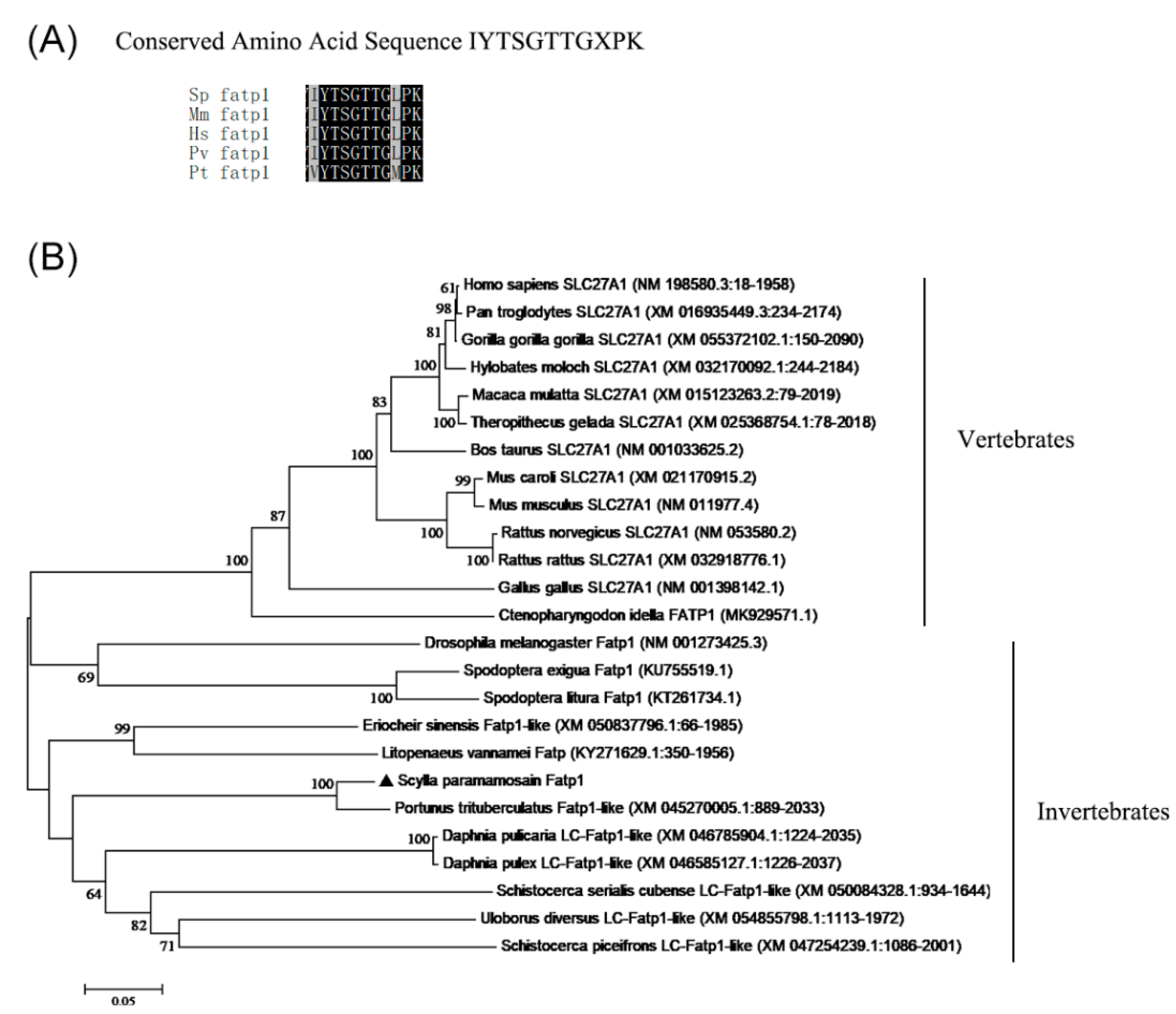
Figure 2.
