
Submitted:
08 August 2024
Posted:
20 August 2024
You are already at the latest version
Abstract
Phytoplankton under the ice of seasonally ice-covered lakes is a key indicator species for maintaining the health of lake ecosystems. However, there is a lack of a unified definition of cold lakes internationally. In addition, there are few studies on the distribution, driving mechanism and simulation prediction of phytoplankton under the ice, which have limited the development of winter limnology and the prevention and control of spring algal blooms. The average temperature of the coldest month, the duration of the ice period and the average annual water temperature were the main indicators for defining cold lakes based on the literatures review. The cross-action of subglacial hydrodynamics and biogeochemistry during the ice period and freeze-thaw period is the key to promoting the phytoplankton growth. The migration and transformation of nutrients and the formation of hypoxic zones of subglacial were driven by physical factors such as available light and water temperature. Cyanobacteria got advantages in the ice period and above the water column through their own gas vesicles and strong adaptation to low temperature and low oxygen. Diatoms are able to multiply rapidly with the increasing of available light and the water temperature in spring. However, the migration and transformation of nutrients induced by physical factors during the freezing-freezing-thawing process, the role of microorganisms in biogeochemical reactions, and the extent and direction of the combined effects of multiple environmental factors on phytoplankton functional groups are still unclear. At present, lake algal blooms have gradually expanded to higher latitudes under the climate change. How to accurately predict the development and response mechanism of algal blooms in cold lakes under the climate change has become a key direction in the development of winter limnology. This review explains the unique physical and chemical processes of phytoplankton growth competition in cold lakes from the perspectives of bibliometrics, mechanism analysis, method construction and future prospects. These findings would provide new insights into the field of winter limnology and provide theoretical support for the future management of spring algal blooms in cold lakes.
Keywords:
phytoplankton growth and competition
; algal blooms
; subglacial hydrodynamics
; climate change
; seasonally ice-covered lakes
1. Introduction
68% of lakes worldwide are experiencing an increasing trend in the scale, frequency, and duration of algal blooms, which seriously serious threat to the health and sustainability of freshwater ecosystems [1]. Algal blooms, especially cyanobacterial blooms, not only reduced the survival space of other aquatic species by competing for dissolved oxygen, nutrients, and available light in the lake, but also produced harmful algal toxins that harm terrestrial organisms and humans [2,3]. In addition, the economic value of surface freshwater will also be reduced due to the turbidity and odorous gases caused by algal blooms [4,5]. Therefore, preventing, controlling and managing algal blooms in lakes is a key and challenge for aquatic ecological management.
The water temperature and nutrient concentration of lakes continue to rise with climate warming and intensified human activities, leading to eutrophication of lakes and the expansion of algal blooms to mid to high latitudes [6]. Relative to mid-low latitude lakes, there was evidence that algal blooms have been found in lakes with low water temperatures, especially cyanobacterial blooms [7]. Cyanobacteria have stronger adaptability to low temperatures than other algae species, which also causesd cyanobacterial blooms to be more frequently in lakes with low water temperatures than other algae [8,9]. Therefore, cyanobacterial blooms may also like cold water based on the area, frequency, physiological shape, and abiotic factors of cyanobacterial blooms [8,9,10,11]. Cyanobacterial blooms were defined as the accumulation of cyanobacteria in the water column or edges, resulting discoloration of the water body or a maximum chlorophyll-a concentrations in the thermocline [36]. Relatively, the definition of cyanobacterial blooms in cold lakes is not clear. Recently, cyanobacterial blooms in cold lakes have been defined as those that occur in lakes with water temperatures below 15°C [12]. According to the above definition, cold lakes included not only mid-high latitude lakes, but also included high-altitude lakes. However, there were few studies on the mechanisms of algal blooms in cold lakes, which have limited the effective prevention and control of algal blooms
The unique water environment with low water temperature, subglacial hydrodynamics, and biogeochemical processes for the survival of phytoplankton in cold lakes was created. [13,14]. However, there was a lack of systematic research on the impact of various environmental factors on the growth of phytoplankton under ice, which made it difficult to support a deeper understanding of the mechanisms of phytoplankton succession [14]. Winter limnology originated in 1963, when the study of phytoplankton ecology in ice-covered lakes was proposed by Wright’s [15]. At present, the nutrient migration pathways and transformation mechanisms of cold lakes during the freezing period, as well as their understanding of lake ecological functions, are limited due to the lack of research data [16]. Since 2016, winter limnology has been developing rapidly abroad. Lake ice dynamics, subglacial biogeochemical reaction processes and subglacial hydrothermal evolution has accelerated the understanding of subglacial environmental factors on phytoplankton [17]. However, the contribution of these processes such as freezing, ice melting, and snowmelt to water circulation, phytoplankton competition, and algal blooms in cold lakes is not yet clear. The biogeochemical processes under the variations in hydrothermal structure of cold lakes have become more complex and variable with the intensification of climate warming [18]. Therefore, in order to enrich the theoretical basis of winter limnology, it is urgent to reveal the definition and mechanisms of algal blooms in cold lakes, and aim to propose adaptive strategies for future climate change [19,20].
Our study summarized domestic and foreign papers of phytoplankton in cold lakes in the past 60 years, which conducted a more detailed analysis related to phytoplankton in ice periods from 1963 to 2023 in the Web of science (WoS) database. Clarifying the development history and current research hotspots based on the visual analysis. Sort out the definition of cold lakes. The focus was on revealing the effects of subglacial fluid dynamics and biogeochemistry on the phytoplankton growth during freezing periods, as well as their responsed to climate change. In view of the problem that current winter limnology research was not systematic, emerging methods for future winter limnology have been proposed to enrich the development of winter limnology theory and control the algal blooms in cold lakes.
2. Data and Hot Spot Analysis
CiteSpace was used to analyze historical documents on the impact of lake subglacial water environment on phytoplankton growth and competition from 1963 to 2023. Research hot spots and trends will be further revealed in our study. The advantage of CiteSpace is the in-depth exploration of literature through mathematical and statistical methods, as well as the visualization of data structures and hotspot relationships [21]. This study mainly used time slicing technology to construct a time-varying model of time series and integrated a single network into an overview network. In addition, this study achieved document visualization through dynamic time series mapping, including keyword identification, research hotspot extraction, correlation between publishing units [6].
2.1. Data Sources
The database for literature search was mainly web of science, and the search period was 60 years of relevant literature from 1963 to 2023. The search keywords were “phytoplankton”, “cold region lakes”, “cyanobacterial blooms”, “ice-covered Lake”, “phytoplankton dynamics”, “winter”, “winter limnology”, “winter convection” and “phytoplankton dynamics”. The “phytoplankton” must be included, which included a total of 8919 papers.
2.2. Hot Spot Analyze
Based on a total of 60 years from 1963 to 2023, the top five countries that have published research on the competitive mechanism of phytoplankton growth in cold lakes were the United States (2,084), China (1654), Canada (747), the United Kingdom (665), and Germany (663) (Figure 1). The number of published articles has increased significantly, especially in the United States and China after 2007. The number of publications in the UK, France, and Canada has remained relatively stable. The number of publications in the United States peaked at 158 in 2019.
In recent years, the number of publications in China has significantly increased, especially since 2018, from 96 to the current 198. In contrast, the number of published articles in the United States has shown a slow downward trend since reaching its peak in 2010. The number of publications in Canada, the United Kingdom, and Germany has been relatively stable in the past few years. Although there have been some fluctuations, the overall change is not significant. 280 hot keywords extracted from 1963 to 2013 based on WOS database for hot spot analysis (Figure 2). The 280 keywords with high frequency were climate change (3359), phytoplankton (1346), temperature (1098), Nitrogen (1095) and ice (1008).

Table 1.
Mapping of keyword frequency, centrality and high-frequency burst.
| order | Sort by frequency of occurrence | Centrality sorting | Emergent analysis | ||||||
| Key words | frequency | Key words | Centrality | Key words | Emergence rate | Start | Over | 1963-2023 | |
| 1 | Phytoplankton | 3359 | Phytoplankton | 0.11 | Carbon | 0.87 | 2007 | 2012 |  |
| 2 | Climate change | 1346 | Water | 0.09 | Plankton | 0.09 | 2007 | 2010 |  |
| 3 | Temperature | 1098 | Great lakes | 0.09 | Dynamics | 0.12 | 2009 | 2015 |  |
| 4 | Nitrogen | 1095 | impacts | 0.08 | Sea ice | 0.85 | 2010 | 2014 |  |
| 5 | ice | 1008 | ecology | 0.07 | Nitrogen | 0.5 | 2013 | 2016 |  |
| 6 | Phosphorus | 970 | ice | 0.06 | diversity | 0.69 | 2014 | 2017 |  |
| 7 | Constructed wetland | 969 | dynamics | 0.06 | winter | 0.88 | 2014 | 2019 |  |
| 8 | Denitrification | 823 | climate | 0.05 | tibetan plateau | 0.55 | 2019 | 2023 |  |
| 9 | Waste water treatment | 762 | trends | 0.05 | northern hemisphere | 0.1 | 2020 | 2023 |  |
| 10 | Dynamics | 707 | diversity | 0.05 | lake ice phenology | 0.66 | 2020 | 2023 |  |
analysis in the WoS database.
