Submitted:
23 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Interactions between human immunodeficiency virus type 1 (HIV-1) and the host factors or restriction factors of its target cells determine the cell’s susceptibility to, and outcome of, infection. Factors intrinsic to the cell are involved at every step of HIV-1’s replication cycle, contributing to productive infection and replication, or severely attenuating HIV-1’s chances of success. Furthermore, factors unique to certain cell types contribute to the differences in infection between these cell types. Understanding the involvement of these factors in HIV-1 infection is a key requirement for the development of anti-HIV-1 therapies. As the list of factors grows, and the dynamic interactions between these factors and the virus are elucidated, comprehensive and up-to-date summaries that recount the knowledge gathered after decades of research are beneficial to the field, displaying what is known so that researchers can build off the groundwork of others to investigate what is unknown. Herein, we aim to provide a review focusing on protein host factors, both well-known and relatively new, that impact HIV-1 replication in a positive or negative manner at each stage of the replication cycle, highlighting factors unique to HIV-1’s various target cell types where appropriate.
Keywords:
Human immunodeficiency virus (HIV-1)
; host factors
; restriction factors
; replication
; infection
; CD4+ T lymphocytes
; myeloid cells
; monocytes
; macrophages
1. Attachment
Human immunodeficiency virus type 1 (HIV-1) is a positive sense, single-stranded (ss) RNA virus. Its spherical virions are comprised of two copies of the viral genome in the center, around which is the protein capsid, which itself is enclosed by an enveloped derived from the host cell from which the virion budded. The HIV-1 genome consists of five genes for structural and regulatory proteins, Gag, Pol, Env, Tat, and Rev; four genes for accessory proteins Nef, Vif, Vpu, and Vpr; and is flanked on both ends by a 5’ and 3’ Long Terminal Repeat (LTR) [1,2]. From the surface of the envelope the viral surface glycoprotein gp120 protrudes, connected to the viral membrane via the transmembrane protein gp41. gp120 is the predominant facilitator of interaction between the virion and HIV-1’s target cells, binding the target receptor to enable a conformational change of gp41 that initiates fusion of the viral envelope with the host cell plasma membrane (PM), releasing the viral capsid into the cytoplasm [3,4,5,6].
The host cell surface protein CD4 is the primary receptor with which gp120 interacts, and is expressed on a wide variety of immune cells, such as CD4+ T lymphocytes (T cells) and myeloid lineage cells such as macrophages, microglia, and dendritic cells (DCs); CD4 is much more prevalent on CD4+ T cells than myeloid cells. [6,7,8,9]. While CD4-independent infection has been demonstrated in vitro, such as via an endocytotic route, the infection is far less efficient than in CD4+ T cells; interestingly, HIV-1 has been seen to bind to CD4-negative cells for cell-cell transfer into T cells for productive infection [10,11,12,13] CD4+ T cells constitute the main target for HIV-1 infection, being the most commonly infected cell type circulating in the blood, with HIV-1 preferentially infecting activated or memory T cells over naïve or quiescent T cells [14,15,16,17,18]. However, activation of T cells is not required for infection to occur [19,20]; indeed, infection in resting T cells is a source of the long-term viral reservoirs that pose a challenge to complete viral clearance [21,22].
Myeloid cells comprise another major target for HIV-1; monocytes, macrophages (both monocyte-derived and tissue resident), microglia, and dendritic cells, have all been observed to be productively infected by HIV-1 [23,24,25,26,27,28,29]. As noted above, myeloid cells have less CD4 available than CD4+ T cells for HIV-1 to bind to [9,30], requiring a higher binding affinity of gp120 for CD4; this constitutes the first major barrier to HIV-1 infection of myeloid cells [31,32,33].
While the presence of CD4 is critical for the binding of HIV-1 to its target cell, the virus requires a co-receptor for the complete membrane-membrane fusion between virion and host cell [12,34]. Once Env binds to CD4, it undergoes a conformational change that engages gp120 with a co-receptor on the host cell surface [3,6,8]. The two main co-receptors used by HIV-1 are the surface proteins CCR5 and CXCR4 [35,36,37,38]. Those HIV-1 variants that use only CCR5 as a co-receptor are denoted as R5 viruses; those that use only CXCR4 are deemed X4 viruses; and there exists variants able to use both CCR5 and CXCR4 aptly categorized as R5X4 viruses [39].
CCR5 is expressed on memory CD4+ T cells, macrophages, microglia, and dendritic cells [9,26,40,41], and is the co-receptor used by “transmitted/founder” (T/F) viruses passed from one individual to another to establish initial infection [42,43,44,45]. Memory T cells express CCR5 at higher levels than naïve T cells [9,45,46,47], and thus are the predominantly infected cells during early infection. Despite the presence of CCR5 on myeloid cells, not all R5-tropic viruses are capable of infecting cells of the myeloid lineage due to the low density of CD4 on these cells; only viruses with a high affinity for CD4 can effectively bind and infect myeloid cells. In fact, it has been found that X4 viruses can also infect both monocyte-derived macrophages (MDMs) and tissue resident macrophages despite their low levels of CXCR4, although replication is less efficient and apoptosis occurs at much higher rates when compared to M-tropic virus infection of MDMs, implying that CD4 affinity, more than co-receptor usage, determines the ability to infect different cell types [48,49,50]. Thus, those that can infect macrophages are deemed “M-tropic”, and the rest are T cell tropic (T-tropic) [51].
As infection continues, CD4+ T cell levels deplete [52,53,54]. T cell populations of long-infected individuals is predominated by naïve and resting T cells, which express high CXCR4 and low CCR5 levels [45]. This change in co-receptor abundance provides selection pressure onto HIV-1, and indeed a rise in X4 T-tropic virus is seen to correlate with chronic infection [46,55,56], as well as in vitro when CCR5-mediated binding is inhibited [57]. Memory and naïve T cells also express L-selectin (CD62L), which can bind gp120 to facilitate fusion and aid their preferential infection of these T cell subsets [58,59].
Dendritic cells express CD4, as well as CCR5 and CXCR4, allowing for virion Env binding to the DC surface, although the low levels of CD4 expression limits efficient CD4-binding to M-tropic variants [26,60]. However, DCs also possess various other receptors that mediate HIV-1 binding, such as the mannose receptor (MR) and DC-SIGN, circumventing the requirement for CD4 affinity [61]. While productive infection of dendritic cells is less common and less efficient than infection of CD4+ T cells [62], DCs provide an effective transmission route to T cells via “trans-infection”, preferentially to activated CD4+ T cells [25,63].
2. Fusion
Fusion of the HIV-1 envelope with the plasma membrane of the host cell requires more than the initial interaction between Env and CD4/CCR5/CXCR4, and various host factors play a part in ensuring a successful fusion occurs. Intercellular Adhesion Molecule 1 (ICAM-1, or CD54) is a glycoprotein expressed on the surface of epithelial and immune cells, and is found to be incorporated in the viral envelope during budding. ICAM-1 incorporation was found to increase the virion infectivity of both primary and lab-generated HIV-1 strains, and also reduces sensitivity to neutralization from anti-Env glycoprotein antibodies; notably this did come at the cost of being sensitive to anti-ICAM-1 antibodies. However, the use of ICAM-1 during fusion appears to be dependent on transmitter and receiver cell-types; similar rates of transmission were observed when comparing virions with or without ICAM-1 that budded from macrophages or DCs to be received by CD4+ T cells [64,65,66]. Adaptor Protein Complex 2 (AP-2) is a clathrin-associated adaptor protein complex involved in endocytosis of the host cell’s surface receptors, and so to plays a role in HIV-1 envelope fusion. The cytosolic domain of gp41 contains two tyrosine-based motifs, and a nearby Glycine residue, that can interact with the mu-2 domain of AP-2 and facilitate internalization of the virion particle [67,68]. A recent study, based on the idea that intracellular signaling prompted by gp120 binding plays a role in HIV-1 fusion, implicated externalized membrane lipid phosphatidylserine (PS), controlled by the lipid scrambalase transmembrane protein 16F (TMEM16F), in contributing to successful fusion. Binding of gp120 to CD4 triggered Ca2+-dependent signaling of TMEM16F, which in turn causes the externalization of PS; blocking PS externalization or inhibiting TMEM16F function resulted in severely attenuated viral fusion [69]. PS found on the surface of virions, acquired from the virion’s prior budding from an earlier infected cell, are a major factor for efficient macrophage infection. Blockage of virion-associated PS by annexin V severely attenuated infection of MDMs and monocytic cell lines but had no effect on CD4+ T cells or T cell lines; binding of virions to MDMs/monocytic cell lines was not impacted, showing that PS does not play a role in binding, but rather in a subsequent fusion step involving a PS receptor on the cell surface [70].
Preventing virion fusion is the first chance a cell has to avoid HIV-1 infection. The interferon-induced transmembrane (IFITM) protein family has demonstrated antiviral activity at multiple steps in the HIV-1 replication cycle, first and foremost by inhibiting entry. In particular, IFITMs 2 and 3 have greater effects against virion entry than IFITM1, most likely attributable to their relatively extended N-terminal region [71]. However, co-receptor usage is a determinant of HIV-1 sensitivity to the different IFITMs; CCR5-using HIV-1, the transmitter/founder viruses, are more sensitive to inhibition by IFITM1 and more resistant to IFITMs 2 and 3, whereas CXCR4-utlizing HIV-1 is more sensitive to IFITMs 2 and 3 than to IFITM1. This is linked to the sub-cellular localization of each IFITM, with IFITM1 being located on the cell surface and IFITMs 2 and 3 in endosomal compartments [72,73,74]. Consistent with this is a study reporting that IFITM3’s anti-entry activity is possibly due to its disruption of endosomal cholesterol homeostasis, creating environments less conducive to enveloped-virion fusion and thus preventing entry [75]. Interestingly, IFITMs can be transferred via extracellular vesicles (EVs) from infected T cells to uninfected T cells that have yet to have their intrinsic immune response stimulated, thus supplying nearby cells with defenses for the oncoming invasion of virions [76].
Ironically, sometimes prevention of HIV-1 entry has the best chance of occurring after HIV-1 has already infected, replicated in, and budded out from, another host cell. During this process, IFITMs can be incorporated into virions, and can enact their anti-entry activities from within [77]. In the same vein, the PM-localized host restriction factor SERINC5 (and to a lesser extent, SERINC3) is incorporated into the virion, and has at least two observed antiviral effects from this position: impairment of fusion pore formation, and an increased sensitivity to neutralizing antibodies. These effects vary in severity depending on the HIV-1 isolate, as some strains’ Envs show enhanced resistance to SERINC5 [78,79,80,81]. Similar to SERINC proteins, P-selectin glycoprotein ligand-1 (PSGL-1) is a host protein highly active during inflammation normally found on leukocytes to enable attachment onto, and migration into, endothelial tissues. PSGL-1 acts as a restriction factor after being packaged into HIV-1 virions, as it then inhibits the ability of the virion to infect the next target cell; its inhibitory effects stem from its extracellular N-terminal region, as deletion of this domain removes the block against virion entry [82,83].
3. Capsid Transport and Uncoating
After membrane fusion, the conical viral capsid is released to the cytosol where it must gain entry to the nucleus. HIV-1’s ability to transport its genome across an intact nuclear membrane is a key factor in productive infection of non-dividing cells, such as resting CD4+ T cells and macrophages, removing the requirement for nuclear dissolution during cell division that other retroviruses may require [84]. While past models of HIV-1 replication involved the capsid uncoating in the cytosol or at the nuclear pore [85,86,87], there is mounting evidence that the capsid may be transported to the nucleus and enters before completely uncoating [88,89,90,91,92]. Beyond facilitating genome transportation, the capsid shields the viral genome from host restriction factors and sensors of foreign DNA that would otherwise raise the alarm of the innate immune system [93,94].
A wide variety of host factors facilitate successful capsid transport, import to the nucleus, and uncoating for viral genome release (Figure 1). For instance: Sec24c and PDZD8 bind the capsid to enhance capsid stability [95,96]; BICD2 and FEZ1, adaptor proteins for the microtubule (MT)-associated proteins dynein and kinesin-1, respectively, bind to the capsid to expedite cytoplasmic trafficking via MTs [97,98,99,100]; Nup358 and Nup 153 mediate nuclear import [101,102]; and transportin-1 (TNPO1), transportin-3 (TNPO3), and Pin1 can all be found in the nucleus, and trigger capsid destabilization and complete uncoating [103,104,105]. It should be noted that while interactions with MTs have been established as an important method of capsid transport to the nucleus, MT-independent trafficking is still a viable route for HIV-1 [106]. Sufficient capsid stability ensures not only proper interactions with these host factors, but prevents recognition of the viral genome by host foreign DNA sensors. Cyclophilin A (CypA) is a host factor with peptidyl prolyl cis-trans isomerase activity, regulates protein folding and trafficking, and is found intracellularly but can be secreted in response to inflammatory signals [107]. CypA is packaged into the virion during viral assembly, binding the HIV-1 Gag polyprotein via the capsid protein (CA) , and has been shown to be a crucial host factor for infectivity [111]. Researchers report that CypA promotes capsid stabilization during transport across the cell [104,112,113,114,115,116] and may provide resistance against DNA sensors and antiviral factors such as TRIM5α [117,118]. Some researchers suggest that CypA is involved in destabilization of the capsid for proper genome release, possibly in a cell type-specific manner [113,119,120]. In either case, CypA is shown to be beneficial to HIV-1 replication, as virions lacking the protein lead to decreased infection productivity. Inositol hexakisphosphate (IP6) was also recently characterized as a host factor packaged into the HIV-1 virion, promoting capsid stability and preventing recognition by DNA sensors [121,122]. Cleavage and polyadenylation specificity factor 6 (CPSF6) is another crucial CA-binding host factor, as it regulates nuclear entry through nuclear pore complexes (NPCs), as well as localization of the pre-integration complex (PIC, discussed later) to open chromatin, the sites of active transcription, ensuring successful generation of vial mRNA [123,124,125,126,127,128,129].
Multiple host restriction factors work to impede HIV-1 capsid functions (Figure 1). Tripartite motif-containing protein 5α (TRIM5α) is an E3-ubituitin ligase and interferon (IFN)-stimulated gene (ISG) that promotes premature virion disassembly, preventing reverse transcription from completing and allowing recognition and destruction of the unprotected viral genome. However, TRIM5α’s antiviral properties function in a species-specific manner; TRIM5α of old world monkeys was found to be effective against HIV-1, being able to bind the capsid and cause uncoating, whereas human TRIM5α in unable to effectively bind the HIV-1 capsid [130,131,132,133,134]. Exceptions to TRIM5α’s species-specific antiviral activity include virions that insufficiently package CypA prior to infection [118], and TRIM5α-mediated degradation via autophagy in Langerhans cells (a subset of dendritic cells located in mucosal epithelia of genitals and considered a barrier against pathogens) [135]. TRIM5α also plays a role in the immune sensing of HIV-1 by forming a “TRIM5 cage” that leads to auto-ubiquitination, promoting viral capsid detection and stimulation of the innate immune response, as well as proteasomal-mediated degradation [136,137,138]. Likewise, cGAS adaptor protein PQBP1 was recently described to enhance immune detection by binding the capsid, facilitating viral genome detection by cGAS once the genome is released [139,140], and another TRIM family protein, TRIM11, causes premature capsid uncoating [141]. MX2, also known as MXB, is an ISG found both within and around the nucleus that binds the virion capsid and prevents nuclear import. MX2 may also inhibit uncoating, preventing viral genome release [142,143,144,145,146,147]. The restriction factor Daxx, found mainly in the nucleus, also inhibits capsid uncoating by interacting with both CypA and CA, in a SUMOylation-dependent manner [148].
4. Reverse Transcription
During the capsid’s trek from membrane to nucleus, the viral genome must be reverse transcribed by RT from positive-sense ssRNA to double-stranded (ds) DNA. The process of reverse transcription, particularly an intermediate step involving first-strand transfer, is shown to be critical for capsid uncoating to occur, as inhibition of RT produced extra-stable capsids that could not disassemble and release their payloads [85,149,150]. The space within the capsid is known as the viral core, and packaged along with the vial genome are the viral RT, nucleocapsid protein (NC), integrase (IN), and Viral Protein R (Vpr); post-fusion, some matrix protein (MA) also associates with the capsid, although whether or not it is incorporated inside the core is unknown. Together these components comprise the reverse transcription complex (RTC) of HIV-1 [151]. Cellular proteins that benefit HIV-1 like CypA, and restriction factors like APOBEC3G (discussed below) are also found within the capsid [152]. Pores in the capsid allow deoxynucleotide triphosphates (dNTPs) to enter and provide the building blocks for RT to create viral cDNA [153]. Reverse transcription is carried out almost entirely by RT’s DNA polymerase and RNAse H abilities, with NC being involved in a strand-transfer step in the process [154].
The host cell cytoskeleton, particularly the actin microfilament network, plays an important role in initiating reverse transcription. MA interacts with actin soon after entry to localize the RTC to the microfilaments, after which efficient reverse transcription occurs. Disruption of actin or the MA-actin interactions directly impacts abundance of newly generated viral DNA [155]. Many host factors that ensure efficient reverse transcription are often packaged into the virion to assist in the process prior to uncoating. Histone deacetylase 1 (HDAC1) is packaged into HIV-1 virions by virtue of the complex it forms with another host factor, sin3A-associated protein 18 (SAP18); SAP18’s interacts with IN, thus leading to the SAP18-HDAC1 complex’s virion packaging. HDAC1 promotes optimal levels of reverse transcription, as knockdown or silencing of HDAC1 results in decreased amounts of reverse transcripts [156,157]. As its name suggests, integrase-interactor 1 (INI1) binds IN and is implicated in efficient RT. When infected with HIV-1, the INI1-deficient rhabdoid cell line MON had reduced virion production, with the virions that were produced displaying reduced infectivity; this debilitation was tracked to the reverse transcription step, as RT productivity was decreased in cells infected with INI1-deficient virions compared to those containing INI1 [158]. Topoisomerase I (TOP1) is a packaged host factor that breaks and rejoins single strands of DNA, relieving tension. TOP1 interacts with the NC of the RTC to ensure efficient RT activity, with inhibition of TOP1 by camptothecin showing a marked reduction in viral cDNA [159]. The RNA helicase Up-frameshift protein 1 (UPF1), ubiquitously found in cells as part of the mRNA Nonsense-Mediated Decay (NMD) pathway, is also packaged in virions and is involved in maximizing the quantity of reverse transcripts produced, a quality linked to its ATPase activity [160,161].
The mRNA-stabilizing ELAV-like protein HuR, while not observed to be packaged into HIV-1 virions, was shown to positively modulate RT activity by protein-protein interaction with RT’s RNase H domain; silencing of HuR resulted in impaired reverse transcription [162]. Similarly, A-kinase anchoring protein 149 (AKAP149, AKAP1), which normally associates with protein kinase A (PKA) to spatially regulate its activity, also interacts with RT via its RNase H region. Silencing of AKAP149 impairs HIV-1 replication at the reverse transcription step, however AKAP149’s exact role in the process is not yet known, though it may relate to its usual associations with PKA, which has also shown to impact HIV-1 replication [163,164]. Survival motor neuron (SMN)-interacting protein 1 (Gemin2) is a mediator of spliceosomal small nuclear ribonucleoprotein (snRNP) assembly, and binds to IN to positively regulate reverse transcription in MDMs, most likely by promoting reassembly of the RTC for continuous production of viral cDNA [165,166]. Interactions between these factors and viral proteins that would theoretically be isolated within capsid lends credence to the possibility that while capsid uncoating may occur in full within the nucleus, as recent evidence suggests (see previous section), it may begin to some degree in the cytoplasm and allow host factor access to these internal proteins.
Reverse transcription’s role as a trigger for capsid uncoating also create more targets of opportunity for the host cell to exploit; without the capsid to act as a bulwark, the threat of viral RNA or DNA detection by cellular sensors becomes a major factor in the viral replication cycle. TREX1 is an exonuclease that degrades HIV-1’s DNA, thus preventing immune sensing of the newly made viral genome [167,168,169]. ADAR1 is an RNA-editing ISG that was found to have pro-virus effects; ADAR1 can edit viral RNA by deaminating adenosine and thus hide it from sensors like MDA5 and RIG-I that would otherwise detect it, and also function as an inhibitor of PKR, an antiviral ISG (discussed below) [170,171,172]. Once the transition to dsDNA is completed, the association of genome and proteins is dubbed the pre-integration complex (PIC). Host factor ADAM10 possess a nuclear localization signal and interacts with the IN of the PIC, and is thought to facilitate shuttling of the PIC to and possibly through the nuclear pore complex, in macrophages and quiescent CD4+ T lymphocytes; this supports the original notion that, to some degree, capsid uncoating occurs, or at least begins, prior to breaching the nucleus [92,173,174,175]. IN is also seen to interact with importin α3 (karyopherin α4) to facilitate PIC nuclear localization and importation [176,177].
Various host restriction factors work to interfere with the creation of viral cDNA and PIC activities. In CD4+ T cells and monocytes/macrophages, the sensing of viral dsRNA is facilitated by RIG-I (with some evidence possibly implicating MDA5), which signals through MAVS to induce an interferon response [178,179,180,181]. Monocytes and macrophages also show viral sensing of ssRNA via toll-like receptor (TLR) 8 [182]. IFI16 and cGAS recognize vial dsDNA and activate the stimulator of interferon genes (STING) pathway, although there is some debate over cGAS’s role [183,184,185,186,187].While myeloid dendritic cells can use cGAS to detect HIV-1, this is dependent on capsid-CypA interactions [188,189]. Plasmacytoid dendritic cells can sense HIV-1 RNA through TLR7 in an endocytosis-dependent manner [190]. Regardless of the RNA/DNA sensing pathway, the end result is expression of interferons that in turn promote expression of antiviral ISGs that work to overcome viral infection [191,192]. APOBEC3 proteins are ISGs found in high levels in nonpermissive cell types, RNA editors that work as cytidine deaminases and introduce excessive mutations, such as premature STOP codons, in the newly reverse transcribed viral DNA. By virtue of binding viral RNA, APOBEC3 proteins are often packaged into virions during assembly to continue harassment of HIV-1 post-budding. Although most members of the APOBEC3 family have anti-HIV-1 abilities to varying degrees, the most prominent seem to be 3G and 3F [193,194,195,196,197,198,199,200,201,202,203,204,205]. Some reports show APOBEC3 antiviral activities are not solely linked to their mutagenic capabilities, and may in fact be attributed to directly binding to RT and acting as a roadblock to reverse transcription [206,207,208,209,210]. PSGL-1 is also an ISG that, outside of its anti-fusion effects, is postulated to negatively impact HIV-1 at a pre-integration step. PSGL-1 was observed binding to F-actin and preventing its depolymerization, thus disrupting the reverse transcription process. However, others have found PSGL-1 to have no impact at this step [83,211,212]. While not an ISG, the PAF1 complex was identified as an antiviral factor that represses HIV-1 replication prior to integration, most likely during the reverse transcription step [213]. Exonuclease 1 (Exo1), a post-replication DNA repair protein that degrades DNA in a 5’ to 3’ manner, serves as a restriction factor by degrading vial DNA [214]. Helicase-like transcription factor (HLTF) is part of DNA repair pathways in macrophages and dividing T cells; HLTF has been shown to bind the 3’ end of viral ssDNA, and most likely acts in concert with proteins like Exo1 to effectively destroy viral DNA [215]. The helicase MOV10 is in the same protein family as UPF1 but has the opposite effect on HIV-1, as it inhibits reverse transcription, and while it can be incorporated into virions this is not required for its antiviral activity [216]. MX2, along with its anti-uncoating capabilities, has been found to impede NUP358-mediated entry of the PIC when the PIC is concurrently interacting with CPSF6 [217]. REAF/RPRD2 is a restriction factor found in dendritic cells and M1/M2 polarized macrophages, and binds reverse transcripts to impede the reverse transcription process [218,219].
Sterile alpha motif and HD-domain containing protein 1 (SAMHD1) can be considered the most crucial post-entry restriction factor against HIV-1 replication in non-dividing cells. It was found that HIV-1 replicated slower in myeloid cells than in activated T cells, and that this retarded growth could be attributed to lower levels of deoxyribonucleoside triphosphates (dNTPs) available for HIV-1 to use for reverse transcription of its RNA genome into DNA [220,221,222]. Interestingly, while HIV-1 was impaired in these nondividing cells, HIV-2 and SIV that expressed viral protein X (Vpx) were not impeded, implying Vpx countered whatever would otherwise block replication. Co-immunoprecipitation experiments for proteins that interacted with Vpx identified SAMHD1, a dNTP triphosphohydrolase that controls dNTP levels when cells are not not preparing for division, as the host restriction factor; subsequent experiments with SAMHD1-deficient non-dividing cells confirmed its importance in HIV-1 restriction [223,224,225,226,227,228]. Later, the observed slower HIV-1 replication in resting T cells was also attributed in large part to SAMHD1 activity [229,230]. Due to the low intracellular dNTP concentrations, possessing an RT with the ability to efficiently reverse transcribe while in a dNTP-low environment is a necessity for HIV-1-1 replication in these non-dividing cells, as HIV-1-1 has no Vpx with which to counter SAMHD1 and has not been shown to alter SAMHD1 function in any capacity; indeed, possession of an extremely efficient RT can be considered the hallmark of M-tropic viruses [226,231,232,233,234]. There is some debate as to the exact nature of SAMHD1’s antiviral properties. Although originally attributed to its function as a dNTPase, experiments involving phosphorylation of SAMHD1 saw a reduction in its antiviral capabilities while no change was seen in dNTP levels [235]. SAMHD1 was also found to have exonuclease activity, acting as an RNAse in the presence of DNA:RNA duplexes such as HIV-1 would produce during reverse transcription [236,237]. Thus, SAMHD1’s antiviral properties may not be attributed solely, if at all, to its depletion of intracellular dNTP levels. Regulation of SAMHD1 in both myeloid and lymphoid cells is controlled by p21, a controller of cell-cycle progression and inhibitor of cyclin dependent kinases, which itself is a downstream gene of the tumor suppressor gene p53 [238,239,240,241]. p21 is also involved in SAMHD1-independent HIV-1 restriction in macrophages, as Fcɣ receptor induction of p21 leads to p21-mediated downregulation of the RNR2 subunit of ribonucleotide reductase, an enzyme involved in dNTP biosynthesis, further limiting the pool of available dNTPs for HIV-1 to use [242,243]. Interestingly, an indirect antiviral effect may be attributed to SAMHD1’s dNTPase activities and p21’s disruption of dNTP biosynthesis; the low dNTP levels cause a low dNTP to ribonucleoside triphosphate (rNTP) ratio in the cell, increasing the chance of incorporations of ribonucleotides into the reverse transcribed viral genome. Not only does this cause delays in reverse transcription, but the reduced RNAse H2 and Fen1 DNA repair capabilities of macrophages results in incomplete repair of these incorporations, the end result being potentially deleterious mutations in the viral genome [244].
5. Integration
Once the PIC has successfully entered the nucleus, HIV-1’s next goal is integration into the host genome [245]. It should be noted that a majority of HIV-1’s newly reverse transcribed genomic DNA will not be integrated, and will continue to exist outside of the host genome. Unintegrated viral DNA can take on a myriad of forms, such as linear DNA, or circularized in 1-LTR or 2-LTR circles; these forms contribute to gene expression, interact with beneficial or detrimental host factors, and can even play a role in “preintegration latency” (reviewed in [246]).
The IN protein of HIV-1’s PIC interacts with a variety of host proteins to ensure successful integration into a quality location for viral transcription to occur. Acetylation of IN greatly enhances its DNA-binding capabilities and enzymatic activities; p300 is a histone acetyl transferases (HAT) that acetylates IN’s C-terminal domain (CTD), thus leading to greater rates of proper integration events [247]. As mentioned earlier, CPSF6, in addition to its role in getting the capsid and PIC to and through the nucleus, is a key player in intranuclear localization of HIV-1 to site of active transcription, euchromatin. Indeed, CPSF6 has been implicated in depth of nuclear penetration of HIV-1 to the gene-rich locations, with CPSF6-KO cells showing reduced proviral formation and decreased levels of integration into sites of active transcription, reducing overall viral infectivity [124,126,129,248]. LEDGF/p75 is another factor involved in HIV-1 integration, previously considered to be the factor that trafficked HIV-1 to sites of active transcription, rather than CPSF6 [249]. However, while LEDGF/p75 was shown to not be responsible for the act of transporting HIV-1 to euchromatin, it plays a very key part in preferential integration into specific parts the active gene bodies to which CPSF6 delivers HIV-1; LEDGF/p75 targets HIV-1 to the transcription units of specific genes, avoiding integration into promoters and CpG islands. Concurrent binding of LEDGF/p75 to IN and the euchromatin promotes integration via “tethering”, with LEDGF/p75-defective mutant models displaying reduced levels of HIV-1 integration into these active gene bodies, and reduced levels of HIV-1 replication [129,245,248,250,251,252,253,254,255,256]. HMG I(Y), of the High Mobility Group “architectural” protein family, co-fractionates with the PIC and helps HIV-1 integrate into the genome [257,258]. Interactions between IN and the SWI/SNF chromatin remodeling complex allows integration into stable nucleosomes that would otherwise be resistant, in vitro [259]. DNA-PK and its DNA-binding component Ku80, a DNA-damage response complex specific for double-strand breaks (DSBs), have been implicated in HIV-1 integration; deletion of DNA-PK leads to abortive-integration-triggered apoptosis, and under-expression of Ku80 results in impaired proviral formation [260,261,262,263]. Similarly, ATM, another protein associated with DSB DNA-damage response, promotes stable integration, with ATM knockdown mutants undergoing cell death as a result of inefficient integration [264].
Prevention of integration is the cell’s last chance to prevent HIV-1’s gaining a foothold in the cell and potentially becoming a reservoir for future reactivation, as this is an irreversible process [265]. Interestingly, INI1, reported to assist in the reverse transcription stage of HIV-1 replication, seems to stall HIV-1 at the integration step in a manner not requiring its prior incorporation into virions. Depletion of INI1 saw an increase in unintegrated 2-LTR circles and integrated HIV-1 DNA, leading to speculation that INI1’s interaction with IN may destabilize the PIC and interrupt IN’s activity [266]. While some DNA repair proteins aid in integration, others are involved in its prevention. RAD52, a DSB repair protein that utilizes the homologous recombination route of repair, binds the long terminal repeat (LTR) portion of HIV-1 cDNA and prevents stable integration. This is done in competition with Ku80, as both these proteins will bind the HIV-1 LTR in a mutually exclusive manner [267]. TFIIH is a protein complex involved in nucleotide exclusion repair, and two of its components, the helicases XPB and XPD, have been implicated in reduction of HIV-1 integration events. Mutant XPB or XPD cell lines showed higher cDNA kinetics than wild-type, and their antiviral activity was deduced to be due to degradation of viral cDNA, rather than interruption of cDNA synthesis [268]. Outside of DNA repair pathway proteins, KAP1 is a TRIM family protein that, similarly to SAP18, complexes with HDAC1. However, this HDAC1 interaction is to HIV-1’s detriment, as KAP1 associates with acetylated IN, promoting HDAC1-mediated deacetylation of IN, disrupting IN’s function and impairing integration [269].
6. Transcription
Whether integrated or existing in the nucleus as unintegrated genomic DNA, HIV-1 has three possible levels of transcription: basal expression, increased expression via the HIV-1 trans-activator protein Tat (which binds the Transactivation Response element (TAR) of nascent RNA transcripts to promote transcriptional elongation [270]), and repressed transcription to maintain a latent state. The 5’ LTR of HIV-1 contains the core promoter (a TATA element, three GC-rich Sp1 binding sites, and an initiator site), upstream elements, enhancer elements, and 5’ UTR/downstream region [271]. Many host factors are involved in modulating expression levels of HIV-1 (comprehensively reviewed in [271,272]). Transcription requires RNA polymerase II (RNAPII) to associate with the viral promoter; this is facilitated by the TATA-binding protein (TBP) portion of the TFIID transcription factor, which binds the HIV-1 TATA element to begin formation of the preinitiation complex that positions RNAPII to begin transcription [271,273,274].
A multitude of host factors, particularly cyclin-dependent kinases (CDKs), work to ensure proper RNAPII function and mRNA processing [275]. Cdk9, part of the positive transcription elongation factor (P-TEFb) and Tat-associated kinase (TAK), can autophosphorylate and thereby activate itself, subsequently phosphorylating serine-2 of RNAPII’s C-terminal Domain (CTD) and facilitating Tat transactivation, promoting transcriptional elongation [276,277,278,279]. Host transcription factor ZASC1 was found to have a DNA binding element upstream of the TAR, recruiting both Tat and P-TEFb in a TAR independent-manner to promote HIV-1 gene expression and transcriptional elongation [280]. Expression of TAK’s components, Cdk9 and cyclin T1, are upregulated in response to T cell activation and monocyte differentiation into macrophages, directly impacting HIV-1 transcription levels [281]. TFIIH has also been shown to associate with the HIV-1 preinitiation complex, through interaction with Tat (although its interactions are not exclusively Tat-mediated), and is implicated in Tat-mediated transcriptional elongation, although there is some dispute over its necessity. In preinitiation complexes lacking Tat, the Cdk7 subunit of TFIIH will phosphorylate serine-5 of RNAPII’s CTD; in the presence of Tat, Cdk9 undergoes Tat-induced autophosphorylation and activation, and its substrate specificity is altered so that it phosphorylates both serine-2 and serine-5 of RNAPII’s CTD, resulting in prolonged CTD phosphorylation even after TFIIH unbinds from the complex. While TFIIH is still associated with the preinitiation complex, Cdk9’s autophosphorylation is forestalled by the XPB subunit of TFIIH, revealing interweaving dynamics between the factors involved in HIV-1 transactivation [282,283,284,285,286]. The RNAPII transcriptional co-activator p15 (PC4), the nuclear DNA-binding protein NUCKS1, and the TAR RNA binding protein (TRBP) are also Tat-associating host factors that play a role in promoting Tat-mediated transcription [287,288,289,290].