The mud crab fatp1 was mainly expressed in tissues with more active metabolism. The relative mRNA expression levels of the fatp1 gene in detected tissues of mud crab as determined by qPCR. Values are means ± S.E.M (n = 6) and represent levels relative to expression level in hemocyte. Bars with different superscript letters indicate significant differences.
Figure 2.
The mud crab fatp1 was mainly expressed in tissues with more active metabolism. The relative mRNA expression levels of the fatp1 gene in detected tissues of mud crab as determined by qPCR. Values are means ± S.E.M (n = 6) and represent levels relative to expression level in hemocyte. Bars with different superscript letters indicate significant differences.
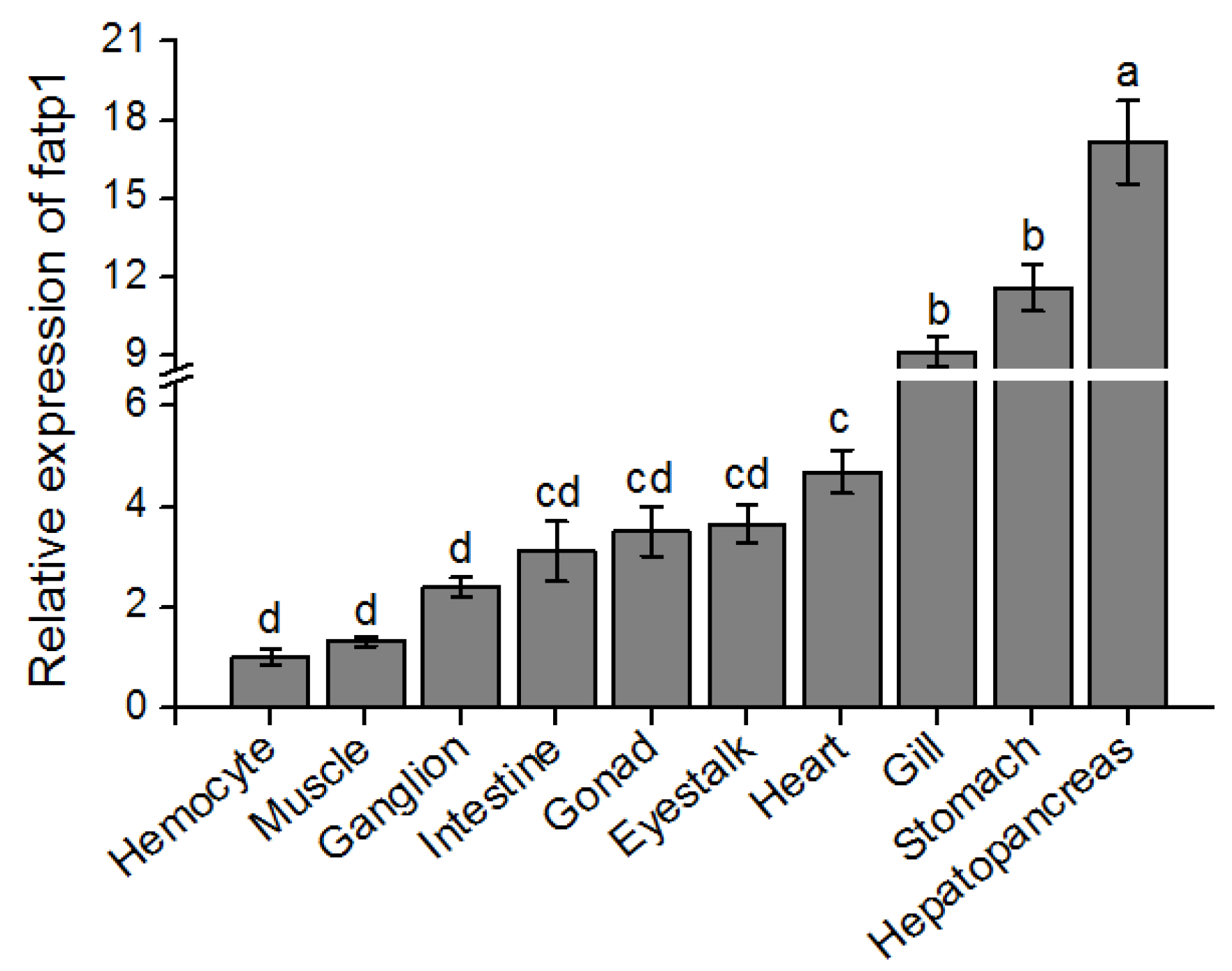
Figure 3.