3. Definition and Driving Mechanism
3.1. Definition of Algal Blooms in Cold Lake
There were differences in the definitions of cold lakes among international scholars. They defined cold lakes based on indicators such as the average temperature of the coldest month, the duration of the freezing period, and the average annual water temperature (Table 2). In 1966, American scholars defined cold lakes based on their average freezing period exceeding 100 days per year and ice depth exceeding 0.3 meters for at least one year every 10 years [22]. In 1997, Yang defined China’s cold lakes based on the fact that the coldest month had a temperature below -3 ° C, the number of months with an average monthly temperature above 10 ° C did not exceed 4 months, the freezing period of lakes exceeded 100 days, and the proportion of solid precipitation received exceeded 50% [22]. In 2000, Yang added accumulated temperature and annual average snowfall days to define cold lakes [107]. In 2013, lakes with an annual average water temperature below 15 ° C were defined as cold lakes by Lurling [22]. Cyanobacterial blooms that occured in lakes with water temperatures well below optimal growth temperatures were defined as cold water cyanobacterial blooms [23].
The United States has conducted a lot of research on phytoplankton in cold lakes at high latitudes. Cyanobacteria (Microcystis, Anabaena, Aphanizomenon, Asterix, Chromococcus, etc.) and diatoms (Icelandic Streptocystis, filamentous diatoms, Baikal Streptocystis, Navicula gracilis, Glacier Navicula, Antarctic Thalassiocystis, Nitzschia fragmenta, etc.) were the dominant algae species in typical cold lakes. The driving factors of dominant cyanobacteria included TP, TN, WT and BOD5, among which nutrients were the main driving factors.
Table 3.
Dominant phytoplankton species and driving factors in typical cold region lakes.
| Lake/country | Representative dominant species | Impact factor |
| Alte Donau/(US) | Raphidiopsis and raciborskii | TN TP |
| Bethel Lake/Canada | Dolichospermum affinis | TN TP BOD5 |
| Brandy Lake/Canada | Aphanizomenonspp.And Dolichospermum | TN TP BOD5 WT |
| Suya Lake Reservoir/China | Cyanobacteria: Microcystis and Anabaena | TN TP |
| Lake Baikal/Russia | Diatom Melosomum baikalense (Russia) | WT TN |
| Lake Erie/North America | Cyanobacteria: Anabaena and Aphanizomenon and MicrocystisDiatoms and Melosceles Icelandica and filamentous diatoms | TN TP |
| Balkan Lakes/Albania | Diatoms and Cyclotella menifolia and Golden algae and Yellow algae | TN TP |
| Lake Luboszki / Latvia | Cyanobacteria | TN TP BOD5 WT |
| Antarctica | Diatoms and Navicula gracilis and Navicula glacierica and Thalassiosira and Nitzschia fragmenta | TN TP NO3- |
| Hulun Lake/China | Cyanobacteria and Chromococcus Chlorella and Fibrous algae and Chlorella and Chlamydomonas | NH3-N TP TN |
| Lake Khanka/China | Cyanobacteria Microcystis and Anabaena | TN TP BOD5 WT |
| Xidayang Reservoir/China | Cyanobacteria Microcystis | BOD5 TN TP |
| Devil’s Lake/US | Dolichospermum circinalis and Aphanizomenon flos-aquae | BOD5 |
| Fernan Lake/US | Microcystis spp and Dolichospermum spp and Gloeotrichia spp. | TN TP |
| LakeStechlin/Germany | Dolichospermum(primary)and Aphanizomenon, and Planktothrix | BOD5 WT |
| Lough Neagh/Northern Ireland | Planktothrix agardhii and Pseudanabaena spp. | TN TP BOD5 WT |
| Neusiedler see/Austria | Aphanocapsa incerta and Oscillatoria and Dolichospermum | TN TP WT chl-a |
| Three Mile Lake/Canada | Aphanizomenonspp.and Dolichospermum spp. | TP BOD5 WT |
| VänernWeyhenmeyer/Sweden | Aphanizomenon sp. | TN BOD5 WT |
3.2. Hydrothermal and Hydrodynamic Conditions
Hydrodynamics and thermal stratification under lake ice were the key physical factors that trigger the spatiotemporal heterogeneity of subglacial phytoplankton [24]. The difference of hydrodynamic processes between ice- period and freeze-thaw period leads to the different migration and transformation of nutrients from other lakes, which further provided an explanation for the competitive mechanism of phytoplankton growth in cold lakes from the perspective of physical environment (Figure 3) [25,26]. Notably, snow on the ice also affected the vertical distribution of physical and chemical properties in the lake by limiting the amount of solar radiation available under the ice [25]. The physical processes of lakes during the freeze-thaw and ice-covered periods are as follows:
- (1)
- Freeze-thaw period (Figure 3c): It mainly occured in early winter and early spring. During this period, there were significant temperature variations and lakes were in a state of both freezing and thawing. In early winter, the freezing process caused nutrients from ice to be released into the water, which triggerred osmotic convection and stratification of the water column [16]. This caused the water quality factors to slow down and formed a stable stratified state [26]. During melting, solar radiation (higher than in winter) penetrating the ice will cause hydrodynamic processes to become more intensity [27]. Radiation-driven (RDC) convection driven vertical convective circulation, which penetrates from the surface boundary layer to the stratified water column below [28]. How water quality factors were affected by increasing water temperature, increasing convection and increasing resussuspension are worthy of further discussion. The melting of ice and snow in early spring can lead to a large amount of freshwater flowing into lakes, exacerbating the resuspension of sediment at the bottom of the lake and carrying a large amount of pollutions into the lake [26]. Wind-induced waves can also cause sediment re-suspension, which induces the release of nutrients (such as nitrogen and phosphorus) from the sediment into the water body [29].
- (2)
- Ice-covered period (Figure 3a-b): mainly occured in midwinter. The presence of snow and ice reduced the available solar radiation under the ice, and the radiation-driven convection has little affected on the hydrodynamic force under the ice (Figure 3a) [26]. Lake convective circulation continued to occur slowly when there was no snow on the ice (Figure 3b) [24].
The variation of hydrodynamic circulation under ice also triggerred vertical variations in water quality indicators (Figure 3). Total phosphorus (TP), nitrate (NO3-), and ammonium (NH4+) concentrations all increased with increasing water depth. Under different conditions, light and temperature have a significant effect on the concentration and distribution of various chemical substances in the water body. The presence of ice will reduce the surface water temperature and dissolved oxygen content, while the concentration of chemical substances at the bottom was generally higher [31]. The increasing of available light underwater would increased the surface temperature, chlorophyll-a concentration, and dissolved oxygen, when the surface of the lake was free of ice and snow [32]. Understanding these impacts will help us better understand and predict the ecological processes and ecosystem functions of phytoplankton. The impact of subglacial hydrodynamics on phytoplankton was multifaceted, including nutrient transport, dissolved oxygen levels, and the special environment of the ice-covered period [33]. These processes controlled subglacial phytoplankton growth by influencing the migration and transformation of N and P between water and sediment [34]. High concentrations of nutrients and low nitrogen-phosphorus ratios can maintain the growth rate of phytoplankton during the ice-covered period, promoting phytoplankton more adaptable to the light conditions during the ice-covered period [35]. Light, temperature, and nutrients work synergistically to maintain the growth of phytoplankton under the ice.
3.3. Nutrient Migration and Transformation During Freezing
The multi-media migration and transformation of nutrients during the freezing-ice-thaw process was the key to affecting the growth competition of phytoplankton under the ice (Figure 4). The ice-covered period changed the physical and chemical processes of the lake, resulting in different nutrient migration and transformation from other seasons [36]. In early winter, the lake has a strong repulsive effect on nutrients during the ice-covered period, resulting in higher nutrient concentrations in the lake water than in the ice [34]. Nutrients continued to migrate and concentrate in the water with lake ice crystals gradually precipitate [37]. During the ice-covered period, the dynamic balance of nutrients between water and sediments was broken, and the nutrients in the water migrates to the sediment under the driving force of the concentrations gradient [38]. Sediments accept a large amount of nutrients and become endogenous pollutants [39]. However, dissolved nitrogen and phosphorus in the interstitial water of endogenous sediments tended to be released into the subglacial water during the ice-covered period, which will also lead to an nutrients increasing in the water [40]. In addition to the uncertainty of the migration direction and process of nutrients, their transformation process was also more complicated. Organic nitrogen in sediments formed ammonium through ammonification, and ammonium salts formed nitrates under nitrification and denitrification (Figure 4). Organic phosphorus in sediments formed phosphates under mineralization [41]. Phytoplankton mainly absorbed carbon dioxide and other nutrients from water through photosynthesis. These nutrients can contribute to the nitrogen fixation activity of nitrogen fixing cyanobacteria [6,7,8,9]. In addition, phytoplankton residues were also the main source of organic nitrogen and phosphorus in sediments [42]. External inputs such as factory wastewater and agricultural wastewater directly provided phosphates and nitrates to phytoplankton. In a word, the direction and amount of multi-media migration and transformation of nutrients in the freezing-thawing process are uncertain, which would increase the difficulty of studying the nutrient driving mechanism of algal blooms in cold lakes.