Both capping and splicing of viral mRNAs require the aid of host factors. Phosphorylation of RNAPII’s CTD’s serine-5 by Cdk9 also causes the recruitment of mRNA capping enzyme (Mce1) and subsequent co-transcriptional 5’ capping of the viral mRNA; Mce1 can also be recruited directly by Tat, although this capping pathway is lower efficiency than the conventional RNAPII-mediated route [291,292,293,294,295]. Alternative splicing of the HIV-1 provirus results in production of over 50 mRNA isoforms; a host of splicing regulatory elements (SREs) such as HNRNPF and SRSF2 are responsible for generating these isoforms (reviewed in [296]). When splicing is not desired, the hnRNP A/B proteins are used to inhibit mRNA splicing [297,298]. Recently, it was found that protein kinase RNA-activated (PKR), which is an antiviral RNA immune sensor, can be tricked into splicing HIV-1 rev/tat mRNA; binding of the mRNA activates PKR and induces splicing of the mRNA, as well as PKR’s antiviral activities (discussed below) [299].
The transcription factor specificity protein 1 (Sp1) binds to the Sp1 binding sites and is needed for basal transcription rates, with HIV-1 transcriptional efficiency showing marked reductions when Sp1 binding sites are mutated, as well as for Tat-activated levels of transcription [300,301]. Other members of the Sp family can also utilize these Sp1 binding sites; Sp4 is also an activator of the LTR whereas SP3 is a repressor [302]. Non-Sp proteins, such as the transcription factor BTEB, can also bind the Sp1 binding sites and activate the LTR [303].
The enhancer site upstream of the Sp1 binding sites is composed of two NF-кB binding sites, wherein NF-кB binds to augment HIV-1 transcription. NF-кB -mediated enhancement occurs in both activated T cells and macrophages; interestingly, infected promonocytes (such as the U937 cell line) only show NF-кB binding once they begin maturing to monocytes [304,305]. Though not required for NF-кB to bind the enhancer element, interactions between NF-кB and Tat promote greater Tat-mediated transactivation of transcription. In fact, Tat can indirectly induce expression of NF-кB to start a perpetual positive feedback loop of enhanced viral transcription [306,307,308]. As the term “enhancer” implies, NF-кB amplifies the rates of transcription, but is not strictly required for HIV-1 to successfully transcribe and replicate [304,309,310,311]. NF-кB -mediated enhancement of HIV-1 transcription requires binding to the TATA element via TBP and TFIIB, as well as to Sp1; Sp1 binding is specific, as neither Sp3 nor Sp4 can interact with NF-кB [302,312,313]. Cytokines such as TNFα, IL1, TGF-β, and IL-10, and phorbol esters like TPA, can all stimulate the enhancer by induction of nuclear factor binding to the NF-кB binding sites [314,315,316,317,318,319]. Binding to the enhancer element is not limited to just NF-кB , nor does it always result in enhancement of transcription; transcription factor Ets can interact with NF-кB while binding the enhancer to stimulate transcription, while cell cycle coordinator EF2-1 prevents NF-кB binding to downregulate enhancer-mediated, and even basal level, transcription [320,321].
Binding sites for factors affecting transcription can be located not just in the 5’LTR of HIV-1, but intragenically as well. An intragenic enhancer element was located in the pol gene of HIV-1, and within this element are 3 binding sites for AP-1 transcription factors (JunD, JunB, c-Fos) that positively regulate HIV-1 transcription. Mutations in these sites lead to marked decrease in HIV-1 replication in promonocytic and T cell lines, as well as MDMs. This decrease was attributed to reduced recruitment of RNAPII to the viral promoter [322,323,324].
Although HIV-1 transcription in myeloid cells and T lymphocytes utilizes many overlapping host factors, cell-type specific transcription factors play a role in controlling HIV-1 transcription. NFAT-1, a T cell-specific factor, binds the upstream region of the LTR to negatively regulate transcription; NFAT has also been seen binding to a downstream site in the 5’UTR to induce positive effects on transcription [325,326]. GATA3 is a factor essential for T cell development, and LEF-1 activates the T-cell receptor; both have been shown to have binding sites in the HIV-1 LTR to enhance transcription [327,328]. Binding sites for CCAAT/enhancer binding proteins (C/EBPs) are found in HIV-1’s 5’ LTR; intact C/EBP sites and the proteins that bind them are essential for both basal and enhanced transcription levels in monocytes and macrophages [329,330,331].
Repression of HIV-1 transcription is a double-edged sword for an infected cell; on the one hand it prevents HIV-1 replication and spread of infection, while on the other it allows latent HIV-1 to stay hidden and persist for years until reactivation. Regardless, and for better or worse, many host factors mediate the repression of HIV-1 transcription. CTIP2 complexes with P-TEFb to act as a negative regulator of P-TEFb activity, thus impairing the elongation of HIV-1 RNA during transcription [332,333]. p53, in addition to its regulation of p21 and ultimately SAMHD1, inhibits transcription by interacting with proteins that associate with the Sp1 and TATA elements of the core promoter, although this inhibition is mitigated when the NF-кB sites are functional [334]. LEDGF/p75, past its role for specific integration of the HIV-1 genome into active gene bodies, is recruited by Spt6 to the HIV-1 promoter to form a complex with Iws1 and silence the HIV-1 provirus and prevent reactivation [335]. Oct1 and Oct2 are factors that can bind to multiple octamer sites in the LTR, repressing basal and Tat-mediated activation of the LTR [336]. Uracil DNA glycosylase 2 (UNG2), a DNA repair protein that excises uracil bases from DNA, negatively regulates LTR transcription when stimulated by Tat [337]. The transcription factor YY1 binds a site near the HIV-1 initiator sequence to downregulate transcription [338]. LBP-1 (LSF) is noteworthy as it has two binding sites in the LTR, one downstream of the promoter and one overlapping the TATA element, with opposing effects on HIV-1 transcription. Binding to the downstream site is implicated in basal promoter activity, whereas binding at the TATA-overlapping site (unless outcompeted for the site by TFIID) repressed the elongation step of transcription; LBP-1 repression of transcription seems to require interaction with YY1. Both LBP-1 binding events happen simultaneously in an LBP-1 concentration-dependent manner, suggesting a balancing act for controlling HIV-1 transcription [339,340,341,342]. IFI16, previously described here as an immune sensor against HIV-1, plays a direct antiviral role (perhaps, in this case, to HIV-1’s benefit) by binding Sp1 to prevent its binding and activation of the LTR, which also suppresses reactivation of latent HIV-1 [343]. TRIM22 similarly prevents transcription, and reactivation from latency, by inhibiting binding of Sp1 to its binding sites [344,345,346]. In addition to its anti-fusion activity, SERINC5 has also been observed to downregulate HIV-1 transcription, although the molecular mechanisms behind it are still unknown [347]. The SMC5-SMC6 complex localization factor 2 (SLF2) specifically targets unintegrated DNA. SLF2 recruits the SMC5/6 complex to unintegrated HIV-1 DNA, in any of its forms, and physically compacts the viral chromatin to decrease its accessibility and reduce its activating histone markers, thereby silencing gene expression by inducing a heterochromatin-like state onto unintegrated HIV-1 DNA [348].
Inhibition of transcription can also be caused by physical barriers; chromatin reassembly factors (CRFs) can restructure HIV-1-integrated DNA to occlude polymerase activity, thereby ensuring a latent state, with depletion of these factors leading to reactivation [349]. HDACs remove acetyl groups on the Lysines of histone tails, causing chromatin remodeling resulting in obstruction of access to associated DNA; this inhibits transcription of HIV-1, and HDACs have long been considered monumental contributors to the latency of HIV-1 [350]. In addition to associating with P-TEFb to repress transcription, CTIP2 is part of a chromatin remodeling complex, and through interactions with lysine methyltransferases and HDACs can restructure the architecture of chromatin to silence HIV-1 expression in microglial cells, unfortunately contributing to HIV-1 reservoirs in the brain [351]. ZBTB2 is recruited to the nucleus by ZASC1, the latter of which was mentioned earlier for its recruitment of Tat and P-TEFb to an element upstream of TAR; ZBTB2 represses HIV-1 gene expression through interactions with HDACs to cause deacetylation of histones, leading to silencing of HIV-1 genes. Interestingly, ZBTB2 is no longer recruited to the nucleus, or is removed from HIV-1’s promoter if already associated, if the ATR DNA damage response pathway is stimulated [352].
Should inhibition of transcription fail, the degradation of viral mRNA serves as a different means to the same end. RNAse L acts to control RNA levels, degrading viral and cellular RNAs alike; interestingly, HIV-1 infection induces RNAse L Inhibitor (RLI, or ABCE1) which helps to counter RNase L-mediated transcript destruction [353,354,355,356]. In CD4+ T cells, the long isoform of ZAP (ZAP-L) is an ISG that, when complexed with TRIM25 and KHNYN, exerts antiviral activity by binding to high-CpG portions of multiply spliced HIV-1 mRNA and recruiting other host factors (poly(A)-specific ribonuclease, RNA exosome, and RNA helicase p72) to facilitate mRNA degradation. It should be noted that only CpG-enriched variants of HIV-1 appear to be significantly affected by ZAP [356,357,358,359,360,361,362,363,364]. N4BP1 is an ISG of both macrophages and CD4+ T cells that degrades viral mRNA [365]. MCPIP1 (otherwise known as regnase-1 or Zc3h12a) is an RNAse that degrades viral mRNAs in quiescent T cells [356,366]. Dicer and components of RNA-induced silencing complexes (RISCs), such as Ago1 and Ago2, are all involved in the creation of microRNAs that target viral mRNA for destruction. Interestingly, high expression of microRNA alone may be a key determinant of monocyte and macrophage susceptibility to HIV-1 infection [367,368,369,370].
7. Nuclear Export of Viral mRNA
Like all mRNA, the successfully transcribed viral RNA must be exported from the nucleus HIV-1 worked so hard to enter before it can be translated into proteins by the cell’s ribosomes. HIV-1 mRNA contains splicing sites, with fully spliced mRNA being exported via the conventional binding by NXF1 and the EJC for transport to the cytoplasm [370,371]. For unspliced and partially spliced mRNA this export route is unavailable, however one of the fully spliced mRNAs is Rev, which can be translated and return to the nucleus via its NLS to rescue the rest of the mRNA. Rev’s NLS promotes interaction with a variety of host factors to facilitate its nuclear localization, such as importins beta, 5, and 7, as well as the transport receptor transportin [372].
Once back in the nucleus, Rev binds to the Rev Response Element (RRE) of unspliced/partially spliced viral mRNA, and contains a leucine-rich Nuclear Export Signal (NES); this NES is where host factor CRM1 (also known as exportin-1 or XPO1) a nuclear export receptor, binds to Rev in a co-transcriptional manner. Binding to the 5’ cap of the mRNA is the nuclear cap-binding complex (CBC), an export complex that associates with CRM1 and unspliced small nuclear RNA (snRNA) to mediate its transport to the cytoplasm, as well as later steps in translation (discussed further below) [373,374,375,376,377]. With Rev bound to the RRE of the unspliced/partially spliced viral mRNA, and with the aid of RanGTP to stabilize the interaction between CRM1 and cargo, the Rev-CRM1-mRNA complex is exported out of the nucleus [378,379,380,381,382,383,384,385]. The proteins PACS1, PSF, MATR3, RAB, RNA helicase A, RBM14, DDX3, DDX1, Sam68, eIF4A, hnRNP A1, and hRIP have all been implicated in the Rev-CRM1 route of nuclear export, as depletion of any of these proteins decreases the quantity of unspliced HIV-1 RNA detectable in the cytoplasm [386,387,388,389,390,391,392,393,394,395,396,397].
Interestingly, unspliced HIV-1 mRNA also recruits various host proteins usually involved in mRNA decay pathways to aid in mRNA export. UPF1, mentioned above as a positive factor in reverse transcription, is involved in NMD of prematurely truncated mRNA, and is recruited by HIV-1 post-transcription to shuttle unspliced mRNA safely from the nucleus to the cytoplasm [370,398,399]. STAU1 and STAU2 are proteins of the Stau-Mediated Decay (SMD) pathway of mRNA degradation that interact with Gag and Rev, respectively, as well as the unspliced mRNA to guarantee nuclear export and translation [370,400,401,402,403].
8. Translation
Translation of the HIV-1 mRNA into proteins is the next step of the actively replicating virus’s life cycle. As with all protein synthesis, this requires effective and accurate recruitment of the host ribosomes, and in this regard an array of viral and host protein interactions must work in tandem. 5’ TAR RNA structures provide extra stability to the unspliced viral mRNA that can interfere with ribosome recruitment and scanning, and for this issue host helicases are required [404,405,406]. RNA helicase A (RHA) is packaged in virions as well as recruited in the host cell, and binds to TAR RNA to enact its helicase functions and promote translation; depletion of RHA or deficiency of RHA-virion packaging greatly reduces HIV-1 translation levels [406,407,408]. Another helicase, DDX3, has been shown to associate with Tat in cytoplasmic stress granules, and is needed for maximal translation [409].
A noteworthy aspect of HIV-1 infection is that the active form of eukaryotic initiation factor 4E (eIF4E) is downregulated by Vpr to induce cell cycle arrest at the G2/M phase [376]. eIF4E is the cap-binding protein component of eIF4F, the protein complex needed for cap-dependent translation [410]. HIV-1 uses the cap-dependent translation route (among others) for its mRNA, so this would seem counterproductive to downregulate eIF4E; however, HIV-1 has workarounds to ensure translation of its own proteins while halting that of the host’s. For instance, CBC-bound unspliced viral mRNAs are still actively translated during this cell-cycle arrest, displaying CBC’s ability to circumvent the need for eIF4E [376]. DDX3 also plays a major role when active eIF4E is depleted; DDX3 can act as a substitute for eIF4E by binding the 5’ end of viral mRNA, and through interactions with eIF4G and poly-A-binding protein cytoplasmic 1 (PABP) it can associate with eIF4F to promote translation in cytoplasmic compartments [409,411,412]. Outside of HIV-1, interactions between DDX3 and the CBC to initiate translation have been observed, particularly in regards to cancer progression; however, to our knowledge there is yet to be any studies directly linking their interactions and translation of HIV-1 [413,414].
Other than cap-dependent translation, HIV-1 also uses internal ribosome entry sites (IRESs) to translate uncapped mRNA [415]. HIV-1 possesses two IRESs; one in the 5’ UTR (HIV-1-1 IRES), and one in gag coding region (HIV-1-1 Gag IRES) [416,417]. Several IRES Trans-Acting Factors (ITAFs) have been identified as amplifiers of HIV-1 IRES translation. Interestingly, various proteins that assist in the Rev-CRM1-mediated nuclear export of HIV-1 mRNA also play roles as ITAFs: depletion of DDX3, eIF4A, or hRIP would result in decreased translation of uncapped HIV-1 mRNA [418]. hnRNP A1 is a nuclear protein involved in a variety of RNA-processing activities like splicing and export; HIV-1 induces the expression of hnRNP A1 and promotes its accumulation in the cytoplasm where it functions as an ITAF for HIV-1 translation [419]. An HIV-1-1IRES pulldown assay identified a variety of proteins that interact with that IRES; PC4, outside of its role in Tat-mediated activation of the 5’LTR, was one of these proteins, revealing a dual role in both transcription and translation [420].
Proteolytic processing of the Gag and Gag-Pol polyproteins is carried out by HIV-1’s protease (PR). PR is initially translated as part of the Gag-Pol polyprotein; during or shortly after virion budding, Gag-Pol undergoes “PR precursor autoprocessing” to release PR, which subsequently cleaves the packaged Gag and Gag-Pol polyproteins into their individual proteins [421]. Cleavage into the individual proteins, and the sequence in which these cleavage events occur, is required for correct maturation of HIV-1 [422]. The Env (gp160) protein is not part of the Gag or Gag-Pol polyproteins, and is not a substrate of PR, but requires proteolytic processing to be split into the gp120 surface and gp41 transmembrane subunits. Unprocessed or incorrectly processed Env gp160, i.e. improper cleavage into gp120 and gp41, can be directed to lysosomes for degradation, or may be assembled into virions and impair infectivity of the virus [423,424]. For Env proteolytic processing, the host endoprotease furin is utilized by HIV-1 to carry out the cleavage of gp160 into gp120 and gp41, the process of which occurs as gp160 is exiting the Golgi [425,426]. PACS-1 is a cytosolic sorting protein that binds furin and localizes it to the trans-Golgi Network (TGN), thus facilitating furin-mediated cleavage of gp160 [427]. Though important for maximal levels of gp160 cleavage, furin is not essential for the process to occur, as gp160 cleavage was observed in furin-deficient cells [428]. Indeed, furin is not the only protein that can cleave gp160, as another endoprotease, Proprotein convertase 1 (PC1), was also found to mediate gp160 cleavage [429].
Restriction factors against HIV-1 translation employ a variety of approaches to prevent viral protein production. The IFITM family of ISGs, previously mentioned for their anti-entry activities, also impair HIV-1 at the translation step. Here, IFITMs intercept viral mRNA transcripts to prevent their interaction with ribosomes, with transcripts encoding an RRE being the most greatly affected. Interestingly, deletion of C-terminal residues of IFITM1 increased its inhibitory effects on Gag synthesis, suggesting regulation of IFITM antiviral activity plays a role in the host immune response [71,421]. HIV-1 requires a specific Gag to Gag-Pol polyprotein ratio, about 20 to 1, for proper replication and infectivity [422]. HIV-1 takes advantage of the Programmed -1 Ribosomal Frameshifting (-1PRF) mechanism of ribosomes; occasionally during the translation of Gag mRNA, this frameshift occurs, generating the Gag-Pol polyprotein and ensuring the proper ratio of viral proteins is kept [423,424]. Shiftless (SFL) is a recently described protein that interferes with the regulation of Gag to Gag-Pol polyproteins in an infected cell. SFL interferes with viral translation by interacting with mRNA containing the -1PRF signal, as well as actively translating ribosomes, to cause premature termination of translation [424]. Schlafen 11 (SLFN11) is an ISG with a fascinating antiviral effect; SLFN11 can selectively inhibit translation of HIV-1 proteins, but not global translation, based on the codons used by the virus. HIV-1 displays a bias towards use of A/T in its genes, namely the rare incorporation of adenine in the third nucleotide position of the codon, making the relatively rare tRNAs that correspond to these codons invaluable to HIV-1 protein synthesis. SLFN11 binds these tRNAs to prevent their use by HIV-1, thus disrupting translation by depleting the available tRNA pool [425,426,427,428]. Schlafen 12 (SLFN12) acts similarly to SLFN11, stalling ribosomes to cause a block in translation in a codon-dependent manner, and has been suggested to play a role in the prevention of reactivation of HIV-1 in CD4+ T cells [429]. HuR, an ELAV-like protein that was previously mentioned as a positive modulator of RT activity, serves to negatively regulate IRES-mediated HIV-1 translation by repressing IRES activity; despite being an RNA-binding protein, HuR’s influence on IRES-mediated translation does not appear to involve direct binding to the viral RNA [430,431].
Recently, caspase recruitment domain-containing protein 8 (CARD8) was described as an immune sensor for HIV-1 PR activity. As mentioned previously, PR is normally released and active during or after virion budding. However, premature PR release and activity, which can occur due to overexpression of the Gag-Pol polyprotein or treatment with non-nucleoside reverse transcriptase inhibitors, causes PR to cleave the N-terminal portion of CARD8, releasing the C-terminal subunit that then triggers the caspase-1 inflammasome pathway of pyroptotic cell death. This occurs soon after viral entry, as the aberrant PR is released into the cell, but prior to establishment of productive infection. CARD8 inflammasome activity was originally observed in myeloid cells, and was later found in resting, but not activated, CD4+ T cells. Taken altogether with the observation that bystander resting T cells showing no evidence of productive HIV-1 infection still undergo programmed cell death, CARD8-mediated pyroptosis is suggested to be a contributing factor to the depletion of resting CD4+ T cells prior to establishment of productive infection [432,433,434,435,436].
Protein Kinase R (PKR) is an ISG that, upon sensing of viral dsRNA, phosphorylates eIF2α, preventing formation of the translation initiation complex, thus halting both viral and cellular translation [170]. Interestingly, during HIV-1 infection multiple host factors work against PKR and greatly undercut its antiviral activity. As mentioned previously, PKR can be countered by another ISG, the adenosine deaminase ADAR1, which edits viral RNA and prevents detection by cellular RNA immune sensors [170,437,438]. The TAR RNA binding protein (TRBP), in addition to its transcriptional enhancement role in HIV-1 infection, inhibits PKR in multiple ways. TRBP can bind dsRNA to mask it from PKR; TRBP can also directly bind PKR to block its phosphorylation of eIF4A; and at high concentrations or when not under stress conditions, TRBP can also bind with the PKR activator (PACT) to prevent activation of PKR [438,439,440,441,442,443]. Normally, PACT phosphorylates PKR to activate it under conditions of stress; however during HIV-1 infection PACT is not only sequestered by TRBP to prevent its interaction with PKR, but has also been seen to have its role reversed, going from a PKR activator to an inhibitor. Though the exact mechanisms are not fully understood, it may involve PACT binding to ADAR1 [444,445,446,447].
The interruption of translation can also be exploited to HIV-1’s benefit. As mentioned previously, HIV-1 requires a specific Gag to Gag-Pol translation ratio for proper replication and infectivity, and the same is true for the ratio of Gag to Env protein, as Env-deficient viral particles are non-infectious [448,449,450]. During translation of the Gag polyprotein, the host factor RuvB-like 2 (RVB2) binds the translated MA portion and the 5’UTR of the translating mRNA to enable mRNA degradation. This anti-translation activity is relieved by Env, which competes with RVB2 for MA-binding, reinforcing the hypothesis that RVB2’s mRNA degradation activities serve HIV-1’s best interests by promoting a proper Gag to Env ratio [451].
Ubiquitination and subsequent degradation via constitutive proteasome (hereafter “proteasome”) or immunoproteasome is the cell’s primary method of disposing of translated viral proteins, and serves a dual role in the immune response. Other than directly halting viral replication in the currently infected cell, proteasome and immunoproteasome degradation is a key step in presenting HIV-1 antigens by antigen-presenting cells (APCs), like macrophages and dendritic cells, via major histocompatibility complexes I and II (MHC I and MHC II, respectively; discussed further in a later section) [452,453,454,455]. The CA, IN, Tat, Nef, and Vif HIV-1 proteins are all targets of ubiquitination and proteasomal degradation [453,456]. Degradation of Tat by the ubiquitin-proteasome system (UPS) is mediated by the long non-coding RNA NRON; NRON binds Tat, and then recruits NRON binding proteins involved in the UPS, specifically CUL4B, PSMD11, and HUWE1, which facilitates UPS-mediated degradation [457]. HIV-1 CA and IN proteins are both antagonized by the E3-ubiquitin ligase TRIM5α, meting out their proteasome-destined destruction [456,458,459]. Nef is degraded via its interactions with ubiquitin specific protease 15 (USP15), although the exact mechanism is not clear since USP15 usually functions to stabilize proteins by deubiquitinating them [460]. Vif is ubiquitinated by the E3-ligase MDM2, and is noteworthy amongst HIV-1’s accessory proteins as it has a notably short half-life, which may suggest Vif’s intense rates of MDM2-mediated degradation are by design and benefit HIV-1 infection [461,462].
Degradation can also be facilitated outside of the UPS pathway. Lysosomal-associated transmembrane protein 5 (LAPTM5) is a major restriction factor against HIV-1 infection of macrophages and DCs, as it is responsible for transporting Env glycoproteins to lysosomes for destruction [463,464]. Interestingly, another heavy hitter in myeloid defenses is mannose receptor (MR), previously mentioned for its benefits to HIV-1 through its aid in initial virion binding to DCs. Env proteins undergo post-translational processing, such as N-linked glycosylation; one addition to Env is the mannose patch, and through this region MR binds Env and Env-containing virions and facilitates their shuttling to lysosomal compartments where they are destroyed [465,466,467].
A consequence of the degradation of HIV-1 proteins is the generation of peptides that can be used by immune cells to generate a large-scale immune response. Antigen-presenting cells such as macrophages and dendritic cells, can present via MHC I to activate CD8+ T cells, or via MHC II to activate naïve CD4+ T cells [26,468,469]. After activation, CD8+ T cells will then kill any infected cell presenting the recognizable antigen via MHC I; nearly all cell types can present via MHC I [470]. Upon activation via MHC II, CD4+ T cells will then differentiate, proliferate, and activate other immune cells to amplify the immune response [471]. Cathepsins are proteases found in endosomes and lysosomes, and play a variety of roles in different immune cells, from processing peptides for MHC II presentation to regulating cytotoxicity of natural killer (NK) and CD8+ T cells [472]. Two cathepsins, D and K, were found to be able to degrade a proteasome-resistant Env gp120 mutant [473]. ER aminopeptidases (ERAPs) are involved in MHC I peptide generation by further degrading proteins post-proteasome; in HIV-1’s case the p17 MA protein and the p24 CA protein are subject to ERAP degradation [474,475].
9. Assembly and Egress
The first step for proper HIV-1 virion assembly and budding, in primary T cells and cell lines, is localization of virus (and host) components to the peripheral PM, and here the Gag polyprotein’s interactions with host factors are critical [476,477,478]. Localization of Gag to the PM is aided by phosphatidylinositol (4,5) bisphosphate [PI(4,5)P2] (PIP2), a phosphoinositide that localizes many host cell factors to the PM [479]. By interacting with basic amino acid residues in the MA domain of Gag, PIP2 binds the Gag polyprotein and transports it to the PM [480]. Assembly sites are not random, but are instead sites where lipid rafts, microdomains composed of cholesterol, sphingolipids, and proteins, can be assembled; cholesterol in particular is a critical necessity, as depletion of cholesterol permeabilizes the virion membrane and greatly impacts incorporations of Gag and Pol proteins [481]. Once at the PM, Gag will bind to the PM, an interaction dependent upon its N-terminally myristoylated MA domain [482,483]. Myristoylation of MA of Gag is carried out by N-myristoyltransferase (NMT) and is required for PM association, proteolytic processing, Gag-Gag associations, and budding, but is not necessary not for localization of Gag to the PM. PIP2 interaction with MA causes a conformational change exposing its N-terminal myristoyl site, which allows PM binding; this “myristoyl switch” is thought to be a method to prevent aberrant MA binding until PIP2 binds Gag and localizes it to the PM [476,484,485,486,487,488,489,489,490]. Basic residues in MA, in addition to the N-terminal myristate, also enable binding to the PM-bound acidic phospholipid PS, further enhancing Gag-PM binding stability, while also contributing to the correct formation of MA hexamers needed for proper virion assembly [491,492].
Env proteins must also be localized to the PM for correct assembly to occur. As mentioned earlier, Env gp160 must be properly processed, by furin or PC1, into gp120 and gp41 for localization to the PM to occur, otherwise it will be targeted to lysosomes where it will be degraded [493]. Post-cleavage, gp120 and gp41 form a complex, which in turn trimerizes with other gp120/gp41 complexes to form an Env complex. gp41 has a long cytoplasmic tail (CT) of around 150 amino acids, and this CT is what interacts with host factors to facilitate Env complex localization to the PM. Rab11-FIP1C/RCP (FIP1C) is from a family of cargo-sorting proteins and, together with its binding partner Rab14, interacts with Env complexes in a CT-dependent manner to localize them to the PM and incorporate Env into the virions [494,495]. Adaptor Protein 1 (AP-1) also appears to play a role in sub-cellular localization of Env after it exits the TGN, and as with FIP1C and Rab14, the gp41 CT is the facilitator of interaction [496]. Interactions between the gp41 CT and MA of Gag also contribute to proper incorporation of Env into budding virions, as mutations in MA or the CT leads to inefficient incorporation of Env into the viral particles [483,497,498].
In myeloid cells, assembly was observed not only at the peripheral PM, but in cytosolic compartments [499]. Originally these were thought to be late endosomes or multi-vesicle bodies, but more recent studies have since shown these compartments to be internalized sections of the PM, marked by the tetraspanins CD9, CD53, and CD81, and are still connected to the cell surface by “virion channels”, tubules only slightly bigger than the virions themselves; these sites were dubbed virus-containing compartments (VCCs) [500,501,502,503,504,505,506]. VCCs are seen during HIV-1 infection, but infection itself is not necessary for VCC formation; the surface lectin Siglec-1, found on macrophages and DCs, was seen to be crucial for formation of VCCs following binding to the gangliosides on virus-like particles, independent of subsequent productive infection [507]. These VCCs also mediate the cell-to-cell transfer of HIV-1 through virological synapses, mentioned earlier in this review [63,508]. Interestingly, the restriction factor tetherin (discussed more later) plays a role beneficial to HIV-1 when it comes to VCC formation, as tetherin-KO macrophages showed impaired formation and distribution of VCCs [509]. As in T cells, PIP2 plays a role in localizing Gag to VCCs, which is consistent with VCCs being comprised of internalized PM; similarly, FIP1C and Rab14 are needed for localization of Env complexes to VCCs [494,510,511].
Once at the PM, Gag sequesters lipid rafts and tetraspanin-containing microdomains to serve as platforms for HIV-1 particle assembly [512,513]. Capsid assembly is aided by ATP-binding cassette E1 (ABCE1, or HP68, or RLI), a transporter protein that binds Gag through the NC domain, though it does not stay attached post-assembly [514,515,516]. Gag-Gag multimerization at the PM is regulated by cellular STAU1 through interactions with the two zinc fingers (ZFs) of Gag’s NC domain; STAU1 also binds genomic RNA, and through this association is packaged into virions [402,517,518,519]. Inositol hexakisphosphate (IP6) also promotes interactions between Gag molecules, and has a conserved role in many lentiviruses; IP6 interacts with the CA domain of Gag to promote assembly of the immature Gag lattice, which then undergoes proteolytic cleavage, after which IP6-CA interactions facilitate formation of the mature capsid structure [520,521]. Gag and Gag-Pol polyproteins are packaged in a radial manner, with the MA domain facing outwards and their C-terminus towards the center [477]. Full-length viral RNA dimers that will serve as the HIV-1 genome are recognized in the cytoplasm by the NC domain of Gag, prior to PM localization, and packaged into the assembling virion [522,523,524]. Nucleolin, an RNA binding chaperone protein that plays a role in ribosome assembly, binds the genomic RNA of HIV-1 and forms a complex with Gag through NC interactions. This leads to virion packaging of both nucleolin and RNA, the latter of which contains the cis-acting RNA element psi, a packaging signal, and enhances virion budding through Gag recognition of psi [525,526,527]. Accessory proteins Vpr, Vif, and Nef are packaged in the virion, with the latter two later undergoing intravirion processing by PR [528,529,530,531,532]. Through a variety of methods, such as mass spectrometry, immunomagnetic capturing, and proteomic profiling, dozens of host proteins have been identified as being incorporated into the virion core or envelope (reviewed in [533,534]). Most packaged proteins are those that beneficially impact a step, or multiple, of the viral replication cycle; examples already mentioned in this review include PSGL-1, CypA, IP6, UPF1, HDAC1, Staufen, RHA, ADAR1, and CBP80 [82,109,121,157,161,376,408,519,535]. Unfortunately for HIV-1, restriction factors such as APOBEC and MOV10 are also incorporated into virions, where they can continue to wreak havoc after the virion has departed from the host cell [199,216].
With HIV-1 genome, HIV-1 proteins, and host proteins in the process of being packaged, the final act of budding proceeds. Once again, Gag interactions with host proteins ensure this process is successful. p6 of the Gag polyprotein contains amino acid motifs dubbed “late domains” (L-domains) that were found to be critical to complete separation of the virion from the cell; deletions of p6 or mutations in this motif would lead to incomplete budding, with virions attached to host cells by thin “stalks” [536,537]. It was discovered that p6 L-domains interact with proteins of the endosomal sorting complex required for transport (ESCRT) pathway at the site of virion assembly to facilitate proper budding [538,539]. The ESCRT pathway is composed of various ESCRT protein complexes (ESCRT-0 to ESCRT-III), and is a trafficking pathway used by cells to sort ubiquitinated membrane proteins via multivesicular bodies (MVBs) to lysosomes for destruction [539,540]. An L-domain of p6 with the PTAP amino acid motif was found to be able to bind the ubiquitin enzyme 2 variant (UEV) region of tumor susceptibility gene 101 (TSG101), a protein of the ESCRT-I protein complex, and recruit TSG101/ESCRT-I to budding sites to initiate ESCRT assembly. Consistent with previous reports that UEV’s bind Ub, ubiquitination of p6 increases its binding affinity with TSG101’s UEV. To the same point, a Gag construct with a deubiquitinating enzyme fused on had impaired TSG101 binding, suggesting ubiquitination of Gag is a key step in efficient recruitment of TSG101. As noted above, anomalous or nonexistent p6 L-domains heavily impairs budding; likewise, depletion of TSG101 attenuates budding but can be rescued once TSG101 is re-introduced [541,542,543,544,545,546,547]. Once bound to Gag and localized at the PM, TSG101/ESCRT-I can recruit ESCRT-II through interactions with ESCRT-II subunits EAP30 and EAP45; EAP20 of ESCRT-II can then recruit ESCRT-III through interactions with charged multivesicular body protein 6 (CHMP6), a subunit of ESCRT-III that can bind to vacuolar protein sorting-associated protein 28 (VPS28) of ESCRT-I. Interestingly, siRNA-mediated knockdown of ESCRT-II component EAP20, or CHMP6 of ESCRT-III, did not significantly impact HIV-1 release or infectivity, implying that HIV-1 budding is not ESCRT-II-dependent [548,549,550,551].
A second L-domain of p6, YPXL (where X is any amino acid), binds the V region of apoptosis-linked gene 2 (ALG-2)-interacting protein (ALIX); ALIX can interact TSG101/ESCRT-I, as well as with CHMP4 proteins of the ESCRT-III complex to facilitate ESCRT-III recruitment independently of TSG101. Interactions with CHMP4 are mediated through ALIX’s Bro1 domain and are regulated by the ATPase VPS4. Impairment of ALIX/p6 interactions are less catastrophic for HIV-1 budding than impaired TSG101/p6 interactions, indicating that the ALIX route of ESCRT-III recruitment is an approach useful in specific scenarios, e.g. cells under-expressing TSG101 or an HIV-1 mutant without a PTAP motif. Interestingly, ALIX-depleted cells showed multiple recruitments of full ESCRT complexes to budding sites, a rarity in wild type scenarios, suggesting that p6/ALIX interactions may facilitate or coordinate correct recruitment of ESCRT machinery [549,552,553,554,555]. The Bro1 domain of ALIX can also interact with the NC domain of Gag, via basic residues in the N-terminal region and ZFs of NC, linking Gag to ESCRT-III in a p6-independent manner [556,557].
