Submitted:
23 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Obesity looms large in the contemporary health landscape, extending its impact beyond physical appearance to influencing overall well-being. This slow intruder subtly weaves its influence through society, affecting individual health and presenting a significant public health dilemma. Beyond the visible weight gain, a web of interconnected health issues emerges, from cardiovascular complications to metabolic disorders, casting a pervasive shadow over the pursuit of a healthy, fulfilling life. In response to the escalating prevalence of obesity and its associated health complexities, there is a growing interest in swift weight management medications. However, current pharmacotherapies targeting crucial weight management pathways often fall short due to substantial side effects outweighing their benefits. Recently, plant extracts have emerged as potential regulators of lipid metabolism and obesity by influencing gut microbiota composition and abundance. This comprehensive review takes deep dive into the potential roles of Salvia spp. phytochemicals as health-promoting agents, with a specific focus on the anti-obesity properties of Salvia spp. by compiling a wealth of in-vitro, in-vivo and clinical studies. Drawing from the latest research, we unravel the potential of Salvia spp. by shedding light on their underlying mechanisms intertwined with gut microbiota dynamics. As scientific inquiry progresses, unraveling the intricate interplay between plant extracts, gut microbiota, and metabolic health holds tremendous promise for addressing the escalating global burden of obesity. This review is a humble attempt to provide a valuable synthesis of current knowledge, so as to pave the way for future advancements in pursuit of effective anti-obesity strategies for clinical interventions.
Keywords:
Clinical studies
; Gut Microbiome
; Obesity
; Salvia spp.
; Therapeutic potential
1. Introduction
1.1. Obesity: A Global Pandemic
Westernization in the lifestyle has inclined people towards the consumption of high-energy foods, but reduced physical activity paves the path for the development of obesity. Currently, obesity has been considered a pandemic, worsening over the last few years. World Health Organization (WHO) defined obesity as the excessive build-up of adipose tissue in the body that poses a significant concern for other health afflictions [1]. Obesity is a major non-communicable disease that is a global threat to health issues in the 21st century. The most recent data put forward by WHO states that over 39% and 13% adults are overweight and obese, respectively [2]. Rates of overweight and obesity continue to grow i.e., from 1975 to 2016, the prevalence of obesity in children and adolescents increased more than four-fold from 4% to 18% globally [3]. The World Obesity Federation’s 2023-World Obesity Atlas projects that the estimated yearly global economic costs associated with overweight and obesity are expected to rise to $4.32 trillion by 2035. This financial burden is comparable to the effects of the COVID-19 pandemic in 2020, accounting for over 3% of the global Gross Domestic Product (GDP) [4].
1.2. Need for Plant Extracts in Curbing Obesity
Hippocrates, the ancient Greek physician from the classical era, believed that “Obesity is not just a metabolic disorder in itself but has been also proclaimed as the precursor for the development of other comorbidities”. This statement still remains relevant in the present world circumstances. A plethora of studies specify that the obese condition contributes to the development of several metabolic disorders, including type-2 diabetes, intestinal inflammation, hypertension, dyslipidemia, and hyperglycemia [5]. Obesity typically results from elevated body fat levels, primarily stemming from unhealthy dietary habits but if integrated into factors like smoking, alcohol consumption, and a lack of physical activity, the condition worsens and exposes the individual to the risk of atherosclerosis, kidney disease, and hypothyroidism [6]. Given the escalating prevalence of obesity and its associated health complications, there is a burgeoning interest in medications for swift weight management. However, existing pharmacotherapies targeting crucial weight management pathways exhibit limited efficacy due to significant side effects that overshadow their benefits [7] (Table 1). Consequently, as the global obesity epidemic persists, researchers are exploring novel treatment strategies, including plant extracts and gut microbiome interventions, as potential solutions to combat obesity. This review seeks to elucidate the current understanding of the use of Salvia spp. in addressing obesity, encompassing their therapeutic potential in both in-vitro and in-vivo settings, along with insights from clinical trials. By delving into the potential mechanisms related to gut microbiota, this review aims to provide an updated comprehension of the intricate relationship among plant extracts, gut microbiota, and obesity. In essence, this review serves as a humble attempt to furnish available information for developing dietary strategies linked to Salvia spp. as a means to treat and prevent obesity and its associated diseases.
2. Salvia: Nature’s Therapeutic Blessing
With about 1000 species, Salvia is the biggest genus in the Lamiaceae family. The genus is widely distributed throughout the world, but is native to temperate and tropical areas [8]. Most of the Sage species are well known for their ornamental nature, distinctive flavour, and bioactive components which is the major reason for its commercial cultivation [9]. Salvia hispanica (chia) is known for its edible seeds while the essential oils from Salvia officinalis have been significantly acknowledged for treating a wide range of ailments associated with the neurological system, cardiovascular health, respiratory system, digestive system, and metabolic and endocrine problems [10]. The Salvia genus encompasses a diverse array of plant species, each distinguished by unique chemical compositions and botanical characteristics. Noteworthy among them are S. officinalis, S. hispanica, Salvia libanotica, Salvia miltiorrhiza, and Salvia plebeian, all of which exhibit promising therapeutic potential and some of the others are discussed in Table 2.
The analgesic potential of Salvia species with antimicrobial properties and mitigating effect against reactive oxidative species has led to their long-standing usage in traditional medicine. Sage species have been used historically for their carminative, spasmolytic, astringent, antibacterial, wound healing, and anti-inflammatory effects for a very long time. Sage has been used in European traditional medicine to treat gastrointestinal diseases, including dyspepsia, flatulence, diarrhoea, abdominal spasms, and intestinal mucosa inflammation [11]. It has also been used to treat inflammation of the mouth and throat, excessive perspiration, coughing, skin inflammations, and galactorrhea. The scientific literature available suggests that Salvia species could be a novel natural therapeutic for the treatment of serious and potentially fatal conditions such as obesity, diabetes, lupus, cardiovascular diseases, cancer, depression and dementia [12].
3. Diving into Diversity: A Comprehensive Exploration of Salvia Species and their Phytochemical Marvels
In the realm of herbal exploration, this section delves into the diverse world of Salvia species, unraveling their phytochemical marvels and their potential implications in the prevention and treatment of obesity and related metabolic disorders. Through an extensive examination, this section reviews the potential role of Salvia species in prevention and treatment of obesity and linked metabolic disorders in various in-vitro (Table 3), in-vivo (Table 4), and human studies (Table 5).
3.1. Salvia Officinalis
S. officinalis goes by many common names, including sage, garden sage, golden sage, culinary sage, dalmatian sage, and broadleaf sage. Native to Mediterranean region, this perennial shrub is commonly grown as a garden plant for its aroma and vibrant red and purple flowers. However, along with the culinary importance, the species has a long history of therapeutic potential associated to it [13]. Traditionally sage had a profound usage in ethnomedicine to treat infertility, inflammation, seizures, hyperglycemia, ulcers, etc. Extensive research has been carried out to investigate novel bioactive components with curative potential. With abundant traditional uses, there are many recent studies that have been focused on antinociceptive potential, antioxidant, antimicrobial properties, antidementia effects along with hypoglycemic and hypolipidemic effects of S. officinalis [14]. Scientific literature also highlights the mitigating effect of sage against other metabolic disorders like obesity, gastroenteritis, diabetes, NAFLD (non-alcoholic fatty liver disease) and cancers related to breast or colon [15].
Many bioactive components have been reported in S. officinalis, such as phenols, terpenes, proteins, carbohydrates, and vitamins C, E, and B. It is also a source of essential minerals such as zinc, iron, copper, and selenium, among other vital components. It is well known that S. officinalis contains a wide range of phytochemical components, the majority of which can be derived via its essential oil, alcoholic and aqueous extracts, butanol fraction, and preparation for infusion [16,17]. The main components of the essential oil include thujone, boreal, camphor, caryophyllene, cineole, elemene, humulene, ledene, and pinene. Because of the high concentration of essential oils and presence of other biologically active components, S. officinalis is one of the most valued herb [18]. Both the aqueous and alcoholic extracts of S. officinalis demonstrate significant concentrations of flavonoids, particularly luteolin-7-glucoside and rosmarinic acid. Furthermore, it has been shown that S. officinalis methanolic extract contains phenolic acids, such as 3-caffeoylquinic acid and caffeic acid [19]. Afonso et al. studied the phytochemical composition of the three different sage species i.e., S. africana, S. officinalis ‘Icterina’ and S. mexicana and it was observed that species had distinct phenolic profile [20]. The glycosidic form of luteolin, apigenin, and scuttelarein were reported in S. officinalis ‘Icterina’, while yunnaneic acid isomers were characterized for S. africana and simple caffeic acid derivatives were obtained from S. mexicana. The study also reported that the rosmarinic acid dominated the phenolic profile and exhibited significant antioxidant potential. In another study, rosmarinic acid was again identified as the main phenolic component among three different plants: lemon balm (Melissa officinalis L.), sage (S. officinalis L.) and spearmint (Mentha spicata L.). The study provided an insight of the active phytochemicals that could also be used as food additive to enhance the shelf life of the food due to the antimicrobial potential of the plant extracts [21]. HPLC analysis identified 12 different polyphenol components in the aqueous extract of S. officinalis (SOE), of which coffeic acid, coumaric acid, ellagic acid, gallic acid, chlorogenic acid, catechin, rutin, and vanillin were the major contributors. The researchers concluded S. officinalis as a promising therapeutic because of its potent antioxidant potential observed against cadmium induced toxicity stress and damage in rats [22]. Jassbi et al. described Salvia as competent natural alternative to the artificial medications available in the market due to the presence of numerous biologically active phytochemicals that can be isolated from shoot and root of the plant and have a great clinical value [23].

Table 4.
In-vivo studies on anti-obesity activity of Salvia species.
| Salvia species | Experimental model, dose, interval period | Experimental outcome | Reference |
|---|---|---|---|
| S. tribola | Male Albino rats N= 20 Methanol extract, 750 mg/kg Gastric intubation |
↓ Plasma triglyceride levels ↑ inhibition of Pancreatic lipase |
[166] |
| S. officinalis | Adult female Wistar rats N= 40 Aqueous extract,10 mg/kg/day Orally 6 weeks |
Group with (OVX + SOE + OR) Body weight & visceral fat ↓ Triglyceride(mg/ml) ↓ TC (mg/ml) ↓ HDL (mg/ml) ↑ LDL (mg/ml) ↓ Glucose levels(mg/ml) ↓ Adiponectin(ng/ml) ↓ Leptin levels(ng/ml) ↓ Pancreatic lipase(ng/ml) ↓ AST(U/ml) ↓ ALT (U/ml) ↓ MDA (nmol/mg protein) ↓ GSH (mmol/mg protein) ↑ TAC (mmol/mg protein) ↑ |
[167] |
| S. hispanica L | C57BL/6J male mice, N= 6 organic solvents: hexane, dichloromethane, ethyl acetate, and ethanol (50 mg/kg/day) Orally 12 weeks |
Loss in Body weight (HFD-BE)- 1.8 ± 0.5 GOT(U/ml) ↓ GPT (U/ml) ↓ ALP (U/ml) ↓ ↓ fasting glucose ↓ fasting insulin ↓ in lipid droplets ↓ in lipid peroxidation degree |
[168] |
| S. officinalis & S. sclarea | Adult white outbred male rats N= 24 5% dry shredded shoots of Salvia officinalis & Salvia sclarea 30 days |
Body weight ↓ ↓ urea concentration ↓ total bilirubin ↓ triglycerides ↑ total protein in blood ↑ alkaline phosphatase activity ↑ Hemoglobin ↑ Erythrocytes ↑ Erythrocyte sedimentation rate (ESR), ↑ WBC, |
[25] |
| S. officinalis L. | Male Wistar rats N=30 Sage Essential oil (2.5 μL/rat) Oral gavage 30 days |
inhibited α-amylase and lipase activities ↓ glycemia ↑ glycogen storage ↓ the ALT ↓ AST ↓ LDH ↓ Creatinine ↓ Uric acid |
[169] |
| S. lavandulifolia | Wistar rats and albino mice Aqueous extract (400 mg/kg) orally |
↓ post-prandial hyperglycemia AUC glucose levels ↓ |
[170] |
Table 5.
Clinical trials (clinicaltrials.gov) linked to anti-obesity activity of Salvia species.
| NCT Number | Study Title | Interventions/ Treatment | Sponsor and Collaborators | Phase | Enrollment | Study type |
|---|---|---|---|---|---|---|
| NCT03233906 | The Effects of Chia on Overweight/Obese Women |
Dietary Supplement: Chia Seeds- Salvia Hispanica |
California State Polytechnic University, Pomona | NA | Female: 60 | Interventional |
| NCT01403571 |
Effectiveness and Safety of Salba on Weight Loss in Overweight Individuals With Type 2 Diabetes (LOSS) |
Dietary Supplement: Salba (Salvia hispanica L.) |
Unity Health Toronto [171] |
Phase II | Male/ Female: 77 |
Interventional |
| NCT02298426 | Body Constitution Classification Based Comprehensive Health Management Intervention on Obese Population |
Dietary Supplement: SCHSANDRA PLUS, YI RUI CAPSULE, Gest Aid Plus YI RUI CAPSULE health product made of Salvia, hawthorn, alisma, perilla oil microcapsules, panax powder, ginkgo biloba and other plant materials |
Hai-Jun Wang, Peking University | Completed | Male/ Female: 1452 | Interventional |
| NCT05068557 | EPICO: (Study for the Pro-resolution of Chronic Inflammation in Obesity. Original Acronym From Spanish) |
Dietary Supplement: Dietary plan along with chia/linseed oil capsules |
University of Guadalajara | NA | Male/ Female: 80 |
Interventional |
| NCT04208308 | Comparison of Effect of Animal and Plant Sources of Omega-3 PUFA in Prevention of Cardiovascular and Metabolic Diseases | Chia seeds supplementation | Pavol Jozef Safarik University | Early phase I | Male/ Female: 300 (Estimated) |
Interventional |
3.1.1. Experimental Evidence of Salvia Officinalis Against Obesity and Metabolic Disorders
A study by Ben Khedher et al. employed an HFD-induced mice model of obesity, inflammation, and insulin resistance to examine the possible anti-obesity benefits of methanol (MeOH) extract of S. officinalis, for five weeks, the trial assessed lipid content, body weight, fat mass, liver glycogen levels, energy expenditure, and food consumption [24]. Furthermore, lipid droplet formation was shown to be reduced in 3T3-L1 cells in in-vitro assays. Notably, adipocyte lipogenesis was inhibited at a lower dosage of the sage MetOH extract. Supplementation of crushed shoots from S. sclarea and S. officinalis to the meals of high-fat diet (HFD) rats showed possible health effects in the study [25]. They suggested that Salvia spp. could be used as a dietary additive for improving complications in obese patients. The results showed a significant reduction in organ weight with potent anti-atherosclerotic activity. The effects of Hail S. officinalis total extract (SOTE) and its high flavonoid fraction (SOHFF) were examined for eight weeks in the research by Othman et al. on obesity in rats produced by HFD [26]. It was noted that along with histological alterations, the administration of SOTE and SOHFF alleviated the substantial increases in body weight, blood insulin, leptin levels, and the atherogenic index. The results implied that SOTE and SOHFF are characterized by hypolipidemic ability and might be used as natural therapeutics to mitigate the problems associated with obesity. This study from Abdulhussein et al. comparatively evaluated the impact of the market-available drug, Xenical and alcohol extract from S. officinalis leaves on the hyperlipidaemic white male mice fed with a high-fat diet for 60 days [27]. The biochemical and histological analysis showed that the alcohol extract from S. officinalis leaves showed beneficial effects in reducing hyperlipidemia. Samah identified the presence of phenols, steroids, terpenoids, flavonoids, tannins, and glycosides in the leaves of S. officinalis L. The methanolic extract was recorded with highest concentration of total phenols [28]. The results showed that the MeOH extracts had the greatest effectiveness against lipase and showed the strongest inhibition of α-amylase activity. In the study by ALsherif, S. officinalis aqueous extract (SAGEAE) showed potential anti-obesity properties against rats with HFD-induced obesity and metformin [29]. Eight weeks later, SAGEAE successfully ameliorated the high body weight, blood sugar, insulin, lipid profiles variables in HFD groups to normal. The extract has shown promising potential as a new pharmaceutical formulation for managing obesity.