3.4. Phytoplankton Characteristics
Phytoplankton is the basis of material circulation and energy flow in lake ecosystems, and plays a very important role in maintaining the balance of lake ecosystems and improving the ecological environment [43,44]. Variations in phytoplankton community structure directly affect water quality. Phytoplankton growth during the ice-covered period was related to the characteristics of phytoplankton cellular structure and the migration and transformation of nutrients [45]. The main reason why cyanobacteria can easily gain an advantage during the ice-covered period was that cyanobacteria have air vesicles and have a stronger ability to adapt to temperature changes [46]. Air vesicles can regulate buoyancy, allowing cyanobacteria to float on the surface of the water body, thereby obtaining more light available under the ice for photosynthesis during the ice-covered period [47]. Therefore, cyanobacteria can survive and reproduce in harsh environments such as low temperature, low oxygen and low nitrogen [35,36,37,38,39,40,41,42,43,44,45,46]. Diatoms have siliceous cell walls (silica shells) and lack air vesicles. Diatoms usually inhabit the lower and middle layers of the water column, relying on water flow and turbulence to maintain suspension [19]. Therefore, diatoms are more adapted to living in environments with abundant light and rich nutrients [61].
4. Advances in Research Methods
Common research methods for the competition mechanism of phytoplankton in cold lakes included remote sensing, field sampling, indoor simulation and numerical simulation [37]. Among them, the conventional methods were remote sensing, field sampling and indoor simulation. With the development of computer technology, data-driven numerical simulation and mechanism-driven numerical simulation were gradually applied to the prediction and prevention of cyanobacterial blooms in lakes.
4.1. Conventional Technical Means
At present, field monitoring is the basis of all monitoring. The advantage of on-site monitoring was that sample testing is the most accurate [38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102]. However, the monitoring frequency, monitoring cost and monitoring scope all limited the effective prevention and control of lake algal blooms [82]. Remote sensing technology was widely used due to its wide coverage and high real-time performance [36]. However, remote sensing technology has the limitations of discontinuous spatiotemporal monitoring and complex monitoring methods due to the existence of rain and fog [91]. In order to predict the mechanism of multiple environmental factors on phytoplankton growth competition, indoor simulation has became a conventional method [39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57]. Experimental simulation experiments can accurately control variables such as temperature, light, nutrients, etc, and have the advantages of strong repeatability and high reliability. Equipment technology and simulation scale limited the accuracy of prediction.
4.2. Numerical Simulation Model
Numerical simulation includes data-driven machine learning methods and mechanism-driven hydrodynamic-ecological integrated model methods. They have significant advantages in the study of competition mechanisms among phytoplankton in cold lakes [49]. The deeper and more comprehensive understanding of the advantages of numerical simulation for research was provided [93]. Machine learning methods can extract complex patterns and relationships from a large amount of observational data, which were suitable for processing high-dimensional and nonlinear data [49]. This method was suitable for real-time or near-real-time applications. The disadvantage of data-driven numerical simulation was that it was highly data-dependent [95]. The model relied more on statistical relationships in the data rather than physical mechanisms [47].
In order to enhance the accuracy and precision of model simulation, it is necessary to construct a numerical simulation model for mechanism characterization [94]. The mechanism model was based on physical and ecological principles, can provide a physical explanation of the system behavior. These models were applicable to lakes of different types and sizes, and has strong extrapolation capabilities [48,100]. Most numerical simulations focused on the characterization of the non-ice-covered period, and lack of the construction of all-season model [Powers et al. 2016]. There were few studies and data on ice-covered lakes, lacking systematicity and in-depthness [44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63]. Therefore, it is urgent to improve the hydrodynamic-algal bloom module based on the principles of lake ice dynamics, mass balance principle and algal competition mechanism, and construct all-season hydrodynamic-water quality-water ecology coupling model (Figure 5). Among them, the hydrodynamic module mainly corrected the hydraulic and thermal elements during the ice-covered period. During the ice-covered period, the lake surface water level used a water gauge and an automatic flow monitor to measure ice thickness (Hi), water depth under ice (d), water level (L), flow velocity (v), and the water level are corrected using the density difference between ice and water and the ice body pressure. Net solar radiation (I), atmospheric radiation (a), convective heat flux (QS), evaporative heat flux (QE), reflectivity (β), and initial temperature (T) are also obtained based on empirical formulas or measurement methods. On the basis of the traditional lake hydrodynamics-water quality-water ecology coupling model, the algal bloom module (BLOOM) is nested to reflect the growth and competition process of algae at the level of phenotypic characteristics. Hydrodynamic, water quality and algal bloom modules are modified as Equation (1)–(6).
The roughness coefficient during the ice-covered period is nc, and the expression is:
nb is the lake basin roughness; ni is the ice cover roughness value.
The expression of the actual water level is:
z is the actual water level, zm is the monitored water level, Hi is the ice thickness, ρi and ρw are the densities of ice and water respectively.
The total heat flux expression of the ice surface is:
where I0 is the net solar radiation (short wave), Qai is the net atmospheric radiation (long wave), Qbr is the reflection (long wave), Qii is the evaporation heat flux (latent heat flux), and Qsi is the convective heat flux (sensible heat flux).
Based on the traditional lake hydrodynamic-water quality-water ecology model, the algal bloom module (BLOOM) is nested to reflect the growth and competition process of algae at the level of algal representation.
Mass balance equation:
C is the concentration of water quality parameters; t is the simulation time; u, v, w represent the flow velocity in the x, y, and z directions respectively; Kx, Ky, Kz represent the turbulent diffusion coefficients in the x, y, and z directions respectively; Sc represents the biochemical transformation of nutrients.
Algae growth and characterization equations:
Phyi is the phytoplankton type, kd is the total extinction coefficient, SN,i is the stoichiometry of nitrogen in type i phytoplankton, and SP,i is the stoichiometry of phosphorus in type i phytoplankton.
5. Response to Climate Change
Global warming has led to rising temperatures, shortening the ice-covered time of cold lakes and advancing the melting time [43]. Phytoplankton is sensitive to environmental changes. Physical and biogeochemical changed during the ice-covered period (such as subglacial solar radiation, temperature, subglacial hydrodynamics, nutrient migration and transformation, nutrient migration and concentration, and dominant phytoplankton species) will affect the structure and function of phytoplankton communities [31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63]. This variations not only affected the seasonal dynamics of lakes, but also had a profound impact on lake ecosystems. The shortening of ice-covered time and the advancement of melting time provide a longer growing season for algae, resulting in the expansion of the range of algal blooms to high-latitude ice-covered lakes [58]. High-latitude regions may not have had algal blooms before, but these areas are also suitable for algal growth with climate warming. Rising water temperatures speed up the circulation of nutrients in the lake, which increases the intensity and frequency of algal blooms [29]. Algal blooms frequently occur in early spring, causing serious impacts on lake water quality, leading to eutrophication of water bodies and destroying the ecological balance of lakes [45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84].
Subglacial physical effected influence subglacial phytoplankton competition by influencing biogeochemistry. The most important physical effects were the dynamic changes of underwater light, lake ice sheet thickness and lake temperature [25,64]. The intensification of fluid dynamics, nutrient migration and transformation under the ice led to the early end of the freezing period due to the rising temperature [102]. The reduction of lake ice cover and the shortening of ice age may lead to increased light intensity and longer photoperiods in subglacial waters [18,19,20,21,22]. This was conducive to the photosynthesis of subglacial phytoplankton, which may lead to an increase in its abundance and productivity.
The melting of lake ice may change the distribution and supply of oxygen, affecting the respiratory process of plankton [97]. The increasing of temperature in subglacial water environment may affect the growth, reproduction and metabolic processes of plankton, thereby affecting its ecological function and survival ability [58]. The reduction of ice cover and the increase in water temperature led to more intense and earlier algal blooms [73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93]. The ice-covered period changed the chemical processes of the lake, resulting in nutrient migration and transformation that were different from other seasons. The nutrient concentrations affected in the water during the ice-covered period may be more significant under global warming. Nutrient may migrate faster from the subglacial water body to the interstitial water, changing the dynamic balance of salt between the water body and the sediment with decreasing ice layer [93]. Variation in nutrient concentrations affected the diffusion and transformation of nutrients. The increasing endogenous nitrogen and phosphorus would lead to lake eutrophication, providing abundant nutrients and promoting the rapid growth and reproduction of phytoplankton [41].
6. Conclusion and Outlook
Understanding the mechanism of algal blooms in cold lakes is an important part of enriching and developing winter limnology, and is also the basis of environmental management and water ecological protection of cold lakes. Based on a literature review of nearly 60 years, this study has summarized the current hot spots and future development directions of phytoplankton growth in cold lakes. Elucidated the mechanisms by which lake ice induced hydrodynamics, nutrient migration and transformation, and the inherent characteristics of phytoplankton compete for growth in cold lakes. The main factors for the frequent occurrence of algal blooms in early spring after ice-melting are analyzed. The existence of ice-melting-ice-covering further made the response of phytoplankton in cold lakes more sensitive to climate change. Variations in lake ice phenology (shortened ice-covered time, early ice-melting time, and increased water temperature) have led to the expansion of the scale, increase in frequency, and prolonged duration of algal blooms in cold lakes. Existing technical such as field monitoring, remote sensing inversion, and indoor simulation can no longer meet the real-time monitoring of algal blooms in cold lakes and effective predictions for future climate change. It is urgent to build a hydrodynamics-water quality-water ecology comprehensive model for cold lakes in all-seasons to provide technological for the effective prevention and control of algal blooms in cold lakes. Based on the existing conclusions, the following prospects are proposed for the study of algal blooms in cold lakes:
- (1)
- Provide a definition of the validity of cold lakes with physical and ecological significance;
- (2)
- Accurately characterize the subglacial hydrodynamics and biogeochemical processes under the action of ice sheet formation and decline, so as to reveal the internal mechanism of algal blooms in cold lakes;
- (3)
- Build a comprehensive simulation technology of hydrodynamics, water quality and water ecology for cold lakes in all seasons, so as to provide technical means for the precise prevention and control of algal blooms in cold lakes;
- (4)
- Strengthen the in-depth understanding of the response of algal blooms in cold lakes to climate change, so as to propose adaptive prevention and control strategies to cope with future climate change.