Another host factor Gag interacts with in the course of virion budding is angiomotin (AMOT), an angiostatin-binding protein. HIV-1 variants with defunct PTAP and YPXL L-domains, or lacking a p6 domain altogether, can still bud efficiently when the HECT ubiquitin E3 ligase NEDD4L is overexpressed; other retroviruses can bind NEDD4L directly through a PPXY L-domain, but HIV-1 Gag lacks this domain. AMOT serves as the connection between Gag and NEDD4L, as it can bind somewhere in residues 278-377 of Gag, and bind NEDD4L through AMOTs PPXY motifs and NEDD4L’s WW domains. Depletion of AMOT or NEDD4L impairs budding of HIV-1 strains with Gags lacking p6 at an early stage in the process; despite these strains being unable to directly bind TSG101, NEDD4L-mediated rescue of the strains requires presence of TSG101, suggesting that NEDD4L acts upstream of TSG101 and connects PTAP-and-YPXL-defective mutants to the ESCRT pathway [558,559,560].
Whether through TSG101, ALIX, or AMOT, the end of Gag’s interactions with these host proteins result is recruitment of ESCRT-III components to the site of the budding virion (interactions summarized in Figure 2). ESCRT-III is critical for membrane fission during the cytokinesis phase of mitosis, and is similarly essential for complete separation of budding virions from the host cell [561]. Of the 12 ESCRT-III-like proteins expressed by humans, only CHMP2 and CHMP4 appear to be indispensable for HIV-1 budding. As noted previously, ALIX recruits ESCRT-III through CHMP4; CHMP2 and CHMP4 also interact directly with each other, even in absence of other ESCRT-III components, and both can bind and recruit VPS4 [552,562]. VPS4 is an AAA-type ATPase whose enzymatic activity is required for disassembly and recycling of membrane-bound ESCRT-III complexes, and is the mediator between the ALIX/CHMP4 interactions. Interruption of the CHMP2-CHMP4 or CHMP2/4-VPS4 interactions impairs HIV-1 budding. [552,561,563,564,565]. Like so many other host proteins, ESCRT-I, ESCRT-III, ALIX, and VPS4 are all incorporated into the virion during the budding process [477]. As mentioned above, host proteins of the PM can be incorporated into the virion as it is budding, with some, such as ICAM-1, playing a role during for that virion/virus during the next round of infection and replication; other host surface proteins taken by the virion during budding include MHCI and MHCII, CD40, LFA-1, and many more [64,65,534].
Even at the final stages of the viral life cycle, the host cell is not ready to give up the fight. The Ras GTPase-activating-like protein IQGAP1 (IQGAP1, or p195) is a scaffolding protein that serves as a regulator of actin-cytoskelton rearrangements, as well as many signaling pathways [566,567]. IQGAP1 pre-emptively restricts virion assembly by binding to NC or p6 of Gag, independent of the L-domains, and preventing Gag from assembling at the PM [568]. The ISG TRIM22 functions similarly, binding Gag and disrupting its trafficking to the PM [569]. Another ISG, ISG15, impairs assembly at multiple steps of the ESCRT recruitment process: ISG15 inhibits both Gag and TSG101 ubiquitination and prohibits interaction between Gag’s PTAP and TSG101, preventing TSG101-mediated recruitment of the rest of ESCRT-I; ISG15 also binds CHMP5 of ESCRT-III, and through this disrupts VPS4 recruitment by blocking VPS4 interactions with its coactivator protein LIP5; and ISG15 can be conjugated onto Gag by the host factor HERC5 to disrupt an early stage in Gag assembly [570,571,572]. PSGL-1, an ISG previously mentioned for its restrictions at the fusion and reverse transcription steps in HIV-1 replication, also impairs HIV-1 budding. PSGL-1 can bind gp41 to restrict Env incorporation into budding virions, reducing subsequent infectivity of the virion as a result. PSGL-1 also interacts with Gag to facilitate its incorporation into the virion, leading to its anti-fusion activities mentioned above [82,211,212]. The T-cell immunoglobulin (Ig) and mucin domain (TIM) family of transmembrane proteins play roles in intracellular signaling, with different TIM proteins being more important in T cells and/or macrophages, e.g. TIM-1 in activated T cells, TIM-4 in myeloid cells, TIM-3 in both [573,574,575]. TIM proteins can all bind the PM-bound PS, which as mentioned earlier is important for Gag-PM binding and MA hexamer assembly. Binding of PS by TIM-1, TIM-3, or TIM-4 results in inhibition of virion release, retaining the virions at the PM of T cells, with similar effects observed with TIM-3 in MDMs [576]. TIM-1 also appears to be stabilized by SERINC5, a restriction factor at the fusion step of the HIV-1 replication cycle (see earlier section) [577].
Viperin is an ISG that is greatly upregulated in macrophages compared to monocyte-derived DCs and CD4+ T cells, and combats HIV-1 infection at the assembly stage by disrupting stable formation of the lipid rafts Gag coalesces to serve as virion assembly sites; however, these antiviral effects are only substantial against certain HIV-1 strains [578,579,580]. GBP5 and GBP2 are ISGs, specifically GTPases, involved in intrinsic immune cell activation, are highly stimulated in MDMs by type I and II IFNs, and can be stimulated in T cells by type II IFN. GBP5 and GBP2 interfere with HIV-1’s Env glycoprotein by localizing at the Golgi and deactivating furin, the host protease that cleaves gp160 into gp120 and gp41. This impairs gp120 trafficking to the PM, increasing virion incorporation of unprocessed gp160 and decreasing that of gp120; this in turn leads to reduced infectivity of progeny virions due to their lack of gp120. Interestingly, HIV-1 strains with defective vpu genes have higher resistance to GBP5, though this is not yet confirmed for GBP2. This is likely due to Vpu’s ability to downregulate NF-кB to restrict ISG expression, as NF-кB also acts as a transcription factor for HIV-1, and thus Vpu-defective mutants will have higher expression of Env that can overwhelm GBP5’s antiviral effects; this may be an explanation for the relatively high frequency of M-tropic strains with a defective vpu gene [581,582,583,584,585]. MARCH8 is a RING-finger E3 ubiquitin ligase highly expressed in MDMs and DCs that normally functions to downregulate host transmembrane proteins, such as MHCII, and directly impacts viral infectivity by sequestering Env proteins from the cell surface, leading to virions lacking Env glycoproteins and thus being noninfectious. Interestingly, despite its normal functions involving UPS-degradation of cellular proteins, MARCH8’s antiviral activity does not alter Env protein levels, revealing that MARCH8-mediated downregulation of Env does not involve proteasomal degradation [586,587,588,589]. From the same protein family of E3 ligases, MARCH1 and MARCH2 have similar anti-Env activities to MARCH8; unlike MARCH8, MARCH1 and MARCH2 are stimulated by type I IFNs, and MARCH2 is upregulated by HIV-1 infection alone [589,590,591].
One of the most extensively studied restriction factors is bone marrow stromal antigen 2 (BST-2), also known as tetherin. Tetherin is constitutively expressed in many cell types but is strongly induced by type I IFN, thus it is considered to be an ISG. Tetherin is a transmembrane protein that is found in foci dotted across the PM, and as the name implies, tetherin’s antiviral activity is the tethering of virions to the outside of the plasma membrane, as well as to each other, after they bud. This is accomplished by incorporation of several dozen tetherin homodimers into the virion, with a preference for C-terminal insertion into the virion in an axial manner, as the C-terminus contains a glycosyl-phosphatidylinositol (GPI) anchor. The cytosolic N-terminus of tetherin, and coiled coil structure adopted by the tetherin homodimer, are critical for retaining virions on the cell surface; as the virion with tetherin’s inserted C-terminus attempts to leave, the N-terminus will stay rooted in the PM of the host cell, thus tethering the virion to the cell [592,593,594,595,596,597,598]. In a twist of irony, this retention of virions may actually benefit HIV-1, as it can facilitate formation of virological synapses and cell-cell transfer of the virions [599]. Finally, tetherin also functions as a watchdog for viral attack: through its cytosolic domain, tetherin acts as an immune sensor that stimulates NF-кB in response to virion binding, leading to type I IFN production [600,601].
10. Concluding Remarks
Since the identification of HIV as the pathogen responsible for Acquired Immunodeficiency Syndrome (AIDS) in the early 1980s, the list of host factors identified as playing a role in its replication cycle has only continued to grow [602]. New factors and new interactions between these factors and HIV, or between the factors themselves during HIV infection, are constantly being discovered and explored further. A wide assortment of techniques potentiates these discoveries: co-immunoprecipitation, yeast-two-hybrid assays, fluorescent microscopy and imaging, mass spectrometry, RNA-hybridization, genome-wide association studies, CRISPR, RNA interference; these methods, and more, have been successful in elucidating the mysteries of what host factors HIV interacts with, and how [603,604,605,606,607,608,609,610,611]. As the complex web of dynamics between all factors that help or hinder HIV over the course of infection grows, so too does our sum-total knowledge on this viral scourge. Every factor identified, interaction studied, and mechanism understood brings us one step closer to our goal: a true and final cure for HIV and AIDS.
Funding
This review received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
The authors wish to thank Michelle Cheng for her assistance in gathering information to put into this review. Figures were created utilizing BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Blut), G.A.C.B. (Arbeitskreis; Blood’, S. ‘Assessment of P.T. by Human Immunodeficiency Virus (HIV). Transfus. Med. Hemotherapy 2016, 43, 203. [CrossRef]
- Ganser-Pornillos, B.K.; Yeager, M.; Pornillos, O. Assembly and Architecture of HIV. In Viral Molecular Machines; Rossmann, M.G., Rao, V.B., Eds.; Springer US: Boston, MA, 2012; pp. 441–465 ISBN 978-1-4614-0980-9.
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell Binding and Entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866. [CrossRef]
- Arrildt, K.T.; Joseph, S.B.; Swanstrom, R. The HIV-1 Env Protein: A Coat of Many Colors. Curr. HIV/AIDS Rep. 2012, 9, 52–63. [CrossRef]
- Yoon, V.; Fridkis-Hareli, M.; Munisamy, S.; Lee, J.; Anastasiades, D.; Stevceva, L. The GP120 Molecule of HIV-1 and Its Interaction with T Cells. Curr. Med. Chem. 2010, 17, 741–749. [CrossRef]
- Negi, G.; Sharma, A.; Dey, M.; Dhanawat, G.; Parveen, N. Membrane Attachment and Fusion of HIV-1, Influenza A, and SARS-CoV-2: Resolving the Mechanisms with Biophysical Methods. Biophys. Rev. 2022, 14, 1109–1140. [CrossRef]
- Sattentau, Q.J.; Weiss, R.A. The CD4 Antigen: Physiological Ligand and HIV Receptor. Cell 1988, 52, 631–633. [CrossRef]
- Doms, R.W.; Moore, J.P. HIV-1 Membrane Fusion. J. Cell Biol. 2000, 151, f9–f14.
- Lee, B.; Sharron, M.; Montaner, L.J.; Weissman, D.; Doms, R.W. Quantification of CD4, CCR5, and CXCR4 Levels on Lymphocyte Subsets, Dendritic Cells, and Differentially Conditioned Monocyte-Derived Macrophages. Proc. Natl. Acad. Sci. 1999, 96, 5215–5220. [CrossRef]
- Clapham, Paul R.; Reeves, Jacqueline D.; Simmons, Graham; Dejucq, Natalie; Hibbitts, S.; Aine, McKnight HIV Coreceptors, Cell Tropism and Inhibition by Chemokine Receptor Ligands. Mol. Membr. Biol. 1999, 16, 49–55. [CrossRef]
- Schaeffer, E.; Geleziunas, R.; Greene, W.C. Human Immunodeficiency Virus Type 1 Nef Functions at the Level of Virus Entry by Enhancing Cytoplasmic Delivery of Virions. J. Virol. 2001, 75, 2993–3000. [CrossRef]
- Clapham, P.R.; McKnight, Á. HIV-1 Receptors and Cell Tropism. Br. Med. Bull. 2001, 58, 43–59. [CrossRef]
- Olinger, G.G.; Saifuddin, M.; Spear, G.T. CD4-Negative Cells Bind Human Immunodeficiency Virus Type 1 and Efficiently Transfer Virus to T Cells. J. Virol. 2000, 74, 8550–8557.
- Schnittman, S.M.; Lane, H.C.; Greenhouse, J.; Justement, J.S.; Baseler, M.; Fauci, A.S. Preferential Infection of CD4+ Memory T Cells by Human Immunodeficiency Virus Type 1: Evidence for a Role in the Selective T-Cell Functional Defects Observed in Infected Individuals. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 6058–6062. [CrossRef]
- Joseph, S.B.; Arrildt, K.T.; Sturdevant, C.B.; Swanstrom, R. HIV-1 Target Cells in the CNS. J. Neurovirol. 2015, 21, 276–289. [CrossRef]
- Blanco, J.; Barretina, J.; Gutiérrez, A.; Armand-Ugón, M.; Cabrera, C.; Clotet, B.; Esté, J.A. Preferential Attachment of HIV Particles to Activated and CD45RO+CD4+ T Cells. AIDS Res. Hum. Retroviruses 2002, 18, 27–38. [CrossRef]
- Spina, C.A.; Prince, H.E.; Richman, D.D. Preferential Replication of HIV-1 in the CD45RO Memory Cell Subset of Primary CD4 Lymphocytes in Vitro. Available online: https://www.jci.org/articles/view/119342/pdf (accessed on 13 May 2024).
- Brenchley, J.M.; Hill, B.J.; Ambrozak, D.R.; Price, D.A.; Guenaga, F.J.; Casazza, J.P.; Kuruppu, J.; Yazdani, J.; Migueles, S.A.; Connors, M.; et al. T-Cell Subsets That Harbor Human Immunodeficiency Virus (HIV) In Vivo: Implications for HIV Pathogenesis. J. Virol. 2004, 78, 1160–1168. [CrossRef]
- Stevenson, M.; Stanwick, T.L.; Dempsey, M.P.; Lamonica, C.A. HIV-1 Replication Is Controlled at the Level of T Cell Activation and Proviral Integration. EMBO J. 1990, 9, 1551–1560. [CrossRef]
- Ostrowski, M.A.; Chun, T.-W.; Justement, S.J.; Motola, I.; Spinelli, M.A.; Adelsberger, J.; Ehler, L.A.; Mizell, S.B.; Hallahan, C.W.; Fauci, A.S. Both Memory and CD45RA+/CD62L+ Naive CD4+ T Cells Are Infected in Human Immunodeficiency Virus Type 1-Infected Individuals. J. Virol. 1999, 73, 6430–6435. [CrossRef]
- Chun, T.W.; Engel, D.; Berrey, M.M.; Shea, T.; Corey, L.; Fauci, A.S. Early Establishment of a Pool of Latently Infected, Resting CD4(+) T Cells during Primary HIV-1 Infection. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 8869–8873. [CrossRef]
- Chavez, L.; Calvanese, V.; Verdin, E. HIV Latency Is Established Directly and Early in Both Resting and Activated Primary CD4 T Cells. PLOS Pathog. 2015, 11, e1004955. [CrossRef]
- Cenker, J.J.; Stultz, R.D.; McDonald, D. Brain Microglial Cells Are Highly Susceptible to HIV-1 Infection and Spread. AIDS Res. Hum. Retroviruses 2017, 33, 1155–1165. [CrossRef]
- Koppensteiner, H.; Brack-Werner, R.; Schindler, M. Macrophages and Their Relevance in Human Immunodeficiency Virus Type I Infection. Retrovirology 2012, 9, 82. [CrossRef]
- Loré, K.; Smed-Sörensen, A.; Vasudevan, J.; Mascola, J.R.; Koup, R.A. Myeloid and Plasmacytoid Dendritic Cells Transfer HIV-1 Preferentially to Antigen-Specific CD4+ T Cells. J. Exp. Med. 2005, 201, 2023–2033. [CrossRef]
- Manches, O.; Frleta, D.; Bhardwaj, N. Dendritic Cells in Progression and Pathology of HIV Infection. Trends Immunol. 2014, 35, 114–122. [CrossRef]
- Gill, V.; Shattock, R.J.; Freeman, A.R.; Robinson, G.; Griffin, G.E.; Gordon-Smith, E.C.; Gibson, F.M. Macrophages Are the Major Target Cell for HIV Infection in Long-Term Marrow Culture and Demonstrate Dual Susceptibility to Lymphocytotropic and Monocytotropic Strains of HIV-1. Br. J. Haematol. 1996, 93, 30–37. [CrossRef]
- Campbell, J.H.; Hearps, A.C.; Martin, G.E.; Williams, K.C.; Crowe, S.M. The Importance of Monocytes and Macrophages in HIV Pathogenesis, Treatment, and Cure. AIDS 2014, 28, 2175. [CrossRef]
- Calantone, N.; Wu, F.; Klase, Z.; Deleage, C.; Perkins, M.; Matsuda, K.; Thompson, E.A.; Ortiz, A.M.; Vinton, C.L.; Ourmanov, I.; et al. Tissue Myeloid Cells in SIV-Infected Primates Acquire Viral DNA through Phagocytosis of Infected T Cells. Immunity 2014, 41, 493–502. [CrossRef]
- Kazazi, F.; Mathijs, J.-M.; Foley, P.; Cunningham, A.L. Variations in CD4 Expression by Human Monocytes and Macrophages and Their Relationship to Infection with the Human Immunodeficiency Virus. J. Gen. Virol. 1989, 70, 2661–2672. [CrossRef]
- Quitadamo, B.; Peters, P.J.; Repik, A.; O’Connell, O.; Mou, Z.; Koch, M.; Somasundaran, M.; Brody, R.; Luzuriaga, K.; Wallace, A.; et al. HIV-1 R5 Macrophage-Tropic Envelope Glycoprotein Trimers Bind CD4 with High Affinity, While the CD4 Binding Site on Non-Macrophage-Tropic, T-Tropic R5 Envelopes Is Occluded. J. Virol. 2018, 92, e00841-17. [CrossRef]
- Joseph, S.B.; Arrildt, K.T.; Swanstrom, A.E.; Schnell, G.; Lee, B.; Hoxie, J.A.; Swanstrom, R. Quantification of Entry Phenotypes of Macrophage-Tropic HIV-1 across a Wide Range of CD4 Densities. J. Virol. 2014, 88, 1858–1869. [CrossRef]
- Arrildt, K.T.; LaBranche, C.C.; Joseph, S.B.; Dukhovlinova, E.N.; Graham, W.D.; Ping, L.-H.; Schnell, G.; Sturdevant, C.B.; Kincer, L.P.; Mallewa, M.; et al. Phenotypic Correlates of HIV-1 Macrophage Tropism. J. Virol. 2015, 89, 11294–11311. [CrossRef]
- Maddon, P.J.; McDougal, J.S.; Clapham, P.R.; Dalgleish, A.G.; Jamal, S.; Weiss, R.A.; Axel, R. HIV Infection Does Not Require Endocytosis of Its Receptor, CD4. Cell 1988, 54, 865–874. [CrossRef]
- Deng, H.; Liu, R.; Ellmeier, W.; Choe, S.; Unutmaz, D.; Burkhart, M.; Marzio, P.D.; Marmon, S.; Sutton, R.E.; Hill, C.M.; et al. Identification of a Major Co-Receptor for Primary Isolates of HIV-1. Nature 1996, 381, 661–666. [CrossRef]
- Dragic, T.; Litwin, V.; Allaway, G.P.; Martin, S.R.; Huang, Y.; Nagashima, K.A.; Cayanan, C.; Maddon, P.J.; Koup, R.A.; Moore, J.P.; et al. HIV-1 Entry into CD4+ Cells Is Mediated by the Chemokine Receptor CC-CKR-5. Nature 1996, 381, 667–673. [CrossRef]
- Alkhatib, G.; Combadiere, C.; Broder, C.C.; Feng, Y.; Kennedy, P.E.; Murphy, P.M.; Berger, E.A. CC CKR5: A RANTES, MIP-1alpha, MIP-1beta Receptor as a Fusion Cofactor for Macrophage-Tropic HIV-1. Science 1996, 272, 1955–1958. [CrossRef]
- Feng, Y.; Broder, C.C.; Kennedy, P.E.; Berger, E.A. HIV-1 Entry Cofactor: Functional cDNA Cloning of a Seven-Transmembrane, G Protein-Coupled Receptor. Science 1996, 272, 872–877. [CrossRef]
- Doranz, B.J.; Rucker, J.; Yi, Y.; Smyth, R.J.; Samson, M.; Peiper, S.C.; Parmentier, M.; Collman, R.G.; Doms, R.W. A Dual-Tropic Primary HIV-1 Isolate That Uses Fusin and the β-Chemokine Receptors CKR-5, CKR-3, and CKR-2b as Fusion Cofactors. Cell 1996, 85, 1149–1158. [CrossRef]
- He, J.; Chen, Y.; Farzan, M.; Choe, H.; Ohagen, A.; Gartner, S.; Busciglio, J.; Yang, X.; Hofmann, W.; Newman, W.; et al. CCR3 and CCR5 Are Co-Receptors for HIV-1 Infection of Microglia. Nature 1997, 385, 645–649. [CrossRef]
- Choe, H.; Farzan, M.; Sun, Y.; Sullivan, N.; Rollins, B.; Ponath, P.D.; Wu, L.; Mackay, C.R.; LaRosa, G.; Newman, W.; et al. The β-Chemokine Receptors CCR3 and CCR5 Facilitate Infection by Primary HIV-1 Isolates. Cell 1996, 85, 1135–1148. [CrossRef]
- Parrish, N.F.; Gao, F.; Li, H.; Giorgi, E.E.; Barbian, H.J.; Parrish, E.H.; Zajic, L.; Iyer, S.S.; Decker, J.M.; Kumar, A.; et al. Phenotypic Properties of Transmitted Founder HIV-1. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, 6626–6633. [CrossRef]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and Characterization of Transmitted and Early Founder Virus Envelopes in Primary HIV-1 Infection. Proc. Natl. Acad. Sci. 2008, 105, 7552–7557. [CrossRef]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. CHEMOKINE RECEPTORS AS HIV-1 CORECEPTORS: Roles in Viral Entry, Tropism, and Disease. Annu. Rev. Immunol. 1999, 17, 657–700. [CrossRef]
- Bleul, C.C.; Wu, L.; Hoxie, J.A.; Springer, T.A.; Mackay, C.R. The HIV Coreceptors CXCR4 and CCR5 Are Differentially Expressed and Regulated on Human T Lymphocytes. Proc. Natl. Acad. Sci. 1997, 94, 1925–1930. [CrossRef]
- Weinberger, A.D.; Perelson, A.S. Persistence and Emergence of X4 Virus in HIV Infection. Math. Biosci. Eng. MBE 2011, 8, 605–626.
- Wu, L.; Paxton, W.A.; Kassam, N.; Ruffing, N.; Rottman, J.B.; Sullivan, N.; Choe, H.; Sodroski, J.; Newman, W.; Koup, R.A.; et al. CCR5 Levels and Expression Pattern Correlate with Infectability by Macrophage-Tropic HIV-1, In Vitro. J. Exp. Med. 1997, 185, 1681–1692. [CrossRef]
- Cashin, K.; Roche, M.; Sterjovski, J.; Ellett, A.; Gray, L.R.; Cunningham, A.L.; Ramsland, P.A.; Churchill, M.J.; Gorry, P.R. Alternative Coreceptor Requirements for Efficient CCR5- and CXCR4-Mediated HIV-1 Entry into Macrophages. J. Virol. 2011, 85, 10699–10709. [CrossRef]
- Jayakumar, P.; Berger, I.; Autschbach, F.; Weinstein, M.; Funke, B.; Verdin, E.; Goldsmith, M.A.; Keppler, O.T. Tissue-Resident Macrophages Are Productively Infected Ex Vivo by Primary X4 Isolates of Human Immunodeficiency Virus Type 1. J. Virol. 2005, 79, 5220–5226. [CrossRef]
- Borrajo, A.; Ranazzi, A.; Pollicita, M.; Bellocchi, M.C.; Salpini, R.; Mauro, M.V.; Ceccherini-Silberstein, F.; Perno, C.F.; Svicher, V.; Aquaro, S. Different Patterns of HIV-1 Replication in MACROPHAGES Is Led by Co-Receptor Usage. Medicina (Mex.) 2019, 55, 297. [CrossRef]
- Bonner, X.; Sondgeroth, A.; McCue, A.; Nicely, N.; Tripathy, A.; Spielvogel, E.; Moeser, M.; Ke, R.; Leiderman, K.; Joseph, S.B.; et al. Stoichiometry for Entry and Binding Properties of the Env Protein of R5 T Cell-Tropic HIV-1 and Its Evolutionary Variant of Macrophage-Tropic HIV-1. mBio 2024, 15, e0032124. [CrossRef]
- Okoye, A.A.; Picker, L.J. CD4+ T Cell Depletion in HIV Infection: Mechanisms of Immunological Failure. Immunol. Rev. 2013, 254, 54–64. [CrossRef]
- Douek, D.C.; Picker, L.J.; Koup, R.A. T Cell Dynamics in HIV-1 Infection. Annu. Rev. Immunol. 2003, 21, 265–304. [CrossRef]
- Blaak, H.; van’t Wout, A.B.; Brouwer, M.; Hooibrink, B.; Hovenkamp, E.; Schuitemaker, H. In Vivo HIV-1 Infection of CD45RA+CD4+ T Cells Is Established Primarily by Syncytium-Inducing Variants and Correlates with the Rate of CD4+ T Cell Decline. Proc. Natl. Acad. Sci. 2000, 97, 1269–1274. [CrossRef]
- Dejucq, N. HIV-1 Replication in CD4+ T Cell Lines: The Effects of Adaptation on Co-Receptor Use, Tropism, and Accessory Gene Function. J. Leukoc. Biol. 2000, 68, 331–337.
- Connor, R.I.; Sheridan, K.E.; Ceradini, D.; Choe, S.; Landau, N.R. Change in Coreceptor Use Correlates with Disease Progression in HIV-1–Infected Individuals. J. Exp. Med. 1997, 185, 621–628. [CrossRef]
- Weinberger, A.D.; Perelson, A.S.; Ribeiro, R.M.; Weinberger, L.S. Accelerated Immunodeficiency by Anti-CCR5 Treatment in HIV Infection. PLoS Comput. Biol. 2009, 5, e1000467. [CrossRef]
- Ivetic, A.; Hoskins Green, H.L.; Hart, S.J. L-Selectin: A Major Regulator of Leukocyte Adhesion, Migration and Signaling. Front. Immunol. 2019, 10. [CrossRef]
- Kononchik, J.; Ireland, J.; Zou, Z.; Segura, J.; Holzapfel, G.; Chastain, A.; Wang, R.; Spencer, M.; He, B.; Stutzman, N.; et al. HIV-1 Targets L-Selectin for Adhesion and Induces Its Shedding for Viral Release. Nat. Commun. 2018, 9, 2825. [CrossRef]
- Sallusto, F.; Schaerli, P.; Loetscher, P.; Schaniel, C.; Lenig, D.; Mackay, C.R.; Qin, S.; Lanzavecchia, A. Rapid and Coordinated Switch in Chemokine Receptor Expression during Dendritic Cell Maturation. Eur. J. Immunol. 1998, 28, 2760–2769. [CrossRef]
- Turville, S.G.; Cameron, P.U.; Handley, A.; Lin, G.; Pöhlmann, S.; Doms, R.W.; Cunningham, A.L. Diversity of Receptors Binding HIV on Dendritic Cell Subsets. Nat. Immunol. 2002, 3, 975–983. [CrossRef]
- McIlroy, D.; Autran, B.; Cheynier, R.; Wain-Hobson, S.; Clauvel, J.P.; Oksenhendler, E.; Debré, P.; Hosmalin, A. Infection Frequency of Dendritic Cells and CD4+ T Lymphocytes in Spleens of Human Immunodeficiency Virus-Positive Patients. J. Virol. 1995, 69, 4737–4745. [CrossRef]
- Felts, R.L.; Narayan, K.; Estes, J.D.; Shi, D.; Trubey, C.M.; Fu, J.; Hartnell, L.M.; Ruthel, G.T.; Schneider, D.K.; Nagashima, K.; et al. 3D Visualization of HIV Transfer at the Virological Synapse between Dendritic Cells and T Cells. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 13336–13341. [CrossRef]
- Fortin, J.F.; Cantin, R.; Lamontagne, G.; Tremblay, M. Host-Derived ICAM-1 Glycoproteins Incorporated on Human Immunodeficiency Virus Type 1 Are Biologically Active and Enhance Viral Infectivity. J. Virol. 1997, 71, 3588–3596. [CrossRef]
- Rizzuto, C.D.; Sodroski, J.G. Contribution of Virion ICAM-1 to Human Immunodeficiency Virus Infectivity and Sensitivity to Neutralization. J. Virol. 1997, 71, 4847–4851. [CrossRef]
- Bounou, S.; Giguere, J.-F.; Cantin, R.; Gilbert, C.; Imbeault, M.; Martin, G.; Tremblay, M.J. The Importance of Virus-Associated Host ICAM-1 in Human Immunodeficiency Virus Type 1 Dissemination Depends on the Cellular Context. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2004, 18, 1294–1296. [CrossRef]
- Ohno, H.; Aguilar, R.C.; Fournier, M.C.; Hennecke, S.; Cosson, P.; Bonifacino, J.S. Interaction of Endocytic Signals from the HIV-1 Envelope Glycoprotein Complex with Members of the Adaptor Medium Chain Family. Virology 1997, 238, 305–315. [CrossRef]
- Boge, M.; Wyss, S.; Bonifacino, J.S.; Thali, M. A Membrane-Proximal Tyrosine-Based Signal Mediates Internalization of the HIV-1 Envelope Glycoprotein via Interaction with the AP-2 Clathrin Adaptor. J. Biol. Chem. 1998, 273, 15773–15778. [CrossRef]
- Zaitseva, E.; Zaitsev, E.; Melikov, K.; Arakelyan, A.; Marin, M.; Villasmil, R.; Margolis, L.B.; Melikyan, G.B.; Chernomordik, L.V. FUSION STAGE OF HIV-1 ENTRY DEPENDS ON VIRUS-INDUCED CELL SURFACE EXPOSURE OF PHOSPHATIDYLSERINE. Cell Host Microbe 2017, 22, 99-110.e7. [CrossRef]
- Callahan, M.K.; Popernack, P.M.; Tsutsui, S.; Truong, L.; Schlegel, R.A.; Henderson, A.J. Phosphatidylserine on HIV Envelope Is a Cofactor for Infection of Monocytic Cells1. J. Immunol. 2003, 170, 4840–4845. [CrossRef]
- Lu, J.; Pan, Q.; Rong, L.; Liu, S.-L.; Liang, C. The IFITM Proteins Inhibit HIV-1 Infection. J. Virol. 2011, 85, 2126–2137. [CrossRef]
- Foster, T.L.; Wilson, H.; Iyer, S.S.; Coss, K.; Doores, K.; Smith, S.; Kellam, P.; Finzi, A.; Borrow, P.; Hahn, B.H.; et al. Resistance of Transmitted Founder HIV-1 to IFITM-Mediated Restriction. Cell Host Microbe 2016, 20, 429–442. [CrossRef]
- Tartour, K.; Nguyen, X.-N.; Appourchaux, R.; Assil, S.; Barateau, V.; Bloyet, L.-M.; Gaillard, J.B.; Confort, M.-P.; Escudero-Perez, B.; Gruffat, H.; et al. Interference with the Production of Infectious Viral Particles and Bimodal Inhibition of Replication Are Broadly Conserved Antiviral Properties of IFITMs. PLOS Pathog. 2017, 13, e1006610. [CrossRef]
- Wang, Y.; Pan, Q.; Ding, S.; Wang, Z.; Yu, J.; Finzi, A.; Liu, S.-L.; Liang, C. The V3 Loop of HIV-1 Env Determines Viral Susceptibility to IFITM3 Impairment of Viral Infectivity. J. Virol. 2017, 91, 10.1128/jvi.02441-16. [CrossRef]
- Amini-Bavil-Olyaee, S.; Choi, Y.J.; Lee, J.H.; Shi, M.; Huang, I.-C.; Farzan, M.; Jung, J.U. The Antiviral Effector IFITM3 Disrupts Intracellular Cholesterol Homeostasis to Block Viral Entry. Cell Host Microbe 2013, 13, 452–464. [CrossRef]
- Silva-Januário, M.E. da; Costa, C.S. da; Tavares, L.A.; Oliveira, A.K.; Januário, Y.C.; Carvalho, A.N. de; Cassiano, M.H.A.; Rodrigues, R.L.; Miller, M.E.; Palameta, S.; et al. HIV-1 Nef Changes the Proteome of T Cells Extracellular Vesicles Depleting IFITMs and Other Antiviral Factors. Mol. Cell. Proteomics 2023, 22. [CrossRef]
- Compton, A.A.; Bruel, T.; Porrot, F.; Mallet, A.; Sachse, M.; Euvrard, M.; Liang, C.; Casartelli, N.; Schwartz, O. IFITM Proteins Incorporated into HIV-1 Virions Impair Viral Fusion and Spread. Cell Host Microbe 2014, 16, 736–747. [CrossRef]
- Inuzuka, M.; Hayakawa, M.; Ingi, T. Serinc, an Activity-Regulated Protein Family, Incorporates Serine into Membrane Lipid Synthesis. J. Biol. Chem. 2005, 280, 35776–35783. [CrossRef]
- Usami, Y.; Wu, Y.; Göttlinger, H.G. SERINC3 and SERINC5 Restrict HIV-1 Infectivity and Are Counteracted by Nef. Nature 2015, 526, 218–223. [CrossRef]
- Sood, C.; Marin, M.; Chande, A.; Pizzato, M.; Melikyan, G.B. SERINC5 Protein Inhibits HIV-1 Fusion Pore Formation by Promoting Functional Inactivation of Envelope Glycoproteins. J. Biol. Chem. 2017, 292, 6014–6026. [CrossRef]
- Beitari, S.; Ding, S.; Pan, Q.; Finzi, A.; Liang, C. Effect of HIV-1 Env on SERINC5 Antagonism. J. Virol. 2017, 91, 10.1128/jvi.02214-16. [CrossRef]
- Grover, J.R.; Veatch, S.L.; Ono, A. Basic Motifs Target PSGL-1, CD43, and CD44 to Plasma Membrane Sites Where HIV-1 Assembles. J. Virol. 2015, 89, 454–467. [CrossRef]
- Fu, Y.; He, S.; Waheed, A.A.; Dabbagh, D.; Zhou, Z.; Trinité, B.; Wang, Z.; Yu, J.; Wang, D.; Li, F.; et al. PSGL-1 Restricts HIV-1 Infectivity by Blocking Virus Particle Attachment to Target Cells. Proc. Natl. Acad. Sci. 2020, 117, 9537–9545. [CrossRef]
- Suzuki, Y.; Craigie, R. The Road to Chromatin — Nuclear Entry of Retroviruses. Nat. Rev. Microbiol. 2007, 5, 187–196. [CrossRef]
- Hulme, A.E.; Perez, O.; Hope, T.J. Complementary Assays Reveal a Relationship between HIV-1 Uncoating and Reverse Transcription. Proc. Natl. Acad. Sci. 2011, 108, 9975–9980. [CrossRef]
- Mamede, J.I.; Cianci, G.C.; Anderson, M.R.; Hope, T.J. Early Cytoplasmic Uncoating Is Associated with Infectivity of HIV-1. Proc. Natl. Acad. Sci. 2017, 114, E7169–E7178. [CrossRef]
- Arhel, N.J.; Souquere-Besse, S.; Munier, S.; Souque, P.; Guadagnini, S.; Rutherford, S.; Prévost, M.; Allen, T.D.; Charneau, P. HIV-1 DNA Flap Formation Promotes Uncoating of the Pre-integration Complex at the Nuclear Pore. EMBO J. 2007, 26, 3025–3037. [CrossRef]
- Müller, T.G.; Zila, V.; Peters, K.; Schifferdecker, S.; Stanic, M.; Lucic, B.; Laketa, V.; Lusic, M.; Müller, B.; Kräusslich, H.-G. HIV-1 Uncoating by Release of Viral cDNA from Capsid-like Structures in the Nucleus of Infected Cells. eLife 2021, 10, e64776. [CrossRef]
- Selyutina, A.; Persaud, M.; Lee, K.; KewalRamani, V.; Diaz-Griffero, F. Nuclear Import of the HIV-1 Core Precedes Reverse Transcription and Uncoating. Cell Rep. 2020, 32, 108201. [CrossRef]
- Gifford, L.B.; Melikyan, G.B. HIV-1 Capsid Uncoating Is a Multistep Process That Proceeds through Defect Formation Followed by Disassembly of the Capsid Lattice. ACS Nano 2024, 18, 2928–2947. [CrossRef]
- Müller, T.G.; Zila, V.; Müller, B.; Kräusslich, H.-G. Nuclear Capsid Uncoating and Reverse Transcription of HIV-1. Annu. Rev. Virol. 2022, 9, 261–284. [CrossRef]
- Rensen, E.; Mueller, F.; Scoca, V.; Parmar, J.J.; Souque, P.; Zimmer, C.; Di Nunzio, F. Clustering and Reverse Transcription of HIV-1 Genomes in Nuclear Niches of Macrophages. EMBO J. 2021, 40, e105247. [CrossRef]
- Sumner, R.P.; Harrison, L.; Touizer, E.; Peacock, T.P.; Spencer, M.; Zuliani-Alvarez, L.; Towers, G.J. Disrupting HIV-1 Capsid Formation Causes cGAS Sensing of Viral DNA. EMBO J. 2020, 39, e103958. [CrossRef]
- Campbell, E.M.; Hope, T.J. HIV-1 Capsid: The Multifaceted Key Player in HIV-1 Infection. Nat. Rev. Microbiol. 2015, 13, 471–483. [CrossRef]
- Rebensburg, S.V.; Wei, G.; Larue, R.C.; Lindenberger, J.; Francis, A.C.; Annamalai, A.S.; Morrison, J.; Shkriabai, N.; Huang, S.-W.; KewalRamani, V.; et al. Sec24C Is an HIV-1 Host Dependency Factor Crucial for Virus Replication. Nat. Microbiol. 2021, 6, 435–444. [CrossRef]
- Guth, C.A.; Sodroski, J. Contribution of PDZD8 to Stabilization of the Human Immunodeficiency Virus Type 1 Capsid. J. Virol. 2014, 88, 4612–4623. [CrossRef]
- Malikov, V.; da Silva, E.S.; Jovasevic, V.; Bennett, G.; de Souza Aranha Vieira, D.A.; Schulte, B.; Diaz-Griffero, F.; Walsh, D.; Naghavi, M.H. HIV-1 Capsids Bind and Exploit the Kinesin-1 Adaptor FEZ1 for Inward Movement to the Nucleus. Nat. Commun. 2015, 6, 6660. [CrossRef]
- Huang, P.-T.; Summers, B.J.; Xu, C.; Perilla, J.R.; Malikov, V.; Naghavi, M.H.; Xiong, Y. FEZ1 Is Recruited to a Conserved Cofactor Site on Capsid to Promote HIV-1 Trafficking. Cell Rep. 2019, 28, 2373-2385.e7. [CrossRef]
- Carnes, S.K.; Zhou, J.; Aiken, C. HIV-1 Engages a Dynein-Dynactin-BICD2 Complex for Infection and Transport to the Nucleus. J. Virol. 2018, 92, 10.1128/jvi.00358-18. [CrossRef]
- Dharan, A.; Opp, S.; Abdel-Rahim, O.; Keceli, S.K.; Imam, S.; Diaz-Griffero, F.; Campbell, E.M. Bicaudal D2 Facilitates the Cytoplasmic Trafficking and Nuclear Import of HIV-1 Genomes during Infection. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E10707–E10716. [CrossRef]
- Dharan, A.; Talley, S.; Tripathi, A.; Mamede, J.I.; Majetschak, M.; Hope, T.J.; Campbell, E.M. KIF5B and Nup358 Cooperatively Mediate the Nuclear Import of HIV-1 during Infection. PLOS Pathog. 2016, 12, e1005700. [CrossRef]
- Opp, S.; Martinez-Lopez, A.; Fricke, T.; Buffone, C.; Severgnini, M.; Cifola, I.; Frabetti, S.; Skorupka, K.; Zadrozny, K.K.; Ganser-Pornillos, B.K.; et al. Nup153 Unlocks the Nuclear Pore Complex for HIV-1 Nuclear Import in Non-Dividing Cells 2018.