The mud crab fatp1 was involved in long-chain polyunsaturated fatty acid (LC-PUFA) metabolism and deposition. (A) The relative mRNA expression levels of the fatp1 gene in hepatopancreas of mud crab fed different dietary levels of n-3 LC-PUFA. Total lipid contents (% dry weight) of (B) hepatopancreas and (C) muscle of mud crab fed different dietary levels of n-3 LC-PUFA. Polar and nonpolar lipid contents (% total lipid) of (D) hepatopancreas and (E) muscle of mud crabs fed different dietary levels of n-3 LC-PUFA. The heatmap visualization of the abundance of total saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), PUFA and LC-PUFA in polar and nonpolar lipids of (F) hepatopancreas and (G) muscle of mud crab fed different dietary levels of n-3 LC-PUFA. Before analysis, all data were checked for homogeneity. Bars with different superscript letters in panels A-E indicate significant differences.
Figure 3.
The mud crab fatp1 was involved in long-chain polyunsaturated fatty acid (LC-PUFA) metabolism and deposition. (A) The relative mRNA expression levels of the fatp1 gene in hepatopancreas of mud crab fed different dietary levels of n-3 LC-PUFA. Total lipid contents (% dry weight) of (B) hepatopancreas and (C) muscle of mud crab fed different dietary levels of n-3 LC-PUFA. Polar and nonpolar lipid contents (% total lipid) of (D) hepatopancreas and (E) muscle of mud crabs fed different dietary levels of n-3 LC-PUFA. The heatmap visualization of the abundance of total saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), PUFA and LC-PUFA in polar and nonpolar lipids of (F) hepatopancreas and (G) muscle of mud crab fed different dietary levels of n-3 LC-PUFA. Before analysis, all data were checked for homogeneity. Bars with different superscript letters in panels A-E indicate significant differences.