Data Availability Statement
Some or all data, models, or code generated or used during the study are available from the corresponding author by request.
Acknowledgements
This research was supported by the National Natural Science Foundation of China ((Grant NO. U23A2008 and 42207088); Jilin Province Science Foundation (YDZJ202401475ZYTS); The Consulting Project Proposal of the Chinese Academy of Engineering (JL2023-17).
Conflict of Interest
There is not any conflict of interest.
References
- Huisman, J.; Codd, G.A.; Paerl, H.W.; et al. Cyanobacterial blooms. NatureReviews Microbiology 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; et al. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environment international 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Sellner, K.G. Physiology, ecology, and toxic properties of marine cyanobacteria blooms. Limnology and oceanography 1997, 42, 1089–1104. [Google Scholar] [CrossRef]
- Falconer, I.R. Toxic cyanobacterial bloom problems in Australian waters: risks and impacts on human health. Phycologia 2001, 40, 228–233. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; et al. The rise of harmful cyanobacteria blooms: the potential roles of eutrophication and climate change. Harmful algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Vincent, W.F.; Hobbie, J.E.; Laybourn-Parry, J. Introduction to the limnology of high-latitude lake and river ecosystems. Polar lakes and rivers: limnology of Arctic and Antarctic aquatic ecosystems 2008, 1–24. [Google Scholar]
- Przytulska, A.; Bartosiewicz, M.; Vincent, W.F. Increased risk of cyanobacterialblooms in northern high-latitude lakes through climate warming and phosphorusenrichment. Freshwater Biology 2017, 62, 1986–1996. [Google Scholar] [CrossRef]
- Vincent, W.F. Cold tolerance in cyanobacteria and life in the cryosphere[M]//Algae and cyanobacteria in extreme environments. Dordrecht: Springer Netherlands 2007, 287-301.
- Pandey, K.D.; Shukla, S.P.; Shukla, P.N.; et al. Cyanobacteria in Antarctica: ecology, physiology and cold adaptation. CELLULAR AND MOLECULAR BIOLOGY-PARIS-WEGMANN- 2004, 50, 575–584. [Google Scholar] [CrossRef]
- Salmaso, N.; Capelli, C.; Shams, S.; et al. Expansion of bloom-forming Dolichospermum lemmermannii (Nostocales, Cyanobacteria) to the deep lakes south of the Alps: colonization patterns, driving forces and implications for water use. Harmful Algae 2015, 50, 76–87. [Google Scholar] [CrossRef]
- Babanazarova, O.; Sidelev, S.; Schischeleva, S. The structure of winter phytoplankton in Lake Nero, Russia, a hypertrophic lake dominated by Planktothrix-like Cyanobacteria. Aquatic Biosystems 2013, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Reinl, K.L.; Harris, T.D.; North, R.L.; et al. Blooms also like it cold. Limnology and Oceanography Letters 2023, 8, 546–564. [Google Scholar] [CrossRef]
- Özkundakci, D.; Gsell, A.S.; Hintze, T.; et al. Winter severity determines functional trait composition of phytoplankton in seasonally ice-covered lakes. Global change biology 2016, 22, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Hampton, S.E.; Moore, M.V.; Ozersky, T.; et al. Heating up a cold subject: prospects for under-ice plankton research in lakes. Journal of plankton research 2015, 37, 277–284. [Google Scholar] [CrossRef]
- Wright, R.T. The Ecology of Phytoplankton in an Ice-covered Lake[D]. Harvard University, 1963.
- Jansen, J.; MacIntyre, S.; Barrett, D.C.; et al. Winter limnology: How do hydrodynamics and biogeochemistry shape ecosystems under ice? Journal of Geophysical Research: Biogeosciences 2021, 126, e2020JG006237. [Google Scholar] [CrossRef]
- Powers, S.M.; Hampton, S.E. Winter limnology as a new frontier. Limnology and Oceanography Bulletin 2016, 25, 103–108. [Google Scholar] [CrossRef]
- Favot, E.J.; Rühland, K.M.; DeSellas, A.M.; et al. Climate variability promotes unprecedented cyanobacterial blooms in a remote, oligotrophic Ontario lake: evidence from paleolimnology. Journal of Paleolimnology 2019, 62, 31–52. [Google Scholar] [CrossRef]
- Favot, E.J.; Rühland, K.M.; DeSellas, A.M.; et al. Climate variability promotes unprecedented cyanobacterial blooms in a remote, oligotrophic Ontario lake: evidence from paleolimnology. Journal of Paleolimnology 2019, 62, 31–52. [Google Scholar] [CrossRef]
- Klanten, Y.; MacIntyre, S.; Fitzpatrick, C.; et al. Regime shifts in lake oxygen and temperature in the rapidly warming high Arctic. Geophysical Research Letters 2024, 51, e2023GL106985. [Google Scholar] [CrossRef]
- Salonen, K.; Leppäranta, M.; Viljanen, M.; et al. Perspectives in winter limnology: closing the annual cycle of freezing lakes. Aquatic Ecology 2009, 43, 609–616. [Google Scholar] [CrossRef]
- Chen, C. CiteSpace II: Detecting and visualizing emerging trends and transient patterns in scientific literature. Journal of the American Society for information Science and Technology 2006, 57, 359–377. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, L.; Li, Y.; et al. Characteristics of the Northern Hemisphere cold regions changes from 1901 to 2019. Scientific Reports 2023, 13, 3879. [Google Scholar] [CrossRef] [PubMed]
- Leppäranta, M. Freezing of lakes[M]//Freezing of Lakes and the Evolution of their Ice Cover. Cham: Springer International Publishing 2023, 17-62.
- Bouffard, D.; Zdorovennova, G.; Bogdanov, S.; et al. Under-ice convection dynamics in a boreal lake. Inland Waters 2019, 9, 142–161. [Google Scholar] [CrossRef]
- Zhao, W.; Huang, W.; Li, R.; et al. Solar radiation transfer in an ice-covered lake at different snow thicknesses. Hydrological Sciences Journal 2024, 1–12. [Google Scholar] [CrossRef]
- Yang, B.; Wells, M.G.; McMeans, B.C.; et al. A new thermal categorization of ice-covered lakes. Geophysical Research Letters 2021, 48, e2020GL091374. [Google Scholar] [CrossRef]
- Jonas, T.; Terzhevik, A.Y.; Mironov, D.V.; et al. Radiatively driven convection in an ice-covered lake investigated by using temperature microstructure technique. Journal of Geophysical Research: Oceans 2003, 108. [Google Scholar] [CrossRef]
- Talling, J.F. Photosynthetic characteristics of some freshwater plankton diatoms in relation to underwater radiation. The New Phytologist 1957, 56, 29–50. [Google Scholar] [CrossRef]
- Rosa, F. Sedimentation and sediment resuspension in Lake Ontario. [CrossRef]
- Journal of Great Lakes Research1985, 11, 13–25. [CrossRef]
- Bengtsson, L.; Malm, J.; Terzhevik, A.; et al. Field investigation of winter thermo-and hydrodynamics in a small Karelian lake. Limnology and oceanography 1996, 41, 1502–1513. [Google Scholar] [CrossRef]
- Leu, E.; Søreide, J.E.; Hessen, D.O.; et al. Consequences of changing sea-ice cover for primary and secondary producers in the European Arctic shelfseas: Timing, quantity, and quality. Progress in Oceanography 2011, 90, 18–32. [Google Scholar] [CrossRef]
- Yang, T.; Hei, P.; Song, J.; et al. Nitrogen variations during the ice-on season in the eutrophic lakes. Environmental Pollution 2019, 247, 1089–1099. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Goldthwait, S.A.; Hansell, D.A. Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea. Deep Sea Research Part I: Oceanographic Research Papers 2002, 49, 1445–1461. [Google Scholar] [CrossRef]
- Zhao, W.; Huang, W.; Li, R.; et al. Solar radiation transfer in an ice-covered lake at different snow thicknesses. Hydrological Sciences Journal 2024, 1–12. [Google Scholar] [CrossRef]
- Yang, B. Physical Processes in Ice-covered Lakes[D]. University of Toronto (Canada), 2022.