- Fernandez, J.; Machado, A.K.; Lyonnais, S.; Chamontin, C.; Gärtner, K.; Léger, T.; Henriquet, C.; Garcia, C.; Portilho, D.M.; Pugnière, M.; et al. Transportin-1 Binds to the HIV-1 Capsid via a Nuclear Localization Signal and Triggers Uncoating. Nat. Microbiol. 2019, 4, 1840–1850. [CrossRef]
- Shah, V.B.; Shi, J.; Hout, D.R.; Oztop, I.; Krishnan, L.; Ahn, J.; Shotwell, M.S.; Engelman, A.; Aiken, C. The Host Proteins Transportin SR2/TNPO3 and Cyclophilin A Exert Opposing Effects on HIV-1 Uncoating. J. Virol. 2013, 87, 422–432. [CrossRef]
- Misumi, S.; Inoue, M.; Dochi, T.; Kishimoto, N.; Hasegawa, N.; Takamune, N.; Shoji, S. Uncoating of Human Immunodeficiency Virus Type 1 Requires Prolyl Isomerase Pin1. J. Biol. Chem. 2010, 285, 25185–25195. [CrossRef]
- McDonald, D.; Vodicka, M.A.; Lucero, G.; Svitkina, T.M.; Borisy, G.G.; Emerman, M.; Hope, T.J. Visualization of the Intracellular Behavior of HIV in Living Cells. J. Cell Biol. 2002, 159, 441–452. [CrossRef]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: A Key Player for Human Disease. Cell Death Dis. 2013, 4, e888–e888. [CrossRef]
- Luban, J.; Bossolt, K.L.; Franke, E.K.; Kalpana, G.V.; Goff, S.P. Human Immunodeficiency Virus Type 1 Gag Protein Binds to Cyclophilins A and B. Cell 1993, 73, 1067–1078. [CrossRef]
- Franke, E.K.; Yuan, H.E.; Luban, J. Specific Incorporation of Cyclophilin A into HIV-1 Virions. Nature 1994, 372, 359–362. [CrossRef]
- Thali, M.; Bukovsky, A.; Kondo, E.; Rosenwirth, B.; Walsh, C.T.; Sodroski, J.; Göttlinger, H.G. Functional Association of Cyclophilin A with HIV-1 Virions. Nature 1994, 372, 363–365. [CrossRef]
- Braaten, D.; Luban, J. Cyclophilin A Regulates HIV-1 Infectivity, as Demonstrated by Gene Targeting in Human T Cells. EMBO J. 2001, 20, 1300–1309. [CrossRef]
- Padron, A.; Prakash, P.; Pandhare, J.; Luban, J.; Aiken, C.; Balasubramaniam, M.; Dash, C. Emerging Role of Cyclophilin A in HIV-1 Infection: From Producer Cell to the Target Cell Nucleus. J. Virol. 2023, 97, e00732-23. [CrossRef]
- Gamble, T.R.; Vajdos, F.F.; Yoo, S.; Worthylake, D.K.; Houseweart, M.; Sundquist, W.I.; Hill, C.P. Crystal Structure of Human Cyclophilin A Bound to the Amino-Terminal Domain of HIV-1 Capsid. Cell 1996, 87, 1285–1294. [CrossRef]
- Perez-Caballero, D.; Hatziioannou, T.; Zhang, F.; Cowan, S.; Bieniasz, P.D. Restriction of Human Immunodeficiency Virus Type 1 by TRIM-CypA Occurs with Rapid Kinetics and Independently of Cytoplasmic Bodies, Ubiquitin, and Proteasome Activity. J. Virol. 2005, 79, 15567–15572. [CrossRef]
- Hulme, A.E.; Kelley, Z.; Okocha, E.A.; Hope, T.J. Identification of Capsid Mutations That Alter the Rate of HIV-1 Uncoating in Infected Cells. J. Virol. 2014, 89, 643–651. [CrossRef]
- Liu, C.; Perilla, J.R.; Ning, J.; Lu, M.; Hou, G.; Ramalho, R.; Himes, B.A.; Zhao, G.; Bedwell, G.J.; Byeon, I.-J.; et al. Cyclophilin A Stabilizes the HIV-1 Capsid through a Novel Non-Canonical Binding Site. Nat. Commun. 2016, 7, 10714. [CrossRef]
- Selyutina, A.; Persaud, M.; Simons, L.M.; Bulnes-Ramos, A.; Buffone, C.; Martinez-Lopez, A.; Scoca, V.; Di Nunzio, F.; Hiatt, J.; Marson, A.; et al. Cyclophilin A Prevents HIV-1 Restriction in Lymphocytes by Blocking Human TRIM5α Binding to the Viral Core. Cell Rep. 2020, 30, 3766-3777.e6. [CrossRef]
- Kim, K.; Dauphin, A.; Komurlu, S.; McCauley, S.M.; Yurkovetskiy, L.; Carbone, C.; Diehl, W.E.; Strambio-De-Castillia, C.; Campbell, E.M.; Luban, J. Cyclophilin A Protects HIV-1 from Restriction by Human TRIM5α. Nat. Microbiol. 2019, 4, 2044–2051. [CrossRef]
- Hatziioannou, T.; Perez-Caballero, D.; Cowan, S.; Bieniasz, P.D. Cyclophilin Interactions with Incoming Human Immunodeficiency Virus Type 1 Capsids with Opposing Effects on Infectivity in Human Cells. J. Virol. 2005, 79, 176–183. [CrossRef]
- Li, Y.; Kar, A.K.; Sodroski, J. Target Cell Type-Dependent Modulation of Human Immunodeficiency Virus Type 1 Capsid Disassembly by Cyclophilin A. J. Virol. 2009, 83, 10951–10962. [CrossRef]
- Mallery, D.L.; Márquez, C.L.; McEwan, W.A.; Dickson, C.F.; Jacques, D.A.; Anandapadamanaban, M.; Bichel, K.; Towers, G.J.; Saiardi, A.; Böcking, T.; et al. IP6 Is an HIV Pocket Factor That Prevents Capsid Collapse and Promotes DNA Synthesis. eLife 2018, 7, e35335. [CrossRef]
- Papa, G.; Albecka, A.; Mallery, D.; Vaysburd, M.; Renner, N.; James, L.C. IP6-Stabilised HIV Capsids Evade cGAS/STING-Mediated Host Immune Sensing. EMBO Rep. 2023, 24, e56275. [CrossRef]
- Bejarano, D.A.; Peng, K.; Laketa, V.; Börner, K.; Jost, K.L.; Lucic, B.; Glass, B.; Lusic, M.; Müller, B.; Kräusslich, H.-G. HIV-1 Nuclear Import in Macrophages Is Regulated by CPSF6-Capsid Interactions at the Nuclear Pore Complex. eLife 2019, 8, e41800. [CrossRef]
- Chin, C.R.; Perreira, J.M.; Savidis, G.; Portmann, J.M.; Aker, A.M.; Feeley, E.M.; Smith, M.C.; Brass, A.L. Direct Visualization of HIV-1 Replication Intermediates Shows That Capsid and CPSF6 Modulate HIV-1 Intra-Nuclear Invasion and Integration. Cell Rep. 2015, 13, 1717–1731. [CrossRef]
- Francis, A.C.; Marin, M.; Singh, P.K.; Achuthan, V.; Prellberg, M.J.; Palermino-Rowland, K.; Lan, S.; Tedbury, P.R.; Sarafianos, S.G.; Engelman, A.N.; et al. HIV-1 Replication Complexes Accumulate in Nuclear Speckles and Integrate into Speckle-Associated Genomic Domains. Nat. Commun. 2020, 11, 3505. [CrossRef]
- Achuthan, V.; Perreira, J.M.; Ahn, J.J.; Brass, A.L.; Engelman, A.N. Capsid-CPSF6 Interaction: Master Regulator of Nuclear HIV-1 Positioning and Integration. J. Life Sci. Westlake Village Calif 2019, 1, 39–45. [CrossRef]
- Bukrinsky, M.I.; Sharova, N.; Dempsey, M.P.; Stanwick, T.L.; Bukrinskaya, A.G.; Haggerty, S.; Stevenson, M. Active Nuclear Import of Human Immunodeficiency Virus Type 1 Preintegration Complexes. Proc. Natl. Acad. Sci. U. S. A. 1992, 89, 6580–6584. [CrossRef]
- Ananth, S.; Ambiel, I.; Schifferdecker, S.; Müller, T.G.; Wratil, P.R.; Mejias-Perez, E.; Kräusslich, H.-G.; Müller, B.; Keppler, O.T.; Fackler, O.T. Spatial Resolution of HIV-1 Post-Entry Steps in Resting CD4 T Cells. Cell Rep. 2024, 43. [CrossRef]
- Sowd, G.A.; Serrao, E.; Wang, H.; Wang, W.; Fadel, H.J.; Poeschla, E.M.; Engelman, A.N. A Critical Role for Alternative Polyadenylation Factor CPSF6 in Targeting HIV-1 Integration to Transcriptionally Active Chromatin. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, E1054-1063. [CrossRef]
- Stremlau, M.; Owens, C.M.; Perron, M.J.; Kiessling, M.; Autissier, P.; Sodroski, J. The Cytoplasmic Body Component TRIM5alpha Restricts HIV-1 Infection in Old World Monkeys. Nature 2004, 427, 848–853. [CrossRef]
- Li, Y.; Li, X.; Stremlau, M.; Lee, M.; Sodroski, J. Removal of Arginine 332 Allows Human TRIM5alpha to Bind Human Immunodeficiency Virus Capsids and to Restrict Infection. J. Virol. 2006, 80, 6738–6744. [CrossRef]
- Black, L.R.; Aiken, C. TRIM5alpha Disrupts the Structure of Assembled HIV-1 Capsid Complexes in Vitro. J. Virol. 2010, 84, 6564–6569. [CrossRef]
- Jimenez-Guardeño, J.M.; Apolonia, L.; Betancor, G.; Malim, M.H. Immunoproteasome Activation Enables Human TRIM5α Restriction of HIV-1. Nat. Microbiol. 2019, 4, 933–940. [CrossRef]
- Perez-Caballero, D.; Hatziioannou, T.; Yang, A.; Cowan, S.; Bieniasz, P.D. Human Tripartite Motif 5alpha Domains Responsible for Retrovirus Restriction Activity and Specificity. J. Virol. 2005, 79, 8969–8978. [CrossRef]
- Ribeiro, C.M.S.; Sarrami-Forooshani, R.; Setiawan, L.C.; Zijlstra-Willems, E.M.; van Hamme, J.L.; Tigchelaar, W.; van der Wel, N.N.; Kootstra, N.A.; Gringhuis, S.I.; Geijtenbeek, T.B.H. Receptor Usage Dictates HIV-1 Restriction by Human TRIM5α in Dendritic Cell Subsets. Nature 2016, 540, 448–452. [CrossRef]
- Pertel, T.; Hausmann, S.; Morger, D.; Züger, S.; Guerra, J.; Lascano, J.; Reinhard, C.; Santoni, F.A.; Uchil, P.D.; Chatel, L.; et al. TRIM5 Is an Innate Immune Sensor for the Retrovirus Capsid Lattice. Nature 2011, 472, 361–365. [CrossRef]
- Lascano, J.; Uchil, P.D.; Mothes, W.; Luban, J. TRIM5 Retroviral Restriction Activity Correlates with the Ability To Induce Innate Immune Signaling. J. Virol. 2016, 90, 308–316. [CrossRef]
- Fletcher, A.J.; Vaysburd, M.; Maslen, S.; Zeng, J.; Skehel, J.M.; Towers, G.J.; James, L.C. Trivalent RING Assembly on Retroviral Capsids Activates TRIM5 Ubiquitination and Innate Immune Signaling. Cell Host Microbe 2018, 24, 761-775.e6. [CrossRef]
- Yoh, S.M.; Schneider, M.; Seifried, J.; Soonthornvacharin, S.; Akleh, R.E.; Olivieri, K.C.; De Jesus, P.D.; Ruan, C.; de Castro, E.; Ruiz, P.A.; et al. PQBP1 Is a Proximal Sensor of the cGAS-Dependent Innate Response to HIV-1. Cell 2015, 161, 1293–1305. [CrossRef]
- Yoh, S.M.; Mamede, J.I.; Lau, D.; Ahn, N.; Sánchez-Aparicio, M.T.; Temple, J.; Tuckwell, A.; Fuchs, N.V.; Cianci, G.C.; Riva, L.; et al. Recognition of HIV-1 Capsid by PQBP1 Licenses an Innate Immune Sensing of Nascent HIV-1 DNA. Mol. Cell 2022, 82, 2871-2884.e6. [CrossRef]
- Yuan, T.; Yao, W.; Tokunaga, K.; Yang, R.; Sun, B. An HIV-1 Capsid Binding Protein TRIM11 Accelerates Viral Uncoating. Retrovirology 2016, 13, 72. [CrossRef]
- Staeheli, P.; Haller, O. Human MX2/MxB: A Potent Interferon-Induced Postentry Inhibitor of Herpesviruses and HIV-1. J. Virol. 2018, 92, 10.1128/jvi.00709-18. [CrossRef]
- Kane, M.; Yadav, S.S.; Bitzegeio, J.; Kutluay, S.B.; Zang, T.; Wilson, S.J.; Schoggins, J.W.; Rice, C.M.; Yamashita, M.; Hatziioannou, T.; et al. MX2 Is an Interferon-Induced Inhibitor of HIV-1 Infection. Nature 2013, 502, 563–566. [CrossRef]
- Smaga, S.S.; Xu, C.; Summers, B.J.; Digianantonio, K.M.; Perilla, J.R.; Xiong, Y. MxB Restricts HIV-1 by Targeting the Tri-Hexamer Interface of the Viral Capsid. Struct. Lond. Engl. 1993 2019, 27, 1234-1245.e5. [CrossRef]
- Fricke, T.; White, T.E.; Schulte, B.; de Souza Aranha Vieira, D.A.; Dharan, A.; Campbell, E.M.; Brandariz-Nuñez, A.; Diaz-Griffero, F. MxB Binds to the HIV-1 Core and Prevents the Uncoating Process of HIV-1. Retrovirology 2014, 11, 68. [CrossRef]
- Melén, K.; Keskinen, P.; Ronni, T.; Sareneva, T.; Lounatmaa, K.; Julkunen, I. Human MxB Protein, an Interferon-Alpha-Inducible GTPase, Contains a Nuclear Targeting Signal and Is Localized in the Heterochromatin Region beneath the Nuclear Envelope. J. Biol. Chem. 1996, 271, 23478–23486. [CrossRef]
- Betancor, G. You Shall Not Pass: MX2 Proteins Are Versatile Viral Inhibitors. Vaccines 2023, 11, 930. [CrossRef]
- Maillet, S.; Fernandez, J.; Decourcelle, M.; El Koulali, K.; Blanchet, F.P.; Arhel, N.J.; Maarifi, G.; Nisole, S. Daxx Inhibits HIV-1 Reverse Transcription and Uncoating in a SUMO-Dependent Manner. Viruses 2020, 12, 636. [CrossRef]
- Yang, Y.; Fricke, T.; Diaz-Griffero, F. Inhibition of Reverse Transcriptase Activity Increases Stability of the HIV-1 Core. J. Virol. 2013, 87, 683–687. [CrossRef]
- Cosnefroy, O.; Murray, P.J.; Bishop, K.N. HIV-1 Capsid Uncoating Initiates after the First Strand Transfer of Reverse Transcription. Retrovirology 2016, 13, 58. [CrossRef]
- Iordanskiy, S.; Bukrinsky, M. Reverse Transcription Complex: The Key Player of the Early Phase of HIV Replication. Future Virol. 2007, 2, 49–64. [CrossRef]
- Arhel, N. Revisiting HIV-1 Uncoating. Retrovirology 2010, 7, 96. [CrossRef]
- Jacques, D.A.; McEwan, W.A.; Hilditch, L.; Price, A.J.; Towers, G.J.; James, L.C. HIV-1 Uses Dynamic Capsid Pores to Import Nucleotides and Fuel Encapsidated DNA Synthesis. Nature 2016, 536, 349–353. [CrossRef]
- Basu, V.P.; Song, M.; Gao, L.; Rigby, S.T.; Hanson, M.N.; Bambara, R.A. Strand Transfer Events during HIV-1 Reverse Transcription. Virus Res. 2008, 134, 19–38. [CrossRef]
- Bukrinskaya, A.; Brichacek, B.; Mann, A.; Stevenson, M. Establishment of a Functional Human Immunodeficiency Virus Type 1 (HIV-1) Reverse Transcription Complex Involves the Cytoskeleton. J. Exp. Med. 1998, 188, 2113–2125. [CrossRef]
- Larguet, F.; Caté, C.; Barbeau, B.; Rassart, E.; Edouard, E. Histone Deacetylase 1 Interacts with HIV-1 Integrase and Modulates Viral Replication. Virol. J. 2019, 16, 138. [CrossRef]
- Sorin, M.; Cano, J.; Das, S.; Mathew, S.; Wu, X.; Davies, K.P.; Shi, X.; Cheng, S.-W.G.; Ott, D.; Kalpana, G.V. Recruitment of a SAP18-HDAC1 Complex into HIV-1 Virions and Its Requirement for Viral Replication. PLoS Pathog. 2009, 5, e1000463. [CrossRef]
- Sorin, M.; Yung, E.; Wu, X.; Kalpana, G.V. HIV-1 Replication in Cell Lines Harboring INI1/hSNF5 Mutations. Retrovirology 2006, 3, 56. [CrossRef]
- Takahashi, H.; Matsuda, M.; Kojima, A.; Sata, T.; Andoh, T.; Kurata, T.; Nagashima, K.; Hall, W.W. Human Immunodeficiency Virus Type 1 Reverse Transcriptase: Enhancement of Activity by Interaction with Cellular Topoisomerase I. Proc. Natl. Acad. Sci. 1995, 92, 5694–5698. [CrossRef]
- Staszewski, J.; Lazarewicz, N.; Konczak, J.; Migdal, I.; Maciaszczyk-Dziubinska, E. UPF1—From mRNA Degradation to Human Disorders. Cells 2023, 12, 419. [CrossRef]
- Serquiña, A.K.P.; Das, S.R.; Popova, E.; Ojelabi, O.A.; Roy, C.K.; Göttlinger, H.G. UPF1 Is Crucial for the Infectivity of Human Immunodeficiency Virus Type 1 Progeny Virions. J. Virol. 2013, 87, 8853–8861. [CrossRef]
- Lemay, J.; Maidou-Peindara, P.; Bader, T.; Ennifar, E.; Rain, J.-C.; Benarous, R.; Liu, L.X. HuR Interacts with Human Immunodeficiency Virus Type 1 Reverse Transcriptase, and Modulates Reverse Transcription in Infected Cells. Retrovirology 2008, 5, 47. [CrossRef]
- Lemay, J.; Maidou-Peindara, P.; Cancio, R.; Ennifar, E.; Coadou, G.; Maga, G.; Rain, J.-C.; Benarous, R.; Liu, L.X. AKAP149 Binds to HIV-1 Reverse Transcriptase and Is Involved in the Reverse Transcription. J. Mol. Biol. 2008, 383, 783–796. [CrossRef]
- Warren, K.; Warrilow, D.; Meredith, L.; Harrich, D. Reverse Transcriptase and Cellular Factors: Regulators of HIV-1 Reverse Transcription. Viruses 2009, 1, 873–894. [CrossRef]
- Meister, G.; Bühler, D.; Pillai, R.; Lottspeich, F.; Fischer, U. A Multiprotein Complex Mediates the ATP-Dependent Assembly of Spliceosomal U snRNPs. Nat. Cell Biol. 2001, 3, 945–949. [CrossRef]
- Hamamoto, S.; Nishitsuji, H.; Amagasa, T.; Kannagi, M.; Masuda, T. Identification of a Novel Human Immunodeficiency Virus Type 1 Integrase Interactor, Gemin2, That Facilitates Efficient Viral cDNA Synthesis In Vivo. J. Virol. 2006, 80, 5670–5677. [CrossRef]
- Mazur, D.J.; Perrino, F.W. Identification and Expression of the TREX1 and TREX2 cDNA Sequences Encoding Mammalian 3′→5′ Exonucleases*. J. Biol. Chem. 1999, 274, 19655–19660. [CrossRef]
- Hasan, M.; Yan, N. Safeguard against DNA Sensing: The Role of TREX1 in HIV-1 Infection and Autoimmune Diseases. Front. Microbiol. 2014, 5, 193. [CrossRef]
- Yan, N.; Regalado-Magdos, A.D.; Stiggelbout, B.; Lee-Kirsch, M.A.; Lieberman, J. The Cytosolic Exonuclease TREX1 Inhibits the Innate Immune Response to Human Immunodeficiency Virus Type 1. Nat. Immunol. 2010, 11, 1005–1013. [CrossRef]
- Radetskyy, R.; Daher, A.; Gatignol, A. ADAR1 and PKR, Interferon Stimulated Genes with Clashing Effects on HIV-1 Replication. Cytokine Growth Factor Rev. 2018, 40, 48–58. [CrossRef]
- Cuadrado, E.; Booiman, T.; van Hamme, J.L.; Jansen, M.H.; van Dort, K.A.; Vanderver, A.; Rice, G.I.; Crow, Y.J.; Kootstra, N.A.; Kuijpers, T.W. ADAR1 Facilitates HIV-1 Replication in Primary CD4+ T Cells. PloS One 2015, 10, e0143613. [CrossRef]
- Liddicoat, B.J.; Piskol, R.; Chalk, A.M.; Ramaswami, G.; Higuchi, M.; Hartner, J.C.; Li, J.B.; Seeburg, P.H.; Walkley, C.R. RNA Editing by ADAR1 Prevents MDA5 Sensing of Endogenous dsRNA as Nonself. Science 2015, 349, 1115–1120. [CrossRef]
- Friedrich, B.M.; Murray, J.L.; Li, G.; Sheng, J.; Hodge, T.W.; Rubin, D.H.; O’Brien, W.A.; Ferguson, M.R. A Functional Role for ADAM10 in Human Immunodeficiency Virus Type-1 Replication. Retrovirology 2011, 8, 32. [CrossRef]
- Endsley, M.A.; Somasunderam, A.D.; Li, G.; Oezguen, N.; Thiviyanathan, V.; Murray, J.L.; Rubin, D.H.; Hodge, T.W.; O’Brien, W.A.; Lewis, B.; et al. Nuclear Trafficking of the HIV-1 Pre-Integration Complex Depends on the ADAM10 Intracellular Domain. Virology 2014, 0, 60–66. [CrossRef]
- Zhang, H.; Dornadula, G.; Orenstein, J.; Pomerantz, R.J. Morphologic Changes in Human Immunodeficiency Virus Type 1 Virions Secondary to Intravirion Reverse Transcription: Evidence Indicating That Reverse Transcription May Not Take Place within the Intact Viral Core. J. Hum. Virol. 2000, 3, 165–172.
- Ao, Z.; Danappa Jayappa, K.; Wang, B.; Zheng, Y.; Kung, S.; Rassart, E.; Depping, R.; Kohler, M.; Cohen, E.A.; Yao, X. Importin A3 Interacts with HIV-1 Integrase and Contributes to HIV-1 Nuclear Import and Replication. J. Virol. 2010, 84, 8650–8663. [CrossRef]
- Gallay, P.; Hope, T.; Chin, D.; Trono, D. HIV-1 Infection of Nondividing Cells through the Recognition of Integrase by the Importin/Karyopherin Pathway. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 9825–9830.