3.2. Salvia hispanica
Chia, scientifically known as S. hispanica and commonly referred to as Mexican chia or salba chia, is a flowering plant of Lamiaceae family and is primarily cultivated for its edible seeds [30]. Native to southern Mexico and northern Guatemala, chia plants are currently well cultivated in Australia, Bolivia, Colombia, Guatemala, Peru, Argentina, and Mexico, the latter country been the largest producer of chia seeds worldwide [31]. S. hispanica has been reported with significant nourishment value that attributes to the well-being of an individual. The nutritional profile demonstrates the chia seeds have a higher protein (18.9%) and lipid (31.2%) content to lower carbohydrate content (3.4%). These are also a great reservoir of Poly unsaturated fatty acids (PUFA) such as α-linolenic acid (19.5%) and α-linoleic acid (5.2%), that are essential for humans but are not synthesized in the body [32]. Prevalent from the pre-Columbian period, the consumption of chia has been ever increasing as the plant is a great source of energy, strength, and endurance. Incorporation of chia to the daily diets have been reported with improved physiological and sensory characteristics due to its higher concentrations of dietary fibers, essential fatty acids, proteins, minerals, antioxidants, vitamins. Due to these diverse nutritional properties, dry chia seeds have made its various food stuffs like smoothies, salads, yogurt, soups, baked products [33]. There are several studies that correlates to the amelioration of obesity related disorders, cardiovascular diseases and oxidative stress with the dietary intervention of chia seeds [34].
Additionally, these seeds serve as a noteworthy reservoir of polyphenols and antioxidants [35]. The phenolic content in the dried chia seeds is around 8.8% which includes certain phenolic acids, including as kaempferol, caffeic acid, chlorogenic acid, quercetin, rosmarinic acid, gallic acid, cinnamic acid, and myricetin. Trace levels of isoflavones such genistein, daidzein, and glycitein have also been reported in the chia seeds [36,37,38]). These bioactive components are believed to have various health benefits, including anti-carcinogenic, antihypertensive, and neuroprotective effects. Rosmarinic acid and daidzein were identified as the major phenolics among the 27 identified using HPLC-DAD-MS/MS. The study also identified the procyanidin dimers in the chia seeds for the first time. Presence of varied phenolic acids, flavonoids and proanthocyanidins in the seeds of S. hispanica points towards an excellent source of nutraceutical [39]. Polyphenolics are major phytochemicals that contributes to the radical scavenging activity, thus, contributing to antioxidant potential. Chia seeds and oils are a great reservoir for natural antioxidants such as polyphenols, phytosterols, carotenoids, tocopherols [40]. Grancieri et al focused on elucidating the bioactive peptides from the chia seeds. The study catalogued 20 different proteins of which 12 were involved in the regular metabolic functioning in the plant cell while, the rest 8 were specifically involved in the production and storage of the plant lipids, explaining high lipid concentrations in chia seeds [41]. Further, amino acid analysis showed the biologically active nature of the peptides that has been correlated with major health benefits specifically antioxidant ability related to the chia seeds. Where chia seeds have been renowned for bioactive potential, chia leaves have also gained attention for the same. Amato et al. tentatively identified 34 polyphenols from the vegetal matrix of the S. hispanica using HPLC-MS [42]. Along the polyphenols already identified, vitexin, orientin, apigenin and some hydroxycinnamic acids were reported for the foliar part of the plant. While, exploring the chia plant for its bioactive potential, Elshafie et al. targeted essential oil extracted from the aerial part of the plant [43]. The study reported that sesquiterpenes, majorly caryophyllenes were the principal constituents in the aerial plant part which could have a potential application for microbial control.
3.2.1. Experimental Evidence of Salvia hispanica Against Obesity and Metabolic Disorders
Fonte- Faria et al examined the effect of S. hispanica seed oil supplemented diet against the obese mice model [44]. The study reported a significant improvement in the blood glucose levels and insulin tolerance with enhanced levels of high-density lipids. It was also observed that there was an increase in lean mass while fat accumulation reduced. In the mice fed with a high-fat diet, digested total proteins (DTP) from chia seeds showed a strong anti-inflammatory and anti-adipogenic properties. The administration of DTP supplements resulted in a decrease in waist circumference, adipocyte size, and plasma total cholesterol, LDL cholesterol, and triglycerides. The study emphasizes the potential therapeutic benefit of digested proteins generated from chia seeds in reducing inflammation and obesity. Singh et al. evaluated the effect of seed extracts of S. hispanica, Ocimum basilicum, and Coriandrum sativum on leptin activation and lipase activity in fat cells derived from chicken liver cells [45] The results point to the possibility of using these seed extracts to treat obesity [46]. A study by Rubavathi et al. assessed S. hispanica, notably the omega-3-rich chia seeds, for its ability to prevent obesity, as an alternative to the synthetic medications which have been reported with possible adverse effects [47] . The study concentrated on lowering cholesterol and body weight. Analysis of various extracts, including hexane, ethyl acetate, methanol, and water, revealed the methanol extract resulted in lowest cholesterol concentration and the highest in water extracts, highlighting chia seeds potential as a natural solution for managing obesity. According to Grancieri et al. the digested proteins (DP) from chia seeds have an anti-inflammatory effect and adipogenesis in mice fed a high-fat diet (HFD) [41]. These mice showed reductions in body fat percentage and plasma cholesterol levels, as well as decreases in adipocyte size, inflammation markers, and the mRNA expression of important adipogenesis-related factors. Dib et al. evaluated the phytochemical makeup and antioxidant capacities of chia seeds and reported high concentrations of the total phenols (19.06±0.14 mg GAE/g DW) and flavonoids (12.3±0.04 mg CE/g DW) [48]. While in another study, methanolic extract of S. hispanica decreased the viability of MCF-7 Breast Cancer cell line with the increase in the plant concentration [49]. Significant antioxidant activity of chia oil supplementation has been observed in a study by Batista et al. where the oil administration ameliorated the increased serum triacylglycerol levels in the high-fat group. An increased expression of antioxidant enzymes such as superoxide dismutase 1 and 2, catalase, and glutathione peroxidase were also observed in the liver [50].
3.3. Salvia miltiorrhiza
S. miltiorrhiza is a perennial plant from the genus Salvia that belongs to the mint family (Lamiaceae). The plant is referred to as Danshen or red rooted sage because of tanshinones, the primary phytochemical that provide the root with its distinctive red color [51]. S. miltiorrhiza is widely distributed in Chinese and Japanese province with damp and warm environment having an average temperature of 17°C and relative humidity of 77% [52]. Shennong’s Classic of Materia Medica (Eastern Han Dynasty) was the first to report the roots of sage as “super grade herb” which has been in clinical use for more than 2000 years. Later, traditional Chinese medicine (TCM) also recorded the red sage as an herb with health promoting benefits as it provided a promising treatment for chronic consumptive disorders, irregular menstruation, waist and spine pain, congestion following delivery, irregular menstruation, rheumatism, and may other disorders [53]. The presence of various bioactive phytochemicals in this species are responsible for the health promoting properties associated with it.
Till date more than 200 compounds have been extracted and identified from S. miltiorhiza. The phytochemical from this Salvia spp. are broadly categorised into two main types : hydrophilic phenolic compounds and lipid-soluble diterpenoid compounds [54]. The tanshinones present are mainly diterpenoid quinones, which could be classified into two series, one is the phenanthro [1,2-b] furan-10,11-diones and other is the phenanthrol [3,2-b] furan-7, 11-diones. A phytochemical analysis conducted by Zhong et al. used ultraperformance liquid chromatography (UPLC) to determine 13 hydrophilic and lipophilic compounds- protocatechuic acid, protocatechuic aldehyde, caffeic acid, ferulic acid, isoferulic acid, rosmarinic acid, salvianolic acid B, salvianolic acid A, dihydrotanshinone I, przewalskin, cryptotanshinone, tanshinone I, and tanshinone IIA from S. militorhiza [55]. Tanshinone IIA (tan-IIA) was reported to protected the macrophages from H2O2-induced cell death in animal models of atherosclerosis. It was also observed that tan-IIA significantly ameliorated the levels of GPx-1 mRNA levels [56]. Zou et al. aimed to isolate and identify salvianolic acids from Danshen to explore its anti-inflammatory potential [57]. The results indicated that two novel bioactive components along with four already known which isosalvianolic acid A-1 (C1) and ethyl lithospermate (C5). These inhibited neutrophil migrations in three zebrafish inflammation models while C1 decreased the secretion of IL-6 and TNF-α and inhibited the expression level of p-IκBα (Ser32) in LPS stimulated RAW 264.7 cells. Another phytochemical caffeic acid derivatives isolated from the roots of S. miltiorrhiza Bunge showed anti-inflammatory potential. The extraction of two new caffeic acid derivatives, and eight known derivatives, were isolated from the ethyl acetate fraction of S. miltiorrhiza [58]. They reported that compounds shimobashiric acid B and rosmarinic acid methyl ester inhibited NO production with IC50 values of 1.4 and 0.6 μM, respectively and were also observed to strongly inhibited the production of iNOS and COX-2. Although major therapeutic potential of S. miltiorhiza is related to its secondary metabolites, but a natural polysaccharide has been extracted from the roots of S. miltiorrhiza, SMP (S. miltiorrhiza polysaccharide) and is an important active component that have gained more attention for their numerous pharmacological uses. The polysaccharides are known for their anti-tumor, hepatoprotection, antioxidant, immunomodulatory, anti-inflammatory activities. Luo et al. revealed that polysaccharides extracted from S. miltiorrhiza shows antioxidant activity [59]. The results showed a significant DPPH and hydroxyl free radical scavenging activity.
3.3.1. Experimental Evidence of Salvia miltiorrhiza Against Obesity and Metabolic Disorders
A significant improvement in body weight management, in insulin resistance, decrease in visceral fat weight, were observed in an 8-week period study by Jung et al. which examined the combined effects of prolonged endurance exercise and S. miltiorrhiza vinegar (SMV) on body composition and insulin resistance in obese rats induced by a high-fat diet [60] . Although, significant benefits were not observed when SMV was supplemented alone, indicating that anti-hyperglycemic and anti-obesity potentials are predominantly derived from the synergistic combination of SMV and endurance exercise. Another study carried out by An et al. showed the efficiency of Salvianolic acid B (Sal B) produced from S. miltiorrhiza in reducing metabolic abnormalities associated with obesity [61]. Illumina Hiseq 4000 was used to analyse the effects of Sal B on the expression of circular RNA and long non-coding RNA in the epididymal white adipose tissue of HFD fed mice. Thet study suggested that Sal B could be used as an effective alternative to treat obesity. The anti-obesity properties of Tanshinone I (Tan I), a diterpenoid derived from S. miltiorrhiza Bunge, were examined in the research conducted by [62]. Administration of Tan I to HFD mice’s decreased adipogenesis by decreasing mitotic clonal expansion (MCE) and early adipogenic transcription factors according to in-vitro experiments. The results showed that Tan I prevents early adipogenesis and mitigated metabolic abnormalities related to obesity. With an emphasis on gut microbiota (GM) and lipid metabolism, Ai et al. examined the anti-obesity effects of S. miltiorrhiza (Sal) in rats with obesity induced by (HFD) [63]. Sal therapy improved gut integrity and lipid factors in liver and adipose tissues, lowering body weight, body fat index, blood lipid levels. The therapy also inhibited Firmicutes and Desulfobacterita while boosting Actinobacteriota and Proteobacteria, that had an impact on GM, which correlated with reduced triglycerides, and cardiolipins and increased ceramides and (O-acyl)-ωhydroxy fatty acids. The results highlighted that Sal therapy has a great potential for GM modulation and lipid metabolism for obesity management. S. miltiorrhiza extract (SME) showed notable anti-obesity benefits in mice fed with a high-fat diet (HFD) [64]. The administration of SME therapy led to a decrease in body weight, fasting plasma glucose, triglyceride levels, and insulin resistance. Additionally, genes linked to adipogenesis and lipogenesis were downregulated. ATF3-mediated downregulation of C/EBPα and modification of the C/EBP homologous protein (CHOP) pathway were the mechanisms by which a novel derivative, ST32db, from S. miltiorrhiza, demonstrated anti-obesity benefits.
3.4. Salvia libanotica
S. libanotica is an East Mediterranean plant known as S. fruticosa, or Greek sage is a perennial herb or sub-shrub. The plant is especially abundant in Palestine, Israel, Lebanon, Southern Italy, the Canary Islands, and North Africa. The herb is also referred to as a three-lobed sage because of its trifoliate leaves, which distinguish it from S. officinalis. The plant has also been given a sacred name ‘Herba sacra’ by the Salerno School of Medicine Valnet, 1990. The herb has been traditionally important in Lebanon because of its remedial potential against ailments such as common colds, headaches, and stomach aches. Besides Lebanon, the plant holds a great traditional value in Greece as well in the fields of beauty, medicine, and culinary, along with its sweet nectar and pollen. The tea from the sage leaves has been reported to provide relief from headaches, stomach aches, and various other disorders [65].
