
Submitted:
08 August 2024
Posted:
08 August 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The COVID-19 pandemic has underscored the intricate interplay between viral infections and opportunistic bacterial pathogens such as Klebsiella pneumoniae. K. pneumoniae harbors various antibiotic resistance determinants like extended-spectrum and plasmid-mediated AmpC β-lactamases and carbapenemases. In the two years following the pandemic, carbapenemase producing K. pneumoniae isolates obtained routinely in outpatient and inpatient setting demonstrated similar resistance patterns which prompted research. The aim was to compare and subsequently analyze carbapenem-resistant hospital and outpatient’s K. pneumoniae isolates’ resistance mechanisms, traits and molecular epidemiology, following the COVID-19 pandemic. Disk diffusion and broth dilution method were the methods of choice to determine antibiotic susceptibility. β-lactamases were screened by phenotypic methods and confirmed with PCR. In total 113 isolates were analysed. Resistance to amoxicillin-clavulanate and ertapenem was confirmed in all strains. High resistance rates (over 90%) were observed for extended-spectrum cephalosporins, and ciprofloxacin. OKNV testing and PCR detected OXA-48 in 106, NDM in 7 and KPC in only one isolate. ESBLs accompanied carbapenemases in 103 isolates. Carbapenem resistance in K. pneumoniae is becoming a global problem, hence careful monitoring of its resistance traits is of utmost importance in order to combat the arising threat. OXA-48 seems to continue predominating in the turbulent period after pandemic.
Keywords:
Klebsiella pneumoniae
; OXA-48
; NDM
; resistance
; COVID-19
1. Introduction
The unprecedented global health crisis triggered by the COVID-19 pandemic has underscored the intricate interplay between viral infections and opportunistic bacterial pathogens [1]. This has led to an exacerbation of the severity of clinical outcomes, prolonged hospitalization periods, and increased mortality [1]. At the top of everything else, it has contributed to the escalation of bacterial resistance, due to selective pressure triggered by indiscriminate use of antibiotics during pandemic.
Of all opportunistic pathogens Klebsiella pneumoniae, alongside Acinetobacter baumannii, is the most important due to its capability to cause severe infections like pneumonia in ventilated patients (VAP), bloodstream infections (BSI), urinary tract infections (UTI) and wound infections in immunocompromised and mechanically ventilated patients. [2]. Higher risk patients are ones with severe underlying illnesses, implanted foreign devices, invasive procedures, malignancies and immunosuppression. In addition, it is an important causative agent of community-acquired UTIs in nursing home residents [3]. The clinical significance of K. pneumoniae is associated with amazing capacity to acquire diverse resistance determinants from other species mediated by horizontal spread of mobile genetic elements [4].
K. pneumoniae isolates harbors a plethora of various antibiotic resistance determinants including extended-spectrum β-lactamases (ESBLs), plasmid-mediated AmpC β-lactamases (p-Amp-C) and also carbapenemases. Colistin is very often the last resort antibiotic, but emergence of colistin resistance in K. pneumoniae limits its therapeutic use. Colistin resistance determinants are usually found in ESBL positive and carbapenem resistant K. pneumoniae (CRKP) resulting in multidrug (MDR) or extensively drug resistance phenotype (XDR) [5]. This poses a challenge to clinicians worldwide who treat these patients and a substantial threat to existing antibiotic armamentarium. The worldwide dissemination of CRKP and its drug resistance transfer poses a global public health threat [5].
In 1983, the first report of plasmid-mediated β-lactamase capable of hydrolyzing expanded-spectrum cephalosporins (ESC) in Klebsiella oxytoca was published [6]. They were named ESBLs and they have since been described worldwide [6]. The first ESBL was named SHV-2 which differed from SHV-1 in one aminoacid. At the beginning, the majority of ESBLs belonged to SHV and TEM family [7,8], but since early 2000s a rapid spread of CTX-M β-lactamases was reported, in particular, CTX-M-15 variant. CTX-M β-lactamases are primarily cefotaximases, which are classified into five clusters (CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9 and CTX-M-25) [9,10]. The fact that carbapenems are the treatment of choice for serious infections caused by ESBL producing organisms, and an increasing trend of fluoroquinolone resistance, has led to an increasing reliance on carbapenems in clinical practice and proliferation of carbapenemases. Genes conferring plasmid-mediated aminoglycoside and fluoroquinolone resistance are often associated with blaCTX-M genes [11]. ESBLs in Croatian K. pneumoniae isolates were first reported in 1998 with predominance of SHV-2, SHV-2a and SHV-5 variants associated with high level ceftazidime resistance [12,13]. Switch to CTX-M family, particularly CTX-M-15 variant, was noted in late 2000s [14].
P-AmpC β-lactamases are derived from the chromosomal β-lactamases of the bacteria belonging to the genus Enterobacter, Serratia, Citrobacter, Pseudomonas and Acinetobacter by the escape of the chromosomal gene to the plasmid. They hydrolyze expanded-spectrum cephalosporins (ESC), monobactams, and cephamycins, but spare fourth generation cephalosporins and carbapenems. Unlike ESBLs, they are not susceptible to the inhibition with clavulanic acid, sulbactam or tazobactam [15].
Resistance to carbapenems in K. pneumoniae is mediated by two main mechanisms. The first involves production of β-lactamases (p-AmpC or ESBL) with a very low level carbapenem hydrolyzing activity combined with decreased permeability due to porin loss or alteration. The second mechanism is attributed to carbapenem hydrolyzing β-lactamases [16]. Carbapenemases involved in acquired resistance to carbapenems in CRKP belong to Ambler class A serin β-lactamases (KPC, GES), class B metallo-β-lactamases (MBL) of the IMP, VIM or NDM family, and OXA-48-like β-lactamases belonging to the class D or carbapenem-hydrolyzing oxacillinases (CHDL) [17,18]. After the first report in Turkey in 2004 [19], OXA-48 spread in Europe, with a remarkable increase of OXA-48-producing organisms, reported in many countries with the highest rates observed in Turkey [20,21], France [22], and Germany [23]. Endemic areas of OXA-48 positive K. pneumoniae are India, Middle East and North Africa [24,25]. The gene encoding OXA-48 is plasmid- borne and located inside a composite transposon composed of two copies of the insertion sequence IS1999 [26]. The enzyme is capable of hydrolyzing penicillins and carbapenems, but possesses poor activity against broad spectrum cephalosporins. Multidrug resistance in OXA-48 producing organisms often results from coproduction of ESBLs or p-AmpC β-lactamases.
Carbapenemases emerged in Croatia in 2008 with NDM-1 in K. pneumoniae as the first published report [27], followed very soon with the emergence of the KPC-in the same hospital [28]. Later studies revealed shift to MBLs belonging to VIM family [29,30]. Rapid diffusion of OXA-48 was noticed after 2016 [31,32]. OXA-48 was the only carbapenemase reported in the outpatient setting in Croatia so far, mostly associated with urinary tract infections in elderly patients [33].
CTX-M β-lactamases were frequently identified as additional β-lactamases in isolates harboring carbapenemases [29,30]. In Croatia, CMY-4 was described in K. pneumoniae as additional β-lactamase to NDM-1 [27,29] and CTX-M-15 and CMY-16 in addition to NDM-1 as well [27].
Plasmid-borne fluoroquinolone resistance is usually mediated by qnr genes which are frequently harbored by the same plasmids encoding CTX-M ESBLs [34]. qnrA and qnrB genes were described in Croatia in earlier studies, linked to NDM metallo-β-lactamases [27,29].
Resistance genes in Klebsiella spp. are carried on conjugative plasmids. A formal scheme of plasmid classification is based on incompatibility (Inc) groups: plasmids with the same replication control are incompatible and cannot reside in the same cell line, whereas plasmids with different replication controls are compatible and can be propagated in the same cell. Classification is based on rep, tra and par genes which are implicated in replication, transfer and partitioning of the plasmids [35].
Colistin is the last resort antibiotic for treatment of serious infections caused by CRKP. The main mechanism of colistin resistance in K. pneumoniae is the inactivation of the mgrB gene, encoding a negative feedback regulator of the PhoQ-PhoP signaling system which activates the pmr system [36] responsible for addition of positively charged phosphoethanolamine and L-arabinose to negatively charged lipid A, and modifying the target for polymixins. Alternatively, the acquisition of plasmid-mediated mcr genes can result in colistin resistance. In Croatia it was observed for the first time in K. pneumoniae isolates with KPC or OXA-48 carbapenemases in 2013. The exact mechanism was not clarified but the isolates exhibited single nucleotide polymorphism (SNP) which was assumed to be related to colistin resistance [37].