- Solis, M.; Nakhaei, P.; Jalalirad, M.; Lacoste, J.; Douville, R.; Arguello, M.; Zhao, T.; Laughrea, M.; Wainberg, M.A.; Hiscott, J. RIG-I-Mediated Antiviral Signaling Is Inhibited in HIV-1 Infection by a Protease-Mediated Sequestration of RIG-I. J. Virol. 2011, 85, 1224–1236. [CrossRef]
- Thoresen, D.; Wang, W.; Galls, D.; Guo, R.; Xu, L.; Pyle, A.M. The Molecular Mechanism of RIG-I Activation and Signaling. Immunol. Rev. 2021, 304, 154–168. [CrossRef]
- Guney, M.H.; Nagalekshmi, K.; McCauley, S.M.; Carbone, C.; Aydemir, O.; Luban, J. IFIH1 (MDA5) Is Required for Innate Immune Detection of Intron-Containing RNA Expressed from the HIV-1 Provirus. BioRxiv Prepr. Serv. Biol. 2023, 2023.11.17.567619. [CrossRef]
- Pujantell, M.; Badia, R.; Ramirez, C.; Puig, T.; Clotet, B.; Ballana, E.; Esté, J.A.; Riveira-Muñoz, E. Long-Term HIV-1 Infection Induces an Antiviral State in Primary Macrophages. Antiviral Res. 2016, 133, 145–155. [CrossRef]
- Chattergoon, M.A.; Latanich, R.; Quinn, J.; Winter, M.E.; Buckheit, R.W.; Blankson, J.N.; Pardoll, D.; Cox, A.L. HIV and HCV Activate the Inflammasome in Monocytes and Macrophages via Endosomal Toll-like Receptors without Induction of Type 1 Interferon. PLoS Pathog. 2014, 10, e1004082. [CrossRef]
- Jakobsen, M.R.; Bak, R.O.; Andersen, A.; Berg, R.K.; Jensen, S.B.; Tengchuan, J.; Laustsen, A.; Hansen, K.; Ostergaard, L.; Fitzgerald, K.A.; et al. IFI16 Senses DNA Forms of the Lentiviral Replication Cycle and Controls HIV-1 Replication. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, E4571-4580. [CrossRef]
- Itell, H.L.; Humes, D.; Overbaugh, J. Several Cell-Intrinsic Effectors Drive Type I Interferon-Mediated Restriction of HIV-1 in Primary CD4+ T Cells. Cell Rep. 2023, 42, 112556. [CrossRef]
- Bosso, M.; Bozzo, C.P.; Volcic, M.; Kirchhoff, F. IFI16 Knockdown in Primary HIV-1 Target Cells. STAR Protoc. 2021, 2, 100236. [CrossRef]
- Gao, D.; Wu, J.; Wu, Y.-T.; Du, F.; Aroh, C.; Yan, N.; Sun, L.; Chen, Z.J. Cyclic GMP-AMP Synthase Is an Innate Immune Sensor of HIV and Other Retroviruses. Science 2013, 341, 903–906. [CrossRef]
- Elsner, C.; Ponnurangam, A.; Kazmierski, J.; Zillinger, T.; Jansen, J.; Todt, D.; Döhner, K.; Xu, S.; Ducroux, A.; Kriedemann, N.; et al. Absence of cGAS-Mediated Type I IFN Responses in HIV-1–Infected T Cells. Proc. Natl. Acad. Sci. 2020, 117, 19475–19486. [CrossRef]
- Lahaye, X.; Satoh, T.; Gentili, M.; Cerboni, S.; Conrad, C.; Hurbain, I.; El Marjou, A.; Lacabaratz, C.; Lelièvre, J.-D.; Manel, N. The Capsids of HIV-1 and HIV-2 Determine Immune Detection of the Viral cDNA by the Innate Sensor cGAS in Dendritic Cells. Immunity 2013, 39, 1132–1142. [CrossRef]
- Manel, N.; Hogstad, B.; Wang, Y.; Levy, D.E.; Unutmaz, D.; Littman, D.R. A Cryptic Sensor for HIV-1 Activates Antiviral Innate Immunity in Dendritic Cells. Nature 2010, 467, 214–217. [CrossRef]
- Beignon, A.-S.; McKenna, K.; Skoberne, M.; Manches, O.; DaSilva, I.; Kavanagh, D.G.; Larsson, M.; Gorelick, R.J.; Lifson, J.D.; Bhardwaj, N. Endocytosis of HIV-1 Activates Plasmacytoid Dendritic Cells via Toll-like Receptor– Viral RNA Interactions. J. Clin. Invest. 2005, 115, 3265–3275. [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [CrossRef]
- Goujon, C.; Malim, M.H. Characterization of the Alpha Interferon-Induced Postentry Block to HIV-1 Infection in Primary Human Macrophages and T Cells. J. Virol. 2010, 84, 9254–9266. [CrossRef]
- Sheehy, A.M.; Gaddis, N.C.; Choi, J.D.; Malim, M.H. Isolation of a Human Gene That Inhibits HIV-1 Infection and Is Suppressed by the Viral Vif Protein. Nature 2002, 418, 646–650. [CrossRef]
- Stopak, K.; de Noronha, C.; Yonemoto, W.; Greene, W.C. HIV-1 Vif Blocks the Antiviral Activity of APOBEC3G by Impairing Both Its Translation and Intracellular Stability. Mol. Cell 2003, 12, 591–601. [CrossRef]
- Zhang, H.; Yang, B.; Pomerantz, R.J.; Zhang, C.; Arunachalam, S.C.; Gao, L. The Cytidine Deaminase CEM15 Induces Hypermutation in Newly Synthesized HIV-1 DNA. Nature 2003, 424, 94–98. [CrossRef]
- Hultquist, J.F.; Lengyel, J.A.; Refsland, E.W.; LaRue, R.S.; Lackey, L.; Brown, W.L.; Harris, R.S. Human and Rhesus APOBEC3D, APOBEC3F, APOBEC3G, and APOBEC3H Demonstrate a Conserved Capacity to Restrict Vif-Deficient HIV-1. J. Virol. 2011, 85, 11220–11234. [CrossRef]
- Harris, R.S.; Bishop, K.N.; Sheehy, A.M.; Craig, H.M.; Petersen-Mahrt, S.K.; Watt, I.N.; Neuberger, M.S.; Malim, M.H. DNA Deamination Mediates Innate Immunity to Retroviral Infection. Cell 2003, 113, 803–809. [CrossRef]
- Harris, R.S.; Petersen-Mahrt, S.K.; Neuberger, M.S. RNA Editing Enzyme APOBEC1 and Some of Its Homologs Can Act as DNA Mutators. Mol. Cell 2002, 10, 1247–1253. [CrossRef]
- Khan, M.A.; Kao, S.; Miyagi, E.; Takeuchi, H.; Goila-Gaur, R.; Opi, S.; Gipson, C.L.; Parslow, T.G.; Ly, H.; Strebel, K. Viral RNA Is Required for the Association of APOBEC3G with Human Immunodeficiency Virus Type 1 Nucleoprotein Complexes. J. Virol. 2005, 79, 5870–5874. [CrossRef]
- Yu, Q.; König, R.; Pillai, S.; Chiles, K.; Kearney, M.; Palmer, S.; Richman, D.; Coffin, J.M.; Landau, N.R. Single-Strand Specificity of APOBEC3G Accounts for Minus-Strand Deamination of the HIV Genome. Nat. Struct. Mol. Biol. 2004, 11, 435–442. [CrossRef]
- Miyagi, E.; Schwartzkopff, F.; Plishka, R.; Buckler-White, A.; Clouse, K.A.; Strebel, K. APOBEC3G-Independent Reduction in Virion Infectivity during Long-Term HIV-1 Replication in Terminally Differentiated Macrophages. Virology 2008, 379, 266–274. [CrossRef]
- Malim, M.H. APOBEC Proteins and Intrinsic Resistance to HIV-1 Infection. Philos. Trans. R. Soc. B Biol. Sci. 2008, 364, 675–687. [CrossRef]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M.; Perrin, L.; Trono, D. Broad Antiretroviral Defence by Human APOBEC3G through Lethal Editing of Nascent Reverse Transcripts. Nature 2003, 424, 99–103. [CrossRef]
- Adolph, M.B.; Ara, A.; Feng, Y.; Wittkopp, C.J.; Emerman, M.; Fraser, J.S.; Chelico, L. Cytidine Deaminase Efficiency of the Lentiviral Viral Restriction Factor APOBEC3C Correlates with Dimerization. Nucleic Acids Res. 2017, 45, 3378–3394. [CrossRef]
- Holmes, R.K.; Malim, M.H.; Bishop, K.N. APOBEC-Mediated Viral Restriction: Not Simply Editing? Trends Biochem. Sci. 2007, 32, 118–128. [CrossRef]
- Wang, X.; Ao, Z.; Chen, L.; Kobinger, G.; Peng, J.; Yao, X. The Cellular Antiviral Protein APOBEC3G Interacts with HIV-1 Reverse Transcriptase and Inhibits Its Function during Viral Replication. J. Virol. 2012, 86, 3777–3786. [CrossRef]
- Bishop, K.N.; Holmes, R.K.; Malim, M.H. Antiviral Potency of APOBEC Proteins Does Not Correlate with Cytidine Deamination. J. Virol. 2006, 80, 8450–8458. [CrossRef]
- Gillick, K.; Pollpeter, D.; Phalora, P.; Kim, E.-Y.; Wolinsky, S.M.; Malim, M.H. Suppression of HIV-1 Infection by APOBEC3 Proteins in Primary Human CD4+ T Cells Is Associated with Inhibition of Processive Reverse Transcription as Well as Excessive Cytidine Deamination. J. Virol. 2013, 87, 1508–1517. [CrossRef]
- Newman, E.N.C.; Holmes, R.K.; Craig, H.M.; Klein, K.C.; Lingappa, J.R.; Malim, M.H.; Sheehy, A.M. Antiviral Function of APOBEC3G Can Be Dissociated from Cytidine Deaminase Activity. Curr. Biol. 2005, 15, 166–170. [CrossRef]
- Guo, F.; Cen, S.; Niu, M.; Saadatmand, J.; Kleiman, L. Inhibition of tRNA₃(Lys)-Primed Reverse Transcription by Human APOBEC3G during Human Immunodeficiency Virus Type 1 Replication. J. Virol. 2006, 80, 11710–11722. [CrossRef]
- Liu, Y.; Fu, Y.; Wang, Q.; Li, M.; Zhou, Z.; Dabbagh, D.; Fu, C.; Zhang, H.; Li, S.; Zhang, T.; et al. Proteomic Profiling of HIV-1 Infection of Human CD4+ T Cells Identifies PSGL-1 as an HIV Restriction Factor. Nat. Microbiol. 2019, 4, 813–825. [CrossRef]
- Liu, Y.; Song, Y.; Zhang, S.; Diao, M.; Huang, S.; Li, S.; Tan, X. PSGL-1 Inhibits HIV-1 Infection by Restricting Actin Dynamics and Sequestering HIV Envelope Proteins. Cell Discov. 2020, 6, 1–15. [CrossRef]
- Liu, L.; Oliveira, N.M.M.; Cheney, K.M.; Pade, C.; Dreja, H.; Bergin, A.-M.H.; Borgdorff, V.; Beach, D.H.; Bishop, C.L.; Dittmar, M.T.; et al. A Whole Genome Screen for HIV Restriction Factors. Retrovirology 2011, 8, 94. [CrossRef]
- Yan, J.; Shun, M.-C.; Hao, C.; Zhang, Y.; Qian, J.; Hrecka, K.; DeLucia, M.; Monnie, C.; Ahn, J.; Skowronski, J. HIV-1 Vpr Reprograms CLR4DCAF1 E3 Ubiquitin Ligase to Antagonize Exonuclease 1-Mediated Restriction of HIV-1 Infection. mBio 2018, 9, e01732-18. [CrossRef]
- Yan, J.; Shun, M.-C.; Zhang, Y.; Hao, C.; Skowronski, J. HIV-1 Vpr Counteracts HLTF-Mediated Restriction of HIV-1 Infection in T Cells. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 9568–9577. [CrossRef]
- Burdick, R.; Smith, J.L.; Chaipan, C.; Friew, Y.; Chen, J.; Venkatachari, N.J.; Delviks-Frankenberry, K.A.; Hu, W.-S.; Pathak, V.K. P Body-Associated Protein Mov10 Inhibits HIV-1 Replication at Multiple Stages. J. Virol. 2010, 84, 10241–10253. [CrossRef]
- Xie, L.; Chen, L.; Zhong, C.; Yu, T.; Ju, Z.; Wang, M.; Xiong, H.; Zeng, Y.; Wang, J.; Hu, H.; et al. MxB Impedes the NUP358-Mediated HIV-1 Pre-Integration Complex Nuclear Import and Viral Replication Cooperatively with CPSF6. Retrovirology 2020, 17, 16. [CrossRef]
- Marno, K.M.; Ogunkolade, B.W.; Pade, C.; Oliveira, N.M.M.; O’Sullivan, E.; McKnight, Á. Novel Restriction Factor RNA-Associated Early-Stage Anti-Viral Factor (REAF) Inhibits Human and Simian Immunodeficiency Viruses. Retrovirology 2014, 11, 3. [CrossRef]
- Jackson-Jones, K.A.; McKnight, Á.; Sloan, R.D. The Innate Immune Factor RPRD2/REAF and Its Role in the Lv2 Restriction of HIV. mBio 2023, 14, e02572-21. [CrossRef]
- Sonza, S.; Maerz, A.; Deacon, N.; Meanger, J.; Mills, J.; Crowe, S. Human Immunodeficiency Virus Type 1 Replication Is Blocked Prior to Reverse Transcription and Integration in Freshly Isolated Peripheral Blood Monocytes. J. Virol. 1996, 70, 3863–3869. [CrossRef]
- O’Brien, W.A.; Namazi, A.; Kalhor, H.; Mao, S.H.; Zack, J.A.; Chen, I.S. Kinetics of Human Immunodeficiency Virus Type 1 Reverse Transcription in Blood Mononuclear Phagocytes Are Slowed by Limitations of Nucleotide Precursors. J. Virol. 1994, 68, 1258–1263. [CrossRef]
- Collin, M.; Gordon, S. The Kinetics of Human Immunodeficiency Virus Reverse Transcription Are Slower in Primary Human Macrophages than in a Lymphoid Cell Line. Virology 1994, 200, 114–120. [CrossRef]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Ségéral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 Is the Dendritic- and Myeloid-Cell-Specific HIV-1 Restriction Factor Counteracted by Vpx. Nature 2011, 474, 654–657. [CrossRef]
- Ballana, E.; Esté, J.A. SAMHD1: At the Crossroads of Cell Proliferation, Immune Responses, and Virus Restriction. Trends Microbiol. 2015, 23, 680–692. [CrossRef]
- Berger, A.; Sommer, A.F.R.; Zwarg, J.; Hamdorf, M.; Welzel, K.; Esly, N.; Panitz, S.; Reuter, A.; Ramos, I.; Jatiani, A.; et al. SAMHD1-Deficient CD14+ Cells from Individuals with Aicardi-Goutières Syndrome Are Highly Susceptible to HIV-1 Infection. PLOS Pathog. 2011, 7, e1002425. [CrossRef]
- Amie, S.M.; Noble, E.; Kim, B. Intracellular Nucleotide Levels and the Control of Retroviral Infections. Virology 2013, 436, 247–254. [CrossRef]
- Franzolin, E.; Pontarin, G.; Rampazzo, C.; Miazzi, C.; Ferraro, P.; Palumbo, E.; Reichard, P.; Bianchi, V. The Deoxynucleotide Triphosphohydrolase SAMHD1 Is a Major Regulator of DNA Precursor Pools in Mammalian Cells. Proc. Natl. Acad. Sci. 2013, 110, 14272–14277. [CrossRef]
- Lahouassa, H.; Daddacha, W.; Hofmann, H.; Ayinde, D.; Logue, E.C.; Dragin, L.; Bloch, N.; Maudet, C.; Bertrand, M.; Gramberg, T.; et al. SAMHD1 Restricts the Replication of Human Immunodeficiency Virus Type 1 by Depleting the Intracellular Pool of Deoxynucleoside Triphosphates. Nat. Immunol. 2012, 13, 223–228. [CrossRef]
- Descours, B.; Cribier, A.; Chable-Bessia, C.; Ayinde, D.; Rice, G.; Crow, Y.; Yatim, A.; Schwartz, O.; Laguette, N.; Benkirane, M. SAMHD1 Restricts HIV-1 Reverse Transcription in Quiescent CD4(+) T-Cells. Retrovirology 2012, 9, 87. [CrossRef]
- Baldauf, H.-M.; Pan, X.; Erikson, E.; Schmidt, S.; Daddacha, W.; Burggraf, M.; Schenkova, K.; Ambiel, I.; Wabnitz, G.; Gramberg, T.; et al. SAMHD1 Restricts HIV-1 Infection in Resting CD4+ T Cells. Nat. Med. 2012, 18, 1682–1688. [CrossRef]
- Diamond, T.L.; Roshal, M.; Jamburuthugoda, V.K.; Reynolds, H.M.; Merriam, A.R.; Lee, K.Y.; Balakrishnan, M.; Bambara, R.A.; Planelles, V.; Dewhurst, S.; et al. Macrophage Tropism of HIV-1 Depends on Efficient Cellular dNTP Utilization by Reverse Transcriptase. J. Biol. Chem. 2004, 279, 51545–51553. [CrossRef]
- Hrecka, K.; Hao, C.; Gierszewska, M.; Swanson, S.K.; Kesik-Brodacka, M.; Srivastava, S.; Florens, L.; Washburn, M.P.; Skowronski, J. Vpx Relieves Inhibition of HIV-1 Infection of Macrophages Mediated by the SAMHD1 Protein. Nature 2011, 474, 658–661. [CrossRef]
- Hasanshahi, Z.; Dehghani, B.; Hashempour, A. Interaction Between Vpx and SAMHD1, Vital for SAMHD1 Inhibition. AIDS Res. Hum. Retroviruses 2023. [CrossRef]
- Plitnik, T.; Sharkey, M.E.; Mahboubi, B.; Kim, B.; Stevenson, M. Incomplete Suppression of HIV-1 by SAMHD1 Permits Efficient Macrophage Infection. Pathog. Immun. 2018, 3, 197–223. [CrossRef]
- White, T.E.; Brandariz-Nuñez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.A.; Kim, B.; Tuzova, M.; Diaz-Griffero, F. The Retroviral Restriction Ability of SAMHD1, but Not Its Deoxynucleotide Triphosphohydrolase Activity, Is Regulated by Phosphorylation. Cell Host Microbe 2013, 13, 441–451. [CrossRef]
- Beloglazova, N.; Flick, R.; Tchigvintsev, A.; Brown, G.; Popovic, A.; Nocek, B.; Yakunin, A.F. Nuclease Activity of the Human SAMHD1 Protein Implicated in the Aicardi-Goutieres Syndrome and HIV-1 Restriction. J. Biol. Chem. 2013, 288, 8101–8110. [CrossRef]
- Choi, J.; Ryoo, J.; Oh, C.; Hwang, S.; Ahn, K. SAMHD1 Specifically Restricts Retroviruses through Its RNase Activity. Retrovirology 2015, 12, 46. [CrossRef]
- Pauls, E.; Ruiz, A.; Riveira-Muñoz, E.; Permanyer, M.; Badia, R.; Clotet, B.; Keppler, O.T.; Ballana, E.; Este, J.A. P21 Regulates the HIV-1 Restriction Factor SAMHD1. Proc. Natl. Acad. Sci. 2014, 111, E1322–E1324. [CrossRef]
- Valle-Casuso, J.C.; Allouch, A.; David, A.; Lenzi, G.M.; Studdard, L.; Barré-Sinoussi, F.; Müller-Trutwin, M.; Kim, B.; Pancino, G.; Sáez-Cirión, A. P21 Restricts HIV-1 in Monocyte-Derived Dendritic Cells through the Reduction of Deoxynucleoside Triphosphate Biosynthesis and Regulation of SAMHD1 Antiviral Activity. J. Virol. 2017, 91, 10.1128/jvi.01324-17. [CrossRef]
- Chen, H.; Li, C.; Huang, J.; Cung, T.; Seiss, K.; Beamon, J.; Carrington, M.F.; Porter, L.C.; Burke, P.S.; Yang, Y.; et al. CD4+ T Cells from Elite Controllers Resist HIV-1 Infection by Selective Upregulation of P21 Available online: https://www.jci.org/articles/view/44539/pdf (accessed on 24 May 2024).
- Shi, B.; Sharifi, H.J.; DiGrigoli, S.; Kinnetz, M.; Mellon, K.; Hu, W.; de Noronha, C.M.C. Inhibition of HIV Early Replication by the P53 and Its Downstream Gene P21. Virol. J. 2018, 15, 53. [CrossRef]
- Bergamaschi, A.; David, A.; Le Rouzic, E.; Nisole, S.; Barré-Sinoussi, F.; Pancino, G. The CDK Inhibitor p21Cip1/WAF1 Is Induced by FcgammaR Activation and Restricts the Replication of Human Immunodeficiency Virus Type 1 and Related Primate Lentiviruses in Human Macrophages. J. Virol. 2009, 83, 12253–12265. [CrossRef]
- Allouch, A.; David, A.; Amie, S.M.; Lahouassa, H.; Chartier, L.; Margottin-Goguet, F.; Barré-Sinoussi, F.; Kim, B.; Sáez-Cirión, A.; Pancino, G. P21-Mediated RNR2 Repression Restricts HIV-1 Replication in Macrophages by Inhibiting dNTP Biosynthesis Pathway. Proc. Natl. Acad. Sci. U. S. A. 2013, 110, E3997-4006. [CrossRef]
- Kennedy, E.M.; Amie, S.M.; Bambara, R.A.; Kim, B. Frequent Incorporation of Ribonucleotides during HIV-1 Reverse Transcription and Their Attenuated Repair in Macrophages. J. Biol. Chem. 2012, 287, 14280–14288. [CrossRef]
- Craigie, R.; Bushman, F.D. HIV DNA Integration. Cold Spring Harb. Perspect. Med. 2012, 2, a006890. [CrossRef]
- Sloan, R.D.; Wainberg, M.A. The Role of Unintegrated DNA in HIV Infection. Retrovirology 2011, 8, 52. [CrossRef]
- Cereseto, A.; Manganaro, L.; Gutierrez, M.I.; Terreni, M.; Fittipaldi, A.; Lusic, M.; Marcello, A.; Giacca, M. Acetylation of HIV-1 Integrase by P300 Regulates Viral Integration. EMBO J. 2005, 24, 3070–3081. [CrossRef]
- Achuthan, V.; Perreira, J.M.; Sowd, G.A.; Puray-Chavez, M.; McDougall, W.M.; Paulucci-Holthauzen, A.; Wu, X.; Fadel, H.J.; Poeschla, E.M.; Multani, A.S.; et al. Capsid-CPSF6 Interaction Licenses Nuclear HIV-1 Trafficking to Sites of Viral DNA Integration. Cell Host Microbe 2018, 24, 392-404.e8. [CrossRef]
- Vranckx, L.S.; Demeulemeester, J.; Saleh, S.; Boll, A.; Vansant, G.; Schrijvers, R.; Weydert, C.; Battivelli, E.; Verdin, E.; Cereseto, A.; et al. LEDGIN-Mediated Inhibition of Integrase-LEDGF/P75 Interaction Reduces Reactivation of Residual Latent HIV. EBioMedicine 2016, 8, 248–264. [CrossRef]
- Cherepanov, P.; Maertens, G.; Proost, P.; Devreese, B.; Van Beeumen, J.; Engelborghs, Y.; De Clercq, E.; Debyser, Z. HIV-1 Integrase Forms Stable Tetramers and Associates with LEDGF/P75 Protein in Human Cells. J. Biol. Chem. 2003, 278, 372–381. [CrossRef]
- Emiliani, S.; Mousnier, A.; Busschots, K.; Maroun, M.; Van Maele, B.; Tempé, D.; Vandekerckhove, L.; Moisant, F.; Ben-Slama, L.; Witvrouw, M.; et al. Integrase Mutants Defective for Interaction with LEDGF/P75 Are Impaired in Chromosome Tethering and HIV-1 Replication. J. Biol. Chem. 2005, 280, 25517–25523. [CrossRef]
- Shun, M.-C.; Raghavendra, N.K.; Vandegraaff, N.; Daigle, J.E.; Hughes, S.; Kellam, P.; Cherepanov, P.; Engelman, A. LEDGF/P75 Functions Downstream from Preintegration Complex Formation to Effect Gene-Specific HIV-1 Integration. Genes Dev. 2007, 21, 1767–1778. [CrossRef]
- Llano, M.; Vanegas, M.; Hutchins, N.; Thompson, D.; Delgado, S.; Poeschla, E.M. Identification and Characterization of the Chromatin-Binding Domains of the HIV-1 Integrase Interactor LEDGF/P75. J. Mol. Biol. 2006, 360, 760–773. [CrossRef]
- De Rijck, J.; Vandekerckhove, L.; Gijsbers, R.; Hombrouck, A.; Hendrix, J.; Vercammen, J.; Engelborghs, Y.; Christ, F.; Debyser, Z. Overexpression of the Lens Epithelium-Derived Growth Factor/P75 Integrase Binding Domain Inhibits Human Immunodeficiency Virus Replication. J. Virol. 2006, 80, 11498–11509. [CrossRef]
- Llano, M.; Saenz, D.T.; Meehan, A.; Wongthida, P.; Peretz, M.; Walker, W.H.; Teo, W.; Poeschla, E.M. An Essential Role for LEDGF/P75 in HIV Integration. Science 2006, 314, 461–464. [CrossRef]
- Marshall, H.M.; Ronen, K.; Berry, C.; Llano, M.; Sutherland, H.; Saenz, D.; Bickmore, W.; Poeschla, E.; Bushman, F.D. Role of PSIP1/LEDGF/P75 in Lentiviral Infectivity and Integration Targeting. PLoS ONE 2007, 2, e1340. [CrossRef]
- Farnet, C.M.; Bushman, F.D. HIV-1 cDNA Integration: Requirement of HMG I(Y) Protein for Function of Preintegration Complexes In Vitro. Cell 1997, 88, 483–492. [CrossRef]
- Li, L.; Yoder, K.; Hansen, M.S.T.; Olvera, J.; Miller, M.D.; Bushman, F.D. Retroviral cDNA Integration: Stimulation by HMG I Family Proteins. J. Virol. 2000, 74, 10965–10974. [CrossRef]
- Lesbats, P.; Botbol, Y.; Chevereau, G.; Vaillant, C.; Calmels, C.; Arneodo, A.; Andreola, M.-L.; Lavigne, M.; Parissi, V. Functional Coupling between HIV-1 Integrase and the SWI/SNF Chromatin Remodeling Complex for Efficient in Vitro Integration into Stable Nucleosomes. PLoS Pathog. 2011, 7, e1001280. [CrossRef]
- Davis, A.J.; Chen, B.P.C.; Chen, D.J. DNA-PK: A Dynamic Enzyme in a Versatile DSB Repair Pathway. DNA Repair 2014, 17, 21–29. [CrossRef]
- Daniel, R.; Katz, R.A.; Skalka, A.M. A Role for DNA-PK in Retroviral DNA Integration. Science 1999, 284, 644–647. [CrossRef]
- Li, L.; Olvera, J.M.; Yoder, K.E.; Mitchell, R.S.; Butler, S.L.; Lieber, M.; Martin, S.L.; Bushman, F.D. Role of the Non-homologous DNA End Joining Pathway in the Early Steps of Retroviral Infection. EMBO J. 2001, 20, 3272–3281. [CrossRef]
- Jeanson, L.; Subra, F.; Vaganay, S.; Hervy, M.; Marangoni, E.; Bourhis, J.; Mouscadet, J.-F. Effect of Ku80 Depletion on the Preintegrative Steps of HIV-1 Replication in Human Cells. Virology 2002, 300, 100–108. [CrossRef]
- Lau, A.; Swinbank, K.M.; Ahmed, P.S.; Taylor, D.L.; Jackson, S.P.; Smith, G.C.M.; O’Connor, M.J. Suppression of HIV-1 Infection by a Small Molecule Inhibitor of the ATM Kinase. Nat. Cell Biol. 2005, 7, 493–500. [CrossRef]
- Anderson, E.M.; Maldarelli, F. The Role of Integration and Clonal Expansion in HIV Infection: Live Long and Prosper. Retrovirology 2018, 15, 71. [CrossRef]
- Maroun, M.; Delelis, O.; Coadou, G.; Bader, T.; Ségéral, E.; Mbemba, G.; Petit, C.; Sonigo, P.; Rain, J.-C.; Mouscadet, J.-F.; et al. Inhibition of Early Steps of HIV-1 Replication by SNF5/Ini1 *. J. Biol. Chem. 2006, 281, 22736–22743. [CrossRef]
- Lau, A.; Kanaar, R.; Jackson, S.P.; O’Connor, M.J. Suppression of Retroviral Infection by the RAD52 DNA Repair Protein. EMBO J. 2004, 23, 3421–3429. [CrossRef]
- Yoder, K.; Sarasin, A.; Kraemer, K.; McIlhatton, M.; Bushman, F.; Fishel, R. The DNA Repair Genes XPB and XPD Defend Cells from Retroviral Infection. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 4622–4627. [CrossRef]
- Allouch, A.; Di Primio, C.; Alpi, E.; Lusic, M.; Arosio, D.; Giacca, M.; Cereseto, A. The TRIM Family Protein KAP1 Inhibits HIV-1 Integration. Cell Host Microbe 2011, 9, 484–495. [CrossRef]
- Marcello, A.; Zoppé, M.; Giacca, M. Multiple Modes of Transcriptional Regulation by the HIV-1 Tat Transactivator. IUBMB Life 2001, 51, 175–181. [CrossRef]
- Roebuck, K.A.; Saifuddin, M. Regulation of HIV-1 Transcription. Gene Expr. 2018, 8, 67–84.
- Van Lint, C.; Bouchat, S.; Marcello, A. HIV-1 Transcription and Latency: An Update. Retrovirology 2013, 10, 67. [CrossRef]
- Pendergrast, P.S.; Morrison, D.; Tansey, W.P.; Hernandez, N. Mutations in the Carboxy-Terminal Domain of TBP Affect the Synthesis of Human Immunodeficiency Virus Type 1 Full-Length and Short Transcripts Similarly. J. Virol. 1996, 70, 5025–5034. [CrossRef]
- Majello, B.; Napolitano, G.; De Luca, P.; Lania, L. Recruitment of Human TBP Selectively Activates RNA Polymerase II TATA-Dependent Promoters*. J. Biol. Chem. 1998, 273, 16509–16516. [CrossRef]
- Rice, A.P. Roles of CDKs in RNA Polymerase II Transcription of the HIV-1 Genome. Transcription 2019, 10, 111–117. [CrossRef]
- Chun, R.F.; Jeang, K.-T. Requirements for RNA Polymerase II Carboxyl-Terminal Domain for Activated Transcription of Human Retroviruses Human T-Cell Lymphotropic Virus I and HIV-1 *. J. Biol. Chem. 1996, 271, 27888–27894. [CrossRef]
- Mancebo, H.S.Y.; Lee, G.; Flygare, J.; Tomassini, J.; Luu, P.; Zhu, Y.; Peng, J.; Blau, C.; Hazuda, D.; Price, D.; et al. P-TEFb Kinase Is Required for HIV Tat Transcriptional Activation in Vivo and in Vitro. Genes Dev. 1997, 11, 2633–2644. [CrossRef]
- Zhu, Y.; Pe’ery, T.; Peng, J.; Ramanathan, Y.; Marshall, N.; Marshall, T.; Amendt, B.; Mathews, M.B.; Price, D.H. Transcription Elongation Factor P-TEFb Is Required for HIV-1 Tat Transactivation in Vitro. Genes Dev. 1997, 11, 2622–2632. [CrossRef]
- Gold, M.O.; Yang, X.; Herrmann, C.H.; Rice, A.P. PITALRE, the Catalytic Subunit of TAK, Is Required for Human Immunodeficiency Virus Tat Transactivation In Vivo. J. Virol. 1998, 72, 4448–4453. [CrossRef]
- Bruce, J.W.; Reddington, R.; Mathieu, E.; Bracken, M.; Young, J.A.T.; Ahlquist, P. ZASC1 Stimulates HIV-1 Transcription Elongation by Recruiting P-TEFb and TAT to the LTR Promoter. PLOS Pathog. 2013, 9, e1003712. [CrossRef]
- Herrmann, C.H.; Carroll, R.G.; Wei, P.; Jones, K.A.; Rice, A.P. Tat-Associated Kinase, TAK, Activity Is Regulated by Distinct Mechanisms in Peripheral Blood Lymphocytes and Promonocytic Cell Lines. J. Virol. 1998, 72, 9881–9888. [CrossRef]
- García-Martínez, L.F.; Mavankal, G.; Neveu, J.M.; Lane, W.S.; Ivanov, D.; Gaynor, R.B. Purification of a Tat-associated Kinase Reveals a TFIIH Complex That Modulates HIV-1 Transcription. EMBO J. 1997, 16, 2836–2850. [CrossRef]
- Schilbach, S.; Hantsche, M.; Tegunov, D.; Dienemann, C.; Wigge, C.; Urlaub, H.; Cramer, P. Structures of Transcription Pre-Initiation Complex with TFIIH and Mediator. Nature 2017, 551, 204–209. [CrossRef]
- Chen, D.; Zhou, Q. Tat Activates Human Immunodeficiency Virus Type 1 Transcriptional Elongation Independent of TFIIH Kinase. Mol. Cell. Biol. 1999, 19, 2863–2871. [CrossRef]
- Zhou, M.; Halanski, M.A.; Radonovich, M.F.; Kashanchi, F.; Peng, J.; Price, D.H.; Brady, J.N. Tat Modifies the Activity of CDK9 to Phosphorylate Serine 5 of the RNA Polymerase II Carboxyl-Terminal Domain during Human Immunodeficiency Virus Type 1 Transcription. Mol. Cell. Biol. 2000, 20, 5077–5086. [CrossRef]
- Zhou, M.; Nekhai, S.; Bharucha, D.C.; Kumar, A.; Ge, H.; Price, D.H.; Egly, J.-M.; Brady, J.N. TFIIH Inhibits CDK9 Phosphorylation during Human Immunodeficiency Virus Type 1 Transcription*. J. Biol. Chem. 2001, 276, 44633–44640. [CrossRef]
- Holloway, A.F.; Occhiodoro, F.; Mittler, G.; Meisterernst, M.; Shannon, M.F. Functional Interaction between the HIV Transactivator Tat and the Transcriptional Coactivator PC4 in T Cells. J. Biol. Chem. 2000, 275, 21668–21677. [CrossRef]
- Kim, H.-Y.; Choi, B.-S.; Kim, S.S.; Roh, T.-Y.; Park, J.; Yoon, C.-H. NUCKS1, a Novel Tat Coactivator, Plays a Crucial Role in HIV-1 Replication by Increasing Tat-Mediated Viral Transcription on the HIV-1 LTR Promoter. Retrovirology 2014, 11, 67. [CrossRef]
- Dorin, D.; Bonnet, M.C.; Bannwarth, S.; Gatignol, A.; Meurs, E.F.; Vaquero, C. The TAR RNA-Binding Protein, TRBP, Stimulates the Expression of TAR-Containing RNAs in Vitro and in Vivo Independently of Its Ability to Inhibit the dsRNA-Dependent Kinase PKR. J. Biol. Chem. 2003, 278, 4440–4448. [CrossRef]
- Christensen, H.S.; Daher, A.; Soye, K.J.; Frankel, L.B.; Alexander, M.R.; Lainé, S.; Bannwarth, S.; Ong, C.L.; Chung, S.W.L.; Campbell, S.M.; et al. Small Interfering RNAs against the TAR RNA Binding Protein, TRBP, a Dicer Cofactor, Inhibit Human Immunodeficiency Virus Type 1 Long Terminal Repeat Expression and Viral Production. J. Virol. 2007, 81, 5121–5131. [CrossRef]
- Cho, E.-J.; Takagi, T.; Moore, C.R.; Buratowski, S. mRNA Capping Enzyme Is Recruited to the Transcription Complex by Phosphorylation of the RNA Polymerase II Carboxy-Terminal Domain. Genes Dev. 1997, 11, 3319–3326.
- Chiu, Y.-L.; Ho, C.K.; Saha, N.; Schwer, B.; Shuman, S.; Rana, T.M. Tat Stimulates Cotranscriptional Capping of HIV mRNA. Mol. Cell 2002, 10, 585–597. [CrossRef]
- Zhou, M.; Deng, L.; Kashanchi, F.; Brady, J.N.; Shatkin, A.J.; Kumar, A. The Tat/TAR-Dependent Phosphorylation of RNA Polymerase II C-Terminal Domain Stimulates Cotranscriptional Capping of HIV-1 mRNA. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 12666–12671. [CrossRef]
- Chiu, Y.L.; Coronel, E.; Ho, C.K.; Shuman, S.; Rana, T.M. HIV-1 Tat Protein Interacts with Mammalian Capping Enzyme and Stimulates Capping of TAR RNA. J. Biol. Chem. 2001, 276, 12959–12966. [CrossRef]
- Wilusz, J. Putting an ‘End’ to HIV mRNAs: Capping and Polyadenylation as Potential Therapeutic Targets. AIDS Res. Ther. 2013, 10, 31. [CrossRef]
- Sertznig, H.; Hillebrand, F.; Erkelenz, S.; Schaal, H.; Widera, M. Behind the Scenes of HIV-1 Replication: Alternative Splicing as the Dependency Factor on the Quiet. Virology 2018, 516, 176–188. [CrossRef]
- Caputi, M.; Mayeda, A.; Krainer, A.R.; Zahler, A.M. hnRNP A/B Proteins Are Required for Inhibition of HIV-1 Pre-mRNA Splicing. EMBO J. 1999, 18, 4060–4067. [CrossRef]
- Bilodeau, P.S.; Domsic, J.K.; Mayeda, A.; Krainer, A.R.; Stoltzfus, C.M. RNA Splicing at Human Immunodeficiency Virus Type 1 3’ Splice Site A2 Is Regulated by Binding of hnRNP A/B Proteins to an Exonic Splicing Silencer Element. J. Virol. 2001, 75, 8487–8497. [CrossRef]
- Namer, L.S.; Harwig, A.; Heynen, S.P.; Das, A.T.; Berkhout, B.; Kaempfer, R. HIV Co-Opts a Cellular Antiviral Mechanism, Activation of Stress Kinase PKR by Its RNA, to Enable Splicing of Rev/Tat mRNA. Cell Biosci. 2023, 13, 28. [CrossRef]
- Jones, K.A.; Kadonaga, J.T.; Luciw, P.A.; Tjian, R. Activation of the AIDS Retrovirus Promoter by the Cellular Transcription Factor, Sp1. Science 1986, 232, 755–759. [CrossRef]
- Suñé, C.; García-Blanco, M.A. Sp1 Transcription Factor Is Required for in Vitro Basal and Tat-Activated Transcription from the Human Immunodeficiency Virus Type 1 Long Terminal Repeat. J. Virol. 1995, 69, 6572–6576. [CrossRef]
- Majello, B.; De Luca, P.; Hagen, G.; Suske, G.; Lania, L. Different Members of the Sp1 Multigene Family Exert Opposite Transcritional Regulation of the Long Terminal Repeat of HIV-1. Nucleic Acids Res. 1994, 22, 4914–4921. [CrossRef]
- IMATAKA, H.; MIZUNO, A.; FUJII-KURIYAMA, Y.; HAYAMI, M. Activation of the Human Immunodeficiency Virus Type 1 Long Terminal Repeat by BTEB, a GC Box-Binding Transcription Factor Available online: https://www.liebertpub.com/doi/10.1089/aid.1993.9.825 (accessed on 29 May 2024).