The plant holds significant therapeutic potential due to the existence of various phytochemical components. More than 100 volatile compounds have been isolated from the essential oils of S. fruticosa, which mainly belonged to the classes of monoterpenes, sesquiterpenes, diterpenes, and non-isoprenoids. The three most prevalent compounds among these were reported to be 1,8-cineole, camphor, and thujone [66]. Furthermore, about 160 polyphenolic compounds, such as flavonoids, anthocyanins, phenolic acids, and distinctive derivatives of caffeic acid like rosmarinic acid, as well as phenolic diterpenes like carnosic acid and other phenolic glycosides, have been found in the non-volatile essential oils [67]. A study conducted by Boukhary et al. isolated eight phenolic compounds from methylene chloride, ethyl acetate, and butanol extracts of S. libanotica [68]. The results showed that among these compounds three flavonoids named apigenin, luteolin, and rutin and three phenolic acids namely, ferulic acid, gallic acid, and rosemarinic acid were reported from the aerial parts of the medicinal plants. In addition to this, the root extract of the plant was reported with carnosol and dehydro-abietic acid. Villalva et al. observed significant antioxidant and anti-inflammatory properties of an extract supplemented with rosmarinic acid [69]. The results showed significant amelioration in the levels of TNF-α, IL-1β, and IL-6 in the human THP-1 macrophage model. Yet another molecular study performed by Subramanya et al. concentrated on 1,8-cineole, a bioactive monoterpenoid that stimulates the colon PPAR-γ transcription factor to reduce inflammation in the colon [70]. The results proved the isolated phytochemical to be a strong activator of the PPAR-γ transcription factor. Hijazi et al. explored the anticancer potential of abietane diterpenes isolated from the roots of S. libanoticum [71]. The results revealed that eight abietane diterpenes were isolated, among them 7α-acetylhorminone, indicated IC50 of 18 and 44 μM on HCT116 and MDA-MB-132 cells, respectively. The study led by Alqudah et al. assessed the potential modes of action and anti-inflammatory abilities of S. fruticosa essential oil (EOSF) [72]. The outcomes of the research showed that carrageenan-induced edema, peritonitis, myeloperoxidase activity, and NOx-peritoneal lavage concentration were all significantly ameliorated. Notably, sodium nitroprusside-stimulated nitric oxide radical generation and Fe2+ induced lipid peroxidation was also inhibited. A promising investigation carried out by Dawra et al. explored the phytochemical composition of S. fruticosa (Mill.) and assessed its antioxidant capacity, anti-proliferative effects and antibacterial activity on the HCT-116 and Caco-2 human colon cancer cell lines [73]. The methanol (MeOH) extract showed significant antioxidant activity, with an IC50 value of 19.4 μg/mL and an inhibition percentage (IP) of 76.1%. Another study by Kyriakou et al. elucidated the possible anti-melanoma properties of S. fruticosa (SF) utilizing an in-vitro model of human malignant melanoma with eight different fractions of the extracts [74]. The results demonstrated significant cytotoxic effects on A375 cells, indicating its potential efficacy in the treatment of melanoma.
3.4.1. Experimental Evidence of Salvia libanotica Against Obesity and Metabolic Disorders
In research led by Bassil et al. examined the rats fed with high-fat diet for the effects of S. libanotica [75]. Following six weeks of increasing dosages of S. libanotica extract, there was an improvement in glucose tolerance, while the levels of insulin and liver glycogen were increased, along with improved HDL/LDL cholesterol ratio and decreased abdominal fat. These results reflect the traditional use of S. libanotica in the prevention of cardiovascular illnesses and type 2 diabetes linked to high-fat diets. In a comparative study carried out by Mervić et al. observed the antioxidant, anti-acetylcholinesterase, and anti-α-glucosidase activities of S. nemorosa, S. officinalis, S. pratensis, S. sclarea, and S. verticillate [8]. All the Sage species were reported with α-glucosidase inhibition, however the results showed that S. fruticosa to be the most prominent inhibitor of the enzyme with an IC50 of 5291.51 ± 335.08. Another investigation by Dawra et al. conducted the phytochemical analysis along with their therapeutic potential and biological properties of the below mentioned medicinal plants Origanum ehrenbergii Boiss., Origanum syriacum L., Stachys nivea Labill., Stachys ehrenbergii Boiss., Cousinia libanotica DC., S. fruticosa Miller, and Berberis libanotica C.K. Schneider [76]. The results reported that the aqueous extract of S. fruticosa had a promising anti-hyperglycemic activity with an IC50 value of 167.4 μg/mL. Ercan and El reported the inhibitory effects of carnosic acid, an important phenolic diterpene extracted from S. fruticosa and Rosmarinus officinalis L. on various digestive enzymes [77]. The results showed that S. fruticosa inhibited α-amylase with IC50 value of 107.65 ± 12.64, and lipase with IC50 6.20 ± 0.63. This study by Drikvandi et al. focused on examining the effects of camphor, a key bioactive component derived from S. libanotica, on oxidative and serum biochemical markers on alloxan-induced diabetic rats [78]. Over a period of 20 days, it was found that consuming camphor raised the level of high-density lipoprotein cholesterol in the blood while lowering blood levels of glucose, total cholesterol, triglycerides, and LDL cholesterol.
3.5. Salvia plebeia
S. plebeia, also known as “Li-Zhi-Cao in Chinese is an annual or biennial plant belonging to the family Labiaceae. It is native to Asia, including China, Korea, Japan, Afghanistan, and India, where it grows in a myriad environments including slopes, stream sides, and damp fields ranging from sea level to heights of 9,200 feet [79]. Though presently appreciated for its medicinal attributes, the herb has been associated with its utility in long traditional health benefits. S. plebeia was employed for traditional herbal treatment in the Chinese Ming Dynasty about 490 years ago, according to the Compendium of Materia Medica. It has been used in TCM (Traditional Chinese Medicine) prescription and home remedies because of its ability to eliminate heat and cleanse the body of impurities. In India, the plant is used as a folk medicine by the name Badarangboya, as well as in South Korea where it has been used to treat common cold, flu, and cough [80]. Certain diseases like hepatitis, asthma, inflammatory illnesses, and diarrhea have been effectively treated with a combination of S. plebeia and Korean red ginseng [81]. Through several clinical trials, it has also been elucidated that the plant possessed antiviral, antitumor, hepatoprotective, hypoglycemic, antioxidant, anti-inflammatory, analgesic, and sedative properties. The plant offers all these significant health benefits due to its variety of beneficial bioactive phytochemicals [82].
Several bioactive compounds were reported to be found in S. plebeia, including flavonoids, phenolic acids, phenylpropanoids, sesquiterpenoids, and diterpenoids [83]. Dai et al. identified 70 metabolites including 46 flavonoids, 16 phenolic acids, seven terpenoids, and one organic acid, out of which 21 were novel phytochemicals reported in S. plebeian [84]. Although the main bioactive compounds, reported as per former studies are flavonoids, especially hispidulin, and homoplantaginin. In a study conducted by Patel and Patel Hispidulin, a naturally occurring phenolic flavonoid derived from S. plebeian was reported as an inhibitor of microsomal lipid peroxidation and scavengers of oxygen free radicals under both in-vitro and in-vivo conditions [85]. Another important phytochemical reported is Homoplantaginin (Hom), a major flavonoid discovered by Fan et al. in S. plebeia R. Br [86]. The phytochemical administration improved the high glucose (HG) conditions on vascular endothelial cells in db/db mice. Results obtained in-vitro showed a strong downregulation of apoptosis with an increase in autophagosome formation. The report also suggested an improved lysosomal activity and increased expression of cathepsin B and LAMP1. While, the in-vivo investigations indicated increased p-AMPK and TFEB protein expression, enhanced autophagy, reduced apoptosis, and vascular damage mitigation. Many studies have demonstrated that hispidulin and homoplantaginin have multiple pharmacological effects, such as anticancer, anti-inflammatory, neuroprotective, anti-epileptic, and thus, alleviated vascular endothelial cell apoptosis. Choi et al. also reported a significantly higher free radical scavenging potential of the phenolic compounds extracted from hairy root cultures of S.plebeia when compared to the wild-type roots [87]. Another promising study performed on the herb by Jang et al. focused on the anti-inflammatory effects of the ethanolic extract of S. plebeia plant [88]. The in-vitro suggested that the ethanolic extract isolated from both aerial parts and roots were effective in ameliorating the pro-inflammatory cytokine (IL-6 and IL-8) in BEAS-2B epithelial cell lines. While the ethanolic extracts from the aerial parts mitigated the LPS induced Nitric Oxide (NO) inflammation.
3.5.1. Experimental Evidence of Salvia plebeia Against Obesity and Metabolic Disorders
In mice fed with high-fat diet, this study by Choi et al. examined the anti-obesity effects of ethanolic extracts from S. plebeia R. Br (SPE) [87]. After eight weeks of administration, a substantial reduction in body and adipose tissue weights were reported along with a drop in blood glucose, lipid levels and reduction in size of adipocyte. A similar study focusing on the antioxidant activity and lipid composition of an aqueous extract of S. plebeia in high-fat-induced rats was conducted by [89]. After a 4-week-long administration of the extract, it resulted in a significant reduction in the serum total lipid, triglyceride, total cholesterol, and LDL-cholesterol levels. Pereira et al evaluated the decoctions from four Salvia spp. (S. elegans Vahl., S. greggii A. Gray, and S. officinalis L.) for their health promoting abilities [90]. The study particularly focused on the antioxidant potential and their ability to inhibit the key enzymes in obesity and diabetes pathogenesis (α-glucosidase, α-amylase and pancreatic lipase). It was observed that where S. elegans was reported as the most potent inhibitor of α-glucosidase, S. officinalis was reported with a good inhibitory activity against xanthine oxidase. High flavones content including glycosidic forms of apigenin, scutellarein and luteolin were reported in S. officinalis while caffeic acid richness in S. elegans decoctions contributed to health benefiting properties. Similarly a study led by Eom et al. investigated the physiological activity and nutritional makeup of plants growing in Chungbuk Province, including Saururus chinensis Baill, Gynura procumbens, Angelica acutiloba, and S. plebeia R. Br [91]. The results showed that S. plebeia exhibited α-glucosidase inhibitory action, however it was not as strong as the other three species.
4. Unveiling the Role of Gut Microbiota in Obesity
The term “gut microbiota” refers to the diverse microbial population found within the gastrointestinal tracts of humans and other animals which includes bacteria, fungi, viruses, and archaea. The domain that is most closely associated with the gut microbiota is the large intestine, specifically the colon which serves as a reservoir for a complex microbial diversity, reported with about 1011 bacteria per gram of intestinal material [92]. The gut flourishes with diverse bacterial groups, but the majority of bacteria belong to two major phyla, Bacteroidetes and Firmicutes which constitute around 70-90% of diversity. At the same time, Proteobacteria, Actinobacteria, Synergistetes, and Fusobacteria are also present. Although nearly all of these species have been reported as obligate anaerobes, different degrees of aerotolerance have been shown by the Actinobacteria and Proteobacteria phyla [93]. There has been a significant influence of the gut microbiota on the metabolic functioning of the host. It helps break down proteins into amino acids and saturates dietary unsaturated fatty acids [94]. Gut microbiota also influences lipid metabolism as it has been reported to modulate the absorption of dietary fats, synthesis of lipids in the liver, and storage of fat in adipose tissue. Concerning the hormones of the host, gut microflora mimics those involved in metabolic control, primarily affecting energy expenditure, glucose homeostasis, and appetite management. The fermentation of complex carbohydrates and non-digestible dietary fibers by the gut microbiota results in the production of short-chain fatty acids (SCFAs), which can be used as an energy source [95]. Over the past 20 years, there has been immense research concerning the beneficial effects of gut microbiota and various diseases linked to the dysbiotic gut [96]. Various disorders such as type 2 diabetes (T2D), obesity, non-alcoholic liver disease (NALD), gastrointestinal disorders like inflammatory bowel disease (IBD) and irritable bowel syndrome (IBS), and cardiovascular diseases all have been linked with the dysbiosis of the gut microbiota [97]. The dietary habits of an individual have a major effect on the microbial composition of their gut. Consuming a high-fat diet alters the gut microflora by negatively impacting the density of Bacteroidetes and enhancing the levels of Firmicutes and Proteobacteria, which eventually has been concerned to gut dysbiosis [98].
Several environmental conditions as well as genetic factors have been correlated to the development of dysbiosis. These host-specific factors include genetic makeup, medical history, dietary habits, the exposure to drugs, food additives, and antibiotics. Also changes in the nutrient consumption are capable of causing significant changes in the makeup of the gut microflora [99]. These variations have profound physiological repercussions, like diets containing significant amounts of simple sugars have been reported to weaken the intestinal barrier, leading to inflammation and ultimately disrupting host metabolism. Although the influence of food additives on gut flora has long been neglected, there has been a recent increase in consciousness regarding this issue. The studies have reported that the gut microbiota is very susceptible to preservatives, which, when subjected, lead to the proliferation of Proteobacteria [59]. Another class of food additives that directly modifies the makeup of the human gut flora are dietary emulsifiers that has also been related to intestinal inflammation [100]. All these facts concerning microbial dysbiosis could become more clearer, by understanding the role of bacteria in the development of various metabolic disorders. A study conducted by Almugadam et al. investigated the relation between intestinal microbiota and type 2 diabetes patients [101]. They observed a significant decrease in the number of beneficial bacteria Faecalibacterium, Fusobacterium, Dialister, and Elusimicrobium. Inflammatory bowel disease (IBD) is also a concerning disorder, which mainly includes Crohn’s disease (CD) and ulcerative colitis (UC). Wiredu Ocansey et al. reported an imbalance in the microbial population during IBD, where the ratio of Bacteriodetes to Firmicutes is decreased while the abundance of gammaproteobacteria was reported in IBD patients [102]. Another study was reported on the faecal samples of NASH (Non-alcoholic Steatohepatitis disease) patients, where Liakina et al. observed the abundance of Parabacteroides and Allisonella but decreased levels of Faecalibacterium and Anaerosporobacter [103].
Sedentary lifestyles have been a major pitfall for the emergence of dysbiotic gut which paves a path for the development of metabolic syndrome where the term “metabolic syndrome” refers to a group of related metabolic disorders that include obesity, insulin resistance, dyslipidemia, hypertension, and insulin resistance. These disorders, either directly or indirectly, degrade the health of the individual. Since all these disorders are linked with imbalanced gut microflora, the main focus is to improve the dysbiotic gut, thus ameliorating various metabolic disorders. Obesity has become a prevalent health issue throughout the Asia, particularly in South Asian countries and India being the most populous country of South Asia, have around 3.5 billion cases of abdominal obesity [104,105]. It is believed that Asians have an inherited tendency to accumulate extra fat around their abdomens [106]. A complex association between the gut microbiota and metabolic syndrome have been reported with obesity. In particular, dysbiosis in the gut microbiota has been marked by distinctive modulations such as relative abundance and scarceness of certain microbial species in the gut [107]. Proteobacteria has been the most consistently reported obesity-associated phylum. Xu et al. comprehensively reported the difference in the microbial species related to the obese and lean individuals in their study [108]. They reported that Faecalibacterium, Akkermansia, and Alistipes were lean-associated genera whereas Prevotella and Ruminococcus were obesity-associated genera in studies from the West while the same were associated with lean in the East. Roseburia and Bifidobacterium were lean-associated genera in the East, whereas Lactobacillus was an obesity-associated genus in the West.
