
Submitted:
30 July 2024
Posted:
31 July 2024
You are already at the latest version
Abstract
Idiopathic pulmonary fibrosis, a chronic and fatal condition with no effective cure, is a challenging disease within the group of interstitial lung diseases. The pathogenic mechanisms of this disease are still not fully understood. In this context, the research on miRNAs, which are small non-coding RNAs that regulate messenger RNAs, is of utmost importance. These miRNAs, released into the circulation outside the cell through exosomes, play a crucial role in the pathogenic pathways and mechanisms involved in IPF development. The analysis of miRNAs in the serum/plasma of IPF patients has opened up new possibilities in the search for biomarkers. This review focuses on circulating miRNAs validated by real-time PCR in IPF and the evidence reported about the pulmonary fibrotic process.
Keywords:
microRNAs
; Biomarkers
; Idiopathic pulmonary fibrosis
1. Introduction
Interstitial lung diseases (ILDs) are a complex set of heterogeneous chronic lung diseases that are difficult to diagnose and classify and affect the lung parenchyma due to inflammation and fibrosis. They can be divided into two groups: ILDs of known cause, such as those related to systemic diseases of connective tissue, environmental exposure, radiation, occupational exposure, or allergens, and those classified as Idiopathic Interstitial Pneumonia (IIP). The most common and with the worst prognosis of these is idiopathic pulmonary fibrosis (IPF), which is characterized by progressive remodeling of the lung parenchyma, excess deposition of the extracellular matrix, and irreversible scarring, which prevents an adequate gas exchange [1,2,3].
2. Idiopathic Pulmonary Fibrosis
IPF is a chronic, progressive, and lethal disease with a survivor mean of 3-5 years after diagnosis. The causes and mechanisms of the development of IPF are unknown. It has been proposed that it is characterized by an epithelial-dependent process that starts with micro-injuries over time in alveolar epithelial cells (AECs), with subsequent aberrant activation of these cells. The disruption of the alveolar epithelial barrier causes hyperplasia of pneumocytes type II (cuboidalization) and apoptosis of AECs. The damage of AECs produces many kinds of profibrotic mediators like transforming growth factor beta 1(TGF-β1), connective tissue growth factor (CTGF), platelet-derived growth factor (PDFG), etc., that promote profibrotic micro ambient in the lung of IPF patients. The above promotes an increase in migration, proliferation of lung fibroblasts, and differentiation in myofibroblast (activation). These cells produce excessive quantities of extracellular matrix (ECM) proteins like collagens type I and III, which finalize with the destruction of the pulmonary architecture (Figure 1). [4,5,6,7,8]
Risk factors associated with the development of IPF include a history of smoking, gastroesophageal reflux, chronic viral infections such as Epstein Barr and hepatitis C, a family history of interstitial lung diseases (rare variants in some genes such as MUC5B, TERT SFTPC, etc.), and aging. Symptoms begin insidiously with chronic exertional dyspnea, cough, inspiratory bibasilar crackles, and digital clubbing without symptoms that suggest a multisystem disease[9,10,11].
The natural history of IPF is heterogeneous; it can present chronic stable symptoms up to an acute respiratory exacerbation, with worsening dyspnea from days to weeks, which is associated with a poor prognosis, causing 40% of the deaths in patients with IPF. [12]
One of the presentations of IPF is rapidly progressive, in which patients present a decrease in forced vital capacity (FVC) and diffusion capacity of the lung for carbon monoxide (DLCO) of ≥10% and ≥15%, respectively, within the first 12 months after diagnosis, whose prognosis is severe. The diagnosis of this disease and its differentiation from other ILDs is challenging. It is essential to differentiate it accurately and promptly since inadequate treatment can increase the morbidity and mortality of patients. [10,13]
3. Serum/Plasma Biomarkers in IPF
Several proteins have been measured in the serum of patients with IPF to discover biomarkers to diagnose and prognostic for IPF. It is necessary to find a biomarker due to the difficulty in diagnosing IPF from other interstitial lung diseases like hypersensitivity pneumonitis, sarcoidosis, etc. In this context, some proteins evaluated as possible biomarkers of IPF include surfactant proteins A and D (SP-A and SP-D), Krebs von den lungen 6 ( KL6), C-C motif chemokines (CCL16, CCL18), matrix metalloproteinases (MMP7, and MMP28), renin and soluble (pro) renin receptor. [14,15,16,17]. The only promising molecule was MMP7, which has different levels between IPF and other ILDs. Another kind of molecule that has been measured in the serum of patients is micro RNAs, which are opening new ways to identify biomarkers for IPF (Figure 2).
4. miRNAs, Biogenesis, and Mechanism of Action
miRNAs are small RNAs of a length of 12-18 nucleotides (mature miRNA). This RNA regulates many RNA messengers (mRNA) post-transcriptionally in two ways: inhibition of mRNA translation or inducing its elimination. The miRNAs are non-coding RNAs found in the genome [18]. The miRNAs are transcribed by RNA polymerase II (RNA pol II), generating an immature miRNA termed pri-miRNA, which needs to be processed until it becomes a mature miRNA[19]. The mechanism described to produce a mature miRNA begins when miRNA is transcripted by RNA pol II. This pri-miRNA is processed by the RNAse type III Drosha and DGCR8 (DiGeorge syndrome critical region 8) complex, producing pre-miRNA of the 70 nucleotides in length [20,21,22]. Pre-miRNA is exported outside the nucleus using the nuclear pore complex RAN GTP/exportin 5[23,24]. In the cytoplasm, this pre-miRNA is taken by a complex formed by Dicer/ protein kinase R/ PACT that cut, generating the mature miRNA (Figure 3). Subsequently, mature miRNA is directed to its
RNAm target through the miRISC protein complex that consists of transactivating response RNA binding protein (TRBP) that, in turn, recruits Argonaute 2 (AGO2), followed by the coupling of GW182[25,26,27,28,29]. The miRISC, through AGO2, selects one of the two strands and then is directed to the RNAm target [30].
miRISC leads to the miRNA at the 3’untranslated region (3’UTR) of the RNAm, where the seed region is joined through Watson and Crick complementarity. This process has two results. 1) Depending on the kind of RNAm, the translation of the RNAm will be inhibited either at the level of preventing the formation of the complex of pre-initiation of translation (eIF4G, ribosomal subunit formation) or once the translation process has been started the miRISC complex plus miRNA joined to the 3’UTR region will promote decoupling of the translation machinery[26]. 2) The interaction of miRNA with target RNAm will produce the degradation of RNAm [25,31].
5. Circulation of Exosomes and Their Mechanism of Action
Exosomes can host different biomolecules, including nucleic acids (mRNA and miRNA) and various proteins, including enzymes, receptors, and transcription factors [32,33,34,35,36].
The biogenesis of exosomes begins through the endocytosis of biomolecules inside the cell through the invagination of the plasma membrane, forming the early-staging endosome (ESE)[37]. However, they can follow two routes; the first is to form “recycling endosomes,” and the second consists of subsequently undergoing a process of maturation to become late-classification endosomes (LSE) [38,39], which finally leads to the formation of multivesicular bodies (MVBs). Its formation consists of a second intussusception within the endosomal membrane, thus forming the intraluminal vesicles (ILVs)[40].
Therefore, MVBs are ILV-rich compartments that will eventually become exosomes upon released into the extracellular space. While there must be a fusion between the MVB and the plasma membrane to release the ILVs, there is also degradation by lysosome fusion or autophagosomes (Figure 4) [41,42].
The molecules internalized in exosomes do not appear random; instead, the load is characteristic of the conditions of origin[43]. We will focus on microRNAs because they are in post-transcriptional regulation[44]. The expression levels of these exosomal microRNAs have been reported in pathophysiologies or different tissues of origin[45].
The machinery by which microRNAs are internalized in exosomes and subsequently released depends on the machinery of the endosomal sorting complex required for transport (ESCRT), a protein complex involved in packaging biomolecules in the LVIs of MVBs [46].
6. Upregulated Serum/Plasma miRNAs in Idiopathic Pulmonary Fibrosis
- miR-21
Many circulating miRNAs have been reported in IPF (serum/plasma); some are overexpressed, and others are downregulated. One of the more constant miRNAs with scientific evidence in IPF is miR-21, which has been found elevated in serum/plasma samples from IPF patients inclusive of proposed like predictor prognosis [48,49,50]. miR-21 The TGF-β1 signaling induces a positive loop because it increases the expression of miR-21; it has been reported that the SMADS (2,3,4) regulate the promoter activity of miR-21[51].
- miR-155
Levels of serum miR-155 in IPF are elevated compared with the control group [48,50]. miR-155 is related to the development of pulmonary fibrosis in several models in vitro and in vivo. It has been reported that miR-155 increases in mice bleomycin model and lung fibroblasts increase migration through Keratinocyte Growth Factor[52]. Besides in the silicosis model in mice, miR-155 has profibrotic effects by inhibition of mephrin α; this protein has antifibrotic effects because stimulation with mephrin α decreases expression of TGF-β1, and the receptors of TGF-β1 (TGFBRI and TGFBRII)[53]. The inhibition of miR-155 in a bleomycin model resulted in diminished profibrotic effects that downregulated the TGF-β1 and IL4 (interleukin 4) [54]. In the same way, as in other types of pulmonary fibrosis derived from systemic sclerosis, miR-155 is also elevated and is required for collagen synthesis during the fibrotic process[55]. It has been proposed that this mechanism is promoted by miR-155 inhibition of the protein forkhead box O3 (FOXO3a), resulting in the activation of the NLR family pyrin domain containing 3 (NLRP3)[56]. However, there is contrary evidence about the role of miR-155 in the development of pulmonary fibrosis, where it has been reported that miR-155 inhibits epithelial-mesenchymal transition (EMT) by targeting the glycogen synthase 3 beta (GSK-3β) and its interaction with liver X receptor (LXR) to exert antifibrotic effects [57].
- miR-590-3p
miR-590-3p is elevated in the plasma of patients with IPF, but there is no direct evidence in some models of pulmonary fibrosis. However, reports in other tissues suggest that the effect of this miRNA depends on the location[58]. For example, in heart fibroblasts, miR-590-3p regulates the proliferation, migration, and collagen synthesis by regulating the protein zinc finger E-box-binding homeobox 1(ZEB1) [59]. In contrast, miR-590-3p inhibits the protein mouse double minute 2 homolog (MDM2) in hepatocellular carcinoma, blocking EMT[60]. It is not clear the effect of miR-590-3p in the development of pulmonary fibrosis. Perhaps the effects depend on the location; this effect has been reported in other molecules in IPF, like matrix metalloproteinase 1 (MMP1), where the cellular location is important. MMP1 is over-expressed in epithelial cells of IPF but not in lung fibroblasts, where it is diminished[61]. It has SMAD7 as a target, an important protein in the signaling pathway of TGF-β-1 due to SMAD7 being an inhibitory protein of the SMAD2/3[62,63]. This regulation is crucial because TGF-β1 signaling can trigger several molecular events in IPF, like an increased expression of ECM proteins (collagen I type I, collagen III, etc.), induce fibroblast proliferation, EMT, etc. [64,65,66]. miR-21 has been consistently observed to promote lung fibrosis in fibroblasts and AECs. Another target of miR-21 is the protein phosphatase and tensin homolog (PTEN), which miR-21 induces EMT in a fibrosis model of ionizing radiation[67,68,69]. Resveratrol has been reported as an antifibrotic agent, and miR-21 could reverse the effects of resveratrol in a bleomycin model in rats, and in a contrary sense, treatment in rats with bleomycin diminished the expression of miR-21[70].