- Nabel, G.; Baltimore, D. An Inducible Transcription Factor Activates Expression of Human Immunodeficiency Virus in T Cells. Nature 1987, 326, 711–713. [CrossRef]
- Griffin, G.E.; Leung, K.; Folks, T.M.; Kunkel, S.; Nabel, G.J. Activation of HIV Gene Expression during Monocyte Differentiation by Induction of NF-kB. Nature 1989, 339, 70–73. [CrossRef]
- Conant, K.; Ma, M.; Nath, A.; Major, E.O. Extracellular Human Immunodeficiency Virus Type 1 Tat Protein Is Associated with an Increase in Both NF-Kappa B Binding and Protein Kinase C Activity in Primary Human Astrocytes. J. Virol. 1996, 70, 1384–1389. [CrossRef]
- Demarchi, F.; d’Adda di Fagagna, F.; Falaschi, A.; Giacca, M. Activation of Transcription Factor NF-kappaB by the Tat Protein of Human Immunodeficiency Virus Type 1. J. Virol. 1996, 70, 4427–4437. [CrossRef]
- Biswas, D.K.; Salas, T.R.; Wang, F.; Ahlers, C.M.; Dezube, B.J.; Pardee, A.B. A Tat-Induced Auto-up-Regulatory Loop for Superactivation of the Human Immunodeficiency Virus Type 1 Promoter. J. Virol. 1995, 69, 7437–7444. [CrossRef]
- Kretzschmar, M.; Meisterernst, M.; Scheidereit, C.; Li, G.; Roeder, R.G. Transcriptional Regulation of the HIV-1 Promoter by NF-Kappa B in Vitro. Genes Dev. 1992, 6, 761–774. [CrossRef]
- Chen, B.K.; Feinberg, M.B.; Baltimore, D. The kappaB Sites in the Human Immunodeficiency Virus Type 1 Long Terminal Repeat Enhance Virus Replication yet Are Not Absolutely Required for Viral Growth. J. Virol. 1997, 71, 5495–5504. [CrossRef]
- Liu, J.; Perkins, N.D.; Schmid, R.M.; Nabel, G.J. Specific NF-Kappa B Subunits Act in Concert with Tat to Stimulate Human Immunodeficiency Virus Type 1 Transcription. J. Virol. 1992, 66, 3883–3887. [CrossRef]
- Schmitz, M.L.; Stelzer, G.; Altmann, H.; Meisterernst, M.; Baeuerle, P.A. Interaction of the COOH-Terminal Transactivation Domain of P65 NF-Kappa B with TATA-Binding Protein, Transcription Factor IIB, and Coactivators. J. Biol. Chem. 1995, 270, 7219–7226. [CrossRef]
- Perkins, N.D.; Edwards, N.L.; Duckett, C.S.; Agranoff, A.B.; Schmid, R.M.; Nabel, G.J. A Cooperative Interaction between NF-Kappa B and Sp1 Is Required for HIV-1 Enhancer Activation. EMBO J. 1993, 12, 3551–3558.
- Duh, E.J.; Maury, W.J.; Folks, T.M.; Fauci, A.S.; Rabson, A.B. Tumor Necrosis Factor Alpha Activates Human Immunodeficiency Virus Type 1 through Induction of Nuclear Factor Binding to the NF-Kappa B Sites in the Long Terminal Repeat. Proc. Natl. Acad. Sci. 1989, 86, 5974–5978. [CrossRef]
- Granowitz, E.V.; Saget, B.M.; Wang, M.Z.; Dinarello, C.A.; Skolnik, P.R. Interleukin 1 Induces HIV-1 Expression in Chronically Infected U1 Cells: Blockade by Interleukin 1 Receptor Antagonist and Tumor Necrosis Factor Binding Protein Type 1. Mol. Med. 1995, 1, 667–677.
- Folks, T.M.; Justement, J.; Kinter, A.; Dinarello, C.A.; Fauci, A.S. Cytokine-Induced Expression of HIV-1 in a Chronically Infected Promonocyte Cell Line. Science 1987, 238, 800–802. [CrossRef]
- Finnegan, A.; Roebuck, K.A.; Nakai, B.E.; Gu, D.S.; Rabbi, M.F.; Song, S.; Landay, A.L. IL-10 Cooperates with TNF-Alpha to Activate HIV-1 from Latently and Acutely Infected Cells of Monocyte/Macrophage Lineage. J. Immunol. Baltim. Md 1950 1996, 156, 841–851.
- Li, J.M.; Shen, X.; Hu, P.P.; Wang, X.F. Transforming Growth Factor Beta Stimulates the Human Immunodeficiency Virus 1 Enhancer and Requires NF-kappaB Activity. Mol. Cell. Biol. 1998, 18, 110–121. [CrossRef]
- Dinter, H.; Chiu, R.; Imagawa, M.; Karin, M.; Jones, K.A. In Vitro Activation of the HIV-1 Enhancer in Extracts from Cells Treated with a Phorbol Ester Tumor Promoter. EMBO J. 1987, 6, 4067–4071. [CrossRef]
- Bassuk, A.G.; Anandappa, R.T.; Leiden, J.M. Physical Interactions between Ets and NF-kappaB/NFAT Proteins Play an Important Role in Their Cooperative Activation of the Human Immunodeficiency Virus Enhancer in T Cells. J. Virol. 1997, 71, 3563–3573. [CrossRef]
- Kundu, M.; Srinivasan, A.; Pomerantz, R.J.; Khalili, K. Evidence That a Cell Cycle Regulator, E2F1, down-Regulates Transcriptional Activity of the Human Immunodeficiency Virus Type 1 Promoter. J. Virol. 1995, 69, 6940–6946. [CrossRef]
- Verdin, E.; Becker, N.; Bex, F.; Droogmans, L.; Burny, A. Identification and Characterization of an Enhancer in the Coding Region of the Genome of Human Immunodeficiency Virus Type 1. Proc. Natl. Acad. Sci. 1990, 87, 4874–4878. [CrossRef]
- Van Lint, C.; Burny, A.; Verdin, E. The Intragenic Enhancer of Human Immunodeficiency Virus Type 1 Contains Functional AP-1 Binding Sites. J. Virol. 1991, 65, 7066–7072. [CrossRef]
- Colin, L.; Vandenhoudt, N.; Walque, S. de; Driessche, B.V.; Bergamaschi, A.; Martinelli, V.; Cherrier, T.; Vanhulle, C.; Guiguen, A.; David, A.; et al. The AP-1 Binding Sites Located in the Pol Gene Intragenic Regulatory Region of HIV-1 Are Important for Viral Replication. PLOS ONE 2011, 6, e19084. [CrossRef]
- Lu, Y.C.; Touzjian, N.; Stenzel, M.; Dorfman, T.; Sodroski, J.G.; Haseltine, W.A. Identification of Cis-Acting Repressive Sequences within the Negative Regulatory Element of Human Immunodeficiency Virus Type 1. J. Virol. 1990, 64, 5226–5229. [CrossRef]
- Van Lint, C.; Amella, C.A.; Emiliani, S.; John, M.; Jie, T.; Verdin, E. Transcription Factor Binding Sites Downstream of the Human Immunodeficiency Virus Type 1 Transcription Start Site Are Important for Virus Infectivity. J. Virol. 1997, 71, 6113–6127. [CrossRef]
- Yang, Z.; Engel, J.D. Human T Cell Transcription Factor GATA-3 Stimulates HIV-1 Expression. Nucleic Acids Res. 1993, 21, 2831–2836. [CrossRef]
- Sheridan, P.L.; Sheline, C.T.; Cannon, K.; Voz, M.L.; Pazin, M.J.; Kadonaga, J.T.; Jones, K.A. Activation of the HIV-1 Enhancer by the LEF-1 HMG Protein on Nucleosome-Assembled DNA in Vitro. Genes Dev. 1995, 9, 2090–2104. [CrossRef]
- Henderson, A.J.; Zou, X.; Calame, K.L. C/EBP Proteins Activate Transcription from the Human Immunodeficiency Virus Type 1 Long Terminal Repeat in Macrophages/Monocytes. J. Virol. 1995, 69, 5337–5344. [CrossRef]
- Henderson, A.J.; Connor, R.I.; Calame, K.L. C/EBP Activators Are Required for HIV-1 Replication and Proviral Induction in Monocytic Cell Lines. Immunity 1996, 5, 91–101. [CrossRef]
- Henderson, A.J.; Calame, K.L. CCAAT/Enhancer Binding Protein (C/EBP) Sites Are Required for HIV-1 Replication in Primary Macrophages but Not CD4+ T Cells. Proc. Natl. Acad. Sci. 1997, 94, 8714–8719. [CrossRef]
- Cherrier, T.; Le Douce, V.; Eilebrecht, S.; Riclet, R.; Marban, C.; Dequiedt, F.; Goumon, Y.; Paillart, J.-C.; Mericskay, M.; Parlakian, A.; et al. CTIP2 Is a Negative Regulator of P-TEFb. Proc. Natl. Acad. Sci. 2013, 110, 12655–12660. [CrossRef]
- Eilebrecht, S.; Le Douce, V.; Riclet, R.; Targat, B.; Hallay, H.; Van Driessche, B.; Schwartz, C.; Robette, G.; Van Lint, C.; Rohr, O.; et al. HMGA1 Recruits CTIP2-Repressed P-TEFb to the HIV-1 and Cellular Target Promoters. Nucleic Acids Res. 2014, 42, 4962–4971. [CrossRef]
- Duan, L.; Ozaki, I.; Oakes, J.W.; Taylor, J.P.; Khalili, K.; Pomerantz, R.J. The Tumor Suppressor Protein P53 Strongly Alters Human Immunodeficiency Virus Type 1 Replication. J. Virol. 1994, 68, 4302–4313.
- Gérard, A.; Ségéral, E.; Naughtin, M.; Abdouni, A.; Charmeteau, B.; Cheynier, R.; Rain, J.-C.; Emiliani, S. The Integrase Cofactor LEDGF/P75 Associates with Iws1 and Spt6 for Postintegration Silencing of HIV-1 Gene Expression in Latently Infected Cells. Cell Host Microbe 2015, 17, 107–117. [CrossRef]
- LIU, Y.-Z.; LATCHMAN, D.S. The Octamer-Binding Proteins Oct-1 and Oct-2 Repress the HIV Long Terminal Repeat Promoter and Its Transactivation by Tat. Biochem. J. 1997, 322, 155–158. [CrossRef]
- Fenard, D.; Houzet, L.; Bernard, E.; Tupin, A.; Brun, S.; Mougel, M.; Devaux, C.; Chazal, N.; Briant, L. Uracil DNA Glycosylase 2 Negatively Regulates HIV-1 LTR Transcription. Nucleic Acids Res. 2009, 37, 6008–6018. [CrossRef]
- Margolis, D.M.; Somasundaran, M.; Green, M.R. Human Transcription Factor YY1 Represses Human Immunodeficiency Virus Type 1 Transcription and Virion Production. J. Virol. 1994, 68, 905–910. [CrossRef]
- Jones, K.A.; Luciw, P.A.; Duchange, N. Structural Arrangements of Transcription Control Domains within the 5’-Untranslated Leader Regions of the HIV-1 and HIV-2 Promoters. Genes Dev. 1988, 2, 1101–1114. [CrossRef]
- Kato, H.; Horikoshi, M.; Roeder, R.G. Repression of HIV-1 Transcription by a Cellular Protein. Science 1991, 251, 1476–1479. [CrossRef]
- Parada, C.A.; Yoon, J.-B.; Roeder, R.G. A Novel LBP-1-Mediated Restriction of HIV-1 Transcription at the Level of Elongation in Vitro(∗). J. Biol. Chem. 1995, 270, 2274–2283. [CrossRef]
- Romerio, F.; Gabriel, M.N.; Margolis, D.M. Repression of Human Immunodeficiency Virus Type 1 through the Novel Cooperation of Human Factors YY1 and LSF. J. Virol. 1997, 71, 9375–9382. [CrossRef]
- Hotter, D.; Bosso, M.; Jønsson, K.L.; Krapp, C.; Stürzel, C.M.; Das, A.; Littwitz-Salomon, E.; Berkhout, B.; Russ, A.; Wittmann, S.; et al. IFI16 Targets the Transcription Factor Sp1 to Suppress HIV-1 Transcription and Latency Reactivation. Cell Host Microbe 2019, 25, 858-872.e13. [CrossRef]
- Kajaste-Rudnitski, A.; Marelli, S.S.; Pultrone, C.; Pertel, T.; Uchil, P.D.; Mechti, N.; Mothes, W.; Poli, G.; Luban, J.; Vicenzi, E. TRIM22 Inhibits HIV-1 Transcription Independently of Its E3 Ubiquitin Ligase Activity, Tat, and NF-kappaB-Responsive Long Terminal Repeat Elements. J. Virol. 2011, 85, 5183–5196. [CrossRef]
- Turrini, F.; Marelli, S.; Kajaste-Rudnitski, A.; Lusic, M.; Van Lint, C.; Das, A.T.; Harwig, A.; Berkhout, B.; Vicenzi, E. HIV-1 Transcriptional Silencing Caused by TRIM22 Inhibition of Sp1 Binding to the Viral Promoter. Retrovirology 2015, 12, 104. [CrossRef]
- Turrini, F.; Saliu, F.; Forlani, G.; Das, A.T.; Van Lint, C.; Accolla, R.S.; Berkhout, B.; Poli, G.; Vicenzi, E. Interferon-Inducible TRIM22 Contributes to Maintenance of HIV-1 Proviral Latency in T Cell Lines. Virus Res. 2019, 269, 197631. [CrossRef]
- Shi, Y.; Simpson, S.; Ahmed, S.K.; Chen, Y.; Tavakoli-Tameh, A.; Janaka, S.K.; Evans, D.T.; Serra-Moreno, R. The Antiviral Factor SERINC5 Impairs the Expression of Non-Self-DNA. Viruses 2023, 15, 1961. [CrossRef]
- Dupont, L.; Bloor, S.; Williamson, J.C.; Cuesta, S.M.; Shah, R.; Teixeira-Silva, A.; Naamati, A.; Greenwood, E.J.D.; Sarafianos, S.G.; Matheson, N.J.; et al. The SMC5/6 Complex Compacts and Silences Unintegrated HIV-1 DNA and Is Antagonized by Vpr. Cell Host Microbe 2021, 29, 792-805.e6. [CrossRef]
- Gallastegui, E.; Millán-Zambrano, G.; Terme, J.-M.; Chávez, S.; Jordan, A. Chromatin Reassembly Factors Are Involved in Transcriptional Interference Promoting HIV Latency. J. Virol. 2011, 85, 3187–3202. [CrossRef]
- Shirakawa, K.; Chavez, L.; Hakre, S.; Calvanese, V.; Verdin, E. Reactivation of Latent HIV by Histone Deacetylase Inhibitors. Trends Microbiol. 2013, 21, 277–285. [CrossRef]
- Le Douce, V.; Cherrier, T.; Riclet, R.; Rohr, O.; Schwartz, C. The Many Lives of CTIP2: From AIDS to Cancer and Cardiac Hypertrophy. J. Cell. Physiol. 2014, 229, 533–537. [CrossRef]
- Bruce, J.W.; Bracken, M.; Evans, E.; Sherer, N.; Ahlquist, P. ZBTB2 Represses HIV-1 Transcription and Is Regulated by HIV-1 Vpr and Cellular DNA Damage Responses. PLOS Pathog. 2021, 17, e1009364. [CrossRef]
- Schröder, H.C.; Wenger, R.; Kuchino, Y.; Müller, W.E. Modulation of Nuclear Matrix-Associated 2’,5’-Oligoadenylate Metabolism and Ribonuclease L Activity in H9 Cells by Human Immunodeficiency Virus. J. Biol. Chem. 1989, 264, 5669–5673.
- Brennan-Laun, S.E.; Ezelle, H.J.; Li, X.-L.; Hassel, B.A. RNase-L Control of Cellular mRNAs: Roles in Biologic Functions and Mechanisms of Substrate Targeting. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2014, 34, 275–288. [CrossRef]
- Martinand, C.; Montavon, C.; Salehzada, T.; Silhol, M.; Lebleu, B.; Bisbal, C. RNase L Inhibitor Is Induced during Human Immunodeficiency Virus Type 1 Infection and Down Regulates the 2-5A/RNase L Pathway in Human T Cells. J. Virol. 1999, 73, 290–296. [CrossRef]
- Nchioua, R.; Bosso, M.; Kmiec, D.; Kirchhoff, F. Cellular Factors Targeting HIV-1 Transcription and Viral RNA Transcripts. Viruses 2020, 12, 495. [CrossRef]
- Zhu, Y.; Chen, G.; Lv, F.; Wang, X.; Ji, X.; Xu, Y.; Sun, J.; Wu, L.; Zheng, Y.-T.; Gao, G. Zinc-Finger Antiviral Protein Inhibits HIV-1 Infection by Selectively Targeting Multiply Spliced Viral mRNAs for Degradation. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 15834–15839. [CrossRef]
- Chen, G.; Guo, X.; Lv, F.; Xu, Y.; Gao, G. P72 DEAD Box RNA Helicase Is Required for Optimal Function of the Zinc-Finger Antiviral Protein. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 4352–4357. [CrossRef]
- Schwerk, J.; Soveg, F.W.; Ryan, A.P.; Thomas, K.R.; Hatfield, L.D.; Ozarkar, S.; Forero, A.; Kell, A.M.; Roby, J.A.; So, L.; et al. RNA-Binding Protein Isoforms ZAP-S and ZAP-L Have Distinct Antiviral and Immune Resolution Functions. Nat. Immunol. 2019, 20, 1610–1620. [CrossRef]
- Zheng, X.; Wang, X.; Tu, F.; Wang, Q.; Fan, Z.; Gao, G. TRIM25 Is Required for the Antiviral Activity of Zinc Finger Antiviral Protein. J. Virol. 2017, 91, e00088-17. [CrossRef]
- Ficarelli, M.; Wilson, H.; Pedro Galão, R.; Mazzon, M.; Antzin-Anduetza, I.; Marsh, M.; Neil, S.J.; Swanson, C.M. KHNYN Is Essential for the Zinc Finger Antiviral Protein (ZAP) to Restrict HIV-1 Containing Clustered CpG Dinucleotides. eLife 2019, 8, e46767. [CrossRef]
- Ficarelli, M.; Antzin-Anduetza, I.; Hugh-White, R.; Firth, A.E.; Sertkaya, H.; Wilson, H.; Neil, S.J.D.; Schulz, R.; Swanson, C.M. CpG Dinucleotides Inhibit HIV-1 Replication through Zinc Finger Antiviral Protein (ZAP)-Dependent and -Independent Mechanisms. J. Virol. 2020, 94, e01337-19. [CrossRef]
- Kmiec, D.; Nchioua, R.; Sherrill-Mix, S.; Stürzel, C.M.; Heusinger, E.; Braun, E.; Gondim, M.V.P.; Hotter, D.; Sparrer, K.M.J.; Hahn, B.H.; et al. CpG Frequency in the 5’ Third of the Env Gene Determines Sensitivity of Primary HIV-1 Strains to the Zinc-Finger Antiviral Protein. mBio 2020, 11, e02903-19. [CrossRef]
- Takata, M.A.; Gonçalves-Carneiro, D.; Zang, T.M.; Soll, S.J.; York, A.; Blanco-Melo, D.; Bieniasz, P.D. CG Dinucleotide Suppression Enables Antiviral Defence Targeting Non-Self RNA. Nature 2017, 550, 124–127. [CrossRef]
- Yamasoba, D.; Sato, K.; Ichinose, T.; Imamura, T.; Koepke, L.; Joas, S.; Reith, E.; Hotter, D.; Misawa, N.; Akaki, K.; et al. N4BP1 Restricts HIV-1 and Its Inactivation by MALT1 Promotes Viral Reactivation. Nat. Microbiol. 2019, 4, 1532–1544. [CrossRef]
- Liu, S.; Qiu, C.; Miao, R.; Zhou, J.; Lee, A.; Liu, B.; Lester, S.N.; Fu, W.; Zhu, L.; Zhang, L.; et al. MCPIP1 Restricts HIV Infection and Is Rapidly Degraded in Activated CD4+ T Cells. Proc. Natl. Acad. Sci. 2013, 110, 19083–19088. [CrossRef]
- Triboulet, R.; Mari, B.; Lin, Y.-L.; Chable-Bessia, C.; Bennasser, Y.; Lebrigand, K.; Cardinaud, B.; Maurin, T.; Barbry, P.; Baillat, V.; et al. Suppression of MicroRNA-Silencing Pathway by HIV-1 During Virus Replication. Science 2007, 315, 1579–1582. [CrossRef]
- Nathans, R.; Chu, C.; Serquina, A.K.; Lu, C.-C.; Cao, H.; Rana, T.M. Cellular MicroRNA and P Bodies Modulate Host-HIV-1 Interactions. Mol. Cell 2009, 34, 696–709. [CrossRef]
- Wang, X.; Ye, L.; Hou, W.; Zhou, Y.; Wang, Y.-J.; Metzger, D.S.; Ho, W.-Z. Cellular microRNA Expression Correlates with Susceptibility of Monocytes/Macrophages to HIV-1 Infection. Blood 2009, 113, 671–674. [CrossRef]
- Toro-Ascuy, D.; Rojas-Araya, B.; Valiente-Echeverría, F.; Soto-Rifo, R. Interactions between the HIV-1 Unspliced mRNA and Host mRNA Decay Machineries. Viruses 2016, 8, 320. [CrossRef]
- Coyle, J.H.; Bor, Y.-C.; Rekosh, D.; Hammarskjold, M.-L. The Tpr Protein Regulates Export of mRNAs with Retained Introns That Traffic through the Nxf1 Pathway. RNA 2011, 17, 1344–1356. [CrossRef]
- Arnold, M.; Nath, A.; Hauber, J.; Kehlenbach, R.H. Multiple Importins Function as Nuclear Transport Receptors for the Rev Protein of Human Immunodeficiency Virus Type 1. J. Biol. Chem. 2006, 281, 20883–20890. [CrossRef]
- Izaurralde, E.; Stepinski, J.; Darzynkiewicz, E.; Mattaj, I.W. A Cap Binding Protein That May Mediate Nuclear Export of RNA Polymerase II-Transcribed RNAs. J. Cell Biol. 1992, 118, 1287–1295. [CrossRef]
- Izaurralde, E.; Lewis, J.; Gamberi, C.; Jarmolowski, A.; McGuigan, C.; Mattaj, I.W. A Cap-Binding Protein Complex Mediating U snRNA Export. Nature 1995, 376, 709–712. [CrossRef]
- Nawroth, I.; Mueller, F.; Basyuk, E.; Beerens, N.; Rahbek, U.L.; Darzacq, X.; Bertrand, E.; Kjems, J.; Schmidt, U. Stable Assembly of HIV-1 Export Complexes Occurs Cotranscriptionally. RNA 2014, 20, 1–8. [CrossRef]
- Sharma, A.; Yilmaz, A.; Marsh, K.; Cochrane, A.; Boris-Lawrie, K. Thriving under Stress: Selective Translation of HIV-1 Structural Protein mRNA during Vpr-Mediated Impairment of eIF4E Translation Activity. PLOS Pathog. 2012, 8, e1002612. [CrossRef]
- Fornerod, M.; Ohno, M.; Yoshida, M.; Mattaj, I.W. CRM1 Is an Export Receptor for Leucine-Rich Nuclear Export Signals. Cell 1997, 90, 1051–1060. [CrossRef]
- Malim, M.H.; Hauber, J.; Le, S.-Y.; Maizel, J.V.; Cullen, B.R. The HIV-1 Rev Trans-Activator Acts through a Structured Target Sequence to Activate Nuclear Export of Unspliced Viral mRNA. Nature 1989, 338, 254–257. [CrossRef]
- Fischer, U.; Meyer, S.; Teufel, M.; Heckel, C.; Lührmann, R.; Rautmann, G. Evidence That HIV-1 Rev Directly Promotes the Nuclear Export of Unspliced RNA. EMBO J. 1994, 13, 4105–4112. [CrossRef]
- Fischer, U.; Huber, J.; Boelens, W.C.; Mattaj, I.W.; Lührmann, R. The HIV-1 Rev Activation Domain Is a Nuclear Export Signal That Accesses an Export Pathway Used by Specific Cellular RNAs. Cell 1995, 82, 475–483. [CrossRef]
- Lai, M.-C.; Lee, Y.-H.W.; Tarn, W.-Y. The DEAD-Box RNA Helicase DDX3 Associates with Export Messenger Ribonucleoproteins as Well asTip-Associated Protein and Participates in Translational Control. Mol. Biol. Cell 2008, 19, 3847–3858. [CrossRef]
- Guo, J.; Zhu, Y.; Ma, X.; Shang, G.; Liu, B.; Zhang, K. Virus Infection and mRNA Nuclear Export. Int. J. Mol. Sci. 2023, 24, 12593. [CrossRef]
- Kuss, S.K.; Mata, M.A.; Zhang, L.; Fontoura, B.M.A. Nuclear Imprisonment: Viral Strategies to Arrest Host mRNA Nuclear Export. Viruses 2013, 5, 1824–1849. [CrossRef]
- Cavazza, T.; Vernos, I. The RanGTP Pathway: From Nucleo-Cytoplasmic Transport to Spindle Assembly and Beyond. Front. Cell Dev. Biol. 2016, 3. [CrossRef]
- Askjaer, P.; Jensen, T.H.; Nilsson, J.; Englmeier, L.; Kjems, J. The Specificity of the CRM1-Rev Nuclear Export Signal Interaction Is Mediated by RanGTP. J. Biol. Chem. 1998, 273, 33414–33422. [CrossRef]
- Liu, H.; Hu, P.-W.; Budhiraja, S.; Misra, A.; Couturier, J.; Lloyd, R.E.; Lewis, D.E.; Kimata, J.T.; Rice, A.P. PACS1 Is an HIV-1 Cofactor That Functions in Rev-Mediated Nuclear Export of Viral RNA. Virology 2020, 540, 88–96. [CrossRef]
- Kula, A.; Gharu, L.; Marcello, A. HIV-1 Pre-mRNA Commitment to Rev Mediated Export through PSF and Matrin 3. Virology 2013, 435, 329–340. [CrossRef]
- Jones, T.; Sheer, D.; Bevec, D.; Kappel, B.; Hauber, J.; Steinkasserer, A. The Human HIV-1 Rev Binding-Protein hRIP/Rab (HRB) Maps to Chromosome 2q36. Genomics 1997, 40, 198–199. [CrossRef]
- Li, J.; Tang, H.; Mullen, T.M.; Westberg, C.; Reddy, T.R.; Rose, D.W.; Wong-Staal, F. A Role for RNA Helicase A in Post-Transcriptional Regulation of HIV Type 1. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 709–714. [CrossRef]
- Budhiraja, S.; Liu, H.; Couturier, J.; Malovannaya, A.; Qin, J.; Lewis, D.E.; Rice, A.P. Mining the Human Complexome Database Identifies RBM14 as an XPO1-Associated Protein Involved in HIV-1 Rev Function. J. Virol. 2015, 89, 3557–3567. [CrossRef]
- Yedavalli, V.S.R.K.; Neuveut, C.; Chi, Y.; Kleiman, L.; Jeang, K.-T. Requirement of DDX3 DEAD Box RNA Helicase for HIV-1 Rev-RRE Export Function. Cell 2004, 119, 381–392. [CrossRef]
- Dayton, A.I. Within You, without You: HIV-1 Rev and RNA Export. Retrovirology 2004, 1, 35. [CrossRef]
- Fang, J.; Kubota, S.; Yang, B.; Zhou, N.; Zhang, H.; Godbout, R.; Pomerantz, R.J. A DEAD Box Protein Facilitates HIV-1 Replication as a Cellular Co-Factor of Rev. Virology 2004, 330, 471–480. [CrossRef]
- Li, J.; Liu, Y.; Kim, B.O.; He, J.J. Direct Participation of Sam68, the 68-Kilodalton Src-Associated Protein in Mitosis, in the CRM1-Mediated Rev Nuclear Export Pathway. J. Virol. 2002, 76, 8374–8382. [CrossRef]
- Schatz, O.; Oft, M.; Dascher, C.; Schebesta, M.; Rosorius, O.; Jaksche, H.; Dobrovnik, M.; Bevec, D.; Hauber, J. Interaction of the HIV-1 Rev Cofactor Eukaryotic Initiation Factor 5A with Ribosomal Protein L5. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 1607–1612. [CrossRef]
- Najera, I.; Krieg, M.; Karn, J. Synergistic Stimulation of HIV-1 Rev-Dependent Export of Unspliced mRNA to the Cytoplasm by hnRNP A1. J. Mol. Biol. 1999, 285, 1951–1964. [CrossRef]
- Fritz, C.C.; Zapp, M.L.; Green, M.R. A Human Nucleoporin-like Protein That Specifically Interacts with HIV Rev. Nature 1995, 376, 530–533. [CrossRef]
- Ajamian, L.; Abrahamyan, L.; Milev, M.; Ivanov, P.V.; Kulozik, A.E.; Gehring, N.H.; Mouland, A.J. Unexpected Roles for UPF1 in HIV-1 RNA Metabolism and Translation. RNA N. Y. N 2008, 14, 914–927. [CrossRef]
- Ajamian, L.; Abel, K.; Rao, S.; Vyboh, K.; García-de-Gracia, F.; Soto-Rifo, R.; Kulozik, A.E.; Gehring, N.H.; Mouland, A.J. HIV-1 Recruits UPF1 but Excludes UPF2 to Promote Nucleocytoplasmic Export of the Genomic RNA. Biomolecules 2015, 5, 2808–2839. [CrossRef]
- Park, E.; Maquat, L.E. Staufen-Mediated mRNA Decay. Wiley Interdiscip. Rev. RNA 2013, 4, 423–435. [CrossRef]
- Dugré-Brisson, S.; Elvira, G.; Boulay, K.; Chatel-Chaix, L.; Mouland, A.J.; DesGroseillers, L. Interaction of Staufen1 with the 5′ End of mRNA Facilitates Translation of These RNAs. Nucleic Acids Res. 2005, 33, 4797–4812. [CrossRef]
- Chatel-Chaix, L.; Clément, J.-F.; Martel, C.; Bériault, V.; Gatignol, A.; DesGroseillers, L.; Mouland, A.J. Identification of Staufen in the Human Immunodeficiency Virus Type 1 Gag Ribonucleoprotein Complex and a Role in Generating Infectious Viral Particles. Mol. Cell. Biol. 2004, 24, 2637–2648. [CrossRef]
- Banerjee, A.; Benjamin, R.; Balakrishnan, K.; Ghosh, P.; Banerjee, S. Human Protein Staufen-2 Promotes HIV-1 Proliferation by Positively Regulating RNA Export Activity of Viral Protein Rev. Retrovirology 2014, 11, 18. [CrossRef]
- Soto-Rifo, R.; Limousin, T.; Rubilar, P.S.; Ricci, E.P.; Décimo, D.; Moncorgé, O.; Trabaud, M.-A.; André, P.; Cimarelli, A.; Ohlmann, T. Different Effects of the TAR Structure on HIV-1 and HIV-2 Genomic RNA Translation. Nucleic Acids Res. 2012, 40, 2653–2667. [CrossRef]
- Parsyan, A.; Svitkin, Y.; Shahbazian, D.; Gkogkas, C.; Lasko, P.; Merrick, W.C.; Sonenberg, N. mRNA Helicases: The Tacticians of Translational Control. Nat. Rev. Mol. Cell Biol. 2011, 12, 235–245. [CrossRef]
- Ohlmann, T.; Mengardi, C.; López-Lastra, M. Translation Initiation of the HIV-1 mRNA. Translation 2014, 2, e960242. [CrossRef]
- Fujii, R.; Okamoto, M.; Aratani, S.; Oishi, T.; Ohshima, T.; Taira, K.; Baba, M.; Fukamizu, A.; Nakajima, T. A Role of RNA Helicase A in Cis-Acting Transactivation Response Element-Mediated Transcriptional Regulation of Human Immunodeficiency Virus Type 1. J. Biol. Chem. 2001, 276, 5445–5451. [CrossRef]
- Bolinger, C.; Sharma, A.; Singh, D.; Yu, L.; Boris-Lawrie, K. RNA Helicase A Modulates Translation of HIV-1 and Infectivity of Progeny Virions. Nucleic Acids Res. 2010, 38, 1686–1696. [CrossRef]
- Lai, M.-C.; Wang, S.-W.; Cheng, L.; Tarn, W.-Y.; Tsai, S.-J.; Sun, H.S. Human DDX3 Interacts with the HIV-1 Tat Protein to Facilitate Viral mRNA Translation. PloS One 2013, 8, e68665. [CrossRef]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The Mechanism of Eukaryotic Translation Initiation and Principles of Its Regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [CrossRef]
- Soto-Rifo, R.; Rubilar, P.S.; Limousin, T.; de Breyne, S.; Décimo, D.; Ohlmann, T. DEAD-Box Protein DDX3 Associates with eIF4F to Promote Translation of Selected mRNAs. EMBO J. 2012, 31, 3745–3756. [CrossRef]
- Soto-Rifo, R.; Rubilar, P.S.; Ohlmann, T. The DEAD-Box Helicase DDX3 Substitutes for the Cap-Binding Protein eIF4E to Promote Compartmentalized Translation Initiation of the HIV-1 Genomic RNA. Nucleic Acids Res. 2013, 41, 6286–6299. [CrossRef]
- Mo, J.; Liang, H.; Su, C.; Li, P.; Chen, J.; Zhang, B. DDX3X: Structure, Physiologic Functions and Cancer. Mol. Cancer 2021, 20, 38. [CrossRef]
- Chen, H.-H.; Yu, H.-I.; Yang, M.-H.; Tarn, W.-Y. DDX3 Activates CBC-eIF3–Mediated Translation of uORF-Containing Oncogenic mRNAs to Promote Metastasis in HNSCC. Cancer Res. 2018, 78, 4512–4523. [CrossRef]
- Ricci, E.P.; Rifo, R.S.; Herbreteau, C.H.; Decimo, D.; Ohlmann, T. Lentiviral RNAs Can Use Different Mechanisms for Translation Initiation. Biochem. Soc. Trans. 2008, 36, 690–693. [CrossRef]
- Brasey, A.; Lopez-Lastra, M.; Ohlmann, T.; Beerens, N.; Berkhout, B.; Darlix, J.-L.; Sonenberg, N. The Leader of Human Immunodeficiency Virus Type 1 Genomic RNA Harbors an Internal Ribosome Entry Segment That Is Active during the G2/M Phase of the Cell Cycle. J. Virol. 2003, 77, 3939–3949. [CrossRef]
- Buck, C.B.; Shen, X.; Egan, M.A.; Pierson, T.C.; Walker, C.M.; Siliciano, R.F. The Human Immunodeficiency Virus Type 1 Gag Gene Encodes an Internal Ribosome Entry Site. J. Virol. 2001, 75, 181–191. [CrossRef]
- Liu, J.; Henao-Mejia, J.; Liu, H.; Zhao, Y.; He, J.J. Translational Regulation of HIV-1 Replication by HIV-1 Rev Cellular Cofactors Sam68, eIF5A, hRIP, and DDX3. J. Neuroimmune Pharmacol. 2011, 6, 308–321. [CrossRef]
- Monette, A.; Ajamian, L.; López-Lastra, M.; Mouland, A.J. Human Immunodeficiency Virus Type 1 (HIV-1) Induces the Cytoplasmic Retention of Heterogeneous Nuclear Ribonucleoprotein A1 by Disrupting Nuclear Import. J. Biol. Chem. 2009, 284, 31350–31362. [CrossRef]
- Vallejos, M.; Deforges, J.; Plank, T.-D.M.; Letelier, A.; Ramdohr, P.; Abraham, C.G.; Valiente-Echeverría, F.; Kieft, J.S.; Sargueil, B.; López-Lastra, M. Activity of the Human Immunodeficiency Virus Type 1 Cell Cycle-Dependent Internal Ribosomal Entry Site Is Modulated by IRES Trans-Acting Factors. Nucleic Acids Res. 2011, 39, 6186–6200. [CrossRef]
- Lee, W.-Y.J.; Fu, R.M.; Liang, C.; Sloan, R.D. IFITM Proteins Inhibit HIV-1 Protein Synthesis. Sci. Rep. 2018, 8, 14551. [CrossRef]
- Shehu-Xhilaga, M.; Crowe, S.M.; Mak, J. Maintenance of the Gag/Gag-Pol Ratio Is Important for Human Immunodeficiency Virus Type 1 RNA Dimerization and Viral Infectivity. J. Virol. 2001, 75, 1834–1841. [CrossRef]
- Jacks, T.; Power, M.D.; Masiarz, F.R.; Luciw, P.A.; Barr, P.J.; Varmus, H.E. Characterization of Ribosomal Frameshifting in HIV-1 Gag-Pol Expression. Nature 1988, 331, 280–283. [CrossRef]
- Wang, X.; Xuan, Y.; Han, Y.; Ding, X.; Ye, K.; Yang, F.; Gao, P.; Goff, S.P.; Gao, G. Regulation of HIV-1 Gag-Pol Expression by Shiftless, an Inhibitor of Programmed −1 Ribosomal Frameshifting. Cell 2019, 176, 625-635.e14. [CrossRef]
- Li, M.; Kao, E.; Gao, X.; Sandig, H.; Limmer, K.; Pavon-Eternod, M.; Jones, T.E.; Landry, S.; Pan, T.; Weitzman, M.D.; et al. Codon-Usage-Based Inhibition of HIV Protein Synthesis by Human Schlafen 11. Nature 2012, 491, 125–128. [CrossRef]
- Kypr, J.; Mrázek, J. Unusual Codon Usage of HIV. Nature 1987, 327, 20–20. [CrossRef]
- Haas, J.; Park, E.C.; Seed, B. Codon Usage Limitation in the Expression of HIV-1 Envelope Glycoprotein. Curr. Biol. CB 1996, 6, 315–324. [CrossRef]
- Kofman, A.; Graf, M.; Bojak, A.; Deml, L.; Bieler, K.; Kharazova, A.; Wolf, H.; Wagner, R. HIV-1 Gag Expression Is Quantitatively Dependent on the Ratio of Native and Optimized Codons. Tsitologiia 2003, 45, 86–93.