Furthermore, it has been observed the metabolites secreted by microbes also have been strongly associated with obesity. The metabolites are essential for controlling body weight, and their imbalance might result in the development of obesity. Adipokines, such as leptin and adiponectin, help in the formation of insulin resistance and abnormal appetite regulation during obesity [109]. Similarly, the link between obesity and Medium-chain fatty acids (MCFAs) has also surfaced. Caproic acid, a major Medium-chain fatty acid present in the gut, helps in maintaining healthy cholesterol levels and regulating body weight and fat deposition. Studies have been observed where obese patients have been reported with a decrease in the level of caproic acid. In a similar study by Patel et al. it was reported that there was a decrease in the levels of caproic acid in patients suffering Clostridium difficile infection [110]. The gut bacteria produce another subset of fatty acids known as Short-chain fatty acids (SCFAs), synthesized by anaerobic microbial fermentation of partially and nondigestible polysaccharides. Several studies have also demonstrated that the levels of SCFA produced by gut bacteria regulate obese conditions [111]. The basic mode of action of SCFA is the stimulation of beige adipogenesis and biosynthesis of mitochondria, reduction in chronic inflammation, and improvement in the breakdown of triglycerides and free fatty acid oxidation [112]. The enteric gut microbiota produces SCFA such as acetate, propionate, and butyrate, which have a range of essential functions in the gut [113]. Butyrate SCFA serves as major source of nutrients for colonocytes, while maintain a barrier integrity of intestinal membrane and decreasing the intestinal inflammation. Collectively acetate, propionate, and butyrate enable G-protein-coupled receptors (GPRs), which regulate fundamental satiety hormones such as GLP-1 and peptide YY (PYY). Studies have shown that supplementation with short-chain fatty acids can help prevent weight gain. Another study on preadipocyte cultures illustrated that the increased expression of proteins like FA binding protein 4 (FABP4) and FA transporter protein (FATP), as well as enzymes like lipoprotein lipase (LPL) and FA synthase (FAS), is all attributed to the addition of acetate, propionate, and butyrate that are involved in lipid metabolism ([114]. Wu et al. (2021) analyzed the diets with a high fiber content of primary SCFAs, namely, acetate and propionate, dramatically lower blood pressure levels in rat models with spontaneous hypertension and deoxycorticosterone acetate salt-induced hypertension [115].
5. Exploring the Mechanisms by which Gut Microbiota Influence Obesity
5.1. Gut Microbiota Interactions with Adipose Tissue
The gut microbiota plays a pivotal role in regulating the adipose cell accumulation in a very intriguing way. It could affect the production of metabolic changes released by white adipose tissues, linked with inflammation, and how brown and beige adipose tissues function in terms of energy expenditure and reducing weight [116]. It facilitates that primarily, through 2 mechanistic pathways. Firstly, the inhibition of lipoprotein lipase (LPL) by reducing the fasting-induced adipose factor (Angptl4/Fiaf) and Angiopoietin-like protein 4, which are mainly released by both white and brown adipose tissue, reduces the rate at which fatty acids are oxidized in muscle and adipose tissues [117]. Therefore, LPL inhibition by Fiaf reduces fat deposition. Studies reported that increased LPL activity is a result of the B. thetaiotaomicron-induced inhibition of Fiaf [118]. Adjacently, the role of beneficial microbiota was discussed in a study conducted by Aronsson et al., where it was observed that mice supplemented with Lactobacillus paracasei ssp. paracasei F19 showed reduced fat accumulation via increasing the levels of circulating fasting-induced adipose factor (Fiaf).
5.2. Microbial Influence on Bile-Acid and Cholesterol Metabolism in the Gut
Apart from the genetic makeup and various environmental factors oh the host, gut microbiota also contributes in playing a critical role in the pathway that regulates the metabolism of cholesterol and bile acids (Figure 1). Mainly the pathway involves the conversion of cholesterol into coprostanol to Bile Acid metabolism. Two main routes are involved in this biotransformation. The first involves the direct, stereospecific reduction of cholesterol’s 5,6-double bond, whereas the second involves an indirect transition that results in the intermediate production of cholestenone and coprostanone. There are many strains of bacteria like Bifidobacterium, Lactobacillus, and Peptostreptococcus found to reduce cholesterol. Interestingly, the upper small intestine is the primary site of cholesterol absorption which is mostly harboured by lactic acid bacteria, namely Lactobacillus, that have the ability to convert cholesterol into coprostanol [119].
Secondly the in the metabolism of bile acids the gut microbiota present in the intestine secretes various microbial that metabolise primary and secondary bile acids. So during this process the primary bile acids enzymes which are Glyco-conjugated and tauro-conjugated Cholic acid (CA) and chenodeoxycholic acid (CDCA) firstly get deconjugated with the help of a catalyst enzyme Bile Salt Hydrolase( BSHs), then get epimerized by hydroxysteroid dehydrogenases released by the intestinal bacteria and then get 7α-dehydroxylated to finally form secondary bile acids namely, deoxycholic acid (DCA) and lithocholic acid (LCA) which are eventually absorbed and utilized by the liver [120].
5.3. Influence of Gut Microbiota on Energy Homeostasis and Satiety Regulation
Another important role of gut microbiota is on the energy production and regulating satiety. These are the factors that support the initiation and progression of metabolic disorders [121]. Importantly, the bacterial metabolites released by the gut microbiota help in this process of regulating metabolism and energy intake. One way is the direct stimulation of neurons which then generate the release of satiety hormones like glucagon-like peptide 1, GLP-1; peptide YY, PYY; and cholecystokinin, CCK and other being the entry into the circulation, thereby directing and influencing both the central and peripheral nervous systems [122]. Furthermore, the gut microbiota regulates intestinal and systemic inflammation and modulates the permeability of the intestinal barrier. There has been a higher rate of neuroinflammation in certain brain areas that are ultimately linked in sustaining satiety and satiation in cases of systemic low-grade inflammation [123]. Another important microbial metabolite that helps in contributing the regulation of satiety and energy intake are the short chain fatty acids [124]. They help in inducing the release of several satiety-related gut hormones like the gut peptides GLP-1 and PYY. Christiansen et al. observed an increase in the amount of PYY and GLP-1 that are produced when administered with intestinal SCFAs which directly or indirectly help in controlling hunger [125].
5.4. Endocannabinoid System
The Endocannabinoid (eCB) system is the system, which regulates appetite and metabolism with the help of the microbiota-gut-brain axis and is considered as important factor which influences obesity [126]. It has a significant impact on energy balance and has been suggested that during obesity there is an imbalance in this system. The eCB system consists of endogenous bioactive lipids that are produced locally, certain G protein-coupled receptors and the regulatory proteins that control their synthesis and breakdown. The microbiota can affect the integrity of the intestinal barrier through the ECS. Therefore, changes in the microbiota could mitigate the signs of illness brought on by a weakened intestinal barrier [127]. One such example is of Irritable bowel syndrome where a study conducted by Jansma et al. observed that when the cannabinoid receptor 1 (CB1) was activated, intestinal peristalsis was decreased by inhibiting ileal contractions and regulating vagal neurotransmission activity [128]. Another study by Xu et al. observed that ingestion of Lactobacillus acidophilus NCFM stimulates the expression of the CB2 receptor in the intestine of mice, which has analgesic properties against visceral discomfort [108].
6. Conclusion and Future Perspective
In conclusion, the investigation into Salvia spp. as a phytomedicine for obesity prevention illuminates a promising pathway in combating the global obesity pandemic. Supported by experimental evidence, Salvia extract exhibit notable potential in ameliorating obesity and related metabolic disorders, owing to their rich array of bioactive compounds. Additionally, delving into the intricate interplay of gut microbiota in obesity elucidates crucial mechanistic insights into how interventions like Salvia spp. extracts may manifest their effects. Looking ahead, the future of utilizing Salvia in the realm of obesity holds great promise. Further research avenues encompass not only optimizing the extraction methods and dosage formulations of Salvia extracts but also exploring synergistic combinations with existing therapeutic modalities. Embracing interdisciplinary collaboration and fostering continued scientific inquiry will be pivotal in harnessing the therapeutic repertoire of nature, particularly Salvia, to confront the multifaceted challenges of obesity on a global scale.
Data availability
Not applicable
Acknowledgments
Declared none.
Conflict of Interests
The authors declare that they have no conflict of interest in the publication.
References
- A. Alice, M. Yadav, R. Verma, M. Kumari, S. Arora, Effect of obesity on balance: A literature review, Ijhs (2022) 3261–3279. [CrossRef]
- WHO, World Health Organization. Obesity and Overweight., (2023). https://www.who.int/news-room/fact-sheets/detail/ obesity-and-overweight.
- WHO, Prevalence of obesity. https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity, (2021). https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity.
- World Obesity Atlas. https://www.indiaenvironmentportal.org.in/content/474617/world-obesity-atlas-2023/, (2023). http://www.indiaenvironmentportal.org.in/content/474617/world-obesity-atlas-2023/.
- N. Hamjane, F. Benyahya, N.G. Nourouti, M.B. Mechita, A. Barakat, Cardiovascular diseases and metabolic abnormalities associated with obesity: What is the role of inflammatory responses? A systematic review, Microvascular Research 131 (2020) 104023. [CrossRef]
- A. De Lorenzo, S. Gratteri, P. Gualtieri, A. Cammarano, P. Bertucci, L. Di Renzo, Why primary obesity is a disease?, J Transl Med 17 (2019) 169. [CrossRef]
- A. Gasmi, P.K. Mujawdiya, A. Nehaoua, M. Shanaida, Y. Semenova, S. Piscopo, A. Menzel, V. Voloshyn, O. Voloshyn, V. Shanaida, G. Bjørklund, Pharmacological Treatments and Natural Biocompounds in Weight Management, Pharmaceuticals 16 (2023) 212. [CrossRef]
- M. Mervić, M. Bival Štefan, M. Kindl, B. Blažeković, M. Marijan, S. Vladimir-Knežević, Comparative Antioxidant, Anti-Acetylcholinesterase and Anti-α-Glucosidase Activities of Mediterranean Salvia Species, Plants 11 (2022) 625. [CrossRef]
- I. Marchioni, B. Najar, B. Ruffoni, A. Copetta, L. Pistelli, L. Pistelli, Bioactive Compounds and Aroma Profile of Some Lamiaceae Edible Flowers, Plants 9 (2020) 691. [CrossRef]
- M.-D. Mot, S. Gavrilaș, A.I. Lupitu, C. Moisa, D. Chambre, D.M. Tit, M.A. Bogdan, A.-M. Bodescu, L. Copolovici, D.M. Copolovici, S.G. Bungau, Salvia officinalis L. Essential Oil: Characterization, Antioxidant Properties, and the Effects of Aromatherapy in Adult Patients, Antioxidants 11 (2022) 808. [CrossRef]
- M. Sharifi-Rad, B. Ozcelik, G. Altın, C. Daşkaya-Dikmen, M. Martorell, K. Ramírez-Alarcón, P. Alarcón-Zapata, M.F.B. Morais-Braga, J.N.P. Carneiro, A.L. Alves Borges Leal, H.D.M. Coutinho, R. Gyawali, R. Tahergorabi, S.A. Ibrahim, R. Sahrifi-Rad, F. Sharopov, B. Salehi, M. Del Mar Contreras, A. Segura-Carretero, S. Sen, K. Acharya, J. Sharifi-Rad, Salvia spp. plants-from farm to food applications and phytopharmacotherapy, Trends in Food Science & Technology 80 (2018) 242–263. [CrossRef]
- M. Hamidpour, R. Hamidpour, S. Hamidpour, M. Shahlari, Chemistry, Pharmacology, and Medicinal Property of Sage (Salvia) to Prevent and Cure Illnesses such as Obesity, Diabetes, Depression, Dementia, Lupus, Autism, Heart Disease, and Cancer, Journal of Traditional and Complementary Medicine 4 (2014) 82–88. [CrossRef]
- A. Soltanbei̇Gi̇, E. Samadpourrigani, PHENOLOGICAL CYCLE AND DIURNAL VARIATION EFFECTS ON THE VOLATILE OIL CHARACTERISTICS OF SAGE (Salvia officinalis L.), Trakya University Journal of Natural Sciences 22 (2021) 59–65. [CrossRef]
- A. Ghorbani, M. Esmaeilizadeh, Pharmacological properties of Salvia officinalis and its components, Journal of Traditional and Complementary Medicine 7 (2017) 433–440. [CrossRef]
- H.-Y. Li, D.-D. Zhou, R.-Y. Gan, S.-Y. Huang, C.-N. Zhao, A. Shang, X.-Y. Xu, H.-B. Li, Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review, Nutrients 13 (2021) 3211. [CrossRef]
- P. Badiee, A.R. Nasirzadeh, M. Motaffaf, Comparison of Salvia officinalis L. essential oil and antifungal agents against candida species, J Pharm Technol Drug Res 1 (2012) 7. [CrossRef]
- M. Couladis, A. Koutsaviti, Chemical composition of the essential oils of Salvia officinalis, S. fruticosa, Melissa officinalis, and their infusions, Ratar i Povrt 54 (2017) 36–41. [CrossRef]
- M. Grdiša, Dalmatian Sage (Salvia officinalis L.): A Review of Biochemical Contents, Medical Properties and Genetic Diversity, (n.d.).