- miR-199a-5p and miR-200c
miR-199a-5p had the more consistent results because it was first reported elevated in a bleomycin model of lung fibrosis and tissues from IPF. Second, serum IPF levels of miR-199a-5p are elevated; interestingly, exosomes extracted from urine have the same result[71]. miR-199a-5p is elevated in the mouse bleomycin model, and the reported targets of miR-199a-5p are caveolin-1 and sestrin-2 (SESN2). Loss of caveolin expression has been associated with lung fibrogenesis, and levels of caveolin-1 are downregulated in the lung tissue of IPF patients[72,73,74,75,76]. SESN2 has been reported to have an anti-fibrotic effect in a model of stimuli with eupatilin; TGFβ1 induces the reduction in the production of the protein of SESN2 sestrin2[77]. In addition, miR-199a-5p has been related to regulating the mesenchymal stem cell senescence in IPF[78]. Another effect was reported in liver fibrosis, in which the inhibition of miR-199a-5p diminished fibrosis[79].
miR-199a-5p has targets like activating transcription factor 6 α (ATF6α) and inositol requiring enzyme 1 (IRE1); both have been reported to be sensors of endoplasmic reticulum stress (ERS) [80,81]. Levels of ATF6α and IRE1 are elevated in lung fibrosis (bleomycin and silica models) and in tissue sections of IPF, which is contradictory according to elevated levels of miR-199a-5p. However, in mice models of cardiac fibrosis, the absence of ATF6α in cardiac fibroblasts contributes to the augment of profibrotic fibroblast markers like collagen 1 and alpha-smooth muscle actin in mice model [82].
miR-200c is elevated in serum from IPF patients and is associated with interstitial lung abnormalities[71]. In contrast, miR-200c is reduced in the lung fibrosis of the bleomycin model, and epithelial cells after treatment with TGF-β1 and diminished of miR-200c was related to the development of EMT and loss of ability to alveolar epithelial cells type II to differentiate into alveolar epithelial cells type I [83,84,85]. The process correlates with another target of mir 200c, cadherin 11 (CDH11), which is elevated in lung fibrosis and is related to fibroblast migration, myofibroblast differentiation, and the EMT process during lung injury [86,87,88]. Another example in which reduced miR-200c expression is relevant is because it regulates fibronectin, a component of the MEC that accumulates in the lung fibrotic process[89,90]. miR-200c seems to participate in regulating EMT through ZEB1[91]. These examples suggest that miR-200c levels are downregulated in IPF.
7. Downregulated Serum/Plasma miRNAs in Idiopathic Pulmonary Fibrosis
- Let-7a
- and Let-7d.
Let-7a and Let-7d are downregulated in serum samples from patients with IPF and have the same targets related to the development of lung fibrosis via TGF-β1 signaling pathway like protein high mobility group AT-hook 2 (HMGA2). HMGA2 is a chromatin protein that regulates gene transcription, participating as a co-factor for other transcription factors. Have been reported that let-7a and let-7d could suppress EMT and fibroblast proliferation by blocking HMGA2[71,92,93,94]. In the same signaling pathway, the targets of let-7a and let-7d are the receptors of TGF-β1 (TGFBR1 and TGFBR3) [95,96,97]. Let-7a and let-7d play important roles in developing pulmonary fibrosis by regulating several components of the TGF-β1 signaling pathway.
Then decreased expression levels of let-7a and let-7d suggest that they participate in promoting pulmonary fibrosis in IPF because they have other profibrotic targets like insulin-like growth factor receptor (IGF1R), platelet-derived growth factor B (PDGFB), both receptors are increased in lung fibrosis. Another profibrotic target of let-7a is the activin A receptor type 1B (ACVR1B), which has been implicated in the pathogenesis of cardiac fibrosis through activation by Activin A. Human lung fibroblasts express the receptors of activin A (ACVR1B included) and have a profibrotic effect by promoting collagen contraction, which is a crucial event inside of development of lung fibrosis [98,99,100,101,102].
- miR-16
Serum miR-16 is downregulated in IPF, and this miRNA has antifibrotic activity[103]. miR-16 inhibits the activity of TGF-β1 and collagen I expression in dermal fibroblast; this inhibition is attributed to the target of miR-16 being SMAD3[104]. In hepatic stellate cells, miR-16 suppresses proliferation and fibrogenesis provoked by TGF- β1 regulating lysyl oxidase homolog 1 (LOX L1)[105]. Other evidence about the antifibrotic effects of miR-16 is found in systemic sclerosis, where it suppresses myofibroblast activation by inhibiting neurogenic locus homolog protein 2(NOTCH2) [106]. HMGA1 and HMGA2 also are targets of miR-16, fibroblast growth factor receptor 1 (FGFR-1), IGF1R [107,108,109]
- miR-25-3p and miR-142-5p
miR-25-3p is downregulated in pulmonary fibrosis, but there is no data about its role in lung fibrosis; it has been reported that miR-25-3p expression is increased in cardiac fibrosis, and its expression is related to the augmentation of alpha-smooth muscle actin[110,111]. In a contrary sense, in hepatic stellate cells, the overexpression of miR25-3p has a positive effect because of the Notch 1 signaling pathway and profibrotic results in the induction of collagen expression through TGF-β1 [112]. Additionally, there is a report that the inhibition of miR-25-3p indirectly caused the elevation of TGF-β1. These findings reinforce the relationship between miR-25 and the TGF-β1 signaling pathway [113]. miR-142-5p is downregulated in serum from IPF, but in macrophages from broncho-alveolar lavage fluid (BALF) of patients with IPF, miR-142-5p seems to be elevated, boosting profibrotic actions in vitro like M2 polarization, production of profibrotic cytokines like C-C motif chemokines (CCL18, CCL17, CCL13), TGF-β1, IL4 that in turn influence neighboring fibroblast[50,114].In the same sense, a blank reported of miR-142-5p is the nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor, which is considered antifibrotic because its actions negatively regulate the TGF-β1 signaling pathway[115,116,117]
- miR-101-3p
miR-101-3p is downregulated in serum and lung tissue sections of patients with IPF. This is important because miR-101-3p has been reported to regulate the WNT5a signaling pathway, which reduces proliferation in lung fibroblasts by regulating the receptor frizzled-4,6 (FZD4/6). WNT5a is elevated in IPF, and its signaling pathway has many profibrotic effects, such as inducing fibroblast proliferation, increasing resistance to apoptosis, and increasing fibronectin expression. miR-101-3p also has like blank to the TGFBRI receptor of the TGF-β1 signaling pathway, and it seems that miR101-3p regulates both signaling pathways WNT5a and TGFβ1 in a coordinated manner [50,118,119,120,121]. Another target of miR-101 with profibrotic effects is endothelin-1; this protein is elevated in the serum of patients with IPF and has many profibrotic effects like inducing production of collagen I and III, overproduction of CTGF, fibronectin, α-smooth-muscle actin[122,123,124,125]
8. Common Targets in TGF beta-1 Signaling Pathways
TGF-β1 is a profibrotic molecule that triggers various molecular events in the development of pulmonary fibrosis, and such events affect both alveolar epithelial cells and lung fibroblasts [126]. In the case of epithelial cells, TGF-β1 induces EMT, and activation of the AECs results in the production of many profibrotic molecules like plasminogen activator inhibitor 1 (PAI-1), CTGF, PDGF, etc.[66,101,127,128]. Lung pulmonary fibroblasts affected by the TGF-β1 signaling pathway are activated, which causes an increase in the expression levels of collagen type I and III, induced proliferation, and fibroblast differentiation to myofibroblast (augmentation of α-smooth muscle actin) that is more aggressive in collagen deposition, and this stimulus gives them resistance to apoptosis[129,130,131,132,133,134,135,136].
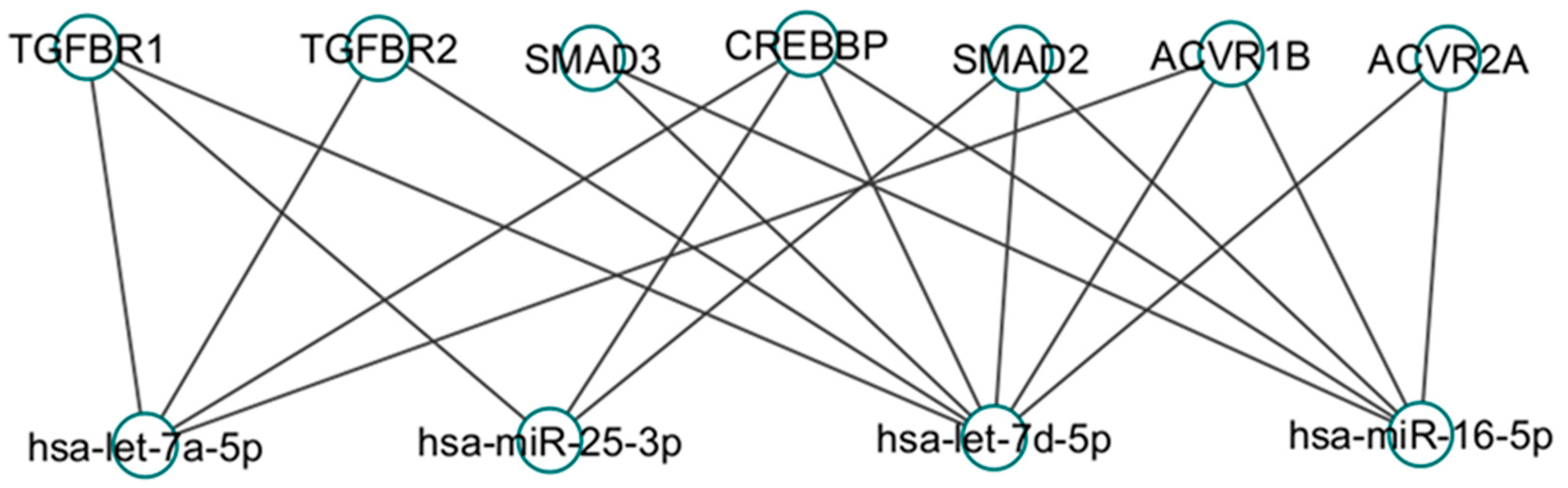
miRNAs let-7a, let-7d, miR-16, and miR-25-3p are downregulated in serum samples from patients with IPF and PCR-validated, according to Tarbase 9.0 is a database to deliver experimentally supported miRNA targets on RNAm from AGO-CLIP-seq protocols that included for example HITS-CLIP, PAR-CLIP. Interestingly, this miRNA shares targets of the TGF-β1 signaling pathway [137]. For example, let-7a, let-7d, miR-16, and miR-25-3p aim at the same target: histone acetyltransferase CREBBP. This protein has been related to promoting an increase in the expression of collagen VI, which is augmented in lung fibrosis and participates in EMT, an important process in developing IPF.[138,139,140]. Let-7d and miR-16 share SMAD3 as a target [70]. SMAD 2 is regulated by let 7d, miR25 and miR 16[141,142]. Let-7d, Let-7a, and miR-25-3p regulate the receptor TGFBR1[97]. Let 7a and Let 7d have like targets of TGFBR2 and AVR1B receptor are regulated by Let 7a, Let 7d and miR16 (Figure 6)[143,144]
9. Discussion and Conclusions
Serum/plasma biomarkers in IPF have been investigated several decades ago[145]. The importance of finding noninvasive biomarkers is due to the characteristics shared between IPF and other ILDs (like hypersensitivity pneumonitis), and therefore, the differential diagnosis is complicated for clinicians[9,146]. miRNAs have emerged as another molecule that could be evaluated in the serum/plasma of IPF patients a few years after the first report about miRNAs and vesicles like exosomes [35,48]. Due to the miRNAs, they could move through the bloodstream protected by exosomes and with protein Argonaute 2 complexes and posteriorly detected by their isolation through the total RNA extraction and its subsequent real-time PCR evaluation. In the field of ILDs, miRNAs in the serum /plasma of IPF patients have only been evaluated compared with the control group[147,148].