Although it is a primarily hospital pathogen, hypervirulent strains can cause life threatening community acquired infections in healthy subjects without significant comorbidities and therefore it has become a worldwide concern [38]. Increasing emergence of hypervirulent, carbapenemase producing strains compromises antimicrobial options for severe infections [39].
In the years following the COVID-19 pandemic, a significant number of CRKP isolates obtained from outpatients shared similar resistance patterns as those obtained in the hospital during routine microbiology diagnostic. This prompted us to initiate a study which aims to compare carbapenem resistant hospital and outpatient’s K. pneumoniae isolates with regards to resistance mechanisms, traits and molecular epidemiology.
2. Materials and Methods
2.1. Bacterial Isolates and Patients
This is a descriptive cross section study conducted two major hospital centers in Zagreb, Croatia: University Hospital Centre Zagreb (UHCZ) and University Hospital Centre Sestre Milosrdnice (UHCSM). The bacterial isolates included in this study were obtained during routine microbiology testing. The 83 non-duplicate K. pneumoniae isolates with reduced susceptibility to carbapenems were collected in surgical intensive care units of the UHCZ and UHCS from 1st October 2022 until 31st December 2023 for the purpose of the study. Thirty outpatient’s isolates were collected only in UHCZ in the same period. The demographic and clinical data (age, gender, underlying diseases, site of infection or colonization and antimicrobial treatment prior and after isolation of resistant strains) were retrospectively analyzed from the internet medical records. The isolates were considered hospital acquired if they were isolated more than 48 h after admission to the hospital. Outpatients were defined as persons not hospitalized in the previous three months. K. pneumoniae isolates were confirmed by MALDI-TOF MS (matrix-assisted laser desorption ionization–time of flight mass spectrometry) Biotyper (Bruker, Daltonik GmbH, Bremen, Germany).
2.2. Antimicrobial Susceptibility Testing and Phenotypic Tests for Detection of ESBLs, Plasmid-Mediated AmpC β-Lactamases and Carbapenemases
The initial antibiotic susceptibility testing was done by the Kirby-Bauer disk-diffusion test according to the EUCAST guidelines [40]. Isolates exhibiting reduced susceptibility to carbapenems were subjected to further analysis. Minimum inhibitory concentrations (MICs) were determined by broth dilution method in Mueller-Hinton broth and 96 wells microtiter plates, according to CLSI standards [41] for the following antibiotics: amoxicillin-clavulanate, piperacillin-tazobactam, cefuroxime, expanded-spectrum cephalosporins or ESC (ceftazidime, cefotaxime, ceftriaxone), cefepime, imipenem, meropenem, gentamicin, amikacin, ciprofloxacin and colistin. Breakpoint of colistin was established by EUCAST, otherwise, we applied those defined by CLSI. Minimum inhibitory concentrations (MICs) were read as the lowest concentration of an antibiotic that inhibited visible growth after 18h at 37º C. E. coli ATCC 25922 and K. pneumoniae 700603 were used as quality control strains for MIC determination. The susceptibility to ceftazidime-avibactam, sulphametoxazole-trimethoprim, tetracycline, and chloramphenicol was determined only by disk-diffusion test. The isolates were classified as MDR, XDR or pandrug-resistant (PDR) as described previously by Magiorakos et al [42].
ESBL production was screened by double disk-synergy test (DDST) using amoxicillin-clavulanic acid disk opposite to ESC disks [43], and confirmed by CLSI-combined disk test using disks with ESC alone and with addition of clavulanic acid [41]. Augmentation of the inhibition zones of cephalosporin disks of at least 5 mm by clavulanic acid, confirmed ESBL production.
Taking into account the susceptibility to expanded-spectrum cephalosporins (ESC: ceftazidime, cefotaxime, ceftriaxone) and cefepime the isolates were assigned to three respective groups: group I (resistant to ceftazidime, cefotaxime, ceftriaxone and cefepime), group II (resistant to ceftazidime, cefotaxime, ceftriaxone, but susceptible to cefepime) and group III (resistant to ceftazidime, but susceptible to cefotaxime, ceftriaxone and cefepime) [44].
A double disk synergy test with a disk supplemented with 500 µg cloxacillin placed between disks containing ceftazidime and cefotaxime on a lawn of the K. pneumoniae isolates with reduced susceptibility to cefoxitin in order to detect p-Amp-C [8]. Distortion of the inhibition zones around ESC disks towards central disk with cloxacillin was considered a positive result [8]. P-AmpC were confirmed by AmpC disk test according to Black [45]. Blank paper disk was impregnated with 20 µl Tris-EDTA to permeabilize bacterial cells. Three to five colonies of the test organism were applied to the surface of the disk. The disk was placed on the surface of MH agar previously inoculated with cefoxitin susceptible E. coli ATCC 25922, nearby cefoxitin disk. Distortion of the inhibition zone around cefoxitin disk indicated enzymatic inactivation of cefoxitin [45].
The modified Hodge test (MHT) [46] and the imipenem EDTA inhibitor- based- test (IDTS) [47] were performed to presumptively identify carbapenemase and MBL, respectively.
For MHT, an overnight culture of carbapenem susceptible indicator strain E. coli ATCC 25922 was inoculated on the surface of Mueller-Hinton (MH) agar plates. After drying, ertapenem disk (10 µg) was placed in the middle of the plate. Overnight CRKP cultures were streaked as a single line from the periphery of the disc to the edge of the plate. The plates were incubated overnight at 37°C. Carbapenemase was suspected if the clover-leaf indentation of the indicator organism was observed towards the ertapenem disc [46].
Isolates proven to possess carbapenemase in MHT were further investigated by IDTS. Overnight CRKP culture was spread on MH agar plate. Imipenem and meropenem disks with and without EDTA were placed on the plate. Cultures were incubated overnight at 37 °C. Augmentation of the inhibition zone around carbapenem disk for at least 7 mm in the presence of EDTA was considered a positive result [47].
All carbapenemase positive CRKP were further tested for enzymatic hydrolysis of ertapenem by CIM test as described previously [48].
Initial screening for carbapenemase type was conducted in the participating centers for the purpose of routine diagnostic with immunochromatographic OKNV test (OXA-48, KPC, NDM, VIM) [49].
2.3. Molecular Detection of Resistance Genes
Total bacterial DNA was extracted by thermal lysis. The primers and protocols used to amplify broad spectrum and extended-spectrum β-lactamases (blaSHV, blaTEM, blaCTX-M,) [50,51,52], and fluoroquinolone resistance genes (qnrA, qnrB, qnrS) [34] were described previously. Multiplex PCR amplification was employed to identify cluster of CTX-M β-lactamase [53], p- AmpC β-lactamase genes [54] and carbapenemase encoding genes of class A, (blaKPC) class B (blaVIM, blaIMP and blaNDM) and carbapenem-hydrolyzing oxacillinases (blaOXA-48-like) [55]. Plasmid-encoded colistin resistance genes mcr-1 and mcr-2 were analyzed in isolate expressing elevated MICs of colistin [56]. PCR reactions were carried out in an AC196-Alpha Cycler (PCR max, UK). PCR mapping was applied to analyze the genetic platform surrounding OXA-48 encoding genes, with primers for IS1999 combined with forward and reverse primers for blaOXA-48 [57]. The presence of insertion sequence preceding the blaCTX-M genes was conducted by PCR mapping with forward primer for ISEcp1 and IS26 combined MA-3 (reverse for blaCTX-M genes) [58]. Representative amplicons were sent to Eurofin Genomics for sequencing. The positive control strains producing TEM-1, TEM-2 and SHV-1 and SHV-2 were kindly provided by Prof. Adolf Bauernfeind (Max von Pettenkofer Institute, Munich, Germany), CTX-M-15 by Prof. Neil Woodford (Health Protection Agency, London, UK), KPC- 2 by Prof. Fred Tenover (Stanford University School of Medicine), and OXA-48 by Dr. Yvonne Pfeifer (Robert Koch Institute, Wernigerode, Germany.