- C.F. Lima, P.C.R. Valentao, P.B. Andrade, R.M. Seabra, M. Fernandes-Ferreira, C. Pereira-Wilson, Water and methanolic extracts of Salvia officinalis protect HepG2 cells from t-BHP induced oxidative damage, Chemico-Biological Interactions 167 (2007) 107–115. [CrossRef]
- A.F. Afonso, O.R. Pereira, Â. Fernandes, R.C. Calhelha, A.M.S. Silva, I.C.F.R. Ferreira, S.M. Cardoso, Phytochemical Composition and Bioactive Effects of Salvia africana, Salvia officinalis ‘Icterina’ and Salvia mexicana Aqueous Extracts, Molecules 24 (2019) 4327. [CrossRef]
- B.N. Silva, V. Cadavez, C. Caleja, E. Pereira, R.C. Calhelha, M. Añibarro-Ortega, T. Finimundy, M. Kostić, M. Soković, J.A. Teixeira, L. Barros, U. Gonzales-Barron, Phytochemical Composition and Bioactive Potential of Melissa officinalis L., Salvia officinalis L. and Mentha spicata L. Extracts, Foods 12 (2023) 947. [CrossRef]
- H.M. Rashwan, H.E. Mohammed, A.A. El-Nekeety, Z.K. Hamza, S.H. Abdel-Aziem, N.S. Hassan, M.A. Abdel-Wahhab, Bioactive phytochemicals from Salvia officinalis attenuate cadmium-induced oxidative damage and genotoxicity in rats, Environ Sci Pollut Res 28 (2021) 68498–68512. [CrossRef]
- A.R. Jassbi, S. Zare, O. Firuzi, J. Xiao, Bioactive phytochemicals from shoots and roots of Salvia species, Phytochem Rev 15 (2016) 829–867. [CrossRef]
- M.R. Ben Khedher, M. Hammami, J.R.S. Arch, D.C. Hislop, D. Eze, E.T. Wargent, M.A. Kępczyńska, M.S. Zaibi, Preventive effects of Salvia officinalis leaf extract on insulin resistance and inflammation in a model of high fat diet-induced obesity in mice that responds to rosiglitazone, PeerJ 6 (2018) e4166. [CrossRef]
- M.A. Lieshchova, A.A. Bohomaz, V.V. Brygadyrenko, Effect of Salvia officinalis and S. sclarea on rats with a high-fat hypercaloric diet, Regul. Mech. Biosyst. 12 (2021) 554–563. [CrossRef]
- M.S. Othman, A.M. Khaled, G.M. Aleid, M.A. Fareid, R.A. Hameed, M.S. Abdelfattah, D.E. Aldin, A.E.A. Moneim, Evaluation of antiobesity and hepatorenal protective activities of Salvia officinalis extracts pre-treatment in high-fat diet-induced obese rats, Environ Sci Pollut Res 29 (2022) 75043–75056. [CrossRef]
- L. Abdulhussein, A. Allaithi, W. Matrood, W.M. Kadhem, STUDY OF THE EFFECT OF SALVIA OFFICINALIS LEAVES EXTRACT AND XENICAL DRUG ON SOME OF THE BIOCHEMICAL AND HISTOLOGICAL PARAMETERS IN THE RATS INDUCED WITH HYPERLIPIDEMIA, Plant Archives 19 (2019) 1111–1112.
- S. Samah, Phytochemical Screening and Anti-Obesity Activity of Salvia Officinalis L, International Journal of Academic Scientific Research 6 (2018) 33–45.
- D.A. ALsherif, M.A. Hussein, S.S. Abuelkasem, S.A. Hassan, M. Wink, Salvia Officinalis Improves Glucose Uptake and Suppresses Ectopic Lipid Deposition in Obese Rats with Metabolic Syndrome, CPB 24 (2023). [CrossRef]
- A.M. Jamshidi, M. Amato, A. Ahmadi, R. Bochicchio, R. Rossi, Chia (Salvia hispanica L.) as a novel forage and feed source: A review, Ital J Agronomy 14 (2019) 1–18. [CrossRef]
- D. Orona-Tamayo, O. Paredes-López, Chia—The New Golden Seed for the 21st Century: Nutraceutical Properties and Technological Uses, in: Sustainable Protein Sources, Elsevier, 2024: pp. 443–470. [CrossRef]
- R.M. Bodoira, M.C. Penci, P.D. Ribotta, M.L. Martínez, Chia (Salvia hispanica L.) oil stability: Study of the effect of natural antioxidants, LWT 75 (2017) 107–113. [CrossRef]
- B. Kulczyński, J. Kobus-Cisowska, M. Taczanowski, D. Kmiecik, A. Gramza-Michałowska, The Chemical Composition and Nutritional Value of Chia Seeds-Current State of Knowledge, Nutrients 11 (2019) 1242. [CrossRef]
- R.D.S. Marineli, S.A. Lenquiste, É.A. Moraes, M.R. Maróstica, Antioxidant potential of dietary chia seed and oil ( Salvia hispanica L.) in diet-induced obese rats, Food Research International 76 (2015) 666–674. [CrossRef]
- M. Knez Hrnčič, M. Ivanovski, D. Cör, Ž. Knez, Chia Seeds (Salvia Hispanica L.): An Overview—Phytochemical Profile, Isolation Methods, and Application, Molecules 25 (2019) 11. [CrossRef]
- N. Mohd Ali, S.K. Yeap, W.Y. Ho, B.K. Beh, S.W. Tan, S.G. Tan, The Promising Future of Chia, Salvia hispanica L., Journal of Biomedicine and Biotechnology 2012 (2012) 1–9. [CrossRef]
- B. De Falco, M. Amato, V. Lanzotti, Chia seeds products: an overview, Phytochem Rev 16 (2017) 745–760. [CrossRef]
- Md.J. Rahman, A.C. De Camargo, F. Shahidi, Phenolic and polyphenolic profiles of chia seeds and their in vitro biological activities, Journal of Functional Foods 35 (2017) 622–634. [CrossRef]
- Md.J. Rahman, A.C. De Camargo, F. Shahidi, Phenolic and polyphenolic profiles of chia seeds and their in vitro biological activities, Journal of Functional Foods 35 (2017) 622–634. [CrossRef]
- B. De Falco, A. Fiore, R. Rossi, M. Amato, V. Lanzotti, Metabolomics driven analysis by UAEGC-MS and antioxidant activity of chia (Salvia hispanica L.) commercial and mutant seeds, Food Chemistry 254 (2018) 137–143. [CrossRef]
- M. Grancieri, H.S.D. Martino, E. Gonzalez De Mejia, Chia Seed ( Salvia hispanica L.) as a Source of Proteins and Bioactive Peptides with Health Benefits: A Review, Comp Rev Food Sci Food Safe 18 (2019) 480–499. [CrossRef]
- M. Amato, M.C. Caruso, F. Guzzo, F. Galgano, M. Commisso, R. Bochicchio, R. Labella, F. Favati, Nutritional quality of seeds and leaf metabolites of Chia (Salvia hispanica L.) from Southern Italy, Eur Food Res Technol 241 (2015) 615–625. [CrossRef]
- H.S. Elshafie, L. Aliberti, M. Amato, V. De Feo, I. Camele, Chemical composition and antimicrobial activity of chia (Salvia hispanica L.) essential oil, Eur Food Res Technol 244 (2018) 1675–1682. [CrossRef]
- T. Fonte-Faria, M. Citelli, G.C. Atella, H.F. Raposo, L. Zago, T. De Souza, S.V. Da Silva, C. Barja-Fidalgo, Chia oil supplementation changes body composition and activates insulin signaling cascade in skeletal muscle tissue of obese animals, Nutrition 58 (2019) 167–174. [CrossRef]
- K. G Singh, A. Kour, A. Mohanty, In-vitro and Ex-vivo studies on Antiobesity Leptin Activation Effects of Saliva hispanica, Ocimum basilicum and Coriandrum sativum Extracts, IJPSRR 65 (2020) 114–120. [CrossRef]
- H. Martino, M. Grancieri, R. Toledo, T. Veridiano, C.T. Sant’Ana, N. Costa, E.G. De Mejia, Chia Seed (Salvia hispanica L.) Digested Total Protein Prevented Adipose Tissue Inflammation and Reduce Obesity Complications in Mice Fed a High-Fat Diet, Current Developments in Nutrition 4 (2020) nzaa045_069. [CrossRef]
- S. Rubavathi, G. Ayyappadasan, N. Sangeetha, T. Harini, D. Saranya, P. Harshapradha, Studies on Antioxidant and Anti-obesity Activity of Salvia hispanica (Chia) Seeds Extracts, J. Drug Delivery Ther. 10 (2020) 98–106. [CrossRef]
- H. Dib, M. Seladji, F.Z. Bencheikh, M. Faradji, C. Benammar, M. Belarbi, Phytochemical Screening and Antioxidant Activity of Salvia hispanica, JPRI (2021) 167–174. [CrossRef]
- H.A. Mutar, J.F.K. Alsadooni, ANTIOXIDANT AND ANTI-CANCER ACTIVITY OF CHIA SEED EXTRACT IN BREAST CANCER CELL LINE, ATMPH 22 (2019) 173–181. [CrossRef]
- A. Batista, F.T. Quitete, T.C. Peixoto, A. Almo, E.B. Monteiro, P. Trindade, L. Zago, M. Citelli, J.B. Daleprane, Chia (Salvia hispanica L.) oil supplementation ameliorates liver oxidative stress in high-fat diet-fed mice through PPAR-γ and Nrf2 upregulation, Journal of Functional Foods 102 (2023) 105462. [CrossRef]
- Z. Jiang, W. Gao, L. Huang, Tanshinones, Critical Pharmacological Components in Salvia miltiorrhiza, Front. Pharmacol. 10 (2019) 202. [CrossRef]
- H. Pang, L. Wu, Y. Tang, G. Zhou, C. Qu, J. Duan, Chemical Analysis of the Herbal Medicine Salviae miltiorrhizae Radix et Rhizoma (Danshen), Molecules 21 (2016) 51. [CrossRef]
- Y. Guo, Y. Li, L. Xue, R.P. Severino, S. Gao, J. Niu, L.-P. Qin, D. Zhang, D. Brömme, Salvia miltiorrhiza: An ancient Chinese herbal medicine as a source for anti-osteoporotic drugs, Journal of Ethnopharmacology 155 (2014) 1401–1416. [CrossRef]
- Q. Jin, X. Hu, Y. Deng, J. Hou, M. Lei, H. Ji, J. Zhou, H. Qu, W. Wu, D. Guo, Four New Depsides Isolated from Salvia miltiorrhiza and Their Significant Nerve-Protective Activities, Molecules 23 (2018) 3274. [CrossRef]
- G.-X. Zhong, P. Li, L.-J. Zeng, J. Guan, D.-Q. Li, S.-P. Li, Chemical Characteristics of Salvia miltiorrhiza (Danshen) Collected from Different Locations in China, J. Agric. Food Chem. 57 (2009) 6879–6887. [CrossRef]
- Y.-I. Li, G. Elmer, R.C. LeBoeuf, Tanshinone IIA reduces macrophage death induced by hydrogen peroxide by upregulating glutathione peroxidase, Life Sciences 83 (2008) 557–562. [CrossRef]
- L. Zou, D. Liu, H. Yang, C. Zhou, S. Deng, N. Xu, X. He, Y. Liu, M. Shao, L. Yu, J. Liu, Salvianolic acids from Salvia miltiorrhiza Bunge and their anti-inflammatory effects through the activation of α7nAchR signaling, Journal of Ethnopharmacology 317 (2023) 116743. [CrossRef]
- H.G. Choi, P.T. Tran, J.-H. Lee, B.S. Min, J.A. Kim, Anti-inflammatory activity of caffeic acid derivatives isolated from the roots of Salvia miltiorrhiza Bunge, Arch. Pharm. Res. 41 (2018) 64–70. [CrossRef]
- M. Luo, D.-D. Zhou, A. Shang, R.-Y. Gan, H.-B. Li, Influences of food contaminants and additives on gut microbiota as well as protective effects of dietary bioactive compounds, Trends in Food Science & Technology 113 (2021) 180–192. [CrossRef]
- D.Y. Jung, J.-H. Kim, M.H. Jung, Anti-Obesity Effects of Tanshinone I from Salvia miltiorrhiza Bunge in Mice Fed a High-Fat Diet through Inhibition of Early Adipogenesis, Nutrients 12 (2020) 1242. [CrossRef]
- T. An, J. Zhang, B. Lv, Y. Liu, J. Huang, J. Lian, Y. Wu, S. Gao, G. Jiang, Salvianolic acid B plays an anti-obesity role in high fat diet-induced obese mice by regulating the expression of mRNA, circRNA, and lncRNA, PeerJ 7 (2019) e6506. [CrossRef]
- I. Jung, H. Kim, S. Moon, H. Lee, B. Kim, Overview of Salvia miltiorrhiza as a Potential Therapeutic Agent for Various Diseases: An Update on Efficacy and Mechanisms of Action, Antioxidants 9 (2020) 857. [CrossRef]
- Z.-L. Ai, X. Zhang, W. Ge, Y.-B. Zhong, H.-Y. Wang, Z.-Y. Zuo, D.-Y. Liu, Salvia miltiorrhiza extract may exert an anti-obesity effect in rats with high-fat diet-induced obesity by modulating gut microbiome and lipid metabolism, World J Gastroenterol 28 (2022) 6131–6156. [CrossRef]
- Y.-L. Wu, H. Lin, H.-F. Li, M.-J. Don, P.-C. King, H.-H. Chen, Salvia miltiorrhiza Extract and Individual Synthesized Component Derivatives Induce Activating-Transcription-Factor-3-Mediated Anti-Obesity Effects and Attenuate Obesity-Induced Metabolic Disorder by Suppressing C/EBPα in High-Fat-Induced Obese Mice, Cells 11 (2022) 1022. [CrossRef]
- Y.-B. Wu, Z.-Y. Ni, Q.-W. Shi, M. Dong, H. Kiyota, Y.-C. Gu, B. Cong, Constituents from Salvia Species and Their Biological Activities, Chem. Rev. 112 (2012) 5967–6026. [CrossRef]
- M.D. Gkioni, K. Zeliou, V.D. Dimaki, P. Trigas, F.N. Lamari, GC-MS and LC-DAD-MS Phytochemical Profiling for Characterization of Three Native Salvia Taxa from Eastern Mediterranean with Antiglycation Properties, Molecules 28 (2022) 93. [CrossRef]
- M. Dawra, J. Bouajila, M. El Beyrouthy, A. Abi Rizk, P. Taillandier, N. Nehme, Y. El Rayess, Chemical Characterization and Antioxidant, Antibacterial, Antiacetylcholinesterase and Antiproliferation Properties of Salvia fruticosa Miller Extracts, Molecules 28 (2023) 2429. [CrossRef]
- R. Boukhary, M. Aboul-Ela, O. Al-Hanbali, A. El-Lakany, Chemical Constituents from Salvia fruticosa libanotica, PJ 10 (2017) 45–48. [CrossRef]
- M. Villalva, L. Jaime, E. Aguado, J.A. Nieto, G. Reglero, S. Santoyo, Anti-Inflammatory and Antioxidant Activities from the Basolateral Fraction of Caco-2 Cells Exposed to a Rosmarinic Acid Enriched Extract, J. Agric. Food Chem. 66 (2018) 1167–1174. [CrossRef]
- S.B. Subramanya, B. Venkataraman, S. Almarzooqi, V. Raj, V.S. Subramanian, B.A. Bhongade, 1,8-Cineole, a bioactive monoterpenoid, mitigates colon inflammation by stimulating colon PPAR-γ transcription factor, The FASEB Journal 36 (2022) fasebj.2022.36.S1.R3110. [CrossRef]
- M. Hijazi, K. Hijazi, K. Bouhadir, Z. Fatfat, M. Aboul-Ela, H. Gali-Muhtasib, A. El-Lakany, Anticancer activity of abietane diterpenoids from Salvia libanoticum grown in Lebanon, Phcog Mag 17 (2021) 127. [CrossRef]