- Whitfield, C.J.; Casson, N.J.; North, R.L.; et al. The effect of freeze-thaw cycles on phosphorus release from riparian macrophytes in cold regions. Canadian Water Resources Journal/Revue Canadienne des Ressources Hydriques 2019, 44, 160–173. [Google Scholar] [CrossRef]
- Fang, Y.; Changyou, L.; Leppäranta, M.; et al. Notable increases in nutrient concentrations in a shallow lake during seasonal ice growth. Water Science and Technology 2016, 74, 2773–2783. [Google Scholar] [CrossRef]
- Cao, X.; Wang, Y.; He, J.; et al. Phosphorus mobility among sediments, water and cyanobacteria enhanced by cyanobacteria blooms in eutrophic Lake Dianchi. Environmental Pollution 2016, 219, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Vallina, S.M.; Cermeno, P.; Dutkiewicz, S.; et al. Phytoplankton functional diversity increases ecosystem productivity and stability. Ecological Modelling 2017, 361, 184–196. [Google Scholar] [CrossRef]
- Borics, G.; Abonyi, A.; Salmaso, N.; et al. Freshwater phytoplankton diversity: models, drivers and implications for ecosystem properties. Hydrobiologia 2021, 848, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Kudoh, S.; Imura, S.; et al. Phytoplankton blooms under dim and cold conditions in freshwater lakes of East Antarctica. Polar Biology 2008, 31, 199–208. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Lovejoy, C.; Vincent, W.F. Global distribution of cyanobacterial ecotypes in the cold biosphere. The ISME Journal 2010, 4, 191–202. [Google Scholar] [CrossRef]
- Priscu, J.C.; Adams, E.E.; Paerl, H.W.; et al. Perennial Antarctic lake ice: a refuge for cyanobacteria in an extreme environment. Life in ancient ice 2005, 22–49. [Google Scholar]
- Neumann, A.; Kim, D.K.; Perhar, G.; et al. Integrative analysis of the Lake Simcoe watershed (Ontario, Canada) as a socio-ecological system. Journal of environmental management 2017, 188, 308–321. [Google Scholar] [CrossRef]
- Wilkins, D.; Yau, S.; Williams, T.J.; et al. Key microbial drivers in Antarctic aquatic environments. FEMS Microbiology Reviews 2013, 37, 303–335. [Google Scholar] [CrossRef]
- Sridhar, R.; Thangaradjou, T.; Kumar, S.S.; et al. Water quality and phytoplankton characteristics in the Palk Bay, southeast coast of India. Journal of environmental biology 2006, 27, 561–566. [Google Scholar]
- Montes-Hugo, M.; Doney, S.C.; Ducklow, H.W.; et al. Recent changes in phytoplankton communities associated with rapid regional climate change along the western Antarctic Peninsula. Science 2009, 323, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Prowse, T.; Alfredsen, K.; Beltaos, S.; et al. Effects of changes in arctic lake and river ice. Ambio 2011, 40, 63–74. [Google Scholar] [CrossRef]
- Laybourn-Parry, J.; Pearce, D.A. The biodiversity and ecology of Antarctic lakes: models for evolution. Philosophical Transactions of the Royal Society B: Biological Sciences 2007, 362, 2273–2289. [Google Scholar] [CrossRef]
- Zepernick, B.N.; Chase, E.E.; Denison, E.R.; et al. Declines in ice cover are accompanied by light limitation responses and community change in freshwater diatoms. The ISME Journal 2024, wrad015. [Google Scholar] [CrossRef] [PubMed]
- Twiss, M.R.; McKay RM, L.; Bourbonniere, R.A.; et al. Diatoms abound in ice-covered Lake Erie: An investigation of offshore winter limnology in Lake Erie over the period 2007 to 2010. Journal of Great Lakes Research 2012, 38, 18–30. [Google Scholar] [CrossRef]
- Anderson, S.P.; Fernald KM, H.; Anderson, R.S.; et al. Physical and chemical characterization of a spring flood event, Bench Glacier, Alaska, USA:evidence for water storage. Journal of Glaciology 1999, 45, 177–189. [Google Scholar]
- Weyhenmeyer, G.A.; Obertegger, U.; Rudebeck, H.; et al. Towards critical white ice conditions in lakes under global warming. Nature communications 2022, 13, 4974. [Google Scholar] [CrossRef]
- Wüest, A.; Pasche, N.; Ibelings, B.W.; et al. Life under ice in Lake Onego(Russia)–an interdisciplinary winter limnology study. Inland Waters 2019, 9, 125–129. [Google Scholar] [CrossRef]
- Hébert, M.P.; Beisner, B.E.; Rautio, M.; et al. Warming winters in lakes: Later ice onset promotes consumer overwintering and shapes springtime planktonic food webs. Proceedings of the National Academy of Sciences 2021, 118, e2114840118. [Google Scholar] [CrossRef] [PubMed]
- Kirillin, G.; Leppäranta, M.; Terzhevik, A.; et al. Physics of seasonally ice-covered lakes: a review. Aquatic sciences 2012, 74, 659–682. [Google Scholar] [CrossRef]
- Peeters, F.; Straile, D.; Lorke, A.; et al. Earlier onset of the spring phytoplankton bloom in lakes of the temperate zone in a warmer climate. Global Change Biology 2007, 13, 1898–1909. [Google Scholar] [CrossRef]
- Huber, V.; Adrian, R.; Gerten, D. Phytoplankton response to climate warming modified by trophic state. Limnology and Oceanography 2008, 53, 1–13. [Google Scholar] [CrossRef]
- O’Reilly, C.M.; Sharma, S.; Gray, D.K.; et al. Rapid and highly variable warming of lake surface waters around the globe. Geophysical Research Letters 2015, 42, 773–10. [Google Scholar] [CrossRef]
- Keller, W. Implications of climate warming for Boreal Shield lakes: a review and synthesis. Environmental Reviews 2007, 15, 99–112. [Google Scholar] [CrossRef]
- Kumar, N.; Grogan, P.; Chu, H.; et al. The effect of freeze-thaw conditionson arctic soil bacterial communities. Biology 2013, 2, 356–377. [Google Scholar] [CrossRef]
- Jawak, S.D.; Kulkarni, K.; Luis, A.J. A review on extraction of lakes from remotely sensed optical satellite data with a special focus on cryospheric lakes. Advances in Remote Sensing 2015, 4, 196–213. [Google Scholar] [CrossRef]
- Siegert, M.J.; Makinson, K.; Blake, D.; et al. An assessment of deep hot-water drilling as a means to undertake direct measurement and sampling ofAntarctic subglacial lakes: Experience and lessons learned from the Lake Ellsworth field season 2012/13. Annals of Glaciology 2014, 55, 59–73. [Google Scholar] [CrossRef]
- Patterson, J.C.; Hamblin, P.F. Thermal simulation of a lake with winter ice cover 1. Limnology and Oceanography 1988, 33, 323–338. [Google Scholar] [CrossRef]
- Toporowska, M.; Pawlik-Skowronska, B.; Krupa, D.; et al. Winter versus summer blooming of phytoplankton in a shallow lake: effect of hypertrophic conditions. Polish Journal of Ecology 2010, 58, 3–12. [Google Scholar]
- Richardson, D.C.; Filazzola, A.; Woolway, R.I.; et al. Nonlinear responses in interannual variability of lake ice to climate change. Limnology and Oceanography 2024, 69, 789–801. [Google Scholar] [CrossRef]
- Ozersky, T.; Bramburger, A.J.; Elgin, A.K.; et al. The changing face of winter: lessons and questions from the Laurentian Great Lakes. 2021.
- Yang, F.; Cen, R.; Feng, W.; et al. Dynamic simulation of nutrient distribution in lakes during ice cover growth and ablation. Chemosphere 2021, 281, 130781. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E.; Hayes, P.K.; Boje, R. The gas vesicles, buoyancy and vertical distribution of cyanobacteria in the Baltic Sea. European Journal of Phycology 1995, 30, 87–94. [Google Scholar] [CrossRef]
- Pozdnoukhov, A.; Foresti, L.; Kanevski, M. Data-driven topo-climatic mapping with machine learning methods. Natural hazards 2009, 50, 497–518. [Google Scholar] [CrossRef]
- Giusti, E.; Marsili-Libelli, S. Spatio-temporal dissolved oxygen dynamics in the Orbetello lagoon by fuzzy pattern recognition. Ecological Modelling 2009, 220, 2415–2426. [Google Scholar] [CrossRef]
- Ralston, D.K.; Moore, S.K. Modeling harmful algal blooms in a changing climate. Harmful Algae 2020, 91, 101729. [Google Scholar] [CrossRef] [PubMed]
- Havens, K.E. Cyanobacteria blooms: effects on aquatic ecosystems. Cyanobacterial harmful algal blooms: state of the science and research needs 2008, 733–747. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Farnsley, S.E.; LeCleir, G.R.; et al. The relationships between nutrients, cyanobacterial toxins and the microbial community in Taihu (Lake Tai), China. Harmful Algae 2011, 10, 207–215. [Google Scholar] [CrossRef]
- Wüest, A.; Lorke, A. Small-scale hydrodynamics in lakes. Annual Review of fluid mechanics 2003, 35, 373–412. [Google Scholar] [CrossRef]
- Bouffard, D.; Wüest, A. Convection in lakes. Annual Review of Fluid Mechanics 2019, 51, 189–215. [Google Scholar] [CrossRef]
- Bengtsson, L.; Malm, J.; Terzhevik, A.; et al. Field investigation of winter thermo-and hydrodynamics in a small Karelian lake. Limnology and oceanography 1996, 41, 1502–1513. [Google Scholar] [CrossRef]
- Hodges, B.R.; Imberger, J.; Laval, B.; et al. Modeling the hydrodynamics of stratified lakes[C]//Hydroinformatics 2000 conference. Iowa Institute of Hydraulics Research 2000, 4, 23–27. [Google Scholar]
- Freeman, E.C.; Creed, I.F.; Jones, B.; et al. Global changes may be promoting a rise in select cyanobacteria in nutrient-poor northern lakes. Global Change Biology 2020, 26, 4966–4987. [Google Scholar] [CrossRef] [PubMed]
- Priscu, J.C.; Adams, E.E.; Paerl, H.W.; et al. Perennial Antarctic lake ice: a refuge for cyanobacteria in an extreme environment. Life in ancient ice 2005, 22–49. [Google Scholar]
- Rühland, K.; Paterson, A.M.; Smol, J.P. Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European lakes. Global Change Biology 2008, 14, 2740–2754. [Google Scholar] [CrossRef]
- Anderson, N.J. Diatoms, temperature and climatic change. European Journal of Phycology 2000, 35, 307–314. [Google Scholar] [CrossRef]
- Elser, J.J.; Wu, C.; González, A.L.; et al. Key rules of life and the fading cryosphere: Impacts in alpine lakes and streams. Global change biology 2020, 26, 6644–6656. [Google Scholar] [CrossRef]
- MacCallum, S.N.; Merchant, C.J. Surface water temperature observations of large lakes by optimal estimation. Canadian Journal of Remote Sensing 2012, 38, 25–45. [Google Scholar] [CrossRef]
- Pernica, P.; North, R.L.; Baulch, H.M. In the cold light of day: The potential importance of under-ice convective mixed layers to primary producers. Inland Waters 2017, 7, 138–150. [Google Scholar] [CrossRef]
- Leppäranta, M. Freezing of lakes and the evolution of their ice cover[M]. Berlin/Heidelberg, Germany: Springer, 2015.