2.4. Detection of Resistance Genes by Inter-Array Kit CarbaResist
Four K. pneumoniae isolates were genotyped by an Inter-array chip according to the manufacturer’s recommendations (Inter-array fzmb GmbH, Bad Langensalza, Germany). The Inter-array genotyping Kit CarbaResist detects broad-spectrum β-lactamases, p-AmpC, ESBLs and carbapenemases and numerous other resistance genes (https://www.inter-array.com/further-genotyping-kits). RNA-free, unfragmented genomic DNA was isolated from pure culture of the test strains, amplified and internally labelled with biotin-dUDP according to the linear PCR amplification protocol using the antisense primer of the different targets only. Single-stranded DNA (ssDNA) reaction products were obtained. The biotin-labelled ssDNA was transferred to the ArrayWell and hybridised to DNA oligonucleotide microarrays with 230 probes for different β-lactam, aminoglycoside, fluoroquinolone, sulphonamide, trimethoprim and colistin resistance genes. HRP-conjugated streptavidin was bound to the hybridised biotin-labelled ssDNA stains and visualised by enzymatic reaction. The INTER-VISION Reader was used to evaluate the spots and their intensities automatically on the basis of a digital image of the microarray. The samples obtained from the strains tested in the study were automatically analysed for the presence or absence of specific probes, cross-checked against a database and then information about existing resistances was output.
2.5. Conjugation
Conjugal transfer of ertapenem, cefotaxime and cefoxitin resistance was carried out in mixed broth cultures at 35°C. E. coli J65 resistant to sodium azide was used as recipient [59]. The ESBL and carbapenemase-producing transconjugants were selected on MacConkey agar containing either ertapenem (0.5 mg/L) or cefotaxime (2 mg/L) and sodium azide (100 mg/L) to ensure that selected isolates were the recipient E. coli containing the transferred resistance gene. The frequency of conjugation was determined relatively to the number of donor cells. Cotransfer of resistance to gentamicin, tetracycline, sulfamethoxazole-trimethoprim, chloramphenicol, and ciprofloxacin was determined in addition to β-lactam resistance transfer. Colonies growing on combined plates were subjected to identification by MALDI-TOF and if confirmed to be E. coli to antibiotic susceptibility testing and PCR for detection of ESBL and carbapenemase encoding genes.
2.6. Characterization of Plasmids
Plasmid DNA of clinical isolates and their transconjugants was extracted with Qiagen Plasmid Mini Kit (Qiagen, Hilden, Germany). After staining with ethidium bromide, the DNA was visualized by ultraviolet light.
PCR-based replicon typing (PBRT) [60] was used for molecular typing of plasmids conferring resistance in Enterobacterales. Eighteen pairs of primers were used, including five multiplex and three simplex PCR in order to assess the plasmid incompatibility group. Since L/M plasmids which usually carry blaOXA-48 genes are difficult to detect with the original protocol, an updated protocol by Carattoli was applied [35].
PCR was performed on transconjugant strains to identify if ESBL and carbapenemase resistance genes were transferred in the conjugation experiment. PBRT was also applied on transconjugants to identify incompatibility groups such as in their respective donors. Positive control strains for PBRT were kindly provided by dr. A. Carattoli (Instituto Superiore di Sanita, Rome, Italy).
2.7. String Test
String test was attempted by stretching a mucoviscous string from the colony using a standard bacteriologic loop, as described previously [5]. If a viscous string >5 mm was formed, isolate was defined as hypermucoviscous.
2.8. Genotyping of the Isolates
Clonal types of two representative K. pneumoniae isolates (35 and 50) were determined by MLST, according to Diancourt et al [61].
3. Results
3.1. Bacterial Isolates and Patients
A total of 113 bacterial isolates were obtained from 113 patients and divided into 3 groups: UHCZ with 37 isolates, UHCSM with 46 isolates and outpatient setting with 30 isolates.
3.2. Antimicrobial Susceptibility and Phenotypic Tests for β-Lactamases
Resistance to amoxicillin/clavulanate and ertapenem was confirmed in all 113 strains. Very high resistance rates (95.6%, n=108) were observed for cefuroxime, ceftazidime, cefotaxime, ceftriaxone and cefepime with MIC values exceeding 128 mg/L. Except ESC, resistance rates were also high for ciprofloxacin (92.9%, n=105), piperacillin-tazobactam (86.7%, n=98) and gentamicin (71.7%, n=81) as shown in Figure 1 and Table 1. The best activity was exerted by ceftazidime-avibactam, colistin and amikacin, with 95.6% (n= 108), 91.1% (n= 103) and 61.9% (n=70) of the isolates being susceptible (Table 1 and Figure 1). Carbapenems exhibited heterogenous susceptibility patterns with ertapenem being resistant against all isolates, and imipenem and meropenem being in the susceptible, intermediate susceptible (susceptible at increased exposure) and resistant range according to CLSI. Meropenem exerted higher resistance rates compared to imipenem (81.4% vs 68.1 %) as illustrated in Table 1. Intermediate susceptibility or susceptibility at increased exposure was observed predominantly with piperacillin-tazobactam (20.3%), imipenem (12.4%) and meropenem (4.4%) as shown in Figure 1. Resistance to cefiderocol was not reported. All three groups of isolates showed similar resistant profiles as shown in Table 1., however, the resistance rates to ESC were higher among hospital isolates and exceeded 98.7% whereas in outpatient isolates the resistant rate was 86.7% (Table 1). According to the resistance pattern to ESC all isolates belonged to group I. The resistance rates to aminoglycosides and fluoroquinolones were also markedly lower in the isolates acquired in outpatient setting compared to the hospital isolates. MIC90 of all cephalosporins, gentamicin and ciprofloxacin exceeded 128 µg/ml. The most efficient antibiotics were imipenem and colistin with MIC90 of 32 µg/ml.
DDST and combined disk test with clavulanic acid confirmed ESBL production in 90.3% (n=102) of the isolates, whereas AmpC detection yielded negative results in all strains. ESBLs were more prevalent among hospital isolates as shown in Table 1 (91.9% and 93.5%) compared to outpatient setting in which the rate of ESBL positivity was 83.3%.
Modified Hodge and CIM test had moderate sensitivity and detected 81.4% (n=92) and 85.8% (n=97) of the isolates positive for carbapenemase in OKNV test. The rates of Hodge and CIM positivity were higher in the outpatient setting (100.0% vs 73.0-76.1%) and (100.0% vs 76.1-86.4%). Inhibitor based test with EDTA produced positive result with all seven strains positive for NDM in OKNV.
3.3. Molecular Detection of Resistance Genes
A complete concordance was reported between routine immunochromatographic OKNV testing and PCR which both detected OXA-48 in 93.8% (n=106), NDM in 6.2% (n=7) and KPC in only one isolate (0.9%). Double carbapenemases (OXA-48+NDM) were identified in one (0.9%) isolate as illustrated in Figure 2.
Following confirmation by multiplex PCR all ESBL positive isolates (n=102) were found to possess CTX-M β-lactamases belonging to cluster 1 as illustrated in Table 1. blaTEM genes tested positive in nine hospital isolates as shown in Table 1 and all generated TEM-1 allelic variant whereas blaSHV genes were found, as expected in all isolates. blaCTX-M genes in 25 isolates were linked to an upstream ISEcp1-like element. PCR for mcr genes gave negative result in all ten colistin resistant isolates.
PCR mapping and sequencing of the flanking regions of the blaOXA-48 genes revealed in 32 isolates the original structure reported by Gianni [57]. Isolates positive for IS1999 displayed elevated MICs of carbapenems and had positive phenotypic tests proving carbapenem hydrolysis.
3.4. Detection of Resistance Genes by Inter-Array Kit CarbaResist
Three OXA-48 producing organisms, according to OKNV and PCR, harbored almost identical resistance genes, as it is shown in Table 2. All possessed blaCTX-M-15, blaSHV, blaOXA-1 and blaOXA-48 genes. Resistance genes to non-β-lactam antibiotics were also identical: aac(6′)-Ib for aminoglycoside resistance, dfrA14 for trimethoprim resistance and oqxA and oqxB encoding efflux pumps. NDM producing organism tested positive for blaCTX-M-15, blaSHV, blaOXA-1 and blaNDM genes, aac(6′)-Ib, dfrA14, oqxA and oqxB as well, and additionaly harbored sul1 for sulphonamide resistance and aphA encoding aminoglycoside resistance.
3.5. Transfer of Resistance Determinants
Twenty- one OXA-48 positive isolates conjugated successfully at a frequency of 10-5 to 10-6 (transconjugant/donor). Other resistance determinants were not co-transferred. Attempts to transfer cefotaxime resistance were unsuccessful. Carbapenem resistance levels were significantly lower in the transconjugants in comparison to the donors.