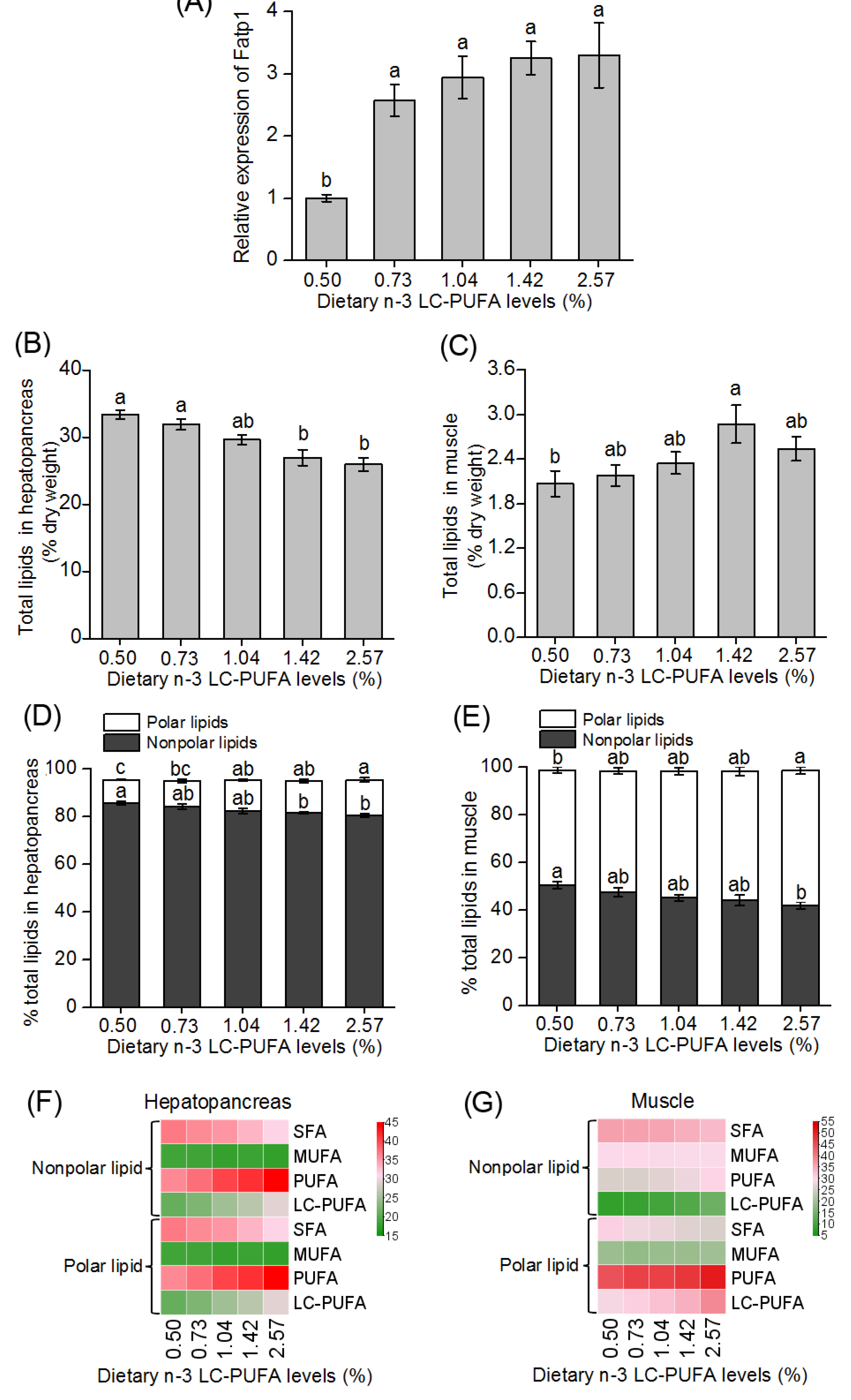
Figure 4.
The mud crab fatp1 was involved in lipid metabolism and deposition in hepatopancreas. (A) Relative mRNA expression levels of fatp1 in hepatopancreas. (B) Total lipid (g/100g dry weight), (C) triglyceride (μmol/g dry weight), and (D) total cholesterol contents (μmol/g dry weight) of hepatopancreas of mud crab after knockdown of fatp1. (E) Mud crab hepatopancreas stained with oil red O after knockdown of fatp1, and (F) Lipid droplet content (area/total cell area) of mud crab hepatopancreas stained with ORO after knockdown of fatp1. Data are presented as means ± S.E.M (n = 4) as fold-change from the negative controls. *, P < 0.05; **, P < 0.01;***, P < 0.001.
Figure 4.
The mud crab fatp1 was involved in lipid metabolism and deposition in hepatopancreas. (A) Relative mRNA expression levels of fatp1 in hepatopancreas. (B) Total lipid (g/100g dry weight), (C) triglyceride (μmol/g dry weight), and (D) total cholesterol contents (μmol/g dry weight) of hepatopancreas of mud crab after knockdown of fatp1. (E) Mud crab hepatopancreas stained with oil red O after knockdown of fatp1, and (F) Lipid droplet content (area/total cell area) of mud crab hepatopancreas stained with ORO after knockdown of fatp1. Data are presented as means ± S.E.M (n = 4) as fold-change from the negative controls. *, P < 0.05; **, P < 0.01;***, P < 0.001.
Figure 5.
Knockdown of fatp1 affected the expression levels of genes related to fatty acid uptake, transportation, oxidation and synthesis. (A) Expression levels of genes related to fatty acid uptake and transportation, and (B) synthesis and oxidation in hepatopancreas of mud crab after fatp1 knockdown were determined by qPCR. Data are presented as means ± S.E.M (n = 4). *, P < 0.05, **, P < 0.01, ***, P < 0.001 vs. the negative control.