- Kobayashi-Ishihara, M.; Frazão Smutná, K.; Alonso, F.E.; Argilaguet, J.; Esteve-Codina, A.; Geiger, K.; Genescà, M.; Grau-Expósito, J.; Duran-Castells, C.; Rogenmoser, S.; et al. Schlafen 12 Restricts HIV-1 Latency Reversal by a Codon-Usage Dependent Post-Transcriptional Block in CD4+ T Cells. Commun. Biol. 2023, 6, 1–15. [CrossRef]
- Rivas-Aravena, A.; Ramdohr, P.; Vallejos, M.; Valiente-Echeverría, F.; Dormoy-Raclet, V.; Rodríguez, F.; Pino, K.; Holzmann, C.; Huidobro-Toro, J.P.; Gallouzi, I.-E.; et al. The Elav-like Protein HuR Exerts Translational Control of Viral Internal Ribosome Entry Sites. Virology 2009, 392, 178–185. [CrossRef]
- López-Ulloa, B.; Fuentes, Y.; Pizarro-Ortega, M.S.; López-Lastra, M. RNA-Binding Proteins as Regulators of Internal Initiation of Viral mRNA Translation. Viruses 2022, 14, 188. [CrossRef]
- Wang, Q.; Gao, H.; Clark, K.M.; Mugisha, C.S.; Davis, K.; Tang, J.P.; Harlan, G.H.; DeSelm, C.J.; Presti, R.M.; Kutluay, S.B.; et al. CARD8 Is an Inflammasome Sensor for HIV-1 Protease Activity. Science 2021, 371, eabe1707. [CrossRef]
- Wang, Q.; Clark, K.M.; Tiwari, R.; Raju, N.; Tharp, G.K.; Rogers, J.; Harris, R.A.; Raveendran, M.; Bosinger, S.E.; Burdo, T.H.; et al. The CARD8 Inflammasome Dictates HIV/SIV Pathogenesis and Disease Progression. Cell 2024, 187, 1223-1237.e16. [CrossRef]
- Linder, A.; Bauernfried, S.; Cheng, Y.; Albanese, M.; Jung, C.; Keppler, O.T.; Hornung, V. CARD8 Inflammasome Activation Triggers Pyroptosis in Human T Cells. EMBO J. 2020, 39, e105071. [CrossRef]
- Park, J.; Morrow, C.D. Overexpression of the Gag-Pol Precursor from Human Immunodeficiency Virus Type 1 Proviral Genomes Results in Efficient Proteolytic Processing in the Absence of Virion Production. J. Virol. 1991, 65, 5111–5117. [CrossRef]
- Finkel, T.H.; Tudor-Williams, G.; Banda, N.K.; Cotton, M.F.; Curiel, T.; Monks, C.; Baba, T.W.; Ruprecht, R.M.; Kupfer, A. Apoptosis Occurs Predominantly in Bystander Cells and Not in Productively Infected Cells of HIV- and SIV-Infected Lymph Nodes. Nat. Med. 1995, 1, 129–134. [CrossRef]
- Clerzius, G.; Gélinas, J.-F.; Daher, A.; Bonnet, M.; Meurs, E.F.; Gatignol, A. ADAR1 Interacts with PKR during Human Immunodeficiency Virus Infection of Lymphocytes and Contributes to Viral Replication. J. Virol. 2009, 83, 10119–10128. [CrossRef]
- Clerzius, G.; Gélinas, J.-F.; Gatignol, A. Multiple Levels of PKR Inhibition during HIV-1 Replication. Rev. Med. Virol. 2011, 21, 42–53. [CrossRef]
- Park, H.; Davies, M.V.; Langland, J.O.; Chang, H.W.; Nam, Y.S.; Tartaglia, J.; Paoletti, E.; Jacobs, B.L.; Kaufman, R.J.; Venkatesan, S. TAR RNA-Binding Protein Is an Inhibitor of the Interferon-Induced Protein Kinase PKR. Proc. Natl. Acad. Sci. U. S. A. 1994, 91, 4713–4717. [CrossRef]
- Benkirane, M.; Neuveut, C.; Chun, R.F.; Smith, S.M.; Samuel, C.E.; Gatignol, A.; Jeang, K. Oncogenic Potential of TAR RNA Binding Protein TRBP and Its Regulatory Interaction with RNA-dependent Protein Kinase PKR. EMBO J. 1997, 16, 611–624. [CrossRef]
- Sanghvi, V.R.; Steel, L.F. The Cellular TAR RNA Binding Protein, TRBP, Promotes HIV-1 Replication Primarily by Inhibiting the Activation of Double-Stranded RNA-Dependent Kinase PKR. J. Virol. 2011, 85, 12614–12621. [CrossRef]
- Laraki, G.; Clerzius, G.; Daher, A.; Melendez-Peña, C.; Daniels, S.; Gatignol, A. Interactions between the Double-Stranded RNA-Binding Proteins TRBP and PACT Define the Medipal Domain That Mediates Protein-Protein Interactions. RNA Biol. 2008, 5, 92–103. [CrossRef]
- Daher, A.; Laraki, G.; Singh, M.; Melendez-Peña, C.E.; Bannwarth, S.; Peters, A.H.F.M.; Meurs, E.F.; Braun, R.E.; Patel, R.C.; Gatignol, A. TRBP Control of PACT-Induced Phosphorylation of Protein Kinase R Is Reversed by Stress. Mol. Cell. Biol. 2009, 29, 254–265. [CrossRef]
- Peters, G.A.; Dickerman, B.; Sen, G.C. Biochemical Analysis of PKR Activation by PACT. Biochemistry 2009, 48, 7441–7447. [CrossRef]
- Li, S.; Sen, G.C. PACT-Mediated Enhancement of Reporter Gene Expression at the Translational Level. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2003, 23, 689–697. [CrossRef]
- Clerzius, G.; Shaw, E.; Daher, A.; Burugu, S.; Gélinas, J.-F.; Ear, T.; Sinck, L.; Routy, J.-P.; Mouland, A.J.; Patel, R.C.; et al. The PKR Activator, PACT, Becomes a PKR Inhibitor during HIV-1 Replication. Retrovirology 2013, 10, 96. [CrossRef]
- Burugu, S.; Daher, A.; Meurs, E.F.; Gatignol, A. HIV-1 Translation and Its Regulation by Cellular Factors PKR and PACT. Virus Res. 2014, 193, 65–77. [CrossRef]
- Gheysen, D.; Jacobs, E.; Foresta, F. de; Thiriart, C.; Francotte, M.; Thines, D.; Wilde, M.D. Assembly and Release of HIV-1 Precursor Pr55gag Virus-like Particles from Recombinant Baculovirus-Infected Insect Cells. Cell 1989, 59, 103–112. [CrossRef]
- Maréchal, V.; Clavel, F.; Heard, J.M.; Schwartz, O. Cytosolic Gag P24 as an Index of Productive Entry of Human Immunodeficiency Virus Type 1. J. Virol. 1998, 72, 2208–2212. [CrossRef]
- Tedbury, P.R.; Freed, E.O. The Role of Matrix in HIV-1 Envelope Glycoprotein Incorporation. Trends Microbiol. 2014, 22, 372–378. [CrossRef]
- Mu, X.; Fu, Y.; Zhu, Y.; Wang, X.; Xuan, Y.; Shang, H.; Goff, S.P.; Gao, G. HIV-1 Exploits the Host Factor RuvB-like 2 to Balance Viral Protein Expression. Cell Host Microbe 2015, 18, 233–242. [CrossRef]
- Rucevic, M.; Boucau, J.; Dinter, J.; Kourjian, G.; Le Gall, S. Mechanisms of HIV Protein Degradation into Epitopes: Implications for Vaccine Design. Viruses 2014, 6, 3271–3292. [CrossRef]
- Lata, S.; Mishra, R.; Banerjea, A.C. Proteasomal Degradation Machinery: Favorite Target of HIV-1 Proteins. Front. Microbiol. 2018, 9. [CrossRef]
- Kimura, H.; Caturegli, P.; Takahashi, M.; Suzuki, K. New Insights into the Function of the Immunoproteasome in Immune and Nonimmune Cells. J. Immunol. Res. 2015, 2015, 541984. [CrossRef]
- Chen, W.; Norbury, C.C.; Cho, Y.; Yewdell, J.W.; Bennink, J.R. Immunoproteasomes Shape Immunodominance Hierarchies of Antiviral Cd8+ T Cells at the Levels of T Cell Repertoire and Presentation of Viral Antigens. J. Exp. Med. 2001, 193, 1319–1326. [CrossRef]
- Seissler, T.; Marquet, R.; Paillart, J.-C. Hijacking of the Ubiquitin/Proteasome Pathway by the HIV Auxiliary Proteins. Viruses 2017, 9, 322. [CrossRef]
- Li, J.; Chen, C.; Ma, X.; Geng, G.; Liu, B.; Zhang, Y.; Zhang, S.; Zhong, F.; Liu, C.; Yin, Y.; et al. Long Noncoding RNA NRON Contributes to HIV-1 Latency by Specifically Inducing Tat Protein Degradation. Nat. Commun. 2016, 7, 11730. [CrossRef]
- Danielson, C.M.; Cianci, G.C.; Hope, T.J. Recruitment and Dynamics of Proteasome Association with rhTRIM5α Cytoplasmic Complexes during HIV-1 Infection. Traffic Cph. Den. 2012, 13, 1206–1217. [CrossRef]
- Lukic, Z.; Hausmann, S.; Sebastian, S.; Rucci, J.; Sastri, J.; Robia, S.L.; Luban, J.; Campbell, E.M. TRIM5α Associates with Proteasomal Subunits in Cells While in Complex with HIV-1 Virions. Retrovirology 2011, 8, 93. [CrossRef]
- Pyeon, D.; Timani, K.A.; Gulraiz, F.; Park, I.-W. Function of Ubiquitin (Ub) Specific Protease 15 (USP15) in HIV-1 Replication and Viral Protein Degradation. Virus Res. 2016, 223, 161–169. [CrossRef]
- Izumi, T.; Takaori-Kondo, A.; Shirakawa, K.; Higashitsuji, H.; Itoh, K.; Io, K.; Matsui, M.; Iwai, K.; Kondoh, H.; Sato, T.; et al. MDM2 Is a Novel E3 Ligase for HIV-1 Vif. Retrovirology 2009, 6, 1. [CrossRef]
- Fujita, M.; Akari, H.; Sakurai, A.; Yoshida, A.; Chiba, T.; Tanaka, K.; Strebel, K.; Adachi, A. Expression of HIV-1 Accessory Protein Vif Is Controlled Uniquely to Be Low and Optimal by Proteasome Degradation. Microbes Infect. 2004, 6, 791–798. [CrossRef]
- Zhao, L.; Wang, S.; Xu, M.; He, Y.; Zhang, X.; Xiong, Y.; Sun, H.; Ding, H.; Geng, W.; Shang, H.; et al. Vpr Counteracts the Restriction of LAPTM5 to Promote HIV-1 Infection in Macrophages. Nat. Commun. 2021, 12, 3691. [CrossRef]
- Ouyang, J.; Xiong, Y.; Shang, H.; Liang, G. LAPTM5 Restricts HIV-1 Infection in Dendritic Cells and Is Counteracted by Vpr. Microbiol. Spectr. 2022, 10, e0138221. [CrossRef]
- Mashiba, M.; Collins, D.R.; Terry, V.H.; Collins, K.L. Vpr Overcomes Macrophage-Specific Restriction of HIV-1 Env Expression and Virion Production. Cell Host Microbe 2014, 16, 722–735. [CrossRef]
- Collins, D.R.; Lubow, J.; Lukic, Z.; Mashiba, M.; Collins, K.L. Vpr Promotes Macrophage-Dependent HIV-1 Infection of CD4+ T Lymphocytes. PLOS Pathog. 2015, 11, e1005054. [CrossRef]
- Lubow, J.; Virgilio, M.C.; Merlino, M.; Collins, D.R.; Mashiba, M.; Peterson, B.G.; Lukic, Z.; Painter, M.M.; Gomez-Rivera, F.; Terry, V.; et al. Mannose Receptor Is an HIV Restriction Factor Counteracted by Vpr in Macrophages. eLife 2020, 9, e51035. [CrossRef]
- MARTíN-OROZCO, N.; ISIBASI, A.; ORTIZ-NAVARRETE, V. Macrophages Present Exogenous Antigens by Class I Major Histocompatibility Complex Molecules via a Secretory Pathway as a Consequence of Interferon-γ Activation. Immunology 2001, 103, 41–48. [CrossRef]
- Reith, W.; LeibundGut-Landmann, S.; Waldburger, J.-M. Regulation of MHC Class II Gene Expression by the Class II Transactivator. Nat. Rev. Immunol. 2005, 5, 793–806. [CrossRef]
- Zhang, N.; Bevan, M.J. CD8+ T Cells: Foot Soldiers of the Immune System. Immunity 2011, 35, 161–168. [CrossRef]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4+T Cells: Differentiation and Functions. Clin. Dev. Immunol. 2012, 2012, 925135. [CrossRef]
- Perišić Nanut, M.; Sabotič, J.; Jewett, A.; Kos, J. Cysteine Cathepsins as Regulators of the Cytotoxicity of NK and T Cells. Front. Immunol. 2014, 5, 616. [CrossRef]
- Steers, N.J.; Ratto-Kim, S.; Souza, M.S. de; Currier, J.R.; Kim, J.H.; Michael, N.L.; Alving, C.R.; Rao, M. HIV-1 Envelope Resistance to Proteasomal Cleavage: Implications for Vaccine Induced Immune Responses. PLOS ONE 2012, 7, e42579. [CrossRef]
- Tenzer, S.; Wee, E.; Burgevin, A.; Stewart-Jones, G.; Friis, L.; Lamberth, K.; Chang, C.; Harndahl, M.; Weimershaus, M.; Gerstoft, J.; et al. Antigen Processing Influences HIV-Specific Cytotoxic T Lymphocyte Immunodominance. Nat. Immunol. 2009, 10, 636–646. [CrossRef]
- Tenzer, S.; Crawford, H.; Pymm, P.; Gifford, R.; Sreenu, V.B.; Weimershaus, M.; de Oliveira, T.; Burgevin, A.; Gerstoft, J.; Akkad, N.; et al. HIV-1 Adaptation to Antigen Processing Results in Population-Level Immune Evasion and Affects Subtype Diversification. Cell Rep. 2014, 7, 448–463. [CrossRef]
- Hermida-Matsumoto, L.; Resh, M.D. Localization of Human Immunodeficiency Virus Type 1 Gag and Env at the Plasma Membrane by Confocal Imaging. J. Virol. 2000, 74, 8670–8679. [CrossRef]
- Freed, E.O. HIV-1 Assembly, Release and Maturation. Nat. Rev. Microbiol. 2015, 13, 484–496. [CrossRef]
- Ghanam, R.H.; Samal, A.B.; Fernandez, T.F.; Saad, J.S. Role of the HIV-1 Matrix Protein in Gag Intracellular Trafficking and Targeting to the Plasma Membrane for Virus Assembly. Front. Microbiol. 2012, 3. [CrossRef]
- Simonsen, A.; Wurmser, A.E.; Emr, S.D.; Stenmark, H. The Role of Phosphoinositides in Membrane Transport. Curr. Opin. Cell Biol. 2001, 13, 485–492. [CrossRef]
- Ono, A.; Ablan, S.D.; Lockett, S.J.; Nagashima, K.; Freed, E.O. Phosphatidylinositol (4,5) Bisphosphate Regulates HIV-1 Gag Targeting to the Plasma Membrane. Proc. Natl. Acad. Sci. 2004, 101, 14889–14894. [CrossRef]
- Graham, D.R.M.; Chertova, E.; Hilburn, J.M.; Arthur, L.O.; Hildreth, J.E.K. Cholesterol Depletion of Human Immunodeficiency Virus Type 1 and Simian Immunodeficiency Virus with β-Cyclodextrin Inactivates and Permeabilizes the Virions: Evidence for Virion-Associated Lipid Rafts. J. Virol. 2003, 77, 8237–8248. [CrossRef]
- Alfadhli, A.; Still, A.; Barklis, E. Analysis of Human Immunodeficiency Virus Type 1 Matrix Binding to Membranes and Nucleic Acids. J. Virol. 2009, 83, 12196–12203. [CrossRef]
- Murphy, R.E.; Saad, J.S. The Interplay between HIV-1 Gag Binding to the Plasma Membrane and Env Incorporation. Viruses 2020, 12, 548. [CrossRef]
- Udenwobele, D.I.; Su, R.-C.; Good, S.V.; Ball, T.B.; Varma Shrivastav, S.; Shrivastav, A. Myristoylation: An Important Protein Modification in the Immune Response. Front. Immunol. 2017, 8. [CrossRef]
- Göttlinger, H.G.; Sodroski, J.G.; Haseltine, W.A. Role of Capsid Precursor Processing and Myristoylation in Morphogenesis and Infectivity of Human Immunodeficiency Virus Type 1. Proc. Natl. Acad. Sci. U. S. A. 1989, 86, 5781–5785. [CrossRef]
- Bryant, M.; Ratner, L. Myristoylation-Dependent Replication and Assembly of Human Immunodeficiency Virus 1. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 523–527. [CrossRef]
- Ono, A.; Freed, E.O. Binding of Human Immunodeficiency Virus Type 1 Gag to Membrane: Role of the Matrix Amino Terminus. J. Virol. 1999, 73, 4136–4144. [CrossRef]
- Li, H.; Dou, J.; Ding, L.; Spearman, P. Myristoylation Is Required for Human Immunodeficiency Virus Type 1 Gag-Gag Multimerization in Mammalian Cells. J. Virol. 2007, 81, 12899–12910. [CrossRef]
- Paillart, J.C.; Göttlinger, H.G. Opposing Effects of Human Immunodeficiency Virus Type 1 Matrix Mutations Support a Myristyl Switch Model of Gag Membrane Targeting. J. Virol. 1999, 73, 2604–2612. [CrossRef]
- Saad, J.S.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural Basis for Targeting HIV-1 Gag Proteins to the Plasma Membrane for Virus Assembly. Proc. Natl. Acad. Sci. 2006, 103, 11364–11369. [CrossRef]
- Dalton, A.K.; Ako-Adjei, D.; Murray, P.S.; Murray, D.; Vogt, V.M. Electrostatic Interactions Drive Membrane Association of the Human Immunodeficiency Virus Type 1 Gag MA Domain. J. Virol. 2007, 81, 6434–6445. [CrossRef]
- Alfadhli, A.; Huseby, D.; Kapit, E.; Colman, D.; Barklis, E. Human Immunodeficiency Virus Type 1 Matrix Protein Assembles on Membranes as a Hexamer. J. Virol. 2007, 81, 1472–1478. [CrossRef]
- Willey, R.L.; Bonifacino, J.S.; Potts, B.J.; Martin, M.A.; Klausner, R.D. Biosynthesis, Cleavage, and Degradation of the Human Immunodeficiency Virus 1 Envelope Glycoprotein Gp160. Proc. Natl. Acad. Sci. 1988, 85, 9580–9584. [CrossRef]
- Qi, M.; Williams, J.A.; Chu, H.; Chen, X.; Wang, J.-J.; Ding, L.; Akhirome, E.; Wen, X.; Lapierre, L.A.; Goldenring, J.R.; et al. Rab11-FIP1C and Rab14 Direct Plasma Membrane Sorting and Particle Incorporation of the HIV-1 Envelope Glycoprotein Complex. PLoS Pathog. 2013, 9, e1003278. [CrossRef]
- Postler, T.S.; Desrosiers, R.C. The Tale of the Long Tail: The Cytoplasmic Domain of HIV-1 Gp41. J. Virol. 2013, 87, 2–15. [CrossRef]
- Wyss, S.; Berlioz-Torrent, C.; Boge, M.; Blot, G.; Höning, S.; Benarous, R.; Thali, M. The Highly Conserved C-Terminal Dileucine Motif in the Cytosolic Domain of the Human Immunodeficiency Virus Type 1 Envelope Glycoprotein Is Critical for Its Association with the AP-1 Clathrin Adapter. J. Virol. 2001, 75, 2982–2992. [CrossRef]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 Envelope Glycoprotein Biosynthesis, Trafficking, and Incorporation. J. Mol. Biol. 2011, 410, 582–608. [CrossRef]
- Murakami, T.; Freed, E.O. Genetic Evidence for an Interaction between Human Immunodeficiency Virus Type 1 Matrix and Alpha-Helix 2 of the Gp41 Cytoplasmic Tail. J. Virol. 2000, 74, 3548–3554. [CrossRef]
- Gousset, K.; Ablan, S.D.; Coren, L.V.; Ono, A.; Soheilian, F.; Nagashima, K.; Ott, D.E.; Freed, E.O. Real-Time Visualization of HIV-1 GAG Trafficking in Infected Macrophages. PLOS Pathog. 2008, 4, e1000015. [CrossRef]
- Raposo, G.; Moore, M.; Innes, D.; Leijendekker, R.; Leigh-Brown, A.; Benaroch, P.; Geuze, H. Human Macrophages Accumulate HIV-1 Particles in MHC II Compartments. Traffic Cph. Den. 2002, 3, 718–729. [CrossRef]
- Pelchen-Matthews, A.; Kramer, B.; Marsh, M. Infectious HIV-1 Assembles in Late Endosomes in Primary Macrophages. J. Cell Biol. 2003, 162, 443–455. [CrossRef]
- Ono, A.; Freed, E.O. Cell-Type-Dependent Targeting of Human Immunodeficiency Virus Type 1 Assembly to the Plasma Membrane and the Multivesicular Body. J. Virol. 2004, 78, 1552–1563. [CrossRef]
- Deneka, M.; Pelchen-Matthews, A.; Byland, R.; Ruiz-Mateos, E.; Marsh, M. In Macrophages, HIV-1 Assembles into an Intracellular Plasma Membrane Domain Containing the Tetraspanins CD81, CD9, and CD53. J. Cell Biol. 2007, 177, 329–341. [CrossRef]
- Tan, J.; Sattentau, Q.J. The HIV-1-Containing Macrophage Compartment: A Perfect Cellular Niche? Trends Microbiol. 2013, 21, 405–412. [CrossRef]
- Lerner, G.; Weaver, N.; Anokhin, B.; Spearman, P. Advances in HIV-1 Assembly. Viruses 2022, 14, 478. [CrossRef]
- Bennett, A.E.; Narayan, K.; Shi, D.; Hartnell, L.M.; Gousset, K.; He, H.; Lowekamp, B.C.; Yoo, T.S.; Bliss, D.; Freed, E.O.; et al. Ion-Abrasion Scanning Electron Microscopy Reveals Surface-Connected Tubular Conduits in HIV-Infected Macrophages. PLOS Pathog. 2009, 5, e1000591. [CrossRef]
- Hammonds, J.E.; Beeman, N.; Ding, L.; Takushi, S.; Francis, A.C.; Wang, J.-J.; Melikyan, G.B.; Spearman, P. Siglec-1 Initiates Formation of the Virus-Containing Compartment and Enhances Macrophage-to-T Cell Transmission of HIV-1. PLOS Pathog. 2017, 13, e1006181. [CrossRef]
- Groot, F.; Welsch, S.; Sattentau, Q.J. Efficient HIV-1 Transmission from Macrophages to T Cells across Transient Virological Synapses. Blood 2008, 111, 4660–4663. [CrossRef]
- Chu, H.; Wang, J.-J.; Qi, M.; Yoon, J.-J.; Chen, X.; Wen, X.; Hammonds, J.; Ding, L.; Spearman, P. Tetherin/BST-2 Is Essential for the Formation of the Intracellular Virus-Containing Compartment in HIV-Infected Macrophages. Cell Host Microbe 2012, 12, 360–372. [CrossRef]
- Mlcochova, P.; Pelchen-Matthews, A.; Marsh, M. Organization and Regulation of Intracellular Plasma Membrane-Connected HIV-1 Assembly Compartments in Macrophages. BMC Biol. 2013, 11, 89. [CrossRef]
- Mariani, C.; Desdouits, M.; Favard, C.; Benaroch, P.; Muriaux, D.M. Role of Gag and Lipids during HIV-1 Assembly in CD4+ T Cells and Macrophages. Front. Microbiol. 2014, 5. [CrossRef]
- Hogue, I.B.; Grover, J.R.; Soheilian, F.; Nagashima, K.; Ono, A. Gag Induces the Coalescence of Clustered Lipid Rafts and Tetraspanin-Enriched Microdomains at HIV-1 Assembly Sites on the Plasma Membrane. J. Virol. 2011, 85, 9749–9766. [CrossRef]
- Ono, A.; Freed, E.O. Plasma Membrane Rafts Play a Critical Role in HIV-1 Assembly and Release. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 13925–13930. [CrossRef]
- Zimmerman, C.; Klein, K.C.; Kiser, P.K.; Singh, A.R.; Firestein, B.L.; Riba, S.C.; Lingappa, J.R. Identification of a Host Protein Essential for Assembly of Immature HIV-1 Capsids. Nature 2002, 415, 88–92. [CrossRef]
- Lingappa, J.R.; Dooher, J.E.; Newman, M.A.; Kiser, P.K.; Klein, K.C. Basic Residues in the Nucleocapsid Domain of Gag Are Required for Interaction of HIV-1 Gag with ABCE1 (HP68), a Cellular Protein Important for HIV-1 Capsid Assembly. J. Biol. Chem. 2006, 281, 3773–3784. [CrossRef]
- Dooher, J.E.; Schneider, B.L.; Reed, J.C.; Lingappa, J.R. Host ABCE1 Is at Plasma Membrane HIV Assembly Sites and Its Dissociation from Gag Is Linked to Subsequent Events of Virus Production. Traffic Cph. Den. 2007, 8, 195–211. [CrossRef]
- Chatel-Chaix, L.; Abrahamyan, L.; Fréchina, C.; Mouland, A.J.; DesGroseillers, L. The Host Protein Staufen1 Participates in Human Immunodeficiency Virus Type 1 Assembly in Live Cells by Influencing pr55Gag Multimerization. J. Virol. 2007, 81, 6216–6230. [CrossRef]
- Chatel-Chaix, L.; Boulay, K.; Mouland, A.J.; DesGroseillers, L. The Host Protein Staufen1 Interacts with the Pr55Gagzinc Fingers and Regulates HIV-1 Assembly via Its N-Terminus. Retrovirology 2008, 5, 41. [CrossRef]
- Mouland, A.J.; Mercier, J.; Luo, M.; Bernier, L.; DesGroseillers, L.; Cohen, E.A. The Double-Stranded RNA-Binding Protein Staufen Is Incorporated in Human Immunodeficiency Virus Type 1: Evidence for a Role in Genomic RNA Encapsidation. J. Virol. 2000, 74, 5441–5451. [CrossRef]
- Dick, R.A.; Xu, C.; Morado, D.R.; Kravchuk, V.; Ricana, C.L.; Lyddon, T.D.; Broad, A.M.; Feathers, J.R.; Johnson, M.C.; Vogt, V.M.; et al. Structures of Immature EIAV Gag Lattices Reveal a Conserved Role for IP6 in Lentivirus Assembly. PLoS Pathog. 2020, 16, e1008277. [CrossRef]
- Dick, R.A.; Zadrozny, K.K.; Xu, C.; Schur, F.K.M.; Lyddon, T.D.; Ricana, C.L.; Wagner, J.M.; Perilla, J.R.; Ganser-Pornillos, B.K.; Johnson, M.C.; et al. Inositol Phosphates Are Assembly Co-Factors for HIV-1. Nature 2018, 560, 509–512. [CrossRef]
- Nikolaitchik, O.A.; Dilley, K.A.; Fu, W.; Gorelick, R.J.; Tai, S.-H.S.; Soheilian, F.; Ptak, R.G.; Nagashima, K.; Pathak, V.K.; Hu, W.-S. Dimeric RNA Recognition Regulates HIV-1 Genome Packaging. PLoS Pathog. 2013, 9, e1003249. [CrossRef]
- Levin, J.G.; Guo, J.; Rouzina, I.; Musier-Forsyth, K. Nucleic Acid Chaperone Activity of HIV-1 Nucleocapsid Protein: Critical Role in Reverse Transcription and Molecular Mechanism. Prog. Nucleic Acid Res. Mol. Biol. 2005, 80, 217–286. [CrossRef]
- Dorfman, T.; Luban, J.; Goff, S.P.; Haseltine, W.A.; Göttlinger, H.G. Mapping of Functionally Important Residues of a Cysteine-Histidine Box in the Human Immunodeficiency Virus Type 1 Nucleocapsid Protein. J. Virol. 1993, 67, 6159–6169. [CrossRef]
- Muriaux, D.; Darlix, J.-L. Properties and Functions of the Nucleocapsid Protein in Virus Assembly. RNA Biol. 2010, 7, 744–753. [CrossRef]
- Bacharach, E.; Gonsky, J.; Alin, K.; Orlova, M.; Goff, S.P. The Carboxy-Terminal Fragment of Nucleolin Interacts with the Nucleocapsid Domain of Retroviral Gag Proteins and Inhibits Virion Assembly. J. Virol. 2000, 74, 11027–11039. [CrossRef]
- Ueno, T.; Tokunaga, K.; Sawa, H.; Maeda, M.; Chiba, J.; Kojima, A.; Hasegawa, H.; Shoya, Y.; Sata, T.; Kurata, T.; et al. Nucleolin and the Packaging Signal, ψ, Promote the Budding of Human Immunodeficiency Virus Type-1 (HIV-1). Microbiol. Immunol. 2004, 48, 111–118. [CrossRef]