3.6. Plasmid Characterization
PBRT showed that blaNDM gene was associated with IncX plasmid in three isolates and blaOXA-48 with IncL plasmid in 48 isolates. IncW, IncY and IncP occurred only sporadically in only 10, 5 and 1 isolate, respectively. No typable plasmids could be found in 30 clinical isolates. PCR performed on transconjugant clones showed that no ESBL-resistance determinants had co-transferred alongside with carbapenem resistance.
3.7. String Test
Hypervirulence was found in two isolates from UHCSM and one from UHCZ. All outpatient isolates were negative.
3.8. Genotyping
One isolate (35) was non- typable (gap-4, pho-4, pgi-57, inf-1, tonB-7, rpoB-1, mdh-2) whereas the other (50) belonged to ST4531 (gap-2, pho-1, pgi-5, inf-6, tonB-6, rpoB-1, mdh-2).
4. Discussion
Medical centers all over the world represent a vast reservoir of antimicrobial resistance genes including ESBLs and carbapenemase-encoding genes and Croatia is not different. The conduction of the study, in two major medical centers in Croatia, was a necessity to elucidate molecular mechanisms of resistance underlying the spread of β-lactamases among K. pneumoniae in the Croatian hospitals and community.
The main finding of the study is that OXA-48 combined with an ESBL belonging to the CTX-M family presented the most important contributor of ertapenem resistance among both hospital and community isolates. However, all except one hospital OXA-48 producing organisms were found to possess an additional ESBL, whereas OXA-48 and ESBL negative strains were identified predominantly in the outpatient setting. ESBL negativity among hospital isolates was linked to KPC and NDM carbapenemases, with only one ESBL negative isolate originating from the hospital. Interestingly, VIM- MBLs and KPC which were the most frequent in Croatian K. pneumoniae isolates at the beginning of carbapenemase spread [28,29], were not found or found only sporadically in case of KPC. Hodge and CIM tests confirmed carbapenemase production in the majority of isolates identified as carbapenemase positive in the routine OKNV test, although phenotypic tests missed some OXA-48 producers.
The resistance phenotype to imipenem and meropenem was inconsistent, ranging from complete susceptibility to plain resistance. On the other hand, uniform resistance to ertapenem was reported. This could be attributed to overproduction of an additional ESBL found in virtually all isolates, combined with porin loss which affects usually only ertapenem. Typically, CTX-M β-lactamases are known for their ability to rapidly hydrolyze cefotaxime yet ceftazidime MICs exceeded 128 mg/L in the majority of the isolates. The explanation is that blaCTX-M genes in all isolates belonged to the cluster 1 with CTX-M-15 as the only allelic variant detected. This widespread variant confers on the producing isolates high level resistance to all ESC, particularly if linked to ISEcp. This is in line with the previous reports from Croatia which described CTX-M group 1 as a dominant ESBL in Croatia with CTX-M-15 as the most prevalent allelic variant [14]. It was also reported as an additional β-lactamase to carbapenemases in the majority of earlier studies [29,30,31]. Additional ESBL provided the isolates high level of resistance to ESC and cefepime. Insertion sequence not only increased the level of resistance to ESC, but also mediated mobilization of the gene. However, conjugation experiments failed to transfer cefotaxime resistance which indicated possible chromosomal incorporation of the genes. The rate of additional ESBL positivity which conferred resistance to ESC was much higher among hospital isolates compared to the community ones. The earlier publications from late 90-ties reported higher resistance rates to ceftazidime as typical for group 3 resistance phenotype and linked to SHV-5 or SHV-12 ESBLs which were dominant at the beginning of ESBL dissemination in Croatia [12,13]. Since the majority of isolates produced OXA-48, which does not hydrolyze cephalosporins, and tested negative for p-AmpC, resistance to ESC was conferred by an additional ESBL. Additional TEM-1 β-lactamase in some isolates may have contributed to resistance to β-lactam inhibitor combinations.
OXA-48 producing isolates exhibited variable MICs for imipenem and meropenem, with a significant proportion of isolates being susceptible. Only ertapenem demonstrated uniform resistance. Resistance to imipenem and meropenem was related to the presence of IS1999 insertion sequence which is in concordance with its role as promotor, driving the expression of the gene and the level of carbapenem resistance. The isolates without detected insertion element showed imipenem and meropenem MICs in the susceptible or intermediate susceptible range. OXA-48 producing organism displayed uniform resistance to amoxicillin-clavulanate and piperacillin-tazobactam, which is in concordance with the substrate profile and inhibitor sensitivity of OXA-48. KPC and MBLs occurred only sporadically in the minority of the isolates. NDM which was very prevalent in the early stage of dissemination [27,29] occurred only sporadically in both hospitals and community, and without any epidemiological link with international travel and therefore could be associated with the Balkan clone [62]. Double carbapenemases found in the previous study from the same center [63], were identified in only one strain. MBLs pose a serious therapeutic problem because they hydrolyze all β-lactam antibiotics except aztreonam which is not available in Croatia and evade all approved β-lactamase inhibitors leaving only a few antibiotics active. Cumulative MIC values and the rate of resistant isolates were higher for meropenem, in spite of the fact that oxacillinases preferentially hydrolyze imipenem. This could be attributed to additional resistance mechanism such as hyperexpression of efflux systems which affect meropenem more than imipenem. The majority of isolates were resistant to almost all β-lactam antibiotics. However, piperacillin-tazobactam had a significant proportion of isolates in the intermediate or increased exposure category although OXA-48 harbored by majority of strains confers high-level resistance to all β-lactam inhibitor combinations. High rate of strains intermediate susceptible to imipenem and meropenem is in line with the predominance of OXA-48 carbapenemase with typical variable susceptibility to carbapenems.
P-AmpC were not found among our isolates. The possible explanation is that cephamycins are not licenced for clinical use in Croatia and thus there is no selection pressure to drive the spread of p-AmpC genes. P-AmpC were in the previous studies found as additional β-lactamase in CRKP [27,28,29,30]. qnr genes were not found in fluoroquinolone resistant isolates, indicating that fluoroquinolone resistance was mostly due to the mutations of chromosomal gyrA and parC genes, but clarification of these resistance traits was beyond this study. Bibliographical data in the past described qnrA and qnrB variants among our K. pneumoniae isolates harboring VIM-1 and NDM-1 [27,28,29,30]. However, in the present study all NDM producing isolates tested negative for plasmid-mediated fluoroquinolone resistance.
PCR for plasmid-mediated mcr genes was negative in all seven colistin resistant strains and therefore the resistance was probably attributed to disruption of mgrB gene as described previously in Italy [36].
L plasmid was responsible for spread of OXA-48 carrying genes in the both settings. This plasmid was previously shown to play a major role in diffusion of OXA-48 coding genes. IncP, IncFIC and IncW plasmids were found only sporadically. In addition to dissemination of resistance traits, they encode proteins involved in replication, transfer and segregational stability of the plasmids [64]. IncX plasmid identified among NDM producing organisms replaced L/M plasmid which carried blaNDM genes in the previous investigations [29]. Interestingly A/C plasmid which was dominant in the period of VIM- MBL domination was not found.
OXA-48 was carried by L type plasmid, reinforcing the hypothesis that the current spread of OXA-48 carrying genes in different species is mainly the consequence of the diffusion of an epidemic plasmid. Previous studies have shown that plasmids carrying blaOXA-48 genes, share similar features, in that they have an incL/M backbone and are very similar in size (60-70kb), suggesting their wide dissemination in different species and countries via an epidemic plasmid [64].
Hypervirulence was associated with OXA-48 producing organisms which is in disconcordance with the previous reports from Croatia in which this trait was associated with KPC carbapenemases [65] This is thought to be the result of acquiring plasmid-mediated resistance and virulence markers, either by transferring resistance plasmid into hypervirulent strain or virulence plasmid into carbapenem-resistant strain. Low prevalence of hypervirulence is in line with the previous reports [66].
Ceftazidime-avibactam and cefiderocol showed best activity with no resistant isolates, observed. Resistance to ceftazidime-avibactam was linked to MBL production. ST4531 identified in one isolate was previously reported in Italy in an outbreak associated with OXA-48 producing K. pneumoniae [67]. Implementation of immunochromatographic OKNV enables rapid identification of the carbapenemase class which enables clinical microbiologist to provide advice for the therapy before susceptibility testing is finished. Ceftazidime-avibactam is recommended if OXA-48 is confirmed while novel inhibitor combinations are ruled out if MBL is identified.