- A. Alqudah, E.Y. Qnais, O. Gammoh, Y. Bseiso, M. Wedyan, Potential anti-inflammatory activity of the Salvia fruticosa Mill. essential oil, J Pharm Pharmacogn Res 12 (2024) 14–26. [CrossRef]
- M. Dawra, J. Bouajila, M. El Beyrouthy, A. Abi Rizk, P. Taillandier, N. Nehme, Y. El Rayess, Chemical Characterization and Antioxidant, Antibacterial, Antiacetylcholinesterase and Antiproliferation Properties of Salvia fruticosa Miller Extracts, Molecules 28 (2023) 2429. [CrossRef]
- S. Kyriakou, V. Tragkola, M. Plioukas, I. Anestopoulos, P.S. Chatzopoulou, E. Sarrou, D.T. Trafalis, M.V. Deligiorgi, R. Franco, A. Pappa, M.I. Panayiotidis, Chemical and Biological Characterization of the Anticancer Potency of Salvia fruticosa in a Model of Human Malignant Melanoma, Plants (Basel) 10 (2021) 2472. [CrossRef]
- M. Bassil, C.F. Daher, M. Mroueh, N. Zeeni, Salvia libanotica improves glycemia and serum lipid profile in rats fed a high fat diet, BMC Complement Altern Med 15 (2015) 384. [CrossRef]
- M. Dawra, N. Nehme, Y.E. Rayess, M.E. Beyrouthy, P. Taillandier, J. Bouajila, Folk medicinal applications, phytochemical composition and biological activities of some Lebanese endemic plants, South African Journal of Botany 150 (2022) 511–527. [CrossRef]
- P. Ercan, S.N. El, Bioaccessibility and inhibitory effects on digestive enzymes of carnosic acid in sage and rosemary, International Journal of Biological Macromolecules 115 (2018) 933–939. [CrossRef]
- P. Drikvandi, S. Bahramikia, M. Alirezaei, Modulation of the antioxidant defense system in liver, kidney, and pancreas tissues of alloxan-induced diabetic rats by camphor, J. Food Biochem. 44 (2020). [CrossRef]
- Y. Liang, X. Wan, F. Niu, S. Xie, H. Guo, Y. Yang, L. Guo, C. Zhou, Salvia plebeia R. Br. : an overview about its traditional uses, chemical constituents, pharmacology and modern applications, Biomedicine & Pharmacotherapy 121 (2020) 109589. [CrossRef]
- H. Kim, J.-Y. Choi, M. Hong, H.S. Suh, Traditional medicine for the treatment of common cold in Korean adults: A nationwide population-based study, Integr Med Res 10 (2021) 100458. [CrossRef]
- S. Bae, Y.-H. Lee, J. Lee, J. Park, W. Jun, Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway, Nutrients 14 (2022) 5379. [CrossRef]
- E.M. Zahran, U.R. Abdelmohsen, H.E. Khalil, S.Y. Desoukey, M.A. Fouad, M.S. Kamel, Diversity, phytochemical and medicinal potential of the genus Ocimum L. (Lamiaceae), Phytochem Rev 19 (2020) 907–953. [CrossRef]
- Y. Liang, X. Wan, F. Niu, S. Xie, H. Guo, Y. Yang, L. Guo, C. Zhou, Salvia plebeia R. Br. : an overview about its traditional uses, chemical constituents, pharmacology and modern applications, Biomedicine & Pharmacotherapy 121 (2020) 109589. [CrossRef]
- Y. Dai, Z. Ye, H. Liu, R. Zhu, L. Sun, S. Li, G. Xie, Y. Zhu, Y. Zhao, M. Qin, The chemical profiling of Salvia plebeia during different growth periods and the biosynthesis of its main flavonoids ingredients, Front. Plant Sci. 14 (2023) 1228356. [CrossRef]
- K. Patel, D.K. Patel, Medicinal importance, pharmacological activities, and analytical aspects of hispidulin: A concise report, J Tradit Complement Med 7 (2017) 360–366. [CrossRef]
- L. Fan, X. Zhang, Y. Huang, B. Zhang, W. Li, Q. Shi, Y. Lin, F. Wu, Homoplantaginin attenuates high glucose-induced vascular endothelial cell apoptosis through promoting autophagy via the AMPK / TFEB pathway, Phytotherapy Research 37 (2023) 3025–3041. [CrossRef]
- S.-I. Choi, I.-H. Cho, S.H. Han, Y.-J. Jeon, J.-G. Choi, J.-S. Kim, J.-H. Lee, Antiobesity Effects of Salvia plebeia R. Br. Extract in High-Fat Diet-Induced Obese Mice, Journal of Medicinal Food 19 (2016) 1048–1056. [CrossRef]
- H.-H. Jang, S.-Y. Cho, M.-J. Kim, J.-B. Kim, S.-H. Lee, M.-Y. Lee, Y.-M. Lee, Anti-inflammatory effects of Salvia plebeia R. Br extract in vitro and in ovalbumin-induced mouse model, Biol Res 49 (2016) 41. [CrossRef]
- H.R. Won, Effect of Salvia plebeia Water Extract on Antioxidant Activity and Lipid Composition of Rats Fed a High Fat-High Cholesterol Diet, KJCLS 27 (2016) 233–243. [CrossRef]
- O. Pereira, M. Catarino, A. Afonso, A. Silva, S. Cardoso, Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes, Molecules 23 (2018) 3169. [CrossRef]
- H.-J. Eom, Y.Y. Jeong, N.R. Kwon, K.H. Kim, E. Yeon, H.-S. Yoon, Y. Ryu, I.J. Kim, Nutritional Components and Physiological Activity of 4 Wild Vegetables (Salvia plebeia R. Br, Angelica acutiloba, Gynura procumbens and Saururus chinensis Baill) Cultivated in Chungbuk Province, The Korean Journal of Food And Nutrition 34 (2021) 398–406. [CrossRef]
- M.A. Senchukova, Microbiota of the gastrointestinal tract: Friend or foe?, World J Gastroenterol 29 (2023) 19–42. [CrossRef]
- A. Andoh, A. Nishida, Alteration of the Gut Microbiome in Inflammatory Bowel Disease, Digestion 104 (2023) 16–23. [CrossRef]
- S. Fujisaka, Y. Watanabe, K. Tobe, The gut microbiome: a core regulator of metabolism, Journal of Endocrinology 256 (2023) e220111. [CrossRef]
- P. Portincasa, L. Bonfrate, M. Vacca, M. De Angelis, I. Farella, E. Lanza, M. Khalil, D.Q.-H. Wang, M. Sperandio, A. Di Ciaula, Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis, IJMS 23 (2022) 1105. [CrossRef]
- B. Madhogaria, P. Bhowmik, A. Kundu, Correlation between human gut microbiome and diseases, Infectious Medicine 1 (2022) 180–191. [CrossRef]
- S.H. Zyoud, M. Shakhshir, A.S. Abushanab, A. Koni, M. Shahwan, A.A. Jairoun, S.W. Al-Jabi, Global research trends on the links between insulin resistance and obesity: a visualization analysis, Transl Med Commun 7 (2022) 18. [CrossRef]
- N. Redondo-Useros, E. Nova, N. González-Zancada, L.E. Díaz, S. Gómez-Martínez, A. Marcos, Microbiota and Lifestyle: A Special Focus on Diet, Nutrients 12 (2020) 1776. [CrossRef]
- T. Hrncir, Gut Microbiota Dysbiosis: Triggers, Consequences, Diagnostic and Therapeutic Options, Microorganisms 10 (2022) 578. [CrossRef]
- M. De Siena, P. Raoul, L. Costantini, E. Scarpellini, M. Cintoni, A. Gasbarrini, E. Rinninella, M.C. Mele, Food Emulsifiers and Metabolic Syndrome: The Role of the Gut Microbiota, Foods 11 (2022) 2205. [CrossRef]
- B.S. Almugadam, Y. Liu, S.-M. Chen, C.-H. Wang, C.-Y. Shao, B.-W. Ren, L. Tang, Alterations of Gut Microbiota in Type 2 Diabetes Individuals and the Confounding Effect of Antidiabetic Agents, J Diabetes Res 2020 (2020) 7253978. [CrossRef]
- D.K. Wiredu Ocansey, S. Hang, X. Yuan, H. Qian, M. Zhou, C. Valerie Olovo, X. Zhang, F. Mao, The diagnostic and prognostic potential of gut bacteria in inflammatory bowel disease, Gut Microbes 15 (2023) 2176118. [CrossRef]
- V. Liakina, S. Strainiene, I. Stundiene, V. Maksimaityte, E. Kazenaite, Gut microbiota contribution to hepatocellular carcinoma manifestation in non-alcoholic steatohepatitis, World J Hepatol 14 (2022) 1277–1290. [CrossRef]
- L. Gao, W. Peng, H. Xue, Y. Wu, H. Zhou, P. Jia, Y. Wang, Spatial–temporal trends in global childhood overweight and obesity from 1975 to 2030: a weight mean center and projection analysis of 191 countries, Global Health 19 (2023) 53. [CrossRef]
- K.W. Tham, R. Abdul Ghani, S.C. Cua, C. Deerochanawong, M. Fojas, S. Hocking, J. Lee, T.Q. Nam, F. Pathan, B. Saboo, S. Soegondo, N. Somasundaram, A.M.L. Yong, J. Ashkenas, N. Webster, B. Oldfield, Obesity in South and Southeast Asia—A new consensus on care and management, Obesity Reviews 24 (2023) e13520. [CrossRef]
- R. Williams, M. Periasamy, Genetic and Environmental Factors Contributing to Visceral Adiposity in Asian Populations, Endocrinol Metab 35 (2020) 681–695. [CrossRef]
- J. Tokarek, J. Gadzinowska, E. Młynarska, B. Franczyk, J. Rysz, What Is the Role of Gut Microbiota in Obesity Prevalence? A Few Words about Gut Microbiota and Its Association with Obesity and Related Diseases, Microorganisms 10 (2021) 52. [CrossRef]
- Z. Xu, W. Jiang, W. Huang, Y. Lin, F.K.L. Chan, S.C. Ng, Gut microbiota in patients with obesity and metabolic disorders - a systematic review, Genes Nutr 17 (2022) 2. [CrossRef]
- T.V. Kirichenko, Y.V. Markina, A.I. Bogatyreva, T.V. Tolstik, Y.R. Varaeva, A.V. Starodubova, The Role of Adipokines in Inflammatory Mechanisms of Obesity, Int J Mol Sci 23 (2022) 14982. [CrossRef]
- M. Patel, D. Fowler, J. Sizer, C. Walton, Faecal volatile biomarkers of Clostridium difficile infection, PLoS ONE 14 (2019) e0215256. [CrossRef]
- K.A. Krautkramer, J. Fan, F. Bäckhed, Gut microbial metabolites as multi-kingdom intermediates, Nat Rev Microbiol 19 (2021) 77–94. [CrossRef]
- Hernández, Canfora, Jocken, Blaak, The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity, Nutrients 11 (2019) 1943. [CrossRef]
- Z.C. Holmes, J.D. Silverman, H.K. Dressman, Z. Wei, E.P. Dallow, S.C. Armstrong, P.C. Seed, J.F. Rawls, L.A. David, Short-Chain Fatty Acid Production by Gut Microbiota from Children with Obesity Differs According to Prebiotic Choice and Bacterial Community Composition, mBio 11 (2020) e00914-20. [CrossRef]
- O. Lange, M. Proczko-Stepaniak, A. Mika, Short-Chain Fatty Acids—A Product of the Microbiome and Its Participation in Two-Way Communication on the Microbiome-Host Mammal Line, Curr Obes Rep 12 (2023) 108–126. [CrossRef]
- Y. Wu, H. Xu, X. Tu, Z. Gao, The Role of Short-Chain Fatty Acids of Gut Microbiota Origin in Hypertension, Front. Microbiol. 12 (2021) 730809. [CrossRef]
- S. Heinonen, R. Jokinen, A. Rissanen, K.H. Pietiläinen, White adipose tissue mitochondrial metabolism in health and in obesity, Obesity Reviews 21 (2020) e12958. [CrossRef]
- D. Wu, H. Wang, L. Xie, F. Hu, Cross-Talk Between Gut Microbiota and Adipose Tissues in Obesity and Related Metabolic Diseases, Front. Endocrinol. 13 (2022) 908868. [CrossRef]
- S.-H. Cho, Y.-J. Cho, J.-H. Park, The human symbiont Bacteroides thetaiotaomicron promotes diet-induced obesity by regulating host lipid metabolism, J Microbiol. 60 (2022) 118–127. [CrossRef]
- M. Kumar, R. Nagpal, R. Kumar, R. Hemalatha, V. Verma, A. Kumar, C. Chakraborty, B. Singh, F. Marotta, S. Jain, H. Yadav, Corrigendum to “Cholesterol-Lowering Probiotics as Potential Biotherapeutics for Metabolic Diseases,” Journal of Diabetes Research 2022 (2022) 1–14. [CrossRef]
- A. Kriaa, M. Bourgin, A. Potiron, H. Mkaouar, A. Jablaoui, P. Gérard, E. Maguin, M. Rhimi, Microbial impact on cholesterol and bile acid metabolism: current status and future prospects, J Lipid Res 60 (2019) 323–332. [CrossRef]
- L.V. Gromova, S.O. Fetissov, A.A. Gruzdkov, Mechanisms of Glucose Absorption in the Small Intestine in Health and Metabolic Diseases and Their Role in Appetite Regulation, Nutrients 13 (2021) 2474. [CrossRef]
- P. Strandwitz, Neurotransmitter modulation by the gut microbiota, Brain Research 1693 (2018) 128–133. [CrossRef]
- O. Contreras-Rodriguez, M. Solanas, R.M. Escorihuela, Dissecting ultra-processed foods and drinks: Do they have a potential to impact the brain?, Rev Endocr Metab Disord 23 (2022) 697–717. [CrossRef]
- B. Van Der Hee, J.M. Wells, Microbial Regulation of Host Physiology by Short-chain Fatty Acids, Trends in Microbiology 29 (2021) 700–712. [CrossRef]
- C.B. Christiansen, S. Veedfald, B. Hartmann, A.M. Gauguin, S. Møller, T. Moritz, S. Madsbad, J.J. Holst, Colonic Lactulose Fermentation Has No Impact on Glucagon-like Peptide-1 and Peptide-YY Secretion in Healthy Young Men, The Journal of Clinical Endocrinology & Metabolism 107 (2022) 77–87. [CrossRef]
- N. Forte, A.C. Fernández-Rilo, L. Palomba, V. Di Marzo, L. Cristino, Obesity Affects the Microbiota–Gut–Brain Axis and the Regulation Thereof by Endocannabinoids and Related Mediators, IJMS 21 (2020) 1554. [CrossRef]
- A. Alharthi, S. Alhazmi, N. Alburae, A. Bahieldin, The Human Gut Microbiome as a Potential Factor in Autism Spectrum Disorder, IJMS 23 (2022) 1363. [CrossRef]
- J. Jansma, F. Brinkman, S. Van Hemert, S. El Aidy, Targeting the endocannabinoid system with microbial interventions to improve gut integrity, Progress in Neuro-Psychopharmacology and Biological Psychiatry 106 (2021) 110169. [CrossRef]
- A.B. Bansal, Y. Al Khalili, Orlistat, in: StatPearls, StatPearls Publishing, Treasure Island (FL), 2024. http://www.ncbi.nlm.nih.gov/books/NBK542202/ (accessed January 23, 2024).