- Cohen, E.A.; Dehni, G.; Sodroski, J.G.; Haseltine, W.A. Human Immunodeficiency Virus Vpr Product Is a Virion-Associated Regulatory Protein. J. Virol. 1990, 64, 3097–3099. [CrossRef]
- Kao, S.; Akari, H.; Khan, M.A.; Dettenhofer, M.; Yu, X.-F.; Strebel, K. Human Immunodeficiency Virus Type 1 Vif Is Efficiently Packaged into Virions during Productive but Not Chronic Infection. J. Virol. 2003, 77, 1131–1140. [CrossRef]
- Khan, M.A.; Akari, H.; Kao, S.; Aberham, C.; Davis, D.; Buckler-White, A.; Strebel, K. Intravirion Processing of the Human Immunodeficiency Virus Type 1 Vif Protein by the Viral Protease May Be Correlated with Vif Function. J. Virol. 2002, 76, 9112–9123. [CrossRef]
- Bukovsky, A.A.; Dorfman, T.; Weimann, A.; Göttlinger, H.G. Nef Association with Human Immunodeficiency Virus Type 1 Virions and Cleavage by the Viral Protease. J. Virol. 1997, 71, 1013–1018. [CrossRef]
- Forshey, B.M.; Aiken, C. Disassembly of Human Immunodeficiency Virus Type 1 Cores in Vitro Reveals Association of Nef with the Subviral Ribonucleoprotein Complex. J. Virol. 2003, 77, 4409–4414. [CrossRef]
- Santos, S.; Obukhov, Y.; Nekhai, S.; Bukrinsky, M.; Iordanskiy, S. Virus-Producing Cells Determine the Host Protein Profiles of HIV-1 Virion Cores. Retrovirology 2012, 9, 65. [CrossRef]
- Burnie, J.; Guzzo, C. The Incorporation of Host Proteins into the External HIV-1 Envelope. Viruses 2019, 11, 85. [CrossRef]
- Orecchini, E.; Federico, M.; Doria, M.; Arenaccio, C.; Giuliani, E.; Ciafrè, S.A.; Michienzi, A. The ADAR1 Editing Enzyme Is Encapsidated into HIV-1 Virions. Virology 2015, 485, 475–480. [CrossRef]
- Göttlinger, H.G.; Dorfman, T.; Sodroski, J.G.; Haseltine, W.A. Effect of Mutations Affecting the P6 Gag Protein on Human Immunodeficiency Virus Particle Release. Proc. Natl. Acad. Sci. U. S. A. 1991, 88, 3195–3199. [CrossRef]
- Huang, M.; Orenstein, J.M.; Martin, M.A.; Freed, E.O. p6Gag Is Required for Particle Production from Full-Length Human Immunodeficiency Virus Type 1 Molecular Clones Expressing Protease. J. Virol. 1995, 69, 6810–6818. [CrossRef]
- Nguyen, D.G.; Booth, A.; Gould, S.J.; Hildreth, J.E.K. Evidence That HIV Budding in Primary Macrophages Occurs through the Exosome Release Pathway. J. Biol. Chem. 2003, 278, 52347–52354. [CrossRef]
- Votteler, J.; Sundquist, W.I. Virus Budding and the ESCRT Pathway. Cell Host Microbe 2013, 14, 10.1016/j.chom.2013.08.012. [CrossRef]
- Raiborg, C.; Stenmark, H. The ESCRT Machinery in Endosomal Sorting of Ubiquitylated Membrane Proteins. Nature 2009, 458, 445–452. [CrossRef]
- VerPlank, L.; Bouamr, F.; LaGrassa, T.J.; Agresta, B.; Kikonyogo, A.; Leis, J.; Carter, C.A. Tsg101, a Homologue of Ubiquitin-Conjugating (E2) Enzymes, Binds the L Domain in HIV Type 1 Pr55(Gag). Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 7724–7729. [CrossRef]
- Garrus, J.E.; von Schwedler, U.K.; Pornillos, O.W.; Morham, S.G.; Zavitz, K.H.; Wang, H.E.; Wettstein, D.A.; Stray, K.M.; Côté, M.; Rich, R.L.; et al. Tsg101 and the Vacuolar Protein Sorting Pathway Are Essential for HIV-1 Budding. Cell 2001, 107, 55–65. [CrossRef]
- Demirov, D.G.; Ono, A.; Orenstein, J.M.; Freed, E.O. Overexpression of the N-Terminal Domain of TSG101 Inhibits HIV-1 Budding by Blocking Late Domain Function. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 955–960. [CrossRef]
- Martin-Serrano, J.; Zang, T.; Bieniasz, P.D. HIV-1 and Ebola Virus Encode Small Peptide Motifs That Recruit Tsg101 to Sites of Particle Assembly to Facilitate Egress. Nat. Med. 2001, 7, 1313–1319. [CrossRef]
- Joshi, A.; Munshi, U.; Ablan, S.D.; Nagashima, K.; Freed, E.O. Functional Replacement of a Retroviral Late Domain by Ubiquitin Fusion. Traffic Cph. Den. 2008, 9, 1972–1983. [CrossRef]
- Bleck, M.; Itano, M.S.; Johnson, D.S.; Thomas, V.K.; North, A.J.; Bieniasz, P.D.; Simon, S.M. Temporal and Spatial Organization of ESCRT Protein Recruitment during HIV-1 Budding. Proc. Natl. Acad. Sci. 2014, 111, 12211–12216. [CrossRef]
- Sette, P.; Nagashima, K.; Piper, R.C.; Bouamr, F. Ubiquitin Conjugation to Gag Is Essential for ESCRT-Mediated HIV-1 Budding. Retrovirology 2013, 10, 79. [CrossRef]
- Martin-Serrano, J.; Neil, S.J.D. Host Factors Involved in Retroviral Budding and Release. Nat. Rev. Microbiol. 2011, 9, 519–531. [CrossRef]
- Schwedler, U.K. von; Stuchell, M.; Müller, B.; Ward, D.M.; Chung, H.-Y.; Morita, E.; Wang, H.E.; Davis, T.; He, G.-P.; Cimbora, D.M.; et al. The Protein Network of HIV Budding. Cell 2003, 114, 701–713. [CrossRef]
- Pineda-Molina, E.; Belrhali, H.; Piefer, A.J.; Akula, I.; Bates, P.; Weissenhorn, W. The Crystal Structure of the C-Terminal Domain of Vps28 Reveals a Conserved Surface Required for Vps20 Recruitment. Traffic 2006, 7, 1007–1016. [CrossRef]
- Langelier, C.; von Schwedler, U.K.; Fisher, R.D.; De Domenico, I.; White, P.L.; Hill, C.P.; Kaplan, J.; Ward, D.; Sundquist, W.I. Human ESCRT-II Complex and Its Role in Human Immunodeficiency Virus Type 1 Release. J. Virol. 2006, 80, 9465–9480. [CrossRef]
- Strack, B.; Calistri, A.; Craig, S.; Popova, E.; Göttlinger, H.G. AIP1/ALIX Is a Binding Partner for HIV-1 P6 and EIAV P9 Functioning in Virus Budding. Cell 2003, 114, 689–699. [CrossRef]
- Fisher, R.D.; Chung, H.-Y.; Zhai, Q.; Robinson, H.; Sundquist, W.I.; Hill, C.P. Structural and Biochemical Studies of ALIX/AIP1 and Its Role in Retrovirus Budding. Cell 2007, 128, 841–852. [CrossRef]
- Fujii, K.; Munshi, U.M.; Ablan, S.D.; Demirov, D.G.; Soheilian, F.; Nagashima, K.; Stephen, A.G.; Fisher, R.J.; Freed, E.O. Functional Role of Alix in Hiv-1 Replication. Virology 2009, 391, 284–292. [CrossRef]
- Gupta, S.; Bendjennat, M.; Saffarian, S. Abrogating ALIX Interactions Results in Stuttering of the ESCRT Machinery. Viruses 2020, 12, 1032. [CrossRef]
- Popov, S.; Popova, E.; Inoue, M.; Göttlinger, H.G. Human Immunodeficiency Virus Type 1 Gag Engages the Bro1 Domain of ALIX/AIP1 through the Nucleocapsid. J. Virol. 2008, 82, 1389–1398. [CrossRef]
- Dussupt, V.; Javid, M.P.; Abou-Jaoudé, G.; Jadwin, J.A.; Cruz, J. de L.; Nagashima, K.; Bouamr, F. The Nucleocapsid Region of HIV-1 Gag Cooperates with the PTAP and LYPXnL Late Domains to Recruit the Cellular Machinery Necessary for Viral Budding. PLOS Pathog. 2009, 5, e1000339. [CrossRef]
- Chung, H.-Y.; Morita, E.; von Schwedler, U.; Müller, B.; Kräusslich, H.-G.; Sundquist, W.I. NEDD4L Overexpression Rescues the Release and Infectivity of Human Immunodeficiency Virus Type 1 Constructs Lacking PTAP and YPXL Late Domains. J. Virol. 2008, 82, 4884–4897. [CrossRef]
- Mercenne, G.; Alam, S.L.; Arii, J.; Lalonde, M.S.; Sundquist, W.I. Angiomotin Functions in HIV-1 Assembly and Budding. eLife 2015, 4, e03778. [CrossRef]
- Rheinemann, L.; Thompson, T.; Mercenne, G.; Paine, E.L.; Peterson, F.C.; Volkman, B.F.; Alam, S.L.; Alian, A.; Sundquist, W.I. Interactions between AMOT PPxY Motifs and NEDD4L WW Domains Function in HIV-1 Release. J. Biol. Chem. 2021, 297, 100975. [CrossRef]
- Hurley, J.H.; Hanson, P.I. Membrane Budding and Scission by the ESCRT Machinery: It’s All in the Neck. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [CrossRef]
- Morita, E.; Sandrin, V.; McCullough, J.; Katsuyama, A.; Hamilton, I.B.; Sundquist, W.I. ESCRT-III Protein Requirements for HIV-1 Budding. Cell Host Microbe 2011, 9, 235–242. [CrossRef]
- Babst, M.; Wendland, B.; Estepa, E.J.; Emr, S.D. The Vps4p AAA ATPase Regulates Membrane Association of a Vps Protein Complex Required for Normal Endosome Function. EMBO J. 1998, 17, 2982–2993. [CrossRef]
- Lata, S.; Schoehn, G.; Jain, A.; Pires, R.; Piehler, J.; Gottlinger, H.G.; Weissenhorn, W. Helical Structures of ESCRT-III Are Disassembled by VPS4. Science 2008, 321, 1354–1357. [CrossRef]
- Wollert, T.; Wunder, C.; Lippincott-Schwartz, J.; Hurley, J.H. Membrane Scission by the ESCRT-III Complex. Nature 2009, 458, 172–177. [CrossRef]
- Watanabe, T.; Wang, S.; Kaibuchi, K. IQGAPs as Key Regulators of Actin-Cytoskeleton Dynamics. Cell Struct. Funct. 2015, 40, 69–77. [CrossRef]
- Abel, A.M.; Schuldt, K.M.; Rajasekaran, K.; Hwang, D.; Riese, M.J.; Rao, S.; Thakar, M.S.; Malarkannan, S. IQGAP1: Insights into the Function of a Molecular Puppeteer. Mol. Immunol. 2015, 65, 336–349. [CrossRef]
- Sabo, Y.; Santos, K. de los; Goff, S.P. IQGAP1 Negatively Regulates HIV-1 Gag Trafficking and Virion Production. Cell Rep. 2020, 30, 4065-4081.e4. [CrossRef]
- Barr, S.D.; Smiley, J.R.; Bushman, F.D. The Interferon Response Inhibits HIV Particle Production by Induction of TRIM22. PLoS Pathog. 2008, 4, e1000007. [CrossRef]
- Okumura, A.; Lu, G.; Pitha-Rowe, I.; Pitha, P.M. Innate Antiviral Response Targets HIV-1 Release by the Induction of Ubiquitin-like Protein ISG15. Proc. Natl. Acad. Sci. 2006, 103, 1440–1445. [CrossRef]
- Pincetic, A.; Kuang, Z.; Seo, E.J.; Leis, J. The Interferon-Induced Gene ISG15 Blocks Retrovirus Release from Cells Late in the Budding Process. J. Virol. 2010, 84, 4725–4736. [CrossRef]
- Woods, M.W.; Kelly, J.N.; Hattlmann, C.J.; Tong, J.G.; Xu, L.S.; Coleman, M.D.; Quest, G.R.; Smiley, J.R.; Barr, S.D. Human HERC5 Restricts an Early Stage of HIV-1 Assembly by a Mechanism Correlating with the ISGylation of Gag. Retrovirology 2011, 8, 95. [CrossRef]
- Umetsu, S.E.; Lee, W.-L.; McIntire, J.J.; Downey, L.; Sanjanwala, B.; Akbari, O.; Berry, G.J.; Nagumo, H.; Freeman, G.J.; Umetsu, D.T.; et al. TIM-1 Induces T Cell Activation and Inhibits the Development of Peripheral Tolerance. Nat. Immunol. 2005, 6, 447–454. [CrossRef]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a Phosphatidylserine Receptor. Nature 2007, 450, 435–439. [CrossRef]
- Kobayashi, N.; Karisola, P.; Peña-Cruz, V.; Dorfman, D.M.; Jinushi, M.; Umetsu, S.E.; Butte, M.J.; Nagumo, H.; Chernova, I.; Zhu, B.; et al. TIM-1 and TIM-4 Glycoproteins Bind Phosphatidylserine and Mediate Uptake of Apoptotic Cells. Immunity 2007, 27, 927–940. [CrossRef]
- Li, M.; Ablan, S.D.; Miao, C.; Zheng, Y.-M.; Fuller, M.S.; Rennert, P.D.; Maury, W.; Johnson, M.C.; Freed, E.O.; Liu, S.-L. TIM-Family Proteins Inhibit HIV-1 Release. Proc. Natl. Acad. Sci. 2014, 111, E3699–E3707. [CrossRef]
- Li, M.; Waheed, A.A.; Yu, J.; Zeng, C.; Chen, H.-Y.; Zheng, Y.-M.; Feizpour, A.; Reinhard, B.M.; Gummuluru, S.; Lin, S.; et al. TIM-Mediated Inhibition of HIV-1 Release Is Antagonized by Nef but Potentiated by SERINC Proteins. Proc. Natl. Acad. Sci. 2019, 116, 5705–5714. [CrossRef]
- Nasr, N.; Maddocks, S.; Turville, S.G.; Harman, A.N.; Woolger, N.; Helbig, K.J.; Wilkinson, J.; Bye, C.R.; Wright, T.K.; Rambukwelle, D.; et al. HIV-1 Infection of Human Macrophages Directly Induces Viperin Which Inhibits Viral Production. Blood 2012, 120, 778–788. [CrossRef]
- Wang, X.; Hinson, E.R.; Cresswell, P. The Interferon-Inducible Protein Viperin Inhibits Influenza Virus Release by Perturbing Lipid Rafts. Cell Host Microbe 2007, 2, 96–105. [CrossRef]
- Lim, E.S.; Wu, L.I.; Malik, H.S.; Emerman, M. The Function and Evolution of the Restriction Factor Viperin in Primates Was Not Driven by Lentiviruses. Retrovirology 2012, 9, 55. [CrossRef]
- McLaren, P.J.; Gawanbacht, A.; Pyndiah, N.; Krapp, C.; Hotter, D.; Kluge, S.F.; Götz, N.; Heilmann, J.; Mack, K.; Sauter, D.; et al. Identification of Potential HIV Restriction Factors by Combining Evolutionary Genomic Signatures with Functional Analyses. Retrovirology 2015, 12, 41. [CrossRef]
- Krapp, C.; Hotter, D.; Gawanbacht, A.; McLaren, P.J.; Kluge, S.F.; Stürzel, C.M.; Mack, K.; Reith, E.; Engelhart, S.; Ciuffi, A.; et al. Guanylate Binding Protein (GBP) 5 Is an Interferon-Inducible Inhibitor of HIV-1 Infectivity. Cell Host Microbe 2016, 19, 504–514. [CrossRef]
- Braun, E.; Hotter, D.; Koepke, L.; Zech, F.; Groß, R.; Sparrer, K.M.J.; Müller, J.A.; Pfaller, C.K.; Heusinger, E.; Wombacher, R.; et al. Guanylate-Binding Proteins 2 and 5 Exert Broad Antiviral Activity by Inhibiting Furin-Mediated Processing of Viral Envelope Proteins. Cell Rep. 2019, 27, 2092-2104.e10. [CrossRef]
- Sauter, D.; Hotter, D.; Van Driessche, B.; Stürzel, C.M.; Kluge, S.F.; Wildum, S.; Yu, H.; Baumann, B.; Wirth, T.; Plantier, J.-C.; et al. Differential Regulation of NF-κB-Mediated Proviral and Antiviral Host Gene Expression by Primate Lentiviral Nef and Vpu Proteins. Cell Rep. 2015, 10, 586–599. [CrossRef]
- Schubert, U.; Bour, S.; Willey, R.L.; Strebel, K. Regulation of Virus Release by the Macrophage-Tropic Human Immunodeficiency Virus Type 1 AD8 Isolate Is Redundant and Can Be Controlled by Either Vpu or Env. J. Virol. 1999, 73, 887–896. [CrossRef]
- Tada, T.; Zhang, Y.; Koyama, T.; Tobiume, M.; Tsunetsugu-Yokota, Y.; Yamaoka, S.; Fujita, H.; Tokunaga, K. MARCH8 Inhibits HIV-1 Infection by Reducing Virion Incorporation of Envelope Glycoproteins. Nat. Med. 2015, 21, 1502–1507. [CrossRef]
- Eyster, C.A.; Cole, N.B.; Petersen, S.; Viswanathan, K.; Früh, K.; Donaldson, J.G. MARCH Ubiquitin Ligases Alter the Itinerary of Clathrin-Independent Cargo from Recycling to Degradation. Mol. Biol. Cell 2011, 22, 3218–3230. [CrossRef]
- Chen, R.; Li, M.; Zhang, Y.; Zhou, Q.; Shu, H.-B. The E3 Ubiquitin Ligase MARCH8 Negatively Regulates IL-1β-Induced NF-κB Activation by Targeting the IL1RAP Coreceptor for Ubiquitination and Degradation. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 14128–14133. [CrossRef]
- Tada, T.; Zhang, Y.; Fujita, H.; Tokunaga, K. MARCH8: The Tie That Binds to Viruses. FEBS J. 2022, 289, 3642–3654. [CrossRef]
- Zhang, Y.; Tada, T.; Ozono, S.; Yao, W.; Tanaka, M.; Yamaoka, S.; Kishigami, S.; Fujita, H.; Tokunaga, K. Membrane-Associated RING-CH (MARCH) 1 and 2 Are MARCH Family Members That Inhibit HIV-1 Infection. J. Biol. Chem. 2019, 294, 3397–3405. [CrossRef]
- Zhang, Y.; Lu, J.; Liu, X. MARCH2 Is Upregulated in HIV-1 Infection and Inhibits HIV-1 Production through Envelope Protein Translocation or Degradation. Virology 2018, 518, 293–300. [CrossRef]
- Van Damme, N.; Goff, D.; Katsura, C.; Jorgenson, R.L.; Mitchell, R.; Johnson, M.; Stephens, E.B.; Guatelli, J. The Interferon-Induced Protein BST-2/CD317 Restricts Release of Virions from Infected Cells and Is down-Regulated from the Cell Surface by HIV-1 Vpu. Cell Host Microbe 2008, 3, 245–252. [CrossRef]
- Neil, S.J.D.; Zang, T.; Bieniasz, P.D. Tetherin Inhibits Retrovirus Release and Is Antagonized by HIV-1 Vpu. Nature 2008, 451, 425–430. [CrossRef]
- Fitzpatrick, K.; Skasko, M.; Deerinck, T.J.; Crum, J.; Ellisman, M.H.; Guatelli, J. Direct Restriction of Virus Release and Incorporation of the Interferon-Induced Protein BST-2 into HIV-1 Particles. PLoS Pathog. 2010, 6, e1000701. [CrossRef]
- Perez-Caballero, D.; Zang, T.; Ebrahimi, A.; McNatt, M.W.; Gregory, D.A.; Johnson, M.C.; Bieniasz, P.D. Tetherin Inhibits HIV-1 Release by Directly Tethering Virions to Cells. Cell 2009, 139, 499–511. [CrossRef]
- Kupzig, S.; Korolchuk, V.; Rollason, R.; Sugden, A.; Wilde, A.; Banting, G. Bst-2/HM1.24 Is a Raft-Associated Apical Membrane Protein with an Unusual Topology. Traffic Cph. Den. 2003, 4, 694–709. [CrossRef]
- Hinz, A.; Miguet, N.; Natrajan, G.; Usami, Y.; Yamanaka, H.; Renesto, P.; Hartlieb, B.; McCarthy, A.A.; Simorre, J.-P.; Göttlinger, H.; et al. Structural Basis of HIV-1 Tethering to Membranes by the BST-2/Tetherin Ectodomain. Cell Host Microbe 2010, 7, 314–323. [CrossRef]
- Venkatesh, S.; Bieniasz, P.D. Mechanism of HIV-1 Virion Entrapment by Tetherin. PLOS Pathog. 2013, 9, e1003483. [CrossRef]
- Jolly, C.; Booth, N.J.; Neil, S.J.D. Cell-Cell Spread of Human Immunodeficiency Virus Type 1 Overcomes Tetherin/BST-2-Mediated Restriction in T Cells. J. Virol. 2010, 84, 12185–12199. [CrossRef]
- Galão, R.P.; Le Tortorec, A.; Pickering, S.; Kueck, T.; Neil, S.J.D. Innate Sensing of HIV-1 Assembly by Tetherin Induces NFκB-Dependent Proinflammatory Responses. Cell Host Microbe 2012, 12, 633–644. [CrossRef]
- Hotter, D.; Sauter, D.; Kirchhoff, F. Emerging Role of the Host Restriction Factor Tetherin in Viral Immune Sensing. J. Mol. Biol. 2013, 425, 4956–4964. [CrossRef]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS Pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [CrossRef]
- Todd, S.; Laboissière, M.C.; Craik, C.S. Yeast Two-Hybrid Assay for Examining Human Immunodeficiency Virus Protease Heterodimer Formation with Dominant-Negative Inhibitors and Multidrug-Resistant Variants. Anal. Biochem. 2000, 277, 247–253. [CrossRef]
- Danielson, C.M.; Hope, T.J. Imaging of HIV/Host Protein Interactions. Curr. Top. Microbiol. Immunol. 2009, 339, 103–123. [CrossRef]
- Saffarian, S. Application of Advanced Light Microscopy to the Study of HIV and Its Interactions with the Host. Viruses 2021, 13, 223. [CrossRef]
- Luo, Y.; Muesing, M.A. Mass Spectrometry-Based Proteomic Approaches for Discovery of HIV–Host Interactions. Future Virol. 2014, 9, 979–992. [CrossRef]
- Knoener, R.A.; Becker, J.T.; Scalf, M.; Sherer, N.M.; Smith, L.M. Elucidating the in Vivo Interactome of HIV-1 RNA by Hybridization Capture and Mass Spectrometry. Sci. Rep. 2017, 7, 16965. [CrossRef]
- van Manen, D.; van ‘t Wout, A.B.; Schuitemaker, H. Genome-Wide Association Studies on HIV Susceptibility, Pathogenesis and Pharmacogenomics. Retrovirology 2012, 9, 70. [CrossRef]
- Montoya, V.R.; Ready, T.M.; Felton, A.; Fine, S.R.; OhAinle, M.; Emerman, M. A Virus-Packageable CRISPR System Identifies Host Dependency Factors Co-Opted by Multiple HIV-1 Strains. mBio 2023, 14, e0000923. [CrossRef]
- Deshiere, A.; Joly-Beauparlant, C.; Breton, Y.; Ouellet, M.; Raymond, F.; Lodge, R.; Barat, C.; Roy, M.-A.; Corbeil, J.; Tremblay, M.J. Global Mapping of the Macrophage-HIV-1 Transcriptome Reveals That Productive Infection Induces Remodeling of Host Cell DNA and Chromatin. Sci. Rep. 2017, 7, 5238. [CrossRef]
- Zhou, H.; Xu, M.; Huang, Q.; Gates, A.T.; Zhang, X.D.; Castle, J.C.; Stec, E.; Ferrer, M.; Strulovici, B.; Hazuda, D.J.; et al. Genome-Scale RNAi Screen for Host Factors Required for HIV Replication. Cell Host Microbe 2008, 4, 495–504. [CrossRef]
Figure 1.
Host factors interact with the HIV-1 capsid to transport it across the cytoplasm it to the nucleus, through the nuclear pore complex, and to facilitate proper uncoating. (1) After attachment and envelope fusion with the plasma membrane, the capsid is released into the cytoplasm. Factors found within the capsid (CypA, red spheres; IP6, green spheres) as well as outside of the capsid (Sec24c, yellow spheres; PDZD8, blue spheres) bind the capsid to promote stability during cytoplasmic transport. Restriction factors (TRIM5α, black squares; TRIM11, purple squares; PQBP1, red triangles) facilitate capsid disassembly and/or immune detection of the viral genome. Transport of the capsid can occur in a microtubule-dependent (2a) or microtubule-independent (2b) manner. Using the adaptor protein BICD2, the microtubule-associated protein dynein moves the capsid inwards towards the nucleus; FEZ1 and kinesin-1 (not shown), though outwards-bound from the nucleus, still positively impact capsid localization to the nucleus (see refs. 97 and 98). (3) At the nucleus, capsid entry through the nuclear pore complex is aided by host factors (Nup358, Nup 153, CPSF6) and prevented by restriction factors (MX2). (4) Once inside the nucleus, host factors promote capsid destabilization and disassembly (TNPO1, TNPO3, Pin1, CypA), ensuring proper viral genome release, followed by localization to sites of open chromatin by CPSF6. At the same time, restriction factors (MX2, Daxx) will attempt to prevent uncoating and genome release.
Figure 1.
Host factors interact with the HIV-1 capsid to transport it across the cytoplasm it to the nucleus, through the nuclear pore complex, and to facilitate proper uncoating. (1) After attachment and envelope fusion with the plasma membrane, the capsid is released into the cytoplasm. Factors found within the capsid (CypA, red spheres; IP6, green spheres) as well as outside of the capsid (Sec24c, yellow spheres; PDZD8, blue spheres) bind the capsid to promote stability during cytoplasmic transport. Restriction factors (TRIM5α, black squares; TRIM11, purple squares; PQBP1, red triangles) facilitate capsid disassembly and/or immune detection of the viral genome. Transport of the capsid can occur in a microtubule-dependent (2a) or microtubule-independent (2b) manner. Using the adaptor protein BICD2, the microtubule-associated protein dynein moves the capsid inwards towards the nucleus; FEZ1 and kinesin-1 (not shown), though outwards-bound from the nucleus, still positively impact capsid localization to the nucleus (see refs. 97 and 98). (3) At the nucleus, capsid entry through the nuclear pore complex is aided by host factors (Nup358, Nup 153, CPSF6) and prevented by restriction factors (MX2). (4) Once inside the nucleus, host factors promote capsid destabilization and disassembly (TNPO1, TNPO3, Pin1, CypA), ensuring proper viral genome release, followed by localization to sites of open chromatin by CPSF6. At the same time, restriction factors (MX2, Daxx) will attempt to prevent uncoating and genome release.
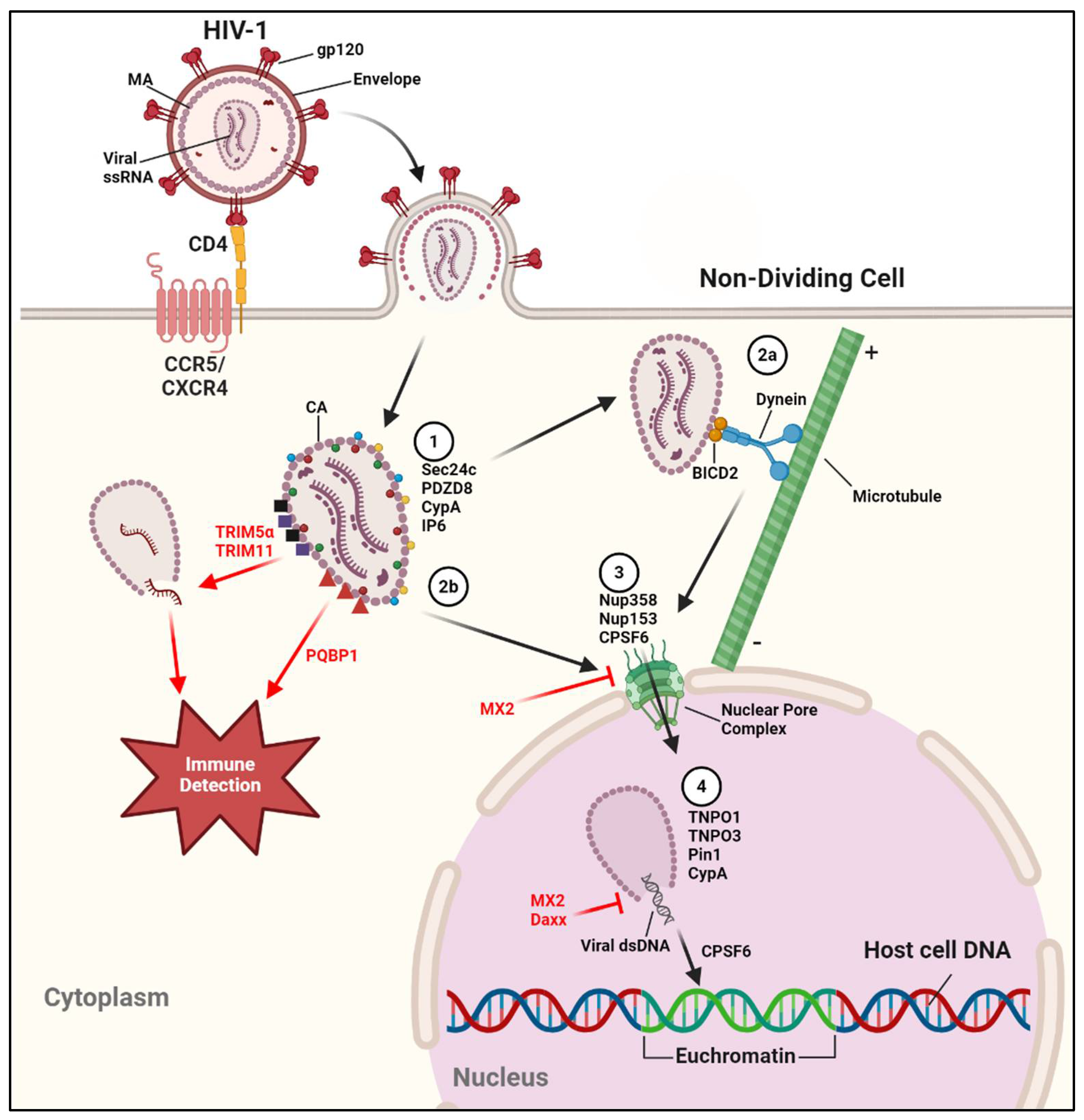
Figure 2.
Various components of the Gag polyprotein interact with host factors to facilitate recruitment of ESCRT pathway machinery, leading to productive and efficient virion budding. See text for further details.
Figure 2.
Various components of the Gag polyprotein interact with host factors to facilitate recruitment of ESCRT pathway machinery, leading to productive and efficient virion budding. See text for further details.
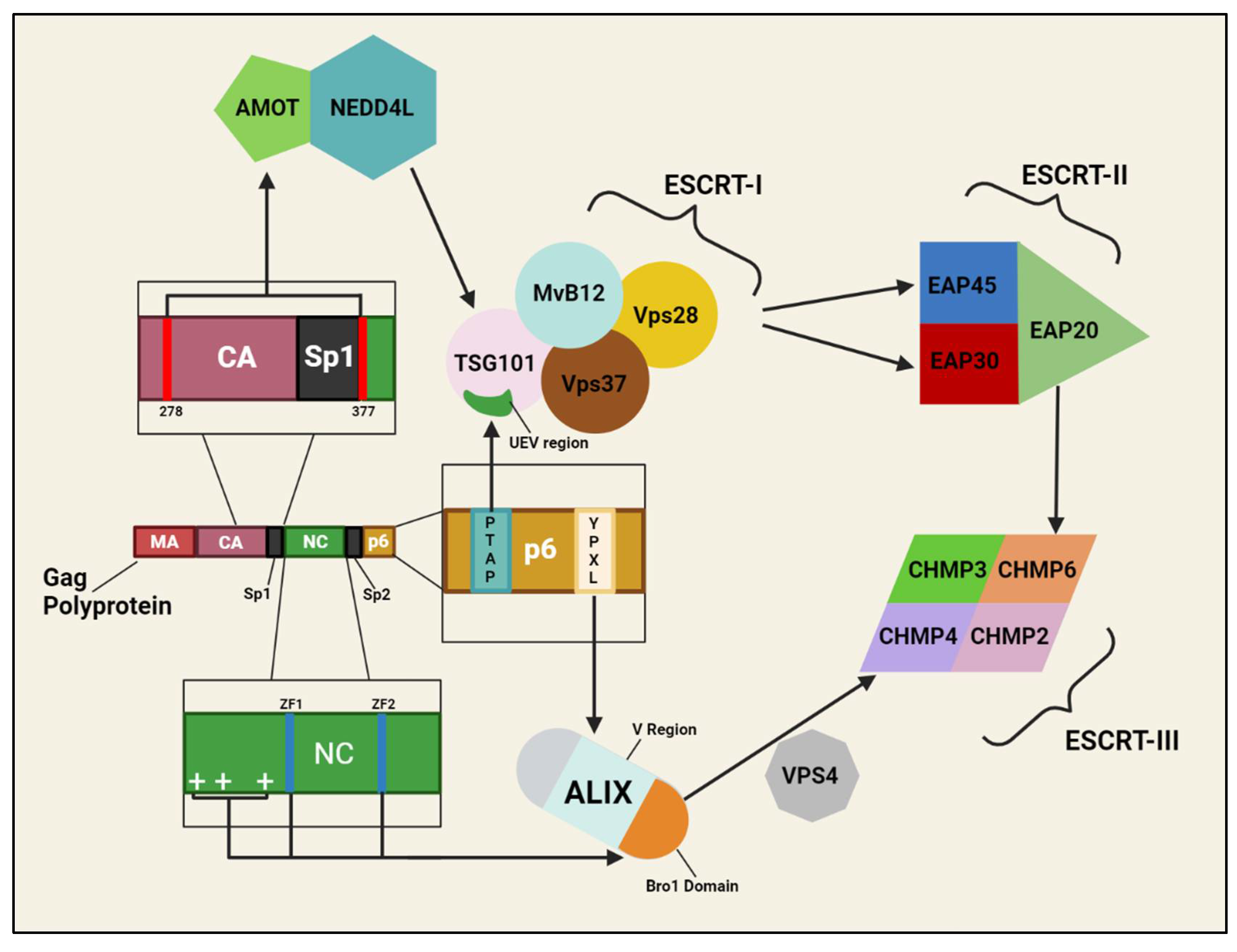
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