Generally, let-7a and let-7d are downregulated, and miR-21 is elevated in IPF. The results have been reported consistently in serum and lung fibroblasts compared to other miRNAs evaluated in serum/plasma. But today, there is no report about serum miRNAs levels in other ILDs like hypersensitivity pneumonitis to find some miRNA differentially expressed with IPF[48,49,50,51,71,92].
The search for miRNAs in serum/plasma is to correlate the expression levels with the clinical characteristics of the patients to find a relationship with the pulmonary state of IPF and to identify a biomarker that could serve as diagnostic or prognostic [149,150]. Additional information obtained indirectly from the serum/plasma miRNAs profile is to link signaling pathways in which they are involved through the in-silico analysis of reported target mRNAs[151]. In addition to the above, the serum miRNA analysis provides new signaling pathways that could participate in the pathogenesis of IPF by using software that predicts new targets of this type of RNA[152].
The contradictory results regarding miRNAs levels in serum/plasma compared with levels in lung tissue sections and fibroblasts from patients with IPF could be explained by the effect of regulation of long non-coding RNAs that have the function of regulating (sequester) the miRNAs[153]. The above responses partially explain why a miRNA is elevated in serum/plasma, but its target is unaffected. Another aspect to consider in regulating the function of miRNAs is the RNA edition mechanism carried out by ADAR proteins. This process could affect 1) the rate of the miRNA processing and redirection of miRNA to another target mRNA[154].
Author Contributions
Conceptualization: E.M.-M. and D.P.R.-R.; writing—original draft preparation, M.D.C.W, X.Q.M., E.M.-M., D.P.R.-R.; writing—review and editing, V.R, E.M.-M. All authors have read and agreed to the published version of the manuscript.
Funding
“This research received no external funding.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nho, R.S. Alteration of Aging-Dependent MicroRNAs in Idiopathic Pulmonary Fibrosis. Drug Dev Res 2015, 76, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Guler, S.A.; Corte, T.J. Interstitial Lung Disease in 2020: A History of Progress. Clin Chest Med 2021, 42, 229–239. [Google Scholar] [CrossRef]
- Samarelli, A.V.; Tonelli, R.; Marchioni, A.; Bruzzi, G.; Gozzi, F.; Andrisani, D.; Castaniere, I.; Manicardi, L.; Moretti, A.; Tabbi, L.; et al. Fibrotic Idiopathic Interstitial Lung Disease: The Molecular and Cellular Key Players. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Selman, M.; Pardo, A. Role of epithelial cells in idiopathic pulmonary fibrosis: from innocent targets to serial killers. Proc Am Thorac Soc 2006, 3, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Velikoff, M.; Canalis, E.; Horowitz, J.C.; Kim, K.K. Activated alveolar epithelial cells initiate fibrosis through autocrine and paracrine secretion of connective tissue growth factor. Am J Physiol Lung Cell Mol Physiol 2014, 306, L786–L796. [Google Scholar] [CrossRef] [PubMed]
- Antoniades, H.N.; Bravo, M.A.; Avila, R.E.; Galanopoulos, T.; Neville-Golden, J.; Maxwell, M.; Selman, M. Platelet-derived growth factor in idiopathic pulmonary fibrosis. J Clin Invest 1990, 86, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Selman, M.; Pardo, A. The leading role of epithelial cells in the pathogenesis of idiopathic pulmonary fibrosis. Cell Signal 2020, 66, 109482. [Google Scholar] [CrossRef] [PubMed]
- Selman, M.; King, T.E.; Pardo, A.; American Thoracic, S.; European Respiratory, S.; American College of Chest, P. Idiopathic pulmonary fibrosis: prevailing and evolving hypotheses about its pathogenesis and implications for therapy. Ann Intern Med 2001, 134, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Remy-Jardin, M.; Richeldi, L.; Thomson, C.C.; Inoue, Y.; Johkoh, T.; Kreuter, M.; Lynch, D.A.; Maher, T.M.; Martinez, F.J.; et al. Idiopathic Pulmonary Fibrosis (an Update) and Progressive Pulmonary Fibrosis in Adults: An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline. Am J Respir Crit Care Med 2022, 205, e18–e47. [Google Scholar] [CrossRef]
- Martinez, F.J.; Collard, H.R.; Pardo, A.; Raghu, G.; Richeldi, L.; Selman, M.; Swigris, J.J.; Taniguchi, H.; Wells, A.U. Idiopathic pulmonary fibrosis. Nat Rev Dis Primers 2017, 3, 17074. [Google Scholar] [CrossRef]
- Pardo, A.; Selman, M. The Interplay of the Genetic Architecture, Aging, and Environmental Factors in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Am J Respir Cell Mol Biol 2021, 64, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Kishaba, T. Acute Exacerbation of Idiopathic Pulmonary Fibrosis. Medicina (Kaunas) 2019, 55. [Google Scholar] [CrossRef] [PubMed]
- Boon, K.; Bailey, N.W.; Yang, J.; Steel, M.P.; Groshong, S.; Kervitsky, D.; Brown, K.K.; Schwarz, M.I.; Schwartz, D.A. Molecular phenotypes distinguish patients with relatively stable from progressive idiopathic pulmonary fibrosis (IPF). PLoS One 2009, 4, e5134. [Google Scholar] [CrossRef] [PubMed]
- Rosas, I.O.; Richards, T.J.; Konishi, K.; Zhang, Y.; Gibson, K.; Lokshin, A.E.; Lindell, K.O.; Cisneros, J.; Macdonald, S.D.; Pardo, A.; et al. MMP1 and MMP7 as potential peripheral blood biomarkers in idiopathic pulmonary fibrosis. PLoS Med 2008, 5, e93. [Google Scholar] [CrossRef] [PubMed]
- Greene, K.E.; King, T.E., Jr.; Kuroki, Y.; Bucher-Bartelson, B.; Hunninghake, G.W.; Newman, L.S.; Nagae, H.; Mason, R.J. Serum surfactant proteins-A and -D as biomarkers in idiopathic pulmonary fibrosis. Eur Respir J 2002, 19, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Hamai, K.; Iwamoto, H.; Ishikawa, N.; Horimasu, Y.; Masuda, T.; Miyamoto, S.; Nakashima, T.; Ohshimo, S.; Fujitaka, K.; Hamada, H.; et al. Comparative Study of Circulating MMP-7, CCL18, KL-6, SP-A, and SP-D as Disease Markers of Idiopathic Pulmonary Fibrosis. Dis Markers 2016, 2016, 4759040. [Google Scholar] [CrossRef] [PubMed]
- Eduardo, M.; Ivette, B.R.; Gabriela, D.P.; Veronica, M.A.; Victor, R. Evaluation of Renin and Soluble (Pro)renin Receptor in Patients with IPF. A Comparison with Hypersensitivity Pneumonitis. Lung 2019, 197, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. MicroRNA maturation: stepwise processing and subcellular localization. EMBO J 2002, 21, 4663–4670. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Radmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, M.T.; Czaplinski, K.; Gorlich, D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.I.; Chendrimada, T.P.; Cooch, N.; Shiekhattar, R. Human RISC couples microRNA biogenesis and posttranscriptional gene silencing. Cell 2005, 123, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, K.; Wakiyama, M.; Yokoyama, S. Mammalian GW182 contains multiple Argonaute-binding sites and functions in microRNA-mediated translational repression. RNA 2009, 15, 1078–1089. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N. The mechanics of miRNA-mediated gene silencing: a look under the hood of miRISC. Nat Struct Mol Biol 2012, 19, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Laine, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a regulator of cellular PKR and HIV-1 virus expression, interacts with Dicer and functions in RNA silencing. EMBO Rep 2005, 6, 961–967. [Google Scholar] [CrossRef]
- Tomari, Y.; Matranga, C.; Haley, B.; Martinez, N.; Zamore, P.D. A protein sensor for siRNA asymmetry. Science 2004, 306, 1377–1380. [Google Scholar] [CrossRef]
- Eulalio, A.; Huntzinger, E.; Izaurralde, E. Getting to the root of miRNA-mediated gene silencing. Cell 2008, 132, 9–14. [Google Scholar] [CrossRef]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Cheng, H.H.; Arroyo, J.D.; Meredith, E.K.; Gallichotte, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc Natl Acad Sci U S A 2014, 111, 14888–14893. [Google Scholar] [CrossRef]
- Lasser, C.; Alikhani, V.S.; Ekstrom, K.; Eldh, M.; Paredes, P.T.; Bossios, A.; Sjostrand, M.; Gabrielsson, S.; Lotvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: uptake by macrophages. J Transl Med 2011, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Didiot, M.C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J Extracell Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: composition, biogenesis and function. Nat Rev Immunol 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Woodman, P.G.; Futter, C.E. Multivesicular bodies: co-ordinated progression to maturity. Curr Opin Cell Biol 2008, 20, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.D.; Donaldson, J.G. Pathways and mechanisms of endocytic recycling. Nat Rev Mol Cell Biol 2009, 10, 597–608. [Google Scholar] [CrossRef]
- Stuffers, S.; Sem Wegner, C.; Stenmark, H.; Brech, A. Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009, 10, 925–937. [Google Scholar] [CrossRef]
- Gruenberg, J.; van der Goot, F.G. Mechanisms of pathogen entry through the endosomal compartments. Nat Rev Mol Cell Biol 2006, 7, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.L.; Urbe, S. The emerging shape of the ESCRT machinery. Nat Rev Mol Cell Biol 2007, 8, 355–368. [Google Scholar] [CrossRef]