- Smetacek, V. Diatoms and the ocean carbon cycle. Protist 1999, 150, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Bopp, L.; Aumont, O.; Cadule, P.; et al. Response of diatoms distribution to global warming and potential implications: A global model study. Geophysical Research Letters 2005, 32. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A. Warmer winters: are planktonic algal populations in Sweden’s largest lakes affected? AMBIO: A Journal of the Human Environment 2001, 30, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Katz, S.L.; Izmest’eva, L.R.; Hampton, S.E.; et al. The “M elosira years” of Lake B aikal: Winter environmental conditions at ice onset predict under-ice algal blooms in spring. Limnology and Oceanography 2015, 60, 1950–1964. [Google Scholar] [CrossRef]
- Coffer, M.M.; Schaeffer, B.A.; Darling, J.A.; et al. Quantifying national and regional cyanobacterial occurrence in US lakes using satellite remote sensing. Ecological Indicators 2020, 111, 105976. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Vincent, W.F. Cyanobacteria in polar and alpine ecosystems. Psychrophiles: from biodiversity to biotechnology 2017, 181–206. [Google Scholar] [CrossRef]
- Eckerstorfer, M.; Bühler, Y.; Frauenfelder, R.; et al. Remote sensing of snow avalanches: Recent advances, potential, and limitations. Cold Regions Science and Technology 2016, 121, 126–140. [Google Scholar] [CrossRef]
- Martynov, A.; Sushama, L.; Laprise, R. Simulation of temperate freezing lakes by one-dimensional lake models: performance assessment for interactive coupling with regional climate models. Boreal environment research 2010, 15, 143. [Google Scholar]
- Surdu, C.M.; Duguay, C.R.; Brown, L.C.; et al. Response of ice cover on shallow lakes of the North Slope of Alaska to contemporary climate conditions (1950–2011): radar remote-sensing and numerical modeling data analysis. The Cryosphere 2014, 8, 167–180. [Google Scholar] [CrossRef]
- Jia, X.; Willard, J.; Karpatne, A.; et al. Physics-guided machine learning for scientific discovery: An application in simulating lake temperature profiles. ACM/IMS Transactions on Data Science 2021, 2, 1–26. [Google Scholar] [CrossRef]
- Guerrero, J.L.; Pernica, P.; Wheater, H.; et al. Parameter sensitivity analysis of a 1-D cold region lake model for land-surface schemes. Hydrology and Earth System Sciences 2017, 21, 6345–6362. [Google Scholar] [CrossRef]
- Hampton, S.E.; Moore, M.V.; Ozersky, T.; et al. Heating up a cold subject: prospects for under-ice plankton research in lakes. Journal of plankton research 2015, 37, 277–284. [Google Scholar] [CrossRef]
- Woolway, R.I.; Merchant, C.J. Amplified surface temperature response of cold, deep lakes to inter-annual air temperature variability. Scientific Reports 2017, 7, 4130. [Google Scholar] [CrossRef]
- Ozersky, T.; Bramburger, A.J.; Elgin, A.K.; et al. The changing face of winter: lessons and questions from the Laurentian Great Lakes. 2021.
- Malmaeus, J.M.; Blenckner, T.; Markensten, H.; et al. Lake phosphorus dynamics and climate warming: A mechanistic model approach. Ecological Modelling 2006, 190, 1–14. [Google Scholar] [CrossRef]
- Yang, B.; Wells, M.G.; Li, J.; et al. Mixing, stratification, and plankton under lake-ice during winter in a large lake: Implications for spring dissolved oxygen levels. Limnology and Oceanography 2020, 65, 2713–2729. [Google Scholar] [CrossRef]
- Arp, C.D.; Jones, B.M.; Whitman, M.; et al. Lake Temperature and Ice Cover Regimes in the Alaskan Subarctic and Arctic: Integrated Monitoring, Remote Sensing, and Modeling 1. JAWRA Journal of the American Water Resources Association 2010, 46, 777–791. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread global increase in intense lake phytoplankton blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef]
Figure 1.
Statistical chart of the top five countries with the most publications on lakes in cold regions, 1963-2023.
Figure 1.
Statistical chart of the top five countries with the most publications on lakes in cold regions, 1963-2023.
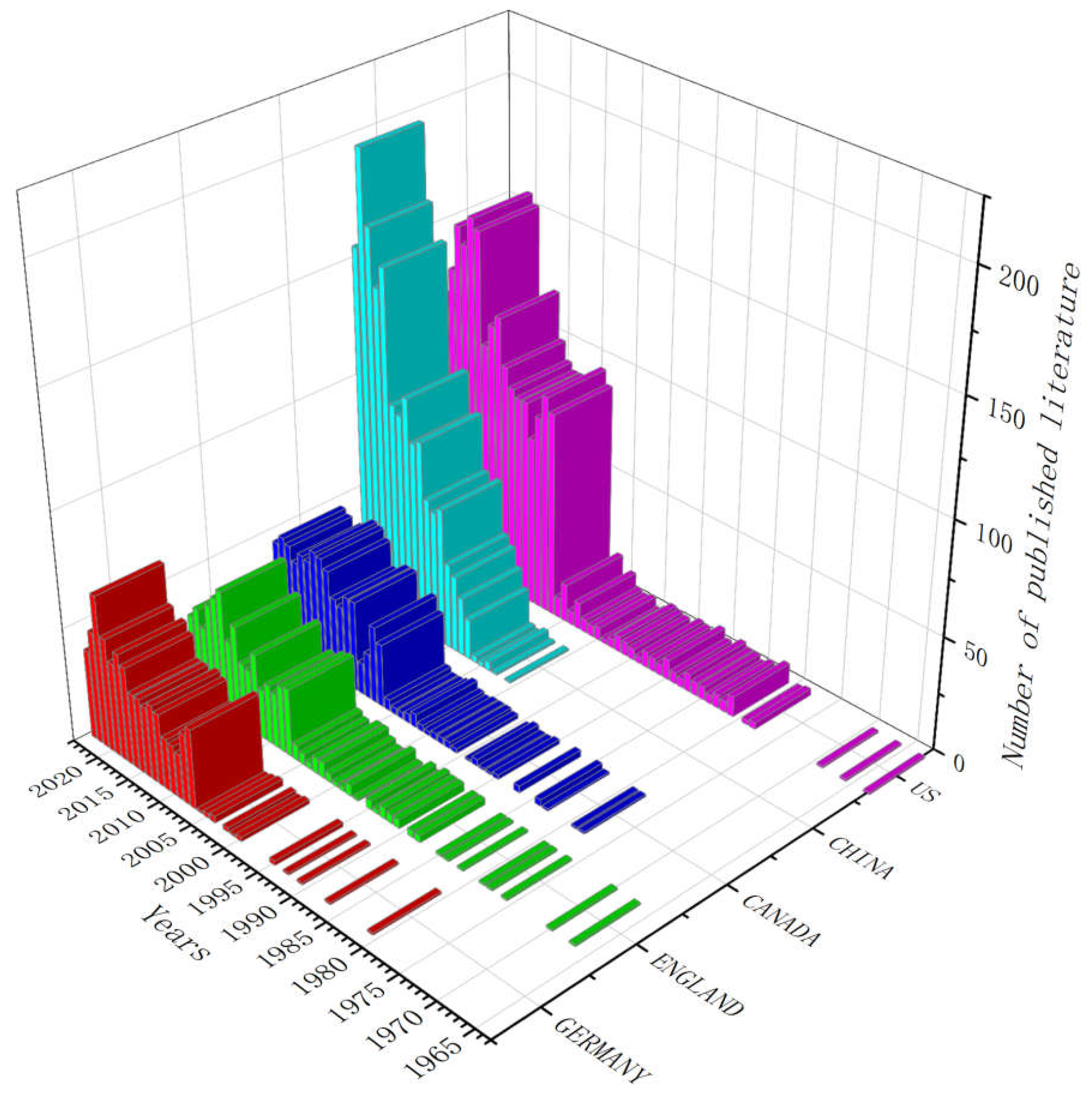
Figure 2.