The study proved a long-lasting persistence of OXA-48 carbapenemase among CRKP in Croatia, beginning in 2015 [31,32]. This is in contrast with KPC and VIM which predominance lasted only a few years. Croatia is facing intensive population mixing and replacement in the last decade due to the war in Ukraine and Middle East alongside labor shortage which prompts immigration from Asian and African countries which were already linked to import of XDR M. tuberculosis. However, these changes did not affect the resistance determinants in CRKP. The reason for such a long persistence of OXA-48 is difficult to explain. It could be attributed to high transmissibility of L plasmid. Croatia was one of the EU countries with the highest mortality rates during COVID-19 pandemic which was in part due to the secondary bacterial infections associated mostly with A. baumannii and CRKP [68]. Increased usage of carbapenems and colistin to treat VAP and BSI caused by CRKP has led to accumulation of resistance traits.
The results presented in the study warrant development of practical containment methods alongside strong and immediate intervention in both major medical centers to prevent spread of resistant bacteria.
Carbapenem resistance in K. pneumoniae is a global phenomenon and it is necessary to monitor resistance traits of this important hospital pathogen and analyze the mobile genetic elements, responsible for the rapid diffusion of its resistance traits. In Croatia, a member of the European Union a national surveillance system exists which monitors carbapenemases in Gram-negative bacteria in order to effectively combat the arising threat.
5. Conclusions
The accumulation of resistance determinants underscores the necessity for stringent infection control measures and robust antimicrobial stewardship programs to curb MDR bacteria. The presence of multiple β-lactamase encoding genes, along with the co-occurrence of two carbapenemase encoding genes, underscores the challenge of antimicrobial resistance, emphasizing the need for targeted antimicrobial stewardship and collaborative efforts.
Author Contributions
Conceptualization, B.B.; validation, M.P.; M.N.; B.B.; H.C.; formal analysis, M.D., M.P.; M.N.; A.G; B.B.; data curation: A.N.H; H.C; J.L; G.Z; writing—original draft preparation, B.B.; writing—review and editing, J.L.; G.Z.; A.G.; M.P.; supervision, B.B. All authors have read and agreed to the published version of the manuscript.
Funding
Grant from the University of Zagreb School of Medicine (Extended-spectrum β-lactamases, plasmid-mediated β-lactamases and carbapenemases in Proteus mirabilis, number: 10106-24-1294.
Data Availability Statement
The data presented in the study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Ethical permission
The study was approved by the Ethical Committee of the University Hospital Centre Zagreb, number 02/013 AG, class 8.1-23/295-2, issued on 4th December 2023. Written consent was not necessary because the isolates were collected in the frames of routine laboratory diagnostic.
References
- Kurra, N.; Woodard, P.I.; Gandrakota, N.; Gandhi, H.; Polisetty, S.R.; Ang, S.P.; Patel, K.P.; Chitimalla, V.; Ali Baig, M.M.; Samudrala, G.O. Opportunistic Infections in COVID-19: A Systematic Review and Meta-Analysis. Cureus. 2022, 31, e23687. [Google Scholar] [CrossRef] [PubMed]
- Antoniadou, A.; Kontopidou, F.; Poulakou, G.; Koratzanis, E.; Galani, I.; Papadomichelakis, E.; Kopterides, P.; Souli, M.; Armaganidis, A.; Giamarellou, H. Colistin-resistant isolates of Klebsiella pneumoniae emerging in intensive care unit patients: First report of a multiclonal cluster. J Antimicrob Chemother. 2007, 59, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front Cell Infect Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Clegg, S.; Murphy, C.N. Epidemiology and Virulence of Klebsiella pneumoniae. Microbiol Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Xiao, X.; Zhout, X.; Zhang, I. Clinical and molecular characteristics of multi-clone carbapenem-resistant hypervirulent isolates in tertiary hospital in Bejing, China. Int. J. Infect. Dis. 2015, 37, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Kliebe, C.; Nies, B.A.; Meyer, J.F.; Tolxdorff-Neutzling, R.M.; Wiedemann, B. Evolution of plasmid-coded resistance to broad-spectrum cephalosporins. Antimicrob Agents Chemother. 1985, 28, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin Microbiol Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A.; Munoz-Price, L.S. The new β-lactamases. N Engl J Med. 2005, 352, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; D’Andrea, M.M.; Mugnaioli, C. The spread of CTX-M-type extended-spectrum beta-lactamases. Clin Microbiol Infect. Erratum in: Clin Microbiol Infect. 2008, (Suppl. 5), 21-24. 2008, 14 (Suppl. S1), 33–41. [Google Scholar] [CrossRef]
- Bonnet, R. Growing group of extended-spectrum β-lactamases: The CTX-M enzymes. Antimicrob Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef]
- Kim, H.B.; Park, C.H.; Kim, C.J.; Kim, E.C.; Jacoby, G.A.; Hooper, D.C. Prevalence of plasmid-mediated quinolone resistance determinants over a 9-year period. Antimicrob Agents Chemother. 2009, 53, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Bedenic, B.; Zagar, Z. Extended-spectrum β-lactamases in clinical isolates of Klebsiella pneumoniae from Zagreb, Croatia. J Chemother. 1998, 10, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Bedenić, B.; Randegger, C.C.; Stobberingh, E.; Hächler, H. Molecular epidemiology of extended-spectrum β-lactamases from Klebsiella pneumoniae strains isolated in Zagreb, Croatia. Eur J Clin Microbiol Infect Dis. 2001, 20, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Vranić-Ladavac, M.; Bosnjak, Z.; Beader, N.; Barisic, N.; Kalenic, S.; Bedenić, B. Clonal spread of CTX-M-15-producing Klebsiella pneumoniae in a Croatian hospital. J Med Microbiol. 2010, 59, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC β-lactamases. J. Clin. Microbiol. 2009, 22, 161–182. [Google Scholar] [CrossRef]
- Poirel, L.; Pitout, J.D.; Nordmann, P. Carbapenemases: Molecular diversity and clinical consequences. Future Microbiol. 2007, 2, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D.D.; Peirano, G.; Kock, M.M.; Strydom, K.A.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin Microbiol Rev. 2019, 33, e00102-19. [Google Scholar] [CrossRef] [PubMed]
- Albiger, B.; Glasner, C.; Struelens, M.J.; Grundmann, H.; Monnet, D.L. European Survey of Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) working group. Carbapenemase-producing Enterobacteriaceae in Europe: Assessment by national experts from 38 countries, May 2015. Euro Surveill. 2015, 20. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Heritier, C.; Tolun, V.; Nordmann, P. Emergence of oxacillinases-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob Agents Chemother. 2004, 48, 15–22. [Google Scholar] [CrossRef]
- Nazik, H.; Aydin, S.; Albayrak, R.; Bilgi, E.A.; Yildiz, I.; Kuvat, N.; Kelesoglu, F.M.; Kelesoglu, F.M.; Pakaştiçali, N.; Yilmaz, F.; et al. Detection and spread of OXA-48-producing Klebsiella oxytoca isolates in Istanbul, Turkey. Southeast Asian J Trop Med Public Health. 2014, 5, 123–129. [Google Scholar]
- Carrer, A.; Poirel, L.; Eraksoy, H.; Cagatay, A.; Badur, S.; Nordmann, P. Spread of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in Istanbul, Turkey. Antimicrob Agents Chemother. 2008, 52, 2950–2954. [Google Scholar] [CrossRef]
- Cuzon, G.; Quanich, J.; Gondret, R.; Naas, T.; Nordmann, P. Outbreak of OXA-48 positive carbapenem-resistant Klebsiella pneumoniae isolates in France. Antimicrob Agents Chemother. 2011, 55, 2420–2423. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, Y.; Schlatterrer, K.; Engelmann, E.; Schiller, R.A.; Frangenberg, H.D.; Holfelder, M.; Witte, W.; Nordmann, P.; Poirel, L. Emergence of OXA-48-type carbapenemase-producing Enterobacteriaceae in German Hospitals. Antimicrob Agents Chemother. 2012, 56, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D β-lactamases. Antimicrob Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [PubMed]
- Aubert, D.; Naas, T.; Héritier, C.; Poirel, L.; Nordmann, P. Functional characterization of IS1999, an IS4 family element involved in mobilization and expression of β-lactam resistance genes. J Bacteriol. 2006, 188, 6506–6514. [Google Scholar] [CrossRef] [PubMed]
- Mazzariol, A.; Bošnjak, Z.; Ballarini, P.; Budimir, A.; Bedenić, B.; Kalenić, S.; Cornaglia, G. NDM-1-producing Klebsiella pneumoniae, Croatia. Emerg Infect Dis. 2012, 18, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Bedenić, B.; Mazzariol, A.; Plečko, V.; Bošnjak, Z.; Barl, P.; Vraneš, J.; Cornaglia, G. First report of KPC-producing Klebsiella pneumoniae in Croatia. J Chemother. 2012, 24, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Zujić-Atalić, V.; Bedenić, B.; Kocsis, E.; Mazzariol, A.; Sardelić, S.; Barišić, M.; Plečko, V.; Bošnjak, Z.; Mijač, M.; Jajić, I.; et al. Diversity of carbapenemases in clinical isolates of Enterobacteriaceae in Croatia-the resu,lts of the multicenter study. Clinical Microbiology and Infection. 2014, 20, O894–O903. [Google Scholar] [CrossRef]