- Guduric-Fuchs, J.; O’Connor, A.; Camp, B.; O’Neill, C.L.; Medina, R.J.; Simpson, D.A. Selective extracellular vesicle-mediated export of an overlapping set of microRNAs from multiple cell types. BMC Genomics 2012, 13, 357. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Zernecke, A.; Bidzhekov, K.; Noels, H.; Shagdarsuren, E.; Gan, L.; Denecke, B.; Hristov, M.; Koppel, T.; Jahantigh, M.N.; Lutgens, E.; et al. Delivery of microRNA-126 by apoptotic bodies induces CXCL12-dependent vascular protection. Sci Signal 2009, 2, ra81. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S.; Lim, J.W.; Tauro, B.J.; Ji, H.; Moritz, R.L.; Simpson, R.J. Proteomics analysis of A33 immunoaffinity-purified exosomes released from the human colon tumor cell line LIM1215 reveals a tissue-specific protein signature. Mol Cell Proteomics 2010, 9, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhao, G.Q.; Chen, T.F.; Chang, J.X.; Wang, H.Q.; Chen, S.S.; Zhang, G.J. Serum miR-21 and miR-155 expression in idiopathic pulmonary fibrosis. J Asthma 2013, 50, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Makiguchi, T.; Yamada, M.; Yoshioka, Y.; Sugiura, H.; Koarai, A.; Chiba, S.; Fujino, N.; Tojo, Y.; Ota, C.; Kubo, H.; et al. Serum extracellular vesicular miR-21-5p is a predictor of the prognosis in idiopathic pulmonary fibrosis. Respir Res 2016, 17, 110. [Google Scholar] [CrossRef]
- Li, P.; Li, J.; Chen, T.; Wang, H.; Chu, H.; Chang, J.; Zang, W.; Wang, Y.; Ma, Y.; Du, Y.; et al. Expression analysis of serum microRNAs in idiopathic pulmonary fibrosis. Int J Mol Med 2014, 33, 1554–1562. [Google Scholar] [CrossRef]
- Zhou, J.; Xu, Q.; Zhang, Q.; Wang, Z.; Guan, S. A novel molecular mechanism of microRNA-21 inducing pulmonary fibrosis and human pulmonary fibroblast extracellular matrix through transforming growth factor beta1-mediated SMADs activation. J Cell Biochem 2018, 119, 7834–7843. [Google Scholar] [CrossRef] [PubMed]
- Pottier, N.; Maurin, T.; Chevalier, B.; Puissegur, M.P.; Lebrigand, K.; Robbe-Sermesant, K.; Bertero, T.; Lino Cardenas, C.L.; Courcot, E.; Rios, G.; et al. Identification of keratinocyte growth factor as a target of microRNA-155 in lung fibroblasts: implication in epithelial-mesenchymal interactions. PLoS One 2009, 4, e6718. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, D.; Yao, J.; Wei, Z.; Li, S.; Gao, X.; Cai, W.; Mao, N.; Jin, F.; Li, Y.; et al. Inhibition of miR-155-5p Exerts Anti-Fibrotic Effects in Silicotic Mice by Regulating Meprin alpha. Mol Ther Nucleic Acids 2020, 19, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Kang, Y.; Xue, S.; Zou, J.; Xu, J.; Tang, D.; Qin, H. In vivo therapeutic success of MicroRNA-155 antagomir in a mouse model of pulmonary fibrosis induced by bleomycin. Korean J Intern Med 2021, 36, S160–S169. [Google Scholar] [CrossRef] [PubMed]
- Christmann, R.B.; Wooten, A.; Sampaio-Barros, P.; Borges, C.L.; Carvalho, C.R.; Kairalla, R.A.; Feghali-Bostwick, C.; Ziemek, J.; Mei, Y.; Goummih, S.; et al. miR-155 in the progression of lung fibrosis in systemic sclerosis. Arthritis Res Ther 2016, 18, 155. [Google Scholar] [CrossRef] [PubMed]
- Artlett, C.M.; Sassi-Gaha, S.; Hope, J.L.; Feghali-Bostwick, C.A.; Katsikis, P.D. Mir-155 is overexpressed in systemic sclerosis fibroblasts and is required for NLRP3 inflammasome-mediated collagen synthesis during fibrosis. Arthritis Res Ther 2017, 19, 144. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Z.; Yan, Z.; Liang, X.; Liu, X.; Liu, Y.; Wang, P.; Bai, C.; Gu, Y.; Zhou, P.K. MiRNA-155-5p inhibits epithelium-to-mesenchymal transition (EMT) by targeting GSK-3beta during radiation-induced pulmonary fibrosis. Arch Biochem Biophys 2021, 697, 108699. [Google Scholar] [CrossRef] [PubMed]
- Dirol, H.; Toylu, A.; Ogus, A.C.; Cilli, A.; Ozbudak, O.; Clark, O.A.; Ozdemir, T. Alterations in plasma miR-21, miR-590, miR-192 and miR-215 in idiopathic pulmonary fibrosis and their clinical importance. Mol Biol Rep 2022, 49, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Pan, J.; Wen, L.; Gong, B.; Li, J.; Gao, H.; Tan, W.; Liang, S.; Zhang, H.; Wang, X. MiR-590-3p regulates proliferation, migration and collagen synthesis of cardiac fibroblast by targeting ZEB1. J Cell Mol Med 2020, 24, 227–237. [Google Scholar] [CrossRef]
- Youssef, A.I.; Khaled, G.M.; Amleh, A. Functional role and epithelial to mesenchymal transition of the miR-590-3p/MDM2 axis in hepatocellular carcinoma. BMC Cancer 2023, 23, 396. [Google Scholar] [CrossRef]
- Pardo, A.; Cabrera, S.; Maldonado, M.; Selman, M. Role of matrix metalloproteinases in the pathogenesis of idiopathic pulmonary fibrosis. Respir Res 2016, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Friggeri, A.; Yang, Y.; Milosevic, J.; Ding, Q.; Thannickal, V.J.; Kaminski, N.; Abraham, E. miR-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosis. J Exp Med 2010, 207, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Abdollah, S.; Qiu, Y.; Cai, J.; Xu, Y.Y.; Grinnell, B.W.; Richardson, M.A.; Topper, J.N.; Gimbrone, M.A., Jr.; Wrana, J.L.; et al. The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGFbeta signaling. Cell 1997, 89, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Fine, A.; Goldstein, R.H. The effect of transforming growth factor-beta on cell proliferation and collagen formation by lung fibroblasts. J Biol Chem 1987, 262, 3897–3902. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Du, Y.; Shen, Y.; He, Y.; Zhao, H.; Li, Z. TGF-beta 1 induced fibroblast proliferation is mediated by the FGF-2/ERK pathway. Front Biosci (Landmark Ed) 2012, 17, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforming growth factor-beta1: potential role in idiopathic pulmonary fibrosis. Am J Pathol 2005, 166, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Kubo, H.; Ota, C.; Takahashi, T.; Tando, Y.; Suzuki, T.; Fujino, N.; Makiguchi, T.; Takagi, K.; Suzuki, T.; et al. The increase of microRNA-21 during lung fibrosis and its contribution to epithelial-mesenchymal transition in pulmonary epithelial cells. Respir Res 2013, 14, 95. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liang, X.; Li, X.; Liu, X.; Zhu, M.; Gu, Y.; Zhou, P. MiRNA-21 functions in ionizing radiation-induced epithelium-to-mesenchymal transition (EMT) by downregulating PTEN. Toxicol Res (Camb) 2019, 8, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xiao, T.; Li, J.; Wang, D.; Sun, J.; Cheng, C.; Ma, H.; Xue, J.; Li, Y.; Zhang, A.; et al. miR-21 in EVs from pulmonary epithelial cells promotes myofibroblast differentiation via glycolysis in arsenic-induced pulmonary fibrosis. Environ Pollut 2021, 286, 117259. [Google Scholar] [CrossRef]
- Jin, W.; Chen, F.; Wang, K.; Song, Y.; Fei, X.; Wu, B. miR-15a/miR-16 cluster inhibits invasion of prostate cancer cells by suppressing TGF-beta signaling pathway. Biomed Pharmacother 2018, 104, 637–644. [Google Scholar] [CrossRef]
- Yang, G.; Yang, L.; Wang, W.; Wang, J.; Wang, J.; Xu, Z. Discovery and validation of extracellular/circulating microRNAs during idiopathic pulmonary fibrosis disease progression. Gene 2015, 562, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Kasper, M.; Reimann, T.; Hempel, U.; Wenzel, K.W.; Bierhaus, A.; Schuh, D.; Dimmer, V.; Haroske, G.; Muller, M. Loss of caveolin expression in type I pneumocytes as an indicator of subcellular alterations during lung fibrogenesis. Histochem Cell Biol 1998, 109, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zhang, Y.; Kim, H.P.; Zhou, Z.; Feghali-Bostwick, C.A.; Liu, F.; Ifedigbo, E.; Xu, X.; Oury, T.D.; Kaminski, N.; et al. Caveolin-1: a critical regulator of lung fibrosis in idiopathic pulmonary fibrosis. J Exp Med 2006, 203, 2895–2906. [Google Scholar] [CrossRef] [PubMed]
- Odajima, N.; Betsuyaku, T.; Nasuhara, Y.; Nishimura, M. Loss of caveolin-1 in bronchiolization in lung fibrosis. J Histochem Cytochem 2007, 55, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Lino Cardenas, C.L.; Henaoui, I.S.; Courcot, E.; Roderburg, C.; Cauffiez, C.; Aubert, S.; Copin, M.C.; Wallaert, B.; Glowacki, F.; Dewaeles, E.; et al. miR-199a-5p Is upregulated during fibrogenic response to tissue injury and mediates TGFbeta-induced lung fibroblast activation by targeting caveolin-1. PLoS Genet 2013, 9, e1003291. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yin, E.; Xu, Y.; Liu, Y.; Li, T.; Dong, Z.; Tai, W. CDKN2B antisense RNA 1 expression alleviates idiopathic pulmonary fibrosis by functioning as a competing endogenouse RNA through the miR-199a-5p/Sestrin-2 axis. Bioengineered 2022, 13, 7746–7759. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Lyu, X.; Liu, Y.; Peng, N.; Tan, S.; Dong, L.; Zhang, X. Eupatilin inhibits pulmonary fibrosis by activating Sestrin2/PI3K/Akt/mTOR dependent autophagy pathway. Life Sci 2023, 334, 122218. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Han, Q.; Hong, Y.; Li, W.; Gong, G.; Cui, J.; Mao, M.; Liang, X.; Hu, B.; Li, X.; et al. Inhibition of miR-199a-5p rejuvenates aged mesenchymal stem cells derived from patients with idiopathic pulmonary fibrosis and improves their therapeutic efficacy in experimental pulmonary fibrosis. Stem Cell Res Ther 2021, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Zheng, J.; Li, N.; Cheng, Q.; Zhu, M.; Wang, Y.; Zhou, X.; Zhang, Z.; Shi, G. DZNep, an inhibitor of the histone methyltransferase EZH2, suppresses hepatic fibrosis through regulating miR-199a-5p/SOCS7 pathway. PeerJ 2021, 9, e11374. [Google Scholar] [CrossRef]
- Dai, B.H.; Geng, L.; Wang, Y.; Sui, C.J.; Xie, F.; Shen, R.X.; Shen, W.F.; Yang, J.M. microRNA-199a-5p protects hepatocytes from bile acid-induced sustained endoplasmic reticulum stress. Cell Death Dis 2013, 4, e604. [Google Scholar] [CrossRef]