Keyword co-occurrence analysis diagram.
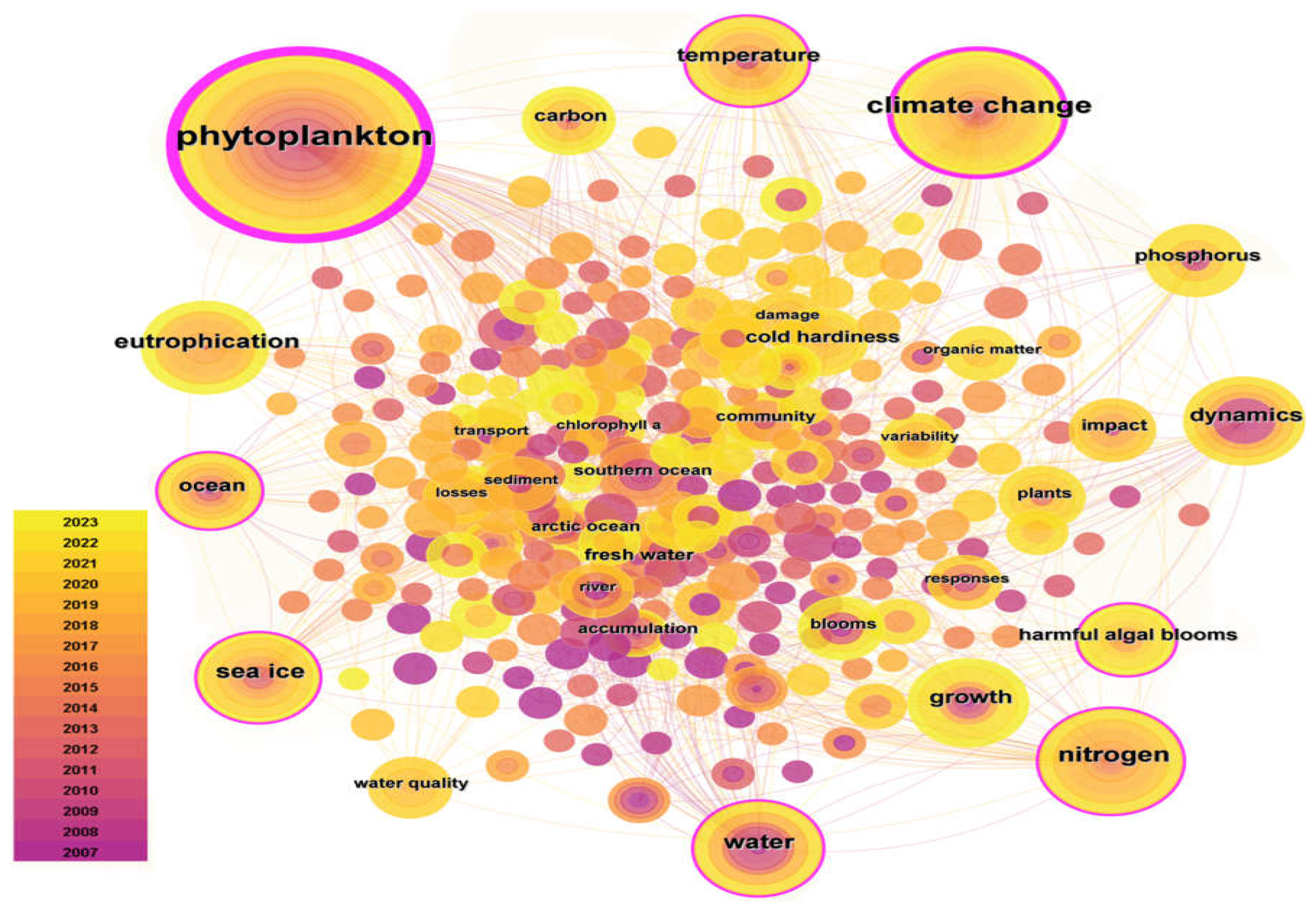
Figure 3.
Schematic diagram of the changes in water heat, hydrodynamics and physical and chemical forms of cold region lakes with water depth (lake ice cover stage diagram). The thermal-hydrodynamic and physicochemical properties of lake water change with water depth when ice and snow coexist in the lake in winter; (b) The thermal-hydrodynamic and physicochemical properties of lake water change with water depth when there is only ice in the lake in winter; (c) Schematic diagram of the thermal-hydrodynamic and physicochemical properties of lake water change with water depth after the early spring ice and snow melt.
Figure 3.
Schematic diagram of the changes in water heat, hydrodynamics and physical and chemical forms of cold region lakes with water depth (lake ice cover stage diagram). The thermal-hydrodynamic and physicochemical properties of lake water change with water depth when ice and snow coexist in the lake in winter; (b) The thermal-hydrodynamic and physicochemical properties of lake water change with water depth when there is only ice in the lake in winter; (c) Schematic diagram of the thermal-hydrodynamic and physicochemical properties of lake water change with water depth after the early spring ice and snow melt.
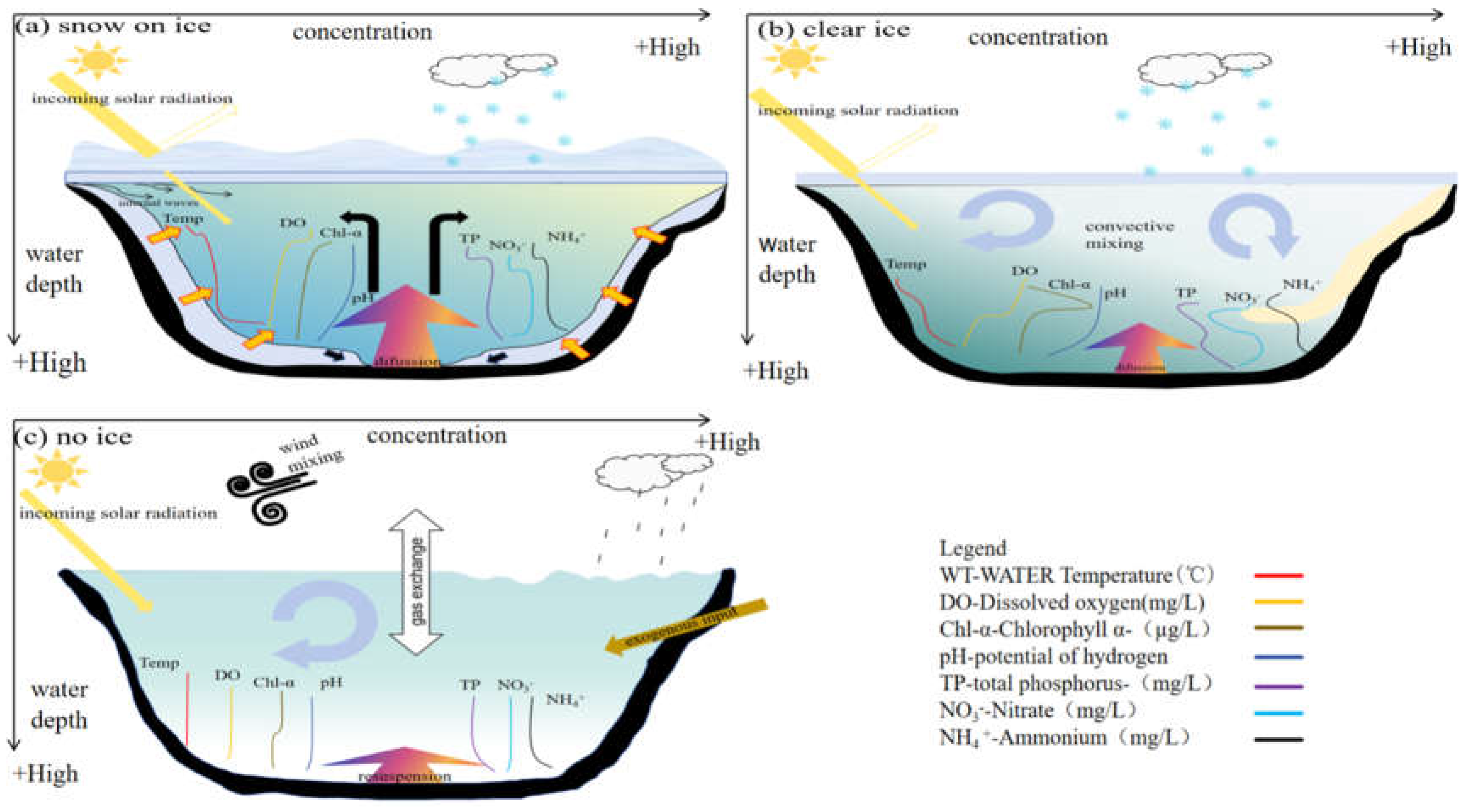
Figure 4.
Schematic diagram of the mechanism of the impact of nutrient migration and transformation on phytoplankton. Note: 1-12 represent ① Nitrogen fixation, ② Nitrification, ③ Denitrification, ④ Amination, ⑤ Assimilation, ⑥ Adsorption, ⑦ Desorption, ⑧ settlement, ⑨ Mineralization, ⑩ Resuspension, ⑪ Adsorption complexation, ⑫ Dissolve.