Figure 5.
Knockdown of fatp1 affected the expression levels of genes related to fatty acid uptake, transportation, oxidation and synthesis. (A) Expression levels of genes related to fatty acid uptake and transportation, and (B) synthesis and oxidation in hepatopancreas of mud crab after fatp1 knockdown were determined by qPCR. Data are presented as means ± S.E.M (n = 4). *, P < 0.05, **, P < 0.01, ***, P < 0.001 vs. the negative control.
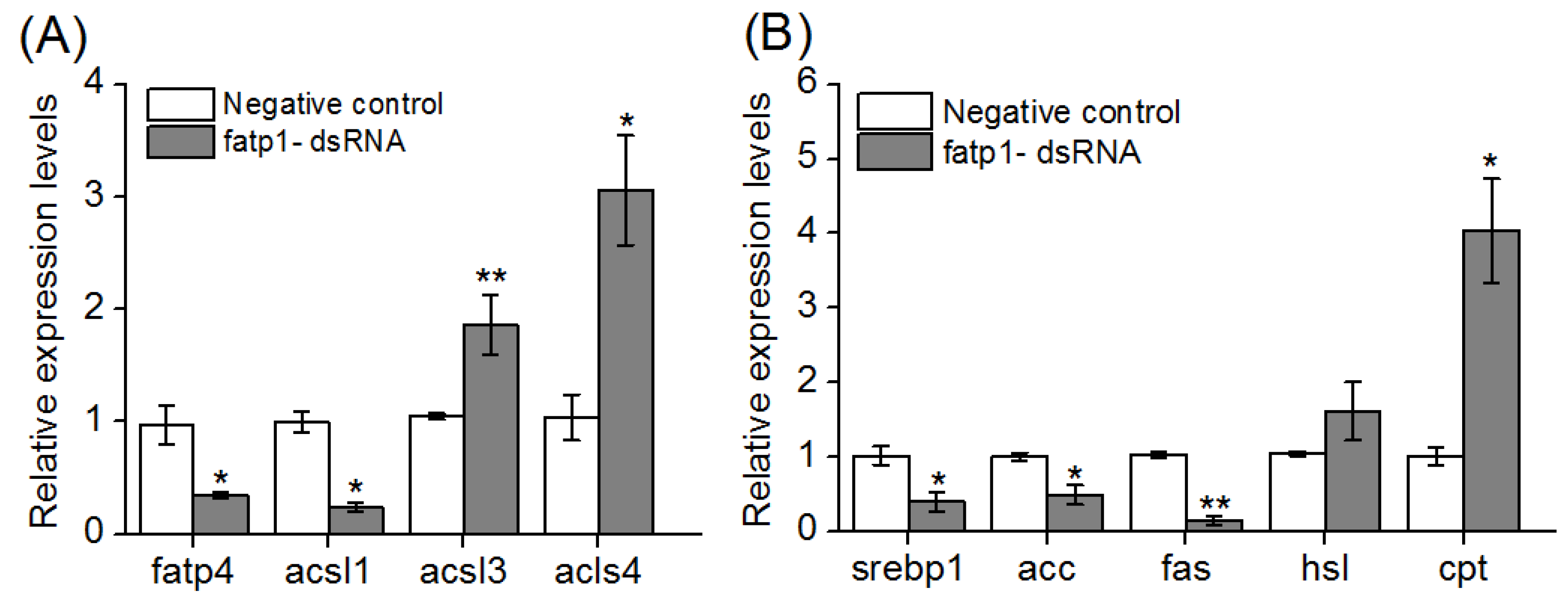

Table 1.
Fatty acid compositions (% total fatty acid) of hepatopancreas and muscle of mud crabs after knockdown of fatp1.
Table 1.