- Bedenić, B.; Sardelić, S.; Luxner, J.; Bošnjak, Z.; Varda-Brkić, D.; Lukić-Grlić, A.; Mareković, I.; Frančula-Zaninović, S.; Krilanović, M.; Šijak, D.; et al. Molecular characterization of class B carbapenemases in advanced stage of dissemination and emergence of class D carbapenemases in Enterobacteriaceae from Croatia. Infect Genetic Evol. 2016, 43, 74–82. [Google Scholar] [CrossRef]
- Bedenić, B.; Slade, M.; Žele-Starčević, L.; Sardelić, S.; Vranić-Ladavac, M.; Benčić, A.; Zujić Atalić, V.; Bogdan, M.; Bubonja-Šonje, M.; Tomić-Paradžik, M.; et al. Epidemic spread of OXA-48 β-lactamase in Croatia. J Med Microbiol. 2018, 67, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Jelić, M.; Škrlin, J.; Bejuk, D.; Košćak, I.; Butić, I.; Gužvinec, M.; Tambić-Andrašević, A. Characterization of Isolates Associated with Emergence of OXA-48-Producing Klebsiella pneumoniae in Croatia. Microb Drug Resist. 2018, 24, 973–979. [Google Scholar] [CrossRef]
- Šuto, S.; Bedenić, B.; Likić, S.; Kibel, S.; Anušić, M.; Tičić, V.; Zarfel, G.; Grisold, A.; Barišić, I.; Vraneš, J. Diffusion of OXA-48 carbapenemase among urinary isolates of Klebsiella pneumoniae in non-hospitalized elderly patients. BMC Microbiol. 2022, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef]
- Carattoli, A.; Seiffert, S.N.; Schwendener, S.; Perreten, V.; Endimiani, A. Differentiation of IncL and IncM plasmids associated with the spread of clinically relevant antimicrobial resistance. PLoS ONE 2015, 10, e0123063. [Google Scholar] [CrossRef] [PubMed]
- Cannateli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M.; COLGRIT Study Group. MgrB inactivation is a common mechanism of colistin resistance in KPC-producing Klebsiella pneumoniae of clinical origin. Antimicrob Agents Chemother. 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, V.; Conzemius, R.; Varda-Brkić, D.; Bogdan, M.; Grisold, A.; Gyssens, I.C.; Barišić, I. Epidemiology of colistin-resistant, carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Croatia. Infect Genet Evol. 2020, 81, 104263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, C.; Wang, Q.; Wang, X.; Chen, H.; Li, H.; Zhang, F.; Li, S.; Wang, R.; Wang, H. High Prevalence of Hypervirulent Klebsiella pneumoniae Infection in China: Geographic Distribution, Clinical Characteristics, and Antimicrobial Resistance. Antimicrob Agents Chemother. 2016, 60, 6115–6120. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12. 2022. Available online: http://www.eucast.org (accessed on 1 October 2023).
- Clinical Laboratory Standard Institution. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Approved Standard M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2002, 18, 268–281. [Google Scholar] [CrossRef]
- Jarlier, V.; Nicolas, M.H.; Fournier, G.; Philippon, A. Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae: Hospital prevalence and susceptibility patterns. Rev Infect Dis. 1988, 10, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Krilanović, M.; Tomić-Paradžik, M.; Meštrović, T.; Beader, N.; Herljević, Z.; Conzemius, R.; Barišić, I.; Vraneš, J.; Elveđi-Gašparović, V.; Bedenić, B.E. Extended-spectrum β-lactamases and plasmid diversity in urinary isolates of Escherichia coli in Croatia: A nation-wide, multicentric, retrospective study. Folia Microbiol (Praha). 2020, 65, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Black, J.A.; Moland, E.S.; Thomson, K.S. AmpC disk test for detection of plasmid-mediated AmpC beta-lactamases in Enterobacteriaceae lacking chromosomal AmpC β-lactamases. J Clin Microbiol. 2005, 43, 3110–3113. [Google Scholar] [CrossRef] [PubMed]
- Amjad, A.; Mirza, I.; Abbasi, S.; Farwa, U.; Malik, N.; Zia, F. Modified Hodge test: A simple and effective test for detection of carbapenemase production. Iran J Microbiol. 2011, 3, 189–193. [Google Scholar] [PubMed]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge test and the imipenem-EDTA-double-disk synergy test for differentiating metallo-β-lactamase-producing isolates of Pseudomonas spp. and Acinetobacter spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef] [PubMed]
- van der Zwaluw, K.; De Haan, A.; Pluister, G.N.; Bootsma, H.J.; de Neeling, A.J. The Carbapenem Inactivation Method (CIM), a simple and low-cost alternative for the carba NP test to assess phenotypic carbapenemase activity in Gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.W.; Chibabhai, V. Evaluation of the RESIST-4 O.K.N.V immunochromatographic lateral flow assay for the rapid detection of OXA-48, KPC, NDM and VIM carbapenemases from cultured isolates. Access Microbiol. 2019, 1, e000031. [Google Scholar] [CrossRef] [PubMed]
- Arlet, G.; Brami, G.; Decre, D.; Flippo, A.; Gaillot, O.; Lagrange, P.H.; Philippon, A. Molecular characterization by PCR restriction fragment polymorphism of TEM β-lactamases. FEMS Microbiol. Lett. 1995, 134, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Nüesch-Inderbinen, M.T.; Hächler, H.; Kayser, F.H. Detection of genes coding for extended-spectrum SHV β-lactamases in clinical isolates by a molecular genetic method, and comparison with the E test. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 398–402. [Google Scholar] [CrossRef]
- Woodford, N.; Ward, M.E.; Kaufmann, M.E.; Turton, J.; Fagan, E.J.; James, D.; Johnson, A.P.; Pike, R.; Warner, M.; Cheasty, T.; et al. Community and hospital spread of Escherichia coli producing CTX-M extended-spectrum β-lactamases in the UK. J. Antimicrob. Chemother. 2004, 54, 735–743. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef]
- Perez-Perez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuveiller, V.; Nordman, P. Multiplex PCR for detection of acquired carbapenemases genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Tian, G.; Dong, B.; Huang, X.; Yu, L.F.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Gianni, T.; Conte, V.; Di Pilato, V.; Aschbacher, R.; Weber, C.; Rossolini, G.M.L. Escherichia coli from Italy producing OXA-48 carbapenemase encoded by a novel Tn1999 Transposon derivative. Antimicrob Agents Chemother. 2012, 56, 2211–2213. [Google Scholar] [CrossRef]
- Saladin, M.; Cao, V.T.B.; Lambert, T.; Donay, J.L.; Hermann, J.; Ould-Hocine, L. Diversity of CTX-M β-lactamases and their promoter regions from Enterobacteriaceae isolated in three Parisian hospitals. FEMS Microbiol. Lett. 2002, 209, 161–168. [Google Scholar] [CrossRef]
- Elwell, L.P.; Falkow, S. The characterization of R plasmids and the detection of plasmid-specified genes. In Antibiotics in Laboratory Medicine, 2nd ed.; Lorian, V., Ed.; Williams and Wilkins: Baltimore, MD, USA, 1986; pp. 683–721. [Google Scholar]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threfall, E.J. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef]
- Livermore, D.M.; Walsh, T.R.; Toleman, M.; Woodford, N. Balkan NDM-1: Escape or transplant? Lancet Infect Dis. 2011, 11, 164. [Google Scholar] [CrossRef]
- Bedenić, B.; Luxner, J.; Car, H.; Sardelić, S.; Bogdan, M.; Varda-Brkić, D.; Šuto, S.; Grisold, A.; Beader, N.; Zarfel, G. Emergence and Spread of Enterobacterales with Multiple Carbapenemases after COVID-19 Pandemic. Pathogens. 2023, 12, 677. [Google Scholar] [CrossRef]
- Kopotsa, K.; Osei Sekyere, J.; Mbelle, N.M. Plasmid evolution in carbapenemase-producing Enterobacteriaceae: A review. Ann N Y Acad Sci. 2019, 1457, 61–91. [Google Scholar] [CrossRef]
- Bedenić, B.; Sardelić, S.; Bogdanić, M.; Zarfel, G.; Beader, N.; Šuto, S.; Krilanović, M.; Vraneš, J. Klebsiella pneumoniae carbapenemase (KPC) in urinary infection isolates. Arch Microbiol 2021, 4, 1825–1831. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, Y.; Wang, G.; Li, C.; Chang, Y.F.; Chen, W.; Mao, Y.; Zhang, J.; Zhong, F.; Zhang, L. blaNDM-5 carried by a hypervirulent Klebsiella pneumoniae with sequence type 29. Antimicrob Resist Infect Control. 2019, 8, 140. [Google Scholar] [CrossRef] [PubMed]
- Achille, G.; Nunzi, I.; Fioriti, S.; Cirioni, O.; Brescini, L.; Giacometti, A.; Teodori, L.; Brenciani, A.; Giovanetti, E.; Mingoia, M.; et al. Clonal dissemination of Klebsiella pneumoniae carrying blaOXA-48 gene in, a central Italy hospital. J Glob Antimicrob Resist. 2024, 18, S2213–S7165. [Google Scholar] [CrossRef]
- Bedenić, B.; Bratić, V.; Mihaljević, S.; Lukić, A.; Vidović, K.; Reiner, K.; Schöenthaler, S.; Barišić, I.; Zarfel, G.; Grisold, A. Multidrug-Resistant Bacteria in a COVID-19 Hospital in Zagreb. Pathogens 2023, 12, 117. [Google Scholar] [CrossRef]
Figure 1.