Figure 4.
Schematic diagram of the mechanism of the impact of nutrient migration and transformation on phytoplankton. Note: 1-12 represent ① Nitrogen fixation, ② Nitrification, ③ Denitrification, ④ Amination, ⑤ Assimilation, ⑥ Adsorption, ⑦ Desorption, ⑧ settlement, ⑨ Mineralization, ⑩ Resuspension, ⑪ Adsorption complexation, ⑫ Dissolve.
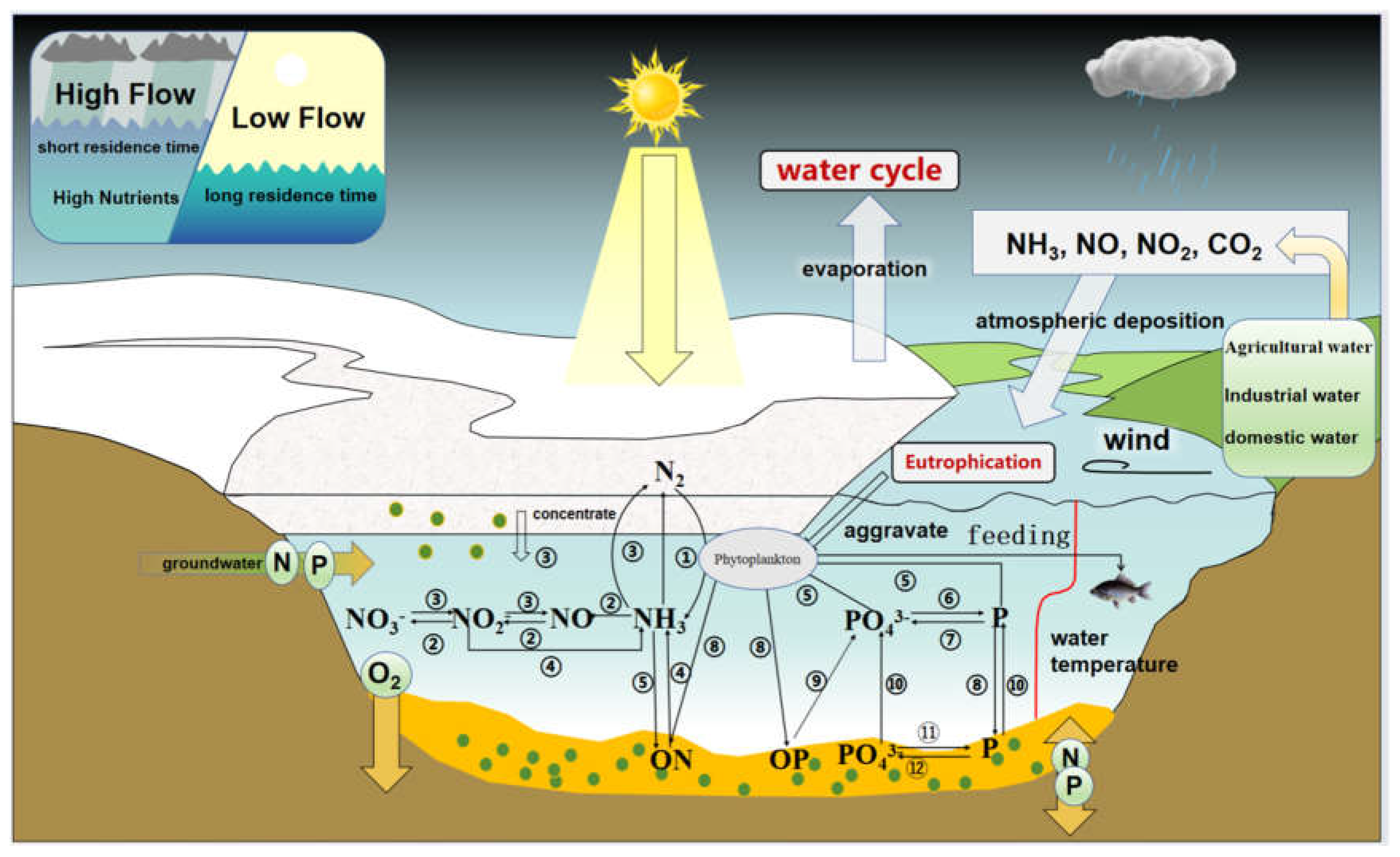
Figure 5.
Schematic diagram of the improved cold region lake hydrodynamic-water ecology model.
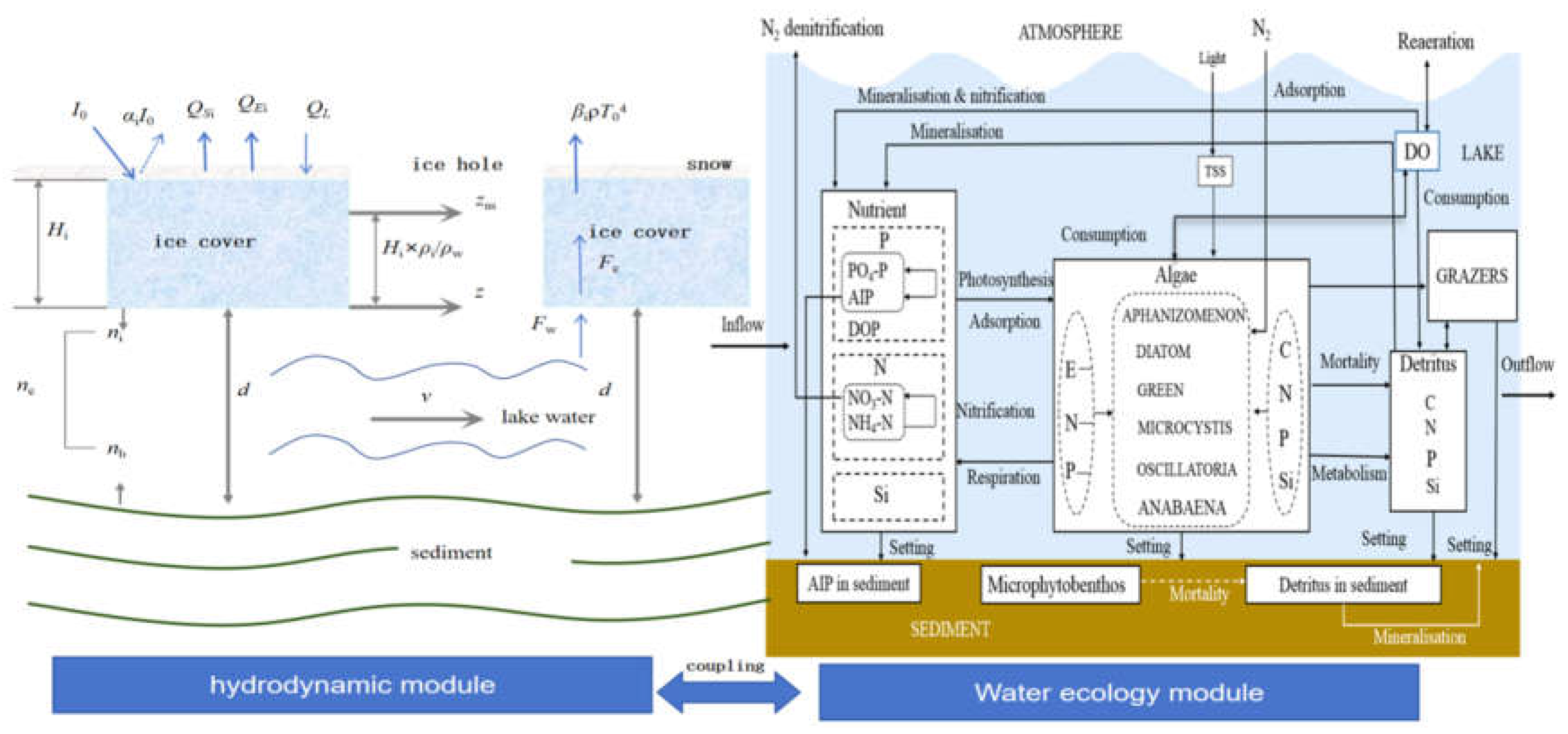
Table 2.
Definitions of lakes in cold regions.
| Name | Country | Identify | References |
| Bates, R. E. And Bilello | US | The maximum snow depth observed on the ground is greater than 0.3m, the average freezing period of rivers and lakes is greater than 100 days per year, and the ice depth is greater than 0.3m in at least one year every 10 years. | Bates, R. E. And Bilello et al. (1966) |
| Yang, Z | China | Yang proposed the criteria for the division of China’s cold regions, including the coldest month temperature below -3°C, the average monthly temperature above 10°C for no more than 4 months, the freezing period of rivers and lakes for more than 100 days, and the proportion of precipitation received in the form of frozen ice exceeding 50%. Yang et al. added the accumulated temperature between 500 and 1000°C and the average number of snow days per year of 30 days to calculate China’s cold regions. | Yang, Z et al. 2000 |
| Paerland and huisman 2008;Lurling et al. 2013 | US | When the annual average water temperature is below 15°C, which is far below the optimal temperature for cyanobacteria to grow, cyanobacterial blooms are observed, which are called cold water cyanobacterial blooms. | Paerland and huisman 2008;Lurling et al. 2013 |
| Maartje 2024 | US | Lake surface ice is defined as cold region lakes | Maartje et al. 2024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