- J.G. Kang, C.-Y. Park, Anti-Obesity Drugs: A Review about Their Effects and Safety, Diabetes Metab J 36 (2012) 13. [CrossRef]
- M. Chakhtoura, R. Haber, M. Ghezzawi, C. Rhayem, R. Tcheroyan, C.S. Mantzoros, Pharmacotherapy of obesity: an update on the available medications and drugs under investigation, eClinicalMedicine 58 (2023) 101882. [CrossRef]
- A. Depaoli, A. Long, G.M. Fine, M. Stewart, S. O’Rahilly, Efficacy of Metreleptin for Weight Loss in Overweight and Obese Adults with Low Leptin Levels, Diabetes 67 (2018) 296-LB. [CrossRef]
- M. Lenharo, Anti-obesity drug also protects against heart disease — what happens next?, Nature 620 (2023) 480–480. [CrossRef]
- S. Ghoneim, Tirzepatide: A New Anti-Obesity Medication, Gastroenterology 164 (2023) 159. [CrossRef]
- H.W.R. Li, K.S.L. Lam, P.C. Ho, Antiobesity drugs for obese women planning pregnancy, in: Obesity and Obstetrics, Elsevier, 2020: pp. 301–306. [CrossRef]
- C. Enright, E. Thomas, D.R. Saxon, An Updated Approach to Antiobesity Pharmacotherapy: Moving Beyond the 5% Weight Loss Goal, Journal of the Endocrine Society 7 (2023) bvac195. [CrossRef]
- E.A. Bohula, S.D. Wiviott, D.K. McGuire, S.E. Inzucchi, J. Kuder, K. Im, C.L. Fanola, A. Qamar, C. Brown, A. Budaj, A. Garcia-Castillo, M. Gupta, L.A. Leiter, N.J. Weissman, H.D. White, T. Patel, B. Francis, W. Miao, C. Perdomo, S. Dhadda, M.P. Bonaca, C.T. Ruff, A.C. Keech, S.R. Smith, M.S. Sabatine, B.M. Scirica, Cardiovascular Safety of Lorcaserin in Overweight or Obese Patients, N Engl J Med 379 (2018) 1107–1117. [CrossRef]
- I.E. Poyraz, G.A. Ciftci, N. Öztürk, Phenolic Contents, in vitro Antioxidant and Cytotoxicity Activities of Salvia aethiopis L. and S. ceratophylla L. (Lamiaceae), Records of Natural Products 11 (2017) 345–355.
- M.B. Bahadori, L. Dinparast, H. Valizadeh, M.M. Farimani, S.N. Ebrahimi, Bioactive constituents from roots of Salvia syriaca L.: Acetylcholinesterase inhibitory activity and molecular docking studies, South African Journal of Botany 106 (2016) 1–4. [CrossRef]
- J.S. Katanić Stanković, N. Srećković, D. Mišić, U. Gašić, P. Imbimbo, D.M. Monti, V. Mihailović, Bioactivity, biocompatibility and phytochemical assessment of lilac sage, Salvia verticillata L. (Lamiaceae) - A plant rich in rosmarinic acid, Industrial Crops and Products 143 (2020) 111932. [CrossRef]
- S. Stankov, H. Fidan, N. Petkova, I. Dincheva, A. Stoyanova, B.C. Senkal, H. Dogan, T. Uskutoglu, Phytochemical Composition of Salvia candidissima Vahl. ssp. occidentalis From Turkey, Journal of Essential Oil Bearing Plants 23 (2020) 710–718. [CrossRef]
- M.S. Abu-Darwish, C. Cabral, Z. Ali, M. Wang, S.I. Khan, M.R. Jacob, S.K. Jain, B.L. Tekwani, F. Zulfiqar, I.A. Khan, H. Taifour, L. Salgueiro, T. Efferth, Salvia ceratophylla L. from South of Jordan: new insights on chemical composition and biological activities, Nat. Prod. Bioprospect. 10 (2020) 307–316. [CrossRef]
- İ. Emre, M. Kurşat, S. Kirbag, P. Erecevi̇T Sönmez, M.Y. Emre, Prof.Dr.Ö. Yilmaz, Ş. Ci̇Velek, The Antioxidant and Antimicrobial Capacities of Phenolic Profiles of Some Salvia L. Seeds Grown in Turkey, International Journal of Secondary Metabolite 8 (2021) 20–30. [CrossRef]
- H. Bardakci, E. Celep, T. Gözet, I. Kurt-Celep, I. Deniz, B. Şen-Utsukarci, G. Akaydin, A comparative investigation on phenolic composition, antioxidant and antimicrobial potentials of Salvia heldreichiana Boiss. ex Bentham extracts, South African Journal of Botany 125 (2019) 72–80. [CrossRef]
- S. Guzel, M. Ulger, A. Kahraman, Chemical Composition and Some Biological Activities of Salvia longipedicellata Hedge Mericarps, Chem Nat Compd 56 (2020) 788–792. [CrossRef]
- S. Bechkri, A. Alabdul Magid, L. Voutquenne-Nazabadioko, D. Berrehal, A. Kabouche, M. Lehbili, H. Lakhal, A. Abedini, S.C. Gangloff, H. Morjani, Z. Kabouche, Triterpenes from Salvia argentea var. aurasiaca and their antibacterial and cytotoxic activities, Fitoterapia 139 (2019) 104296. [CrossRef]
- M. Doğan, N. Akıcı, M.E. Diken, S. Doğan, B. Yilmaz Kardas, T. Dirmenci, Biological activities of some Salvia species, Zeitschrift Für Naturforschung C 77 (2022) 133–143. [CrossRef]
- M. Shanaida, N. Hudz, M. Białoń, M. Kryvtsowa, L. Svydenko, A. Filipska, P. Paweł Wieczorek, Chromatographic profiles and antimicrobial activity of the essential oils obtained from some species and cultivars of the Mentheae tribe (Lamiaceae), Saudi Journal of Biological Sciences 28 (2021) 6145–6152. [CrossRef]
- M. Yılar, Y. Bayar, A. Bayar, N. Genc, Chemical composition of the essential oil of Salvia bracteata banks and the biological activity of its extracts: Antioxidant, total phenolic, total flavonoid, antifungal and allelopathic effects, Bot Serb 44 (2020) 71–79. [CrossRef]
- G. Zengin, E.J. Llorent-Martínez, M.L.F. Córdova, M.B. Bahadori, A. Mocan, M. Locatelli, A. Aktumsek, Chemical composition and biological activities of extracts from three Salvia species: S. blepharochlaena, S. euphratica var. leiocalycina, and S. verticillata subsp. amasiaca, Industrial Crops and Products 111 (2018) 11–21. [CrossRef]
- M. Akdeniz, I. Yener, M.A. Yilmaz, S. Irtegun Kandemir, F. Tekin, A. Ertas, A potential species for cosmetic and pharmaceutical industries: Insight to chemical and biological investigation of naturally grown and cultivated Salvia multicaulis Vahl, Industrial Crops and Products 168 (2021) 113566. [CrossRef]
- S.M. Talebi, M. Askary, N. Khalili, A. Matsyura, M. Ghorbanpour, K. Kariman, Genetic structure and essential oil composition in wild populations of Salvia multicaulis Vahl., Biochemical Systematics and Ecology 96 (2021) 104269. [CrossRef]
- G.G. Toplan, M. Kürkçüoğlu, F. Göger, T. Taşkın, A. Civaş, G. İşcan, G. Ecevit-Genç, A. Mat, K.H.C. Başer, Phytochemical screening and biological evaluation of Salvia hydrangea DC. ex Benth. growing in eastern Anatolia, South African Journal of Botany 147 (2022) 799–807. [CrossRef]
- M. Ben Farhat, J.A. Sotomayor, M.J. Jordán, Salvia verbenaca L. essential oil: Variation of yield and composition according to collection site and phenophase, Biochemical Systematics and Ecology 82 (2019) 35–43. [CrossRef]
- F. Noorbakhsh, S. Zare, O. Firuzi, A. Sakhteman, J.N. Chandran, B. Schneider, A.R. Jassbi, Phytochemical Analysis and Biological Activity of Salvia compressa Vent., Iran J Pharm Res 21 (2022). [CrossRef]
- A.F. Afonso, O.R. Pereira, Â.S.F. Fernandes, R.C. Calhelha, A.M.S. Silva, I.C.F.R. Ferreira, S.M. Cardoso, The Health-Benefits and Phytochemical Profile of Salvia apiana and Salvia farinacea var. Victoria Blue Decoctions, Antioxidants 8 (2019) 241. [CrossRef]
- A. Mocan, M. Babotă, A. Pop, I. Fizeșan, A. Diuzheva, M. Locatelli, S. Carradori, C. Campestre, L. Menghini, C.R. Sisea, M. Sokovic, G. Zengin, R. Păltinean, S. Bădărău, D. C. Vodnar, G. Crișan, Chemical Constituents and Biologic Activities of Sage Species: A Comparison between Salvia officinalis L., S. glutinosa L. and S. transsylvanica (Schur ex Griseb. & Schenk) Schur, Antioxidants 9 (2020) 480. [CrossRef]
- İ. Gülçin, A.Z. Tel, A.C. Gören, P. Taslimi, S.H. Alwasel, Sage (Salvia pilifera): determination of its polyphenol contents, anticholinergic, antidiabetic and antioxidant activities, Food Measure 13 (2019) 2062–2074. [CrossRef]
- Etsassala, Badmus, Waryo, Marnewick, Cupido, Hussein, Iwuoha, Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities of Novel Abietane Diterpenes from Salvia africana-lutea, Antioxidants 8 (2019) 421. [CrossRef]
- N.G.E.R. Etsassala, J.A. Badmus, J.L. Marnewick, E.I. Iwuoha, F. Nchu, A.A. Hussein, Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities, Molecular Docking, and Antioxidant Capacities of Salvia aurita Constituents, Antioxidants 9 (2020) 1149. [CrossRef]
- M.S. Kocak, C. Sarikurkcu, M. Cengiz, S. Kocak, M.C. Uren, B. Tepe, Salvia cadmica: Phenolic composition and biological activity, Industrial Crops and Products 85 (2016) 204–212. [CrossRef]
- G. Zengin, E.J. Llorent-Martínez, M.L.F. Córdova, M.B. Bahadori, A. Mocan, M. Locatelli, A. Aktumsek, Chemical composition and biological activities of extracts from three Salvia species: S. blepharochlaena, S. euphratica var. leiocalycina, and S. verticillata subsp. amasiaca, Industrial Crops and Products 111 (2018) 11–21. [CrossRef]
- G. Zengin, E.J. Llorent-Martínez, M.L.F. Córdova, M.B. Bahadori, A. Mocan, M. Locatelli, A. Aktumsek, Chemical composition and biological activities of extracts from three Salvia species: S. blepharochlaena, S. euphratica var. leiocalycina, and S. verticillata subsp. amasiaca, Industrial Crops and Products 111 (2018) 11–21. [CrossRef]
- O. Pereira, M. Catarino, A. Afonso, A. Silva, S. Cardoso, Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes, Molecules 23 (2018) 3169. [CrossRef]
- M.B. Bahadori, B. Asghari, L. Dinparast, G. Zengin, C. Sarikurkcu, M. Abbas-Mohammadi, S. Bahadori, Salvia nemorosa L.: A novel source of bioactive agents with functional connections, LWT 75 (2017) 42–50. [CrossRef]
- S. Arabiyat, A. Al-Rabi’ee, H. Zalloum, M. Hudaib, M. Mohammad, Y. Bustanji, Antilipolytic and hypotriglyceridemic effects of dietary Salvia triloba Lf (Lamiaceae) in experimental rats, Trop. J. Pharm Res 15 (2016) 723. [CrossRef]
- B. Abd El-Motelp, H. Mohamed, Hanan. Abd El-Latief, M. Hassen, In Vivo Salvia officinalis Extract and Orlistat Provides a Multimechanistic Strategy for Preventing Obesity, Egyptian Academic Journal of Biological Sciences, B. Zoology 15 (2023) 83–104. [CrossRef]
- G. Maturana, J. Segovia, C. Olea-Azar, E. Uribe-Oporto, A. Espinosa, M.C. Zúñiga-López, Evaluation of the Effects of Chia (Salvia hispanica L.) Leaves Ethanolic Extracts Supplementation on Biochemical and Hepatic Markers on Diet-Induced Obese Mice, Antioxidants 12 (2023) 1108. [CrossRef]
- S. Belhadj, O. Hentati, M. Hammami, A. Ben Hadj, T. Boudawara, M. Dammak, S. Zouari, A. El Feki, Metabolic impairments and tissue disorders in alloxan-induced diabetic rats are alleviated by Salvia officinalis L. essential oil, Biomedicine & Pharmacotherapy 108 (2018) 985–995. [CrossRef]
- F. Remok, S. Saidi, A.A. Gourich, K. Zibouh, M. Maouloua, F.E. Makhoukhi, N.E. Menyiy, H. Touijer, M. Bouhrim, S. Sahpaz, A.M. Salamatullah, M. Bourhia, T. Zair, Phenolic Content, Antioxidant, Antibacterial, Antihyperglycemic, and α-Amylase Inhibitory Activities of Aqueous Extract of Salvia lavandulifolia Vahl, Pharmaceuticals 16 (2023) 395. [CrossRef]
- V. Vuksan, A.L. Jenkins, C. Brissette, L. Choleva, E. Jovanovski, A.L. Gibbs, R.P. Bazinet, F. Au-Yeung, A. Zurbau, H.V.T. Ho, L. Duvnjak, J.L. Sievenpiper, R.G. Josse, A. Hanna, Salba-chia (Salvia hispanica L.) in the treatment of overweight and obese patients with type 2 diabetes: A double-blind randomized controlled trial, Nutrition, Metabolism and Cardiovascular Diseases 27 (2017) 138–146. [CrossRef]
Figure 1.