Comparison of antibiotic susceptibility between hospital and outpatient setting isolates. Resistant, red; Intermediate, yellow, Susceptible, green. Abbreviations: AMC-amoxycilin/clavulanic acid; TZP-piperacillin/tazobactam; CXM-cefuroxime; CAZ-ceftazidime; CTX-cefotaxime; CRO-ceftriaxone; FEP-cefepime; IMI-imipenem; MEM-meropenem; GM-gentamicin; CIP-ciprofloxacin; COL-colistin; R-resistance, UHCZ-University Hospital Centre Zagreb; UHCSM-University Hospital Centre Sestre Milosrdnice.
Figure 1.
Comparison of antibiotic susceptibility between hospital and outpatient setting isolates. Resistant, red; Intermediate, yellow, Susceptible, green. Abbreviations: AMC-amoxycilin/clavulanic acid; TZP-piperacillin/tazobactam; CXM-cefuroxime; CAZ-ceftazidime; CTX-cefotaxime; CRO-ceftriaxone; FEP-cefepime; IMI-imipenem; MEM-meropenem; GM-gentamicin; CIP-ciprofloxacin; COL-colistin; R-resistance, UHCZ-University Hospital Centre Zagreb; UHCSM-University Hospital Centre Sestre Milosrdnice.
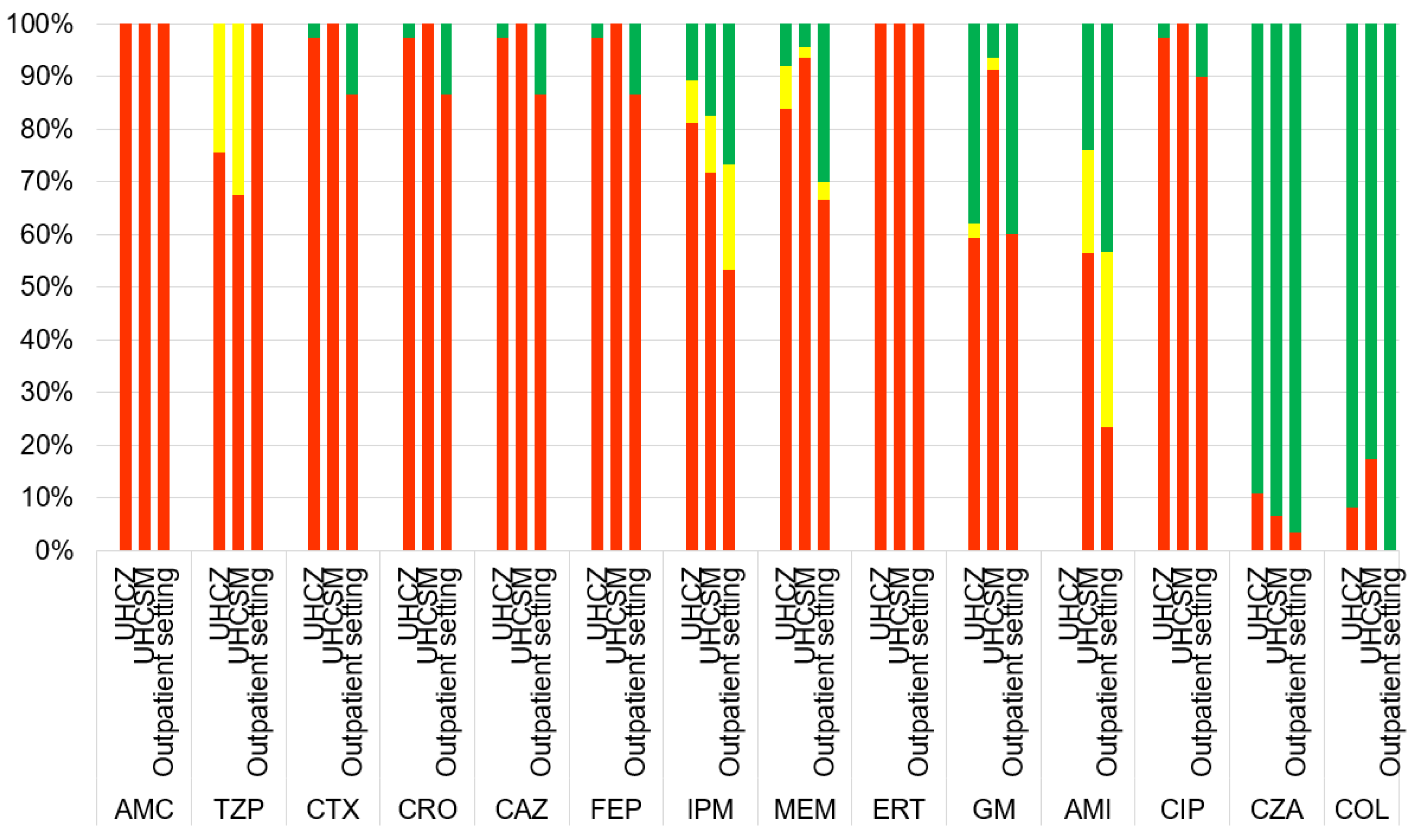
Figure 2.
Distribution of carbapenemases in bacterial isolates according to the sampling location. OXA-48, blue; KPC, brown; NDM, gray; OXA-48 + NDM, yellow.
Figure 2.
Distribution of carbapenemases in bacterial isolates according to the sampling location. OXA-48, blue; KPC, brown; NDM, gray; OXA-48 + NDM, yellow.