- Lei, Y.; Yu, H.; Ding, S.; Liu, H.; Liu, C.; Fu, R. Molecular mechanism of ATF6 in unfolded protein response and its role in disease. Heliyon 2024, 10, e25937. [Google Scholar] [CrossRef]
- Stauffer, W.T.; Blackwood, E.A.; Azizi, K.; Kaufman, R.J.; Glembotski, C.C. The ER Unfolded Protein Response Effector, ATF6, Reduces Cardiac Fibrosis and Decreases Activation of Cardiac Fibroblasts. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Banerjee, S.; de Freitas, A.; Sanders, Y.Y.; Ding, Q.; Matalon, S.; Thannickal, V.J.; Abraham, E.; Liu, G. Participation of miR-200 in pulmonary fibrosis. Am J Pathol 2012, 180, 484–493. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Xu, Q.; Yao, W.; Wu, Q.; Yuan, J.; Yan, W.; Xu, T.; Ji, X.; Ni, C. Long non-coding RNA-ATB promotes EMT during silica-induced pulmonary fibrosis by competitively binding miR-200c. Biochim Biophys Acta Mol Basis Dis 2018, 1864, 420–431. [Google Scholar] [CrossRef]
- Moimas, S.; Salton, F.; Kosmider, B.; Ring, N.; Volpe, M.C.; Bahmed, K.; Braga, L.; Rehman, M.; Vodret, S.; Graziani, M.L.; et al. miR-200 family members reduce senescence and restore idiopathic pulmonary fibrosis type II alveolar epithelial cell transdifferentiation. ERJ Open Res 2019, 5. [Google Scholar] [CrossRef]
- Luo, D.; Wilson, J.M.; Harvel, N.; Liu, J.; Pei, L.; Huang, S.; Hawthorn, L.; Shi, H. A systematic evaluation of miRNA:mRNA interactions involved in the migration and invasion of breast cancer cells. J Transl Med 2013, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.J.; Wu, M.; Le, T.T.; Cho, S.H.; Brenner, M.B.; Blackburn, M.R.; Agarwal, S.K. Cadherin-11 contributes to pulmonary fibrosis: potential role in TGF-beta production and epithelial to mesenchymal transition. FASEB J 2012, 26, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Black, M.; Milewski, D.; Le, T.; Ren, X.; Xu, Y.; Kalinichenko, V.V.; Kalin, T.V. FOXF1 Inhibits Pulmonary Fibrosis by Preventing CDH2-CDH11 Cadherin Switch in Myofibroblasts. Cell Rep 2018, 23, 442–458. [Google Scholar] [CrossRef]
- Howe, E.N.; Cochrane, D.R.; Richer, J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res 2011, 13, R45. [Google Scholar] [CrossRef]
- Rennard, S.I.; Crystal, R.G. Fibronectin in human bronchopulmonary lavage fluid. Elevation in patients with interstitial lung disease. J Clin Invest 1982, 69, 113–122. [Google Scholar] [CrossRef]
- Siemens, H.; Jackstadt, R.; Hunten, S.; Kaller, M.; Menssen, A.; Gotz, U.; Hermeking, H. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle 2011, 10, 4256–4271. [Google Scholar] [CrossRef] [PubMed]
- Pandit, K.V.; Corcoran, D.; Yousef, H.; Yarlagadda, M.; Tzouvelekis, A.; Gibson, K.F.; Konishi, K.; Yousem, S.A.; Singh, M.; Handley, D.; et al. Inhibition and role of let-7d in idiopathic pulmonary fibrosis. Am J Respir Crit Care Med 2010, 182, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, X.; Chen, D.; Ma, C. Let-7a suppresses glioma cell proliferation and invasion through TGF-beta/Smad3 signaling pathway by targeting HMGA2. Tumour Biol 2016, 37, 8107–8119. [Google Scholar] [CrossRef] [PubMed]
- Sgarra, R.; Pegoraro, S.; Ros, G.; Penzo, C.; Chiefari, E.; Foti, D.; Brunetti, A.; Manfioletti, G. High Mobility Group A (HMGA) proteins: Molecular instigators of breast cancer onset and progression. Biochim Biophys Acta Rev Cancer 2018, 1869, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhou, H.; Wu, D.; Ni, H.; Chen, Z.; Chen, C.; Xiang, Y.; Dai, K.; Chen, X.; Li, X. MicroRNA let-7a regulates angiogenesis by targeting TGFBR3 mRNA. J Cell Mol Med 2019, 23, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Wen, L.; Peng, R.; Li, H.; Liu, H.; Peng, H.; Sun, Y.; Wu, T.; Chen, L.; Duan, Q.; et al. Naringenin Ameliorated Kidney Injury through Let-7a/TGFBR1 Signaling in Diabetic Nephropathy. J Diabetes Res 2016, 2016, 8738760. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Gao, Y.M.; Zhang, Y.C.; Jia, M.W.; Peng, F.; Meng, Q.H.; Wang, Y.C. Low let-7d exosomes from pulmonary vascular endothelial cells drive lung pericyte fibrosis through the TGFbetaRI/FoxM1/Smad/beta-catenin pathway. J Cell Mol Med 2020, 24, 13913–13926. [Google Scholar] [CrossRef] [PubMed]
- Homma, S.; Nagaoka, I.; Abe, H.; Takahashi, K.; Seyama, K.; Nukiwa, T.; Kira, S. Localization of platelet-derived growth factor and insulin-like growth factor I in the fibrotic lung. Am J Respir Crit Care Med 1995, 152, 2084–2089. [Google Scholar] [CrossRef]
- Wang, L.N.; Chen, W.W.; Zhang, J.; Li, C.Y.; Liu, C.Y.; Xue, J.; Zhang, P.J.; Jiang, A.L. The miRNA let-7a1 inhibits the expression of insulin-like growth factor 1 receptor (IGF1R) in prostate cancer PC-3 cells. Asian J Androl 2013, 15, 753–758. [Google Scholar] [CrossRef]
- Wyss, C.B.; Duffey, N.; Peyvandi, S.; Barras, D.; Martinez Usatorre, A.; Coquoz, O.; Romero, P.; Delorenzi, M.; Lorusso, G.; Ruegg, C. Gain of HIF1 Activity and Loss of miRNA let-7d Promote Breast Cancer Metastasis to the Brain via the PDGF/PDGFR Axis. Cancer Res 2021, 81, 594–605. [Google Scholar] [CrossRef]
- Kasai, H.; Allen, J.T.; Mason, R.M.; Kamimura, T.; Zhang, Z. TGF-beta1 induces human alveolar epithelial to mesenchymal cell transition (EMT). Respir Res 2005, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, Y.; Zhang, P.; Chen, Y.; Li, C.; Chen, J.; Wang, Y.; Li, Y. The crucial role of activin A/ALK4 pathway in the pathogenesis of Ang-II-induced atrial fibrosis and vulnerability to atrial fibrillation. Basic Res Cardiol 2017, 112, 47. [Google Scholar] [CrossRef] [PubMed]
- Lacedonia, D.; Scioscia, G.; Soccio, P.; Conese, M.; Catucci, L.; Palladino, G.P.; Simone, F.; Quarato, C.M.I.; Di Gioia, S.; Rana, R.; et al. Downregulation of exosomal let-7d and miR-16 in idiopathic pulmonary fibrosis. BMC Pulm Med 2021, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Bo, Y.; Liu, B.; Yang, L.; Zhang, L.; Yan, Y. Exosomes derived from miR-16-5p-overexpressing keratinocytes attenuates bleomycin-induced skin fibrosis. Biochem Biophys Res Commun 2021, 561, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, J.; Xiao, E.; Ning, H.; Li, K.; Shang, J.; Kang, Y. MiR-15b and miR-16 suppress TGF-beta1-induced proliferation and fibrogenesis by regulating LOXL1 in hepatic stellate cells. Life Sci 2021, 270, 119144. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Xing, Y.; Wang, Z.; Liang, J.; Lin, Q.; Huang, M.; Chen, Y.; Lin, B.; Xu, X.; Chen, W. MiR-16-5p suppresses myofibroblast activation in systemic sclerosis by inhibiting NOTCH signaling. Aging (Albany NY) 2020, 13, 2640–2654. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, D.; D’Angelo, D.; Valentino, T.; De Martino, I.; Ferraro, A.; Wierinckx, A.; Fedele, M.; Trouillas, J.; Fusco, A. Downregulation of HMGA-targeting microRNAs has a critical role in human pituitary tumorigenesis. Oncogene 2012, 31, 3857–3865. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Jorganes, A.; Araldi, E.; Penalva, L.O.; Sandhu, D.; Fernandez-Hernando, C.; Suarez, Y. MicroRNA-16 and microRNA-424 regulate cell-autonomous angiogenic functions in endothelial cells via targeting vascular endothelial growth factor receptor-2 and fibroblast growth factor receptor-1. Arterioscler Thromb Vasc Biol 2011, 31, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Q.; Wang, G.D.; Wang, H.S.; Huang, Y.; Liu, X.M.; Cai, X.H. miR-16 inhibits cell proliferation by targeting IGF1R and the Raf1-MEK1/2-ERK1/2 pathway in osteosarcoma. FEBS Lett 2013, 587, 1366–1372. [Google Scholar] [CrossRef]
- Min, H.; Fan, S.; Song, S.; Zhuang, Y.; Li, H.; Wu, Y.; Cai, H.; Yi, L.; Dai, J.; Gao, Q. Plasma microRNAs are associated with acute exacerbation in idiopathic pulmonary fibrosis. Diagn Pathol 2016, 11, 135. [Google Scholar] [CrossRef]
- Zeng, N.; Wen, Y.H.; Pan, R.; Yang, J.; Yan, Y.M.; Zhao, A.Z.; Zhu, J.N.; Fang, X.H.; Shan, Z.X. Dickkopf 3: a Novel Target Gene of miR-25-3p in Promoting Fibrosis-Related Gene Expression in Myocardial Fibrosis. J Cardiovasc Transl Res 2021, 14, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Genz, B.; Coleman, M.A.; Irvine, K.M.; Kutasovic, J.R.; Miranda, M.; Gratte, F.D.; Tirnitz-Parker, J.E.E.; Olynyk, J.K.; Calvopina, D.A.; Weis, A.; et al. Overexpression of miRNA-25-3p inhibits Notch1 signaling and TGF-beta-induced collagen expression in hepatic stellate cells. Sci Rep 2019, 9, 8541. [Google Scholar] [CrossRef] [PubMed]
- Hirota, N.; McCuaig, S.; O’Sullivan, M.J.; Martin, J.G. Serotonin augments smooth muscle differentiation of bone marrow stromal cells. Stem Cell Res 2014, 12, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Guiot, J.; Cambier, M.; Boeckx, A.; Henket, M.; Nivelles, O.; Gester, F.; Louis, E.; Malaise, M.; Dequiedt, F.; Louis, R.; et al. Macrophage-derived exosomes attenuate fibrosis in airway epithelial cells through delivery of antifibrotic miR-142-3p. Thorax 2020, 75, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, M.; Patel, D.; Vedpathak, D.; Rathinam, M.; Henderson, G.; Mahimainathan, L. Identification of novel microRNAs in post-transcriptional control of Nrf2 expression and redox homeostasis in neuronal, SH-SY5Y cells. PLoS One 2012, 7, e51111. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yang, H.; Pan, L.; Wei, S.; Li, Z.; Zhang, P.; Li, R.; Wu, Y.; Liu, M.; Liu, X. Ginkgo biloba Extract 50 (GBE50) Exerts Antifibrotic and Antioxidant Effects on Pulmonary Fibrosis in Mice by Regulating Nrf2 and TGF-beta1/Smad Pathways. Appl Biochem Biotechnol 2023. [Google Scholar] [CrossRef] [PubMed]