A diagrammatical representation of gut microbiota’s regulatory control over obesity.
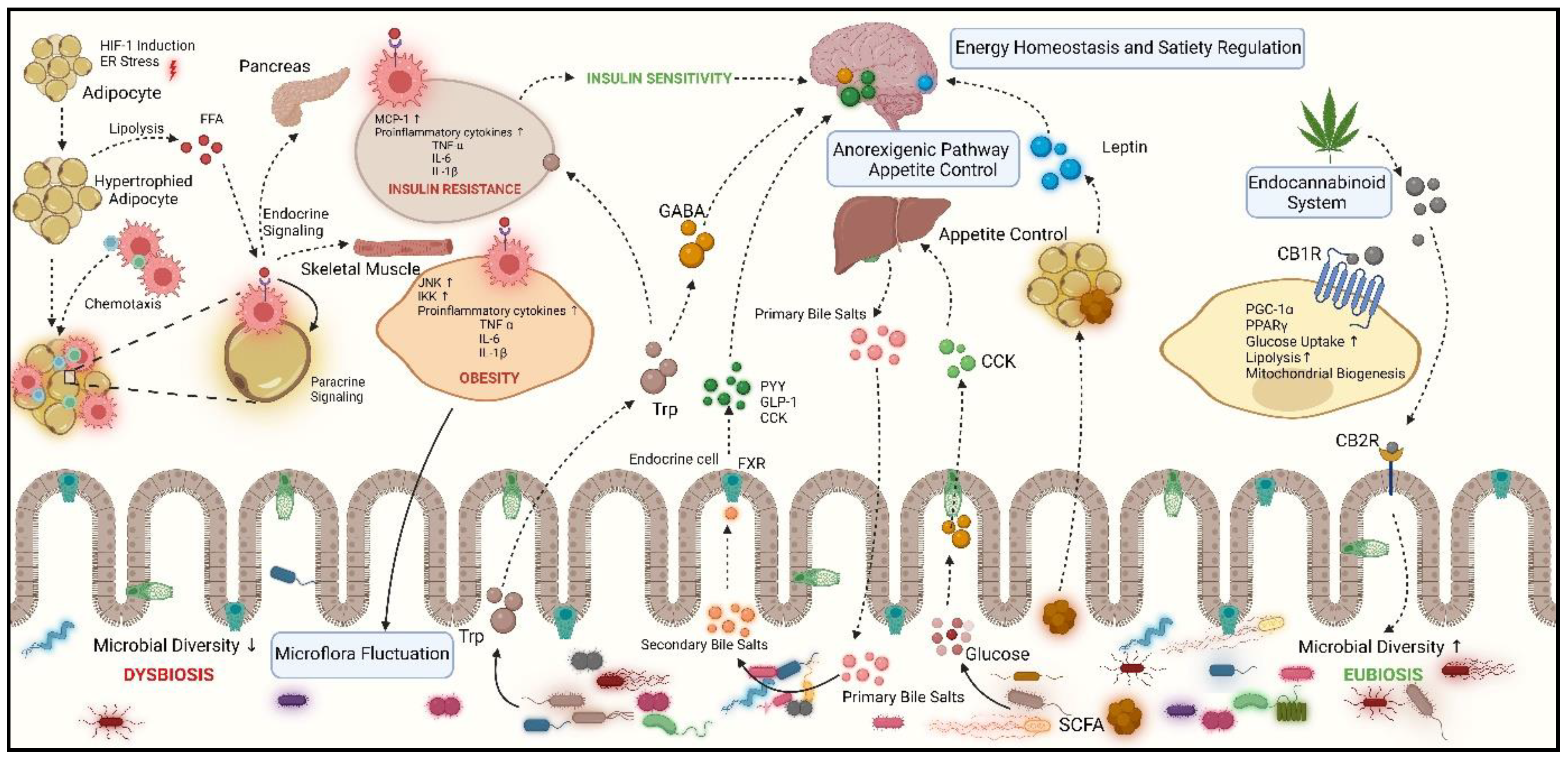
Table 1.
Market available drugs against obesity and their side-effects.
| DRUG | MECHANISM OF ACTION | SIDE EFFECTS | REFERENCES |
|---|---|---|---|
|
Orlistat (Xenical, Alli) |
Inhibits fat absorption in the digestive tract: inhibiting gastric and pancreatic lipases | Steatorrhea, diarrhoea, and anal fissures, hepatotoxicity, acute kidney injury, osteoporosis, colorectal cancer |
[129] |
|
Phentermine (Qsymia) |
Appetite suppressant | Insomnia, dry mouth, dizziness, palpitation, hand tremor, and elevation in blood pressure and pulse rate | [130] |
| Setmelanotide (Imcivree) | melanocortin-4 (MC4) receptor agonist | Increased heart rate, emotional and sleep issues, memory loss, metabolic acidosis, paraesthesia, and dry mouth | [131] |
|
Metreleptin (Myalept) |
leptin analogue | Anxiety, blurred vision, body aches burning, itching, numbness, chills/fever, dizziness, increased hunger and sore throat | [131,132] |
|
Semaglutide (Wegovy) |
GLP-1 analogue | Abdominal pain, constipation, diarrhoea, vomiting, and headache | [133] |
| Tirzepatide (Mounjaro®) | GLP-1 and GIP receptor agonist | Vomiting, diarrhoea, nausea, dyspepsia, reduced appetite, and stomach discomfort. | [134] |
|
Liraglutide (Saxenda and Victoza) |
Mimics glucagon-like peptide-1 (GLP-1 agonist) | Elevated pulse rate, hypoglycemia, diarrhoea, constipation, nausea, vomiting, and headache | [145] |
| Naltrexone/Bupropion (Contrave®) | Modulates brain pathways related to appetite, reuptake inhibitor/ opioid antagonist | Nausea, headache, and constipation, insomnia, dizziness, or dry mouth | [136] |
| Lorcaserin | Activates serotonin receptors in the brain | Headache, dizziness, nausea | [137] |
Table 2.
Pharmacological potential of various bioactive components derived from Salvia spp.
| SPECIES | NATIVE AREA | EXTRACTS AND PLANT PART | MAJOR BIOACTIVE COMPONENTS | THERAPEUTIC POTENTIAL | REFERENCES |
|---|---|---|---|---|---|
| S. aethiopis | Turkey | Methanol, ethyl acetate extracts |
Rosmarinic acid |
Anti-oxidant and Cytotoxic | [138] |
|
S. syriaca L. |
Urmia, West Azarbaijan province | Acetone extract of powdered roots | ursolic acid corosolic acid β-sitosterol urs-12-en-2α,3β-diol daucosterol (1–5) β-sitosterol daucosterol |
Antioxidant, Anticholinesterase inhibition | [139] |
| S. verticillata | Western Serbia | Methanol extract from aerial parts | Rosmarinic acid, Caffeic acid, Salvianolic acid, Yunnaneic acids, Flavonoids |
Antimicrobial, Antibacterial, Antifungal, Antioxidant, Anti-inflammatory |
[140] |
|
S. candidissima subsp. occidentalis |
Turkey | Ethanol extract | Sclareol, Spathulenol, Caryophyllene oxide |
Antiviral, Cytotoxic activity, Antioxidant |
[141] |
| S. ceratophylla | South of Jordan | Methanol extract from aerial parts | Linalool, Germacrene-D, Bicyclogermacrene, Bicyclic diterpene- alcoholsclareol |
Antiviral, Cytotoxic activity, Anti-inflammatory, Antiprotozoal activitiy |
[142] |
|
S. frigida Boiss., S. candidissima subsp. candidissima Vahl., S. virgata Jacq., S. verticillata L. var. verticillate, S. russellii Benth |
Turkey | Methanol, Acetonitrile, Water extract from plant seeds |
Catechin, Rosmarinic acid, vanilic acid |
Antiviral, Cytotoxic activity, Antimicrobial, Antioxidant |
[143] |
| S. heldreichiana | Turkey | Methanol, Chloroform, ethyl acetate from Aerial parts | p-coumaric acid, caffeic acid, chlorogenic acid, rosmarinic acid, apigenin, apigenin-7-O-glucoside, vitexin, isoorientin, luteolin-7-O-glucoside, hyperoside | Antioxidant, Antimicrobial | [144] |
| S. longipedicellata | East Anatolia (Turkey) | Ethanol, Hexane extracts from mericarps |
α-linolenic acid, β-sitosterol, γ-tocopherol, potassium |
Antioxidant, Antimicrobial | [145] |
| S. argentea | Mediterranean region | Acetone extract from aerial parts | 1β,3β-dihydroxyurs-9(11)-12-diene, 3β-hydroxy-urs-9(11),12-diene, 1β,3β,15α,28-tetrahydroxyurs-12-en, 1β,3β,11α,15α-tetrahydroxy-urs-12-ene, 3β-hydroxy-olean9(11),12-diene |
Antibacterial, Cytotoxic activity | [146] |
|
S. macrochlamys S. kronenburgii, S. euphratica Montbret, S. huberi, S. kurdica |
Turkey | rutin hydrate, luteolin-7-glucoside, Vanillic acid |
Antioxidant, Antibacterial, Cytotoxic activity | [147] | |
|
S. sclarea |
Italy, Ukraine | Aerial, Fresh raw, Leaves |
Linalol, linaliyl acetate, germacrene D, α-terpineol |
Anti-inflammatory | [148] |
| S. bracteata | Province of Kırşehir, Turkey | Methanol, Ethyl acetate, Hexane extracts, Essentials oil, from flowers, shoots, leaves |
ledol, camphor, valencene |
Antioxidant, Antifungal | [149] |
| S. blepharochlaena | Central Türkiye | Dichloromethane, methanol, water extracts |
Rosmarinic acid | Antioxidant, Cytotoxic activity | [150] |
| S. multicaulis | Iran | Essential oil and ethanol extracts from leaf, roots, flower, |
α-pinene, camphene, 1,8-cineole, Camphor, Caryophllene, rosmarinic acid |
Anti-inflammatory, Antimicrobial, Antioxidant Analgesic effects | [151] [152] |
| S. hydrangea | Eastern region of Turkey | Water, n-hexane, chloroform, methanol from Aerial parts |
Camphor, 1,8-cineole, Camphene, Limonene, β-pinene, α-pinene, rosmarinic acid, luteolin glycoside |
Antioxidant, Antimicrobial, Anti-inflammatory, Antispasmodic, Aypolipidemic |
[153] |
| S. verbenaca | Tunisia | Aerial | Viridiflorol, α-pinene, β-caryophyllene, p-cymene |
Antibacterial, Anticancer, Antioxidant, Antidiabetic | [154] |
| S. compressa | Iran | Dichloromethane extract from shoot extract | Citrostadienol, β-sitosterol, two glyceride esters of linolenic, linoleic, and palmitic acids geraniol |
Cytotoxic, Antibacterial |
[155] |
|
S. apiana S. farinacea var. Victoria Blue |
Portugal | Aqueous extract from Aerial parts | Rosmanol, Hydroxycarnosic acid, derivative of sageone |
Antioxidant Anti-inflammatory Antimicrobial Cytotoxic |
[156] |
|
S. transsylvanica S. glutinosa S. officinalis |
Schenk Romania |
Ethanol extract from aerial parts | Rutin, Catechin, Epicatechin kaempferol, Apigenin, Quercetin derivatives, Caffeic acid, p-coumaric acid, naringenin |
Antimicrobial, Antifungal, Cytotoxic | [157] |
| S. pilifera | Turkey | Methanol, Water extract from aerial parts |
Salvigenin quercetagetin-3,6-dimethylether, caffeic acid, epicatechin, ferulic acid, fumaric acid |
Anticholinergic, Antidiabetic, Antioxidant | [158] |
Table 3.
In-vitro studies on anti-obesity activity of Salvia species .
| Salvia species | Experimental plant extracts–Extract form / Constituents, dose | Phytochemicals (PY) isolated and Experimental outcome (EO) | Reference |
|---|---|---|---|
| S. africana-lutea | MeOH extract 20 μL |
PY: Oleanolic (OlA) Ursolic acids 11,12-dehydroursolic acid lactone β-amyrin EO: Inhibition of α-glucosidases Inhibition of α-Amlyase |
[159] |
| S. aurita | MeOH extracts 20 μL |
PY: 4,7-dimethylapigenin ether 7-methoxyrosmanol 12-methoxycarnosic acid Rosmanol Acarbose EO: Inhibition of α-glucosidases Inhibition of α-Amlyase |
[160] |
| S. cadmica |
Aqueous extract (H2O) Methanol extract (MeOH) ethyl acetate extract (EA) |
Inhibition of α-glucosidases: MeOH > EA > H2O Inhibition of α-Amlyase MeOH > EA > H2O |
[161] |
| S. blepharochlaena | Aqueous extract (H2O) Dichloromethane extract (DCM), Methanol (MeOH) extract |
Inhibition of α-Amlyase: DCM > MeOH > H2O Inhibition of α-glucosidase : MeOH > H2O > DCM Inhibition of Lipase: DCM > MeOH > H2O |
[162] |
| S. euphratica var. leiocalycina | Aqueous extract (H2O) Dichloromethane extract (DCM), Methanol (MeOH) extract |
Inhibition of α-glucosidase:- H2O > MeOH > DCM Inhibition of α-Amlyase DCM > MeOH > H2O Inhibition of Lipase: DCM > MeOH |
[163] |
| S. greggii | Aqueous (H2O) extract |
PY: Luteolin-C-hexoside Apigenin -C-exoside EO: Inhibited α-glucosidase Inhibited α-Amlyase Inhibited Lipase |
[164] |
| S. nemorosa |
n-hexane extract (n-hex) Dichloromethane extract (DCM) Methanol (MeOH) extract |
PY: Caffeic acid Quercetin EO: Inhibition of α-glucosidase: n-hex > DCM > MeOH |
[165] |
| S. officinalis | Aqueous (H2O) extract |
PY: Apigenin-O-glucuronide Rosmarinic acid Scutellarein-O- glucuronide EO: Inhibited α-glucosidase Inhibited lipase |
[164] |
| S. hispanica(Chia) | organic solvent (Hexane extract, Ethyl acetate extract, Methanol extract, Water extract) | AgNPS + methanolic extract showed lipase inhibition ↓ Cholesterol levels |
[47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