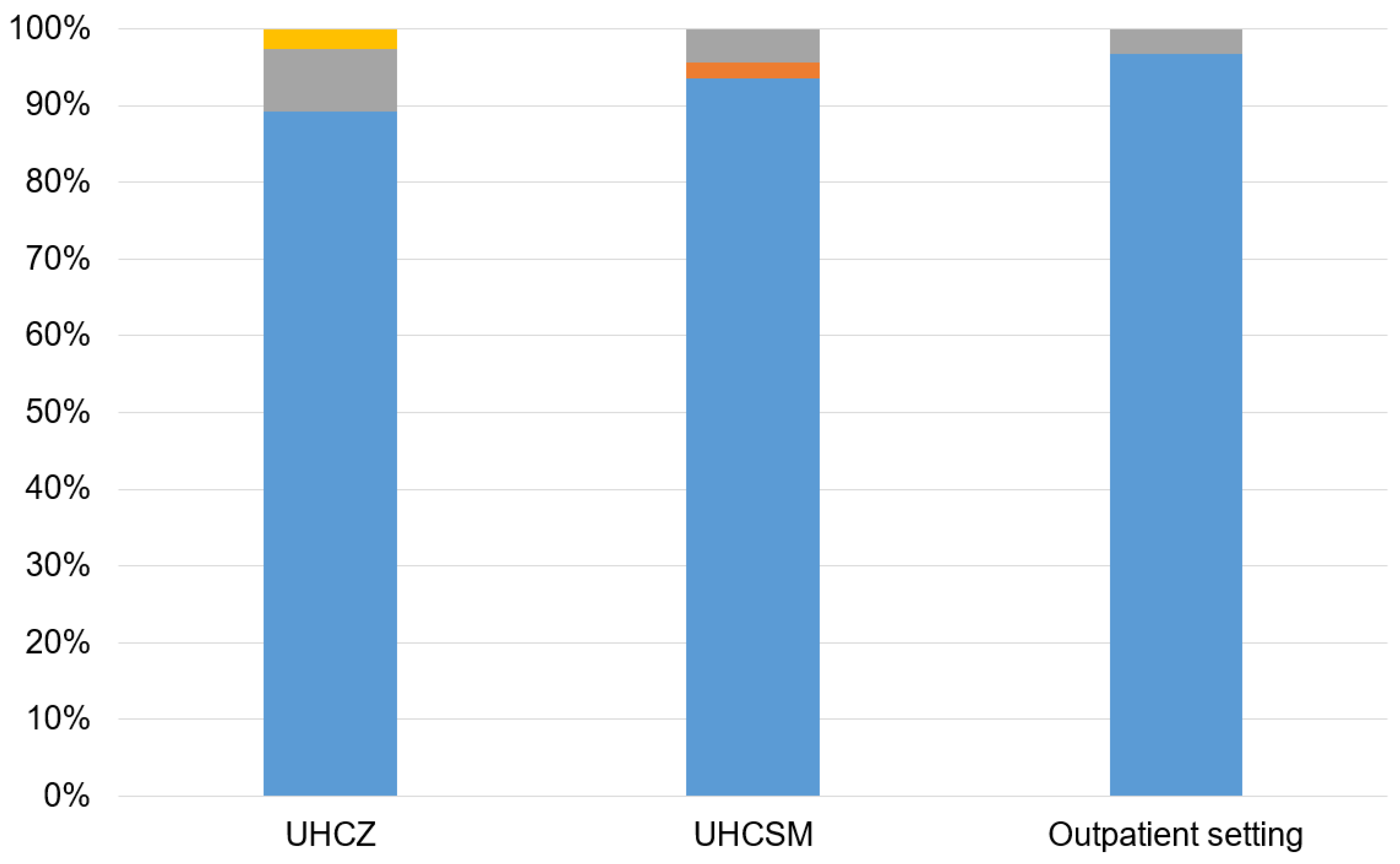

Table 1.
Comparison of antibiotic susceptibility, β-lactamase production and plasmid incompatibility groups of K. pneumoniae isolates between hospital and community isolates.
Table 1.
Comparison of antibiotic susceptibility, β-lactamase production and plasmid incompatibility groups of K. pneumoniae isolates between hospital and community isolates.
| Characteristics | UHCZ | UHCSM | Community isolates |
| Number of isolates | 37 | 46 | 30 |
| Number of ESBL positive isolates | 91.9% (34/37) | 93.5% (43/46) | 83.3% (25/30) |
| Number of AmpC positive isolates | 0.0% (0/37) | 0.0% (0/46) | 0.0% (0/30) |
| HODGE test | 73.0% (27/37) | 76.1% (35/46) | 100.0% (30/30) |
| CIM test | 86.4% (32/37) | 76.1% (35/46) | 100.0% (30/30) |
| AMC -R | 100.0% (37/37) | 100.0% (46/46) | 100.0% (30/30) |
| TZP-R | 100.0% (37/37) | 67.4% (31/46) | 100.0% (30/30) |
| CXM-R | 97.3% (36/37) | 100.0% (46/46) | 86.7% (26/30) |
| CAZ-R | 97.3% (36/37) | 100.0% (46/46) | 86.7% (26/30) |
| CTX-R | 97.3% (36/37) | 100.0% (46/46) | 86.7% (26/30) |
| CRO-R | 97.3% (36/37) | 100.0% (46/46) | 86.7% (26/30) |
| FEP-R | 97.3% (36/37) | 100.0% (46/46) | 86.7% (26/30) |
| IMI-R | 75.7% (28/37) | 71.7% (33/46) | 53.3% (16/30) |
| MEM-R | 78.4% (29/37) | 93.5% (43/46) | 66.7% (20/30) |
| ERT-R | 100.0% (37/37) | 100.0% (46/46) | 100.0% (30/30) |
| GM-R | 62.1% (23/37) | 89.1% (41/46) | 56.7% (17/30) |
| AMI-R | 32.4% (12/37) | 56.5% (26/46) | 16.7% (5/30) |
| CIP-R | 94.6% (35/37) | 100.0% (46/46) | 80.0% (24/30) |
| COL-R | 8.1% (3/37) | 15.2% (7/46) | 0.0% (0/30) |
| CZA-R | 8.1% (3/37) | 17.4% (8/46) | 0.0% (0/30) |
| blaCTX-M | 91.9% (34/37) | 93.4% (43/46) | 83.3% (25/30) |
| blaTEM | 5.4% (2/37) | 15.2% (7/46) | 0.0% (0/30) |
| blaoxa-48 | 91.9% (34/37) | 93.4% (43/46) | 96.7% (29/30) |
| blaKPC | 0.0% (0/37) | 2.2% (1/46) | 0.0% (0/30) |
| BlaNDM | 10.8% (4/37) | 4.3% (2/46) | 3.3% (1/30) |
| String test | 2.7% (1/37) | 4.3% (2/46) | 0.0% (0/30) |
| Inc L plasmid | 48.6% (18/37) | 43.5% (20/46) | 33.3% (10/30) |
| IncY plasmid | 2.7% (1/37) | 8.7% (4/46) | 0.0% (0/30) |
| IncW plasmid | 5.4% (2/37) | 17.4% (8/46) | 0.0% (0/30) |
| IncP plasmid | 0.0% (0/37) | 2.2% (1/46) | 0.0% (0/30) |
Abbreviations: AMC-amoxycilin/clavulanic acid; TZP-piperacillin/tazobactam; CXM-cefuroxime; CAZ-ceftazidime; CTX-cefotaxime; CRO-ceftriaxone; FEP-cefepime; IMI-imipenem; MEM-meropenem; GM-gentamicin; CIP-ciprofloxacin; COL-colistin; R-resistance, UHCZ-University Hospital Centre Zagreb; UHCSM-University Hospital Centre Sestre Milosrdnice.
Table 2.
Interaray chip results of f K. pneumoniae isolates.
| Isolate and protocol number | Centre | β-lactam | AG | SUL | THR | efflux |
|
K. pneumoniae 36 (47168) |
UHCSM |
blaCTX-M-15 blaSHV blaOXA-1 blaOXA-48 |
aac(6’)-Ib | dfrA14 | oqxA, oqxB | |
|
K. pneumoniae 38 (39118) |
UHCSM |
blaCTX-M-15 blaSHV bla0XA-48 |
aac(6’)-Ib | dfrA14 | oqxA, oqxB | |
|
K. pneumoniae 39 (23199) |
UHCSM |
blaCTX-M-15 blaSHV blaOXA-1 blaOXA-48 |
aac(6’)-Ib | dfrA14 | oqxA, oqxB | |
|
K. pneumoniae 40 (152854) |
UHCZ |
blaCTX-M-15 blaSHV blaOXA-1 blaNDM |
aac(6’)-Ib aphA |
sul1 | oqxA, |
Abbreviations: UHCZ – University Hospital Centre Zagreb, UHCSM – University Hospital Centre Sestre Milosrdnice, AG-aminoglycosides; SUL – sulphonamides; THR – trimethoprim.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