- Hua, Q.; Ren, L. The SIRT1/Nrf2 signaling pathway mediates the anti-pulmonary fibrosis effect of liquiritigenin. Chin Med 2024, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xiao, X.; Yang, Y.; Mishra, A.; Liang, Y.; Zeng, X.; Yang, X.; Xu, D.; Blackburn, M.R.; Henke, C.A.; et al. MicroRNA-101 attenuates pulmonary fibrosis by inhibiting fibroblast proliferation and activation. J Biol Chem 2017, 292, 16420–16439. [Google Scholar] [CrossRef] [PubMed]
- Vuga, L.J.; Ben-Yehudah, A.; Kovkarova-Naumovski, E.; Oriss, T.; Gibson, K.F.; Feghali-Bostwick, C.; Kaminski, N. WNT5A is a regulator of fibroblast proliferation and resistance to apoptosis. Am J Respir Cell Mol Biol 2009, 41, 583–589. [Google Scholar] [CrossRef]
- Newman, D.R.; Sills, W.S.; Hanrahan, K.; Ziegler, A.; Tidd, K.M.; Cook, E.; Sannes, P.L. Expression of WNT5A in Idiopathic Pulmonary Fibrosis and Its Control by TGF-beta and WNT7B in Human Lung Fibroblasts. J Histochem Cytochem 2016, 64, 99–111. [Google Scholar] [CrossRef]
- Yang, J.; Lu, Y.; Lin, Y.Y.; Zheng, Z.Y.; Fang, J.H.; He, S.; Zhuang, S.M. Vascular mimicry formation is promoted by paracrine TGF-beta and SDF1 of cancer-associated fibroblasts and inhibited by miR-101 in hepatocellular carcinoma. Cancer Lett 2016, 383, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Pulito-Cueto, V.; Genre, F.; Lopez-Mejias, R.; Mora-Cuesta, V.M.; Iturbe-Fernandez, D.; Portilla, V.; Sebastian Mora-Gil, M.; Ocejo-Vinyals, J.G.; Gualillo, O.; Blanco, R.; et al. Endothelin-1 as a Biomarker of Idiopathic Pulmonary Fibrosis and Interstitial Lung Disease Associated with Autoimmune Diseases. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Shahar, I.; Fireman, E.; Topilsky, M.; Grief, J.; Schwarz, Y.; Kivity, S.; Ben-Efraim, S.; Spirer, Z. Effect of endothelin-1 on alpha-smooth muscle actin expression and on alveolar fibroblasts proliferation in interstitial lung diseases. Int J Immunopharmacol 1999, 21, 759–775. [Google Scholar] [CrossRef]
- Rizvi, M.A.; Katwa, L.; Spadone, D.P.; Myers, P.R. The effects of endothelin-1 on collagen type I and type III synthesis in cultured porcine coronary artery vascular smooth muscle cells. J Mol Cell Cardiol 1996, 28, 243–252. [Google Scholar] [CrossRef]
- Marini, M.; Carpi, S.; Bellini, A.; Patalano, F.; Mattoli, S. Endothelin-1 induces increased fibronectin expression in human bronchial epithelial cells. Biochem Biophys Res Commun 1996, 220, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Hu, Y. TGFbeta1: Gentlemanly orchestrator in idiopathic pulmonary fibrosis (Review). Int J Mol Med 2021, 48. [Google Scholar] [CrossRef]
- Shi, L.; Dong, N.; Fang, X.; Wang, X. Regulatory mechanisms of TGF-beta1-induced fibrogenesis of human alveolar epithelial cells. J Cell Mol Med 2016, 20, 2183–2193. [Google Scholar] [CrossRef]
- Rana, T.; Jiang, C.; Liu, G.; Miyata, T.; Antony, V.; Thannickal, V.J.; Liu, R.M. PAI-1 Regulation of TGF-beta1-induced Alveolar Type II Cell Senescence, SASP Secretion, and SASP-mediated Activation of Alveolar Macrophages. Am J Respir Cell Mol Biol 2020, 62, 319–330. [Google Scholar] [CrossRef]
- Heine, U.I.; Munoz, E.F.; Flanders, K.C.; Roberts, A.B.; Sporn, M.B. Colocalization of TGF-beta 1 and collagen I and III, fibronectin and glycosaminoglycans during lung branching morphogenesis. Development 1990, 109, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Grande, J.P.; Melder, D.C.; Zinsmeister, A.R. Modulation of collagen gene expression by cytokines: stimulatory effect of transforming growth factor-beta1, with divergent effects of epidermal growth factor and tumor necrosis factor-alpha on collagen type I and collagen type IV. J Lab Clin Med 1997, 130, 476–486. [Google Scholar] [CrossRef]
- Kenyon, N.J.; Ward, R.W.; McGrew, G.; Last, J.A. TGF-beta1 causes airway fibrosis and increased collagen I and III mRNA in mice. Thorax 2003, 58, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Krupsky, M.; Fine, A.; Kuang, P.P.; Berk, J.L.; Goldstein, R.H. Regulation of type I collagen production by insulin and transforming growth factor-beta in human lung fibroblasts. Connect Tissue Res 1996, 34, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Masta, S.; Meyers, D.; Narayanan, A.S. Collagen synthesis by normal and fibrotic human lung fibroblasts and the effect of transforming growth factor-beta. Am Rev Respir Dis 1989, 140, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Desmouliere, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J Cell Biol 1993, 122, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Phan, S.H. Inhibition of myofibroblast apoptosis by transforming growth factor beta(1). Am J Respir Cell Mol Biol 1999, 21, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Montesano, R.; Orci, L. Transforming growth factor beta stimulates collagen-matrix contraction by fibroblasts: implications for wound healing. Proc Natl Acad Sci U S A 1988, 85, 4894–4897. [Google Scholar] [CrossRef] [PubMed]
- Skoufos, G.; Kakoulidis, P.; Tastsoglou, S.; Zacharopoulou, E.; Kotsira, V.; Miliotis, M.; Mavromati, G.; Grigoriadis, D.; Zioga, M.; Velli, A.; et al. TarBase-v9.0 extends experimentally supported miRNA-gene interactions to cell-types and virally encoded miRNAs. Nucleic Acids Res 2024, 52, D304–D310. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M.; McCann, F.E.; Cabrita, M.A.; Layton, T.; Cribbs, A.; Knezevic, B.; Fang, H.; Knight, J.; Zhang, M.; Fischer, R.; et al. Identifying collagen VI as a target of fibrotic diseases regulated by CREBBP/EP300. Proc Natl Acad Sci U S A 2020, 117, 20753–20763. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Liu, Y.; Kahn, M.; Ann, D.K.; Han, A.; Wang, H.; Nguyen, C.; Flodby, P.; Zhong, Q.; Krishnaveni, M.S.; et al. Interactions between beta-catenin and transforming growth factor-beta signaling pathways mediate epithelial-mesenchymal transition and are dependent on the transcriptional co-activator cAMP-response element-binding protein (CREB)-binding protein (CBP). J Biol Chem 2012, 287, 7026–7038. [Google Scholar] [CrossRef] [PubMed]
- Specks, U.; Nerlich, A.; Colby, T.V.; Wiest, I.; Timpl, R. Increased expression of type VI collagen in lung fibrosis. Am J Respir Crit Care Med 1995, 151, 1956–1964. [Google Scholar] [CrossRef]
- Diaz-Pina, G.; Rubio, K.; Ordonez-Razo, R.M.; Barreto, G.; Montes, E.; Becerril, C.; Salgado, A.; Cabrera-Fuentes, H.; Aquino-Galvez, A.; Carlos-Reyes, A.; et al. ADAR1 Isoforms Regulate Let-7d Processing in Idiopathic Pulmonary Fibrosis. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Xie, H.; Jin, Q.; Zhao, X.; Shi, Y.; Zhou, Y.; Hu, Z.; Ye, Y.; Huang, X.; Sun, Y.; et al. Extracellular vesicles derived from neural EGFL-Like 1-modified mesenchymal stem cells improve acellular bone regeneration via the miR-25-5p-SMAD2 signaling axis. Bioact Mater 2022, 17, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Chen, Y.; Suo, L.; Chen, H.; Zhu, L.; Wan, G.; Han, X. Activin a promotes myofibroblast differentiation of endometrial mesenchymal stem cells via STAT3-dependent Smad/CTGF pathway. Cell Commun Signal 2019, 17, 45. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.D.; Plumers, R.; Fischer, B.; Schmidt, V.; Hendig, D.; Kuhn, J.; Knabbe, C.; Faust, I. Activin A-Mediated Regulation of XT-I in Human Skin Fibroblasts. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Liu, C.; Tan, C.; Zhang, J. Predictive biomarkers of disease progression in idiopathic pulmonary fibrosis. Heliyon 2024, 10, e23543. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Remy-Jardin, M.; Ryerson, C.J.; Myers, J.L.; Kreuter, M.; Vasakova, M.; Bargagli, E.; Chung, J.H.; Collins, B.F.; Bendstrup, E.; et al. Diagnosis of Hypersensitivity Pneumonitis in Adults. An Official ATS/JRS/ALAT Clinical Practice Guideline. Am J Respir Crit Care Med 2020, 202, e36–e69. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A 2011, 108, 5003–5008. [Google Scholar] [CrossRef]
- Moldovan, L.; Batte, K.; Wang, Y.; Wisler, J.; Piper, M. Analyzing the circulating microRNAs in exosomes/extracellular vesicles from serum or plasma by qRT-PCR. Methods Mol Biol 2013, 1024, 129–145. [Google Scholar] [CrossRef]
- Tomos, I.; Roussis, I.; Matthaiou, A.M.; Dimakou, K. Molecular and Genetic Biomarkers in Idiopathic Pulmonary Fibrosis: Where Are We Now? Biomedicines 2023, 11. [Google Scholar] [CrossRef]
- Stainer, A.; Faverio, P.; Busnelli, S.; Catalano, M.; Della Zoppa, M.; Marruchella, A.; Pesci, A.; Luppi, F. Molecular Biomarkers in Idiopathic Pulmonary Fibrosis: State of the Art and Future Directions. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: deciphering microRNA function with experimental support. Nucleic Acids Res 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Quillet, A.; Saad, C.; Ferry, G.; Anouar, Y.; Vergne, N.; Lecroq, T.; Dubessy, C. Improving Bioinformatics Prediction of microRNA Targets by Ranks Aggregation. Front Genet 2019, 10, 1330. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yang, Y.; Liu, L. Interaction of long noncoding RNAs and microRNAs in the pathogenesis of idiopathic pulmonary fibrosis. Physiol Genomics 2015, 47, 463–469. [Google Scholar] [CrossRef]
- Shevchenko, G.; Morris, K.V. All I’s on the RADAR: role of ADAR in gene regulation. FEBS Lett 2018, 592, 2860–2873. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Model of developing IPF. A) Normal alveolar epithelium composed of alveolar epithelial.