Fatty acid compositions (% total fatty acid) of hepatopancreas and muscle of mud crabs after knockdown of fatp1.
| Fatty acids |
Hepatopancreas | Muscle | |||||
| Negative control |
fatp1 dsRNA | P - value | Negative control |
fatp1 dsRNA | P - value | ||
| 12:00 | 3.45 ± 0.20 | 2.67 ± 0.23 | 0.044 | 1.04 ± 0.19 | 0.40 ± 0.03 | 0.027 | |
| 14:00 | 4.40 ± 0.23 | 3.92 ± 0.42 | 0.352 | 2.24 ± 0.29 | 1.23 ± 0.04 | 0.028 | |
| 16:00 | 15.97 ± 0.63 | 17.62 ± 1.88 | 0.436 | 17.83 ± 0.33 | 18.88 ± 0.81 | 0.296 | |
| 18:00 | 5.63 ± 0.30 | 3.73 ± 0.94 | 0.103 | 10.89 ± 0.25 | 11.45 ± 0.43 | 0.319 | |
| 16:1n-9 | 3.52 ± 0.28 | 4.24 ± 0.80 | 0.427 | 2.20 ± 0.26 | 2.26 ± 0.35 | 0.894 | |
| 16:1n-7 | 0.67 ± 0.07 | 1.52 ± 0.37 | 0.066 | 0.89 ± 0.19 | 4.89 ± 1.17 | 0.028 | |
| 18:1n-9 | 14.34 ± 0.31 | 14.70 ± 0.32 | 0.453 | 11.00 ± 0.54 | 10.39 ± 0.80 | 0.558 | |
| 20:1n-9 | 0.85 ± 0.10 | 1.85 ± 0.34 | 0.03 | 0.55 ± 0.12 | 0.91 ± 0.06 | 0.054 | |
| 18:2n-6 | 13.12 ± 0.58 | 13.28 ± 0.74 | 0.87 | 7.19 ± 0.74 | 6.50 ± 0.62 | 0.509 | |
| 18:3n-3 | 4.32 ± 0.12 | 3.87 ± 0.14 | 0.051 | 1.59 ± 0.18 | 1.44 ± 0.28 | 0.679 | |
| 20:4n-6 | 6.94 ± 0.24 | 6.68 ± 0.48 | 0.649 | 12.48 ± 0.94 | 10.81 ± 0.31 | 0.168 | |
| 22:4n-6 | 0.45 ± 0.12 | 0.44 ± 0.03 | 0.948 | 1.06 ± 0.20 | 1.03 ± 0.23 | 0.919 | |
| 20:5n-3 | 6.58 ± 0.27 | 6.08 ± 0.46 | 0.385 | 14.22 ± 0.19 | 11.97 ± 1.31 | 0.165 | |
| 22:5n-3 | 1.40 ± 0.08 | 1.42 ± 0.06 | 0.866 | 1.35 ± 0.06 | 0.98 ± 0.14 | 0.071 | |
| 22:6n-3 | 6.41 ± 0.34 | 6.27 ± 0.42 | 0.806 | 7.18 ± 0.40 | 5.98 ± 0.85 | 0.268 | |
| ΣSFA1 | 29.44 ± 1.32 | 27.94 ± 1.60 | 0.496 | 31.99 ± 0.68 | 31.96 ± 1.29 | 0.985 | |
| ΣMUFA2 | 19.39 ± 0.49 | 22.31 ± 1.04 | 0.044 | 14.64 ± 0.69 | 18.45 ± 0.95 | 0.032 | |
| ΣPUFA3 | 39.21 ± 1.54 | 38.04 ± 1.59 | 0.615 | 45.07 ± 1.02 | 38.70 ± 3.14 | 0.126 | |
| ΣLC-PUFA4 | 21.78 ± 0.85 | 20.89 ± 1.27 | 0.584 | 36.28 ± 0.70 | 30.76 ± 2.57 | 0.107 | |
Data are presented as means ± S.E.M (n = 4). 1 SFA, saturated fatty acids. 2 MUFA, mono-unsaturated fatty acids. 3 PUFA, polyunsaturated fatty acids. 4 LC-PUFA, long-chain polyun-saturated fatty acids, including 20:4n-6, 22:4n-6, 20:5n-3, 22:5n-3 and 22:6n-3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