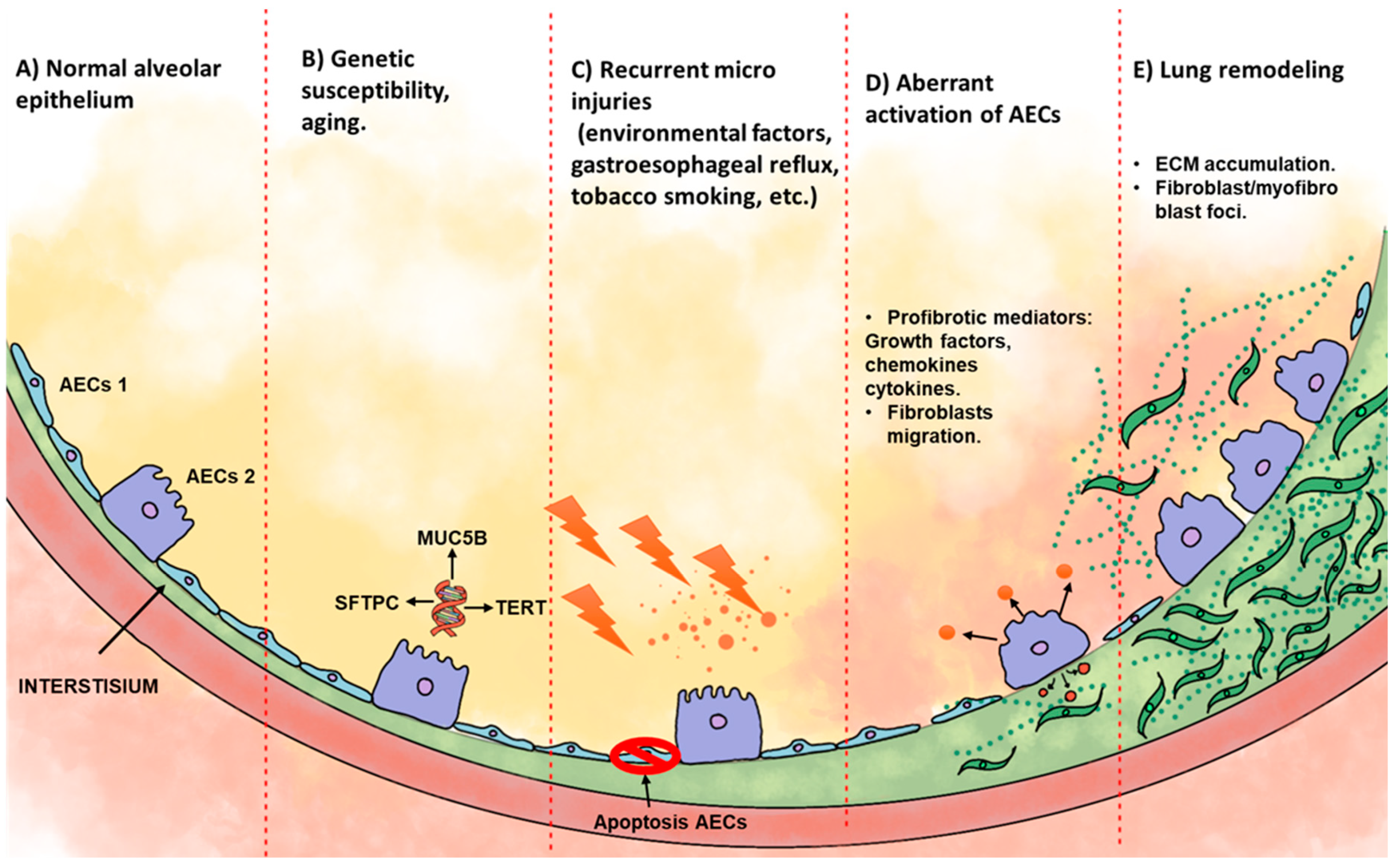
Figure 2.
The miRNAs reported in the serum/plasma of patients with IPF are summarized below; the list includes those miRNAs with real-time PCR-validated.
Figure 2.
The miRNAs reported in the serum/plasma of patients with IPF are summarized below; the list includes those miRNAs with real-time PCR-validated.
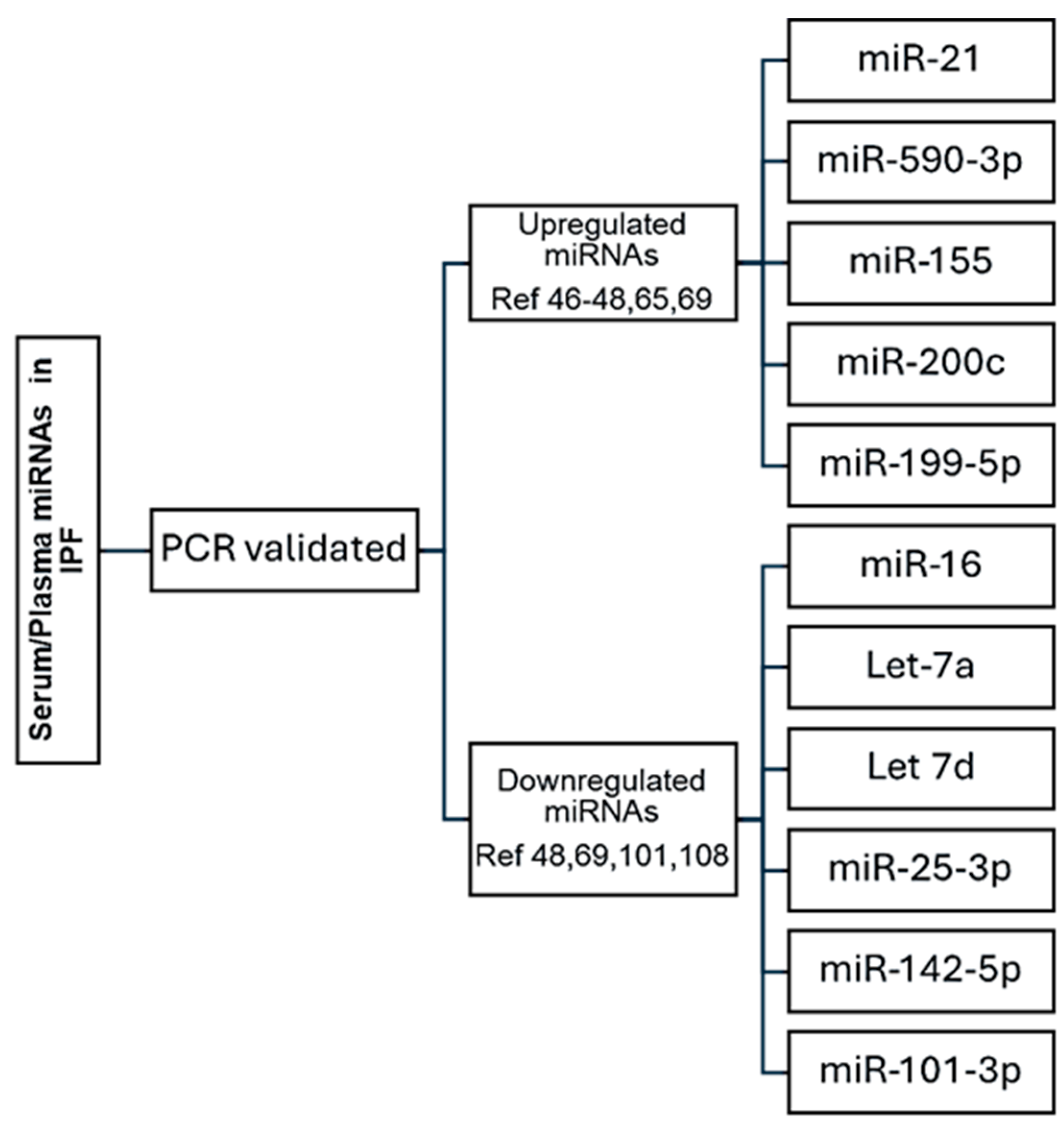
Figure 3.
miRNA biogenesis pathway. 1) The miRNA gene is transcribed through RNA polymerase II/III; this transcript is named pri-miRNA. 2) The pri-miRNA is matured by the protein complex formed by Drosha/DGCR8, producing a pre-miRNA. 3) The pre-miRNA is directed from the nucleus via the nuclear pore complex exportin-5 to the cytoplasm. 4) The Dicer/TRBP complex processes the pre-miRNA. 5) The mature miRNA is directed to its target mRNA through the RNA-induced silencing complex (miRISC), formed by the miRNA and the AGO2 protein.
Figure 3.
miRNA biogenesis pathway. 1) The miRNA gene is transcribed through RNA polymerase II/III; this transcript is named pri-miRNA. 2) The pri-miRNA is matured by the protein complex formed by Drosha/DGCR8, producing a pre-miRNA. 3) The pre-miRNA is directed from the nucleus via the nuclear pore complex exportin-5 to the cytoplasm. 4) The Dicer/TRBP complex processes the pre-miRNA. 5) The mature miRNA is directed to its target mRNA through the RNA-induced silencing complex (miRISC), formed by the miRNA and the AGO2 protein.
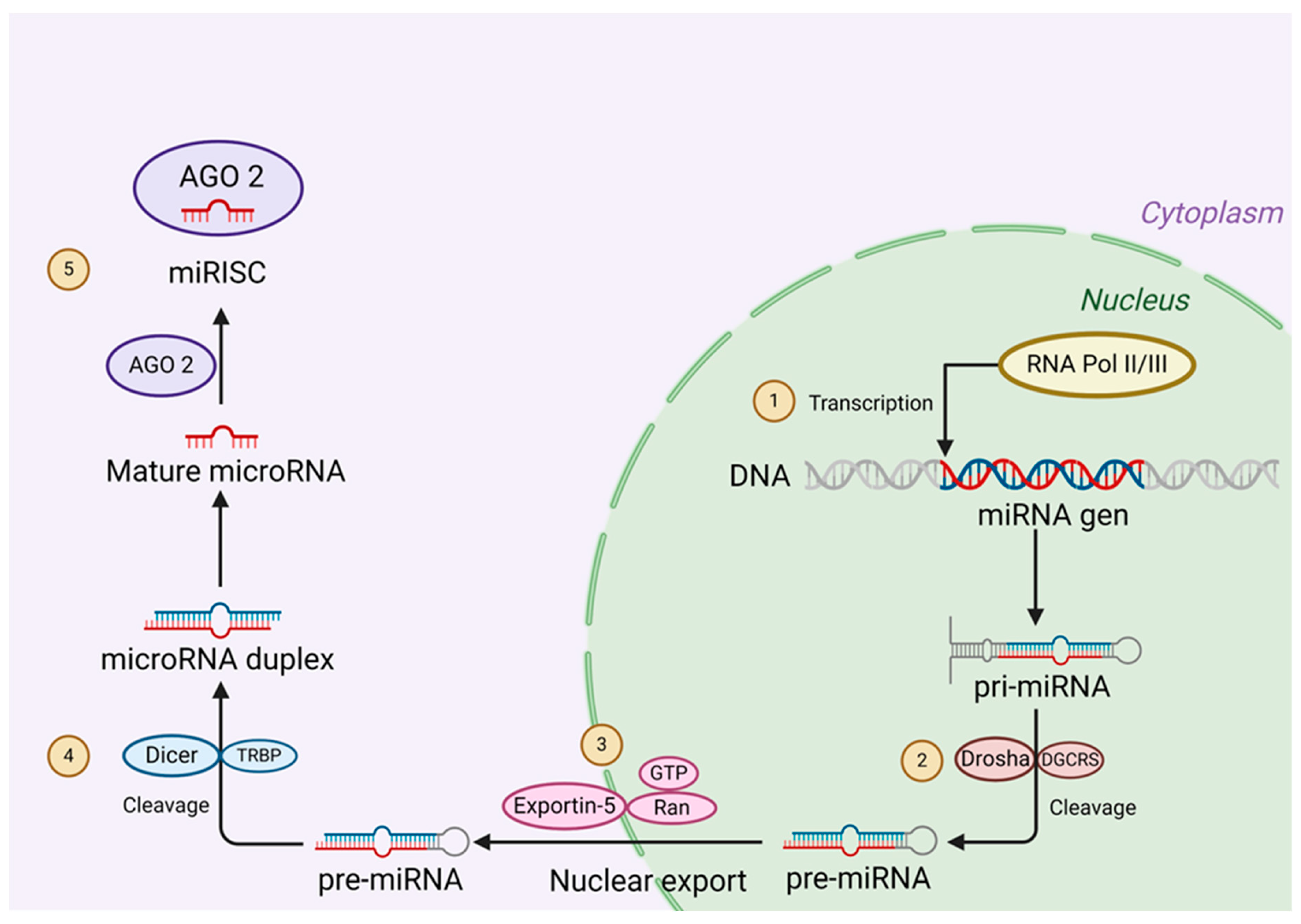
Figure 4.
1, 2) The biogenesis of exosomes begins with the endocytosis of biomolecules, forming the early endosome (ESE). 3, 4) Formation of late endosome (LSE), 5) maturation of multivesicular bodies (MVB), and formation of intraluminal vesicles (ILVs). 6) Fusion of ILVs with the plasma membrane and release of ILVs to the extracellular space as exosomes or 7) degraded by fusion with lysosomes or autophagosomes.
Figure 4.
1, 2) The biogenesis of exosomes begins with the endocytosis of biomolecules, forming the early endosome (ESE). 3, 4) Formation of late endosome (LSE), 5) maturation of multivesicular bodies (MVB), and formation of intraluminal vesicles (ILVs). 6) Fusion of ILVs with the plasma membrane and release of ILVs to the extracellular space as exosomes or 7) degraded by fusion with lysosomes or autophagosomes.
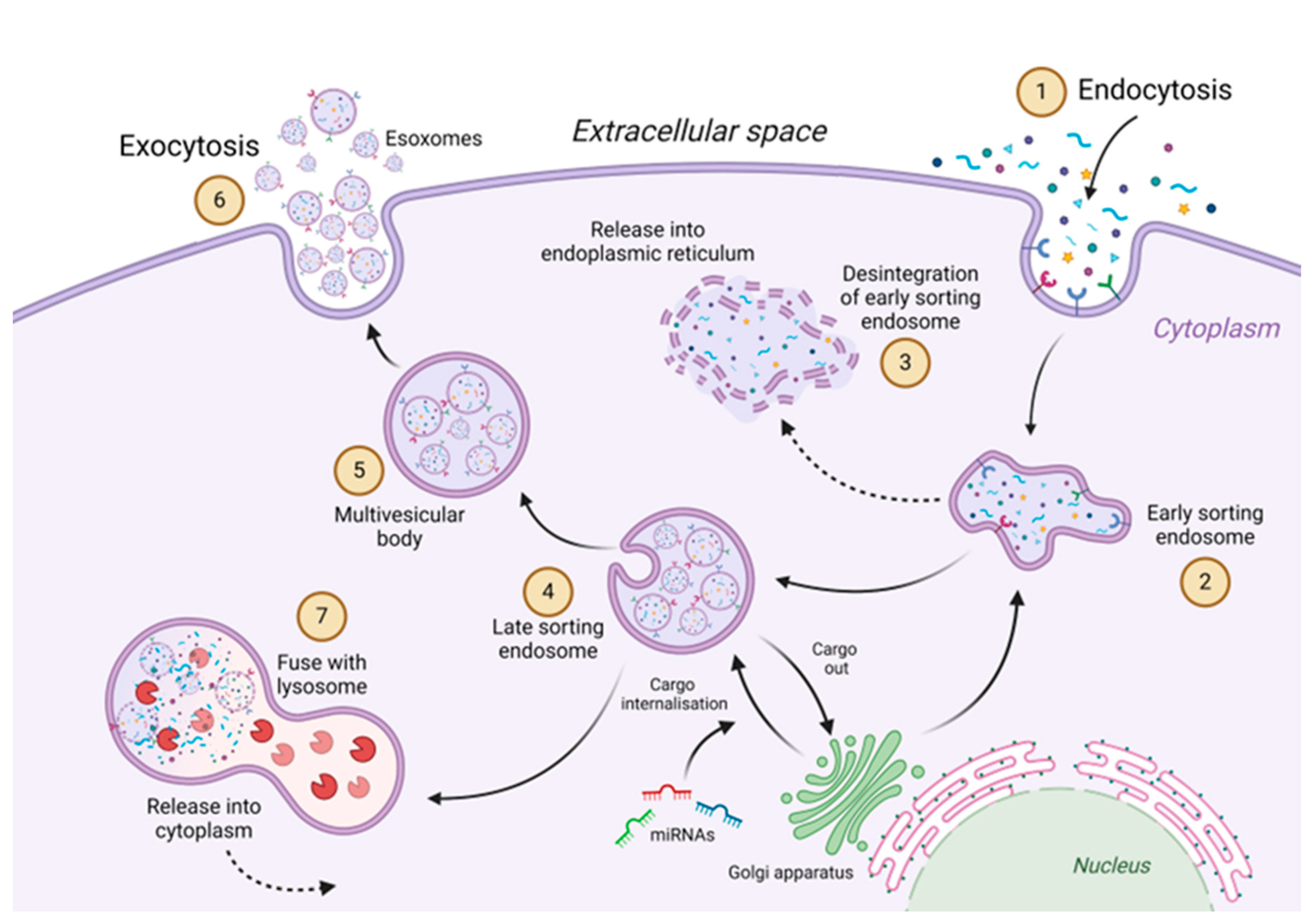
Figure 5.
Release of exosomes. 1) Exosomes are released into the extracellular space and could internalize their contents to the recipient cell through 2) Endocytosis, 3) membrane fusion between the exosome and the recipient cell, and 4) receptor-ligand interaction. 5 and 6) The miRNAs released from the exosome will continue in the miRNA pathway, binding to the AGO2 protein (miRISC), where the miRNA will be directed to the target mRNA.
Figure 5.
Release of exosomes. 1) Exosomes are released into the extracellular space and could internalize their contents to the recipient cell through 2) Endocytosis, 3) membrane fusion between the exosome and the recipient cell, and 4) receptor-ligand interaction. 5 and 6) The miRNAs released from the exosome will continue in the miRNA pathway, binding to the AGO2 protein (miRISC), where the miRNA will be directed to the target mRNA.
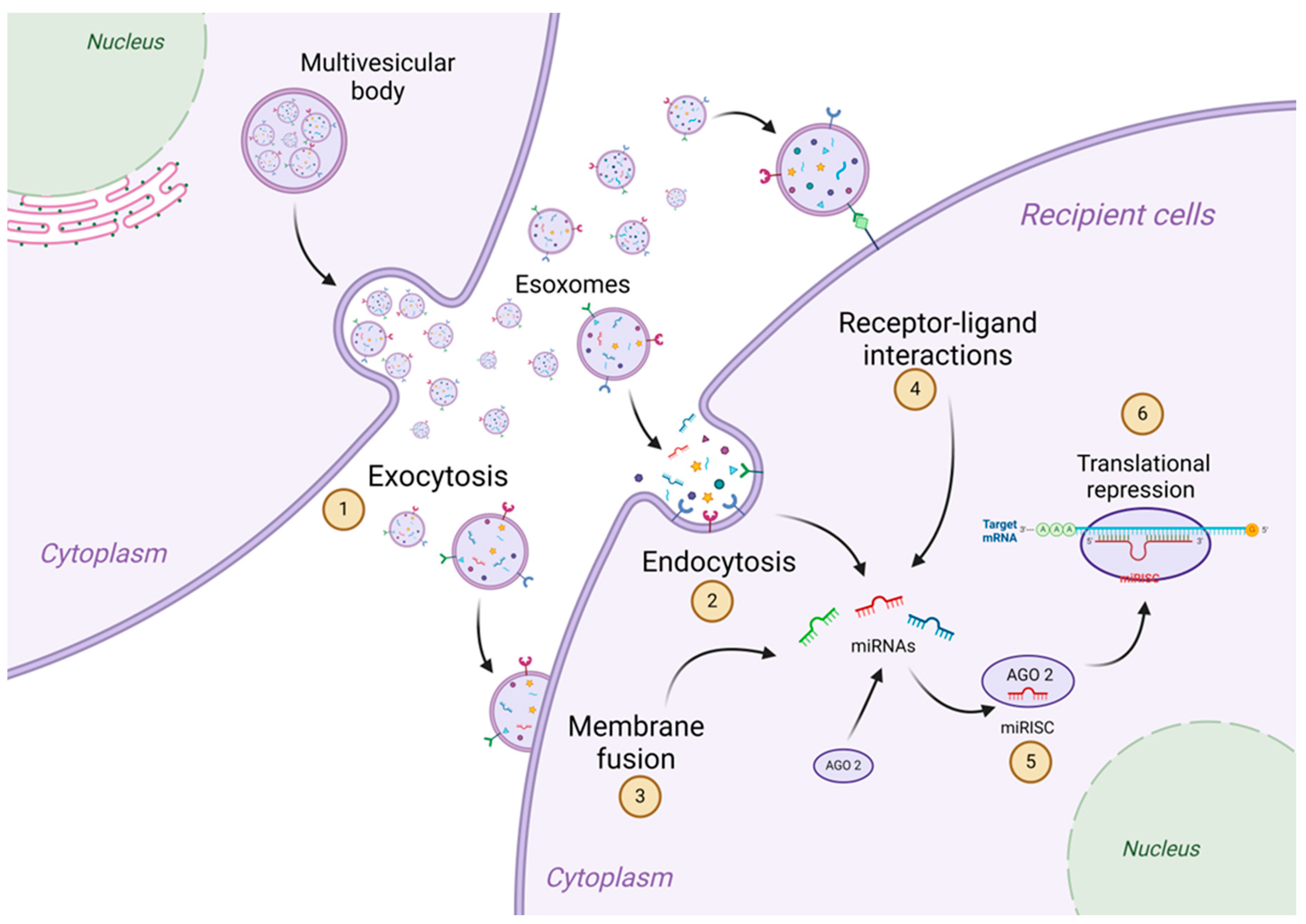
Figure 6.
Four miRNAs decreased in the serum of patients with IPF share target messenger RNAs that have an important role in developing IPF. .
Figure 6.
Four miRNAs decreased in the serum of patients with IPF share target messenger RNAs that have an important role in developing IPF. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



